developmental origins of aggression
TRANSCRIPT


DEVELOPMENTAL ORIGINS OF AGGRESSION

This page intentionally left blank

DEVELOPMENTAL ORIGINSOF AGGRESSION
Edited by
RICHARD E. TREMBLAYWILLARD W. HARTUP
JOHN ARCHER
THE GUILFORD PRESSNew York London

© 2005 The Guilford PressA Division of Guilford Publications, Inc.72 Spring Street, New York, NY 10012www.guilford.com
All rights reserved
No part of this book may be reproduced, translated, stored ina retrieval system, or transmitted, in any form or by any means,electronic, mechanical, photocopying, microfilming, recording,or otherwise, without written permission from the Publisher.
Printed in the United States of America
This book is printed on acid-free paper.
Last digit is print number: 9 8 7 6 5 4 3 2 1
Library of Congress Cataloging-in-Publication Data
Developmental origins of aggression / edited by Richard E. Tremblay, WillardW. Hartup, John Archer.
p. cm.Includes bibliographical references and index.ISBN 1-59385-110-3 (hardcover : alk. paper)
1. Aggressiveness. 2. Aggressiveness—Longitudinal studies. 3. Childpsychology. I. Tremblay, Richard Ernest. II. Hartup, Willard W. III. Archer,John, 1944–
BF575.A3D45 2005155.2′32—dc22
2004025158

In memory ofJoan McCord(1930–2004),
colleague, role model, and friend
(left to right): Richard E. Tremblay, Joan McCord, Willard W. Hartup, and John Archer.

This page intentionally left blank

About the EditorsAbout the Editors
About the Editors
Richard E. Tremblay, PhD, is Professor of Pediatrics, Psychiatry, and Psychologyat the University of Montreal and Director of the Inter-University ResearchUnit on Children’s Psychosocial Maladjustment. He is also Director of theCentre of Excellence for Early Childhood Development, Canada ResearchChair in Child Development, Molson Fellow of the Canadian Institute forAdvanced Research, Fellow of the Academy of Experimental Criminology, andFellow of the Royal Society of Canada. For over 20 years, he has conducted aprogram of longitudinal and experimental studies addressing the physical,cognitive, emotional, and social development of children from conceptiononward to understand the development and prevention of antisocial behavior.
Willard W. Hartup, EdD, is Regents’ Professor Emeritus and former Directorof the Institute of Child Development at the University of Minnesota. Dr.Hartup has spent many years researching friendship and peer relations in childdevelopment, antipathies and their significance, and conflict and aggression inchildhood and adolescence. He obtained an EdD degree from Harvard Uni-versity, and has received the G. Stanley Hall Award for DistinguishedContributions to Developmental Psychology from the American PsychologicalAssociation and Distinguished Scientific Contribution Awards from both theSociety for Research in Child Development and the International Society forthe Study of Behavioural Development.
John Archer, PhD, is Professor of Psychology at the University of CentralLancashire, Preston, United Kingdom. He received a PhD from the Universityof Bristol in 1970. A Fellow of the British Psychological Society and Presidentof the International Society for Research on Aggression, his research is con-cerned with human aggression, grief and loss, and sex differences. Dr. Archeris also author of several books, including Sex and Gender (with Barbara Lloyd)and The Nature of Grief; numerous book chapters; and over 100 articles inrefereed journals covering psychology, medicine, and biology. In recent years,he has published a number of meta-analytic reviews on topics connected withsex differences in aggression.
vii

This page intentionally left blank

ContributorsContributors
Contributors
John Archer, PhD, Department of Psychology, University of CentralLancashire, Preston, United Kingdom
Chawki Benkelfat, MD, Department of Psychiatry, McGill University,Montreal, Quebec, Canada
Michel Boivin, PhD, Research Unit on Children’s PsychosocialMaladjustment, School of Psychology, Laval University, Quebec City,Quebec, Canada
Mara Brendgen, PhD, Department of Psychology, University of Quebec atMontreal, Montreal, Quebec, Canada
Sylvana Côté, PhD, School of Psychoeducation, University of Montreal,Montreal, Quebec, Canada
Ginette Dionne, PhD, School of Psychology, Laval University, Quebec City,Quebec, Canada
Joseph L. Flanders, BSc, Department of Psychology, McGill University,Montreal, Quebec, Canada
Afra Foroud, MSc, Canadian Centre for Behavioural Neuroscience,Department of Psychology and Neuroscience, University of Lethbridge,Lethbridge, Alberta, Canada
Uberto Gatti, MD, Section of Criminology and Forensic Psychiatry,University of Genoa, Genoa, Italy
Paul L. Gendreau, PhD, School of Psychoeducation, University of Montreal,Montreal, Quebec, Canada
Willard W. Hartup, EdD, Institute of Child Development, University ofMinnesota, Minneapolis, Minnesota
Dale F. Hay, PhD, School of Psychology, Cardiff University, Cardiff, Wales,United Kingdom
ix

D. Lynn Homish, BSc, Western Psychiatric Institute and Clinic, Universityof Pittsburgh Medical Center, Pittsburgh, Pennsylvania
David Joubert, MA, MCA, Department of Psychology, University ofQuebec at Montreal, Montreal, Quebec, Canada
Eric Lacourse, PhD, Department of Sociology, University of Montreal,Montreal, Quebec, Canada
Rolf Loeber, PhD, Western Psychiatric Institute and Clinic, University ofPittsburgh Medical Center, Pittsburgh, Pennsylvania
Tania Mazzarello, BSc, Department of Psychology, University of Quebec atMontreal, Montreal, Quebec, Canada
Daniel S. Nagin, PhD, H. J. Heinz III School of Public Policy andManagement, Carnegie Mellon University, Pittsburgh, Pennsylvania
Tomá� Paus, MD, PhD, Departments of Neurology and Neurosurgery, andPsychology, McGill University, Montreal, Quebec, Canada
Sergio M. Pellis, PhD, Canadian Centre for Behavioural Neuroscience,Department of Psychology and Neuroscience, University of Lethbridge,Lethbridge, Alberta, Canada
Vivien C. Pellis, PhD, Canadian Centre for Behavioural Neuroscience,Department of Psychology and Neuroscience, University of Lethbridge,Lethbridge, Alberta, Canada
Daniel Pérusse, PhD, Department of Anthropology, University of Montreal,Montreal, Quebec, Canada
Jordan B. Peterson, PhD, Department of Psychology, University of Toronto,Toronto, Ontario, Canada
Robert O. Pihl, PhD, Departments of Psychology and Psychiatry, McGillUniversity, Montreal, Quebec, Canada
François Poulin, PhD, Department of Psychology, University of Quebec atMontreal, Montreal, Quebec, Canada
Elisa Romano, PhD, Department of Psychology, University of Ottawa,Ottawa, Ontario, Canada
Jean Richard Séguin, PhD, Research Unit on Children’s PsychosocialMaladjustment, Department of Psychiatry, University of Montreal,Montreal, Quebec, Canada
Stephen J. Suomi, PhD, Laboratory of Comparative Ethology, NationalInstitute of Child Health and Human Development, National Institutesof Health, Bethesda, Maryland
Richard E. Tremblay, PhD, Departments of Pediatrics, Psychiatry, andPsychology, University of Montreal, Montreal, Quebec, Canada
x Contributors

Tracy Vaillancourt, PhD, Department of Psychology, McMaster University,Hamilton, Ontario, Canada
Stephanie H. M. Van Goozen, PhD, School of Psychology, CardiffUniversity, Cardiff, Wales, United Kingdom
Frank Vitaro, PhD, Research Unit on Children’s PsychosocialMaladjustment, School of Psychoeducation, University of Montreal,Montreal, Quebec, Canada
Philip David Zelazo, PhD, Department of Psychology, University ofToronto, Toronto, Ontario, Canada
Mark Zoccolillo, MD, Department of Psychiatry, Montreal Children’sHospital, Montreal, Quebec, Canada
Contributors xi

This page intentionally left blank

PrefacePreface
Preface
It has been over 30 years since the publication of a book giving a detailedoverview of the state of knowledge on the developmental origins of aggressivebehavior (de Wit & Hartup, 1974). Much has changed during this time. Mostof the developmental work published in the 1974 book was based on cross-sectional studies of small samples of school-age children. The focus of atten-tion was on aggressive events and the situations that instigate them.
Over the past 30 years, investigators have conducted longitudinal studiesof large samples of children assessed repeatedly from birth to adulthood. Re-sults from these studies are telling unexpected stories. For example, childrenappear not to be learning to use physical aggression as they grow older; ratherthey appear to be learning not to use physical aggression. As expected, girlsuse physical aggression less often than boys from infancy to adolescence; how-ever, they use another form of aggression (indirect aggression) more often thanboys from early childhood to adulthood.
Such findings are clearly of interest for our general understanding of hu-man nature. Philosophers have argued for centuries on the origins of antiso-cial behavior. Rousseau defended the thesis that humans are created good andbecome evil through the influence of society, and Hobbes argued that awicked person was simply a child who had not grown up. In 1973, AlbertBandura wrote, “People are not born with preformed repertoires of aggressivebehavior; they must learn them in one way or another” (p. 61).
The findings from research on the development of aggressive behavior arealso extremely important for the prevention and treatment of violent behavior.Violence is still a serious problem in our modern societies. To help childrenlearn alternatives to violent behavior, we need to understand the developmentof aggression and its alternatives. Every professional who works with childrenand adolescents needs to understand the developmental origins of aggression.
Technological progress over the past 30 years has enabled investigators tocombine longitudinal studies of large samples of children over their lifespanswith molecular assessments of their genetic material and assessments of
xiii

hormones and neuromodulators through blood and saliva samples and brainimaging. More sophisticated statistical techniques provide researchers withtools to make better use of twin samples and repeated measurements overlong periods of time.
This volume provides a timely overview of the best research and the keyquestions at the beginning of a new era, which will lead to a synthesis of ge-netic, brain, behavior, and environmental research.
REFERENCES
Bandura, A. (1973). Aggression: A social learning analysis. New York: Holt.de Wit, J., & Hartup, W. W. (Eds.). (1974). Determinants and origins of aggressive be-
havior. The Hague: Mouton.
xiv Preface

ContentsContents
Contents
I. INTRODUCTION
1 The Development of Aggression: Where Do We Stand? 3Willard W. Hartup
II. THE DEVELOPMENT OF AGGRESSION IN ANIMALS AND HUMANS
2 Subtypes of Aggression in Humans and Animals 25Paul L. Gendreau and John Archer
3 Play Fighting: Aggression, Affiliation, and the Developmentof Nuanced Social Skills
47
Sergio M. Pellis, Vivien C. Pellis, and Afra Foroud
4 Genetic and Environmental Factors Influencing the Expressionof Impulsive Aggression and Serotonergic Functioningin Rhesus Monkeys
63
Stephen J. Suomi
5 The Developmental Origins of Physical Aggression in Humans 83Richard E. Tremblay and Daniel S. Nagin
6 The Beginnings of Aggression in Infancy 107Dale F. Hay
7 Play and the Regulation of Aggression 133Jordan B. Peterson and Joseph L. Flanders
8 Indirect Aggression among Humans: Social Constructor Evolutionary Adaptation?
158
Tracy Vaillancourt
9 Proactive and Reactive Aggression:A Developmental Perspective
178
Frank Vitaro and Mara Brendgen
10 Homicide, Violence, and Developmental Trajectories 202Rolf Loeber, Eric Lacourse, and D. Lynn Homish
xv

III. DETERMINANTS OF AGGRESSION
11 Genetics and the Development of Aggression 223Daniel Pérusse and Paul L. Gendreau
12 Mapping Brain Development and Aggression 242Tomá� Paus
13 Neuromodulators in the Development and Expressionof Inhibition and Aggression
261
Robert O. Pihl and Chawki Benkelfat
14 Hormones and the Developmental Origins of Aggression 281Stephanie H. M. Van Goozen
15 Executive Function in Early Physical Aggression 307Jean Richard Séguin and Philip David Zelazo
16 Language Development and Aggressive Behavior 330Ginette Dionne
17 The Intergenerational Transmission of Aggressionand Antisocial Behavior
353
Mark Zoccolillo, Elisa Romano, David Joubert,Tania Mazzarello, Sylvana Côté, Michel Boivin,Daniel Pérusse, and Richard E. Tremblay
18 Peer Relationships and the Development of AggressiveBehavior in Early Childhood
376
Michel Boivin, Frank Vitaro, and François Poulin
19 Social Capital and Physical Violence 398Uberto Gatti and Richard E. Tremblay
20 Sex Differences in Aggressive Behavior:A Developmental and Evolutionary Perspective
425
John Archer and Sylvana Côté
IV. CHALLENGES FOR THE FUTURE
21 The Developmental Origins of Aggression:Where Are We Going?
447
Richard E. Tremblay and Sylvana Côté
Index 465
xvi Contents

Part I
INTRODUCTION

This page intentionally left blank

INTRODUCTIONThe Development of Aggression
1
The Development of AggressionWhere Do We Stand?
WILLARD W. HARTUP
Three changes have occurred in research on the development of aggression.First, both theoretical and empirical attention have shifted from the aggressiveact, on one hand, to the development of aggressive individuals, on the other;that is, from aggressive events and the situations that instigate them to aggres-sive children assessed within ecological systems. Investigators no longer exam-ine the effects of exposure to aggressive models (Bandura & Walters, 1963),for example, but rather the developmental consequences of reciprocities in socialinteraction involving individuals, bilateral and transactional processes in fam-ily and peer interaction, gene–environment permutations, and contextual com-ponents (including exposure to aggression and violence in the community).
A second change is the emerging synthesis between research on aggres-sion and research on antisocial behavior (Tremblay, 2000). Everyone knowsthat aggression and antisocial behavior are overlapping but not identical con-structs. Nevertheless, because aggressive behavior is relatively stable as an in-dividual characteristic, and because stable aggression predicts antisocial be-havior during adolescence and adulthood for males at least (Moffitt, Caspi,Rutter, & Silva, 2001), it no longer seems wise to separate these terms inthinking about social development and adaptation.
Third, a trend toward more developmentally oriented studies of aggres-sion has emerged. What was known about aggression in childhood and ado-lescence in the late 1960s was quite nondevelopmental: The relevant workdealt with contingencies in observation or social interaction that increased ag-gression in the child, usually in constrained situations and with little attentionto age differences. Normative studies encompassed only a few years (in early
3

childhood), and the events associated with individual differences in the realworld were not much remarked on (see Feshbach, 1970).
To be concerned with development, however, is to be concerned with con-tinuities and discontinuities in harm-doing activity across the lifespan, espe-cially during the years before maturity, along with changes across time in themorphology and functioning of aggressive behavior, the reorganization of var-ious mental structures as these involve aggression, and the role of violence inthe development of attitudes toward oneself and others (Hartup & DeWit,1974).
By the 1980s, new ideas were percolating through the entire field ofsocial and emotional development even though long-term longitudinal datawere scarce. Studies linking aggression in early childhood with aggression inadolescence and adulthood were extremely rare; even cross-sectional studieswere not abundant. But the zeitgeist was shifting. Ross Parke and RonaldSlaby (1983) knew it, and called their chapter for the Handbook of Child Psy-chology, “The Development of Aggression.” Ahead of its time, this chapterled the way to fundamental changes in research and research strategy high-lighting developmental processes in the emergence of aggression and antisocialbehavior. Nowadays, the major studies of aggression and antisocial behaviorare definitely developmental, and most use longitudinal data to search for de-terminants, dynamics, and outcomes of these behaviors.
Against the background of these comments, several issues are now dis-cussed in terms of how things have changed and what we might expect in thenear future in understanding the development of aggression: First, whatchanges? Three issues are considered: (1) construct selection, (2) normativechange versus individual differences, and (3) child development versus lifespanperspectives. Second, what are the determinants of developmental change inaggression and antisocial behavior? Again, three issues are addressed: (1) na-tivism versus empiricism, (2) socialization and relationships, and (3) the eco-logical context.
WHAT CHANGES?
Aggression, like most human activity, changes with age both in its incidenceand in the processes leading to its instigation. But which changes are meaning-ful? Not every change in language and cognitive activity at every level of anal-ysis is meaningful, nor is every change in aggression and antisocial behavior.Our first task, then, is to identify those changeworthy behaviors that havesome significance in behavioral adaptation across time.
Construct Selection
One must proceed cautiously through the thicket of constructs available in thecurrent aggression literature. Merely specifying a workable construct that can
4 I N T R O D U C T I O N

be called “aggression” is difficult. Saying that aggression is “intentional harmdoing” or “harm doing for its own sake” may be reasonable for the ordinaryuser of the English language, but it is fraught with difficulties for scientistswho want to describe relevant phenomena with precision: Intentions cannotbe observed easily; instigating conditions are difficult to separate from thestructure of the aggressive act; outcomes are difficult to specify; and we can’talways tell whether the act has been aversive to the victim (Bandura, 1973;Hartup & De Wit, 1974; Coie & Dodge, 1998; Tremblay, 2000). Moreover,similar measures across age (if not equivalent ones) must be used in manyforms of developmental analysis (e.g., specifying developmental trajectoriesand pathways), and one cannot always count on this equivalence.
Researchers have made much progress by becoming more global in defin-ing aggression and antisocial behavior, that is, by making increasingly greateruse of global constructs drawn from natural language. Many investigators relyon ordinary observers (parents, teachers, and children) to tell them when ag-gression occurs and what some of its implications are. For example, elemen-tary school children are asked to tell us which classmates “start fights.” Whenthey answer, the reliability and validity of these nominations turn out to besubstantial even though we do not know the circumstances under which thetarget child may start fights, the behaviors used in fighting, the consequencesto the child’s companions, or the goals achieved by the target child.
At certain times, however, these “down and dirty” measurement strate-gies will not work, that is, for determining whether babies “start fights” orwhether 3-year-old and 13-year-old aggression reflect the same functional dy-namics. Particulars about instigation, motor patterns, and consequences(functions) are needed in those instances even if it is difficult to obtain them.
One endeavor needs to be encouraged strongly: construct selection at in-termediate levels of conceptual complexity. First, certain older constructs needto be maintained. Consider that physical and verbal aggression have been dis-tinguished from one another for many years (Goodenough, 1931), and severalexamples will show that this distinction remains useful:
1. Neither physical aggression nor most forms of antisocial behavior oc-cur as frequently among females as among males, and gender differ-ences in aggression cannot be understood without taking this into con-sideration.
2. Physical aggression among females during childhood does not predictadolescent antisocial behavior as it does for males (Broidy et al.,2003), and we do not completely understand this.
Other midlevel constructs must be preserved, for example, violent versusnonviolent aggression, instrumental versus hostile aggression, and proactiveversus reactive aggression. Bullying and victimization are also uniquely usefulconstructs. It may be time to resurrect other old constructs such as prosocialversus antisocial aggression; this one has languished since Robert Sears and
The Development of Aggression 5

his colleagues could not find differential correlates in the doll play of kinder-garten children more than 40 years ago (Maccoby, 1992). But we must putthis old wine in new bottles.
New midlevel constructs are badly needed. Relational (or indirect) ag-gression has evoked considerable interest in the last decade (Crick et al., 1998;Vaillancourt, Chapter 8, this volume) Research shows that relational aggres-sion is more frequent among females than among males, although its sequelaeare not remarkably different (Crick et al., 1998; Vaillancourt, Chapter 8, thisvolume). It is not really surprising that harm doing may occur in normativelydifferent ways in males and females; it is only surprising that it took us so longto realize this.
Natural language may or may not furnish us with the complete concep-tual armamentarium needed for research on aggression and antisocial behav-ior. As a consequence, the research community needs to look beyond the dic-tionary for constructs that will invigorate their work.
Normative Change or Individual Differences?
Once upon a time, the following item appeared regularly on the exams for mygraduate course: “Discuss the similarities and differences between the study ofnormative issues in developmental psychology and individual differences:Demonstrate how each of these issues informs the other about developmentalprocess.” Historically, developmental psychology has been much concernedwith both issues, with students of cognitive development emphasizing norma-tive continuities and change rather than the emergence of individual differ-ences (except for investigators interested in the IQ); students of social and per-sonality development, however, have been interested mainly in individualdifferences.
Normative change, of course, refers to age-related changes that are uni-versal or characteristic of major population groups rather than developmentalvariations within or between groups. Aggression researchers, especially earlierones, were interested in normative questions ranging from the ontogeny of an-ger regulation (Goodenough, 1931), to delinquency (Glueck & Glueck, 1950),to verbal mediation in the child’s response to aggressive modeling (Coates &Hartup, 1969).
Contemporary researchers remain interested in normative assessment ofaggression, as revealed in Figure 1.1 which shows data on hitting, biting, andkicking in a Canadian sample (Tremblay et al., 1996). These behaviors peakat about 27 months for both boys and girls, with slightly more boys than girlsshowing them “sometimes” or “frequently.” Frequencies decline steadilythereafter and are not often evident (but not completely absent) by age 12.These data represent one of the most comprehensive normative reports onphysical aggression existing anywhere.
Normative studies, however, are increasingly difficult to distinguish fromdifferential studies of aggression. Normative and differential research on ag-
6 I N T R O D U C T I O N

gression have blurred together, underscoring something we have known allalong but have chosen to ignore: Most normative changes in aggression areconstrained by variations among individuals. Recent studies show dramati-cally that individual-to-individual variations in developmental pathways areubiquitous. Some of these developmental variations are associated with gen-der, social class, and ethnicity. Indeed, one can describe many relevant differ-ences between the sexes in developmental trajectories, even though the basicprocesses by which children acquire certain types of aggression may not differbetween boys and girls (Moffitt et al., 2001).
Even among children of a single race or gender, however, normativechanges encompass a range of individual differences. Nowhere is this conclu-sion more clearly revealed than in a recent manuscript that brings togetherdata from six well-known longitudinal studies (Broidy et al., 2003). In everyinstance, semiparametric analyses reveal multiple types of developmentalchange. For example, four trajectories in physical aggression can be identifiedwithin a male sample from Montreal studied by Nagin and Tremblay (1999;Tremblay & Nagin, Chapter 5, this volume). Two are similar to those identi-fied in a number of longitudinal studies: (1) high levels of aggression among arelatively small group of boys that persist across childhood and into adoles-cence and (2) low aggression that persists across childhood and characterizes amuch larger group. Two other trajectories were also identified in these data:
The Development of Aggression 7
FIGURE 1.1. Hitting, biting, and kicking (boys and girls ages 2–11 years). From Tremblay etal. (1996). Statistics Canada information is used with the permission of the Minister of In-dustry, as Minister responsible for Statistics Canada. Information on the availability of thewide range of data from Statistics Canada can be obtained from Statistics Canada’s Re-gional Offices, its website at www.statcan.ca, and its toll-free access number 1-800-263-1136.

(3) one that begins with high levels of aggression and declines steadily as timegoes on (boys sometimes called “desistors” or “recovered” aggressors) and (4)one that begins at more moderate levels and also declines.
One other example included in this six-site investigation, the so-calledDunedin (New Zealand) studies of Moffitt and her associates (Moffitt, Caspi,Rutter, & Silva, 1996; Broidy, et al., 2003), revealed three distinct trajectoriesinstead of four: (1) a small group of consistently aggressive boys, (2) a largegroup of consistently nonaggressive boys, and (3) another stable group ofmoderately aggressive boys. No cluster of decliners was identified in thisstudy. The remaining studies in the investigation show either three- or four-cluster patterns, with only one (based on a sample from the United States)showing a trajectory of physical aggression that increases across childhood(Broidy et al., 2003).
Developmental trajectories derived from longitudinal studies of girls aresimilar to those obtained with boys but also different: For example, four dis-tinctive patterns were obtained in another Quebec study that are similar tothose obtained for boys, but with the following exceptions: (1) Scores weredecidedly lower than those for boys across the board, and (2) the desistorsamong the females declined more sharply across childhood into adolescencethan those among the boys (see Broidy et al., 2003). Once again, a smallgroup of chronic aggressors emerged, as well as a larger group of consistentlynonaggressive girls. Among the girls in the Dunedin (New Zealand) sample,again, the trajectories are much like the boys’, except scores are lower and thedecline in two groups is a bit greater (see Broidy et al., 2003).
Normatively, then, the existence of two particular trajectories seemsubiquitous—a high level of aggression that is continuous throughout child-hood and into adolescence and a low level of aggression that is equally consis-tent across age. Other patterns, including those characterized by aggressionthat declines during childhood and those typified by increases around the timeof puberty, occur in some data sets but not others. No one is entirely certain asto why these sample-to-sample variations occur in the types of trajectoriesidentified. Moreover, clear evidence does not yet establish the reasons for thevariations that exist. Some investigators believe that persistent and high levelsof aggression across childhood (much more common in boys than in girls)may have biological determinants, whereas “adolescence-limited” aggressionmay be derived from social experience, especially experience with aggressivepeers (Moffitt et al., 2001). Other investigators, however, believe that dys-functional family relationships in early childhood differentiate persistently ag-gressive boys from those showing other developmental patterns (Aguilar,Sroufe, Egeland, & Carlson, 2000). These questions are now at center stage inthe research on the development of aggression. One conclusion, however, can-not be questioned: Normative change in aggressive behavior must be evalu-ated in the context of individual differences. The obverse is also true, namely,that individual differences must be evaluated in the context of normativechange.
8 I N T R O D U C T I O N
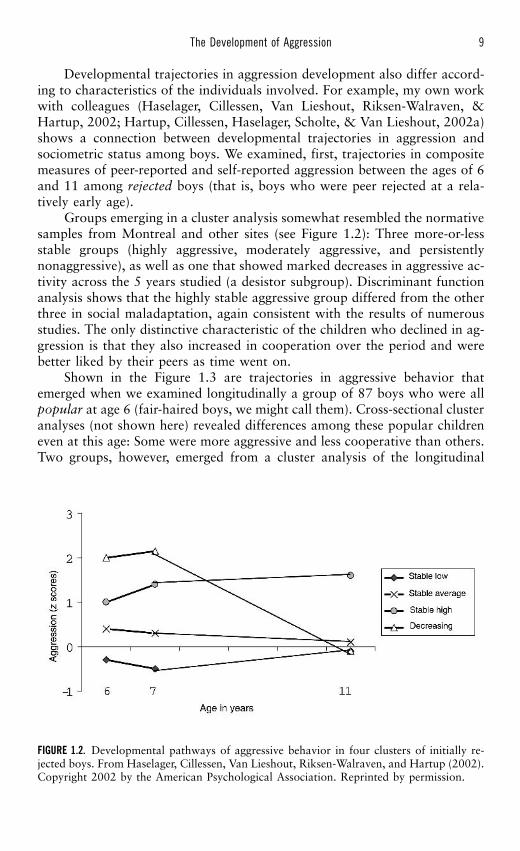
Developmental trajectories in aggression development also differ accord-ing to characteristics of the individuals involved. For example, my own workwith colleagues (Haselager, Cillessen, Van Lieshout, Riksen-Walraven, &Hartup, 2002; Hartup, Cillessen, Haselager, Scholte, & Van Lieshout, 2002a)shows a connection between developmental trajectories in aggression andsociometric status among boys. We examined, first, trajectories in compositemeasures of peer-reported and self-reported aggression between the ages of 6and 11 among rejected boys (that is, boys who were peer rejected at a rela-tively early age).
Groups emerging in a cluster analysis somewhat resembled the normativesamples from Montreal and other sites (see Figure 1.2): Three more-or-lessstable groups (highly aggressive, moderately aggressive, and persistentlynonaggressive), as well as one that showed marked decreases in aggressive ac-tivity across the 5 years studied (a desistor subgroup). Discriminant functionanalysis shows that the highly stable aggressive group differed from the otherthree in social maladaptation, again consistent with the results of numerousstudies. The only distinctive characteristic of the children who declined in ag-gression is that they also increased in cooperation over the period and werebetter liked by their peers as time went on.
Shown in the Figure 1.3 are trajectories in aggressive behavior thatemerged when we examined longitudinally a group of 87 boys who were allpopular at age 6 (fair-haired boys, we might call them). Cross-sectional clusteranalyses (not shown here) revealed differences among these popular childreneven at this age: Some were more aggressive and less cooperative than others.Two groups, however, emerged from a cluster analysis of the longitudinal
The Development of Aggression 9
FIGURE 1.2. Developmental pathways of aggressive behavior in four clusters of initially re-jected boys. From Haselager, Cillessen, Van Lieshout, Riksen-Walraven, and Hartup (2002).Copyright 2002 by the American Psychological Association. Reprinted by permission.

data—one group composed of individuals who were consistently nonaggres-sive and another group of individuals (15% of the subjects) who changed de-cisively in the direction of greater aggressiveness. These increasingly aggressivechildren were distinctive. Concomitant with their increasing aggression weredecreases in cooperativeness and increases in peer rejection. Most interesting,though, are data available for 47 of these boys when they were 14 years ofage: Discriminant function analysis identified two functions that successfullydiscriminated these boys from the other groups: (1) they were more sociable,were less inhibited, and achieved more poorly in school and (2) they were lesswell liked and more likely to engage in aggression, as well as in more bullyingand victimization, than members of the other groups. These results suggestthat synergies involving sociometric status and aggressive trajectories must betaken into account in order to describe, on one hand, “model” boys (stable,nonaggressive, popular boys) and, on the other hand, “toughs” (popular kidswho are increasingly antisocial and not turned on by school) (Rodkin, Farmer,Pearl, & Van Acker, 2000).
The message is clear: Normative change in aggression and antisocial be-havior is meaningful only in the context of differential analysis. And the re-verse may also be true: individual differences can be understood only in thecontext of normative analysis. Consideration of these combined forces is nec-essary to understanding both the development of aggression and social adap-tation more generally.
Child Development versus Lifespan Perspectives
Owing mainly to child-oriented societal forces, over most of the last centuryprimary attention was given to developmental changes occurring in childhood
10 I N T R O D U C T I O N
FIGURE 1.3. Developmental pathways of aggression in three initially popular clusters. FromHartup, Cillessen, Haselager, Scholte, and Van Lieshout (2002a).

rather than to changes occurring in adolescence or adulthood. Most studies ofaggression and its development at the beginning of the child developmentmovement were devoted to young children (Goodenough, 1931; Dawe, 1934);juvenile delinquency was studied among adolescents by sociologists and psy-chiatrists (Thrasher, 1927; Glueck & Glueck, 1934), but other aspects of ado-lescent aggressive behavior were largely ignored. These truncated perspectivesdominate developmental science even today, although they have been modi-fied in various ways—i.e., the doctrine of early experience is questioned in im-portant aspects (Schaffer, 2002) and lifespan perspectives are promulgated asframeworks for examining developmental changes (Baltes, Lindenberger, &Staudinger, 1998).
The bias toward childhood studies of behavioral development stems fromtheoretical/scientific sources as well as the nature of the early child welfaremovement in the United States and elsewhere (Hartup, Johnson, & Weinberg,2001). First, developmental science was heavily influenced in the early 20thcentury by psychoanalysis, on one hand, and learning theories, on the other,including their assumptions that development during the early years has per-vasive and long-lasting consequences in both cognitive and social develop-ment. At the same time, social activists came to believe that the road to gen-eral societal progress begins with changes made in the lives of children andfamilies, especially during a child’s early years. Parent education, social wel-fare services, and early childhood education were expanded greatly in manycountries to assist in this effort. Services to adolescents were also improved,especially those relating to juvenile delinquency, but community welfare in thelarger sense did not encompass lifespan considerations.
Meanwhile, criminologists were discovering that childhood antecedentsaccount for a large amount of variance in adolescent and adult crime (Glueck& Glueck, 1950; McCord, 1979). Clearly, short-term studies of children oradolescents were not going to be able to furnish an adequate basis for socialintervention. More recent investigators utilize increasingly longer-term studiesof aggression and antisocial behavior. Although infancy and early childhoodare not encompassed by most of these studies, longitudinal analysis beginningin childhood and ending in early adulthood teaches us a great deal. We are indebt to David Farrington, Rolf Loeber, Robert Cairns, Leonard Eron andRowell Husemann, John Coie, Joan McCord, and many others for work likethis.
It has been difficult to extend these studies through the life course, begin-ning in infancy and ending in middle or old age. Moffitt et al.’s (2001) work isnotable for the fact that the data set begins with measurements at age 3, andnewer studies (e.g., Tremblay, 2000) begin prenatally and remain open-ended.Studies ranging from middle childhood to young adulthood are still morecommon than those covering a longer term, and cradle-to-grave studies do notexist. Nevertheless, the message of lifespan developmentalists is being takenwith increasing seriousness: Understanding adaptational problems requiresdevelopmental perspectives that extend beyond childhood and adolescence,
The Development of Aggression 11

and the coherence of human development, whether outcomes are good or bad,can be assessed only in terms of both growth and decline in behavioral func-tion.
Must we wait for geriatric data on the Montreal subjects or the Dunedinsample before studying the decline in aggression and antisocial behavior thatis known to mark middle and old age? Probably not. Overlapping longitudi-nal studies—some beginning in adolescence, some beginning later—can un-ravel the conditions responsible for decline in antisocial behavior with age.One can argue, then, that normative change must be studied not only withinthe context of individual differences, but also within the context of the lifecourse.
WHAT ARE THE DETERMINANTS OF BEHAVIOR CHANGE?
Our ultimate goal is to identify the conditions responsible for normative de-velopment of aggression within the context of individual differences. As every-one knows, explanation in the behavioral sciences is extraordinarily difficult.Most developmental change is multidetermined, and causation is burieddeeply within the genome and the mind. Moreover, the time separating“cause” from “effect” in human development is frequently decades, not mo-ments, and change within one behavioral domain is entangled with changes inothers. One cannot synthesize everything that has been written about the de-terminants of aggression in the space available here. Instead, several “ten-sions” or “dialectics” remaining in this field are discussed.
Nativism versus Empiricism
The notion that human beings begin life with “innate ideas” and “inborn pro-clivities” appears in our most ancient religious and philosophical texts. Alsoexpressed in classical philosophical writing is extreme empiricism, the argu-ment being that nearly everything about human behavior—including aggres-sion—derives from the individual’s commerce with the world. Most modernwriters, though, view the development of aggression (like many other traits) asconstrained by intrinsic structures (these may include the physiological andneural structures necessary to arousal, anger, learning, attention, language,and emotional regulation). The structures in question are species-wide andboth determine the success of individual adaptation and make individual dif-ferences possible. Although relatively little is known about the processesthrough which these innate structures are deployed in behavioral develop-ment, especially in social and emotional behavior, the evidence suggests thatrelationships with others (including caretakers, friends, and enemies) are im-portant vectors. The task, then, is to specify both the intrinsic and experientialcontributions to developmental acquisitions and the manner in which they“synthesize” or “interact.”
12 I N T R O D U C T I O N

Recent studies in molecular genetics suggest that allelic variations maycommonly interact with social and relationship experience in determining theaggressive phenotype. Suomi (Chapter 4, this volume) discovered that varia-tions in the serotonin transporter gene (5-HTT) interact with rearing condi-tions to affect a variety of outcomes, including aggression and position in thedominance hierarchy, among rhesus monkeys. So-called “short” allelic varia-tions are associated with nonaggressive maladaptations—but for peer-rearedindividuals and not mother-reared individuals. In other words, attachment toa maternal figure moderates the effects of the genetic variations. The findingsalso partially explain why peer rearing does not produce extremely aggressiveanimals in all cases.
In recent work with human beings, Caspi et al. (2002) discovered an in-teraction between parental maltreatment and levels of monoamine oxidase A(MAOA) genetic expression (MAOA is a gene located on the X chromosomethat encodes an enzyme that metabolizes various neurotransmitters, renderingthem inactive). Maltreated boys in a large longitudinal sample possessing thegenotype associated with high levels of MAOA expression were less likely tohave antisocial problems in the course of their development than those withgenotypes associated with low levels of MAOA expression. Results of thisstudy help to account for the fact that, although harsh parental discipline andmaltreatment of young boys is related to aggressive and antisocial outcomes,not all maltreated children grow up to victimize others.
In addition to these studies, process-oriented prospective studies areneeded (beginning earlier than most existing studies) to explain the manner inwhich so-called biological variations contribute to individual differences in ag-gressiveness and antisocial behavior. One hopes that investigators will notsimplistically consider factors like maternal IQ, maternal age, and maternalsocial adjustment as well as child IQ, child memory, attention disabilities, andchild temperament to be exclusively biological indicators (Moffitt et al.,2001), nor attachment histories to be exclusively social indicators. These con-structs are only the roughest indications of intrinsic or social structures, re-spectively, and the manner in which they work. We need new expertise inpsychoendocrine measurement in infancy and beyond, temperament, emo-tional processes in infancy and early childhood (Thompson, 1998), and be-havioral regulation in early relationships between the child and his or hercaretakers. Implications of the neuropsychological history for the child’s socialand emotional history must be studied along with experience. Overall, refine-ments in methods for studying psychological processes in the development ofaggression are just as important as advances in gene mapping.
Socialization: From Unilateral to Bilateral Constructions
Although there were many indications in earlier writings that children are “so-cializing creatures” as well as “socialized ones” (Baldwin, 1897), a consensusconcerning a two-way or bilateral view of childhood socialization was
The Development of Aggression 13

reached only recently (Bell, 1968). The notion that social adaptation is theoutcome of interaction between the child and others, rather than some sort ofone-way social-molding process, is now accepted by almost everybody.
Parent–Child Relationships
With respect to the development of aggression, one does not find many studiesthat deal directly with social interaction or social relationships within the fam-ily. Most commonly, investigators enter measures like the following into theirregression models as predictors of child or adolescent outcomes: (1) childcharacteristics—temperament, intelligence, troublesomeness, memory and at-tention problems, impulsivity, and personality and (2) parent characteristics—warmth versus negativity, family coercion and inconsistent discipline, punish-ment and punitiveness, along with abusive parenting (Coie & Dodge, 1998).The ensuing regressions allow us to examine main and interaction effectsamong these child and maternal characteristics, but not the social exchangesbetween parents and children from which changes over time actually emanate.
One of the most brilliant achievements of the last century is the “perfor-mance theory” formulated and tested by Gerald Patterson and his associates.This socialization model specifies the parameters and contingencies occurringin family interaction that instantiate troublesomeness among young boys,maintain it, and supply the basis for the attitudes that characterize these sameboys in their early interactions with other children outside the home. These re-lationships, in turn, involve them in increasingly violent behavior (Patterson,Reid, & Dishion, 1992). In other studies, certain mileage can be gained by us-ing global measures of family conflict, maternal punitiveness, and harsh disci-pline, but the “performance model” makes clear that such measures are onlyproxies for measures of social interaction. One concludes that microanalyticapproaches to socialization studies of the development of aggression areneeded as well as macroanalytic ones.
Most parent–child measures also do not pass muster as relationships con-structs. Relationships are usually defined as aggregates of interaction betweenindividuals who have formed expectations about one another that guide inter-personal behavior over time (Hinde, 1997). It so happens that relationshipsare the social contexts within which most development occurs: (1) basic skillsin language and communication, (2) competence in coping with both socialand nonsocial problems, and (3) the attitudes and abilities needed for con-structing other relationships. Relationships theory, however, has not pene-trated very deeply into research on the development of aggression. Parentalpunitiveness or harsh discipline (common measures) are parent characteristics,not dimensions of the parent–child relationship. Similarly, maternal warmthdoes not describe a relationship between mother and child. Exactly what thisomission means for research is not really known. Diana Baumrind (1971) hadconsiderable success earlier in describing permissive relationships betweenparents and children and showing that these were associated with early ag-
14 I N T R O D U C T I O N

gression. Some investigators have consistently reported cross-time correlationsbetween disorganized attachment patterns in infancy and early childhood withmanifestations of aggression and antisocial behavior in middle childhood andadolescence (Renken, Egeland, Marvinney, Mangelsdorf, & Sroufe, 1989).Others report similar findings, especially among vulnerable children, for ex-ample, children who live in chaotic, poor families with a single parent (seeCoie & Dodge, 1998). And, in the Dunedin data set (Moffitt et al., 2001), arelationships measure (“relationship with parents,” based on items like trust,communication, and lack of alienation between adolescents and their parents)turns out to be a better predictor of adolescent aggression (–.44, –.47) thanmeasures of harsh discipline (.19, .27). No one would argue that relationshipconstructs are the only ones that account for significant variance in adolescentaggression. We have sufficient evidence to argue, however, that relationshipsmeasurement definitely belongs in studies of the family antecedents of aggres-sion, especially as we look forward to research dealing with gene–environ-ment interactions.
Peer Relationships
Until recently, the significance of peer relations in the development of aggres-sion and antisocial behavior was considered to be mostly a unidirectional pro-cess. That is, the prevailing view was that kids get into trouble because theyare exposed to the exhortations and examples of deviant peers and are re-jected by better-adjusted associates, an occurrence that lowers self-esteem andrestricts access to well-socialized companions. Sometimes the interaction be-tween child characteristics (e.g., earlier disruptiveness or troublesomeness)and peer experience (e.g., social rejection by other children) is examined in re-lation to aggressive outcomes. In most instances, significant variance in childor adolescent aggression flows additively from both child sources (includingearlier aggression or compliance difficulties) and the attitudes and behaviorsof other children (Coie & Dodge, 1998). Additional variance in the develop-ment of aggression, however, may be traced to close relationships with otherchildren, including (1) friends, (2) enemies, and (3) bully–victim relationships(see Boivin, Vitaro, & Poulin, Chapter 18, this volume).
Friends
Friendship dyads, in general, are neither more nor less aggressive or antiso-cial than “neutral” dyads (Hartup, Verhoeven, DeBoer, Scholte, & VanLieshout, 2002b). Among individual children and adolescents, those whohave friends are not as troubled as those who do not, but “not havingfriends” does not predict aggression very well either concurrently or overtime (Hartup, 1996).
Friends, however, are more similar to one another in both aggressivenessand antisocial behavior than nonfriends (Haselager, Hartup, Van Lieshout, &
The Development of Aggression 15

Risken-Walraven, 1998; Hartup et al., 2002b), a situation that can also be ob-served in social networks or cliques (Cairns & Cairns, 1994). Moreover, thesesimilarities reflect more than “birds of a feather flock together.” The day-to-day interaction that occurs between antisocial friends actually increases theirantisocial behavior over time, especially among children already identified asaggressive and rejected (Dishion, 1990; Tremblay, Masse, Vitaro, & Dobkin,1995). Most likely, this happens because the interaction between aggressivechildren and their friends is more contentious and conflict-ridden than inter-action between nonaggressive friends (Dishion, Andrews, & Crosby, 1995). Inaddition, overtly aggressive children are not notably intimate with one an-other nor as exclusive in their relationship attitudes as their nonaggressivecounterparts (Grotpeter & Crick, 1996). Thus, aggressive friends are aggres-sive risks.
Friendship quality also contributes significant variance to the develop-ment of aggression in childhood and adolescence. In general, unharmoniousfriendships exacerbate the individual child’s problems in school, create ad-justment difficulties, and increase aggressiveness. In many instances, how-ever, friendship quality has a greater effect on children who are at risk insocial development than on those who are not. For example, Poulin,Dishion, and Haas (1999) found significant interaction effects betweenboys’ antisocial behavior (delinquency) at age 13–14 and friendship qualityat that same age (harmoniousness) in relation to antisocial behavior oneyear later, at age 14–15. The greatest effects of low-quality friendships onsubsequent delinquency occurred when baseline delinquency levels werehigh rather than low. In other words, the friendship construct was morestrongly related to developmental outcome among vulnerable individualsthan among less vulnerable ones.
Overall, friends may or may not support good developmental outcome.Empirical studies tell us that aggressive friends (or unharmonious relation-ships) are risk factors inasmuch as the children are not well socialized, insti-gate aggressive behavior (e.g., train one another in deviancy), and do not instigateeffective regulatory controls. In addition, friendships that are not harmoniousinstigate increases in aggressive behavior over time, possibly for some of thesame reasons, and mainly among children who are at risk (Boivin et al., Chap-ter 18, this volume).
Enemies
Social networks also include relationships that are not based on social attrac-tion but, rather, are rooted in antipathy, animosity, and enmity. The term“mutual antipathies” describes this relationships category; the word encom-passes “being enemies” as well as other relationships maintained on the basisof social aversion. Recent studies (Hartup et al., 2002b) show that differencesin aggression and antisocial behavior between adolescents who do not likeeach other are greater than differences between nonenemies (individuals in
16 I N T R O D U C T I O N

neutral dyads). This confirms the “repulsion hypothesis,” which states that in-dividuals dislike those who are different from themselves (Rosenbaum, 1986).Adolescents in mutual antipathies are also substantially more antisocial andaggressive than in “control” or “neutral” dyads, as well as more internalizingand victimized by others.
These differences are consistent with earlier analyses of individual chil-dren and adolescents in this same data set, which suggest that involvement inmutual antipathies is a risk marker. That is, both children and adolescentswho are involved in mutual antipathies show more fighting and bullying, so-cial ineffectiveness, and victimization than those who are not so involved. Peerrejection was a covariate in these analyses, so the variance associated withmutual antipathies is unique. These results characterize both boys and girlswho are involved in same-sex antipathies, and boys, but not girls, who are in-volved in mixed-sex antipathies (Abecassis, Hartup, Haselager, Scholte, &Van Lieshout, 2002). Results from several other studies of mutual antipathiesand their correlates are not entirely consistent, although these results havebeen replicated in three instances (see Hodges & Card, 2003).
Bully–Victim Relationships
Some bullies and victims dislike each other, some are neutral toward one an-other, and some are friends. The correlation between bully–victim status andfriendship attraction is negative but moderate in magnitude (Perry, Finnegan,Hodges, Kennedy, & Malone, 1993). Although the literature on bullying andvictimization is substantial, most of the relevant studies are monadically ori-ented, that is, centered on questions such as “What makes a bully?” or “Whatmakes a victim?” rather than on ongoing bully–victim relationships. Greatprogress has been made in understanding bullying and victimization duringthe last 30 years (see Rigby, 2002), but relationships-oriented ideas are onlyslowly making their way into this area.
As it turns out, both friends and enemies bear on these situations: For ex-ample, increases in victimization are inversely related to the number ofexternalizing problems evinced by friends, suggesting that victims with exter-nalizing friends may well be protected by them (Hodges & Perry, 1999). A sig-nificant relation also exists between characteristics of children’s enemies andvictimization: The aggressiveness, physical strength, and victimization experi-enced by a child’s enemy uniquely predicts the child’s own victimization (Card& Hodges, 2003). Much more, however, remains to be learned about relation-ships between bullies and victims, their varieties, and their vicissitudes.
Relations among Relationships
The social networks of most children and adolescents include friends, neutralassociates, enemies, and sometimes bullies or victims. Do these relationshipexperiences moderate one another in the development of aggression? Does in-
The Development of Aggression 17

volvement in one relationship affect correlates of the other? Recently, my col-leagues and I (Hartup et al., 2002b) examined adolescent friendship dyads asa function of whether one or both individuals is also involved in a mutual an-tipathy (i.e., has an enemy). We also asked whether antipathy dyads vary inpsychological adjustment as a function of whether one or both individuals hasa mutual friend.
First, we gathered together all dyads in a large data set in which the ado-lescents (14-year-olds) mutually nominated each other as “someone you espe-cially like,” as well as all dyads in which the participants mutually nominatedeach other as “someone you don’t like at all.” Results show that friends withmutual antipathies are both more antisocial and more aggressive than friendswithout mutual antipathies or those in which only one child has an enemy.Peer victimization and internalizing behavior are also greater among friends inwhich one or both children have mutual antipathies. Second, we examinedpairs of adolescents who did not like one another as well as whether either ad-olescent had a mutual friend. Antisocial behavior is less frequent among theadolescent enemies who also have friends than among those who do not. Peervictimization is lower, too, when one or both enemies have a friend. But differ-ences are not significant for aggressive behavior according to whether enemiesalso have friends.
Overall, then, friends are more antisocial when they also have enemies;concomitantly, enemies have lower antisocial behavior scores when they alsohave friends. Aggressiveness, however, is not moderated in the same way:Friends with enemies are more aggressive than friends without, but friend-ships do not appear to be strong enough to lessen the aggressiveness that ac-companies involvement in a mutual antipathy.
Does Context Make a Difference?
Currently, the most advanced ideas being pursued about contextualism anddevelopment do not include the notion that person and environment can beconceived dualistically. Gone are notions that persons cause environmentalchange and vice versa. Instead, the modern view treats the person and the en-vironment as integrated or fused; neither is an entity that can be described sen-sibly without reference to the other, and development is the story of changeover time in this integrated system. One must consider development to becharacteristic of biological and social systems, not an attribute of either indi-viduals or contexts.
Conventional (dualistic) contextualism is well represented in research onthe development of aggression. Cross-national studies show differences in de-velopmental trajectories (Broidy et al., 2003); cross-cultural studies, the same(Whiting & Whiting, 1975). There is also clear evidence that ecological fac-tors like neighborhood violence have a great deal to do with the early onset ofaggressive behavior. These scenarios, however, are not simple. As Schwartzand colleagues (Schwartz, Gorman, Toblin, & Abou-ezzedine, 2003) demon-
18 I N T R O D U C T I O N

strate, exposure to community violence is consistently related to childhood ag-gression only among children who are involved in a relatively large number ofmutual antipathies with other children. Once again, a relationship factor in-creases risk in the development of aggression among children who are alreadyvulnerable. These results move us a step or two from dualistic notions aboutindividuals and the community because, in this case, relationship systems areseen to moderate the effects of external conditions. Community and societalvariables will remain in research on the development of aggression, but no-tions about the “environment” need to be rethought in terms of ecologicalsystems theory (Gatti & Tremblay, Chapter 19, this volume).
CONCLUSION
Conclusions to this survey of research on aggression and its development arerelatively straightforward: Conceptualization and empirical studies dealingwith the development of aggression need to organized as a series of “nests.”
1. Normative change needs to be nested theoretically in the context of in-dividual differences and within long-term (i.e., lifespan) perspectives.
2. The development of aggression can be understood only when the earli-est genetic expressions are examined as nested in early social experi-ence, especially individual social histories and close relationships.
3. The aggressive act must be considered as nested within a broad andcomplex ecology of harm doing and antisocial behavior.
We stand at the edge of the nest, then, regarding what we know about thedevelopment of aggression.
REFERENCES
Abecassis, M., Hartup, W. W., Haselager, G. J. T., Scholte, R. J. H., & Van Lieshout, C. F.M. (2002). Mutual antipathies and their significance in childhood and adolescence.Child Development, 73, 1543–1556.
Aguilar, B., Sroufe, L. A., Egeland, B., & Carlson, E. (2000). Distinguishing the early-onset-persistent and adolescent-onset antisocial behavior types: From birth to 16years. Development and Psychopathology, 12, 109–132.
Baldwin, J. M. (1897). Social and ethical interpretations in mental development. NewYork: Macmillan.
Baltes, P. B., Lindenberger, U., & Staudinger, U. M. (1998). Life-span theory in develop-mental psychology. In W. Damon (Series Ed.), R. M. Lerner (Vol. Ed.), Handbook ofchild psychology: Vol. 1. Theoretical models of human development (5th ed., pp.1029–1144). New York: Wiley.
Bandura, A. (1973). Aggression: A social learning analysis. Englewood Cliffs, NJ:Prentice-Hall.
The Development of Aggression 19

Bandura, A., & Walters, R. H. (1963). Social learning and imitation. New York: Holt,Rinehart & Winston.
Baumrind, D. (1971). Current patterns of parental authority. Developmental PsychologyMonographs, 4(1), 1–103.
Bell, R. Q. (1968). A reinterpretation of direction of effects in studies of socialization.Psychological Review, 75, 81–95.
Broidy, L. M., Nagin, D. S., Tremblay, R. E., Bates, J. E., Brame, B., Dodge, K. A.,Fergusson, D., Horwood, J. L., Loeber, R., Laird, R., Lynam D. R., Moffitt, T. E.,Pettit, G. S., & Vitaro, F. (2003). Developmental trajectories of childhood disrup-tive behaviors and adolescent delinquency: A six site, cross-national study. Devel-opmental Psychology, 39(2), 222–245.
Cairns, R. B., & Cairns, B. D. (1994). Lifelines and risks. Cambridge, UK: CambridgeUniversity Press.
Card, N. A., & Hodges, E. V. E. (2003) Parent–child relationships and enmity with peers:The role of avoidant and preoccupied attachment. In E. V. E. Hodges & N. A. Card(Eds.), Enemies and the darker side of peer relations (pp. 23–37). San Francisco:Jossey-Bass.
Caspi, A., McClay, J., Moffitt, T. E., Mill, J., Martin, J., Craig, I. W., Taylor, A., &Poulton, R. (2002). Role of genotype in the cycle of violence in maltreated children.Science, 297, 851–854,
Coates, B., & Hartup, W. W. (1969). Age and verbalization in observational learning.Developmental Psychology, 1, 556–562.
Coie, J. D., & Dodge, K. A. (1998). Aggression and antisocial behavior. In W. Damon (Se-ries Ed.), N. Eisenberg (Vol. Ed.), Handbook of child psychology: Vol. 3. Social,emotional, and personality development (pp. 779–862). New York: Wiley.
Crick, N. R., Werner, N. E., Casas, J. F., O’Brien, K. M., Nelson, D. A., Grotpeter, J. K., &Markon, K. (1998). Childhood aggression and gender: A new look at an old prob-lem. In D. Bernstein (Ed.), Nebraska Symposium on Motivation: Vol. 45. Genderand motivation. Lincoln: University of Nebraska Press.
Dawe, H. C. (1934). An analysis of two hundred quarrels of preschool children. ChildDevelopment, 5, 139–157.
Dishion, T. J. (1990). The peer context of troublesome child and adolescent behavior. InP. Leone (Ed.), Understanding troubled and troublesome youth (pp. 128–153).Newbury Park, CA: Sage.
Dishion, T. J., Andrews, D. W., & Crosby, L. (1995). Anti-social boys and their friends inearly adolescence: Relationship characteristics, quality, and interactional process.Child Development, 66, 139–151.
Feshbach, S. (1970). Aggression. In P. H. Mussen (Ed.), Carmichael’s manual of childpsychology (Vol. 2, pp. 159–259). New York: Wiley.
Glueck, S., & Glueck, E. T. (1934). One thousand juvenile delinquents. Cambridge, MA:Harvard University Press.
Gleuck, S., & Glueck, E. T. (1950). Unraveling juvenile delinquency. Cambridge, MA:Harvard University Press.
Goodenough, F. L. (1931). Anger in young children. Minneapolis: University of Minne-sota Press.
Grotpeter, J. K., & Crick, N. R. (1996). Relational aggression, overt aggression, andfriendship. Child Development, 67, 2328–2338.
Hartup, W. W. (1996). The company they keep: Friendships and their developmental sig-nificance. Child Development, 67, 1–13.
20 I N T R O D U C T I O N

Hartup, W. W., Cillessen, A. H. N., Haselager, G. J. T., Scholte, R. J., & Van Lieshout, C.F. M. (2002a). Heterogeneity among popular boys: Subtypes and developmentaltrajectories. Unpublished manuscript, University of Minnesota.
Hartup, W. W., & De Wit, J. (1974). The development of aggression: Problems and per-spectives. In J. De Wit & W. W. Hartup (Eds.), Determinants and origins of aggres-sive behavior (pp. 595–620). The Hague: Mouton.
Hartup, W. W., Johnson, A., & Weinberg, R. A. (2001). The Institute of Child Develop-ment: Pioneering in science and application. In W. W. Hartup & R. A. Weinberg(Eds.), Child psychology in retrospect and prospect: The 32nd Minnesota Sympo-sium on Child Psychology (pp. 227– 257). Mahwah, NJ: Erlbaum.
Hartup, W. W., Verhoeven, M., DeBoer, R., Scholte, R. J. H., & Van Lieshout, C. F. M.(2002b, August). Heterogeneity of mutual friendships and mutual antipathies: Across-sectional study. Paper presented at the biennial meeting of the InternationalSociety for the Study of Behavioural Development, Ottawa (Canada).
Haselager, G. J. T., Cillessen, A. H. N., Van Lieshout, C. F. M., Riksen-Walraven, J. M. A.,& Hartup, W. W. (2002). Heterogeneity of social behavior in peer rejected boysacross middle childhood: Developmental pathways and their correlates. Develop-mental Psychology, 38, 446–456.
Haselager, G. J. T., Hartup, W. W., Van Lieshout, C. F. M., & Riksen-Walraven, J. M. A.(1998). Similarities between friends and nonfriends in middle childhood. Child De-velopment, 69, 1198–1208.
Hinde, R. A. (1997). Relationships: A dialectical perspective. Hove, UK: PsychologyPress.
Hodges, E. V. E., & Card, N. A. (Eds.) (2003). Enemies and the darker side of peer rela-tions. San Francisco: Jossey-Bass.
Hodges, E. V. E., & Perry, D. G. (1999). Personal and interpersonal antecedents and con-sequences of victimization by peers. Journal of Personality and Social Psychology,76, 677–685.
Maccoby, E. E. (1992). The role of parents in the socialization of children: An historicaloverview. Developmental Psychology, 28, 1006–1017.
McCord, J. (1979). Some child-rearing antecedents of criminal behavior in adult men.Journal of Personality and Social Psychology, 37, 1477–1486.
Moffitt, T. E., Caspi, A., Dickson, N., Silva, P., & Stanton, W. (1996). Childhood-onsetversus adolescent-onset antisocial conduct problems in males: Natural history fromages 3 to 18 years. Development and Psychopathology, 8, 399–424.
Moffitt, T. E., Caspi, A., Rutter, M., & Silva, P. A. (2001). Sex differences in antisocial be-havior. Cambridge, UK: Cambridge University Press.
Nagin, D., & Tremblay, R. E. (1999). Trajectories of boys’ physical aggression, opposi-tion, and hyperactivity on the path to physically violent and nonviolent juvenile de-linquency. Child Development, 70(5), 1181–1196.
Parke, R. D., & Slaby, R. (1983). The development of aggression. In P. H. Mussen (SeriesEd.), E. M. Hetherington (Vol. Ed.), Handbook of child psychology: Vol. 4. Social-ization, personality, and social development (pp. 547–641). New York: Wiley.
Patterson, G. R., Reid, J., & Dishion, T. J. (1992). Antisocial boys. Eugene, OR: CastaliaPress.
Perry, D. G., Finnegan, R. A., Hodges, E. V. E., Kennedy, E., & Malone, M. (1993, Au-gust). Aggressive and victimized children’s relations with parents and peers. Paperpresented at the annual meeting of the American Psychological Association, To-ronto.
The Development of Aggression 21

Poulin, F., Dishion, T. J., & Haas, E. (1999). The peer influence paradox: Friendshipquality and deviancy training within male adolescent friendships. Merrill-PalmerQuarterly, 45, 42–61.
Renken, B., Egeland, B., Marvinney, D., Mangelsdorf, S., & Sroufe, L. A. (1989). Earlychildhood antecedents of aggression and passive-withdrawal in early elementaryschool. Journal of Personality, 57, 257–281.
Rigby, K. (2002). Bullying in childhood. In P. K. Smith & C. H. Hart (Eds.), Blackwellhandbook of childhood social development (pp. 549–568). Oxford, UK: Blackwell.
Rodkin, P. C., Farmer, T. W., Pearl, R., & Van Acker, R. (2000). Heterogeneity of popularboys: Antisocial and prosocial configurations. Developmental Psychology, 36, 14–24.
Rosenbaum, M. E. (1986). The repulsion hypothesis: On the nondevelopment of rela-tionships. Journal of Personality and Social Psychology, 51, 1156–1166.
Schaffer, H. R. (2002). The early experience assumption: Past, present, and future. In W.W. Hartup & R. K. Silbereisen (Eds.), Growing points in developmental science (pp.24–46). Hove, UK: Psychology Press.
Schwartz, D., Gorman, A. H., Toblin, R., & Abou-ezzedine, T. (2003). Mutual antipa-thies in the peer group as a moderating factor in the association between communityviolence exposure and psychosocial maladjustment. In E. V. E. Hodges and N. A.Card (Eds.), Enemies and the darker side of peer relations (pp. 39–54). San Fran-cisco: Jossey-Bass.
Thompson, R. A. (1998). Early sociopersonality development. In W. Damon (Series Ed.),N. Eisenberg (Vol. Ed.), Handbook of child psychology: Vol. 3. Social, emotional,and personality development (pp. 25–104). New York: Wiley.
Thrasher, F. M. (1927). The gang. Chicago: University of Chicago Press.Tremblay, R. E. (2000). The development of aggressive behaviour during childhood:
What have we learned in the past century? International Journal of Behavioral De-velopment, 24, 129–141.
Tremblay, R. E., Boulerice, B., Harden, P. W., McDuff, P., Perusse, D., Pihl, R. O., &Zoccolillo, M. (1996). Do children in Canada become more aggressive as they ap-proach adolescence? In Human Resources Development Canada & Statistics Can-ada (Eds.), Growing up in Canada: National longitudinal survey of children andyouth (Catalogue 89-550, pp. 127–137). Ottawa: Statistics Canada.
Tremblay, R. E., Masse, L. C., Vitaro, F., & Dobkin, P. L. (1995). The impact of friends’deviant behavior on early onset of delinquency: Longitudinal data from 6 to 13years of age. Development and Psychopathology, 7, 649–667.
Whiting, B. B., & Whiting, J. W. M. (1975). Children of six cultures. Cambridge, MA:Harvard University Press.
22 I N T R O D U C T I O N

Part II
THE DEVELOPMENTOF AGGRESSION
IN ANIMALS AND HUMANS

This page intentionally left blank

THE DEVELOPMENT OF AGGRESSIONSubtypes of Aggression
2
Subtypes of Aggressionin Humans and Animals
PAUL L. GENDREAU and JOHN ARCHER
From the elementary fencing behavior of fruit flies to the lunging attack ofmice or rats, and from the biting of dogs to the more subtle form of indirectaggression in humans, the expression of aggressive behavior becomes increas-ingly sophisticated and diversified as animals increase in neural complexity.Comparable transformation is observed during the course of human ontog-eny. Children first bite, hit, push, and kick (Tremblay et al., 1999); then grad-ually, they may come to choose more indirect, socially oriented forms of ag-gression such as spreading negative rumors about a classmate or attempting toalter relationships (Björkqvist, Lagerspetz, & Kaukiainen, 1992). The expres-sion of aggression undoubtedly reaches a peak of complexity in adulthood,not only in terms of expression but also with regard to its multiple causes andconsequences (Loeber & Day, 1997).
The fact that aggression is not a unitary or homogeneous phenomenon isclearly not in dispute among scientists. More challenging, however, is the needto provide a definition of aggression that would satisfy researchers from most(or, it is hoped, all) spheres of expertise (Archer & Browne, 1989). Some au-thors have argued that this is an impossible mission because the concept of ag-gression involves too many variables and determinants and, more particularly,a significant part of social and moral judgment (e.g., Johnson, 1972). Othersavoid using the term aggression because of its diffuse meaning (e.g., Patterson& Cobb, 1973), or declare that reaching a consensus on a definition is notnecessary to perform a meticulous analysis of aggression (e.g., Cairns, 1979).Finally, some researchers have maintained that aggression is essentially an in-adequate concept that ought to be replaced by more precise terminology (e.g.,coercive control; Tedeschi & Felson, 1994; Felson, 2002).
25

Before reaching a potential agreement about a definition, a first step is todescribe and classify the different types of aggression. As Moyer (1968)argued, “progress in understanding the general phenomenon of aggressive be-havior can only be made when the various aggressions are carefully and oper-ationally defined” (p. 65). Hence, several attempts have been made to sort thedifferent forms of aggression into discrete categories. Various taxonomieshave been offered for both animal and human aggression, based on the simi-larity of behavioral expression, contextual characteristics of the eliciting stim-ulus, functional significance, motivational underpinning, or underlying neuro-physiological mechanisms.
In this chapter we review the diverse forms of aggression in both animalsand humans in an attempt to integrate the current knowledge into a coherenttheoretical and practical framework. We argue that a better understanding ofthe concept of aggression requires a systematic examination of the variousproximal and distal forces that induce, facilitate, or maintain the developmentand expression of the different subtypes of aggression in both humans and an-imals. Comparing the similarities and dissimilarities of these multiple forces ina variety of species, and among individuals within a given species, is the onlypossible strategy for formulating a universal taxonomy of aggression.
COMPARING DIFFERENT LEVELS OF PHYLOGENETICAND ONTOGENETIC ORGANIZATION
The Risk of Anthropomorphism and Zoomorphism
To recognize the phylogenetic sophistication of aggression, it is important todiscriminate between subtypes of aggression that are unique to humans andthose that are also present in other species. The ontogenetic sophistication ofaggression will become manifest only by differentiating subtypes of aggressionthat are expressed during a specific developmental period from those that oc-cur throughout ontogeny. Distinctions should be articulated behaviorally,functionally, and neurobiologically. At the behavioral level, a number ofaggression-related behaviors are strikingly similar among many species, evenin those that are extremely distant in phylogenetic terms. For instance, a“boxing” stance (an upright posture observed during fighting) is shown byfruit flies, rats, and humans. Although the behavioral repertoire of flies androdents is more complex than one would expect (Chen, Lee, Bowens, Huber,& Kravitz, 2002; Barnett & Marples, 1981; Gendreau, Gariépy, Petitto, &Lewis, 1997), it does not reach the range of human behavioral expression.Nevertheless, one may ask how is it possible that such unrelated species ex-hibit similar forms of aggression. What do flies, rodents, and humans have incommon besides possessing pairs of limbs and being able to display a similarmotor-expressive pattern? At the functional level, fruit flies and rats do notfight for money or pride, but both can fight for sexual access and territoriality,as humans sometimes do. At the neurobiological level, the divergence is con-
26 T H E D E V E L O P M E N T O F A G G R E S S I O N

siderable. The rudimentary nervous system of a fruit fly is made up of a mere250,000 neurons, and the brain of an adult rat weighs more or less 2 g. Withits 100 billion neurons, the 1.4-kg human brain is at a different level of com-plexity. Nevertheless, rats and humans share similar brain structures and path-ways, and, like fruit flies, they can develop addiction to cocaine, nicotine, andalcohol (Bainton, Tsai, Singh, Moore, Neckameyer, & Heberlein, 2000), sug-gesting a certain degree of neurobiological similarity.
Cross-species comparisons call for differentiating behavioral and neuro-physiological characteristics that are homologous (i.e., having a common evo-lutionary origin) from those that are simply analogous (i.e., likeness in func-tion but not in evolutionary origin). When analyzing behavioral and/orphysiological similarity between species, we run the risk of anthropomor-phism, that is, viewing animals as having human-like qualities, or zoomorph-ism, that is, mistakenly perceiving human behavior as the mirror image of ani-mal behavior (Cairns, 1979). For instance, contrary to the controversial andsensationalist opinion of authors in the past (e.g., Ardrey, 1966), predatorybehavior in animals should not be equated with hunting or warfare in hu-mans. Although these behaviors may share some common neuroevolutionaryprocesses, the function of predatory behavior is to supply food, thus enhanc-ing the probability of survival. Human hunting, at least in modern society, isoften motivated only by the pleasure that the activity provides, and not by thenecessity to gain food. Predation targets other species, whereas warfare is di-rected at other humans.
Likewise, matching children’s and adults’ behavior on the basis of theirexpressive similarity, something one could adventurously call “adultomorph-ism” or “pedomorphism,” depending on the direction of the comparison, isnot uncommon. A child hitting another child in kindergarten and an adult hit-ting a colleague at work are events that—albeit similar in their expression andto some extent in their immediate consequences (i.e., physical hurt)—clearlydiffer in terms of antecedents and long-term consequences. In sum, any defini-tion of aggression that does not address the issue of phylogenetic andontogenetic similarity and contrast encourages a static or reified view of acomplex phenomenon.
Early Taxonomies of Aggression
Animal models of aggression provide a strong conceptual base for approach-ing the study of human aggression. Moyer (1968) established seven categoriesof aggressive behavior based on their functional values and stimulus-boundcharacteristics: predatory aggression, intermale aggression, fear-induced ag-gression, irritable aggression, territorial defense, maternal aggression, and in-strumental aggression. A possible eighth category, sex-related aggression,was also suggested. As Moyer himself mentioned five years later, however,“definition of the kinds of aggression on that basis alone now appears too re-strictive: the kinds of aggression vary on a number of different dimensions,
Subtypes of Aggression 27

and all of them must be considered in the definition of each kind” (Moyer,1973, p. 12). The dimensions that Moyer referred to were the specificity of theeliciting stimulus, the presence or absence of an emotional display, the sex ofthe attacker, and the neurophysiological correlates. With these four “dimen-sions” in mind, Moyer (1973) finally rejected territorial defense as a subtypeof aggression. Although Moyer’s taxonomy was primarily targeting animalaggression, reference was also made to human behavior; however, the unique-ness and complexity of human aggression was not specifically addressed. Theuse of the categories to describe and differentiate human aggression undoubt-edly leans toward zoomorphism.
A similar comment can be made regarding Wilson’s sociobiological tax-onomy (Wilson, 1980). Although most of the subtypes of aggression (i.e., ter-ritorial, dominance, sexual, parental disciplinary, weaning, moralistic, preda-tory, and antipredatory aggression) were meant to be theoretically applicableto both animals and humans, there are obvious exceptions. Weaning aggres-sion, for example—that is, occurring when parents “gently attack” their off-spring to make them stop begging for food—is clearly more relevant to animalbehavior. The opposite can be said of moralistic aggression, which for Wilson(1980) represents advanced forms of reciprocal altruism that reduce the mani-festation of aggression via rules, codes of punishment, or enforced conformity.
In conjunction with the risk of zoomorphism and/or anthropomorphism,a major problem with Moyer’s and Wilson’s taxonomies is that they are basedon an eclectic set of criteria. These involve a specific target (e.g., prey, preda-tor, another male), a specific context (e.g., territorial, maternal, weaning), aspecific function (e.g., to dominate, to obtain sexual access) or the concomi-tant occurrence of an emotional state (e.g., fear, anger), or a learning process(e.g., instrumental, moralistic). As argued by Archer and Browne (1989), oneshould try to be more explicit, and perhaps more restrictive, in terms of whichcriteria are used to divide aggression into distinct classes. Archer (1988) pro-posed a simpler classification on the basis of function by distinguishing formsof aggression that are “competitive,” “protective,” and “parental.” Parentalaggression can be viewed as an extension of protective aggression, involvingthe protection of those recognized as genetically related and dependent. Preda-tion was omitted from this scheme because, as argued in an earlier article (Ar-cher, 1976), Moyer (1968) actually presented good evidence for the neuraland motivational separation of the two forms of behavior. The argument wasthat the motivational systems underlying aggression and predation hadevolved independently of one another; a good example is provided by cats,who show very different types of behavior and emotional states when fightingone another and when stalking prey. However, it is undoubtedly the case thatthe systems controlling predation and aggression are more related in somecases, for example, in the case of mouse killing by rats (e.g., Karli, 1956), incannibalism in chimpanzees (Bygott, 1972), and more widely in the case ofmale infanticide. In humans, the link between hunting and aggression may bemore complex.
28 T H E D E V E L O P M E N T O F A G G R E S S I O N

At about the same time that Moyer proposed his taxonomy and sug-gested four dimensional criteria to discriminate subtypes of aggression in ani-mals, similar attempts were made to categorize the different forms of aggres-sion in humans. Buss (1961) suggested that aggression be dichotomized alongthree behavioral dimensions: physical–verbal, active–passive, and direct–indi-rect. In 1969, Pulkkinen formulated a bidimensional (and bicriteria) modelcharacterizing human aggression (see Pulkkinen, 1987). A first dimension wasrelated to the expression of aggression, ranging from indirect forms to moredirect ones. A second dimension concerned the defensive–offensive dichotomy,which was determined by the presence or absence of proximal threateningstimuli. Although these models have the merit of being parsimonious, they arenot truly pertinent to animal aggression, as indirect forms of aggression areexclusive to humans and, possibly, nonhuman primates (Campbell, 1999). Inaddition, a model depicting behavioral or contextual features in terms of acontinuum (poles or axes) may not be appropriate for some distinctions, be-cause it postulates quantitative differences between the different forms of ag-gression. An aggressive act can fluctuate in intensity, but it cannot be half di-rect and half indirect.
A MULTILEVEL ANALYSIS OF AGGRESSION IN ANIMALS AND HUMANS
Aggression is a dynamic, multifaceted social-emotional process that calls for acareful examination of its various antecedents, expressions, and consequences(or functions). The adoption of a dynamic and multifactorial perspectivewhen investigating the foundations of aggression and other types of social be-havior in humans and animals was ardently advocated by Cairns (1979).Cairns stressed the importance of using adequate criteria in order to generalizefrom one phylogenetic level to another and from one developmental stage toanother. Observing a similar feature in two or more species (e.g., boxing) or ina child and an adult (e.g., hitting) is not enough to conclude that these behav-iors serve the same function and derive from the same proximal or distal ante-cedents. This principle has a significant impact on how aggression is ulti-mately defined and how the different forms of aggression can be grouped.
Figure 2.1 summarizes the major domains or themes of investigation inaggression research. This is an extended representation of Cairns’s model ofwhat he called “polythetic analysis,” that is, the examination of aggressionfrom multiple conceptual angles. The first three domains encompass the prox-imal (eliciting context and neurophysiological mechanisms) and the more dis-tal antecedents of aggression, which are perceptible only through a longitudi-nal, developmental analysis. The fourth domain relates to the expression ofaggression, whereas the fifth and the sixth cover the consequences of aggres-sion to others (harm) and to self and society (social/emotional function), re-spectively.
One must recognize that there are important differences between, as well
Subtypes of Aggression 29

as within, species in terms of variables that influence the development and ex-pression of aggression. Nevertheless, this thematic framework probably cov-ers most subtypes of aggression found in the scientific literature and can begenerally applied to both animal and human research. Obviously, dichoto-mizing aggression as direct versus indirect or as producing physical versusmental harm may be more appropriate and applicable to human behaviorthan to rat or fruit fly aggression. It is also important to note that even if mostsubtypes of aggression have been established on the basis of a single criterion,it is always possible to relate them to other criteria. Therefore, overlap be-tween criteria is clearly the rule rather than the exception. For example,parental aggression in animals serves an obvious long-term function (i.e., sur-vival of offspring), but it can also be described in terms of antecedents, eitherproximal (e.g., perception of a prey, underlying neurophysiological mecha-nism) or more distal (e.g., genetic predisposition).
Common sense would dictate that we follow the temporal sequence ofaggression, beginning with its antecedents, then considering its expression,and finishing with its consequences for others and the self. We decided to gothe opposite way, starting with the first and most obvious outcome of aggres-sion (its consequences for others and self) and then considering elements thatnecessitate a more thorough examination of the problem (e.g., distal anteced-ents and developmental issues). This upward presentation of the differentcomponents of aggression ends with a provisional integrative representationof the most significant subtypes of aggression and their underlying processes.
30 T H E D E V E L O P M E N T O F A G G R E S S I O N
FIGURE 2.1. Polythetic analysis of aggression. Based on Cairns (1979).

SUBTYPING AGGRESSION ON THE BASISOF ITS CONSEQUENCES ON OTHERS AND SELF
Based on Harm and Injury
Harm or injury to others is the foremost indicator that an aggressive act hasoccurred. This is perhaps why it has been a common criterion for qualifyingaggression (Berkowitz, 1981; Rule, 1974). In its most simplified form, aggres-sion has been defined as the delivery of noxious or painful stimuli to anotherindividual (Buss, 1961) or as a response resulting in injury (Rule, 1974) or as“damage or destruction of some goal entity” (Moyer, 1968). Some authorsspecified that the targeted individual ought to “be motivated to avoid suchtreatment,” in order to exclude cases of sadomasochism (Baron, 1977). Re-search has traditionally focused on physical harm, but with increasing fre-quency, has been including mental or psychological harm as a potential conse-quence of aggression, as in the case of indirect aggression or persistentbullying.
A number of issues have been raised concerning the validity of harm as acriterion for assessing or qualifying aggression. First, as potential harm, eitherphysical or psychological, seems inherent to all forms of aggression, it has lit-tle discriminatory value. In addition, assessment of harm is not only contin-gent upon the nature of the aggressive action (a hit by a 2-year-old may nothurt an adult), but it is also conditional on the victim’s sensibility. Some indi-viduals may be hypersensitive to external stimulation and may be more proneto feel physically and/or mentally harmed. Assessment of harm is indeed ahighly subjective matter. Even from a behaviorist perspective that targets theobservable behavior of the aggressor (delivery of noxious stimuli), a judgmentmust be made to determine whether pain or harm has been inflicted or not.Furthermore, the harm criterion has little value for subtyping aggression inanimals. Automated bite-recording devices to measure the intensity of noxiousdelivery by aggressive primates and rodents have been designed (Ulrich,Dulaney, Arnett, & Mueller, 1973) but the devices turned out to be too im-practical and restrictive (Knutson, 1973).
Based on Intent, Motivation
Descriptive models of aggression have often emphasized the cognitive/motiva-tional antecedent of aggression. An influential paper by Feshbach (1964) wascritical in establishing two major types of aggression based on harm, or, morespecifically, on the motivation to harm. If injury (to a person or object) wasthe primary goal of the action (pleasure or satisfaction following injury beingthe main reward), it was labeled hostile aggression. If injury was not the mainpurpose and the action was executed for reward other than the pleasure of in-juring, then it was termed instrumental aggression. More specifically, Fesh-bach (1964) defined instrumental aggression as any act that produces harmand that “is directed toward the achievement of nonaggressive goals.” These
Subtypes of Aggression 31

“nonaggressive” goals include getting attention and acquiring an object or aresource.
Although differentiating aggression based on the presence of hostile in-tention has been a dominant and relatively valid conceptual dichotomy in ag-gression research (Atkins, Stoff, Osborne, & Brown, 1993), it raises theoreti-cal problems. First, aggression can be both hostile and instrumental (Bushman& Anderson, 2001; Hartup & De Wit, 1974). Second, labeling aggression ashostile is rather tautological. Are there nonhostile, friendly forms of aggres-sion? This semantic glitch should not hinder the fact that some people maygain a considerable satisfaction or pleasure from hitting and injuring some-one, whereas others may benefit more from the social or materialistic conse-quences of their aggressive action. It is important to note that getting pleasurefrom producing harm to someone may not be the most common motivationalantecedent of aggression. Indeed, for many explanatory models (e.g., Archer,1976; Berkowitz, 1993; Dollard, Doob, Miller, Mowrer, & Sears, 1939),bringing an end to a situation that is annoying and removing an irritant or adiscrepancy from what is expected are frequent motivational precursors of ag-gression. Nonetheless, both pleasure-motivated aggression and relief-moti-vated aggression imply an emotional outcome or reward that is either pleasur-able (positive reinforcement) or a release from a previously noxious state(negative reinforcement). Whether we call Feshbach’s categories of aggressionhostile versus instrumental, annoyance-motivated versus incentive-motivated(Zillman, 1978), or emotionally rewarding versus materialistically/socially re-warding, does not really matter if an unambiguous definition is provided.
Researchers with a social-interactionist perspective (Tedeschi & Felson,1994) view aggression (which they term “coercive power”) as being motivatedby interpersonal goals—to control others, to maintain justice, or to defend so-cial identity. From the perspective of the actor, all aggression is therefore in-strumental in that it pursues one of these social goals. However, these situa-tions can just as readily be viewed as removing an irritant (i.e., negativereinforcement), and it may be confusing to regard such situations—which in-evitably involve anger—as instrumental in nature. For the purposes of clarity,it is probably better to restrict the term “instrumental” to cases involving pos-itive reinforcement of aggression by a reward unconnected with activation ofthe aggression system. This would highlight the parallels between such humancases as robbery and rape (Felson, 2002) and the use of conventional reinforc-ers such as food or water to facilitate animal aggression (e.g., Ulrich,Johnston, Richardson, & Wolff, 1963).
Feshbach (1971) and Rule (1974) proposed a different taxonomy by sub-dividing instrumental aggression into personally motivated aggression and so-cially motivated aggression. Personally motivated aggression is different fromhostile aggression in that the primary goal is not to hurt but to obtain rewardvia inflicting injury. Yet aggression is said to be socially motivated when theprimary goal is to gain a social advantage. Even in the authors’ views, how-ever, the distinction between these subtypes of aggression was problematic,
32 T H E D E V E L O P M E N T O F A G G R E S S I O N

because of the difficulty in assessing motivation, intent, or feeling. Sears(1961) suggested the term prosocial aggression to distinguish aggressive be-havior considered as socially and morally acceptable. However, to the sameextent that the term hostile aggression is certainly a truism, the expression“prosocial aggression” appears to be an oxymoron. Naming an aggressive actas prosocial or antisocial depends too much on individual viewpoints to be auseful categorization criterion. Physically punishing a child to prevent the oc-currence of a life-threatening situation (e.g., crossing a high-traffic road) mayhave a justifiable intention and long-term benefits. From the child’s perspec-tive, however, the immediate consequences of being spanked by a parent orbeing slapped by a peer may not differ much. Moreover, any act of terrorismor war can be considered legitimate and morally acceptable from the aggres-sor’s perspective. From the victim’s point of view, the immediate and harmfulconsequences do not depend on how good the intention was.
It has been claimed that behavior should be at once intentional (motiva-tion to injure) and harmful to be classified as being aggressive (Berkowitz,1993; Dollard et al., 1939). As with the nature and intensity of harm, how-ever, intent is a concept that is difficult to prove and easy to deny (Loeber &Hay, 1997). It also imposes serious limitations in investigating aggression inchildren or animals (Tremblay, 2000). For these reasons, although intent iscertainly central to the definition of aggression, it may not be an adequate cri-terion in practical terms for establishing a taxonomy of aggression applicableto both animals and humans (children and adults). Intention may be alsoproblematic in cases of indirect aggression where the perpetrator seeks tocover up his or her actions.
SUBTYPING AGGRESSION ON THE BASIS OF ITS EXPRESSION
Categorizing aggression on the basis of its behavioral expression has been apopular criterion as it circumvents the pitfalls of intrinsic notions such asinstinct, motivation, drive, or intent to harm. Unfortunately, too many behav-ioral responses that have little in common have been subsumed under the labelof aggression. Display of aggressive behavior is species-typical and age-dependent, as it is manifestly constrained by morphogenetic, neurophysiologi-cal, and maturational factors unique to each species and developmental stage.Topographical descriptions of aggressive interactions in animals have beenperformed systematically (Barnett & Marples, 1981; Grant & Mackintosh,1963; see also Pellis, Pellis, & Foroud, Chapter 3, this volume). Although at-tempts have been made to extend the ethological approach in children(Blurton-Jones, 1967; McGrew, 1972) much less has been achieved regardingthe operationalization of specific aggressive behavior in humans (Knutson,1973; Tremblay, 2000). Buss (1961) was among the first to offer a behavioraltaxonomy of human aggression by dichotomizing it along the physical–verbal,active–passive, and direct–indirect dimensions. The active–passive dimension is
Subtypes of Aggression 33

debatable, however, because it is difficult to determine whether behavioral in-activity (e.g., not helping someone in need) was intended or not. The physical–verbal and the direct–indirect dimensions are more defendable. They are moreuseful when applied to humans, although animals often begin aggressive en-counters with threat displays, including vocalizations (Archer, 1988).
In animals, direct aggression is straightforward, involving bodily contactsuch as biting, hitting, or pushing. Some behavioral elements that do not in-volve any physical contact, such as threat and thrust (Grant & Mackintosh,1963), are clearly direct and aggressive, as their objective is to intimidate andthey are often precursors of genuine attack. In humans, Blurton-Jones (1972)also labeled as aggressive a number of nonphysical behaviors that were tem-porally related to other more obvious aggressive actions. In humans, however,the expression of aggression is far more complex and includes a broader as-sortment of direct and indirect actions. Direct forms of aggression comprisephysical assault and a range of verbal behaviors that may be hostile in contentor in tone. These verbal behaviors may occur with or without physical attack.In humans, direct aggression is somehow equivalent to overt aggression(Crick, 1996).
Indirect aggression is at the other end of the behavioral dichotomy, con-sisting of actions aimed at harming others that involve some kind of social in-termediary between the aggressor and the victim. The course of action is oftendelayed and subtle, as there is no physical contact. The psychological effects,however, may persist over a long period. This can be achieved through gossip-ing and ostracizing, which ultimately may damage peer relationships or socialstatus, or through actions aiming at reducing accessibility to resources. Re-search on indirect aggression has grown substantially in the last few years, asthis may be the most prevalent form of aggression during adolescence andadulthood (Björkqvist et al., 1992). It is the only form of aggression that maybe more prevalent in females than in males (Campbell, 1999). Indirect aggres-sion overlaps with two further categories, relational (Crick, Grotpeter, &Bigbee, 2002) and social (Galen & Underwood, 1997) aggression. Relationalaggression emphasizes damage to relationships as a way of harming someone.As this can include face-to-face statements such as “I’m not friends with youanymore,” relational aggression is therefore not always direct (Coyne, Archer,& Eslea, 2004). Social aggression is a wider category that includes both rela-tional and indirect forms, along with expressions such as negative facial ex-pressions or body movements.
SUBTYPING AGGRESSION ON THE BASIS OF ITS ANTECEDENTS
Proximal Contextual and Emotional Elicitors
Proximal antecedents, either contextual or emotional, are the most frequentlyused criteria for discriminating subtypes of aggression, particularly in animals.A wide range of social and nonsocial contextual/situational variables have
34 T H E D E V E L O P M E N T O F A G G R E S S I O N

been used to elicit aggressive behavior in animals (Archer, 1976, 1988). Socialelicitors include the presence of a prey or exposure to a same-species, same-sex conspecific either in the home cage (resident–intruder paradigm) or in anovel environment. These social encounters are often combined with prior orconcomitant experimental manipulations (e.g., social isolation, food depriva-tion, electric shock) that alter the neurobehavioral state of the potential ag-gressor. These predisposing factors, which influence the potency of theelicitors (Hinde, 1974), represent an important issue in aggression research.
A decisive factor is the presence or absence of a threatening elicitor. Whena situation is clearly threatening or perceived as threatening, an arsenal of be-havioral responses is put in motion to protect the individual from pain, injury,and, possibly, death. Responses to threat range from harmless reactions, suchas heightened immobility (i.e., freezing, crouching), escape, and holding offthe threatening assailant with stretching of the upper limbs (boxing), to fierceattack and delivery of noxious stimuli. This constellation of responses hasbeen traditionally subsumed under the label “agonistic behavior” (Scott &Fredericson, 1951).
The notion of aggression as a response to potential threat to an animal’swelfare is central to theories that conceive of aggression as an aversion,whereby the animal is motivated to avoid a situation. This contrasts with anappetite, whereby the animal is motivated to approach or seek out a situation(Craig, 1928). Aversion-based theories seek to identify the mechanism throughwhich animals or humans detect threats, and they include the frustration–aggression hypothesis (Dollard et al., 1939), the discrepancy model of animalaggression (Archer, 1976), and the theory of threatened egotism (Baumeister,Smart, & Boden, 1996).
Pain is another “emotional” antecedent of aggressive behavior. Animalsexposed to painful stimulation (e.g., electrical shocks, sudden heat, bite) willstart fighting against each other or will attack an inanimate object if testedalone (Archer, 1989/1990; Ulrich & Azrin, 1962). Blanchard and Blanchard(1981) have convincingly argued that such pain-induced aggression is analo-gous to fear-induced defensive aggression. Archer (1989/1990) suggested that,although this is the case in laboratory setups involving repeated painfulshocks, an isolated single painful stimulus may under natural conditionsevoke anger-induced offensive aggression. Overall, aggressive behavior can begenerated by a variety of external stimuli that are threatening or painful, andthese stimuli activate, to different degrees, both the peripheral and the centralnervous systems. In pain-induced aggression, activation of the nociceptorsand/or thermal receptors sets in motion neural pathways that control the ex-pression of aggressive behavior, and, in this case, the threshold for fear is eas-ily reached. Then, determining whether aggression is mediated by fear or an-ger will depend on which brain regions are activated during attack.
Under threatening circumstances, aggression may be offensive or defen-sive (Blanchard & Blanchard, 1989), that is, anger or fear motivated. In hu-mans, responses to provocation broadly similar to threats in animals have
Subtypes of Aggression 35

been termed reactive aggression. Thus, reactive aggression is an impulsive,negatively valenced act displayed in response to a threat or provocation(Dodge, & Coie, 1987; Vitaro, Gendreau, Tremblay, & Oligny, 1998). Itstems from the frustration–aggression hypothesis that viewed aggression as a“primordial reaction . . . whenever pleasure-seeking or pain-avoiding behav-ior is blocked” (Dollard et al., 1939, p. 21). Interestingly, it is not so muchcontextual stimuli per se that determine whether aggression will be labeled asreactive, but the perception that the individual has and what he or she makesof it. Indeed, an interesting finding in human research is that reactively aggres-sive children more readily perceive an ambiguous situation as threatening orhostile (Dodge & Coie, 1987; Vitaro, Brendgen, & Tremblay, 2002). Thisattributional/perceptual bias and behavioral hypersensitivity to what typicallyshould be mild, nonthreatening stimulation is reminiscent of what is observedin animals after prolonged social isolation (Gendreau, Gariépy, Petitto, &Lewis, 1998) or after successive defeat experiences (Keeney & Hogg, 1999).These animal paradigms may be more relevant to human reactive aggressionthan paradigms using clear life-threatening conditions. Conversely, proactiveaggression occurs with more forethought and does not seem to be associatedwith any apparent proximal elicitor. It is more controlled, more premeditated,and less emotionally reactive. It can be understood in terms of social learning,as a result of previous external reinforcement (Bandura, 1973; Patterson &Cobb, 1973).
Discriminant and convergent validity of these subtypes of aggression hasbeen provided by an increasing number of empirical reports (Dodge & Coie,1987; Poulin & Boivin, 2000; Pulkkinen, 1996; Vitaro et al., 1998, 2002).Contrary to the dichotomy hostile–instrumental, which is determined by thenature of the motivation (goal is to hurt vs. goal is to gain), reactive andproactive forms of aggression are differentiated by the presence of a prior pro-vocative event and the short delay before enactment. The distinction betweenreactive and proactive aggression, however, is problematic, as it depends onno provocation being identified, and what constitutes a provocation variesconsiderably between individuals. Looking at someone the wrong way may beclassed as a provocation in a subculture where everyone is on the lookout forsigns of disrespect. Having said this, the category proactive aggression (i.e.,aggression without obvious provocation or threat) may alert us to cases, pri-marily among young men, where fights are sought for no apparent reasonother than to inflict a physical defeat on another person.
Brain Mechanisms
The use of psychic or inner states to qualify aggression is not necessary whenexamining the direct and observable effects of lesions and stimulations of spe-cific areas of the brain. The brains of humans and other mammals share manystructures, pathways, and neurochemical properties, reflecting the long andprogressive transformation of the brain during evolution. In the last four de-
36 T H E D E V E L O P M E N T O F A G G R E S S I O N

cades an increasing body of evidence has accumulated describing the role ofspecific neural circuits in the expression of different types of aggression. Nu-merous sites within the neocortex and subcortical structures have been identi-fied as central in mediating aggressive behavior. Moyer (1968, 1973) wasamong the first to relate specific neural circuitries and distinctive neuro-endocrinological status to subtypes of aggression. Particular attention wasgiven to the amygdaloid complex, the various hypothalamic nuclei, and otherparts of the so-called limbic system (e.g., septum, cingulate cortex). Moyer(1973) argued that each of the subtypes of aggression that he had previouslyidentified (except for instrumental aggression) had its own set of neural cir-cuits. Functions associated with the expression of specific forms of aggressivebehavior have indeed been localized in the brain (Valzelli, 1981; Panksepp,1998). As Panksepp pointed out, however, there are more subtypes of aggres-sion based on the proximal contextual antecedents than there are subtypesbased on neural processes. In other words, similar brain circuitry may allowdifferent types of aggression to be expressed; what differs are the eliciting cir-cumstances and the perceptual processing. As we will see, this has a significantimpact on our attempt to produce a parsimonious taxonomy of aggression.
Panksepp (1998) denoted distinct neural circuitry for no more than threesubtypes of aggression in the brains of rats and cats: predatory aggression, af-fective or rage-like aggression (which may coincide with defensive aggressionidentified by Blanchard & Blanchard, 1989); and intermale aggression (whatthe Blanchards called offensive aggression). So-called predatory aggression (orquiet-biting attack in the laboratory) can be generated by stimulation of thedorsolateral hypothalamus and involves activation of the ventral part of theperiaqueductal gray. It is believed to be primarily mediated by what Panksepp(1998) termed the seeking or the appetitive motivational system of the brain.If so, this contrasts with the emphasis on aversive motivation in most theoriesof aggression (see above). This system involves not only the lateral hypothala-mus but also most structures that are innervated by neurons releasing dopa-mine and serotonin. Abnormal development of the prefrontal cortex has beenassociated with antisocial personality disorder (Raine, Lencz, Bihrle, LaCasse,& Colletti, 2000), and people diagnosed with this disorder have been shownto display more proactive forms of aggression (Chase, O’Leary, & Heyman,2001). This would be consistent with their seeking out situations in which todisplay aggressive actions.
In contrast, the affective subtype of aggression is mediated by differentand more primitive (Archer, 1988) interactive neural circuits, what Panksepp(1998) termed the fear system and the rage system. Affective aggression in-volves the ventrolateral–medial hypothalamus, several amygdaloid nuclei, andthe dorsal part of the periaqueductal gray, to cite just a few (Gregg & Siegel,2001). It is important to note that this subcortical circuitry operates relativelyindependently of neocortical input. The neurophysiological basis of intermale(or intrasexual) aggression has not been well established. Panksepp (1998)mentioned that the brain circuitry of intermale aggression may interact with
Subtypes of Aggression 37

both the seeking and rage systems, but is somehow relatively independent.The evidence for this is clearly not overwhelming, but Panksepp (1998) re-ported that brain lesions that impair predatory and affective aggression do notaffect intermale aggression.
Obviously, this is an extremely simplified account of very complex brainmechanisms involving many more structures and neurotransmitter systems(see Gregg & Siegel, 2001). Although primitive forms of aggression, such asthose elicited by stimuli perceived as painful, threatening or annoying, may beprocessed through brain circuitry highly similar to that of other species, thesophistication and diversification of aggression in humans necessarily impliesthe existence of brain processes unique to our species. Our neocortex, whichdoes not complete its maturation before adulthood (Giedd et al., 1999), playsan important inhibitory role in more primitive brain structures, therefore con-trolling the expression of social/emotional reactivity. The gradual transforma-tion of the brain during development may explain the progressive shift in theexpression of aggression from early childhood to adulthood, that is, direct, re-active, and physical forms of aggressive behavior being gradually substitutedby more indirect, controlled, and nonphysical forms (see Vaillancourt, Chap-ter 8, this volume).
Developmental Issues
What differentiates humans from animals is not only the increased complexityof the human brain, the advanced cognitive skills and social world, but thelonger developmental period that is necessary for organizing behavior (Cairns,1979; Hinde, 1974). As mentioned in the preceding section, the expression ofaggression changes throughout ontogeny, with physical aggression reaching itspeak in early childhood (Tremblay et al., 1999) and relationally orientedforms of aggression gradually emerging in late childhood/early adolescence(Björkqvist et al., 1992). Hartup (1974) observed that aggression developsfrom being first object-oriented (instrumental) to being more person-oriented(hostile) aggression (see also Caplan, Vespo, Pedersen, & Hay, 1991).
Hinde (1992) proposed one of the few categorizations of aggression ex-clusive to childhood. He discriminated instrumental (or specific) aggression(aiming at gaining or retrieving an object or situation), teasing aggression (ag-gression unrelated to acquiring a specific object or situation), defensive ag-gression (in response to an attack), and game aggression (that results fromrough-and-tumble play). Hinde suggested that childhood instrumental and de-fensive aggression were somehow equivalent to reactive–impulsive aggressionas observed in adulthood. Teasing aggression had its adult counterpart underthe label of “spontaneous aggression.” No further distinction between impul-sive and spontaneous aggression and no adult correspondence for game ag-gression were provided.
A social–developmental analysis of the functional and expressive charac-teristics of aggression from early childhood to adulthood is indispensable for
38 T H E D E V E L O P M E N T O F A G G R E S S I O N

our understanding of the different ontogenetic pathways to more sophisti-cated forms of aggression. Hinde’s distinction between subtypes of aggressionin childhood was clearly a step toward establishing a developmental topologyof aggression. One may question, however, the need to use different taxonom-ical labels for children and adult aggression, especially when there is corre-spondence between the different subtypes. In addition, characterizing oneform of aggression as impulsive and another as spontaneous is confusing.Finally, game aggression (or rough-and-tumble play) is clearly motivationallydistinct from other forms of aggression (Blurton-Jones, 1972; Smith, 1974). Inthe end, we need to agree on a general taxonomy that is applicable throughoutdevelopment, not only to a specific period. Once this is achieved, the develop-mental trajectory of the different subtypes of aggression can be investigated.
The onset and stability of aggression during ontogeny has been an impor-tant issue in developmental and clinical sciences. Moffitt (1993) proposed adevelopmental taxonomy of conduct disorders based on the onset and persis-tence of antisocial behavior (which relates to aggression). Specifically, conductdisorders could be described as life-course persistent (behavioral problemsstart during childhood and persist throughout childhood up to adolescenceand adulthood) or as adolescence-limited (when problems emerge and endduring adolescence) Discriminating between early- versus late-onset conductdisorders has been a valuable clinical classification (American Psychiatric As-sociation, 1994).
The relationship between individual development and the transformationof aggression over time in boys and girls, in both expression and function, iscentral to the science of aggression. We now recognize the gender-dependentdevelopment and use of physical aggression (Tremblay et al., 1999) and thegradual emergence of hostile (Caplan et al., 1991; Hartup, 1974) and indirect(Björkqvist et al., 1992) aggression. Similarly, reactive aggression seems to ap-pear first (as tantrums) or may be a more prevalent form of aggression earlyon (Loeber & Hay, 1997). However, children prone to display proactive formsof aggression may be more at risk of exhibiting delinquent activities in adoles-cence (Vitaro et al., 2002). Finally, recent evidence suggests that reactive andproactive aggression in boys and girls may have different etiological anteced-ents (Connor, Steingard, Anderson, & Melloni, 2003). Many important issuesremain to be investigated regarding the onset and developmental trajectory ofthe different forms of aggression across gender and sociocultural background.
AN INTEGRATIVE MODEL OF AGGRESSION
The goal of this chapter is to provide a summary of conceptual and theoreticalissues related to the categorization of different subtypes of aggression in hu-mans and animals. Finding an appropriate taxonomy seems to be a recurrentproblem in aggression research. A taxonomy is, by definition, a classificationbased on similarities of a specified characteristic. Suggesting a taxonomy for
Subtypes of Aggression 39

subtypes of aggression based on a single criterion or for a single species is onething, but conceiving one that encompasses the multiple ontogenetic andphylogenetic aspects of this phenomenon is more challenging. Aggression isnot a behavior, or a set of genes, or a brain mechanism. It is a complex socialphenomenon that conveys so many meanings, takes so many forms, resultsfrom so many proximal and distal antecedents, and has so many consequencesthat a universal taxonomy seems an unachievable puzzle. It has identifiableunderlying brain processes, but those are dependent upon context, emotionalstate, and previous learning experience (reinforcement). It is important to notethat aggression is a phenomenon in constant transformation, both phylogen-etically and ontogenetically. Hinde (1974) summarized well the challenge ofelaborating a taxonomy for subtypes of aggression in both humans and ani-mals: “The range validity of any generalizations we make is inversely relatedto their precision. As more diverse phenomena are included within our cate-gory of aggressive behavior, our generalizations inevitably become less pre-cise” (p. 4).
The primary problem with respect to categorizing aggression into differ-ent subtypes is to select a parsimonious set of criteria that integrate mostforms of aggression. In this regard, some criteria, despite being relevant to ourbetter understanding of aggression, do not appear to be essential for establish-ing a categorization. Harm to others, for instance, either physically or psycho-logically induced, may be the first, more expeditious consequence of aggres-sion, but it has little taxonomic value. Therefore, we settled on a simplifiedtaxonomic model that encompasses the most significant and discriminativefeatures of aggression (Figure 2.2). These include the antecedent (to determinewhether aggression is proactive or reactive), the expression (to determinewhether aggression is direct or indirect), and the function (to determinewhether aggression was produced only to hurt or to benefit socially). Otherimportant elements of aggression, such as distal antecedents (past experience,biological or genetic predisposition), aggressor’s emotional–perceptual bias,basic neurobiological mechanisms, and learning, were also integrated.
The first step in identifying the form of aggression should be to determinewhether there is a proximal contextual elicitor. If there is no apparent proxi-mal antecedent, then aggression is proactive. This form of aggression is pri-marily controlled by the neocortex, more particularly the prefrontal cortexand the descending corticolimbic pathways. When a proximal antecedent canbe identified, then aggression is said to be reactive. This behavioral output in-volves various motivational/emotional states (e.g., fear, pain, anger, or annoy-ance) that are under the control of subcortical, primarily limbic, structures. Inreactive aggression, perception of the eliciting stimulus is the key for judgingthe pathological nature of the response. Hypersensitivity to certain contextsand stimulations may indicate heightened anxiety or heightened fearfulness(paranoia). Both proactive and reactive aggression can be expressed directlyor indirectly, and they can target either the source of stimulation or a substi-tute person/object (displaced aggression). Then the aggressive action may pro-
40 T H E D E V E L O P M E N T O F A G G R E S S I O N

vide a pleasurable reward from harming (hostile aggression), a social/material-istic reward (instrumental aggression), or both. Following these consequences,learning (reinforcement) will take place and certainly influence individual de-velopment.
CONCLUDING REMARKS
Aggression is always a timely topic. The news media provide an incessant flowof information on the shocking nature of human behavior. We were just fin-ishing this chapter when we heard the news that three teenage girls were fac-ing charges of attempted murder for allegedly trying to poison a classmate byadding copper sulfate, a highly toxic substance, to her drink. What aggressivecategory does poisoning fit into? It undoubtedly relates to physical harm, butwithout a clear, direct physical action. It is a physical action but with an indi-rect, concealed behavioral twist. Although it is a planned, proactive-like ag-gressive action, it is likely a retaliatory response to some prior event or situa-tion. This is a good example of the difficulty in fitting every act of aggressioninto a definite category or model. Do we need more research to produce abetter taxonomy? Reaching a consensus on this issue may not be as necessaryas continuing our thorough analysis of the wide range of proximal and distal
Subtypes of Aggression 41
FIGURE 2.2. A simplified taxonomical model of aggression based on context, neuroaffectiveprocesses, consequences on self, and learning.

forces that influence the development and expression of aggression (Cairns,1979). It is important to refine our knowledge of the many genetic, biological,and social factors that induce, facilitate, or maintain aggression throughoutdevelopment. Eventually, a universally agreed-upon taxonomy, at once parsi-monious and comprehensive and integrating both animal and human aggres-sion, may emerge.
REFERENCES
American Psychiatric Association. (1994). Diagnostic and statistical manual of mentaldisorders (4th ed.). Washington, DC: Author.
Archer, J. (1976). The organization of aggression and fear in vertebrates. In P. P. G.Bateson & P. Klopfer (Eds.), Perspectives in ethology (Vol. 2, pp. 231–298). NewYork: Plenum Press.
Archer, J. (1988). The behavioural biology of aggression. Cambridge, UK: CambridgeUniversity Press.
Archer, J. (1989/1990). Pain-induced aggression: An ethological perspective. CurrentPsychology: Research and Reviews, 8, 298–306.
Archer, J., & Browne, K. (1989). Concepts and approaches to the study of aggression. InJ. Archer & K. Browne (Eds.), Human aggression: Naturalistic approaches (pp. 3–24). London: Routledge.
Ardrey, R. (1966). The territorial imperative. New York: Atheneum.Atkins, M. S., Stoff, D. M., Osborne, M. L., & Brown, K. (1993). Distinguishing instru-
mental and hostile aggression: Does it make a difference? Journal of AbnormalChild Psychology, 21, 355–365.
Bainton, R. J., Tsai L. T., Singh, C. M., Moore, M. S., Neckameyer, W. S., & Heberlein, U.(2000). Dopamine modulates acute responses to cocaine, nicotine and ethanol inDrosophila. Current Biology, 10, 187–194.
Bandura, A. (1973). Social learning theory and aggression. In J. F. Knutson (Ed.), Thecontrol of aggression: Implications from basic research (pp. 201–250). Chicago: Al-dine.
Barnett, S. A., & Marples, T. G. (1981). The “threat posture” of wild rats: A social signalor an anthropomorphic assumption? In P. F. Brain & D. Benton (Eds.), Multidisci-plinary approaches to aggression research (pp. 39–52). Amsterdam: Elsevier/NorthHolland Biomedical Press.
Baron, R. A. (1977). Human aggression. New York: Plenum Press.Baumeister, R. F., Smart, L., & Boden, J. M. (1996). Relation of threatened egotism to vi-
olence and aggression: The dark side of high self-esteem. Psychological Review,103, 5–33.
Berkowitz, L. (1981). The concept of aggression. In P. F. Brain & D. Benton (Eds.),Multidisciplinary approaches to aggression research (pp. 3–15). Amsterdam: Elsevier/North Holland Biomedical Press.
Berkowitz, L. (1993). Aggression: Its causes, consequences and control. New York:McGraw-Hill.
Björkqvist, K., Lagerspetz, K. M. J., & Kaukiainen, A. (1992). Do girls manipulate andboys fight?: Developmental trends in regard to direct and indirect aggression. Ag-gressive Behavior, 18, 117–127.
42 T H E D E V E L O P M E N T O F A G G R E S S I O N

Blanchard, D. C., & Blanchard, R. J. (1989). Experimental animal models of aggression:What do they say about human behavior? In J. Archer & K. Browne (Eds.), Humanaggression: Naturalistic approaches (pp. 94–121). London: Routledge.
Blanchard, R. J., & Blanchard, D. C. (1981). The organization and modelling of animalaggression. In P. F. Brain & D. Benton (Eds.), The biology of aggression (pp. 529–562). Rockville, MD: Sijhoff & Noordhoff.
Blurton-Jones, N. G. (1967). An ethological study of some aspects of social behaviour ofchildren in nursery school. In D. Morris (Ed.), Primate ethology (pp. 347–368).Chicago: Aldine.
Blurton-Jones, N. G. (1972). Categories of child–child interaction. In N.G. Blurton Jones(Ed.), Ethological studies of child behaviour (pp. 97–127). London: CambridgeUniversity Press.
Bushman, B. J., & Anderson, C. A. (2001). Is it time to pull the plug on the hostile vs. in-strumental aggression dichotomy? Psychological Review, 108, 273–279.
Buss, A. H. (1961). The psychology of aggression. New York: Wiley.Bygott, J. D. (1972). Cannibalism among wild chimpanzees. Nature, 238, 410–411.Cairns, R. B. (1979). Social development: The origins and plasticity of interchanges. San
Francisco: Freeman.Campbell, A. (1999). Staying alive: Evolution, culture, and women’s intrasexual aggres-
sion. Behavioral and Brain Sciences, 22, 203–252.Caplan, M., Vespo, J. E., Pedersen, J., & Hay, D. F. (1991). Conflict and resolution in
small groups of one- and two-year-olds. Child Development, 62, 1513–1524.Chase, K. A., O’Leary, K. D., & Heyman, R. E. (2001). Categorizing partner-violent men
within the reactive-proactive typology model. Journal of Consulting and ClinicalPsychology, 69, 567–572.
Chen, S., Lee, A. Y., Bowens, N. M., Huber, R., & Kravitz E. A. (2002). Fighting fruitflies: A model system for the study of aggression. Proceedings of the National Acad-emy of Science USA, 99, 5664–5668.
Connor, D. F., Steingard, R. J., Anderson, J. J., & Melloni, R. H., Jr. (2003). Gender dif-ferences in reactive and proactive aggression. Child Psychiatry and Human Devel-opment, 33, 279–294.
Coyne, S. M., Archer, J., & Eslea, M. (2004). We’re not friends anymore! Unless . . . ”:The frequency and harmfulness of relational, indirect and social aggression. Manu-script submitted for publication.
Craig, W. (1928). Why do animals fight ? International Journal of Ethics, 31, 264–278.Crick, N. R. (1996). The role of overt aggression, relational aggression, and prosocial be-
havior in the prediction of children’s future social adjustment. Child Development,67, 2317–2327.
Crick, N. R., Grotpeter, J. K., & Bigbee, M. A. (2002). Relationally and physically ag-gressive children’s intent attributions and feelings of distress for relational and in-strumental peer provocations. Child Development, 73,1134–1142.
Dodge, K. A., & Coie, J. D. (1987). Social-information-processing factors in reactive andproactive aggression in children’s peer groups. Journal of Personality and SocialPsychology, 53, 1146–1158.
Dollard, J., Doob, L. Miller, N., Mowrer, O., & Sears, R. (1939). Frustration and aggres-sion. New Haven, CT: Yale University Press.
Felson, R. B. (2002). Violence and gender reexamined. Washington, DC: American Psy-chological Association.
Subtypes of Aggression 43

Feshbach, S. (1964). The function of aggression and the regulation of aggressive drive.Psychological Review, 71, 257–272.
Feshbach, S. (1971). Dynamics and morality of violence and aggression. American Psy-chologist, 26, 281–292.
Galen, B. R., & Underwood, M. K. (1997). A developmental investigation of social ag-gression among children. Developmental Psychology, 33, 589–600.
Gendreau, P. L., Gariépy, J. L., Petitto, J. M., & Lewis, M. H. (1997). D1 dopamine recep-tor mediation of social and nonsocial emotional reactivity in mice: Effects of hous-ing and strain difference in motor activity. Behavioral Neuroscience, 111, 424–434.
Gendreau, P. L., Petitto, J. M., Gariépy, J. L., & Lewis, M. H. (1998). D2-like dopaminereceptor mediation of social-emotional reactivity in a mouse model of anxiety:Strain and experience effects. Neuropsychopharmacology, 18, 210–221.
Giedd, J. N., Blumenthal, J., Jeffries, N. O., Castellanos, F. X., Liu, H., Zijdenbos, A.,Paus, T., Evans, A. C., & Rapoport, J. L. (1999). Brain development during child-hood and adolescence: A longitudinal MRI study. Nature Neuroscience, 2, 861–863.
Grant, E. C., & Mackintosh, J. H. (1963). A comparison of the social postures of somecommon laboratory rodents. Behaviour, 21, 246–259.
Gregg, T. R., & Siegel, A. (2001). Brain structures and neurotransmitters regulating ag-gression in cats: Implications for human aggression. Progress in Neuro-Psycho-pharmacology and Biological Psychiatry, 25, 91–140.
Hartup, W. W. (1974). Aggression in childhood: Developmental perspectives. AmericanPsychologist, 29, 336–341.
Hartup, W. W., & De Wit, J. (1974). The development of aggression: Problems and per-spectives. In J. De Wit & W. W. Hartup (Eds.), Determinants and origins of aggres-sive behavior (pp. 595–620). The Hague: Mouton.
Hinde, R. A. (1974). The study of aggression: Determinants, consequences, goals andfunctions. In J. De Wit & W. W. Hartup (Eds.), Determinants and origins of aggres-sive behavior (pp. 3–27). The Hague: Mouton.
Hinde, R. A. (1992). Some complexities in aggressive behaviour. In A. Fraczek & H.Zumkley (Eds.), Socialization and aggression (pp. 3–10). Berlin: Springer-Verlag.
Johnson, R. N. (1972). Aggression in man and animals. Philadelphia: Saunders.Karli, P. (1956). The Norway rat’s killing response to the while mouse: An experimental
analysis. Behaviour, 10, 81–103.Keeney, A. J., & Hogg, S. (1999). Behavioural consequences of repeated social defeat in
the mouse: Preliminary evaluation of a potential animal model of depression. Be-havioral Pharmacology, 10, 753–764.
Knutson, J. F. (1973). Aggression as manipulatable behavior. In J. F. Knutson (Ed.), Thecontrol of aggression: Implications from basic research. (pp. 253–295). Chicago:Aldine.
Loeber, R., & Hay, D. (1997). Key issues in the development of aggression and violencefrom childhood to early adulthood. Annual Review of Psychology, 48, 371–410.
McGrew, W. C. (1972). An ethological study of children’s behaviour. London: AcademicPress.
Moffitt, T. E. (1993). Adolescent-limited and life-course-persistent antisocial behavior: Adevelopmental taxonomy. Psychological Review, 100, 674–701.
Moyer, K. E. (1968). Kinds of aggression and their physiological basis. Communicationsin Behavioral Biology, 2, 65–87.
44 T H E D E V E L O P M E N T O F A G G R E S S I O N

Moyer, K. E. (1973). The physiological inhibition of hostile behavior. In J. F. Knutson(Ed.), The control of aggression: Implications from basic research (pp. 9–38). Chi-cago: Aldine.
Panksepp, J. (1998). Affective neuroscience: The foundations of human and animal emo-tions. New York: Oxford University Press.
Patterson, G. R., & Cobb, J. A. (1973). Stimulus control for classes of noxious behavior.In J. F. Knutson (Ed.), The control of aggression: implications from basic research(pp. 145–199). Chicago: Aldine.
Poulin F., & Boivin, M. (2000). Reactive and proactive aggression: Evidence of a two-fac-tor model. Psychological Assessment, 12, 115–122.
Pulkkinen, L. (1987). Offensive and defensive aggression in humans: A longitudinal per-spective. Aggressive Behavior, 13, 197–212.
Pulkkinen, L. (1996). Proactive and reactive aggression in early adolescence as precur-sors to anti- and prosocial behavior in young adults. Aggressive Behavior, 22, 241–257.
Raine, A., Lencz, T., Bihrle, S., LaCasse, L., & Colletti, P. (2000). Reduced prefrontalgray matter volume and reduced autonomic activity in antisocial personality disor-der. Archives of General Psychiatry, 57, 119–127.
Rule, B. G. (1974). The hostile and instrumental functions of human aggression. In J. DeWit & W. W. Hartup (Eds.), Determinants and origins of aggressive behavior (pp.121–141). The Hague: Mouton.
Scott, J. P., & Fredericson, E. (1951). The causes of fighting in mice and rats. Physiologi-cal Zoology, 24, 273–309.
Sears, R. R. (1961). Relation of early socialization experiences to aggression in earlychildhood. Journal of Abnormal and Social Psychology, 63, 466–492.
Smith, P. K. (1974). Aggression in a preschool playgroup: Effects of varying physical re-sources. In J. De Wit & W. W. Hartup (Eds.), Determinants and origins of aggressivebehavior (pp. 97–105). The Hague: Mouton.
Tedeschi, J. T., & Felson, R. B. (1994). Violence, aggression, and coercive actions. Wash-ington, DC : American Psychological Association.
Tremblay, R. E. (2000). The development of aggressive behaviour during childhood:What have we learned in the past century? International Journal of Behavioral De-velopment, 24, 129–141.
Tremblay, R. E., Japel, C., Pérusse, D., Boivin, M., Zoccolillo, M., Montplaisir, J., &McDuff, P. (1999). The search for the age of “onset” of physical aggression:Rousseau and Bandura revisited. Criminal Behavior and Mental Health, 9, 24–39.
Ulrich, R. E., & Azrin, N. H. (1962). Reflexive fighting in response to aversive stimula-tion. Journal of the Experimental Analysis of Behavior, 5, 511–520.
Ulrich, R. E, Dulaney, S., Arnett, M., & Mueller, K. (1973). An experimental analysis ofnonhuman and human aggression. In J. F. Knutson (Ed.), The control of aggression:Implications from basic research (pp. 79–111). Chicago: Aldine.
Ulrich, R. E., Johnston, M., Richardson, J., & Wolff, P. (1963). The operant conditioningof fighting behaviour in rats. Psychological Record, 13, 465–470.
Valzelli, L. (1981). Psychobiology of aggression and violence. New York: Raven Press.Vitaro, F., Brendgen, M., & Tremblay, R. E. (2002). Reactively and proactively aggres-
sive children: Antecedents and subsequent characteristics. Journal of Child Psychol-ogy and Psychiatry, 43, 495–505.
Subtypes of Aggression 45

Vitaro, F., Gendreau, P. L., Tremblay, R. E., & Oligny, P. (1998). Reactive and proactiveaggression differentially predict later conduct problems. Journal of Child Psychol-ogy and Psychiatry, 39, 377–385.
Wilson, E. O. (1980). Sociobiology: The abridged edition. Cambridge, MA: HarvardUniversity Press.
Zillman, D. (1978). Hostility and aggression. Hillsdale, NJ: Erlbaum.
46 T H E D E V E L O P M E N T O F A G G R E S S I O N

THE DEVELOPMENT OF AGGRESSIONPlay Fighting
3
Play FightingAggression, Affiliation, and the
Development of Nuanced Social Skills
SERGIO M. PELLIS, VIVIEN C. PELLIS, and AFRA FOROUD
In traditional New Guinean society, if two men—strangers to each other—were to meet on a path between their respective villages, they would thenlikely, if they wished to avoid bloodshed, begin to list all the people to whomthey were related in the hope of discovering some common ground betweenthem (Diamond, 1997). Although having the skill to win in a potentially ag-gressive encounter is important, this New Guinean practice illustrates thathaving the skill to avoid unnecessary aggression may be just as important. It isbecoming clear that aggression is but a part of a larger tool kit for gaining ad-vantage in a complex social world. This appears to be true not just for hu-mans, but for many social mammals (de Waal & Aureli, 2000). In such aworld, knowing who and when to fight becomes important, and thus so doeshaving the cognitive skills to gather the information required to make the ap-propriate decisions. Placing aggression into a broader social context has impli-cations for the understanding of the developmental processes that shape ag-gression. The problem lies not only in learning how and when to useaggression, but also in knowing how to assess the situation so as to determinewhether a nonaggressive approach would be a more suitable solution. In thischapter we show that play fighting can provide a vehicle for the developmentof such skills.
Play fighting resembles serious fighting in that the partners compete foraccess to some advantage. Most often, this advantage takes the form of bitingor striking another individual on a particular body target (Aldis, 1975). Thetargets competed over during play fighting can be those that in adulthood are
47

used in conspecific aggression, sex, or predation. Despite these differences intargets, all forms of play fighting have play-typical rules of attack and defensein common (Pellis & Pellis, 1998a). In contrast, during conspecific aggressiveencounters, in the early stages of sexual encounters and in predatory encoun-ters, the opponents follow the rules of attack and defense typical of seriousfighting. For play fighting to remain playful, it needs to follow the 50:50 rule(Aldis, 1975; Altmann, 1962). That is, each pairmate has to win close to 50%of the playful encounters. Such a win:loss ratio requires that the rules of at-tack and defense differ from those that apply to serious fighting (Pellis &Pellis, 1998a).
When an attack is launched during a serious fight, the attacking animalhas to guard against retaliation from the opponent. Typically, in order to doso, the attacker simultaneously incorporates some defensive tactic into its at-tack, which limits the defender’s opportunity to counterattack (Pellis, 1997).Similarly, when defending itself against a serious attack, the defender uses anintensity in its defense that reduces the likelihood of a successful penetrationby the attacker (Pellis, 1997). In contrast, during play fighting, the attackerrarely incorporates defensive maneuvers in its attack, thus facilitating success-ful counterattacks by the defender (Pellis & Pellis, 1998a). In addition, whendefending itself against a playful attack, the defender will use an intensity ofdefense that is lower than that in serious fights, increasing the likelihood of asuccessful contact by the attacker (Pellis & Pellis, 1998a). That is, in playfighting, the tactics of attack and defense are decoupled so as to ensure thateach animal can successfully contact the partner. This contrasts sharply withserious fighting, in which an animal may deliver injurious, or even lethal, at-tacks if the opponent leaves its target exposed (Blanchard, Blanchard, Taka-hashi, & Kelly, 1977; Geist, 1971; Pellis & Pellis, 1992).
In the following text, we show that given its rule structure, play fightingcan provide fertile ground for learning a variety of social skills. Detailed stud-ies of the development of play fighting in rats (Pellis, 2002b) provide a modelfor understanding how play fighting may be used for sharpening the socialskills needed to straddle the subtle divide between competition and coopera-tion effectively (Pellis, 2002a).
PLAY FIGHTING IN RATS1
In rats, play fighting involves attack and defense of the nape (Figure 3.1). Suc-cessful contact with the nape involves gently rubbing the nape with the nose(Pellis & Pellis, 1987; Siviy & Panksepp, 1987). Nape contact is a target typi-cal of adult sexual encounters (Pellis, 1988, 1993), whereas agonistic attacksinvolve bites directed at the lower dorsum or the face (Blanchard et al., 1977;Pellis, 1997). To defend themselves against playful nape contact, rats use thesame tactics of defense that are used in serious aggression, but modify them tomatch both the differences in targets (Pellis & Pellis, 1987) and the differences
48 T H E D E V E L O P M E N T O F A G G R E S S I O N

in the rules of execution of those tactics (Pellis & Pellis, 1998a). Even thoughmost common in the juvenile phase (Meaney & Stewart, 1981; Panksepp,1981; Thor & Holloway, 1984), play fighting in rats persists well into youngadulthood (Adams & Boice, 1983, 1989; Pellis & Pellis, 1991b). Indeed, playfighting between adults occurs among many mammalian species, in both sex-ual and nonsexual encounters (Pellis & Iwaniuk, 1999b, 2000). However,whether the context is sexual or nonsexual, such play fighting appears to beused to assess and manipulate the other animal, be it human or nonhuman(Breuggeman, 1978; Pellis & Iwaniuk, 1999b). When it occurs in subadultsand adults, play fighting appears to be used in two general contexts—for so-cial bonding and for social testing (Pellis & Pellis, 1996; Pellis & Iwaniuk,1999b, 2000; Smith, Fantella, & Pellis, 1999). Social bonding may includecourtship and pair bonding, as well as nonsexual social affiliation. Social test-ing most typically involves some form of jostling for social status. The distri-bution of these functions vary across species, with some species using themonly for affiliation, some using them only for dominance testing, and some us-ing them for both (Pellis, 2002a). In rats, play fighting is used for both thesepurposes (Pellis, Pellis, & McKenna, 1993; Smith et al., 1999).
It should be noted that in rats, as in many other mammals, play fightingis more frequent among males (Pellis & Pellis, 1990, 1997), although prefer-ences for which sex is the most attractive as a play partner may change during
Play Fighting 49
FIGURE 3.1. A sequence of play fighting is shown for a pair of 31-day-old rats. Note that theattacking rat (on the left), lunges toward its partner’s nape. The defending rat avoids napecontact by turning to supine. From this on-top/on-bottom position, both rats make attemptsto contact each other’s napes. Adapted from Pellis and Pellis (1987). Copyright 1987 byWiley. Adapted by permission.

development (Meaney & Stewart, 1981). With regard to the social uses ofplay fighting, only male rats show the context-specific modulation of contentthat is consistent with such uses (Pellis, Field, Smith, & Pellis, 1997). In someother species of mammals, females also appear to use play fighting for socialassessment and manipulation (Pellis, 2002a; Pellis & Iwaniuk, 2000). For thepresent purposes, in this discussion of rats, males are the focus of consider-ation.
Play fighting in rats has several features useful for the analysis of the de-velopment of social skills. First, the difference in targets between play fightingand serious fighting makes it easy to differentiate between these two forms offighting. This is very useful for identifying cases where play fights escalate intoserious fights. By using a switch in target as a marker, it was found that play-ful encounters escalate into serious ones if one of the partners breaks the play-typical rules of attack and defense. That is, if one of the pair stops allowingthe partner an opportunity for reciprocal contact, the partner may switch toan aggressive attack (Pellis & Pellis, 1998a). Second, adults use play fightingin clearly functional contexts, allowing us to identify the social skills neededand so evaluate play fighting earlier in development to ascertain whether theopportunity to acquire such skills is present. To explain what those skills en-tail, we describe the social uses of play fighting among adult male rats. Thisunderstanding then sets the stage for interpreting the developmental changesin play fighting present in this species.
THE ADULT USES OF PLAY FIGHTING
Within a colony, adult male rats form a dominance hierarchy (Blanchard &Blanchard, 1990; Calhoun, 1962; Flannery & Lore, 1977) in which subordi-nates initiate more playful encounters with dominants than they do with eachother (Pellis et al., 1993). When playfully contacted by a dominant, subordi-nates are more likely to roll over onto their backs, a response typical of juve-niles (Panksepp, 1981; Pellis & Pellis, 1987), whereas when contacted by an-other subordinate, they are more likely to remain standing and push againstthe attacker with their flanks (Pellis et al., 1993), a response typical of a domi-nant male (Pellis & Pellis, 1991a, 1992). In the absence of dominance rela-tionships, postpubertal males are more likely to stand and push than to rollover to supine (Pellis & Pellis, 1990; Smith, Field, Forgie, & Pellis, 1996;Smith, Forgie, & Pellis, 1998, Smith et al.,1999). Thus, the increased solicitingof playful contact by subordinates toward dominants, and the juvenile-like re-sponse toward those dominants, suggests that the play fighting is used as ameans of maintaining “friendly” relations with dominants (Pellis et al., 1993).However, not all subordinates are equally obsequious.
The greater the dominance asymmetry between pairmates, the morejuvenile-like the playful responses by the subordinate (Pellis & Pellis, 1992).
50 T H E D E V E L O P M E N T O F A G G R E S S I O N

Indeed, it is those pairmates that are the least asymmetrical in their play re-lations that are the most likely to escalate the playful encounter into a seri-ous fight (Pellis & Pellis, 1991a). Similarly, when male rats that are unfamil-iar with each are placed into a neutral arena, it is the dominant–dominantcombination that is the most likely to escalate into a serious fight (Smith etal., 1999). Close inspection of these escalations suggests that the play fight-ing preceding the serious fight is “rougher.” These observations also suggestthat when animals are testing each other in regard to their opponent’s capa-bility to maintain or gain a position of superior dominance, the play canthen be escalated into a quasi-aggressive intensity. Therefore, there are twoextremes in the style of play that are available to an individual—a gentlerform, which is seen when a subordinate is maintaining familiarity with adominant, and a rougher form, which is seen when one rat is probing an-other one for its weaknesses.
As previously described, these differences in play intensity among adultmale rats can be explicitly converted into formal rules of engagement and canprovide a rat with the basis for judging how such play can be used to assessand manipulate its partner. For example, when a subordinate rat uses playfighting as a means of social bonding with the dominant male in the colony, itbends the 50:50 rule in the dominant’s favor, whereas when a subordinateuses play fighting to probe the dominant for weakness, or when it encountersan unfamiliar rat, it bends the rule in its own favor. In the latter case, the ratcan assess how much of a deviation from equality the opponent will toleratebefore retaliating aggressively. A simple rule of thumb may operate in such ascenario. If the dominant or unfamiliar rat tolerates the bending of the 50:50rule in the opponent’s favor, the opponent may then bend the rule further, un-til the relative status of the pair is reversed. However, if the dominant starts torespond forcefully, there is still time for the opponent to back down before theencounter escalates into a serious fight. Many primates and carnivores thathave affiliative signals that can be used to indicate playful intention may havea capacity for greater flexibility along this playful–serious gradient, as suchsignals can be used to diffuse unwanted escalations (see Bekoff, 1995;Boulton, 1994; Pellis & Pellis, 1996, 1997).
What all these social uses of play fighting have in common is that they in-volve assessment and manipulation of the partner. As such, they are consistentwith evolutionary game theory models that show that if an animal can assessthe opponent’s fighting ability relative to its own, potentially damaging fightscan be avoided (Parker, 1974). The benefits derived from such assessmentcould thus have been the selective advantage for the evolution of play fightingas a social tool in rats and other mammals. What the present analysis offers isa window into the causal mechanisms that make such an assessment possible.Furthermore, given the cognitive demands associated with such functions, thequestion arises as to whether play in the juvenile phase is organized so as tofacilitate the development of these skills.
Play Fighting 51

IMPLICATIONS FOR OUR UNDERSTANDING OF THE PLAY OF JUVENILES
All muroid rodents studied thus far appear to have a form of play fighting thatresembles the species-typical patterns of adult precopulatory behavior (Pellis,1993). The resemblance is in both the body targets attacked and in the defen-sive maneuvers that are used to block these attacks. During postweaning de-velopment, these playful interactions grow increasingly like the adult sexualencounters (Pellis & Pellis, 1998b). The rat, however, exhibits a strikingly dif-ferent developmental pattern. Although in the rat the target competed overduring play fighting is the same as for sexual encounters (Pellis & Pellis,1987), the pattern of defense differs. During sexual encounters the female ratprimarily evades the male’s contact, while during play fighting the attackedpartner does so in only 20–25% of contacts (Pellis & Pellis, 1987). Moreover,the form of the contact promoting defense also differs. During precopulatoryencounters, female rats rarely flip over onto their backs (<10%), typically do-ing so only when mounted (work in progress). However, during play fighting,juvenile rats flip over onto their backs in response to more than 50% of con-tacts. With the onset of puberty, the males switch to partially rotating theirbody axes and pushing the attacker with their flanks (Pellis & Pellis, 1987,1990).
When play fighting emerges before weaning, the predominant pattern ofdefense is the same as that seen in postpubescent male rats—the partially ro-tated defense. This is striking because the partially rotated defensive posturerequires the rat to shift its body weight backward onto its hind feet and to bal-ance its forequarters in a horizontal, cantilevered position, a feat of posturalcoordination beyond the capacity of preweanling rats, which typically fallover. In the days following weaning, the full rotation to supine defensive tacticbecomes predominant (Pellis & Pellis, 1997) and remains so until puberty(Pellis & Pellis, 1990). Therefore, in rats, not only is the defensive componentof play fighting different from that which is present in adulthood at all devel-opmental stages, but the juvenile phase is also marked by a reversal of the pat-tern of defense initiated before weaning and regained at puberty (Pellis,2002b). These highly structured developmental changes suggest that the playpresent in the juvenile phase is organized so as to maximize specific experi-ences. The question is, what are those experiences?
When, in the juvenile phase, a rat is introduced to a partner followingshort-term social isolation (6–24 hours), there is an increase in the level ofplay relative to other social behavior, such as social investigation (Panksepp,1981; Panksepp & Beatty, 1980; Pellis et al., 1997). Increasing the length ofthe isolation (1–2 weeks) also leads to a preferential increase in play(Varlinskaya, Spear, & Spear, 1999), but such longer-term deprivation also re-sults in long lasting deficiencies of social behavior in adulthood (Hol, van denBerg, van Ree, & Spruijt, 1999; van den Berg et al., 1999a). These deficitsseem to be more in the motivation to engage in social behavior than in theability to engage in appropriately sequenced bouts of the social interaction
52 T H E D E V E L O P M E N T O F A G G R E S S I O N

(van den Berg, van Ree, & Spruijt, 1999b). More prolonged social depriva-tion, lasting for all of the juvenile phase, leads to more generalized deficits insocial and nonsocial behavior (DaSilva, Ferreira, Carobrez, & Morato, 1996;Einon, Morgan, & Kibbler, 1978). With regard to social and sexual behavior,these deficits appear to involve a reduced ability to respond to the movementsof the partner (e.g., Larsson, 1978; Einon et al., 1978; Pellis, Field, &Whishaw, 1999). It is important to note that rats that have been socially iso-lated as juveniles have deficits in regulating the intensity of their defensive re-sponses to social partners (Byrd & Briner, 1999; Potegal & Einon, 1989; vanden Berg et al., 1999a) and to the stresses arising from such contact (VonFrijtag, Schot, van des Bos, & Spruijt, 2002). How, then, does wrestling,which arises from supine defense and leads to one partner standing on top andthe other lying on its back (on-top/on-bottom), provide for the development ofvaluable social and nonsocial skills?
When in this on-top/on-bottom configuration, the on-top rat has twoways of standing and thus supporting its weight (Foroud & Pellis, 2003). Inboth cases, the on-top rat stands on the on-bottom rat with its forepaws anduses them to restrain its partner’s movements. The difference between the twopostures is in the placement of the hind paws. In the first posture, the on-toprat places its hind paws on the ground (see Figure 3.1e), whereas in the secondposture the hind paws are placed on its supine partner’s body. However, in thelatter case, the wriggling and squirming supine partner provides an unstablebase; this makes it difficult for the on-top rat to maintain its balance and suc-cessfully restrain its supine partner. When the on-top/on-bottom configurationfirst appears, at about the time of weaning, the on-top rat adopts a more sta-ble on-the-ground placement of its hind paws in approximately 80% of cases(Foroud & Pellis, 2002). This is true for puberty and thereafter (Foroud &Pellis, 2003). Surprisingly, during the juvenile phase, the rate of adopting themore stable placement declines to about 30–40% (Foroud & Pellis, 2002,2003). This decline does not arise as a byproduct of the greater level of play-fulness in the juvenile phase (Foroud & Pellis, 2002), nor is it dependent onsocial experience (Foroud, Whishaw, & Pellis, 2004). Rather, just as in thecase of the pubertal switch to using the complete rotation tactic more often inthe juvenile phase, the switch to the less stable on-top configuration appearsto be a programmed pattern of development.
However, there is a major consequence associated with adopting the lessstable postural configuration—it facilitates the partner’s ability to counterat-tack and so gain the upper hand. The context of 22 role reversals from the on-top/on-bottom position in 11 pairs of male juvenile rats was analyzed. Rolereversals arise when the on-bottom rat successfully counterattacks and re-verses its position relative to its partner. In 70.5% of the role reversals, the on-top rat adopted the less stable postural configuration. That is, role reversalsare significantly more likely when the on-top rat stands on its partner with itshind feet (sign test: X = 9, p < .05). Thus, by standing in the less stable posi-tion, the on-top rat increases its chances of losing the encounter, or perhaps,
Play Fighting 53

more accurately, increases the opportunity for allowing the partner to recipro-cate the contact. These data show that the juvenile phase is peculiar not onlybecause of the increase in the use of the supine defense by defenders (Pellis &Pellis, 1990, 1997), but also because of the change that occurs at this time inthe behavior of the attacking partner (Foroud & Pellis, 2002, 2003). It is as ifby combining an increased likelihood of rolling over to supine by the defenderwith an increased likelihood of standing on the supine partner by the attacker,the rats are exaggerating the possibility for reciprocation by placing them-selves in a more disadvantageous position. This combination is unique to thejuvenile phase. That is, when play fighting is at its most gentle (defense mostlyinvolves rotation to supine and the on-top attacker mostly adopts the unstablestanding position), it is at its most frequent. Conversely, when play fighting isat its most frequent, it is at its least rough (Figure 3.2). Not only may such anexaggerated reciprocation allow for an elevated level of play fighting to occurin the juvenile phase, but it also may provide the learning experiences usefulfor adults to make fine-grain judgments about themselves in relation to theirconspecifics.
The on-top/on-bottom configuration with the less stable standing postureprovides an experience of continuous change. That is, the on-top rat mustcontinuously recalibrate its own position because of the changes that occur inits stability. These changes arise from a combination of its own movementswith those of its supine partner. But at what level of vigor in the supine ani-mal’s squirming should the on-top animal increase or decrease its own vigor in
54 T H E D E V E L O P M E N T O F A G G R E S S I O N
FIGURE 3.2. The developmental changes in “roughness,” as measured by whether the rats usethe partial rotation defense tactic and adopt the stable on-top posture, are contrasted withthe developmental changes in the frequency of play fighting in rats. During the infantile andadult phases, when the frequency of play fighting is low, play is rougher. In contrast, duringthe juvenile phase, when the frequency of play fighting is at its highest, play is less rough.

its attempt to gain the advantage? By having to make these judgments in thecontext of having its own sensorimotor skills compromised, the rat may belearning to integrate cues from its own body with those of its partner and so,at the same time, be learning how to apply appropriate levels of force to ahighly dynamic situation. Several lines of evidence support this possibility.
THE MODIFIED PATTERN OF PLAY FIGHTING IN JUVENILE RATSAS A TOOL FOR ENHANCING SOCIAL–COGNITIVE SKILLS
We have shown that the rat is a species in which the adults use play fightingfor various social functions and the juveniles have a highly modified form ofplay. This is not true for all species that play as juveniles (Pellis & Iwaniuk,1999a). Most other rodents that have been studied do not use play fighting asadults. Indeed, the play fighting of these juveniles mimics that of the adult-typical context from which the play elements are derived (Pellis & Pellis,1998b). Thus, for example, voles not only attack and defend the targets typi-cal of sexual encounters when they play fight, but they also use the tactics ofattack and defense with the same frequencies as are typical of adults (Pellis,1993). Furthermore, species differences between voles, such as the use of thesetactics during play fighting, are consistent with adult differences in the use ofthese same tactics in sexual encounters (Pellis & Pellis, 1998b). As has beennoted, in rats, the discordance between the use of tactics in play fighting andtheir use in adult sexual encounters cannot be so explained. This implies thatthe play fighting present in extant species cannot be viewed as having the samecontent for all species. Rather, with the advent of novel functions, new controlsystems must have evolved so as to regulate the behavior appropriately (Pellis,1993). Again, the rat provides some clues as to how such layering of functionsand control systems may have evolved. Rats decorticated at birth engage inplay as juveniles (Panksepp, Normansell, Cox, & Siviy, 1994; Pellis, Pellis, &Whishaw, 1992) and are able to follow the 50:50 rule. However, they fail toexhibit the juvenile reversal in complete and partial rotation. That is, theypreferentially use the partial rotation tactic before weaning and on into adult-hood (Pellis et al., 1992). Decorticated rats also fail to show the modulation ofthe on-top rat in its standing posture, in that at all ages they most frequentlyuse the least stable position (Foroud et al., 2004). Furthermore, as adults,decorticates do not modulate their defensive tactics with a change in partner.Rather, they mostly use the partial rotation tactic whether confronted by adominant or a subordinate pairmate (unpublished observations based on datacollected in Pellis et al., 1992).
These findings suggest that interacting with a partner using the 50:50 ruleinvolves subcortical neural mechanisms, but that modulating the pattern ofdefense with age, or with different partners, involves cortical systems. Further-more, we have found that these two forms of cortical modulation are, inthemselves, dissociable. Although neonatal ablation of the prefrontal cortex
Play Fighting 55

does not disrupt the age-related modulation, it does appear to disrupt the rela-tionship-based modulation (work in progress). We have also found that ratsthat are reared in isolation during the juvenile phase fail to exhibit the rela-tionship-based modulation (work in progress). Given that the prefrontal cor-tex is not fully mature until puberty (van Eden, Kros, & Ulings, 1990)—thatis, after the peak period of play in the juvenile phase—it is highly likely thatthis structure is involved in learning the subtle skills needed to assess and ma-nipulate other rats during social interactions. The rat thus provides a basis fora model for the evolution of play fighting.
As has been argued, for play fighting to occur, an animal must have theability to use the 50:50 rule. As the neural control systems needed for this ap-pear to be subcortical, they may provide the basis for commonality in thepresence of such play in mammals, birds (Fagen, 1981), and, perhaps, reptiles(Burghardt, 1998).2 Most species that play fight do not exhibit the same rangeof elaboration of the 50:50 rule as do rats; most have only the basic,subcortical reciprocation mechanism. In rats, the developmental and partner-based modifications of the 50:50 rule appear to involve additional corticalsystems. Furthermore, cortical mechanisms appear to be needed to gain thecognitive benefits of such play (Pellis et al., 1992). Therefore, we predict thatonly species with a modified form of play, such as that which exists in rats,would suffer from cognitive deficiencies if deprived of play experience in thejuvenile phase. Data from experiments by Einon and her colleagues appear tosupport this prediction. Rats, but not three other commonly used laboratoryrodents, suffer long lasting cognitive deficits if socially isolated in the juvenilephase (Einon, Humphreys, Chivers, Field, & Naylor, 1981). Clearly, for a va-riety of species, there is a necessity for detailed comparisons of the organiza-tion, development, and functions of play fighting to be mapped against the ef-fects of social isolation.
Another aspect of the comparative distribution of play fighting is thateven for species that have the additional rat-like mechanisms for modulatingthe 50:50 rule, the pattern of juvenile play experience is likely to vary withspecies-specific demands in the subtlety needed to modulate social interac-tions. We predict that the degree of rehearsal in moving along the 50:50 gradi-ent should correlate with the degree of flexibility (and hence uncertainty) inadult social relationships. A preliminary comparison of the play fighting oftwo species of macaques suggests such a possibility (work in progress). Al-though the targets and tactics of the two species are the same, the magnitudeof the deviations from the 50:50 rule that are tolerated before play fights areterminated differ. Whereas Japanese macaques (Macaca fuscata) tolerate onlya narrow range of rule attenuation, in Tonkean macaques (Macaca tonkeana)the range is quite large. The social organization of Japanese macaques is rig-idly hierarchical, with subordinates usually accepting punishment from adominant. In contrast, the social hierarchy of Tonkean macaques is more re-laxed, with subordinates likely to retaliate against transgressions by a domi-
56 T H E D E V E L O P M E N T O F A G G R E S S I O N

nant (Thierry, 2000). Thus, the need to assess subtle cues and to be more cog-nizant of the boundaries of manipulation is likely to be more important inTonkean than in Japanese macaques.
Individual differences also offer an opportunity to study the developmen-tal, social, and neural processes that regulate reciprocity during play fighting.For example, some rats are more playful than others, and there are individualdifferences in regard to responsiveness to the movements of a partner. Some ofthese individual differences are related to endogenous levels of dopamine ac-tivity (Pellis & McKenna, 1992). A more striking example has been shown forpopulations of rhesus macaques (Macaca mulatta). About 5–10% of the juve-nile males are hyperactive and impulsive. After a few playful interactions withthese animals, other juveniles avoid further contact with them. It appears thatbecause of their impulsiveness, their play quickly becomes rough, often esca-lating into serious aggression. These impulsive individuals have deficits intheir serotonin metabolism (Suomi, 2002). Thus, both for rats and monkeys,individual differences in behavior can be linked to differences in the activity ofparticular neurochemical systems.
In regard to behavior, the example of the rhesus monkeys (described ear-lier) serves to underscore the importance of following the rules governing playfighting so as to avoid escalation into full-blown aggression (Boulton, 1994).Also of relevance to the present discussion is the finding that rearing condi-tions with appropriate social experiences can attenuate the impulsiveness ofthese individuals (Suomi, 2002). These data thus reinforce the claim madehere, that for mammals with at least the level of play organization that existsin rats, emotional and cognitive skills that are relevant to social behavior canbe influenced by the experiences gained from play fighting as juveniles.
The neural and comparative data presented here support the idea thatwhen play fighting is used by adults for social assessment and manipulation,juveniles’ play fighting is modified so as to enhance the opportunity to maxi-mize the experiences that refine the social–cognitive skills useful for adult-hood. Rats show all facets of these combined changes—play is retained foradult purposes, the content of juvenile play is modified relative to that of pre-and postjuvenile play, and the addition of novel cortical mechanisms enableage- and context-specific modulation of the tactics of attack and defense, andthus of the 50:50 rule. Furthermore, there is evidence that during the juvenilephase, social experiences, such as those provided by play fighting, are criticalfor rats to learn how to modify their responses so as to take into account themovements of conspecifics (Pellis et al., 1999), and that such learning is de-pendent on cortical mechanisms (Pellis et al., 1992). As can be seen, there are,in rats, nonlearned, maturational changes that occur in the content of playfighting during development. Further, there appear to be neural mechanismsthat permit learning to occur from the experiences arising from these changesin play. These changes may be present in a variety of species, including hu-mans. Several lines of evidence support this suspicion.
Play Fighting 57

CONCLUSION
The sequential use of attack and defense during play fighting ensures that the50:50 rule is followed. Having the ability to modify the sequential pattern ofuse of these tactics to exaggerate the win:loss ratio either in the partner’s favoror in the performer’s favor provides a means for using play fighting as a toolfor social assessment and manipulation. Social assessment and manipulationcan occur in two seemingly opposite functional contexts: Play fighting can beused to maintain the status quo or to overturn the status quo. To maintain un-changing relationships, repeated play fights can reveal that no change in thewin:loss ratio has occurred. That is, the relative advantage in play has re-mained unchanged. Such use of play fighting for the reinforcement of existingrelationships has been described in a variety of species, including rats (Pellis,2002a) and New World deer (Geist, 1981), and among familiar boys (Pellegrini,1994, 1995). To change social relationships, repeated play fights can be usedto alter the win:loss ratio. That is, the relative advantage to one of the partici-pants is changed; this permits the one that is doing the testing to ascertainwhether the partner is able to tolerate such a change; this may then providethe testing partner with an opportunity to reverse the current relationship.This use of play fighting, for testing and reversing existing relationships, hasalso been described in a variety of species, including rats (Pellis, 2002a), OldWorld deer (Geist, 1982), adolescent chimpanzees (Paquette, 1994), and boysin unstable social relationships (Pellegrini, 1994, 1995).
The social–cognitive skills that enable play fighting to be used to assessand manipulate conspecifics are the ones that are probably of general use insocial interactions. Most critically, these skills are the ones that enable animalsto make subtle decisions about the correct course of action to take—an actionthat carefully straddles competition and cooperation so as to maximize the ad-vantage to the performer. We have suggested here that the experiences gainedby juveniles when play fighting can enhance the development of these skills insome species, such as rats. If, in this regard, humans are found to be similar torats—and there is some evidence that they are (Pellegrini & Smith, 1998)—then it may be necessary to reconsider policies that prevent children, especiallyboys, from engaging in playful contests.
NOTES
1. The work described here is mostly based on domesticated Norway rats (Rattus norvegicus).Even though domestication can greatly modify developmental processes and behavior (Cop-pinger & Coppinger, 2001), the little evidence available from wild rats observed in free-livingand captive conditions suggests that the basic patterns of play described for the domestic formare also present in the wild type (Boreman & Price, 1972; Robitaille & Bovet, 1976; Seward,1945).
2. The origin of the 50:50 rule is beyond the scope of the present chapter, but it is likely to havearisen from the reciprocity needed for copulation to occur. That is, at its most primitive, play
58 T H E D E V E L O P M E N T O F A G G R E S S I O N

fighting may be a precocial expression of sexual behavior by juveniles (Kramer & Burghardt,1998; Pellis, 1993).
REFERENCES
Adams, N., & Boice, R. (1983). A longitudinal study of dominance in an outdoor colonyof domestic rats. Journal of Comparative Psychology, 97, 24–33.
Adams, N., & Boice, R. (1989). Development of dominance in rats in laboratory andseminatural environments. Behavioural Processes, 19, 127–142.
Aldis, O. (1975). Play fighting. New York: Academic Press.Altmann, S. A. (1962). Social behavior of anthropoid primates: Analysis of recent con-
cepts. In E. L. Bliss (Ed.), Roots of behavior (pp. 277–285). New York: Harper.Bekoff, M. (1995). Play signals as punctuation: The structure of social play in canids. Be-
haviour, 132, 419–429.Blanchard, D. C., & Blanchard, R. J. (1990). Behavioral correlates of chronic domi-
nance–subordination relationships of male rats in a semi-natural situation. Neuro-science and Biobehavioral Reviews, 14, 455–462.
Blanchard, R. J., Blanchard, D. C., Takahashi, L. K., & Kelly, M. J. (1977). Attack anddefensive behaviour in the albino rat. Animal Behaviour, 25, 622–634.
Boreman, J., & Price, E. (1972). Social dominance in wild and domestic Norway rats(Rattus norvegicus). Animal Behaviour, 20, 534–542.
Boulton, M. J. (1994). The relationship between playful and aggressive fighting in chil-dren, adolescents and adults. In J. Archer (Ed.), Male violence (pp. 23–41). London:Routledge.
Breuggeman, J. A. (1978). The function of adult play in free-ranging Macaca mulatta. InE. O. Smith (Ed.), Social play in primates (pp. 169–192). New York: AcademicPress.
Burghardt, G. M. (1998). Play. In G. Greenberg & M. M. Haraway (Eds.), Comparativepsychology: A handbook (pp. 725–735). New York: Garland.
Byrd, K. R., & Briner, W. E. (1999). Fighting, nonagonistic social behavior, and explora-tion in isolation-reared rats. Aggressive Behavior, 25, 211–223.
Calhoun, J. B. (1962). A “behavioral sink.” In E. L. Bliss (Ed.), Roots of behavior (pp.295–315). New York: Harper.
Coppinger, R., & Coppinger, L. (2001). Dogs: The startling new understanding of canineorigins, behavior and evolution. New York: Scribner.
DaSilva, N. L., Ferreira, V. M. M., Carobrez, A. D. P., & Morato, G. S. (1996). Individualhousing from rearing modifies the performance of young rats on the elevated plus-maze apparatus. Physiology and Behavior, 60, 1391–1396.
de Waal, F. B. M., & Aureli, F. (Eds.). (2000). Natural conflict resolution. Berkeley, CA:University of California Press.
Diamond, J. (1997). Guns, germs and steel: The fates of human societies (pp. 271–272).New York: Norton.
Einon, D., Humphreys, A. P., Chivers, S. M., Field, S., & Naylor, V. (1981). Isolation haspermanent effects upon the behaviour of the rat, but not mouse, gerbil, or guineapig. Developmental Psychobiology, 14, 343–355.
Einon, D., Morgan, M. J., & Kibbler, C. C. (1978). Brief periods of socialization and laterbehavior in the rat. Developmental Psychobiology, 11, 213–225.
Fagen, R. (1981). Animal play behavior. New York: Oxford University Press.
Play Fighting 59

Flannery, K., & Lore, R. (1977). Observations of the subterranean activity of domesti-cated and wild rats (Rattus norvegicus): A descriptive study. Psychological Record,2, 315–329.
Foroud, A., & Pellis, S. M. (2002). The development of “anchoring” in the play fightingof rats: Evidence for an adaptive age-reversal in the juvenile phase. InternationalJournal of Comparative Psychology, 15, 11–20.
Foroud, A., & Pellis, S. M. (2003). The development of “roughness” in the play fightingof rats: A Laban Movement Analysis perspective. Developmental Psychobiology,42, 35–43.
Foroud, A., Whishaw, I. Q., & Pellis, S. M. (2004). Experience and cortical control overthe pubertal transition to rougher play fighting in rats. Behavioural Brain Research,149, 69–76.
Geist, V. (1971). Mountain sheep. Chicago: University of Chicago Press.Geist, V. (1981). Behavior: Adaptive strategies in mule deer. In O. C. Wallmo (Ed.), Mule
and black-tailed deer of North America (pp. 157–223). Lincoln: University of Ne-braska Press.
Geist, V. (1982). Adaptive behavioral strategies. In J. W. Thomas & D. E. Toweill (Eds.),Elk of North America (pp. 214–277). Harrisburg, PA: Stackpole Books.
Hol, T., van den Berg, C. L., van Ree, J. M., & Spruijt, B. M. (1999). Isolation during theplay period in infancy decreases adult social interactions in rats. Behavioural BrainResearch, 100, 91–97.
Kramer, M., & Burghardt, G. M. (1998). Precocious courtship and play in emydid tur-tles. Ethology, 104, 38–56.
Larsson, K. (1978). Experiential factors in the development of sexual behaviour. In J. B.Hutchinson (Ed.), Biological determinants of sexual behavior (pp. 55–86). NewYork: Wiley.
Meaney, M. J., & Stewart, J. (1981). A descriptive study of social development in the rat(Rattus norvegicus). Animal Behaviour, 29, 34–45.
Panksepp, J. (1981). The ontogeny of play in rats. Developmental Psychobiology, 14,327–332.
Panksepp, J., & Beatty, W. W. (1980). Social deprivation and play in rats. Behavioral andNeural Biology, 30, 197–206.
Panksepp, J., Normansell, L., Cox, J. F., & Siviy, S. M. (1994). Effects of neonataldecortication on the social play of juvenile rats. Physiology and Behavior, 56, 429–443.
Paquette, D. (1994). Fighting and play fighting in captive adolescent chimpanzees. Ag-gressive Behavior, 20, 49–65.
Parker, G. A. (1974). Assessment strategy and the evolution of fighting behavior. Journalof Theoretical Biology, 47, 223–243.
Pellegrini, A. D. (1994). The rough play of adolescent boys of differing sociometric sta-tus. International Journal of Behavioral Development, 17, 525–540.
Pellegrini, A. D. (1995). A longitudinal study of boys’ rough-and-tumble play duringearly adolescence. Journal of Applied Developmental Psychology, 16, 77–93.
Pellegrini, A. D., & Smith, P. K. (1998). Physical activity play: The nature and function ofa neglected aspect of play. Child Development, 69, 577–598.
Pellis, S. M. (1988). Agonistic versus amicable targets of attack and defense: Conse-quences for the origin, function and descriptive classification of play-fighting. Ag-gressive Behavior, 14, 85–104.
60 T H E D E V E L O P M E N T O F A G G R E S S I O N

Pellis, S. M. (1993). Sex and the evolution of play fighting: A review and a model based onthe behavior of muroid rodents. Journal of Play Theory and Research, 1, 56–77.
Pellis, S. M. (1997). Targets and tactics: The analysis of moment-to-moment decisionmaking in animal combat. Aggressive Behavior, 23, 107–129.
Pellis, S. M. (2002a). Keeping in touch: Play fighting and social knowledge. In M. Bekoff,C. Allen, & G. M. Burghardt (Eds.), The cognitive animal: Empirical and theoreti-cal perspectives on animal cognition (pp. 421–427). Cambridge, MA: MIT Press.
Pellis, S. M. (2002b). Sex differences in play fighting revisited: Traditional and nontradi-tional mechanisms of sexual differentiation in rats. Archives of Sexual Behaviour,31, 17–26.
Pellis, S. M., Field, E. F., Smith, L. K., & Pellis, V. C. (1997). Multiple differences in theplay fighting of male and female rats: Implications for the causes and functions ofplay. Neuroscience and Biobehavioral Reviews, 21, 105–120.
Pellis, S. M., Field, E. F., & Whishaw, I. Q. (1999). The development of a sex-differenti-ated defensive motor-pattern in rats: A possible role for juvenile experience. Devel-opmental Psychobiology, 35, 156–164.
Pellis, S. M., & Iwaniuk, A. N. (1999a). The roles of phylogeny and sociality in the evolu-tion of social play in muroid rodents. Animal Behaviour, 58, 361–373.
Pellis, S. M., & Iwaniuk, A. N. (1999b). The role of adult play: A comparative analysis ofplay and courtship in primates. Ethology, 105, 783–806.
Pellis, S. M., & Iwaniuk, A. N. (2000). Adult–adult play in primates: Comparative analy-ses of its origin, distribution and evolution. Ethology, 106, 1083–1104.
Pellis, S. M., & McKenna, M. M. (1992). Intrinsic and extrinsic influences on play fight-ing in rats: Effects of dominance, partner’s playfulness, temperament and neonatalexposure to testosterone propionate. Behavioural Brain Research, 50, 135–145.
Pellis, S. M., & Pellis, V. C. (1987). Play fighting differs from serious fighting in both tar-get of attack and tactics of fighting in the laboratory rat Rattus norvegicus. Aggres-sive Behavior, 13, 227–242.
Pellis, S. M., & Pellis, V. C. (1990). Differential rates of attack, defense and counterattackduring the developmental decrease in play fighting by male and female rats. Devel-opmental Psychobiology, 23, 215–231.
Pellis, S. M., & Pellis, V. C. (1991a). Role reversal changes during the ontogeny of playfighting in male rats: Attack versus defense. Aggressive Behavior, 17, 179–189.
Pellis, S. M., & Pellis, V. C. (1991b). Attack and defense during play fighting appear to bemotivationally independent behaviors in muroid rodents. Psychological Record,41, 175–184.
Pellis, S. M., & Pellis, V. C. (1992). Juvenilized play fighting in subordinate male rats. Ag-gressive Behavior, 18, 449–457.
Pellis, S. M., & Pellis, V. C. (1996). On knowing it’s only play: The role of play signals inplay fighting. Aggression and Violent Behavior, 1, 249–268.
Pellis, S. M., & Pellis, V. C. (1997). The pre-juvenile onset of play fighting in rats (Rattusnorvegicus). Developmental Psychobiology, 31, 193–205.
Pellis, S. M., & Pellis, V. C. (1998a). The play fighting of rats in comparative perspective:A schema for neurobehavioral analyses. Neuroscience and Biobehavioral Reviews,23, 87–101.
Pellis, S. M., & Pellis, V. C. (1998b). The structure-function interface in the analysis ofplay fighting. In M. Bekoff & J. A. Byers (Eds.), Animal play: Evolutionary, compar-ative and ecological aspects (pp. 115–140). Cambridge, UK: Cambridge UniversityPress.
Play Fighting 61

Pellis, S. M., Pellis, V. C., & McKenna, M. M. (1993). Some subordinates are more equalthan others: Play fighting amongst adult subordinate male rats. Aggressive Behav-ior, 19, 385–393.
Pellis, S. M., Pellis, V. C., & Whishaw, I. Q. (1992). The role of the cortex in play fightingby rats: Developmental and evolutionary implications. Brain, Behavior and Evolu-tion, 39, 270–284.
Potegal, M., & Einon, D. (1989). Aggressive behavior in adult rats deprived of play-fighting experience as juveniles. Developmental Psychobiology, 22, 159–172.
Robitaille, J. A., & Bovet, J. (1976). Field observations on the social behaviour of theNorway rat, Rattus norvegicus (Berkenhout). Biology of Behavior, 1, 289–308.
Seward, J. P. (1945). Aggressive behavior in the rat: I. General characteristics: Age andsex differences. Journal of Comparative Psychology, 38, 175–197.
Siviy, S. M., & Panksepp, J. (1987). Sensory modulation of juvenile play in rats. Develop-mental Psychobiology, 20, 39–55.
Smith, L. K., Fantella, S.-L., & Pellis, S. M. (1999). Playful defensive responses in adult maleratsdependon the statusof theunfamiliaropponent.AggressiveBehavior,25, 141–152.
Smith, L. K., Field, E. F., Forgie, M. L., & Pellis, S. M. (1996). Dominance and age-relatedchanges in the play fighting of intact and post-weaning castrated male rats (Rattusnorvegicus). Aggressive Behavior, 22, 215–226.
Smith, L. K., Forgie, M. L., & Pellis, S. M. (1998). Mechanisms underlying the absence ofthe pubertal shift in the playful defense of female rats. Developmental Psycho-biology, 33, 147–156.
Suomi, S. J. (2002). How gene–environment interactions can shape the development ofsocioemotional regulation in rhesus monkeys. In B. S. Zuckerman, A. F. Zuckerman,& N. A. Fox (Eds.), Emotional regulation and developmental health: Infancy andearly childhood (pp. 5–26). New Brunswick, NJ: Johnson & Johnson Pediatric In-stitute, L.L.C.
Thierry, B. (2000). Covariation of conflict management patterns across macaque species.In F. Aureli & F. B. M. De Waal (Eds.), Natural conflict resolution (pp. 106–128).Berkeley, CA: University of California Press.
Thor, D. H., & Holloway, W. R., Jr. (1984). Social play in juvenile rats: A decade of meth-odological and experimental research. Neuroscience and Biobehavioral Reviews, 8,455–464.
van den Berg, C. L., Hol, T., van Ree, J. M., Spruijt, B. M., Everts, H., & Koolhaas, J. M.(1999a). Play is indispensible for an adequate development of coping with socialchallenges in the rat. Developmental Psychobiology, 34, 129–138.
van den Berg, C. L., van Ree, J. M., & Spruijt, B. M. (1999b). Sequential analysis of juve-nile isolation-induced decreased social behavior in the adult rat. Physiology and Be-havior, 67, 483–488.
van Eden, C. J., Kros, J. M., & Ulings, H. B. M. (1990). The development of the ratprefrontal cortex. In H. B. M. Ulings, C. G. van Eden, M. A. Corner, & M. G. P.Feenstra (Eds.), Progress in brain research: Vol. 85. The prefrontal cortex. Its struc-ture, function and pathology (pp. 169–183). Amsterdam: Elsevier.
Varlinskaya, E. I., Spear, L. P., & Spear, N. E. (1999). Social behavior and social motiva-tion in adolescent rats: Role of housing conditions and partner’s activity. Physiologyand Behavior, 67, 475–482.
Von Frijtag, J. C., Schot, M., van des Bos, R., & Spruijt, B. M. (2002). Individual housingduring the play period results in changed responses to and consequences of apsychosocial stress situation in rats. Developmental Psychobiology, 41, 58–69.
62 T H E D E V E L O P M E N T O F A G G R E S S I O N

THE DEVELOPMENT OF AGGRESSIONGenetic and Environmental Factors in Rhesus Monkeys
4
Genetic and Environmental FactorsInfluencing the Expression of Impulsive
Aggression and SerotonergicFunctioning in Rhesus Monkeys
STEPHEN J. SUOMI
This chapter is dedicated to the memory of Markku Linnoila, who passedaway 5 years ago following a sudden recurrence of cancer. At the time of hisdeath at age 50, Dr. Linnoila was the Scientific Director of the National Insti-tute on Alcohol Abuse and Alcoholism (NIAAA) and was clearly expandingan already extraordinary research career. I first met him more than 20 yearsago; at that time he was a recently appointed laboratory chief in the NIAAAand I was being recruited from the University of Wisconsin to develop a newlaboratory in a different institute of the National Institutes of Health (NIH),the National Institute of Child Health and Human Development (NICHD).Within minutes of being introduced to each other, we decided to form a long-term collaboration, wherein his laboratory would analyze cerebrospinal fluid(CSF) concentrations of monoamine metabolites collected from rhesus mon-keys that would be reared in different physical and social environments at thenew primate facility that was being developed in the Maryland countryside.Moreover, he proposed to provide postdoctoral support for my senior gradu-ate student from Wisconsin, J. D. Higley, in order to maintain our collabora-tion. At that time I had no idea how unusual this collaboration was, given thetradition of intense competition for resources between the different instituteswithin the NIH. Higley is now a tenured scientist in the NIAAA, our collabo-rative efforts actually have expanded over the years, and although Markku isno longer with us, his remarkable vision and his spirit persist in full force.
63

This chapter summarizes a program of research inspired by Linnoila thathas focused on the development of excessive physical aggression in a subgroupof young male rhesus monkeys, with an emphasis on findings linking suchpatterns of behavior to an apparent deficit in central serotonin metabolism. Ibegin by describing the social organization of rhesus monkeys in natural set-tings and how aggression typically emerges and is subsequently socialized dur-ing the juvenile years. Next, I summarize evidence that demonstrates a strikingassociation between excessive physical aggression and deficits in central sero-tonin metabolism for rhesus monkeys, as has been found in humans and otherprimate species. I then characterize the biobehavioral developmental trajecto-ries of individuals prone to engage in inappropriate and excessive aggressionand present evidence regarding the relative heritability of these phenomena.Next, studies of the long-term consequences of excessive aggression for mon-keys growing up in free-ranging environments are examined, followed by asummary of the ubiquitous effects of differential early social rearing experi-ences. Finally, I present some recent findings demonstrating that allelic varia-tion in a specific gene implicated in serotonin metabolism can have dramati-cally different developmental consequences for monkeys as a function of theirearly social rearing history.
SOCIAL ORGANIZATION, SOCIAL DEVELOPMENT,AND THE SOCIALIZATION OF AGGRESSION IN RHESUS MONKEYS
The primate facility that was built by NICHD in the Maryland countrysidenear the town of Poolesville has enabled us to study rhesus monkeys growingup under conditions that simulate most basic aspects of their natural physicaland social environments. In nature, rhesus monkeys reside in large socialgroups (troops) that can range in size from several dozen to several hundredindividuals. Each troop is composed of several different female-headed fami-lies (matrilines) that each span several generations of kin, plus numerous im-migrant males. This pattern of social organization derives from the fact thatrhesus monkey females stay in the troop in which they were born for their en-tire lives, whereas virtually all rhesus monkey males emigrate from their nataltroop around the time of puberty, usually in their 4th or 5th year of life, andmost eventually join other troops.
Rhesus monkey troops in nature are also characterized by multiple socialdominance relationships, including distinctive hierarchies both between andwithin families, as well as a hierarchy among the immigrant adult males(Lindburg, 1971). Although male status superficially appears to be a functionof relative tenure—that is, the longer a male has been living in the troop, themore likely he is to be high in rank—in point of fact male dominance statusseems to be more a function of social skill in joining and maintaining coali-tions, not only with other males but especially with high-ranking females inthe troop (Berard,1989). In sum, the dominance status of a rhesus monkey de-
64 T H E D E V E L O P M E N T O F A G G R E S S I O N

pends not so much on how big and strong it is, but rather who its family andfriends are—and the latter is clearly dependent on the development of com-plex social skills during ontogeny.
Rhesus monkey infants spend virtually all of their initial days and weeksof life in physical contact with their biological mother, during which time theyform a strong and enduring specific attachment bond with her. In their 2ndmonth of life, infant monkeys begin to explore their immediate physical andsocial environment, using their mother as a secure base to support such explo-ration (cf. Suomi, 1995). Over the next few months these infants spend in-creasing amounts of time engaging in extensive social interactions with othergroup members, especially peers. Social play with peers soon becomes theirpredominant social activity and remains so until puberty. During this timeplay interactions become increasingly gender-segregated and involve patternsof behavior that appear to simulate virtually all adult social activities, includ-ing various forms of aggression (Ruppenthal, Harlow, Eisele, Harlow, &Suomi, 1974; Suomi & Harlow, 1975).
Aggression is a normal and necessary part of every rhesus monkey’s over-all behavioral repertoire—indeed, most primatologists consider rhesus mon-keys to be among the most aggressive of all primate species. Rhesus monkeyaggression can range in intensity from mere facial threats and vocalizations tovigorous chases and actual physical attacks, including slapping, hitting, hairpulling, and biting with sufficient intensity to produce lasting tissue damage oreven death. The capability for aggression is crucial for survival in the wild, notonly from the standpoint of defending one’s self and offspring from predatorsand nonspecific competitors, but also in maintaining social order and enforc-ing the complex dominance hierarchies characteristic of all rhesus monkeytroops. However, uncontrolled, unpredictable, and violent aggression withinany troop could drive members apart and destroy it as a social unit. There-fore, aggression must be socialized—it must be minimized or at least largelyritualized in intragroup interactions, but it must also remain a viable responsein order to counter external threats or other dangers.
Socialization of aggression for young rhesus monkeys involves not onlylearning in which circumstances and toward what targets aggressive behaviormight be appropriate, but also gauging the relative intensity of the attack orresponse called for and the appropriate time and means for terminating an ag-gressive bout or for avoiding it altogether. Indeed, learning whom not to at-tack, as well as how to moderate aggressive impulses, is as important in thesocialization process as is honing one’s fighting skills.
Aggression typically emerges in a rhesus monkey’s behavioral repertoireat about 6 months of age, and it initially appears in the context of rough-and-tumble play (Symonds, 1978). Biting, hair pulling, wrestling, and other formsof physical contact are basic components of rough-and-tumble play directedtoward peers, which occurs with increasing frequency among males in the sec-ond half of their first year of life and becomes the predominate type of playfor the rest of their juvenile years (Harlow & Lauersdorf, 1974). Although
Genetic and Environmental Factors in Rhesus Monkeys 65

some form of virtually all behavioral components of adult aggressive ex-changes can be seen in the rough-and-tumble play bouts of young males, theintensity of such interactions is usually quite controlled and seldom escalatesto the point of potential physical injury—if it does, the play bout is almost al-ways terminated immediately, either via adult intervention or by one or moreof the participants backing away themselves. The importance of these playbouts with peers for the socialization of aggression becomes apparent whenone considers that rhesus monkey infants reared in laboratory environmentsthat denied them regular access to peers during their initial months consis-tently exhibited excessive and socially inappropriate aggression later in life(e.g., Alexander & Harlow, 1966; Harlow & Harlow, 1969).
INDIVIDUAL DIFFERENCES IN AGGRESSION AND SEROTONIN METABOLISM
Both laboratory and field studies of rhesus monkeys and other nonhuman pri-mate species have reported striking individual differences in the levels andtypes of aggression exhibited by monkeys of like age and gender, especiallywhen displayed in circumstances where the aggression is neither provoked norsocially appropriate (e.g., Bernstein, Williams, & Ramsay, 1983; Steklis,Brammen, Raleigh, & McGuire, 1985), much as has been the case in recenthuman surveys and clinical investigations (cf. Lahey, Moffitt, & Caspi, 2003).Over the past two decades numerous studies, including several by Linnoila(e.g., Linnoila, 1988; Linnoila, DeJong, & Virkkunen, 1989; Linnoila et al.,1983), of child, adolescent, and adult males who engaged in unusually and vi-olently aggressive activities have reported links between such aggression andapparent deficits in serotonergic functioning, such that those aggressive indi-viduals tended to have unusually low CSF concentrations of the primary cen-tral serotonin metabolite 5-hydroxyindoleacetic acid (5-HIAA). Given thesefindings from the human clinical literature and with the strong encouragementof Linnoila, my colleagues and I decided to test the so-called serotonin hy-pothesis (cf. Coccaro & Murphy, 1990) in the rhesus monkey colony. Accord-ingly, we collected and assayed CSF samples for concentrations of 5-HIAAfrom a cross-sectional sample of infant, juvenile, and young adult monkeysrepresenting both genders and three different early-rearing backgrounds.
We also had considerable background data regarding levels of aggressionexhibited by monkeys of comparable age, gender, and rearing history, al-though nobody had previously put those two data sets together. When wethen did so, what seemed striking was the degree to which differences in CSF5-HIAA concentrations were inversely related to differences in the incidenceof aggression, whether the comparison involved age, gender, or rearing condi-tion differences. For example, infant monkeys display minimal levels of ag-gression, juvenile monkeys show some, typically in the context of play, andadolescents and young adults, especially males, tend to exhibit the highest lev-els of aggressive behavior for any age-gender subgroup. We found exactly the
66 T H E D E V E L O P M E N T O F A G G R E S S I O N

opposite to be true for 5-HIAA concentrations: Infants had the highest levels,juveniles’ concentrations were significantly lower, and postpubertal monkeys(especially males) had the lowest concentrations (Higley, Suomi, & Linnoila,1990b). Equally striking was the finding that short-term longitudinal CSF 5-HIAA data obtained from a subset of these monkeys, collected under vastlydifferent environmental circumstances, revealed remarkable stability acrossboth situations and individuals, suggesting a trait-like characteristic for theCSF 5-HIAA measure (Higley, Suomi, & Linnoila, 1990a).
Given the findings, we began a concentrated effort to explore thisapparently inverse relationship more fully in our Poolesville rhesus monkeypopulation. A series of prospective longitudinal studies replicated and ex-tended our basic findings from the initial cross-sectional analyses. We foundthat approximately 5–10% of our monkeys exhibited impulsive behavioraltendencies and general social incompetence. Males in this subgroup becameinvolved in a disproportionate number of aggressive exchanges with othergroup members. Females in this subgroup typically engaged in fewer overtacts of physical aggression than those males, but more than most other fe-males. They also were less likely to engage in positive affiliative behaviorssuch as social grooming. Monkeys in this subgroup also consistently hadlower CSF 5-HIAA concentrations than their peers of like age and gender(Higley & Suomi, 1996).
DEVELOPMENTAL TRAJECTORIES FOR IMPULSIVELY AGGRESSIVE MONKEYS
We now know that rhesus monkeys with chronically low CFS 5-HIAA con-centrations typically begin to distinguish themselves from their peers in theirearly play interactions. This is especially true for low–5-HIAA males: Theyseem to lack the ability (or perhaps the motivation) to moderate their behav-ioral responses to playful initiations from peers, and by late childhood theirrough-and-tumble play bouts often escalate into more serious aggressive ex-changes. Not surprisingly, most of these males soon come to be avoided bythose around them, and as a result they become increasingly isolated socially(Figure 4.1).
Moreover, low–5-HIAA juvenile males often seem unable (or unwilling)to follow social rules inherent in rhesus monkey dominance hierarchies(Higley, Suomi, & Linnoila, 1996d). For example, they may directly challengea dominant adult male, a foolhardy act that often leads to serious injury, espe-cially when they seemingly refuse to back down or exhibit appropriate sub-missive behavior once defeat becomes obvious. Juvenile females with chroni-cally low CSF 5-HIAA concentrations tend to exhibit low levels of socialgrooming, and although they are generally involved in fewer overtly aggres-sive acts than their male counterparts, they too tend to become relatively iso-lated socially as they are growing up, typically drifting to the bottom of thedominance hierarchy within their social group (Higley et al., 1996a).
Genetic and Environmental Factors in Rhesus Monkeys 67

Longitudinal studies (e.g., Higley, Suomi, & Linnoila, 1992) have demon-strated that individual differences in CSF 5-HIAA concentrations tend to beremarkably stable from infancy to early adulthood, despite steady and signifi-cant developmental decreases in overall CSF 5-HIAA concentrations through-out childhood and adolescence. Given this stability and the inverse relation-ship between CSF 5-HIAA concentrations and impulsive aggressiveness (cf.Higley et al., 1996d), it appears possible to predict an individual’s likelihoodof becoming impulsively aggressive later in life from its CSF 5-HIAA valuesexhibited during infancy. Moreover, as previously noted, individual differ-ences in CSF 5-HIAA concentrations remain relatively stable across a varietyof both stressful and benign environmental circumstances, unlike most indicesof central nervous system activity. These findings therefore suggest that theCSF concentration of 5-HIAA can serve as a marker of an individual’s propen-sity to engage in impulsively aggressive actions, even when assessed under cir-cumstances in which no overt aggression is being displayed (Higley, Suomi, &Linnoila, 1991b).
Recent research has shown that individual differences in impulsiveaggressiveness and CSF 5-HIAA concentrations are also predictive of individ-ual differences in the propensity to consume alcohol in a “happy hour situa-tion.” Over the past decade Higley and his colleagues have developed an ex-perimental paradigm in which group-living rhesus monkeys are given theopportunity to consume an aspartame-flavored 7% alcohol (ETOH) bever-age, a nonalcoholic aspartame-flavored beverage, and/or plain tap water for
68 T H E D E V E L O P M E N T O F A G G R E S S I O N
FIGURE 4.1. Aggressive male rhesus monkey.

daily 1-hour periods within their familiar social group (e.g., Higley et al.,1991a). Impulsive adolescent monkeys who have low CSF 5-HIAA concen-trations tend to consume excessive amounts of alcohol when placed in thishappy hour experimental paradigm (Higley et al., 1996c). Researchers havealso demonstrated a significant relationship between degree of alcohol intoxi-cation and serotonin transporter availability in these monkeys (Heinz et al.,1998).
GENETIC FACTORS AND STRAIN DIFFERENCES
Several studies involving rhesus monkeys whose genetic backgrounds havebeen well characterized and who have been raised from birth in controlledlaboratory environments have demonstrated significant heritability for indi-vidual differences in CSF 5-HIAA concentrations. One investigation com-pared paternal half-siblings (same father, different mothers, with no physicalexposure to their biological father at any time) and unrelated individualsreared under identical conditions. Heritability analyses revealed significantlygreater concordance of CSF 5-HIAA concentrations among same-gender pa-ternal half-siblings than among unrelated same-sex control subjects matchedfor age and rearing history. Another study utilized cross-fostering procedures,in which infants were separated from their biological mothers within the firstweek of life and raised thereafter by an unrelated multiparous female. CSFconcentrations for the cross-fostered infants, assessed at 6 and 18 months ofage, more closely resembled the CSF 5-HIAA concentrations of their biologi-cal mothers than those of their foster mothers (Higley et al., 1993). Thesefindings provide compelling (albeit indirect) evidence of heritable influenceson central serotonin metabolism, as well as risk for the development of impul-sive aggressiveness, in laboratory-reared rhesus monkeys.
A completely different line of research linking low serotonin metabolismwith impulsive behavioral tendencies originated from serendipitous observa-tions by the personnel caring for and testing rhesus monkey infants in thePoolesville facility’s neonatal nursery. Our colony has had a self-sufficientbreeding program since its inception; that is, we have not had to bring in anyrhesus monkeys from outside sources in order to carry out our basic researchprogram. All of our breeders were either born in the laboratory or were partof the original colony that had been moved to the Poolesville facility from theUniversity of Wisconsin Harlow Primate Laboratory in the mid-1980s; all ofthe Wisconsin monkeys, in turn, were descendents of wild rhesus monkeys im-ported from India prior to 1971. In contrast, some other primate facilities atthe NIH have imported rhesus monkeys from China over the past two de-cades. Anecdotal reports by researchers and animal caretaking staff alike havesuggested that rhesus monkeys of Chinese origin were unusually aggressive,difficult to train, and otherwise generally intractable.
One NIH laboratory using rhesus monkeys imported from China was
Genetic and Environmental Factors in Rhesus Monkeys 69

carrying out contraception research; because that laboratory lacked nurseryfacilities, unwanted infants that were the product of contraceptive failureswere sent to us to be raised in our neonatal nursery. Maribeth Champoux,who oversees the nursery research and caretaking staff, soon noticed thatthose infants with parents of Chinese origin were unusual in several respects,not the least of which was their tendency to bite nursery staff while beinghand-fed during their first week of life, a phenomenon we had never previ-ously experienced with our own infants. Champoux subsequently tested otherChinese-origin infants with the lab’s standardized neonatal test battery andfound significant deficits in their activity state control and their visual orient-ing capabilities during their first month of life, relative to the normative stan-dards established for infants whose parents came from the lab’s breeding col-ony (Champoux, Suomi, & Schneider, 1994).
CSF samples were obtained from Chinese-derived infants throughouttheir first 6 months of life, and their CSF 5-HIAA concentrations were com-pared with those of the lab’s Indian-derived infants. The Chinese-derivedinfants were found to have significantly lower CSF 5-HIAA concentrationsfrom 3 to 4 months onward (Champoux, Higley & Suomi, 1997). Moreover,behavioral observations of these youngsters of Chinese origin revealed higherfrequencies of impulsive and aggressive behaviors during their first 2 years oflife than those observed in like-reared peers born into the lab colony (follow-up observations are currently under way). In addition to providing an empiri-cal basis for insights based on anecdotal observations, these studies comparingIndian- and Chinese-derived infants established a predictive relationship be-tween measures of basic biobehavioral characteristics during the neonatal pe-riod and patterns of serotonergic function and behavioral tendencies later inlife.
FIELD STUDIES
Given that low CSF 5-HIAA is associated with a host of behavioral tendenciesthroughout development that seem, at best, problematic for rhesus monkeysmaintained in captive environments, a reasonable question is whether similarphenomena can be found in wild populations of rhesus monkeys—and if so,what the adaptive consequences for such individuals might be. ThroughLinnoila’s good offices, we established a collaboration with David Taub andPatrick Mehlman, and more recently continued with Greg Westergaard, thatgave us access to a free-ranging population of approximately 4,000 rhesusmonkeys that have been living for many years on Morgan Island, a sea islandon the South Carolina Atlantic coast. These monkeys are provisioned but re-ceive no special veterinary treatment, and except for periodic trapping of spec-ified individuals for biological sampling, these monkeys are not subject to anyexperimental interventions.
Analyses of CSF samples obtained from free-ranging juvenile, adolescent,
70 T H E D E V E L O P M E N T O F A G G R E S S I O N

and adult male rhesus monkeys residing on this sea island found 5-HIAAconcentrations closely resembling those obtained from males in the Poolesvillecolony in terms of absolute values, age-related differences, and stability of in-dividual differences across repeated sampling and across major developmentaltransitions (e.g., natal troop emigration). Moreover, the strong inverse rela-tionship between CSF 5-HIAA concentrations and the rate of escalating ag-gression consistently reported in laboratory studies was at least as strong inthis field population of rhesus monkey males and, similarly, first appeared inthe context of play with peers. Like their captive counterparts, these wild rhe-sus monkey juveniles with low CSF 5-HIAA concentrations frequently turnedroutine rough-and-tumble play bouts into episodes involving serious aggres-sive exchanges, and they were also more likely to commit obvious social blun-ders that typically elicit physical attacks from larger and more socially domi-nant troop members (Higley et al., 1992a). Juvenile males with low CSF 5-HIAA concentrations were also more likely to take long, potentially danger-ous leaps from treetop to treetop, sometimes falling to the ground and injur-ing themselves. We found a significant inverse linear relationship between therelative frequency of such leaps and CSF 5-HIAA concentrations that paral-leled the negative relationship between CSF 5-HIAA values and the incidenceof escalating aggression (Mehlman et al., 1994; Figure 4.2).
The long-term outlook for young rhesus monkey males with low CSF 5-HIAA concentrations is not very promising in free-ranging environments suchas Morgan Island. Ostracized by their peers and frequently attacked by adultsof both genders, most of these young males are physically driven out of their
Genetic and Environmental Factors in Rhesus Monkeys 71
FIGURE 4.2. Impulsive juvenile male monkey taking dangerous leap.

natal troop prior to 3 years of age, long before the onset of puberty (Mehlmanet al., 1995). These impulsive young males generally lack the social skills to beable to join another troop; indeed, they even seem unable to join an all-malegang. As a result, most of these males become solitary, and virtually all ofthem perish within a year, long before becoming physically capable of repro-duction (Higley et al., 1996b).
Given the previously noted finding that individual differences in CSF 5-HIAA concentrations appear to be highly heritable, along with the fact thatmost male rhesus monkeys with unusually low CSF 5-HIAA concentrationsfail to survive to reproductive age, one might reasonably ask how it could bepossible for as many as 10% of rhesus monkeys in wild populations to havechronically low CSF 5-HIAA concentrations. Three possible explanationsreadily come to mind. One possibility is that the few males with low CSF 5-HIAA that do survive to adulthood sire a disproportionately large number ofoffspring (the “silver bullet” hypothesis). Recent observational data obtainedfrom the Morgan Island population suggest that this hypothesis is exceed-ingly unlikely. In point of fact, the low CSF 5-HIAA males who do survivetend to be actively avoided by most females. In the few circumstances in whichconsort behavior with a female actually occurs, those males tend to engage infewer mounts per copulatory sequence (rhesus monkeys are serial mounters),are less likely to ejaculate before the sequence is terminated, and have a lowerprobability of inseminating the females when they do ejaculate than do othermales in the population (Mehlman et al., 1997).
A more plausible possibility is that the propensity for developing chroni-cally low central serotonin metabolism is passed on to the next generation ofrhesus monkeys primarily through the female, rather than the male, genome.Young female rhesus monkeys with low–5-HIAA concentrations are indeedimpulsive and aggressive and tend to be generally incompetent socially(Westergaard et al., 2003). Unlike males, however, they are not expelled fromtheir natal troop but instead remain in their respective families for the rest oftheir lives. Moreover, they engage in relatively normal reproductive behavior,and they clearly produce and rear offspring—offspring that therefore maywell be at increased genetic risk for low CSF 5-HIAA concentrations.
EFFECTS OF DIFFERENTIAL EARLY SOCIAL EXPERIENCE
A third possible mechanism for cross-generational transmission of deficits inserotonergic functioning is derived from recent studies of maternal behavior incaptive rhesus monkey groups. We have found that females with chronicallylow CSF 5-HIAA concentrations tend to exhibit significant abnormalities intheir maternal behavior, often resulting in the development of insecure and/ordisorganized attachment relationships with their infants. Other recent datasuggest that infants who develop the least secure attachment relationshipswith their mothers are also the most likely to develop deficits in their central
72 T H E D E V E L O P M E N T O F A G G R E S S I O N

serotonin metabolism (cf. Suomi, 1999). Thus, an alternative possible meansof cross-generational transmission of the propensity for having chronicallylow CSF 5-HIAA concentrations may be via aberrant maternal care, or otherforms of social stimulation, that offspring might experience during develop-ment. According to this view, the mother’s behavior toward offspring may bemore relevant for cross-generational transmission of the phenomenon thanher genes. Of course, these two potential pathways are not necessarily mutu-ally exclusive (Suomi & Levine, 1998).
In fact, an increasing body of data from laboratory studies of monkeysreared under different social circumstances has demonstrated that early socialexperiences can profoundly affect both the development of impulsive aggres-sive tendencies and central serotonin metabolism alike. Some of the most com-pelling findings have come from longitudinal studies of peer-reared monkeys,that is, infants that were permanently separated from their mothers at birth,hand-reared in the neonatal nursery for their first month of life, and thenplaced in small social groups with like-reared age mates (cf. Harlow &Harlow, 1969). In most of the relevant studies the peer-reared subjects weremoved into larger social groups containing both peer-reared and mother-reared age mates at 7 months of age, and they remained in these larger socialgroups at least until puberty, if not beyond (e.g., Higley & Suomi, 1996). Dur-ing their initial months these infants readily developed strong social attach-ment bonds to each other, much as mother-reared infants develop attachmentrelationships with their own mothers. However, because peers are not nearlyas effective as a normal monkey mother in reducing fear in the face of stress orin providing a secure base for exploration, the attachment relationships thatthese peer-reared infants developed were almost always “anxious” or “inse-cure” in nature (Suomi, 1995). As a consequence, whereas the peer-rearedmonkeys exhibited relatively normal physical and motor development, theirearly exploratory behavior was somewhat limited. They seemed reluctant toapproach novel objects, and they tended to be shy in their initial encounterswith unfamiliar peers.
Moreover, even when peer-reared infants interacted with their same-agecage mates in familiar settings, their emerging social play repertoires were gen-erally retarded in both frequency and complexity. For example, peer-rearedmonkeys were more likely to play with only one partner at a time rather thanwith multiple partners simultaneously, as mother-reared youngsters quicklycome to prefer, and their play bouts were usually limited to relatively brief ex-changes rather than the extended interactions that may go on for several min-utes at a time among mother-reared peers. One explanation for the relativelypoor play performance of peer-reared monkeys is that their cage mates had toserve both as attachment objects and as playmates, a dual role that neithermothers nor mother-reared peers have to fulfill. Perhaps as a result, they typi-cally dropped to the bottom of their respective dominance hierarchies whenthey were eventually grouped with mother-reared monkeys their own age(Higley et al., 1996d).
Genetic and Environmental Factors in Rhesus Monkeys 73

Early peer-rearing also tended to make young rhesus monkey males moreimpulsive. Like the previously described impulsive monkeys growing up in thewild, peer-reared males initially exhibited aggressive tendencies in the contextof juvenile play; and as they approached puberty, the frequency and severity oftheir aggressive episodes exceeded that of mother-reared group members ofsimilar age. Peer-reared females tended to groom (and be groomed by) othersin their social group less frequently and for shorter durations than theirmother-reared counterparts and, as before, they usually remained at the bot-tom of their respective dominance hierarchies. These differences between peer-reared and mother-reared age mates in aggression, grooming, and dominancecontinued to be relatively robust when the monkeys were subsequently movedinto new social groups, and they generally remained quite stable throughoutthe juvenile and adolescent years.
An important finding was that early peer-rearing was also associated withdiminished central serotonin metabolism throughout development. Significantgroup differences between peer-reared infants and their mother-reared coun-terparts appeared within the first month of life, and these differences weremaintained throughout the rest of the 7-month period of differential rearing,even though CSF 5-HIAA concentrations dropped dramatically for both rear-ing groups during this period. Moreover, peer-reared subjects continued tohave significantly lower CSF 5-HIAA concentrations than their mother-reared counterparts after they were moved into the same large social groups,and these differences persisted through puberty. Finally, peer-reared adolescentmonkeys consistently consumed larger amounts of alcohol under comparableconditions than their mother-reared age mates (Higley, Hassert, Suomi, &Linnoila, 1991a). Follow-up studies also demonstrated that the peer-rearedsubjects quickly develop a greater tolerance for alcohol, which can be pre-dicted by their CNS serotonin turnover rates, which in turn appear to be asso-ciated with differential serotonin transporter availability (Heinz et al., 1998).
GENE–ENVIRONMENT INTERACTIONS
The previously described association between turnover rate and serotonintransporter availability led Markku Linnoila to help us establish a collabora-tion with Peter Lesch at the University of Wurzburg. Lesch and his colleaguesrecently characterized the serotonin transporter gene (5-HTT), a candidategene for impaired serotonergic function in that it mediates serotonin neuro-transmission, plays a critical role in brain serotonin homeostasis, and is a tar-get for both selective serotonin reuptake inhibiting (SSRI) antidepressant com-pounds and certain drugs of abuse (Lesch et al., 1996). They have shown thatlength variation in the 5-HTT gene-linked polymorphic region (5-HTT-LPR)results in allelic variation in 5-HTT expression. The short allele of the 5-HTT-LPR confers low transcription efficiency to the 5-HTT gene promoter, andlower 5-HTT mRNA levels are found in humans with the short allele, suggest-
74 T H E D E V E L O P M E N T O F A G G R E S S I O N

ing that the low 5-HTT expression may result in decreased serotonergic func-tion (Heils et al., 1996). Although this genetic polymorphism was first de-tected in humans, it also appears in rhesus monkeys and, in fact, is founduniquely among simian primates and humans (Lesch et al., 1997).
We recently have characterized a total of 237 rhesus monkeys in ourPoolesville colony using polymerase chain reaction (PCR)-based genotypeanalysis in order to determine allelic frequency involving the short (398bp)and long (419bp) PCR fragments in the promoter region of the 5-HTT gene.These analyses revealed that the relative frequency of the homozygous long al-lele (LL) was approximately three times that of the relative frequency of theheterozygous short allele (LS). Curiously, only two subjects in the entire sam-ple had the homozygous short allele (SS). However, among the 237 monkeysthat were genotyped, 34 were of Chinese descent or were Chinese–Indian hy-brids, and among these monkeys the relative frequency of the LS allele wasmore than twice as high (55% vs. 23%) as it was for monkeys of exclusivelyIndian origin. Moreover, consistent with our previous findings, as a group theChinese/hybrid monkeys had significantly lower concentrations of CSF 5-HIAA than their Indian-derived counterparts when age and gender factorswere statistically partialled out. The suggestion that these subgroup differ-ences in 5-HIAA concentrations, not to mention the behavioral differences de-scribed earlier, might be in part attributable to strain differences in the relativefrequency of 5-HTT genetic polymorphisms represents an attractive, albeit asyet unproven, hypothesis.
Some of the monkeys in the previously described Indian-derived sub-group had been peer-reared, whereas others had been reared by their biologi-cal mothers since birth. We found that the relative frequency of subjects pos-sessing the short 5-HTT allele did not differ significantly between the tworearing groups, which was not surprising given that these monkeys had beenmore or less randomly preassigned to their respective rearing conditions atbirth. Because we had collected CSF samples from those monkeys during their2nd and 4th years of life under comparable experimental conditions (andwhile they were all living in comparable social groups), it was possible todetermine whether their 5-HIAA concentrations differed as a function of their5-HTT polymorphic status, as might be expected from the extant literature.Interestingly, we did find such a predicted relationship, with individuals pos-sessing the LS allele having significantly lower 5-HIAA concentrations—butonly among peer-reared subjects. For mother-reared subjects, 5-HIAA concen-trations were essentially identical (and well within the normative range) formonkeys possessing either allele (Bennett et al., 2002; see Figure 4.3).
Thus, these analyses demonstrated a significant genotype–environment in-teraction, wherein the ultimate effect of a polymorphism in a specific gene fora given individual appeared to be highly dependent on the specific early expe-rience of that individual. In this case, mother-rearing appeared to buffer theinfants from potentially deleterious effects of the LS allele on serotonin metab-olism.
Genetic and Environmental Factors in Rhesus Monkeys 75

A different form of gene–environment interaction was suggested by theanalysis of alcohol consumption data: Whereas peer-reared monkeys with theLS allele consumed more alcohol than peer-reared monkeys with the LL allele,the reverse was true for mother-reared subjects, with individuals possessingthe LS allele actually consuming less alcohol than their LL counterparts (Ben-nett, Lesch, Heils, & Linnoila, 1998). A similar reversal was found for relativelevels of alcohol intoxication (Barr et al., 2004). In other words, the LS allelewas a risk factor for excessive alcohol consumption among peer-rearedmonkeys but was apparently a protective factor for mother-reared subjects.
We are currently carrying out additional analyses involving other behav-ioral and physiological measures that had already been collected on thesesame mother- and peer-reared monkeys of known 5-HTT allelic status. Forexample, Champoux et al. (2002) examined the relationship between earlyrearing history and 5-HTT allelic status on measures of neonatal neuro-behavioral development during the first month of life and found further evi-dence of maternal buffering. Specifically, infants possessing the LS allele whowere being reared in the laboratory neonatal nursery showed significant defi-cits in measures of attention, activity, and motor maturity relative to nursery-reared infants possessing the LL allele, whereas both LS and LL infants whowere being reared by competent mothers exhibited normal values for each ofthese measures. Other analyses (e.g., Barr et al., 2003) have yielded similargene–environment interactional patterns of apparent maternal buffering of ag-gressive behavior. We believe that this buffering pattern will prove to be ubiq-uitous across a wide range of measures of rhesus monkey behavioral and bio-logical functioning throughout development.
76 T H E D E V E L O P M E N T O F A G G R E S S I O N
FIGURE 4.3. CSF concentrations of 5-HIAA in peer-reared LL, peer-reared LS, mother-rearedLL, and mother-reared LS monkeys. The gene x rearing environment interaction is the resultof significantly lower CSF 5-HIAA concentrations in peer-reared LS monkeys than the otherthree subject groups. From Bennett et al. (2002).

SUMMARY AND PERSPECTIVES
This chapter has outlined a program of research investigating the developmentof excessive and socially inappropriate aggression in a subgroup of rhesusmonkeys, biological correlates, and factors that can shape such development.The chapter described how aggression is typically socialized during the infantand juvenile years and contrasted this developmental pattern with that of thesubgroup of individuals who consistently exhibit such aggression, as well asother behavioral manifestations of impulsivity and consistent deficits in ameasure of central serotonin metabolism. These biobehavioral patterns emergeearly in development, are maintained through puberty if not beyond, and are atleast in part heritable. Male monkeys that develop these patterns in nature sel-dom survive beyond puberty, whereas females developing parallel patterns ofinappropriate aggression and serotonergic dysfunctioning usually survive tobear and rear offspring—unfortunately, often in less than optimal fashion.Early rearing experiences can clearly influence the development of thesebiobehavioral patterns, even those with demonstrated heritability. Indeed, itappears that genetic and early experiential factors can and do interact toshape individual developmental trajectories. In particular, good mothering ap-pears to somehow protect offspring at genetic risk for developing problems inbehavioral and biological functioning when they are reared in less sociallysupportive settings (see Figure 4.4).
My colleagues and I believe that the implications of these most recentfindings are of considerable importance in a variety of domains. Beyond dem-onstrating that whether or not a particular genetic polymorphism has specificbiobehavioral consequences may depend on certain early experiences, thepresent findings readily suggest several potentially promising avenues forfuture studies of impulsive aggressiveness in monkeys. One approach wouldbe to become increasingly molecular in scope, focusing on demonstrations ofspecific gene expression, as well as delineating possible mechanisms by whichspecific early rearing experiences might trigger and/or otherwise influencegene expression. Indeed, this approach is currently being followed in an ele-gant program of research with rodents by Meaney and his colleagues (cf.Francis, Champagne, Liv, & Meanen, 1999).
A second approach might focus on determining the generality of this andother types of gene–environment interaction with respect to other genes asso-ciated with particular human psychopathologies, as an increasing number ofcandidate genetic polymorphisms become identified and functionally charac-terized. Recently, Caspi et al. (2002) reported a specific gene–environment in-teraction involving a polymorphism in one monoamine oxidase A (MAOA)receptor gene and adverse early social experiences across a large cohort of ad-olescents and young adults in New Zealand. Individuals who possessed oneform of the polymorphism were disproportionately likely to have been seri-ously involved in the juvenile justice system, but only if they had been abused
Genetic and Environmental Factors in Rhesus Monkeys 77

as children. We have basically replicated the overall pattern of these findingsin our Poolesville rhesus monkey colony in a very recent study focusing on afunctionally similar MAOA polymorphism (Newman et al., in press), and weare currently examining potential gene–environment interactions, as well aspossible gene–gene interactions, in several other candidate genes.
A third possible avenue for study involves identifying potentially optimalrearing environments that might serve to minimize the risk for developingpsychopathology, or actually turn that risk into a potentially adaptive ad-vantage. For example, we are currently in the middle of a cross-fostering ex-periment, in which infants selectively bred to have low versus high CSF 5-HIAA concentrations are being foster-reared by mothers who differ system-atically in their own CSF 5-HIAA concentrations. We are also studying thematernal behavior of females with chronically high versus low CSF 5-HIAA concentrations at the Morgan Island field site, with the goal ofdetermining the long-term adaptive consequence for offspring of differinggenotypes.
It is truly unfortunate that Markku Linnoila is no longer with us to helpadvance these endeavors. I think he would have been pleased with what hasbeen found to date, and I’m certain he would be eager to press onward in theeffort he urged us to begin so many years ago—using research with rhesusmonkeys to better understand the basis for the relationship between seroton-ergic functioning and the expression of impulsive aggression in humansthroughout development.
78 T H E D E V E L O P M E N T O F A G G R E S S I O N
FIGURE 4.4. Rhesus monkey mothers, each holding an infant.

REFERENCES
Alexander, B. K., & Harlow, H. F. (1966). Social behavior of juvenile rhesus monkeyssubjected to different rearing conditions during the first 6 months of life. ZoologiaJahrb Physiologia, 60, 167–174.
Barr, C. S., Newman, T. K., Becker, M. L., Parker, C. C., Champoux, M., Lesch, K. P.,Goldman, D. A., Suomi, S. J, & Higley, J. D. (2003). The utility of the non-humanprimate model for studying gene by environment interactions in behavioral re-search. Genes, Brain, and Behavior, 2, 336–340.
Barr, C. S., Newman, T. K., Lindell, S. G., Champoux, M., Lesch, K. P., Suomi, S. J.,Goldman, D. A., & Higley, J. D. (2004). Interaction between serotonin transportergene variation and rearing condition in alcohol preference and consumption in fe-male primates. Archives of General Psychiatry, 61, 1146–1152.
Bennett, A. J., Lesch, K. P., Heils, A., &., Linnoila, M. (1998). Serotonin transporter genevariation, CSF 5–HIAA concentrations, and alcohol-related aggression in rhesusmonkeys (Macaca mulatta). American Journal of Primatology, 45, 168–169.
Bennett, A. J., Lesch, K. P., Heils, A., Long, J., Lorenz, J., Shoaf, S. E., Champoux, M.,Suomi, S. J., Linnoila, M. V., & Higley, J. D. (2002). Early experience and serotonintransporter gene variation interact to influence primate CNS function. MolecularPsychiatry, 7, 118–122.
Berard, J. (1989). Male life histories. Puerto Rican Health Sciences Journal, 8, 47–58.Bernstein, I. S., Williams, L., & Ramsay, M. (1983). The expression of aggression in Old
World monkeys. International Journal of Primatology, 4, 113–125.Caspi, A., McClay, J., Moffitt, T. E., Mill, J., Marin, J., Craig, I. W., Taylor, A., &
Poulton, R. (2002). Role of genotype in the cycle of violence in maltreated children.Science, 297, 851–854.
Champoux, M., Bennett, A. J., Shannonm C., Higley, J. D., Lesch, K. P., & Suomi, S. J.(2002). Serotonin transporter gene polymorphism, differential early rearing, andbehavior in rhesus monkey neonates. Molecular Psychiatry, 7, 1158–1163.
Champoux, M., Higley, J. D., & Suomi, S. J. (1997). Behavioral and physiological char-acteristics of Indian and Chinese-Indian hybrid rhesus macaque infants. Develop-mental Psychobiology, 31, 49–63.
Champoux, M., Suomi, S. J., & Schneider, M. L. (1994). Temperamental differences be-tween captive Indian and Chinese-Indian hybrid rhesus macaque infants. Labora-tory Animal Science, 44, 351–357.
Coccaro, E. M., & Murphy, D. L. (1990) Serotonin in major psychiatric disorders. Wash-ington, DC: American Psychiatric Press.
Francis, D. D., Champagne, F. A., Liu, D., & Meaney, M. J. (1999). Maternal care, geneexpression, and the development of individual differences in stress reactivity. An-nals of the New York Academy of Science, 896, 66–84.
Harlow, H. F., & Harlow, M. K. (1969). Effects of various mother–infants relationshipson rhesus monkey behaviors. In B. M. Foss (Ed)., Determinants of infant behaviour(Vol. 4, pp. 15–36). London: Metheun.
Harlow, H. F., & Lauersdorf, H. E. (1974). Sex differences in passions and play. Perspec-tives in Biology and Medicine, 17, 348–360.
Heils, A. Teufel, A., Petri, S., Stober, G., Riederer, P. Bengel, B., & Lesch, K. P. (1996).Allelic variation of human serotonin transporter gene expression. Journal of Neuro-chemistry, 6, 2621–2624.
Heinz, A., Higley, J. D., Gorey, J. G., Saunders, R. C., Jones, D. W., Hommer, D., Zajicek,
Genetic and Environmental Factors in Rhesus Monkeys 79

K., Suomi, S. J., Weinberger, D. R., & Linnoila, M. (1998). In vivo association be-tween alcohol intoxication, aggression, and serotonin transporter availability innonhuman primates. American Journal of Psychiatry, 155, 1023–1028.
Higley, J. D., Hasert, M. L., Suomi, S. J., & Linnoila, M. (1991a). A new nonhuman pri-mate model of alcohol abuse: Effects of early experience, personality, and stress onalcohol consumption. Proceedings of the National Academy of Sciences, 88, 7261–7265.
Higley, J. D., King, S. T., Hasert, M. F., Champoux, M., Suomi, S. J., & Linnoila, M.(1996a). Stability of interindividual differences in serotonin function and its rela-tionship to severe aggression and competent social behavior in rhesus macaque fe-males. Neuropsychopharmacology, 14, 67–76.
Higley, J. D., Mehlman, P. T., Taub, D. M., Higley, S., Fernald, B., Vickers, J. H., Suomi, S.J., & Linnoila, M. (1996b). Excessive mortality in young free-ranging male nonhu-man primates with low CSF 5–HIAA concentrations. Archives of General Psychia-try, 53, 537–543.
Higley, J. D., Mehlman, P. T., Taub, D. M., Higley, S. B., Vickers, J. H., Suomi, S. J., &Linnoila, M. (1992a). Cerebrospinal fluid monoamine and adrenal correlates of ag-gression in free-ranging rhesus monkeys. Archives of General Psychiatry, 49, 436–441.
Higley, J. D., & Suomi, S. J. (1996). Reactivity and social competence affect individualdifferences in reaction to severe stress in children: Investigations using nonhumanprimates. In C. R. Pfeffer (Ed.), Intense stress and mental disturbance in children(pp. 3–58). Washington, DC: American Psychiatric Press.
Higley, J. D., Suomi, S. J., & Linnoila, M. (1990a). Developmental influences on theserotonergic system and timidity in the nonhuman primate. In E. M. Coccaro & D.L. Murphy (Eds.), Serotonin in major psychiatric disorders (pp. 27–46). Washing-ton, DC: American Psychiatric Press.
Higley, J. D., Suomi, S. J., & Linnoila, M. (1990b). Parallels in aggression and serotonin:Consideration of development, rearing history, and sex differences. In H. van Praag,R. Plutchik, & A. Apter (Eds.), Violence and suicidality (pp. 245–256). New York:Bruner/Mazel.
Higley, J. D., Suomi, S. J., & Linnoila, M. (1991b). CSF monoamine metabolite concen-trations vary according to age, rearing, and sex, and are influenced by the stressor ofsocial separation in rhesus monkeys. Psychopharmacology, 103, 551–556.
Higley, J. D., Suomi, S. J., & Linnoila, M. (1992b). A longitudinal assessment of CSFmonoamine metabolite and plasma cortisol concentrations in young rhesus mon-keys. Biological Psychiatry, 32, 127–145.
Higley, J. D., Suomi, S. J., & Linnoila, M. (1996c). A nonhuman primate model of Type IIexcessive alcohol consumption? Part 1: Low CSF 5–HIAA concentrations and di-minished social competence correlate with excessive alcohol consumption. Alco-holism: Clinical and Experimental Research, 20, 629–642.
Higley, J. D., Suomi, S. J., & Linnoila, M. (1996d). A nonhuman model of Type II alco-holism? Part 2: Diminished social competence and excessive aggression correlateswith low CSF 5–HIAA concentrations. Alcoholism: Clinical and Experimental Re-search, 20, 643–650.
Higley, J. D., Thompson, W. T., Champoux, M., Goldman, D., Hasert, M. F., Kraemer, G.W., Scanlan, J. M., Suomi, S. J., & Linnoila, M. (1993). Paternal and maternal ge-netic and environmental contributions to CSF monoamine metabolites in rhesusmonkeys (Macaca mulatta). Archives of General Psychiatry, 50, 615–623.
80 T H E D E V E L O P M E N T O F A G G R E S S I O N

Lahey, B. B., Moffitt, T. E., & Caspi, A. (2003). Causes of conduct disorder and juveniledelinquency. New York: Guilford Press.
Lesch, K. P., Bengel, D., Heils, A., Sabol, S. Z., Greenberg, B. D., Petri, S., Benjamin, J.,Muller, C. R., Hamer, D. H., & Murphy, D. L. (1996). Association of anxiety-re-lated traits with a polymorphism in the serotonin transporter gene regulatory re-gion. Science, 274, 1527–1531.
Lesch, L. P., Meyer, J., Glatz, K., Flugge, G., Hinney, A., Hebebrand, J., Klauck, S. M.,Poustka, A., Poustka, F., Bengel, D., Mossner, R., Riederer, P., & Heils, A. (1997).The 5–HT transporter gene-linked polymorphic region (5–HTTLPR) in evolution-ary perspective: Alternative biallelic variation in rhesus monkeys. Journal of NeuralTransmission, 104, 1259–1266.
Lindburg, D. G. (1971). The rhesus monkey in north India: An ecological and behavioralstudy. In L. A. Rosenblum (Ed.), Primate behavior: Developments in field and labo-ratory research (Vol. 2, pp. 1–106). New York: Academic Press.
Linnoila, M. (1988). Monoamines and impulse control. In J. A. Swinkels & W. Blijeven(Eds.), Depression, anxiety, and aggression (pp. 167–172). Houten, The Nether-lands: Medidact.
Linnoila, M., DeJong, J., & Virkkunen, M. (1989). Monoamines, glucose metabolism,and impulse control. Psychopharmacy Bulletin, 25, 404–406.
Linnoila, M., Virkkunen, M., Scheinin, M., Nuutila, A., Rimon, R., & Goodwin, F. K.(1983). Low cebrospinal fluid 5–hydroxyindoleacetic acid concentration differenti-ates impulsive from nonimpulsive violent behavior. Life Sciences, 33, 2609–2614.
Mehlman, P. T., Higley, J. D., Faucher, T., Lilly, A. A., Taub, D. M., Vickers, J., Suomi, S.J., & Linnoila, M. (1994). Low cerebrospinal fluid 5–hydroxyindoleacetic acid con-centrations are correlated with severe aggression and reduced impulse control infree-ranging nonhuman primates (Macaca mulatta). American Journal of Psychia-try, 151, 1485–1491.
Mehlman, P. T., Higley, J. D., Faucher, I., Lilly, A. A., Taub, D. M., Vickers, J. M., Suomi,S. J., & Linnoila, M. (1995). CSF 5–HIAA concentrations are correlated withsociality and the timing of emigration in free-ranging primates. American Journal ofPsychiatry, 152, 907–913.
Mehlman, P. T., Higley, J. D., Fernald, B. J., Sallee, F. R., Suomi, S. J., & Linnoila, M.(1997). CSF 5–HIAA, testosterone, and sociosexual behaviors in free-ranging malemacaques during the breeding season. Psychiatric Research, 72, 89–102.
Newman, T. K., Syagailo, Y., Barr, C. S., Wendland, J., Champoux, M., Graessle, M.,Suomi, S. J., Highley, J. D., & Lesch, K. P. (in press). Monoamine oxidase: A genepromoter polymorphism and infant rearing experience interact to influence aggres-sion and injuries in rhesus monkeys. Biological Psychiatry.
Ruppenthal, G. C., Harlow, M. K., Eisele, C. D., Harlow, H. F., & Suomi, S. J. (1974).Development of peer interactions of monkeys reared in a nuclear family environ-ment. Child Development, 45, 670–682.
Steklis, H. D., Brammen, G. L., Raleigh, M. J., & McGuire, M. T. (1985). Serum testos-terone, male dominance, and aggression in captive groups of vervet monkeys(Cercopithecus aethiops sabacus). Hormones and Behavior, 19, 156–165.
Suomi, S. J. (1995). Influence of Bowlby’s attachment theory on research on nonhumanprimate biobehavioral development. In S. Goldberg, R. Muir, & J. Kerr (Eds.), At-tachment theory: Social, developmental, and clinical perspectives (pp. 185–201).Hillsdale, NJ: Analytic Press.
Suomi, S. J. (1999). Attachment in rhesus monkeys. In J. Cassidy & P. R. Shaver (Eds.),
Genetic and Environmental Factors in Rhesus Monkeys 81

Handbook of attachment: Theory, research, and clinical applications (pp. 181–197). New York: Guilford Press.
Suomi, S. J., & Harlow, H. F. (1975). The role and reason of peer friendships. In M. Lewis& L. A. Rosenblum (Eds.) Friendships and peer relations (pp. 310–334). New York:Basic Books.
Suomi, S. J., & Levine, S. (1998). Psychobiology of intergenerational effects of trauma:Evidence from animal studies. In Y. Daniele (Ed.), International handbook ofmultigenerational legacies of trauma (pp. 623–637.) New York: Plenum Press.
Symonds, D. (1978). Play and aggression: A study of rhesus monkeys. New York: Co-lumbia University Press.
Westergaard, C. G., Suomi, S. J., Chavanne, T. J., Houser, L., Hurlye, A. C., Cleveland,A., Snoy, P., & Higley, J. D. (2003). Physiological correlates of aggression andimpulsivity in free-ranging female primates. Neuropsychopharmacology, 28, 1045–1055.
82 T H E D E V E L O P M E N T O F A G G R E S S I O N

THE DEVELOPMENT OF AGGRESSIONPhysical Aggression in Humans
5
The Developmental Originsof Physical Aggression in Humans
RICHARD E. TREMBLAY and DANIEL S. NAGIN
We here saw the native Fuegan; an untamed savage is I reallythink one of the most extraordinary spectacles in the world.—thedifference between a domesticated & wild animal is far morestrikingly marked in man . . . with difficulty we see a fellow-creature.
—DARWIN (1832/1985, pp. 302–303)
The aim of this chapter is to summarize our present knowledge on the devel-opment of physical aggression from early childhood to adulthood, and to dis-cuss the mechanisms that could explain this development. We define physicalaggression as the use of physical force against another person with an object(e.g., stick, rock, bullet) or without (e.g., slap, push, punch, kick, bite). Theverbs fight, assault, attack, and bully are often used to summarize the actionsduring a physical aggression. Antecedents and consequences can be used tosubdivide this subcategory of aggressions (see Gendreau & Archer, Chapter 2,this volume). However, inasmuch as most developmental studies of physicalaggression have not made these distinctions, we will generally refer to an over-all category of physical aggression.
THE SOCIAL–HISTORICAL–PHILOSOPHICAL–MORAL CONTEXTOF RESEARCH ON THE DEVELOPMENT OF PHYSICAL AGGRESSION
Physical aggression is a socially “hot” topic. An act of physical aggression canmake the difference between life and death. Physical violence is omnipresentdaily in the mass media of modern societies (Bushman & Anderson, 2001) in
83

the same way that it is a constant preoccupation for animals living in the wild.Physical aggression is of concern as much to the health professionals (WorldHealth Organization, 2002) as to the social–psychological well-being profes-sionals (U.S. Human Capital Initiative, 1997).
Concerted efforts to suppress physical aggression in human interactions isone of the major historical social transformations observed in countries of theWestern world in the later part of the 20th century. A half-century ago, it wasgenerally considered normal for elementary school teachers to physically pun-ish students for spelling errors. Elementary school principals used leatherstraps to discipline disruptive students. Caning was still in use in English pub-lic and state schools. In other words, some forms of physical force against oth-ers were considered morally acceptable, and were even recommended. Notethat in all of these examples an adult uses physical force against a child. Twolearning specialists who spent their careers studying nonhuman primates con-clude that physical punishment is the natural pedagogical tool (Premack &Premack, 2003). There is now a strong movement to make these practicescriminal acts. Some go as far as to argue that parents should not take theirchildren in their arms without asking the children for their permission. Zerotolerance has become the rule concerning physical aggression on the schoolplayground, in the neighborhood, and at home, between spouses and betweenparents and children. Not using physical force against another human has be-come the hallmark of a civilized human.
The difference in attitudes between 1950 and 2000 is certainly striking.But the difference is still more impressive when we compare late-20th-centuryattitudes to those of past centuries. Consider the educational practices in oneof the great eras of the English public schools. When the 24-year-old CharlesDarwin described to his sister the marked difference between “an untamedsavage” and his civilized compatriots, the famous educationist and scholarThomas Arnold had been appointed headmaster of Rugby for less than 4years. Harold Nicolson (1955) writes that Arnold was convinced that the boysshould be whipped to learn their moral responsibilities. To enforce his “mus-cular Christianity,” he organized a disciplinary system in which older boysruled by fagging and by beating the younger boys. All accounts of 19th-century English public schools indicate that bullying was not the exceptionbut the rule. Nicolson cites the following description from Thackeray: “Thereare at this present writing 500 boys at Eton, kicked and licked and bullied byanother 100” (1955, p. 263). Previous centuries were not less violent. Con-sider the general use of torture and homicide by kings and priests during themost civilized times of Erasmus, Luther, Galileo, Mozart, and Voltaire. In-deed, Eisner (2003) documents that homicide rates in western Europe declined50- to 100-fold over the past 5 centuries. And what did the civilized Romansenjoy most on a nice leisurely afternoon? A bloody battle among gladiatorsand starved lions!
From an evolutionary perspective, aggression is a solution to a problem(Archer, 1988). From a social, historical, and moral perspective, humans de-
84 T H E D E V E L O P M E N T O F A G G R E S S I O N

cide at different places and times which solutions are acceptable. Concerningthe use of the word aggress, the American Heritage Dictionary (1985) says:“Though the verb aggress has a long and honorable history, it has lately cometo be associated primarily with the jargon of psychology and is often objectedto.” Indeed, there seems to have been much change in our attitudes toward theuse of physical force by humans. But how deeply rooted is this change? Thetwo world wars of the 20th century killed close to 30 million soldiers, Hitlermanaged to exterminate approximately 6 million Jews, and nearly 200,000Japanese civilians were killed by two atomic bombs on Hiroshima and Naga-saki. The United States saw a huge increase in homicides and other violentcrimes during the 1990s, mainly among adolescents and young adults (Blum-stein & Wallman, 2000; McCord, Widom, & Crowell, 2001). On December31, 2002, the number of persons in U.S. prisons was equivalent to approxi-mately half the population of Norway (2 million), and a year earlier the rich-est country in the history of humanity had close to 70 prisoners waiting to beexecuted for crimes they committed before they were 18 years of age (Bureauof Justice Statistics, 2003; Steinberg & Scott, 2003). Planes are hijacked to de-stroy skyscrapers filled with thousands of people. We are still killing tens ofthousands of people to gain control of a country, and in some European citiesconflicts force children to go to school under the protection of the army.
The physical violence described here victimizes millions of humans, butonly a very small minority of the inhabitants of the wealthy countries will beamong these victims. Paradoxically, this fortunate minority, who are less andless at risk of being physically aggressed by a parent, a teacher, a spouse, aneighbor, a stranger, or a soldier, are using a large part of their financial re-sources and leisure time to consume fictive depictions of physical violence ontelevision, in movies, and in electronic games.
This behavior of the most civilized humans in the history of humanity iscertainly quite a puzzlement. But still more puzzling is the contrast betweenthis strange addiction to fictive violence by humans who have been educatednot to use physical violence, and the results of research published in the topscientific journals, which conclude that the more individuals look at physicalviolence, the more they become physically aggressive (Anderson & Bushman,2002; Johnson, Cohen, Smailes, Kasen, & Brook, 2002). Thanks to moderntechnology, citizens of present-day industrialized nations have, during theirlives, seen more physical aggressions causing death than any Roman who hada lifelong Coliseum season ticket. By what peculiar process are these hungry-for-physical-violence modern humans passing laws to eradicate physical vio-lence from their daily lives?
THE AGE–VIOLENT CRIME CURVE
Two important scientific committees published reports in the 1990s on thecauses and prevention of violence. The U.S. National Research Council panel
Physical Aggression in Humans 85

(Reiss & Roth, 1993) concluded that “modern psychological perspectives em-phasize that aggressive and violent behaviors are learned responses to frustra-tion, that they can also be learned as instruments for achieving goals, and thatthe learning occurs by observing models of such behaviors. Such models maybe observed in the family, among peers, elsewhere in the neighborhood,through the mass media, or in violent pornography, for example” (p. 7, em-phasis in original). Four years later the report of the U.S. Human Capital Ini-tiative (1997) coordinating committee on reducing violence concluded:
In short, watching violent movies and television shows year after year and lis-tening to brutal lyrics set to throbbing music can change one’s attitudes aboutantisocial, aggressive behavior. In children it can lead to more aggressive be-havior and also can evoke unwarranted fears and defensive actions. What-ever the violent content, movies and television exert powerful influencesthrough visual imagery and dramatic characterizations; video games mayhave similar effects. (p. 12)
Figure 5.1 is a representation of the age-specific rate (per 100,000) of individ-uals arrested (United States) and accused (Canada) for violent offenses (physi-cal aggressions) in 2001. It can be seen that violent offenses appear inpreadolescence, increase sharply during adolescence, and then decrease slowly.If young humans learn to physically aggress by imitation of aggressions ob-served in the home, in the neighborhood, and in the media, the increase inphysical violence during adolescence makes sense. The older they are, themore they have observed acts of physical aggression, and the more likely theyare to do what they have seen. The beauty of these data is not only that theyconfirm the prevalent psychological theory, but they replicate the data thatAdolphe Quetelet (1833), the Belgian astronomer/statistician, started collect-ing in the late 1820s. This was at the time Thomas Arnold was becomingheadmaster of Rugby and Charles Darwin was a student at Cambridge. WhileDarwin was traveling around the world to catch beetles and observe volca-noes, the Belgian astronomer/statistician decided to quantify the physical,cognitive, and moral development of humans.
The age–crime curve is probably the most robust criminological observa-tion (see Sampson & Laub, 2003). However, if social learning explains the in-crease in physical violence during adolescence, it does not explain the decreaseduring adulthood. Quetelet, who was interested in moral development, butalso in cognitive and physical development (he invented the Body Mass In-dex), used a biological explanation (intensity of physical strength and pas-sions), which is close to the modern biological explanation that challenges thesocial learning hypothesis. The modern explanation is testosterone (see Ellis& Coontz, 1990; Harris, Rushton, Hampson, & Jackson, 1996; Tremblay etal., 1997; and Van Goozen, Chapter 14, this volume). T, as it is known by en-docrinologists, has been shown to correlate with physical violence and notonly increases substantially from early to late adolescence, but also decreases
86 T H E D E V E L O P M E N T O F A G G R E S S I O N

during adulthood, hence following the age–violent-crime curve. Testosteronehas the added power to explain the sex difference in physical aggression. In-deed, T is the male hormone. Although the adrenal glands and ovaries ofwomen produce T and this production increases during adolescence, the levelsof T at the end of adolescence are 20 times higher in males than in females.From this perspective, in his explanation of the causes of antisocial behavior,Quetelet would have been closer to the truth than Jean-Jacques Rousseau, themost famous advocate of the theory of social learning as an explanation ofbad behavior (Rousseau, 1762/1979). T feeds muscles and thus increasesstrength: the fuel for Quetelet’s strength and passion hypothesis.
AGGRESSION BEFORE PUBERTY
The main problem with the age–violent-crime curve studies is that they focuson adolescents and adults. Criminologists have tended to neglect humans be-fore the start of adolescence because they cannot be considered legally delin-quent. Since Quetelet, at least, criminologists have fed on data produced bythe legal system. Only very recently have they started to study precursors ofcriminal behavior before adolescence. Do humans physically aggress eachother before the start of adolescence? The age–violent-crime curve figures givethe impression that physical aggressions appear with the legal age for criminal
Physical Aggression in Humans 87
FIGURE 5.1. Age–crime curve in 2001 in the United States and Canada.

responsibility as if lawmakers had chosen the age for criminal responsibilityafter detailed studies of child development. Those who decided to studyelementary school children to understand the precursors of adolescent delin-quency discovered that elementary school children use physical aggressionbefore puberty increases their production of T, and before they have accumu-lated 12 or 13 years of social learning.
The Columbia County Study, initiated in the state of New York in 1960,was one of the rare longitudinal studies of elementary school children specifi-cally focused on aggression. More than 800 third-grade schoolchildren ratedeach other on different behavioral dimensions, including aggression, to testthe social learning hypothesis. Following the analysis of the first wave of datawhen the children were 8 years old, the investigators gave the following sum-mary of their findings: “It is fair to say that all the major findings of that anal-ysis were consistent with the hypothesis that aggression may be learned by achild from his interactions with his environment” (Lefkowitz, Eron, Walder,& Huesmann, 1977, p. 37). Note that this was a conclusion drawn fromcross-sectional data. Later analyses of data collected at ages 8, 19, and 30 re-vealed relatively high correlations between aggression assessments at these dif-ferent points in time (Huesmann, Eron, Lefkowitz, & Walder, 1984).
These results led Eron (1990), in his presidential address to the Interna-tional Society for Research on Aggression in 1990, to conclude that aggressioncrystallizes at 8 years of age. Thus, the social learning of aggression would oc-cur much earlier than the start of the age–violent-crime curve. But there weretwo important limitations with the Columbia County Study. First, the aggres-sion scale used at age 8 should have been labeled a “disruptive behavior” scale(e.g., who does not obey the teacher, who gives dirty looks, who does thingsthat bother others, who is always getting in trouble). Only 2 of the 10 itemsreferred specifically to physical aggressions (who starts a fight over nothing;who pushes and shoves children). The second limitation is that the data collec-tion points had 10-year gaps, making it impossible to study developmentaltrajectories.
Fortunately, a similar U.S. study was initiated 20 years later and collectedannual data on physical aggression. The Carolina Longitudinal Study fol-lowed 220 boys and girls from 4th to 12th grade and obtained assessments ofphysical aggression from the students and the teachers (Cairns & Cairns,1994). Analyses of these data painted an unexpected picture in regard to the“social learning” and “testosterone” hypotheses. The information from teach-ers, as well as from students, showed a clear mean decrease in the frequency ofphysical aggression from 10 to 18 years of age, and this was true for males aswell as for females (Cairns, Cairns, Neckerman, Ferguson, & Gariépy, 1989).How could longitudinal data from a population sample of American adoles-cents, living in a southern state, show a physical aggression curve exactly the re-verse of the age–violent-crime curve?
One explanation could be that although the majority of youth reduce thefrequency of physical aggressions as they grow older, a minority are increasing
88 T H E D E V E L O P M E N T O F A G G R E S S I O N

their frequency of physical violence and are being processed through the legalsystem that generates the statistics for the age-crime curve. To test this hypoth-esis, we needed to go beyond a description of the mean developmental trajec-tories of a given sample and identify the different types of developmental tra-jectories that children were following. Nagin and Tremblay (1999) used asemiparametric mixture model with a longitudinal sample of more than 1,000kindergarten boys in schools from low socioeconomic areas of Montreal todescribe these different trajectories. Why use males from low socioeconomicareas to study the development of physical aggression? The answer can befound in the report “Reducing Violence” from the U.S. Human Capital Initia-tive (1997) coordinating committee: “Certain environmental conditions notonly trigger violence, they can also seem to teach aggressive or violent behav-ior patterns. For example, poverty is associated with both sudden violent out-bursts and long-term, habitual aggression” (p. 11).
Results (Figure 5.2) from teacher ratings confirmed the North Carolinadata. The large majority of boys from the poorest inner city areas of Canadawere using physical aggression less frequently as they grew older. Only a verysmall group of boys (4%) did not show the declining trend; these were theboys who had the highest levels of physical aggression in kindergarten, whichremained at the highest level until adolescence. When interviewed at ages 15and 17, they were the boys who reported the highest frequency of physical vi-olence and those the ones most frequently found guilty of infractions before
Physical Aggression in Humans 89
FIGURE 5.2. Physical aggression trajectories. From Nagin and Tremblay (1999). Copyright1999 by the Society for Research in Child Development. Reprinted by permission.

18 years of age. Thus, the increase in the age-crime curve during adolescencefor physical aggressions would be produced by the fact that, during this pe-riod, the police and judicial system start arresting and convicting individualswho have been physically aggressing others at least since kindergarten. An in-ternational team of investigators replicated these finding using five other lon-gitudinal studies in Canada, New Zealand, and the United States (Broidy et al.2003).
AGGRESSION BEFORE SCHOOL ENTRY
Figure 5.2 clearly shows that all the boys tended to be at their peak level infrequency of physical aggression at 6 years of age, when they were at the endof their kindergarten year. If this is the case, when do such children start to usephysical aggression? Is the social learning of physical aggression occurringduring the kindergarten year? Studies of preschool children suggest that if hu-mans need to learn to physically aggress, the learning occurs before school en-try.
There is a long history of case studies of physical aggression in youngchildren. One of the first publications of a “developmental origins of aggres-sion” study appeared during the reign of the Roman Emperor Arcadius (A.D.395–408). The 46-year-old Augustine of Thagaste (397/1960, p. 49), laterknown as St. Augustine, when writing his Confessions, realized that he couldnot remember his first sins. Instead of concluding, like Sigmund Freud, that heneeded to “psycho-analyze” himself, he decided to observe babies, reasoning,in all humility, that they must be doing what he did at their age. Followingthese observations he writes: “Were these things good . . . [when] my parents,and many other prudent people who would not indulge in my whims, when Istruck at them and tried to hurt them as far as I could because they did notobey orders that would be obeyed only to my harm?” One millenium and ahalf later, in the “Rage” section of the The Expression of the Emotions inMan and Animals, Darwin (1872, p. 243) wrote: “Every one who has hadmuch to do with young children must have seen how naturally they take tobiting, when in a passion. It seems as instinctive in them as in young croco-diles, who snap their little jaws as soon as they emerge from the egg.”
Fifty-nine years after this other jewel by Darwin, two books were pub-lished by two women who pioneered 20th-century quantitative studies ofearly human social-emotional development. The two books used differentmethodologies. Katharine M. Bantham Bridges was a professor of psychologyin the McGill department of psychology and a psychologist in the university’snursery school. Her book (Bridges, 1931) presented social development andemotional development scales created by daily observations of 2–5-year-oldchildren in the nursery school. One of her conclusions was that children, morethan adults, were susceptible to anger expressed by physical aggression. Inter-estingly, 2 years later, she introduced a paper on the observation of 62 chil-
90 T H E D E V E L O P M E N T O F A G G R E S S I O N

dren, between 3 weeks and 2 years of age, by writing that “the trends of devel-opment” between 2 and 5 years of age described in her previous book “musthave had their beginnings in early infancy” (Bridges, 1933, p. 36). Her 3-month cross-sectional observation of infant interactions led her to the conclu-sion that by 1 year of age children start “to struggle slightly for possession” ofobjects, and by 15 months “aggressive attacks are indulged in at this stage fortheir own sake and are not out-grown till well after two years of age” (p. 48).
At exactly the same time, Florence Goodenough (1931), a professor atthe University of Minnesota Institute of Child Welfare, was studying anger inyoung children by asking “intelligent parents,” mostly college graduates, tokeep records of their children’s anger outbursts during a 1-month period. Twochildren were not yet 1 year old (7 and 11 months), 9 were between 17 and 23months, 13 were between 26 and 35 months, 10 were between 36 and 46months, and the 11 others ranged from 53 to 94 months. One thousand eighthundred and seventy-eight anger outbursts were recorded, for a mean of ap-proximately 1 outburst per 10 hours of observation. As shown in Figure 5.3a,the 17- to 23-month-olds had the highest mean, with 1.3 outbursts per 10hours. Interestingly, data from a longitudinal study of temper tantrums re-ported by the mothers of a sample of 126 Belgian children at the end of the1950s led to very similar results (Sand, 1966; Figure 5.3b). Goodenough andSand specifically studied the frequency of physical and verbal aggressions dur-ing the angry outbursts. They both concluded that as they grow older, childrenuse fewer physical aggressions. Goodenough specifically concluded that withage they attempt more often to hurt the feelings of others (see Vaillancourt,Chapter 8, this volume).
An observational study of French children’s social interactions in daycareduring the 1970s reported on the frequency of physical aggressions, as com-pared with other forms of social interactions, for children of different age groups(Restoin et al., 1985). A total of 30 hours of filmed interactions were coded. Re-sults represented in Figure 5.3c show a trend that is very similar to those in par-ent reports in Figures 5.3a and 3b. The relative frequency of physical aggressionincreases from the first year after birth to the second, and then decreases.Holmberg (1977) also found that physical attacks increased from 12 to 30months, and then decreased from 30 to 42 months, when she observed 16 boysand girls interacting with peers in the United States. Recently, similar resultswere obtained with parent reports in two large Canadian-based longitudinaland cross-sectional studies. From 9 months to 30 months of age the proportionof children who are reported to use physical aggressions increases substantially(Tremblay, 2004); in addition, the frequency of use appears to increase up tonearly the end of the third year after birth (Tremblay et al., 2004). This remark-able increase is then followed by a continuous decline in frequency (CôtéVaillancourt, LeBlanc, Nagin, & Tremblay, 2004; Tremblay et al., 1996).
Clearly, the same general developmental picture is drawn whether we usedata from different times (1920s, 1970s, or 1990s), data from different coun-tries (Belgium, Canada, France or the United States), data from different re-
Physical Aggression in Humans 91

porting sources (filmed interactions in day care, parents’ detailed records, orparents’ recall of behavior in the past months), or data from different method-ologies (cross-sectional or longitudinal, calculated as an absolute frequencyover a given period of time, a relative percentage of social behaviors, a per-centage of individuals using the behavior, or a general estimate of the fre-quency of the behavior). Frequency of anger outbursts and physical aggressionincrease rapidly from the first year after birth to approximately the third, andthen the frequency decreases. Unfortunately, none of the longitudinal studiesthat started in early childhood have yet reported on physical aggression up tothe end of adolescence.
From the Restoin et al. (1985) data it appears that at the peak frequencyof physical aggression in a day care environment, physical aggression repre-sents one in four social interactions. This is certainly a high frequency of ag-
92 T H E D E V E L O P M E N T O F A G G R E S S I O N
FIGURE 5.3a. Frequency of anger outbursts for 10 hours of observation. Data from Good-enough (1931).
FIGURE 5.3b. Percentage of children reported to have a temper tantrum at least every day.Data from Sand (1966).

gressions, but even at the worst of times, there are more prosocial or neutralsocial interactions than there are physically aggressive ones. This can probablybe said for any social group, whether it is composed of nonhuman primates(see de Waal, 2000; also see Suomi, Chapter 4, this volume) or Hell’s Angels.Social life would probably be impossible within a group where physical ag-gression is the most frequent form of social interaction. However, it does ap-pear clear that most humans have used physical aggression before they reach36 months of age, that humans use physical aggression most often between 18and 42 months after birth, and that if humans are learning to physically ag-gress through imitation, this learning is happening in the first 2 years afterbirth, not by watching television and playing video games in middle childhoodor adolescence. After many decades of training chimpanzees to use “lan-guage,” David and Ann Premack (2003, p. 72) wrote: “Biting, hitting, kick-ing, scratching, pushing are natural reactions to the offensive behavior of an-other individual. These require no training.”
CAUSES OF PHYSICAL AGGRESSION DURING EARLY CHILDHOOD
The causes of physical aggression during early childhood can be consideredfrom many different perspectives, both with reference to the sequence ofevents (e.g., proximal and distal causes) and with reference to the level ofanalysis (e.g., behavioral, psychological, physiological, genetic). For example,from the proximal perspective we can investigate the time, the place, and theevents that appear to trigger physical aggression by an infant. We can also fo-cus on the emotional characteristics of the child prior to, during, and after anagonistic interaction. Such investigations could target the external expression(e.g., facial expression) or the physiological expression (e.g., heart rate,cortisol levels).
Physical Aggression in Humans 93
FIGURE 5.3c. Relative frequency of aggressive behavior between peers from age 6 to 36months in day care centers. Data from Restoin et al. (1985).

Most observers of proximal causes of physical aggressions by infants andtoddlers have concluded that anger is the main “emotional” antecedent, andthat this anger is provoked either by something that prevents the child fromgetting what he or she wants, or by a request for the child to do something heor she does not want to do (e.g., Bridges, 1933; Goodenough, 1931; Hay,Chapter 6, this volume). The most frequent situational “cause” of physical ag-gressions among young children is competition for desirable objects or space.Note that these motivations appear not very different from those that leadadult humans to war between countries, inasmuch as these tend to be insti-gated by competition over desirable resources (e.g., oil) or territories (e.g., foraccess to the sea). Thus, in humans, as in other species, from the beginning oflife onward, the function of many instances of physical aggressions is competi-tion for resources (see Archer, 1988).
The developmental trajectories described earlier in this chapter do nothelp much in identifying proximal causes of physical aggressions, but they dohelp in understanding distal causes. The distal perspective tends to look forthe mechanisms that are put in place over a long period of time and explainsat least two different mechanisms: (1) how an individual acquires the abilityto physically aggress and (2) why he or she tends to use this ability. For exam-ple, the social learning hypothesis appears to argue that a human who hasnever witnessed a physical aggression in his life could not use physical aggres-sion, even when extremely angry, because he or she would not have “learned”to physically aggress. It is a very different argument from the one stating thatinfants do not need to learn to physically aggress, but the more they witnessphysical aggressions the more they are likely to use them when they becomeangry. The first part of the prior sentence refers to mechanism (1), and the sec-ond part refers to mechanism (2).
The developmental data we have presented in this chapter appear to indi-cate that children do not need to have seen models of physical aggression inorder to start using physical aggression (onset), unless this learning throughmodels occurs for almost every child during the first 9–15 months after birth.Indeed, by 17 months of age most children are reported by their parents tohave used some form of physical force to get what they want or to expresstheir anger (Tremblay et al., 1999a).
The developmental trajectories of physical aggression also indicate thathaving frequently observed models of physical aggression generally does notlead to an increase in the frequency of physical aggression. This can be con-cluded from the fact that, as they grow older, the majority of children haveseen more and more instances of physical aggression, and their frequency ofphysical aggression decreases. If models of physical aggression have an im-pact, it is probably by reducing the speed at which children learn not to usephysical aggression. Indeed, the declining frequency of physical aggressionswith age indicates that the vast majority of children are learning not to usephysical aggression as they grow older. It could be argued that the reason forthe decline is the reduction of occasions for conflict as children grow older.
94 T H E D E V E L O P M E N T O F A G G R E S S I O N

However, the increase in verbal and indirect aggressions with age (seeVaillancourt, Chapter 8, this volume) suggests that reasons for physical ag-gressions remain, but that children are learning other ways to deal with them.
The developmental trajectories described in this chapter indicate that liv-ing with others, even in violent environments, forces individuals to learn, asthey grow older, to gain some control over their social behavior. Thus, thereseems to be much more pressure to learn not to physically aggress than tolearn to physically aggress. Even children in “high-risk” families are likely toobserve a greater proportion of nonphysically aggressive interactions in theirenvironment, because there is a high probability that no social group couldsurvive for long if the majority of their interactions were physically aggressive.
There has been much research over the past 2 decades on the “onset” ofchildren’s disruptive behaviors, such as physical aggressions, property “of-fenses” (e.g., stealing), and hyperactivity. Most of that work has focused onschool-age children (e.g., Lahey et al., 1999; Loeber, Green, Keenan, & Lahey,1995; Tremblay, Pihl, Vitaro, & Dobkin, 1994). The longitudinal data wehave examined from infancy onward leads us to think that physical aggressionis not a behavior children learn, like reading or writing, nor an illness children“catch,” like poliomyelitis or smallpox. It is rather a behavior like crying, eat-ing, sleeping, grasping, throwing, and running, which young humans exhibitwhen the physiological structure is in place, but then learn to control with ex-perience. The learning-to-control process implies regulating one’s needs to ac-commodate those of others, and this process can be labeled “socialization”(see Paus, Chapter 12, as well as Séguin & Zelazo, Chapter 15, this volume).
It is not hard to understand why the evolutionary process would give hu-mans a genetic program that codes for all the basic mechanisms to react tohunger and to threat. Young children’s muscles are activated to run, push,kick, grab, hit, throw, and yell with extreme force when hungry, when angry,or when they are strongly attracted by something. In his Expression of theEmotions, Darwin (1872) anticipates present-day brain imaging techniqueswhen he describes the reddening of the skin during a state of rage and givesthis example: “With one of my own infants, under four months old, I repeat-edly observed that the first symptom of an approaching passion was the rush-ing of the blood into his bare scalp” (p. 240). Stating that humans are geneti-cally programmed to be able to physically aggress when needed, is differentfrom stating that the frequency of the physical aggression they use is geneti-cally programmed. Thus, although all 18-month-olds who have developednormally can, and possibly do, physically aggress, not all do so at the samefrequency and with the same vigor.
The trajectories shown in Figure 5.2 clearly indicate that these individualdifferences exist at any given point, but also over time. The gene–environmentmechanisms are probably very similar to the developmental trajectories ofgrowth in height. Genes code for the growth mechanisms, but there are indi-vidual differences in this coding, as well as environmental differences (e.g., ac-cess to food) that lead to individual differences in growth in height. Thus,
Physical Aggression in Humans 95

these individual differences in the frequency of physical aggressions can be dueto a large number of causes—for example, individual differences in the geneticcoding for serotonin (see Pihl & Benkelfat, Chapter 13, this volume), testos-terone (Van Goozen, Chapter 14, this volume), or language development (seeDionne, Chapter 16, this volume), or cognitive development (see Séguin &Zelazo, Chapter 15, this volume), or to environmental differences such as to-bacco use during pregnancy (see Wakschlag, Pickett, Cook, Benowitz, &Leventhal, 2002), birth complications (see Arseneault, Tremblay, Boulerice, &Saucier, 2002), parental care (see Gatti & Tremblay, Chapter 19, this volume;Raine, Brennan, & Mednick, 1997; Zoccolillo et al., Chapter 17, this vol-ume), and peer characteristics (see Boivin, Vitaro, & Poulin, Chapter 18, thisvolume). However, the individual differences that we observe are very likely tobe due to interactions between many of these mechanisms, and hence toepigenetic mechanisms (Francis, Diorio, Plotsky, & Meaney, 2002; Francis,Szegda, Campbell, Martin, & Insel, 2003; Raine, 2002; Tremblay, 2003;Weaver et al., 2004).
Dionne, Tremblay, Boivin, Laplante, and Pérusse (2003) showed, with alarge sample of 19-month-old female and male twins, that variance in moth-ers’ reports of physical aggression was explained about equally by genetic andenvironmental factors. Similar results were observed with an older sample oftwins for assessments of antisocial behavior (e.g., Arseneault et al., 2003;Slutske et al., 1997), but the Dionne et al. study shows that both the geneticand environmental distal causes of physical aggression are in place by 19months of age, long before the violent video games and the delinquent peers.A similar story is being told by the molecular genetic studies of physical vio-lence. With a longitudinal study of a New Zealand male birth cohort, Caspi etal. (2002) observed that children of abusive and neglectful parents who had agenotype leading to high levels of monoamine oxidase A (MAOA) expressionwere at much less risk of having a disposition toward violence as young adultsand being convicted for violent crimes.
We need molecular genetic and twin studies to identify which environ-mental factors interact with which genetic factors to explain the individualdifferences in developmental trajectories of physical aggression from earlychildhood to adulthood. The following is an example of the first few steps in aprogram of longitudinal studies that pursues this aim. A large population sam-ple of newborns (singletons and twins) are assessed annually from 5 monthsonward, with physical aggression assessed annually from 17 months (Dionneet al., 2003; Tremblay et al., 2004). Semiparametric statistical analyses ofphysical aggression data at 17, 30, and 42 months with the singletons indi-cated that there were three different developmental trajectories (Figure 5.4).Variables measuring parental characteristics before birth were used as predic-tors of the probability that a child would be on the high-level physical aggres-sion trajectory. Controlling for all other measured variables, the best predic-tors were presence of a sibling (odds ratio [OR] = 4.00), mother’s history ofantisocial behavior before the end of high school (OR = 3.1), mother’s age
96 T H E D E V E L O P M E N T O F A G G R E S S I O N

when her first child was born (OR= 3.1), low family income (OR= 2.6), and amother who smoked (OR= 2.2). Two variables measured at 5 months of agealso contributed to the prediction: mother’s coercive parenting (OR = 2.3) andfamily dysfunction (OR = 2.2).
This list of predictors for high levels of physical aggression from infancyto toddlerhood is amazingly similar to the variables that have traditionallybeen shown to predict physical aggression during the school years and delin-quency during adolescence (Bovet, 1951; Brennan, Grekin, & Mednick.,1999; Loeber & Dishion, 1983; Nagin & Tremblay, 2001; Tremblay &LeMarquand, 2001). This would be expected if those who have high levels ofphysical aggression during childhood and adolescence were frequent users ofphysical aggression during early childhood. Some of the predictors point tocausal mechanisms, and others are markers of unclear mechanisms. For exam-ple, the sibling variable appears to indicate that young children who have atarget are much more likely to use physical aggression (Dunn & Munn, 1985).The smoking variable replicates the increasing number of longitudinal studiessuggesting that nicotine has a negative effect on fetal brain development(Brennan, 2003; Fergusson, Woodward, & Horwood, 1998; Wakschlag et al.,2002). Antisocial behavior of the mother during her adolescence could be amarker for many causal mechanisms (see Keenan & Shaw, 1994). First, itcould be a marker for some form of genetic transmission of a propensity toimpulsive and aggressive behavior (see Caspi et al., 2002; Suomi, Chapter 4,
Physical Aggression in Humans 97
FIGURE 5.4. Trajectories of physical aggression from 17 to 42 months of age. From Tremblayet al. (2004).

this volume; and Pihl & Benkelfat, Chapter 13, this volume). It could also be amarker for the quality of the environment the mother is creating for the childboth during the prenatal (e.g., quality of nutrition, stress) and postnatal (e.g.,maternal care) periods (Francis et al., 2003). Finally, it could be a marker forboth the genetic and environmental contribution of the father, inasmuch asthere is much assortative mating (e.g., Rowe & Farrington, 1997). Similarly,family income is probably a marker for many different causal mechanisms,some of which could be quality of nutrition during the prenatal and postnatalperiods The two predictors measured at 5 months of age (maternal coercivebehavior toward the child and family dysfunction) suggest that children whouse physical aggression more frequently than others during infancy are the vic-tims and witnesses of a disproportionately large amount of physical aggres-sions during their first year after birth. We need more detailed observationaland physiological studies of parent–child interactions in these families duringthe first 12–18 months to understand the mechanisms by which the adult be-haviors have an impact on the child’s propensity to use physical aggression.
Cross-fostering experiments like those done by Francis et al. (2003),Weaver et al. (2004), and Suomi and his colleagues (see Chapter 4, this vol-ume) would give better insights into causal mechanisms than studies that arelimited to observations. Obviously, it would be hard to do such studies withhumans. However, prevention experiments that aim to help high-risk familiesto give better care to their children during early childhood (e.g., Campbell &Ramey, 1994; Olds et al. 1998) could be used to understand the mechanismsthat prevent children from behaving as if physical aggression were the best op-tion for adjusting to their environments.
CONCLUSIONS
I can say in my own favour that I was as a boy humane, but Iowed this entirely to the instruction and example of my sisters. Idoubt indeed whether humanity is a natural or innate quality.
—DARWIN (1876, p. 11)
The first aim of this chapter was to summarize our present knowledge ofthe development of physical aggression from early childhood to adulthood.Unfortunately, none of the large longitudinal studies of humans followed fromearly childhood to adulthood seem to have collected repeated measures ofphysical aggression throughout the study. Thus, to describe the lifespan intra-individual changes in the physical aggression of humans, we had to put to-gether a patchwork of longitudinal studies that covered different age periodswith samples from different populations. Figure 5.5 is a rough representationof this patchwork. The physical aggression trajectories from adolescence toadulthood are modeled on the data from White, Bates, and Buyske (2001);those from childhood to adolescence are inspired by data from Broidy et al.
98 T H E D E V E L O P M E N T O F A G G R E S S I O N

(2003) and Lacourse et al. (2002); and the preschool to elementary school tra-jectories are adapted from Côté et al. (2004), and Tremblay et al. (2004).
The patchwork shows that the peak in frequency of physical aggressionsfor the large majority of humans is most probably between the end of the 2ndand 4th year after birth. This representation of physical aggression develop-ment has a shape similar to the traditional age–violent-crime curve. There is asharp increase in frequency, a peak that does not last very long, and a slow de-cline. Note that even those who can be considered on a chronic physical ag-gression trajectory are slowly declining in physical aggression, with a possiblebrief increase during adolescence. By adulthood the individual differences infrequency are possibly the smallest.
Although the shapes of the curves are similar, moving the peak in fre-quency of physical aggression from 18 months to 3 years of age should quali-tatively change the way we think about the second aim of this chapter, defin-ing the mechanisms involved in the development of physical aggression. St.Augustine (397/1960) can be paraphrased to express one of the obvious dif-ferences: Thank God (or nature!) that humans have weak limbs at the agewhen they most often use physical aggression. We believe that the observeddevelopmental trajectories seriously question the social learning hypothesis asit continues to be expressed, especially with reference to media violence (e.g.,Johnson et al., 2002). There is also no evidence that aggression is like an appe-tite that needs to be satisfied (Archer, 1988; Lorenz, 1966), but it appears veryunlikely that humans need to observe physical aggression to initiate its use. Ifthey do, the learning must be occurring in the first 15 months after birth, andthat learning would be universal. What appears more certain, from the ob-served developmental trajectories, is that humans learn not to use physical ag-
Physical Aggression in Humans 99
FIGURE 5.5. Age–physical aggression curves (hypothesized). From Tremblay (2003). Copy-right 2003 by The Guilford Press. Reprinted by permission.

gression by learning alternatives. Some children learn more quickly than oth-ers, and girls are overrepresented in this group (see Archer & Côté, Chapter20, this volume). However, if humans learn more or less rapidly to use alterna-tives to physical aggression, it is important to note that this behavior remainsin the behavioral repertoire and can be used when the need arises. Because wetend to forget this, there is always much surprise when a quiet, law-abidingcitizen suddenly physically aggresses a neighbor, a spouse, or a work mate.
Most investigators, and indeed, most humans, forget that physical aggres-sion is not in itself a “bad,” “destructive” behavior. It is a solution for secur-ing resources and surviving in a given environment (Archer, 1988). Humanshave inherited the ability to use that solution, but they have also inherited theability to use many other strategies, including cooperation and conflict resolu-tion, that are not based on physical force (see Peterson & Flanders, Chapter 7,this volume, and de Waal, 2000). We are slowly creating a social environmentin which the physical aggression solution generally becomes a much less adap-tive strategy than its alternatives. Young children will spontaneously cooper-ate and fight, but they need to learn which of the alternatives is the most effec-tive in which situation. Some, because of their genetic makeup, will moreeasily learn some solutions than others. Similarly, some environments willtend to facilitate some solutions more than others. The gene–environment in-teractions will favor different developmental trajectories. The simple law thatDarwin put forward was that those who will thrive are those who will bemost able to learn to use the solutions that are most appropriate for the prob-lems they face. In a social group that does not tolerate physical aggression, itis better to use a form of aggression that is less sanctioned. This does not meanthat a punch in the nose is more harmful than an indirect aggression to a per-son’s reputation. It simply means that a group of humans have decided thatconflicts should not be solved by physical force. There was a time, not longago, when conflicts between men of honor were solved by a duel using pistols,swords, or fists; in our 21st-century civilization there is an increasing trend tosettle conflicts with words.
The fact that physical aggression peaks in early childhood when limbs areweak, and when the brain is at its peak flexibility for learning, leads us to twoimportant conclusions concerning research and education during early child-hood. First, it is clear that we need to better understand the mechanisms thatunderlie the rapid increase in frequency of physical aggression in infancy, aswell as the mechanisms that lead to the large differences among infants. Itmust be noted that a similar conclusion was reached 30 years ago. Hartup andDe Wit (1974, p. 607) highlighted the fact that “the role of tantrums in the de-velopment of aggression can be inferred from existing studies only by piecingtogether bits and pieces of data. The origins of aggression in human childrencould be elucidated by several types of longitudinal studies (which might coverno more than 12 or 18 months).” In the same book, Hamburg and vanLawick-Goodall (1974, p. 72) also concluded: “There is a great need for di-rect studies, utilizing systematic observational techniques of aggressive behav-
100 T H E D E V E L O P M E N T O F A G G R E S S I O N

ior and its precursors in human infancy and early childhood.” Unfortunately,not much progress has been made on that front during the 30 years since.
Our second conclusion is that because early childhood appears to be theoptimal period to learn alternatives to the use of physical aggression, it couldbe the optimal time to link education and research. Over the past 4 decadesthere has been an important movement to enrich the educational experienceduring the preschool years. A number of randomized control experimentstested the effects of different types of preventive programs for children at riskof learning and behavior problems (Olds et al., 1999; Tremblay, LeMarquand,& Vitaro, 1999b). To our knowledge, none of these experiments included aspecific component for learning to regulate physical aggression. We believethat such experiments would not only help us learn to what extent we canchange the developmental course of physical aggression, but would also helptest hypotheses concerning the developmental origins of aggression.
We end this chapter with a brief allegoric answer to a question we askedat its beginning when we referred to the huge appetite for media violence inmodern societies. The question was: “By what peculiar process are thesehungry-for-physical-violence modern humans passing laws to eradicate physi-cal violence from their daily lives?” In the book of Genesis we learn thathumans were created to dominate planet Earth, and that soon after Adam andEve were chased from Paradise there was a homicide: Cain killed Abel.Whether we believe that story or its revision by Darwin, we are told that hu-mans have inherited a biological machine that has the capacity to physicallyaggress. As humans, we dream of the Paradise where we will not fear beingphysically aggressed; however, Paradise will be a boring place if the aggressionmachinery (perceptions, cognitive–emotional processing, and action) is left torust. Hence, the invention of the horror story, the violent video games, andcontact sports. The transformation of real physical violence into fiction orgame should help keep Paradise a safe place to live in, as long as we under-stand what we are doing and why we are doing it.
REFERENCES
American Heritage Dictionary. (1985). (2nd college ed.). Boston: Houghton Mifflin.Anderson, C. A., & Bushman, B. J. (2002). The effects of media violence on society. Sci-
ence, 295, 2377–2379.Archer, J. (1988). The behavioural biology of aggression. Cambridge, UK: Cambridge
University Press.Arseneault, L., Moffitt, T. E., Caspi, A., Taylor, A., Rijsdijk, F. V., Jaffee, S. R., Ablow, J.
C., & Measelle, J. R. (2003). Strong genetic effects on cross-situational antisocialbehaviour among 5-year-old children according to mothers, teachers, examiner–observers, and twins’ self-reports. Journal of Child Psychology and Psychiatry,44(6), 832–848.
Arseneault, L., Tremblay, R. E., Boulerice, B., & Saucier, J.-F. (2002). Obstetrical compli-
Physical Aggression in Humans 101

cations and violent delinquency: Testing two developmental pathways. Child De-velopment, 73(2), 496–508.
Blumstein, A., & Wallman, J. (Eds.). (2000). The crime drop in America. Cambridge, UK:Cambridge University Press.
Bovet, L. (1951). Psychiatric aspects of juvenile delinquency. Geneva: World Health Or-ganization.
Brennan, P. A. (2003). Tobacco consumption during pregnancy and its impact onpsychosocial child development. Centre of Excellence for Early Childhood Develop-ment Encyclopedia. http://www.excellence-earlychildhood.ca/theme.asp?id=1&lang=EN (March 27)
Brennan, P. A., Grekin, E. R., & Mednick, S. A. (1999). Maternal smoking and criminaloutcomes. Archives of General Psychiatry, 56, 215–219.
Bridges, K. M. B. (1931). The social and emotional development of the pre-school child.London: Kegan Paul.
Bridges, K. M. B. (1933). A study of social development in early infancy. Child Develop-ment, 4, 36–49.
Broidy, L. M., Nagin, D. S., Tremblay, R. E., Brame, B., Dodge, K., Fergusson, D.,Horwood, J., Loeber, R., Laird, R., Lynam, D., Moffitt, T., Bates, J. E., Pettit, G. S.,& Vitaro, F. (2003). Developmental trajectories of childhood disruptive behaviorsand adolescent delinquency: A six site, cross national study. Developmental Psy-chology, 39(2), 222–245.
Bureau of Justice Statistics. U.S. Department of Justice, Office of Justice Programs. 7 Au-gust 2003. http://www.ojp.usdoj.gov/bjs/
Bushman, B. J., & Anderson, C. A. (2001). Media violence and the American public: Sci-entific facts versus media misinformation. American Psychologist, 56(6–7), 477–489.
Cairns, R. B., & Cairns, B. D. (1994). Life lines and risks: Pathways of youth in our time.New York: Cambridge University Press.
Cairns, R. B., Cairns, B. D., Neckerman, H. J., Ferguson, L. L., & Gariépy, J. L. (1989).Growth and aggression: 1. Childhood to early adolescence. Developmental Psy-chology, 25(2), 320–330.
Campbell, F. A., & Ramey, C. T. (1994). Effects of early intervention on intellectual andacademic achievement: A follow-up study of children from low-income families.Child Development, 65(2), 684–698.
Caspi, A., McClay, J., Moffitt, T., Mill, J., Martin, J., Craig, I. W., Taylor, A., & Poulton,R. (2002). Role of genotype in the cycle of violence in maltreated children. Science,297, 851–854.
Côté, S., Vaillancourt, T., LeBlanc, J., Nagin, D. S., & Tremblay, R. E. (2004). The devel-opment of physical aggression during childhood: A nation wide longitudinal studyof Canadian children. Manuscript submitted for publication.
Darwin, C. (1832/1985). Letter to Caroline Darwin. In F. Burkhardt & S. Smith (Eds.),The correspondence of Charles Darwin (Vol. 1). London : Cambridge UniversityPress.
Darwin, C. (1872). The expression of the emotions in man and animals. London: JohnMurray, reprinted by Gryphon Editions 1989.
Darwin, C. (1876/1983). Autobiography. Oxford: Oxford University Press.De Waal, F. B. (2000). Primates: A natural heritage of conflict resolution. Science, 289,
586–590.Dionne, G., Tremblay, R. E., Boivin, M., Laplante, D., & Pérusse, D. (2003). Physical ag-
102 T H E D E V E L O P M E N T O F A G G R E S S I O N

gression and expressive vocabulary in 19-month-old twins. Developmental Psy-chology, 39(2), 261–273.
Dunn, J., & Munn, P. (1985). Becoming a family member: Family conflict and the devel-opment of social understanding in the second year. Child Development, 56, 480–492.
Eisner, M. (2003). Long-term historical trends in violent crime. In M. Tonry (Ed.), Crimeand justice: A review of research (Vol. 30, pp. 83–142). Chicago: University of Chi-cago Press.
Ellis, L., & Coontz, P. D. (1990). Androgens, brain functionning, and criminality: Theneurohormonal foundations of antisociality. In L. Ellis & H. Hoffman (Eds.), Crimein biological, social and moral contexts (pp. 162–193). New York: Praeger.
Eron, L. D. (1990). Understanding aggression. Bulletin of the International Society forResearch on Aggression, 12, 5–9.
Fergusson, D. M., Woodward, L. J., & Horwood, L. J. (1998). Maternal smoking duringpregnancy and psychiatric adjustment in late adolescence. Archives of General Psy-chiatry, 55(8), 721–727.
Francis, D. D., Diorio, J., Plotsky, P. M., & Meaney, M. J. (2002). Environmental enrich-ment reverses the effects of maternal separation on stress reactivity. Journal of Neu-roscience, 22, 7840–7843.
Francis, D. D., Szegda, K., Campbell, G., Martin, W. D., & Insel, T. (2003). Epigeneticsources of behavioral differences in mice. Nature Neuroscience, 6, 445–446.
Goodenough, F. L. (1931). Anger in young children. Westport, CT: Greenwood Press.Hamburg, D. A., & Van Lawick-Goodall, J. (1974). Factors facilitating development of
aggressive behavior in chimpanzees and humans. In J. de Wit & W. W. Hartup(Eds.), Origins of aggressive behavior (pp. 59–86). The Hague, Netherlands:Mounton.
Harris, J. A., Rushton, J. P., Hampson, E., & Jackson, D. N. (1996). Salivary testosteroneand self-report aggressive and pro-social personality characteristics in men andwomen. Aggressive Behavior, 22, 321–331.
Hartup, W. W., & DeWit, J. (1974). The development of aggression: Problems and per-spectives. In J. deWit & W. W. Hartup (Eds.), Determinants and origins of aggres-sive behavior (pp. 595–620). The Hague, Netherlands: Mouton.
Holmberg, M. C. (1977). The development of social interchange patterns from 12 to 42months: Cross-sectional and short-term longitudinal analyses [Ph.D. Thesis]. TheUniversity of North Carolina at Chapel Hill: Microfilms International 78–7137.
Huesmann, L. R., Eron, L. D., Lefkowitz, M. M., & Walder, L. O. (1984). Stability of ag-gression over time and generations. Developmental Psychology, 20(6), 1120–1134.
Johnson, J. G., Cohen, P., Smailes, E. M., Kasen, S., & Brook, J. S. (2002). Televisionviewing and aggressive behavior during adolescence and adulthood. Science, 295,2468–2471.
Keenan, K., & Shaw, D. S. (1994). The development of aggression in toddlers: A study oflow-income families. Journal of Abnormal Child Psychology, 22(1), 53–77.
Lacourse, E., Côté, S., Nagin, D. S., Vitaro, F., Brendgen, M., & Tremblay, R. E. (2002).A longitudinal-experimental approach to testing theories of antisocial behavior de-velopment. Development and Psychopathology, 14, 909–924.
Lahey, B. B., Goodman, S. H., Waldman, I. D., Bird, H., Canino, G., Jensen, P., Regier, D.A., Leaf, P. J., Gordon, R. A., & Applegate, B. (1999). Relation of age of onset to thetype and severity of child and adolescent conduct problems. Journal of AbnormalChild Psychology, 27(4), 247–260.
Physical Aggression in Humans 103

Lefkowitz, M. M., Eron, L. D., Walder, L. O., & Huesmann, L. R. (1977). Growing up tobe violent. A longitudinal study of the development of aggression. New York:Pergamon Press.
Loeber, R., & Dishion, T. J. (1983). Early predictors of male delinquency: A review. Psy-chological Bulletin, 94, 68–99.
Loeber, R., Green, S. M., Keenan, K., & Lahey, B. B. (1995). Which boys will fare worse?Early predictors of the onset of conduct disorder in a six-year longitudinal study.Journal of the American Academy of Child and Adolescent Psychiatry, 34(4), 499–509.
Lorenz, K. (1966). On aggression. New York: Harcourt, Brace and World.McCord, J., Widom, C. S., & Crowell, N. E. (2001). Juvenile crime, juvenile justice.
Washington, DC: National Academy Press.Nagin, D., & Tremblay, R. E. (1999). Trajectories of boys’ physical aggression, opposi-
tion, and hyperactivity on the path to physically violent and non violent juvenile de-linquency. Child Development, 70(5), 1181–1196.
Nagin, D., & Tremblay, R. E. (2001). Parental and early childhood predictors of persis-tent physical aggression in boys from kindergarten to high school. Archives of Gen-eral Psychiatry, 58, 389–394.
Nicolson, H. (1955). Good behavior. London: Constable.Olds, D., Henderson, C. R., Cole, R., Eckenrode, J., Kitzman, H., Luckey, D., Pettitt, L.,
Sidora, K., Morris, P., & Powers, J. (1998). Long-term effects of nurse home visita-tion on children’s criminal and antisocial behavior: Fifteen-year follow-up of a ran-domized controlled trial. Journal of the American Medical Association, 280(14),1238–1244.
Olds, D. L., Henderson, C. R., Jr., Kitzman, H., Eckenrode, J., Cole, R., & Tatelbaum, R.C. (1999). Prenatal and infancy home visitation by nurses: Recent findings. The Fu-ture of Children, 9(1), 44–65.
Premack, D., & Premack, A. (2003). Original intelligence. New York: McGraw-Hill.Quetelet, A. (1833). Research on the propensity for crime at different ages. Brussels: M.
Hayez.Raine, A. (2002). Annotation: The role of prefrontal deficits, low autonomic arousal,
and early health factors in the development of antisocial and aggressive behavior inchildren. Journal of Child Psychology and Psychiatry, 43, 417–434.
Raine, A., Brennan, P., & Mednick, S. A. (1997). Interaction between birth complicationsand early maternal rejection in predisposing individuals to adult violence: Specific-ity to serious, early-onset violence. American Journal of Psychiatry, 154, 1265–1271.
Reiss, A. J., & Roth, J. A. (Eds.). (1993). Understanding and preventing violence. Wash-ington, DC: National Academy Press.
Restoin, A., Montagner, H., Rodriguez, D., Girardot, J. J., Laurent, D., Kontar, F.,Ullmann, V., Casagrande, C., & Talpain, B. (1985). Chronologie des comporte-ments de communication et profils de comportement chez le jeune enfant [Chronol-ogy of communicative behavior and behavior profiles in early childhood]. In R. E.Tremblay, M. A. Provost, & F. F. Strayer (Eds.), Ethologie et développement del’enfant (pp. 93–130). Paris: Editions Stock/Laurence Pernoud.
Rousseau, J.-J. (1762/1979). Emile or On education. New York: Basic Books.Rowe, D. C., & Farrington, D. P. (1997). The familial transmission of criminal convic-
tions. Criminology, 35, 177–201.
104 T H E D E V E L O P M E N T O F A G G R E S S I O N

Sampson, R. J., & Laub, J. H. (2003). Life-course desisters? Trajectories of crime amongdelinquent boys followed to age 70. Criminology, 40, 555–592.
Sand, E. A. (1966). Contribution à l’étude du développement de l’enfant. Aspectsmédico-sociaux et psychologiques [Contribution to the study of child development.Medical-social and psychological effects]. Bruxelles: Éditions de l’Institut de Socio-logie de l’Université Libre de Bruxelles.
Slutske, W. S., Heath, A. C., Dunne, M. P., Statham, D. J., Dinwiddie, S. H., Madden, P.A. F., Martin, N. G., & Bucholz, K. K. (1997). Modeling genetic and environmentalinfluences in the etiology of conduct disorder: A study of 2,682 adult twin pairs.Journal of Abnormal Psychology, 106(2), 266–279.
St. Augustine. (397/1960). Confessions. New York: Doubleday.Steinberg, L., & Scott, E. S. (2003). Less guilty by reason of adolescence: Developmental
immaturity, diminished responsibility, and the juvenile death penalty. AmericanPsychologist, 58, 1009–1018.
Tremblay, R. E. (2003). Why socialization fails: The case of chronic physical aggression.In B. B. Lahey, T. E. Moffitt, & A. Caspi (Eds.), Causes of conduct disorder and juve-nile delinquency (pp. 182–224). New York: Guilford Press.
Tremblay, R. E. (2004). The development of human physical aggression: How importantis early childhood? In L. A. Leavitt & D. M. B. Hall (Eds.), Social and moral devel-opment: Emerging evidence on the toddler years (pp. 221–238). New Brunswick,NJ: Johnson and Johnson Pediatric Institute.
Tremblay, R. E., Boulerice, B., Harden, P. W., McDuff, P., Pérusse, D., Pihl, R. O., &Zoccolillo, M. (1996). Do children in Canada become more aggressive as they ap-proach adolescence? In Human Resources Development Canada & Statistics Can-ada (Eds.), Growing up in Canada: National Longitudinal Survey of Children andYouth (pp. 127–137). Ottawa: Statistics Canada.
Tremblay, R. E., Japel, C., Pérusse, D., McDuff, P., Boivin, M., Zoccolillo, M., &Montplaisir, J. (1999a). The search for the age of “onset” of physical aggression:Rousseau and Bandura revisited. Criminal Behavior and Mental Health, 9, 8–23.
Tremblay, R. E., & LeMarquand, D. (2001). Individual risk and protective factors. In R.Loeber & D. Farrington (Eds.), Child delinquents: Development, interventions andservice needs (pp. 137–164). Thousand Oaks, CA: Sage.
Tremblay, R. E., LeMarquand, D., & Vitaro, F. (1999b). The prevention of ODD andCD. In H. C. Quay & A. E. Hogan (Eds.), Handbook of disruptive behavior disor-ders (pp. 525–555). New York: Kluwer Academic/Plenum.
Tremblay, R. E., Nagin, D. S., Séguin, J. R., Zoccolillo, M., Zelazo, P., Boivin, M.,Pérusse, D., & Japel, C. (2004). Physical aggression during early childhood: Trajec-tories and predictors. Pediatrics, 114(1), e43–e50.
Tremblay, R. E., Pihl, R. O., Vitaro, F., & Dobkin, P. L. (1994). Predicting early onset ofmale antisocial behavior from preschool behavior. Archives of General Psychiatry,51, 732–738.
Tremblay, R. E., Schaal, B., Boulerice, B., Arseneault, L., Soussignan, R., & Pérusse, D.(1997). Male physical aggression, social dominance and testosterone levels at pu-berty: A developmental perspective. In A. Raine, P. Brennan, D. P. Farrington, & S.A. Mednick (Eds.), Biosocial bases of violence (pp. 271–291). New York: PlenumPress.
U.S. Human Capital Initiative. (1997, November). Reducing violence: A research agenda[Special issue]. APS Observer.
Wakschlag, L., Pickett, K. E., Cook, E., Benowitz, N. L., & Leventhal, B. (2002). Mater-
Physical Aggression in Humans 105

nal smoking during pregnancy and severe antisocial behavior in offspring: A review.American Journal of Public Health, 92(6), 966–974.
Weaver, I. C. G., Cervoni, N., Champagne, F. A., D’Alessio, A. C., Sharma, S., Sheckl, J.R., Dymov, S., Szyf, M., & Meaney, M. J. (2004). Epigenetic programming by ma-ternal behavior. Nature Neuroscience, doi: 10.1038/nn1276.
White, H. R., Bates, M. E., & Buyske, S. (2001). Adolescence-limited versus persistentdelinquency: Extending Moffitt’s hypothesis into adulthood. Journal of AbnormalPsychology, 110(4), 600–609.
World Health Organization. (2002). World report on violence and health. Geneva: Au-thor.
106 T H E D E V E L O P M E N T O F A G G R E S S I O N

THE DEVELOPMENT OF AGGRESSIONThe Beginnings of Aggression in Infancy
6
The Beginnings of Aggressionin Infancy
DALE F. HAY
Aggression is a ubiquitous feature of human social life, which makes it allthe more surprising that its developmental course is not more clearly known(see Tremblay, 2000). The aim of this chapter is to trace the earliest begin-nings of the human capacity for aggression, as manifested in the first 3years of life. Two general questions are considered, reflecting the fact thatany developmental analysis requires attention to both change and continuityover time (see Appelbaum & McCall, 1983). First, when do infants in gen-eral reveal the ability to direct aggression against their companions? Second,do some infants show stable and coherent tendencies to be more aggressivethan their peers?
To address either question means getting to grips with the thorniest prob-lem in studying aggression, namely, the question of intent. Determining theintent of any action becomes particularly difficult when observing preverbalinfants, who cannot be questioned about their intentions and whose motivesare usually given the benefit of the doubt. I suggest that the intentionality ofsocial action is itself a developmental phenomenon, and therefore the inten-tional nature of early aggression becomes an empirical question. I return tothis issue later in the chapter. First, I consider some definitional issues, pro-pose operational definitions of the use of force against one’s companions, andnote when behaviors meeting these criteria can first be identified and howthey change over time.
107

DEFINITIONAL ISSUES
Defining Aggression
Hardly any construct in psychology is more difficult to define than aggression(see Cairns, 1979). Broadly speaking, the term aggression implies harm donewith malevolent intent; physical harm designed to do good (e.g., surgical pro-cedures) and unintentional assault (e.g., elbowing someone in the face by acci-dent) do not qualify. Even when assaults are intentional, they may be deemedmore or less serious on the basis of whether they are reactive rather thanproactive and decided upon in the heat of the moment or in a cool, premedi-tated fashion—hence the legal distinctions among self-defense, manslaughter,and murder.
The concept of harm is itself open to varying interpretations: Aggressionis often synonymous with physical assault, though it need not be. Intentionallydelivered malicious comments about another person’s appearance, actions, orcharacter could so qualify; yet, with notable exceptions (e.g., Muste &Sharpe, 1947), there have been few attempts to explore the early developmentof verbal aggression. It can also be argued that the failure to act in a friendly,sociable manner can, in some cases, qualify as intentional harm, as in the phe-nomenon of “relational aggression” in childhood (e.g., Crick, 1995). Thus, ingeneral, definitions of aggression are multidimensional, requiring informationabout the specific intent of an action, its form and intensity, its provocation,and the interpersonal history of aggressor and victim. It is hardly surprisingthat such a quicksilver construct is hard to study. How, then, can one proceedto study the developmental origins of something so hard to define?
Although human infants may not usually be described as aggressive,some are thought to be “difficult.” Infants show anger (Bridges, 1932;Goodenough, 1931; Lewis, Alessandri, & Sullivan, 1990; Stenberg, Campos,& Emde, 1983). Older toddlers may be “jealous” and “rivalrous” with theiryounger siblings (Levy, 1939). All of these constructs suggest a potentiallyfraught relationship between some infants and their companions; infants’worlds are clearly not free of conflict. I propose that to seek the origins of ag-gression, we need to look within infants’ conflicts with adults and with otherchildren. Social conflict can be operationally defined as a social interaction inwhich there is opposition, with one person’s action being protested, resisted,or retaliated against by another person (Hay & Ross, 1982). This definition isat a sufficient level of abstraction that it can be applied across ages and acrossdifferent sorts of personal relationships.
By using conflict as a basic framework, one can identify and chart the de-velopmental course of various phenomena of relevance to aggression through-out the life course. These include (1) emotional expressions of anger and theircontrol, (2) resistance to the actions of others, in terms of gaze avoidance orwithdrawal from interaction, (3) vocal and, eventually, verbal protest of theactions of others, (4) use of physical force against another person, (5) defen-
108 T H E D E V E L O P M E N T O F A G G R E S S I O N

sive reactions, and (6) name calling and other forms of verbal insult. One canalso chart infants’ use of prosocial actions to resolve their disputes, and studythe development of related phenomena such as moral condemnation of an-other’s actions and justification of one’s own behavior. By using conflict as thebasic framework for investigating the origins of aggression, infants’ expres-sions of anger and use of force can be examined in social context, and thegrowing intentionality of forceful actions can be studied as a developmentalphenomenon in its own right. Furthermore, some additional distinctions canbe made.
Personally Directed versus Instrumental Use of Force
Two characteristic means of directing physical force against one’s companionscan be distinguished in early life. The first can be described as personally di-rected force (e.g., hitting, kicking, pushing, batting at, or otherwise directingone’s body against another’s in a forceful manner). The second is instrumentalforce (e.g., snatching, grabbing, or otherwise wresting an object away fromanother person into one’s own possession). These categories roughly corre-spond to Hartup’s (1974) classic distinction between hostile and instrumentalaggression.
This basic distinction between force used as an end in itself versus ameans to gain access to an object, position, or territory is present in most ob-servational coding systems that have been applied to young children’s socialinteractions (e.g., Bridges, 1933; Eckerman, Whatley, & Kutz, 1975; Hay &Ross, 1982; Rubin, Hastings, Chen, Stewart, & McNichol, 1998; Strayer &Strayer, 1976). Although these two classes of behavior are often combinedinto a single category, they appear to have somewhat different developmentalcourses and sets of correlates, and so I consider them separately in this chap-ter.
Proactive versus Reactive Use of Force
Several investigators have distinguished between aggression used proactivelyand reactively, and, as we have seen, the extent to which aggression is premed-itated or used in self-defense is also an important legal distinction. Longitudi-nal research has suggested that proactive aggression in early childhood is aserious predictor of later violence and delinquency (Vitaro, Gendreau, Tremblay,& Oligny, 1998). It should be noted, however, that many aggressive individu-als use both (Vitaro, Brendgen, & Tremblay, 2002). Furthermore, angry, reac-tive aggression is linked to social–cognitive problems (Schwartz et al., 1998)and is responded to negatively by peers, which fosters further aggression(Price & Dodge, 1989). It is important to assess the extent to which veryyoung children use force proactively and reactively and whether there are con-sistent individual differences in this regard.
The Beginnings of Aggression in Infancy 109

The Recipients of the Young Child’s Acts of Force
Different investigators tend to study infants’ interactions with parents, sib-lings, peers, and other adults, and so it is rare to find studies that compare theuse of force with different types of companions (for exceptions to this generaltrend, see Smetana, 1984, 1989). Although we may assume that infants directforce against their caregivers when in the midst of a temper tantrum, or disci-plinary encounter, such events seem rarely to have been recorded and are notusually defined as instances of aggression. Rather, most accounts of the earlyorigins of aggression, and its developmental course, focus on children’s use offorce in conflicts with other children, that is, their siblings, friends, acquain-tances, and newly met peers. However, the initial steps in the development ofaggression are probably taken in the context of interactions with adults, inparticular with primary caregivers. I thus include these early features ofparent–infant relations in a proposed model of the normative development ofaggression.
THE NORMATIVE COURSE OF CONFLICT AND AGGRESSION
A review of the existing literature on infants’ interactions with adults, siblings,and peers suggests the following model of the normative development of ag-gression over the first 3 years of life.
The Capacity for Anger
A developmental account of the origins of aggression begins with the study ofanger (Goodenough, 1931). Recognizable signs of specific emotions emerge inthe 1st year of life. Frustrating experiences, such as an adult’s yanking acookie away from the baby (Stenberg et al., 1983), or extinction of a previ-ously rewarded behavior (Lewis et al., 1990), provoke these expressions.
It is, of course, not completely clear whether the facial expressions re-corded in these studies are mirrored by the phenomenological experience ofanger as felt by adults. Rather little is known about the manifestations of an-ger in nonexperimental situations and still less about its rate of occurrence inordinary interactions with parents and other people. Measures of infant tem-perament do not rate anger per se, although Rothbart’s (1981) Infant Behav-iour Questionnaire measures “distress to limitations,” which predicts ratingsof “anger proneness” in the toddler years (Goldsmith, 1996). Furthermore,Goldsmith (1996, pp. 225–226) argued that “anger expression appears to bethe emotional core of difficulty” as rated by other temperament instruments.In Goldsmith’s (1996) analysis of 11 samples of toddlers, anger-proneness isstable and coherent, but somewhat bound up with activity level. Much workis needed to chart the normative development of anger and its regulation inthe early years.
110 T H E D E V E L O P M E N T O F A G G R E S S I O N

Resistance and Protest
I have defined social conflict as an encounter in which one person’s actionmeets with an objection from another person, which takes the form of protest,resistance, or retaliation (Hay & Ross, 1982). Early signs of resistance andprotest predate the use of retaliation in development. A simple form of con-flict occurs when infants avert their gaze in response to unduly intense or pro-longed overtures from adults. Young infants also show objections to the ac-tions of other people by wriggling, leaning away, pushing away a caregiver’shands or face, and, eventually, moving away from the other person. Distressthat is contingent upon another person’s actions is similar to an early form ofprotest that defines a conflict.
In the early months of life infants seem remarkably forbearing vis-à-vistheir companions. For example, 6-month-old infants touch each other andtouch toys they each are holding; these events, which provoke conflict be-tween older infants (e.g., Caplan, Vespo, Pedersen, & Hay, 1991), do not leadto visible signs of protest or resistance (Hay, Nash, & Pedersen, 1983). How-ever, gaze avoidance, distress, and withdrawal from interaction are likely tooccur in response to intrusive or very withdrawn maternal behavior, such asthat shown by women suffering from postnatal depression (e.g., Field et al.,1988) or mothers who are merely simulating depressed affect in a laboratoryparadigm (Cohn & Tronick, 1983). This form of resistance to intrusive or in-consistent caregiving may be the genesis of insecure attachment relationships(Ainsworth, Blehar, Waters, & Wall, 1978; De Wolff & van IJenzdoorn,1997). Less is known about the normative use of resistance in infants’ socialrelationships with adults and other children.
Autonomy, Initiative, and Active Defense
Studies of infants’ more positive social interaction—their engagement in con-ventional social games—have demonstrated that infants become increasinglyactive and creative in the second half of the first year of life (Gustafson,Green, & West, 1979; Ross & Kay, 1980). For example, in the well-studiedgame of “peek-a-boo,” the infant’s role is first simply to express affect—tolaugh when its face is covered with a cloth. Next, at a slightly older age, theinfant begins actively to pull off the cloth; then to put the cloth over his or herown face; and finally to cover up the adult’s face with the cloth. I propose thatan analogous progression from passive resistance to active defense and the in-tentional use of force takes place in the context of infants’ conflicts with otherpeople. This development is likely to take place between 9 and 12 months ofage, when infants begin to move about the world autonomously. Relation-ships between infants and their parents become more problematic when theinfants learn to walk, particularly when boys walk early (Biringen, Emde,Campos & Appelbaum, 1995).
An early scholar suggested that infants’ actions in conflict indeed become
The Beginnings of Aggression in Infancy 111

more active around 11 or 12 months of age. It is fitting that the conference pa-per on which this chapter is based was presented in Montreal, where, in the1920s, one of the earliest and most systematic studies of infants’ peer relationswas conducted (Bridges, 1932, 1933, 1936). Bridges observed several groupsof physically healthy infants and toddlers residing at two foundling hospitalsin Montreal. (She noted that the prospects for cognitive and emotional devel-opment might be restricted, because of the institutional upbringing of thesechildren, but also suggested that in view of the amount of time they spentplaying with peers, their social development might be precocious.) Bridgeswrote that the 10-month-old infant seemed “rather indifferent when anotherchild in the [playpen] grasps his toy. He takes hold of some other object or sitsand rocks. He scarcely struggles at all for the possession of anything.” In con-trast, a month later, “If another child takes it from him he cries. In a fewweeks’ time he makes some attempt to hold on to his possessions; he strugglesand squeaks. He also squeals if another baby takes hold of some object hecoveted.”
The 1-year-old’s more active engagement in conflict is also facilitated byimprovements in cognitive abilities. Particular strides are made in infants’ un-derstanding of means–end relationships, with consequences for their apprecia-tion of the intentionality of action (e.g., Frye, 1993). Furthermore, in this lastquarter of the first year of life infants’ reactions to potentially disturbingevents become conditional. They interpret ambiguous events in terms of care-givers’ expressed emotion (Feinman, 1982) and their own degree of controlover the situation (Rheingold & Eckerman, 1970; Gunnar, 1980). It is in thiscontext of increased capacities for interpretation and understanding of thethings that happen to them that infants can be said to show focused protestand self-defense, not just undifferentiated distress. Unlike younger infants(Hay et al., 1983), 12-month-old infants do notice when their hair is pulled ortheir toys are taken away by their peers, and react accordingly (Caplan et al.,1991).
The Intentional and Strategic Use of Force
Force against Family Members
We know relatively little about infants’ use of force against their parents. Forexample, in one study of small boys in Pennsylvania, the boys’ defiance wassubsumed into general measures of “family trouble” and the developmentalcourse of the boys’ aggression against parents was not charted in its own right(Belsky, Woodworth, & Crnic, 1996). The means suggest, however, that defi-ant behavior on the part of the boys (which might include the use of force)was shown in approximately one-fifth of the conflicts recorded between par-ents and children.
In an observational study of 2-year-olds with their mothers and elder sib-lings in a laboratory apartment, family conflict was common but the use of
112 T H E D E V E L O P M E N T O F A G G R E S S I O N

force was not (Hay, Vespo, & Zahn-Waxler, 1998). However, disputes thatentail aggression were observed in home settings when toddlers interactedwith mothers as well as with familiar peers (Smetana, 1989).
Force against Peers
Bridges (1933) suggested, based on her observations of children in foundlinghospitals, that the active use of personal force, in the form of biting or hitting,was in place by 14 or 15 months of age. Despite her belief that institutional-ized children might be socially precocious, this is probably a conservative esti-mate of the age of onset. Both striking others and taking of toys have been re-corded, at low levels, in infants between 10 and 12 months (Eckerman et al.,1975).
Does the Use of Force Decline with Age?
Given that the use of force emerges around the time of the first birthday, isthere then a steady decline in its use thereafter? When force begins to be usedin infants’ conflicts with peers, it occurs at quite low rates. Thereafter, the useof force over the next 2 years appears to ebb and flow, rather than showing aclear decline. For example, in our experimental study of previously unac-quainted children, observed in a laboratory playroom with their mothers, ingroups of three mother–infant pairs, both personally directed and instrumen-tal force occurred at 12 months of age (Caplan et al., 1991). There was no sig-nificant difference in the mean rate of force between the 12- and 24-month-old children observed in that study, nor did the rate of conflict with peers dif-fer between the two age groups.
It is somewhat difficult to weigh the evidence for a general decline in ag-gression, as different investigators have used slightly different definitions, dif-ferent units of analysis, time- versus event-sampling strategies, and specific ex-perimental procedures. However, our own program of work suggests that,rather than showing a monotonic decline in the use of force from the 1st tothe third birthdays, there appear to be peaks around 1½ and 2½–3 years ofage. For this chapter, I examined cross-sectional data from several studies mycolleagues and I conducted in Canada, the United States, and the United King-dom, in which the occurrence of instrumental and personally directed forceagainst peers was recorded, using equivalent operational definitions (Caplanet al., 1991; Ross & Hay, 1977; Ishikawa, 2003; Pedersen, 1985).
In these experiments children were tested in dyads or triads for varyingperiods of time. To adjust for differences in experimental procedures, I calcu-lated the rate of forceful acts per hour, divided by the number of individualsobserved (see Ishikawa, 2003, for an extended justification of this way of ad-justing for group size). The results are presented in Figure 6.1. For clarity ofpresentation, the data are presented as a line graph, although the differentdata points represent separate studies. These studies show an initial peak in
The Beginnings of Aggression in Infancy 113

both personal and instrumental aggression around 18 months of age, and thenanother around the 3rd birthday.
We sought further evidence of change in the rate of force by undertakinglongitudinal observations of British children at home with familiar peers (Hay,Castle, & Davies, 2000). The toddlers were observed initially at 18, 24, or 36months of life and then again 6 months later. Thus, two samples were ob-served at 24 and 30 months of life, but only one sample each at 18 and 36months. Instrumental force was reliably more common than hitting, kicking,or biting; many toddlers in the sample never hit their peers. When the pairs ofchildren were observed again 6 months later, the rate of grabbing toys had de-clined and was now significantly less frequent than spontaneous sharing oftoys and other objects. In contrast, personally directed force remained at sta-ble, low rates, shown by a minority of the children (see Figure 6.2). The pat-terns of change revealed in the figure are broadly comparable to those foundin the cross-sectional observations, with peaks in the use of force at 18 and 30months of life. It is of interest to note that a cross-sectional study of infantsand toddlers attending day care centers similarly found peak levels of “asser-tive interchanges” (combining instrumental force, personal force, and negativecommands) at 18 and 30 months of age (Holmberg, 1980); on average, thetoddlers made 11 such initiations per hour at 18 months and 7.6 per hour at30 months.
Both the cross-sectional and the longitudinal comparisons show thatforce is not the “default” strategy that toddlers use in interacting with theirpeers. Rather, force against peers is used selectively and strategically. For ex-ample, laboratory observations of Canadian 21-month-olds showed that tod-dlers hardly ever began their conflicts with the use of force; the tendency touse force increased the longer a conflict remained unresolved (Hay & Ross,
114 T H E D E V E L O P M E N T O F A G G R E S S I O N
FIGURE 6.1. Cross-sectional data from a series of laboratory studies of interaction betweenunfamiliar peers showed a peak in both instrumental and personal use of force at 21months.

1982). Two decades later, a new generation of Canadian toddlers showed asimilar reluctance to use force, the majority doing so only once, if at all (Ru-bin, Burgess, Dwyer, & Hastings, 2003). Most conflict initiations are not ex-plicitly aggressive (Green, 1933; Rubin et al., 1998). Furthermore, toddlersdemonstrate an implicit awareness of “social rules” governing the use offorce; they are less likely to resist instrumental force if the child trying to graba toy had possessed it previously (Bakeman & Brownlee, 1982). Thus, ratherthan being a very common state of engagement between young peers, forceappears to be used on selected occasions.
Attributions about Companions’ Desires and Intentions
In the 2nd and 3rd years of life, toddlers become increasingly aware of otherpeople’s feelings and possible actions. In later childhood, aggression and con-duct problems have been linked to social–cognitive deficits and biases (e.g.,Dodge & Coie, 1987; Hughes, White, Sharpen, & Dunn, 2000; for an alter-native view, see Sutton, Smith, & Swettenham, 1999).
The normative understanding of other people’s desires and intentions isthought to develop over the course of the preschool years, with performancein early primary school approaching adult norms (Schult, 2002; Wellman,Phillips, & Rodriguez, 2000). However, our observational studies of children’sconflicts with peers indicate that a rudimentary understanding of intent ispresent by the 2nd year of life. In our longitudinal study of toddlers observedat home with familiar peers, when one child gestured toward a toy, the peerattempted to defend the object on more than one-third (38%) of the occasions(Hay et al., 2000). In contrast, sharing in response to peers’ gestures occurredon 8% of occasions (Hay, Castle, Davies, Demetriou, & Stimson, 1999a). Inboth cases, the toddlers were demonstrating their awareness of the peer’s de-
The Beginnings of Aggression in Infancy 115
FIGURE 6.2. Longitudinal analyses of a sample of British toddlers observed with familiar peersshowed peaks in both instrumental and personal use of force at 18 and 30 months.

sire for the object. This growing understanding of other people’s desires andintentions is likely to affect the circumstances in which toddlers choose to useforce.
Conflict Avoidance and Resolution
Toddlers also develop ways of avoiding conflict or terminating it, once begun.Clearly children do not always protest the actions of their companions.Pedersen (1985) recorded a variety of possible “provocations” in the behaviorof 2-year-olds, tested in dyads with familiar peers from a nursery school class-room. Although a great variety of possible provocations were recorded, only aminority of these possibly provoking actions led to conflict and overt aggres-sion was very rare indeed. Similar findings emerged in a study of 2-year-oldswith unfamiliar peers, where behaviors that might initiate conflict often failedto do so (Rubin et al., 1998).
Furthermore, even when one child protests the action of a sibling or peer,the first actor may simply let it go, and the conflict does not proceed beyondtwo initial moves (Hay & Ross, 1982; Hay et al., 1998). Even in longer con-flicts, one of the antagonists may simply walk away. For example, after a sus-tained conflict between two 21-month-olds, which involved the use of force,one child ended the dispute by wriggling out from under his antagonist, goingover to a mirror and trying on a hat (Hay & Ross, 1982). Toddlers sometimesuse explicit prosocial actions to end their conflicts with peers (Caplan et al.,1991; Sackin & Thelen, 1984), but yielding to the antagonist and leaving thesituation are rather more common ways of ending disputes.
The extent to which a toddler will yield to a peer depends on the identityof that peer. In an experimental study of the effects of acquaintance on peerinteraction, pairs of toddlers met on 3 consecutive days; on the 4th day, halfthe pairs were rearranged so that toddlers met unfamiliar peers (Hay & Ross,1982). The probability of initiating conflict on the 4th day was predictablefrom the toddlers’ prior behavior for both groups, but the probability of yield-ing to the peer’s demands was predictable only if the toddlers remained withtheir former partners. Such patterns of yielding to particular peers consolidateinto structured dominance relationships in peer groups (Bakeman & Brownlee,1982; Strayer & Strayer, 1976).
Very young children’s avoidance or termination of conflicts with theirparents has traditionally been studied in a different intellectual framework,namely, the study of compliance, self-regulation and conscience (e.g., Koch-anska, 1993; Kochanska, Akson, & Koenig, 1995a; Kopp, 1982; Kuczynski& Kochanska, 1990). Prompt and cheerful compliance to a parent’s commandor prohibition would constitute conflict avoidance (e.g., Kochanska et al.,1995a; Rheingold, Cook, & Kolowitz, 1987); in contrast, resistance, pro-longed protest, or defiance would produce conflict, which would then have tobe resolved by either the parent or the child. Conflicts between parents andtoddlers, which are variously described as “child behavior management
116 T H E D E V E L O P M E N T O F A G G R E S S I O N

events” (Belsky et al., 1996) or “testings of will” (Biringen et al., 1995), in-crease in frequency between 9 and 12 months of age (Biringen et al., 1995).During the first 3 years, mothers are increasingly likely to characterize theirchildren in negative terms (Stimson, Castle, & Hay, 1997). A substantial num-ber of “moral encounters” between mothers and children do involve thechild’s use of aggression (Smetana, 1989).
Within the family circle there are rather distinct features of sibling con-flicts as opposed to parent–child conflicts. Positive sibling relationships some-times follow from fraught relations between the mother and the firstborn(Dunn, 1993). If elder children’s actions are often objected to by their moth-ers, they are less likely than other children to make objections to behaviorshown by their younger siblings, and thus avoid sibling conflict (Hay et al.,1998). Much more attention needs to be paid to the emergence of conflictavoidance and resolution strategies, and the control of aggression, across thechild’s different relationships.
Emergence of a Moral Vision
By the end of the preschool years, children have firm opinions about aggres-sion and talk about aggression in a moral context. Five-year-olds condemn ag-gression, especially if it is proactive and involves personal assault (Ferguson &Rule, 1988; Hay, Zahn-Waxler, Cummings, & Iannotti, 1992). This capacityfor moral judgment is fostered in the context of children’s conflicts with par-ents, siblings, and peers (Dunn, 1988; Ross, Tesla, Kenyon, & Lollis, 1990;Smetana, 1989). Particularly in dealing with their siblings, 2- and 3-year-oldchildren begin to justify their own actions and cast aspersions on other peo-ple’s motives (Dunn, 1988). Parents’ moral teachings take place under theseemotionally fraught circumstances (Ross et al., 1990), and parental interven-tion in their children’s quarrels reduces the use of physical aggression(Perlman & Ross, 1997).
Direct moral education takes place against a background of toddlers’own increasing concerns with order and social arrangements. Over the courseof the 2nd year of life, children begin to show an awareness of “standards” insocial and aesthetic life (Kagan, 1981). For example, they express distaste formess and broken objects (Kochanska, Casey, & Fukomoto, 1995b) and be-come distressed when they create mess or break things (Cole, Barrett, &Zahn-Waxler, 1992; Kochanska et al., 1995b). In the early years children alsobecome increasingly aware of the concept of personal property. In the contextof sibling quarrels, children show an understanding of principles of entitle-ment that come to govern their designs on other people’s possessions (Ross,1996). Rudimentary understanding of the rights of possessors can be dis-cerned in 1-year-olds’ interactions with peers (Bakeman & Brownlee, 1982).
The capacity for moral evaluation also emerges in the toddler years. Ourlongitudinal observations of children with familiar peers indicated that even2-year-olds condemn each other’s actions as “naughty” or “bad” (Hay et al.,
The Beginnings of Aggression in Infancy 117

2000). In the context of this general concern for order, a child’s own aggres-sive actions require justification, and the aggressive actions of others may pro-voke fear and dislike (Denham, McKinley, Couchoud, & Holt, 1990).
Summary of the Developmental Trends
The foregoing section set out a developmental progression from unfocused ac-tivity, reactivity and interactivity through the development of the capacity foranger, focused protest and conflict, with the deployment of instrumental andpersonal force, to an eventual weighing of the justifications for aggression andthe use of alternative strategies to avoid or resolve conflict. The infant’s gen-eral approach to conflict moves from general irritation and passive resistanceto a capacity for focused protest and explicit use of force. However, aggres-sion never seems to occur at very high rates, even among notoriously “terri-ble” 2-year-olds. I have proposed that this general developmental model willhold across all the different interpersonal relationships in which infants andtoddlers engage. However, to my knowledge, there are no available longitudi-nal data on which to test that assumption.
EVIDENCE FOR INDIVIDUAL DIFFERENCES IN TODDLERS’ USE OF FORCE
Individual differences presumably will manifest themselves at each step in thedevelopmental progression I have described. Of primary interest, however, areindividual differences in the use of force. I now examine the evidence for sta-ble and coherent individual differences in young children’s use of force andbriefly speculate about possible factors that might underlie these differences,placing particular infants at risk for stable violent careers. First, however, onemust briefly consider the vexing question of gender differences.
Most studies of infants and toddlers reveal few gender differences in ag-gression or externalizing problems (for reviews, see Keenan & Shaw, 1997;Loeber & Hay, 1997). In the observational literature there are few strikingdifferences in the use of force by girls and boys. In some studies aggression oc-curs so infrequently that tests for sex differences are impossible (e.g., Brooks-Gunn & Lewis, 1979; Castell, 1970; Vespo, Pedersen, & Hay, 1995); in othercases, no significant sex differences are observed (e.g., Bakeman & Brownlee,1982).
The absence of significant sex differences in much observational researchmay partly be due to lack of statistical power. In a somewhat larger study of104 2-year-olds (Rubin et al., 1998), girls had lower scores on a compositemeasure of observed aggression. However, the girls and boys did not differsignificantly on “angry aggression,” which comprised physical aggression (hit-ting, pushing, punching, and kicking), verbal threats and insults, and angry af-fect. Nor were there gender differences in adult-rated externalizing problems.
Where gender appears to make the most difference is not in the absolute
118 T H E D E V E L O P M E N T O F A G G R E S S I O N

rate of force, but in qualitative as well as quantitative patterns of individualdifferences. Thus, with respect to the following evidence for stable and coher-ent individual differences in the early development of aggression, a number offindings are qualified by gender.
A Forceful Minority?
It is generally thought that the use of force is so common in toddlers’ interac-tions with other children that individual differences would be difficult to dis-cern. This is not true. Although the capacity for forceful action is present inthe second year of life, not all toddlers display the use of force during observa-tion sessions. Indeed, most toddlers are observed to use personal force seldomif at all, and in many cases it occurs too infrequently to be analyzed (e.g.,Brooks-Gunn & Lewis, 1979; Castell, 1970; Rubin et al., 1998; Vespo et al.,1995).
In our experimental studies the use of force is less common than otherpeer-directed behaviors. For example, over the course of four days in a labora-tory playroom, only 35% of the sample engaged in conflicts where force wasused (Hay & Ross, 1982). Similar findings emerged over a year of observa-tions of experimental play groups, where, on average across trimesters, 40%of the toddlers used force instrumentally and 20% showed “belligerent as-saults” (Bronson, 1981). A higher number of British toddlers used force athome with familiar peers (Hay et al., 2000). Most of those toddlers grabbedobjects held by peers (80% at the first visit and 77% six months later). How-ever, the majority refrained from the use of personally directed force, whichwas shown by 48% at the first visit and 44% 6 months later.
Thus, it appears that fewer children use personal than instrumental force.Particular forms of force are even less common. For example, only a minorityof boys who hit their peers used “hard hits” (Brownlee & Bakeman, 1981).The proactive use of physical force is relatively uncommon. In our longitudi-nal study, only a minority of children began an interaction by proactivelystriking the peer (27% at each visit).
Consistency across Different Measures of Aggression
There appears to be moderate consistency across different measures of aggres-sion in very early childhood, to the extent that composite measures show ac-ceptable levels of internal consistency (e.g., Fagot & Leve, 1998; NICHDEarly Childcare Research Network, 2001; Rubin et al., 1998, 2003). Tod-dlers’ observed use of force correlates significantly though modestly withmore general measures of externalizing problems (e.g., Rubin et al., 1998).
In our longitudinal study of toddlers observed at home with familiarpeers, the association between instrumental and personally directed acts offorce was positive, but only approached statistical significance. Further analy-ses showed that the association between the two types of forceful actions was
The Beginnings of Aggression in Infancy 119

significant for girls, but not for boys (Hay et al., 2000). Indeed, at the firstvisit, boys’ uses of instrumental and personal force were completely uncor-related. Mothers’ ratings of aggressiveness were associated with girls’ use ofpersonal force at both visits, and with boys’ instrumental force at the time ofthe second visit. Thus, girls’ aggressive tendencies were more consistent acrosstypes of behaviors and types of measurement.
Continuity of Individual Differences in the Use of Force over Time
To the extent that early aggression is seen as random, or trivial, or due merelyto situational factors, we would not expect any degree of stability of individ-ual differences over time. However, a number of studies have documented sta-tistically significant continuities over time in phenomena related to aggression.For example, toddlers’ tendencies to initiate conflict are stable over days andacross peer partners, r’s = .57–.66 (Hay & Ross, 1982). “Agonistic” interac-tions between toddlers are stable from the first to the third trimester of thesecond year of life, r = .44 (Bronson, 1981). Aggressive ratings during an ex-perimental paradigm where adults simulated anger were found to be stableover one month, r = .48 (Cummings, Iannotti, & Zahn-Waxler, 1985). Tod-dlers’ “negative interactions” with peers predicted teachers’ ratings of exter-nalizing problems at 5 years of age, r = .31; the observed measure of aggres-sion was a stronger predictor of teacher-rated externalizing problems thanmeasures of temperament, security of attachment, or parent-rated behavioralproblems (Fagot & Leve, 1998).
There is considerable debate as to whether such continuities are more evi-dent for boys or for girls. Two-year-old children’s “dysregulated aggression”with familiar peers, measured in a laboratory situation, predicted aggressionwith peers for boys but not for girls three years later (Cummings, Iannotti, &Zahn-Waxler, 1989). This finding is in line with other longitudinal researchsuggesting continuities over the lifespan in male aggression (e.g., Olweus,1979). However, in a study of toddlers in lower-income families, continuitiesover time were observed for girls as well as boys, r’s = .35 and .23, respectively(Keenan & Shaw, 1994).
In some samples, continuity in aggressive tendencies is more evident forgirls than for boys. For example, in our longitudinal study of toddlers with fa-miliar peers (Hay et al., 2000), both mothers’ ratings of aggressiveness andobserved measures of personally directed force showed significant levels ofstability over a 6-month period for girls (r’s = .61 and .47, respectively), butnot for boys. Similar patterns are also seen in sibling interaction, where thefirstborn child’s hostility to the younger sibling is persistent over a 6-monthperiod for girls (r = .66), but not for boys (Kendrick & Dunn, 1983). Two-year-olds’ observed initiation of conflict and aggression predicted ratings ofexternalizing problems two years later for girls, but not for boys (Rubin et al.,2003).
Thus, just as female toddlers’ aggressiveness seems more consistent across
120 T H E D E V E L O P M E N T O F A G G R E S S I O N

measures and informants, so their forceful and aggressive behavior showscontinuities over the years of early childhood. It thus becomes of considerableinterest to inquire why there appears to be a discontinuity thereafter. Girls’ an-tisocial behavior is often linked to a wide range of problems in later childhoodand adolescence (Loeber & Hay, 1994; Rutter, Giller, & Hagell, 1998). It ispossible that girls who are overtly aggressive in early childhood desist fromthe overt use of force but continue to manifest other types of adjustment prob-lems thereafter (Rubin et al., 2003); it is certainly possible that they turn to re-lational aggression (Crick, 1995).
Links with Attributions of Intent
Highly aggressive children often make misattributions about their compan-ions’ intentions, and come to be rejected by their peers through a series of mis-understandings (Coie & Dodge, 1998). Children who are rejected by theirpeers in childhood are at risk for later conduct problems and delinquency,even if by that time they have formed close ties with other deviant teenagers(Cairns & Cairns, 1994). Analogous links between attributions about peers’intentions and aggression can be discerned in the first years of life.
We have seen that a rudimentary anticipation of peers’ intentions can bediscerned in the course of toddlers’ conflicts over objects, when one childwithdraws or defends an object that is being gestured to by the peer. In ourlongitudinal study of British toddlers, those children who often responded topeers’ possible designs on their possessions in this way were significantly moreaggressive than other toddlers. They were more likely to use personal forceagainst the peer, and, especially, to use it proactively, r’s = .60 to .64. Indeed,defensive reactions to peers’ gestures during the first home visit significantlypredicted proactive force 6 months later, r’s = .35 to .45 (Hay et al., 2000).The association between defensive reactions to peers’ possible intentions andthe use of force was especially striking for boys. Thus, individual differencesin toddler boys’ aggression may be linked to this dimension of social under-standing.
RISK FACTORS AND THE EARLY DEVELOPMENT OF AGGRESSION
We have seen that behavioral precursors to aggression appear around thetime of the first birthday and that stable and coherent individual differencesemerge in the 2nd and 3rd years of life. We know that a variety of biologi-cal and social risk factors are associated with aggression at later points inthe lifespan. Are there any links between the early forms of aggression dis-cussed in this chapter and known sources of risk? In this final section of thechapter, I speculate about some possible risk factors that might operate inearly life and therefore influence toddlers’ use of force against their compan-ions.
The Beginnings of Aggression in Infancy 121

Inheritance
Behavioral genetics studies have provided only modest evidence for theheritability of overt aggression, as opposed to broader categories of conductproblems and behavioral dysregulation; aggression has been observed directlyin only a few such studies, which have shown little evidence for heritability(DiLalla, 2002). One exception is a study of 19-month-olds’ physical aggres-sion (Dionne, Tremblay, Boivin, Laplante, & Pérusse, 2003), which suggeststhat the striking individual differences in early aggression reported in the stud-ies just reviewed are partly due to genetic factors. Studies that have focused at-tention on parents’ reports of externalizing problems have shown higher levelsof heritability, though also considerable evidence for environmental effects(DiLalla, 2002). It should be noted that measures of externalizing and inter-nalizing problems are highly correlated in the preschool years (e.g., Hay et al.,1999b) and that externalizing scores included items relevant to attention-defi-cit/hyperactivity disorder (ADHD), which has been shown to be influenced bygenetic factors (Thapar, Holmes, Poulton, & Harrington, 1999). Thus, find-ings regarding the heritability of externalizing problems do not provide directevidence concerning the genetics of aggression.
One possibility for future research is to examine links between psycho-metric and physiological measures of temperament and children’s overt ag-gression. Infant temperament has been demonstrated to be heritable (Arcus &Kagan, 1995; Goldsmith, 1996). Individual differences in temperament areunder intense scrutiny, particularly in terms of their correlations with patternsof frontal lobe activation (Fox et al., 1995; Calkins, Fox, & Marshall, 1996),production of cortisol (Lewis & Ramsay, 1995; Hertsgaard, Gunnar, Erick-son, & Nachmias, 1995; Nachmias, Gunnar, Mangelsdorf, Parritz, & Buss,1996), and vagal tone (Eisenberg et al., 1995).
This literature is complex. Studies of older individuals suggest that bothextremes of underreactivity and overreactivity may be associated with aggres-sion. For example, antisocial behavior is sometimes predicted by low heartrate (Raine, 2002) and hyposecretion of cortisol (McBurnett, Lahey, Rathouz,& Loeber, 2000). However, externalizing problems in early childhood aresometimes associated with hypersecretion of cortisol (Gunnar, Tout, de Haan,& Pierce, 1997). This suggests that there may be two distinct pathways to se-rious aggression; early temperamental features may predict an aggressive ca-reer that is characterized by reactive versus proactive aggression (Vitaro et al.,2002).
Less is known about the contribution of these temperamental features tovery early aggression. It has been noted that aggressive toddlers do not showthe same patterns of vagal suppression in response to experimental challengesshown by their agemates (Calkins & Fox, 2002). This was seen both with re-spect to parental ratings of externalizing problems and overt aggressive behav-ior with unfamiliar peers.
122 T H E D E V E L O P M E N T O F A G G R E S S I O N

Prenatal Insults
Prospective longitudinal studies have demonstrated links between prenatal in-sults and later antisocial behavior (for a review, see Raine, 2002). Forexample, analyses of large British cohort studies have demonstrated links be-tween maternal smoking and early-onset conduct disorder (Maughan, Taylor,Taylor, Butler, & Bynner, 2001). Similar links between smoking in pregnancyand conduct symptoms have been observed across a range of studies (Wak-schlag & Hans, 2002).
Mothers’ mental health problems during pregnancy also put infants atrisk. In a large British cohort study, maternal anxiety during pregnancy pre-dicted the child’s later externalizing problems (O’Connor, Heron, Golding,Beveridge, & Glover, 2002). Analyses of our own longitudinal data set, theSouth London Child Development Study, have shown that children whosemothers are depressed during pregnancy are seven times as likely to be diag-nosed with conduct disorder at age 11, even when the mother’s later history ofdepression is taken into account.
The Caregiving Environment
Whatever the heritable risk for aggression, it is clear that factors in thecaregiving environment potentiate risk. For example, insecure attachment re-lationships are associated with externalizing problems in the preschool years,especially for boys (Turner, 1991); secure attachment seems especially impor-tant for the optimal development of irritable infants (van den Boom, 1995).Avoidant attachment patterns are associated with aggressive behavior in high-risk samples; however, across a wide range of samples, the link between disor-ganized patterns of attachment and aggression is evident (for a review, seeLyons-Ruth, 1996). The effects of disorganized attachment relationships maybe bound up with multiple risk factors, including biological ones. Moleculargenetic studies have demonstrated associations between disorganized attach-ment and the DRD4-7 allele of the dopamine receptor gene (e.g., Lakatos etal., 2003).
Structural features of the caregiving environment, such as the mother’smarital status, are also associated with young children’s externalizing prob-lems (e.g., Fagot & Leve, 1998). The impact of family arrangements is oftenmediated by parental psychopathology and parental behavior (for a review,see Hay & Nash, 2002). For example, analyses of a large British communitysample have shown that links between family structure and children’s exter-nalizing problems are mediated by parental depression (Dunn, Deater-Deckard,Pickering, O’Connor, & Golding, 1998). The timing of episodes of depressionis important. In our longitudinal study in South London, violent behavior atage 11 was predicted by the mother’s depression during the child’s firstmonths of life, even when family structure and economic status, and the
The Beginnings of Aggression in Infancy 123

mother’s subsequent mental health problems, were taken into account (Hay,Pawlby, Angold, Harold, & Sharp, 2003).
There has been relatively little attention paid to the impact of parentalpsychopathology on aggression in the first years of life. However, two corre-lates of maternal depression—lowered responsiveness and irritability—clearlyinfluence early aggression. Maternal responsiveness to sibling conflicts de-creases the chance of aggression being used (Perlman & Ross, 1997). Mater-nal negativity increases toddlers’ aggression with peers (Rubin et al., 1998). Ina large multisite analysis of the impact of infant day care on peer relation-ships, toddlers’ aggression against peers was predicted by maternal sensitivity,as well as by the day caregiver’s behavior (NICHD Early Childcare ResearchNetwork, 2001).
The amount of time spent being cared for by someone other than themother appears to have a modest but significant effect on children’s subse-quent externalizing problems, even when factors such as maternal smokingand depression are taken into account (NICHD Early Childcare ResearchNetwork, 2003). It is not clear, however, what dimensions of the child care ex-perience foster aggression. Several possibilities deserve testing. For example,to the extent that adult intervention in children’s quarrels reduces aggression(Perlman & Ross, 1997), it is important to know whether child care workersare inclined to intervene, or whether they instead believe it is important forchildren to work out their own problems. Second, and more controversial, al-though harsh discipline is generally associated with children’s externalizingproblems, some evidence suggests that there may be positive effects of punish-ment in the toddler years (Baumrind, Larzelere, & Cowan, 2002). To the ex-tent that toddlers’ aggression is not punished or otherwise dealt with by childcare workers, it may stay within the children’s repertoires. Finally, peer influ-ences foster aggression when children are aggregated into groups. There isconsiderable evidence for the ill effects of group-based interventions for ag-gressive children in middle childhood and adolescence (e.g., Dishion, Bullock,& Granic, 2002). It is time to investigate whether similar effects of spendingthe day with peers who are at the peak age for aggression (i.e., between 2 and3) similarly encourages externalizing problems in toddlers. However, it shouldbe noted that physical aggression is shown more often if children are cared forby their mothers in high-risk homes than if children from low-risk homes at-tend day care (Borge, Rutter, Côté, & Tremblay, 2004).
CONCLUSIONS
The behavioral precursors to aggression in early life are neither random nortrivial. Yet these behaviors, for the most part, occur at low rates. In severalsamples, personal aggression occurs too seldom to be included in statisticalanalyses. Instrumental aggression is rather more common, but shows a cleardecline over time. Some toddlers, however, use force at higher rates than their
124 T H E D E V E L O P M E N T O F A G G R E S S I O N

peers and do so proactively and with greater intensity. These individual differ-ences in the rate at which force is used do not derive solely from situationalfactors. There is consistency across different measures, and stability over time.Individual differences in early forms of aggression are associated with biologi-cal and social risk factors. This has important implications for theory andpractice.
With respect to developmental theory, additional research on the early or-igins of aggression would help resolve current debates. For example, classicsocial learning accounts of aggression (e.g., Bandura & Walters, 1963), whichlong held sway in developmental theory, have been challenged; children donot need to learn how to be aggressive. At the same time, the search for bio-logical underpinnings of aggression has been hampered by problems of mea-surement, with only a few genetic studies attempting to assess aggression di-rectly. This is partly because it is neither convenient nor safe for psychologiststo be lurking about when serious violence is taking place; they rely on self-reports, informants’ testimony, projective tests, and laboratory simulations.However, as we have seen, toddlers are not loath to use direct force againsttheir companions. Their use of force can be easily and reliably measuredwithin genetically informative designs (e.g., Dionne et al., 2003).
There is accumulating evidence that aggression is influenced by both bio-logical and environmental factors. Gene–environment correlations and gene–environment interactions are both likely to be important. This point has beenunderscored by the recent molecular genetic evidence concerning the risk forantisocial behavior in genetically vulnerable individuals who have been mis-treated (Caspi et al., 2002). Normative biological processes underpin the de-velopment of aggression, but individual differences are affected by social aswell as biological factors.
Thus, I argue that the early development of aggression entails the gradualattainment of an “aggressive competence,” that is, the ability to deploy physi-cal and verbal aggression effectively in conflict situations in a species-charac-teristic manner, using the advantages of our fists, our opposable thumbs—sogood for pinching and pulling hair—and our powers of hurtful speech. Theappearance of distinctive forms of human aggression is then followed by theflowering of individual differences in “aggressive performance,” influenced byan interaction between the child’s own temperament and social input. Per-sonal aggression soon becomes even more of a minority phenomenon, and thedevelopment of other, more socially acceptable ways of pursuing one’s self-interest leads to a decline in the boorish seizing of other people’s things. Girlsas well as boys develop the basic “aggressive competence,” but they are likelyto desist sooner. Indeed, what happens to girls may qualify as one of the mosteffective social intervention strategies ever attempted.
With respect to social policy and clinical practice, it is vitally important totake on board the fact that some toddlers are already showing serious aggres-sive tendencies. The belief that early aggression is normative, and perhapseven desirable, may blind us to important individual differences. Some 2-year-
The Beginnings of Aggression in Infancy 125

olds use force against their companions at rates considerably higher than thenorm. This bodes poorly for adjustment to preschool groups and for latersocial life. We know that some preschool children are already showingoppositional behavior and conduct problems of clinical concern (Keenan &Wakschlag, 2002). It is important to support infants’ and toddlers’ early de-velopment and thus prevent the consolidation of early individual differencesinto entrenched aggressive careers.
REFERENCES
Ainsworth, M. D. S., Blehar, M. C., Waters, E., & Wall, S. (1978). Patterns of attachment:Assessed in the strange situation and at home. Hillsdale, NJ: Erlbaum.
Appelbaum, M., & McCall, R. (1983) Design and analysis in developmental psychology.In P. H. Mussen (Series Ed.) & W. Kessen (Ed.), Handbook of child psychology: Vol.1. History, theory, and methods (pp. 415–476). New York: Wiley.
Arcus, D., & Kagan, J. (1995). Temperament and craniofacial variation in the first twoyears. Child Development, 66, 1529–1540.
Bakeman, R., & Brownlee, J. R. (1982). Social rules governing object conflicts in toddlersand preschoolers. In K. H. Rubin & H. S. Ross (Eds.), Peer relations and social skillsin childhood (pp. 99–110). New York: Springer-Verlag.
Bandura, A., & Walters, R. H. (1963). Social learning and personality development. NewYork: Holt, Rinehart, & Winston.
Baumrind, D., Larzelere, R. E., & Cowan, P. A. (2002). Ordinary physical punish-ment: Is it harmful? Comment on Gershoff (2002). Psychological Bulletin, 128,580–589.
Belsky, J., Woodworth, S., & Crnic, K. (1996). Trouble in the second year: Three ques-tions about family interaction. Child Development, 67, 556–578.
Biringen, Z., Emde, R. N., Campos, J. J., & Appelbaum, M. I. (1995). Affective reorgani-zation in the infant, the mother, and the dyad: The role of upright locomotion and itstiming. Child Development, 66, 499–514.
Borge, A. I. H., Rutter, M., Côté, S., & Tremblay, R. (2004). Early child care and physicalaggression: Differentiating social selection and social causation. Journal of ChildPsychology and Psychiatry, 45, 367–376.
Bridges, K. M. B. (1932). Emotional development in early infancy. Child Development,3, 324–341.
Bridges, K. M. B. (1933). A study of social development in early infancy. Child Develop-ment, 4, 36–49.
Bridges, K. M. B. (1936). The development of the primary drives in infancy. Child Devel-opment, 7, 40–56.
Bronson, W. C. (1981). Toddlers’ behaviors with agemates: Issues of interaction, cogni-tion, and affect. Norwood, NJ: Ablex.
Brooks-Gunn, J., & Lewis, M. (1979). The effects of age and sex on infants’ playroom be-haviour. Journal of Genetic Psychology, 134, 99–105.
Brownlee, J., & Bakeman, R. (1981). Hitting in toddler peer interaction. Child Develop-ment, 52, 1076–1079.
Cairns, R. (1979). Social development: The origins and plasticity and interchanges. SanFrancisco: Freeman.
126 T H E D E V E L O P M E N T O F A G G R E S S I O N

Cairns, R., & Cairns, B. D. (1994). Lifelines and risks: Pathways of youth in our time.Cambridge, UK: Cambridge University Press.
Calkins, S. D., & Fox, N. A. (2002). Self-regulatory processes in early personality devel-opment: A multilevel approach to the study of childhood social withdrawal and ag-gression. Development and Psychopathology, 14, 477–498.
Calkins, S. D., Fox, N. A., & Marshall, T. R. (1996). Behavioral and physiological ante-cedents of inhibited and uninhibited behavior. Child Development, 67, 523–540.
Caplan, M., Vespo, J. E., Pedersen, J., & Hay, D. F. (1991). Conflict and its resolution insmall groups of one- and two-year-olds. Child Development, 62, 1513–1524.
Caspi, A., McClay, J., Moffitt, T., Mill, J., Martin, J., Craig, I. W., et al. (2002). Role ofgenotype in the cycle of violence in maltreated children. Science, 297, 851–854.
Castell, R. (1970). Effect of familiar or unfamiliar environment on proximal behaviourof young children. Journal of Experimental Child Psychology, 9, 342–370.
Cohn, J. F., & & Tronick, E. Z. (1983). Three-month-old infants’ reactions to simulatedmaternal depression. Child Development, 54, 185–193.
Coie, J. D., & Dodge, K. A. (1998). Aggression and antisocial behavior. In W. Damon(Ed.), Handbook of child psychology (Vol. 3., pp. 779–862). Chichester, UK:Wiley.
Cole, P. M., Barrett, K. C., & Zahn-Waxler, C. (1992). Emotion displays in two-year-oldsduring mishaps. Child Development, 63, 314–324.
Crick, N. R. (1995). Relational aggression: The role of intent attributions, feelings of dis-tress and provocation type. Development and Psychopathology, 7, 313–322.
Cummings, E. M., Iannotti, R. J., & Zahn-Waxler, C. (1985). Influence of conflict be-tween adults on the emotions and aggression of young children. DevelopmentalPsychology, 21, 495–507.
Cummings, E. M., Iannotti, R. J., & Zahn-Waxler, C. (1989). Aggression between peersin early childhood: Individual continuity and change. Child Development, 60, 887–895.
Denham, S. A., McKinley, M., Couchoud, E. A., & Holt, R. (1990). Emotional and be-havioral predictors of preschool peer ratings. Child Development, 61, 1145–1152.
De Wolff, M. S., & van IJzendoorn, M. H. (1997). Sensitivity and attachment: A meta-analysis on parental antecedents of infant attachment. Child Development, 68,571–591.
DiLalla, L. F. (2002). Behavior genetics of aggression in children: Review and future di-rections. Developmental Review, 22, 593–622.
Dionne, G., Tremblay, R. E., Boivin, M., Laplante, D., & Pérusse, D. (2003). Physical ag-gression and expressive vocabulary in 19–month-old twins. Developmental Psy-chology, 39, 261–273.
Dishion, T. J., Bullock, B. M., & Granic, I. (2002). Pragmatism in modeling peer influ-ence: Dynamics, outcomes, and change processes. Development and Psycho-pathology, 14, 969–981.
Dodge, K. A., & Coie, J. D. (1987). Social information processing factors in reactive andproactive aggression in children’s peer groups. Journal of Personality and SocialPsychology, 53, 1146–1158.
Dunn, J. (1988). The beginnings of social understanding. Cambridge, MA: Harvard Uni-versity Press.
Dunn, J. (1993). Children’s relationships: Beyond attachment. London: Sage.Dunn, J., Deater-Deckard, K., Pickering, K., O’Connor, T., & Golding, J. (1998).
Children’s adjustment and prosocial behavior in step- single-parent and non-
The Beginnings of Aggression in Infancy 127

stepfamily settings: Findings from a community sample. Journal of Child Psychol-ogy and Psychiatry, 38, 965–980.
Eckerman, C. O., Whatley, J., & Kutz, S. L. (1975). Growth of social play with peers dur-ing the second year of life. Developmental Psychology, 11, 42–49.
Eisenberg, N., Fabes, R. A., Murphy, B., Maszk, P., Smith, M., & Karbon, M. (1995).The role of emotionality and regulation in children’s social functioning: A longitudi-nal study. Child Development, 66, 1360–1384.
Fagot B., & Leve, L. D. (1998). Teacher ratings of externalizing behaviour at school entryfor boys and girls: Similar early predictors and different correlates. Journal of ChildPsychology and Psychiatry, 39, 555–546.
Feinman, S. (1982). Social referencing in infancy. Merrill–Palmer Quarterly, 28, 445–470.
Ferguson, T. J., & Rule, B. (1988). Children’s attributions of retaliatory aggression. ChildDevelopment, 59, 961–968.
Field, T., Healy, B., Goldstein, S., Perry, S., Bendell, D., Schanberg, S., Zimmerman, E. A.,& Kuhn, C. (1988). Infants of depressed mothers show “depressed” behavior evenwith nondepressed adults. Child Development, 59, 1569–1579.
Fox, N., Coplan, R. J., Rubin, K. H., Porges, S. W., Calkins, S. D., Long, J. M., Marshall,T. R., & Stewart, S. (1995). Frontal activation asymmetry and social competence atfour years of age. Child Development, 66, 1770–1784.
Frye, D. (1993). Causes and precursors of children’s theories of mind. In D. F. Hay & A.Angold (Eds.), Causes and precursors of development and psychopathology (pp.145–168). Chichester, UK: Wiley.
Goldsmith, H. H. (1996). Studying temperament via construction of the Toddler Behav-ior Assessment Questionnaire. Child Development, 67, 218–235.
Goodenough, F. L. (1931). Anger in young children. Minneapolis: University of Minne-sota.
Green, E. H. (1933). Group play and quarrelling among preschool children. Child Devel-opment, 4, 302–307.
Gunnar, M. R. (1980). Control, warning signals, and distress in infancy. DevelopmentalPsychology, 16, 281–299.
Gunnar, M. R., Tout, K., de Haan, M., & Pierce, S. (1997). Temperament, social compe-tence, and adrenocortical activity in preschoolers. Developmental Psychobiology,31, 65–85.
Gustafson, G. E., Green, J. A., & West, M. J. (1979). The infant’s changing role inmother–infant games. Infant Behavior and Development, 2, 301–308.
Hartup, W. W. (1974). Aggression in childhood: Developmental perspectives. AmericanPsychologist, 29, 336–341.
Hay, D. F., Castle, J., & Davies, L. (2000). Toddlers’ use of force against familiar peers: Aprecursor of serious aggression? Child Development, 71, 457–467.
Hay, D. F., Castle, J., Davies, L., Demetriou, H., & Stimson, C. (1999a). Prosocial ac-tion in very early childhood. Journal of Child Psychology and Psychiatry, 40,905–916.
Hay, D. F., & Nash, A. (2002). Social development in different family arrangements. In P.K. Smith & C. H. Hart (Eds.), Blackwell handbook of childhood social develop-ment (pp. 238–261). Oxford, UK: Blackwell.
Hay, D. F., Nash, A., & Pedersen, J. (1983). Interaction between six-month-old peers.Child Development, 54, 557–562.
Hay, D. F., Pawlby, S., Angold, A., Harold, G., & Sharp, D. (2003). Pathways to violence
128 T H E D E V E L O P M E N T O F A G G R E S S I O N

in the children of mothers who had postnatal depression. Developmental Psychol-ogy, 39, 1083–1094.
Hay, D. F., Pawlby, S., Sharp, D., Schmücker, G., Mills, A., Allen, H., & Kumar, R.(1999b). Parents’ judgements about young children’s problems: Why mothers andfathers might disagree yet still predict later outcomes. Journal of Child Psychologyand Psychiatry, 40, 1249–1258.
Hay, D. F., & Ross, H. S. (1982). The social nature of early conflict. Child Development,53, 105–111.
Hay, D. F., Vespo, J. E., & Zahn-Waxler, C. (1998). Young children’s conflicts with theirsiblings and mothers: Links with maternal depression and bipolar illness. BritishJournal of Developmental Psychology, 16, 519–538.
Hay, D. F., Zahn-Waxler, C., Cummings, E. M., & Iannotti, R. (1992). Young children’sviews about conflict with peers: A comparison of the daughters and sons of de-pressed and well women. Journal of Child Psychology and Psychiatry, 33, 669–683.
Hertsgaard, L., Gunnar, M., Erickson, M. F., & Nachmias, M. (1995). Adrenocorticalrespones to the strange situation in infants with disorganized/disoriented attach-ment relationships. Child Development, 66, 1100–1106.
Holmberg, M. C. (1980). The development of social interchange patterns from 12 to 42months. Child Development, 51, 448–456.
Hughes, C., White, A., Sharpen, J., & Dunn, J. (2000). Antisocial, angry and unsympa-thetic: “Hard-to-manage” preschoolers’ peer problems and possible cognitive influ-ences. Journal of Child Psychology and Psychiatry, 41, 169–179.
Ishikawa, F. (2003). Dyadic and triadic interaction between newly acquainted two-year-olds. Unpublished doctoral dissertation, Cambridge University, Cambridge, UK.
Kagan, J. (1981). The second year. Cambridge, MA: Harvard University Press.Keenan, K., & Shaw, D. (1994). The development of aggression in toddlers: A study of
low-income families. Journal of Abnormal Child Psychology, 22, 53.Keenan, K., & Shaw, D. (1997). Developmental and social influences on young girls’
early problem behavior. Psychological Bulletin, 121, 95–113.Keenan, K., & Wakschlag, L. S. (2002). Can a valid diagnosis of disruptive behavior dis-
order be made in preschool children? American Journal of Psychiatry, 159, 351–358.
Kendrick, C., & Dunn, J. (1983). Sibling quarrels and maternal responses. Developmen-tal Psychology, 19, 62–70.
Kochanska, G. (1993). Toward a synthesis of parental socialization and child tempera-ment in early development of conscience. Child Development, 64, 325–347.
Kochanska, G., Aksan, N., & Koenig, A. L. (1995a). A longitudinal study of the roots ofpreschoolers’ conscience: Committed compliance and emerging internalization.Child Development, 66, 1752–1769.
Kochanska, G., Casey, R. J., & Fukomoto, A. (1995b). Toddlers’ sensitivity to standardviolations. Child Development, 66, 643–656.
Kopp, C. B. (1982). Antecedents of self-regulation: A developmental perspective. Devel-opmental Psychology, 18, 199–214.
Kuczynski, L., & Kochanska, G. (1990). The development of children’s noncompliancestrategies from toddlerhood to age 5. Developmental Psychology, 26, 398–408.
Lakatos, K., Nemoda, Z., Ronai, Z., Ney, K., Sasvari-Szekeley, M., & Gervai, J. (2003).Further evidence for the role of the dopamine DR receptor (DRD4) gene in attach-ment disorganization: Interaction of the exon III 48-bp repeat and the –521 C/Tpromoter polymorphisms. Molecular Psychiatry, 7, 27–31.
The Beginnings of Aggression in Infancy 129

Levy, D. M. (1939). Sibling rivalry studies in children of primitive groups. AmericanJournal of Orthopsychiatry, 9, 205–215.
Lewis, M., Alessandri, S. M., & Sullivan, M. W. (1990). Violation of expectancy, loss ofcontrol, and anger expressions in young infants. Developmental Psychology, 26,745–751.
Lewis, M., & Ramsay, D. S. (1995). Developmental change in infants’ responsiveness tostress. Child Development, 66, 657–670.
Loeber, R., & Hay, D. F. (1994). Developmental approaches to aggression and conductproblems. In M. L. Rutter and D. F. Hay (Eds.), Development through life: A hand-book for clinicians. (pp. 488–516) Oxford, UK: Blackwell Scientific.
Loeber, R., & Hay, D. F. (1997). Key issues in the development of aggression and violencefrom childhood to early adulthood. Annual Review of Psychology, 48, 371–410.
Lyons-Ruth, K. (1996). Attachment relationships among children with aggressive behav-ior problems: The role of disorganized early attachment patterns. Journal of Con-sulting and Clinical Psychology, 64, 64–73.
Maughan, B., Taylor, C., Taylor, A., Butler, N., & Bynner, J. (2001). Pregnancy smokingand childhood conduct problems: A causal association? Journal of Child Psychol-ogy and Psychiatry, 42, 1021–1028.
McBurnett, K., Lahey, B. B., Rathouz, P. J., & Loeber, R. (2000). Low salivary cortisoland persistent aggression in boys referred for disruptive behavior. Archives of Gen-eral Psychiatry, 57, 38–43.
Muste, M. J., & Sharpe, D. I. (1947) Some influential factors on the determination of ag-gressive behavior in preschool children. Child Development, 51, 823–829.
Nachmias, M., Gunnar, M., Mangelsdorf, S., Parritz, R., & Buss, K. (1996). Behavioralinhibition and stress reactivity: The moderating role of attachment security. ChildDevelopment, 67, 508–522.
NICHD Early Child Care Research Network. (2001). Child care and children’s peer in-teraction at 24 and 36 months. Child Development, 72, 1478–1500.
NICHD Early Child Care Research Network. (2003). Does amount of time spent in childcare predict socioemotional adjustment during the transition to kindergarten?Child Development, 74, 976–1005.
O’Connor, T., Heron, J., Golding, J., Beveridge, M., & Glover, V. (2002). Maternal ante-natal anxiety and children’s behavioural/emotional problems at 4 years. BritishJournal of Psychiatry, 180, 502–508.
Olweus, D. (1979). Stability of aggressive reaction patterns in males: A review. Psycho-logical Bulletin, 86, 852–857.
Pedersen, J. (1985). Young girls’ and boys’ conflicts with their preschool peers. Unpub-lished doctoral dissertation, State University of New York at Stony Brook.
Perlman, M., & Ross, H. S. (1997). The benefits of parent intervention in children’s dis-putes: An examination of concurrent changes in children’s fighting styles. Child De-velopment, 68, 690–700.
Price, J. M., & Dodge, K. A. (1989). Reactive and proactive aggression in childhood: Re-lations to peer status and social context dimensions. Journal of Abnormal ChildPsychology, 17, 455–471.
Raine, A. (2002). Annotation: The role of prefrontal deficits, low autonomic arousal,and early health factors in the development of antisocial and aggressive behavior inchildren. Journal of Child Psychology and Psychiatry, 43, 417–434.
Rheingold, H. L., Cook, K., & Kolowitz, V. (1987). Commands activate the behavior andpleasure of two-year-old children. Developmental Psychology, 23, 146–151.
130 T H E D E V E L O P M E N T O F A G G R E S S I O N

Rheingold, H. L., & Eckerman, C. O. (1970). The infant separates himself from hismother. Science, 168, 78–83.
Ross, H. S. (1996). Negotiating principles of entitlement in sibling property disputes. De-velopmental Psychology, 32, 90–101.
Ross, H. S., & Hay, D. F. (1977). Conflict and conflict resolution between 21-month-oldpeers. Paper presented at the biennial meeting of the Society for Research in ChildDevelopment, New Orleans.
Ross, H. S., & Kay, D. A. (1980). The origins of social games. In K. H. Rubin (Ed.),Children’s play (pp. 17–31). San Francisco: Jossey-Bass.
Ross, H. S., Tesla, C., Kenyon, B., & Lollis, S. P. (1990). Maternal intervention in toddlerpeer conflict: The socialization of principles of justice. Developmental Psychology,26, 994–1003.
Rothbart, M. K. (1981). Measurement of temperament in infancy. Child Development,52, 569–578.
Rubin, K. H., Burgess, K. B., Dwyer, K. M., & Hastings, P. D. (2003). Predicting pre-schoolers’ externalizing behaviors from toddler temperament, conflict, and mater-nal negativity. Developmental Psychology, 39, 164–176.
Rubin, K. H., Hastings, P., Chen, X., Stewart, S., & McNichol, K. (1998). Intrapersonaland maternal correlates of aggression, conflict, and externalizing problems in tod-dlers. Child Development, 69, 1614–1629.
Rutter, M., Giller, H., & Hagell, A. (1998). Antisocial behavior by young people. Cam-bridge, UK: Cambridge University Press.
Sackin, S., & Thelen, E. (1984). An ethological study of peaceful associative outcomes toconflict in preschool children. Child Development, 55, 1098–1102.
Schult, C. A. (2002). Children’s understanding of the distinction between intentions anddesires. Child Development, 73, 1727–1747.
Schwartz, D., Dodge, K. A., Coie, J. D., Hubbard, J. A., Cillessen, A. H., Lemerise, E. A.,& Bateman. (1998). Social-cognitive and behavioral correlates of aggression andvictimization in boys’ play groups. Journal of Abnormal Child Psychology, 26,431–440.
Smetana, J. (1984). Toddlers’ social interactions regarding moral and conventional trans-gressions. Child Development, 35, 1767–1776.
Smetana, J. (1989). Toddlers’ social interaction in the context of moral and conventionaltransgressions in the home. Developmental Psychology, 25, 499–508.
Stenberg, C. R., Campos, J. J., & Emde, R. N. (1983). The facial expression of anger inseven-month-old infants. Child Development, 54, 178–184.
Stimson, C. A., Castle, J., & Hay, D. F. (1997). Mothers’ construction of their toddlers’personalities. Child Study Journal, 27, 21–39.
Strayer, F. F., & Strayer, J. (1976). An ethological analysis of social agonsim anddominance relations among preschool children. Child Deveolopment, 47, 980–989.
Sutton, J., Smith, P. K., & Swettenham, J. (1999). Bullying and “theory of mind”: A cri-tique of the “social skills deficit” view of antisocial behaviour. Social Development,8, 117–127.
Thapar, A., Holmes, J., Poulton, K., & Harrington, R. (1999). The genetic basis of atten-tion deficit and hyperactivity. British Journal of Psychiatry, 174, 105–111.
Tremblay, R. E. (2000). The development of aggressive behaviour during childhood:What have we learned in the past century? International Journal of Behavioural De-velopment, 24, 129–141.
The Beginnings of Aggression in Infancy 131

Turner, P. J. (1991). Relations between attachment, gender, and behavior with peers inpreschool. Child Development, 62, 1475–1488.
van den Boom, D. (1995). Do first-year intervention effects endure? Followup duringtoddlerhood of a sample of Dutch irritable infants. Child Development, 66, 1798–1816.
Vespo, J. E., Pedersen, J., & Hay, D. F. (1995). Young children’s conflicts with peers andsiblings: Gender effects. Child Study Journal, 25, 189–212.
Vitaro, F., Brendgen, M., & Tremblay, R. (2002). Reactively and proactively aggressivechildren: Antecedent and subsequent characteristics. Journal of Child Psychologyand Psychiatry, 43, 495–505.
Vitaro, F., Gendreau, P. L., Tremblay, R. E., & Oligny, P. (1998). Reactive and proactiveaggression differentially predict later conduct problems. Journal of Child Psychol-ogy and Psychiatry, 39, 377–385.
Wakschlag, L., & Hans, S. L. (2002). Maternal smoking during pregnancy and conductproblems in high-risk youth: A developmental framework. Development andPsychopathology, 14, 351–369.
Wellman, H. H., Phillips, A. T., & Rodriguez, T. (2000). Young children’s understandingof perceptions, desire, and emotion. Child Development, 71, 895–912.
132 T H E D E V E L O P M E N T O F A G G R E S S I O N

THE DEVELOPMENT OF AGGRESSIONPlay and the Regulation of Aggression
7
Play and the Regulation of Aggression
JORDAN B. PETERSON and JOSEPH L. FLANDERS
INTRODUCTION: AGGRESSION, EMPATHY, AND SOCIAL BEING ARE ALL INNATE
With what simplicity would I have demonstrated that man isnaturally good and that it is through these institutions alone thatmen become bad.
—ROUSSEAU, Discourse on the Arts and Sciences (1750/1974)
Hereby it is manifest, that during the time men live without acommon Power to keep them all in awe, they are in thatcondition which is called War; and such a war, as is of everyman, against every man.
—HOBBES, The Leviathan (1651/1909)
Since the 1960s, our culture has leaned powerfully toward the developmentalphilosophy of Rousseau: Children are naturally self-regulating, creative, posi-tive, and good. Only the arbitrary forces of culture make them bad. Carl Rog-ers (1989, p. x), a psychologist perhaps more responsible than any other forthe modern promotion of Rousseau’s viewpoint, makes the following observa-tion: Man is
[a] basically trustworthy member of the human species, whose deepest char-acteristics tend toward development, differentiation, cooperative relation-ships . . . whose impulses tend naturally to harmonize into a complex andchanging pattern of self-regulation; whose total character is such as to tend to
133

preserve and enhance himself and his species, and perhaps to move it towardits further evolution.
But is it really true that in our more aggressive moments we are all merely in-nocent victims of our cultures? Hobbes (1651/1909) believed very much theopposite: In his state of nature,
there is no place for industry, because the fruit thereof is uncertain: and conse-quently no culture of the earth; no navigation, nor use of the commoditiesthat may be imported by sea; no commodious building; no instruments ofmoving and removing such things as require much force; . . . no account oftime; no arts; no letters; no society; and which is worst of all, continual fear,and danger of violent death; and the life of man, solitary, poor, nasty, brutish,and short.
Many cultures tilt strongly toward Hobbes: The child is a force of nature,willful, destructive, capable of self-harm, in dire need of careful, cautious, andintense socialization, and damned in the absence of social order (Fischer,1989).
What might a modern psychologist say about this debate, given the prog-ress in the behavioral sciences over the last century? Is the human being a no-ble savage, corrupted by the stresses of civilized social being, or a beast ofprey, selfish and cruel? To answer this question properly, it is necessary to con-sider evidence derived from a diverse sampling of the behavioral sciences andto reframe the argument: What tendencies to aggression, if any, characterizethe human species, and what mechanisms, individual and social, regulate andconstrain those tendencies?
It should first be noted that the psychoanalytic viewpoint is not a simplederivative of the Hobbesian perspective, in the same way that the humanisticview is of Rousseau’s. Freud proposed that aggression was innate, part of theid, noting that aggression emerged as a consequence of socially induced frus-tration in the form of conflict between the pleasure and reality principles. Inthe psychoanalytic world, the id (nature), the ego (the individual and his orher subjectivity) and the superego (culture) are all good and bad, simulta-neously. In keeping with this conceptualization, it appears that aggression is anatural component of our behavioral repertoire, emerging far back in the se-quence of development, and not something added secondarily to an essentiallypeaceful temperament. Young children appear fundamentally egocentric (Piaget,1932). They hold their own intrinsic desires paramount and exist in a worldwhere those desires are bounded only by their immediate consequences. Theyreliably begin to manifest aggressive behaviors such as pushing, hitting, kick-ing, and throwing about 18 months of age, although there is wide individualvariability in the frequency with which these behaviors are manifested (Nagin& Tremblay, 1999). Such aggression, manifested in defensive or instrumentalform (Vitiello & Stoff, 1997), appears dependent on the operation of very low
134 T H E D E V E L O P M E N T O F A G G R E S S I O N

level, early-maturing brain structures, such as the hypothalamus or periaque-ductal grey (reviewed in Peterson & Shane, 2004). The incidence of aggressivebehavior peaks, surprisingly, in kindergarten, and then declines over time(Nagin & Tremblay, 1999). By age 4, most children have become social. Thesmall number who have not (Nagin & Tremblay, 1999) tend to be aggressivefor the rest of their lives (Coie & Dodge, 1998). Chronically aggressive chil-dren, then adults, lack empathy, are suspicious, narcissistic, and self-centered(Coie & Dodge, 1998), and are characterized by inappropriate and brittlehigh self-esteem (Olweus, 1994; Bushman & Baumeister, 1998). Few interven-tions appear helpful.
Chimpanzees, our surprisingly close cousins (Sibley & Ahlquist, 1984),appear primally aggressive within their social groups, in the same manner aschildren. Most of their within-group aggression appears related to dominance-hierarchy maneuvering, as it does in the human case (Wilson & Daly, 1997).Such maneuvering appears to initially manifest itself in the innocuous and eas-ily overlooked form of teasing. De Waal (1996, p. 114) states:
[Chimp youngsters] throw handfuls of dirt or pebbles at their elders, hit themwith sticks, splash them with water, jump on their heads when they are doz-ing, and so on. Much of the time, the individual thus bothered takes it re-markably well, tickles the youngster, or makes a mock chase that turns thewhole incident into a game.
Teasing techniques transform with age, becoming less frequent, but more se-vere. The infant engages in little pushes from behind, jumping away when theadult turns around. The adolescent male, in contrast, manifests full-fledgedcharging displays, seeking to dominate his peers, the adult females that sur-round him and, eventually, higher-ranking adult males. As adults, chimpsform sophisticated coalitions, jockeying for position and, upon occasion,physically engage and dominate or subordinate themselves to other individu-als in conflicts that can become violent (De Waal, 1996).
Dominance-hierarchy position appears to be a vital determinant of sur-vival and reproductive success. In consequence, little is more important to asocial animal than accurate representation of who rules and who is subordi-nate under what circumstances (Abbott et al., 2003; Virgin & Sapolsky,1997). The establishment and maintenance of a predictable dominance hierar-chy allows for the emergence of orderly access to desirable resources, so thatevery attempt at consummation within the social environment does not imme-diately escalate into an aggressive encounter. Tracking dominance and othersocial information is so important that group size appears as an importantcorrelate of neocortical size in primates, particularly in regard to brain sys-tems devoted to analysis of complex relationships (Joffe & Dunbar, 1997).This all means, of course, that advancement is frequently worth fighting for.
The fact of innate dominance striving, buttressed by the mechanisms ofaggression, does not mean that chimps or humans lack social feeling and sim-
Play and the Regulation of Aggression 135

ply learn to inhibit their aggression through fear or through cognitively medi-ated calculation of the potential consequences of aggressive behavior. Primatesare as gregarious as they are aggressive—even in the immediate aftermath ofintense agonistic encounters (De Waal, 1989b). It appears, therefore, thatagonistic and cooperative behaviors are not necessarily opposed to one an-other, at least not in any simple manner. First, more innately aggressive socialcreatures may also have to be more innately affiliative (De Waal, 1989b), inorder to find and maintain social support, which is even more important tothem than the objective safety of their environment (Abbott et al., 2003). Sec-ond, at any given time or place, individual action and social interaction can becharacterized by cooperation at one level, and competition at another. Amongintensely social animals, the social group, the dominance hierarchy, thesuperordinate level, clearly constitutes a form of extended cooperation.Within that group, however—that cooperative space, the subordinate level—dominance striving takes place. It appears, therefore, that the essentially ag-gressive instincts appear complexly counterbalanced by the interplay of twoequally powerful domains of regulation, one internal and innate; the other, so-cial and emergent.
The internal process that regulates aggression (in addition to simple fear)seems to be empathy or, perhaps, identification—the ability to feel the experi-ences or to adopt the viewpoint of another, respectively. Whether such abilityemerges as a consequence of conditioning, emotional contagion, or cognitivelymediated understanding, the evidence for its existence is strong (Preston & DeWaal, 2002). The circuitry that governs empathy—or its close variants, love,affiliation, and nurturance—is arguably as archaic and deeply rooted as thatmotivating aggression (Preston & De Waal, 2002; Panksepp, 1998a) and ap-pears to play a modulatory role, regulating the intensity of response to thosedeemed kin. A wide range of animals exhibit sophisticated reactions to thedistress or actions of their conspecifics: Rats appear visibly upset by the sightof another rat receiving electric shocks (Rice & Gainer, 1962; Rice, 1964); hy-enas can be primed to eat and drink by the sight of their group mates doingthe same, even when they are not visibly attending (Yoerg, 1991); and rhesusmonkeys will starve themselves if they learn that their food gathering effortsculminate in the shock of a conspecific (Masserman, Wechkin, & Terris,1964). Human infants, in turn, respond with crying to the crying of other in-fants (Zahn-Waxler, Radke-Yarrow, & King, 1979) and, after the first year,imitate the distress behaviors of others and spontaneously manifest helpingbehaviors (Zahn-Waxler, Radke-Yarrow, & Brady-Smith, 1977). Furthermore,Miller, Eisenberg, Fabes, and Shell (1996) have noted that older children whomanifest facial expressions of concern when exposed to the suffering of othersare characterized by higher levels of moral reasoning and increased prosocialbehavior.
The social process that regulates aggression appears more integrally asso-ciated, as mentioned earlier, with dominance-hierarchy structure. Chimps areperfectly capable of killing while hunting and during raiding parties con-
136 T H E D E V E L O P M E N T O F A G G R E S S I O N

ducted on foreign conspecifics (Wrangham & Peterson, 1996), so there isclearly no necessary internal limit on their aggressive behavior. It appears, aswell, that the tendency toward dominance striving among chimps can at leasttemporarily override any innate tendency toward empathy during intenseagonistic within-troupe disputes. De Waal (1989a) has suggested that undersuch conditions it is the whole chimp troupe that constrains the “ambition” ofthe individual, becoming agitated en masse and interfering, actively, with anydominance battle that goes too far. Preston and De Waal (2002) have takenpains to outline the nature of those factors that modulate the expression ofempathy: Familiarity and perceived similarity, as well as factors such as learn-ing, past experience with the cause of suffering, and the salience of the ob-served suffering all affect empathic responding. What this means is that socialforces can alter the probability that empathy will inhibit aggression by alteringthe nature of the factors that modulate both. The consequences of extendedsocial being, however, are more indirectly associated with aggression regula-tion and appear related to the function of neural circuits that mature later in apredictable, regulated, and orderly social environment. If human children aresocialized within such an environment, they learn socially acceptable but morecomplex alternatives to violence. They begin to integrate their own proximatedesires with distal wishes and to consider and allow for the wants and needsof others.
The human child appears to face the world with a basic set of functionalmotivational states, mediated by low-level but sophisticated brain circuitsgoverning action, setting the frame for perception, emotion, and cognition(Gregg & Siegel, 2001; Peterson & Flanders, 2002; Swanson, 2000). The op-eration of these circuits enables the child to identify and pursue valuable goalssuch as food, water, warmth, social affiliation, self-protection, and the explor-ing of new territory (Swanson, 2000; Gregg & Siegel, 2001). Each primarymotivational circuit sets a unidimensional goal for behavior (Swanson, 2000),so that the child’s first developmental requirement is to learn how to attainthat goal. Second, however, he or she must learn how to balance all those pri-mary goals and to determine how they might find their fulfillment within acomplex interpersonal environment. The emergence of the emotional circuits,one stage above those governing motivation, helps fulfill this second set of re-quirements, as does the development of prefrontal circuitry, designed to mod-ulate motivation and emotion and to extend comprehension of the conse-quences of action across broad spans of time and place (Oatley & Johnson-Laird, 1987; Oatley, 1999; Peterson, 1999).
Initially, the child’s mother provides what is needed, with minimal de-mands for reciprocation. However, as the child grows, expectations for reci-procity grow. He or she enters a world characterized by long-term consider-ations and by the presence of other people who have goals and feelings oftheir own. These goals and feelings have to be taken into account, as theymanifest themselves in the same environment that provides opportunity forthe developing child. This means that the child must solve not only the prob-
Play and the Regulation of Aggression 137

lem of his or her own instincts and their interactions, but also the problem ofthe instincts of others, in combination with his or her own.
The movement toward such solution appears to be mediated, at least inpart, by empathy, and then by play. The child appears intrinsically able to ex-perience the motivations and emotions of others. He or she is capable ofaffiliative instincts and is prepared to be a social animal, cooperative and sup-portive, as well as competitive and agonistic. The development of social un-derstanding appears to take place from the bottom up, in a kind of bootstrap-ping process. Rough-and-tumble (R&T) play, mediating and regulating directphysical contact, allows the child to attune his or her body to the embodiedpresence of others. More abstract forms of play allow for the attuning of mo-tivational states, emotional reactions, and the contents of consciousness overincreasingly large spans of space and time. Role play and fantasy mediate ab-stract forms of identification and the consequent extension of empathy tothose beyond the immediately familiar. Finally, the adoption of a role, part fic-tion and part genuine being, constitutes the establishment of a functional posi-tion within a real-world hierarchy of cooperation and dominance.
Numerous researchers have sketched the developmental effects of child-hood play. Such effects include enhanced physical fitness and improved cog-nitive, emotional, and social function (Pellegrini & Smith, 1998). Less atten-tion has been paid to the manner in which play cultivates self-regulation.Play, however, might well be regarded as early social cognition: If I can playwith you, I can adapt my actions and reactions to yours. I can allow yourmotivational and emotional states, your reference frame, to modulate mine.I can start to act out your frame, to understand and to embody it. Even-tually, perhaps, we can share the same perspective and use the fact of thatsharing to work cooperatively toward a common goal. This means that wecan start to share identity, predicated on voluntary compliance with thesame set of values, and benefit mutually from the consequent control ofaggression—not so much because of inhibition, but because of the align-ment of mutual desire. This ability unfolds, over time, in a lengthy develop-mental process—one that appears to start not so much with the mind, asmight be initially predicted, but with the body, in direct, physical contactwith the bodies of others.
ROUGH-AND-TUMBLE PLAY AND THE EMBODIEDNEGOTIATION OF DOMINANCE AND COOPERATION
Pellis and Iwaniuk (2000, p. 136) state that “species with a greater proportionof their growth occurring postnatally play more and have more complex playthan do species with more of their growth occurring prenatally,” in keepingwith Bruner’s (1976) suggestion that the prolonged infancy of humans pro-vides more time for play and more time to develop sophisticated cognitiveabilities. Panksepp (1998a) has argued, specifically, that the mammalian brain
138 T H E D E V E L O P M E N T O F A G G R E S S I O N

is hard-wired for play—at least for rough-and-tumble (R&T) play. Juvenilerats, for example, exhibit R&T play beginning at 17 days of age, even whenprevented from engaging in any prior play experiences, and will play morevigorously if intermittently deprived of the opportunity to do so. However,these early play impulses appear to manifest themselves only under the appro-priate conditions. Fear and hunger and associated states of deprivationquickly eliminate play. Young rats must also have a secure home environment,with abundant parental involvement, to play.
R&T play is different from exploratory activity and from aggression—twoforms of behavior with which it can easily be confused. R&T play and explor-atory activity share the fact that both are enjoyable. Habitual R&T play win-ners and losers will engage in effortful instrumental tasks to gain an opportu-nity to play, for example, indicating that play episodes are reinforcing, andboth will run toward the play arena at equal speeds. (The winners enter confi-dently, however, while the losers move in more timidly and slowly.) Such en-joyment appears mediated in part by the same dopaminergic incentive rewardcircuits that underly exploratory behavior, although play also activates wide-spread release of opiates, particularly in those areas characterized by circuitsfor sexual, maternal, and other affiliative behaviors (Panksepp, 1998a). How-ever, dopamine (DA) agonists such as amphetamines invigorate exploratoryactivity, but markedly reduce play.
Finally, formal behavioral analysis clearly discriminates R&T play fromgenuine aggression. A playful rat chases his or her partner around in a “flurryof dynamic, carefree rambunctiousness” (Panksepp, 1998a, p. 284), pouncingon the partner, pinning him or her, in the consummatory stage of each play ep-isode. Such pinning is clearly a gesture of dominance, but not one that breaksthe rules of play. In a real fight, rats box, and prance sideways, postures andgestures accompanied by piloerection. Furthermore, in genuine dominancebouts, the resident animal consistently wins if the activity occurs in the ani-mal’s home territory. This is not the case in play fighting (Panksepp, 1998a).Finally, pins during a real battle are more sustained and menacing than theyare in playful contexts.
Play fighting and genuine aggression appear as distinguishable within thecontext of human behavior as they are among rats. Blurton Jones (1972), forexample—taking a cue from Harlow and Harlow’s (1965) observations ofplay fighting in young rhesus monkeys—clearly differentiated R&T play fromphysical aggression among preschool children. Children involved in a playfight wrestle, grapple, jump, tumble, and run, while laughing and exhibitingfacial expressions of enjoyment. In addition, in play fighting contexts, chil-dren spend most of their time in close proximity, whereas they come togetherbriefly for a genuinely aggressive act and then pull away rapidly. Positive emo-tional expressions are also markedly absent on such an occasion. Despite theserelatively subtle differences, children are highly adept at distinguishing playfulfrom aggressive fighting (Smith & Boulton, 1990; Boulton & Smith, 1992),regardless of their culture (Costabile et al., 1991), and play fighting gestures
Play and the Regulation of Aggression 139

exchanged between experienced children seldomly elicit a hostile response(Boulton, 1991). Children who become skilled at R&T play learn directlywhat forms of agonistic interaction will be tolerated by others, and carefullyand judiciously limit the social expression of their aggression.
Mothers are the first to initiate R&T play cycles with their infants,through tickling bouts and mocked acts of aggression. As motor coordinationdevelops and children become more active, fathers, who play more robustly,become the play partners of choice (Roopnarine, Hooper, Ahmeduzzaman, &Pollack, 1993). Maternal depression and paternal absence are therefore asso-ciated with childhood externalizing behavior problems (Bates, Bayles, Ben-nett, Ridge, & Brown, 1991; Pagani, Boulerice, Tremblay, & Vitaro, 1997;Patterson, Reid, & Dishion, 1992). Depressed mothers are less likely to tickleand play peek-a-boo with their infants (Field, 1998). Older children of singlemothers have fewer opportunities to learn to regulate aggressive behavior, in-asmuch as fathers tend to engage in play fighting after the end of the first year.Such children appear awkward when invited to engage in R&T play. Acci-dentally, or motivated by frustration, they more frequently hurt their playpartners. The victims respond with rejection. Aggression emerges in responseto this rejection (Asher & Coie, 1990; Smith, Hunter, Carvalho, & Costabile,1992), and a detrimental positive feedback cycle establishes itself.
The broader nature and significance of R&T play, as well as its role as ascaffold for more sophisticated social cognition, may best be revealed within abroader conceptual framework, including both Piagetian and neuropsycho-logical components. As the child develops, he or she experiments, stage bystage, with the construction of small-scale motor patterns, designed to attainsmall-scale, motivated ends. Piaget (1932, pp. 16–18) points out, for example,that in the initial, primary stages of play, a child handles objects at the dictatesof his or her “desires and motor habits.” Because “play is purely individual”at this stage, “ritualized schemas” develop—skilled play habits—but no col-lective patterns, much less rules. The child first plays by him- or herself, con-structing a repertoire of functional actions, then conceptions, from the bottomup. Swanson (2000, p. 115–116) describes the physiology underlying the con-struction of such a functional hierarchy:
The lowest or first level of the locomotor system is formed . . . by a subset ofmotoneuron pools in the spinal cord ventral horn that innervates the limb mus-cles responsible for locomotor behavior. The second major level is referred to asthe locomotor pattern generator, which lies entirely within the spinal cord, nearthe motoneuron pools that it regulates. In fact, it is itself a hierarchy of increas-ingly complex motor pattern generators that coordinate and time muscle con-tractions across individual joints, then across multiple joints within a particu-lar limb, and finally amongst all four limbs. . . . A third major level isrepresented, at least in part, by an ill-defined region of the dorsal tegmentumknown as the mesencephalic locomotor region, and rostroventral to this is afourth major level in an ill-defined region of the caudal hypothalamus/rostralmidbrain—the so called subthalamic or hypothalamic locomotor region.
140 T H E D E V E L O P M E N T O F A G G R E S S I O N

This hypothalamic locomotor region, a locomotor pattern controller, gener-ates downward outputs to the spinal locomotor pattern generator, and is thenext control system to develop, as the individual constructs increasingly com-plex hierarchies of motor behavior. It is of interest to note, in this regard, (1)that tactile stimulation during infancy—an important aspect of R&T play,and one linked to it through the specialized skin receptors involved in suchplay (Panksepp, 1998a)—has an important organizing and stabilizing effecton different brain structures, including the hypothalamic–pituitary–adrenal(HPA) axis (Lande, Scarr, & Gunzenhauser, 1989; Meaney, Aitken, VanBerkel, Bhatnager, & Sapolsky, 1988), and (2) that children with chronic ag-gression problems are frequently characterized by dysregulated HPA activity(McBurnett, Lahey, Rathouz, & Loeber, 2000; Van Goozen, Matthys, Cohen-Kettenis, Thijssen, & Van Engeland, 1998). Traumatic experience during in-fancy is one of the factors that can cause permanent alterations to the HPAaxis by detrimentally affecting steroid receptor function in the hippocampusand the prefrontal cortex.
Piaget’s emphasis on embodiment and procedural knowledge (given phys-iological grounding by Swanson) is particularly clear in his description of theearliest play stages. Before stateable rules emerge, there are behavioral pat-terns. These arise first under the control of internal motivation, and then as aconsequence of social interaction. As the child progresses toward Piaget’s sec-ond stage—the first point at which the social world has any major impact—more complex understanding emerges. First, the child starts to copy him- orherself, using procedure to map procedure, at the initial but still embodiedstage of genuine representation. The child experiments, initially, using trialand error to attain his or her goals. Any successful action is immediately imi-tated and practiced. In this manner, the child builds a repertoire of voluntarilyaccessible, practiced, and automatized motor schema (Piaget, 1932). The imi-tative process then extends itself to interpersonal action, and the child be-comes capable of imitating others. It should be noted, however, that even atthis second, imitative stage, the child is still not genuinely playing with others.He or she engages in socially constructed and possibly sanctioned means ofplaying, but is neither trying to win nor attempting to unify the various modesof playing individually developed or imitated. Nonetheless, at this stage, pat-terned social interactions can arise spontaneously as a consequence of theinteraction of motivated and emotionally driven participants, who are con-stantly exchanging information about which actions and reactions are accept-able and mutually shaping shared behavior. Although such primary socialinteraction may look “rule governed” to an observer, because of its regularity,it is still instantiated at a purely procedural, implicit level.
It is possible, nonetheless, to see the emergence of a procedural moral-ity at this level. Panksepp (1998a) describes the manner in which rats learnto govern their pinning behavior: Stable patterns of play dominance, corre-sponding to the establishment of complex, socially modified motor behav-iors, rapidly emerge during R&T play. One rat ends up on top more often
Play and the Regulation of Aggression 141

during pinning, in repeatedly matched play pairs. However, if the dominantrat pins its playmate more than 70% of the time, the subordinate, who typi-cally initiates the play sequence, begins to ignore the victor, and playfulactivity gradually diminishes (Panksepp, 1998a). This means that the domi-nant rat must learn to respond carefully to the behavioral cues of the subor-dinate. If the subordinate breaks the shared play frame by escaping or bit-ing, as a consequence of undue frustration or anxiety, then its value as aplaymate rapidly decreases. Whether this is morality, or merely condition-ing, is beside the point. Such modulation still constitutes the beginning ofsocial behavior, laying the basis for the development of the higher-order mo-rality that keeps aggression properly regulated.
The child is alone, at the first stage of play, constructing the basic ele-ments of motor competence—grasping, letting go, extending and contractinglimbs—adapting him- or herself to his or her own motivations and their inter-actions in an increasingly complex world of objects. Then the child starts tocombine those actions, sequencing multiple motivated patterns of action, un-der the guidance of higher-order control systems. The child’s isolated mannerof being takes on a social aspect with the onset of R&T play, and he or she be-gins to establish socially modulated behavioral patterns. R&T play, in turn,shades into dramatic play, laid down in its most fundamental aspects over thesensorimotor substructure constructed first by the individual and then modi-fied by R&T play. As play becomes increasingly dramatic, increasingly ab-stract, the substructure for the highest stages of social cognition begins toestablish itself.
ABSTRACT PLAY AND THE COOPERATIVEESTABLISHMENT OF JOINT FICTIONAL WORLDS
During R&T play children use their bodies in playful dominance interactions,modifying and constructing motor schemas that take the other’s qualities intoaccount, learning to control anxiety, frustration, and the thrill of victory.Some of this control is best understood as inhibition. Whalen & Henker(1991) reported, for example, that theories of impaired behavioral control ap-peared more relevant to understanding the interpersonal problems of youngchildren with attention-deficit/hyperactivity disorder than theories of impairedsocial cognition. Hughes, White, Sharpen, and Dunn (2000) noted, likewise,that conduct disorder in 4-year-olds was associated with problems in execu-tive function (associated with inhibitory control), but not with problems inperformance on higher-order theory of mind tasks. Séguin, Zelazo, andTremblay (1999) associated physical aggression among preschoolers with def-icits in self-regulatory cognitive skills, and Séguin, Pihl, Harden, Tremblay,and Boulerice (1995) associated aggression among older children with im-paired executive or prefrontal function.
142 T H E D E V E L O P M E N T O F A G G R E S S I O N

However, theories of inhibition appear insufficient to account for morecomplex forms of aggression regulation, because they do not take into ac-count the emergence of cooperative behaviors and conceptions, designed tomaximize the utility of social being. These are better understood, perhaps, assophisticated alternatives to aggression, instead of mechanisms that inhibit,govern, or regulate aggression. To cooperate means to establish a mode ofoccupying the same space as other individuals in a manner that makes aggres-sion positively counterproductive. The groundwork for this ability appears tobe scaffolded at the sensorimotor level—mediated, perhaps, by R&T play—but the ability itself is something different: something more akin to the under-standing of narrative, drama, and fiction; something more like the develop-ment of explicit theory of mind.
Higher-order, more explicit, cooperative morality appears to emergearound Piaget’s (1932) third developmental stage, appearing at about age 7.At this stage, each child playing a given game starts to try to win—tries todominate the narrow hierarchy of achievement specified and constrained bythe rules of the game. At first glance, this appears as something essentiallycompetitive. However, for individual victory to occur, the modes of playingbetween children have to be unified so that all players share the same goal.This means that any disagreements about the game have to be resolved, andthat aggression emerging as a consequence of those disagreements must berendered unnecessary, before any attempt to play, let alone win, can even be-gin. Piaget notes that this more complex form of play appears to emerge pro-cedurally as well, rather than explicitly. Within the confines of a given groupgame, where each child can check him- or herself against the behavior of theothers, conflict-free, stable, patterned playing quickly emerges. However, ifthe playing children are separated and interviewed individually, they give dis-parate or even contradictory accounts of the emergent game’s formal “rules.”At this third stage, children are just beginning to map the contours of theirstructured social behavior into truly representational linguistic patterns. Theystill need the information provided by the presence of the others to maintainadherence to the predictable pattern of the game.
Once a joint social ritual or game is firmly established, however, its na-ture can be explicitly described and codified. As a consequence of emergingcognitive ability, and because the child can test his or her explicit verbalizablehypotheses against those of others, the patterns that actually constitute thegame and the explicit description of the game come into alignment. At thispoint, the child has successfully “mapped” his or her own socially modifiedsensorimotor output, as well as that of the play partners, and becomes a con-scious player (Piaget, 1932). Such convergence means that children playing thesame game come to inhabit the same fantasy-predicated fictional or dramaticworld. It appears to be the ability to establish such joint fictional habitation—more than alignment of motor patterning, more than inhibition of aggressiveresponding—that constitutes cooperation, and that allows for the modulation
Play and the Regulation of Aggression 143

of motivation and emotion toward some shared end. In a good game, every-one has fun. There is no need to be defensive. There are many opportunitiesfor joint gain. In consequence, there is little need for violence.
To understand the organization characterizing complex social play, it isimportant to note that innate motivational systems are not simple determinis-tic systems of drive. Nor do these systems merely set goals, although they dothat as well (Swanson, 2000). Instead, states of motivation serve as axioms orpredicates of experience, providing a delimited, bounded, but flexible, framefor perception, emotion, cognition, and action (Barsalou, 1983; Peterson &Flanders, 2002). With the establishment of such a frame, the more sophisti-cated goal-oriented individual can strive toward necessary goals in multiplenonreflexive manners, instead of mindlessly heading in a single, familiar, butpotentially counterproductive, direction. Such frames appear to be governedby low-level brain circuitry, primarily within the hypothalamus, but also de-pend on the extended processing resources of the limbic system and cortex.
Motivationally bounded states or frames are manifold in number, as thereare qualitatively different states of motivation (Rolls, 1999; Swanson, 2000),and they manifest themselves singly and sequentially, as processes of percep-tion, emotion, cognition, and action must be directed toward specified, lim-ited and sequential targets (Miller, 1956; Cowan, 2001). Each frame appearsto contain particularized conceptualizations of the current state of affairs, aswell as the desired end (Peterson, 1999; Peterson & Flanders, 2002). The indi-vidual or individuals operating within the confines of a given story move frompresent to future in a linear track. Two points define such a track, or line. Apresent position cannot be defined without a point of future contrast. Like-wise, a potential future cannot be evaluated—judged affectively as better—except in terms of a present position. A verbal description of such a conceptu-alization can be regarded as the most basic form of fiction—drama, narrative,and, not infrequently, game (Peterson, 1999).
The construction of a fiction-like frame, simplified by the momentarydomination of a single motivational state, helps specify what ends actionshould pursue and what phenomena might be considered as objects in thatpursuit (Hacking, 1999; Lakoff, 1987; Tranel, Logan, Frank, & Damasio,1997; Wittgenstein, 1968). The immature individual or child, pursuing his orher purely individual goals, acts and perceives in a solipsistic world, estab-lished in accordance with those goals. As the developing individual becomesmore complex, however, control over the contents of the goal-frameworkstarts to become more differentiated, so that although fundamental underlyingmotivational states still have access to it (in cases of mounting hunger orthirst, etc.), so do emergent systems of even more complex control. The hippo-campus, for example, allows for determination by context or situation(LeDoux, 1996), while the orbitofrontal–prefrontal cortex allows previouslyunvalued but learned goals to attain the status of true value (Krawczyk,2002). The dorsolateral–prefrontal cortex, finally, allows complex, abstractedframeworks to govern behavior, removing the developing individual from the
144 T H E D E V E L O P M E N T O F A G G R E S S I O N

incessant short-term demands of basic motivational states (Pochon et al.,2002). This sequentially emergent access by higher-order systems to motiva-tional framing appears to allow the individual to formulate goals that takemultiple states of motivation and the vagaries of external context into ac-count. “External context” also constitutes social being: the fact of motivatedothers, singly and in groups, who are also rank ordering their values hierarchi-cally, and implementing their motivational worlds, while constantly exchang-ing motivational and emotional information with one another.
The individual construction of a motivationally predicated frame or storyallows an individual to specify starting place, goal, objects of perception, andimplication for emotion, and to therefore deal with those bits of the world rel-evant to a particular need or desire. The joint construction of such a frame,such a story, integrates perception across individuals, placing them in the sameworld of objects at the same time it aligns their emotions. Such constructionallows more than one individual to inhabit the same experiential goal-directedspace and to thereby cooperate, voluntarily, to reach the goal and to maintainthe integrity of the space. This is the process by which fundamental agree-ments, nullifying the very necessity of aggression (rather than merely inhibit-ing its expression), come into being.
The specific circuitry mediating the establishment of such agreement hasbeen recently outlined, provisionally, at the prefrontal–cortical level. Rizzo-latti, Fogassi, and Gallese (2001) describe a localized class of visuomotor neu-rons constituting the “mirror neuron system.” First described in monkeys,mirror neurons are located in area F5 of the ventral premotor cortex. Thisarea contains neurons that code “goal-related” motor acts, such as graspingby hand and by mouth. Rizzolatti et al. (2001) state, in regard to these neu-rons: “Some of these cells are motor neurons, others also respond to visualstimuli. Some of them are activated by the presentation of three dimensionalobjects, whereas others—mirror neurons—require action observation for theiractivation” (p. 661). Mirror neurons, part of the system that uses motor out-put patterning as part of what occurs during an act of “perception,” have anumber of remarkable properties. First, they do not respond to the presence ofa motivationally significant object (say, an apple) in isolation. Nor do they re-spond to the sight of a conspecific engaged in a context-independent action(making a grasping action). But they do respond to the sight of a conspecificmaking a grasping action in the presence of a motivationally significant ob-ject. More to the point, their pattern of action when observing such a moti-vated sequence precisely matches their pattern of action when that sequence isundertaken by the observer.
Rizzolatti et al. (2001, p. 662) note that this neural “congruence is some-times extremely strict. In such cases, the effective motor action and the effec-tive observed action coincide both in terms of goal (for example, grasping)and in terms of how the goal is achieved (for example, precision grip).” Inother cases, however, the congruence is broader, matched more to the broadgoal of the action. The action of these less specific neurons is of even greater
Play and the Regulation of Aggression 145

interest, because their broader response pattern is not a sign of inaccuracy, butof increased sophistication: They are capable of “imitating” more approxi-mate patterns of motor output—those that
generalize the goal of the observed action across many instances of it. . . .[T]he novelty of these findings is the fact that, for the first time, a neuralmechanism that allows a direct matching between the visual description of anaction and its execution has been identified. Such a matching system consti-tutes a parsimonious solution to the problem of translating the results of thevisual analysis of an observed action… into an account that the individual isable to understand. (p. 662)
Such understanding appears complete and comprehensive, stretchingfrom the abstract, through the emotional, to the physical. The motor systemunderlying mirroring accepts neural inputs from systems governing sensation,cognition, and circadian state and has three primary divisions of output:somatic, endocrine, and neuroendocrine (Swanson, 2000). Descending controlsystems from higher-order, “limbic” structures such as the amygdala govern-ing affective response have also been well described (LeDoux, 1996). Further-more, area F5, which contains the mirror neurons, shares connectivity to infe-rior parietal lobe with area “a” of the superior temporal sulcus—a brain areathat is part of a circuit including the amygdala and orbitofrontal cortex(Amaral, Price, Pitkanen, & Carmichael, 1992). This combination of facts im-plies that the matching described by Rizzolatti extends past action patterns tothe associated emotional, motivational, cognitive, and neuroendocrine conse-quences and concomitants of those action patterns. To understand someonetherefore begins to appear more and more like walking a mile in his or herskin, rather than in his or her shoes.
This a hypothesis appears even more compelling once careful attention ispaid to the additional neuroanatomical and functional significance of area F5,the monkey homologue of Broca’s area in humans—the prefrontal area gov-erning voluntary speech. This anatomical equivalence suggests that the pri-mary purpose of verbal communication could well be the exchange of moti-vated patterns of action, instead of the description of the objective world(Peterson, 1999). Furthermore, it appears to be the association between themirror neuron system and speech that allows verbal communication to takeon its embodied, imitation-provoking meaning. These ideas appear criticallyimportant in regard to understanding and considering the importance of dra-matic social play and associated forms of narrative. Imagine that the develop-mental elaboration of the mirror neuron system allows the maturing child toembody the action and motivational states of those he or she observes first,directly, with more or less faithful and precise mimicry. Then imagine that theinterrelationship between the linguistic abilities of Broca’s area and the mirrorneuron circuitry allow communicating children to verbally instantiate sharedmotivated or goal-oriented states, not at the level of precise imitation, but at a
146 T H E D E V E L O P M E N T O F A G G R E S S I O N

higher, generalized state. This means that cooperating and communicatingchildren, engaging in pretend play, can jointly establish fictional worlds andthen coordinate their motivations, their actions, their emotions—their veryobject-perceptions—within those worlds. Finally, imagine that this process ofcoordination within the confines of a fictional world constitutes the process ofscaffolding that underlies (1) the understanding of narrative and fiction, intheir verbal forms, (2) the capacity to engage in large-scale, cooperative socialenterprises (which have a pronounced fiction-like aspect prior to their mani-festation as “completed projects” within the world), and (3) the ability to en-gage in understanding abstract and even more disembodied semantic thought.A plan, after all—including a shared plan—is nothing but the projection of acompelling fiction onto extant and agreed-upon objects and contexts. Thejoint establishment of such a plan, motivationally significant to all, emotion-ally gripping, eliminates the very necessity for conflict.
Sophisticated mother–child conversations about emotions appear to laythe groundwork both for children’s discussions of affective states and theemergence of the ability to adopt the perspective of another (reviewed brieflyin Oppenheim, Nir, Warren, & Emde, 1997). Oppenheim et al. (1997) havedemonstrated, as well, that preschool children able to engage in less emotion-ally disrupted and more coherent pretend play representations of a hypotheti-cal parent–child separation event with their mothers were also able to producesentence-stem completion narratives that were more prosocial, less aggressive,and more coherent, and were rated by their mothers as characterized by fewerbehavior problems concurrently and 1 year later. Wolf (1990) has suggested,likewise, that the development of narrative ability increases children’s abilityto view interpersonal situations from multiple perspectives, including those oftheir potential selves (their individual selves, in a different motivational state)and those of others. Hughes and Dunn (1997) provide direct support for thenotion of social collaboration in pretend play by (1) noting a strong positiverelationship between mental state talk and turn-taking and (2) noting that therelationship between mental state talk on the part of one child is as stronglyaffected by mental state talk on the part of his or her partner as it is by indi-vidual variability in verbal ability. Seja and Russ (1999) have demonstrated,similarly, that fantasy play ability is significantly related to emotional under-standing, over and above individual variation in verbal ability, and Taylor andCarlson (1997) have demonstrated that children whose fantasy and pretendplay abilities was more sophisticated also did better on theory of mind tasks,independently of their verbal intelligence. Finally, Hughes and Dunn (1997)note that theory of mind task performance is positively associated with refer-ences to mental state, independently of age.
Brown, Donelan-McCall, and Dunn (1996, p. 847) note, in this regard,that the maintenance of interactive games and the construction of fantasyworlds “must surely provide multiple opportunities for the fledgling theoristto appreciate the workings of the mind” (from Hughes & Dunn, 1997).Eckler and Weininger (1989) have paid particular attention to the elaboration
Play and the Regulation of Aggression 147

of such pretense worlds, pointing out that younger children tend toward “pre-episodic” and older children toward “episodic play,” which is very dramaticand story-like. Furthermore, these authors also clearly distinguish two aspectsof pretense, “setting-up” and “play”—processes that appear to correspondvery much to the establishment of a shared frame and action within thatframe, respectively. Children setting up a pretend play episode appear to beconstructing a joint fictional frame, a delimited subsection of the “realworld,” too complex to fully model. This delimited subsection has a consum-matory element, which is the complex goal of the fictional world (Rumelhart,1977) and also contains shared objects of perception—objects which, in thecase of pretense, may not even be there (as a child is perfectly capable of act-ing as if something is there, and as if an object that is there is in fact somethingelse). Children who are “setting up” a play episode therefore appear not onlyto be negotiating the nature of the actions that will occur in that space, butmutually regulating each other’s perception (as suggested previously). Pretendplay thus logically appears associated with talk about subjective mental stateat rates much higher than chance (Hughes & Dunn, 1997) and, more specifi-cally, with talk directed at another’s inferred mental state, desired or actual.
Children with autism, characterized by fundamental deficits in the sub-structural elements of social cognition, also fail to use social gaze, which typi-cally serves to specify the target of joint motivation and perception during em-pathy and joint attention tasks (Charman et al., 1997). This observation addsto the knowledge base already established demonstrating similar impairmentsamong autistic children in empathy, pretend play, and joint attention (re-viewed briefly in Charman et al., 1997; Baron-Cohen, Allen, & Gillberg,1992). The gaze dysfunction is particularly interesting, given that gaze moni-toring constitutes a procedural technique for determining intent, or goal-ori-entation, on the part of another, and is clearly part of the social–cognitive sys-tem that enables human beings to establish shared goal-oriented conceptualframes and associated behaviors and emotions (Adolphs, 2001; Hobson,1990). Autistic children also appear delayed, if not fundamentally impaired,in imitation (Charman et al., 1997).
The fact that individuals who have sustained left or right prefrontal dam-age in adulthood appear impaired on theory of mind tasks, even when theirdeficits on classic executive–cognitive tests have been controlled statistically,may also be relevant to understanding the emergent neuropsychological con-trol over shared motivation (Rowe, Bullock, Polkey, & Morris, 2001), andhelps to elucidate, physiologically, the distinction between voluntary coopera-tion and pure inhibition of aggression. Similar patterns of deficits also appearto characterize non-Alzheimer’s frontal variant frontotemporal dementia, fre-quently characterized by emergent suspiciousness, dysregulation of formerlysocialized behavior, and antisociality (Lough & Hodges, 2002; Lough, Greg-ory, & Hodges, 2001). In the case of fronto-temporal dementia, emergentantisociality also appears associated specifically with a breakdown of socialcognition, but not classic executive function (Lough et al., 2001) (perhaps as a
148 T H E D E V E L O P M E N T O F A G G R E S S I O N

consequence of deterioration in orbitofrontal–ventromedial circuitry, whichappears specifically activated during social cooperation [McCabe, Houser,Ryan, Smith, & Trouard, 2001; Rilling et al., 2002]).
Dunn and Hughes (2001) demonstrated a clear distinction between hard-to-manage children and normal children in regard to violent fantasy play, witha clearly higher proportion of the former group, both boys and girls, consis-tently engaging in pretend games that involved killing, fighting, or beating (de-spite the fact that no overall difference in frequency of pretend play as suchwas observed). Such interest in violent fantasy was itself related to poor exec-utive control and language ability, impaired theory of mind (related as well tolanguage ability), frequent antisocial behavior, displays of anger and refusal tohelp a friend, poor communication and coordination of play, more conflictwithin friendships, and decreases in empathic moral sensibility 2 years later.Hughes, Cutting, and Dunn (2001) then attempted to determine whetherconduct-disordered children were more likely to respond negatively to thethreat of losing a game because (1) they were more preoccupied with aggres-sion, in general, as indexed by violent themes in their pretend play, (2) theywere delayed or deviant in regard to reading the intentions of others (poten-tially manifesting a hostile attribution bias), or (3) they were characterized bya problem with executive function, resulting in difficulties in inhibiting inap-propriate behaviors. At age 4, violent pretend play theme frequency, theory ofmind performance, and executive function all appeared significantly associ-ated with negative behavior in response to threat of loss at age 5 at the zero-order correlation level of analysis, and all appeared to be of significant ornear-significant statistical import when entered simultaneously in a regressionanalysis. The picture of multidimensional causality portrayed by this studywas marred somewhat in its clarity by the results of longer-term follow-up: Byage 7, only the age-4 violent pretend play theme measure remained as a signif-icant predictor (accounting for 25% of the variance). Nonetheless, there issuggestive evidence that many different forms of regulation of aggression ex-ist, at very different levels of neurological instantiation.
Perhaps the effects of various pathologies, heightening the probability ofaggression, may be additive or even interactive: Aggressive children may lackbasic inhibition (first instantiated under normal conditions, as a consequenceof R&T play, governing motor output in the presence of others). They mayalso be impaired in regard to theory of mind and capacity to cooperate.Finally, as Crick and Dodge (1996) and Happe and Frith (1996) have sug-gested, such children may actively infer hostile intent on the part of others,and plot revenge (thus making even their limited capacity for social under-standing into something positively counterproductive). Jenkins and Green-baum (1999) have argued, in this regard, that disruptive children develop anovert “adversarial goal structure” that leads to frequent anger and aggression.Such a structure could be a generalized hypothesis such as “cooperation is im-possible” (a theme that makes antisociality more or less logically inevitable),or “individual victory trumps cooperative action” (a theme logically associ-
Play and the Regulation of Aggression 149

ated with narcissism, characterized by high levels of extraversion and low lev-els of agreeableness). Rose and Asher (1999) have noted, in keeping with thisgeneral hypothesis, that children who pursue the explicit goal of revenge to-ward a friend after conflict within a friendship were (1) more likely to use ag-gressive strategies, such as assertion of self-interest and hostility, (2) less likelyto use prosocial strategies, such as relationship maintenance and accommoda-tion or compromise, (3) more likely to lack friends, and (4) more likely tohave poor-quality friendships.
It is interesting to note more broadly in this regard, first, that socialstrategies or algorithms such as strike back once, then forgive and forget,are the most likely to win iterative repetitive tit-for-tat cooperative exchangegames such as Prisoner’s Dilemma (Wedekind & Milinski, 2000) and, second,that the combination of yoked goal-and-strategy implementation soundsvery much like a social game (not least because of the emergence of con-stant reciprocity). It is also very relevant in regard to the potential develop-mental origins of an essentially adversarial game that maltreated childrenare more likely than their nonmaltreated peers to develop more negative,constricted, and less coherent narrative or story-like representations of theircaregivers. These more negative representations, although potentially accu-rate in the circumstances in which they emerged, generalize poorly and areassociated with emotional dysregulation, aggression, and peer rejection. Pos-itive and coherent representations of caregivers, by contrast, are relatedboth to prosocial behavior and to preference by peers (Shields, Ryan, &Cicchetti, 2001).
Maltreated children appear to see the social surround as “angry, malevo-lent, punitive, exploitative, and conflictual” (reviewed briefly in Shields et al.,2001). It is particularly interesting, therefore, to note that R&T play initiatedwith popular, prosocial young school-age children generally turns into playwith rules or more advanced pretend play, whereas such play initiated withaggressive children degenerates rapidly into violence (Pellegrini & Smith,1998). Given the aggressive child’s vengeful and mistrustful view of the world,it is not unreasonable to suppose that exploratory R&T play forays on thepart of one child with regard to another allow that child to assess the basic so-cial strategies employed by that other—to investigate, very rapidly and di-rectly, whether the other has established a basic capacity for inhibition, andcan therefore be trusted not to harm, or if he or she is hostile and aggressive,and unfit for continued interaction. Finally it is useful to note that violentvideo game play—a form of scaffolded pretend play that highlights themes ofrevenge and destruction—does in fact appear causally associated with aggres-sive behavior, thoughts and feelings, and with a decrease in prosociality(Bushman & Anderson, 2002).
Kochanska and colleagues (Murray & Kochanska, 2002) have recentlydeveloped a statistically coherent and longitudinally stable battery of “effort-ful control tasks,” which measure the capacity to delay gratification and tovoluntarily suppress or initiate actions (performance on which can be assessed
150 T H E D E V E L O P M E N T O F A G G R E S S I O N

as early as 2.5 years). Lesser performance on this task battery was associatedwith some elements of conduct-disordered behavior, such as impaired atten-tional control. Forman and Kochanska (2001) have also demonstrated thatchildren who were more imitative and responsive during pretend-play se-quences modeled by mothers were also more positively responsive to maternalcontrol, and less likely to manifest noncompliance in a typical discipline con-text. This appears to mean that children who are more capable of adopting ashared frame of reference in a play-like context are also those who are morewilling to “play the right game” when attention is called to their transgres-sions or rule-breaking behaviors. Stipek, Recchia, and McClintic (1992) note,in this regard, that young children do not really distinguish between teachingand disciplinary contexts, reacting with pride to doing well and with shame todoing badly in both contexts. Kochanska, Aksan, and Koenig (1995) andKochanska, Tjebkes, and Forman (1998) have therefore proposed the conceptof committed compliance, something opposite to that of “the adversarial goalstructure,” distinguished from externally imposed obedience by its “enthusias-tic and self-sustaining quality, unmediated by ongoing parental control”(Forman & Kochanska, 2001, p. 199), and described further as “visible em-bracing of the parent’s agenda” and as “a behavioral tendency consistentacross situations shown by the child’s continued restraint when the [parent] isno longer present.” This sounds very much like the adoption of a shared moti-vational frame of reference, constructed on the basis of true social coopera-tion, and not merely the inhibition of aggression.
CONCLUSION
First, it is clear that both Hobbes and Rousseau were correct. The individualbrings to the world a set of inborn motivations, including those that underlieaggression, and these motivations are brought under control—or not—as aconsequence of socialization. This control appears at least twofold. The directinhibition and regulation of aggression appears established as a consequenceof R&T play and also appears associated, in principle, with the developmentof some forms of executive control. The formal adoption of a prosocial stancetoward the world, by contrast, mediated by emergent trust in the trustworthi-ness of self and other, culminates not so much in the capacity to obey rulesand to stay on track (associated perhaps with inhibitory and executive con-trol), but in the willingness to voluntarily enter into complex, cooperative so-cial games with others, mediated by shared, goal-directed frames of reference(Peterson, 1999; Peterson & Flanders, 2002). These tendencies toward the so-cial good seem as predicated on innate capability, interest, and inborn empa-thy as do the tendencies toward aggression. There is no shortage of evidencefor innate human good, and much suggestion that it might even reasonably beconsidered the norm. Furthermore, it is clear that pathological socializationexperiences, first in the context of the family and second in the context of
Play and the Regulation of Aggression 151

early peer experiences (variants of the institutional sickness described byRousseau) can produce and then reinforce in a child the conviction that theworld is a cruel and sadistic place, fit only for interpretation through lensescolored by the desire for revenge.
Finally, it is clear that complex processes of play, beginning with R&Tplay and culminating in the production of sophisticated, abstract sociallyshared frames of reference, play an important role in the modulation of ag-gression, both in regard to its inhibition and in regard to its integration intofully functional individual and social identities. The first problem that lifepresents, so to speak, is the necessity of satisfying basic motivational states.The next emergent problem is the necessity to construct and integrate tech-niques designed to satisfy these motivational states across different states,across different time frames and in a wide variety of contexts. Given the in-tensely social nature of the natural human environment, this problem of inte-gration must eventually expand to include the other, the motivated other.
REFERENCES
Abbott, D. H., Keverne, E. B., Bercovitch, F. B., Shively, C. A., Mendoza, S. P., Saltzman,W., Snowdon, C. T., Ziegler, T. E., Banjevic, M., Garland, T., Jr., & Sapolsky, R. M.(2003). Are subordinates always stressed? Hormones and Behavior, 43, 67–82.
Adolphs R. (2001). The neurobiology of social cognition. Current Opinions in Neuro-biology, 11, 231–39.
Amaral, D. G., Price, J. L., Pitkanen, A., & Carmichael, S. T. (1992) Anatomical organi-zation of the primate amygdaloid complex. In J. P. Aggleton (Ed.), The amygdala(pp. 1–66). New York: Wiley-Liss.
Asher, S. R., & Coie, J. D. (1990). Peer rejection in childhood. New York: CambridgeUniversity Press.
Baron-Cohen, S., Allen, J., & Gillberg, C. (1992). Can autism be detected at 18 months?British Journal of Psychiatry, 161, 839–843.
Barsalou, L. W. (1983). Ad hoc categories. Memory and Cognition, 11, 211–227.Bates, J. E., Bayles, K., Bennett, D. S., Ridge, B., & Brown, M. M. (1991). Origins of
externalizing behavior problems at eight years of age. In D. J. Pepler & K. H. Rubin(Eds.), The development and treatment of childhood aggression (pp. 93–120).Hillsdale, NJ: Erlbaum.
Blurton Jones, N. (1972). Categories of child–child interaction. In N. B. Jones (Ed.),Ethological studies of child behavior (pp. 97–129). New York: Cambridge Univer-sity Press.
Boulton, M. J. (1991). A comparison of structural and contextual features of middleschool children’s playful and aggressive fighting. Ethology and Sociobiology, 12,119–145.
Boulton, M. J., & Smith, P. K. (1992). The social nature of play fighting and play chasing.In J. H. Barkow (Eds.), The adapted mind. New York: Oxford University Press.
Brown, J. R., Donelan-McCall, N., & Dunn, J. (1996). Why talk about mental states?Child Development, 67, 836–849.
Bruner, J. S. (1976). Play: Its role in development and evolution. New York: Basic Books.
152 T H E D E V E L O P M E N T O F A G G R E S S I O N

Bushman, B. J., & Anderson, C. A. (2002). Violent video games and hostile expectations.Personality and Social Psychology Bulletin, 28, 1679–1686.
Bushman, B. J., & Baumeister, R. F. (1998). Threatened egotism, narcissism, self-esteem,and direct and displaced aggression. Journal of Personality and Social Psychology,75, 219–229.
Charman, T., Swettenham, J., Baron-Cohen, S., Cox, A., Baird, G., & Drew, A. (1997).Infants with autism. Developmental Psychology, 33, 284–294.
Coie, J. D., & Dodge, K. A. (1998). Aggression and antisocial behavior. In W. Damon &N. Eisenberg (Eds.), Handbook of child psychology (Vol. 3, pp. 779–862). Toronto:Wiley.
Costabile, A., Smith, P. K., Matheson, L., Aston, J., Hunter, T., & Boulton, M. (1991).Cross-national comparison of how children distinguish serious and playful fighting.Developmental Psychology, 27, 881–887.
Cowan, N. (2001). The magical number 4 in short-term memory. Brain and BehavioralSciences, 24, 154–176.
Crick, N. R., & Dodge, K. A. (1996). Social information-processing mechanisms in reac-tive and proactive aggression. Child Development, 67, 993–1002.
De Waal, F. (1989a). Chimpanzee politics. London: Johns Hopkins University Press.De Waal, F. (1989b). Peacemaking among primates. New York: Harvard University
Press.De Waal, F. D. (1996). Good natured. Cambridge: Harvard University Press.Dunn, J., & Hughes, C. (2001). “I got some swords and you’re dead!” Child Develop-
ment, 72, 491–505.Eckler, J. A., & Weininger, O. (1989). Structural parallels between pretend play and nar-
ratives. Developmental Psychology, 25, 736–743.Field, T. (1998). Maternal depression effects on infants and early interventions. Preven-
tive Medicine, 27, 200–203.Fischer, D. H. (1989). Albion’s seed. New York: Oxford University Press.Forman, D. R., & Kochanska, G. (2001). Viewing imitation as child responsiveness: A
link between teaching and socialization domains of socialization. DevelopmentalPsychology, 37, 198–206.
Gregg, T. R, & Siegel, A. (2001). Brain structures and neurotransmitters regulating ag-gression in cats: Implications for human aggression. Progress in Neuropsycho-pharmacology and Biological Psychiatry, 25, 91–140.
Hacking, I. (1999). The social construction of what? Cambridge: Harvard UniversityPress.
Happe, F., & Frith, U. (1996). The neuropsychology of autism. Brain, 119, 1377–1400.Harlow, H., & Harlow, M. (1965). The affectional systems. Behavior of Non-Human
Primates, 2, 287–334.Hobbes, T. (1909). The leviathan. Oxford, UK: Clarendon Press. (Original work pub-
lished 1651)Hobson, R. P. (1990). Concerning knowledge of mental states. British Journal of Medical
Psychology, 63, 199–213.Hughes, C., Cutting, A. L., & Dunn, J. (2001). Acting nasty in the face of failure? Journal
of Abnormal Child Psychology, 29, 403–416.Hughes, C., & Dunn, J. (1997). “Pretend you didn’t know. ” Cognitive Development, 12,
381–403.Hughes, C., White, A., Sharpen, J., & Dunn, J. (2000). Antisocial, angry, and unsympa-
thetic. Journal of Child Psychology and Psychiatry, 41, 169–179.
Play and the Regulation of Aggression 153

Jenkins, J. M., & Greenbaum, R. (1999). Intention and emotion in child psycho-pathology. In P. D. Zelazo, J. W. Astington, & D. R. Olson (Eds.), Developing theo-ries of intention (pp. 269–291). Mahwah, NJ: Erlbaum.
Joffe, T. H., & Dunbar, R. I. (1997) Visual and socio-cognitive information processing inprimate brain evolution. Proceedings of the Royal Society of London: BiologicalScience, 264, 1303–1307.
Kochanska, G., Aksan, N., & Koenig, A. L. (1995). A longitudinal study of the roots ofpreschoolers’ conscience: Committed compliance and emerging internalization.Child Development, 66, 1752–1769.
Kochanska, G., Tjebkes, T. L., & Forman, D. R. (1998). Children’s emerging regulationof conduct: Restraint, compliance, and internalization from infancy to the secondyear. Child Development, 69, 1378–1389.
Krawczyk, D. C. (2002). Contributions of the prefrontal cortex to the neural basisof human decision making. Neuroscience and Biobehavioral Reviews, 26, 631–664.
Lakoff, G. (1987). Women, fire, and dangerous things. Chicago: University of ChicagoPress.
Lande, J., Scarr, S. W., & Gunzenhauser, N. (1989). Caring for children. Hillsdale, NJ:Erlbaum.
LeDoux, J. (1996) The emotional brain. New York: Touchstone Press.Lough, S., Gregory, C., & Hodges, J. R. (2001). Dissociation of social cognition and ex-
ecutive function in frontal variant frontotemporal dementia. Neurocase, 7, 123–130.
Lough, S., & Hodges, J. R. (2002). Measuring and modifying abnormal social cognitionin frontal variant frontotemporal dementia. Journal of Psychosomatic Response,53, 639–646.
Masserman, J. H., Wechkin, S., & Terris, W. (1964). “Altruistic” behavior in rhesus mon-keys. American Journal of Psychiatry, 121, 584–585.
McBurnett, K., Lahey, B. B., Rathouz, P. J., & Loeber, R. (2000). Low salivary cortisoland persistent aggression in boys referred for disruptive behavior. Archives of Gen-eral Psychiatry, 57, 38–43.
McCabe, K., Houser, D., Ryan, L., Smith, V., & Trouard, T. (2001). A functional imagingstudy of cooperation in two-person reciprocal exchange. Proceedings of the Na-tional Academy of Sciences USA, 98, 11832–11835.
Meaney, M. J., Aitken, D. H., Van Berkel, C., Bhatnagar, S., & Sapolsky, R. M. (1988).Effect of neonatal handling on age-related impairments associated with the hippo-campus. Science, 239, 766–768.
Miller, G. A. (1956). The magical number seven, plus or minus two. Psychological Re-view, 63, 81–97.
Miller, P. A., Eisenberg, N., Fabes, R. A., & Shell, R. (1996). Relations of moral reasoningand vicarious emotion to young children’s prosocial behavior toward peers andadults. Developmental Psychology, 32, 210–219.
Murray, K. T., & Kochanska, G. (2002). Effortful control: Factor structure and relationto externalizing and internalizing behaviors. Journal of Abnormal Child Psychol-ogy, 30, 503–514.
Nagin, D., & Tremblay, R. E. (1999). Trajectories of boys’ physical aggression, opposi-tion, and hyperactivity on the path to physically violent and non violent juvenile de-linquency. Child Development, 70, 1181–1196.
154 T H E D E V E L O P M E N T O F A G G R E S S I O N

Oatley, K. (1999). Why fiction may be twice as true as fact. Review of General Psychol-ogy, 3, 101–117.
Oatley, K., & Johnson-Laird, P. N. (1987). Towards a cognitive theory of emotion. Cog-nition and Emotion, 1, 29–50.
Olweus, D. (1994). Bullying at school. New York: Blackwell.Oppenheim, D., Nir, A., Warren, S., & Emde, R. N. (1997). Emotion regulation in
mother–child narrative co-construction. Developmental Psychology, 33, 284–294.Pagani, L., Boulerice, B., Tremblay, R. E., & Vitaro, F. (1997). Behavioral development in
children of divorce and remarriage. Journal of Child Psychology and Psychiatry, 38,769–781.
Panksepp, J. (1998a). Affective neuroscience. New York: Oxford.Panksepp, J. (1998b). Attention deficit hyperactivity disorder, psychostimulants, and in-
tolerance of childhood playfulness: A tragedy in the making? Current Directions inPsychological Science, 7, 91–98.
Patterson, G. R., Reid, J. B., & Dishion, T. J. (1992). Antisocial boys. Eugene, OR:Castalia.
Pellegrini, A. D., & Smith, P. K. (1998). Physical activity play. Child Development, 69,577–598.
Pellis, S. M., & Iwaniuk, A. N. (2000). Comparative analyses of the role of postnatal de-velopment on the expression of play fighting. Developmental Psychobiology, 36,136–147.
Peterson, J. B. (1999). Maps of meaning: The architecture of belief. New York: Routledge.Peterson J. B., & Flanders, J. L. (2002) Complexity management theory. Cortex, 38,
429–458.Peterson, J. B., & Shane, M. (2004). The functional neuroanatomy and psycho-
pharmacology of predatory and defensive aggression. In J. McCord (Ed.), Beyondempiricism: Vol. 13. Advances in criminological theory. Picataway, NJ: TransactionBooks.
Piaget, J. (1932). The moral judgement of the child. London: Kegan, Paul, Trench,Trubner.
Pochon, J. B., Levy, R., Fossati, P., Lehericy, S., Poline, J. B., Pillon, B., Le Bihan, D., &Dubois, B. (2002). The neural system that bridges reward and cognition in humans.Proceedings from the National Academy of Sciences USA, 99, 5669–5674.
Preston, S. D., & De Waal, F. B. M. (2002). Empathy. Behavioral Brain Research, 25, 1–72.Rice, G. E. J. (1964). Aiding behavior vs. fear in the albino rat. Psychological Record, 14,
165–170.Rice, G. E. J., & Gainer, P. (1962). Altruism in the albino rat. Journal of Comparative and
Physiological Psychology, 55, 123–125.Rilling, J., Gutman, D., Zeh, T., Pagnoni, G., Berns, G., & Kilts, C. (2002). A neural basis
for social cooperation. Neuron, 35, 395–405.Rizzolatti, G., Fogassi, L., & Gallese, V. (2001). Neurophysiological mechanisms under-
lying the understanding and imitation of action. Nature Reviews: Neuroscience, 2,661–670.
Rogers, C. R. (1989). The Carl Rogers reader. New York: Houghton Mifflin.Rolls, E. (1999). The brain and emotion. New York: Oxford University Press.Roopnarine, J. L., Hooper, F. H., Ahmeduzzaman, M., & Pollack, B. (1993). Gentle play
partners. In K. MacDonald (Ed.), Parent–child play (pp. 287–304). Albany, NY:SUNY Press.
Play and the Regulation of Aggression 155

Rose, A. J., & Asher, S. R. (1999). Children’s goals and strategies in response to conflictswithin a friendship. Developmental Psychology, 35, 69–79.
Rousseau, J. J. (1974). The essential Rousseau: The social contract, discourse on the artsand sciences. The creed of a savoyard priest. New York: New America Library.(Original work published 1750)
Rowe, A. D., Bullock, P. R., Polkey, C. E., & Morris, R. G. (2001). “Theory of mind” im-pairments and their relationship to executive functioning following frontal lobe ex-cisions. Brain, 124, 600–616.
Rumelhart, D. E. (1977). Understanding and summarizing brief stories. In D. LaBerge &J. Samuels (Eds.), Basic process in reading: Perception and comprehension (pp.265–303). Hillsdale, NJ: Erlbaum.
Séguin, J. R., Pihl, R. O., Harden, P. W., Tremblay, R. E., & Boulerice, B. (1995). Cogni-tive and neuropsychological characteristics of physically aggressive boys. Journal ofAbnormal Psychology, 104, 614–624.
Séguin, J. R., Zelazo, P. D., & Tremblay, R. E. (1999). Executive function in early physi-cal aggression and hyperactivity: Progress report on a longitudinal study of pre-schoolers. Paper presented at the inaugural meeting of the Cognitive DevelopmentSociety, Chapel Hill, NC.
Seja, A. L., & Russ, S. W. (1999). Children’s fantasy play and emotional understanding.Journal of Clinical Child Psychology, 28, 269–277.
Shields, A., Ryan, R. M., & Cicchetti, D. (2001). Narrative representations of caregiversand emotion dysregulation as predictors of maltreated children’s rejection by peers.Developmental Psychology, 37, 321–337.
Sibley, C. G., & Ahlquist, J. E. (1984). The phylogeny of the hominoid primates, asindicated by DNA-DNA hybridization. Journal of Molecular Evolution, 20, 2–15.
Smith, P. K., & Boulton, M. (1990). Rough-and-tumble play, aggression and dominance.Human Development, 33, 271–282.
Smith, P. K., Hunter, T., Carvalho, A. M., & Costabile, A. (1992). Children’s perceptionsof playfighting, playchasing and real fighting. Social Development, 1, 211–229.
Stipek, D., Recchia, S., & McClintic, S. (1992). Self-evaluation in young children. Mono-graphs of the Society for Research on Child Development, 57, 1–98.
Swanson, L. W. (2000). Cerebral hemisphere regulation of motivated behavior. Brain Re-search, 886, 113–164.
Swanson, L. W. (2003). Brain architecture. New York: Oxford University Press.Taylor, M., & Carlson, S. M. (1997). The relationship between individual differences in
fantasy and theory of mind. Child Development, 68, 436–455.Tranel, D., Logan, C. G., Frank, R. J., & Damasio, A. R. (1997). Explaining category-
related effects in the retrieval of conceptual and lexical knowledge for concrete enti-ties. Neuropsychologia, 35, 1329–1139.
Van Goozen, S. H. M., Matthys, W., Cohen-Kettenis, P. T., Thijssen, J. H. H., & vanEngeland, H. (1998). Adrenal androgens and aggression in conduct disorderedprepubertal boys and normal controls. Biological Psychiatry, 43, 156–158.
Virgin, C. E., Jr., & Sapolsky, R. M. (1997). Styles of male social behavior and their endo-crine correlates among low-ranking baboons. American Journal of Primatology,42, 25–39.
Vitiello, B., & Stoff, D. M. (1997) Subtypes of aggression and their relevance to child psy-chiatry. Journal of the American Academy of Child and Adolescent Psychiatry, 36,307–315.
156 T H E D E V E L O P M E N T O F A G G R E S S I O N

Wedekind, C., & Milinski, M. (2000). Cooperation through image scoring in humans.Science, 288, 850–852.
Whalen, C. K., & Henker, B. (1991). Social impact of stimulant treatment for hyperactivechildren. Journal of Learning Disabilities, 24, 231–241.
Wilson, M., & Daly, M. (1997). Life expectancy, economic inequality, homicide, and re-productive timing in Chicago neighbourhoods. British Medical Journal, 314, 1271–1274.
Wittgenstein, L. (1968). Philosophical investigations. New York: Macmillan.Wolf, D. P. (1990). Being of several minds: Voices and versions of the self in early child-
hood. In D. Cicchetti & M. Beeghly (Eds.), The self in transition: Infancy to child-hood (pp. 183–212). Chicago: University of Chicago Press.
Wrangham, R., & Peterson, D. (1996). Demonic males. New York: Houghton Mifflin.Yoerg, S. I. (1991). Social feeding reverses learned flavor aversions in spotted hyenas.
Journal of Comparative Psychology, 105, 185–189.Zahn-Waxler, C., Radke-Yarrow, M., & Brady-Smith, J. (1977) Perspective-taking and
prosocial behavior. Developmental Psychology, 13, 87–88.Zahn-Waxler, C., Radke-Yarrow, M., & King, R. A. (1979) Child rearing and children’s
prosocial initiations toward victims of distress. Child Development, 50, 319–330.
Play and the Regulation of Aggression 157

THE DEVELOPMENT OF AGGRESSIONIndirect Aggression among Humans
8
Indirect Aggression among HumansSocial Construct or Evolutionary Adaptation?
TRACY VAILLANCOURT
During the course of the 20th century, a considerable growth in knowledgetook place concerning the functions, correlates, and developmental patterns ofaggression among humans. The bulk of these studies focused on physical ag-gression, which was found to be used more by males than females (e.g.,Loeber & Hay, 1997; Maccoby & Jacklin, 1974, 1980; Hyde, 1984) and todecrease with age (e.g., Broidy et al., 2003; Cote et al., 2004; Kingston &Prior, 1995; Loeber & Stouthamer-Loeber, 1998; Tremblay et al., 1999). Asthe turn of the century approached, scientists demonstrated that aggression inhumans extended beyond its physical form and beyond the Y chromosome.Indeed, a wave of new studies appeared, showing that females were also capa-ble of aggressing against another, but that they expressed aggression in a morecovert manner (i.e., indirect aggression), taking the form of rumor spreading,peer group exclusion, and silent treatments, to name a few strategies (e.g.,Björkqvist, Lagerspetz, & Kaukiainen, 1992a; Björkqvist, Osterman, &Kaukiainen, 1992b; Cairns, Cairns, Neckerman, Ferguson, & Gariepy, 1989;Crick, 1995, 1996; Crick & Grotpeter, 1995; Lagerspetz, Björkqvist, &Peltonen, 1988; Galen & Underwood, 1997). These studies also revealed thatindirect aggression, in contrast to physical aggression, increased with age (e.g.,Vaillancourt et al., 2004; Vaillancourt, Brendgen, Boivin, & Tremblay, 2003b)and was associated with its own unique set of correlates/predictors (seeUnderwood, 2003, for review).
Although our knowledge relating to indirect aggression is growing, thegreater part of this knowledge is informed by an ontogenetic perspective, withlittle attention paid to its possible evolutionary significance. Twenty-five years
158

ago, Robert Cairns (1979) noted that “the phenomena of evolutionary adap-tations can no longer be ignored in developmental analyses, even if they donot readily fit existing ideas” (p. 370). This argument stemmed from the ideathat social patterns can be “seen as developing in an individual’s life time(over years) or they may be viewed as evolving over a species ‘lifetime’ (overthousands or millions of years)” (p. 370). Through the integration of researchfrom a variety of different fields such as psychology, sociology, anthropology,ethology, and biology, the purpose of this chapter is to put forth a phylogen-etic argument concerning the function of indirect aggression among humanfemales. Toward this end, indirect aggression is first defined, followed by a re-view of the correlates of indirect aggression, with particular focus on age- andsex-related differences associated with its use. Next, early theories of aggres-sion are presented to place in context the current hypotheses concerningfemales’ use of indirect aggression. Finally, an evolutionary hypothesis is pro-posed on why females use indirect aggression.
WHAT IS INDIRECT AGGRESSION?
To understand what we have learned and what we need to learn,we first need to agree on the topic.
—TREMBLAY (2000, p. 130)
Naming the behavior that entails the use of socially conniving acts such asgetting others to dislike a person, becoming friends with another as a formof revenge, telling a person’s secrets to another, saying bad things about aperson behind his or her back, telling others to avoid a person, using deri-sive body language, and the like, should, in theory, be relatively straightfor-ward. The problem, however, is rooted in that fact that there are currentlythree terms being employed to describe this phenomenon: social aggressionis used by Underwood and colleagues (e.g., Galen & Underwood, 1997) todescribe behavior that is “directed toward damaging another’s self-esteem,social status, or both” (p. 589; see also Cairns et al., 1989; Xie, Cairns, &Cairns, 2002a); relational aggression is used by Crick and colleagues (Crick,1995, 1996; Crick & Grotpeter, 1995) to describe behavior that harms others“through purposeful manipulation and damage of their peer relationships”(Crick & Grotpeter, 1995, p. 711); and indirect aggression is used byBjörkqvist and colleagues to describe a set of behaviors that are socially ma-nipulative and circuitous in nature, such as spreading invidious rumorsabout another person (e.g., Björkqvist et al., 1992a; Björkqvist et al.,1992b; Lagerspetz et al., 1988).
Although these three “types” of aggression seem to overlap considerably(Björkqvist, 2001), it has nevertheless been argued that they are in fact con-ceptually different from one another (Underwood, 2003; Underwood, Galen,& Paquette, 2001). For instance, Underwood and colleagues (Underwood,
Indirect Aggression among Humans 159

2003; Underwood et al., 2001) argue that social aggression is different fromrelational aggression in that social aggression involves damaging a person’sself-esteem and/or social status, whereas relational aggression involves damag-ing a person’s peer relations. Indirect aggression is similar to relational aggres-sion in that it involves the manipulation of peers and damage to these liaisons,but differs in that it includes only behaviors that are covert. Both social ag-gression and relational aggression include a mélange of covert and overt be-haviors. These nuances aside, what these so-called distinct types of aggressionhave in common is that they involve hurtful behaviors that are socially schem-ing. Using different terms to describe a certain phenomenon is problematic(Björkqvist, 2001; Vaillancourt & Hymel, 2003) in that it impedes progress inthe area by confusing, among others, new scholars coming to the field.
According to Walter Mischel, researchers should create and go beyondtheories that have already been developed rather than renaming them (Ra-basca, 2000). In an attempt to circumvent another “renaming of the term” in-direct aggression is used throughout this chapter in recognition of the pioneerwork by Feshbach (1969, 1971) and Björkqvist and colleagues (Björkqvist etal., 1992a, 1992b; Lagerspetz et al., 1988) in this area of study. Once a studyhas been conducted demonstrating that social, relational, and indirect aggres-sion are distinct forms of aggression, this position can be reevaluated. How-ever, it is likely that this stance will remain, in that this future study will notonly need to demonstrate the statistical uniqueness of these “different” typesof aggression, but will also have to demonstrate that social, relational, and in-direct aggression are differentially associated with outcomes.
WHO USES INDIRECT AGGRESSION?
Age Differences
It has been suggested that as children age their mode of aggressing against an-other changes and that the use of indirect aggression is the more sophisticatedform of hurtful behavior (Björkqvist, 1994; Björkqvist et al., 1992a, 1992b;Lagespetz et al., 1988). Specifically, Björkqvist and colleagues hypothesizethat children switch from being physically aggressive to being verbally aggres-sive as their language skills develop, and, in turn, they change from being ver-bally aggressive to being indirectly aggressive as their social-cognitive skillsadvance. Cross-sectional and longitudinal studies of aggression appear to sup-port this claim. For example, in contrast to the use of physical aggression inhumans, which has been shown to peak at around the age of 30 months anddecline steadily thereafter (e.g., Broidy et al., 2003; Cote et al., 2004; Kingston& Prior, 1995; Loeber & Stouthamer-Loeber, 1998; Tremblay et al., 1999), theuse of indirect aggression has been shown to increase with age (Björkqvist etal., 1992a; Björkqvist et al., 1992b; Cairns et al., 1989; Osterman et al., 1998;Tremblay et al., 1996; Vaillancourt et al., 2004). Some have suggested that in-direct aggression peaks during early adolescence (Björkqvist et al., 1992a;
160 T H E D E V E L O P M E N T O F A G G R E S S I O N

Björkqvist et al., 1992b), and others have argued that it continues to rise wellinto late adolescence (e.g., Owens, 1996). To date, precious few studies haveexamined, longitudinally, the development of indirect aggression, and the fewstudies that have, focused primarily on children and preadolescents (Cairns etal., 1989; Vaillancourt et al., 2003b; Vaillancourt et al., 2004). Thus, at thispoint in time, a complete picture of the developmental sequence of indirect ag-gression is not available. Nevertheless, based on these few longitudinal stud-ies, as well as on cross-sectional results, it appears that as children age, theiruse of indirect aggression increases (Björkqvist et al., 1992a; Björkqvist et al.,1992b; Osterman et al., 1998; Tremblay et al., 1996; Vaillancourt et al.,2003b; Vaillancourt et al., 2004). However, it is still unclear as to when indi-rect aggression is at its highest or lowest across the lifespan. Currently, what isknown is that humans as young as preschoolers (e.g., Crick, Casas, &Mosher, 1997; McNeilly-Choque, Hart, Robinson, Nelson, & Olsen, 1996;Vaillancourt et al., 2003b) and as old as seniors (Walker, Richardson, &Green, 2000) use indirect aggression.
Sex Differences
Although at present there is little information concerning developmentalchanges associated with the use of indirect aggression, there is a considerableamount of information regarding sex differences in the expression of aggres-sion. These studies have consistently demonstrated that females are typicallymore indirectly aggressive than males, whereas males are invariably morephysically aggressive than females (Craig, Peters & Willms, 2002; Crick,1995, 1996; Crick et al., 1997; Crick & Grotpeter, 1995; Björkqvist et al.,1992a; Björkqvist et al., 1992b; Feshbach, 1969, 1971; French, Jansen,Pidada, 2002; Hines & Fry, 1994; McNeilly-Choque et al., 1996; Lagerspetzet al., 1988; Osterman et al., 1998; Prinstein & Cillessen, 2003; Tremblay etal., 1996; Vaillancourt et al. 2003b). Although this sex difference in the ex-pression of indirect aggression has been reported often, it is important to notethat in some studies no sex differences in the use of indirect aggression havebeen found (Hart, Nelson, Robinson, Olsen, & McNeilly-Choque, 1998; Rys& Bear, 1997; Willoughby, Kupersmidt, & Bryant, 2001), and in others,males have been shown to be more indirectly aggressive than females (David& Kistner, 2000; Hennington, Hughes, Cavell, & Thompson, 1998; Tomada& Schneider, 1997). It is worthwhile to clarify these discrepancies concerningwho uses indirect aggression more—females or males—and what these find-ings mean. The fact that a few studies have shown males to be more indirectlyaggressive than females does not detract from the overarching finding, whichis that females’ preferred mode of aggressing is indirect. Further, the fact thatseveral studies have demonstrated that females use indirect aggression morethan males does not mean that females are globally more aggressive than, orequally aggressive as, males. Indeed, the literature to date points in a definitivedirection, suggesting that males use aggression more than females. However,
Indirect Aggression among Humans 161

these findings highlight two important points that have historically been over-looked in the aggression literature: (1) females’ use of aggression is not a rarephenomenon, and (2) when females do aggress, they tend to do so in a circu-itous manner. The evolutionary significance of these differences is consideredshortly.
WHAT ARE OTHER CORRELATES OF INDIRECT AGGRESSION?
Although physical aggression has been linked to an extensive list of individual(e.g., Lahey, McBurnett, & Loeber, 2000; Moore, Scarpa, & Raine, 2002;Plomin, 1983; Raine, 1993; Schaal, Tremblay, Soussignan, & Susman, 1996;Tremblay & LeMarquand, 2001), early environmental (e.g., Arseneault,Tremblay, Boulerice, & Saucier, 2002; Fergusson, Woodward, & Horwood,1998; Raine, 2002), familial (e.g., Keenan & Shaw, 1994; Lykken, 2001;Tremblay, 1999), and parent–child factors (see Coie & Dodge, 1998 andPatterson, 1995, for reviews), the study of indirect aggression is still in its in-fancy. Accordingly, there have been only a small number of studies that haveexamined its correlates and even fewer that have examined its predictors. Fur-ther, the majority of studies that have examined the correlates of indirect ag-gression have tended to focus on the same few variables—social status (e.g.,Crick, 1997; Crick et al., 1997; Crick & Grotpeter, 1995; Hennington et al.,1998; McNeilly-Choque et al., 1996; Rys & Bear, 1997; Salmivalli, Kau-kiainen, & Lagespetz, 2000; Vaillancourt & Hymel, 2004; Werner & Crick,1999) and psychological maladjustment (e.g., Crick, 1997; Crick & Grot-peter, 1995; Owens, Slee, & Shute, 2000b; Richardson & Green, 1997; seeCrick et al., 1999 for a review). These studies have demonstrated that indirectaggression is associated with increased peer rejection and psychological im-pairment, such as greater loneliness and depression for both females andmales, although girls seem to be more adversely affected than boys (e.g., Crick& Grotpeter, 1995; Hennington et al., 1998; Rys & Bear, 1997). It is also im-portant to note that the direction of these relations has yet to be established.For example, it is not clear as to whether peer rejection precedes the use of in-direct aggression or whether peer rejection is a consequence associated with itsuse. Accordingly, more longitudinal work in this area is needed. Moreover, itis also not clear as to whether the association between indirect aggression andpsychosocial maladjustment is curvilinear. That is, for some, using indirectaggression may not be problematic because it is used sparingly and in a goal-directed way. However, for others, diminished returns are likely to ensue inthat indirect aggression is used often and impulsively.
Studies that have investigated the link between indirect aggression andother markers of social status emphasize the idea that for some, indirect ag-gression may be related to competency. For example, several researchers haverecently noted a positive relation between the use of indirect aggression and
162 T H E D E V E L O P M E N T O F A G G R E S S I O N

peer perceptions of popularity (i.e., high visibility and influence among peers)(e.g., Cillessen & Mayeux, 2002; Prinstein & Cillessen, 2003; Vaillancourt &Hymel, 2004) and peer group centrality (e.g., Xie et al., 2002a; Xie, Swift,Cairns, & Cairns, 2002b). Ethnographic work by Merten (1997) and Adlerand Adler (1998) elucidate this seemingly contradictory link by describinghow girls use indirect aggression to achieve and maintain hegemony. In theiraggressive strive for popularity, girls are often disliked by their lower-statusand victimized peers. Thus, girls can be perceived as popular and powerfulwhile simultaneously being rejected by the larger peer network (Vaillancourt& Hymel, 2004).
Beyond sex, age, social status, and psychological maladjustment, the listof variables examined in relation to indirect aggression is rather short (seeUnderwood, 2003, for a more comprehensive review). For example, Kau-kiainen et al. (1999) found that among adolescent males and females, highlevels of social intelligence were associated with the use of indirect aggression,whereas the relation between physical aggression and social intelligence wasvirtually orthogonal. Vaillancourt and Hymel (2004) reported that adoles-cents’ use of indirect aggression was positively associated with the possessionof peer-valued characteristics such as attractiveness and stylish clothing.Walker et al. (2000) noted that adults who used indirect aggression were moremasculine than nonaggressive adults. These few studies certainly do not depicta clear profile regarding who typically uses indirect aggression but, rather,highlight how much there is to be done in this area of study.
WHY DO FEMALES USE INDIRECT AGGRESSION?
Women are naturally less prone to react aggressively than aremen . . . such variations . . . are largely socially determined.Physically smaller and weaker and periodically incapacitated dur-ing an important period of their lives by pregnancy and, to a lessextent, by menstruation, women are taught to accept a protectedalbeit subservient position within the group.
—DOLLARD, DOOB, MILLER, MOWRER, &SEARS (1939, p. 130)
Early Theories of Aggression
In conjecturing about why humans use aggression, many early theorists inad-vertently wrote about physically aggressive males and thus incorrectly as-sumed that females were less likely to respond aggressively (e.g., Bandura,1973; Dollard, Doob, Miller, Mowrer, & Sears, 1939; Freud, 1915; Lorenz,1966). These early (male) scholars of aggression failed to recognize (or per-haps consider) that females did aggress and did so in a circuitous manner. Thisoversight may be due to the fact that there were not a lot of data on female ag-
Indirect Aggression among Humans 163

gression to attend to. More likely, however, the omission was due to the factthat they held the assumption that the typical female was characteristicallydocile and nurturing, not aggressive.
The failure of early aggression theorists to notice or contemplate females’use of aggression was not overlooked by Maccoby and Jacklin (1974), who intheir review of the literature did address the “different modes hypothesis”(i.e., females and males aggress in dissimilar ways), only to dismiss it in favorof a biological argument, which again concentrated on physical aggression.More recently, Campbell (1993) appeared to be aware that females and malesexpressed aggression differently, with males using physical aggression and fe-males using indirect aggression. As a matter of fact, she stated that “boys arenot simply more aggressive than girls; they are aggressive in a different way”(p. 19), suggesting a recognition of the different modes hypothesis. In actual-ity, however, what Campbell meant by “in a different way” was that malesexpress their aggression directly whereas females suppress their aggressivefeelings entirely. “Girls do not learn the right way to express aggression; theysimply learn not to express it” (p. 20).
Naturally, since the initial writings of scholars such as Freud (1915),Dollard et al. (1939), Lorenz (1966), Bandura (1973) and Maccoby andJacklin (1974), our understanding of aggression in humans has developedconsiderably, with many of these progressions evolving in terms of our un-derstanding of how females aggress. For example, researchers have shownthat sex differences in the mode of aggressing are robust (see the precedingsection) and several researchers have put forth hypotheses of why this maybe.
Current Hypotheses Concerning Indirect Aggression
Although Feshbach (1969, 1971) was the first to empirically examine differ-ent modes of aggression in relation to sex, she did not comment on why it wasthat girls directed more indirect aggression against newcomers than did boys.In fact, it took another 2 decades for researchers to validate these findings andto hypothesize why this pattern of results existed. The first researchers to ar-ticulate a hypothesis concerning sex differences in the mode of aggressingwere Lagerspetz et al. (1988), who postulated that the organization of chil-dren’s peer groups, in particular the closeness of girls’ friendships, facilitatesthe use of indirect aggression by females by creating more “opportunities” toaggress in this manner. Lagerspetz et al. also argued that the social sanctionsplaced against the use of physical aggression by females contribute to thesesex differences. Like Lagespetz et al. (1988), Crick and Grotpeter (1995) ar-gued that sex differences in the mode of aggressing exist because childrenaggress in ways that are most salient and valued by their respective gendersand peer groups (see also Campbell, 1999). For boys, dominance and submis-sion are important themes in their social lives, but for girls, inclusion and ex-clusion are important themes (see Block, 1983, for a review). Consequently,
164 T H E D E V E L O P M E N T O F A G G R E S S I O N

boys use physical aggression to harm their peers, and girls rely on indirect ag-gression.
Whereas Lagerspetz et al. (1988) and Crick and Grotpeter (1995) at-tempt to explain sex differences in the ways children aggress in relation topeer group structures, Björkqvist (1994) proposed that such sex differencesare the result of a cognitive assessment strategy he termed the “effect/dangerratio” in which the aggressor considers the situation and makes a decision asto the type of aggressive tactic to employ. This determination is based on thejudgment of its effectiveness and on the potential danger involved. Because fe-males are usually involved in conflict with other females, targeting social rela-tionships would be the most successful way of inflicting harm (see also Crick& Grotpeter, 1995). Further, because indirect aggression is circuitous, it alsorepresents the least dangerous strategy inasmuch as the aggressor is often notidentified as the perpetrator. The use of indirect aggression by females thusrepresents an aggressive strategy that yields the maximum effect with the min-imum amount of hazard (see Björkqvist, 1994).
As mentioned previously, ethnographers such as Merten (1997) postulatethat girls, in particular adolescent females, use indirect aggression as a way ofachieving and maintaining popularity within their social reference group (seealso Adler & Adler, 1998; Vaillancourt, 2001). Being popular is an importantgoal for many adolescents (see Gavin & Furman, 1989). Studies that have ex-amined the link between indirect aggression and peer perceived popularityprovide support for this hypothesis by demonstrating its strong associationwith peer recognized power and visibility (e.g., Vaillancourt & Hymel, 2004;Vaillancourt & McDougall, 2003).
What is striking about the aforementioned hypotheses concerning sex dif-ferences in the expression of aggression is that they almost invariably fail toexplain the function of indirect aggression (with the exception of Adler &Adler, 1998; Merten, 1997; Vaillancourt, 2001). In other words, why aggressindirectly in the first place? More than 30 years ago, Bandura (1973) statedthat “a complete theory of aggression, whatever its orientation, must explainhow aggressive patterns of behavior are developed, what provokes people tobehave aggressively, and what maintains their aggressive actions” (p. 43).Within the study of indirect aggression, it can be said that we have succeededonly somewhat in explaining how it develops. Perhaps, then, it is time to con-sider alterative hypotheses regarding why females use indirect aggression. Onesuch hypothesis, which to date has not been fully considered, is that there maybe an evolutionary history associated with its use. That is, indirect aggressionmay be an inherent strategy that is employed by females to secure access to de-sirable mates and resources.
Evolutionary Theory of Direct Aggression
From an evolutionary perspective, males and females share similar reproduc-tive goals—the passing of their genes to subsequent generations. However, de-
Indirect Aggression among Humans 165

spite similarities in their ultimate reproductive objectives, males and femalesapproach this endeavor differently. This sex difference in procreative strate-gies had been attributed to differential parental investment, whereby (typi-cally) females across mammalian species, including humans, invest moreresources in their offspring than do males (Trivers, 1972). The result of thisimbalance is that females are choosy when selecting a reproductive partner,and males compete for access to these selective females.
The idea that females select whereas males contend is one of the centraltenets of Daly and Wilson’s (1988) evolutionary theory of violence. Spe-cifically, they argue that aggression is an evolved adaptation. This position issupported by numerous examples of aggression being initiated in circum-stances where increases to fitness (i.e., reproductive success) are to be reaped,as well as by data demonstrating that aggression comes to pass in situations inwhich resources (i.e., territory, food, etc.) are limited or unavailable, or threatsto self and/or family are perceived (Daly & Wilson, 1988; see also Archer,1988). These contexts, in which aggression is regularly displayed, illustrateswell the two processes believed to be involved in adaptation—natural selec-tion, which arises from competition for survival (i.e., access to food), andsexual selection, which arises from competition for reproduction (Darwin,1859, 1871).
Toward an Evolutionary Theory of Indirect Aggression among Females
Although Daly and Wilson’s (1988) evolutionary theory of aggression is ap-pealing insofar as it addresses the functions aggression might serve (i.e.,increased fitness and increased survival), one complication is the centered at-tention on direct forms of aggression and what it means to the lives of males.Such a concentration on male antagonism has a long tradition among evolu-tionary thinkers, including Darwin, who hardly referenced female–femalecompetition but rather focused on the “struggle between individuals of onesex, generally males, for the possession of the other sex” (1859, p.88). It is cu-rious that well over a hundred years after Darwin (1859, 1871) wrote his sem-inal books on the evolution of species, few scientists have considered the pos-sibility that there may be an intrinsic function associated with females’ use ofindirect aggression (Campbell, 1999). This is especially intriguing given thathuman females are not randomly choosing mates but rather, are often activelyselecting partners (Fisher, 1930; Zahavi, 1975; see Kokko, Brooks, Jennions,& Morley, 2003, for review) whom they perceive to be of superior quality—typically meaning higher in occupational and/or economic status (e.g., Ber-eczkei, Voros, Gal, & Bernath, 1997; Buss, 1989; Feingold, 1992). Thesehigh-status males can presumably provide females with greater resources,which in turn translates into greater reproductive advantage such as an in-creased number of surviving offspring (Daly & Wilson, 1983). Studies frompreindustrial (Voland & Engel, 1990) and industrial (Bereczkei & Csanaky,
166 T H E D E V E L O P M E N T O F A G G R E S S I O N

1996) societies support this idea, showing that females married to high-statusmales produced more surviving offspring than other females.
Considering that a female’s overall fitness may be positively augmentedby choosing a male with viability characteristics, and considering that not allmales will enjoy such positive qualities, intrasexual competition among fe-males should be expected (Paul & Baenninger, 1991). Such competition couldtake the form of attracting mates (Darwin, 1871), for example, by makingoneself more appealing through the use of tactics like manipulating appear-ance (Buss, 1988) or by intimidating competitors (Darwin, 1871) by makingthe adversary seem less attractive through, for example, the use of derogationstrategies (Buss & Dedden, 1990). Fittingly, then, it may be that females useindirect aggression because it represents an effective and less dangerous(Björkqvist, 1994) intrasexual strategy that ameliorates access to qualitymates and their resources (Campbell, 1995, 1999). Circumlocutory evidencesupporting this idea follows.
First, several researchers have shown that indirect aggression is especiallydamaging to female victims (e.g., Crick, 1996; Crick, Bigbee, & Howes, 1996;Galen & Underwood, 1997; Paquette & Underwood, 1999), who experiencedifficulties such as decreased self-esteem, increased anxiety and depression,school departure, and even suicide (e.g., Munro, 2000; Owens et al., 2000b).The plight of these victims highlights the efficacy of such agonistic tactics. Infact, when undergraduates were asked to rate the usefulness of derogation tac-tics in the context of intrasexual competition (Buss & Dedden, 1990), femalesselected items that were markedly consistent with those cited by developmen-tal psychologists (e.g., Crick & Grotpeter, 1995; Galen & Underwood, 1997;Lagerspetz et al., 1988) when describing the acts of indirect aggression. Forinstance, females rated spreading rumors (i.e., calling a competitor promiscu-ous or a “tease”) and peer group exclusion as effective ways to disparage a fe-male rival. In considering these findings, it seems likely that female victims ofindirect aggression would be less willing or able to vie for a male’s affection ifthey were suffering psychologically or if they had removed themselves entirelyfrom the competition (i.e., suicide).
In the nonhuman primate literature, it has been remarked that high-statusfemales harass low-status females indirectly. This pestering has been linked toestrus suppression and miscarriages among the victims. The reproductive suc-cess of the victim is thus obviously reduced and, accordingly, the resourcesavailable to the aggressor and her offspring increased (Hrdy, 1981, 1999; seeCampbell, 1999). Among humans, several studies have demonstrated thatprominent and powerful females use indirect aggression more than lower-status females (e.g., Vaillancourt & Hymel, 2004). Like their primate sisters(Wildman, Uddin, Liu, Grossman, & Goodman, 2003), high-status femalesare more likely to succeed in reducing a rival, given their social standingamong peers (see Merten, 1997). It is perhaps not so peculiar that girls use in-direct aggression to achieve and maintain hegemony, and do so at an early
Indirect Aggression among Humans 167

age, for this jostling for elevated status may be fundamentally linked to fit-ness.
Second, the notion that the use of indirect aggression by females may rep-resent an effective means for securing access to mates (direct benefit) andhence increase reproductive success (ultimate benefit) is supported by theethnographic work of Artz (1998a, 1998b), Campbell (1982, 1995), andMarsh and Paton (1986). In these studies, adolescent females, without fail, ac-cused other females of being “sluts.” This tactic of exaggerating another’s sex-ual history is consistent with the evolutionary idea that a female’s success inobtaining a desirable partner, who is committed, is largely determined by hisappraisal of her fidelity (Buss, 1988; Wilson & Daly, 1992). The adolescentfemales in these studies also reported that they would defame and even beatanother female if she slandered the aggressor’s reputation or showed an inter-est in her current or desired partner. In 1997, a 14-year-old Canadian girl wassavagely beaten and than drowned by a gang of females (and one male). Thereason for this murderous assault—supposedly, the victim had spread rumorsabout one of her attackers and was perceived to have shown an interest in theboyfriend of one of her assailants (Pemberton, 1997). It appears from theseaccounts that indirect aggression and physical aggression are used by femalesto secure access to mates, although the use of indirect aggression is far morecommon among females than the use of physical aggression. The evolutionarysignificance of this sex difference in aggressing likely has to do with the risk ofinjury involved to self and/or offspring when physical methods are used (seeBjörkqvist, 1994; Campbell, 1999, regarding humans; see Hrdy, 1981, 1999,regarding nonhuman primates). Indeed, physically aggressive encounters mayresult in injury or death, which in turn may reduce a female’s offspring’s abil-ity to survive, thus decreasing her fitness. Historically, among humans, andcurrently among most other species, offspring survival was/is inextricablylinked to maternal survival (see Hrdy, 1999)
Third, the idea that indirect aggression may be an evolved adaptation isfurther supported by cross-culture studies that have reliably demonstrated abias for females with respect to the use of indirect aggression. Specifically, fe-males from such diverse cultural backgrounds such as Indonesia (French et al.,2002), Finland (Lagerspetz et al., 1988), Poland (Osterman et al., 1994,1998), Italy (Osterman et al., 1994, 1998), North America (Crick & Grot-peter, 1995; Vaillancourt, 2001), Israel (Osterman et al., 1994, 1998), Austra-lia (Owens, Shute, & Slee, 2000a), and Argentina (Hines & Fry, 1994) werefound to express their aggression in a similarly covert and socially manipula-tive fashion. These consistent findings challenge theories of socialization inthat “culture” is no longer a fixed variable. It will be interesting to seewhether this pattern of results remains robust when indirect aggression isstudied more diversely (i.e., in more varied ethnic and religious cultures).
Fourth, according to Campbell (1999), females are more likely to useintrasexual strategies (i.e., indirect aggression) when their reproductive valueis at its peak (i.e., around puberty; ages 11–15) (Tanner, 1989), because this
168 T H E D E V E L O P M E N T O F A G G R E S S I O N

represents a time in their lives in which they have a “competitive advantageover younger and older women” (p. 109). This evolutionary hypothesis is con-sistent with several cross-sectional reports of indirect aggression that haveshown that early- and middle-adolescent females use indirect aggression morethan preadolescent and late-adolescent females (e.g., Björkqvist et al., 1992a,1992b; Osterman et al., 1998; Xie et al., 2002a, 2002b). This hypothesis isalso consistent with results from a recent nationally representative study of2,123 adolescent girls aged 10 to 16 in which early-maturing girls reported us-ing significantly more indirect aggression than average- and late-maturinggirls (Vaillancourt, Balshine, & Clark, 2003a). Maturation status was assessedby having participants indicate the age at which they began to menstruate,and the three groups were created based on a ±1 standard deviation split.
Fifth, coherent with the hypothesis that indirect aggression represents aneffective strategy to access and secure romantic partners, Vaillancourt et al.(2003a) also found that self-reported use of indirect aggression was positivelyassociated with number of romantic partners and negatively related to the ageat which participants had their first boyfriends. One limitation to this studywas that in these analyses, maturation status was not controlled for statisti-cally. However, similar results were also reported by Pellegrini and Long(2003), who found in their longitudinal study of adolescent males and females(grades 6–8) that the use of indirect aggression was positively related to datingat all four assessment point for girls only. For boys, “dating popularity” wasrelated to dominance-related strategies. Informed by sexual selection theory,Pellegrini and Long concluded that males and females use different “agonisticstrategies in the service of dating popularity,” with females using indirect ag-gression and males using dominance (p. 271).
Perhaps young females’ use of indirect aggression is akin to Wilson andDaly’s (1985) “young male syndrome,” albeit less extreme and risky. “Youngmale syndrome” refers to the intense intrasexual competition (violence) thatoccurs more often among young, poor males who, presumably, experience themost “intense reproductive competition (fitness variance) during the species’evolutionary history, and in those individuals whose present circumstances arepredictive of reproductive failure” (p. 59). Using 1972 homicidal data fromDetroit, Wilson and Daly (1985) found that most murders were committedover status disputes by young, socially disadvantaged, unwed males. Theadaptive function of engaging in these high-risk tactics is that they impart tofemales information about their resource-holding capability and their “goodgenes” (see Kokko et al., 2002, 2003). With respect to the use of indirect ag-gression, studies have shown that adolescents (ages 10 and 11) report usingindirect aggression more when they come from families of lower socioeco-nomic status (SES) that are headed by single mothers (Craig et al., 2002; seealso Tremblay, 1999). Interestingly, increased reported use of indirect aggres-sion was also found to be inversely related to the age of the participant’smothers at the time of his or her birth. Unfortunately, in these analyses, possi-ble interactions between sex, SES, and indirect aggression were not reported,
Indirect Aggression among Humans 169

so it is not clear whether these relations are stronger for females than formales. Nevertheless, these findings highlight important future areas of inquirythat are informed by a phylogentic perspective. For instance, Campbell (1995)argues that “the degree of female economic and social dependence on men isrelated to the intensity with which women are prepared to fight to secure highstatus males” (p. 116). Accordingly, then, less economically advantaged fe-males should engage in more intrasexual competition than their more re-sourced peers, because their “fitness” is rather dependent on securing andretaining a partner with resources (Bereczkei & Csanaky, 1996; Voland &Engel, 1990). The strength of these associations should be greater in nonin-dustrialized countries, where dependency on males for economic and social se-curity is still very much a reality.
In considering the aforementioned studies, there is some evidence to sup-port the idea that use of indirect aggression by females represents an effective(and less dangerous) means of intimidating potential rivals, hence increasingaccess to desirable partners and ultimately increasing fitness. From an evolu-tionary perspective, females and males have similar reproductive goals. How-ever, the ways in which they achieve their goals are different, with males rely-ing on overtly aggressive tactics and females relying on covertly aggressivetactics (see Pellegrini & Long, 2003). It has been argued that for females, therisks involved in engaging in physical aggression outweigh the benefits ac-crued, and thus females use indirect aggression to secure their goals (Björk-qvist, 1994; Campbell, 1995, 1999). It has also been argued that indirect ag-gression is less appealing to males than females because it does not promotetheir social goals of achieving dominance within the peer group (Campbell,1999; Pellegrini & Long, 2003). To do this, males should use visible tacticsthat signal to others their strength and bravado (see also Wilson & Daly,1985). It certainly appears that males and females are using different strategiesto access romantic partners. The question that remains, however, is “whetherthe two sexes have evolved distinctive modes of adaptation or, more likely,whether there has been a single adaptation with sex-specific threshold set-tings” (Campbell, 1999, p. 120).
CONCLUSION
Aggression is an organized, goal-directed activity, not the kindof event that could come from a random malfunction.
—PINKER (2002, p. 316)
Because the concentrated focus of research on aggression has been on physicalaggression, the terms male and aggression have effectively become synony-mous with one another. The review of the indirect aggression literature chal-lenges this assumption by demonstrating that females are capable of aggress-ing, but do so in a more covert manner. The review also highlights that, with
170 T H E D E V E L O P M E N T O F A G G R E S S I O N

few exceptions, the bulk of our research efforts to date have been on explain-ing indirect aggressing from an ontogenetic perspective, with little consider-ation of the fact that ontogeny is inextricably linked to phylogenetic selection(Cairns, 1979; Gaulin & McBurney, 2001). Accordingly, then, there still re-mains a need to consider the function of indirect aggression, as there shouldbe a reason for its consistent use by females across the planet. One such expla-nation is that, for females, indirect aggression may pay off in evolutionaryterms by increasing access and retention of high-quality males and their re-sources, and thus increasing their reproductive fitness.
ACKNOWLEDGMENTS
I wish to thank Amanda Krygsman, Brynn Winegard, and Sigal Balshine for their help-ful comments regarding this chapter. I also wish to thank Richard Tremblay, BillHartup, and John Archer for their careful review of, and suggestions for, this chapter.
REFERENCES
Adler, P. A., & Adler, P. (1998). Peer power: Preadolescent culture and identity.Piscataway, NJ: Rutgers University Press.
Archer, J. (1988). The behavioural biology of aggression. New York: Cambridge Univer-sity Press.
Arseneault, L., Tremblay, R. E., Boulerice, B., & Suacier, J.-F. (2002). Obstetrical compli-cations and violent delinquency : Testing two developmental pathways. Child De-velopment, 73, 496–508.
Artz, S. (1998a). Where have all the school girls gone? Violent girls in the school yard.Child and Youth Care Forum, 27, 77–109.
Artz, S. (1998b). Sex, power and the violent school girl. Toronto: Trifolium Books.Bandura, A. (1973). Aggression—A social learning analysis. Englewood Cliffs, NJ:
Prentice-Hall.Bereczkei, T., & Csanaky, A. (1996). Mate choice, marital status, and reproduction in a
modern society. Ethology and Sociobiology, 17, 23–35.Bereczkei, T., Voros, S., Gal, A., & Bernath, L. (1997). Resources, attractiveness, family
commitment; Reproductive decisions in human mate choice. Ethology, 103, 681–699.
Björkqvist, K. (1994). Sex differences in physical, verbal, and indirect aggression: A re-view of recent research. Sex Roles, 30, 177–188.
Björkqvist, K. (2001). Different names, same issue. Social Development, 10(2), 272–274.Björkqvist, K., Lagerspetz, K. M. J., & Kaukiainen, A. (1992a). Do girls manipulate and
boys fight? Developmental trends in regard to direct and indirect aggression. Ag-gressive Behavior, 18, 117–127.
Björkqvist, K., Osterman, K., & Kaukiainen, A. (1992b). The development of direct andindirect aggressive strategies in males and females. In K. Björkqvist & P. Niemela(Eds.), Of mice and women: Aspects of female aggression (pp. 51–64). San Diego,CA: Academic Press.
Indirect Aggression among Humans 171

Block, J. H. (1983). Differential premises arising from differential socialization of thegenders: Some conjectures. Child Development, 54, 1335–1354.
Broidy, L. M., Nagin, D. S., Tremblay, R. E., Brame, B., Dodge, K., Fergusson, D.,Horwood, J., Loeber, R., Laird, R., Lynam, D., Moffitt, T., Bates, J. E., Pettit, G. S.,& Vitaro, F. (2003). Developmental trajectories of childhood disruptive behaviorsand adolescent delinquency: A six site, cross-national study. Developmental Psy-chology, 39, 222–245.
Buss, D. M. (1988). The evolution of human intrasexual competition: Tactics of mate at-traction. Journal of Personality and Social Psychology, 54, 616–628.
Buss, D. M. (1989). Sex difference in human mate preferences: Evolutionary hypothesestested in 37 cultures. Behavioral and Brain Sciences, 12, 1–49.
Buss, D. M., & Dedden, L. A. (1990). Derogation of competitors. Journal of Social andPersonal Relationships, 7, 395–422.
Cairns, R. B. (1979). Social Development: The origins and plasticity of interchanges. SanFrancisco: Freeman.
Cairns, R. B., Cairns, B. D., Neckerman, H., Ferguson, L. L., & Gariepy, J. (1989).Growth and aggression: 1. Childhood to early adolescence. Developmental Psy-chology, 25, 320–330.
Campbell, A. (1982) Female aggression. In P. Marsh & A. Campbell (Eds.), Aggressionand violence. Oxford, UK: Blackwell.
Campbell, A. (1993). Men, women, and aggression. New York: Basic Books.Campbell, A. (1995). A few good men: Evolutionary psychology and female adolescent
aggression. Ethology and Sociobiology, 16, 99–123.Campbell, A. (1999). Staying alive: Evolution, culture and women’s intrasexual aggres-
sion. Behavioral and Brain Sciences, 22, 203–252.Cillessen, A. H. N., & Mayeux, L. (2002). From censure to reinforcement: Developmen-
tal changes in the role of aggression in peer relations. In T. Vaillancourt & P. C.Rodkin (Chairs), Functions of aggression in children’s peer relations. Symposiumconducted at the 15th World Meeting of the International Society for Research inAggression, Montreal, Canada.
Coie, J. D., & Dodge, K. A. (1998). Aggression and antisocial behavior. In W. Damon (Se-ries Ed.) & N. Eisenberg (Vol. Ed.), Handbook of child psychology (5th ed.): Vol. 3.Social, emotional and personality development (pp. 779–862). New York: Wiley.
Cote, S., Vaillancourt, T., Farhat, A., LeBlanc, J. C., Nagin, D., & Tremblay, R. (2004).The development of physical aggression during childhood. Manuscript submittedfor publication.
Craig, W. M., Peters, R. D., & Willms, J. D. (2002). The role of the peer group in pre-ado-lescent behaviour. In J. D. Willms (Ed.), Vulnerable children: Findings from Can-ada’s National Longitudinal Survey of Children and Youth (pp. 317–327). Edmon-ton: University of Alberta Press.
Crick, N. R. (1995). Relational aggression: The role of intent attributions, feelings of dis-tress, and provocation type. Development and Psychopathology, 7, 313–322.
Crick, N. R. (1996). The role of overt aggression, relational aggression, and prosocial be-havior in the prediction of children’s future social adjustment. Child Development,67, 2317–2327.
Crick, N. R. (1997). Engagement in gender normative versus nonnormative forms of ag-gression: Links to social-psychological adjustment. Developmental Psychology,33(4), 610–617.
Crick, N. R., Bigbee, M. A., & Howes, C. (1996). Gender differences in children’s norma-
172 T H E D E V E L O P M E N T O F A G G R E S S I O N

tive beliefs about aggression: How do I hurt thee? Let me count the ways. Child De-velopment, 67(3), 1003–1014.
Crick, N. R., Casas, J. F., & Mosher, M. (1997). Relational and over aggression in pre-school. Developmental Psychology, 33(4), 579–588.
Crick, N. R., & Grotpeter, J. K. (1995). Relational aggression, gender and social-psycho-logical adjustment. Child Development, 66, 710–722.
Crick, N. R., Werner, N. E., Casas, J. F., O’Brien, K. M., Nelson, D. A., Grotpeter, J. K., &Markon, K. (1999). Childhood aggression and gender: a new look at an old prob-lem. In Bernstein, D. (Ed.), Nebraska Symposium on Motivation: Vol. 45. Genderand motivation (pp. 75–141). Lincoln: University of Nebraska Press.
Daly, M., & Wilson, M. (1983). Sex, evolution, and behavior: Adaptations for reproduc-tion (2nd ed.). Boston: Willard Grant Press.
Daly, M., & Wilson, M. (1988). Homicide. New York: Aldine de Gruyter.Darwin, C. (1859). On the origins of species by means of natural selection. London:
Cambridge University Press.Darwin, C. (1871). The decent of man, and selection in relation to sex. New York: D.
Appleton.David, C. F., & Kistner, J. A. (2000). Do positive self-perceptions have a “dark side”?:
Examination of the link between perceptual bias and aggression. Journal of Abnor-mal Child Psychology, 28(4), 327–337.
Dollard, J., Doob, L. W., Miller, N. E., Mowrer, O. H., & Sears, R. R. (1939). Frustrationand aggression. New Haven, CT: Yale University Press.
Feingold. A. (1992). Gender differences in mate selection preferences: A test of parentalinvestment model. Psychological Bulletin, 112, 125–139.
Fergusson, D. M., Woodward, L. J., & Horwood, L. J. (1998). Maternal smoking duringpregnancy and psychiatric adjustment in late adolescence. Archives of General Psy-chiatry, 55(8), 721–727.
Feshbach, N. D. (1969). Sex differences in children’s modes of aggressive responses to-ward outsiders. Merrill–Palmer Quarterly, 15(3), 249–258.
Feshbach, N. D. (1971). Sex differences in adolescent reactions toward newcomers. De-velopmental Psychology, 4(3), 381–386.
Fisher, R. A. (1930). The genetical theory of natural selection. Oxford, UK: ClarendonPress.
French, D. C., Jansen, E. A., & Pidada, S. (2002). US and Indonesian children’s and ado-lescent’s reports of relational aggression by disliked peers. Child Development, 73,1143–1150.
Freud, S. (1915). The interpretation of dreams (authorized translation of 3rd ed. with in-troduction by A. A. Brill). London: G. Allen.
Galen, B. R., & Underwood, M. K. (1997). A developmental investigation of social ag-gression among children. Developmental Psychology, 33, 589–600.
Gaulin, S. J. C., & McBurney, D. (2001). Psychology: An evolutionary approach.Englewood Cliffs, NJ: Prentice-Hall.
Gavin, L. A., & Furman, W. (1989). Age differences in adolescents’ perceptions of theirpeer groups. Developmental Psychology, 25(5), 827–824.
Hart, C. H., Nelson, D. A., Robinson, C. C., Olsen, S. F., & McNeilly-Choque, M. K.(1998). Overt and relational aggression in Russian nursery-school-age children:Parenting style and marital linkages. Developmental Psychology, 34(4), 687–697.
Hennington, C., Hughes, J. N., Cavell, T. A., & Thompson, B. (1998). The role of rela-
Indirect Aggression among Humans 173

tional aggression in identifying boys and girls. Journal of School Psychology, 36,457–477.
Hines, N. J., & Fry, D. P. (1994). Indirect modes of aggression among women of BuenosAires, Argentina. Sex Roles, 30, 213–236.
Hrdy, S. B. (1981). The woman that never evolved. Cambridge, MA: Harvard UniversityPress.
Hrdy, S. B. (1999). Mother nature. New York: Ballantine.Hyde, J. S. (1984). How large are gender differences in aggression? A developmental
meta-analysis. Developmental Psychology, 20, 722–736.Kaukiainen, A., Björkqvist, K., Lagerspetz, K., Osterman, K., Salmivalli, C., Rothberg,
S., & Ahlbo, A. (1999). The relationship between social intelligence, empathy andthree types of aggression. Aggressive Behavior, 25, 81–89.
Keenan, K., & Shaw, D. S. (1994). The development of aggression in toddlers: A study oflow-income families. Journal of Abnormal Child Psychology, 22, 53–77.
Kingston, L., & Prior, M. (1995). The development of patterns of stable, transient, andschool-age onset aggressive behavior in young children. Journal of American Acad-emy of Child and Adolescent Psychiatry, 34, 348–358.
Kokko, H., Brooks, R., Jennions, M. D., & Morley, J. (2003). The evolution of matechoice and mate biases. Proceedings of the Royal Society of London. Section B: Bi-ology, 270, 653–664.
Kokko, H., Brooks, R., McNamara, J. M., & Houston, A., I. (2002). The sexual selectioncontinuum. Proceedings of the Royal Society of London. Section B: Biology, 269,1331–1340.
Lagerspetz, K. M. J., Björkqvist, K., & Peltonen, T. (1988). Is indirect aggression typicalof females? Gender differences in aggressiveness in 11 to 12–year-old children. Ag-gressive Behavior, 14, 403–414.
Lahey, B. B., McBurnett, K., & Loeber, R. (2000). Are attention-deficit hyperactivity dis-order and oppositional defiant disorder developmental precursors to conduct disor-der? In A. Sameroff, M. Lewis, & S. Miller (Eds.), Handbook of developmentalpsychopathology (pp. 431–446). New York: Plenum Press.
Loeber, R., & Hay, D. (1997). Key issues in the development of aggression and violencefrom childhood to early adulthood. Annual Review of Psychology, 48, 371–410.
Loeber, R., & Stouthamer-Loeber, M. (1998). Development of juvenile aggression and vi-olence: Some common misconceptions and controversies. American Psychologist,53, 242–259.
Lorenz, K. (1966). On aggression. New York: Harcourt, Brace and World.Lykken, D. T. (2001). Parental licensure. American Psychologist, 56, 885–894.Maccoby, E. E., & Jacklin, C. N. (1974). The psychology of gender differences. Stanford,
CA: Stanford University Press.Maccoby, E. E., & Jacklin, C. N. (1980). Sex differences in aggression: A rejoinder and
reprise. Child Development, 51, 964–980.Marsh, P., & Paton, R. (1986). Gender, social class and conceptual schemas of aggres-
sion. In A. Campbell & J. Gibbs (Eds.), Violent transactions: The limits of personal-ity (pp. XX). Oxford, UK: Basil Blackwell.
McNeilly-Choque, M. K., Hart, C. H., Robinson, C. C., Nelson, L. J., & Olsen, S. F.(1996). Overt and relational aggression on the playground: Correspondence amongdifferent informants. Journal of Research in Childhood Education, 11, 47–67.
Merten, D. E. (1997). The meaning of meanness: Popularity, competition, and conflictamong junior high school girls. Sociology of Education, 70, 175–191.
174 T H E D E V E L O P M E N T O F A G G R E S S I O N

Moore, T. M., Scarpa, A., & Raine, A. (2002). A meta-analysis of serotonin metabolite5–HIAA and antisocial behavior. Aggressive Behavior, 28, 299–316.
Munro, H. (2000, December 2). Dawn-Marie’s tragic last days. The Vancouver Sun, p.B1.
Osterman, K., Björkqvist, K., Lagerspetz, K. M. J., Kaukiainen, A., Huesmann, L. R., &Fraczek, A. (1994). Peer and self-estimated aggression and victimization in 8–year-old children from five ethnic groups. Aggressive Behavior, 20, 411–428.
Osterman, K., Björkqvist, K., Lagerspetz, K. M. J., Kaukiainen, A., Landau, S. F.,Fraczek, A., & Caprara, G. V. (1998). Cross-cultural evidence of female indirect ag-gression. Aggressive Behavior, 24, 1–8.
Owens, L. D. (1996). Sticks and stones and sugar and spice: Girls’ and boys’ aggression inschools. Australian Journal of Guidance and Counseling, 6, 45–55.
Owens, L., Shute, R., & Slee, P. (2000a). “Guess what I just heard”: Indirect aggressionamong teenage girls in Australia. Aggressive Behavior, 26(1), 67–83.
Owens, L., Slee, P., & Shute, R. (2000b). It hurts a hell of a lot . . . : The effects of indirectaggression on teenage girls. School Psychology International, 21, 359–376.
Paquette, J. A., & Underwood, M. K. (1999). Gender differences in young adolescents’experiences of peer victimization: Social and physical aggression. Merrill–PalmerQuarterly, 45, 242–266.
Patterson, G. R. (1995). Coercion—A basis for early age of onset for arrest. In J. McCord(Ed.), Coercion and punishment in long-term perspective (pp. 81–105). New York:Cambridge University Press.
Paul, L., & Baenninger, M. A. (1991). Aggression by women: Mores, myths and methods.Oxford, UK: North-Holland.
Pellegrini, A. D., & Long, J. D. (2003). A sexual selection theory longitudinal analysis ofsexual segregation and integration in early adolescence. Journal of ExperimentalChild Psychology, 85, 257–278.
Pemberton, K. (1997, November 26). Slain girl suffered teasing, rejection in troubled life.The Vancouver Sun, p. A3.
Pinker, S. (2002). The blank slate: The modern denial of human nature. New York: Vi-king Press.
Plomin, R. (1983). Childhood temperament. In B. B. Lahey & A. E. Kazdin (Eds.), Ad-vances in clinical child psychology (Vol. 6, pp. 45–92) New York: Plenum Press.
Prinstein, M. J., & Cillessen, A. H. N. (2003). Forms and functions of adolescent peer ag-gression associated with high levels of peer status. Merrill–Palmer Quarterly, 49,310–342.
Rabasca, N. (2000). Michel sees Psychological Review as forum to connect with newfields. Monitor on Psychology, 31(1). Retrieved August 26, 2004, from www.apa.org/monitor/jan00/as3.html.
Raine, A. (1993). The psychopathology of crime: Criminal behavior as a clinical disorder.San Diego, CA: Academic Press.
Raine, A. (2002). Annotation: The role of prefrontal deficits, low automatic arousal, andearly health factors in the development of antisocial and aggressive behavior in chil-dren. Journal of Child Psychology and Psychiatry, 43, 417–434.
Richardson, D. R., & Green, L. R. (1997). Circuitous harm: Determinants and conse-quences of nondirect aggression. In Kowalski (Ed.), Aversive interpersonal behav-iors. New York: Plenum Press.
Rys, G. S., & Bear, G. G. (1997). Relational aggression and peer relations: Gender and de-velopmental issues. Merrill–Palmer Quarterly, 43, 87–106.
Indirect Aggression among Humans 175

Salmivalli, C., Kaukiainen, A., & Lagerspetz, K. (2000). Aggression and sociometric sta-tus among peers: Do gender and type of aggression matter? Scandinavian Journal ofPsychology, 41, 17–24.
Schaal, B., Tremblay, R. E., Soussignan, R., & Susman, E. J. (1996). Male testosteronelinked to high school dominance but low physical aggression in early adolescence.Journal of the American Academy of Child and Adolescent Psychiatry, 35, 1322–1330.
Tanner, J. M. (1989). Foetus into man: Physical growth from conception to maturity.London: Castlemead.
Tomada, G., & Schneider, B. (1997). Relational aggression, gender, and peer acceptance:Invariance across culture, stability over time, and concordance among informants.Developmental Psychology, 33, 601–609.
Tremblay, R. E. (1999). When children’s social development fails. In D. P. Keating & C.Hertzman (Eds.), Developmental health and the wealth of nations: Social, biologi-cal, and educational dynamics (pp. 55–71). New York: Guilford Press.
Tremblay, R. E. (2000). The development of aggressive behavior during childhood: Whathave we learned in the past century? International Journal of Behavioral Develop-ment, 24, 129–141.
Tremblay, R. E., Boulerice, B., Harden, P. W., McDuff, P., Peruse, D., Pihl, R. O., &Zoccolillo, M. (1996). Do children in Canada become more aggressive as they ap-proach adolescence? In Human Resources Development Canada and Statistics Can-ada (Eds.), Growing up in Canada: National Longitudinal Survey of Children andYouth (pp. 127–137). Ottawa: Statistics Canada.
Tremblay, R. E., Japel, C., Perusse, D., McDuff, P., Boivin, M., Zoccolillo, M., &Montplaisir, J. (1999). The search for the age of “onset” of physical aggression:Rousseau and Bandura revisited. Criminal Behavior and Mental Health, 9, 8–23.
Tremblay, R. E., & LeMarquand, D. (2001). Individual risk and protective factors. In R.Loeber & D. Farrington (Eds.), Child delinquents: Development, interventions andservice needs (pp. 137–164). Thousand Oaks, CA: Sage.
Trivers, R. L. (1972). Parental investment and sexual selection. In B. Campbell (Ed.),Sexual selection and the descent of man 1871–1971 (pp. 136–179). Chicago:Aldine.
Underwood, M. K. (2003). Social aggression among girls. New York: Guilford Press.Underwood, M. K., Galen, B. R., & Paquette, J. A. (2001). Top ten challenges for under-
standing gender and aggression in children: Why can’t we all just get along? SocialDevelopment, 10, 248–266.
Vaillancourt, T. (2001). Competing for hegemony during adolescence: A link betweenaggression and social status. Unpublished doctoral dissertation, University of Brit-ish Columbia, Vancouver, BC, Canada.
Vaillancourt, T., Balshine, S., & Clark, A. (2003a). Puberty, dating and the use of indirectaggression: Evolution or social construct? Paper presented at the Biennial Meetingof the Canadian Psychological Association, Hamilton, ON, Canada.
Vaillancourt, T., Brendgen, M., Boivin, M., & Tremblay, R. (2003b). Longitudinal con-firmatory factor analysis of indirect and physical aggression: Evidence of two fac-tors over time? Child Development, 74(6), 1628–1638.
Vaillancourt, T., Cote, S., Farhat, A., LeBlanc, J. C., Boivin, M., & Tremblay, R. (2004).The development of indirect aggression during childhood. Manuscript submittedfor publication.
Vaillancourt, T., & Hymel. S. (2003). The social context of children’s aggression. In M.
176 T H E D E V E L O P M E N T O F A G G R E S S I O N

Moretti, M. Jackson, & C. Odgers (Eds.), Girls and aggression: Contributing fac-tors and intervention principles (pp. 57–73). New York: Kluwer Academic.
Vaillancourt, T., & Hymel, S. (2004). Competing for hegemony during adolescence:Gender, aggression and social status. Manuscript submitted for publication.
Vaillancourt, T., & McDougall, P. (2003). Rejected by whom?: A look at the relationshipbetween same-sex and other-sex perceptions of physical and relational aggression.Poster presented at the Biennial Meeting of the Society for Research in Child Devel-opment, Tampa, FL.
Voland, E., & Engel, C. (1990). Female choice in humans: A conditional mate selectionstrategy of the Krumhorm women (Germany, 1720–1874). Ethology, 84, 144–154.
Walker, S., Richardson, D. S., & Green, L. R. (2000). Aggression among older adults: Therelationship of interaction networks and gender role to direct and indirect re-sponses. Aggressive Behavior, 26, 145–154.
Werner, N. E., & Crick, N. R. (1999). Relational aggression and social-psychological ad-justment in a college sample. Journal of Abnormal Psychology, 108, 615–623.
Wildman, D. E., Uddin, M., Liu, G., Grossman, L. I., & Goodman, M. (2003). Implica-tions of natural selection in shaping 99.4% nonsynonymous DNA identity betweenhumans and chimpanzees: Enlarging genus Homo. Proceedings of the NationalAcademy of Science, 100, 7181–7188.
Willoughby, J., Kupersmidt, J. B., & Bryant, D. (2001). Overt and covert dimensions ofantisocial behavior. Journal of Abnormal Child Psychology, 29, 177–187.
Wilson, M., & Daly, M. (1985). Competitiveness, risk taking, and violence: The youngmale syndrome. Ethology and Sociobiology, 6, 59–73.
Wilson, M., & Daly, M. (1992). The man who mistook his wife for a chattel. In J. H.Barkow, L. Cosmides, & J. Tooby (Eds.), The adapted mind (p. XX). New York:Oxford University Press.
Xie, H., Cairns, R. B., & Cairns, B. D. (2002a). The development of social aggression andphysical aggression: A narrative analysis of interpersonal conflicts. Aggressive Be-havior, 28, 341–355.
Xie, H., Swift, D. J., Cairns, B. D., & Cairns, R. B. (2002b). Aggressive behaviors in so-cial interaction and developmental adaptation: A narrative analysis of interpersonalconflicts during early adolescence. Social Development, 11, 205–224.
Zahavi, A. (1975). Mate selection: A selection for handicap. Journal of Theoretical Biol-ogy, 53, 205–214.
Indirect Aggression among Humans 177

THE DEVELOPMENT OF AGGRESSIONProactive and Reactive Aggression
9
Proactive and Reactive AggressionA Developmental Perspective
FRANK VITARO and MARA BRENDGEN
TERMINOLOGIES AND DEFINITIONS
Different subtypes of aggressive behaviors may be defined on the basis of theirform. For example, based on form, several authors have suggested distinctionsbetween physical and nonphysical aggression (Tremblay, 2000) or between di-rect and indirect/relational aggression (Björkqvist, Lagerspetz, & Kaukiainen,1992; Crick & Grotpeter, 1995). Others proposed subtyping of aggressive be-haviors on the basis of the underlying goal (i.e., function, motivation). Onesuch subtyping with respect to the underlying goal that has received increasingattention is the distinction between reactive and proactive aggression (Dodge,1991; Dodge & Coie, 1987).
The concept of reactive aggression has its roots in the frustration–angertheory of aggression (e.g., Berkowitz, 1963, 1993; Dollard, Doob, Miller,Mowrer, & Sears, 1939). According to this perspective, aggression is a re-sponse triggered by goal blocking and is accompanied by high autonomicarousal (i.e., anger). Reactive aggression thus occurs as a consequence of an-tecedent conditions of real or perceived provocation, frustration, or threatand is usually accompanied by the expression of anger. Its main goal is toreact to the anger–frustration stimulus and hurt the perpetrator of the prov-ocation or the threat. Finally, it is rather immediate and impulsive in re-sponse to the source of provocation or threat. As such, reactive aggressioncan be linked conceptually to behavior displayed by Type A personalities,who are easily irritable and angered by provocation, competition, or deni-gration (Holmes & Will, 1985). In line with the frustration—aggression
178

model, reactively aggressive behaviors and anger can be viewed as uncondi-tioned temperamentally based reactions or conditioned emotional responsesto external events (Berkowitz, 1989). This view, however, does not implythat external contingencies cannot influence the use of reactively aggressiveacts or moderate their links to anger feelings. Conceptually, reactive aggres-sion can be used as a synonym for “defensive,” “angry,” “hot blooded,”“impulsive,” “emotional,” or “retaliatory” aggression (Berkowitz, 1993;Buss, 1961; Feshback, 1964).
Conversely, the concept of proactive aggression is more in line with thesocial learning model of aggression (Bandura, 1973, 1983), which postulatesthat aggression is an acquired behavior governed by reinforcement contin-gencies. According to this notion, proactive aggression is thought to bedriven by the anticipated rewards that follow the perpetration of aggressiveacts. For example, proactive aggression can be used as an instrumentalmeans to secure goods from others or to dominate others. Synonyms forproactive aggression are “offensive,” “predatory,” and “instrumental” ag-gression.
Following these two theoretical concepts of aggression, a substantialnumber of researchers have differentiated between reactive acts of aggression,committed out of anger or fear, and goal-directed proactive aggression(Berkowitz, 1989, 1993; Buss, 1961; Dodge, 1991; Feshback, 1964; Parke &Slaby, 1983; Pulkkinen, 1987; Zillman, 1979). Others, however, have chal-lenged the view that the distinction between reactive and proactive aggressionin humans is reliable, valid, and useful (Bushman & Anderson, 2001). Thequestion is thus whether reactive and proactive aggression are indeed two dis-tinct types of aggressive behaviors, that is, whether they can be distinguishednot only theoretically but also empirically. If the answer is yes, then importantimplications follow. At the theoretical level, two distinct etiological modelsmay be necessary to account for the origins and the developmental course ofeach type of aggressive behavior. At the practical level, it will be necessary tomeasure the two types of aggressive behaviors separately. At the clinical level,evidence for a distinction between reactive and proactive aggression might ne-cessitate specific treatments and prevention programs for each type of aggres-sive behavior.
The purpose of this chapter is to more closely examine the empirical evi-dence in support of, or against, this distinction. Because the main focus of thisbook is the investigation of the developmental origins of aggressive behavior,we review studies of proactive and reactive aggression that are based on childand adolescent samples; adult data are also discussed in some cases. The firstpart of the chapter is devoted to measurement and construct validity issues re-garding proactive and reactive aggression. The second part reviews evidencesupporting or contesting the distinction between the two types of aggressivebehaviors by examining the personal, social, behavioral, and physiologicalcorrelates of proactive and reactive aggression. In the third part, we attemptto integrate the existing findings into a tentative developmental model of
Proactive and Reactive Aggression 179

proactive and reactive aggression and point out specific areas that merit fur-ther research.
MEASUREMENT AND VALIDITY ISSUES
Measurement
With some notable exceptions (i.e., Chaux, Arboleda, & Rincón, 2003; Coie,Dodge, Terry, & Wright, 1991; Little, Jones, Henrich, & Hawley, 2003b;Pulkkinen, 1996), the majority of studies on proactive and reactive aggressionin human children used questionnaire-based measures that are usually evalu-ated by teachers, or in some instances, by parents or peers. With the exceptionof the aforementioned investigations, the studies have used six items originallydeveloped by Dodge and Coie (1987) to operationalize the two different typesof aggression. The proactive items are, “This child uses physical force, orthreatens to use force, to dominate other children,” “This child threatens orbullies others in order to get his/her way,” “This child gets other children togang up on a peer he/she does not like.” The reactive items are, “When thischild has been teased or threatened, he/she gets angry easily and strikes back,”“When a peer accidentally hurts this child, this child assumes that the peermeant to do it and then overreacts with anger and fighting,” This child claimsthat other children are to blame for a fight and feels like they started the trou-ble.”
On the basis of these items, some authors then analyzed the continuoustotal scores of reactive and proactive aggression, respectively. Others createdgroups of individuals who scored high (e.g., one standard deviation above themean or more) or low (e.g., less than one standard deviation above the mean)on one or both types of aggressive behaviors to identify proactive-only, reac-tive-only, proactive–reactive, and nonaggressive individuals. For example, inone study, about 53% of children were found to be both proactively and reac-tively aggressive, about 32% were only reactively but not proactively aggres-sive, and about 15% were only proactively but not reactively aggressive(Dodge, Lochman, Harnish, Bates, & Pettit, 1997). Despite slight variationsdue to different cutoff criteria, these percentages are very similar across differ-ent studies (e.g., Pulkkinen, 1996; Vitaro, Gendreau, Tremblay, & Oligny,1998). The first strategy is called the “variable-centered approach,” and thesecond is known as the “person-centered or grouping approach.” The vari-able-centered approach is mostly used (1) to test the factor structure ofproactive and reactive aggression or (2) to examine the unique relations of thecontinuous reactive and proactive aggression scores (i.e., controlling for therespective other type of aggression) with some other continuous variable suchas, for example, self-esteem. In contrast, the person-centered approach is usedto examine differences between the aggression groups (i.e., proactive-only,reactive-only, proactive–reactive, and nonaggressive individuals) in regard to agiven set of variables. The use of continuous scores of proactive and reactive
180 T H E D E V E L O P M E N T O F A G G R E S S I O N

aggression has the obvious advantage of using of the full variability of thesebehaviors. However, given that aggression scores are often non-normally dis-tributed, with most individuals receiving low scores and a few individuals re-ceiving high scores, the person-centered approach is also reasonable and doesnot necessarily diminish statistical power (Farrington & Loeber, 2000).
Nevertheless, the use of arbitrary cutoffs to create groups of reactivelyand/or proactively individuals remains an important and difficult-to-solveproblem for the person-centered approach, which might affect the interpret-ability and comparability of findings across studies. Despite their methodolog-ical differences, however, the two approaches basically yield the same infor-mation in regard to the question of whether the distinction between proactiveand reactive aggression is valid. Therefore, we include both studies using avariable-centered approach and studies using a person-centered approach inthis chapter.
Factor Structure
The first question that needs to be answered in regard to the proactive–reactivedichotomy is whether the two can be at all distinguished on a measurementlevel. Individuals obviously can exhibit both reactive and proactive aggression(Berkowitz, 1993; Dodge, 1991), and continuous proactive and reactive aggres-sion scores indeed correlate on average at r = .70 (± .15) in variable-centeredstudies. Similarly, a majority of participants identified as aggressive in person-centered studies score above the cutoffs on both reactive and proactive aggres-sion, whereas only a minority are only reactive and even fewer are onlyproactive (Dodge, Lochman, Harnish, Bates, & Pettit, 1997; Pulkkinen, 1996;Vitaro et al., 1998). These results seem to speak against a clear distinction be-tween reactive and proactive aggression at the empirical level and are in linewith arguments by Bushman and Anderson (2001) that it is often not possibleto conclude whether an aggressive act is reactively or proactively motivated.Despite the high overlap between the two types of aggression, exploratory andconfirmatory factor analyses nevertheless yield two distinct factors in linewith the reactive–proactive dichotomy (Brown, Atkins, Osborne, & Milma-now, 1996; Crick & Dodge, 1996; Day, Bream, & Paul, 1992; Dodge & Coie,1987; Pellegrini, Bartini, & Brooks, 1999; Poulin & Boivin, 2000a; Salmivalli& Nieminen, 2002). For example, Poulin and Boivin (2000a) used confirma-tory factor analysis to test whether a one- or two-factor solution would bestfit the data. They found that a two-factor model showed better fit indices thana single-factor model, based on teacher and parent ratings of children’sproactive and reactive aggression, but were unable to distinguish between re-active and proactive aggression when using peer data. However, other authorsdid find a two-factor structure based on peer-rated reactive and proactive ag-gression using exploratory factor analysis (e.g., Salmivalli & Nieminen,2002).
Even stronger evidence for the factorial distinctiveness of proactive and
Proactive and Reactive Aggression 181

reactive aggression comes from a recent study by Little et al. (2003b). Previ-ous studies on proactive and reactive aggression have focused exclusively ondirect, especially physical, forms of aggression. Little and colleagues, how-ever, distinguished not only between the different functions of aggression(i.e., between proactivity and reactivity, labeled instrumental and reactive intheir study), but also between the different forms that aggressive behaviorcan take (i.e., overt/direct versus relational/indirect). Based on self-reports ofaggressive behavior from children in grades 5–10, confirmatory factor anal-ysis revealed a clear four-factor structure. Specifically, two distinct con-structs referring to the function of aggression were identified—proactive andreactive aggression—as well as two distinct constructs referring to the formof aggression—overt and relational aggression. Notably, by taking into ac-count any confounds between proactive and reactive aggression that mightbe due to a similar form of aggression, Little et al. (2003b) showed thatproactive and reactive aggression were basically uncorrelated (r = –.10).This obvious distinctiveness of proactive and reactive aggression was thesame across age (grades 5–7 vs. grades 8–10), gender, and ethnic groups(i.e., in German and Turkish children).
Construct Validity of the Proactive–Reactive Distinction
As we have seen, it seems possible to reliably measure two distinct factors ofproactive aggression and reactive aggression in children and in adolescents.The question emerges, however, as to whether these measures indeed capturethe proactive–reactive distinction on a conceptual level. Dodge & Coie (1987)were the first to examine the convergent validity of their teacher-rated mea-sure of reactive and aggressive aggression by comparing it with direct observa-tions of proactive and reactive aggression during children’s interactions insmall play groups. These authors found that teacher ratings and direct obser-vations of the same type of aggressive behavior were moderately correlated(i.e., r = .27 for both types of aggressive behavior), indicating that question-naire-based measures of proactive and reactive aggression are indeed rooted inreal-life behavior. A similar conclusion can be drawn from research linkingpsychopathy and proactive versus reactive aggressive tendencies in adult of-fenders. For example, in two ethnically different samples of incarceratedadults, proactive but not reactive aggression was significantly correlated withFactor 1 of the Psychopathy Checklist (PCL; Harpur, Hakstian, & Hare,1989) (Joseph P. Newman, personal communication). Factor 1 of the PCLcovers personality traits such as callousness, a lack of affect, guilt, remorse,and empathy, habitual lying and manipulation, and superficiality, whichclosely resemble the definition of proactive aggression as instrumental and“cold-blooded.” Similar findings were obtained in a person-centered studyshowing that incarcerated proactive–reactive adult offenders are more psycho-pathic than either reactive-only violent offenders or nonviolent offenders(Cornell et al., 1996).
182 T H E D E V E L O P M E N T O F A G G R E S S I O N

Other Validity Issues: The Underlying Social Cognitions of Proactiveand Reactive Aggression
The view of reactive aggression as an angry response to real or perceived prov-ocation, and of proactive aggression as an instrumental means driven by theanticipated rewards, implies that the underlying social–cognitive processes ofthese two types of aggression should be very different. In line with this notion,several studies have shown that reactive but not proactive aggression is relatedto hostile attributional biases in response to ambiguous provocation (Dodge& Coie, 1987; Crick & Dodge, 1996; Dodge et al., 1997; Hubbard, Dodge,Cillesen, Coie, & Schwatz, 2001; Schippell, Vasey, Cravens-Brown, & Bret-veld, 2003). Reactive but not proactive aggression is also related to biasedattention for rejection, ridicule, and failure cues (Schippell et al., 2003). More-over, as compared with nonaggressive children, only reactively aggressive chil-dren manifest problem solving deficits in difficult social situations (Dodge,1991; Kolko & Brown, 1997; Price & Dodge, 1989). In contrast, proactivelyaggressive children value aggression as an effective means to achieve desiredgoals more than other children (Crick & Dodge, 1996; Dodge et al., 1997).Similarly, proactive but not reactive aggression was related to positive out-come expectancies for aggressive behavior in a sample of incarcerated maleadolescents (Smithmyer, Hubbard, & Simons, 2000). Although these findingsdo not reveal the directionality of effects between social cognitions andproactive or reactive aggression, they nevertheless support the notion of twoforms of aggressive behavior that are used in different contexts and for differ-ent purposes.
Validity of Proactive and Reactive Aggression: A Word of Caution
The empirical evidence reviewed here indicates that the distinction betweenproactive and reactive aggression is internally valid. However, it is importantnot to equate the proactive and reactive distinction with other terminologies.For example, a distinction between instrumental and hostile aggression mightbe used as a synonym for the proactive–reactive aggression distinction (e.g.,Bushman & Anderson, 2001), but this equation may not necessarily be appro-priate. Thus, in some studies, instrumental aggression has been operational-ized by the rate at which a child pressed a button, which created an obstacle tovictory for a (fictitious) opponent during a competitive game. Hostile aggres-sion was defined by the rate at which the child pressed a different button,which caused the opponent to hear an aversive noise (Atkins & Stoff, 1993;Atkins, Stoff, Osborne, & Brown, 1993b; Hoving, Wallace, & La Forme,1979). This type of aggressive behavior was termed “hostile” because it gaveno apparent advantage to the child with respect to the game. It is obviousfrom this description that not only instrumental but also hostile aggressioncan be proactive (i.e., initiated by the aggressor without a triggering provoca-tion from the opponent). In addition, hostile aggression does not necessarily
Proactive and Reactive Aggression 183

include the emotional features (i.e., anger) that characterize reactive aggres-sion. Empirical evidence for the notion that the proactive–reactive distinctionmay not be synonymous with instrumental and hostile aggression, respec-tively, comes from a study examining the links between teacher-rated proactive–reactive aggression and instrumental–hostile aggression using the competitiveparadigm described earlier (Atkins et al., 1993a). The results indicated thatboys who engaged in high rates of instrumental aggression were likely to berated by teachers as high in proactive aggression, but this was true only forboys who were rejected by their peers. In contrast, boys who engaged in highrates of hostile aggression were rated by teachers as high in both proactive andreactive aggression. Again, this result applied only to rejected boys. Obviously,although there is a parallel between instrumental and proactive aggression, re-active aggression cannot be equated with hostile aggression. These subtle butimportant differences between different terminologies need to be kept in mindwhen interpreting empirical findings from different studies.
CORRELATES OF PROACTIVE AND REACTIVE AGGRESSION
So far, we have seen that the distinction between proactive and reactive ag-gression seems to be sufficiently reliable and also shows reasonable constructand convergent validity. Another way to examine whether proactive and reac-tive aggression are two distinct constructs is to test whether reactive andproactive aggression have different concurrent or longitudinal correlates(Campbell & Fiske, 1959). As discussed in the following paragraphs, empiri-cal findings indeed support the notion that the two types of aggression are dif-ferently related to various personal, social–environmental, behavioral, andphysiological factors.
Temperament
So far, only one study has examined whether proactive and reactive aggressionare differentially related to temperament (Vitaro, Brendgen, & Tremblay,2002). The findings are in line with the view of reactive—but not proactive—aggression as an intense temperament-based response to an aversive stimulus.By age 6, reactively aggressive preadolescents (whether also proactively ag-gressive or not) already seem to be temperamentally more reactive and moreinattentive than proactive-only or nonaggressive children, who actually do notdiffer from each other in this respect. Notably, reactively aggressive pre-adolescents in the Vitaro et al. (2002) study also showed increased reactivityto unconditioned stimuli such as light or pain by age 6, suggesting that theseyoungsters might be prone to react to any disturbance from a social or anonsocial source. These findings concur with the image of reactively aggres-sive children as more excitable than proactively aggressive or nonaggressivechildren, because of a lower response threshold in regard to disturbing stimuli.
184 T H E D E V E L O P M E N T O F A G G R E S S I O N

However, because temperament was assessed at age 6, temperament measuresmay reflect socialization influences as well personal dispositions at that age.As shown next, empirical evidence indeed suggests that proactive and reactiveaggression are related to very different socialization experiences within thefamily and with peers.
Family History
Dodge hypothesized that a history of social–environmental hostility and mal-treatment is implicated in reactive aggression, whereas a history of exposureto successful aggressive models and a positively reinforced experience with co-ercive behavior are implicated in proactive aggression (Dodge, 1991; Dodge& Coie, 1987). Existing studies seem to confirm this hypothesis. Thus, par-ents of reactively aggressive schoolchildren have been found to be more con-trolling and punitive during the preschool years than parents of proactivelyaggressive or nonaggressive children (Bowen & Vitaro, 1998). Similarly, in asample of young adolescent males treated for severe behavior problems, thereactively aggressive youngsters demonstrated histories of physical abuse,whereas proactively-only and proactively–reactively aggressive individuals didnot (Dodge et al., 1997). In contrast, proactively aggressive adolescents seemto enjoy rather positive family relations as compared with their proactively–reactively aggressive or reactively aggressive counterparts (Poulin & Dishion,2000). At the same time, proactively aggressive youth report less parentalmonitoring and fewer household rules than the reactively aggressive groupand the nonaggressive group (Poulin & Dishion, 2000).
Although suggestive of a unidirectional causal influence of differentparenting practices on proactive and reactive aggression, these results may beinterpreted in more than one way. For example, if indeed reactive aggressionhas its roots in a highly reactive, difficult temperament, parents may soon be-come exasperated by their reactively aggressive child’s behavior and develop anegative attitude toward him or her as a consequence. What may seem like acause may thus well be a consequence of the child’s reactive behavior. Onceagain, only the study of a birth cohort of children and their parents may helpsolve this chicken-and-egg problem and possibly reveal a transactional insteadof a unidirectional process between children’s and parents’ behaviors.
Peer Relations
In addition to different family experiences, proactive and reactive aggressionalso seem to entail different experiences with peers, both in terms of children’slevel of acceptance in the peer group and in terms of their dyadic friendships.Overall, existing data indicate that reactive aggression seems to be less toler-ated by other children than proactive aggression. For example, reactive butnot proactive aggression has been linked to low social preference during thepreschool years as well as in late childhood and adolescence, both when using
Proactive and Reactive Aggression 185

continuous measures of aggression (Alvarez & Olson, 1999; Price & Dodge,1989; Prinstein & Cillessen, 2003) and when employing a person-centered ap-proach (Dodge et al., 1997; Poulin & Boivin, 2000a). Further evidence thatreactive but not proactive aggression is related to peer difficulties comes fromresearch showing that reactive children are also more victimized than pro-active children (Poulin & Boivin, 2000a). It is clear from these results thatreactively aggressive children are at high risk for maltreatment by peers, in ad-dition to maltreatment by adults (Dodge et al., 1997). In turn, maltreatmentby peers may aggravate their reactive tendencies and their propensity for hos-tile attributions. This hypothesis is supported by recent data showing that peerrejection during the early elementary years predicted later reactive aggressionmore than later proactive aggression (Dodge et al., 2003).
Proactive aggression also seems to be more tolerated and even reinforcedby peers at the dyadic level. Thus, proactively aggressive children not onlyhave more friends than reactively aggressive children, but the former also havea greater tendency to have similarly aggressive friends than the latter (Poulin& Boivin, 2000b). Moreover, friends’ proactive aggressiveness has beenshown to increase children’s own proactive aggressiveness over the course of aschool year, whereas friends’ reactive aggressiveness has no influence on chil-dren’s own aggressiveness profile (Poulin & Boivin, 2000b). Once more, theseresults concur with the view that proactive aggression is supported by positivesocialization experiences whereas reactive aggression is related to or resultsfrom aversive experiences. However, the greater acceptability of proactiveaggression among peers seems to depend on the context in which proactivelyaggressive behavior is used. Although object-oriented proactive aggression isindeed unrelated to peer acceptance, person-oriented proactive aggression(i.e., bullying) has been found to be negatively related to popularity amongpeers (Price & Dodge, 1989). These latter findings suggest that the link be-tween each type of aggressive behavior and peer status may be conditioned bycontextual and dispositional factors, such as local norms regarding use of ag-gression, children’s age, and possibly other methodological elements such asthe type of specific measures or instruments used.
Other Related Adjustment Problems
In light of the negative experiences in the family and peer contexts associatedwith reactive aggression, it is perhaps not surprising that reactive aggressionhas also been linked with other concurrent behavioral, emotional, and cogni-tive adjustment problems to a greater extent than proactive aggression. Forexample, reactive aggression, but not proactive aggression, is significantlycorrelated with disruptive classroom behaviors (Waschbusch, Willoughby, &Pelham, 1998). Reactive-only boys and proactive–reactive (but not proactive-only) boys are also less skilled than nonaggressive children at responding toteasing, sharing, negotiating, handling failure, compromising, and displayingsportsmanship, and perform more poorly in school (Day et al., 1992). More-
186 T H E D E V E L O P M E N T O F A G G R E S S I O N

over, reactive but not proactive aggression has been linked to internalizingproblems such as withdrawal, anxiety, depression, and psychosomatic symp-toms, as well as ADHD diagnosis and personality disorder (Day et al., 1992;Dodge et al., 1997; Vitaro et al., 2002). In contrast, proactive aggression hassometimes even been associated with reduced anxiety (Scarpa, Hurley, &Hirai, 2002). An extensive test of the notion that reactive aggression but notproactive aggression is linked with a host of concurrent adjustment difficultieshas been performed in a recent study of primary and secondary school chil-dren (Little et al., 2003a). In addition to the three usual groups of highly reactive-only, highly proactive-only, and highly reactive–proactive individuals, theseauthors distinguished between two groups of nonaggressive participants,namely, children with a typical/average level of proactive and/or reactive ag-gression and children with very low levels of both proactive and reactive ag-gression. Interestingly, the proactive-only group resembled the typical/averagegroup in many ways. Proactive-only youth did well in school, had positiveself-concepts, did not harbor hostile feelings toward others, and were able toregulate feelings of frustration. In contrast, the reactive-only group and thereactive–proactive group showed considerable frustration, intolerance, andelevated levels of hostility, as well as low levels of school performance and in-tellectual skills, as compared with nonaggressive children.
In light of these findings, one might be tempted to conclude that reac-tively aggressive children experience considerably more adjustment difficultiesthan proactively aggressive children in a variety of domains. Empirical evi-dence suggests, however, that not only reactive but also proactive aggressioncarries substantive risks for developmental adjustment problems. Specifically,proactively aggressive children seem more at risk than reactively aggressivechildren for some forms of externalizing problems. Thus, teacher-ratedproactive but not reactive aggression during childhood has been found to pre-dict later delinquency and conduct disorders during adolescence in low SESboys (Vitaro et al., 1998). These results were replicated in another study witha representative sample of children that also included girls (Vitaro et al.,2002). Proactive-only children were mostly at risk for later overt delinquency(e.g., violent bullying and gang fights), whereas reactive-only children werenot more involved in overt or covert delinquent behaviors than nonaggressivechildren. Other researchers also found that proactive aggression but not reac-tive aggression predicts later criminal behavior (Pulkkinen, 1996). Finally,there is evidence with adult criminals (Cornell et al., 1996), as well as with ad-olescent offenders (Connor, Steingard, Anderson, & Melloni, 2003) and nor-mal adolescent samples (Poulin & Dishion, 2000), that proactive but not reac-tive aggression is concurrently related to criminal behaviors and substanceuse.
Part of the explanation of why proactively aggressive children are more atrisk of later delinquency-related problems involving other co-offenders may liein the fact that proactively aggressive children often have friends who are sim-ilarly proactively aggressive, whereas reactively aggressive children are often
Proactive and Reactive Aggression 187

friendless (Poulin and Boivin, 2000b). This tentative explanation is furthersupported by recent findings by Poulin, Dishion, and Boivin (2002) indicatingthat proactive but not reactive aggression predicts involvement with a deviantpeer group from grade 6 to grade 7 even when controlling for gender, currentdeviant peers, parental monitoring, and peer rejection. Finally, the high anxi-ety of reactively aggressive children may also help explain why they are not ata similar risk for delinquency-related acts as proactively aggressive children.Anxiety might reflect high harm avoidance, according to Cloninger’s personal-ity model (Cloninger, 1986), or a high inhibition system, according to Gray’smodel (Gray, 1982). This disposition to inhibition has been shown to operateas a protective factor against delinquency for aggressive boys (Kerr, Tremblay,Pagani & Vitaro, 1997). Recent evidence suggests, however, that it is not onlythe proactively aggressive individuals who are at risk for violence and criminalbehavior (Brendgen, Vitaro, Tremblay, & Lavoie, 2001). Although confirmingthat proactive aggression uniquely predicted delinquency-related violence(e.g., gang fights), these authors showed that reactive aggression uniquely pre-dicted dating violence during late adolescence, after controlling for proactiveaggression. Findings from Cornell et al. (1996) and Cornell, Benedek, andBenedek (1987) also support the notion that reactively aggressive individualsare more prone to violence in intimate relationships, whereas proactively ag-gressive counterparts are more inclined to violence in delinquency-related situ-ations such as robbery, gang fights, or drug dealing. The fact that violenceagainst intimate partners often occurs as a defense against feelings of frustra-tion or vulnerability in situations of conflict with the partner (Prince & Arias,1994) may explain why reactive aggression is specifically related to this typeof behavior.
Emotions and Biological Correlates
Virtually all the studies reported until now support the validity of the distinc-tion between reactive and proactive aggression by showing different links withsocial–cognitive, temperamental, behavioral, school-related, peer-related, andfamily-related correlates. In comparison, only a few studies used biological oraffective/emotional correlates to establish the validity of the reactive–proactivedichotomy, and the few existing studies show equivocal findings. For example,the link between reactive and proactive aggression to physiological and obser-vational indicators of anger was examined in a sample of second-grade chil-dren (Hubbard et al., 2002). As mentioned previously, anger is, by definition,considered a concomitant (and possibly a cause) of reactive but not proactiveaggression and should thus be strongly related to the former but unrelated tothe latter. In order to test this notion, angry feelings were induced by exposingthe children to a confederate who cheated during a competitive game. Physio-logical, behavioral, and subjective indicators of anger (i.e., skin conductanceand heart rate, angry facial expressions, and angry nonverbal behaviors) werecollected during the game and self-reports of anger collected during the subse-
188 T H E D E V E L O P M E N T O F A G G R E S S I O N

quent viewing of the game on videotape. Two aspects of each measure wereconsidered: an aggregate score throughout the game and an increased scoreover the course of the game. Two of the five measures (i.e., angry nonverbalbehaviors and skin conductance) were positively related to teacher-rated reac-tive aggression while controlling for proactive aggression. Moreover, angrynonverbal behaviors were negatively related to proactive aggression. The rateof increase of these behaviors over the course of the game was also positivelyrelated to reactive aggression but unrelated to proactive aggression. An aggre-gate score of skin conductance throughout the game was also uniquely andpositively related to reactive aggression. Furthermore, the skin conductancelevels of children who were high in reactive aggression increased continuouslythroughout the game, whereas it did not for children who were low in reactiveaggression.
Although these results support the distinction between reactive andproactive aggression by suggesting a unique link between reactive aggressionand anger, other results from the same study did not support this distinction.Thus, the total number of angry expressions and their evolution during thegame were not related to reactive (nor to proactive) aggression. Moreover,self-reports of anger were not related to reactive aggression when averagedacross the game, but, unexpectedly, they were related to proactive aggressionwhen examined over the course of the game. Finally, the pattern of results forheart rate reactivity did not support an exclusive link with reactive aggression.When averaged across the game, heart rate reactivity was negatively related toreactive aggression but not to proactive aggression. The authors explain theseresults by stating the possibility that the game may have elicited as much (ifnot more) interest as anger in the participants. The fact that heart rate isdriven upward by emotional arousal, but downward by interest or orientationto a triggering event, may explain the results with respect to the links betweenheart rate and proactive and reactive aggression. Other results indicating thatteacher-rated reactively aggressive children are not viewed by their classmatesas more angry than proactively–reactively and proactively-only aggressivechildren also cast doubt on the exclusive link between reactive aggression andanger (Dodge & Coie, 1987).
Another physiological correlate that may differ for proactive and reactiveaggression is cortisol. Cortisol is a marker of the activity of the hypothalamic–pituitary–adrenal (HPA) axis, which is sensitive to physical and psychologicalstress. Most researchers have reported an inverse relationship between cortisollevel and antisociality at all ages (King, Jones, Scheurer, Curtis, & Zarcone,1990; Woodman, Hinton, & O’Neill, 1978). One study, however, reportedhigh levels of cortisol in conduct-disordered children, provided that they alsomanifested anxiety problems (McBurnett et al., 1991). Based on behavioralcorrelates of reactive aggression, these children may well resemble reactivelyaggressive children. A direct test of this hypothesis was performed recently byvan Bokhoven et al. (2003). These authors found that, when controlling forproactive aggression, boys who were high in teacher-rated reactive aggression
Proactive and Reactive Aggression 189

manifested significantly higher levels of cortisol than boys low or average inreactive aggression. Like the majority of studies on cortisol, this study wasbased on measurements of resting cortisol concentrations. The few other stud-ies that tested for the link between aggression and cortisol in children or ado-lescents during stress or anger-provoking situations unfortunately did not dis-tinguish between reactive and proactive aggression (Klimes-Dougan, Hastings,Granger, Usher, & Zahn-Waxler, 2001; Susman, Dorn, Inoff-Germain, Nottel-mann, & Chrousos, 1997; Van Goozen, Matthys, Cohen-Kettenis, Buitelaar,& Van Engeland, 2000).
CONNECTING THE PIECES: A DEVELOPMENTAL MODELOF PROACTIVE AND REACTIVE AGGRESSION
In light of the studies reviewed in this chapter, can we conclude that the dis-tinction between reactive and proactive aggression is reliable, valid, and use-ful? Despite a high correlation between the two types of aggressive behaviorsand their co-occurrence in a majority of aggressive individuals, reactive andproactive aggression can be distinguished at the factorial level, which speaksto the reliability of the proactive–reactive distinction. In addition, despitesome equivocal findings, most existing evidence suggests that reactive andproactive aggression do not seem to share the same correlates at the personal,social, academic, behavioral, and physiological levels. This speaks to the va-lidity of the proactive–reactive distinction. Perhaps most important, person-centered studies consistently show that, in addition to individuals who displayboth types of aggressive behavior, individuals can be identified who showmostly reactive but not proactive aggression, even though proactively-onlyaggressive individuals seem to be rare. In light of the different correlates ofproactive and reactive aggression, this person-centered distinction may even-tually be important in identifying at-risk children for prevention purposesaimed at specific maladjustment outcomes. As such, the existing evidence sug-gests that the distinction between reactive and proactive aggression can beuseful for both theoretical and practical purposes.
Obviously, much still remains to be learned about the developmental ori-gins and outcomes of proactive and reactive aggression. For one thing, mostof the existing research is based on concurrent data. This makes it impossibleto disentangle the directionality of effects between proactive and reactive ag-gression and their respective associated correlates. Moreover, studies usuallyfocus on only a limited number of correlates, which makes it difficult to assessthe relative contribution of a given risk factor to the development of proactiveversus reactive aggression. Despite these shortcomings, we believe that the ex-isting data already allow the formulation of a tentative model of the develop-mental pathways associated with reactive and proactive aggression. In thissection we attempt to integrate the current knowledge into such a theoretical
190 T H E D E V E L O P M E N T O F A G G R E S S I O N

framework. The starting point is the theoretical model proposed by Dodge(1991), in which reactive and proactive aggression originate from differentexperiences and develop independently. According to this model, reactiveaggression develops in reaction to a harsh, threatening, and unpredictable en-vironment or abusive and cold parenting. Conversely, proactive aggressionthrives in supportive environments that foster the use of aggression as a meansto achieve one’s goals. Individuals who are high in both types of aggressive be-haviors likely experience both types of environment. Many empirical studiesreviewed in this chapter support these different etiological pathways for reac-tive and proactive aggression. This first theoretical model may be termed theparallel development model, because it views reactive and proactive aggres-sion as originating from different etiological factors and as developing in aparallel, almost independent manner.
There is, however, another possible model in which temperamental fac-tors operate as main (i.e., initial) effects and in which different psychosocialfactors serve as moderators. We call this second model the sequential develop-ment model. It is based on the findings, discussed previously, that tempera-mental and neurophysiological elements seem to play a role in reactive aggres-sion but not in proactive aggression. Conversely, environmental factors seemto play a role mostly in proactive aggression. According to this alternativemodel, children with certain temperamental or neurophysiological character-istics are initially disposed to display aggressive behavior as a means of inter-acting with their environment. Initially, this tendency might be expressedmainly in a reactive manner, such as in high irritability and a difficult tempera-ment in early infancy. If early outbursts of anger (screaming, kicking) lead tothe desired outcome (food, toys, etc.), these children may learn that such be-havior is a successful means not only of alleviating a stressful situation butalso of obtaining a desired goal in a nonstressful situation. Gradually, aggres-sion may also be used in a “purely” proactive manner to achieve goals, partic-ularly in a permissive familial or peer environment, which may not only fail toinhibit but even foster this type of behavior. These personal and environmen-tal contingencies related to proactive aggression may eventually open the wayto other externalizing behavior problems, in particular delinquency. Suchoversimplifications do not imply that proactive aggression has no neurophys-iological or genetic correlates or that reactive aggression is not partly depen-dent on environmental contingencies and learning experiences. Nevertheless,the idea that reactive aggression might precede and open the way to proactiveaggression is supported by data showing that elements of reactive aggressionsuch as anger and irritability seem to appear already in early infancy. In con-trast, dominating and instrumental proactive aggression directed toward peersdoes not emerge until about 12 months of age, when conflicts become ex-tremely frequent, characterizing as much as half of peer exchanges between 12and 18 months (Caplan, Vespo, Peterson, & Hay, 1991). Also in support ofthis notion, longitudinal data from Lansford, Dodge, Pettit, and Bates (2002)
Proactive and Reactive Aggression 191

showed that reactive aggression in one year is predictive of proactive aggres-sion in the next over a period of several years (i.e., from kindergarten throughgrade 7), whereas proactive aggression does not predict subsequent reactiveaggression.
Why do not all children with an initial disposition for aggressive behaviordisplay both reactive and proactive aggression later on? More specifically,why are some individuals mainly reactively but not proactively aggressive andwhy are there so few who seem to use aggression in a proactive but not in a re-active manner? Some aggressive children may be less successful than others incontrolling resources and dominating through aggression, perhaps becausethey are more frequently and severely punished for aggressive acts or becausethey are more anxious and sensitive to punishment. To avoid negative conse-quences, these aggressive children may stop using or never start using (physi-cally) aggressive behavior in a proactive fashion and may adopt alternativestrategies for goal achievement. Nevertheless, because of their high sensitivityto stressful stimuli and their lack of impulse control, these children may re-main reactively aggressive and use aggression when provoked or threatenedand would therefore be called the “reactive-only” group. Because of their highreactivity to stress, these children may eventually also develop internalizingproblems, which may be exacerbated by other problems associated with reac-tive aggression, such as peer rejection, victimization, or low school achieve-ment.
The question then remains as to how some children come to be proactive-only if aggressive behavior is indeed initially used mainly in a reactive manner.One possible explanation is that these children may experience very few prov-ocations from others, perhaps because of their greater physical strength or be-cause they possess other personal characteristics that make them an unlikelytarget of attacks from others. In their case, there may not be much need to useaggression in a reactive manner. As a consequence, outside observers such asteachers, parents, or peers, who are often used to evaluate children’s aggres-sion, may hardly ever observe reactive aggression in these children. The use ofproactive aggression in these children is shaped through reinforcement contin-gencies and maintained through beliefs in its power and deference of its effectson the victims. This is not to say that these children would not use aggressionin a reactive manner if they were provoked. In fact, given that their proactiveaggression is a very successful strategy of goal achievement, it is highly likelythat these children will avert a provocation or threat from another childthrough aggressive means with equal success. Thus, there may in fact not re-ally be a group of “proactive-only” children, a notion that is fostered by theextremely small number of children identified as such in empirical studies(e.g., Dodge et al., 1997; Pulkkinen, 1996; Vitaro et al., 1998). Of course, thedescribed developmental processes of reactive and proactive aggression re-main highly speculative at this point. However, this (temporary, it is hoped)situation should not weaken the conclusion that subtypes of aggressive behav-
192 T H E D E V E L O P M E N T O F A G G R E S S I O N

iors based on their function can and should be distinguished in future researchand that pertinent instruments should be developed in that direction. In therest of this chapter, we identify areas in need of investigation and offer possi-ble directions for future research.
OPEN QUESTIONS AND ADDITIONAL ISSUES
Developmental Trajectories
As mentioned earlier, many open questions remain that preclude a full un-derstanding of the etiology of proactive and reactive aggression. An obviousissue in need of clarification is the question of whether the “age of onset”and the developmental course of proactive and reactive aggression differ. Inour tentative model, we have speculated that reactive aggression precedesproactive aggression, but no study (to our knowledge) has examined the de-velopmental trajectories of reactive and proactive aggression from birth on-ward to examine the developmental course of the two types of aggressivebehavior. Some support of the notion that reactive aggression seems to startat an earlier age than proactive aggression is provided by findings that, ac-cording to mothers’ reports, clinically referred reactively aggressive boyswere 4.4. years old when their behavior problems began, whereas pro-actively aggressive boys were 6.8 years old when their behavior problemsbegan (Dodge et al., 1997). Given the retrospective nature of the assess-ments, the characteristics of the sample, and the possibility that reactive ag-gression was reported earlier because it was more salient than proactive ag-gression, these results are difficult to interpret. There is only one study, toour knowledge, that has directly tested the mean level development ofproactive and reactive aggression over an extended period of time, that is,from kindergarten through grade 7, through growth curve analysis (Lans-ford et al., 2002). In this study, children displayed higher levels of reactiveaggression than of proactive aggression during kindergarten. On one hand,this finding might indicate that proactive aggression develops at a later agethan reactive aggression. On the other hand, because both types of aggres-sion decreased with age, it is also possible that both types of aggressionemerge at the same time but that socialization experiences succeed morequickly in teaching children to abandon the use of aggression for instrumen-tal purposes. This may be more difficult to achieve with the highly emotion-driven reactive aggression. Obviously, more longitudinal research is neededto clarify the developmental course of proactive and reactive aggression. Inthis context, the use of new clustering techniques for longitudinal data, suchas Nagin’s (1999) trajectories procedure would be especially useful to iden-tify groups of children with different longitudinal profiles of proactive and/or reactive aggression and to examine the longitudinal course of these chil-dren’s aggressive behaviors.
Proactive and Reactive Aggression 193

In Search of Causality: Experimental Manipulation of Reactiveand Proactive Aggression
Apart from mapping the longitudinal developmental course of behaviors suchas proactive and reactive aggression, the big challenge in developmental re-search is to detect the causal links between these behaviors and associatedvariables. As mentioned earlier, most of the discussed studies on proactive andreactive aggression are based on concurrent data, although some used longitu-dinal designs either (1) to investigate potential antecedents of proactive andreactive aggression while controlling for previous levels of the two types of ag-gression or (2) to assess the putative outcomes of proactive and reactive ag-gression while controlling for previous levels of the outcome measures. Evenin longitudinal studies, however, effects may be difficult to interpret. For ex-ample, the contribution of a putative environmental factor such as parentalbehavior on the etiology of proactive and reactive aggression is difficult to in-terpret because the effect may also be due, for example, to the genetic trans-mission of aggressive behaviors. The use of twin samples provides a bettercontrol of this problem because it allows testing additive models of variancedecomposition, which distinguish between heritable or genetic influences,shared environmental influences, and nonshared environmental influences onthe phenotypic (i.e., behavioral) similarity between twins (Neale & Cardon,1992). Although, to our knowledge, no studies to date have used a twin de-sign to examine the antecendents (or outcomes) of proactive and reactive ag-gression, this approach represents a promising avenue for future research.
Although superior to studies producing cross-sectional or simple longitu-dinal data, even a twin design does not provide clear information about thecausality of effects between proactive and reactive aggression, on the onehand, and associated variables, on the other hand. A test of causality can beachieved only through experimental manipulation. One such way to test, forexample, the specific causal link between reactive or proactive aggression, andtheir putative antecedents, would be to reduce the hypothesized antecedentrisk factor through intervention and see whether proactive or reactive aggres-sion is decreased subsequently. For example, the notion that a lack of angermanagement skills represents a risk factor for reactive but not for proactiveaggression could be tested by improving anger management skills in aggres-sive children and testing whether reactive—but not proactive—aggression isreduced as a consequence. Similarly, potential outcomes of proactive and reac-tive aggression could be examined by reducing either proactive or reactive ag-gression through intervention and investigating whether the outcomes that areconsidered specific to one type of aggression or the other (e.g., delinquent be-havior or depression) are reduced subsequently.
To date, only one study (to our knowledge) has attempted a manipulationof putative specific antecedents of proactive and reactive aggression throughintervention (Phillips & Lochman, 2003). A normative sample of 10–12-year-old boys from low SES families was divided into a reactive-anger manipula-
194 T H E D E V E L O P M E N T O F A G G R E S S I O N

tion group and a proactive-instrumental manipulation group. The children inthe reactive-anger manipulation group were trained to recognize angry feel-ings, to control impulsive responses, and to relax and refocus attention whenangry. Children in the proactive-instrumental manipulation condition learnedto recognize domineering behavior and to think about the positive conse-quences of using nonaggressive acts. Results indicated that the reactive manip-ulation was indeed successful in specifically reducing reactive aggression andanger in a competition game. Contrary to what would be expected, however,the proactive manipulation resulted in lowered levels of both reactive andproactive aggression.
Lochman and collaborators used a similar approach to test outcomesspecific to proactive and reactive aggression. The first study used a child-centered small-group-format program aimed at teaching aggressive boyswith anger management problems to regulate their angry feelings and im-prove their social problem-solving strategies (Lochman, 1992). Although re-active and proactive aggression were not measured specifically, it is reason-able to assume that the program impacted mostly on reactive aggression,given its aim and the participants’ characteristics. Interestingly, later delin-quency was not reduced despite a proximal positive effect of the programon reactive aggression and social-problem–solving skills. This finding is thusin line with the previously discussed correlational studies showing that reac-tive aggression does not predict subsequent delinquency (e.g., Pulkkinen,1996; Vitaro et al., 2002). In a more recent prevention program called thePower Coping Program, a significant proximal effect of the child-centeredprogram on angry feelings and hostile attributions in the aggressive partici-pants was found (Lochman & Wells, 2002; Lochman & Wells, 2004).Again, no effect was found on delinquency and substance use one year afterthe end of the program. Interestingly, however, the data show that the com-bination of the Coping Power Program with a Parent Training Componentand a universal prevention program was successful in improving parent dis-ciplinary strategies and in reducing children’s positive outcome expectanciesregarding aggression. Parent-rated and teacher-rated proactive aggressionalso decreased. Moreover, the combined program successfully reduced sub-stance use and delinquency one year later. These results suggest that reduc-ing proactive aggression and associated positive expectations regarding theuse of violence has an impact on later delinquency and later substance use.In contrast, reducing anger-driven reactive aggression seems less effective inthis regard. It should be noted, however, that no analysis was performed inthese studies to directly test this mediational assumption.
These studies provide excellent examples of how experimental manipula-tion in an intervention can be used for testing causal linkages betweenproactive and reactive aggression, respectively, and putative antecedents oroutcomes. To be sure, intervention studies would be a costly strategy if thesole purpose were the testing of theories. However, the potential benefits forthe developmental adjustment of proactively and/or reactively aggressive chil-
Proactive and Reactive Aggression 195

dren makes the intervention approach an attractive option and more, similarstudies are needed.
Gender, Ethnicity, and Culture
A final issue in need of further research is the question of potential differencesbetween boys and girls, and between ethnic or cultural groups, with respect tothe prevalence or etiology of proactive and reactive aggression. As mentioned,many of the previously discussed studies on proactive and reactive aggressionhave been conducted with boys only, and the few studies that did examinegender differences reported equivocal results. For example, in a sample ofGerman and Turkish participants in grades 5–10, boys were found to displaymore proactive aggression than girls, whereas girls were more reactively ag-gressive than boys (Little et al., 2003b). In a sample of American adolescentsin grades 6–8, however, boys reported only higher levels of reactive aggres-sion, but not of proactive aggression, than girls at the first time of measure-ment (Little, 2002). However, reactive aggression in that study decreased inboys over an 18-month interval, whereas it increased in girls, so that boys andgirls did not differ any more in regard to reactive aggression at the final pointof measurement. In yet another study with a clinically referred adolescentsample, no gender differences were found in regard to both proactive and re-active aggression (Connor et al., 2003). An even greater lack of knowledge ex-ists with respect to a potential difference in proactive and reactive aggressionbetween different ethnic groups and cultures. So far, only one study—to ourknowledge—has examined potential differences between different ethnic groupswith respect to proactive and reactive aggression (Little et al., 2003b). Theseauthors not only examined potential gender differences but also comparedTurkish and German children with respect to the prevalence of proactive andreactive aggression. The Turkish children reported more reactive but not moreproactive aggression than the German children.
To be sure, the detection of differences between genders or culturalgroups in regard to proactive and reactive aggression has limited value unlesswe try to understand the underlying mechanisms that may explain these dif-ferences. For example, the dissimilarity between Turkish and German childrenwith respect to reactive aggression may hint at possible differences in tempera-mental and potentially heritable characteristics between ethnic groups, ormight reflect different cultural norms concerning the expression of reactive ag-gression. Only through a comprehensive comparison of the interplay betweenbehavioral, cognitive, environmental, and biological factors in boys and girlsor between ethnic groups can we hope to gain a better understanding of thecultural or gender differences in proactive and reactive aggression. At thesame time, such research might provide valuable insights into the etiology ofproactive and reactive aggression, especially when based on longitudinal dataor combined with powerful methodological approaches such as twin designsor experimental manipulations.
196 T H E D E V E L O P M E N T O F A G G R E S S I O N

In this chapter, we have attempted to summarize and integrate the exist-ing knowledge regarding the reliability, the validity, and the theoretical andpractical usefulness of the distinction between proactive and reactive aggres-sion. Although limited and not always consistent, the available evidence sup-ports this distinction. Nevertheless, we are still a long way from a full under-standing of the developmental origins and course of proactive versus reactiveaggression. With new longitudinal research and innovative measurement andanalytical tools, the next decade promises to open the door to a greater insightinto these different aspects of aggressive behavior.
REFERENCES
Alvarez, L. M., & Olson, S. L. (1999). The relation between proactive and reactive ag-gression and peer social status in preschool children. Poster session presented at thebiennial meeting of the Society for Research in Child Development, Albuquerque,NM.
Atkins, M. S., Osborne, M. L., Brown, K. S., Milnamow, M., Bennett, D., & Kerwin, M.L. (1993a). Proactive and reactive aggression: Instrumental or hostile? Paper pre-sented at the biennial meeting of the Society for Research in Child Development,New Orleans.
Atkins, M. S., & Stoff, D. M. (1993). Instrumental and hostile aggression in childhooddisruptive behavior disorders. Journal of Abnormal Child Psychology, 21, 165–178.
Atkins, M. S., Stoff, D. M., Osborne, M. L., & Brown, K. (1993b). Distinguishing instru-mental and hostile aggression: Does it make a difference? Journal of AbnormalChild Psychology, 21, 355–365.
Bandura, A. (1973). Aggression: A social learning analysis. Englewood Cliffs, NJ:Prentice-Hall.
Bandura, A. (1983). Psychological mechanisms of aggression. In R. G. Green & E. I.Donnerstein (Eds.), Aggression: Theoretical and empirical views (Vol. 1, pp. 1–40).New York: Academic Press.
Berkowitz, L. (1963). Aggression: A social learning analysis. New York: McGraw-Hill.Berkowitz, L. (1989). Frustration–aggression hypothesis: Examination and reformula-
tion. Psychological Bulletin, 106, 59–73.Berkowitz, L. (1993). Aggression: Its causes, consequences, and control. New York:
McGraw-Hill.Björkqvist, K., Lagerspetz, K., & Kaukiainen, A. (1992). Do girls manipulate and boys
fight? Developmental trends in regard to direct and indirect aggression. AggressiveBehavior, 18, 117–127.
Bowen, F., & Vitaro, F. (1998, July). Reactively and proactively aggressive children: An-tecedent and subsequent characteristics. Paper presented at the International Soci-ety for the Study of Behavioral Development, Berne, Switzerland.
Brendgen, M., Vitaro, F., Tremblay, R. E., & Lavoie, F. (2001). Reactive and proactive ag-gression: Predictions to physical violent in different contexts and moderating effectsof parental monitoring and caregiving behavior. Journal of Abnormal Child Psy-chology, 29, 293–304.
Brown, K., Atkins, M. S., Osborne, M. L., & Milmanow, M. (1996). A revised teacher
Proactive and Reactive Aggression 197

rating scale for reactive and proactive aggression. Journal of Abnormal Child Psy-chology, 24, 473–480.
Bushman, B. J., & Anderson, C. A. (2001). Is it time to pull the plug on the hostile versusinstrumental aggression dichotomy? Psychological Review, 108, 273–279.
Buss, A. H. (1961). The psychology of aggression. New York: Wiley.Campbell, D. T., & Fiske, D. W. (1959). Convergent and discriminant validation by the
multi-trait/multi-method matrix. Psychological Bulletin, 56, 81–105.Caplan, M., Vespo, J., Peterson, J., & Hay, D. F. (1991). Conflict and its resolution in
small groups of one- and two-year-olds. Child Development, 62, 1513–1524.Chaux, E., Arboleda, J., & Rincón, C. (2003, April). A self-reported measure of reactive
and proactive aggressive behavior. Paper presented at the biennial meeting of the So-ciety for Research in Child Development, Tampa, FL.
Cloninger, C. R. (1986). A unified biosocial theory of personality and its role in the devel-opment of anxiety states. Psychiatric Development, 3, 167–226.
Coie, J. D., Dodge, K. A., Terry, R., & Wright, V. (1991). The role of aggression in peer re-lations: An analysis of aggression episodes in boys’ play groups. Child Develop-ment, 62, 812–826.
Connor, D. F., Steingard, R. J., Anderson, J. J., & Melloni, R. H. (2003). Gender differ-ences in reactive and proactive aggression. Child Psychiatry and Human Develop-ment, 33, 279–294.
Cornell, D. G., Benedek, E. P., & Benedek, D. M. (1987). Juvenile homicide: Prior adjust-ment and a proposed topology. American Journal of Orthopsychiatry, 53, 383–393.
Cornell, D. G., Warren, J., Hawk, G., Stafford, E., Oram, G., & Pine, D. (1996). Psy-chopathy in instrumental and reactive violent offenders. Journal of Consulting andClinical Psychology, 64, 783–790.
Crick, N. R., & Dodge, K. A. (1996). Social information-processing mechanisms in reac-tive and proactive aggression. Child Development, 67, 993–1002.
Crick, N. R., & Grotpeter, J. K. (1995). Relational aggression, gender, and social psycho-logical adjustment. Child Development, 66, 710–722.
Day, D. M., Bream, L. A., & Paul, A. (1992). Proactive and reactive aggression: An analy-sis of subtypes based on teacher perceptions. Journal of Clinical Child Psychology,21, 210–217.
Dodge, K. A. (1991). The structure and function of reactive and proactive aggression. InD. J. Pepler & K. H. Rubin (Eds.), The development and treatment of childhood ag-gression (pp. 201–218). Hillsdale, NJ: Erlbaum.
Dodge, K. A., & Coie, J. D. (1987). Social information processing factors in reactive andproactive aggression in children’s peer groups. Journal of Personality and SocialPsychology, 53, 1146–1158.
Dodge, K. A., Lansford, J. E., Salzer Burks, V., Bates, J. E., Pettit, G. S., Fontaine, R., &Price, J. M. (2003). Peer rejection and social information-processing factors in thedevelopment of aggressive behavior problems in children. Child Development, 74,374–393.
Dodge, K. A., Lochman, J. E., Harnish, J. D., Bates, J. E., & Pettit, G. S. (1997). Reactiveand proactive aggression in school children and psychiatrically impaired chroni-cally assaultive youth. Journal of Abnormal Psychology, 106, 37–51.
Dollard, D. J., Doob, L. W., Miller, N. E., Mowrer, O. H., & Sears, R. R. (1939). Frustra-tion and aggression. New Haven, CT: Yale University Press.
Farrington, D. P., & Loeber, R. (2000). Some benefits of dichotomization in psychiatricand criminological research. Criminal Behavior and Mental Health, 10, 100–122.
198 T H E D E V E L O P M E N T O F A G G R E S S I O N

Feshback, S. (1964). The function of aggression and the regulation of aggressive drive.Psychological Review, 71, 257–272.
Gray, J. A. (1982). The neuropsychology of anxiety. New York: Oxford University Press.Harpur, T. J., Hakstian, A. R., & Hare, R. D. (1989). Two-factor conceptualization of
psychopathy: Construct validity and assessment implications. Psychological As-sessment, 1, 6–17.
Holmes, D. S., & Will, M. J. (1985). Expression of interpersonal aggression by angeredand nonangered persons with the type A and type B behavior patterns. Journal ofPersonality and Social Psychology, 48, 723–727.
Hoving, K. L., Wallace, J. R., & La Forme, G. L. (1979). Aggression during competition:Effects of age, sex, and amount and type of provocation. Genetic PsychologyMonographs, 99, 251–289.
Hubbard, J. A., Dodge, K. A., Cillessen, A. H., Coie, J. D., & Schwartz, D. (2001). Thedyadic nature of social information processing in boys’ reactive and proactive ag-gression. Journal of Personality and Social Psychology, 80, 268–280.
Hubbard, J. A., Smithmyer, C. M., Ramsden, S. R., Parker, E. H., Flanagan, K. D.,Dearings, K. F., Relyea, N., & Simons, R. F. (2002). Observational, physiological,and self-report measures of children’s anger: Relations to reactive versus proactiveaggression. Child Development, 73, 1101–1118.
Kerr, M., Tremblay, R. E., Pagani, L., & Vitaro, F. (1997). Boys’ behavioral inhibitionand the risk of later delinquency. Archives of General Psychiatry, 54, 809–816.
King, R. J., Jones, J., Scheurer, J. W., Curtis, D., & Zarcone, V. P. (1990). Plasma–cortisolcorrelates of impulsivity and substance abuse. Personality and Individual Differ-ences, 11, 287–291.
Klimes-Dougan, B., Hastings, P. D., Granger, D. A., Usher, B. A., & Zahn-Waxler, C.(2001). Adrenocortical activity in at-risk and normally developing adolescents: In-dividual differences in salivary cortisol basal levels, diurnal variation, and responsesto social challenges. Development and Psychopathology, 13, 695–719.
Kolko, D. J., & Brown, E. J. (1997, June). Reactive and proactive aggression in clinicallyreferred disruptive children: Social and cognitive characteristics. Paper presented atthe International Society for Research in Child and Adolescent Psychopathology,Paris, France.
Lansford, J. E., Dodge, K. A., Pettit, G. S., & Bates, J. E. (2002). Developmental trajecto-ries of reactive and proactive aggression: Similarities and differences over time. Pa-per presented at the International Society For Research on Aggression, Montreal,QC.
Little, T. D. (2002). Pathways of the forms and functions of aggression during adoles-cence. Paper presented at the biennial meeting of International Society for Researchon Aggression, Montreal, QC.
Little, T. D., Braumer, J., Jones, S. M., Nock, M. K., & Hawley, P. H. (2003a). Rethinkingaggression: A typological examination of the functions of aggression. Merrill-Palmer Quarterly, 49, 343–369.
Little, T. D., Jones, S. M., Henrich, C. C., & Hawley, P. H. (2003b). Disentangling the“whys” from the “whats” of aggressive behavior. International Journal of Behav-ioral Development, 27, 122–133.
Lochman, J. E. (1992). Cognitive-behavioral intervention with aggressive boys: Three-year follow-up and preventive effects. Journal of Consulting and Clinical Psychol-ogy, 60, 426–432.
Lochman, J. E., & Wells, K. C. (2002). The Coping Power Program at the middle school
Proactive and Reactive Aggression 199

transition: Universal and indicated prevention effects. Psychology of Addictive Be-haviors, 16, 40–54.
Lochman, J. E., & Wells, K. C. (2004). Effectiveness of the Coping Power Program and ofclassroom intervention with aggressive children: Outcomes at a 1-year follow-up.Behavior Therapy, 34, 493–515.
McBurnett, K., Lahey, B. B., Frick, P. J., Risch, C., Loeber, R., Hart, E. L., Christ, M. A.G., & Hanson, K. S. (1991). Anxiety, inhibition, and conduct disorder in children:II. Relation to salivary cortisol. Journal of the American Academy of Child and Ad-olescent Psychiatry, 30, 192–196.
Nagin, D. S. (1999). Analyzing developmental trajectories: A semi-parametric, group-based approach. Psychological Methods, 4, 139–177.
Neale, M. C., & Cardon, L. R. (1992). Methodology for genetic studies of twins and fam-ilies. New York: Kluwer Academic/Plenum Press.
Parke, R. D., & Slaby, R. G. (1983). The development of aggression. In E. M. Hethering-ton (Ed.), Socialization, personality, and social development (4th ed.: Vol. 4, pp.547–641). New York: Wiley.
Pellegrini, A. D., Bartini, M., & Brooks, F. (1999). School bullies, victims, and aggressivevictims: Factors relating to group affiliation and victimization in early adolescence.Journal of Educational Psychology, 91, 216–224.
Phillips, N. C., & Lochman, J. E. (2003). Experimentally manipulated change in children’sproactive and reactive aggressive behavior. Aggressive Behavior, 29, 215–227.
Poulin, F., & Boivin, M. (2000a). Reactive and proactive aggression: Evidence of a two-factor model. Psychological Assessment, 12, 115–122.
Poulin, F., & Boivin, M. (2000b). The role of proactive and reactive aggression in the for-mation and development of boys’ friendships. Developmental Psychology, 36, 233–240.
Poulin, F., & Dishion, T. J. (2000). The peer and family experiences of proactively and re-actively aggressive pre-adolescents. Paper presented at the biennial meeting of theSociety for Research on Adolescence, Chicago, IL.
Poulin, F., Dishion, T. J., & Boivin, M. (2002). Proactive/reactive aggression and adjust-ment problems in adolescence: A look at peer and family processes. Paper presentedat the biennial meeting of the International Society for Research on Aggression,Montreal, QC.
Price, J. M., & Dodge, K. A. (1989). Reactive and proactive aggression in childhood: Re-lations to peer status and social context dimensions. Journal of Abnormal ChildPsychology, 17, 455–471.
Prince, J. E., & Arias, I. (1994). The role of perceived control and the desirability of con-trol among abusive and nonabusive husbands. American Journal of Family Ther-apy, 22, 126–134.
Prinstein, M. J., & Cillessen, A. H. N. (2003). Forms and functions of adolescent peer ag-gression associated with high levels of peer status. Merrill-Palmer Quarterly, 49,310–342.
Pulkkinen, L. (1987). Offensive and defensive aggression in humans: A longitudinal per-spective. Aggressive Behavior, 13, 197–212.
Pulkkinen, L. (1996). Proactive and reactive aggression in early adolescence as precur-sors to anti- and prosocial behaviors in young adults. Aggressive Behavior, 22, 241–257.
Salmivalli, C., & Nieminen, E. (2002). Proactive and reactive aggression among schoolbullies, victims, and bully-victims. Aggressive Behavior, 28, 30–44.
200 T H E D E V E L O P M E N T O F A G G R E S S I O N

Scarpa, A., Hurley, J. D., & Hirai, M. (2002). Emotional and behavioral correlates of re-active and proactive aggression in children. Paper presented at the 15th WorldMeeting of the International Society for Research on Aggression, Montreal, QC.
Schippel, P. L., Vasey, M. W., Cravens-Brown, L. M., & Bretveld, R. A. (2003). Sup-pressed attention to rejection, ridicule, and failure cues: A unique correlate of reac-tive but not proactive aggression in youth. Journal of Clinical Child and AdolescentPsychology, 32, 40–55.
Smithmyer, C. M., Hubbard, J. A., & Simons, R. F. (2000). Proactive and reactive aggres-sion in delinquent adolescents: Relations to aggression outcome expectancies. Jour-nal of Clinical Child Psychology, 29, 86–93.
Susman, E. J., Dorn, L. D., Inoff-Germain, G., Nottelmann, E. D., & Chrousos, G. P.(1997). Cortisol reactivity, distress behavior, and behavioral and psychologicalproblems in young adolescents: A longitudinal perspective. Journal of Research onAdolescence, 7, 81–105.
Tremblay, R. E. (2000). The development of aggressive behavior during childhood: Whathave we learned in the past century? International Journal of Behavioral Develop-ment, 24, 129–141.
Van Bokhoven, I., Van Goozen, S. H. M., van Engeland, H., Schaal, B., Arseneault, L.,Séguin, J. R., Nagin, D. S., Vitaro, F., & Tremblay, R. E. (2003). Salivary cortisoland aggression in a population based sample of adolescent males. Manuscript sub-mitted for publication.
van Goozen, S. H. M., Matthys, W., Cohen-Kettenis, P. T., Buitelaar, J. K., & VanEngeland, H. (2000). Hypothalamic–pituitary–adrenal axis and autonomic ner-vous system activity in disruptive children and matched controls. Journal of theAmerican Academy of Child and Adolescent Psychiatry, 39, 1438–1445.
Vitaro, F., Brendgen, M., & Tremblay, R. E. (2002). Reactively and proactively aggres-sive children: Antecedent and subsequent characteristics. Journal of Child Psychol-ogy and Psychiatry, 43, 495–505.
Vitaro, F., Gendreau, P. L., Tremblay, R. E., & Oligny, P. (1998). Reactive and proactiveaggression differentially predict later conduct problems. Journal of Child Psychol-ogy and Psychiatry and Allied Disciplines, 39, 377–385.
Waschbusch, D. A., Willoughby, M. T., & Pelham, W. E. (1998). Criterion validity andthe utility of reactive and proactive aggression: Comparisons to attention deficithyperactivity disorder, oppositional defiant disorder, conduct disorder, and othermeasures of functioning. Journal of Clinical Child Psychology, 27, 396–405.
Woodman, D. D., Hinton, J. W., & O’Neill, M. T. (1978). Cortisol secretion and stress inmaximum security hospital patients. Journal of Psychosomatic Research, 22, 133–136.
Zillman, D. (1979). Hostility and aggression. Hillsdale, NJ: Erlbaum.
Proactive and Reactive Aggression 201

THE DEVELOPMENT OF AGGRESSIONHomicide, Violence, and Developmental Trajectories
10
Homicide, Violence,and Developmental Trajectories
ROLF LOEBER, ERIC LACOURSE, and D. LYNN HOMISH
The title of this book, Developmental Origins of Aggression, necessarily focuseson childhood as the period in which many children display aggression and out-grow aggression (Tremblay et al., 2004). More contentious is whether persistentaggression can first emerge after the first few years of life and how important it isto study escalation in the severity of aggression from childhood onward. In thischapter we do not deal with the onset of aggression in the preschool period.Instead, we focus on two aspects of violence processes: late onset and escalationprocesses, especially processes leading to violence and homicide.
Typically, the onset of violence (such as rape, robbery, and aggravated as-sault) and homicide takes place in late adolescence and early adulthood(Snyder & Sickmund, 1999). It is vastly different in form as compared withtypical forms of aggression during childhood, involving more force, moretrauma to the victim’s body, and often the use of a lethal weapon such as agun. Homicide is the most serious forms of aggression. We are particularly in-terested in tracing the developmental origins of homicide between childhoodand early adulthood. An important assumption about individuals who com-mit homicide is that they have shown violent behaviors consistently earlier inlife. However, this assumption has been challenged (Heide, 2003), and we re-examined it with prospectively collected data rather than using retrospectivedata, which are typical of almost all homicide studies.
In this chapter we address three key questions:
1. Are there developmental precursors to homicide?2. Which developmental trajectories in violence are particularly relevant
202

to homicide, and are most homicide offenders positioned on the high-est and most stable trajectory of violent individuals?
3. What is the relevance of the findings for interventions and the preven-tion of aggression, violence, and homicide?
We address these questions in the context of two theoretical conceptualiza-tions of the explanation of delinquency: population heterogeneity and statedependency.
VIOLENCE AND HOMICIDE
In the United States alone, during 1999 there were 46 homicides, 1,050 sexualassaults and 2,220 robberies committed each day. Homicide is known to bethe second leading cause of death for persons ages 15–24 (Federal Bureau ofInvestigation, 2001). During the past two decades the overall rate of homicideand violence has varied. From the early 1980s through the early 1990s, the ju-venile homicide rate increased substantially to reach a peak of 14 homicidesper 100,000 persons in 1993. The increase in violence during the 1990s wasso spectacular that it was described by many scholars as an unprecedented ep-idemic (Cook & Laub, 1998; Cook & Laub, 2002; Blumstein, 2002). This ep-idemic was mostly characterized by an increase in gun-related homicides. Sub-sequently, the homicide rate dropped dramatically to around 6 per 100,000 inthe United States and 1.9 per 100,000 in Canada in the early 2000s. Althoughthis apparent decline in overall homicide offending is encouraging, the homi-cide rate today is at approximately the same rate as it was in the early 1980s.Strikingly, this increase and drop in violence affected specific age segments ofthe population, teenagers and young adults. Although this apparent decline inoverall homicide offending in youth is encouraging, it remains a very impor-tant issue inasmuch as the youth homicide and violence rate today remainshigh. More research is needed to better understand the developmental hetero-geneity of violence in the population and the environmental and proximal fac-tors that can increase the escalation and lethality of violence in individuals.
HETEROGENEITY OF HOMICIDE OFFENSES AND HOMICIDE OFFENDERS?
The field of homicide studies is torn by opposites. On one hand, each homi-cide is unique in the circumstance in which the act is committed and the char-acteristics, motivations, and setting in which the homicide offender meets hisor her victim. Sometimes, the difference between who will be the homicide of-fender rather than the homicide victim will be decided at the last moment anddepends on strength, the weapon used, accuracy, and other circumstances. Onother hand, criminologists are seeking principles and classifications behindmultiple instances of murder (Heide, 2003). For example, homicides have typ-
Homicide, Violence, and Developmental Trajectories 203

ically been classified on the basis of the victim characteristics, such as infanti-cide and parricide, or on the basis of frequency of homicides committed (sin-gle vs. serial homicide offenders). Other classification schemes concern themotives for homicide, such as gang-related homicide, homicide in retaliationto prior homicide, and family–intimate partners homicide. Other typologieshave focused on the circumstances of the offense, including psychotic epi-sodes, conflict, and crime circumstances leading to homicide (Heide, 2003).What these classifications have in common is the reliance on the basis of a sin-gle event (leaving aside multiple or serial murderers, who are usually veryuncommon). We argue that a classification based on a single incident at onepoint in time is hazardous because it contains too little information to revealthe following key aspects: (1) the individual’s propensity for violence and (2)the etiology of violence and homicide.
To concentrate on the first point, the key forensic question is whether ju-venile homicide offenders from inner cities with high levels of community vio-lence have higher levels of propensity for violence other than homicide. To an-swer this question, it is necessary to obtain information about the offender’shistory of violence and aggression (such as bullying, physical fighting withoutcausing wounds) as a child or adolescent. Several possibilities exist. First, onemight suppose that most homicide offenders have been highly aggressive andviolent for years, and that committing a homicide results from a process ofgradual escalation in violence, in which early forms of aggression (such as bul-lying) set the stage for physical fighting, which in turn set the stage for vio-lence. Another train of thought is that some homicide offenders are notknown for a highly violent career, but even at an intermittent or moderatelevel of violence they still are at risk of killing someone. A third possibility isthat even among those juveniles without a history of violence, there are somewho may “lose it” on a rare occasion and murder somebody. Heide (2003)concluded in her review of homicide offenders that all three possibilities mightapply and that “findings regarding whether young killers have had a lengthyhistory of fighting and other acts of violence have been mixed” (p. 13). Thisconclusion, however, is mostly based on case studies and chart reviews ratherthan prospective follow-up studies.
Developmental approaches to the classification of violent offenders thattake individuals’ past history of aggression and violence into account have ad-dressed two aspects: developmental pathways leading to violence and seriousproperty crime, and developmental trajectories of violence and property of-fenders throughout the life course.
Developmental Pathways
A developmental pathway is defined as the behavioral development of a groupof individuals that is different from the behavioral development of othergroup(s) of individuals (Loeber, DeLamatre, Keenan, & Zhang, 1998a). Thefirst question is what dimensions should be used to plot individuals’ progres-
204 T H E D E V E L O P M E N T O F A G G R E S S I O N

sion to serious property crime and violence. The second question is whetherthe development toward violence occurs in an orderly way or randomly. Andthe third question pertains to the particular stages in the development of vio-lence and serious property crime.
We addressed these issues in our research on developmental pathways toviolence and serious property offenses. We aimed at creating a model thatwould specify heterotypic continuity of disruptive behaviors (Loeber et al.,1993), particularly from minor to serious forms of delinquency.
There has been considerable debate about whether all juvenile problembehaviors should be considered sufficiently similar to be covered by a singleconstruct. Some have advocated a general deviance or problem construct (e.g.,Jessor, Donovan, & Costa, 1991; Robins, 1966), whereas others have advo-cated a more differentiated approach (e.g., Loeber, 1988; McCord, 1990;Osgood, Johnston, O’Malley, & Bachman, 1988). The latter position is sup-ported by accumulating empirical evidence for a distinction between overt(e.g., aggression and violence) and covert (e.g., theft and fraud) behaviorproblems (e.g., Frick et al., 1993; Loeber & Schmaling, 1985; Loeber et al.,1993).
We found that antisocial behavior was best represented by a model ofthree pathways (Figure 10.1). The steps in each pathway are developmentallyarranged by average age of onset. The pathways are as follows: (1) an author-ity conflict pathway prior to the age of 12, which started with stubborn be-havior, had defiance/disobedience at a second stage, and authority avoidance(e.g., truancy) at a third stage, (2) a covert pathway emerging before age 15,
Homicide, Violence, and Developmental Trajectories 205
FIGURE 10.1. Developmental pathways to serious and violent offending.

which started with minor covert acts, had property damage at a second stage,and moderate to serious delinquency at a third stage, and (3) an overt path-way, which started with minor aggression, had physical fighting at a secondstage, and more severe violence at a third stage. Typically, individuals who es-calated to the most serious delinquent acts progressed along the steps specifiedin the model. The results of these studies showed that with age, a proportionof boys progressed on two or three pathways, indicating an increasing varietyof problem behavior over time (Kelley, Loeber, Keenan, & DeLamatre, 1997;Loeber et al., 1993; Loeber, Keenan, & Zhang, 1997).
We found some evidence that development in more than one pathwaywas asymmetric; that is, boys who were escalating in the overt pathway wereat risk of also escalating in the covert pathway, but not vice versa. Further, es-calation in either the overt or covert pathway was often preceded by boys’ es-calation in the authority conflict pathway (Loeber et al., 1993). In otherwords, conflict with authority figures was either a precursor or a concomitantof boys’ escalation in overt or covert acts. Moreover, an early age of onset ofproblem behavior or delinquency, as compared with onset at a later age, wasassociated with boys’ escalation to more serious behaviors (Tolan, Gorman-Smith, & Loeber, 2000). The pathway findings have been replicated in sam-ples of Caucasian, African American, and Hispanic males in several cities(Loeber et al., 1998a; Loeber, Wei, Stouthamer-Loeber, Huizanga, & Thorn-berry, 1999; Loeber et al., 1993; Tolan et al., 2000). In summary, the pathwaymodel allows several conceptualizations of development toward increasinglyserious disruptive and delinquent behavior, such as escalation within a singleor multiple pathway(s), heterotypic continuity, and age of onset of problembehaviors.
Developmental Trajectories
Major work has been undertaken in the past decade to document individuals’trajectories of crime and delinquency (Nagin & Land, 1993; Osgood &Rowe, 1994; Horney, Osgood, & Marshall, 1995). For example, aggressionand violence have been studied from early childhood to adulthood (Brame,Nagin, & Tremblay, 2001; Broidy et al., 2003; Lacourse et al., 2002; Laub &Sampson, 2003; Nagin & Tremblay, 1999; Tremblay et al., 2004). Trajec-tories approach developmental aspects of violence from a different angle.There have been two major trends in the study of developmental trajectories.One trend, random effect growth modeling, is related to important advancesin structural equation and multilevel modeling during the past decades (Bryk& Raudenbush, 1987; Meredith & Tisak, 1990; Rogosa & Willet, 1985).Random effect growth modeling has been a very popular analytical techniquein developmental psychology and criminology since it provides analyticaltools that favor more dynamic explanations of repeated measures data ascompared with more traditional approaches such as analysis of variance orsimplex regression. A shortcoming of these methods is that they are not well
206 T H E D E V E L O P M E N T O F A G G R E S S I O N

suited for testing theories that postulate qualitatively different developmentaltrajectories of antisocial behaviors resulting from possibly distinct causal pro-cesses (Moffitt, 1993; Patterson & Yoerger, 1993, 1997). In recent years sta-tistical analyses have been developed to identify developmental trajectories ofunobserved subgroups in the population, described as latent trajectory classes(Jones, Nagin, & Roeder, 2001; Muthen, 1989, 2001; Nagin, 1999; Nagin &Land, 1993). Typically, group-based trajectory analyses assume that a popula-tion of individuals “is composed of a mixture of groups with distinct develop-mental trajectories” (Nagin & Tremblay, 2001, p. 21). Trajectory analyses ofaggressive and violent behaviors have usually been based on few repeatedlymeasured indicators of disruptive behavior and on the frequency of behavior(thus, different than the diversity of behaviors considered in the pathwaysmodel). They have also been less focused on qualitative changes over time.The analyses, primarily done with data on the period between childhood andadolescence, usually led to the identification of four more easily interpretabletrajectories: individuals whose aggressive and violent behavior remains highover time, individuals who remain nonaggressive, and individuals whose vio-lent behavior increases or decreases with development (e.g., Brame et al.,2001; Broidy et al., 2003; Fergusson & Horwood, 1995; Lacourse et al.,2002; Nagin & Tremblay, 1999). The key advantage of group-based trajec-tory analyses is that a statistical estimation procedure using mixture modelingidentifies the shape of the trajectory for each group, and the prevalence in thepopulation that constitutes each group (Nagin, 1999). However, few studiesusing trajectory analyses have shed light on heterotypic continuity of differentproblem behaviors as they unfold over time (see Brame et al., 2001; Nagin &Tremblay, 2001, for examples of heterotypic continuity models). Furthermore,homicidal behavior is not often measured in surveys using self-reported delin-quency questionnaires (e.g., Elliott, Huizinga, & Ageton, 1985; Loeber et al.,in press). For these reasons, there are no studies yet that have shown the loca-tion of homicide offenders on developmental trajectories of violence that varyin terms of severity.
DEVELOPMENTAL ASPECTS OF HOMICIDE
The notion that delinquency and violence develop gradually in individuals hasgained widespread acceptance (e.g., Blumstein, Cohen, Roth, & Visher, 1986;Loeber & Le Blanc, 1990; Loeber & Farrington, 1998). There is also a con-sensus that individual differences in offending are quite stable and that initialindividual differences persist over time (e.g., Moffitt, 1993). Although wehave described how manifestations of aggression change during adolescence,it is also important to understand how manifestations of aggression are re-lated to each other during different age periods. It could be important to dis-tinguish between different types of violence trajectories that have differentbeginnings and differently unfold over time. It appears clear from the research
Homicide, Violence, and Developmental Trajectories 207

in psychology and criminology that there is a moderate-to-strong positive cor-relation between past and future aggressive or violent behaviors (Olweus,1979). Although this correlation has been acknowledged by researchers, theydiffer as to its meaning. Drawing from the work of Heckman (1981), Naginand Paternoster (1991, 2000) concluded that criminal behaviors that includeviolent offends could be related to two processes, population heterogeneityand state dependence.
Population Heterogeneity Theories
Theories of population heterogeneity suggest that there is an initial pronenessto commit violent offenses and that this early difference in the development ofan individual remains quite stable over time (Gottfredson & Hirschi, 1990;Moffitt, 1993; Caspi, Lynam, Moffitt, & Silva, 1994). An individual with nu-merous risk factors before birth, in toddlerhood, or in childhood would bemore likely to act with violence during adolescence and adulthood. This is notthe place to review the variety of risk factors that appear associated with vio-lence (see, for example, Loeber & Farrington, 1998; Raine, 2002; Thornberry,Freeman-Gallant, Lizotte, Krohn, & Smith, 2003). Yet, it is important toknow that research on population heterogeneity for explanations of crime andviolence is broad, focusing on topics such as early biological differences, neu-rological deficits, temperament, personality, parenting, and early child care. Acommonality in population heterogeneity models of violence is that “any ob-served correlation between past and future criminal offending is due to endur-ing differences between individuals in an initial proneness of propensity tocommit crime” (Nagin & Paternoster, 2000, p. 124).
State Dependence and Escalation Theories
The state dependence position suggests that actions and events that occur in aperson’s life affect the probability of subsequent actions or events. State de-pendence is a dynamic process that evolves in two ways: (1) aggressive behav-ior can open opportunities for more aggressive behavior while closing oppor-tunities for other kinds of behaviors and (2) nonaggressive behaviors couldopen up possibilities for other nonaggressive behaviors and close opportuni-ties for aggressive behaviors. The state dependence position suggests that lifeevents that occur later in the development of an individual have a direct causaleffect on violence while controlling for prior proneness to commit violent acts.
There are many classical and modern theories in criminology and psy-chology that use state dependence as a primary explanation for crime and vio-lence. Labeling and symbolic interaction theories (Lemert, 1951; Becker,1963; Matsueda & Heimer, 1997), social learning theory (Akers, 1985;Bandura, Ross, & Ross, 1961; Coie & Dodge; 1988), general strain theory(Agnew, 1992) and social interactional theory (Thornberry, 1987) are all ex-amples of theories that focus on state dependence. For example, labeling theo-
208 T H E D E V E L O P M E N T O F A G G R E S S I O N

rists argue that the likelihood of future criminal or violent behavior is closelyrelated to the reaction of social control agencies to the first aggressive or vio-lent behavior of the individual. If an individual is labeled as aggressive, he orshe will define himself as aggressive and will act accordingly. Social learningand interactional theories focus their research on the reinforcement contingen-cies in the environment that will maintain or extinguish the aggressive or violentbehavior. Family and peer interactions are of primary importance in these the-ories. Coercive family interactions and deviancy training by peers are thoughtto be important state-dependent factors that explain stability and change in vi-olent behavior over time. Preventive intervention with high-risk children canalso be thought of as a state dependence process that can modify a person’sdevelopmental trajectory (see Lacourse et al., 2002, for an example).
Mixed Theories
Although population heterogeneity and state dependence appear as incompat-ible processes, some theories have tried to explain violence using a mixture ofthese two processes. The blending of developmental psychology and criminol-ogy during the past 10 years and the strong interest in the age–crime curvehave helped to set aside the more traditional focus on interindividual differ-ences to put more focus on intra-individual development and the life course ofoffending. The confrontation between ontogenetic and sociogenetic theoriesor, in other words, between population heterogeneity and state dependencetheories fostered the integration of theories that could be considered mixed. Incriminology the blend of these two perspectives was pioneered by Le Blancand Fréchette (1989), Loeber and Le Blanc (1990), and Sampson and Laub(1993), whereby the link between childhood and adolescent experiences andadulthood life circumstances were combined into one theoretical framework.For example, Sampson and Laub (1993), in their age-graded theory, showedthat the self-selection thesis, or early individual difference model, was sup-ported, but that important life events in late adolescence such as military en-rollment, a good marriage, employment, parenthood, and an advantagedneighborhood (low crime, high employment opportunities) were also thoughtto reduce crime and violence. Recent studies using the semiparametric trajec-tory models have confirmed the tenability of mixed theories of crime and vio-lence (Lacourse et al., 2002; Lacourse, Nagin, Tremblay, Vitaro, & Claes,2003; Laub, Nagin, & Sampson, 1998; Laub & Sampson, 2003; Nagin,Pagani, Tremblay, & Vitaro, 2003). The mixed model appears to apply as wellto homicide.
PREDICTORS OF HOMICIDE
Using data from the Pittsburgh Youth Study (Loeber, Farrington, Stouthamer-Loeber, & Van Kammen, 1998b), we briefly present a mixed model predicting
Homicide, Violence, and Developmental Trajectories 209

violence and homicide among young urban males. Virtually all the studies thathave tried to understand causes or factors that could explain homicide offend-ing in youth during the past 50 years have relied on case studies and studiesusing a retrospective design (Cornell, 1989; Ewing, 1990; Heide, 2003). Mostof the case studies were done with male youth who were referred to psychiat-ric facilities for evaluation and treatment following their perpetration of ho-micide (Heide, 2003). Given the professional background of the clinician/re-searchers, it is clear that most descriptive explanations of homicidal behaviorwere related to population heterogeneity factors such as mental illness, neuro-logical deficits, intelligence, and childhood trauma and abuse. In this sectionwe briefly describe recent findings from the Pittsburgh Youth Study, a longitu-dinal empirical study in which three samples of schoolboys were followed upfrom childhood to early adulthood. The study sheds light on population heter-ogeneity and state dependence factors that could explain differences betweenviolent offenders, homicide offenders, and nonviolent delinquent youth (Loeberet al., in press).
By the middle of 2003, 129 participants had been convicted of vio-lence, and another 306 self-reported violence. Thirty-three participants hadbeen convicted in court of homicide (2.2%). Normally, it is impossible topredict low-base-rate events such as homicide, because almost all individualsdo not commit homicide, and the best prediction would be that 97.8%would not commit homicide. A way to overcome the low-base-rate prob-lem, however, is to use knowledge of state dependency to improve the pre-diction. We first looked at which delinquent behavior was shared by mostof the homicide offenders. The results showed that almost all (93.9%) ofthe homicide offenders had been violent prior to the homicide (a probableindication of state dependence). Based on this finding, we decided that thebest strategy would be to first predict who would become violent and thenpredict who would become a homicide offender among the violent offend-ers. The results of the prediction of violence showed support for the popula-tion heterogeneity hypothesis, in that predictors of violence were observedin the domains of the individual child, child attitudes, family, school, anddemographics. The following factors predicted violence in univariate analy-ses: Factors evident early in life (such as the mother’s cigarette or alcoholuse during pregnancy), onset of delinquency prior to age 10, and acting outproblem behavior, including serious markers such as physical aggression,cruelty, and callous/unemotional behavior. In addition, cognitive factors(such as low school motivation) predicted violence. Poor and unstable childrearing factors also contributed to the prediction of violence, including twoor more caretaker changes prior to age 10, physical punishment, poor su-pervision and communication, and several other family factors. Undesirableor delinquent peer behavior, according to either parent report or self-report,predicted violence. Poor school performance and truancy were also amongthe predictors of violence. Finally, demographic factors indicative of familydisadvantage (low family SES, welfare, teenage motherhood) and residence
210 T H E D E V E L O P M E N T O F A G G R E S S I O N

in a disadvantaged neighborhood also predicted violence. Among the proxi-mal factors associated with violence were weapon carrying, weapon use,gang membership, drug selling, and persistent drug use. Those who scoredpositively for three or more risk factors out of a total violence risk score of12 were six times more likely to later commit violence as compared withthose with fewer risk factors (odds ratio = 6.30).
The next step was to predict those who would commit homicide amongthe violent offenders. Homicide offenders, as compared with violent offend-ers, were more likely to carry a weapon, to qualify for a diagnosis of conductdisorder, and to sell hard drugs. In addition, homicide offenders were morelikely to have a positive attitude to substance use, to have delinquent peers,and to be held back in school. Homicide offenders were more likely to have ahigh risk score at the beginning of the study (i.e., at ages 7, 10, or 13), have atleast one disruptive behavior diagnosis, and to be African American. The ho-micide risk index was based on the presence or absence of nine significant riskfactors for homicide offenders (screening risk score, positive attitude to sub-stance use, conduct disorder, carrying a weapon, gang fight, selling harddrugs, peer delinquency, being held back in school, and having a family onwelfare). A homicide risk score based on these factors (excluding race) showedthat those with 4 or more of the risk factors were 14 times (odds ratio =14.33) more likely to be convicted for homicide than violent offenders with alower risk index.
The results underscore the applicability of the state dependency model tothe prediction of both violence and homicide. However, the population heter-ogeneity model applied better to the prediction of violence than to the predic-tion of homicide among the violent offenders. In the latter instance, more indi-vidual factors, rather than family and neighborhood factors, contributed tothe prediction of homicide as compared with violence. Less distinctive werepeer factors, which predicted both homicide and violence.1
HOMICIDE OFFENDERS AND VIOLENCE TRAJECTORIES
The observation that almost all of the homicide offenders had been violentearlier in life does not tell us whether most homicide offenders derive fromthose who were characterized by a consistently high level of violence overtime. Recall that Heide’s (2003) review of homicide offenders spoke of mixedevidence that all homicide offenders were characterized by a lengthy history offighting and other violent behavior.
To test this important issue, we used a violence seriousness scale to firstmodel the number and the shape of developmental trajectories. Rating on thisscale had three levels: (0) no violence; (2) moderate violence: gang fighting orcarrying a weapon; (3) serious violence: attack to seriously hurt or kill. Ratingwas based on information provided by the teacher, the parents, and the youthhimself. Each individual’s score on this scale represents the most serious vio-
Homicide, Violence, and Developmental Trajectories 211

lent offense committed during a given time interval (thus, giving less weight topresence of less serious offenses). The best model was chosen using the BICstatistic, which identifies the optimal number of groups in mixture models ofdevelopmental trajectories (D’Unger, Land, McCall, & Nagin, 1998). Asshown in Figure 10.2, a four-group model was selected as the best-fittingmodel to explain the relationships between violence scores over the 12 assess-ment waves. A main finding is that violence seriousness trajectories stronglydecline and become close to zero between the ages of 20 and 24. Another im-portant finding is that the mixture model did not identify any adolescent- oradulthood-onset trajectories and there were more trajectories than the numbergenerally theoretically speculated (Moffitt, 1993; Patterson & Yoerger, 1993,1997). One group representing 4.7% of the sample appears to increase itslevel of violence from ages 14–20 and to slowly decline until age 24, slightlybelow the initial level at age 14 (called the chronic group). A second group,22.4%, starts off at the same level but shows a steep decline that reaches zeroby age 24 (called the late desister group). A third group, 21.9%, starts off at alower level and shows a slow decline that also reaches zero by age 24 (calledthe early desister group). A last group displays, on average, no violence duringthe whole time period (the low group). This specific group represents 51.0%of the overall sample. These results are in line with previous research thatshows a strong decline in trajectories of violent crimes during the transitionbetween late adolescence and adulthood (Laub & Sampson, 2003).
How do these data look at the individual level? Figure 10.3 presents thetrajectories of four randomly selected homicide offenders. The trajectories
212 T H E D E V E L O P M E N T O F A G G R E S S I O N
FIGURE 10.2. Trajectories of violence between ages 14 and 24.

show that these homicide offenders have experienced violence during adoles-cence. Most of these trajectories show some decline during adulthood, whichfollows the general trend found in other samples (Laub & Sampson, 2003). Asexpected, 81.8% of the homicide offenders and 61.7% of the violent index of-fenders (with a court conviction of rape, robbery, or aggravated assault) fol-lowed the two highest trajectories of violence seriousness (the chronic groupand the late desister group; see Table 10.1). Most court-identified homicideoffenders (63.6%) and violent index offenders (53.2%) followed a trajectorythat shows declining levels of violence over time (the late desister group). Wefirst thought that these results were due to the impact of incarceration, butpreliminary results suggest that incarceration did not have a large impact onthe level of violence of individuals in this representative sample. Similar resultswere found by Laub and Sampson (2003).
What the study clearly demonstrates is that most homicide and index vio-lent offenders are individuals on developmental trajectories that are alreadypresent in early adolescence. There is substantive agreement among studiesthat the majority of serious and violent offenders experience an early onset ofoffending of any type (Loeber & Farrington, 1998, 2001). Analyses of path-ways to serious delinquency suggest that onset patterns of predelinquent prob-lems often take place during the elementary school age period, if not earlier(Loeber et al., 1993; Loeber et al., 1998a; Moffitt, 1993).
The study has several limitations. Only 11 convicted homicide offenderswere available for the analyses. Moreover, the unavailability of data prior toage 13 and after age 25, means that the analyses of trajectories were limitedby left- and right-censored data. It is possible that membership in certain tra-
Homicide, Violence, and Developmental Trajectories 213
FIGURE 10.3. Trajectories of violence for four homicide offenders.

jectories, based on the currently available data, would have been different ifearlier or later assessments had been undertaken (Laub & Sampson, 2003;Eggelston, Laub, & Sampson, 2004). The study did not attempt to determinewhich risk factors were associated with which trajectory group. Neither didthe study attempt to explain different ages of desistance for those trajectoriesthat decreased with age. Trajectory analyses, although very valuable, arepostdictive rather than predictive in that individuals’ assignment to a trajec-tory group is dependent on knowledge of their offending over a full period oftime. Instead, we need to move to predictive assignment of individuals to tra-jectories in a probabilistic manner before they reach the most serious offenselevels. This is particularly relevant for the prevention of homicide and violencein a proactive manner.
PREVENTION
We think that the findings are relevant for prevention. First, there is evidencefor state dependency and escalation, as evident from developmental pathwaysfrom less serious forms of aggression to violence and, eventually, homicide.Second, almost all homicide offenders had been violent earlier in life, whichfurther supports the state dependency hypothesis. At the same time, however,we think that there is strong support for the notion of escalation within thestate dependency paradigm, in that it takes years for aggression processes toculminate in homicide for a minority of the population. Models of escalationcan be best conceptualized as funnels (Figure 10.1) in that most individualswith the lower-level forms of aggression do not escalate to homicide or evenviolence. However, those who took the next one or two steps in the develop-mental sequence (such as physical fighting and violence) were at much greaterrisk of committing homicide later.
We also propose that as individuals escalate from minor aggression to vi-olence and homicide, population heterogeneity and diversity of risk factors/domains diminishes. Specifically, population heterogeneity is highest at thelowest level of developmental pathways (i.e., at the base of the pyramid
214 T H E D E V E L O P M E N T O F A G G R E S S I O N
TABLE 10.1. Percentage of Court-Reported Homicide and Violence Index Offenders perViolent Offending Trajectories
Violence seriousnesstrajectories
Homicide offenders (%)(n = 11)
Violence index offenders (%)(n = 47)
1 Low 0.0 23.4
2 Early desister 18.2 14.9
3 Late desister 63.6 53.2
4 Chronic 18.2 8.5

shown in Figure 10.1), but decreases as certain individuals progress and esca-late to the more serious behaviors. Thus, population heterogeneity is inverselyrelated to escalation, or, stated otherwise, population homogeneity increasesas some individuals desist from lesser forms of antisocial and delinquent be-havior, whereas others rise to become serious offenders.
In conjunction with these processes, we also postulate a narrowing of thedomains of risk factors that play a role in antisocial and delinquent behavioras individuals escalate to violence and homicide. We saw that individual, fam-ily, peer, school, and neighborhood factors all contribute to the prediction ofviolence. In contrast, however, individual factors especially account for theprediction of homicide (leaving aside peer factors, which probably are, tosome extent, confounded with individual factors).
What are the implications for prevention? First, the process of escalationusually takes years to be completed, which therefore provides ample opportu-nity for preventing individuals to commit more serious antisocial and delin-quent acts (Loeber & Farrington, 2001). Second, intervention research (Loeber& Farrington, 1998) shows that the best interventions address at least tworisk domains (and are less effective if aimed at a single risk domain). This rein-forces the notion that interventions should take place early in the delinquencycareers rather than later, when it is probable that other individual risk factorsprevail. However, because the transition of individuals’ aggression to violenceoften involves increasingly more lethal weapons and increasingly more dan-gerous activities (such as gang membership and drug dealing), other importantavenues for the prevention of violence (and homicide) should consist of the re-striction of weapons, especially guns, for those who are known to be highlyaggressive, and the encouragement of alternatives to gang membership anddrug dealing.
ACKNOWLEDGMENTS
This chapter was prepared with financial assistance of Grant Nos. 86-JN-CX0009 and96-JN-CX-0003 from the Office of Juvenile Justice and Delinquency Prevention(OJJDP), and of Grant Nos. MH48890 and MH50778 from the National Institute ofMental Health (NIMH). Points of view or opinions in this chapter are those of the au-thors and do not necessarily represent the official position of OJJDP, the Departmentof Justice, or NIMH.
NOTE
1. However, we are not certain that this is valid, because peer factors may have reflected co-offending rather than independent effects. This seems probable because information aboutpeer delinquency and peer substance use was based on the reports of the participants in thestudy.
Homicide, Violence, and Developmental Trajectories 215

REFERENCES
Agnew, R. (1992). Foundation for a general strain theory of crime and delinquency.Criminology, 30, 47–87.
Akers, R. (1985). Deviant behavior: A social learning approach (3rd ed.). Belmont, CA:Wadsworth.
Bandura, A., Ross, D., & Ross, S. A. (1961). Transmission of aggression through imita-tion of aggressive models. Journal of Abnormal and Social Psychology, 63, 575–582.
Becker, H. S. (1963). Outsiders. New York: Free Press.Blumstein A. (2002) Youth, guns, and violent crime. Future of Children, 12, 38–53.Blumstein, A., Cohen, J., Roth, J. A., & Visher, C. A. (Eds.). (1986). Criminal careers and
“career criminals. ” Washington, DC: National Academy Press.Brame, B., Nagin, D. S., & Tremblay, R. E. (2001). Developmental trajectories of physi-
cal aggression from school entry to late adolescence. Journal of Child Psychologyand Psychiatry and Allied Disciplines, 42, 503–512.
Broidy, L. M., Nagin, D. S., Tremblay, R. E., Bates, J. E., Brame, B., Dodge, K. A.,Fergusson, D., Horwood, J. L., Loeber, R., Laird, R., Lynam, D. R., Moffitt, T. E.,Pettit, G. S., &Vitaro, F. (2003). Developmental trajectories of childhood disruptivebehaviors and adolescent delinquency: A six-site, cross-national study. Develop-mental Psychology, 39, 222–245.
Bryk, A. S., & Raudenbush, S. W. (1987). Application of hierarchical linear models to as-sessing change. Psychological Bulletin, 101, 147–158.
Caspi, A., Lynam, D. R., Moffitt, T. E., & Silva, P. A. (1994). Are some people crimeprone? Replications on the personality–crime relationship across counties, genders,races, and methods. Criminology, 32, 163–195.
Coie, J. D., & Dodge, K. A. (1988). Multiple sources of data on social behavior and socialstatus in the school: A cross-age comparison. Child Development, 59, 815–829.
Cook, P. J., & Laub, J. H. (1998). The unprecedented epidemic in youth violence. In M.Tonry & M. H. Moore (Eds.), Youth violence, crime and justice: A review of re-search (Vol. 24, pp. 27–64). Chicago: University of Chicago Press.
Cook, P. J., & Laub, J. H. (2002). After the epidemic: Recent trends in youth violence inthe United States. In M. Tonry (Ed.), Crime and justice: A review of research (pp.117–153). Chicago: University of Chicago Press.
Cornell, D. G. (1989). Causes of juvenile homicide: A review of the literature. In E. P.Benedek & D. G. Cornell (Eds.), Juvenile homicide (pp. 3–36). Washington, DC:American Psychiatric Press.
D’Unger, A. V., Land, K. C., McCall, P. L., & Nagin, D. S. (1998). How many latentclasses of delinquent/criminal careers?: Results from mixed Poisson regression anal-yses of the London, Philadelphia, and Racine cohorts studies. American Journal ofSociology, 103, 1593–1630.
Eggelston, E. P., Laub, J. H., & Sampson, R. J. (2004). Methodological sensitivities to la-tent class analysis of long-term criminal trajectories. Journal of Quantitative Crimi-nology, 20, 1–26.
Elliott, D. S., Huizinga, D., & Ageton, S. S. (1985). Explaining delinquency and drug use.Beverly Hills, CA: Sage.
Ewing, C. P. (1990). When children kill. Lexington, MA: Lexington Books.Federal Bureau of Investigation. (2001). Crime in the United States (1984–2000). Wash-
ington, DC: U.S. Government Printing Office.
216 T H E D E V E L O P M E N T O F A G G R E S S I O N

Fergusson, D. M., & Horwood, L. J. (1995). Early disruptive behavior, IQ, and laterschool achievement and delinquent behavior. Journal of Abnormal Child Psychol-ogy, 23, 183–199.
Frick, P. J., Lahey, B. B., Loeber, R., Tannenbaum, L., Van Horn, Y., Christ, M. A. G., etal. (1993). Oppositional defiant disorder and conduct disorder: A meta-analytic re-view of factor analyses and cross-validation in a clinic sample. Clinical PsychologyReview, 13, 319–340.
Gottfredson, M. R., & Hirschi, T. (1990). A general theory of crime. Stanford, CA: Stan-ford University Press.
Heckman, J. (1981). Statistical models for discrete panel data. In C. F. Manski & D.McFadden (Eds.), Structural analysis of discrete data with econometric applica-tions. Cambridge, MA: MIT Press.
Heide, K. M. (2003). Youth homicide: A review of the literature and a blueprint for ac-tion. International Journal of Offender Therapy and Comparative Criminology, 47,6–36.
Horney, J. D., Osgood, W., & Marshall, I. H. (1995). Criminal careers in the short-term:Intra-individual variability in crime and its relation to local life circumstances.American Sociological Review, 60, 655–673.
Jessor, R., Donovan, J. E., & Costa, F. M. (1991). Beyond adolescence: Problem behaviorand young adult development. Cambridge, UK: Cambridge University Press.
Jones, B. L., Nagin, D. S., & Roeder, K. (2001). A SAS procedure based on mixture mod-els for estimating developmental trajectories. Sociological Methods and Research,29, 374–393.
Kelley, B. T., Loeber, R., Keenan, K., & DeLamatre, M. (1997). Developmental pathwaysin disruptive and delinquent behavior. OJJDP Juvenile Justice Bulletin. www. ncjrs.org
Lacourse, E., Côté, S., Nagin, D. S., Vitaro, F., Brendgen, M., & Tremblay, R. E. (2002).A longitudinal-experimental approach to testing theories of antisocial behavior de-velopment. Development and Psychopathology, 14, 909–924
Lacourse, E., Nagin, D., Tremblay, R. E., Vitaro, F., & Claes, M. (2003). Developmentaltrajectories of boys’ delinquent group membership and facilitation of violent behav-iors during adolescence. Development and Psychopathology, 15, 183–197.
Laub, J. H., Nagin, D. S., & Sampson, R. J. (1998). Trajectories of change in criminal of-fending: Good marriages and the desistance process. American Sociological Review,63, 225–238.
Laub, J. H., & Sampson, R. J. (2003). Shared beginnings, divergent lives: Delinquentboys to age 70. Cambridge, MA: Harvard University Press.
Le Blanc, M., & Fréchette, M. (1989). Male criminal activity from childhood throughyouth: Multilevel and developmental perspectives. New York: Springer.
Lemert, E. M. (1951). Social pathology; A systematic approach to the theory ofsociopathic behavior. New York: McGraw-Hill.
Loeber, R. (1988). Natural histories of conduct problems, delinquency, and associatedsubstance use: Evidence for developmental progressions. In B. B. Lahey & A. E.Kazdin (Eds.), Advances in clinical child psychology (Vol. 11, pp. 73–124). NewYork: Plenum Press.
Loeber, R., DeLamatre, M., Keenan, K., & Zhang, Q. (1998a). A prospective replicationof developmental pathways in disruptive and delinquent behavior. In R. Cairns, L.Bergman, & J. Kagan (Eds.), Methods and models for studying the individual (pp.185–215). Thousand Oaks, CA: Sage.
Homicide, Violence, and Developmental Trajectories 217

Loeber, R., & Farrington, D. P. (Eds.). (1998). Serious and violent juvenile offenders:Risk factors and successful interventions. Thousand Oaks, CA: Sage.
Loeber, R., & Farrington, D. P. (Eds.). (2001). Child delinquents: Development, interven-tion and service needs. Thousand Oaks, CA: Sage.
Loeber, R., Farrington, D. P., Stouthamer-Loeber, M., & Van Kammen, W. B. (1998b).Antisocial behavior and mental health problems: Explanatory factors in childhoodand adolescence. Mahwah, NJ: Erlbaum.
Loeber, R., Homish, D. L., Wei, E. H., Crawford, A. M., Farrington, D. P., Stouthamer-Loeber, M., Creemers, J., Koehler, S., & Rosenfeld, R. (in press). The prediction of vi-olence and homicide in young males. Journal of Consulting and Clinical Psychology.
Loeber, R., Keenan, K., & Zhang, Q. (1997). Boys’ experimentation and persistence indevelopmental pathways toward serious delinquency. Journal of Child and FamilyStudies, 6, 321–357.
Loeber, R., & Le Blanc, M. (1990). Toward a developmental criminology. In M. Tonry &N. Morris (Eds.), Crime and Justice (Vol. 12, pp. 375–473). Chicago: University ofChicago Press.
Loeber, R., & Schmaling, K. B. (1985). Empirical evidence for overt and covert patternsof antisocial conduct problems: A metaanalysis. Journal of Abnormal Child Psy-chology, 13, 337–353.
Loeber, R., Wei, E., Stouthamer-Loeber, M., Huizanga, D., & Thornberry, T. P. (1999).Behavioral antecedents to serious and violent offending: Joint analyses from theDenver Youth Survey, Pittsburgh Youth Study and the Rochester Youth Develop-ment Study. Studies on Crime and Crime Prevention, 8, 245–263.
Loeber, R., Wung, P., Keenan, K., Giroux, B., Stouthamer-Loeber, M., Van Kammen, W.B., & Maughan, B. (1993). Developmental pathways in disruptive child behavior.Development and Psychopathology, 5, 101–132.
Matsueda, R. L., & Heimer, K. (1997). A symbolic interactionist theory of role-transi-tions, role-commitments, and delinquency. In T. P. Thornberry (Ed.), Developmen-tal theories of crime and delinquency. Advances in criminological theory (Vol. 7, pp.163–213). Lincoln: University of Nebraska Press.
McCord, J. (1990). Problem behaviors. In S. S. Feldman & G. R. Elliott (Eds.), At thethreshold: The developing adolescent (pp. 441–430). Cambridge, MA: HarvardUniversity Press.
Meredith, W., & Tisak, J. (1990). Latent curve analysis. Psychometrika, 55, 107–122.Moffitt, T. E. (1993). Adolescence-limited and life-cycle-persistent antisocial behavior: A
developmental taxonomy. Psychology Review, 100, 674–701.Muthen, B. (1989). Latent variable modeling in heterogeneous populations. Psycho-
metrika, 54, 557–585.Muthen, B. (2001). Second-generation structural equation modeling with a combination
of categorical and continuous latent variables: New opportunities for latent class-latent growth modeling. In L. M. Collins & A. G. Sayer (Eds.), New methods for theanalysis of change: Decade of behavior (pp. 291–322). Washington, DC: AmericanPsychological Association.
Nagin, D. S. (1999). Analyzing developmental trajectories: A semiparametric, group-based approach. Psychological Methods, 4, 139–157.
Nagin, D. S., & Land, K. C. (1993). Age, criminal careers and population heterogeneity:Specification and estimation of a nonparametric, mixed Poisson model. Criminol-ogy, 31, 327–362.
218 T H E D E V E L O P M E N T O F A G G R E S S I O N

Nagin, D. S., Pagani, L., Tremblay, R. E., & Vitaro, F. (2003). Life course turning points:The effect of grade retention on physical aggression. Development and Psycho-pathology, 15, 343–361.
Nagin, D., & Paternoster, R. (1991). On the relationship of past and future participationin delinquency. Criminology, 29, 163–190.
Nagin, D., & Paternoster, R. (2000). Population heterogeneity and state dependence:State of the evidence and directions for future research. Journal of QuantitativeCriminology, 16, 117–144.
Nagin, D. S., & Tremblay, R. E. (1999). Trajectories of boys’ physical aggression, opposi-tion, and hyperactivity on the path to physically violent and non-violent delin-quency. Child Development, 70, 1181–1196.
Nagin, D. S., & Tremblay, R. E. (2001). Analyzing developmental trajectories of distinctbut related behaviors: A group-based method. Psychological Methods, 6, 18–34.
Olweus, D. (1979). Stability of aggressive reaction patterns in males: A review. Psycho-logical Bulletin, 86, 852–875.
Osgood, D. W., Johnston, L. D., O’Malley, P. M., & Bachman, J. G. (1988). The general-ity of deviance in late adolescence and early adulthood. American Sociological Re-view, 53, 81–93.
Osgood, D. W., & Rowe, D. C. (1994). Bridging criminal careers, theory, and policythrough latent variable models of individual offending. Criminology, 32, 517–554.
Patterson, G. R., & Yoerger, K. (1993). Developmental models for delinquent behavior.In S. Hodgins (Ed.), Mental disorder and crime (pp. 140–172). Newbury Park, CA:Sage.
Patterson, G. R., & Yoerger, K. (1997) A developmental model for late-onset delin-quency. In D. W. Osgood (Ed.), Motivation and delinquency (Vol. 44, pp. 119–177).Lincoln: Nebraska Symposium on Motivation.
Raine, A., (2002). Biosocial studies of antisocial and violent behavior in children andadults: A review. Journal of Abnormal Child Psychology, 30, 311–326.
Robins, L. (1966). Deviant children grown up. Baltimore: Williams & Wilkins.Rogosa, D. R., & Willett, J. B. (1985). Understanding correlates of change by modeling
individual differences in growth. Psychometrika, 50, 203–228.Sampson, R. J., & Laub, J. H. (1993). Crime in the making: Pathways and turning points
through life. Cambridge, MA: Harvard University Press.Snyder, H. N., & Sickmund, M. (1999). Juvenile offenders and victims: 1999 national re-
port. Washington, DC: U.S. Department of Justice, Office of Juvenile Justice andDelinquency Prevention.
Thornberry, T. P. (1987). Toward an interactional theory of delinquency. Criminology,25, 863–891.
Thornberry, T. P., Freeman-Gallant, A., Lizotte, A. J., Krohn, M. D., & Smith, C. A.(2003). Linked lives: The intergenerational transmission of antisocial behavior.Journal of Abnormal Child Psychology, 31, 171–184.
Tolan, P. H., Gorman-Smith, D., & Loeber, R. (2000) Developmental timing of onsets ofdisruptive behaviors and later delinquency of inner-city youth. Journal of Child andFamily Studies, 9, 203–220.
Tremblay, R. E., Nagin, D. S., Séguin, J. R., Zoccolillo, M., Zelazo, P., Boivin, M.,Pérusse, D., & Japel, C. (2004). Physical aggression during early childhood: Trajec-tories and predictors. Pediatrics, 114, 43–50.
Homicide, Violence, and Developmental Trajectories 219

This page intentionally left blank

Part III
DETERMINANTSOF AGGRESSION

This page intentionally left blank

DETERMINANTS OF AGGRESSIONGenetics and the Development of Aggression
11
Genetics and the Developmentof Aggression
DANIEL PÉRUSSE and PAUL L. GENDREAU
The overall goal of this chapter is to assess the evidence for genetic influenceson the developmental origins of aggression. Unlike simple qualitative Mende-lian traits, which are generally mediated by a single gene and are relatively in-sensitive to environmental influences, quantitative traits, such as aggressive-ness, vary continuously across a population and derive from a large number ofadditive and/or interactive genetic and environmental influences. As this is avast and complex issue, we focus on the major findings and their limitations.We first describe the most significant approaches to investigate the genetic ba-sis of aggression in animals. Then we review evidence from quantitative genet-ics concerning the genetic–environmental etiology of aggression in humans.Finally, we examine the relatively recent and still scarce evidence from molec-ular genetic studies of aggressive behavior.
Both animal and human models of aggression have contributed consider-ably to progress in understanding the complex interaction of genetic and envi-ronmental factors in the emergence of aggressiveness. Different approachescan be used to investigate the genetic basis of complex traits. Some are termedgenotype driven, as the initial step is to alter the genome in animal models byknocking out or “knocking in” specific genes. Once the desired developmentalstage has been reached, the phenotypic outcome is observed. With this “re-verse genetics” (Picciotto & Wickman, 1998), an infinite number of geneti-cally altered animal strains (primarily mouse strains) characterized by distinctneurobehavioral phenotypes can be produced. This approach is particularlyuseful to dissect the function of specific genes and proteins on brain and be-havior, as well as the compensatory mechanisms by which neurobehavioraladjustment occurs (for reviews, see Phillips et al., 2002; Picciotto & Wick-
223

man, 1998). Despite their considerable scientific interest, however, geneknockouts have little value in explaining naturally occurring variations in be-havioral phenotypes such as aggression. In addition, gene-targeted approachesmay induce gross morphological and neurophysiological anomalies that maymask more subtle behavioral effects (Lathe, 1996; Nelson & Young, 1998).
Some approaches are more applicable to normal—and human—development,as the first step is to characterize the neurobehavioral phenotype; then an at-tempt is made to trace back the genes associated with the expression of thisphenotype. These approaches are termed phenotype driven. In animals, crossing/backcrossing of two strains—each with its own genetic makeup but with littleor no intrastrain genetic variability (inbred)—and selective breeding of indi-viduals with high genetic variability (outbred) have been the traditionalphenotype-driven methods. These methods, however, tell us only that a phe-notype of interest is heritable and that genes contribute in a dominant or re-cessive fashion. To circumvent these limitations and to identify genes or DNAregions potentially associated with a phenotype, quantitative trait locus (QTL)mapping can be employed. This method is extremely valuable to examinecomplex gene–environment interactions and the development of specificneurobehavioral phenotypes in both animals and humans. However, despitethe fact that a particular locus (chromosomal region that encloses a definitegene) can be associated to a phenotype, QTLs typically yield only small ge-netic effects and do not provide a fine resolution for detecting relevant geneunits (Darvasi, 1998). Therefore, once a region of interest has been deter-mined, the task to identify the putative influential DNA sequences among thelarger region still remains.
GENETIC–DEVELOPMENTAL MODELS OF AGGRESSION IN ANIMALS
Mus musculus (mouse) has been the species of choice for genetic studies of ag-gressive behavior. Mouse models provide many advantages over human stud-ies for examining the relative influences of genes and environment on the de-velopment of complex behavioral traits, such as aggressiveness. One suchadvantage is that the genotypes of several strains have been well characterized,therefore facilitating the analysis of gene–environment interactions. In addi-tion, the mouse genome has about 30,000 protein-coding genes, a numberbroadly similar to that in the human genome. Approximately 80% of the twogenomes are organized similarly (orthologue), and only 1% of the human ge-nome does not have a homologue mouse gene (Mouse Genome SequencingConsortium, 2002).
Inbred Mouse Strains
The use of inbred mouse strains has been a popular methodological choice toexamine the genetic basis of biobehavioral phenotypes. This approach stems
224 D E T E R M I N A N T S O F A G G R E S S I O N

from a long history in mammalian genetics, dating from the beginning of thecentury. Scott (1942) and Ginsburg and Allee (1942) were among the first toexamine differences in aggressiveness among inbred mouse strains. Inbredmouse strains are produced using brother–sister mating over several genera-tions (20 generations are sufficient to produce very low levels of heterozygos-ity at all loci; see Taylor et al., 1999). Inbred strains such as Balb/c and ABGhave been shown to be extremely aggressive under a variety of testing condi-tions, whereas other strains such as A/J, CBA/ca, and AB//Halle rarely exhibitaggressive behavior, even under experimental conditions, known to inducehigh levels of aggression (e.g., prolonged social isolation, resident–intruderparadigm; see Gendreau, Petitto, Petrova, Gariépy, & Lewis, 2000; Kessler,Elliott, Orenberg, & Barchas, 1977; Southwick & Clark, 1968).
Two strains that differ both genetically and behaviorally (but with lowwithin strain variability) provide an excellent starting point for the geneticanalysis of complex traits, particularly those that have low heritability (Taylor,1989). The crossing of two inbred strains results in genetically identical pups,the so-called F1 animals, which receive one allele at every locus from each par-ent. These animals are not of much interest in respect to genetic analysis, buttheir offspring, the F2 generation individuals, are, because each animal of thissecond generation has a unique combination of alleles. With this genetic diver-sity, it becomes possible to correlate genetic markers with the differential ex-pression of aggressiveness. Typically, the genetic analysis is limited to the ani-mals that are at the two extremes of the continuum—that is, animals thatshow no aggression and those that display the highest levels. The effects of re-ciprocal backcrosses of the F2 offspring with one or both of the parentalstrains can also be informative. Kessler et al. (1977) performed crossing andbackcrossing of the aggressive Balb/c strain with the nonaggressive A/J strain.Their results suggested the presence of a single autosomal recessive trait forthe transmission of high aggressiveness. Others have argued that aggressive-ness may be transmitted through the Y chromosome (Maxson, 1996). But asCarlier, Roubertoux, Kottler, and Degrelle (1990) claimed, “the Y correlatesprobably are nothing more than possible genetic variants being expressed atdifferent levels of physiological functions and acting on aggression” (p. 151).
Selective Breeding of Aggressive Behavior
Selective breeding, also known as artificial selection or psychogenetic selection(Broadhurst, 1958), is clearly the most ancient method used to produce variet-ies or subspecies of animals that exhibit a particular phenotypic characteristic.Selective breeding is a relatively simple method that allows the gradual segre-gation of genes contributing to the phenotype of interest. Numerous behav-ioral characteristics, from locomotor activity to emotionality and aggression,have been investigated through selective breeding (Fuller & Thompson,1978).
Since the 1960s there have been three major selective breeding programs
Genetics and the Development of Aggression 225

of aggressive behavior in male mice. All three programs were based on the ani-mal’s propensity to exhibit either high or low levels of aggression against amale opponent. The earliest program was initiated by Kirsti Lagerspetz in Fin-land (Lagerspetz, 1961, 1964; Lagerspetz, Tirri, & Lagerspetz, 1968; see alsoNyberg, Vekovischeva, & Sandnabba, 2003). Two lines of mice, the TurkuAggressive (TA) and the Turku Nonaggressive (TNA) were produced in just afew generations. Aggression was measured following a period of social isola-tion. Then Van Oortmerssen set in motion another breeding program in theNetherlands, producing two lines of mice that differed in their latency to at-tack another male conspecific, but without using social isolation prior to test-ing (Van Oortmerssen & Bakker, 1981; see also Sluyter, Korte, Bohus, & VanOortmerssen, 1996).
At about the same time in North America, Cairns initiated his own pro-gram of selective breeding for isolation-induced aggression in Institute forCancer Research (ICR) mice (Cairns, MacCombie, & Hood 1983). For induc-ing intermale aggression, mice were singly caged from 21 days of age to ap-proximately 45–50 days, as this specific period of ontogeny and this particularlength of isolation (sensitive period) were shown to produce the highest levelsof aggression (Cairns, Hood, & Midlam, 1985). Hence, by altering the socialenvironment during a restricted window of time, two lines of mice that dif-fered markedly in behavioral reactivity were produced: one line of animalsthat promptly attacked and a second line that exhibited high levels of “freez-ing” (for a review see Cairns, Gariépy, & Hood, 1990). Selection effects ap-peared promptly, with statistically significant line differences in isolation-in-duced aggression obtained as early as the fourth (S4) generation of selection.In this breeding program, no brother–sister mating was allowed. Perhaps as aresult, aggressive behavior continued to appear, albeit extremely rarely, in thelow-aggressive line even after the 30th generation (Gariépy, Rodriguiz, &Jones, 2002; Gariépy, Gendreau, Mailman, Tancer, & Lewis, 1995).
In addition, although the breeding program had targeted intermale ag-gression, females of the highly aggressive line were also found to be moreaggressive than females of the low-aggressive line when assessed underpostpartum conditions (Hood & Cairns, 1988). One breeding program specif-ically targeted interfemale aggression (Ebert & Hyde, 1976). As in the otherbreeding programs, the effects of selection appeared quickly (S2). These effectswere not paralleled by aggression in males, however (Hyde & Ebert, 1976).Sandnabba, Lagerspetz, and Jensen (1994) showed that females of the TurkuAggressive line also exhibited more aggression after repeated exposure to tes-tosterone than those of the Turku Nonaggressive line. During female–femaledyadic interactions, however, no such difference was observed (Van Oort-merssen & Baker, 1981). In their selective breeding program based on attacklatency in males, Van Oortmerssen and Baker (1981) estimated a heritabilityof 30% for intermale aggression. In the selective breeding program performedby Lagerspetz and colleagues, heritability was estimated at 36% (McClearn &
226 D E T E R M I N A N T S O F A G G R E S S I O N

DeFries, 1973, in Van Oortmerssen and Baker, 1981). Ebert and Hyde (1976)estimated the heritability of female aggression to be between 43 and 55%.
A significant contribution of these selective breeding programs was topoint out the importance of developmental timing for the expression of ag-gressive behavior and the specificity of the context for eliciting specific formsof aggression. In Cairns’s program, for instance, line differences in aggressionwere dependent on the developmental stage in which comparisons were made;line differences in aggression were not observed earlier in ontogeny and werenot as pronounced later in development (Cairns et al., 1985, 1990), whichsuggests that the influence of genes on aggression varies throughout the life-span.
What do animal models tell us about the genetic basis of human aggres-sion? QTL mapping in conjunction with selective breeding would provide aninteresting approach to examining the genetic basis of aggression. For in-stance, once an association with a genetic locus and a specific form of aggres-sion has been reported, selective breeding on this phenotype can be conducted.If QTL analysis shows that the frequency of this fragment of DNA changes asselection is progressing, support can be acquired for the establishment of agenotype–phenotype link (Phillips et al., 2002). However, QTL analyses ofaggressive behavior in mammals or even in insects—for instance, defensivestinging behavior in honeybees (Hunt, Guzman-Novoa, Fondrk, & Page,1998)—do not contribute much to understanding the neurobiological and be-havioral processes by which aggression develops in humans. In addition,despite obvious genetic and neurobiological similarities, the phylogenetic gapbetween rodents and humans is important, therefore making phenotypic andetiologic comparisons rather precarious.
Nevertheless, developmental models of mouse aggression such as thoseusing selective breeding provided a significant contribution by pointing outthe importance of developmental timing for genetic influences on aggressivebehavior. These selective breeding programs have also stressed the key inputof environmental–experimental factors in the expression of aggression. For in-stance, rearing animals in groups rather than isolating them prevented the ex-pression of aggressive behavior (Gariépy et al., 1995; Gendreau, Gariépy,Petitto, & Lewis, 1997). Comparable gene–environment interaction for theexpression of aggression in monkeys was found recently by Suomi and col-leagues (Suomi, Chapter 4, this volume). These authors showed that the ge-netic predisposition to exhibit aggressiveness, which was linked to a polymor-phism of the gene coding for the serotonin transporter, was conditional tobeing maternally deprived. Genetically predisposed animals that were rearedwith their mothers early in life did not show aggressiveness, as compared withthose that were reared only with same-age conspecifics. These findings consti-tute the beginning of a promising era of research for determining the complexdevelopmental interaction of specific environmental–experiential factors withspecific genes and gene products.
Genetics and the Development of Aggression 227

GENETIC STUDIES OF HUMAN AGGRESSION
The genetic–developmental theory behind a complex trait like aggression isbased on the multifactorial polygenic threshold model of heredity. This modelholds that individual differences in a continuous phenotype such as aggressionresult from the action of a large number of genes, each exerting a small effect,acting in conjunction with nongenetic (environmental) factors. Above a cer-tain threshold of genetic liability and environmental risk, an extreme, oftenpathological manifestation of the trait (e.g., violent crime) develops.
In humans, the study of the genetic and environmental causes of pheno-typic variation depends on genetically informative designs, that is, twin andadoption studies. The magnitude of genetic influence is inferred from compar-ing the phenotypic resemblance between groups of individuals that vary intheir degree of genetic relatedness (e.g., monozygotic [MZ] twins sharing alltheir genes vs. dizygotic [DZ] twins sharing on average half of their genes), ev-erything else being assumed equal. Similarly, the magnitude of environmentalinfluence is estimated by comparing groups of individuals differing, or not, intheir exposure to common environmental factors (e.g., biological siblings vs.adoptive siblings). One should not be concerned merely with quantifying thegenetic–environmental contribution to aggression, however, but also withidentifying specific genes and specific environmental factors that make upthese contributions. On the genetic side, this has recently been explored usingmolecular genetic techniques. On the environmental side, twin and adoptionstudies offer the opportunity of “controlling” for genetic factors in order toidentify unambiguously environmental influences on aggression.
Quantitative Genetics of Human Aggression
More than 100 twin and adoption studies have been conducted in the pastthree decades to examine the genetic basis of aggressive behavior and relatedconstructs (e.g., externalizing behavior problems, conduct disorder, antisocialpersonality disorder (ASPD), delinquency, criminality). Not surprisingly, sev-eral reviews have concluded that both genetic and environmental factors wereimportant in the etiology of these heterogeneous operationalizations of ag-gression assessed throughout the lifespan (e.g., Gottesman & Goldsmith,1994; Carey, 1994).
Four meta-analyses have gone further by providing quantitative estimatesof the genetic and environmental contributions to individual differences inaggression-related constructs across studies. Walters (1992) reviewed 11 fam-ily, 14 twin, and 13 adoption studies of criminality and concluded that geneticinfluences on criminality were low to moderate. Mason and Frick (1994) re-viewed 12 twin and 3 adoption studies of antisocial behavior and concludedthat approximately half of the variance in various measures of antisocialbehavior could be attributed to genetic influences. Miles and Carey (1997) re-viewed 20 twin and 4 adoption studies of behavioral aggression and also con-
228 D E T E R M I N A N T S O F A G G R E S S I O N

cluded that genetic influences accounted for approximately 50% of the vari-ance.
A more recent and more exhaustive meta-analysis of various constructsof antisocial behavior was conducted by Rhee and Waldman (2002). A totalof 51 twin and adoption studies comprising 87,152 individuals were re-viewed, bearing on antisocial behavior operationalized in four ways: (1) psy-chiatric diagnoses such as conduct disorder and ASPD, (2) delinquency andcriminality, (3) antisocial behavior combining delinquency and aggression,and (4) behavioral aggression per se. Across these four operationalizations,broad heritability, that is, the sum of additive (termed “A” in the genetic liter-ature) and nonadditive (“D”) genetic effects reached 41%. Common environ-mental influence (“C,” i.e., environmental effects that are shared betweentwins and siblings) was 16%, and unique environmental influence (“E,” i.e.,environmental effects that are not shared between twins and siblings) reached43%. According to this latest meta-analysis, genetic variation thus appears tocause approximately 40% of individual differences across various constructsof aggressiveness, whereas about 60% of the variance is explained by environ-mental factors.
Interestingly, Rhee and Waldman (2002) examined whether these resultsvaried according to the different assessment methods used to measure the con-structs of aggression (i.e., official records, behavioral reaction to stimuli,objective assessment, self-report, and report by others). In the case of officialrecords, assessment method and operationalization were completely con-founded, as the assessment by records was used only in studies of criminality,and the potential moderating effect of this method could therefore not be ex-amined as such. In the case of criminality, however, a distinction needs to bemade between violent crimes involving physical aggression toward others andnonviolent crimes such as theft. This suboperationalization was not per-formed by Rhee and Waldman (2002), but Mednick, Gabrielli, and Hutchings(1984), as well as Bohman, Cloninger, Sigvardsson, and von Knorring (1982),found evidence that genetic influences were significant for property crimes,but not for violent crimes.
Most studies reviewed by Rhee and Waldman (2002) used self-reportand/or report by others for assessing aggressiveness. Interestingly, significantdifferences in the magnitude of genetic and environmental effects were foundbetween these two methods, with heritability reaching 39% and 53%, respec-tively. Behavioral reactivity to stimuli and objective assessment were used inonly three studies. The moderating effects of these methods could thereforenot be estimated. The single study using direct observation ratings of labora-tory behavior reviewed by Rhee and Waldman (2002) was that of Plomin,Foch, and Rowe (1981), who examined aggression in young twins using anobjective test validated against peer ratings and teacher ratings. No heritabil-ity was found for this measure. Only three published studies have actuallymeasured children’s and adolescents’ aggressive behaviors in the laboratory(Owen & Sines, 1970; Plomin et al., 1981; O’Connor, Deater-Deckard,
Genetics and the Development of Aggression 229

Fulker, Rutter, & Plomin, 1998). None of these studies found evidence for sig-nificant genetic influence on aggression.
What to make of these conflicting results? Depending on how aggressionis measured, genetic influences appear to range from 0% (objective assess-ment) to more than 50% (report by others). We grouped twin studies of overtaggression reviewed in Rhee and Waldman (2002), DiLalla (2002), Miles andCarey (1997) as well as two recently published studies (Dionne, Tremblay,Boivin, Laplante, & Pérusse, 2003; Arseneault et al., 2003) in three categoriesaccording to the degree of twin-wise dependency in information resultingfrom the assessment method used: (1) complete dependency (i.e., report byother: The same informant (parent or teacher) provides information for bothmembers of a twin pair), (2) partial dependency (i.e., self-report: Each twin re-ports on him- or herself, but knows the other twin, and (3) no dependency(i.e., different raters for each twin, official records). As shown in Figure 11.1,the heritability is found to decrease linearly with information dependency,from 45% for complete dependency to 29% for partial dependency to 5% forno dependency.
Rhee and Waldman (2002) also found that twin studies yielded higherheritability estimates than adoption studies across the four operationalizationsof aggressiveness. This may be due to overestimates of heritability resultingfrom methodological shortcomings specific to the twin design, such as theinformant-dependency bias suggested earlier and possible violations of theequal environment assumption of twin studies. To our knowledge, the require-ment that MZ twins are not exposed to more similar environmental influences
230 D E T E R M I N A N T S O F A G G R E S S I O N
FIGURE 11.1. Heritability estimates of aggression in relation to twin-wise dependency in thesource of information for the assessment of aggression.

than DZ twins has never been tested in the case of aggression, although it hasbeen found to be supported in a number of studies of other phenotypes.
Adoption studies also have a number of methodological problems thatlimit their interpretation. One problem is that it has been shown in severalcountries that children given away for adoption have a higher rate of antiso-cial behavior as compared with the general population (e.g., New Zealand(Fergusson, Lynskey, & Horwood, 1995), the Netherlands (Verhulst, Versluis-den-Bieman, Van der Ende, Berden, & Sanders-Woudstra, 1990), the UnitedStates (Sharma, McGue, & Benson, 1998)). It is likely that the parents whogive their children away for adoption also exhibit higher levels of antisocialbehavior than parents in the general population, and certainly as comparedwith adoptive parents. The resulting correlation between adoptees and theirbiological parents in antisocial behavior can then hypothetically be generatedby two different processes: (1) the transmission of antisocial genes from bio-logical parents to their adopted-away children or (2) a purely environmentalcausal process, progressing from parental antisociality to giving away childrenfor adoption to adoptive child antisociality due to abandonment. These twocompeting hypotheses appear confounded by the very design that is supposedto enable their disentangling. A more adequate adoptive design, correspond-ing to the animal cross-fostering method, would involve comparing adopted-away children (AAC) with children who would have been given away by theAAC’s adoptive parents, and with children who would have been adopted bythe AAC’s biological parents—a “natural experiment” that, perhaps fortu-nately, does not occur in reality.
Stoolmiller (1999) has shown that genetic influences may be overesti-mated and shared environmental influences underestimated in adoption stud-ies if the sample’s range of environments is restricted—for instance, in terms ofsocioeconomic status, which is generally the case in adoptive families. Clerget-Darpoux, Goldin, and Gershon (1986) demonstrated how a genetic effect issimulated in adoption studies when a positive correlation exists between theadoptive and biological parents for an etiologic environmental variable, whichis likely the case for selective placement (i.e., where environmental features ofthe biological and adoptive parents’ households tend to be matched by place-ment agencies). In addition, prenatal and early postnatal environments are ev-idently shared to a greater extent by adopted-away children and their biologi-cal siblings than by adopted-away children and their adoptive siblings. Thiswould be an important control, as there is mounting evidence from animaland human studies that both prenatal and perinatal risk factors are associatedwith the development of later aggressive and antisocial behavior (e.g., Raine,2002). Such unmeasured correlation in environmental risk between adopteesand their biological siblings, if any, will artificially inflate heritability esti-mates.
Finally, both twin and adoption designs use analytical models that as-sume the independence, and therefore additivity, of genetic and environmentalinfluences. If the two types of influences are not perfectly independent, the ap-
Genetics and the Development of Aggression 231

plication of an additive model may result in systematic overestimation ofheritability and underestimation of environmentality. In the case of aggressionand antisocial behavior, evidence precisely points to the dependence betweengenetic and environmental influences (e.g., Cadoret, Cain, & Crowe, 1983;Ge et al., 1996; O’Connor et al., 1998; Caspi et al., 2004; Jaffee, Caspi,Moffitt, & Taylor, 2004). As discussed in the next section, genotype-by-envi-ronment interaction for aggressive and antisocial behavior has now recentlybeen traced at the molecular level (Caspi et al., 2002).
Molecular Genetics of Aggression
In recent years a number of molecular genetic studies have targeted differentneurobehavioral correlates of aggression. Researchers have generally used acandidate gene approach, that is, attempting to locate genes that influenceneurophysiological and hormonal systems (i.e., endophenotypes) thought tobe involved in aggressiveness. Most studies have examined various polymorphismsof serotonin receptors, dopamine receptors, and the neurotransmitter-metabolizing enzyme monoamine oxidase A (MAOA). The serotonergic ab-normalities related to aggressiveness in humans have motivated linkage andassociation studies of serotonin-related genes and impulsive aggression. Knock-out of the HTR1B (formerly 5-HT1B) serotonin receptor gene in mice resultsin increased aggression, as well as alcohol and cocaine consumption. Relation-ships between various measures of aggressiveness and HTR1B, which regu-lates the synthesis and release of serotonin from presynaptic neuron terminals,have thus been tested in a number of samples.
In a study of Finnish alcoholic criminal offenders and a large multi-generational family of southwestern American Indians with a high rate of al-coholism, Lappalainen et al. (1998) investigated whether the HTR1B gene waslinked with the psychiatric diagnoses of antisocial personality disorder and in-termittent explosive disorder comorbid with alcoholism. Significant linkageswere found for one of the two polymorphisms studied in the Finnish sample,and for both polymorphisms in the American Indian tribe. However, the func-tional significance of the polymorphism found to be linked in both popula-tions has not yet been established.
Given the comorbidity of aggression, suicide, alcoholism, and substanceabuse, Huang, Grailhe, Arango, Hen, and Mann (1999) studied the relation-ship of these psychopathological problems to the human HTR1B receptorgene. Contrary to the findings of Lappalainen et al. (1998), no relationshipwas found between two polymorphisms of the HTR1B gene and suicide, ma-jor depression, alcoholism, or pathological aggression. Similarly, in a sampleof 89 Caucasian patients with personality disorder, no association between theHTR1B genotype and self-reported measures of impulsive aggression wasfound (New, Goodman, Mitropoulou, & Siever, 2002).
Transcription of the serotonin transporter gene (SLC6A4) is modulatedby a common polymorphism in its promoter region, with demonstrated func-
232 D E T E R M I N A N T S O F A G G R E S S I O N

tional significance in coding for high and low transporter production. In amuch-publicized study, association analyses in two independent samples re-vealed that the S allele was associated with increased anxiety-related personal-ity scores, including impulsivity and harm avoidance (Lesch et al., 1996). Asubsequent study using a different but related personality questionnaire failedto replicate this finding in a normal cohort (Ebstein et al. 1997). In anotherreplication attempt, Gelernter, Kranzler, Coccaro, Siever, and New (1998) in-vestigated the same SLC6A4 promoter polymorphism and an additional poly-morphism in intron 2, among a sample of American subjects of European andAfrican ancestry with diagnoses of a personality disorder or substance de-pendence and normal subjects. Again, they were unable to replicate the associ-ation finding.
Tryptophan hydoxylase is the rate-limiting enzyme in the synthesis of se-rotonin from tryptophan and is coded for by the TPH gene (see Pihl &Benkelfat, Chapter 13, this volume). As low turnover rates of serotonin havebeen associated with impaired impulse control, polymorphisms of the TPHgene were investigated in a sample of Finnish impulsive and nonimpulsive, al-coholic, violent offenders and normal control subjects. The TPH genotypewas associated with suicidality, which may involve impaired impulse control,but not with impulsive behavior itself (Nielsen et al., 1994). In a subsequentstudy, association and linkage analyses were conducted on a polymorphism inintron 7 of the TPH gene with suicidality, alcoholism, and a personality ques-tionnaire among a larger sample of Finnish alcoholic offenders, controls, andtheir relatives (Nielsen et al., 1998). This polymorphism showed significantevidence of linkage with suicidality and severe suicide attempts.
An association between various polymorphisms of the TPH gene and sui-cide by violent means has been replicated in some studies (e.g., Mann et al.,1997; Abbar et al., 2001), but not in others (e.g., Kunugi et al., 1999; New etal., 1998). These latter authors reported that the LL genotype of the TPH genewas associated with higher scores of impulsive aggression personality traits ascompared with the UL or UU genotypes in Caucasian males, whereas Manucket al. (1999) found exactly the opposite, with aggression and anger-relatedtraits associated with the UL or UU genotype as compared with the LL geno-type. It should be noted that, being intronic, these polymorphisms do not codefor any gene products.
Schmidt, Fox, Rubin, Hu, and Hamer (2002) investigated three candidatepolymorphisms (dopamine receptor gene DRD4, serotonin transporter 5-HTT,and serotonin receptor 5-HT2C) in relation to aggression and shyness in astudy of 4-year-old children. A significant association was found between theDRD4 genotype and maternal report of problems with aggression. However,there were no significant associations of the DRD4 genotype with observed be-havioral measures of aggression. In addition, no significant associations ofeither of the 5-HTT or 5-HT2C serotonin genotype were found with any of thematernal report and observed behavioral measures.
Brunner, Nelen, Breakefield, Ropers, and van Oost (1993) showed that a
Genetics and the Development of Aggression 233

mutation in the MAOA gene could cause a syndrome that includes violenceand impulsivity in a large Dutch family. This mutation is extremely rare, how-ever. Furthermore, an association between MAOA and aggressive behavior inthe general population has not been established. Using data from a longitudi-nal study of 500 adult males, Caspi et al. (2002) found higher levels of antiso-cial behavior in men who had a low-activity MAOA genotype if they had alsoexperienced childhood maltreatment. However, antisocial behavior was not aselevated among men who had the more common high-activity MAOA geno-type even if they had experienced maltreatment as children. Although thesefindings suggest a genotype-by-environment interaction for aggression in hu-mans, a main statistical effect was found for maltreatment but not for theMAOA genotype.
Finally, many studies of testosterone activity show a relation betweenhigh plasma levels and a tendency toward aggression (see Van Goozen, Chap-ter 14, this volume). Jonsson et al. (2001) investigated the association betweena trinucleotide (CAG) repeat polymorphism in the androgen receptor geneand various personality traits. There were tendencies in some scales indicatingpossible relationships between the androgen receptor allele length and person-ality traits related to dominance and aggression. After correction for multipletesting, however, no significant differences were found.
To summarize, linkage and association studies of aggression have so farlargely resulted in contradictory findings characterized by failed attempts atreplication. Most research has focused on aggression assessed as a personalitytrait, as opposed to a behavior. Studies generally report simple tests of associa-tion, with or without correction for multiple testing. Explained phenotypicvariance is rarely reported, and potential confounding variables, such as pop-ulation stratification and environmental risk factors, are seldom taken intoaccount. It is still premature to provide a definite conclusion concerning themolecular-genetic causes of individual differences in aggression. Studies maybe generally underpowered to detect polygenes of very small effects, and manycandidate genes remain to be investigated. Nevertheless, the inconclusive re-sults observed so far are compatible with the equally disappointing findingsobtained for most other complex behavioral phenotypes (Goldman & Maz-zanti, 2002). These weak findings appear somehow difficult to reconcile withthe otherwise strong heritability estimates found for those phenotypes.
CONCLUSION
More than two decades ago, Plomin (1981) noted that parental ratings of anycharacteristic tend to yield high heritability estimates, suggesting that parentstend to rate identical twins more similarly than fraternal twins on all mea-sures. Concerning the lack of heritability found for objective assessments ofaggressive behavior, Miles and Carey concluded at the end of their 1997review: “Given that almost all substantive conclusions about the genetics of
234 D E T E R M I N A N T S O F A G G R E S S I O N

personality have been drawn from self or parental reports, this last finding hasobvious and important implications for both aggression research in particularand personality research in general.” In her review of the genetics of aggres-sion in children, DiLalla (2002) concluded that “although a large number ofstudies support the heritability of childhood aggression, it is still inconclu-sive.”
To interpret the mixed findings presented in this chapter, it is importantto probe a number of theoretical issues on the genetic and developmental ori-gins of aggression. First, what does the term genetic influence entail concern-ing the development of a phenotype like aggression? As such, the term doesnot refer simply to whether genes play a part in the production of aggression,because it is taken for granted that aggression (like all other behavioral pheno-types) is the outcome of a neurobiological machinery that is unquestionablyunderwritten by genes, as is amply documented by the studies reviewed here.Rather, what behavior geneticists mean when evoking a genetic influence onaggression is that variation in aggression between individuals is partiallycaused by genetic differences between these individuals.
The distinction is important because although we posit a genetic sub-strate for aggression (along with nongenetic factors), this in no way impliesthat this substrate varies between individuals. Nor, if it does, that the geneticvariation contributes to phenotypic variation in the development of aggressivebehavior. It is clear, by analogy, that we posses two arms and two legs becauseof genetic factors. The phenotype “limb number” is unquestionably specifiedin our DNA. Differences between individuals in this phenotype, however, areoverwhelmingly caused by nongenetic factors (e.g., teratogens, accidents, sur-gical amputations) and the heritability (i.e., the magnitude of “genetic influ-ence”) of number of arms and legs thus approaches zero.
Given this meaning of genetic influences on aggression (i.e., genetic vari-ants causing individual differences in aggression), a fundamental questionmust be addressed. Theoretically, should we expect genetic variation betweenindividuals for a trait like aggressive behavior? The general answer to thisquestion is, no, we should not. This is because several well-known processesgovern the evolution of gene frequencies and variation among living orga-nisms. Genetic variation is fueled by mutations and can be temporarily main-tained by certain phenomena such as heterozygote vigour, genetic drift, andevolutionarily stable strategies. However, the overarching process guiding ge-netic evolution is natural selection. And natural selection, inexorably, stampsout genetic variation. The phenotypic outcome of natural selection is adapta-tion, the ultimate measure of which being Darwinian fitness. There is a truismamong plant and animal geneticists that “one cannot breed for fitness.” Thereason is simple: With fitness being the result of natural selection, whichweeds out genetic variation, breeding for fitness is impossible, for lack of ge-netic variation. In natural populations, it has indeed been shown that fitness-related traits such as size, longevity, and mating behavior have very low (nearthe mutational rate) or zero heritability. Thus, the data reviewed here on arti-
Genetics and the Development of Aggression 235

ficial breeding for aggression-related traits in laboratory animals does nottruly inform us on the existence of genetic variants that would account fornaturally occurring variation in aggression in these species.
Is aggressiveness a fitness-related trait? In elephant seals, for instance,intermale aggression and success in the monopolization of ovulating females isdirectly proportional to reproductive success. Studies covering the lifespan ofelephant seals, however, show that individual males do not differ significantlyin their lifetime reproductive success (LeBoeuf & Laws, 1994). This is becausemale aggression is directly proportional to size, which itself depends on age:With time, each male passes through the optimal period in terms of develop-ment and exhibits approximately the same level of aggression pertaining to itsage and size—thereby achieving roughly the same lifetime reproductive suc-cess. In this case, genes affecting the development of aggression seem to havelong been “fixed” by natural selection, suboptimal genetic variants havingbeen selected out.
It is uncertain whether aggression in humans was a crucial fitness-relatedtrait during our evolutionary past, but there is no reason to believe that we areunique among mammals or, for that matter, among living organisms. The in-conclusive results reviewed here concerning the quantitative and molecularquest for genetic variants associated with individual differences in the devel-opment of aggression appear, in this light, less than surprising.
The recent studies of Suomi (2003) and Caspi and colleagues (2002)point to a new era of behavior genetics in which specific genes and specific en-vironmental risk will be measured in longitudinal designs to assess theirdynamic interplay during development. This search should be guided by quan-titative genetic assessments of the magnitude of genetic–environmental influ-ences as they unfold during development from birth to adulthood, taking careto document environmental risk directly and prospectively, to test designfeatures such as the equal environment assumption, and to ensure unbiasedassessment of aggression phenotypes themselves.
REFERENCES
Abbar, M., Courtet, P., Bellivier, F., Leboyer, M., Boulenger, J. P., Castelhau, D., Ferreira,M., Lambercy, C., Mouthon, D., Paoloni-Giacobino, A., Vessaz, M., Malafosse, A.,& Buresi, C. (2001). Suicide attempts and the tryptophan hydroxylase gene. Molec-ular Psychiatry, 6, 268–273.
Arseneault, L., Moffitt, T. E., Caspi, A., Taylor, A., Rijsdijk, F. V., Jaffee, S., Ablow, J. C.,& Measelle, J. R. (2003). Strong genetic effects on cross situational antisocial be-havior among 5–year-old children according to mothers, teachers, examiner-ob-servers, and twins’ self-reports. Journal of Child Psychology and Psychiatry, 44,832–848.
Bohman, M., Cloninger, C. R., Sigvardsson, S., & Von Knorring, A.-L. (1982). Predispo-sition to petty criminality in Swedish adoptees: I. Genetic and environment hetero-geneity. Archives of General Psychiatry, 39, 1233–1241.
236 D E T E R M I N A N T S O F A G G R E S S I O N

Broadhurst, P. L. (1958). Determinants of emotionality in the rat: III. Strain differences.Journal of Comparative Physiology and Psychology, 51, 55–59.
Brunner, H, G., Nelen, M., Breakefield, X. O., Ropers, H. H., & Van Oost, B. A. (1993).Abnormal behavior associated with a point mutation in the structural gene formonoamine oxidase. Science, 262, 578–580.
Cadoret, R. J., Cain, C. A., & Crowe, R. R. (1983). Evidence for gene–environment inter-action in the development of adolescent antisocial behavior. Behavior Genetics, 13,301–310.
Cairns, R. B., Gariépy, J.-L., & Hood, K. E. (1990). Development, microevolution, andsocial behavior. Psychological Review, 97, 49–65.
Cairns, R. B., Hood, K. E., & Midlam, J. (1985). On fighting in mice: Is there a sensitiveperiod for isolation effects? Animal Behavior, 33, 166–180.
Cairns, R. B., MacCombie, D. J., & Hood, K. E. (1983). A developmental–genetic analy-sis of fighting in mice: I. Behavioral outcome. Journal of Comparative Physiologyand Psychology, 97, 69–89.
Carey, G. (1994). Genetics and violence. In A. J. Reiss, K. A. Miczek, & J. A. Roth (Eds.),Understanding and preventing violence (Vol. 2, pp. 21–58). Washington, DC: Na-tional Academy Press.
Carlier, M., Roubertoux, P. L., Kottler, M. L, & Degrelle, H. (1990). Y chromosome andaggression in strains of laboratory mice. Behavior Genetics, 20, 137–156.
Caspi, A., McClay, J., Moffitt, T. E., Mill, J., Martin, J., Craig, I. W., Taylor, A., &Poulton, R. (2002). Role of genotype in the cycle of violence in maltreated children.Science, 297, 851–854.
Caspi, A., Moffitt, T. E., Morgan, J., Rutter, M., Taylor, A., Arseneault, L., Tully, L.,Jacobs, C., Kim-Cohen, J., & Polo-Tomas, M. (2004). Maternal expressed emotionpredicts children’s antisocial behavior problems: Using MZ-twin differences toidentify environmental effects on behavioral development. Developmental Psychol-ogy, 40(2), 149–161.
Clerget-Darpoux, F., Goldin, L. R., & Gershon, E. S. (1986). Clinical methods in psychi-atric genetics: III. Environmental stratification may simulate a genetic effect inadoption studies. Acta Psychiatrica Scandinavica, 74, 305–311.
Darvasi A. (1998). Experimental strategies for the genetic dissection of complex traits inanimal models. Nature Genetics, 18, 19–24.
DiLalla, L. F. (2002). Behavior genetics of aggression in children: Review and future di-rections. Developmental Review, 22, 593–622.
Dionne, G., Tremblay, R. E., Boivin, M., Laplante, D., & Pérusse, D. (2003). Physical ag-gression and expressive vocabulary in 19–month-old twins. Developmental Psy-chology, 37, 261–273.
Ebert, P. D., & Hyde, J. S. (1976). Selection for agonistic behavior in wild female Musmusculus. Behavior Genetics, 6, 291–304.
Ebstein, R. P., Gritsenki, I., Nemanov, L., Fisher, A., Osher, Y., & Belmaker, R. H. (1997).No association between the serotonin transporter gene regulatory region polymor-phism and the tridimensional personality questionnaire (TPQ) temperament ofharm avoidance. Molecular Psychiatry, 2, 224–226.
Fergusson, D. M., Lynskey, M., & Horwood, L. J. (1995). The adolescent outcomes ofadoption: A 16–year longitudinal study. Journal of Child Psychology and Psychia-try, 36, 597–615.
Fuller, J. L., & Thompson, W. R. (1978). Foundations of behavior genetics. St. Louis,MO: Mosby.
Genetics and the Development of Aggression 237

Gariépy, J.-L., Gendreau, P. L., Mailman, R. B., Tancer, M. E., & Lewis, M. H. (1995).Rearing conditions alter social reactivity and D1 dopamine receptors in high- andlow-aggressive mice. Pharmacology, Biochemistry and Behavior, 51, 767–773.
Gariépy J.-L., Rodriguiz, R. M., & Jones, B. C. (2002). Handling, genetic and housing ef-fects on the mouse stress system, dopamine function, and behavior. Pharmacology,Biochemistry, and Behavior, 73, 7–17.
Ge, X., Conger, R. D., Cadoret, R. J., Neiderhiser, J. M., Yates, W., Troughton, E., &Stewart, M. A. (1996). The developmental interface between nature and nurture: Amutual influence model of child antisocial behavior and parent behaviors. Develop-mental Psychology, 32, 574–589.
Gelernter, J., Kranzler, H., Coccaro, E. F., Siever, L. J., & New, A. F. (1998). Serotonintransporter protein gene polymorphism and personality measures in African Ameri-can and European American subjects. American Journal of Psychiatry, 155, 1332–1338.
Gendreau, P. L., Gariépy, J. L., Petitto, J. M., & Lewis, M. H. (1997). D1 dopamine recep-tor mediation of social and nonsocial emotional reactivity in mice: Effects of hous-ing and strain difference in motor activity. Behavioral Neuroscience, 111, 424–434.
Gendreau, P. L., Petitto, J. M., Petrova, A., Gariépy, J.-L., & Lewis, M. H. (2000). D3 andD2 dopamine receptor agonists differentially modulate social-emotional reactivityin mice. Behavioural Brain Research, 114,107–117.
Ginsburg, B., & Allee, W. C. (1942). Some effects of conditioning on social dominanceand subordination in inbred strains of mice. Physiological Zoology, 15, 485–506.
Goldman, D., & Mazzanti, C. (2002). From phenotypes to genes and back: A critical ap-praisal of progress so far. In J. Benjamin, R. P. Epstein, & R. H. Belmaker (Eds.),Molecular genetics and the human personality (pp. 273–291). Washington, DC:American Psychiatric Press.
Gottesman, I. I., & Goldsmith, H. H. (1994). Developmental psychopathology of antiso-cial behavior: Inserting genes into its ontogenesis and epigenesist. In C. A. Nelson(Ed.), Threats to optimal development: Integrating biological, psychological, andsocial risk factors (pp. 69–104). Hillsdale, NJ: Erlbaum.
Hood, K. E., & Cairns, R. B. (1988). A developmental-genetic analysis of aggressive be-havior in mice. II. Cross-sex inheritance. Behavior Genetics, 18, 605–619.
Huang, Y., Grailhe, R., Arango, V., Hen, R., & Mann, J. J. (1999). Relationship ofpsychopathology to the human serotonin 1B genotype and receptor binding kineticsin postmortem brain tissue. Neuropsychopharmacology, 21, 238–246.
Hunt, G. J., Guzman-Novoa, E., Fondrk, M. K., & Page, R. E., Jr. (1998). Quantitativetrait loci for honey bee stinging behavior and body size. Genetics, 148, 1203–1213.
Hyde, J. S., & Ebert, P. D. (1976). Correlated response in selection for aggressiveness infemale mice. I. Male aggressiveness. Behavior Genetics, 6, 421–427.
Jaffee, S. R., Caspi, A., Moffitt, T. E., & Taylor, A. (2004). Physical maltreatment victimto antisocial child: Evidence of an environmentally mediated process. Journal ofAbnormal Psychology, 113(1), 44–55.
Jonsson, E. G., von Gretten, C., Gustavsson, J. P., Yuan, Q.-P., Lindblad-Toh, K.,Forslund, K., Rylander, G., Mattila-Evenden, M., Asberg, M., & Schalling, M.(2001). Androgen receptor trinucleotide repeat polymorphism and personalitytraits. Psychiatric Genetics, 11, 19–23.
Kessler, S., Elliott, G. R., Orenberg, E. K., & Barchas, J. D. (1977). A genetic analysis ofaggressive behavior in two strains of mice. Behavior Genetics, 7, 313–321.
Kunugi, H., Ishida, S., Kato, T., Sakai, T., Tatsumi, M., Hirose, T., & Nanko, S. (1999).
238 D E T E R M I N A N T S O F A G G R E S S I O N

No evidence for an association of polymorphisms of the tryptophan hydroxylasegene with affective disorders or attempted suicide among Japanese patients. Ameri-can Journal of Psychiatry, 156, 774–776.
Lagerspetz, K. M. J. (1961). Genetic and social causes of aggressive behavior in mice.Scandinavian Journal of Psychology, 2, 167–173.
Lagerspetz, K. M. J. (1964). Studies on the aggressive behavior in mice. AnnalesAcadamiae Scientiarum Fennicae, Sarja-series B, 1–131.
Lagerspetz, K. Y. H., Tirri, R., & Lagerspetz, K. M. J. (1968). Neurochemical andendocrinological studies of mice selectively bred for aggressiveness. ScandinavianJournal of Psychology, 9, 157–160.
Lappalainen, J., Long, J. C., Eggert, M., Ozaki, N., Robin, R. W., Brown, G. L., et al.(1998). Linkage of antisocial alcoholism to the serotonin 5-HT1B receptor gene in 2populations. Archives of General Psychiatry, 55, 989–994.
Lathe, R. (1996). Mice, gene targeting and behaviour: More than just genetic back-ground. Trends in Neurosciences, 19, 183–186.
LeBoeuf, B. J., & Laws, R. M. (Eds.). (1994). Elephant seals: Population ecology, behav-ior and physiology. Berkeley: University of California Press.
Lesch, K., Bengel, D., Heils, A., Sabol, S. Z., Greenberg, B. D., Petri, S., et al. (1996).Association of anxiety-related traits with a polymorphism in the serotonin trans-porter gene regulatory region. Science, 274, 1527–1531.
Mann, J. J., Malone, K. M., Nielsen, D. A., Goldman, D., Erdos, J., & Gelernter, J.(1997). Possible association of a polymorphism of the tryptophan hydroxylase genewith suicidal behavior in depressed patients. American Journal of Psychiatry, 154,1451–1453.
Manuck, S. B., Flory, J. D., Ferrell, R. E., Dent, K. M., Peters, D. G., & Muldoon, M. F.(1999). Aggression and anger-related traits associated with a polymorphism of thetryptophan hydroxylase gene. Biological Psychiatry, 45, 603–614.
Mason, D. A., & Frick, P. J. (1994). The heritability of antisocial behavior: A meta-analy-sis of twin and adoption studies. Journal of Psychopathology and Behavioral As-sessment, 16, 301–323.
Maxson, S. C. (1996). Searching for candidate genes with effects on an agonistic behav-ior, offense, in mice. Behavior Genetics, 26, 471–476.
Mednick, S. A., Gabrielli, W. F., & Hutchings, B. (1984). Genetic influences in criminalconvictions: Evidence from an adoption cohort. Science, 224, 891–894.
Miles, D. R., & Carey, G. (1997). Genetic and environmental architecture of human ag-gression. Journal of Personality and Social Psychology, 72, 207–217.
Mouse Genome Sequencing Consortium. (2002). Initial sequencing and comparativeanalysis of the mouse genome. Nature, 410, 520–562.
Nelson, R. J, & Young, K. A. (1998). Behavior in mice with targeted disruption of singlegenes. Neuroscience and Biobehavioral Reviews, 22, 453–462.
New, A. S., Gelernter, J., Yovell, Y., Trestman, R. L., Nielsen, D. A., Silverman, J., et al.(1998). Increases in irritable aggression associated with “LL” genotype at thetryptophan hydroxylase locus. American Journal of Medical Genetics, 81, 13–17.
New, A. S., Goodman, M., Mitropoulou, V., & Siever, L. (2002). Genetic polymorphismsand aggression. In J. Benjamin, R. P. Epstein, & R. H. Belmaker (Eds.), Moleculargenetics and the human personality (pp. 231–244). Washington, DC: American Psy-chiatric Press.
Nielsen, D. A., Goldman, D., Virkkunen, M., Tokola, R., Rawlings, R., & Linnoila, M.
Genetics and the Development of Aggression 239

(1994). Suicidality and 5–hydroxyindolacetic acid concentration associated with atryptophan hydroxylase polymorphism. Archives of General Psychiatry, 51, 34–38.
Nielsen, D. A., Virkkunen, M., Lappalainen, J., Eggert, M., Brown, G. L., Long, J. C., etal. (1998). A tryptophan hydroxylase gene marker for suicidality and alcoholism.Archives of General Psychiatry, 55, 593–602.
Nyberg, J. M., Vekovischeva, O., & Sandnabba, N. K. (2003). Anxiety profiles of miceselectively bred for intermale aggression. Behavior Genetics, 33, 503–511.
O’Connor, T. G., Deater-Deckard, K., Fulkner, D., Rutter, M., & Plomin, R. (1998). Ge-notype-environment correlations in late childhood and early adolescence: Antiso-cial behavioral problems and coercive parenting. Developmental Psychology, 34,970–981.
Owen, D., & Sines, J. O. (1970). Heritability of personality in children. Behavior Genet-ics, 1, 235–248.
Phillips, T. J., Belknap, J. K., Hitzemann, R. J., Buck, K. J., Cunningham, C. L., &Crabbe, J. C. (2002). Harnessing the mouse to unravel the genetics of human dis-ease. Genes, Brain and Behavior, 1, 14–26.
Picciotto, M. R, & Wickman, K. (1998). Using knockout and transgenic mice to studyneurophysiology and behavior. Physiological Review, 78, 1131–1163.
Plomin, R. (1981). Heredity and temperament: A comparison of twin data for self-reportquestionnaires, parental ratings, and objectively assessed behavior. In L. Gedda, P.Parisi, & W. E. Nance (Eds.), Twin research: Vol. 3. Intelligence, personality, and de-velopment (pp. 269–278). New York: Alan R. Liss.
Plomin, R., Foch, T. T., & Rowe, D. C. (1981). Bobo clown aggression in childhood: En-vironment, not genes. Journal of Research in Personality, 15, 331–342.
Raine, A. (2002). Annotation: The role of prefrontal deficits, low autonomic arousal,and early health factors in the development of antisocial and aggressive behavior inchildren. Journal of Child Psychology and Psychiatry, 43, 417–434.
Rhee, S. H., & Waldman, I. D. (2002). Genetic and environmental influences on antiso-cial behavior: A meta-analysis of twin and adoption studies. Psychological Bulletin,128, 490–529.
Sandnabba, N. K., Lagerspetz, K. M., & Jensen, E. (1994). Effects of testosterone expo-sure and fighting experience on the aggressive behavior of female and male mice se-lectively bred for intermale aggression. Hormones and Behavior, 28, 219–231.
Schmidt, L. A., Fox, N. A., Rubin, K. H., Hu, S., & Hamer, D. H. (2002). Molecular ge-netics of shyness and aggression in preschoolers. Personality and Individual Differ-ences, 33, 227–238.
Scott, J. P. (1942). Genetic differences in the social behavior of inbred strains of mice.Journal of Heredity, 33, 11–15.
Sharma, A. R., McGue, M. K., & Benson, P. L. (1998). The psychological adjustment ofUnited States adopted adolescents and their nonadopted siblings. Child Develop-ment, 69, 791–802.
Sluyter, F., Korte, S. M., Bohus, B., & Van Oortmerssen, G. A. (1996). Behavioral stressresponse of genetically selected aggressive and nonaggressive wild house mice in theshock-probe/defensive burying test. Pharmacology, Biochemistry, and Behavior,54, 113–116.
Southwick, C. H., & Clark, L. H. (1968). Interstrain differences in aggressive behaviorand exploratory activity in inbred mice. Communications in Behavioral Biology, 1,49–59.
Stoolmiller, M. (1999). Implications of the restricted range of family environments for es-
240 D E T E R M I N A N T S O F A G G R E S S I O N

timates of heritability and nonshared environment in behavior-genetic adoptionstudies. Psychological Bulletin, 125, 392–409.
Suomi, S. J. (2003). Gene–environment interactions and the neurobiology of social con-flict. Annals of the New York Academy of Science, 1008, 132–139.
Taylor, B. A. (1989). Recombinant inbred strains. In M. L. Lyon & A. G. Searle (Eds.),Genetic variants and strains of the laboratory mouse (2nd ed., pp 773–796). Ox-ford, UK: Oxford University Press.
Taylor, B. A., Wnek, C., Kotlus, B. S., Roemer, N., MacTaggart, T., & Phillips, S. J.(1999). Genotyping new BXD recombinant inbred mouse strains and comparisonof BXD and consensus maps. Mammalian Genome, 10, 335–348.
Van Oortmerssen, G. A., & Baker, T. C. M. (1981). Artificial selection for short and longattack latencies in wild Mus musculus domesticus. Behavior Genetics, 11, 115–126.
Verhulst, F. C., Versluis-den Bieman, H., Van der Ende, J., Berden, G. F. M. G., &Sanders-Woudstra, J. A. R. (1990). Problem behavior in international adoptees: III.Diagnosis of child psychiatric disorders. Journal of the American Academy of Childand Adolescent Psychiatry, 29, 420–428.
Walters, G. D. (1992). A meta-analysis of the gene–crime relationship. Criminology, 30,595–613.
Genetics and the Development of Aggression 241

DETERMINANTS OF AGGRESSIONMapping Brain Development and Aggression
12
Mapping Brain Developmentand Aggression
TOMÁŠ PAUS
BRAIN–BEHAVIOR RELATIONSHIPS
This chapter provides an overview of the basic principles guiding research onbrain–behavior relationships in general, and as applied to studies of aggres-sion during human development in particular. The emphasis is on the use ofstructural and functional neuroimaging to measure age- and behavior-relatedvariations in brain structure and function. It begins with a brief introductionto the methodology of techniques used to map the developing brain, includingmagnetic resonance imaging (MRI), electroencephalography (EEG), and mag-netoencephalography (MEG). I then review the current knowledge of struc-tural maturation, assessed by MRI, of the human brain during childhood andadolescence; given the importance of structural connectivity for interregionalcommunication, the emphasis of this section is on the maturation of whitematter. The next part describes some of the results of neuroimaging studiesaimed at identifying neural circuits involved in various aspects of aggression.The chapter concludes by discussing the potential and limitations of theneuroimaging approach in this field.
The chief motivation for brain mapping studies is identification—in timeand space—of neural circuits associated with particular sensory, motor, andcognitive functions. Although the initial research has focused on the healthybrain of an adult individual, functional neuroimaging is also beginning tomake inroads into the research of normal and abnormal brain development;unraveling the details of structural and functional maturation of various neu-ral systems represents a first step toward the understanding of the biological
242

underpinnings of cognitive development in health and disease. A wide rangeof brain-mapping techniques are available today, including positron emissiontomography (PET), functional MRI (fMRI), EEG, MEG, and transcranialmagnetic stimulation (TMS). These methods allow the researcher to measure(PET, fMRI, EEG, MEG) and to manipulate (TMS) neural activity in the hu-man brain; they differ in spatial and temporal resolution, in the nature of themeasured signal, as well as in the extent of brain coverage. Given theirnoninvasive nature, MRI, EEG, and MEG represent the most common tech-niques employed for developmental studies and are considered in this chapter.The reader interested in the other techniques is referred to other publications(e.g., Paus, 2003; Walsh & Cowey, 2000).
PRINCIPLES OF BRAIN MAPPING (MRI, FMRI, EEG, AND MEG)
Several noninvasive techniques are available today to map the structure andfunction of the human brain during development; those most frequently usedare MRI, EEG, and MEG.
MRI is the only technique that allows the researcher to measure both thestructure and function of the brain. MRI is based on the following principles.Nuclei that have an odd number of nucleons (protons and neutrons) possessboth a magnetic moment and an angular momentum (or spin). In the presenceof an external magnetic field such nuclei precess (or wobble) around their axisat a rate proportional to the strength of the magnetic field, emitting electro-magnetic energy in the process. The hydrogen atom contains only a single pro-ton and therefore precesses when exposed to a magnetic field. In the majorityof MR imaging studies, precessing nuclei of hydrogen associated with waterand fat are the source of the signal.
Structural MRI
Contrast in structural MR images is based on local differences in (1) protondensity (i.e., in the number of hydrogen nuclei per unit of tissue volume) or in(2) either of the following two relaxation times: (a) longitudinal relaxationtime (T1); and (b) transverse relaxation time (T2). Local differences in relax-ation times are reflected in the image contrast because, at a given measure-ment time, the MR signal has already recovered (T1) or decayed (T2) more inregions with short T1 (or T2), and vice versa. For this reason, tissue withshort T1 (white matter) shows a high MR signal and appears bright on T1-weighted images, whereas tissue with long T2 (grey matter) shows high signaland is bright in T2-weighted images (see Figure 12.1).
Although proton density reflects the amount of signal-emitting nucleipresent in the tissue (primarily water), relaxation times and therefore tissue“brightness” on T1- and T2-weighted images depend on a variety of biologi-cal and structural properties of the brain tissue, all of which vary as a function
Mapping Brain Development and Aggression 243

of age. Content of water is one of the most important determinants influenc-ing T1 in the brain; to simplify, the more water there is in a given tissue com-partment, the longer the T1 and the lower the signal on a T1-weighted imagewill be in that compartment. In the adult brain, T1 is the longest incerebrospinal fluid, intermediate in grey matter, and the shortest in white mat-ter. The content of hydrophobic lipids in white matter may also influence T1through magnetic interactions with the hydrogen nuclei of the lipids. The con-tent of iron is another important factor, influencing primarily local magneticinhomogeneities; the higher the content of iron, the shorter the T2. Finally, theanatomic arrangement of axons may influence the amount of interstitial waterand, in turn, T1 values; more tightly bundled axons would have shorter T1and therefore appear brighter on T1-weighted images. Several other MR tech-niques are used to image specific structural properties of the brain. Thus, thepreferred direction of movement of water molecules along the axons is the ba-sis of diffusion tensor imaging (DTI). The chemical composition of the brain isstudied with magnetic resonance spectroscopy (MRS).
Computational Analysis of MR Images
Structural images are but color-coded representations of MR, the signal mea-sured throughout the brain and localized in individual three-dimensional (3-D) elements of the brain image (i.e., voxels). It is therefore relatively straight-forward to apply various computational approaches to quantify a variety ofmorphometric features using fully automatic procedures. In this way, we canmeasure volumes of grey and white matter in the whole brain and its mainsubdivisions (e.g., frontal lobe), the size/volume of well-demarcated brain re-gions (e.g., corpus callosum or caudate nucleus), variations in the shape oflarge (e.g., frontal pole) or small (e.g., cingulate sulcus) morphological entities
244 D E T E R M I N A N T S O F A G G R E S S I O N
FIGURE 12.1. MR images acquired in a 15-year-old individual (sagittal sections).

or those in the cortical thickness. Voxel-wise comparisons of various age (orclinical) groups are used to assess the “density” of grey and white matter (seeFigure 12.2).
Functional MRI
The chief motivation for functional neuroimaging is identification—in timeand space—of neural circuits associated with particular sensory, motor, andcognitive functions and their development, from infancy through adolescence.The high spatial resolution of functional MRI (fMRI) makes this techniqueideal for determining where in the brain changes in neural activity occur. Inthe majority of fMRI studies, task-related changes in local hemodynamics areinferred from changes in the so-called blood-oxygenation–level dependent(BOLD) signal. As its name suggests, the BOLD signal is based on our abilityto detect, with MRI, changes in blood concentration of deoxyhemoglobin.Deoxyhemoglobin is paramagnetic and, as such, acts as an endogenous con-trast agent that creates microscopic magnetic-field inhomogeneities in thesampled tissue. These local inhomogeneities increase the rate of decay oftransverse magnetization, that is, shorten T2 and T2* relaxation times. Uponneural activation, the disproportionate increase in local blood flow “dilutes”the amount of deoxyhemoglobin in the small veins leaving the “activated” tis-sue and, in turn, reduces the amount of local inhomogeneities; this leads to apositive signal on T2*-weighted images (“red” veins in Figure 12.3).
Mapping Brain Development and Aggression 245
FIGURE 12.2. Age-related changes in white matter density in the internal capsule (left) and theleft arcuate fasciculus (right). The thresholded maps of t-statistic values (t > 4.0) are super-imposed on axial (capsule) and sagittal (arcuate) sections through the magnetic resonanceimage of a single subject. The images depict the exact brain locations that showed statisti-cally significant correlations between white matter density and the subject’s age (n = 111;age: 4–17 years). From Paus et al. (1999). Copyright 1999 by the American Association forthe Advancement of Science. Reprinted by permission.

EEG and MEG
Compared with fMRI, more direct and real-time measurements of electricaland magnetic signals generated by brain tissue are recorded by EEG and MEG(Niedermeyer & Lopes da Silva, 1999). EEG and MEG are the methods ofchoice for determining when changes in neural activity occur. The mainsources of these signals are intra- and extracellular currents, or field poten-tials, elicited by the activation of excitatory and inhibitory synapses. Spatialand temporal summation is necessary, however, to generate signals strongenough to be detected outside the head. Such a summation occurs most oftenduring simultaneous excitatory inputs onto apical dendrites of pyramidalcells; the apical dendrites are for the most part oriented in parallel with eachother and are perpendicular to the cortical surface. EEG detects field poten-tials regardless of their orientation relative to the skull. In contrast, MEG canmeasure only magnetic fields perpendicular to the skull. Such fields are gener-ated by tangential current dipoles and, because of the orientation of pyramidalcells and their apical dendrites in the cortex, reflect primarily postsynaptic activ-ity occurring in the cerebral sulci. Analysis of EEG and MEG data for brain map-ping purposes focuses typically on two domains: (1) event-related potentialsand (2) event-related synchronization or desynchronization of brain activity.
MRI STUDIES OF BRAIN DEVELOPMENT
Brain weight reaches adult values (about 1.45 kg) between 10 and 12 years ofage. The fastest growth occurs during the first three years of life, so that by the
246 D E T E R M I N A N T S O F A G G R E S S I O N
FIGURE 12.3. Principles of functional MRI (the blood oxygenation-level dependent signal).Courtesy of Dr. Bruce Pike, Montreal Neurological Institute.

age of 5 years the infant’s brain weighs about 90% of the adult value(Dekaban, 1978). Clearly, the changes in brain morphology in childhood andadolescence are more subtle that those in the first four years of life (see Paus etal., 2001, for a review of MR studies carried out during infancy). Qualitativeevaluation of MR images is of little value at this point, and the ability to ob-tain quantitative measurements is of the essence if we are to detect brain mat-uration during this “late” period of brain development. Several approacheshave been used to obtain such measurements, including (1) semiautomatic orautomatic classification of brain tissue and the subsequent “count” of imageelements classified as a particular tissue type (e.g., white matter), (2) manualoutlining of a structure of interest (e.g., the corpus callosum) on an MR imageand the subsequent calculation of its volume/area, (3) voxel-wise analysis oflocal growth using deformation fields, and (4) voxel-wise statistical analysis ofwhite matter “density.”
Classification of brain tissue into grey and white matter and cerebro-spinal fluid allowed several investigators to calculate the absolute and/or rela-tive brain volume occupied by white matter. In the most extensive study todate, Giedd et al. (1999a) reported age-related changes in 145 children andadolescents (age 4–20 years) in volumes of grey and white matter of the fron-tal, parietal, temporal, and occipital lobes; the study used a cross-sectional (n= 145) and longitudinal (65/145 subjects) design. Volumes were quantified bycombining a technique using an artificial neural network to classify tissuesbased on voxel intensity, with a technique performing nonlinear registrationto a template brain for which the four lobes had been manually defined (Col-lins, Zijdenbos, Baare, & Evans, 1999). A significant increase in the absolutevolume of white matter was found in this study, with the volume increasingsteadily across ages 4–22 by about 12%; this increase was similar in the fourdifferent lobes but was steeper in boys than in girls. These recent findings areconsistent with the previous observations of other authors, including the age-related increase in the cerebral white matter : grey matter ratio (Jernigan &Tallal, 1990; 8–10 years [n = 9] vs. 25–39 years [n = 15]); the increase in abso-lute and relative volumes of “cortical” white matter (Pfefferbaum et al., 1994;3 months–30 years [n = 88]); and increase in the relative volume of cerebralwhite matter (Reiss, Abrams, Singer, Ross, & Denckla, 1996; 5–17 years [n =85]). In the Pfefferbaum et al. (1994) cross-sectional study, cortical white mat-ter continued to increase from birth to about 20 years, after which it leveledoff.
In the majority of the aforementioned studies, age-related increases inwhite-matter volume are accompanied by decreases in grey-matter volume.This observation is best illustrated in a study by Steen, Ogg, Reddick, andKingsley (1997; 4–10 years [n = 19], 10–20 years [n = 31], 20–30 years [n =20]), who measured T1 relaxation times and used these to classify all voxelsinto those containing either “pure” grey or white matter; the number ofwhite-matter and grey-matter voxels increased and decreased with age, respec-tively, with the inverse relationship between the number of “white” and
Mapping Brain Development and Aggression 247

“grey” voxels (r = .64; p < .0001) (Steen et al., 1997). This finding raises thepossibility that a decrease in (cortical) grey-matter volumes may, at least inpart, reflect intracortical myelination (and the resulting partial volume effects)rather than loss of grey-matter tissue due to “pruning.” Recent observationsby Zilles and collaborators are consistent with this view; using high-resolutionMR imaging and post-mortem histological analysis of the (adult) human cere-bral cortex, they could explain up to 80% of variance in T1 relaxation timesby differences in myeloarchitecture in deep cortical layers (Zilles et al., unpub-lished observations).
These changes in the volume of white matter, calculated using varioustissue-classification algorithms, reflect, to a great extent, subtle age-relatedchanges in relaxation times. Steen et al. (1997; see earlier for sample size andage) measured T1 relaxation times in nine regions of interest (ROIs), includingthe four placed in the white matter: (1) the genu of the corpus callosum, (2)frontal white matter, (3) optic radiation, and (4) occipital white matter. Themean values of T1 decreased from childhood (4–10 years) to adulthood (20–30 years) in all four white-matter structures. Significant decreases were alsoobserved in comparing children with adolescents (10–20 years) in all white-matter structures except the genu of the corpus callosum, and in comparingadolescents with young adults in all regions except occipital white matter.Transverse relaxation times (T2) were measured by Hassink, Hiltbrunner,Muller, and Lutschg (1992; 8–10 years [n = 9] and 24–25 years [n = 8]) ineight ROIs, including the frontal subcortical white matter, the corpus callo-sum and the genu of the internal capsule. Although limited by a small sample,significant decreases in T2 values were found from childhood to adulthood inall white-matter regions but the corpus callosum. These two quantitative stud-ies clearly illustrate significant age-related changes in white matter, most likelycaused by small but consistent increases in the degree of myelination.
The volumetric assessment of age-related changes in white matter is oftena global rather than a regional one. This is due to the absence of readily de-tectable boundaries between different fiber tracts constituting the white mat-ter of the four cerebral lobes. This is not the case for the corpus callosum, thelargest fiber tract in the human brain interconnecting the cerebral cortex ofthe left and right hemispheres. The corpus callosum can be easily delineatedon a midsagittal section and, therefore, its area can be measured with greatprecision. Even though the corpus callosum acquires an “adult” appearanceby about the age of 1 year (Barkovich & Kjos, 1988), its growth continues un-til early adulthood. In one of the first MR-based morphometric studies, Pujol,Vendrell, Junque, Marti-Vilalta, and Capdevila (1993) measured the area ofthe corpus callosum in adolescents (11–19 years, n = 14), young adults (20–29years, n = 45) and older adults (30–61 years, n = 31) using a longitudinal de-sign; two scans were acquired in each subject with an average between-scaninterval of 2 years. Not surprisingly, the largest growth in the 2-year periodwas observed in the youngest group (an increase of about 10%). But the areaof the corpus callosum increased significantly even in the group of young
248 D E T E R M I N A N T S O F A G G R E S S I O N

adults (24–25 years, an increase of about 5% over 2 years); no change wasobserved from the age of 27 years on. In a cross-sectional study, Giedd et al.(1999a; 4–18 years, n = 114) confirmed the significant age-related increase inthe total area of the corpus callosum. They were also able to demonstrate thatmost of this growth occurs in the posterior half of the corpus callosum and, inparticular, in the splenium; no growth was observed in the genu. These find-ings were corroborated by the same authors, who followed up their originalsample and rescanned a large number of the subjects (n = 75) at least once 2years later (Giedd et al., 1999b). Again, the growth of the splenium far out-performed that of any other subdivision of the corpus callosum. This findingis somewhat surprising in light of the caudal-to-rostral time sequence ofmyelination of this structure after birth and the presumed late development ofthe prefrontal cortex, a region interconnected through the (rostral) genu of thecorpus callosum. Further studies are necessary to establish whether changes inthe splenium reflect possible ongoing maturation of the inferior temporal andoccipital cortices, the main sources of the callosal fibers in the splenium(Pandya & Seltzer, 1986).
The rostro–caudal wave of growth of the corpus callosum was recentlydemonstrated by a computational analysis of so-called deformation fields(Thompson et al., 2000; 3–15 years, n = 6). The 3-D deformation fields spec-ify, at each voxel, the vector of forces applied to bring the local anatomy ofthe subject’s brain in alignment with that of the template brain. Thompson etal. (2000) rescanned each subject within 1–4 years and, by subtracting localdeformation fields obtained in each subject at Time 1 and Time 2, were able tovisualize changes in the local growth of the corpus callosum. Consistent withthe previous findings, changes in the genu were observed in the youngest sub-ject (scanned at 3 and 6 years of age), whereas changes in the splenium weremost pronounced in rescanning during adolescence (Time 1: 9 years, Time 2:13 years). In order to detect more subtle variations, however, group analysisof deformation fields and its statistical evaluation will be necessary. Chung etal. (2001) have developed a novel statistical analysis of local growth that willallow investigators to evaluate the statistical significance of age-relatedchanges in deformation fields throughout the brain.
Subtle regional variations in white matter can also be evaluated using avoxel-by-voxel analysis of the images. This approach borrows from the con-cepts developed in the context of functional neuroimaging: It is based on theuse of standardized stereotaxic space and voxel-based statistics. In a study ofage-related changes in white matter, we (Paus et al., 1999; 4–18 years, n =111) have observed significant age-related changes in white-matter “density”in the posterior limb of the internal capsule and in the left arcuate fasciculus;the former is likely to contain fibers connecting the motor cortex and spinalcord, and the latter those connecting the anterior (Broca’s) and posterior(Wernicke’s) language areas. The analysis of images obtained in 111 childrenand adolescents included the following steps: (1) nonlinear transformation ofimages into standardized stereotaxic space to remove global and local differ-
Mapping Brain Development and Aggression 249

ences in the size and shape of the individual brains, (2) classification of braintissue into white matter, grey matter, and cerebrospinal fluid, (3) blurring ofwhite-matter binary masks to generate 3-D maps of white-matter “density,”and (4) correlation between voxel values of white-matter density and the sub-ject’s age. Such voxel-based analyses of age-related variations in white- andgrey-matter (Sowell et al., 1999) densities complement the volumetric ap-proach in allowing for subtle local differences to emerge in regions that maynot be delineated as a single volume. However, relatively large numbers of in-dividuals and rather conservative statistical criteria need to be applied to sepa-rate signal from noise in a reliable fashion.
Overall, there is ample evidence that white matter continues to matureduring childhood and adolescence, increasing its volume and becoming moremyelinated. Most of the changes are occurring throughout this period of de-velopment, with no pronounced differences in the rate of maturation duringpuberty. The absence of such developmental “spurs” could, however, reflectthe lack of high-density longitudinal data for this period of life. Several large-scale longitudinal MR studies of brain maturation are under way at the mo-ment; these are likely to provide important insights regarding the structure–function relationship during infancy, childhood, and adolescence in healthyindividuals.
BRAIN MAPPING STUDIES OF AGGRESSION
Most of the literature on the neural underpinnings of aggressive behavior re-flects a theoretical framework that views aggression in the context of antiso-cial behavior, such as violence and delinquency, rather than as one of the es-sential (adaptive) motivational states, such as hunger or maternal instinct (forrecent criticism, see Tremblay, 2003). It is therefore not surprising that thevast majority of previous studies are based on two methodological ap-proaches: (1) clinical studies of overt aggressive behavior in patients withbrain lesions and (2) neuroimaging studies of brain abnormalities in individu-als who had committed acts incompatible with societal norms, such as mur-derers. In the first part of this section, I review this literature and, given its em-phasis on the frontal lobes, I also provide a brief synopsis of the structure andfunction of this part of the primate cerebral cortex. In the second part, I sug-gest an alternative approach for studying aggression and its elements inhealthy subjects; this approach is based on a theoretical framework that viewsaggression as a social interaction between familiar individuals and, conse-quently, includes conflict resolution (reconciliation) as an integral part of ag-gressive behavior (De Waal, 2000).
The search for a neural substrate of aggression has a long history thatstarted with clinical observations of patients with brain lesions. In the 19thcentury, several reports described individuals who had had an injury to thefrontal lobes and, subsequently, showed changes in their “personality.” The
250 D E T E R M I N A N T S O F A G G R E S S I O N

most famous example is Phineas Gage, who suffered a large penetrating injuryto the ventromedial and orbitofrontal cortex, after which he became fitful,irreverent, profane, indifferent, inpatient, obstinate, and unable to plan(Harlow, 1868; Damasio, Grabowski, Frank, Galaburda, & Damasio, 1994).In the 20th century, several studies of war veterans confirmed a link betweenlesions to frontal lobes and an increased risk of aggressive behavior (e.g.,Grafman et al., 1996, reviewed in Brower and Price, 2001; Kleist, 1934, citedin Brower & Price, 2001; Lishman, 1968; Virkkunen, Nuutila, & Huusko,1976). In the majority of these studies, the injury occurred in young maleadults, who were studied many years after their injuries. Several case studiesalso described a high incidence of aggressive behavior in adults who sufferedlesions to the frontal cortex early (before the age of 8 years) in their lives (An-derson, Bechara, Damasio, Tranel, & Damasio, 1999; Eslinger, Grattan,Damasio, & Damasio, 1992; Price, Daffner, Stowe, & Mesulam, 1990). To-gether, these lesion studies suggest that an intact frontal cortex is importantfor adequate control of aggressive behavior in adulthood; the early-lesioncases suggest that remaining brain structures cannot compensate for the lossof brain tissue in childhood. Which parts of the frontal lobes are critical in thisrespect? A reconstruction of the skull injury suffered by Phineas Gage ledDamasio and colleagues to conclude that “the lesion favored the ventromedialregion of both frontal lobes while sparing the dorsolateral” (Damasio et al.,1994). The Vietnam Head Injury Study found a similar association betweenaggressive behavior and lesions in the orbital and medial aspects of the frontallobes (Grafman et al., 1996). To understand the significance of this pattern, abrief review of the organization of the primate frontal cortex can be helpfulhere.
The frontal cortex constitutes about 35% of all cerebral cortex in hu-mans and great apes (Semendeferi, Lu, Schenker, & Damasio, 2002). It con-tains the primary motor cortex, premotor cortex (dorsal and ventral premotorcortex on the lateral convexity; supplementary motor area (SMA) and thecingulate motor areas (CMA) on the medial convexity), and the prefrontalcortex. The prefrontal cortex is further subdivided into the mid-dorsolateraland mid-ventrolateral frontal cortex, the frontopolar cortex, the orbitofrontalcortex, and the mesial frontal cortex; the latter also includes the anteriorcingulate cortex surrounding the anterior part of the corpus callosum. Each ofthese gross morphological subdivisions consists of a number of areas that dif-fer in the exact arrangement of cells in the six cortical layers (cytoarchitecture;see Figure 12.4) and in their connectivity with other cortical and subcorticalregions. These anatomical differences determine to a great extend what typeof information is being processed in a given region, and which other regionsshare this information. Such regional specialization is reflected, in turn, in thespecific contributions made by a given “node” of a particular neural networkin support of a specific behavior.
Given the structural heterogeneity of the frontal cortex and the presenceof extensive connectivity of frontocortical regions with other parts of the cere-
Mapping Brain Development and Aggression 251

bral cortex, most prominently with the temporal and parietal cortex, it is notsurprising that the frontal cortex supports a multitude of motor, sensory, andcognitive functions. For example, the premotor areas on the medial convexitymay be essential for sequencing behavior (SMA) and willed initiation and sup-pression of movement (CMA; reviewed in Paus, 2001). The mid-dorsolateraland mid-ventrolateral frontal cortex play an important role in working memoryand retrieval, respectively (Petrides, 1996). Maturation of the mid-dorsolateralfrontal cortex underlies the emergence of working memory during infancy(e.g., A-not-B test) and a further increase in working memory and planningduring childhood. Maturations of the lateral prefrontal and/or cingulate corti-ces may be critical for the dramatic increase in the child’s ability to suppressexternal interference observed just before the onset of puberty (e.g., suppressionof reflexive eye movements; Munoz, Broughton, Goldring, & Armstrong, 1998;Paus, Babenko, & Radil, 1990). Maturation of the orbitofrontal, medio-frontal, and cingulate cortex may be associated with the development of de-layed gratification and other aspects of intentional behavior, including the controlof motivation states such as aggression. Underlying the latter control of motiva-tional states are extensive connections between the orbitofrontal/cingulatecortex and the amygdala (Barbas, 1995; Morecraft & Van Hoesen, 1998).
The second “classical” approach to studies of the neural underpinningsof aggressive behavior is that of imaging the brains of individuals who showdifferent forms of violent or antisocial behavior; unlike the neurological pa-tients described earlier, these individuals have not suffered any known exter-nal damage to their brains. Using structural MR imaging, two recent studiesdescribed reductions of the volume of grey matter in different parts of thefrontal lobes in aggressive patients with temporal lobe epilepsy (Woermann etal., 2000) and individuals with antisocial personality disorder (Raine, Lencz,Bihrle, LaCasse, & Colletti, 2000); the latter study described a 11% reductionin the volume of the orbitofrontal cortex in 21 community volunteers with an-
252 D E T E R M I N A N T S O F A G G R E S S I O N
FIGURE 12.4. Schematic diagrams of the lateral (left), medial (middle), and inferior (right) sur-faces of the human frontal lobe to illustrate its cytoarchitectonic parcellation. From Petridesand Pandya (1994). Copyright 1994 by Elsevier. Reprinted by permission.

tisocial personality; like changes were not found in two different controlgroups. Using functional imaging, Raine, Buchsbaum, and LaCasse (1997) de-scribed hypometabolism in the prefrontal cortex in 41 individuals chargedwith murder or manslaughter; in subsequent reanalyses of this sample, Raineand colleagues found that this hypometabolism was present only in those indi-viduals who did not have history of psychosocial deprivation (Raine, Pihl,Stoddard, Bihrle, & Buchsbaum, 1998b) and those who committed impulsive/emotional rather than purposeful aggression (Raine et al., 1998a). Similarfindings were obtained in other clinical populations (reviewed in Brower &Price, 2001).
Given the above lesion and imaging findings, we may ask: What is hap-pening in the human brain during early infancy and childhood that “enables”aggressive behavior, which arguably peaks between 2 and 4 years of age(Tremblay, 2003; Tremblay et al., 2004) and then provides adaptive mecha-nisms, allowing its expression and resolution in a socially acceptable manner?Admittedly, we know too little about structural and functional developmentof the human brain to answer these questions at this point. It is clear that themost dramatic phase of brain growth takes place during the first 2 years oflife: brain weight more than doubles, the overall volume of white matter andthe degree of myelination increases sharply, with the overall increases in greymatter being less dramatic (e.g., Matsuzwa et al., 2001; Paus et al., 2001;Utsunomia, Takano, Okazaki, & Mitsudome, 1999). It is important to notethat some interesting regional differences are beginning to emerge. For exam-ple, the volume of grey matter constituting the hippocampal formation showsa fast increase during the first 2 years of life and continues to increase slowlyduring childhood and adolescence (Utsunomyia et al., 1999). However, thevolume of prefrontal grey matter appears to increase slowly until the age of 8years, followed by rapid growth between 8 and 14 years (Kanemura, Aihara,Aoki, Araki, & Nakazawa, 2003). Although these patterns are based oncross-sectional studies of a relatively small number of infants and children,they provide a preview of what we may learn from several large-scale MR-based developmental studies that are under way. Returning to our question, itis possible that the development of structures in the mesial temporal lobes,such as the hippocampus and amygdala, underlie the initial development ofaggressive behavior, whereas the relatively late development of cortical re-gions, such as the prefrontal and parietal cortex, enables adaptive regulationof aggression in the social context. Let us now examine some of the processes,and their neural substrates, that may be important for the latter.
As suggested in the opening paragraph of this section, most acts of ag-gression take place in the context of social interactions among familiar indi-viduals, be it at home, daycare, school, or the workplace. As with any otherforms of communication, each side of the conflict emits and receives a numberof cues, both verbal and nonverbal, that are essential components of the over-all dynamics of their (aggressive) interaction. I suggest that the study of theneural substrates of such interactions, and their normal development, will
Mapping Brain Development and Aggression 253

benefit from the use of the experimental paradigms utilized over the past de-cade by cognitive neuroscientists interested in the brain mechanisms of inter-personal communication carried by gaze, face, body, and voice. Over the pastseveral years a number of brain-mapping studies carried out with PET, fMRI,and EEG identified a key set of brain regions involved in the processing of “bi-ological motion” generated by the movements of the eyes, mouth, and body.In such studies the subjects view stimuli such as eye and mouth movements(e.g., Puce, Allison, Bentin, Gore, & McCarthy, 1998), hand movements/actions (e.g., Beauchamp, Lee, Haxby, & Martin, 2002; Decety et al., 1997;Grezes, Costes, & Decety, 1999), or body movements conveyed by pointlights attached to the body (e.g., Bonda, Petrides, Ostry, & Evans, 1996).Among other brain regions, biological motion leads consistently to the in-creases in neural activity along the superior temporal sulcus, (STS) (Figure12.5; reviewed in Allison, Puce, & McCarthy, 2000); the exact location maydepend on the moving body part (e.g., mouth vs. eyes vs. hands) and thesource of movement (tool vs. hand; Beauchamp et al,. 2002).
Although most of the previous studies of biological motion used emotion-ally “neutral” stimuli, several investigators observed changes in brain activityin structures involved in processing emotional salience (amygdala) or rewardvalue (orbitofrontal cortex). As suggested by Allison et al. (2000), feedfor-ward and feedback interactions between the STS and the amygdala may becritical for the discrimination of various facial expressions and for the
254 D E T E R M I N A N T S O F A G G R E S S I O N
FIGURE 12.5. Regions along the left (a) and right (b) superior temporal sulcus of the humanbrain “activated” by the movement of various parts of the body. From Allison, Puce, andMcCarthy (2000). Copyright 2000 by Elsevier. Reprinted by permission.

attentional enhancement of the neural response to socially salient stimuli.Consistent with such an “amplification” mechanism, Kilts, Egan, Gideon, Ely,and Hoffman (2003) observed significantly stronger neural response to dy-namic, as compared with static, facial expressions of anger in both the STSand the amygdala. Developmentally, the basic aspects of face perception are inplace shortly after birth (Goren, Sarty, & Wu, 1975), but both the quantityand quality of face processing continues to increase all the way through ado-lescence (e.g., Carey, 1992; McGivern, Andersen, Byrd, Mutter, & Reilly,2002; Taylor, McCarthy, Saliba, & Degiovanni, 1999); perhaps one of thestructural correlates of such continuing maturation of face processing in an in-crease in white matter density along the occipitotemporal pathway (Watkinset al., 2002).
Seeing and hearing signals of upcoming aggression is often enough to pre-vent it; in the monkey, this can be achieved by quickly averting the gaze (e.g.,Capitanio, 2002). The frontal cortex and its various subregions (see the pre-ceding discussion) are likely involved in evaluating the overall context andsignificance of the detected visual (eyes, face) and auditory (vocalization) dis-plays of aggression and in generating the appropriate behavioral response.This response could, nonetheless, be an aggressive act. If this is the case, whathappens next? In many primate species, aggression is often followed by “rec-onciliation” or “conflict resolution” (Aureli & De Waal, 1997). Rather thanviewing aggression as serving a “spacing” function, De Waal argues that “in-dividuals try to ‘undo’ the social damage inflicted by aggression; hence, theywill actively seek contact, specifically with former opponents” (De Waal,2000). Several studies are beginning to document different forms of recon-ciliatory behavior in children observed in naturalistic settings (e.g., Ljungberg,Westlund, & Lindqvist Forsberg, 1999).
Overall, careful studies of the neural substrate underlying variousaggression-relevant elements of social cognition will undoubtedly further ourunderstanding of human aggression, its normal development, and its sociallyunacceptable expressions.
CONCLUSIONS
Structural and functional neuroimaging is making strides in elucidating thestructure–function relationships underlying aggression, as well as providingnovel information on brain maturation in children and adolescents. Previousresearch has focused on studies of adult individuals and overt aggressive be-havior; lesions to the frontal lobes seem to increase the incidence of certainelements of aggressive behavior, whereas having aggression-relevant behav-ioral traits appears to correlate with functional or structural abnormalitiesin this brain region. Very little is known, however, about the relationshipbetween aggression and the individual’s ability to detect and interpret non-verbal cues carried by the gaze, the face, and the body in a social context. I
Mapping Brain Development and Aggression 255

suggest that the study of brain–behavior relationships in this domain wouldbe very helpful when combined, for example, with a detailed assessment ofthe perceptual and cognitive skills of individuals with different developmen-tal histories of aggressive behavior. Finally, it is likely that future researchon the role of the frontal lobes in aggression will shift from studying the ac-tual act of aggression to investigating the brain substrate of reconciliationand conflict resolution.
Although neuroimaging is a valuable tool in this endeavor, one shouldnot forget about its limitations. For example, although T1-weighted MR im-ages provide an impressive contrast between the (cortical) gray and white mat-ter, we are not sure about the biological source of signal differences betweenvarious clinical or age groups; many factors may influence the signal, includ-ing the density of neurons, glia, or vascularization, or the degree of (intra-cortical) myelination. Functional imaging poses different challenges; forexample, are age-related differences in the fMRI signal the cause or conse-quence of age-related changes in a given behavior? In other words, how do wemake sure that the “stimulus” is held constant across different populations sothat we can evaluate the magnitude of neural response to the same probe,rather than use the fMRI signal as a mere correlate of different behavior?Finally, structure–function correlations observed with structural and func-tional neuroimaging are just that, correlations. Other experimental ap-proaches must be used to confirm whether a given region is necessary for agiven behavior; such an inference can be made only when irreversible (lesion)or reversible (electrical or pharmacological stimulation) manipulations lead tothe predicted behavioral consequences. Although some of these techniques canbe used in (adult) human subjects, the ultimate tests would require animalmodels.
ACKNOWLEDGMENTS
My research is supported by the Canadian Institutes of Health Research, the CanadianFoundation for Innovation, and the National Science and Engineering Research Coun-cil of Canada. I thank Drs. Richard Tremblay and Gabriel Leonard for their commentson the early version of this chapter.
REFERENCES
Allison, T., Puce, A., & McCarthy, G. (2000). Social perception from visual cues: Role ofthe STS region. Trends in Cognitive Science, 4, 267–278.
Anderson, S. W., Bechara, A., Damasio, H., Tranel, D., & Damasio, A. R. (1999). Impair-ment of social and moral behavior related to early damage in human prefrontal cor-tex. Nature Neuroscience, 2, 1032–1037.
Aureli, F., & De Waal, F. B. (1997). Inhibition of social behavior in chimpanzees underhigh-density conditions. American Journal of Primatology, 41, 213–228.
256 D E T E R M I N A N T S O F A G G R E S S I O N

Barbas, H. (1995). Anatomic basis of cognitive-emotional interactions in the primateprefrontal cortex. Neuroscience and Biobehavioral Reviews, 19, 499–510.
Barkovich, A. J., & Kjos, B. O. (1988). Normal postnatal development of the corpus cal-losum as demonstrated by MR imaging. American Journal of Neuroradiology, 9,487–491.
Beauchamp, M. S., Lee, K. E., Haxby, J. V., & Martin, A. (2002). Parallel visual motionprocessing streams for manipulable objects and human movements. Neuron, 34,149–159.
Bonda, E., Petrides, M., Ostry, D., & Evans, A. (1996). Specific involvement of humanparietal systems and the amygdala in the perception of biological motion. Journal ofNeuroscience, 16, 3737–3744.
Brower, M. C., & Price, B. H. (2001). Neuropsychiatry of frontal lobe dysfunction in vio-lent and criminal behaviour: a critical review. Journal of Neurology, Neurosurgery,and Psychiatry, 71, 720–726.
Capitanio, J. P. (2002). Sociability and responses to video playbacks in adult male rhesusmonkeys (Macaca mulatta). Primates, 43, 169–177.
Carey, S. (1992). Becoming a face expert. Philosophical Transactions of the Royal Societyof London. Series B: Biological Sciences, 335, 95–102.
Chung, M. K., Worsley, K. J., Paus, T., Cherif, C., Collins, D. L., Giedd, J. N., Rapoport,J. L., & Evans, A. C. (2001). A unified statistical approach for deformation-basedmorphometry. NeuroImage, 14, 595–606.
Collins, D. L., Zijdenbos, A. P., Baare, W. F. C., & Evans, A. C. (1999). Animal + insect:Improved cortical structure segmentation. In A. Kuba, M. Samal, & A. Tod (Eds.),Proceedings of the 16th International Conference on Information Processing inMedical Imaging (IPMI) (pp. 210–223). New York: Springer-Verlag.
Damasio, H., Grabowski, T., Frank, R., Galaburda, A. M., & Damasio, A. R. (1994).The return of Phineas Gage: Clues about the brain from the skull of a famous pa-tient. Science, 264, 1102–1105.
Decety, J., Grezes, J., Costes, N., Perani, D., Jeannerod, M., Procyk, E., Grassi, F., &Fazio, F. (1997). Brain activity during observation of actions. Influence of actioncontent and subject’s strategy. Brain, 120, 1763–1777.
Dekaban, A. S. (1978). Changes in brain weights during the span of human life: Relationof brain weights to body heights and body weights. Annals of Neurology, 4, 345–356.
De Waal, F. B. (2000). Primates: A natural heritage of conflict resolution. Science, 289,586–590.
Eslinger, P. J., Grattan, L. M., Damasio, H., & Damasio, A. R. (1992). Developmentalconsequences of childhood frontal lobe damage. Archives of Neurology, 49(7),764–769.
Giedd, J. N., Blumenthal, J., Jeffries, N. O., Castellanos, F. X., Liu, H., Zijdenbos, A., Paus,T., Evans, A. C., & Rapoport, J. L. (1999a). Brain development during childhood andadolescence: A longitudinal MRI study. Nature Neuroscience, 2, 861–863.
Giedd, J. N., Blumenthal, J., Jeffries, N. O., Rajapakse, J. C., Vaituzis, A. C., Liu, H.,Berry, Y. C., Tobin, M., Nelson, J., & Castellanos, F. X. (1999b). Development ofthe human corpus callosum during childhood and adolescence: A longitudinal MRIstudy. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 23, 571–588.
Goren, C. C., Sarty, M., & Wu, P. Y. (1975). Visual following and pattern discriminationof face-like stimuli by newborn infants. Pediatrics, 56, 544–549.
Mapping Brain Development and Aggression 257

Grafman, J., Schwab, K., Warden, D., Pridgen, A., Brown, H. R., & Salazar, A. M.(1996). Frontal lobe injuries, violence, and aggression: A report of the VietnamHead Injury Study. Neurology, 46, 1231–1238.
Grezes, J., Costes, N., & Decety, J. (1999). The effects of learning and intention on theneural network involved in the perception of meaningless actions. Brain, 122,1875–1887.
Harlow, J. M. (1868). Recovery from the passage of an iron bar through the head. Publi-cations of the Massachusetts Medical Society, 2, 327–347.
Hassink, R. I., Hiltbrunner, B., Muller, S., & Lutschg, J. (1992). Assessment of brain mat-uration by T2–weighted MRI. Neuropediatrics, 23, 72–74.
Jernigan, T. L., & Tallal, P. (1990). Late childhood changes in brain morphology observ-able with MRI. Developmental Medicine and Child Neurology, 32, 379–385.
Kanemura, H., Aihara, M., Aoki, S., Araki, T., & Nakazawa, S. (2003). Development ofthe prefrontal lobe in infants and children: A three-dimensional magnetic resonancevolumetric study. Brain and Development, 25(3), 195–199.
Kilts, C. D., Egan, G., Gideon, D. A., Ely, T. D., & Hoffman, J. M. (2003). Dissociableneural pathways are involved in the recognition of emotion in static and dynamic fa-cial expressions. NeuroImage, 18, 156–168.
Lishman, W. A. (1968). Brain damage in relation to psychiatric disability after head in-jury. British Journal of Psychiatry, 114, 373–410.
Ljungberg, T., Westlund, K., & Lindqvist Forsberg, A. J. (1999). Conflict resolution in 5-year-old boys: Does postconflict affiliative behaviour have a reconciliatory role?Animal Behavior, 58, 1007–1016.
Matsuzawa, J., Matsui, M., Konishi, T., Noguchi, K., Gur, R., Bilker, W., & Miyawaki, T.(2001). Age-related volumetric changes of brain gray and white matter in healthyinfants and children. Cerebral Cortex, 11, 335–342.
McGivern, R. F, Andersen, J., Byrd, D., Mutter, K. L., & Reilly, J. (2002). Cognitive effi-ciency on a match to sample task decreases at the onset of puberty in children. Brainand Cognition, 50, 73–89.
Morecraft, R. J., & Van Hoesen, G. W. (1998). Convergence of limbic input to thecingulate motor cortex in the rhesus monkey. Brain Research Bulletin, 45, 209–232.
Munoz, D. P., Broughton, J. R., Goldring, J. E., & Armstrong, I. T. (1998). Age-relatedperformance of human subjects on saccadic eye movement tasks. ExperimentalBrain Research, 121, 391–400.
Niedermeyer, E., & Lopes da Silva, F. (Eds.). (1999). Electroencephalography. Basic prin-ciples, clinical applications, and related fields (4th ed.). Baltimore: Williams &Wilkins.
Pandya, D. N., & Seltzer, B. (1986). The topography of commissural fibers. In F. Lepore,M. Ptito, & H. Jasper (Eds.), Two hemispheres, one brain: Functions of the corpuscallosum (pp. 47–73). New York: Liss.
Paus, T. (2001). Primate anterior cingulate cortex: Where motor control, drive and cogni-tion interface. Nature Reviews Neuroscience, 2, 417–424.
Paus, T. (2003). Principles of functional neuroimaging. In R. B. Schiffer, S. M. Rao, & B.S. Fogel (Eds.), Neuropsychiatry (2nd ed., pp. 63–90). New York: Lippincott, Wil-liams & Wilkins.
Paus, T., Babenko, V., & Radil, T. (1990). Development of an ability to maintain verballyinstructed central gaze fixation studied in 8 to 10 year children. International Jour-nal of Psychophysiology, 10, 53–61.
Paus, T., Collins, D. L., Evans, A. C., Leonard, G., Pike, B., & Zijdenbos, A. (2001). Mat-
258 D E T E R M I N A N T S O F A G G R E S S I O N

uration of white matter in the human brain: A review of magnetic-resonance stud-ies. Brain Research Bulletin, 54, 255–266.
Paus, T., Zijdenbos, A., Worsley, K., Collins, D. L., Blumenthal, J., Giedd, J. N.,Rapoport, J. L., & Evans, A. C. (1999). Structural maturation of neural pathways inchildren and adolescents: In vivo study. Science, 283, 1908–1911.
Petrides, M. (1996). Lateral frontal cortical contribution to memory. Seminars in theNeurosciences, 8, 57–63.
Petrides, M., & Pandya, D. N. (1994). Comparative architectonic analysis of the humanand the macaque frontal cortex. In F. Boller & J. Grafman (Eds.), Handbook ofneuropsychology (Vol. 9, pp. 17–58). Amsterdam: Elsevier Science.
Pfefferbaum, A., Mathalon, D. H., Sullivan, E. V., Rawles, J. M., Zipursky, R. B., & Lim,K. O. (1994). A quantitative magnetic resonance imaging study of changes in brainmorphology from infancy to late adulthood. Archives of Neurology, 51, 874–887.
Price, B. H., Daffner, K. R., Stowe, R. M., & Mesulam, M. M. (1990). The compart-mental learning disabilities of early frontal lobe damage. Brain, 113, 1383–1393.
Puce, A., Allison, T., Bentin, S., Gore, J. C., & McCarthy, G. (1998). Temporal cortex ac-tivation in humans viewing eye and mouth movements. Journal of Neuroscience,18, 2188–2199.
Pujol, J., Vendrell, P., Junque, C., Marti-Vilalta, J. L., & Capdevila, A. (1993).When doeshuman brain development end? Evidence of corpus callosum growth up to adult-hood. Annals of Neurology, 34, 71–75.
Raine, A., Buchsbaum, M., & LaCasse, L. (1997). Brain abnormalities in murderers indi-cated by positron emission tomography. Biological Psychiatry, 42, 495–508.
Raine, A., Lencz, T., Bihrle, S., LaCasse, L., & Colletti, P. (2000). Reduced prefrontalgray matter volume and reduced autonomic activity in antisocial personality disor-der. Archives of General Psychiatry, 57, 119–127.
Raine, A., Meloy, J. R., Bihrle, S., Stoddard, J., LaCasse, L., & Buchsbaum, M. S.(1998a). Reduced prefrontal and increased subcortical brain functioning assessedusing positron emission tomography in predatory and affective murderers. Behav-ioral Sciences and the Law, 16, 319–332.
Raine, A., Pihl, D., Stoddard, J., Bihrle, S., & Buchsbaum, M. (1998b). Prefrontal glucosedeficits in murderers lacking psychosocial deprivation. Neuropsychiatry, Neuro-psychology, and Behavioral Neurology, 11(1), 1–7.
Reiss, A. L., Abrams, M. T., Singer, H. S., Ross, J. L., & Denckla, M. B. (1996). Brain de-velopment, gender and IQ in children. A volumetric imaging study. Brain, 119,1763–1774.
Semendeferi, K., Lu, A., Schenker, N., & Damasio, H. (2002). Humans and great apesshare a large frontal cortex. Nature Neuroscience, 5, 272–276.
Sowell, E. R., Thompson, P. M., Holmes, C. J., Batth, R., Jernigan, T. L., & Toga, A. W.(1999). Localizing age-related changes in brain structure between childhood andadolescence using statistical parametric mapping. NeuroImage, 9, 587–597.
Steen, R. G., Ogg, R. J., Reddick, W. E., & Kingsley, P. B. (1997). Age-related changes inthe pediatric brain: Quantitative MR evidence of maturational changes during ado-lescence. American Journal of Neuroradiology, 18, 819–828.
Taylor, M. J., McCarthy, G., Saliba, E., & Degiovanni, E. (1999). ERP evidence of devel-opmental changes in processing of faces. Clinical Neurophysiology, 110, 910–915.
Thompson, P. M., Giedd, J. N., Woods, R. P., MacDonald, D., Evans, A. C., & Toga, A.W. (2000). Growth patterns in the developing brain detected by using continuummechanical tensor maps. Nature, 404, 190–193.
Mapping Brain Development and Aggression 259

Tremblay, R. E. (2003). Why socialization fails: The case of chronic physical aggression.In B. B. Lahey, T. E. Moffitt, & A. Caspi (Eds.), Causes of conduct disorder and juve-nile delinquency (pp. 182–224). New York: Guilford Press.
Tremblay, R. E., Nagin, D. S., Séguin, J. R., Zoccolillo, M., Zelazo, P., Boivin, M.,Pérusse, D., & Japel, C. (2004). Physical aggression during early childhood: Trajec-tories and predictors. Pediatrics, 114, e43–e50.
Utsunomiya, H., Takano, K., Okazaki, M., & Mitsudome A. (1999). Development of thetemporal lobe in infants and children: Analysis by MR-based volumetry. AmericanJournal of Neuroradiology, 20, 717–723.
Virkkunen, M., Nuutila, A., & Huusko, S. (1976). Effect of brain injury on social adapt-ability: Longitudinal study on frequency of criminality. Acta Psychiatrica Scandin-avica, 53, 168–172.
Walsh, V., & Cowey, A. (2000). Transcranial magnetic stimulation and cognitive neuro-science. Nature Reviews. Neuroscience, 1, 73–79.
Watkins, K. E., Paus, T., Mangin, J.-F., Zijdenbos, A. P., Collins, D. L., Lerch, J., Worsley,K. J., Blumenthal, J., Giedd, J., Rappoport, J., & Evans, A. C. (2002). Maturation ofwhite matter tracts during adolescence: A longitudinal MRI study. Paper presentedat the 8th International Conference on Functional Mapping of the Human Brain,June 2–6, Sendai, Japan. Available on CD-Rom in NeuroImage 16(2).
Woermann, F. G., Van Elst, L. T., Koepp, M. J., Free, S. L., Thompson, P. J., Trimble, M.R., & Duncan, J. S. (2000). Reduction of frontal neocortical grey matter associatedwith affective aggression in patients with temporal lobe epilepsy: An objective voxelby voxel analysis of automatically segmented MRI. Journal of Neurology, Neuro-surgery and Psychiatry, 68, 162–169.
260 D E T E R M I N A N T S O F A G G R E S S I O N

DETERMINANTS OF AGGRESSIONNeuromodulators in Aggression
13
Neuromodulators in the Developmentand Expression of Inhibition
and Aggression
ROBERT O. PIHL and CHAWKI BENKELFAT
Lack of behavioral control/regulation is known to predispose to aggressionand substance abuse/dependency (Pihl, Peterson, & Finn, 1990). Conversely,the ability to inhibit/regulate behavior is a basic characteristic of healthy de-velopment. It is the assumption of this chapter that much of inhibition repre-sents learned behavior. For example, a lack of inhibition is fundamental inimpulsive–reactive aggression, and its frequency and form at various ages isaffected by how biology and experience alter particular neuromechanismsinvolved in behavioral control. Tremblay et al., (1996), among others, have il-lustrated how aggression typically decreases with age, steadily declining froma peak around age 2. Clearly, the form of this behavior changes with age andsex, with the declining frequency likely representing the incorporation by theindividual of societal restrictions and thus behavioral control. To drasticallyrestate Rousseau, we begin not as noble savages but as savages who may ormay not be “nobilized.” Thus, how the ability to inhibit develops and whatmechanisms are operative is a preeminent developmental question.
The purpose of this chapter is to present information about variousneuromodulators that fuel brain development and functioning as it pertains toinhibition and aggression. Foremost among these are the neurotransmitters se-rotonin, dopamine, norepinephrine, and gamma-aminobutyric acid (GABA),as well as their interactions and the myriad of factors that modulate their ac-tivity. We first state some important caveats and then review the evidence sup-porting the involvement of these neurotransmitters and their interactions in
261

inhibition. Finally, we examine how these systems change with developmentand specifically affect the development and functioning of the prefrontal cor-tex and its putative “executive control.”
A FEW ESSENTIAL CAVEATS
That neuromodulators affect inhibition and aggression is a given. The specifi-cation of which neuromodulators play the key roles, and how, is, however, adifferent matter. Generalizations and conclusions must be qualified by at leastfour caveats. First, although we discuss various neurotransmitters and theirrespective systems as separate entities, nothing in the brain functions inde-pendently. The serotonin system, for example, interacts with most other majorsystems affecting behaviors ranging from aggression to sleep. In fact, the con-clusion that the serotonin system’s main role is to modulate the activity ofother systems has much support. Further, pharmacological studies, particu-larly, from which many conclusions regarding how these systems operate aredrawn, rarely impact one system, one set of receptors, and so forth. Thus,findings often presented as specific and clear are in fact suspect.
Second, although we argue for particular functions of certain neuro-transmitters, these functions are neither static nor stable. For example, GABA,well recognized as the major inhibitory neurotransmitter in the brain, hasbeen shown to be a major stimulus for neuronal excitation in the developingbrain (McCarthy, Auger, & Perrot-Sinal, 2002). It is only with brain matura-tion that the more traditional function of neuronal inhibition becomes domi-nant. Facts like this, of course, raise the general issue of plasticity. Nelson(1999) has analogized neural plasticity as a “subtle but orchestrated dancethat occurs between the brain and the environment” (p. 42). The brain shapeshow one reacts to the environment, which in turn shapes how the brain affectsanatomical, neurochemical, and metabolic change. As Nelson notes, thesechanges can be both adaptive and maladaptive for the organism, and for ourpurposes this phenomenon greatly complicates discussion of generic neuro-chemical systems.
Third, consideration of sex differences in the functioning of neuro-chemicals is rare, but is important. Such differences are especially relevant, asa wealth of studies have reported significant differences between men andwomen in levels and types of aggression (see Archer & Lloyd, 2002). Further,laboratory studies of alcohol-related aggression typically display a muchgreater effect for men than women (Hoaken & Pihl, 2000). How hormonesmay alter gene expression, and thus subsequent brain development, particu-larly during adolescence, may have direct ramifications for sex differences inmood disorders and aggression, as well as in personality traits like introver-sion and sensation seeking. The frontal and temporal corticies, as well as theamygdala and hippocampus in particular, reflect significant postpubertal
262 D E T E R M I N A N T S O F A G G R E S S I O N

development. Walker (2002) has recently pointed out the significance of thistime for brain development, which is reflected in the fact that the heritabilityof some behaviors and disorders increase with age, particularly during adoles-cence. Fourth, any attempt to delineate the neurochemical precursors ofinihibition and aggression implies that we have a clear definition and subse-quent measures for these phenomena. We have previously examined the rela-tionship between self-report and behavioral measures of inhibition and havefound it to be low to nonexistent (Helmers, Young, & Pihl, 1995). Similarly,definitions of aggression proliferate, as do measures, both lacking, respec-tively, in operationism and validity.
EVIDENCE FOR THE DIFFERENTIAL INVOLVEMENTOF NEUROTRANSMITTERS IN DISINHIBITION
Serotonin
The contention that altered serotonergic neurotransmission might play an im-portant role in the pathophysiology of impulse-control disorders/aggressivityis not new. Using measurements in lumbar cerebrospinal fluid (CSF) of themain metabolite of serotonin (5-HT), 5-hydroxyindoleacetic acid (5-HIAA),investigators in the 1970s and 1980s proposed theoretical models implicatinglow serotonergic neurotransmission as a critical biological substrate for be-havioral dyscontrol, often associated with impulsive, suicidal, and/or aggres-sive behaviors. These observations were fully consistent at the time with thefindings that neurochemical lesions of the ascending serotonergic fibers werereported to facilitate aggressive behavior in rodents and with the subsequentdiscovery that tricyclic antidepressants enhanced serotonergic neurotransmis-sion in rodents. Some researchers were then prompt to close the circle andsuggest that these medications were effective in preventing suicide and attenu-ating aggressivity–impulsivity through their purported ability to correct for aneurochemical imbalance. This model of a deficit in serotonergic neurotrans-mission being primary in the causation of impulsivity and/or aggressivity, andpredictive of therapeutic response to drugs shown to enhance serotonergicneurotransmission, is now obsolete. For example, not all impulsive–aggressivepatients show these abnormalities and benefit from drugs enhancing seroton-ergic neurotransmission. Further, several drugs devoid of major effects onserotonergic neurotransmission are known to be effective in controllingaggressive behavior/impulsivity. The categorical view has gradually been re-placed by a dimensional approach, that is, that low 5-HT explains in part thevulnerablity to impulsivity across various disorders. Hence, the thinking aboutthis system has progressively evolved, which is the case for all systems de-scribed in this chapter, from an absolute deficit to a high-risk model. In otherwords, low serotonin neurotransmission is now thought to operate as a bio-logical risk factor, resulting from genetic inheritance and/or early environmen-
Neuromodulators in Aggression 263

tal insult, neither sufficient nor necessary, yet when combined with othersalient genetic factors and/or environmental stressors, may play an importantrole in the triggering and maintenance of disorders of impulse control and ag-gressive behavior(s).
Most indices suggest that 5-HT metabolism is a relatively stable trait.Over time (months to years), CSF concentrations of 5-HIAA are highly corre-lated in studies in both human and nonhuman primates. This stability of CSF5-HIAA concentrations likely reflects genetic contributions. Evidence for theexistence of a low 5-HT neurotransmission trait is manifold; 5-HT1B receptorknockout mice are more aggressive (Hen, 1996); aggressive–impulsive pri-mates followed longitudinally in their own ecosystem for years have stablelow CSF 5-HIAA over time (Higley et al., 1996), and low CSF 5-HIAA hasalso been found in patients who reattempted suicide within 5 years as com-pared with those who did not (Roy, Pickar, De Jong, & Karoum, 1989). Fur-ther, it has been reported that newborns from families with antisocial person-ality disorder show lower CSF 5-HIAA concentrations than those fromfamilies devoid of a history of antisocial behavior (Constantino, Morris, &Murphy, 1997), and low central 5-HT neurotransmission in probands hasbeen associated with an increased risk for impulsive aggression in their first-degree relatives (Coccaro, Kavoussi, Hauger, Cooper, & Ferris, 1998). Wehave shown that subjects at genetic risk for mood and/or impulse controldisorders are more vulnerable to the effects of transient lowering of seroton-ergic neurotransmission by means of dietary manipulations than age- and sex-matched controls devoid of family history for depression, substance abuse,and/or violent behavior (Benkelfat, Le Marquand, & Pihl, 1994; LeMarquand,Benkelfat, Pihl, Palmour, & Young, 1999). Impulsive aggression, includingimpulsive suicidal behavior, has been shown to be associated with specificpolymorphisms of tryptophan hydroxylase (TPH) (the rate-limiting enzymefor 5-HT synthesis) and the serotonin transporter (5-HTT) (the protein re-sponsible for the neuronal reupake of 5-HT and responsible for the recyclingof up to 85% of synaptic 5-HT) (Nielsen et al., 1998). In addition, postmor-tem brain studies and in vivo imaging studies have demonstrated greater5-HT1A receptor binding in females as compared with males (Parsey et al.,2002). Accordingly, there are several reports of an inverse relationship be-tween aggression and 5-HT1A receptors (Cleare & Bond, 2000). Although theunderlying cause for this sex difference is unclear, sex steroids are known tomodulate 5-HT1A binding and it has been suggested that estradiol increases5-HT1A binding (Biegon & McEwan, 1982). Furthermore, several lines ofnonhuman evidence exist that suggest a functional difference in 5-HT1A recep-tors between males and females. For example, 5-HT transporter knockoutmice have lower 5-HT1A receptor binding, which is more extensive in femalesthan in males (Li, Wichems, Heils, Lesch, & Murphy, 2000). Finally, in thehippocamus, 5-HT and/or 5-HIAA concentrations are higher in female ratsthan in males (Haleem, Kennett, & Curzon, 1990). These serotonergic differ-
264 D E T E R M I N A N T S O F A G G R E S S I O N

ences may help to explain the higher incidence of mood disorders and depres-sion among females, being twice that of males, and the lower reported overallincidence of overt female aggression.
The concept of a low 5-HT trait being a predisposing factor for abnormalbehavior has generated considerable interest and lots of supporting indirectevidence. Yet this concept is still much of a working hypothesis. Most of thefindings establishing a link between low 5-HT neurotransmission and impuls-ivity and/or aggressive behavior were obtained in psychiatric patients, col-lected from nonepidemiological samples, studied as adults typically once, andcompared with controls. In general, no information could be inferred fromthis type of design as to whether the purported abnormalities are trait- orstate-related. Moreover, studies of clinical samples, though essential, have lim-itations of their own: sample heterogeneity, small sample size, not representa-tive, retrospective, confounded by uncontrolled random environmental events(i.e., drugs), and so on. Further, there is no consensus of the operationaldefinition of the corresponding behavioral phenotype for the proposed traitwith the clinical syndrome complex, combining symptoms of mood liability/explosiveness-irritability, increased impulsivity, quarrelsomeness, a predisposi-tion for binge eating and substance abuse, with unclear boundaries, and withcharacter cluster B pathology. More germane is the consideration of involve-ment and interaction of 5-HT with other neurotransmitters.
Dopamine
The relationship between 5-HT and dopamine (DA) is particularly relevant. Ina study in which microdialysis probes were used to measure these neurotrans-mitters in the nucleus accumbens (NAC) and medial prefrontal cortex(MPFC) in rats during intruder-precipitated aggressive behavior, it was foundthat DA levels increased significantly in the NAC and 5-HT decreased 80% inthe prefrontal cortex (PFC; Van Erp & Miczek, 2000). Increased DA levels inthe NAC have also been linked to many motivational and motor functions,including drug taking, food-reinforced behavior, sexual behavior, foot shock,social defeat, and attention to novel environmental stimuli. This range of be-haviors reflects the fact that DA ascending pathways from this area project toand innervate more than 20 different structures, thus reflecting general ratherthan specific functions, and, in effect, neuromodulating various areas of incen-tive motivation (Le Moal & Simon, 1991). In particular, this system respondsto cues of incentive reward, including novelty, receiving differential afferentsfrom many sources, including the basolateral amygdala, the extended amyg-dala, the hippocampus, and the prefrontal medial orbital cortex.
DA’s activity is related to its activation of two families of DA receptors:the D1 and D2 subtypes. Both are linked to G-protein activation, thus their sig-nals are generally slow and long lasting rather than fast (Nieoullon, 2002). D1receptors appear to underlie DA’s tonic activity, whereas D2 receptors underlie
Neuromodulators in Aggression 265

DA’s phasic activity. A tonic response implies that the neurons fire continu-ously throughout the duration of a stimulus, which appears to relate to DA’sability to increase the stability of maintained representations. Stabilization in-volves increasing the signal-to-noise ratio of background versus evoked activ-ity in the prefrontal cortex (Cohen, Braver, & Brown, 2002) D1 receptors ap-pear to interact with other receptors, especially fast-signaling ones such asglutamate (Nieoullon, 2002), and have been implicated in the working mem-ory functions of the dorsolateral PFC.
Conversely, a phasic response implies that the neurons fire briefly at aparticular phase of the stimulus (usually the onset) and likely relates to DA’srole as a gating signal. The DA system is responsible for indicating when newinputs should be encoded and maintained, or when currently maintained rep-resentations should be updated in response to salient, reward-predicting infor-mation. DA’s phasic response appears to involve activation of D2 receptors(Cohen et al., 2002). These receptors have been implicated in the reward cir-cuits of the brain (Blum, Cull, Braverman, & Comings, 1996), as well as inthe PFC’s regulation of the subcortical DA system.
The subcortical DA system is involved in signaling reward-predicting in-formation and, as such, is involved in motivational aspects of behavior. Thus,increases in DA transmission lead to more information being allowed to enterthe “motive circuit” (ventral tegmental area–accumbens–ventral pallidum), sothat this information becomes more motivationally relevant (Spear, 2000). Inseeking out reward-related information, the DA system helps the organism toadapt to its present situation by facilitating the initiation of new responses andswitching between alternate responses (Spoont, 1992). Serotonin and GABAappear to modulate DA’s reward-related activity. Blum et al. (1996) have pro-posed a “cascade theory” of the reward circuitry of the brain, implicatingdopamine, serotonin, and GABA. In particular, they propose that the releaseof serotonin in the hypothalamus causes a release of enkephalin (an opioidpeptide) in the ventral tegmental area, which, in turn, inhibits GABAergic neu-rons in the substantia nigra. This disinhibits DA neurons in the ventraltegmental area, allowing them to release DA in the nucleus accumbens andhippocampus. The released DA then activates D2 receptors in these regions,producing rewarding effects. Deficits in this reward pathway lead to whatBlum et al. (1996) refer to as the “reward deficiency syndrome,” characterizedby addictive, impulsive, and compulsive behavior. Thus, hypoactivity of thelimbic DA system may be related to reduced levels of 5-HT or increased levelsof GABA, whereas hyperactivity of this system should be associated with in-creased levels of 5-HT and decreased levels of GABA. As stated previously, re-duced 5-HT has been associated with disorders of impulsivity, addiction, andaggression. Furthermore, GABA agonists have been suggested as a possibletreatment for drug addiction, and aggression (Eichelman, 1990). In particular,GABA appears to dampen DA neurons via inhibitory GABAB receptors. Stim-ulation of GABAB receptors in the ventral tegmental area has been shown todampen DA release in the nucleus accumbens produced by abused drugs
266 D E T E R M I N A N T S O F A G G R E S S I O N

(Cousins, Roberts, & De Wit, 2002), thereby reducing the rewarding effectsof these drugs.
Sex differences also exist in the regulation of dopaminergic neurotrans-mission. There is substantial evidence, primarily from nonhumans, that go-nadal hormones modulate behavioral and neurochemical indices of activity inthe striatum and nucleus accumbens. For example, female rats have higherstriatal dopamine and homovanillic acid (HVA) levels than males, and agingreduces DA and HVA in males but not in females. Endogenous ovarian hor-mones, but not testicular hormones, modulate extracellular striatal DA con-centrations in rats (Xiao & Becker, 1994). These differences may be related tothe observations that female rats appear to be more sensitive than males to thetoxic and reinforcing effects of psychostimulants that increase the DA system,an idea that is supported by research on gender differences in the behavioralresponse to cocaine, in which males and females do not differ in cocaine me-tabolism but female rats exhibit greater locomotor activation in response tococaine than do male rats (Bowman et al., 1999).
A contrasting view of serotonin’s interaction with dopamine has alsobeen proposed. Spoont (1992) has suggested that 5-HT constrains DA’sreward-related activity, such that the organism’s behavioral output becomesmore controlled. Thus, 5-HT’s role would be to increase the threshold bywhich exogenous stimuli become motivationally relevant, by decreasing thesignal-to-noise ratio. Indeed, 5-HT has been shown to decrease the signal-to-noise ratio by inhibiting spontaneous cell activity to a lesser extent than spe-cific evoked responses (Tassin, 1992). This would suggest that, through 5-HT’s activity, a greater evoked response would be required in order for a stim-ulus to cause the cell to fire. This would ensure that only signals of sufficientstrength or of sufficient relevance to the organism are able to interrupt currentinformation flow and would prevent overshoots in the system by constrainingthe intensity of signals once they gain access to the system (overshoots in thesystem are associated with an increased magnitude of response and decreasedsensitivity to cues of suppression). Supporting evidence for this view is drawnfrom studies showing decreases in 5-HT associated with increased locomotoractivity and increased exploratory behavior signifying a release of normallyinihibited operant behavior in diverse contexts, as well as increased food in-take, sexual behavior, and aggression. Furthermore, behavioral evidenceshowing serotonergic modulation of the startle reflex is cited, with 5-HT de-creasing the signal-to-noise ratio, thereby protecting the animal from interfer-ence from nonsalient environmental cues. Spoont (1992) further argues thatDA normally functions to initiate new responses as well as to switch betweenalternative responses (a role that would appear to be related to DA’s reward-signaling activity), which 5-HT constrains by making it more difficult forsignals to gain access to the system. A similar pathway could be postulated toexplain the effects of decreased serotonin on operant behavior, food intake,sexual behavior, and aggression. Furthermore, in considering the evidence that5-HT normally constrains operant behavior and protects the organism from
Neuromodulators in Aggression 267

interference from nonsalient environmental cues, as well as evidence that de-creasing 5-HT increases the propensity to switch between alternative re-sponses while decreasing the integrity of those responses, one could argue thatthese are indicators of 5-HT’s effect on PFC function. As the section“Execu-tive Control” discusses further, 5-HT appears to enhance PFC function, al-though the effect may depend on the particular characteristics of the situationin which behavior is assessed.
Norepinephrine
The noradrenergic system serves many different functions, all of which areknown to affect behavior profoundly. One comprehensive review argued thatnoradrenaline (NA) centrally and indirectly affects aggression on three differ-ent levels: the hormonal system, the sympathetic nervous system, and the cen-tral nervous system, in different, but functionally synergistic ways (Haller,Makara, & Kruk, 1998). All three components react very rapidly to environ-mental changes, catecholamine hormones being amongst the fastest to react.Behavior, of course, may involve a very rapid response to environmental chal-lenges and engages the whole organism. Behavior, including aggressive behav-ior, is also energy dependent and therefore requires the rapid mobilization ofenergy stores. The peripheral catecholamines are important in ensuring the“energy” of behavior, whereas central noradrenaline is activated by offensiveand defensive aggression in males (Higley et al., 1992). As such, external stim-uli anticipating a demanding behavior or the demanding behavior itself acti-vates the central nervous system noradrenergic system. Thus, even if NAwould have no effect on mechanisms directly involved in aggression, it wouldnonetheless have profound influence on the expression of aggressive behaviorthrough its indirect effects on other mechanisms vital during social challengessuch as arousal and attention, pain sensitivity, olfaction, and memory.
The NA neurotransmitter system has long been implicated as a modula-tor of normal arousal, vigilance, and engagement with the environment. It hasalso been shown that agents that increase brain noradrenaline improve atten-tion in rats (Sirvio et al., 1993). Furthermore, pain sensitivity, beyondopiodergic control, is decreased by medullar descending noradrenergic path-ways, and changing pain perception has an obvious function in aggressive be-havior. The NAergic system is also involved in memory/recognition processesand in learning, mainly when olfaction is also involved. In summary, varioussocially relevant stimuli elicit a central release of norepinephine, which in turnpromotes arousal and shifts attention toward relevant stimuli, induces analge-sia, and enhances olfaction, learning, and memory. Such specific changes maytogether significantly change an animal’s reaction to social challenges.
In general, studies have demonstrated an increase in aggressive behaviorwhen norepinephine (NE) activity surges, and a decrease in aggression whenNE activity is decreased. Increased NE function is positively correlated withthe number of shock-induced aggressive episodes in rodents (Stolk, Connor,
268 D E T E R M I N A N T S O F A G G R E S S I O N

Levine, & Barchas, 1974), and pharmacological agents that increase NE func-tion also increase shock-induced fighting in rodents. Mice that have been iso-lated for prolonged periods of time show increased aggressive behavior whengiven desipramine (which increases NE), an effect that is not observed whenthey are pretreated with a selective NE neurotoxin (Matsumoto, Ojima, &Watanabe, 1995). Stimulating postsynaptic noradrenergic receptors in cats’hypothalami facilitates aggression; in humans, clinical treatment with tricyclic/monoamine oxidase inhibitor antidepressants (which increase NE function) isassociated with agitation and irritability, particularly in patients with border-line personality disorder. Thus, NE seems to increase/facilitate aggression.However, there are also contradictory studies. For example, positive correla-tions have been reported between the CSF concentration of NE’s majormetabolite, 3-methoxy-4-hydroxyphenylglycol (MHPG), and life history ofaggressive events in men confined to a military forensic unit (Brown, Good-win, Ballenger, Goyer, & Major, 1979), and to impulsivity ratings in humans(Roy et al., 1989). Yet Virkkunen, Nuutila, Goodwin, and Linnoila (1987)have found lower CSF MHPG in violent offenders and in impulsive arsonistsas compared with healthy volunteers, and a study by Coccaro et al. (1998) re-ported a negative correlation between plasma MHPG and life history of ag-gression in personality-disordered patients.
The most extensive review on the relationship between norepinephrineand aggression offers an explanation for this discrepancy in results by suggest-ing that NA’s contradictory effects on aggressive behavior depends on whatkind of aggressive paradigm is used (Haller et al., 1998). According to Hallerand colleagues (1998), as far as isolation-induced or social (rank associated)aggression is concerned, a reduction in NA is followed by a parallel inhibitionof aggressive behavior, but NA depletion appears to increase shock-inducedand chemically elicited aggression. In these latter instances, the increase in ag-gression after NA depletion may be caused by parallel changes in other neuro-transmitter systems, such as dopamine and/or serotonin and/or alterations inreactivity to environmental challenges. Haller and colleagues (1998) also con-clude that when noradrenaline is in excess (i.e., when release is stimulated orreuptake is blocked), it influences aggression biphasically, such that a slightactivation increases aggression whereas a strong activation decreases it.
GABA
Similarly elusive is the role of gamma-aminobutyric acid (GABA) in aggres-sion. It is well known that GABA serves as an inhibitory neurotransmitter atmany central nervous system synapses. Yet increased GABAA receptor activa-tion by alcohol suggests that GABA transmission can facilitate aggression(Miczek, De Bold, van Erp, & Tomatzky, 1997). In addition, one study sur-prisingly showed that when GABA was injected into medial hypothalamic de-fensive rage sites in cats, response thresholds were actually lowered followingdrug infusion (Nakao, Tashiro, Kono, & Araki, 1979). However, interven-
Neuromodulators in Aggression 269

tions that increase GABA receptor activation nonselectively do inhibit aggres-sion. For example, microinjections of the GABA agonist muscimol into defen-sive rage sites within the dorsal periaqueductal gray matter inhibits aggression(Siegel, Roeling, Gregg, & Kruk, 1999). Similarly, aggressive mice have lowerbrain GABA concentrations as compared with nonaggressive mice, and agentsthat inhibit GABA metabolism dose-dependently decrease fighting in conjunc-tion with increases in brain GABA (Simler, Puglisi-Allegra, & Mandel, 1982).Furthermore, both clonidine and lithium, which have anti-aggressive behav-ioral effects in some patients, have reportedly increased peripheral measuresof GABA (Kemph, De Vane, Levin, Jarecke, & Miller, 1993). A study by Bjorket al. (2001) found that plasma GABA levels were lower in first-degree rela-tives of patients with major depressive disorder—who are also shown to havelow plasma GABA—as compared with controls, thus indicating that plasmaGABA is under genetic control. This study went on to demonstrate thatplasma GABA levels and self-reported aggressiveness correlated negatively,but that this was more specific to subjects with a family history of depression.
DEVELOPMENT
At each of the developmental stages, from genes to old age, the functioningand density of these neurotransmitters critical to behavioral control are opera-tive. For example, regarding dopamine, a recent review (Di Maio, Grizenko,& Joober, 2003) has detailed the involvement of dopamine genes in attention-deficit/hyperactivity disorder (ADHD). These authors marshal considerableevidence to support the strong involvement of the dopamine transporter gene(SLC3A6) and the dopamine receptor 4 gene (DRD4) in ADHD. A mousewith the dopamine transporter gene knockout has been developed, with theresultant animals displaying spontaneously increased activity and higher reac-tions to stress, which are significantly calmed by the administration of stimu-lants. This review further details how extra repeat copies of this gene havebeen reported in diagnosed ADHD populations. A similar repeat polymor-phism has been reported for the alleles of the DRD4 gene, with particular fo-cus on the 7 repeat allele. In a recent meta-analysis (Faraone, Doyle, Mick, &Biederman, 2001) it was reported that an odds ratio of 1.9 existed betweenthe repeat allele and ADHD for family-based studies, suggesting a small butsignificantly increased risk. Another study of DA receptor density in 16 micestrains found threefold differences for the nucleus accumbens and a sevenfolddifference among strains for the caudate-putamen between strains, demon-strating remarkable genetic variability (Jones, Hou, Cook, & Melloni, 1996).
Excess dopamine prenatally, such as occurs when a mother is cocaine de-pendent, likely results in abnormalities, which studies suggest affect the differ-entiation of neurotransmitter systems, cell function, migration, and neuronalgrowth. Cocaine levels in the fetal brains of rats have been found to be greaterthan the 100% of those in the maternal blood. As shown in longitudinal stud-
270 D E T E R M I N A N T S O F A G G R E S S I O N

ies, infants born to cocaine-using mothers typically display poor behavior reg-ulation, habituation, and orientation (Chasonoff et al., 1998). At age 6 this isseen in difficulty in managing impulses, frustration, and arousal. Studies ofmonkeys have shown that this prenatal exposure results in reduced expressionof the rate-controlling enzyme in dopamine synthesis, as well as an elevationin dopamine receptors, particularly in frontal brain areas (Ronnekleiv, Fang,Choi, & Chai, 1998).
Similar behavioral profiles to that produced by prenatal cocaine insulthave been reported for many other drugs, with alcohol predominating. Prena-tal alcohol exposure affects the functioning of many neurotransmitters andneuromodulators. Recent work focusing on serotonin has shown that alcoholfed to pregnant mice resulted in a 20–30% reduction in serotonin neurons(Zhou, Sari, Zhang, Goodlett, & Li, 2001) and a decrease in the density of se-rotonin fibers, particularly in the forebrains of the offspring (Sari, Powrozek,& Zhou, 2001). These effects likely permanently decrease serotonergic func-tioning in the mature animal, thus affecting behavioral regulation.
The effect of prenatal malnutrition on the developing brain is another ex-ample of altered neurochemical function that has been extensively reviewed(Galler & Tonkiss, 1998). Depending on the time, type, and severity of malnu-trition, the basic cytoarchitectine of the brain can be affected. Protein deficien-cies in particular are significant, as proteins are precursors of enzymes, peptidehormones, and neurotransmitters. Morgane et al. (1993) have demonstratedin rats long-term inhibition in certain hippocampal GABA interneuron sys-tems following prenatal protein malnutrition, which affects a myriad of othersystems and is possibly responsible for many attentional and learning prob-lems.
In terms of neonates, rearing rats in isolation produces a behavioral syn-drome that includes hyperactivity and an increased response to reinforcers, re-sponse perseveration, decreased pain thresholds, and a heightened response todopamine agonists. Further, basal levels of dopamine are higher, and a downregulation of D2 receptors in the nucleus accumbens is affected (Hall et al.,1998). In the turbulent years of adolescence there is a concomitant shift indopaminergic activity from subcortical to cortical areas. As one further ages,so does neurotransmitter functioning. In a PET study of individuals ages 24–86 correlations were conducted between the density of D2 receptors andneuropsychological test performance. It was found that D2 receptor availabil-ity in the caudate and putamen declined with age and was correlated withdecreased performance on a motor task, a measure of abstraction, and aresponse inhibition task (Volkow et al., 1998).
As the preceding examples illustrate, throughout childhood, adolescence,and even into old age, changes occur in the circuitry of the brain that likelyaccount for age-related cognitive and behavioral changes. For example, in ad-olescence there is a massive loss of synapses in the neocortex, with the mostintense period of pruning occurring between 7 and 16 years of age. Althoughthis pruning occurs for many different kinds of receptors, the majority of syn-
Neuromodulators in Aggression 271

apses lost are those that are excitatory in nature. Such pruning occursthroughout the brain, including the PFC and limbic regions. During adoles-cence there is also an increase in DA fiber density in the PFC, which may becompensated for later in adolescence by a decline in DA synthesis and/or turn-over in this region. Developmental changes also occur in the GABAergic sys-tem. Studies in rats have shown increased responsiveness of the GABAB sys-tem and of the GABA/benzodiazepine receptor complex from adolescence toadulthood, but a decrease in responsiveness of the cortical GABAA system.Based on studies of knockout mice, it has been suggested that dysfunctionalGABAA receptors lead to increased emotional reactivity (Mohler, Fritschy,Benke, Benson, & Luscher, 1996). In light of GABAB’s previously mentionedinvolvement in the reward circuit of the brain (in particular, dampening DArelease produced by drugs), it could be inferred that this gradual increase inresponsiveness of the GABAB system may reduce one’s susceptibility to the re-warding effects of certain drugs. Thus, it may be postulated that from child-hood into adulthood, there is a decrease in the responsiveness to cues of re-ward. With respect to the serotonergic system, 5-HT binding appears to belower during adolescence than during childhood or adulthood (Spear, 2000).Considering the myriad studies linking low serotonin levels to disorders ofbehavioral control and impulsivity, this lowered serotonin binding duringadolescence may account for the impulsive behavior often seen at this devel-opmental period.
As concerns the relative balance between subcortical and cortical DA sys-tems, there appears to be a shift toward greater predominance of cortical DAduring early adolescence. With a peak in cortical DA activity at this time, aconcomitant loss in excitatory drive to the cortex may result. Both rat andhuman studies have found a decrease of one-third to one-half in dopamine re-ceptors in the striatum during adolescence (Seeman, Bzowej, Guan, &Bergeron, 1987). Spear (2000) has postulated that this adolescent-associatedloss in excitatory drive to the cortex may create a mini-“reward–deficiencysyndrome.” Thus, too little DA stimulation caused by overactivity of the PFCmay lead to a state of underarousal. It is further thought that this under-arousal may account for the increase in impulsive behavior often seen duringadolescence. In addition, these mesocortical DA projections are seen as moresensitive to activation by stressors than are mesolimbic DA systems. Con-sequently, stress may exacerbate this adolescent-associated mini-reward–deficiency syndrome by increasing the extent to which cortical DA inhibitsexcitatory drive to the cortex.
Adolescence is also a time when there is a shift toward greater efficiencyof processing information. MRI studies have shown an inverse relationshipbetween cortical grey matter loss and brain growth in the frontal cortex of ad-olescents, suggesting that the synaptic losses may be concomitant with in-creased myelination. Thus, maturation of the frontal cortex seems to involve ashift toward fewer and faster connections, a change that presumably increasesthe speed and efficiency of processing. These changes involve use-dependent
272 D E T E R M I N A N T S O F A G G R E S S I O N

patterns of connectivity, implying that experience plays an important role inthe development of the PFC.
EXECUTIVE CONTROL
Russian neuropsychologist Luria (1980) conceptualized the operation of thePFC more wholistically and realistically than the current focus on specificfunctions and their localization. Conceptually, he saw working memory,attentional control, verbal organization, and so forth, operating collectively toascribe meaning to stimuli, in effect creating the context for possible re-sponses. In this view, the frontal lobes are seen as responsible for planning,controlling, and verifying behavior in the presence of goals, working within acontext and providing control over the more automatic subcortical systems.As reviewed earlier, the major neurotransmitters are integral to the develop-ment and functioning of the PFC.
Dopamine is highly concentrated in the prefrontal cortex, in fact, moreconcentrated than in any other cortical region. One study showed that as in-fant monkeys improved their performance on a delayed response task requir-ing inhibition, the level of dopamine in the prefrontal cortex increased, as didthe density of dopamine receptors (Brown & Goldman, 1977). Further,lesioning of the dorsolateral prefrontal cortex interferes greatly with perfor-mance on delayed response tasks, as does blocking dopamine receptors inthese areas. On these tasks delay of responding is critical, which is why thisarea has been related to sustained attention and working memory. What isalso noteworthy is that the pattern of responses in lesioned and pharmacologi-cally manipulated animals is not random. Specifically, what occurs is aperseveration of the previous response. On tasks requiring memory with inhi-bition, little is known about dopaminergic functioning in children between theages of 3 and 6. However, Diamond (2000) has been studying children whohave been treated early and continuously for phenylketonuria, and who thushave reduced levels of dopamine in the prefrontal cortex, but have otherwisenormal brains. These children typically reflect cognitive deficits and score in asimilar IQ range to children with prefrontal damage (Diamond, 2000).Treated phenylketonuric children have a reduced level of the dopamine pre-cursor tyrosine in the brain, which is particularly significant for the prefrontalcortex because it has a higher rate of dopamine turnover than any other brainarea. The nature of the deficits in these treated children involve deficits inworking memory and in inhibitory control. These deficits are seen even whenIQ, gender, and other variables are controlled and are evident in all agegroups: infants, toddlers, and young children. It is also notable that it is specif-ically deficits in working memory and inhibitory tasks, rather than other cog-nitive measures, that reflect the determined deficiencies.
Cognitive and behavioral inhibition have long been linked to DA activ-ity in the dorsolateral and ventrolateral PFC. Braver et al. (2001) have sug-
Neuromodulators in Aggression 273

gested that one of the key components in the ability to exert control overthoughts and actions is context processing, using subsets of representationswithin working memory that govern how other representations are used.Thus, in order to understand behavioral control and aggression, it is helpfulto examine the mechanisms underlying working memory. Within the PFC,DA enhances working memory by modulating excitatory neurotransmission.D1 receptors, being colocalized with glutamate receptors on the spines ofpyramidal neurons in the PFC, appear to enhance working memory by in-hibiting excitatory input by activating the neuron. Thus, too little DA maylead to cognitive dysfunction through excessive stimulation, whereas toomuch DA may lead to deficits through insufficient stimulation (Goldman-Rakic, 1999). This would be consistent with the theory of schizophrenia,which postulates that hypodopaminergic activity in the frontal cortex (alongwith hyperdopaminergic activity in the limbic system) leads to overstimu-lation and disorganized thought. This is also consistent with Arnsten’s(2000) finding that, in the PFC, DA has an inverted U-shaped dose–responsecurve, whereby either very high or very low levels of D1 stimulation impaircognitive function.
The activity of the subcortical DA system appears to be partially underthe control of the PFC. Deutch (1992) has proposed that DA tonically inhibitscorticostriatal projection neurons, such that glutamate’s release in the striatalcomplex, in turn, inhibits DA from being released in the subcortical site. Thishypothesis implies that diminishing the activity of DA in the PFC results in in-creased subcortical DA being released. Indeed, levels of DA activity in the PFCappear to be inversely related to DA release in subcortical regions (Spear,2000). For example, it has been proposed that in children with ADHD, re-duced activity in the PFC may lead to low tonic DA activity in limbic regions.This low tonic stimulation of DA autoreceptors may lead to high phasic activ-ity in the nucleus accumbens and other subcortical sites. Thus, reduced PFCactivity leads to low tonic DA activity, which leads to hyperresponsiveness ofthe DA system to reward-related cues. This model would account for the ben-eficial effects of stimulants in treating ADHD.
DA’s inhibitory action may be mediated through direct activation of D2receptors, or may occur indirectly by facilitating GABA release from inter-neurons. D2 receptors have been implicated in a number of addictive, impul-sive, and compulsive disorders (Blum et al., 1996), and drugs that enhanceGABAergic function have been used in clinical settings to control aggressivebehavior. GABA and 5-HT have also been implicated in working memory inthe PFC. GABAergic interneurons stabilize representations by inhibiting py-ramidal neurons with opposite “best directions” (Goldman-Rakic, 1999).Thus, by inhibiting contradictory signals, the GABAergic system providesfeedforward inhibition, thereby stabilizing maintained representations andpreventing distractors from disturbing the system. In fact, postmortem andother biological studies have supported the suggestion of a defective GABAergic
274 D E T E R M I N A N T S O F A G G R E S S I O N

system in the frontal cortex of patients with schizophrenia (Blum & Mann,2002). With respect to the 5-HT system, Williams, Srinivas, & Goldman-Rakic, 2002) have found that in rhesus monkeys, 5-HT enhanced PFCfunction via activation of the 5-HT2A receptor, which facilitates both pyrami-dal cells and interneurons. In particular, direct facilitation of pyramidal cells,as well as indirect feedforward inhibition of pyramidal cells via GABAinterneurons, provides tonic facilitation of the neuronal network (increasedsignal-to-noise ratio). However, in light of behavioral findings linking 5-HT todeficits in PFC function (Sasaki-Adams & Kelley, 2001), 5-HT’s facilitory ef-fects may apply to only very constrained situations. Thus, 5-HT, by increasingthe signal-to-noise ratio, may improve PFC function when the organism ishighly motivated and when only task-relevant stimuli are present in theenvironment. However, in more realistic situations where there are many pos-sible behaviors in response to a wide variety of stimuli, 5-HT’s facilitory effectmay impair PFC function by increasing the amount of “noise” entering thesystem.
NE has also been implicated in the functioning of the PFC. It appears toexert its influence through activation of alpha-1, alpha-2, and beta-noradrenergicreceptors, all of which have been located in the PFC, and may also act throughD4 receptors. Studies with rats, humans, and primates have indicated oppos-ing effects of alpha-1 and alpha-2 receptor stimulation. Activation of alpha-2Asubtype appears to enhance PFC functioning, as indexed by improvements inworking memory tasks, response inhibition, and planning (Arnsten, 2000).Conversely, activation of alpha-1 receptors appears to impair performance ontasks subserved by the PFC. Furthermore, there appears to be an optimal levelof NE, as NE exhibits higher affinity for alpha-2 than for alpha-1 adreno-ceptors. Thus, moderate levels of NE (for example, as induced by stress)engage alpha-1 receptors and impair PFC function (Arnsten, 2000). Further-more, there are NE–DA interactions, for example, cortical alpha-1 adrenergicand D1 receptors may be located on the same cells, and “the message inducedby the stimulation of cortical D1 receptors is inhibited by alpha-1 adrenergicreceptors” (Tassin, 1992, p. 145). NE’s functioning in the PFC also seems tobe consistent with clinical research. Administration of beta-adrenergic recep-tor antagonists appears to attenuate aggressiveness in organically impairedchildren and adults, as well as in schizophrenic patients prone to violent out-bursts. Moreover, lithium (which describes the functional availability of NE)reduces aggressiveness in personality-disordered adults and conduct-disor-dered children. Furthermore, one study found a positive correlation betweenCSF levels of the NE metabolite (MHPG, 3-methoxy-4-hydroxyphenylglycol)and a life history of aggressive behavior (Eichelman, 1990). Thus, in individu-als with unusually high levels of NE, PFC function may be impaired by in-creased activation of alpha-1 adrenoceptors, which may lead to impaired be-havioral control. In these populations, decreasing available NE seems toreduce aggressiveness, perhaps by reducing activation of alpha-1 receptors.
Neuromodulators in Aggression 275

CONCLUSION
The development of behavioral regulation is essential to the control ofimpulsivity and most forms of aggressive behavior. This development requiresthe normative functioning of a wide range of neurochemical activities thataffect and are affected by genetic variation and by pre- and postnatal experi-ence. These interactions shape the brain and its functioning in that orches-trated dance between brain and environment. Although many neuromodula-tors are active participants, featured performers are serotonin, dopamine,norepinephrine, and GABA. Low serotonergic neurotransmission has been re-peatedly linked to impulsive–aggressive behaviors. The mechanisms of thelinkage, however, remain at issue, with enhanced vulnerability most likelyassociated with serotonin’s interaction with dopamine—specifically, hypoac-tivity at the limbic level, an involvement not unlike that of GABA’s. With re-duced serotonin and GABA activity at this level weak, even irrelevant signalsproduce system activation. The inhibiting potential of the prefrontal cortexcan limit this overactivation, again, through neurochemical functioning.Prefrontal serotonin levels, which appear reduced during adolescence, seemgermane specifically to situations where highly motivated and task-relevantstimuli are present. Dopamine’s role, which is initiated when limibically ac-tive, appears inhibiting when active in the prefrontal cortex. This brain areabasic to the control of behavior is a structure that develops through adoles-cence, is programmed by experience but fueled by the interaction of basicbrain neuromodulators. Finally, an unmentioned but most significant caveat isrelevant. That is, with the remarkable development of new technologies anddaily flurry of scientific study, “all of the above is subject to change.”
REFERENCES
Archer, J., & Lloyd, B. (2002). Sex and gender. London: Cambridge University Press.Arnsten, A. F. T. (2000). Genetics of childhood disorders: XVIII. ADHD, Part 2:
Norepinephrine has a critical modulatory influence on prefrontal cortical function.Journal of the American Academy of Child and Adolescent Psychiatry, 39, 1201–1203.
Benkelfat, C., LeMarquand, D., & Pihl, R. O. (1994). Serotonin and alcohol intake,abuse, and dependence: Clinical evidence. Biological Psychiatry, 1(36), 327–337.
Biegon, A., & McEwan, B. S. (1982). Modulation by estradiol of serotonin receptors inbrain. Journal of Neuroscience, 2, 199–205.
Bjork, J. M., Moeller, F. G., Kramer, G. L., Kram, M., Suris, A., Rush, A. J., & Petty, F.(2001). Plasma GABA levels correlate with aggressiveness in relatives of patientswith unipolar depressive disorder. Psychiatry Research, 101, 131–136.
Blum, B. P., & Mann, J. J. (2002). The GABAergic system in schizophrenia. InternationalJournal of Neuropsychopharmacology, 5(2), 159–179.
Blum, K., Cull, J. G., Braverman, E. R., & Comings, D. E. (1996). Reward deficiency syn-drome. American Scientist, 84, 132–145.
276 D E T E R M I N A N T S O F A G G R E S S I O N

Bowman, B., Vaughan, S., Walker, Q., Davis, S., Little, P., Scheffler, N., Thomas, B., &Kuhn, C. (1999). Effects of gender and gonadectomy on cocaine metabolism in therat. Journal of Pharmacology and Experimental Therapeutics, 297, 114–120.
Braver, T. S., Barch, D. M., Keys, B. A., Cohen, J., Taylor, S. F., Carter, C. S., Kaye, J. A.,Janowsky, J. S., Yesavage, J. A., & Mumenthaler, M. S. (2001). Context processingin older adults: Evidence for a theory relating cognitive control to neurobiology inhealthy aging. Journal of Experimental Psychology: General, 130(4), 746–763.
Brown, G. L., Goodwin, F. K., Ballenger, J. C., Goyer, P. F., & Major, L. F. (1979). Aggres-sion in humans correlates with cerebrospinal fluid amine metabolites. PsychiatryResearch, 1, 131–139.
Brown, R. M., & Goldman, P. S. (1977). Catecholamines in neocortex of rhesus mon-keys: Regional distribution and ontogenetic development. Brain Research, 168,133–150.
Chasnoff, I., Anson, A., Hatcher, R., Stenson, H., Iankea, K., & Randolph, L. (1998).Prenatal exposure to cocaine and other drugs. Annals of the New York Academy ofScience, 846, 314–328.
Cleare, A. J., & Bond, A. J. (2000). Ipsapirone challenge in aggressive men shows an in-verse correlation between 5-HT1A receptor function and aggression. Psycho-pharmacology, 148, 344–349.
Coccaro, E. F., Kavoussi, R. J., Hauger, R. L., Cooper, T. B., & Ferris, C. F. (1998).Cerebrospinal fluid vasopressin levels correlate with aggression and serotonin func-tion in personality disordered subjects. Archives of General Psychiatry, 55, 708–714.
Cohen, J. D., Braver, T. S., & Brown, J. W. (2002). Computational perspectives on dopa-mine function in prefrontal cortex. Current Opinion in Neurobiology, 12, 223–229.
Constantino, J. N., Morris, J. A., & Murphy, D. L. (1997). CSF 5-HIAA and family his-tory of antisocial personality disorder in newborns. American Journal of Psychiatry,154, 1771–1773.
Cousins, M. S., Roberts, D. C. S., & De Wit, H. (2002). GABAB receptor agonists for thetreatment of drug addiction: A review of recent findings. Drug and Alcohol De-pendence, 65(3), 209–220.
Deutch, A. Y. (1992). The regulation of subcortical dopamine systems by the prefrontalcortex: Interactions of central dopamine systems and the pathogenesis of schizo-phrenia. Journal of Neural Transmission, 36, 61–89.
Diamond, A. (2000). Close interrelation of motor development and cognitive develop-ment and of the cerebellum and prefrontal cortex. Child Development, 71, 44–56.
Di Maio, S., Grizenko, N., & Joober, R. (2003). Dopamine genes and attention-deficithyperactivity disorder: A review. Journal of Psychiatry and Neuroscience, 28, 27–38.
Eichelman, B. S. (1990). Neurochemical and psychopharmacologic aspects of aggressivebehavior. Annual Review of Medicine, 41, 149–158.
Faraone, S. V., Doyle, A. E., Mick, E., & Biederman, J. (2001). Meta-analysis of the asso-ciation between the 7–repeat allele of the dopamine D-sub-4 receptor gene and at-tention deficit hyperactivity disorder. American Journal of Psychiatry, 158, 1052–1057.
Galler, J. R., & Tonkiss, J. (1998). The effects of prenatal protein malnutrition and co-caine on the development of the rat. Annals of the New York Academy of Sciences,846, 29–39.
Neuromodulators in Aggression 277

Goldman-Rakic, P. S. (1999). The “psychic” neuron of the cerebral cortex. Annals of theNew York Academy of Sciences, 868, 13–26.
Haleem, D. J., Kennett, G. A., & Curzon, G. (1990). Hippocampal 5–hydroxytryptaminesynthesis is greater in female rats than in males and more decreased by the 5-HT1Aagonist 8-OH-DPAT. Journal of Neural Transmission, 79, 93–101.
Hall, F., Wilkinson, L., Humby, T., Inglis, W., Kendall, D., Marsden, C., & Robbins, T.(1998). Isolation rearing in rats: Pre- and postsynaptic changes in striatal dopamin-ergic systems. Pharmacology, Biochemistry and Behavior, 59, 859–872.
Haller, J., Makara, G., & Kruk, M. (1998). Catecholaminergic involvement in the con-trol of aggression: Hormones, the peripheral sympathetic, and central noradren-ergic systems. Neuroscience and Biobehavioral Reviews, 22(1), 85–97.
Helmers, K., Young, S., & Pihl, R. O. (1995). Assessment of measures of impulsivity inhealthy male volunteers. Personality and Individual Differences, 19, 927–935.
Hen, R. (1996). Mean Genes, Neuron, 16, 17–21.Higley, J., Mehlman, P., Taub, D., Higley, S., Suomi, S., & Linnoila, M. (1996). Stability
of interindividual differences in serotonin function and its relationship to severe ag-gression and competent social behavior in rhesus macaque females. Neurospsycho-pharmacology, 7, 67–76.
Higley, J., Mehlman, P., Taub, D., Higley, S., Suomi, S., Vickers, J., & Linnoila, M.(1992). Cerebrospinal fluid monoamine and adrenal correlates of aggression infree-ranging rhesus monkeys. Archives of General Psychiatry, 49, 436–441.
Hoaken, P. N. S., & Pihl, R. O. (2000). The effects of alcohol intoxication on aggressiveresponses in men and women. Alcohol and Alcoholism, 35, 471–477.
Jones, B., Hou, X., Cook, M., & Melloni, N. (1996). Effect of exposure to novelty onbrain monoamines in C57BL/6 and DBA/2 mice. Physiology and Behavior, 59,361–367.
Kemph, J. P., DeVane, C. L., Levin, G. M., Jarecke, R., & Miller, R. L. (1993). Treatmentof aggressive children with clonidine: Results of an open pilot study. Journal of theAmerican Academy of Child and Adolescent Psychiatry, 32, 577–581.
LeMarquand, D., Benkelfat, C., Pihl, R., Palmour. R., & Young, S. (1999). Behavioraldisinhibition induced by tryptophan depletion in nonalcoholic young men withmultigenerational family histories of paternal alcoholism. American Journal of Psy-chiatry, 15, 1771–1779.
Le Moal, M., & Simon, H. (199l). Mesocorticolimbic dopaminergic network: Functionaland regulatory roles. Physiological Review, 71, 155–234.
Li, Q., Wichems, C., Heils, A., Lesch, K. P., & Murphy, D. L. (2000). Reduction in thedensity and expression, but not G-protein coupling, of serotonin receptors (5-HT1A) in 5-HT transporter knock-out mice: Gender and brain region differences.Journal of Neuroscience, 20, 7888–7895.
Luria, A. R. (1980). Higher cortical functions in man. Moscow: Moscow UniversityPress.
Matsumoto, K., Ojima, K., & Watanabe, H. (1995). Noradrenergic denervation attenu-ates desipramine enhancement of aggressive behavior in isolated mice. Pharmacol-ogy, Biochemistry and Behavior, 50, 481–484.
McCarthy, M., Auger, A., & Perrot-Sinal, T. (2002). Getting excited about GABA andsex differences in the brain. Trends in Neuroscience, 25, 307–312.
Miczek, K. A., DeBold, J. F., Van Erp, A. M., & Tornatzky, W. (1997). Alcohol, GABA-a-benzodiazepine receptor complex, and aggression. Recent Developments in Alco-hol, 13, 139–171.
278 D E T E R M I N A N T S O F A G G R E S S I O N

Mohler, H., Fritschy, J. M., Benke, D., Benson, J., & Luscher, B. (1996). Genetics andfunction of GABAA-receptor subtypes. European Neuropsychopharmacology, 6,S4.
Morgane, P. J., Austin-LaFrance, R., Bronzino, J. D., Tonkiss, J., Diaz-Cintra, S., &Cintra, L., et al. (1993). Prenatal malnutrition and development of the brain. Neu-roscience and Biobehavioral Reviews, 17(1), 91–128.
Nakao, H., Tashiro, N., Kono, R., & Araki, R. (1979). Effects of GABA and glycine onaggressive-defense reaction produced by electrical stimulation of the ventromedialhypothalamus in cats. In M. Itoh (Ed.), Integrative control functions of the brain(pp. 332–334). Amsterdam: Elsevier.
Nelson, C. (1999). Neural plasticity and human development. Current Directions in Psy-chological Science, 8, 42–45.
Nielsen, D. A., Virkkunen, M., Lappalainen, J., Eggert, M., Brown, G. L., Long, J. C.,Goldman, D., & Linnoila, M. (1998). A tryptophan hydroxylase gene marker forsuicidality and alcoholism. Archives of General Psychiatry, 55(7), 593–602.
Nieoullon, A. (2002). Dopamine and the regulation of cognition and attention. Progressin Neurobiology, 571, 1–31.
Parsey, R. V., Oquendo, M. A., Simpson, N. R., Ogden, R. T., Van Heertum, R., Arango,V., & Mann, J. J. (2002). Effects of sex, age, and aggressive traits in man on brain se-rotonin 5-HT1A receptor binding potential measured by PET using [C-11]WAY-100635. Brain Research, 954, 173–182.
Pihl, R. O., Peterson, J., & Finn, P. (1990). The inherited predisposition to alcoholism:Characteristics of sons of male alcoholics. Journal of Abnormal Psychology, 99,291–301.
Ronnekleiv, O., Fang, Y., Choi, W., & Chai, L. (1998). Changes in the midbrain-rostralforebrain dopamine circuitry in the cocaine-exposed primate fetal brain. Annals ofthe New York Academy of Sciences, 846, 165–181.
Roy, A., Pickar, D., De Jong, J., & Karoum, F. (1989). Suicidal behavior in depression:Relationship to noradrenergic function. Biological Psychiatry, 25, 341–350.
Sari, Y., Powrozek, T., & Zhou, F. C. (2001). Alcohol deters the outgrowth of seroton-ergic neurons at midgestation. Journal of Biomedical Science, 8, 119–125.
Sasaki-Adams, D., & Kelley, A. (2001). Serotonin-dopamine interactions in the controlof conditioned reinforcement and motor behavior. Neuropsychopharmacoloogy,25, 440–452.
Seeman, P., Bzowej, N., Guan, H., & Bergeron, C. (1987). Human brain D-sub-1 and D-sub-2 dopamine receptors in schizophrenia, Alzheimer’s, Parkinson’s, and Hunting-ton’s diseases. Neuropsychopharmacology, 1, 5–15.
Siegel, A., Roeling, T., Gregg, T., & Kruk, M. (1999). Neuropharmacology of brain-stim-ulation-evoked aggression. Neuroscience and Biobehavioral Reviews, 23, 359–389.
Simler, S., Puglisi-Allegra, S., & Maudel, P. (1982). Gamma-aminobutyric acid in brainareas of isolated aggressive or non-aggressive inbred strains of mice. Pharmacology,Biochemistry and Behavior, 16, 57–61.
Sirvio, J., Jakala, P., Mazurkiewicz, M., Haapalinna, A., Riekkinen, P., Jr., & Riekkinen,P. J. (1993). Dose- and parameter-dependent effects of atipamezole, an alpha 2–an-tagonist, on the performance of rats in a five-choice serial reaction time task. Phar-macology, Biochemistry and Behavior, 45, 123–129.
Spear, L. P. (2000). The adolescent brain and age-related behavioral manifestations. Neu-roscience and Biobehavioral Reviews, 24, 417–463.
Neuromodulators in Aggression 279

Spoont, M. R. (1992). Modulatory role of serotonin in neural information processing:Implications for human psychopathology. Psychological Bulletin, 112, 330–350.
Stolk, J. M., Connor, R. L., Levine, S., & Barchas, J. D. (1974). Brain norepinephrine me-tabolism and shock-induced fighting behavior in rats. Journal of Pharmacology andExperimental Therapeutics, 190, 193–209.
Tassin, J. P. (1992). NE/DA interactions in prefrontal cortex and their possible roles asneuromodulators in schizophrenia. Journal of Neural Transmission, 36, 135–162.
Tremblay, R., Boulerice, B., Harden, P., McDuff, P., Pérusse, D., Pihl, R. O., & Zoccolillo,M. (1996). Do children in Canada become more aggressive as they approach ado-lescence? Growing up in Canada: The National Longitudinal Survey of Children(pp. 127–136). Ottawa: Statistics Canada.
Van Erp, A. M. M., & Miczek, K. A. (2000). Aggressive behavior, increased accumbal do-pamine, and decreased cortical serotonin in rats. Journal of Neuroscience, 20(24),9320–9325.
Virkkunen, M., Nuutila, A., Goodwin, F. K., & Linnoila, M. (1987). Cerebrospinal fluidmonoamine metabolite levels in male arsonists. Archives of General Psychiatry, 44,241–247.
Volkow, N., Gur, R., Wang, G., Fowler, J., Moberg, P., Ding, Y., Hitzemann, R., Smith,G., & Logan, J. (1998). Association between decline in brain dopamine activitywith age and cognitive and motor impairment in healthy individuals. AmericanJournal of Psychiatry, 155, 344–349.
Walker, E. (2002). Adolescent neurodevelopmental and psychopathology. Current Di-rections in Psychological Science, 11, 24–28.
Williams, G., Srinivas, G., & Goldman-Rakic, P. (2002). The physiological role of 5-HT2A receptors in working memory. Journal of Neuroscience, 22, 2843–2854.
Xiao, L., & Becker, J. B. (1994). Quantitative microdialysis determination of extra-cellular striatal dopamine concentration in male and female rats: Effects of estrouscycle and gonadectomy. Neuroscience Letters, 180(2), 155–158.
Zhou, F. C., Sari, Y., Zhang, J. K., Goodlett, C. R., & Li, T. K. (2001). Prenatal alcoholexposure retards the migration and development of serotonin neurons in fetalC57BL mice. Brain Research Developmental Brain Research, 126, 147–155.
280 D E T E R M I N A N T S O F A G G R E S S I O N

DETERMINANTS OF AGGRESSIONHormones and Aggression
14
Hormones and the DevelopmentalOrigins of Aggression
STEPHANIE H. M. VAN GOOZEN
In considering biological influences on the development of behavior, in thiscase aggression, one should (and this cannot be emphasized enough) alwayssee the behavior as the outcome of a complex interplay of individual, develop-mental, and social factors. Some of these factors might play a more or less im-portant role as a function of the age of a child. For example, with respect tosocial factors, harsh parental discipline with cruel punishment plays a causalrole in the development of antisocial behavior in childhood, whereas the ab-sence of parental supervision is an important factor in antisocial behavior inlate childhood or adolescence (Lahey, Waldman, & McBurnett, 1999). To givean example of a more individual factor, the early (perinatal) hormonal envi-ronment is assumed to play a role in shaping temperament and its develop-ment before the onset of important social influences such as peer interactions;at puberty, when pronounced hormonal changes occur, these individual fac-tors may again become more relevant.
When we consider young children, it is most likely that the origin ofaggressive behavior lies in a combination of a toddler/child with a difficulttemperament and a nonoptimal environment in which ineffective socializationplays a key role: A difficult child elicits harsh, inconsistent, and negativesocialization behaviors, as a result of which a difficult temperament ultimatelydevelops into aggressive or disruptive behavior. Although there are factorsthat contribute to antisocial behavior in childhood becoming chronic, not allantisocial children become antisocial adolescents, and not all antisocial ado-lescents become antisocial adults. A study of the individual factors in the de-velopment of aggression could help explain why these behavioral patternschange over time.
281

In this chapter the role of two hormones—testosterone and cortisol—inthe development and maintenance of aggressive and/or antisocial behavior arediscussed, namely the androgen testosterone and the stress hormone cortisol,the primary reason for their selection being that most research on human ag-gression has focused on these two hormones.
Before considering the evidence of hormonal factors in aggression, it isimportant to keep in mind that aggression can occur in both normal andpathological contexts. In the first case we mean normal individual variationsin aggressive behavior, whereas in the second case aggression occurs more fre-quently and in more serious forms and is usually more persistent. In the lattercase we are dealing with evidence from psychopathological studies. Becausethere have been very few studies addressing the influence of hormones on thenormal development of aggression, evidence in support of the influence ofhormones on both normal variations in aggression and more serious and per-sistent aggression are discussed.
Another complex issue concerns the behavior of interest, aggression, and,in particular, its multiple causations. Many aggressive, antagonistic, or hostileacts are elicited by events, for instance, frustrating or otherwise aversiveevents. Yet aggression results from the individual’s disposition to react aggres-sively to such events. Such a disposition can be conceptualized in variousways, and problems have always existed with the predictive validity of instru-ments that measure such traits. One reason for such problems may be thatmuch aggressive behavior results from emotional responses and that individu-als differ in their proneness to emotional responding of particular kinds.
It is not difficult to find examples from everyday life showing that aggres-sion and emotion are intimately connected phenomena. Aggression can resultfrom fear, hate, or even love. Still, the most common emotion related to theoccurrence of aggressive behavior is anger. Whether a certain situation leadsto anger, and perhaps to aggressive behavior, depends to an important extenton individual differences, such as personality characteristics, and these can bepartly influenced by hormonal (organizing or activating) factors. Moreover,such individual differences can be of a short-term (temporary mood changesin irritability and quick-temperedness) or long-term character. It is clear thatitems in most aggression questionnaires, whether the focus is on adults or chil-dren, measure the occurrence of overt aggressive acts without reference to rel-evant antecedents, such as emotions and situational cues. Although there isimportant research examining the relationship between aggression and anger(Berkowitz, 1989), little has been done to study possible relationships betweenaggression and individuals’ proneness to anger, and as far as I know, nobodyhas studied hormonal influences on anger proneness in young children.
A final comment concerns the type of aggression under investigation. Anemotional or reactive form of aggression (with high levels of emotionalarousal) is commonly distinguished from a more instrumental, proactive, orcalculated (low emotional arousal) form (Vitiello & Stoff, 1997). Within thesecategories aggression can be of a more direct or indirect nature, it can be phys-
282 D E T E R M I N A N T S O F A G G R E S S I O N

ical or verbal, and other subcategories can also be distinguished. It is beyondthe scope of this chapter to discuss these differences, but it should be clear thathighly emotional and impulsive aggression and cold-blooded, calculatedaggression have different neuropsychological and/or neurobiological under-pinnings. For present purposes it is important to keep in mind that studiesinvestigating the relationship between hormones and aggression have rarelyaddressed this issue, focusing usually on one type of aggressive behavior inparticular.
TESTOSTERONE
Why Is Testosterone an Interesting Hormone?
Studies of aggressive behavior in animals indicate that the exchange of antagonis-tic and hostile acts is predominately a male attribute, exhibited betweenmales. Criminological studies reveal a similar clear-cut sex difference: Men usefar more physical violence and are more often the victims of aggression. Bio-logical explanations have linked these findings to the social dominance of themale and to testosterone, the most important male sex hormone. Whereasmany animal studies have demonstrated that the presence of this hormone is aprerequisite for the display of aggressive behavior, results of such studies inprimates (Dixson, 1980; Eberhart, Keverne, & Meller, 1980) and humans areless clear-cut and at best suggestive of a positive association (Albert, Walsh, &Jonik, 1993; Archer, 1991; Archer, Graham-Kevan, & Davies, in press;Harris, 1999; Mazur & Booth, 1998). However, the interest in the aggression–testosterone relationship remains a strong one and can most probably beexplained by the extensive literature on gender differences in aggressive be-havior.
If one investigates behaviors that clearly show differences between menand women, it is clear from a biopsychological perspective that the role of go-nadal hormones is an important one on which to reflect. Indeed, gonadal hor-mones have been shown to be a relevant factor in the origin and expression ofsex differences in behavior: The so-called “organizing” and “activating” ef-fects have been well established in studies on neuroendocrine functioning, inanatomical studies, and in studies of behavior (Goy & McEwen, 1980). Thedevelopment of internal and external genitalia, of sex differences in figure andappearance, and of various classes of sex-specific behaviors, such as the occur-rence of sexual behavior or aggression (but by no means only these!) are to animportant extent related to the presence or absence of testosterone during par-ticular prenatal and postnatal developmental phases (Collaer & Hines, 1995).In adulthood both males and females are exposed to different classes andpatterns of gonadal hormone stimulation, which, again, affect brain and be-havior. However, understanding the origin of sex differences in behavior iscomplicated by the fact that such differences are not only manifestations ofthe organizing and activating effects of hormones; they are also the result of
Hormones and Aggression 283

interactions between these hormonal influences on behavior, on one hand, andsocial influences on the individual during various phases of development, onthe other.
Organizing and Activating Hormonal Effects
The long-term developmental and structural effects of pre- and postnatal ex-posure to sex hormones on sex-dimorphic behavior have been termed “orga-nizing” effects. These developmental effects are distinguishable from “activat-ing” effects of gonadal hormones in that activating effects are reversible,repeatable, and not limited to a critical phase of development. It is, however,generally assumed that organizing, and activating hormone effects interact,and the distinction is therefore not as easy to make as the definition of theterms suggests. Activating hormone effects presumably act in concert withperinatal hormonal and early developmental experiential factors to perpetuatesex-dependent and individual behavioral characteristics (Van de Poll & VanGoozen, 1992). Organizing effects are usually tested by manipulating the ex-posure to sex hormones during fetal development, whereas activating effectsare investigated at the time of puberty or adulthood. For obvious reasons, ex-perimental research on these effects has focused almost exclusively on ani-mals, rodents in particular.
Androgenic gonadal hormones have been shown to be involved in vari-ous aspects of fighting behavior, including “intermale” aggression (Moyer,1976). Studies of female rodents also demonstrate that the female, when ex-posed to testosterone prenatally or in adulthood, may exhibit high levels ofaggressive behavior equal to those of males (Van de Poll, Van Zanten, &DeJonge, 1986). Although animal studies can teach us a great deal about therole of testosterone in behavior, and in aggression in particular, extrapolatingthe results to humans and implicitly postulating similar mechanisms inhumans may present us with an unduly simplified and misleading picture ofhuman functioning. The important issue to be addressed next is whether indi-vidual differences in human aggression have anything to do with differences inhormone levels or hormonal functioning.
Organizing Effects: Sex Differences in Aggression
Evidence concerning the development of aggression is obviously important forthe study of sex differences in aggression. One can assume that socializingprocesses start early, so that if one wants to research gender differences, thesehave to be studied at an early age. Boys and girls differ in play styles and toypreference: Boys are more energetic and active and show more aggressive anddestructive behavior, involving so-called rough-and-tumble play, a form ofplay-aggression that has also been observed to be sex-dependent in monkeys(Hines & Green, 1991). Some of these sex differences have been reported in
284 D E T E R M I N A N T S O F A G G R E S S I O N

children as early as 12 months of age. Recently, Hines et al. (2002), in a largepopulation-based study, found a relationship between testosterone levels, asmeasured in the blood samples of pregnant women, and gender role behaviorin 3.5 year old girls (n = 337), such that as maternal testosterone levels in-creased, so did masculine-typical gender role behavior. No relationships be-tween hormones and gender role behavior were found in boys (n = 342). Thelargest gender differences, however, are manifested in the years that follow,and this is a strong argument for the influence of socialization, modeling, andnorms and values.
A fair amount of evidence also exists for gender differences in aggressivefeelings and behaviors during adolescence. Most of these associations involvethe androgens and, as expected, larger effect sizes have been reported for boysthan for girls. Several studies found evidence of a mediating role for impa-tience and impulse control in the association between hormonal levels andaggression (Susman et al., 1987; Warren & Brooks-Gunn, 1989). Olweus,Mattson, Schalling, and Low (1988) reported a causal effect of pubertal hor-mones on aggressive behavior via increased impatience and irritability.
Hormonal influences on the development of the human brain and behav-ior can be investigated in individuals who have been exposed to unusual go-nadal hormone environments, pre- or neonatally, due to genetic disorders orbecause their mothers were prescribed hormones during pregnancy. Reinisch(1981) concluded on the basis of her study that the development of aggressivebehavior in humans is partly determined by prenatal androgens. Aggressionproneness in children was measured with a response hierarchy test, consistingof a series of drawings picturing daily situations (e.g., a child takes away yourtoy) together with a number of possible reactions (hitting, shouting, go to par-ents, or crying); the number of aggressive reactions was the aggression score.Reinisch found a clear sex difference in (physical) aggression between boysand girls. Moreover, the administration of progestins to the mother duringpregnancy (i.e., progestin being a synthetic sex hormone with androgenic ac-tion, prescribed to avoid the risk of possible miscarriage) had an aggression-enhancing influence on exposed boys and girls. Animal research on mice hasdemonstrated that sex differences in aggression can be counteracted by expos-ing female mice to testosterone or by castrating male mice postnatally. Laterstudies of similar effects in girls with congenital adrenal hyperplasia (CAH: anautosomal recessive disorder associated with an enzyme defect in the pathwayof cortisol and aldosterone biosynthesis), who had been exposed prenatally toelevated levels of androgens, have not shown clear effects on aggressive behav-ior, although different aspects of gender role behavior were affected. Thesegirls were more energetic, played with male peers and with boys’ toys more,and were identified by themselves and others as tomboys (Ehrhardt & Meyer-Bahlburg, 1981; Berenbaum & Resnick, 1997; Berenbaum, Duck, & Bryk,2000).
These studies of the prenatal effects of androgens on aggression used
Hormones and Aggression 285

small samples and different research methods. For example, Reinisch (1981)measured aggression proneness as a more or less stable way of reacting to anumber of standard situations. In studies in which no effects were found, amore or less standardized interview was often conducted with the children,their parents, and sometimes also their teachers. The differences in findingsmay be due to the different research methods used. Gender differences in ag-gression seem to be found most often when the subjects are offered responsealternatives, for instance, a choice between verbal versus physical aggression.
Reviewing the results of these studies of humans exposed prenatally toexcessive androgens, one can conclude that certain aspects of gender-relatedbehavior are influenced by prenatal hormones. The influence of these hor-mones is primarily on “tomboyism” or over-energetic and playful behavior. Ingirls, interest in maternity may be affected, but many androgenized womenwho are able to have children do become mothers. Clear-cut differences ingender identity and sexual orientation have not been found (Ehrhardt &Meyer-Bahlburg, 1981; Ehrhardt et al., 1985; Gooren, Fliers, & Courtney,1990), although androgenized females sometimes show a delay in exhibitingnormative teenage sexual experience and motivation, possibly because thesegirls look “different” and are less likely than their normal counterparts to de-velop a typically female body shape and may be masculinized if not treated byestrogens. Thus, whereas clinical studies with humans suggest organizationaleffects of excessive androgens on gender role behavior (specifically, opposite-sex relationships and activity level), they do not appear to have a clear influ-ence on aggression or other gender-typed behavior such as sexual behavior orsexual orientation (Paikoff & Brooks-Gunn, 1990).
In sum, there are not many studies that have addressed the issue ofwhether prenatal androgen levels influence human aggression. The limitedsupport for hormonal effects on aggression comes from studies with good ex-perimental designs, good selection of control groups, and standardized mea-sures of aggression (Berenbaum & Resnick, 1997). The literature reviewedearlier in this section suggests that there is no good evidence of a link betweenandrogens and aggression in children, apart from Reinisch (1981). The recentstudy by Hines et al. (2002) and the research on girls with CAH were cited asevidence of effects of androgens on energy levels, play activity, and gender rolebehavior, not aggression.
Activating Effects of Testosterone on Aggression in Adolescents and Adults
Correlations in men between androgens and self-reports of aggressive behav-ior or scores on aggression inventories are generally low (Meyer-Bahlburg,Boon, Sharma, & Edwards, 1974). For example, in one study a correlation of0.28 was found between total testosterone and scores on a physical aggressionsubscale; the correlation with free testosterone was even lower, at 0.14(Gladue, 1991). A study relevant to the issue of whether pubertal increases in
286 D E T E R M I N A N T S O F A G G R E S S I O N

testosterone levels produce an increase in physical aggression is that byHalpern, Udry, Campbell, and Suchindran (1994). They found no evidence ofeither an increase in aggression or an association between testosterone and ag-gression in a 3-year longitudinal study of 100 teenage boys (ages 12–13 atstudy entry) going through puberty. Recently, in a high-quality multitrait,multimethod, and multisampling study, Granger et al. (2003) also found norelationships between (saliva) testosterone and aggression, delinquency, orsymptoms of disruptive behavior disorders in a relatively large sample of ado-lescent boys (n = 106). However, present or past aggressive behavior hassometimes been found to correlate positively with testosterone concentrations.For instance, Dabbs, Frady, Carr, and Besch (1987) measured saliva testoster-one in 89 male prison inmates. They found that testosterone concentrationwas related to the type of crime: Men convicted for violent crimes (murder,rape, robbery) had higher testosterone concentrations than men convicted fortheft or burglary. Most of the studies on activating effects have beencorrelational in design, thereby limiting the possibility of establishing a causalrole for testosterone in eliciting aggression. A more compelling line of researchis one that examines the effects of testosterone administration or depletion onaggression. Finkelstein et al. (1997) investigated the effects of testosterone ad-ministration on aggression in hormone-deficient (delayed puberty) adoles-cents. The data demonstrated significant testosterone effects, specifically onphysical aggression and aggressive impulses in boys (and similar effects werefound after estrogen administration in teenage girls). O’Connor, Archer, Hair,and Wu (2002) studied the effects of exogenous testosterone on aggressionand mood in 30 eugonadal and 8 hypogonadal men. No significant increasesin a range of aggression measures and no testosterone-related mood effectswere observed in the eugonadal men. However, significant positive mood ef-fects were found in the hypogonadal group. Another group of human subjectsthat is interesting from the standpoint of the study of activating effects of tes-tosterone on aggression are transsexuals. In the process of their sex reassign-ment procedure, these patients are treated with high doses of so-called oppo-site sex hormones. In two studies it was found that when female-to-maletranssexuals received high doses of androgens, their anger and aggressionproneness increased, whereas the reverse happened in male-to-female trans-sexuals receiving anti-androgens and estrogens (Van Goozen, Frijda, Van dePoll, 1994; Van Goozen, Cohen-Kettenis, Gooren, Frijda, & Van de Poll,1995). It is nevertheless clear that the evidence of a link between androgensand aggression is much weaker in humans than in animals. The results seem tobe clearer when the relation is studied in younger age groups, as shown instudies of adolescents (Olweus et al., 1988; Schaal, Tremblay, Soussignan, &Sussman, 1996). This could be because social restrictions on aggression arestill limited at a younger age and behavior has yet to be shaped. One thereforewonders what evidence there is for a relation between testosterone and aggres-sion in prepubertal children.
Hormones and Aggression 287

Androgens and the Development of Aggression in Young,Prepubertal Children
Testosterone
Given that there has been little research on testosterone and aggression inolder age groups, it does not come as a surprise that there is even less informa-tion on young children. Until very recently it was not easy to measure testos-terone levels in children. This is because prepubertal plasma levels are verylow,1 and if one wants to sample testosterone in saliva, which is the least com-plicated way, only the unbound fraction of testosterone is available. To myknowledge, only four studies have measured testosterone levels (two inplasma and two in saliva) in preschool or schoolaged children and relatedthese levels to aggression. Constantino et al. (1993) measured testosterone inplasma of 18 highly aggressive boys and 11 controls, all children being be-tween 4 and 10 years old, and found neither a difference in testosterone be-tween the groups nor a relationship between testosterone and the children’sChild Behavior Checklist (CBCL; Achenbach, 1991) aggression scores. Theauthors concluded that these findings raise questions about inferences fromadult studies suggesting that testosterone may play a causal role in the devel-opment of aggression. However, an explanation for the negative results maylie in the composition of the groups, because 10 out of 18 aggressive boys hada comorbid diagnosis of psychotic disorder. Psychosis has been considered anexclusion criterion (Kruesi et al., 1990), because aggression as a temporarysymptom of a psychotic episode is clearly different from aggression as ob-served in children with a psychiatric diagnosis of oppositional–defiant disor-der (ODD) (American Psychiatric Association, 1994) or conduct disorder(CD) (American Psychiatric Association, 1994), let alone in normal, healthychildren. Moreover, the age range in the subgroups was large (between 4 and10 years), and the investigators had to divide the groups into smaller sub-groups (i.e., an adrenarchal subgroup consisting of 6 psychotic–aggressive, 4nonpsychotic–aggressive, and 6 normal controls; and a preadrenarchal sub-group of 4 psychotic–aggressive, 4 nonpsychotic–aggressive, and 5 normalcontrols). Van Goozen, Matthys, Cohen-Kettenis, Thijssen, and Van Engeland(1998b) also found no difference in plasma testosterone between boys withCD (n = 15) and normal controls (n = 25; all participants were between 8 and12 years old), and there were no significant relationships between testosteroneand CBCL ratings of aggression and delinquency. A third study focused onnormal, healthy 4-year-olds and measured testosterone in saliva (Sánchez-Martín et al., 2000).2 No difference in testosterone was observed betweenboys (n = 28) and girls (n = 20), but testosterone in boys was positively corre-lated with observations of more serious aggression in social interaction. Nosuch relations were observed for more playful aggression, and no relationsbetween testosterone and behavior were observed in girls. Finally, Scerbo andKolko (1994) measured saliva testosterone in forty 7–14-year-old clinic-
288 D E T E R M I N A N T S O F A G G R E S S I O N

referred disruptive children and found a significant positive relationship be-tween testosterone and staff-rated aggression, after taking age and physicalcharacteristics into account, but no differences in mean levels for childrenwith or without CD, ODD, or ADHD.
DHEA/DHEAS
From about age 6, children exhibit a gradual increase in androgens of adrenalorigin, a period called the adrenarche, and it is not until puberty that gonadalandrogens, such as testosterone, become more important. Research in pre-pubertal children should therefore not only focus on testosterone, but shouldalso consider adrenal androgens, such as dehydroepiandrosterone (DHEA), itssulphate (DHEAS), and androstenedione. Of these, DHEA and DHEAS areparticularly interesting because they are also endogenously synthesized by thebrain and act as neurosteroids (Robel & Baulieu, 1995), they increaseneuronal excitability, enhance neuronal plasticity, and have neuroprotectiveproperties (Wolf & Kirschbaum, 1999) and, finally, they appear to have po-tent antagonistic effects on central gamma-aminobutyric acid (GABAA) mech-anisms, which are involved in aggressiveness (Majewska, 1992). One wouldtherefore expect to find elevated DHEA/DHEAS levels in individuals who areaggressive.
Until now, very few studies have focused on the role of adrenal androgensin children. Interestingly, a pattern of DHEA hyposecretion has been found inchildren and adolescents with major depression (Goodyer et al., 1996). So far,three studies have examined DHEAS levels in prepubertal aggressive boys.The previously mentioned study by Constantino et al. (1993) found no rela-tionship with aggression, but, as noted earlier, there are some grounds fortreating this null finding with caution. Van Goozen et al. (1998b) found thatDHEAS levels were significantly higher in boys with CD than in normal con-trols and were strongly related to aggression and delinquency scores; differ-ences between the two groups in androstenedione and testosterone were,respectively, marginally significant and not significant. However, no data ondevelopmental status were collected and no psychiatric control group was in-cluded. One could speculate that if higher adrenal androgen levels reflect adre-nal response to stress, and if chronic stress stimulates adrenal developmentand secretion (Sapolsky, 1997), it is possible that elevated DHEAS levelswould also be found in other psychiatric groups as a result of stress associatedwith the psychiatric disorders. The aim of a subsequent study (Van Goozen etal., 2000b) was to investigate DHEAS levels in a new group of prepubertalchildren with ODD and normal and psychiatric controls, including a group ofchildren with attention-deficit/hyperactivity disorder (ADHD). DHEAS levelswere examined in relation to developmental status, psychiatric diagnosis, andintensity of aggression and delinquency during the preceding 6 months asrated by parents/care workers. The results again showed that the ODD group
Hormones and Aggression 289

had significantly higher levels of DHEAS than both the normal and psychiat-ric control groups.
At present it is not clear how best to explain these results. It may be thatthe DHEAS levels of the children with ODD were higher from early childhoodonward; alternatively, it may be that these children had higher levels onlysince the beginning of adrenarche. However, by measuring the development ofthe secondary sex characteristics and finding that the different groups of chil-dren were similar with respect to physical development, Van Goozen andcolleagues could rule out the possibility that the children with ODD werephysically more mature than their counterparts in the two control groups.
The positive relation observed between ODD and/or CD, on one hand,and plasma levels of DHEAS, on the other, is of interest. As mentioned earlier,higher endogeneous levels of this neuroexcitatory steroid may contribute tomore aggression because of its GABAA antagonistic actions (Majewska,1992). It is also possible that higher DHEAS levels contribute to a larger poolof endogeneous testosterone. However, in our own study measuring differentandrogenic hormones (i.e., testosterone, DHEAS, and androstenedione) inchildren with CD, we found that the relation between DHEAS and aggressivebehavioral problems is stronger than that between testosterone and suchproblems, suggesting a more important role of CNS excitatory mechanisms inaggressive behavior of children (Van Goozen et al., 1998b). These results dem-onstrate that adrenal androgen functioning is an important subject for futureresearch on the developmental origin of aggression in children.
Methodological Considerations
A large part of human aggression research has been questionnaire-based, andit is worthwhile to reflect on the potential for measuring the effects of hor-mones on aggression by means of this methodology. If one accepts the evi-dence that hormonal effects on aggression are rather subtle, and admits thatthe assessment of the type of behavior under consideration is complicated, notonly because there are many different ways of expression, but also becauseone is dealing with a sensitive (and rather negatively valued) issue, it followsthat asking people to report on their own aggression or asking informants fortheir observations about other people’s aggression is fraught with difficulties.The evidence shows that the assessment of the frequency and intensity of chil-dren’s aggression is, to a large extent, dependent on whether one asks the par-ents, the teachers, or the children themselves, and that there are considerablecross-informant discrepancies (see, e.g., Loeber, Green, Lahey, & Stouthamer-Loeber, 1991). If the assessment of the target behavior is so variable, how canone expect to find a reliable relationship with hormonal parameters?
It is safe to conclude that questionnaire studies of the relationship be-tween testosterone levels in humans and aggressive disposition have generateda conflicting body of evidence and that, on the whole, directly observationalresearch or studies that treat differences in hormonal condition as an inde-
290 D E T E R M I N A N T S O F A G G R E S S I O N

pendent variable (e.g., Finkelstein et al., 1997; Reinisch, 1981; Sánchez-Martín et al., 2000; Van Goozen, Frijda, Wiegant, Endert, & Van de Poll,1996) have been better able to reveal the rather subtle hormonal effects onovert angry and aggressive behavior.3 Given that there are very few data onchildren’s aggression, it is our belief that future studies on the development ofaggression in children, in focusing on the involvement of hormonal parame-ters, should rely on such methods to explore this issue further.
CORTISOL
A second hormonal factor contributing to individual differences in antisocialbehavior is cortisol. Cortisol is related to the experience of stress, and itseems likely, on the basis of observing overt aggressive behavior, that thestress system is involved in displaying aggression. Specifically, one could hy-pothesize that people who engage in aggression are, relatively speaking, un-afraid of the negative consequences of their actions or, stated differently, areless sensitive to punishment (Matthys, Van Goozen, De Vries, Cohen-Kettenis, & Van Engeland, 1998; Van Goozen et al., 2004). One importantbiological stress system is the hypothalamic–pituitary–adrenal (HPA) axis,which plays an important role in the regulation of both physical and psy-chological stress.
It has been proposed that a reduced activity of the HPA system is part ofthe biological predisposition to antisocial behavior (Van Goozen, Matthys,Cohen-Kettenis, Buitelaar, & Van Engeland, 2000a). Low activity or arousalhas been linked to fearlessness, sensation seeking, and/or risk-taking behavior(Raine, 1996). Specifically, one can predict that individuals who are less sensi-tive to stress (i.e., stress hyporesponsive) are more likely to engage in aggres-sion because they do not fear the negative consequences of their actions be-cause of a lack of fear conditioning. A lack of fear of punishment would alsoreduce the effectiveness of social conditioning and could therefore explainwhy this pattern of behavior, especially when observed in disruptive and anti-social clinical cases, can be so persistent and hard to treat.
The Stress System
There are clear indications that stress plays an important role in the explana-tion of individual differences in antisocial behavior, and in this connection it isuseful to draw on concepts such as stress sensitivity, stress regulation, andcoping with stress. The activity of the neuroendocrine HPA system is centralto stress resistance and can be assessed by using measures of its end products,the glucocorticoids. The primary glucocorticoid in humans is cortisol, and thishormone can be relatively easily assessed in saliva (Dettling, Gunnar, & Don-zella, 1999). A well-functioning HPA system is necessary for survival, buthyperactivity of this system may have deleterious effects on immune system
Hormones and Aggression 291

activity and brain areas involved in memory and attentional processes andmay influence the expression of defensive behaviors (Gunnar, Tout, de Haan,Pierce, & Stansbury, 1997). Thus, healthy adaptation involves efficient regula-tion of HPA system activity.
Generally speaking, the expression of an adrenocortical stress response isdependent on (1) whether the situation is perceived as potentially threatening,(2) whether the outcome is important to the individual, and (3) whether theindividual expects to have the resources necessary to manage the threat (Gun-nar et al., 1997). Factors such as control, sense of success and efficacy, andpredictability are therefore very important in the activity of the HPA system.
The starting point of research on the relationship between stress and anti-social or aggressive behavior is the hypothesis that aggressive individuals areless fearful or sensitive to stress. This can be deduced from the fact that theseindividuals are less inhibited and more prone to engage in risky, stressful, ordangerous situations than other people are. If this is true, there are two possi-ble explanations for a relationship between a lower stress sensitivity and ag-gressive or antisocial behavior. One theory claims that antisocial or aggressiveindividuals have low levels of fear (Raine, 1996). A relative lack of fear or aless inhibited temperament would lead to aggression or antisocial behavior be-cause the person is less sensitive to the negative consequences of his or herown or other people’s behavior in general, and the receipt of punishment inparticular, and because the person overestimates his or her own coping strate-gies. If this is the case, the implication for the treatment of serious aggressionor antisocial behavior is clear. Thus, aggressive individuals, including children,have problems in conditioning, so trying to point out the negative conse-quences of behavior, or punishing unacceptable behavior, is likely to have littleor no effect.
A different stress theory focuses on sensation seeking (Zuckerman, 1991).Here it is argued that a certain level of stress is needed in order to feel pleasantand that too little or too much stress is experienced as unpleasant. Aggressiveindividuals are supposed to have an elevated threshold for stress. They areeasily bored and are not put off by situations that most people find too excit-ing, stressful, or dangerous.
Human Studies on Cortisol and Aggression
What evidence is there that a dysfunctional stress system plays a role inaggressive behavior? Several studies have found that antisocial and aggressiveadults have lower levels of cortisol. There are also indications of inverse rela-tionships: The more aggressive the behavior, or the more serious the antisocialbehavior disorder, the lower the biological stress levels (King, Jones, Scheurer,Curtis, & Zarcone, 1990; Virkkunen, 1985; Woodman, Hinton, & O’Neill,1978). This type of research has also been conducted on children with antiso-cial behavior, albeit less frequently. Although the results are less consistent
292 D E T E R M I N A N T S O F A G G R E S S I O N

than those from the adult studies, the predicted relations have been found. Forexample, Tennes and Kreye (1985) and Tennes, Kreye, Avitabe, and Wells(1986) found a relationship between lower cortisol levels in urine and the in-tensity of aggressive behavior as expressed toward peers or a teacher. In otherstudies researchers have found a negative relation between conduct disorderseverity and cortisol (McBurnett, Lahey, Rathouz, & Loeber, 2000; Pajer,Gardner, Rubin, Perel, & Neal, 2001; Van Goozen et al., 1998a; Van Goozenet al., 2000a; Vanyukov et al., 1990).
However, the findings for children and adolescents are not as unequivocalas those for antisocial adults, in that positive relationships between cortisoland antisocial behavior have also been found in some studies. In normal,healthy adolescents a positive relationship was found between aggression andcortisol response level during an experimentally induced aggression task(Gerra et al., 1997) and during an emotion-arousing and painful procedure(Susman, Dorn, Inoff-Germain, Nottelmann, & Chrousos, 1997). Moreover,McBurnett et al. (1991) reported high levels of cortisol in conduct-disordered(CD) children, but only when they had a comorbid anxiety disorder. Finally,some studies have found no relationship between cortisol and antisocial or ag-gressive behavior (Klimes-Dougan, Hastings, Granger, Usher, & Zahn-Waxler,2001; Kruesi, Schmidt, Donnelly, Hibbs, & Hamburger, 1989; Scerbo &Kolko, 1994; Schulz, Halperin, Newcorn, Sharma, & Gabriel, 1997; Stoff etal., 1992; Susman et al., 1999).
Apart from the direct relationship between stress and antisocial oraggressive behavior, cortisol also seems to play a part in the prediction of anti-social behavior in the long term (McBurnett et al., 2000). At present is notknown whether a reverse relationship also exists, that is, that aggressive chil-dren with an elevated sensitivity to stress are more fearful and therefore have alower risk of persisting in their antisocial behavior. Such data already exist forautonomic nervous system measures (Brennan et al., 1997). An importantfuture line of research is therefore to establish whether the development of ag-gression over time is related to young children’s stress sensitivity or level offearfulness, in the sense that children with elevated HPA axis activity levels arebiologically protected against acting aggressively on a frequent and/or moreserious basis.
Methodological Considerations
The mixed findings in regard to cortisol and aggression in children and ado-lescents may be due to important methodological differences between thesestudies. First, the label “aggression” or “antisocial” has been used for be-haviors as different as physical aggression, running away from home, steal-ing, and drug use (Coie & Dodge, 1998; Tremblay, 2000, 2003). Further-more, studies that specifically assess physical aggression have generally nottaken into account whether it is of a reactive or proactive type. Reactive
Hormones and Aggression 293

and proactive aggression have been observed in children, adolescents, andadults (Brendgen, Vitaro, Tremblay, & Lavoie, 2001; Dodge, Lochman,Harnish, Bates, & Pettit, 1997). Reactive aggression is impulsive, often ac-companied by disinhibition and affective instability, but not necessarily byantisocial tendencies; it is characterized by high levels of bodily arousal.Proactive aggression, however, is nonimpulsive and controlled and occurs inthe context of persistent antisocial behavior. Proactively aggressive individu-als are less likely to have unstable affect, their aggression is goal directed,and the level of arousal is usually low (Vitiello & Stoff, 1997). It seems the-oretically plausible that proactive aggression is likely to be associated withlow levels of cortisol, whereas reactive aggression is accompanied by ele-vated levels of cortisol.
Differences in assessments of cortisol may also explain the diversity of re-sults. Some studies measured cortisol under resting conditions (Kruesi et al.,1989; McBurnett et al., 2000; Tennes & Kreye, 1985), and other studies mea-sured cortisol before the occurrence of an anticipated stressful event (Dawes etal., 1999; Moss, Vanyukov, & Martin, 1995), during an aggression-provokingtask (Gerra et al., 1997), or under highly stressful conditions (Van Goozen etal., 1998a, 2000a). Moreover, most studies on children’s cortisol involvedmeasurements varying over the day, without controlling for the clear circadianrhythm in cortisol secretion or for the kinds of activity in which participantshad been engaged before the start of the study (McBurnett et al., 2000); only afew studies measured cortisol at specific time points, having kept the condi-tions constant for all participants (Susman et al., 1997; Van Goozen et al.,1998a, 2000a). Finally, some studies used only clinical samples (Scerbo &Kolko, 1994; McBurnett et al., 2000), some compared clinical samples withnonclinical samples (Kruesi et al., 1989; Pajer et al., 2001; Stoff et al., 1992;Van Goozen et al., 1998a, 2000a), and others used community samples (Gerraet al., 1997; Klimes-Dougan et al., 2001).
To summarize, research on aggressive child psychiatric patients (i.e., chil-dren with a psychiatric diagnosis of CD or ODD) demonstrates that thesechildren are characterized by a lower sensitivity to stress, as reflected by alower secretion of cortisol during experimental conditions involving stress ex-posure (Van Goozen et al., 2000a). These data support the fearlessness theoryof aggression as described by Raine (1996). The situation is less clear with re-spect to the issue of whether persistently aggressive children also have lowerresting levels of cortisol.
Furthermore, normal variations in HPA system (re-) activity seem to playa clear role in temperamental differences between children, with higher activ-ity being linked to a shy, inhibited, and anxious temperament and low activitywith a more impulsive and aggressive temperament (Gunnar et al., 1997;Kagan, Resnick, & Snidman, 1988; Susman, Schmeelk, Ponirakis, & Gariepy,2001). However, there has been hardly any research directly studying the rela-tionship between cortisol and the occurrence and development of aggression.This is clearly an important issue for future research.
294 D E T E R M I N A N T S O F A G G R E S S I O N

The Role of Early Experiences
We do not currently know why aggressively acting individuals have lowerHPA-axis activity levels, although it is, of course, possible that genetic factorsplay a role (Bakshi & Kalin, 2000). However, there is also evidence that stress-ful events—by which is meant serious stressors such as neglect, abuse, andtrauma—in the first years of life play an important role in “programming” theHPA axis. The evidence comes mainly from animal studies, but it seems likelythat the neurobiological consequences of the severe stress that can be manipu-lated in animal studies also occur in humans.
Interactions between a person’s biological makeup, on one hand, andthe environment in which that person is raised, on the other, determine hisor her physical and psychological development. Physical and biologicalproblems during important phases in pre- or postnatal development (e.g.,birth complications, intense stress or illnesses during the mother’s preg-nancy), together with aversive early psychosocial experiences (such as mal-nutrition, poverty, neglect, and abuse) contribute importantly to the devel-opment of personality and psychopathology (Gottlieb & Halpern, 2002).There is increasing evidence that such interactions between biological andenvironmental factors affect the developing brain both prenatally and post-natally, including the functioning of the HPA axis (Francis, Caldji, Cham-pagne, Plotsky, & Meaney, 1999; Kaufman, Plotsky, Nemeroff, & Charney,2000; Liu et al., 1997) as well as the serotonergic system (see Suomi, Chap-ter 4, this volume).
Although preclinical studies offer important suggestions regarding theway in which early experiences can influence the development of specific bio-logical systems that are involved in aggressive behavior, the conclusions arebased on animal data and it is obvious that such experimental studies cannotbe done on humans. There is, however, some preliminary evidence that similarprocesses can occur in humans.
In a study conducted in an orphanage in Romania, Carlson and Earls(1996) measured the cortisol levels of children who had been severely ne-glected. They found that these children failed to exhibit a normal diurnalcortisol rhythm. In another phase of the study they exposed some of thesechildren to an intervention. In this intervention the children lived in smallergroups, and they were positively stimulated by their caretakers and were giventoys. After 13 months their cortisol levels were measured again in order to es-tablish whether they exhibited a normal diurnal pattern. This was not thecase. Such a finding is consistent with the view that a long and intense periodof neglect had a permanent effect on cortisol functioning, although it is, ofcourse, also possible that the intervention was too short or too weak to allowthe children to recover normal functioning.
Although there is presently not much solid evidence that processes similarto those observed in animals also occur in humans, it is known that abuse andneglect are serious psychosocial stressors, that such experiences can occur at a
Hormones and Aggression 295

young age, and that a high proportion of children with aggressive behavioralproblems have had such experiences.
FUTURE RESEARCH ON ADRENAL STEROIDS
Testosterone has been found, albeit inconsistently, to play a role in aggression,but levels are very low in prepubertal children and it is therefore difficult toassess its role in the development of this behavior from an early age. Thischapter reviewed the evidence that children with aggressive or antisocial be-havior have increased plasma levels of the adrenal androgen DHEAS and ei-ther a decreased basal level of another adrenal steroid, cortisol, or a decreasedcortisol reactivity to stress. It is not unusual to observe these steroids changingin opposite directions. Adrenocorticotropic hormone (ACTH) stimulates thesecretion of cortisol, DHEA, and DHEAS, but there are many instances of dis-sociation between the secretion of cortisol and adrenal androgens, indicatingthat they are governed by separate regulatory mechanisms and that other fac-tors may modulate their biosynthesis. Moreover, although both are secretedby the adrenal cortex, they have different sites of origin. Clarke, Fearon,Cunningham, and McKenna (1996), in their research on the hypothalamic–pituitary–adrenal (HPA) axis, demonstrated that beta-endorphins (the levelsof which rise at adrenarche at the same time as the secretion of DHEA/DHEAS increases) and joining peptide stimulated adrenal androgen produc-tion but inhibited ACTH-stimulated cortisol production. One could inferfrom these results that a dissociation between cortisol and adrenal androgensecretion might be found in people with antisocial behavior, but it is also pos-sible that other regulatory mechanisms will be uncovered in future research.
Clearly, adrenal androgens in young children should be studied moreextensively, and preferably in combination rather than separately. The obser-vation of an association between increased DHEAS levels, decreased cortisollevels, and a current diagnosis of ODD or CD in children (Van Goozen et al.,1998a, 1998b, 2000a, 2000b) suggests that it would be worthwhile to studythe mechanisms underlying adrenal steroids alterations in childhood. If futurestudies replicate the findings of an aberrant secretion of adrenal steroids (i.e.,an excess of DHEAS and a deficit of cortisol reactivity), this would confirmthe idea that such a hormonal profile contributes to aggressive or antisocialtendencies.
Such a hormonal line of research on the developmental origins of aggres-sion should focus on children as young as possible, preferably in the prenatalperiod, and should extend from the immediate postnatal period into early andlate childhood. The implication is that hormonal samples would be taken fre-quently and controlled for various internal (e.g., diurnal rhythm) and external(e.g., life events) influencing factors. Not only is such a line of research time-consuming and expensive, it is also difficult to carry out for practical reasons.Large sample sizes are required, and these have to be followed up over time.
296 D E T E R M I N A N T S O F A G G R E S S I O N

Moreover, as far as the prenatal part of such a project is concerned, recentevidence from my own research group suggests that it is not steroid levelsmeasured in the umbilical cord or maternal serum (assessed over the three tri-mesters of pregnancy) that provide the best source of information if one wantsto assess androgen exposure during fetal development (Van de Beek, Thijssen,Cohen-Kettenis, Van Goozen, & Buitelaar, 2004). Rather, it is the analysis ofthe amniotic fluid that provides the most valid assessment of fetal androgenexposure during a very sensitive period of development (about 16–18 weeksinto pregnancy). Specifically, it was found that the mother’s own androgenlevels were unrelated to the androgen exposure of her child as reflected in theamniotic fluid. If one accepts these results, this poses severe restrictions onconducting this type of research, because it is not easy to collect amniotic fluidsimply for scientific purposes. Nevertheless, if one wants to investigate hor-monal influences on the very early development of temperament in general,and aggression in particular, this seems to be the way to go about it.
The picture is somewhat different for cortisol. An adequate physiologicalstress response, characterized by a rapid increase and subsequent decrease instress hormones, is essential in adapting to and coping with stress. As dis-cussed earlier, variations in both basal activity and stress responses have beenassociated with the existence of problem behavior and the development ofpsychopathology. Long-term changes in stress hormonal functioning play akey role not only in the pathophysiology of aggressive behavior but also in de-pression (Goodyer et al., 1996). Adequate physiological stress responses arealso important in infants. Neonates (4–7 days old) who were alert during aphysical examination and well orientated had relatively higher baselinecortisol values and weaker cortisol responses (Spangler & Scheubeck, 1993).Insecure–avoidant and disorganized 1-year-old children showed a clear in-crease in cortisol during the Strange Situation Test (SST) as compared with se-curely attached children, who showed an adequate behavioral response andno increase in cortisol (Spangler & Grossmann, 1993). These examples illus-trate the relationship between physiological and behavioral responses tostress. However, the precise relationship between behavioral and physiologicalstress responses is complex and dependent on age and developmental phase,type of stressor, and the state the child is in.
Given that physiological stress responses play an important role innormal and deviant forms of adaptation, it is important to establish what de-termines individual differences in reactivity. Animal research has shown thatprenatal and early postnatal stress results in long-term changes in the regula-tion of the offspring’s stress hormonal (HPA) axis (Francis, Diorio, Plotsky, &Meaney, 2002; Francis, Szegda, Campbell, Martin, & Insel, 2003). Thus, itcan be assumed that individual differences in maternal hormones and emo-tions can have a significant impact on fetal neural development and, specifi-cally, that the amount of prenatal stress the mother experiences has a pro-gramming effect on the reactivity of the child’s HPA axis (Anisman, Zaharia,Meaney, & Merali, 1998). In humans, prospective research examining the in-
Hormones and Aggression 297

fluence of prenatal stress on the HPA axis of mother and fetus, and later HPAaxis functioning and development in the child, is virtually absent (but seeSusman et al., 2001). At present it is therefore unclear how prenatal and earlypostnatal stress in the mother affect the infant’s HPA axis and temperament.
Following birth, maternal hormone concentrations and emotions can stillcontribute to the development of the infant’s temperament as mediated by thequality, quantity, and regularity of maternal interactions. The postpartumperiod is for many women a time of intense emotions attributable to the dra-matic psychological and role-requirement changes that accompany the transi-tion into motherhood. New mothers have to assume responsibility for a help-less infant, often without adequate preparation. If these challenging roles andresponsibilities occur in a family context without adequate social or economicsupport, negative effects on the mothers and their children may well occur.Postpartum maternal emotions are in many women characterized by dys-phoria or other depressive symptoms, and the rapid decrease in gonadal andadrenal hormone concentrations in this period is likely to be a contributingfactor. It has been found that children of affectively ill mothers have earlierand significantly more depressive and disruptive behavior problems, and it islikely that a suboptimal mother–child relationship plays an important role inthis effect (Susman et al., 2001). On the basis of the substantial literature onanimals, one can expect maternal stress and alterations in human prenatalhormones to affect fetal development and later infant behavior. For example,one can hypothesize that infants born from pregnancies with elevated levels ofprenatal and/or postnatal stress will have a more reactive stress system andproblems with coping and adaptation, resulting in behavioral problems suchas aggression.
CONCLUSIONS
To avoid any misunderstanding, it is worth emphasizing that biological contri-butions to human behavior should not be regarded as deterministic. Rather,the hormonal environment may affect behavior by biasing the neural systemin such a way as to create behavioral dispositions. It is then that environmentand culture add their influence by rewarding, punishing, and/or ignoring be-haviors, thereby exaggerating, diminishing, distorting, or permitting the ex-pression of biologically based behaviors (Reinisch & Sanders, 1992). Aggres-sion is, of course, an optional behavioral strategy: As humans, we areexquisitely sensitive to our social environment and we are usually capable ofinhibiting this form of behavior, although biological factors may play a role inhow easy it is for us to do so.
Any consideration and reflection of the biological—in this case thehormonal—influence on aggression is controversial. On one hand, it is obvi-ous that individual differences in aggression originate, at least partly, in differ-ences in the biological substrate and the central nervous system. On the other
298 D E T E R M I N A N T S O F A G G R E S S I O N

hand, we find it hard to accept that these differences, and the way in whichthey are established, may also involve specific possibilities and restrictions. Inlight of this debate, it is important to remember that the evidence that hor-mones play a role in individual differences in aggression, certainly when wefocus on normal aggression, is small. Nevertheless, research on the influenceof biological factors on the origin and development of behavior in humans isimportant, not in order to demonstrate or reject the importance of biologicalfactors, but rather because these biological factors are part and parcel of aprocess of growth and development that can be influenced and disturbed innumerous ways.
NOTES
1. The Constantino et al. (1993) study provides some information about pre-adrenarchal plasmatestosterone values: in one study the mean T value for 3- to 5-year-olds was found to be 6.0 ±2.0 ng/dl; the mean T value for 6- to 8-year-olds was 8.0 ± 2.8 ng/dl (data published by Forest,1989); in another study by Lashansky, Saenger, & Fishman (1991) a mean T value for 1- to 5-year-olds of 1.98 ± 0.85 ng/dl and a mean T value for 6- to 12-year-olds of 6.8 ± 6.0 ng/dl wasobserved. Data from our own study on testosterone in 8- to 12-year-old boys found a mean Tvalue of 0.86 ± 1.5 nmol/l (range = 0.1–6.8), with a mean T value of 0.2 nmol/l in 8-year-oldboys (Van Goozen, Matthys, Cohen-Kettenis, Thijssen, & Van Engeland, 1998b). With respectto saliva, Scerbo and Kolko (1994) found a mean T level of 0.037 ng/ml in those younger than9 years old (n = 6), and a mean T level of 0.057 ng/ml in those between 9 and 14 years old (n =34). Clearly, T values in prepubertal children are low, and reliable norms for serum and salivaandrogen levels are needed.
2. Testosterone can be reliably detected in saliva using immunoassay in boys and girls ages 8 andolder (Granger, Schwartz, Booth, & Arentz, 1999). It is therefore unclear how seriously oneshould regard the testosterone values and findings in the study published by Sánchez-Martín etal. (2000).
3. It is beyond the scope of this chapter to discuss in any detail the possibility that the influence oftestosterone on aggression could be largely non-neural and indirect, and work primarilythrough its effects on the body (increased physical size, muscle mass, and appearance of sec-ondary sex characteristics) and activity. For example, Mazur and Booth (1998) conclude thattestosterone affects adolescent behavior mostly through indirect social responses elicited bymaturation rather than through direct activation of target receptors in organs or the nervoussystem. Similarly, the activity differences between girls and boys seem to be associated withplay and toy preferences that are at least partly influenced by testosterone (Hines et al., 2002),leading to sex segregation and to different socialization experiences in girls and boys.
REFERENCES
Achenbach, T. M. (1991). Manual for the Child Behavior Checklist and 1991 Profile.Burlington, VT: University of Vermont Department of Psychiatry.
Albert, D., Walsh, M., & Jonik, R. (1993). Aggression in humans: What is its biologicalfoundation? Neuroscience and Biobehavioral Reviews, 17, 405–425.
American Psychiatric Association. (1994). Diagnostic and statistical manual of mentaldisorders (4th ed.). Washington, DC: Author.
Anisman, H., Zaharia, M. D., Meaney, M. J., & Merali, Z. (1998). Do early-life events
Hormones and Aggression 299

permanently alter behavioral and hormonal responses to stressors? InternationalJournal of Developmental Neuroscience, 16, 149–164.
Archer, J. (1991). The influence of testosterone on human aggression. British Journal ofPsychology, 82, 1–28.
Archer, J., Graham-Kevan, N., & Davies, M. (in press). Testosterone and aggression: Are-analysis of Book, Starzyk and Quinsey (2001). Aggression and Violent Behavior.
Bakshi, V. P., & Kalin, N. H. (2000). Corticotropin-releasing hormone and animal mod-els of anxiety: Gene–environment interactions. Biological Psychiatry, 48, 1175–1198.
Berenbaum, S. A., Duck, S. C., & Bryk, K. (2000). Behavioral effects of prenatal versuspostnatal androgen excess in children with 21–hydroxylase-deficient congenital ad-renal hyperplasia. Journal of Clinical Endocrinology and Metabolism, 85(2), 727–733.
Berenbaum, S. A., & Resnick, S. M. (1997). Early androgen effects on aggression in chil-dren and adults with congenital adrenal hyperplasia. Psychoneuroendocrinology,22(7), 505–515.
Berkowitz, L. (1989). Frustration–aggression hypothesis: Examination and reformula-tion. Psychological Bulletin, 106, 59–73.
Brendgen, M., Vitaro, F., Tremblay, R. E., & Lavoie, F. (2001). Reactive and proactive ag-gression: Predictions to physical violence in different contexts and moderating ef-fects of parental monitoring and caregiving behavior. Journal of Abnormal ChildPsychology, 29, 293–304.
Brennan, P. A., Raine, A., Schulsinger, F., Kirkegaard-Sorensen, L., Knop, J., Hutchings,B., et al. (1997). Psychophysiological protective factors for male subjects at highrisk for criminal behavior. American Journal of Psychiatry, 154, 853–855.
Carlson, M., & Earls, F. (1996). Psychological and neuroendocrinological sequelae ofearly social deprivation in institutionalized children in Romania. Annals of the NewYork Academy of Sciences, 807, 419–428.
Clarke, D., Fearon, U., Cunningham, S. K., & McKenna, T. J. (1996). The steroidogeniceffects of b- endorphin and joining peptide: A potential role in the modulation of ad-renal androgen production. Journal of Endocrinology, 151, 301–307.
Coie, J. D., & Dodge, K. A. (1998). Aggression and antisocial behavior. In W. Damon &N. Eisenberg (Eds.), Handbook of child psychology: Social, emotional, and person-ality development (pp. 779–862). New York: Wiley.
Collaer, M. L., & Hines, M. (1995). Human behavioural sex differences: A role forgondal hormones during early development? Psychological Bulletin, 118, 55–107.
Constantino, J. N., Grosz, D., Saenger, P., Chandler, D. W., Nandi, R., & Earls, F. J.(1993). Testosterone and aggression in children. Journal of the Academy of Childand Adolescent Psychiatry, 32, 1217–1222.
Dabbs, J. M., Frady, R. L., Carr, T. S., & Besch, N. F. (1987). Saliva testosterone and crim-inal violence in young adult prison inmates. Psychosomatic Medicine, 49, 174–182.
Dawes, M. A., Dorn, L. D., Moss, H. B., Yao, J. K., Kirisci, L., Ammerman, R. T., &Tarter, R. E. (1999). Hormonal and behavioral homeostasis in boys at risk for sub-stance abuse. Drug and Alcohol Dependence, 55, 165–176.
Dettling, A. C., Gunnar, M. R., & Donzella, R. (1999). Cortisol levels of young childrenin full-day childcare centers: Relations with age and temperament. Psychoneuro-endocrinology, 24, 519–536.
Dixson, A. F. (1980). Androgens and aggressive behavior in primates: A review. Aggres-sive Behavior, 6, 37–67.
300 D E T E R M I N A N T S O F A G G R E S S I O N

Dodge, K. A., Lochman, J. E., Harnish, J. D., Bates, J. E., & Pettit, G. S. (1997). Reactiveand proactive aggression in school children and psychiatrically impaired chroni-cally assaultive youth. Journal of Abnormal Child Psychology, 106, 37–51.
Eberhart, J. A., Keverne, E. B., & Meller, R. E. (1980). Social influences on plasma testos-terone levels in male Talapoin monkeys. Hormones and Behavior, 14, 247–266.
Ehrhardt, A. A., & Meyer-Bahlburg, H. F. L. (1981). Effects of prenatal sex hormones ongender-related behavior. Science, 211, 1312–1318.
Ehrhardt, A. A., Meyer-Bahlburg, H. F. L., Rosen, L. R., Feldman, J. F., Veridiano, N. P.,Zimmerman, I., & McEwen, B. S. (1985) Sexual orientation after prenatal exposureto exogeneous estrogen. Archives of Sexual Behavior, 14, 57–75.
Finkelstein, J. W., Susman, E. J., Chinchilli, V. M., Kunselman, S. J., D’Arcangelo, M. R.,Schwab, J., Demers, L. M., Liben, L. S., Lookingbill, G., & Kulin, H. E. (1997). Es-trogen or testosterone increases self-reported aggressive behaviors in hypogonadaladolescents. Journal of Clinical Endocrinology and Metabolism, 82(8), 2433–2438.
Forest, M. G. (1989). Physiological changes in circulating androgens. Pediatric Adoles-cent Endocrinology, 19, 104–129.
Francis, D. D., Caldji, C., Champagne, F., Plotsky, P. M., & Meaney, M. J. (1999). Therole of corticotropin-releasing factor-norepinephrine systems in mediating the ef-fects of early experience on the development of behavioral and endocrine responsesto stress. Biological Psychiatry, 46, 1153–1166.
Francis, D. D., Diorio, J. F., Plotsky, P. M., & Meaney, M. J. (2002). Environmental en-richment reverses the effects of maternal separation on stress reactivity. Journal ofNeuroscience, 22(18), 7840–7843.
Francis, D. D., Szegda, K., Campbell, G., Martin, D., & Insel, T. R. (2003). Epigeneticsources of behavioral differences in mice. Nature Neuroscience, 6(5), 445–446.
Gerra, G., Zaimovic, A., Avanzini, P., Chittolini, B., Giucastro, G., Caccavari, R.,Palladino, M., Maestri, D., Monica, C., Delsignore, R., & Brambilla, F. (1997).Neurotransmitter-neuroendocrine responses to experimentally induced aggressionin humans: Influence of personality variable. Psychiatry Research, 66, 33–43.
Gladue, B. A. (1991). Aggressive behavioral characteristics, hormones, and sexual orien-tation in men and women. Aggressive Behavior, 17, 313–326.
Goodyer, I. M., Herbert, J., Altham, P. M. E., Pearson, J., Secher, S., & Shiers, S. (1996).Adrenal secretion during major depression in 8 to 16 year olds: I. Altered diurnalrhythms in salivary cortisol and dehydroepiandrosterone (DHEA) at presentation.Psychological Medicine, 26, 245–256.
Gooren, L., Fliers, E., & Courtney, K. (1990). Biological determinants of sexual orienta-tion. Annual Review of Sex Research, 1, 175–196.
Gottlieb, G., & Halpern, C. T. (2002). A relational view of causality in normal and ab-normal development. Development and Psychopathology, 14(3), 421–435.
Goy, R. W., & McEwen, B. S. (1980). Sexual differentiation of the brain. Cambridge,MA: MIT Press.
Granger, D. A., Schwartz, E. B., Booth, A., & Arentz, M. (1999). Salivary testosterone de-termination in studies of child health and development. Hormones and Behavior,35, 18–27.
Granger, D. A., Shirtcliff, E. A., Zahn-Waxler, C., Usher, B., Klimes-Dougan, B., &Hastings, P. (2003). Salivary testosterone diurnal variation and psychopathology inadolescent males and females: Individual differences and developmental effects. De-velopment and Psychopathology, 15, 431–449.
Hormones and Aggression 301

Gunnar, M. R., Tout, K., de Haan, M., Pierce, S., & Stansbury, K. (1997). Temperament,social competence, and adrenocortical activity in preschoolers. DevelopmentalPsychobiology, 31, 65–85.
Halpern, C. T., Udry, R., Campbell, B., & Suchindran, C. (1994). Relationships betweenaggression and pubertal increases in testosterone: A panel analysis of adolescentmales. Social Biology, 40, 8–24.
Harris, J. A. (1999). Review and methodological considerations in research on testoster-one and aggression. Aggression and Violent Behavior, 4(3), 273–291.
Higley, J. D., Mehlman, P. T., Poland, R. E., Taub, D. M., Vickers, J., Suomi, S. J., &Linnoila, M. (1996). CSF testosterone and 5–HIAA correlate with different types ofaggressive behaviors. Biological Psychiatry, 40, 1067–1082.
Hines, M., Golombok, S., Rust, J., Johnston, K. J., Golding, J., & the Avon LongitudinalStudy of Parents and Children Study Team. (2002). Testosterone during pregnancyand gender role behavior of preschool children: A longitudinal population study.Child Development, 73(6), 1678–1687.
Hines, M., & Green, R. (1991). Human hormonal and neural correlates of sex-typed be-haviors. Review of Psychiatry, 10, 536–555.
Kagan, J., Reznick, J. S., & Snidman, N. (1988). Biological bases of childhood shyness.Science, 240, 167–173.
Kaufman, J., Plotsky, P. M., Nemeroff, C. B., & Charney, D. S. (2000). Effects of early ad-verse experiences on brain structure and function: Clinical implications. BiologicalPsychiatry, 48, 778–790.
King, R. J., Jones, J., Scheurer, J. W., Curtis, D., & Zarcone, V. P. (1990). Plasma-cortisolcorrelates of impulsivity and substance abuse. Personality and Individual Differ-ences, 11, 287–291.
Klimes-Dougan, B., Hastings, P. D., Granger, D. A., Usher, B. A., & Zahn-Waxler, C.(2001). Adrenocortical activity in at-risk and normally developing adolescents: In-dividual differences in salivary cortisol basal levels, diurnal variation, and responsesto social challenges. Development and Psychopathology, 13, 695–719.
Kruesi, M. J. P., Rapoport, J. L., Hamburger, S., Hibbs, E., Potter, W. Z., Lenane, M., &Brown, G. L. (1990). Cerebrospinal fluid monoamine metabolites, aggression, andimpulsivity in disruptive behavior disorders of children and adolescents. Archives ofGeneral Psychiatry, 47, 419–426.
Kruesi, M. J. P., Schmidt, M. E., Donnelly, M., Hibbs, E. D., & Hamburger, S. D. (1989).Urinary free cortisol output and disruptive behavior in children. Journal of theAmerican Academy of Child and Adolescent Psychiatry, 28, 441–443.
Lahey, B. B., Waldman, I. D., & McBurnett, K. (1999). Annotation: The development ofantisocial behavior: An integrative causal model. Journal of Child Psychology andPsychiatry, 40, 669–682.
Lashansky, G., Saenger, P., & Fishman, K. (1991). Normative data for adrenal steroido-genesis in a healthy pediatric population: Age and sex-related changes after ACTHstimulation. Journal of Endocrinology and Metabolism, 73, 674–686.
Liu, D., Diorio, J., Tannenbaum, B., Caldji, C., Francis, D., Freedman, A., Sharma, S.,Pearson, D., Plotsky, P. M., & Meany, M. J. (1997). Maternal care, hippocampalglucocorticoid receptors, and hypothalamic-pituitary-adrenal responses to stress.Science, 277, 1659–1662.
Loeber, R., Green, S. M., Lahey, B. B., & Stouthamer-Loeber, M. (1991). Differences andsimilarities between children, mothers, and teachers as informants on disruptivechild behavior. Journal of Abnormal Child Psychology, 19, 75–95.
302 D E T E R M I N A N T S O F A G G R E S S I O N

Majewska, M. D. (1992). Neurosteroids: Endogeneous bimodal modulators of theGABAA receptor: Mechanisms of action and physiological significance. Progress inNeurobiology, 38, 379–395.
Matthys, W., Van Goozen, S. H. M., De Vries, H., Cohen-Kettenis, P. T., & Van Engeland,H. (1998). The dominance of behavioral activation over behavioral inhibition inconduct disordered boys with or without attention deficit hyperactivity disorder.Journal of Child Psychology and Psychiatry, 39, 643–651.
Mazur, A., & Booth, A. (1998). Testosterone and dominance in men. Behavioral andBrain Sciences, 21, 353–397.
McBurnett, K., Lahey, B. B., Frick, P. J., Risch, C., Loeber, R., Hart, E. L., Christ, M. A.G., & Hanson, K. S. (1991). Anxiety, inhibition, and conduct disorder in children:II. Relation to salivary cortisol. Journal of the American Academy of Child and Ad-olescent Psychiatry, 30, 192–196.
McBurnett, K., Lahey, B. B., Rathouz, P. J., & Loeber, R. (2000). Low salivary cortisoland persistent aggression in boys referred for disruptive behavior. Archives of Gen-eral Psychiatry, 57, 38–43.
Meyer-Bahlburg, H. F. L., Boon, D. A., Sharma, M., & Edwards, J. A. (1974). Aggres-siveness and testosterone measures in man. Psychosomatic Medicine, 36, 269–274.
Moss, H. B., Vanyukov, M. M., & Martin, C. S. (1995). Salivary cortisol responses andthe risk for substance abuse in prepubertal boys. Biological Psychiatry, 38, 547–555.
Moyer, K. E. (1976). The psychobiology of aggression. New York: Harper & Row.O’Connor, D. B., Archer, J., Hair, W. M., & Wu, F. C. W. (2002). Exogenous testosterone,
aggression, and mood in eugonadal and hypogonadal men. Physiology and Behav-ior, 75, 557–566.
Olweus, D., Mattsson, A., Schalling, D., & Low, H. (1988). Circulating testosterone lev-els and aggression in adolescent males: A causal analysis. Psychosomatic Medicine,50, 261–272.
Paikoff, R., & Brooks-Gunn, J. (1990). Physiological processes: What role do they playduring the transition to adolescence? In P. Montemayor, G. Adams, & T. Gullota(Eds.), Advances in adolescent development: Transition from childhood to adoles-cence (Vol. 2, pp. 63–81). Newbury Park, CA: Sage.
Pajer, K., Gardner, W., Rubin, R. T., Perel, J., & Neal, S. (2001). Decreased cortisol levelsin adolescent girls with conduct disorder. Archives of General Psychiatry, 58, 297–302.
Raine, A. (1996). Autonomic nervous system activity and violence. In D. M. Stoff & R. B.Cairns (Eds.), Aggression and violence (pp. 145–168). Mahwah, NJ: Erlbaum,
Reinisch, J. M. (1981). Prenatal exposure to synthetic progestins increases potential foraggression in humans. Science, 211, 561–562.
Reinisch, J. M., & Sanders, S. A. (1992). Prenatal hormonal contributions to sex differ-ences in human cognitive and personality development. In A. A. Gerall, H. Moltz,& I. L. Ward (Eds.), Handbook of behavioral neurobiology: Sexual differentiation(Vol. 11, pp. 221–243). New York: Plenum Press.
Robel, P., & Baulieu, E. E. (1995). Dehydroepiandrosterone (DHEA) is a neuroactiveneurosteroid. Annals of the New York Academy of Sciences, 774, 82–110.
Sánchez-Martín, J., Fano, E., Ahedo, L., Cardas, J., Brain, P., & Azpíroz, A. (2000). Re-lating testosterone levels and free play social behavior in male and female preschoolchildren. Psychoneuroendocrinology, 25, 773–783.
Hormones and Aggression 303

Sapolsky, R. M. (1997). The importance of a well-groomed child. Science, 277, 1620–1621.
Scerbo, A. S., & Kolko, D. J. (1994). Salivary testosterone and cortisol in disruptivechildren: Relationship to aggressive, hyperactive and internalizing behaviors.Journal of the American Academy of Child and Adolescent Psychiatry, 33, 1174–1184.
Schaal, B., Tremblay, R., Soussignan, R., & Susman, E. (1996). Male testosterone linkedto high social dominance but low physical aggression in early adolescence. Journalof the Academy of Child and Adolescent Psychiatry, 34, 1322–1330.
Schulz, K. P., Halperin, J. M., Newcorn, J. H., Sharma, V., & Gabriel, S. (1997). Plasmacortisol and aggression in boys with ADHD. Journal of the American Academy ofChild and Adolescent Psychiatry, 36, 605–609.
Spangler, G., & Grossmann, K. E. (1993). Biobehavioral organization in securely and in-securely attached infants. Child Development, 64(5), 1439–1450.
Spangler, G., & Scheubeck, R. (1993). Behavioral organization in newborns and itsrelation to adrenocortical and cardiac activity. Child Development, 64(2), 622–633.
Stoff, D. M., Pasatiempo, A. P., Yeung, J., Cooper, T. B., Bridger, W. H., & Rabinovich, H.(1992). Neuroendocrine responses to challenge with dl-fenfluramine and aggres-sion in disruptive behavior disorders of children and adolescents. Psychiatry Re-search, 43, 263–276.
Susman, E. J., Dorn, L. D., Inoff-Germain, G., Nottelmann, E. D., & Chrousos, G. P.(1997). Cortisol reactivity, distress behavior, and behavioral and psychologicalproblems in young adolescents: A longitudinal perspective. Journal of Research onAdolescence, 7, 81–105.
Susman, E. J., Inoff-Germain, G., Nottelmann, E. D., Loriaux, D., Cutler, G., &Chrousos, G. P. (1987). Hormones, emotional dispositions, and aggressive attrib-utes in young adolescents. Child Development, 58, 1114–1134.
Susman, E. J., Schmeelk, K. H., Ponirakis, A., & Gariepy, J. L. (2001). Maternal prenatal,postpartum, and concurrent stressors and temperament in 3–year-olds: A personand variable analysis. Development and Psychopathology, 13, 629–652.
Susman, E. J., Schmeelk, K. H., Worrall, B. K., Granger, D. A., Ponirakis, A., &Chrousos, G. P. (1999). Corticotropin-releasing hormone and cortisol: Longitudi-nal associations with depression and antisocial behavior in pregnant adolescents.Journal of the American Academy of Child and Adolescent Psychiatry, 38, 460–467.
Tennes, K., & Kreye, M. (1985). Children’s adrenocortical responses to classroom activi-ties and tests in elementary school. Psychosomatic Medicine, 47, 451–460.
Tennes, K., Kreye, M., Avitable, N., & Wells, R. (1986). Behavioral correlates of excretedcatecholamines and cortisol in second-grade children. Journal of the AmericanAcademy of Child and Adolescent Psychiatry, 25, 764–770.
Tremblay, R. E. (2000). The development of aggressive behaviour during childhood:What have we learned in the past century? International Journal of Behavioral De-velopment, 24, 129–141.
Tremblay, R. E. (2003). Why socialization fails: The case of chronic physical aggression.In B. B. Lahey, T. E. Moffitt, & A. Caspi (Eds.), Causes of conduct disorder and juve-nile delinquency (pp. 182–224). New York: Guilford Press.
Van de Beek, C., Thijssen, J. H. H., Cohen-Kettenis, P. T., Van Goozen, S. H. M., &Buitelaar, J. K. (2004). Sex steroids in pregnant women: Relationships between hor-
304 D E T E R M I N A N T S O F A G G R E S S I O N

mones assessed in amniotic fluid, and maternal and umbilical cord serum. Hor-mones and Behavior, 46(5), 663–669.
Van de Poll, N. E., & Van Goozen, S. H. M. (1992). Hypothalamic involvement in sexu-ality and hostility: Comparative psychological aspects. Progress in Brain Research,93, 343–361.
Van de Poll, N. E., Van Zanten, S., & De Jonge, F. H. (1986) Effects of testosterone, estro-gen and dihydrotestosterone upon aggressive and sexual behavior of female rats.Hormones and Behavior, 20, 418–431.
Van Goozen, S. H. M., Cohen-Kettenis, P. T., Gooren, L. J. G., Frijda, N. H., & Van dePoll, N. E. (1995). Gender differences in behaviour: Activating effects of cross-sexhormones. Psychoneuroendocrinology, 20, 343–363.
Van Goozen, S. H. M., Cohen-Kettenis, P. T., Snoek, H., Matthys W., Swaab-Barneveld,H., & Van Engeland, H. (2004). Executive functioning in children: A comparison ofhospitalized ODD and ODD/ADHD children and normal controls. Journal ofChild Psychology and Psychiatry, 45(2), 284–292.
Van Goozen, S. H. M., Frijda, N. H., & Van de Poll, N. E. (1994). Anger and aggressionin women: Influence of sports choice and testosterone administration. AggressiveBehavior, 20, 213–222.
Van Goozen, S. H. M., Frijda, N. H., Wiegant, V. M., Endert E., & Van de Poll, N. E.(1996). The premenstrual phase and reactions to aversive events: A study of hor-monal influences on emotionality. Psychoneuroendocrinology, 21, 479–497.
Van Goozen, S. H. M., Matthys, W., Cohen-Kettenis, P. T., Buitelaar, J. K., & VanEngeland, H. (2000a). Hypothalamic–pituitary–adrenal axis and autonomic ner-vous system activity in disruptive children and matched controls. Journal of theAmerican Academy of Child and Adolescent Psychiatry, 39, 1438–1445.
Van Goozen, S. H. M., Matthys, W., Cohen-Kettenis, P. T., Gispen-deWied, C., Wiegant,V. M., & Van Engeland, H. (1998a). Salivary cortisol and cardiovascular activityduring stress in oppositional defiant disorder boys and normal controls. BiologicalPsychiatry, 43, 531–539.
Van Goozen, S. H. M., Matthys, W., Cohen-Kettenis, P. T., Thijssen, J. H. H., & VanEngeland, H. (1998b). Adrenal androgens and aggression in prepubertal boys. Bio-logical Psychiatry, 43, 156–158.
Van Goozen, S. H. M., Van den Ban, E., Matthys, W., Cohen-Kettenis, P. T., Thijssen, J.H. H., & Van Engeland, H. (2000b). Increased adrenal androgen functioning inchildren with oppositional defiant disorder: A comparison with psychiatric andnormal controls. Journal of the American Academy of Child and Adolescent Psychi-atry, 39(11), 1446–1451.
Vanyukov, M. M., Moss, H. B., Plail, J. A., Blackson, T., Mezzich, A. C., & Tarter, R. E.(1990). Antisocial symptoms in preadolescent boys and in their parents: Associa-tions with cortisol. Psychiatry Research, 46, 9–17.
Virkkunen, M. (1985). Urinary free cortisol secretion in habitually violent offenders.Acta Psychiatrica Scandinavica, 72, 40–44.
Vitiello, B., & Stoff, D. M. (1997). Subtypes of aggression and their relevance to childpsychiatry. Journal of the American Academy of Child and Adolescent Psychiatry,36(3), 307–315.
Warren, M. P., & Brooks-Gunn, J. (1989). Mood and behavior at adolescence: Evidencefor hormonal factors. Journal of Clinical Endocrinology and Metabolism, 69, 77–83.
Wolf, O. T., & Kirschbaum, C. (1999). Actions of dehydroepiandrosterone and its sulfate
Hormones and Aggression 305

in the central nervous system: Effects on cognition and emotion in animals and hu-mans. Brain Research Review, 30(3), 264–288.
Woodman, D. D., Hinton, J. W., & O’Neill, M. T. (1978). Cortisol secretion and stress inmaximum security hospital patients. Journal of Psychosomatic Research, 22, 133–136.
Zuckerman, M. (1991). Psychobiology of personality. Cambridge, MA: Cambridge Uni-versity Press.
306 D E T E R M I N A N T S O F A G G R E S S I O N

DETERMINANTS OF AGGRESSIONExecutive Function in Aggression
15
Executive Functionin Early Physical Aggression
JEAN RICHARD SÉGUIN and PHILIP DAVID ZELAZO
Physical aggression is characteristic of early childhood but typically becomesless frequent with development (Tremblay et al., 1996). During this sameperiod of their development, there are marked age-related increases in chil-dren’s executive function (Zelazo & Müller, 2002). Executive function gener-ally refers to the self-regulation of thought, action, and emotion—processesthat depend on the integrity of neural systems involving prefrontal cortex (e.g.Owen et al., 1999; Robbins, 1996; Stuss, 1992). We propose that this patternof declining aggression and increasing executive function reflects the fact that,during development, children typically acquire the executive function skillsneeded to regulate physical aggression. Moreover, we propose that in thosecases (about 4–6% of the population) where children’s levels of physical ag-gression remain high (Nagin & Tremblay, 1999; Tremblay et al., 2004), thedevelopment of executive function will be atypical.
The literature is replete with studies that have examined the relation be-tween global cognitive measures and global indices of antisocial or external-izing behavior problems. To date, however, very few studies have tested thespecific hypothesis that executive function development is particularly rele-vant to the regulation of physical aggression. Executive function is a specificaspect of cognitive function that is rarely captured by global measures. Simi-larly, physical aggression is a specific aspect of aggression that may not becaptured by externalizing behavior problems or global scales of aggression,disruptiveness, antisocial behavior, or delinquency. Thus, a test of the hypoth-
307

esis requires taking into account (1) that executive function is a specific subsetof more global cognitive abilities and (2) that physical aggression is a specificbehavior within externalizing, aggressive, disruptive, antisocial behavior prob-lems or delinquency.
PHYSICAL AGGRESSION AMONG GLOBAL MEASURES OF BEHAVIOR PROBLEMS
Physical aggression can be found in the form of hitting, kicking, biting, as-saults, confrontation with a weapon, getting involved in fights, threatening touse physical force, rape, and homicide. Physical aggression is also studied indi-rectly, via its relation to various externalizing disorders, and we now knowfrom retrospective and longitudinal studies that externalizing disorders begina chronic course in early childhood (Tremblay et al., 1999). Physically aggres-sive behavior is found in conduct disorder (CD), and it is related to, but not adiagnostic feature of, attention-deficit/hyperactivity disorder (ADHD) andoppositional defiant disorder (ODD). ADHD itself can be subdivided into asubtype in which attention deficits predominate and another subtype charac-terized by hyperactive and impulsive behavior (Milich, Balentine, & Lynam,2001). Within ADHD, it is the hyperactive–impulsive subtype that is morestrongly associated with ODD and CD (Willcutt, Pennington, Chhabildas,Friedman, & Alexander, 1999), and possibly with physical aggression, al-though early ADHD behavior problems are not typically related to laterviolence (Farrington & Loeber, 2000; Lahey, Loeber, Burke, Rathouz, &McBurnett, 2002; Loeber, Burke, & Lahey, 2002; Nagin & Tremblay, 1999;Stouthamer-Loeber, Loeber, Wei, Farrington, & Wikström, 2002). CD, ADHD,and ODD co-occur at a greater than chance level, and this has been a growingsource of concern in understanding the development of externalizing disorders(Lilienfeld, 2003).
In a longitudinal study of a community sample of more than 1,000 boysfrom low-socioeconomic-status (SES) schools (Séguin, Boulerice, Harden,Tremblay, & Pihl, 1999), close to 90% of adolescents who met criteria forADHD or CD between the ages of 14 and 16 were found to have had a his-tory of occasional or chronic physically aggressive behavior (based on teacherratings of physical aggression between the ages of 6 and 12 years). This is incontrast to 61% of boys without such a diagnosis. It should be noted, how-ever, that although a history of physical aggression during childhood is rela-tively common in most adolescents with ADHD or CD, only about a quarterof boys with a history of physical aggression meet the criteria for ADHD orCD (Séguin et al., 1999).
Finally, besides externalizing disorders, physical aggression is also foundamong other global categories such as “disruptive disorders,” in children de-fined as “hard to manage,” among “antisocial disorders,” and in “delin-quency” and “criminality.”
308 D E T E R M I N A N T S O F A G G R E S S I O N

Developmental Course of Physical Aggression
Research on the developmental course of physical aggression per se suggests(Robins, 1991) that the frequency of physical aggression actually peaks ataround 2 years of age (See Tremblay & Nagin, Chapter 5, this volume). Mostchildren who begin with high levels of physical aggression gradually desist atvarious rates. However, there is a small but significant proportion of children(about 4–6%) for whom frequencies of physical aggression remain very highthroughout childhood and adolescence (Broidy et al., 2003). We propose thatthese children will display early impairments in executive function.
EXECUTIVE FUNCTION AND ITS DEVELOPMENT
Executive function is a popular topic in contemporary research, but defini-tions of executive function differ widely. A traditional approach is to treat ex-ecutive function as a higher order cognitive mechanism or ability, as whenDenckla and Reiss (1997) suggest that “executive function refers to a cogni-tive module consisting of effector output elements involving inhibition, work-ing memory, and organizational strategies necessary to prepare a response”(p. 283, emphasis in original). Unfortunately, this approach (see also Baddeley& Della Sala, 1996; Norman & Shallice, 1986) essentially invokes a homun-culus, or little man in the head, rather than explaining in detail how executivefunction, including inhibition, working memory, and strategy use, is accom-plished (Parkin, 1998; Zelazo & Müller, 2002).
An alternative and more common approach is more empirically driven.This approach involves devising comprehensive neuropsychological batteriesand using factor analysis to reveal the underlying structure of executive func-tion (see Zelazo & Müller, 2002, for a review). Such studies generally revealthree or four factors—a result that has been taken to suggest that there aredissociable dimensions of executive function, consistent with efforts to “frac-tionate” executive function based on lesion studies in nonhuman animals(Robbins, 1996). Although the results of factor-analytic studies are potentiallymisleading, because providing labels for factors may give the erroneous im-pression that researchers actually understand the cognitive processes underly-ing performance on various tasks, this approach does suggest that there areboth common and diverse aspects of the measures that have been used to as-sess executive function (Miyake et al., 2000).
Research on executive function has demonstrated that executive functionfirst emerges early in development, probably around the end of the first yearof life, and it develops across a wide range of ages, with important changes oc-curring between about 2 and 5 years of age. Adult-level performance on manystandard tests of executive function is reached at about 12 years of age, andperformance on some measures continues to change into adulthood (see
Executive Function in Aggression 309

Zelazo & Müller, 2002, for review). One of the most widely studied examplesof executive function in infants is performance on the A-not-B task. In thistask, an infant watches as an object is hidden at one location (A). Following abrief delay (e.g., 3 seconds), the infant is permitted to search for the object.After a number of A trials, the object is then hidden conspicuously at a differ-ent location (B). Despite witnessing the hiding event at B, most 9-month-oldscontinue to search perseveratively at A on the first B trial, thereby committingthe A-not-B error (for a recent meta-analysis, see Marcovitch & Zelazo,1999). However, by about 12 months of age, most infants search flexibly onthis task.
The changes occurring between about 2 and 5 years have been carefullydocumented using a variety of measures of rule use. The ability to use a rulesystematically to control behavior seems to be acquired at about 2.5 years ofage. However, children at this age are highly susceptible to perseverative re-sponding, and it is not until 3 years that most children are able to use tworules simultaneously (Luria, 1973; Zelazo, Frye, & Rapus, 1996a; Zelazo &Reznick, 1991). Then, although 3-year-olds can use a pair of rules, it not untilabout 5 years that most children can switch flexibly between two incompati-ble pairs of rules, or two different perspectives on a situation, as seen in tasksused in assessing causal reasoning (e.g., Frye, Zelazo, Brooks, & Samuels,1996; Zelazo, Reznick, & Piñon, 1995), behavioral prediction (Zelazo,Helwig, & Lau, 1996b), card sorting (e.g., Zelazo et al., 1996a), and understand-ing false beliefs (see Wellman, Cross, & Watson, 2001, for a meta-analysis).
The Wisconsin Card Sorting Test (WCST) has frequently been used to ex-plore the development of executive function beyond about 5 years of age. Inthe WCST (Grant & Berg, 1948; Robinson, Heaton, Lehman, & Stilson,1980), participants are presented with four target cards that differ on three di-mensions (number, color, and shape) and asked to sort a series of test cardsthat match different target cards on different dimensions. Participants mustdiscover the sorting rule by trial and error, and after a certain number of con-secutive correct responses, the sorting rule is changed. In a representativestudy, Chelune and Baer (1986) documented a linear increase in performanceon the WCST between the ages of 6 and 10 years, with 6-year-olds performingat the level of patients with prefrontal cortical damage (e.g., Milner, 1963)and 10-year-olds performing like healthy adults. This pattern was obtainedfor the three variables assessed: number of perseverative errors, number ofcategories achieved, and failures to maintain set.
THE RELATION OF EXECUTIVE FUNCTIONTO EXTERNALIZING OR ANTISOCIAL DISORDERS
Most research on the relation between executive function and physical aggres-sion has relied on relatively global measures of both executive function andaggression. In most cases, the measures of executive function correspond to
310 D E T E R M I N A N T S O F A G G R E S S I O N

standard neuropsychological tests that are either taken directly from the adultliterature or adapted from adult tasks. For example, several studies have usedthe WCST. The WCST taps numerous aspects of executive function, and, as aresult, the origin of errors on this task is difficult to determine (e.g., see Delis,Squire, Bihrle, & Massman, 1992). To perform correctly on the WCST, onemust first construct a representation of the problem space, which includesidentifying the relevant dimensions. Then, one must choose a promisingplan—for example, sorting according to shape. After selecting a plan, onemust (1) keep the plan in mind long enough for it to guide one’s thought or ac-tion and (2) actually carry out the prescribed behavior. Keeping a plan in mindto control behavior is referred to as intending; translating a plan into action isrule use. Finally, after acting, one must evaluate one’s behavior, which includesboth error detection and error correction. Inflexibility can occur at eachphase, so there are several possible explanations of perseverative performanceon the WCST—and on global executive function tasks more generally. For ex-ample, perseveration can occur after a rule change in the WCST either becausea new plan was not formed (one type of representational inflexibility; Zelazoet al., 1995) or because the plan was formed but not carried out (an exampleof lack of response control; Zelazo et al., 1995).
Nonetheless, research using the WCST has proven useful in documentingthat executive function is diminished in groups of children who are likely highin physical aggression. Indeed, relatively high levels of perseveration on theWCST have been documented in individuals with antisocial behavior (Kandel& Freed, 1989), in children with externalizing behavior problems (Matson &Fisher, 1991), in children with ADHD (Pennington, 1997), and in adolescentswith CD (Toupin, Déry, Pauzé, Mercier, & Fortin, 2000). Performance on theWCST was also related to childhood social abilities (Bonino & Cattelino,1999). Perseveration on other card-playing tasks that also involve error detec-tion and correction has also been shown in adult psychopathic populations(Newman & Wallace, 1993) and in our work on adolescents with a well-documented history of physical aggression (Séguin, Arseneault, Boulerice,Harden, & Tremblay, 2002).
Several studies along these lines have shown that poor executive functionis involved in ADHD and antisocial disorders. These have recently been re-viewed, respectively, by Pennington and Ozonoff (1996) and Morgan andLilienfeld (2000). This latter review covered studies published between 1942and 1997 for ages 13–40 years and revealed a medium-sized effect (meanstandard difference) of about 0.6 in comparing antisocial and nonantisocialgroups. Unfortunately, however, the review could not focus on violentantisociality because most studies lacked information about violence. This isindeed a serious problem in the field (Tremblay, 2000). Further, Morgan andLilienfeld (2000) and Pennington and Ozonoff (1996) point out that the stud-ies they reviewed have typically not taken into account the comorbidity be-tween ADHD and antisocial disorders such as CD. A third recent review fur-ther examined problems associated with tests for the specificity of executive
Executive Function in Aggression 311

function to the three externalizing disorders, and to Tourette’s syndrome andhigh-functioning autism (Sergeant, Geurts, & Oosterlaan, 2002).
The problem of examining physical aggression per se (instead of antiso-cial disorders or CD/ODD) has been addressed, however, in several recentstudies that have supported the hypothesis that executive function problemsare related to physical aggression when observable physical aggression wasentered into statistical models even after controlling for ADHD (see Giancola,Mezzich, & Tarter, 1998; Séguin et al., 1999; Toupin et al., 2000). The failureto take into account physical aggression and hyperactivity at the outset hastypically resulted in a weaker assessment of one or the other behavior prob-lem. In our earlier work we found that boys from a community sample with ahistory of physical aggression performed most poorly on tests of executivefunction even after controlling for verbal and spatial abilities as well as cere-bral dominance (Séguin, Pihl, Harden, Tremblay, & Boulerice, 1995). In thatstudy and others that followed, the core executive function tasks were derivedfrom the neuropsychological literature and had been validated with brain im-aging techniques. The Self-Ordered Pointing task and a number randomiza-tion task involved primarily rule use, and Conditional Association tasks alsoinvolved error detection and correction in addition to rule use (Petrides,Alivisatos, Evans, & Meyer, 1993a; Petrides, Alivisatos, Meyer, & Evans,1993b). In addressing a question raised by Pennington and Ozonoff (1996)about that study, we further controlled for chronic ADHD. We also added acontrol for IQ and still found poor executive function in the physically aggres-sive boys (Séguin et al., 1999). Moreover, in an attempt to understand morefully the relation between executive function and physical aggression andhyperactivity, we selected study participants based on the histories of both be-havior problems. We chose to use the narrow-band behavior of hyperactivityinstead of the broader-band construct of ADHD because hyperactivity wasfound to attenuate the relation between physical aggression and card persev-eration (Séguin et al., 2002), although ADHD, as a whole, was not related toexecutive function in our previous study (see Séguin et al., 1999). Using thismethodology also allowed testing for additive and interactive effects of physi-cal aggression and hyperactivity on cognitive function in general and execu-tive function in particular (Waschbusch, 2002). We did not find an interac-tion, but did find clear additive effects of both behavior problems on executivefunction, although the effects were clearer for physical aggression than theywere for hyperactivity (Séguin, Nagin, Assaad, & Tremblay, 2004).
THE RELATION OF PHYSICAL AGGRESSIONAND EXECUTIVE FUNCTION IN EARLY CHILDHOOD
Given that, for most children, the decline in physical aggression occurs be-tween ages 24 and 60 months (see Tremblay & Nagin, Chapter 5, this vol-ume) and that this period is marked by major changes in executive function, it
312 D E T E R M I N A N T S O F A G G R E S S I O N

seems especially important to examine the relation between aggression andexecutive function during the preschool years. Early identification of potentialproblems also has implications for the success of any interventions, becauseearly childhood may be a period of relative plasticity and it may be easier toalter behavior before it becomes entrenched.
Studies of this relation in preschoolers have addressed a broad range ofrelevant issues. Some studies focused more specifically on the relation betweenearly hyperactivity and aspects of cognitive function relevant to, but not iso-morphic with, executive function. For example, one community study foundthat three-quarters of the 2% of children who had hyperactivity problems andwere “very hard to manage” in preschool still had problems at age 15(McGee, Partridge, Williams, & Silva, 1991). They were worse off as adoles-cents on both behavioral and cognitive measures than preschoolers who wereonly “very hard to manage” (3% of that sample) and developmental controlchildren (95% of that sample). However, it is not clear whether early poorcognitive abilities contributed to the maintenance of the behavior problems.Other studies of preschool hyperactivity have also failed to take into accountconcurrent conduct problems, let alone physical aggression. Such was the casein one study of hyperactive preschoolers from the community (vs. the clinic)who were found to have deficits in executive function (Mariani & Barkley,1997).
Studies Using Global Measures of Cognition
Only a handful or so of studies have examined more directly the relation be-tween preschool cognitive function and behavior problems not restricted tohyperactivity. For example, poor intelligence assessed by the McCarthy Scales(McCarthy, 1970) was related to mother-rated Child Behavior Check List(CBCL; Achenbach, Edelbrock, & Howell, 1987) psychopathology indices inpreschoolers recruited from pediatricians’ practices (Dietz, Lavigne, Arend, &Rosenbaum, 1997). However, because performance on the McCarthy Scaleswas related to internalizing as well as externalizing problems and because theMcCarthy score was taken either as a whole or only broken down into verbaland performance IQ, the study does not provide information about the speci-ficity of behavior problems in relation to cognitive function. Another longitu-dinal study with assessments at about 55 and 80 months of age found thatpreschool IQ was related to CBCL teacher ratings of externalizing problems atfollow-up and that preschool expressive language abilities were related toCBCL maternal ratings of externalizing problems at follow-up (Heller, Baker,Henker, & Hinshaw, 1996). However, these associations were accounted forby maternal education and socioeconomic status. This again suggests thatparental characteristics may influence both cognitive development and exter-nalizing problems, but it is not clear that this poor cognitive development rep-resents an incremental risk factor beyond parenting effects, because interac-tions with cognitive abilities were not tested. Interestingly, preschool ratings of
Executive Function in Aggression 313

externalizing behavior problems did not attenuate first-grade IQ and languageabilities over and above the variance explained by preschool cognitive abili-ties. Although preschool language abilities predicted stability and pervasive-ness of externalizing disorder across assessments, this was not the case for IQ.
Another prospective longitudinal study failed to support Moffitt’s (1993)hypothesis that adolescents with early-onset/persistent conduct problemswould show early neuropsychological deficits (Aguilar, Sroufe, Egeland, &Carlson, 2000). Based on CBCL externalizing scores, the authors contrastedfour groups (i.e., never antisocial, child-limited, adolescent-limited, and earlyonset/persistent) in a one-factor (group) design instead of a 2 × 2, child byadolescent, factorial design. Adolescent scores differentiated the adolescent-onset and early-onset/persistent groups, as in Moffitt’s earlier work (seeMoffitt, 1990; Moffitt & Henry, 1989; Moffitt & Silva, 1988b; Moffitt &Silva, 1988a), but the effect seems to be the result of having had behaviorproblems as a child, because those children who recovered from behaviorproblems (i.e. child-limited) appear to have performed as poorly as those thatdid not (i.e. early-onset/persistent). Nonetheless, early cognitive scores wereassociated with later ones, but stability of cognitive performance was not usedto identify those children who performed poorly in childhood and adoles-cence.
Despite the many merits of that study, its test of Moffitt’s hypothesis maybe severely limited. First, most children in that sample were at high risk for be-havior problems. Second, about a third of the sample was assigned to theearly-onset/persistent category, when we would expect about 4% of chroniccases in normative samples (Broidy et al., 2003). Third, the study included aroughly equal number of males and females, yet trajectories of early-onsetproblems do not predict the same maladaptive outcomes in males and females(Broidy et al., 2003; see also Archer & Côté, Chapter 20, this volume).Fourth, Moffitt suggested that the presence of aggression and hyperactivityshould be used as a criterion for early onset, and it is not clear that this is thecase here. Finally, there were no tests of executive function.
Raine and colleagues (Raine, Yaralian, Reynolds, Venables, & Mednick,2002) have proposed a different cognitive interpretation of persistent antiso-cial behavior. Their early-starter spatial impairment model of antisocial be-havior suggests that early visuospatial (right hemisphere) impairments canpredispose a child to persistent antisocial behavior by interfering with earlyattachment via emotion recognition and regulation. The model is admittedlyspeculative, but is reasonably well based on a series of community studies, in-cluding a longitudinal study of children from ages 3–17 years. Unlike moststudies reviewed so far, this study included stability of behavior as an impor-tant design feature. As in the Aguilar et al. (2000) study, four groups wereformed, based on whether children were above or below the top 33rd percen-tile of an antisocial behavior problem index at ages 8 and 17 years: a compari-son group, a child-limited group, an adolescence-onset group, and a persistentgroup. Results show that spatial abilities at age 3 years were poorest in the
314 D E T E R M I N A N T S O F A G G R E S S I O N

persistent group, and that this effect appeared to be stronger than the effectsof groups on verbal deficits. This effect was robust to statistical control forlevel of hyperactivity and test motivation at age 3 years. However, by the timethe children reached age 11 years, spatial and verbal deficits were equally im-paired in this group and social adversity attenuated considerably the groupeffect on spatial deficits. There appears to be a loss of specificity of spatial def-icits over time; the persistent children now seemed generally impaired.Although a spatial deficit hypothesis was formulated, it is difficult to arguethat the tasks that formed that spatial index (which represents essentially per-formance IQ tasks) were not contaminated by several other abilities. Theproblem of task purity is found everywhere in this type of research, and, un-fortunately, tasks that more explicitly and specifically involve executive func-tion were not administered in that study. A close examination of the batterydoes not suggest that any tasks could have been construed in terms of execu-tive function either. Although the Wisconsin Intelligence Scale for Children(WISC) Mazes subtest, which involves some degree of planning, was adminis-tered, individual subtest scores were not available. Further, physical aggres-sion was not examined. Thus, the question of the specific relevance of pre-school executive function to the concurrent or later regulation of physicalaggression remains unanswered.
Studies Using Specific Measures of Executive Functionin Community Samples
Moving more closely toward the concept of executive function, “disruptive-ness” in preschoolers, aged about 4½ years, was found to be negatively associ-ated with verbal and visuospatial abilities (Cole, Usher, & Cargo, 1993).“Executive function” was related to another measure of behavioral control(i.e., following the rule not to play with a forbidden but attractive object) butnot to disruptiveness. In that study verbal and visuospatial abilities were as-sessed with the McCarthy Scales (Kaufman & Kaufman, 1977; McCarthy,1970), as well as with the Florida Kindergarten Screening Battery (Satz &Fletcher, 1982), which includes the Beery Visual-Motor integration test (Beery,1982) and the Peabody Picture Vocabulary Test (Dunn & Dunn, 1981). Theexecutive function composite was made up of a tapping test (Becker, Isaac, &Hynd, 1987), a rapid-alternating-stimulus-naming test (Wolf, 1986), a handmovement test (Kaufman, 1983), a block sort (based on the WCST; Heaton,1981), and a visual search test (adapted from Plude & Doussard-Roosevelt,1989). All five tests loaded onto one principal component accounting for 42%of the variance, but the sum of scale scores was used instead of the principalcomponent score. Individual scores correlated significantly with one anotherbetween r = .27 and r = .64. Cognitive risk was defined as scores below onestandard deviation from the mean. Behavior was assessed with four scales.The mothers completed the CBCL (Achenbach, 1999; Achenbach & Edel-brock, 1983) and the Eyberg Child Behavior Inventory (Eyberg & Robinson,
Executive Function in Aggression 315

1983), and teachers completed the Teacher Report Form (TRF; Achenbach &Edelbrock, 1986) and the Preschool Behavior Questionnaire (Behar & String-field, 1974). In regard to behavior, three risk levels had been identified in thisstudy on the basis of combined informants and scale scores (i.e., low < 1 SDbelow mean for all informants, high > 1 SD above mean according to at leastone informant or T score > 70 on CBCL total score or > 85th percentile on theTRF, and moderate for the rest of the sample). We note that these criteria forbehavior risk definition were very liberal and may have increased the chancefor false positive identification of risk. Analyses contrasted a combination ofthe two high groups (moderate and high) with the low-risk group. Therefore,we do not know the relative positions of performance scores between thethree groups. It was also clear that the “high” group was elevated not only indisruptive behavior but also in other behavior problems, which may limit fur-ther the specificity of the finding to disruptive behavior.
Another study also referring explicitly to the concept of executive func-tion found that “hard-to-manage” children performed more poorly than con-trol children not only on executive function tasks, but also on theory of mindand emotion understanding tasks (Hughes, Dunn, & White, 1998). Interest-ingly, the relation between number of successful executive function tasks andbehavior problems was partly accounted for by the introduction of socioeco-nomic status and verbal abilities to the model, although the executive functionbattery was explicitly designed to require minimal verbal abilities. In thatstudy, executive function was measured with six tasks, and the authors wereguided by a model of executive function put forward by Welsh, Pennington,and Groisser (1991). This model emphasized three components: workingmemory and planning (measured with the Tower of London task ([Shallice,1982] and a “noisy book” auditory sequencing task [Hughes, 1998]), inhibi-tion of maladaptive prepotent responses (measured with Luria’s hand game[Hughes, 1998] and a “detour-reaching box” [Hughes & Russell, 1993]), andself-monitoring/attentional flexibility (measured with a pattern reproductiontask [Frith, 1971] and a card sort [Hughes, 1998] similar to the WCST). Alltest scores were converted to success/failure scores following Hughes (1998),and significant differences were observed on four of the six tasks, excludingthe “noisy book” and card sort tasks. In the first of two follow-up studieswith these same children, poor executive function and poor language abilitywere both related to violent fantasy (Dunn & Hughes, 2000). A secondfollow-up study of these “hard-to-manage” children involved playing with ateacher-nominated best friend in a room with an attractive toy (Hughes,White, Sharpen, & Dunn, 2000). Scores on the “detour-reaching box” and theTower of London tasks, seen as the two tasks most sensitive to antisocialbehavior by Hughes et al. (1998), were used to estimate executive function.Although the “hard-to-manage” children did not display more verbal aggres-sion relative to the control group, they snatched toys, teased, bullied, brokerules or the other child’s toy, hurt the other child, and engaged in sex play (i.e.,with a doll, themselves, or the other child in the room) more often. Verbal
316 D E T E R M I N A N T S O F A G G R E S S I O N

abilities were negatively related to hurting the other child (an observational in-dex of physical aggression) within the “hard-to-manage” group. The execu-tive function scores were associated with frequency of observed antisocial be-havior in this group even after controlling for verbal abilities and maternaleducation, but this was not the case for theory of mind scores. This latterstudy suggests that executive function problems, as opposed to verbal abilitiesor theory of mind, may be specifically related to antisocial behavior in chil-dren observed in the laboratory. However, this study does not focus on physi-cal aggression in the analyses involving executive function nor does it takeinto account the potential behavior problems of the teacher-nominated bestfriend. Thus, it is not clear whether antisocial peers, which are more likely toassociate with one another, have contributed to the escalation of behaviorproblems in the “hard-to-manage” group. Peer dynamics is a different but rel-evant issue (see Boivin, Vitaro, & Poulin, Chapter 18, this volume).
Studies Using Specific Measures of Executive Function in Clinic Samples
Most of the preschool studies reviewed so far have focused on communitysamples. One study examined clinic-referred preschool boys (ages 47–68months) with ODD and ADHD, an ODD group without ADHD, and amatched nonclinic control group (Speltz, DeKlyen, Calderon, Greenberg, &Fisher, 1999). In addition to being referred and having a primary DSM-III-R(American Psychiatric Association, 1987) diagnosis of ODD, these childrenalso had a T score > 65 on the CBCL aggression scale. Executive function wasoperationalized with two measures, one of motor planning (adapted from theLuria–Nebraska Battery for Adults) and the other of verbal fluency (takenfrom the McCarthy Scales). Verbal abilities were measured with the Expres-sive One Word Picture Vocabulary Test—Revised (Gardner, 1990), the Pea-body Picture Vocabulary Test (Dunn & Dunn, 1981), the Wechsler Preschooland Primary Scales of Intelligence—Revised (WPPSI-R; Wechsler, 1989) arith-metic and comprehension subtests, a Test of Early Reading Ability, and a testof vocabulary for affective states. Visual–motor abilities were measured withthe Beery Visual-Motor Integration Test (Beery, 1982) and the WPPSI-R blockdesign and picture completion subtests. Although confirmatory factor analy-ses showed that this dimensional grouping of variables was appropriate, testscores were summed within dimension and reliabilities were ranging frompoor to moderate, that is, from .41 for executive function, .52 for visual–motor, and .68 for verbal abilities. Clinic-referred boys combined did morepoorly on all dimensions and on test behavior in univariate analyses. How-ever, contrary to the authors’ hypothesis, the effect for executive function wasaccounted for by test behavior (even after removing test inattention andpersistence) and by verbal abilities. However, when breaking down the clinic-referred group into its subtypes, univariate comparison of the groups withODD-ADHD and ODD-only revealed poorer verbal (about 0.4 mean stan-dard difference) and executive function abilities (about 0.6 mean standard dif-
Executive Function in Aggression 317

ference, a moderate effect size) in the comorbid group. Although it was nottested for significance, the ODD-only and comparison groups differed byabout 0.5 SD in verbal abilities and 0.4 SD on executive function. Further, thecontrast for executive function was not controlled for performance in verbalabilities. In the absence of an ADHD-only group, support for a specific rela-tion of ODD-ADHD to poor executive function remains elusive.
In a second follow-up study, these same clinic-referred boys also gener-ated more aggressive solutions to social information–processing vignettes thannonclinic boys, independently from ADHD (Coy, Speltz, DeKlyen, & Jones,2001). However, verbal abilities at age 57 months, which were then worse inclinic-referred boys, were no longer associated with clinic status at age 82months. Unfortunately, executive function at age 57 months was not used as apredictor of later cognitive abilities or behavior in this follow-up study. Inter-estingly, and surprisingly, several physically aggressive behaviors (e.g. occa-sional shoving or hitting, or hitting of parent or sibling during a tantrum)were explicitly discounted for establishing the diagnosis of CD in that study sothat the diagnosis would emphasize deliberate confrontation. If most expres-sions of aggression are reactive instead of proactive, and poor executive func-tion is more likely to be associated with reactive aggression (Giancola, Moss,Martin, Kirisci, & Tarter, 1996), then these studies may be missing importantopportunities to understand the cognitive underpinnings of CD. (See Vitaro &Brendgen, Chapter 9, this volume, for a more detailed discussion of reactiveand proactive aggression.) Further, despite the longitudinal design of the sec-ond study, we do not know anything about the stability of problems acrossthe three measurement times. The longitudinal study did not examine whetherthe children with comorbid ODD-ADHD at time 1 were more likely to havean externalizing disorder at time 3. The effects of having been referred andpresumably being under professional care are also unknown.
THE UTILITY OF A PROBLEM-SOLVING FRAMEWORKFOR UNDERSTANDING AGGRESSION
We have already emphasized the need for greater specificity of behavior prob-lems that are the object of study instead of broad-band categories such as de-linquency, externalizing disorders, or even CD. We also suggest that measure-ments across time may be more powerful in determining children at highestrisk for later violence. This concept is becoming the standard, as many of thestudies reviewed thus far indicate. Similar methodological features need to besystematically implemented for the study of cognitive aspects involved in thebehavior problems of interest. In their empirical review, Pennington andOzonoff (1996) point to difficulties (1) in isolating components of executivefunction and (2) in considering also the many factors that may disrupt execu-tive function performance on traditional tasks. Because executive functionappears to be more strongly and specifically involved in physical aggression
318 D E T E R M I N A N T S O F A G G R E S S I O N

than does general cognitive development, indexed, for example, by IQ tests, atleast in adolescence and young adulthood, we propose that executive functionin preschoolers be examined from a functional perspective according to aproblem-solving framework.
This approach, which follows Luria (e.g., 1973), views executive functionas a functional construct that makes reference to (but cannot be equated with)the psychological processes involved in self-regulated, goal-directed problemsolving (Zelazo, Carter, Reznick, & Frye, 1997). Like all functions, executivefunction is defined solely in terms of what it accomplishes—not the mecha-nisms that accomplish the function. From this perspective, the varioussubfunctions of problem solving, from initially representing a problem, plan-ning a strategy, executing the plan, to eventually evaluating the adequacy ofan attempted solution, can all be seen as contributing to the function of goal-directed problem solving. For example, when searching for a hidden toy, chil-dren need to represent the problem, select a plan for action, actually executethe plan, and then evaluate the outcome. Treating executive function as afunctional construct does not explain executive function (it remains somethingto be explained), but it does lay the groundwork for an explanation by facili-tating the formulation of specific hypotheses regarding the role of basic cogni-tive processes (e.g., attention, memory, action monitoring) in different aspectsof executive function. It also provides a framework for devising more precisemeasures of executive function (e.g., measures of planning that do not also re-quire rule use).
A major advantage of the problem-solving framework is that it integratesdiverse executive function processes (and disparate lines of research) into asingle, coherent model of components. The problem-solving framework al-lows one to understand executive function in a way that suggests specific ex-ecutive function assessments (see Zelazo et al., 1997, for an exhaustive list)and clarifies the way in which different aspects of executive function work to-gether to fulfill the higher-order function of problem solving while avoidingconceptualizing executive function as a homuncular ability (e.g., as a supervi-sory attentional system; Shallice, 1988). Further, by designing tasks in light ofthis framework, one can discover more precisely when performance breaksdown in the process of problem solving, and this, in turn, bears on specific is-sues regarding the cognitive bases of unwarranted physical aggression.
1. Do some children engage in physical aggression because they fail torepresent a problem adequately? For example, some children may bebiased to represent certain situations as threatening because they havean underlying difficulty in attaining a more objective perspective andflexibly reinterpreting several situations more adaptively.
2. Alternatively, or in addition, some children may fail to plan or thinkahead properly. They may fail to anticipate the negative consequencesof a strategy that can be aggressive, for example.
3. In some cases, children may understand the rules that govern a social
Executive Function in Aggression 319

situation (e.g., “I should not hit others” or “I should be quiet duringclass” or “I should do as I am asked by my teacher”) but fail to usethese rules much in the same way that children fail to use rules thatthey know on tests of rule use (Zelazo & Frye, 1998).
4. Finally, difficulties with action evaluation may contribute to inflexibil-ity and persistence of inappropriate responding despite relevant feed-back (e.g., failing to learn from negative consequences of behavior).
THE UTILITY OF DISTINGUISHING BETWEEN HOT AND COLD EXECUTIVE FUNCTION
In addition to the distinctions between different phases of problem solving,one might usefully differentiate between the relatively “hot” affective aspectsof executive function associated more with ventral and medial regions ofprefrontal cortex and the more purely cognitive “cool” aspects associatedmore with dorsolateral prefrontal cortex (Zelazo & Müller, 2002). Whereascool executive function is more likely to be elicited by relatively abstract,decontextualized problems, hot executive function is required for problemsthat involve the regulation of affect and motivation (i.e., regulation of basiclimbic system functions). This characterization of hot executive function incontradistinction to cool executive function is consistent with several recentproposals regarding the function of orbitofrontal cortex (e.g., Damasio, 1994;Rolls, 1999). For example, based on single-cell recordings of neurons inorbitofrontal cortex together with neuroimaging data and evidence that dam-age to orbitofrontal cortex impairs performance on simple tests of object re-versal and extinction, Rolls (e.g., 1999) suggests that orbitofrontal cortex isrequired for the flexible representation of the reinforcement value of stimuli.Damasio proposed a rather different theory, the somatic marker theory (e.g.,1994). According to this theory, orbitofrontal cortex is required for processinglearned associations between affective reactions and specific scenarios, andthis processing plays a crucial but often overlooked role in decision making.Despite their differences, however, both approaches capture the fact that thecontrol of thought and the control of action depend on different cortical sys-tems, depending on whether or not such control occurs in motivationally sig-nificant contexts. Although a strict orbitofrontal cortex account of antisocialbehavior, and particularly physical aggression, is questionable (Séguin, 2004),the relevance of hot executive function is not.
Traditionally, research on executive function in human beings has fo-cused almost exclusively on cool executive function, using measures such asthe WCST. Recently, however, there has been growing interest in hot executivefunction. The hot aspect of executive function appears to be involved, for ex-ample, in theory of mind (Zelazo & Müller, 2002), delay of gratification(Mischel, Shoda, & Rodriguez, 1989), and affective decision making (Damasio,1994). An experimental approach to hot executive function using tasks suchas delay of gratification or the children’s gambling task (Kerr & Zelazo, 2004)
320 D E T E R M I N A N T S O F A G G R E S S I O N

might reveal that hot executive function may also play an important role inregulating physical aggression. However, an approach that integrates hot andcool executive function is likely to be most informative. As shown earlier, coolexecutive function was related to behavioral control—that is, following therule not to play with a forbidden but attractive object (Cole et al., 1993). Thetoy’s attractiveness invokes hot executive function, but it seems likely thatchildren regulate their behavior in this type of situation in part by translatingthe problem into a more cool, decontextualized problem (Mischel et al.,1989). We also found that boys with a history of physical aggression playedmore cards associated with rewards and punishments than nonaggressive boyseven after controlling for cool executive function (in this case working mem-ory; Séguin et al., 2002). Whereas one group of stable aggressive boys did soperhaps because of an underlying attentional problem, card perseveration wasrelated to emotional regulation (in this case neuroticism) in a group of unsta-ble aggressive boys. Individuals high in neuroticism are more likely to be re-sponsive to the hot or motivationally significant aspects of task demands,which increases their likelihood of dysregulated behavior.
CONCLUDING COMMENTS
Implicit in the problem-solving framework is the idea that deficits in physi-cally aggressive preschoolers are not necessarily global, although they may befor a subset of children. Much research in childhood and adolescence has at-tempted to specify the nature of the cognitive deficit in physical aggression(e.g., Séguin et al., 1999; Séguin et al., 1995) and hyperactivity (e.g., Penning-ton, Bennetto, McAleer, & Roberts, 1996). As this review shows, only a hand-ful of studies of preschoolers have attempted to break down cognition intosubcomponents (e.g., Raine et al., 2002) and isolate executive function deficitsfrom more global deficits (e.g., Cole et al., 1993; Dunn & Hughes, 2000;Hughes et al., 1998; Speltz et al., 1999). Still, many of the poor executivefunction abilities identified in physical aggression (i.e., working memory;Giancola, 1995; Séguin et al., 1995) and in hyperactivity (i.e., planning, inhi-bition; Pennington & Ozonoff, 1996) can be reformulated in terms of theproblem-solving framework. For example, much research on child aggression(not specifically physical) has focused on the way in which children representproblems. This research emphasizes distortions, biases, beliefs, and schemas(Huesmann, 1997), such as those that are the object of cognitive therapies(Lochman, Magee, & Pardini, 2003) and that often fall under the label ofsocial information processing (Pardini, Lochman, & Frick, 2003). In theirformulation of an information-processing model, Crick and Dodge (1994)emphasized four stages: encoding, attribution, problem solving, and responseevaluation. This model is strongly based on distortions and biases that predis-pose an individual to automatically construe problems in preconceived ways.Yet there are deficit models that emphasize more fundamental processing diffi-
Executive Function in Aggression 321

culties (Séguin et al., 1995; Wallace & Newman, 1997), which could be moredirectly based in brain anatomy or physiology but still interact with individualmotivational predispositions and which may be influenced greatly by early ex-perience (Mezzacappa, Kindlon, & Earls, 2001). Thus the problem-solvingframework suggests that a child’s inability to take another person’s perspectivein a relatively cool task (underlying deficit) is unlikely to change a more emo-tional bias. Conversely, the child who can show flexibility on a relatively cooltask (no underlying deficit) is more likely to be able to change perspective on aproblem that is laden with affective tone and for which that child shows amaladaptive response bias.
Several studies have also examined family characteristics and early expe-riences that influence not only preschoolers’ behavior problems but also theircognitive development (e.g., Heller et al., 1996; Mezzacappa et al., 2001;Morrell & Murray, 2003; Pierrehumbert, Ramstein, Karmaniola, & Halfon,1996). As such studies refine their measurement of executive function and be-havior problems, we will not only be able to see how family characteristics af-fect specific as opposed to global behavior problems but will also be able toidentify family characteristics that may more specifically influence the sub-phases of the problem-solving framework. As they become embedded in longi-tudinal designs, a study of the components of executive function will betterinform early intervention strategies that will target families or the child at risk.
In conclusion, there is growing evidence that poor executive function isinvolved in externalizing problems as early as the preschool years. However,the findings lack consistency largely because there is considerable variabilityin study objectives, and in cognitive and behavioral assessments across stud-ies. Further, very few studies have explicitly examined preschool executivefunction or focused on physical aggression. However, on the basis of studiesspanning childhood to adulthood, we would expect that physical aggressionalone, or in combination with other externalizing problems, becomes morespecifically related to poor executive function. There is thus a need for studiesthat explicitly chart the joint development of physical aggression, its comorbidcharacteristics, and executive function. Those studies would also do well toconsider predisposing and maintenance factors that affect this joint develop-ment in order to better inform prevention and intervention studies.
REFERENCES
Achenbach, T. M. (1999). The Child Behavior Checklist and related instruments. InM.E.Maruish (Ed.), The use of psychological testing for treatment planning andoutcomes assessment (2nd ed., pp. 429–466). Mahwah, NJ: Erlbaum.
Achenbach, T. M., & Edelbrock, C. S. (1983). Manual for the Child Behavior Checklistand Revised Child Behavior Profile. Burlington, VT: University Associates in Psy-chiatry.
Achenbach, T. M., & Edelbrock, C. S. (1986). Manual for the Teacher’s Report Form and
322 D E T E R M I N A N T S O F A G G R E S S I O N

the teacher version of the Child Behavior Profile. Burlington, VT: University Associ-ates in Psychiatry.
Achenbach, T. M., Edelbrock, C. S., & Howell, C. T. (1987). Empirically based assess-ment of the behavioral/emotional problems of 2- and 3-year-old children. Journal ofAbnormal Child Psychology, 15, 629–650.
Aguilar, B., Sroufe, L. A., Egeland, B., & Carlson, E. (2000). Distinguishing the early-on-set/persistent and adolescence-onset antisocial behavior types: From birth to 16years. Development and Psychopathology, 12, 109–132.
American Psychiatric Association (1987). Diagnostic and statistical manual of mentaldisorders (3rd ed., rev.). Washington, DC: Author.
Baddeley, A., & Della Sala, S. (1996). Working memory and executive control. Philo-sophical Transactions of the Royal Society of London, 351, 1397–1403.
Becker, M. G., Isaac, W., & Hynd, G. W. (1987). Neuropsychological development ofnonverbal behaviors attributed to “frontal lobe” functioning. DevelopmentalNeuropsychology, 3, 275–298.
Beery, K. E. (1982). Revised administration, scoring, and teaching manual for the devel-opmental test of visual–motor integration. Cleveland, OH: Modern CurriculumPress.
Behar, L., & Stringfield, S. (1974). A behavior rating scale for the preschool child. Devel-opmental Psychology, 10, 601–640.
Bonino, S., & Cattelino, E. (1999). The relationship between cognitive abilities and so-cial abilities in childhood: A research on flexibility in thinking and co-operationwith peers. International Journal of Behavioral Development, 23, 19–36.
Broidy, L., Nagin, D. S., Tremblay, R. E., Bates, J. E., Brame, R., Dodge, K. A., et al.(2003). Developmental trajectories of childhood disruptive behaviors and adoles-cent delinquency: A six site, cross-national study. Developmental Psychology, 39,222–245.
Chelune, G. J., & Baer, R. A. (1986). Developmental norms for the Wisconsin CardSorting Test. Journal of Clinical and Experimental Neuropsychology, 3, 219–228.
Cole, P. M., Usher, B. A., & Cargo, A. P. (1993). Cognitive risk and its association withrisk for disruptive behavior disorder in preschoolers. Journal of Clinical Child Psy-chology, 22, 154–164.
Coy, K., Speltz, M. L., DeKlyen, M., & Jones, K. (2001). Social-cognitive processes inpreschool boys with and without oppositional defiant disorder. Journal of Abnor-mal Child Psychology, 29, 107–119.
Crick, N. R., & Dodge, K. A. (1994). A review and reformulation of social information-processing mechanisms in children’s social adjustment. Psychological Bulletin, 115,74–101.
Damasio, A. R. (1994). Descartes’ error. New York: Harper & Collins.Delis, D. C., Squire, L. R., Bihrle, A., & Massman, P. (1992). Componential analysis of
problem-solving ability: Performance of patients with frontal lobe damage and am-nesic patients on a new sorting test. Neuropsychologia, 30, 683–697.
Denckla, M. B., & Reiss, A. L. (1997). Prefrontal–subcortical circuits in developmentaldisorders. In N. A. Krasnegor, G. R. Lyon, & P. Goldman-Rakic (Eds.), Develop-ment of the prefrontal cortex: Evolution, neurobiology, and behavior (pp. 283–293). Baltimore: Paul H. Brookes.
Dietz, K. R., Lavigne, J. V., Arend, R., & Rosenbaum, D. (1997). Relation between intel-ligence and psychopathology among preschoolers. Journal of Clinical Child Psy-chology, 26, 99–107.
Executive Function in Aggression 323

Dunn, J., & Hughes, C. (2000). “I got some swords and you’re dead!” Violent fantasy,antisocial behavior, friendship and moral sensibility in young children. Child Devel-opment, 72, 491–505.
Dunn, L. M., & Dunn, L. M. (1981). PPVT: Peabody Picture Vocabulary Test—Revised:Manual for forms L and M. Circle Pines, MN: American Guidance Services.
Eyberg, S. M., & Robinson, E. A. (1983). Conduct problem behavior: Standardization ofa behavioral rating scale. Journal of Clinical Child Psychology, 12, 347–354.
Farrington, D. P., & Loeber, R. (2000). Epidemiology of juvenile violence. Child and Ad-olescent Psychiatric Clinics of North America, 9, 733.
Frith, U. (1971). Studies in pattern detection in normal and autistic children. II. Repro-duction and production of colour sequences. Journal of Experimental Child Psy-chology, 10, 120–135.
Frye, D., Zelazo, P. D., Brooks, P. J., & Samuels, M. C. (1996). Inference and action inearly causal reasoning. Developmental Psychology, 32, 120–131.
Gardner, M. F. (1990). Expressive One Word Picture Vocabulary Test—Revised. Novato,CA: Academic Therapy.
Giancola, P. R. (1995). Evidence for dorsolateral and orbital prefrontal cortical in-volvement in the expression of aggressive behavior. Aggressive Behavior, 21,431–450.
Giancola, P. R., Mezzich, A. C., & Tarter, R. E. (1998). Executive cognitive functioning,temperament, and antisocial behavior in conduct-disordered adolescent females.Journal of Abnormal Psychology, 107, 629–641.
Giancola, P. R., Moss, H. B., Martin, C. S., Kirisci, L., & Tarter, R. E. (1996). Executivecognitive functioning predicts reactive aggression in boys at high risk for substanceabuse: A prospective study. Alcoholism: Clinical and Experimental Research, 20,740–744.
Grant, D. A., & Berg, E. A. (1948). A behavioral analysis of degree of reinforcement andease of shifting to new responses in a Weigl-type card-sorting problem. Journal ofExperimental Psychology, 38, 404–411.
Heaton, R. K. (1981). A manual for the Wisconsin Card Sorting Test. Odessa, FL: Psy-chological Assessment Resources.
Heller, T. L., Baker, B. L., Henker, B., & Hinshaw, S. P. (1996). Externalizing behaviorand cognitive functioning from preschool to first grade: Stability and predictors.Journal of Clinical Child Psychology, 25, 376–387.
Huesmann, L. R. (1997). The role of social information processing and cognitive schemain the acquisition and maintenance of habitual aggressive behavior. In R.G.Geen &E. Donnerstein (Eds.), Human aggression: Theories, research, and implications forpolicy. New York: Academic Press.
Hughes, C. (1998). Executive function in preschoolers: Links with theory of mind andverbal ability. British Journal of Developmental Psychology, 16, 233–253.
Hughes, C., Dunn, J., & White, A. (1998). Trick or treat? Uneven understanding of mindand emotion and executive dysfunction in “hard-to-manage” preschoolers. Journalof Child Psychology and Psychiatry, 39, 981–994.
Hughes, C., & Russell, J. (1993). Autistic children’s difficulty with mental disengagementfrom an object: Its implications for theories of autism. Developmental Psychology,29, 498–510.
Hughes, C., White, A., Sharpen, J., & Dunn, J. (2000). Antisocial, angry, and unsympa-thetic: “Hard to manage” preschoolers’ peer problems, and possible cognitive influ-ences. Journal of Child Psychology and Psychiatry, 41, 169–179.
324 D E T E R M I N A N T S O F A G G R E S S I O N

Kandel, E., & Freed, D. (1989). Frontal-lobe dysfunction and antisocial behavior: A re-view. Journal of Clinical Psychology, 45, 404–413.
Kaufman, A. S. (1983). Kaufman Assessment Battery for Children (K-ABC). CirclePines, MN: American Guidance Services.
Kaufman, A. S., & Kaufman, N. (1977). Clinical evaluation of young children with theMcCarthy Scales. Boston: Allyn & Bacon.
Kerr, A., & Zelazo, P. D. (2004). Development of “hot” executive function: TheChildren’s Gambling Task. Brain and Cognition, 55, 148–157.
Lahey, B. B., Loeber, R., Burke, J. D., Rathouz, P. J., & McBurnett, K. (2002). Waxingand waning in concert: Dynamic comorbidity of conduct disorder with other dis-ruptive and emotional problems over 17 years among clinic-referred boys. Journalof Abnormal Psychology, 111, 556–567.
Lilienfeld, S. O. (2003). Comorbidity between and within childhood externalizing andinternalizing disorders: Reflections and directions. Journal of Abnormal Child Psy-chology, 31, 285–291.
Lochman, J. E., Magee, T. N., & Pardini, D. A. (2003). Cognitive behavioral interven-tions for children with conduct problems. In M. Reinecke & D. A. Clark (Eds.),Cognitive therapy over the lifespan: Theory, research and practice (pp. 441–476).Cambridge, UK: Cambridge University Press.
Loeber, R., Burke, J. D., & Lahey, B. B. (2002). What are adolescent antecedents toantisocial personality disorder? Criminal Behaviour and Mental Health, 12, 24–36.
Luria, A. R. (1973). The working brain. New York: Basic Books.Marcovitch, S., & Zelazo, P. D. (1999). The A-not-B error: Results from a logistic meta-
analysis. Child Development, 70, 1297–1313.Mariani, M. A., & Barkley, R. A. (1997). Neuropsychological and academic functioning
in preschool boys with attention deficit hyperactivity disorder. DevelopmentalNeuropsychology, 13, 111–129.
Matson, D. E., & Fisher, M. (1991). A comparison of internalizers, externalizers, andnormals using the WISC—R and Wisconsin Card Sorting Test. Journal of Psycho-educational Assessment, 9, 140–151.
McCarthy, D. (1970). McCarthy Scales of children’s abilities. New York: PsychologicalCorporation.
McGee, R., Partridge, F., Williams, S., & Silva, P. A. (1991). A twelve-year follow-up ofpreschool hyperactive children. Journal of the American Academy of Child and Ad-olescent Psychiatry, 30, 224–232.
Mezzacappa, E., Kindlon, D., & Earls, F. J. (2001). Child abuse and performance task as-sessments of executive functions in boys. Journal of Child Psychology and Psychia-try, 42, 1041–1048.
Milich, R., Balentine, A. C., & Lynam, D. R. (2001). ADHD combined type and ADHDpredominantly inattentive type are distinct and unrelated disorders. Clinical Psy-chology Science and Practice, 8, 463–488.
Milner, B. (1963). Effects of different brain lesions on card sorting. Archives of Neurol-ogy, 9, 90–100.
Mischel, W., Shoda, Y., & Rodriguez, M. L. (1989). Delay of gratification in children.Science, 244, 933–938.
Miyake, A., Friedman, N. P., Emerson, M. J., Witzki, A. H., Howerter, A., & Wager, T. D.(2000). The unity and diversity of executive functions and their contributions tocomplex “frontal lobe” tasks. Cognitive Psychology, 41, 49–100.
Executive Function in Aggression 325

Moffitt, T. E. (1990). Juvenile delinquency and attention deficit disorder: Boys’ develop-mental trajectories from age 3 to age 15. Child Development, 61, 893–910.
Moffitt, T. E. (1993). Adolescence-limited and life-course-persistent antisocial behavior:A developmental taxonomy. Psychological Review, 100, 674–701.
Moffitt, T. E., & Henry, B. (1989). Neuropsychological assessment of executive func-tions in self-reported delinquents. Development and Psychopathology, 1, 105–118.
Moffitt, T. E., & Silva, P. A. (1988a). Neuropsychological deficit and self-reported delin-quency in an unselected birth cohort. Journal of the American Academy of Childand Adolescent Psychiatry, 27, 233–240.
Moffitt, T. E., & Silva, P. A. (1988b). Self-reported delinquency, neuropsychological defi-cit, and history of attention deficit disorder. Journal of Abnormal Child Psychology,16, 553–569.
Morgan, A. B., & Lilienfeld, S. O. (2000). A meta-analytic review of the relation betweenantisocial behavior and neuropsychological measures of executive function. Clini-cal Psychology Review, 20, 113–136.
Morrell, J., & Murray, L. (2003). Parenting and the development of conduct disorder andhyperactive symptoms in childhood: A prospective longitudinal study from 2months to 8 years. Journal of Child Psychology and Psychiatry, 44, 489–508.
Nagin, D. S., & Tremblay, R. E. (1999). Trajectories of boys’ physical aggression, opposi-tion, and hyperactivity on the path to physically violent and nonviolent juvenile de-linquency. Child Development, 70, 1181–1196.
Newman, J. P., & Wallace, J. F. (1993). Diverse pathways to deficient self-regulation: Im-plications for disinhibitory psychopathology in children. Clinical Psychology Re-view, 13, 699–720.
Norman, D. A., & Shallice, T. (1986). Attention to action: Willed and automatic controlof behavior. In R.J.Davidson, G. E. Schwartz, & D. Shapiro (Eds.), Consciousnessand self-regulation (pp. 4–18). New York: Plenum Press.
Owen, A. M., Herrod, N. J., Clark, J. C., Downey, S. P., Carpenter, T. A., Minhas, P. S., etal. (1999). Redefining the functional organization of working memory processeswithin human lateral prefrontal cortex. European Journal of Neuroscience, 11,567–574.
Pardini, D. A., Lochman, J. E., & Frick, P. J. (2003). Callous/unemotional traits and so-cial-cognitive processes in adjudicated youths. Journal of the American Academy ofChild and Adolescent Psychiatry, 42, 364–371.
Parkin, A. J. (1998). The central executive does not exist. Journal of the InternationalNeuropsychological Society, 4, 518–522.
Pennington, B. F. (1997). Dimensions of executive functions in normal and abnormal de-velopment. In N.A.Krasnegor, G. R. Lyon, & P. Goldman-Rakic (Eds.), Develop-ment of the prefrontal cortex: Evolution, neurobiology, and behavior (pp. 265–281). Toronto: Brookes.
Pennington, B. F., Bennetto, L., McAleer, O., & Roberts, R. J. (1996). Executive func-tions and working memory: Theoretical and measurement issues. In G. R. Lyon &N. A. Krasnegor (Eds.), Attention, memory, and executive function (pp. 327–348).Baltimore: Paul H. Brookes.
Pennington, B. F., & Ozonoff, S. (1996). Executive functions and developmentalpsychopathology. Journal of Child Psychology and Psychiatry, 37, 51–87.
Petrides, M., Alivisatos, B., Evans, A. C., & Meyer, E. (1993a). Dissociation of humanmid-dorsolateral from posterior dorsolateral frontal cortex in memory processing.
326 D E T E R M I N A N T S O F A G G R E S S I O N

Proceedings of the National Academy of Sciences of the United States of America,90, 873–877.
Petrides, M., Alivisatos, B., Meyer, E., & Evans, A. C. (1993b). Functional activation ofthe human frontal cortex during the performance of verbal working memory tasks.Proceedings of the National Academy of Sciences of the United States of America,90, 878–882.
Pierrehumbert, B., Ramstein, T., Karmaniola, A., & Halfon, O. (1996). Child care in thepreschool years: Attachment, behaviour problems and cognitive development. Eu-ropean Journal of Psychology of Education, 11, 201–214.
Plude, D. J., & Doussard-Roosevelt, J. A. (1989). Aging, selective attention, and featureintegration. Psychology and Aging, 4, 98–105.
Raine, A., Yaralian, P. S., Reynolds, C., Venables, P. H., & Mednick, S. A. (2002). Spatialbut not verbal cognitive deficits at age 3 years in persistently antisocial individuals.Development and Psychopathology, 14, 25–44.
Robbins, T. W. (1996). Dissociating executive functions of the prefrontal cortex. Philo-sophical Transactions of the Royal Society of London, 351, 1463–1470.
Robins, L. N. (1991). Conduct disorder. Journal of Child Psychology and Psychiatry, 32,193–212.
Robinson, A. L., Heaton, R. K., Lehman, A. W., & Stilson, D. W. (1980). The utility ofthe Wisconsin Card Sorting Test in detecting and localizing frontal lobe lesions.Journal of Consulting and Clinical Psychology, 48, 605–614.
Rolls, E. T. (1999). The brain and emotion. Oxford, UK: Oxford University Press.Satz, P., & Fletcher, J. M. (1982). Florida Kindergarten Screening Battery. Odessa, FL:
Psychological Assessment Resources.Séguin, J. R. (2004). Neurocognitive elements of antisocial behavior: Relevance of an
orbitofrontal cortex account. Brain and Cognition, 55, 185–197.Séguin, J. R., Arseneault, L., Boulerice, B., Harden, P. W., & Tremblay, R. E. (2002). Re-
sponse perseveration in adolescent boys with stable and unstable histories of physi-cal aggression: The role of underlying processes. Journal of Child Psychology andPsychiatry, 43, 481–494.
Séguin, J. R., Boulerice, B., Harden, P., Tremblay, R. E., & Pihl, R. O. (1999). Executivefunctions and physical aggression after controlling for attention deficit hyperactiv-ity disorder, general memory, and IQ. Journal of Child Psychology and Psychiatry,40, 1197–1208.
Séguin, J. R., Nagin, D. S., Assaad, J.-M., & Tremblay, R. E. (2004). Cognitive-neuropsychological function in chronic physical aggression and hyperactivity. Jour-nal of Abnormal Psychology, 113, 603–613.
Séguin, J. R., Pihl, R. O., Harden, P. W., Tremblay, R. E., & Boulerice, B. (1995). Cogni-tive and neuropsychological characteristics of physically aggressive boys. Journal ofAbnormal Psychology, 104, 614–624.
Sergeant, J. A., Geurts, H., & Oosterlaan, J. (2002). How specific is a deficit of executivefunctioning for attention-deficit/hyperactivity disorder? Behavioural Brain Re-search, 130, 3–28.
Shallice, T. (1982). Specific impairments of planning. Philosophical Transactions of theRoyal Society of London, B298, 199–209.
Shallice, T. (1988). From neuropsychology to mental structure. New York: CambridgeUniversity Press.
Speltz, M. L., DeKlyen, M., Calderon, R., Greenberg, M. T., & Fisher, P. A. (1999).
Executive Function in Aggression 327

Neuropsychological characteristics and test behaviors of boys with early onset con-duct problems. Journal of Abnormal Psychology, 108, 315–325.
Stouthamer-Loeber, M., Loeber, R., Wei, E., Farrington, D. P., & Wikström, P.-O. H.(2002). Risk and promotive effects in the explanation of persistent serious delin-quency in boys. Journal of Consulting and Clinical Psychology, 70, 111–123.
Stuss, D. T. (1992). Biological and psychological development of executive functions.Brain and Cognition, 20, 8–23.
Toupin, J., Déry, M., Pauzé, R., Mercier, H., & Fortin, L. (2000). Cognitive and familialcontributions to conduct disorder in children. Journal of Child Psychology and Psy-chiatry, 41, 333–344.
Tremblay, R. E. (2000). The development of aggressive behaviour during childhood:What have we learned in the past century? International Journal of Behavioral De-velopment, 24, 129–141.
Tremblay, R. E., Boulerice, B., Harden, P. W., McDuff, P., Pérusse, D., Pihl, R. O., et al.(1996). Do children in Canada become more aggressive as they approach adoles-cence? In Growing up in Canada (pp. 127–137). Ottawa: Statistics Canada.
Tremblay, R. E., Japel, C., Pérusse, D., Boivin, M., Zoccolillo, M., & Montplaisir, J.(1999). The search for the age of “onset” of physical aggression: Rousseau andBandura revisited. Criminal Behavior and Mental Health, 9, 24–39.
Tremblay, R. E., Nagin, D. S., Séguin, J. R., Zoccolillo, M., Zelazo, P. D., Boivin, M., etal. (2004). Physical aggression during early childhood: Trajectories and predictors.Pediatrics, 114, e43–e50.
Wallace, J. F., & Newman, J. P. (1997). Neuroticism and the attentional mediation ofdysregulatory psychopathology. Cognitive Therapy and Research, 21, 135–156.
Waschbusch, D. A. (2002). A meta-analytic examination of comorbid hyperactive–im-pulsive–attention problems and conduct problems. Psychological Bulletin, 128,118–150.
Wechsler, D. (1989). Manual for the Wechsler Preschool and Primary Scale of Intelli-gence—Revised. Toronto: Psychological Corporation.
Wellman, H. M., Cross, D., & Watson, J. (2001). Meta-analysis of theory of mind devel-opment: The truth about false belief. Child Development, 72, 655–684.
Welsh, M. C., Pennington, B. F., & Groisser, D. B. (1991). A normative-developmentalstudy of executive function: A window on prefrontal function in children. Develop-mental Neuropsychology, 7, 131–149.
Willcutt, E. G., Pennington, B. F., Chhabildas, N. A., Friedman, M. C., & Alexander, J.(1999). Psychiatric comorbidity associated with DSM-IV ADHD in a nonreferredsample of twins. Journal of the American Academy of Child and Adolescent Psychi-atry, 38, 1355–1362.
Wolf, M. (1986). Rapid alternating stimulus naming in the developmental dyslexias.Brain and Language, 27, 360–379.
Zelazo, P. D., Carter, A., Reznick, J. S., & Frye, D. (1997). Early development of execu-tive function: A problem-solving framework. Review of General Psychology, 1,198–226.
Zelazo, P. D., & Frye, D. (1998). Cognitive complexity and control: II. The developmentof executive function in childhood. Current Directions in Psychological Science, 7,121–126.
Zelazo, P. D., Frye, D., & Rapus, T. (1996a). An age-related dissociation between know-ing rules and using them. Cognitive Development, 11, 37–63.
328 D E T E R M I N A N T S O F A G G R E S S I O N

Zelazo, P. D., Helwig, C. C., & Lau, A. (1996b). Intention, act, and outcome in behav-ioral prediction and moral judgment. Child Development, 67, 2479–2492.
Zelazo, P. D., & Müller, U. (2002). Executive function in typical and atypical develop-ment. In U. Goswami (Ed.), Handbook of childhood cognitive development (pp.445–469). Oxford, UK: Blackwell.
Zelazo, P. D., & Reznick, J. S. (1991). Age-related asynchrony of knowledge and action.Child Development, 62, 719–735.
Zelazo, P. D., Reznick, J. S., & Piñon, D. E. (1995). Response control and the executionof verbal rules. Developmental Psychology, 31, 508–517.
Executive Function in Aggression 329

DETERMINANTS OF AGGRESSIONLanguage Development and Aggressive Behavior
16
Language Developmentand Aggressive Behavior
GINETTE DIONNE
Aggression has been associated with low language proficiency as early as thesecond year of life and throughout the lifespan. Although language deficits arenot sufficient or necessary correlates of aggressive behavior, they have receivedmuch attention as frequently co-occurring with behavior problems (for re-views, see Beitchman, Cohen, Konstantareas, & Tannock, 1996; Cantwell &Baker, 1977, 1987; Cohen, 2001; Donahue, Cole & Hartas, 1994; Gallagher,1999; Goodyer, 2000; Howlin & Rutter, 1987; Silva, 1987; Stevenson, 1996).This has led to a consensus around the view that the link between languageand behavior is functional, not spurious. Just what this functional link is,however, remains unclear.
Previous reviews of the association between language and behavior havemainly focused on the variety of behavioral and emotional correlates of lan-guage deficits in childhood. One difficulty in attempting to write a chapterspecifically on aggression and language is that aggression is rarely directly as-sessed in developmental psychopathology. Yet aggression, possibly leading toviolence and criminality, has a huge social impact. As such, it is important thatwe understand how it comes about, and language should not be disregardedas an associated factor.
This chapter examines empirical evidence and surveys theoretical expla-nations of the association between language deficits and aggression. The goalis to look at the numbers, then search for the story (or, possibly, stories)behind the numbers. For the sake of brevity, it does not attend to specific syn-dromes in which aggression and pervasive language deficits are severely inca-pacitating, as in general developmental delay, pervasive developmental disor-ders, autism, childhood-onset schizophrenia, psychosis, or structural braininjuries. Where aggression is not assessed per se, as is the case in most studies
330

of early development, the review is extended to include conduct disorders.Early-onset conduct disorders have been shown to be particularly character-ized by the use of physical aggression (Lahey et al., 1998). Finally, morebroadly defined externalizing behavior problems are also included, but withan added caution in regard to making inferences about aggression.
In line with the developmental focus of this book, this chapter takes a de-velopmental approach to the review of prevalence data from clinical, epidemi-ological, and longitudinal studies. As the picture unfolds from descriptivedata, theoretical models of the language–aggression association are outlined,with a focus on the empirical evidence available to support these models. Asthis empirical evidence still remains scarce, this chapter concludes with a lookat some considerations that research designs could address to take on the cur-rent challenges in understanding the functional links between language andaggression.
THE HISTORICAL PERSPECTIVE
The notion that language and behavior are intrinsically linked in humans ishardly new. Although it can be traced back to ancient philosophers, the func-tional role of language in moral action and social order became a favoritetopic of late 17th- and 18th-century philosophers such as Locke and Rous-seau. By the early 20th century, the renewed interest from an empirical per-spective largely stemmed from observations in clinical settings. British lan-guage and literacy specialist Samuel T. Orton concluded, in his classic 1937essay on dyslexia and language, that communication problems led to second-ary behavior and social problems. A few years later, Stella Chess (1944)echoed this view in an early paper in which she presented language disabilitiesas a factor in “personality distortions” in children.
On another front, early Russian psychologists were speculating on possi-ble causal processes linking language to behavior regulation. Pavlov’s (1927)concept of language as a second signaling system, on which excitatory and in-hibitory processes operate to direct and control behavior, pointed to the possi-ble role of language in the inhibition of aggressive impulses. Both Luria (1961)and Vygotsky (1962) later formalized the notion that language, spoken or asan inner process, was a necessary means to behavior regulation. Althoughmuch of this earlier work was speculative in nature, it created the basis for thenext four decades of theorizing and research.
In the early 1970s, Montare and Boone (1973) formulated an explicit“language–aggression hypothesis”: They proposed that language was relatedto physical aggression in two ways—low levels of proficiency linked to higherlevels of observable aggression, and high proficiency linked to lower levels ofaggression. Their empirical work on a group of children and teenagers fromdiverse ethnic backgrounds provided mitigated support for the hypothesis(Boone & Montare, 1976), and subsequent investigations of a specific link be-
Language Development and Aggressive Behavior 331

tween aggression and language proficiency have been few and far between(Cole, 2001; Dionne, Tremblay, Boivin, Laplante, & Pérusse, 2003; Mack &Warr-Leeper, 1992; Stattin & Klackenberg-Larsson, 1993; Piel, 1990). Duringthis time frame, general intelligence became the focus of many studies as a cor-relate of delinquency and criminality, low intelligence more often characteriz-ing violent offenders (see Moffitt, Gabrielli, Mednick, & Schulsinger, 1981).However, as Stattin and Klackenberg-Larsson (1993) point out, early lan-guage delays have been shown to be highly correlated with lower verbal andnonverbal intelligence. In this regard, they hypothesize that “the early differ-ences between future delinquents and nondelinquents might be found in thearea of language development” (p. 370). Is there support for this assertion?The next sections of this chapter look into empirical evidence of concurrentand longitudinal links between language development and aggression at dif-ferent periods of development. But first, the terms and criteria used to defineatypical development in both fields are addressed and the normative develop-mental course of language and socialized behavior is outlined to provide abackground against which empirical data may be understood.
DEFINING LANGUAGE DEFICITS AND AGGRESSION
To study the interface between language and behavior in psychopathology,one must be aware of the variety of terms and criteria used to define atypicaldevelopment in both fields. In psycholinguistics, a deficit may permeate thewhole language system or may affect specific aspects of language differen-tially. On one hand, general deficits are often referred to as specific languageimpairments (SLI)—although what is specific to SLI remains somewhat con-troversial—indicating language proficiency below that which is expected foran otherwise normally developing child’s IQ range. Speech and language diffi-culties (SLD), on the other hand, is a more general construct that refers tomore or less severe nonoptimal functioning in many language-related skills. Avariety of general and specific measures of language deficits are used in theprevalence studies reviewed. Basically, language skills may be affected at threedifferent levels: production, content, and use of language. Production prob-lems linked with speech processes such as stuttering or articulation have, forthe most part, not been linked with a higher occurrence of conduct problems(Beitchman et al.,1999; Cantwell & Baker, 1987; Gualtieri, Koriath, VanBourgondien, & Saleeby, 1983; Love & Thompson, 1988). The content anduse of language however, have been linked with behavioral problems. Thecontent of language refers to the mastery of a sufficient number of words (lex-ical development) and their meaning (semantic development) and of the rulesthat govern how they come together to form sentences and convey meaning.To understand and create these sentences, a child must master phoneme recog-nition, be able to segment the stream of speech to decipher basic units, and beable to memorize and use these units effectively. Deficits at this level, whether
332 D E T E R M I N A N T S O F A G G R E S S I O N

expressive or receptive, are more readily diagnosed because the resulting com-munication impairment is often obvious. In addition, they are assessed moresystematically in screening procedures. Although emotional and social prob-lems appear to be the most prevalent forms of psychopathology in childrenwith deficits in the content of language (Cantwell & Baker, 1977), a few stud-ies suggest that there may be long-term effects of early deficits in regard to vi-olent behavior (Beitchman et al., 1999; Stattin & Klackenberg-Larson, 1993).Finally, use of language comprises the notions of turn taking in communica-tion and use of contextually appropriate and socially efficient language. Defi-cits in the pragmatics of language, as they are called, are more readily missedbecause they do not audibly impair everyday communication. Yet, over time,they result in a breakdown in communication with others and unrewardingcommunication experiences. A few studies point to pragmatics as the prevail-ing language deficit in aggressive children (Cole, 2001; Mack & Warr-Leeper,1992; Minutti, 1991). As pragmatics are often overlooked unless audible lan-guage presents some concern, it is not surprising to find that a majority ofchildren referred to clinics for externalizing behavior problems have undiag-nosed language deficits (Cohen, 2001).
Aggression, on the other hand, is rarely assessed directly in most large-scale studies dealing with the language–behavior interface. For one thing, ag-gression is not a psychiatric diagnosis, but rather a criterion associated withconduct or behavior disorder. Most large-scale studies reviewed have dealtwith either psychiatric outpatient clinic populations or populations ascer-tained for language deficits. When behavior problems are considered, theymay be labeled as conduct disorders or externalizing problems (including orexcluding ADHD) in preschool and school-age children, antisocial personal-ity, or delinquency or registered criminality in adolescents and adults. Any ofthese labels may or may not imply overt aggression. For instance, although 7of 15 criteria for conduct disorder refer to overt aggression, a diagnosis ofconduct disorder may be given in the absence of an aggression component. Inaddition, although aggression studies point to different developmental trajec-tories involving overt, covert, and verbal aggression as well as reactive versusproactive types of aggression, these distinctions have never been addressed instudies looking into comorbid language deficits. Inferences are thus limitedwith respect to specific forms of aggression. Nonetheless, the convergence orabsence thereof, between studies of aggression and those looking at more gen-eral forms of externalizing behavior problems, may be of particular interest todecipher what, if any, developmental processes they may share.
PREVALENCE THROUGHOUT DEVELOPMENT
How the Developmental Story Unfolds
These conceptual limitations aside, the first question to address is the preva-lence of these co-occurring problems throughout development. As with many
Language Development and Aggressive Behavior 333

other considerations in developmental psychopathology, most of what weknow about co-occurring language and behavior problems is documented forschool-age children, and to a much lesser extent, adolescents and adults, oftenin follow-up studies. Although the exact proportions of overlap vary fromstudy to study, it is estimated that more than 50% of children with behaviorproblems have language deficits, many of them undiagnosed (Cohen, 1998).Similar proportions of language-impaired children are reported to display be-havior problems severe enough to warrant intervention. It is safe to assumethat a significant proportion of these behavior problems would include someforms of aggression. Whether the initial focus is on behavior or language, thegeneral outlook is the same: A child with one of these problems is up to threeor four times more likely to have the other, than a child in the community atlarge (Goodyer, 2000; Stevenson, 1996). This childhood snapshot, however,says little about what leads to these compelling statistics and even less aboutwhen and how the story unfolds.
Which came first? In any attempt to understand co-occurring problems,this question inevitably arises. And as with hens and eggs, the answer in thiscase may be equally problematic. Decades of research on aggression in hu-mans still leaves the century-old question of the onset of physical aggressionunresolved or, at best, with controversial answers (Tremblay, 2000). Psycho-linguists, however, have less difficulty in agreeing that early infancy is the ves-sel of early language development. The first spoken words typically appeararound the first birthday, but a child is able to comprehend words and linkthem to objects somewhat earlier. By the end of the second year most childrenexperience a vocabulary spurt, which generally precedes the onset of wordcombinations into sentences. By age 3, simple grammatical forms are used tomake complete sentences, albeit language remains somewhat telegraphic atthis age. At any point in time, a child’s understanding of language, and thushis or her receptive abilities, precede the ability to use the same grammaticalforms in expressive language. At ease with the rudiments of spoken language,most 3-year-olds are able to make demands, give information verbally, andhave an active role in conversation. With the availability of normed instru-ments of early language development, we are able to diagnose early delays inthe lexical and grammatical content of spoken language, as well as in a child’sability to understand spoken language. Although most delays before age 3appear transient (Dale, Price, Bishop, & Plomin, 2003), some 40% of slowtalkers still show signs of delay by the end of the preschool years and beyond.Those with receptive delays, who show early difficulties in understanding lan-guage, tend to have a worse prognosis.
Theoretical disputes aside, the developmental story of aggression sharessome interesting milestones with language development. As Goodenough(1931) illustrated years ago, the frequency of aversive types of behavior, fromfussing and crying in the first year, to kicking, biting, hitting, and tantrumswhen physical ability enables such behavior, shows a marked increase from 6
334 D E T E R M I N A N T S O F A G G R E S S I O N

months onward to reach a peak between the ages of 18 and 36 months.Goodenough, and others after her (Vygotsky, 1962; Luria, 1961), have arguedthat a child’s growing verbal ability is instrumental in the following decreaseof physically disruptive behaviors after 36 months. This milestone does coin-cide with the onset of efficient communicative skills in most children, as statedearlier.
In an attempt to prospectively track the onset of physical aggression in arepresentative birth cohort, Tremblay and colleagues (Tremblay et al., 1996)showed that by 17 months, 80% of children were reported to exhibit someform of physically aggressive behavior. From the third year onward, physicalaggression was shown to decrease (Tremblay et al., 1996), with a second peakaround midadolescence, mainly for boys. As with early language deficits, highlevels of physical aggression in infancy tend to remain stable in a proportionof children (Hay, Castle, & Davies, 2000; Keenan & Shaw, 1993), placingthem most at risk of following a high aggression trajectory leading to laterviolence and criminality.
Some forms of physical aggression and rough-and-tumble play may benormative during the period fondly known to parents as the “terrible twos.”In most children, after this period, physical aggression is replaced by less overtforms of aggression, namely, verbal and indirect aggression (Bjorkvist, Oster-man, & Kaukiainen, 1992). Although minimal language ability is required touse verbal aggression, it remains unclear whether the use of verbal aggressionis highest among verbally proficient toddlers. If, as is the case in school-agechildren, rates of verbal and physical aggression remain associated (Pepler,Craig, & Roberts, 1998), it may be that some physical aggressors add verbalaggression to their repertoires and others move from physical to verbal formsof aggression. In time (it is hoped), language becomes for most children asocial tool for increased prosocial interactions. Whatever the case may be,from both the behavior and language perspectives, the early toddler yearsserve as the center stage for emergent individual differences that may have aprofound impact on later adjustment.
From that point onward, language abilities continue to progress andphysically aggressive behaviors diminish in frequency. At no other moment inlife will the frequency of aversive-type behaviors reach the peak and subse-quent sharp decline observed in the toddler years; similarly, at no other pointin time will language abilities show a growth as staggering as that evidenced inthe first 3 years of life. For some children, however, language has not pro-gressed as anticipated at this milestone, and for some, physical aggression con-tinues to attract increasing concern. The question is to what extent are thesethe same children? Studies offer different answers, using different methodolo-gies at various periods of development. Two separate concerns are generallyaddressed in regard to prevalence: (1) concurrent behavior and language prob-lems and (2) longitudinal outcomes of earlier language deficits. The followingsections offer a review of prevalence studies throughout development.
Language Development and Aggressive Behavior 335

Concurrent Prevalence in Infancy and the Preschool Years
The youngest age at which epidemiological data are available on the language–behavior association comes from two recent twin studies addressing the ge-netic etiology of this association and a small community sample. We returnto the etiological answers provided by the genetic studies later. A first ques-tion they attend to pertains to the age of onset of this association. Howearly is there evidence of an association between language deficits and ag-gression?
In the Québec Newborn Twin Study (QNTS), a modest but significantcorrelation of –.20 was found between expressive vocabulary assessed via aparent checklist and parent reports of physical aggression at 19 months(Dionne et al., 2003). Although there were no sex differences in either vocabu-lary or aggression at this age, the correlation was slightly higher for boys thanfor girls. In contrast, when more general conduct problems were considered inrelation to vocabulary in the Twin Early Development Study (TEDS), Plominand colleagues (Plomin, Price, Eley, Dale & Stevenson, 2002) found correla-tions of –.05 at 24 months for both boys and girls. In addition, language-delayed children (below the 5th and 10th percentiles) in this sample did notshow a higher incidence of conduct problems. There was also no significantassociation between language and behavior in a small community samplecomparing language-delayed (n = 11) and language normal (n = 53) 24-month-olds on a CBCL measure of total externalizing behavior (Carson, Klee,Perry, Muskina, & Donaghy, 1998). Subsequent analyses of the Québec sam-ple, however, show this not to be the case when physical aggression is consid-ered. Indeed, posthoc analyses on this sample of 900 twins indicate a signifi-cantly higher incidence of physical aggression in 19-month-olds below the15th percentile on vocabulary (p < .05). Although there appears to be an asso-ciation, albeit very modest, of delayed language with aggression before age 2,this does not seem to be the case when more general conduct problems areconsidered.
By age 3, prevalence reports markedly differ. In the TEDS sample, corre-lations at follow-up assessments at 36 and 48 months remain modest: –.13 forboys and –.09 for girls at 36 months and –.18 for boys and –.15 for girls by 48months. Again, the incidence of conduct problems and total behavior prob-lems is not higher in the delayed children at both ages. This is in sharp con-trast with results from an earlier U.K. study by Stevenson and Richman(1978), in which half of the language-delayed children, from a sample of 8283-year-olds, showed behavior problems. A majority of these problems per-tained to general immaturity and overactive behavior. The most frequent lan-guage delay pertained to difficulty with language structure. In a later study,Silva, Williams, and McGee (1987) assessed a normative sample of 1,0373-year-olds in New Zealand. In this study, a higher incidence of behaviorproblems, as rated by parents and teachers on the Rutter Scale, was found for
336 D E T E R M I N A N T S O F A G G R E S S I O N

children with general language delay and comprehension delay but not forthose with expressive delay.
The variety of language and behavior measures make comparisons andconclusions difficult before age 3, but the general trends in the U.K. and N.Z.studies of 3-year-olds appear sharper by age 4–5. In the late 1970s, Cantwelland Baker (1977) studied behavioral outcomes in incoming patients to a com-munity speech and hearing clinic. Two-thirds of these children were of pre-school age (median 4.8 years). The 600 children identified within a 3-year pe-riod were divided into three language attainment categories: pure speechdisorder, speech and language disorder, and pure language disorder, based ona comprehensive speech and language assessment. Speech-disordered-onlychildren showed fewer concurrent psychiatric disorders. Overt behavioral dis-orders were identified in 30% of the speech and language group, and in 47%of the pure language group. However, approximately two-thirds of the overtbehavioral disorder were related to attention deficits, and the incidence ofconduct disorder was only slightly more elevated in these groups than wouldbe expected in the normal population. Cantwell and Baker (1977) cautionedthat because a conduct disorder diagnosis requires a long pattern of behavior,it was rarely given for the younger children. This may account for the low inci-dence. However, as there is a high prevalence of physical aggression in attention-deficit disorders, the co-occurrence of language deficits and aggression in thelate preschool period may have been underestimated in this sample.
A stronger association has been shown in one of the most extensive longi-tudinal investigations of psychopathology in language-delayed children. Thisstudy was initiated in a community-based sample of 5-year-olds in Ottawa,Canada, by Joseph Beitchman and his colleagues (Beitchman, Nair, Clegg,Ferguson, & Patel, 1985). At initial assessment, the boys (n = 90) among the142 children in the language-delayed group showed a higher incidence(13.6% versus 3.4% in controls) of conduct disorder than controls, but notthe girls. A recent community study of 4- and 5-year-old preschoolers also re-ports this gender difference (Ortiz, Stowe, & Arnold, 2001). Lower levels ofexpressive and receptive vocabulary skills in this sample (n = 56) werestrongly associated with disruptive behavior as coded from video observationsand from teacher ratings, but for boys only. Although none of these studies fo-cused specifically on aggression, the gender-biased results are consistent withthose of previous studies (Beitchman et al., 1985; Dionne et al., 2003).
Thus, during infancy and the preschool years, some studies show evi-dence of a higher incidence of aggressive behaviors in preschoolers with lowerlanguage proficiency, particularly for boys. However, other studies report nosuch evidence. In the studies where there is a significant association in this agerange, pure speech impairments are not the culprit. Some studies report astronger association for receptive deficits, whereas other studies report associ-ations for both receptive and expressive language deficits, mainly for vocabu-lary. Overall, however, these associations remain very modest.
Language Development and Aggressive Behavior 337

Concurrent Prevalence in Childhood and Adolescence
By the early school years, the specificity of language deficits linked with ag-gression emerges. In one of the earlier childhood studies, Camarata, Hughes,and Ruhl (1988) showed that the language performance of children identifiedas having mild to moderate behavior disorder was typically 2 or more stan-dard deviations below the normative mean. These children displayed prob-lems with expressive syntax while having a satisfactory knowledge of vocabu-lary. Results from Cohen and colleagues (Cohen, Davine, Horodezky, Lipsett,& Isaacson, 1993) point to similar language deficiencies linked with use oflanguage more than content of language in aggressive children. They system-atically screened 399 psychiatric outpatients ages 4–12 years and found thatone-third of the referred children had an unsuspected language impairment.Overall, more than 50% of these children exhibited some form of languageimpairment, previously identified or not. Although semantics, syntax, andphonology were more affected in the group with previously identified lan-guage impairment, a higher proportion of children from the unsuspected lan-guage impairment group showed problems with receptive syntax, auditorymemory, and fluency. Cohen (1998) later reported that although the prevail-ing behavior diagnosis for children with language impairments was ADHD,the children with unsuspected language impairment were rated as more ag-gressive and having more problems with delinquency. These two studies sug-gest that aggressive children may have more problems with the use and under-standing of complex language structures than with lexical knowledge.
The specificity of pragmatic difficulties associated with aggression or be-havior problems is reported in other studies on smaller samples. In a study of54 special education second- and third-grade children, Minutti (1991) foundthat among children in special classes, with either behavioral problems orlearning problems, language deficiencies of more than 2 standard deviationsbelow the normative mean identified those having more behavior problems,based on teacher ratings. The most important deficit in this subgroup was as-sociated with difficulty in formulating coherent sentence structures to conveymeaning. A more thorough investigation of pragmatic language developmentin association with physical aggression was conducted in two separate studiesby Cole (2001) and Mack and Warr-Leeper (1992). In the study by Cole(2001), aggressive (n = 19) and nonaggressive (n = 26) 8–13-year-old boyswere compared on standardized receptive and expressive language measures,as well as measures of syntactic and narrative complexity. Although thegroups did not differ on the composite scores of expressive and receptive lan-guage, the aggressive boys did perform more poorly on both the syntactic andnarrative complexity measures. Mack and Warr-Leeper (1992) had previouslyidentified similar problems in chronic-behavior-disordered boys, even once IQlevel was taken into account. In their sample, 16 of 20 boys scored below theaverage range on at least 4 of 20 language assessments. The main areas of lan-guage compromised were use of complex linguistic structures, use of abstract
338 D E T E R M I N A N T S O F A G G R E S S I O N

language, and use of concepts. More important, however, for half of theseboys, language assessments were below expected levels based on IQ. This sug-gests that pragmatic and sentence structure difficulties may be associated withthe more severe forms of behavior problems and aggression in childhood andearly adolescence, over and beyond IQ level.
Although these studies offer compelling results, inferences remain limitedby the fact that they are based on clinical populations of behaviorally compro-mised children and that, for the most part, these are mainly boys (from 75–100% of the samples). Do normative samples of boys and girls replicate theassociation between language deficits and conduct disorder or aggression inthis age range? For the most part, they do. Although total prevalence is higherin boys, the co-occurence rates with language impairments appear to be simi-lar for boys and girls (Stevenson, Richman, & Graham, 1985). In one of thefew studies looking simultaneously at physical and verbal aggression, Piel(1990) added another consideration to gender differences. He studied lan-guage maturity in association with responses on an apperceptive test of ag-gressive behavior in 108 second- and third-grade children. Although children’sbehaviors were not assessed directly, the best predictor of physically aggressiveresponses to vignettes was low language maturity, accounting for 20% of thevariance in both boys and girls. Sex and social class were not associated withthe selection of physically aggressive responses. Interestingly, girls did use ver-bally aggressive responses more often than boys, and language maturity wasassociated positively with this form of aggressive response in girls, albeit lessstrongly (4% of the variance). The more verbally proficient girls were moreprone to offer verbally aggressive responses to the vignettes. Even thoughapperceptive tests have important methodological limitations in generalizingto behavior, these results suggest that the developmental shift from physicalaggression to verbal aggression may be mediated by language maturity, yetdifferently in boys and girls.
Tomblin and colleagues (Tomblin, Zhang, Buckwalter, & Catts, 2000)offered yet another consideration in looking at the co-occurence of behaviorand language problems in school-age children. They studied language, read-ing, and behavior in a normative sample of second-grade boys and girls. Theoverall association between externalizing behavior problems assessed byteachers and parents and a composite language score (including expressiveand receptive vocabulary and pragmatic complexity) was –.30. Both were neg-atively associated with reading problems. Subgroups of children were thenidentified as language impaired or reading impaired. When investigating thepattern of association between reading, language, and behavior problems atthis age, log-linear model fitting indicated that behavior problems in the chil-dren with language impairments were mediated by reading problems whereasbehavior problems in the reading-impaired group were not mediated by lan-guage problems. This suggests that by the early school years, reading difficul-ties may act as an additional risk factor for behavioral problems, either linkedwith early language impairment or as a novel liability. Written forms of lan-
Language Development and Aggressive Behavior 339

guage may have specific links with aggression and externalizing behaviors thatneed to be addressed separately.
In sum, by the early school years and adolescence, many studies havedocumented the link between language proficiency and behavior problems.Although both problems are more prevalent in boys at this age, their rate ofco-occurrence appears generally similar across genders. Contrary to what wasobserved in the preschool years, studies specifically pertaining to aggressionshow patterns of co-occurences similar to those shown in studies of generalbehavior problems or conduct disorders. But although the co-occurence maybe of the same magnitude, these studies suggest that low pragmatic languageskills and difficulties with sentence structure may be more specifically associ-ated with aggression during this period.
Longitudinal Data: Do Early Language Deficits Predict Later Aggression?
Probably the most striking findings of the association between language andaggression are those reported in longitudinal lifespan studies. Although lan-guage has been a continuous focus in studies of early development, studies inadulthood shifted to low IQ as a concurrent correlate of physical aggression,delinquency, and criminality (Moffitt et al., 1981). Language proficiency mayhave appeared as an unlikely concurrent candidate by adulthood, as, for themost part, adults seem to have mastered the basic linguistic skills, even the lesseducated adults. Prospective longitudinal studies, however, tell a differentstory when early language skills are associated with adulthood outcomes.
But first, we consider longitudinal results during childhood. In three stud-ies described earlier, children assessed as preschoolers were followed into laterchildhood. In all of these studies, early behavior problems (ages 3–4) predictedlater behavior problems (at age 8 in Stevenson et al., 1985, and Benasich,Curtiss, & Tallal, 1993; at ages 7, 9, and 11 in Silva et al., 1987). However,early language delays were limited in predicting later behavior problems. InBenasich and colleagues’ U.S. sample of language-impaired preschoolers andcontrols, neither degree of language impairment at the initial assessment northe language improvement between assessments predicted behavioral out-comes at age 8. In Stevenson and colleagues’ U.K. sample, low language struc-ture at age 3 did not predict antisocial behavior scores at age 8 but did predictneurotic problems. In the N.Z. sample of Silva and colleagues, however, thegeneral language delay group at age 3 did score significantly higher for totalbehavior problems on the Rutter Teacher Scale at age 9 and on the Rutter Par-ent Scale at 11. Although behavior problems remained quite stable, the predic-tive value of early language impairment appears mitigated in these samples ofboys and girls.
The story, when looking at late adolescence and early adulthood follow-ups, is quite different. Beitchman’s Canadian sample of language-impairedchildren initially assessed at age 5 were followed well into late adolescence.Boys and girls with initial language impairement had strikingly divergent
340 D E T E R M I N A N T S O F A G G R E S S I O N

trajectories by age 19 (Beitchman et al., 1999). Both speech- and language-impaired girls showed a higher prevalence of psychiatric disorders thancontrols, predominantly affective and anxiety disorders. However, antisocialpersonality disorder was the most prevalent diagnosis for the boys in thelanguage-impaired group. Boys in this group were up to four times more likelyto have this diagnosis than controls. Similar results are shown for boys in rela-tion to registered criminality in a longitudinal follow-up of a sample of 122Swedish boys (Stattin & Klackenberg-Larsson, 1993). Infant measures of lan-guage development were conducted at 3, 6, 9, 12, 18, and 24 months. Of the122 boys, 36 were sporadic (n = 22) or frequent (n = 14) offenders by age17. Registered criminality was significantly but modestly correlated with6-, 18-, and 24-month infant language measures (correlations ranging from–.16 to –.19). Moreover, language ratings by a psychologist were obtained atages 3 and 5. Registered criminality was both related to boys language matu-rity at age 3 (r = –.16, p < .05) and age 5 (r = –.17, p < .05) and to themother’s self-reported difficulty in understanding the speech of the child atages 4 and 5. Intelligence measures were also obtained (ages 3, 5, 8, 11, 14,and 17 years), and all except one (age 8) predicted later registered criminality.However, once SES was controlled for, only the age 3 intelligence measure dif-ferentiated offenders from nonoffenders, but 18- and 24-month language abil-ity, 3-year comprehension of language, and 5-year maturity of language useand comprehension of language remained significant predictors of criminality.Thus, early language development appeared as the early aspect of these boys’lives that better distinguished later offenders from nonoffenders, even onceSES and IQ were taken into account.
The longitudinal picture these studies paint appears to vary according totwo criteria: gender and the specificity of the behavior measures. Longitudinaloutcomes for children with early language delays seem to differ considerablyas a function of gender. The antisocial trajectory, possibly more directly asso-ciated with the use of aggression, is up to 4 times more likely in boys with lan-guage impairments than in control boys, but almost 10 times more likely thanin girls with language impairments. Girls with language impairments at age 5do not even reach the rate of antisocial disorder in control boys. The specificbehavior measure also distinguishes the predictive value of early language def-icits. Results in childhood studies assessing general behavior problems aremore inconsistent. However, when criminality and antisocial disorders weretargeted, early language impairments distinguished offenders from nonof-fenders, beyond SES measures, whereas intelligence measures in childhood didnot.
Overall, the striking feature of the prevalence studies reviewed is not thestrength of the association between language deficits and aggression. Thecoprevalence may be high in clinical populations, but the linear association re-mains modest, with correlations ranging at best from –.17 to –.33. Rather, it isthe consistency with which results appear in many studies based on a varietyof designs, populations, age ranges, and even construct definitions.
Language Development and Aggressive Behavior 341

Is the Association between Language Proficiency and Aggression Linear?
A major question arises from the consistency of the modest association be-tween low language proficiency and aggression throughout development.Why are correlations so low when the prevalence of one problem can reach asmuch as 80% when children have the other? Low correlations, aside from in-dicating a modest association, could also stem from one of two other situa-tions: Either good language skills are not associated with lower aggressionscores—that is, the relationship is not linear—or heterogeneous subgroups,with respect to the association between language skills and aggression, createcompeting effects that outweigh each other in the assessment of a linear trend.
Although studies have looked at the higher co-occurrences of the prob-lematic aspects of language and aggression, few have considered looking atwhether good language skills predicted lower aggression scores. This questionis important, because if the trend is not linear in nature, the resulting correla-tions may have been underestimations of the true underlying relationship be-tween language impairment and aggression. In addition, if better languageskills are not associated with less aggression and more prosocial skills, thenthere is little to infer from these studies in terms of prevention. Post hoc analy-sis of the Québec sample offers some reassurances in that respect. At 19months, the infants above the 85th percentile for vocabulary development didshow a lower incidence of physically aggressive behaviors (p < .05), but moreimportant, they also showed a higher incidence of prosocial behaviors (p <.05). This trend persists at 30 months and is true for both boys and girls.Thus, language proficiency may be playing a protective role in the process ofearly socialization of behavior.
The other possibility for the modest correlations is that physical aggres-sion occurs in some verbally proficient individuals, just as some language-impaired individuals show no signs of using aggression at higher rates. Inother words, language deficits, although they occur more frequently in aggres-sive individuals, may lead to other behavioral and emotional problems not in-volving aggression, or to no other specific problem. In our attempts to makesense of co-occurring milestones of language and socialized behavior intoddlerhood, we may have overlooked the possibility that distinct subgroupsof aggressive individuals with differing levels of language proficiency mayemerge in early development. In an examination of communicative strategiesin preschool classroom disputes, Danby and Baker (2001) have shown howdominant boys in such group disputes use a variety of verbal strategies to as-sert their identity, construct group affiliations, and generate terror in targetedrejected children. Although it is unclear whether these boys are in fact moreproficient verbally, it is clear that they are able to use language to attain goalsinvolving some forms of aggression. If such different subgroups exist, the eti-ology of aggression accompanied by low language proficiency may differ fromthe etiology of aggression in verbally proficient individuals. Different statisti-cal strategies are needed if we are to distinguish these nonlinear associations
342 D E T E R M I N A N T S O F A G G R E S S I O N

and, more important, identify individuals following at-risk developmental tra-jectories for aggression with or without language impairment. This possiblemissing piece to the puzzle suggests that we may only be starting to under-stand the complexity of the interface between language and aggression, and tosome degree, between language and socialized behavior.
THEORETICAL MODELS OF THE LANGUAGE–AGGRESSION ASSOCIATION
There have been numerous theoretical propositions put forth to explain thelanguage–aggression association. By and large, these fall within two sets of de-velopmental models. Models of the first type are those that assume languagedeficits and aggression share similar origins. The second type of models arethose that contend that one problem leads to the other as a secondary conse-quence, either via distinct pathways or through a combination of multiplepathways acting in transaction, or that both problems feed on each other dur-ing development, creating a consolidation of the co-occurence in time.
Shared Etiology Pathways
There are three types of shared etiology models that have been suggested toexplain the language–aggression association. First, there are models that implythat both language deficits and aggression may stem from the same geneticand biologically based predispositions (Stevenson, 1996). The origins of bothproblems would then be organic in nature. Stevenson (1996) has demon-strated that ADHD and reading problems do share a common genetic etiologyin school-age children. However, even though language and reading problemsoften co-occur, and aggression and ADHD often co-occur, there are no studiesshowing that this generalizes to language deficits and aggression. In fact, boththe Québec Newborn Twin Study and the U.K. Twin Early DevelopmentStudy have found no evidence of shared genetic influences—between physicalaggression and delayed vocabulary in the Québec sample, and between gen-eral conduct problems and delayed vocabulary in the U.K. sample. The sharedgenetic liability pathway thus seems unlikely in early childhood. However, asgenetic influences on most traits tend to increase with time, a possible sharedgenetic etiology may emerge only later. In addition, empirical tests of thismodel should take into account the possibility that the etiology of the associa-tion of behavior and language may vary, depending on the specific behaviorand language deficits.
Second, there are models that suggest at-risk environmental features canlead to both unsocialized behavior and poor cognitive-intellectual develop-ment, including or secondary to poor language skills. In this macro-socialpathway, parenting styles and harsh economic conditions are viewed as envi-ronmental liabilities that steer children into trajectories of poorly socializedbehavior and leave them poorly stimulated in language development. The link
Language Development and Aggressive Behavior 343

between aggression and parenting has been documented in numerous studies,such as in work by Diana Baumrind (1993) and Gerald Patterson and his col-leagues (Patterson, 1982; Patterson, Reid, & Dishion, 1992) at the OregonSocial Learning Center. These have generally demonstrated that inconsistantand harsh parenting, with poor child supervision, create a context withinwhich a child is more susceptible to develop behavior problems. In a similarfashion, but within a completely different field, psycholinguists have demon-strated how parental input (moderated by such factors as socioeconomic sta-tus, parental education levels, and neighborhood safety, to name a few) has aprofound effect on children’s language development (Storch & Whitehurst,2001). Michael Tomasello (2003) offered compelling empirical evidence of theeffect of input in tasks of non-word learning in helping a child learn newwords to designate novel objects. Others have shown how parents’ reading tochildren (Lyytinen, Laakso, & Poikkeus, 1998; Sénéchal, LeFèvre, Thomas, &Daley, 1998), and parents’ rephrasing of early language mistakes (Bohannon& Stanowicz, 1989) also play a role in the development of language in chil-dren. The absence of these forms of early language stimulation is more likelyto be accompanied by parental discipline lacking in attention to a child’sneeds. Such parental discipline may be characterized by harshness and lack ofappropriate supervision, often associated with behavior problems in children,and may account for the association of problems regarding both language andbehavior.
Finally, the co-occurence of both traits has been suggested to be linked tothe same cognitive deficit, such that poor judgment and/or lack of ability tounderstand others’ perspectives may be accompanied by language difficultiesbut also heighten the risk for social exclusion and deviant behaviors. The nextsection of this chapter explores in more detail the theoretical propositions in-volving a causal contribution of language to the development of aggression.
Causal Pathways
Causal models of the language–behavior association have received more atten-tion than their shared etiology counterparts. The prevailing hypothesis in thisrespect suggests that low language proficiency makes one more liable to useaggression (Cohen, 2001). Two studies have conducted direct tests of this hy-pothesis, the Québec Newborn Twin Study (Dionne et al., 2003) and an inter-vention study conducted more than three decades ago (Slaby & Crowley,1977). In the Québec study, the investigators compared models specifyingshared etiological contributions and models testing the significance of unidi-rectional and reciprocal contribution paths from language to aggression at19 months. These analyses were conducted using structural equation model-ing and the additional statistical flexibility offered by genetically informativedata. Results indicated no significant shared etiology component, whether ge-netic or linked to environments shared by twins at this age, except for aunique environmental component specific to twins within families. However,
344 D E T E R M I N A N T S O F A G G R E S S I O N

causal genetic models revealed that a causal path from vocabulary to physicalaggression could account totally for the covariance between the two. Assump-tions of the genetic methodology should, however, be taken into considerationin generalizing these results. In addition, the results remain specific to the timeframe for which models were tested.
The most compelling evidence of causal effects comes from experimentaldesigns using and assessing interventions. Many preschool programs targetingat-risk children have an incorporated intervention focus on language and liter-acy, yet these rarely supply a proper control group to assess the efficacy of in-terventions. In a study conducted in the late 1970s (Slaby & Crowley, 1977),when the language–aggression hypothesis attracted attention, disruptive chil-dren were identified in classroom settings. Half of them received an interven-tion based on increasing their linguistic skills in social contexts, the other halfdid not. Teachers reported that the children having received the interventionhad significantly decreased their use of disruptive behavior in class and aggres-sion in the schoolyard. There was no significant change in the noninterventiongroup. The authors concluded that these results were indicative of a causalcontribution of enhanced language proficiency toward more socialized behav-ior. Because there was not a separate group receiving intervention targetingthe behavior problems, it is difficult to conclude that regulating behavior doesnot also contribute to better language learning. Although there were manymethodological limitations to this small study, it serves to highlight the neces-sity of conducting experimental intervention studies to assess causal path-ways.
Another limitation of these two studies is that although they suggest thatlanguage proficiency can affect behavior, they do not address the mechanismspossibly involved. Three major views on how language deficits can lead to be-havior problems, including aggression but not specific to it, have been out-lined. They are referred to here as the micro-social pathway, the self-regulationpathway, and the social-cognition pathway.
The first of these views assume that language plays a significant role insustaining fulfilling social interactions. This micro-social pathway, along withothers reviewed later, has been formalized and explored by Brinton, Fujiki,and colleagues (Brinton & Fujiki, 1993; Brinton, Fujiki & McKee, 1998). Inone study, language-delayed school-age children were compared with controlsin regard to their negotiation skills in a series of negotiating sequences withtwo other peers. As compared with controls, the language-delayed childrencontributed significantly less and developmentally lower-level negotiationstrategies to their teams. Interestingly, the language-delayed children did notspeak less often than controls. Rather, the maturity level of their verbal contri-butions to the team was what distinguished them from their peers. The au-thors caution that lack of efficient negotiation skills may place the language-delayed children at risk for confrontations with peers and lead them to adoptmore disruptive behaviors.
Brinton and Fujiki have also documented the fact that language-delayed
Language Development and Aggressive Behavior 345

children show more emotion regulation problems (Fujiki, Brinton, & Clarke,2002). Cohen (2001) points out that language is an important means bywhich one controls one’s behavior and emotions. This notion goes back toearly propositions by Luria (1961) and Vygotsky (1962) to the effect that in-ner language is a necessary tool to inhibit antisocial impulses, among otherthings. Emotion regulation and self-regulation are generally viewed as requir-ing complex linguistic tools such as the ability to analyze social situations, or-ganize thoughts about one’s own emotions, and plan behavior according tosocial rules. Given this central role of language in emotion regulation, in theself-regulation pathway, children with language difficulties are expected to beat a disadvantage (Gallagher, 1999). There is some evidence showing thatlanguage-delayed children are more at risk of also having regulation prob-lems. In a study of 82 school-age children, half of whom were language de-layed, Fujiki and colleagues (Fujiki et al., 2002) reported lower scores overallfor the language-delayed children on the Emotion Regulation Checklist, espe-cially for boys. Although emotion regulation problems may stem fromoverregulation as well as underregulation (Cole, Michel, & O’Donnell-Teti,1994), it may be expected that the underregulated children would be the onesat risk for aggression. Empirical evidence is, however, not yet available to sup-port the assumption that the contribution of language delays to aggression ismediated by underregulation problems.
Social cognition pathways offer yet other explanations for possible medi-ators of language deficits leading to aggressive behaviors. Among these,Dodge’s social information-processing model suggests that aggression devel-ops as a consequence of deficits in social problem-solving abilities. The modelassumes that there are six steps involved in the social problem-solving process.Once a child has (1) encoded cues, the child must (2) interpret those cues be-fore deciding (3) what his or her goal is, (4) constructing his or her response,(5) choosing it, and (6) acting on it (Crick & Dodge, 1994). Although aggres-sive children tend to construct more aggressive responses and act on them,Lochman and Dodge (1994) have shown that what mainly distinguishes themfrom nonaggressive children is the tendency to perceive hostile intent in am-biguous situations. This presumably causes them to act on these perceivedhostile intents by choosing aggressive responses. It is less clear, however, howlanguage delay may lead a child into this attribution biases pathway. If, asCohen (2001) points out, language-delayed children have had more negativesocial encounters, and have more difficulty in making complex analysesbecause of their linguistic limitations, they may in fact be more at risk offorming hostile attributions. However, this needs to be documented empiri-cally. In addition, the attribution biases pathway seems an unlikely explana-tion for aggression that appears as early as late infancy.
The same can be said about another approach to the social cognitionpathway. Theory of mind has been receiving much attention in recent years asa possible central cognitive impairment, linked to autism in particular. Theoryof mind involves understanding that individuals may have different views
346 D E T E R M I N A N T S O F A G G R E S S I O N

about a given situation, and that sometimes one may act on the basis of an as-sumption that appears true, which is in fact a false belief (Dunn, 1996).Children begin to be able to consider another person’s perspective, and under-stand that thoughts, beliefs, and emotions are determined by a person’s uniqueperspective, at about age 4. Autistic children do not master this task (Wimmer& Perner, 1983). Happé and Frith (1996) have shown that children withsocial/emotional problems have more problems mastering this task as well,and Cutting and Dunn (1999) have shown theory of mind mastery to be moredifficult for language-delayed children. However, there is no evidence to sup-port a theory of mind pathway by which the effects of language delay onsocio-emotional problems and/or aggression are mediated.
Finally, although it has received much less attention, the alternativecausal pathway, whereby aggression leads to secondary language delays, hasnot been ruled out. For instance, aggressive individuals could be less sensitiveto linguistic stimulation in their environment or too disrupted by their ownbehavior to benefit from this stimulation, leaving them with linguistic deficitsthey are unable to overcome. As Cantwell and Baker (1977) pointed outdecades ago, the main impediment to this hypothesis has been that languagedelays have always appeared to precede the onset of aggressive behaviors indevelopment. But because overt aggression has rarely been assessed aroundthe time language appears, their conclusion may have been premature. Untilproven otherwise, we should consider the possibility that aggression may playthe causal role at one moment of development, and not at another, or bothmay influence each other reciprocally. For example, language-based formalschooling represents a specific time frame within which specific demands areplaced on children, requiring both good language skills and social skills. If lowlanguage skills appear to drive aggressive behavior in early childhood, can weinfer that low language skills drive aggressive behavior once schooling starts?Or does aggression lead to social interactions and choices of peers that haveadverse consequences on schooling and limit the sophistication of languageskills that comes with schooling? If this is the case, concurrent language skillsmay not be the best focus for intervention with adult delinquents or violentcriminals, but may be targeted in early development to promote prosocial de-velopment. For now, we are unable to discern whether any or all of these al-ternatives are at play.
LOOKING FOR ANSWERS
As this review illustrates, research designs of the past decades have beenmainly concerned with documenting the prevalence of and, to a lesser extent,exploring the etiology of the language–behavior association. The initial clini-cal observations of Orton (1937) and Chess (1944) hold true across manysamples, large and small, from the normative to the clinical, concurrently andlongitudinally. Research on aggression, although more scarce, also suggests
Language Development and Aggressive Behavior 347

language and aggression are associated. That much we know. Any answers tothe question concerning how language and aggression become linked duringdevelopment, however, are at best, tentative for the time being. For the mostpart, we are left to speculate, given incomplete correlational information.
To be more efficient, research designs will need to incorporate measuresof mediating variables, such as economic considerations, concurrent measuresof IQ, literacy, and discipline-oriented parental practices, to assess their spe-cific contributions in developmental pathways. In addition, aggression shouldbe targeted specifically if designs seek to understand the specificity of thelanguage–aggression association. Similarly, a survey of language skills shouldinclude the full array of linguistic abilities. To assess possible shared physio-logical contributions to both problems, lower-level cortical functioning, suchas executive functions and time processing, could be assessed. Obviously, gen-der differences in both prevalence and developmental mechanisms need toconsidered. Finally, the true test of causality rests on experimental designs. Toscan the full array of possibilities, such designs should divide groups to con-sider the etiology and course of aggression and language deficits separately, aswell as in association, and study the impact of interventions on both aspectslongitudinally.
REFERENCES
Baumrind, D. (1993). The average expectable environment is not good enough: A re-sponse to Scarr. Child Development, 64, 1299–1317.
Beitchman, J. H., Cohen, N. J., Konstantareas, M. M., & Tannock, R. (1996). Language,learning, and behavior disorders: Developmental, biological, and clinical perspec-tives. New York: Cambridge University Press.
Beitchman, J. H., Douglas, L., Wilson, B., Johnson, C., Young, A., Atkinson, L., et al.(1999). Adolescent substance use disorders: Findings from a 14-year follow-up ofspeech and language impaired and control children. Journal of Clinical Child Psy-chology, 28, 312–321.
Beitchman, J. H., Nair, R., Clegg, M., Ferguson, B., & Patel, P. G. (1985). Prevalence ofpsychiatric disorders in children with speech and language disorders. Journal ofAmerican Academy of Child Psychiatry, 25, 528–535.
Benasich, A. A., Curtiss, S., & Tallal, P. (1993). Language, learning, and behavioral dis-turbances in childhood: A longitudinal perspective. Journal of the American Acad-emy for Child and Adolescent Psychiatry, 32, 585–594.
Bjorkqvist, K., Osterman, K., & Kaukiainen, A. (1992). The development of direct andindirect aggressive strategies in males and females. In K. Bjorkqvist & P. Niemela(Eds.), Of mice and women: Aspects of female aggression (pp. 51–64). San Diego:Academic Press.
Boone, S. L., & Montare, A. (1976). Test of the language–aggression hypothesis. Psycho-logical Reports, 39, 851–857.
Brinton, B., & Fujiki, M. (1993). Clinical forum: Language and social skills in the school-age population: Language, social skills, and socioemotional behavior. Language,Speech, and Hearing Services in Schools, 24, 194–198.
348 D E T E R M I N A N T S O F A G G R E S S I O N

Brinton, B., Fujiki, M., & McKee, L. (1998). Negotiation skills of children with specificlanguage impairment. Journal of Speech, Language, and Hearing Research, 41,927–940.
Camarata, S., Hughes, C., & Ruhl, K. (1988). Mild/moderate behaviorally disorderedstudents: A population at risk for language disorders. Language, Speech, and Hear-ing Services in Schools, 19, 191–200.
Cantwell, D. P., & Baker, L. (1977). Psychiatric disorder in children with speech and lan-guage retardation: A critical review. Archives of General Psychiatry, 34, 583–591.
Cantwell, D. P., & Baker, L. (1987). Clinical significance of children with communicationdisorders: Perspectives from a longitudinal study. Journal of Child Neurology, 2,257–264.
Carson, D. K., Klee, T., Perry, C. K., Muskina, G., & Donaghy, T. (1998). Comparison ofchildren with delayed and normal language at 24 months of age on measures of be-havioral difficulties, social and cognitive development. Infant Mental Health Jour-nal, 19, 59–75.
Chess, S. (1944). Developmental language disability as a factor in personality distortionin childhood. American Journal of Orthopsychiatry, 14, 483–490.
Cohen, N. J. (1998). Language impairments and psychopathology [Letter to the edi-tor]. Journal of the American Academy for Child and Adolescent Psychiatry, 37,461–462.
Cohen, N. J. (2001). Language development and psychopathology in infants, childrenand adolescents. Thousand Oaks, CA: Sage.
Cohen, N. J., Davine, M., Horodezky, N., Lipsett, L., & Isaacson, L. (1993). Unsus-pected language impairment in psychiatrically disturbed children: Prevalence andlanguage and behavioral characteristics. Journal of American Academy for Childand Adolescent Psychiatry, 32, 595–603.
Cole, D. (2001). Narrative development in aggressive boys. Behavioral Disorders, 26,332–343.
Cole, P. M., Michel, M. K., & O’Donnell-Teti, L. (1994). The development of emotionregulation and dysregulation: A clinical perspective. Monographs of the Society forResearch in Child Development, 59(Serial No. 240).
Crick, N. R., & Dodge, K. A. (1994). A review and reformulation of social information-processing mechanisms in children’s social adjustment. Psychological Bulletin, 115,74–101.
Cutting, A. L., & Dunn, J. (1999). Theory of mind, emotion understanding, language,and family background: Individual differences and interrelations. Child Develop-ment, 70, 853–865.
Dale, P. S., Price, T. S., Bishop, D. V. M., & Plomin, R. (2003). Outcome of early languagedelay: I. Predicting persistent and transient delay at 3 and 4 years. Journal of SpeechLanguage and Hearing Research, 46(3), 544–560.
Danby, S., & Baker, C. D. (2001). Escalating terror: Communicative strategies in a pre-school classroom dispute. Early Education and Development, 12, 343–358.
Dionne, G., Tremblay, R., Boivin, M., Laplante, D., & Pérusse, D. (2003). Physical ag-gression and expressive vocabulary in 19–month-old twins. Developmental Psy-chology, 39, 261–273.
Donahue, M., Cole, D., & Hartas, D. (1994). Links between language and emotional/be-havioral disorders. Education and Treatment of Children, 17, 246–254.
Dunn, J. (1996). Children’s relationships: Bridging the divide between cognitive and so-cial development. Journal of Child Psychology and Psychiatry, 37, 507–518.
Language Development and Aggressive Behavior 349

Fujiki, M., Brinton, B., & Clarke, D. (2002). Emotion regulation in children with specificlanguage impairment. Language, Speech, and Hearing Services in Schools, 33, 102–111.
Gallagher, T. M. (1999). Interrelationships among children’s language, behavior, andemotional problems. Topics in Language Disorders, 19, 1–15.
Goodenough, F. L. (1931). Anger in young children. Minneapolis: University of Minne-sota Press.
Goodyer, I. M. (2000). Language difficulties and psychopathology. In D. V. M. Bishop &L. B. Leonard (Eds.), Speech and language impairments in children: Causes, charac-teristics, intervention, and outcome (pp. 227–244). Philadelphia: Psychology Press.
Gualtieri, T., Koriath, U., Van Bourgondien, M., & Saleeby, N. (1983). Language disor-ders in children referred for psychiatric services. Journal of the American Academyof Child and Adolescent Psychiatry, 22, 165–171.
Happé, F., & Frith, L. (1996). Theory of mind and social impairment in children withconduct disorder. British Journal of Developmental Psychology, 14, 385–398.
Hay, D. F., Castle J., & Davies L. (2000). Toddlers’ use of force against familiar peers: Aprecursor of serious aggression? Child Development, 71, 457–467.
Howlin, P., & Rutter, M. (1987). The consequences of language delay for other aspects ofdevelopment. In W. Yule & M. Rutter (Eds.), Language development and disorders(pp. 271–294). Oxford, UK: Blackwell.
Keenan, K., & Shaw, D. S. (1993). The development of aggression in toddlers: A study oflow income families. Journal of Abnormal Child Psychology, 22, 53–77.
Lahey, B. B., Pelham, W. E., Stein, M. A., Loney, J., Trapani, C., Nugent, K., Kipp, H.,Schmidt, E., Lee, S., Cale, M., Gold, E., Hartung, C. M., Willcutt, E., & Baumann,B. (1998). Validity of DSM-IV attention-deficit/hyperactivity disorder for youngerchildren. Journal of the American Academy for Child and Adolescent Psychiatry,37, 695–702.
Lochman, J. E., & Dodge, K. A. (1994). Social-cognitive processes of severely violent,moderately aggressive, and nonaggressive boys. Journal of Consulting and ClinicalPsychology, 62, 366–374.
Love, A., & Thompson, M. G. G. (1988). Language disorders and attention deficit disor-ders in young children referred for psychiatric services. Journal of Orthopsychiatry,58, 52–63.
Luria, A. R. (1961). The role of speech in the regulation of normal and abnormal behav-ior. Oxford, UK: Pergamon Press.
Lyytinen, P., Laasko, M.-L., & Poikkeus, A.-M. (1998) Parental contribution tochild’s early language and interest in books. European Journal of Psychology ofEducation, 13(3), 297–308.
Mack, A. E., & Warr-Leeper, G. A. (1992). Language abilities in boys with chronicbehavior disorders. Language, Speech, and Hearing Services in Schools, 23, 214–223.
Miniutti, A. M. (1991). Language deficiencies in inner-city children with learning and be-havioral problems. Language, Speech, and Hearing Services in Schools, 22, 31–38.
Moffitt, T. E., Gabrielli, W. F., Mednick, S. A., & Schulsinger, F. (1981). Socioeconomicstatus, IQ, and delinquency. Journal of Abnormal Psychology, 90, 152–156.
Montare, A., & Boone, S. L. (1973, February). Language and aggression, an exploratorystudy among black, Puerto Rican youth. Paper presented at the meeting of the Edu-cational Research Association, New Orleans.
350 D E T E R M I N A N T S O F A G G R E S S I O N

Ortiz, C., Stowe, R., & Arnold, D. H. (2001). Parental influence on child interest inshared picture book reading. Early Childhood Research Quarterly, 16, 263–281.
Orton, S. T. (1937). Reading, writing and speech problems in children. New York:Norton.
Patterson, G. R. (1982). Coercive family processes. Eugene, OR: Castalia.Patterson, G. R., Reid, J. B., & Dishion, T. (1992). Antisocial boys. Eugene, OR:
Castalia.Pavlov, I. P. (1927). Conditioned reflexes (G. Anrep, Trans.). London: Oxford University
Press.Pepler, D. J., Craig, W. M., & Roberts, W. L. (1998). Observations of aggressive and
nonaggressive children on the school playground. Merrill-Palmer Quarterly, 44,55–76.
Piel, J. A. (1990). Unmasking sex and social class differences in childhood aggression:The case for language maturity. Journal of Educational Research, 84, 100–106.
Plomin, R., Price, T. S., Eley, T. C., Dale, P. S., & Stevenson, J. (2002). Associations be-tween behaviour problems and verbal and nonverbal cognitive abilities and disabili-ties in early childhood. Journal of Child Psychology and Psychiatry, 43, 619–633.
Sénéchal, M., LeFèvre, J.-A., Thomas, E., & Daley, K. (1998). Differential effects ofhome literacy experiences on the development of oral and written language. Read-ing Research Quarterly, 32, 96–116.
Silva, P. A. (1987). Epidemiology, longitudinal course and some associated factors: Anupdate. In W. Yule & M. Rutter (Eds.), Language development and disorders (pp.1–15). Oxford, UK: Blackwell.
Silva, P. A., Williams, S., & McGee, R. (1987). A longitudinal study of children with de-velopmental language delay at age three: Later intelligence, reading and behaviourproblems. Developmental Medicine and Child Neurology, 29, 630–640.
Slaby, R. G., & Crowley, C. G. (1977). Modification of cooperation and aggressionthrough teacher attention to children’s speech. Journal of Experimental Child Psy-chology, 23, 442–458.
Stattin, H., & Klackenberg-Larsson, I. (1993). Early language and intelligence develop-ment and their relationship to future criminal behavior. Journal of Abnormal Psy-chology, 102, 369–378.
Stevenson, J. (1996). Developmental changes in the mechanisms linking language dis-abilities and behavior disorders. In J. Beitchman, N. Cohen, M. Konstantareas, &R. Tannock (Eds.), Language, learning and behaviour disorders (pp. 78–100). Cam-bridge, UK: Cambridge University Press.
Stevenson, J., & Richman, N. (1978). Behaviour, language and development in three-year-old children. Journal of Autism and Childhood Schizophrenia, 8, 299–313.
Stevenson, J., Richman, N., & Graham, P. (1985). Behaviour problems and languageabilities at three years and behavioural deviance at eight years. Journal of Child Psy-chology and Psychiatry, 26, 215–230.
Storch, S., & Whitehurst, G. J. (2001). The role of family and home in the literacy devel-opment of children from low-income backgrounds. In P. R. Britto & J. Brooks-Gunn (Eds.), The role of family literacy environments in promoting young chil-dren’s emerging literacy skills: New directions for child and adolescent development(pp. 53–77). San Francisco: Jossey-Bass.
Tomasello, M. (2003). Constructing a language: A usage-based theory of language acqui-sition. Cambridge, MA: Harvard University Press.
Tomblin, J. B., Zhang, X., Buckwalter, P., & Catts, H. (2000). The association of reading
Language Development and Aggressive Behavior 351

disability, behavioral disorders, and language impairment among second-gradechildren. Journal of Child Psychology and Psychiatry, 41, 473–482.
Tremblay, R. E. (2000). The development of aggressive behaviour during childhood:What have we learned in the past century? International Journal of Behavioural De-velopment, 24, 129–141.
Tremblay, R. E., Boulerice, B., Harden, P. W., McDuff, P., Pérusse, D., Pihl, R. O., &Zoccolillo, M. (1996). Do children in Canada become more aggressive as they ap-proach adolescence? In Human Resources Development Canada and Statistics Can-ada (Eds.), Growing up in Canada: National Longitudinal Survey of Children andYouth (pp. 127–137). Ottawa: Statistics Canada.
Vygotsky, L. S. (1962). Thought and language. New York: Wiley.Wimmer, H., & Perner, J. (1983). Beliefs about beliefs: Representation and the constrain-
ing function of wrong beliefs in young children’s understanding of deception. Cog-nition, 13, 103–128.
352 D E T E R M I N A N T S O F A G G R E S S I O N

DETERMINANTS OF AGGRESSIONIntergenerational Transmission of Aggression
17
The Intergenerational Transmissionof Aggression and Antisocial Behavior
MARK ZOCCOLILLO, ELISA ROMANO, DAVID JOUBERT, TANIA MAZZARELLO,SYLVANA CÔTÉ, MICHEL BOIVIN, DANIEL PÉRUSSE, and RICHARD E. TREMBLAY
In this chapter the intergenerational transmission of childhood-onset persis-tent aggression (CPA) is reviewed. The evidence for both genetic and environ-mental risk is presented. The association between parental antisocial behaviorand environmental risk factors is examined, and some new data from a largelongitudinal study are presented. The difficulties posed by the strong asso-ciation between parental antisocial behavior and environmental risk factorsand the lack of adequate control for parental antisocial behavior andgene–environment correlation in past studies are also discussed. The chapterconcludes with a discussion of how future studies of risk factors for childhood-onset persistent aggression can be improved by incorporating assessments ofparental antisocial behavior when examining environmental risk and by stud-ies of the early environment of children with antisocial parents.
CHILDHOOD-ONSET PERSISTENT AGGRESSIONAND THE PROBLEM OF DEFINING AGGRESSION
A recent review of the development of aggressive behavior has noted thatdefining aggression in studies has been problematic: “The aggregation of dif-ferent types of aggressive behaviours, and the aggregation of aggressive behav-iours with different forms of antisocial behaviour clearly creates an importantproblem for a developmental science aiming to understand the origin and de-velopment of these behaviours” (Tremblay, 2000, p. 130). For example, theaggression scale for one of the few longitudinal and intergenerational studies
353

of aggressive behavior included the following items: disobeys teacher, givesdirty looks, makes up stories and lies, does things that bothers others, gets introuble (Huesmann, Eron, Lefkowitz, & Walder, 1984). Another problem inregard to studying aggression has been requiring an intent to harm as part ofthe definition. This would preclude studies of aggressive behavior in infantswhere intent to harm cannot be assessed, and of aggression following anger orfear as “anger and fear [that] lead to reactions which are clearly not under thecontrol of anyone’s will” (Tremblay, 2000, p. 131).
Nonetheless, despite these problems with defining aggression, several lon-gitudinal studies of physical aggression in children (rigorously defined asfighting, bullying, or threatening others) have all concluded that there is asmall group of children who are persistently aggressive from preschool intoadolescence or adulthood (Broidy et al., 2003). Furthermore, children in thishigh-aggression group are much more likely to engage in violent delinquencythan nonaggressive children. It is this childhood-onset persistent aggression(CPA) that is the focus of this chapter.
Studies of the intergenerational transmission of rigorously defined CPAare few. There are many more studies of more broadly defined childhood-onset and persistent antisocial behavior (CPASB), which includes both aggres-sive and nonaggressive antisocial behaviors (Zoccolillo, Price, Hwu, & Ji,1998; Moffitt, Caspi, Rutter, & Silva, 2001; Robins, 1966). This group hasbeen given different names depending on the domain of research, includingthe diagnoses of antisocial personality disorder (APD) (within the DSM-IV(American Psychiatric Association, 1994) diagnostic framework) and childhood-onset and life course persistent antisocial disorder (Moffitt et al., 2001).
Individuals who are severely and persistently aggressive also engage inless serious and nonviolent antisocial behaviors (Zoccolillo et al., 1998;Robins, 1996; Moffitt et al., 2001; Loeber, Burke, Lahey, Winters, & Zera,2000). Conversely, among both men and women who met full DSM-III diag-nostic criteria for APD in a general population survey of the prevalence of psy-chiatric disorders in the United States, 85% of men and 83% of women withAPD met the criterion for violent behavior defined as follows: hit or threwthings at partner more than once, spanked a child hard enough to bruise, hadmore than one physical fight (other than with spouse) (Robins & Regier,1991, p. 261). Children with rigorously defined CPA are also more likely toengage in nonviolent delinquency (Broidy et al., 2003). Studies that focus onthe more broadly defined CPASB are also informative in regard to CPA andare included in this chapter. Studies of rigorously defined CPA are highlighted.
WHAT CAUSES CHILD-ONSET PERSISTENTANTISOCIAL BEHAVIOR OR AGGRESSION?
Two recent studies have focused specifically on risk factors for childhood per-sistent aggression. In one study, risk factors for high versus low trajectories of
354 D E T E R M I N A N T S O F A G G R E S S I O N

physical aggression, defined using three items (fights with other children;kicks, bites, or hits other children; bullies or intimidates other children), wereexamined in boys of ages 6–15 from a high-risk sample (Nagin & Tremblay,2001). Maternal low education status, teenage mother, and family not intactwere environmental variables that were significantly associated with trajecto-ries of high aggression. In a study of infants’ trajectories of aggression from 17to 42 months of age in a random population sample, using the same items foraggression, three trajectories of aggression were found, including one groupwith stable high aggression (Tremblay et al., 2004). Risk factors for this tra-jectory of high stable aggression included young siblings, low family income,early motherhood, mother’s own history of conduct problems, maternalsmoking during pregnancy, high family dysfunction, and maternal coerciveparenting.
Loeber and Farrington (2000) have recently reviewed the literature onyoung children who commit crime, and listed 40 factors across five domains(child, family, school, peers, and neighborhood). Hinshaw has reviewed is-sues related to process and mechanism in regard to externalizing behavior(Hinshaw, 2002). In commenting on the list of risk factors generated byLoeber and Farrington, he notes “The sheer size of this list, which must beconsidered only a partial consideration of potential risk factors, betrays thefield’s lack of ability to synthesize or to tell a fully coherent story about thedevelopment and maintenance of externalizing behavior.” In regard tomethodologic issues and promising practices for the field, he warns investi-gators of psychosocial influences: “Environmental factors may well accountfor independent variance in the field; but the typical means of inferring theireffects (i.e., correlating a socialization variable with a child outcome) aresufficiently confounded and uninformative that considerably more sophisti-cated tests are in order.” As to promising avenues of investigation, he high-lights the importance of genetically informative designs and conceptuallydriven investigations of interactions and transactions across multiple levelsof influence.
CPASB/CPA IS A FAMILIAL DISORDER
A major, and often neglected, aspect of CPASB/CPA is that it is a familial dis-order. A recent meta-analysis has examined the association separately betweenmaternal and paternal antisocial personality disorder and offspring external-izing disorders and found significant associations between both mother andfather antisocial personality disorder and child externalizing disorder (Connell& Goodman, 2002). The weighted mean r for mothers was .17 and for fathers.16, or small effect sizes. A number of intergenerational studies have foundthat CPASB/CPA runs in families. As noted earlier, maternal history of con-duct problems was a significant risk factor for persistent high levels of aggres-sion in infants.
Intergenerational Transmission of Aggression 355

Why Does Antisocial Behavior and Aggression Run in Families?
Familial transmission can be due to genes, environment, or both. Geneticallyinformative designs can disentangle genetic and environmental effects throughtwin and adoption studies (Plomin, 1997; Lahey et al., 1998). Environmentalrisk (e.g., hostile parenting) in parents with antisocial disorder is likely to af-fect all children in a family. If shared environment effects are found, this sug-gests (but does not prove) that early-onset antisocial disorder runs in families,in part due to cultural transmission.
A longitudinal twin study has attempted to address the specific contribu-tions of genetic influences, shared environment influences, and nonshared en-vironmental influences separately for aggressive and nonaggressive antisocialbehavior (Eley, Lichtenstein, & Moffitt, 2003). This study is of particular in-terest because both aggression and nonaggressive antisocial behavior, childversus adolescent onset, and persistence were examined. Twin pairs (n = 1,232)from the population-based Swedish Twin Registry were assessed at two differ-ent ages (8–9 years and 13–14 years). Aggressive antisocial behavior was ratedby a parent using the Aggression subscale of the CBCL, and nonaggressive an-tisocial behavior was rated using the Delinquency subscale of the CBCL. Atboth ages aggressive and nonaggressive antisocial behavior were highly corre-lated. At age 8–9 aggressive behavior was highly heritable, with little evidencefor shared environment. Nonaggressive antisocial behavior was influenced byboth genes and shared environment. At age 13–14 both aggressive andnonaggressive antisocial behavior were influenced by genes and shared envi-ronment. Continuity in aggression was largely mediated by genetic influences,and continuity in nonaggressive antisocial behavior was mediated by bothgenes and shared environment. A limitation of this study is that the measureof aggression included nonaggressive items such as arguing a lot, stubborn,and jealous.
A meta-analysis of 51 twin and adoption studies was conducted recently(Rhee & Waldman, 2002). Antisocial behavior was operationalized four dif-ferent ways: (1) diagnosis of antisocial personality disorder or conduct disor-der by DSM criteria, (2) criminality or delinquency by official records, (3) be-havioral aggression, and (4) antisocial behavior (antisocial behaviors but notoperationalized as diagnoses). For diagnosis, aggression, and antisocial behav-ior, the models that included additive genetic effects, shared environmental ef-fects, and nonshared environmental effects were the best-fitting models, withgenetic effects explaining between 44 and 47% of the variance.
CPASB/CPA and Environmental (Nongenetic) Risk
Given the evidence that there is a shared environment effect on CPASB and,possibly, aggression, what might the risk factors be? This is a difficult questionto answer because of possible gene–environment correlations. In regard to en-vironmental risk factors, three questions can be asked:
356 D E T E R M I N A N T S O F A G G R E S S I O N

1. What are the potential environmental (nongenetic) mechanisms bywhich antisocial behavior may be transmitted within families andwhich occur early enough (prenatally to kindergarten) to precede orcoincide with the first signs of early-onset antisocial disorder or ag-gression?
2. Are these risk factors confounded with genetic risk for early antisocialdisorder or aggression, as measured by either parental antisocial be-havior or aggression or through genetically informative designs?
3. Have researchers adequately controlled for gene–environment correla-tions in examining environmental risk factors for child antisocial be-havior?
Five specific environmental risk factors are discussed briefly in thischapter: having a young or teen mother, maternal cigarette smoking duringpregnancy, maternal sensitivity and attachment, coercive parenting practices,and child maltreatment. A composite risk of multiple adversity is also dis-cussed. These risk factors are chosen because they are believed to be the finalpathway through which other factors operate, are supported by developmen-tal theory, are potentially modifiable and therefore candidates for interventionstudies, or, if causal, open up a new field of investigation (e.g., prenatal expo-sure to tobacco smoke). In addition, they also illustrate the difficulties of dis-entangling gene from environment effects in studying the intergenerationaltransmission of early-onset antisocial disorder.
Young or Teenage Parenting
In two longitudinal studies of trajectories of physical aggression, teen mother-hood was a significant risk factor for trajectories of high aggression. This wasthe case for aggression trajectories between ages 6 and 15 years (Nagin &Tremblay, 2001) and 7 to 42 months of age (Tremblay et al., 2004). For boysages 6–15, having a teen mother also predicted which boys would be in achronic high aggression path versus a high but declining path.
Young maternal age at birth is associated with serious offspring antisocialbehavior (Nagin, Pogarsky, & Farrington, 1999; Wakschlag et al., 2000; Mar-tin & Burchinal, 1992; Jaffee, Caspi, Moffitt, Belsky, & Silva, 2001a; Hardyet al., 1997; Christ et al., 1990; Fergusson & Lynskey, 1993). This associationis strongest for maternal age at first birth rather than maternal age per se(Wakschlag et al., 2000; Jaffee et al., 2001a).
Maternal Cigarette Smoking during Pregnancy
Maternal smoking was a significant risk factor for stable high aggression tra-jectories in infancy (Tremblay et al., 2004). A number of research studies havesuggested that maternal prenatal tobacco smoking is a risk factor for conductdisorder or related constructs (criminality, delinquency, and violent criminal-
Intergenerational Transmission of Aggression 357

ity) (Olds, 1997; Fergusson, Woodward, & Horwood, 1998; Rasanen et al.,1999; Weissman, Warner Wickramaratne, & Kandel, 1999; Brennan, Grekin,& Mednick, 1999; Wakschlag, Pickett, Cook, Benowitz, & Leventhal, 2002;Wakschlag et al., 1997). A recent study has found an interaction between ma-ternal sensitivity and maternal smoking on the development of conduct disor-der in offspring (Wakschlag & Hans, 2002).
Maternal Responsiveness and Attachment
Maternal responsiveness to infant behaviors is broadly defined as action bythe mother that is contingent on a cue from the infant and is appropriate in itstype, timing, and intensity to the needs of the infant. Unlike coercive parentingbehaviors, which are dependent on response to oppositional behaviors seen intoddlers and older children, maternal responsiveness can be assessed nearbirth and in the context of infant behaviors that are unrelated to aggression oroppositional behavior. It is also believed that maternal responsiveness pro-vides the foundation for the development of behavioral regulation and socialcompetence in children (Wakschlag & Hans, 1999).
Maternal responsiveness during infancy (rated at 4, 12, and 24 months)has been shown in one study to predict offspring disruptive behavior (diag-noses of DSM-III-R conduct disorder or oppositional defiant disorder or asymptom count of criteria from these disorders) in middle childhood. Themothers were either opiate addicts or controls for low-income neighbor-hoods (Wakschlag & Hans, 1999). In a subsequent study on the same sam-ple, maternal responsiveness was found to interact with exposure to ciga-rette smoke prenatally in predicting offspring conduct problems (Wakschlag& Hans, 2002).
Although early research on the role of attachment in the development ofaggression has highlighted the importance of the avoidant category (Renken,Egeland, Marvinney, Mangelsdorf, & Sroufe, 1989), more recent work hasuncovered an association between attachment disorganization, specifically,and aggressive behaviors in children. In a sample of high-risk infants, clini-cally elevated levels of aggressive behavior at age 5 were found in 44% of dis-organized children but in only 5% of the secure group (Lyons-Ruth, Alpern,& Repacholi,1993). The association between attachment disorganization andaggression was observed in the same sample 2 years later (age 7). Anotherstudy found that the majority (60%) of children classified as disorganized alsoshowed clinically elevated levels of aggressive behavior, as compared with17% of their peers classified as secure (Shaw, Owens, Vondra, & Keenan,1996).
Although attachment disorganization seems to be a significant contribu-tor to the development of aggression in children, recent evidence indicates thatit is better considered in interaction with other risk factors. In one study, at-tachment disorganization predicted child aggression only in the context ofproblems in information processing (“mild mental lag”) (Lyons-Ruth, Easter-
358 D E T E R M I N A N T S O F A G G R E S S I O N

brooks, & Cibelli, 1997). Infants both classified as disorganized at age 1 andperceived by their mothers as having a difficult temperament at age 2 wereparticularly at risk for manifesting high levels of aggression at school age(Shaw et al., 1996).
Coercive Parenting Practices
A test of one of the most comprehensive models in the field of child aggression(Patterson, 1982; Reid & Patterson, 1989, Eddy, Leve, & Fagot, 2001) pro-vided evidence for a link between coercive maternal behavior and aggressivebehavior for both boys and girls. Maternal coercive behavior and lack ofaffectionate behavior predicted both high and increasing levels of aggressivebehavior, especially in boys (McFadyen-Ketchum, Bates, Dodge, & Pettit,1996). In research on aggression in toddlers, maternal negative dominance ininteraction with male gender and difficult (dysregulated) temperament was apredictor of aggression in the child (Rubin, Hastings, Chen, Stewart, &McNichol, 1998). Observed aggression and mother-reported externalizingproblems were associated significantly with dysregulated temperament onlyfor boys with mothers who demonstrated relatively high levels of negativedominance. In a study designed to clarify the influence of socioeconomic fac-tors (operationalized as mother’s and father’s years of education and occupa-tional status) in the development of conduct problems and aggressiveness inyoung children, several socialization indicators predicted externalizing and ag-gressive behavior (Dodge, Pettit, & Bates, 1994). Of this set, the strongest in-dividual contribution to the prediction of aggressive behavior in the childrencame from the level of exposure to harsh discipline as experienced by thechild. In a study of 46 parent-referred problem children, negative and directivematernal behaviors were predictive of maternal ratings of aggressive behaviorin the child (Campbell, Breaux, Ewing, & Szumoski, 1986). The authors con-cluded that negative and coercive mother–child interactions are significantcontributors to the long-term maintenance of early-onset aggressive behavior.Maternal coercive parenting at 5 months predicted a stable high trajectory ofinfant aggression from 17 to 42 months (Tremblay et al., 2004).
Child Maltreatment
Child maltreatment has been associated with a number of problematic devel-opmental outcomes, especially physical aggression and aggression-related be-haviors. A number of researchers have found that physical abuse, in particu-lar, is predictive of elevated levels of aggression in toddlers, preschoolers, andschool-age children (Dodge, Bates, & Pettit, 1990; Dodge, Pettit, Bates, &Valente, 1995; Downey & Walker, 1992; Erickson, Egeland, & Pianta, 1989;Malinosky-Rummel & Hansen, 1993). Moreover, physical abuse duringchildhood has been linked to aggression-related behaviors, such as delin-quency, antisocial behavior, and criminal activity, during adolescence and
Intergenerational Transmission of Aggression 359

adulthood (Herrenkohl, Egolf, & Herrenkohl, 1997; Muller & Diamond,1999; Widom, 2001).
A recent longitudinal birth cohort study suggests that child maltreatmentmay interact with a common gene on the X chromosome for monoamineoxidase A (MAOA) to produce CPA, at least in boys (Caspi et al., 2002). Inthat study, 36.8% of the males had the low-MAOA-activity form of the gene.By itself, the low form did not confer risk for antisocial behavior or for beingmaltreated, as compared with the high-MAOA-activity form. Maltreatmentalone, however, did confer some increased risk. The low-MAOA gene, in in-teraction with maltreatment, had a significant and meaningful increased riskfor antisocial behavior, defined four different ways: conduct disorder, convic-tion for violent offenses, a continuous score of disposition toward violence,and a continous score measure of antisocial personality disorder.
Multiple Adversity
Some studies suggest that it is the cumulation of risk factors that is importantin the development of early-onset antisocial disorder (Rutter, Tizard, Yule,Graham, & Whitmore, 1976; Shaw, Vondra, Hommerding, Keenan, & Dunn,1994; Fergusson, Horwood, & Lynskey, 1994). The evidence is strong thatthe more adversity, the greater the probability of developing CPA/CPASB. Theeffect of multiple risks is above and beyond that of the additive effects of eachrisk, suggesting that it is multiple risk itself that is possibly causal.
The Association between Parental Antisocial Behavior and Risk Factors
Do these risk factors occur more commonly in families in which parents ex-hibit antisocial behavior? The evidence for the association between the afore-mentioned risk factors and parental antisocial behavior comes from two typesof studies—studies of adult outcomes of children with CPASB and studies thathave assessed the relationship between antisocial disorder in parents and therisk factors.
The adult outcomes of children with early-onset antisocial disorder in-clude the very outcomes that are also considered risk factors for early-onsetantisocial disorder. Girls and boys with antisocial behavior are more likely tobecome teen parents. In a recent adult outcome study of a birth cohort of chil-dren in Dunedin, New Zealand, the authors stated: “Research into the conse-quences of teenage parenthood should begin to address the implications of thestrong link between antisocial behaviour and teenage parenthood. Most of theinfants born to teenage parents have at least one antisocial parent, and manyare born to two [antisocial] parents” (p. 197) (Moffitt et al., 2001). There arealso strong associations between conduct disorder in girls and smoking (Boyleet al., 1993; Hawkins, Catalano, & Miller, 1992; Kandel et al., 1997).
Multiple adversity in adult life appears to be particularly associated withchildhood histories of conduct problems. Among a sample of adults who in
360 D E T E R M I N A N T S O F A G G R E S S I O N

childhood were either in the care of child welfare authorities or from a poorneighborhood, multiple social adversity was found almost exclusively in thosewith a history of conduct disorder (Zoccolillo, Pickles, Quinton, & Rutter,1992). The current diagnostic construct of antisocial personality disorder isbased on the empirical observation that adults with pervasive dysfunctionacross several domains and who have antisocial behavior almost always havehistories of child conduct problems (Robins, 1966).
Turning to studies on parents, there is again good evidence for an associa-tion between antisocial behavior and the five risk factors examined here.Coercive parenting has been found to be associated with parental antisocialbehavior (Verlaan & Schwartzman, 2002; Patterson, DeGarmo, & Knutson,2000; Bosquet & Egeland, 2000; Johnson, Cohen, Kasen, Smailes, & Brook,2001). Adults who report child neglect have higher rates of antisocial person-ality disorder (Robins & Regier, 1991, Table 11-1, p. 261), and infants andtoddlers of women who were aggressive in childhood have higher rates of hos-pitalization for accidents (Serbin, Peters, & Schwartzman, 1996). Maternalsensitivity has been found in three separate studies to be associated withmaternal conduct disorder (Cassidy, Zoccolillo, & Hughes, 1996), a history ofmaternal aggression (Serbin, Peters, McAffer, & Schwartzman, 1991), ormaternal antisocial personality disorder (Hans, Bernstein, & Henson, 1999).Maternal smoking during pregnancy is associated with paternal antisocial per-sonality disorder and parental criminality (Fergusson et al., 1998; Wakschlaget al., 1997).
Parental Antisocial Behavior and Risk Factors for CPASB/CPA in Québec
Although many studies have found an association between parental antisocialbehavior and/or disorder and risk factors for CPASB/CPA, there has been nopopulation-based study of child development that has prospectively assessedboth maternal and paternal antisocial behavior and risk factors. This hasmade it difficult to estimate the strength of the association between risk vari-ables and parental antisocial behavior. To supplement these data, we presentfindings from the Longitudinal Study of Child Development in Québec(LSCDQ) (Jette, Desrosiers, Tremblay, & Thibault, 2000; Zoccolillo, 2000).The LSCDQ assessed maternal and paternal antisocial behavior in a sample of2,223 infants who were 5 months old. These infants were selected from birthcertificate records using a three-stage sampling design in order for them to berepresentative of all infants of the province of Québec, Canada.
Maternal and paternal conduct problems, specified as behaviors occur-ring before the end of high school, were assessed by self-report. Maternal be-haviors included stealing more than once (17.8%), starting more than onefight (3.3%), involvement with the police or youth protection because of mis-behavior (4%), skipping school more than twice in one year (47.6%), andrunning away from home overnight (9.6%). Paternal behaviors included steal-ing more than once (27.1%), starting more than one fight (10%), involvement
Intergenerational Transmission of Aggression 361

with the police or youth protection because of misbehavior (8.7%), andsuspension/expulsion from school (20%).
The associations between the number of maternal conduct problems anda variety of potential risk factors for adverse child development are shown inTable 17.1. Note that maternal conduct problems predicted whether the bio-logic father was missing from the home. Table 17.2 shows the association be-tween the number of paternal conduct problems for biological fathers living inthe home and potential risk factors.
Tables 17.1 and 17.2 illustrate very clearly, in this population-based sam-ple, the association between a measure of parental antisocial behavior (num-ber of conduct problems) and a variety of possible risk factors. The risk ismost pronounced at the level of 3–5 conduct problems, which is similar to theDSM-IV cutoff for a diagnosis of conduct disorder.
What proportion of infants with multiple risks come from homes withantisocial parents? An index of multiple risk was created by a sum score of
362 D E T E R M I N A N T S O F A G G R E S S I O N
TABLE 17.1. Risk Factors by the Number of Maternal Conduct Symptoms
Number of conduct symptoms
0(n = 955)
1(n = 759)
2(n = 275)
3–5(n = 129) p
Sociodemographic status
Had first child when age19 or younger
7.3 12.9 13.4* 24.7* < .001
No high school degree 13.5 18.5 18.8 32.2 < .001
Biological father nolonger in the home
7.1 8.6 10.2* 19.3 * < .05
Insufficient householdincome (Statistics Canadadefinition)
24.5 24.0 28.4 41.7 < .01
Substance use
Used illegal drugs inthe past 12 month
1.2** 3.7 4.2** 13.0 * < .001
Ever drunk (5 or moredrinks on one occasion)in past 12 months
9.2 16.2 23.5 31.8 < .001
Smoked during pregnancy 15.0 29.4 38.8 50.2 < .001
Used illegal drugs duringpregnancy
0.2** 2.1** 2.0** 5.9 ** < .001
Note. Data from Institut de la Statistique du Québec, ELDEQ 1998–2002.*Coefficient of variation between 15% and 25%; interpret with caution.**Coefficient of variation higher than 25%; imprecise estimate for descriptive purposes only.

four risk factors: (1) mother does not have high school diploma, (2) family inlowest income category, (3) mother smoked during pregnancy, (4) mother hadfirst child before age 21. A dichotomous measure of parent antisocial disorderwas created. Because one conduct problem was relatively common in themothers and studies have suggested that it is the presence of multiple conductproblems that is particularly associated with pervasive and persistent antiso-cial disorder (Zoccolillo et al., 1992; Robins, 1966), infants were dichoto-mized into two groups where data on antisocial disorder was available forboth parents—infants who had mothers and fathers who had no or one con-duct symptom versus all other infants. Studies have suggested that biologicalfathers not living in the home are more antisocial than nonmissing fathers(Caspi et al., 2001; Jaffee, Caspi, Moffitt, Taylor, & Dickson, 2001b), and inour sample infants with the most antisocial mothers were also the most likelyto have biological fathers not living in the home. Therefore, infants withbiologic fathers not living in the home were also placed in the “all others”group. Finally, some mothers and fathers living in the home did not return theself-administered questionnaires that contained the questions on antisocial be-havior, and a small subset of infants not participating in the longitudinal follow-up (103) were removed from the analyses, leaving a total of 1986 infants.
Intergenerational Transmission of Aggression 363
TABLE 17.2. Risk Factors by the Number of Paternal Conduct Symptomsin Two-Parent Families
Number of conduct symptoms
0(n = 1,067)
1(n = 457)
2(n = 208)
3–5(n = 102) p
No high school degree 13.2 17.6 24.6 25.6* < .001
College degree 43.2 35.4 26.9 12.5** < .001
Unemployed 11.6 11.4 18.0* 20.3* < .05
Currently daily smoker 21.5 33.8 42.8 55.3 < .001
Used illegal drugs in thepast 12 month
4.6* 7.6* 11.0* 20.6* < .001
Mean number of timesdrunk (5 or more drinkson one occasion)
4.2 7.4 7.0 13.1 < .001
Maternal behaviors of partner
Mother had 2 or moreconduct symptoms
12.9(n = 1,048)
20.0(n = 451)
30.8(n = 204)
30.7*(n = 101)
< .001
Mother smoked duringpregnancy
19.4(n = 1,062)
26.1(n = 456)
31.3(n = 208)
42.7(n = 103)
< .001
Note. Data from Institut de la Statistique du Québec, ELDEQ 1998–2002.*Coefficient of variation between 15% and 25%; interpret with caution.**Coefficient of variation higher than 25%; imprecise estimate for descriptive purposes only.

The association between the number of risks was cross-classified by thedichotomous parental antisocial disorder/father not in the home measure(Table 17.3). There was a strong and significant association between multiplerisk and parental antisocial behavior/father not in the home. Most infantswithout risk do not have antisocial parents, and most infants with multiplerisk have at least one parent with at least two conduct problems or have fa-thers not living in the home. The association between multiple risk andCPASB/CPA is confounded by parental antisocial behavior.
Controlling for Parental Antisocial Behavior When ExaminingRisk Factors for CPASB/CPA
To summarize the findings: (1) Parental antisocial behavior and offspringearly antisocial behavior are associated; (2) parenting practices, maternal sen-sitivity, child maltreatment, young motherhood, maternal smoking duringpregnancy, and multiple adversity are all predictors of CPASB and or CPA; (3)these same risk factors are also associated with parental antisocial behavior;(4) genetically informative designs have consistently found evidence for bothshared environment and genetic effects for antisocial behavior and aggression.Given these strong relationships between environmental risk and parental an-tisocial behavior, have previous studies adequately controlled for gene–envi-ronment correlations in examining environmental risk factors for child antiso-cial behavior?
An important question is how can the confound of genetic association be
364 D E T E R M I N A N T S O F A G G R E S S I O N
TABLE 17.3. Proportion of Infants with NonantisocialParents by Level of Adversity
Adversity indexa
Mother and father have noor one conduct problemand father is not missingb
None (n = 1,117) 75.1%
One (n = 453) 56.5%
Two (n = 179) 50.3%
Three (n = 107) 36.6%
Four (n = 40) 21.4%
Note. n = 1,896; χ2 = 121.34, 4 df, p < .000 (corrected for design effectof 1.3).aSum of 4 risks: Mother does not have high school degree; in lowest in-come category; mother smoked during pregnancy; mother had firstchild before age 21.bInfants classified into two parent groups: (1) Mother and father haveno or one conduct problem and father is not missing; (2) all other com-binations of maternal and paternal conduct problems and those with fa-thers not living in the home. Homes where mother or father was presentbut did not return valid questionnaire on antisocial behavior were ex-cluded.

controlled for when testing a specific risk factor? A critical question iswhether genetically informative designs (twin and adoption studies) are neces-sary to control for parental antisocial disorder when testing a specific risk fac-tor. The adoption design is not feasible to examine the critical question of whychildren exposed to maltreatment and multiple adversities are more likely togrow up to be antisocial, because adopted children are generally not placed inadoptive homes where maltreatment and severe adversity will occur (Rutter,1997). Another, related, problem with adoption studies is that the maternalprenatal environment is often not well assessed. Given the association be-tween maternal and paternal antisocial disorder, maternal prenatal smoking,and offspring antisocial behavior, this could lead to an overestimate of geneticeffects.
Twin studies are limited because the families most of interest are those oftwins with one or more antisocial parents and/or twins with severe adversity,maltreatment, or other risk factors. Furthermore, the ideal studies are pro-spective studies and studies based on observational data collected before thetwins show aggressive or antisocial behavior. This requires recruiting from thepool of twin births. Because approximately 1 in 100 deliveries is a twin birth,recruiting 100 twin pairs requires screening 10,000 births (Vogel & Motulsky,1996). However, if twins with an antisocial parent are required (assuming10% of twins have an antisocial mother or father), then 100,000 births willhave to be screened and 1,000 twin pairs recruited. Even this is limiting, as theend result will be approximately 30 monozygotic and 70 dizygotic twin pairswith an antisocial parent.
A recently developed twin study of high-risk infants is the EnvironmentalRisk (E-Risk) Longitudinal Twin Study (Jaffee, Moffitt, Caspi, & Taylor,2003). This sample of twins was based on the Twins Early DevelopmentStudy, a register of all twins born in England and Wales in 1994–1995. Duringthis time 15,906 twin pairs were born, of whom 71% joined the register. Fromthis pool, a subset of 1,116 twin pairs was drawn, of whom one-third werethose of mothers whose first birth was less than 20 years of age and the restwere representative of all twins. Of note is that this twin sample was specifi-cally designed to look at environmental risk and includes assessments of anti-social behavior in the parents. Few researchers, however, are able to accesssuch large twin samples.
Can the twin design be useful in identifying specific environmental ef-fects? First, as noted earlier, it is important to study twins with antisocialparents so as to have the twins exposed to the type of risk seen in infants ofantisocial parents. Apart from the large number of twins needed to find twinsmeeting this criterion, the twin design itself poses a particular problem forstudying children of antisocial parents. Mothers of dizygotic twins are consid-erably older than mothers of monozygotic twins (Vogel & Motulsky, 1996).Furthermore, mothers who postpone childbearing to later in life, because ofeducation and/or career, and who then require fertility treatment are likelyhave dizygotic twins as a result. Because antisocial mothers have children at a
Intergenerational Transmission of Aggression 365

younger age, this raises the possibility that identical twins will be more likelyto have antisocial parents than dizygotic twins. Differences in environment be-tween monozygotic and dizygotic twins must be properly examined and ac-counted for, as the key assumption of twin studies is that the environments ofmonozygotic and dizygotic twins are similar. Nonetheless, a twin high-riskdesign is a powerful tool for examining environmental effects. In the E-Riskstudy noted earlier, combining behavior genetic and epidemiologic analysisshowed that “children who resided with antisocial fathers received a ‘doublewhammy’ of genetic and environmental risk for conduct problems” (Jaffee etal., 2003, p. 109).
Because adoptive designs are not feasible, and twin designs suitable foraddressing questions such as maltreatment or multiple risk are methodologi-cally intensive and require large populations of twin births, is there another al-ternative? The answer is a qualified yes. The solution is to control for antiso-cial behavior in both biological parents when measuring the effects of thesuspected risk factor.
It is important to note that this method can be used to control for genetictransmission but cannot be used to address the question of proportion of vari-ance of the outcome explained by genes or environment. It is also importantto note that the “genotype” measured is very imperfectly measured throughparental phenotype. If the wrong phenotype is measured, then it will not be anadequate control. An example of how this can occur is provided by the adop-tion and family studies examining the association between unexplained so-matic complaints in women and antisocial behavior or disorders in their malerelatives. The evidence is strong that multiple somatic complaints in womenare genetically related to antisocial behavior in men, seen most dramatically inthe excess of somatic complaints in the adopted daughters of violent alcoholicfathers (Bohman, Cloninger, von Knorring, & Sigvardsson, 1984). Con-trolling for overtly antisocial behaviors in the mother may still not adequatelycontrol for genotype.
Before turning to the literature to see whether studies of suspected riskfactors for offspring early-onset antisocial disorder have controlled for paren-tal antisocial disorder, some issues of measurement need to be addressed: (1)classification of parents for use as a control variable, (2) assessing both par-ents, and (3) modeling both parents. These issues are discussed in turn in thefollowing paragraphs.
Classifying parents for use as a control variable is problematic becausethere is no clear boundary between “antisocial” and “not antisocial.” For ex-ample, even one child conduct-disorder symptom in childhood is associatedwith an increased risk for antisocial behavior in adult life (Robin, 1966). Be-cause the reason for controlling for parental antisocial behavior is to removeany covariance between parental antisocial behavior and child antisocial be-havior in order to test the independent effects of other variables, the measureof parental antisocial behavior should be broad enough to capture the fullcontinuum of parental antisocial behavior.
366 D E T E R M I N A N T S O F A G G R E S S I O N

Two common measures used as a control for parental antisocial behaviorare a diagnosis of antisocial personality disorder and criminality (e.g., arrests,convictions). Both are problematic because they are dichotomous measures,and many parents without a diagnosis or without criminality are still likely tohave antisocial behavior. With regard to DSM-IV antisocial personality disor-der (APD), three child symptoms and three adult symptoms are required tomeet the criteria (American Psychiatric Association, 1994). This means thatthose with three child symptoms and two adult symptoms, or two child symp-toms and three adult symptoms, would be classified as “No disorder.” In pre-vious work, it was shown that DSM-III (American Psychiatric Association,1980) criteria for APD identified only about half of young adults with childhood-onset persistent and pervasive antisocial behavior (Zoccolillo et al., 1992). Itis highly likely that many parents with some antisocial symptoms, but notenough to meet DSM criteria for APD, will have childhood-onset persistentand pervasive antisocial behavior. Classifying these parents as not antisocialwhen investigating risk factors for offspring CPASB/CPA does not adequatelycontrol for the possibility of confounding by parental antisocial behavior. Sim-ilarly, criminality is also a poor control because, in population samples ofadults with DSM-III diagnoses of APD, almost half the men and most womenwith APD did not have major criminal convictions (Robins & Regier, 1991).
Both biological parents need to be assessed for antisocial disorder. Theheritability for antisocial behavior was not found to differ for men andwomen (Rhee & Waldman, 2002), and the effects of parental APD on off-spring externalizing disorder were also the same (Connell & Goodman,2002). Assessing both parents is particularly important because environmen-tal risk factors may be associated with antisocial disorder in one or the otherparent. For example, antisocial men may be more prone to mate with teenagefemales, independent of whether the female also had an antisocial disorder.Although there is good evidence for assortative mating, it is not complete(Krueger, Moffitt, Caspi, Bleske, & Silva, 1998). This means that there will bea large proportion of families in which only one parent has an antisocial disor-der.
Another major concern is missing biological fathers. Missing biologicalfathers are likely to be more antisocial than fathers who are present in thehome (Jaffee et al., 2001b). Asking the biological mother about the missing bi-ological father is essential and appears to give reliable information (Caspi etal., 2001). Alternatively, missing biological fathers can be incorporated in thetheoretical model being tested as a separate category.
Both parents also need to be included in the analyses with the child as theunit of analysis. For example, in the LSCDQ there was a strong associationbetween maternal smoking during pregnancy and the number of paternal con-duct symptoms (Table 17.2). Controlling only for maternal antisocial behav-ior when evaluating the effect of maternal prenatal smoking would notadquately control for the child’s genetic background.
We have been unable to find any studies of the environmental risk factors
Intergenerational Transmission of Aggression 367

that meet all the aforementioned criteria, except for one recent study of infantaggression (Tremblay et al., 2004) and the Environmental Risk (E-Risk) Lon-gitudinal Twin Study (Jaffee et al., 2003). Although some investigators haveattempted to measure antisocial behavior in the parent, most used a diagnosticcategory of parental antisocial personality disorder versus no disorder (John-son et al., 2001; Wakschlag et al., 1997), relied on parental criminality (some-times specified only as familial criminality) (Moffitt et al., 2001; Patterson etal., 2000; Fergusson et al., 1998; Rutter et al., 1976), or used personality dis-order scales from the Minnesota Multiphasic Personality Inventory (Pattersonet al., 2000; Wakschlag et al., 1997).
To summarize, none of the studies have demonstrated good control of pa-rental antisocial disorder. It would be difficult to conclude that maternalsmoking during pregnancy, teenage motherhood, maternal sensitivity, parent-ing practices, maltreatment, or multiple adversity is causal for the develop-ment of early-onset antisocial behavior, as these factors are confounded byparental antisocial disorder.
DIRECTIONS FOR FUTURE RESEARCH
It is critical for studies that are attempting to identify the causes of antisocialbehavior to take into account the familial transmission of antisocial disorderand that antisocial disorder in parents is strongly associated with a multiplic-ity of environmental risk factors for their offspring. At the population level,the association between parental antisocial disorder and adversity, particularlymultiple adversity, is large. Researchers cannot ignore that fact that a signifi-cant proportion of children living in adversity he found to have an antisocialparent. There are several implications of these findings.
First, it is critical to study families with antisocial parents. Although thedifficulties of studying these families and the complexity of multiple risk fac-tors may appear to be daunting, these families are not rare and account for alarge proportion of antisocial youth, particularly the most severely affectedchildren.
Second, children with antisocial parents are exposed to multiple risk fac-tors that start very early. Teasing out which risk factors are causal for antiso-cial disorder in the child is the challenge. Tests of hypotheses need to be wellstructured to control for confounds and to test for interactions. Of critical im-portance are studies of the developmental sequence of risk factors and behav-iors. Priority should be given to studies that start prenatally or at birth.
To test whether poor parenting practices are a causal risk factor for earlyantisocial disorder requires demonstrating that these parenting practices pre-cede the emergence of early-onset antisocial disorder. This is not an easy task,inasmuch as coercive parenting practices may emerge only when the child de-velops normal opposition behaviors in the toddler years, making it difficult todetermine the time order of effects. Controlling for parental antisocial disor-
368 D E T E R M I N A N T S O F A G G R E S S I O N

der is critical in such design because the same genetic mechanism could poten-tially lead to an antisocial parent, poor parenting practices, and offspring anti-social disorder (Ge et al., 1996; O’Connor, Deater-Deckard, Fulker, Rutter, &Plomin, 1998).
Third, as much attention needs to be paid to defining “antisocial behav-ior” in the parent as in the child. Future studies should avoid using criminalityas the sole measure of parental antisocial disorder, as it is neither sensitive norspecific enough to be useful. The LSCDQ study has shown that parents in apopulation sample will answer questions on their own child and adult antiso-cial behaviors. An advantage of assessing a broad range of child and adult an-tisocial behaviors in parents is that the researcher can then use latent variablemethods to cluster parents into homogenous groups or to create a latentcontinous variable. It also becomes possible to test whether there are distinctfamilial types of antisocial behavior—for example, early-onset versus adolescent-onset. Ideally, the same dimensions of aggression and antisocial behavior as-sessed in the child should be assessed in the adult. Little is known aboutwhether parental aggression is specifically associated with child aggression,for example.
Fourth, both biological parents must be assessed. Controlling for oneparent only is not adequate, as assortative mating is incomplete and antisocialdisorder is more common in males than in females. Missing biological fathersneed to be accounted for, either by proxy interview of other informants or byhaving a separate category of “missing father.” Analytic strategies for incor-porating data by proxy interview on missing fathers with data from presentfathers need to be refined.
The analysis of data that incorporates measurements of antisocial be-havior in both parents is more complex than treating maternal and paternalantisocial disorder as separate variables in a logistic or linear regressionmodel. For example, fathers are more likely to be missing if the mother isantisocial and are therefore not missing at random which regard to manyanalyses of interest. This poses considerable problems for most statisticalanalyses in that maximum likelihood estimates for parameters often cannotbe estimated (Little & Rubin, 1987). Analyses of interactions between pa-rental antisocial disorder and other variables (as proxy measures for gene–environment interactions) are also complex. For example, a parent contrib-utes only half of his or her genes to offspring, and path models of parent ef-fects on offspring would need to take this into account (Vogel & Motulsky,1996).
Fifth, randomized controlled trials of interventions to prevent CPA/CPASB should assess parental antisocial behavior. Little is known aboutwhether promising prevention interventions are also effective in families withantisocial parents. Furthermore, if modifying a risk factor is effective in pre-venting CPA/CPASB even among children with parents with antisocial behav-ior, then this strongly suggests a causal role for that factor.
In a recent review of studies of conduct disorder, the authors noted,
Intergenerational Transmission of Aggression 369

“Parsing out the contributions of genetic effects from environmental risk fac-tors to familial antisocial behavior is a significant task that remains largely in-complete” (Burke, Loeber, & Birmaher, 2002, p. 1277). A major challenge ofthe next 10 years is to fill in the holes in our knowledge of genetic and envi-ronmental contributions to the development of early-onset antisocial behaviorand aggression through new and better research designs.
REFERENCES
American Psychiatric Association. (1980). Diagnostic and statistical manual of mentaldisorders (3rd ed.). Washington, DC: Author.
American Psychiatric Association. (1994). Diagnostic and statistical manual of mentaldisorders (4th ed.). Washington, DC: Author.
Bohman, M., Cloninger, C. R., von Knorring, A. L., & Sigvardsson, S. (1984). An adop-tion study of somatoform disorders. III. Cross-fostering analysis and genetic rela-tionship to alcoholism and criminality. Archives of General Psychiatry, 41, 872–878.
Bosquet, M., & Egeland, B. (2000). Predicting parenting behaviors from Antisocial Prac-tices Content Scale scores of the MMPI-2 administered during pregnancy. Journalof Personality Assessment, 74, 146–162.
Boyle, M. H., Offord, D. R., Racine, Y. A., Fleming, J. E., Szatmari, P., & Links, P. S.(1993). Predicting substance use in early adolescence based on parent and teacherassessments of childhood psychiatric disorder: Results from the Ontario ChildHealth Study follow-up. Journal of Child Psychology and Psychiatry and AlliedDisciplines, 34, 535–544.
Brennan, P. A., Grekin, E. R., & Mednick, S. A. (1999). Maternal smoking during preg-nancy and adult male criminal outcomes [see comments]. Archives of General Psy-chiatry, 56, 215–219.
Broidy, L. M., Nagin, D. S., Tremblay, R. E., Bates, J. E., Brame, B., Dodge, K. A.,Fergusson, D., Horwoood, J. L., Loeber, R., Laird, R., Lynam, D. R., Moffitt, T. E.,Petit, G. S., & Vitaro, F. (2003). Developmental trajectories of childhood disruptivebehaviors and adolescent delinquency: A six-site, cross-national study. Develop-mental Psychology, 39, 222–245.
Burke, J. D., Loeber, R., & Birmaher, B. (2002). Oppositional defiant disorder and con-duct disorder: A review of the past 10 years. Part II. Journal of the American Acad-emy of Child and Adolescent Psychiatry, 41, 1294–1305.
Campbell, S. B., Breaux, A. M., Ewing, L. J., & Szumowski, E. K. (1986). Correlates andpredictors of hyperactivity and aggression: A longitudinal study of parent-referredproblem preschoolers. Journal of Abnormal Child Psychology, 14, 217–234.
Caspi, A., McClay, J., Moffitt, T. E., Mill, J., Martin, J., Craig, I. W., Taylor, A., &Poulton, R. (2002). Role of genotype in the cycle of violence in maltreated children.Science, 297, 851–854.
Caspi, A., Moffitt, T. E., Newman, D. L., & Silva, P. A. (1996). Behavioral observationsat age 3 years predict adult psychiatric disorders: Longitudinal evidence from abirth cohort. Archives of General Psychiatry, 53, 1033–1039.
Caspi, A., Taylor, A., Smart, M., Jackson, J., Tagami, S., & Moffitt, T. E. (2001). Canwomen provide reliable information about their children’s fathers? Cross-infor-
370 D E T E R M I N A N T S O F A G G R E S S I O N

mant agreement about men’s lifetime antisocial behaviour. Journal of Child Psy-chology and Psychiatry and Allied Disciplines, 42, 915–920.
Cassidy, B., Zoccolillo, M., & Hughes, S. (1996). Psychopathology in adolescent moth-ers and its effects on mother–infant interactions: A pilot study. Canadian Journal ofPsychiatry [Revue Canadienne de Psychiatrie], 41, 379–384.
Christ, M. A., Lahey, B. B., Frick, P. J., Russo, M. F., McBurnett, K., Loeber, R.,Stouthamer-Loeber, M., & Green, S. (1990). Serious conduct problems in the chil-dren of adolescent mothers: Disentangling confounded correlations. Journal ofConsulting and Clinical Psychology, 58, 840–844.
Connell, A. M., & Goodman, S. H. (2002). The association between psychopathology infathers versus mothers and children’s internalizing and externalizing behavior prob-lems: A meta-analysis. Psychological Bulletin, 128, 746–773.
Dodge, K. A., Bates, J. E., & Pettit, G. S. (1990). Mechanisms in the cycle of violence. Sci-ence, 250, 1678–1683.
Dodge, K. A., Pettit, G. S., & Bates, J. E. (1994). Socialization mediators of the relationbetween socioeconomic status and child conduct problems. Child Development,65, 649–665.
Dodge, K. A., Pettit, G. S., Bates, J. E., & Valente, E. (1995). Social information-process-ing patterns partially mediate the effect of early physical abuse on later conductproblems. Journal of Abnormal Psychology, 104, 632–643.
Downey, G., & Walker, E. (1992). Distinguishing family-level and child-level influenceson the development of depression and aggression in children at risk. Developmentand Psychopathology, 4, 81–95.
Eddy, J. M., Leve, L. D., & Fagot, B. I. (2001). Coercive family processes: A replicationand extension of Patterson’s Coercion Model. Aggressive Behavior, 27, 14–25.
Eley, T. C., Lichtenstein, P., & Moffitt, T. E. (2003). A longitudinal behavioral geneticanalysis of the etiology of aggressive and nonaggressive antisocial behavior. Devel-opment and Psychopathology, 15, 383–402.
Erickson, M. F., Egeland, B., & Pianta, R. (1989). The effects of maltreatment on the de-velopment of young children. In D. Cicchetti & V. Carlson (Eds.), Child maltreat-ment: Theory and research on the causes and consequences of child abuse and ne-glect (pp. 647–684). New York: Cambridge University Press.
Fergusson, D. M., Horwood, L. J., & Lynskey, M. (1994). The childhoods of multipleproblem adolescents: A 15-year longitudinal study. Journal of Child Psychologyand Psychiatry and Allied Disciplines, 35, 1123–1140.
Fergusson, D. M., & Lynskey, M. T. (1993). Maternal age and cognitive and behaviouraloutcomes in middle childhood. Paediatric and Perinatal Epidemiology, 7, 77–91.
Fergusson, D. M., Woodward, L. J., & Horwood, L. J. (1998). Maternal smoking duringpregnancy and psychiatric adjustment in late adolescence. Archives of General Psy-chiatry, 55, 721–727.
Ge, X., Conger, R., Cadoret, R., Neiderhiser, J., Yates, W., Troughton, E., & Stewart, M.A. (1996). The developmental interface between nature and nurture: A mutual in-fluence model of child antisocial behavior and parent behaviors. DevelopmentalPsychology, 32, 574–589.
Hans, S. L., Bernstein V. J., & Henson L. G. (1999). The role of psychopathology in theparenting of drug-dependent women. Development and Psychopathology, 11, 957–977.
Hardy, J. B., Shapiro, S., Astone, N. M., Miller, T. L., Brooks-Gunn, J., & Hilton, S. C.(1997). Adolescent childbearing revisited: The age of inner-city mothers at delivery
Intergenerational Transmission of Aggression 371

is a determinant of their children’s self-sufficiency at age 27 to 33. Pediatrics, 100,802–809.
Hawkins, J. D., Catalano, R. F., & Miller, J. Y. (1992). Risk and protective factors for al-cohol and other drug problems in adolescence and early adulthood: Implications forsubstance abuse prevention. Psychological Bulletin, 112, 64–105.
Herrenkohl, R. C., Egolf, B. P., & Herrenkohl, E. C. (1997). Preschool antecedents of ad-olescent assaultive behaviour: A longitudinal study. American Journal of Ortho-psychiatry, 67, 422–432.
Hinshaw, S. P. (2002). Process, mechanism, and explanation related to externalizing be-havior in developmental psychopathology. Journal of Abnormal Child Psychology,30, 431–446.
Huesmann, L. R., Eron, L. D., Lefkowitz, M. M., & Walder, L. O. (1984). Stability of ag-gression over time and generations. Developmental Psychology, 20, 1120–1134.
Jaffee, S., Caspi, A., Moffitt, T. E., Belsky, J., & Silva, P. (2001a). Why are children bornto teen mothers at risk for adverse outcomes in young adulthood? Results from a20–year longitudinal study. Development and Psychopathology, 13, 377–397.
Jaffee, S. R., Caspi, A., Moffitt, T. E., Taylor, A., & Dickson, N. (2001b). Predicting earlyfatherhood and whether young fathers live with their children: Prospective findingsand policy reconsiderations. Journal of Child Psychology and Psychiatry and AlliedDisciplines, 42, 803–815.
Jaffee, S. R., Moffitt, T. E.,Caspi, A., & Taylor, A. (2003). Life with or without father:The benefits of living with two biological parents depend on the father’s antisocialbehavior. Child Development, 74, 109–126.
Jette, M., Desrosiers, H., Tremblay, R. E., & Thibault, J. (2000). Longitudinal study ofchild development in Quebec (ELDEQ 1998–2002) (Vol. 1). Québec: Institut de laStatistique du Québec.
Johnson, J. G., Cohen, P., Kasen, S., Smailes, E., & Brook, J. S. (2001). Association ofmaladaptive parental behavior with psychiatric disorder among parents and theiroffspring. Archives of General Psychiatry, 58, 453–460.
Kandel, D., Johnson, J., Bird, H., Canino, G., Goodman, S. H., Lahey, B. B., et al. (1997).Psychiatric disorders associated with substance use among children and adoles-cents: Findings from the Methods for the Epidemiology of Child and AdolescentMental Disorders (MECA) Study. Journal of Abnormal Child Psychology, 25, 121–132.
Krueger, R. F., Moffitt, T. E., Caspi, A., Bleske, A., & Silva, P. A. (1998). Assortative mat-ing for antisocial behavior: Developmental and methodological implications. Be-havior Genetics, 28, 173–186.
Lahey, B. B., Loeber, R., Quay, H. C., Applegate, B., Shaffer, D., Waldman, I., Hart, E. L.,McBurnett, K., Frick, P. J., Jensen, P. S., Dulcan, M. K., Canino, G., & Bird, H. R.(1998). Validity of DSM-IV subtypes of conduct disorder based on age of onset. Jour-nal of the American Academy of Child and Adolescent Psychiatry, 37, 435–442.
Little R. A., & Rubin, D. R. (1987). Statistical analysis with missing data. New York:Wiley.
Loeber, R., Burke, J. D., Lahey, B. B., Winters, A., & Zera, M. (2000). Oppositional defi-ant and conduct disorder: A review of the past 10 years. Part I. Journal of the Ameri-can Academy of Child and Adolescent Psychiatry, 39, 1468–1484.
Loeber, R., & Farrington, D. P. (2000). Young children who commit crime: Epidemiol-ogy, developmental origins, risk factors, early interventions, and policy implica-tions. Development and Psychopathology, 12, 737–762.
372 D E T E R M I N A N T S O F A G G R E S S I O N

Lyons-Ruth, K., Alpern, L., & Repacholi, B. (1993). Disorganized infant attachmentclassification and maternal psychosocial problems as predictors of hostile-aggres-sive behavior in the preschool classroom. Child Development, 64, 572–585.
Lyons-Ruth, K., Easterbrooks, A., & Cibelli, C. (1997). Infant attachment strategies, in-fant mental lag, and maternal depressive symptoms : Predictors of internalizing andexternalizing problems at age 7. Developmental Psychology, 33, 681–692.
Malinosky-Rummel, R., & Hansen, D. J. (1993). Long-term consequences of childhoodphysical abuse. Psychological Bulletin, 114, 68–79.
Martin, S. L., & Burchinal, M. R. (1992). Young women’s antisocial behavior and thelater emotional and behavioral health of their children. American Journal of PublicHealth, 82, 1007–1010.
McFadyen-Ketchum, S. A., Bates, J. E., Dodge, K. A., & Pettit, G. S. (1996). Patterns ofchange in early childhood aggressive-disruptive behavior: Gender differences inpredictions from early coercive and affectionate mother–child interactions. ChildDevelopment, 67, 2417–2433.
Moffitt, T. E., Caspi, A., Rutter, M., & Silva, P. (2001). Sex differences in antisocial be-haviour. Cambridge, UK: Cambridge University Press.
Muller, R. T., & Diamond, T. (1999). Father and mother physical abuse and child aggres-sive behaviour in two generations. Canadian Journal of Behavioural Science, 31,221–228.
Nagin, D. S., Pogarsky, G., & Farrington, D. P. (1999). Adolescent mothers and the crim-inal behavior of their children. Law and Society Review, 31, 137–162.
Nagin, D. S., & Tremblay, R. E. (2001). Prenatal and early childhood predictors of persis-tent physical aggression in boys from kindergarten to high school. Archives of Gen-eral Psychiatry, 58, 389–394.
O’Connor, T. G., Deater-Deckard, K., Fulker, D., Rutter, M., & Plomin, R. (1998). Geno-type–environment correlations in late childhood and early adolescence: Antisocialbehavioral problems and coercive parenting. Developmental Psychology, 34, 970–981.
Olds, D. (1997). Tobacco exposure and impaired development: A review of the evidence.Mental Retardation and Developmental Disabilities Review, 3, 257–269.
Patterson, D. R., DeGarmo, D. S., & Knutson, N. (2000). Hyperactive and antisocial be-haviors: Comorbid or two points in the same process? Development and Psycho-pathology, 12, 91–106.
Patterson, G. R. (1982). Coercive family processes. Eugene, OR: Castalia.Plomin, R. (1997). Behavioral genetics (3rd ed.). New York: Freeman.Rasanen, P., Hakko, H., Isohanni, M., Hodgins, S., Jarvelin, M. R., & Tiihonen, J.
(1999). Maternal smoking during pregnancy and risk of criminal behavior amongadult male offspring in the Northern Finland 1966 Birth Cohort. American Journalof Psychiatry, 156, 857–862.
Reid, J., & Patterson, G. (1989). The development of antisocial behaviour patterns inchildhood and adolescence. European Journal of Personality, 3, 107–119.
Renken, B., Egeland, B., Marvinney, D., Mangelsdorf, S., & Sroufe, A. (1989). Earlychildhood antecedents of aggression and passive-withdrawal in early elementaryschool. Journal of Personality, 57, 257–281.
Rhee, S. H., & Waldman, I. (2002). Genetic and environmental influences on antisocialbehavior: A meta-analysis of twin and adoption studies. Psychological Bulletin,128, 490–529.
Robins, L. N. (1966). Deviant children grown up. Baltimore: Williams & Wilkins.
Intergenerational Transmission of Aggression 373

Robins, L. N. (1996). Deviant children grown up. European Child and Adolescent Psy-chiatry, 5(Suppl. 6).
Robins, L. N., & Regier, D. A. (1991). Psychiatric disorders in America: The epidemio-logic catchment area study. New York: Free Press.
Rubin, K. H., Hastings, P., Chen, X., Stewart, S., & McNichol, K. (1998). Intrapersonaland maternal correlates of aggression, conflict, and externalizing problems in tod-dlers. Child Development, 69, 1614–1629.
Rutter, M. (1997). Nature–nurture integration. The example of antisocial behavior.American Psychologist, 52, 390–398.
Rutter, M., Tizard, J., Yule, W., Graham, P., & Whitmore, K. (1976). Research report:Isle of Wight studies. Psychological Medicine, 126, 493–509.
Serbin, L., Peters, P., McAffer, V., & Schwartzman, A. (1991). Childhood aggression andwithdrawal as predictors of adolescent pregnancy, early parenthood, and environ-mental risk for the next generation. Canadian Journal of Behavioural Science, 23,318–331.
Serbin, L. A., Peters, P. L., & Schwartzman, A. E. (1996). Longitudinal study of earlychildhood injuries and acute illnesses in the offspring of adolescent mothers whowere aggressive, withdrawn, or aggressive-withdrawn in childhood. Journal of Ab-normal Psychology, 105, 500–507.
Shaw, D. S., Owens, E. B., Vondra, J. I., & Keenan, K. (1996). Early risk factors and path-ways in the development of early disruptive behavior problems. Development andPsychopathology, 8, 679–699.
Shaw, D. S., Vondra, J. I., Hommerding, K. D., Keenan, K., & Dunn, M. (1994). Chronicfamily adversity and early child behavior problems: A longitudinal study of low in-come families. Journal of Child Psychology and Psychiatry and Allied Disciplines,35, 1109–1122.
Tremblay, R. E. (2000). The development of aggressive behaviour during childhood:What have we learned in the past century? International Journal of Behavioral De-velopment, 24, 129–141.
Tremblay, R. E., Nagin, D. S., Seguin, J. R., Zoccolillo, M., Zelazo, P. D., Boivin, M.,Pérusse, D., & Japel, C. (2004). Predictors of high level physical aggression from 17to 42 months after birth. Pediatrics, 114, e43–e50.
Verlaan, P., & Schwartzman, A. E. (2002). Mother’s and father’s parental adjustment:Links to externalizing behaviour problems in sons and daughters. InternationalJournal of Behavioral Development, 26, 214–224.
Vogel, F., & Motulsky, A. (1996). Human genetics: Problems and approaches (3rd ed.).Berlin: Springer.
Wakschlag, L. S., Gordon, R., Lahey, B. B., Loeber, R., Green, S., & Leventhal, B. L.(2000). Maternal age at first birth and boys’ risk for conduct disorder. Journal ofResearch on Adolescence, 10, 417–441.
Wakschlag, L. S., & Hans, S. L. (1999). Relation of maternal responsiveness during in-fancy to the development of behavior problems in high-risk youths. DevelopmentalPsychology, 35, 569–579.
Wakschlag, L. S., & Hans, S. L. (2002). Maternal smoking during pregnancy and con-duct problems in high-risk youth: A developmental framework. Development andPsychopathology, 14, 351–369.
Wakschlag, L. S., Lahey, B. B., Loeber, R., Green, S. M., Gordon, R. A., & Leventhal, B.L. (1997). Maternal smoking during pregnancy and the risk of conduct disorder inboys. Archives of General Psychiatry, 54, 670–676.
374 D E T E R M I N A N T S O F A G G R E S S I O N

Wakschlag, L. S., Pickett, K. E., Cook, E., Jr., Benowitz, N. L., & Leventhal, B. L. (2002).Maternal smoking during pregnancy and severe antisocial behavior in offspring: Areview. American Journal of Public Health, 92, 966–974.
Weissman, M., Warner V., Wickramaratne, P., & Kandel, D. B. (1999). Maternal smok-ing during pregnancy and psychopathology in offspring followed to adulthood.Journal of the American Academy of Child and Adolescent Psychiatry, 38, 892–899.
Widom, C. S. (2001). Child abuse and neglect. In S. O. White (Ed.), Handbook of youthand justice (pp. 31–47). New York: Kluwer Academic/Plenum Press.
Zoccolillo, M. (2000). Parents’ health and social adjustment. Part II. Social adjustment.In M. Jette, H. Desrosiers, R. E. Tremblay, & J. Thibault (Eds.), Longitudinal studyof child development in Québec (ELDEQ 1998–2002). Québec: Institut de laStatistique du Québec.
Zoccolillo, M., Pickles, A., Quinton, D., & Rutter, M. (1992). The outcome of childhoodconduct disorder: Implications for defining adult personality disorder and conductdisorder. Psychological Medicine, 22, 971–986.
Zoccolillo, M., Price, R. K., Hwu, H. G., & Ji, T. (1998). Antisocial personality disorder:Comparisons of prevalence, symptoms, and correlates in four countries. In C.Slomkowski, P. Cohen, & L. N. Robins (Eds.), Where and when: Historical and geo-graphic effects on psychopathology. Mahwah, NJ: Erlbaum.
Intergenerational Transmission of Aggression 375

DETERMINANTS OF AGGRESSIONPeer Relationships and Aggressive Behavior
18
Peer Relationshipsand the Development of Aggressive
Behavior in Early Childhood
MICHEL BOIVIN, FRANK VITARO, and FRANÇOIS POULIN
Children’s peer relationships have been the object of persistent attention bythe developmental research community in the last decades. The prevalent viewamong members of this community is that children’s peer relationships serveimportant developmental functions (Asher & Coie, 1990; Hartup, 1983,1996; Rubin, Bukowski, & Parker, 1998). They provide contexts where chil-dren may acquire new social skills, learn to self-regulate and solve interper-sonal conflict, expand and validate their self-knowledge, and discover thesocial roles, norms, and processes involved in interpersonal relationships (Ru-bin et al., 1998).
Unfortunately, peer relationships are not always beneficial to the child, aswe discuss in this chapter; there are many ways in which specific features ofpeer relations may impede child development. The present review focuses onpeer relationship problems as they relate to aggressive behaviors, inasmuch asaggression has been identified as the prime behavioral correlate of a variety ofpeer relationship problems in childhood (Rubin et al., 1998; Coie & Dodge,1998). However, a comprehensive understanding of the association betweenaggressive behaviors and peer relation problems requires a differentiated viewof childhood peer relationships.
THE MANY FACES OF PEER RELATIONSHIPS
In examining children’s peer relationships, it is important to recognize thatchildhood peer relations are multifaceted. Children experience peer relations
376

through the group aggregate—for example, positive peer status, peer rejec-tion, and peer victimization—through peer networks (Cairns, Leung, Bu-chanan, & Cairns, 1995), and through dyadic relationships, generally withinfriendship relations (Hartup, 1996). The distinction between these broad-band classes of peer experiences is especially relevant when examining the as-sociation between aggressive behaviors and peer relationships.
At least two main streams of research have extensively documented thelink between aggressive behaviors and peer relationships in childhood. Thefirst line of research is on peer rejection. Indeed, aggressive behavior is themost commonly cited behavioral correlate of peer rejection, an index of nega-tive peer status derived from aggregated sociometric nominations (Rubin etal., 1998). Peer rejection is assumed to reflect difficulties in social integrationwithin the peer group. The second stream of research focuses on the nature ofthe peer affiliations of aggressive children. According to this line of research,aggressive children, mostly boys, are not socially left out by their peers, butrather tend to associate with each other (Boivin & Vitaro, 1995; Cairns,Cairns, Neckerman, Gest, & Gariépy, 1988; Dishion, Andrews, & Crosby,1995a).
Thus, aggressive children may suffer from exclusion and ostracism bynormative peers, as well as from problematic associations with deviantpeers. More fundamentally, this distinction in peer experiences also pertainsto putative processes of peer influence (i.e., peer exclusion processes), whichare mostly derived from aggregate measures, versus deviant peer social-ization processes, which mainly stem from dyadic relationship assessments.In other words, processes of peer influence are heterogeneous; that is,peers may marginalize, as well as provide support to, aggressive children.This review is organized to reflect this distinction in peer experience andprocess.
Over the past 50 years there has been substantial research aimed at docu-menting, and then at understanding, the nature and meaning of the link be-tween aggressive behaviors and peer relation problems (Hartup, 1983; Rubinet al., 1998). However, most of the more recent research effort has been cen-tered on school-age children. To our knowledge, no one has specificallyreviewed the evidence concerning the association between aggressive behav-iors and peer relation problems during the preschool period. This lack existsdespite evidence that peer relationships are already well established and quitedifferentiated in the preschool years (see the following section), and notwith-standing their potential influence on the early developmental course of aggres-sive behaviors. Furthermore, this question is especially timely and relevant tosocial policy issues because a growing number of children are exposed to peersearly in their lives through daycare (NICHD Early Child Care Research Net-work, 2003).
The purpose of this chapter is to provide an overview of the researchbearing on the issue of peer relationships and aggressive behaviors in earlychildhood, that is, during the preschool years and in kindergarten. We start by
Peer Relationships and Aggressive Behavior 377

reviewing some of the basic milestones in the early development of peer rela-tionships and aggressive behaviors.
THE FOUNDATIONS OF PEER INTERACTIONSAND PEER RELATIONSHIPS IN EARLY CHILDHOOD
Early toddlerhood witnesses the development of many social skills underlyingpeer interactions. Social interest in peers is clearly present in the first year oflife (Buhler, 1931; Vandell, Wilson, & Buchanan, 1980), but it is really duringthe second year that important gains in interactive skills can be observed. Bythe end of that year, toddlers have the ability to coordinate behavior in gameswith play partners. They can reciprocally imitate each other and progressivelylearn to alternate roles in play, thus indicating that they have acquired expec-tations about the behaviors of the other (Ross, Lollis, & Elliot, 1982; Strayer,1989). With the onset of language, peer interactions become more refined andelaborate. Between age 3 and age 5, there is a dramatic increase in pretendplay and in positive dyadic social behaviors, reflecting the augmented capacityof the child to adopt the perspective of the play partner (Rubin, Watson, &Jambor, 1978).
These emerging social interactive skills are the foundation of early peerrelationships. Signs of these relationships are seen in toddlers’ increasing be-havioral preference for specific peers (Howes, 1987; Strayer, 1989), and thosepreferences culminate in stable preschool friendships. These early friendshipsare often, but not exclusively, embedded in extensive affiliative networks(Strayer & Santos, 1996) and progressively become sex-segregated (Lafrenière,Strayer, & Gauthier, 1984; Maccoby, 1998; Pellegrini, in press). By age 3–4years, children initiate more play behavior and display more mutual activitywith friends than with nonfriends. Further, behavioral homophily, that is, thetendency to befriend or affiliate with peers whose behavior is similar to theirown, is already present by the end of preschool. Nevertheless, preschoolfriendships are not without disagreements, quarrels, and conflicts; there is in-deed more conflict among preschool friends than among preschool non-friends. However, these conflicts are resolved more equally and more often bythe use of negotiations and disengagement among friends than amongnonfriends (Hartup & Laursen, 1991).
Preschoolers also progressively internalize their perceptions about theirfriends, as well as about other peers with whom they are in contact. At leastby age 4, and when asked appropriately, preschoolers will readily and reliablyidentify best friends, as well as peers they like and peers they dislike. The ag-gregation of these perceptions reveals a coherent peer status structure withinthe larger group (Boivin & Bégin, 1986; Boivin, Tessier, & Strayer, 1985;Howes, 1987).
Physically aggressive behaviors are also manifested very early in life(Caplan, Vespo, Pederson, & Hay, 1991; Goodenough, 1931; Tremblay,
378 D E T E R M I N A N T S O F A G G R E S S I O N

2000). They become prevalent by the age of 2, peaking around age 3, beforedeclining steadily with age (Strayer, 1989; Tremblay, 2000; see also Tremblay& Nagin, Chapter 5, and Archer & Coté, Chapter 20, this volume). Theemergence of sex differences in aggressive behaviors seems to coincide withthe time children start interacting more regularly in preschool groups (see Ar-cher & Coté, Chapter 20, this volume). This transition is also characterized bya shift toward affiliation in the dyadic structure of social participation. Socialdominance hierarchies based on agonistic behaviors are established very early,even among groups of infants. However, the ratio of initiated agonistic activ-ity to initiated affiliation is much lower among older preschoolers (Strayer,1989).
Many factors may explain the decline in physical aggression, includinglanguage development (Dionne, Chapter 16, this volume), an augmented ca-pacity for perspective taking and empathy (Selman, 1980; Zahn-Waxler,Radke-Yarrrow, & King, 1979), and an increasing awareness of dominancehierarchies (Strayer & Trudel, 1984). There is also the possible emergence ofpeer group norms and implicit rules with respect to aggressive behaviors.These norms and rules are likely to curb aggressive behaviors by providing aconvergent feedback from group members to aggressors. All of this suggeststhat maturational changes and peer experiences compel a majority of youngchildren to progressively develop a capacity to use other means than overtphysical aggression to meet their goals. However, not all of them do.
Physical aggression is one specific form of aggressive behavior, and wealso need to attend to the other types and functions of aggressive behaviors tofully understand the nature of the developing association between these be-haviors and peer relationships. Of special interest here is the distinctionbetween direct and indirect or relational aggressive behaviors, as well as be-tween reactive and proactive aggression, as they reflect changes in thematurational and social processes underlying aggression. For instance, Bjork-qvist, Lagerspetz, and Kaukiainen (1992) have argued that the prevalent formof aggressive behaviors is likely to change over the years, with physically ag-gressive behaviors giving way to more verbal forms of aggression, and then tomore subtle and indirect forms (see Archer & Coté, Chapter 20, andVaillancourt, Chapter 8, this volume). According to Bjorkqvist and colleagues(1992), these changes would be brought about by the parallel maturationalchanges in language and social cognitive skills. It is important to determinewhether there is heretotypic stability in this normative change (i.e., the samechildren displaying these different forms of aggressive behaviors over theyears) and to assess to what extent peer relationships play a role in the variousdevelopmental trajectories of aggression.
Clearly, the social lives of preschoolers are elaborate and refined, and thedevelopment of peer relationships and aggressive behaviors are intertwined inthe preschool years. It is thus important to document and understand the na-ture of the early association between peer relations and aggressive behaviors.Thus, in the following section of this chapter, we focus on experiences of seg-
Peer Relationships and Aggressive Behavior 379

regation by peers in early childhood, specifically peer rejection or ostracism,as they relate to aggressive behaviors. Then we turn our attention to the roleof affiliating with aggressive peers in the development of aggressive behaviorsduring the same age period. In doing so, we adopt an ecological perspective;we consider different contexts of peer influence (e.g., friends and cliques,larger peer groups, daycare centers and schools) and examine the potential in-fluence of specific nonpeer agents (parents, teachers) on children’s relation-ships with peers.
AGGRESSIVE BEHAVIORS AND PEER REJECTIONAND OSTRACISM IN THE PRESCHOOL YEARS
Between 5 and 10% of children experience chronic rejection by peers (Coie &Dodge, 1983). These children are at risk for a variety of future adjustmentproblems, including both internalizing and externalizing difficulties, droppingout of school, and delinquency (Rubin et al., 1998; McDougall, Hymel,Vaillancourt, & Mercer, 2001). Peer rejection could be a cause of these adjust-ment problems, but this predictive association could also result from anunderlying factor, such as the child’s tendency to aggress or another relatedfactor (see Parker & Asher, 1987). There has been much valuable researchaimed at confirming the truly causal contribution of peer rejection to latermaladjustment (see McDougall et al., 2001). However, evidence substantiat-ing the causal model has been mixed, especially in studies examining the lon-gitudinal associations between peer relationships and aggressive behaviors.This is why the nature of the association between peer rejection and aggressivebehaviors has been, and is still, the focus of much interest.
There has been abundant research linking aggressive behaviors and nega-tive peer status. Some of this research has been conducted with preschool chil-dren (e.g., Boivin, Dorval, & Bégin, 1990; Hartup, Glazer, & Charlesworth,1967; Masters & Furman, 1981). This research indicates that preschool nega-tive peer status is not only associated with a negative aggressive behaviorstyle, but is also linked to more negative peer experiences (Boivin et al., 1990;Masters & Furman, 1981). These negative peer experiences could reflect thegive-and-take of aggressive exchanges. They could also antecede, or followfrom, the child’s aggression. However, because behavior and status were mea-sured concurrently in these studies, the causal nature of the relation was indoubt.
The question of the direction of the relation between aggressive behaviorsand peer rejection was tested more straightforwardly in a series of experimen-tal play-group studies (Coie & Kupersmidt, 1983; Dodge, 1983). In thesestudies, unacquainted boys were typically assembled and observed in smallgroups, where their emerging peer status was also assessed. Verbal and physi-cal aggressive behaviors were shown to antecede the emergence of peer rejec-tion, which suggested that these behaviors could be considered as a proximal
380 D E T E R M I N A N T S O F A G G R E S S I O N

determinant of these difficulties (Coie, 1990). However, because these studieswere conducted with school-age children, it is not clear whether this could bethe case in preschool.
Dodge, Coie, Pettit, and Price (1990) came close to providing an answerto this question in a study that looked at the play-group behaviors of first-grade children and those of third-grade children in order to investigate age dif-ferences in the behavioral predictors of peer status. They also looked at differ-ent forms of aggressive behaviors, including reactive aggression, instrumentalaggression, bullying, and rough play. Both angry reactive aggression and in-strumental aggression, but not rough play, were associated with negative peerstatus at the end of the sessions at both ages. Bullying was negatively relatedto status, but only in the third-grade groups. In the first-grade groups, themore popular boys displayed more bullying against aggressive boys, as if toestablish their dominance, whereas for older boys, this goal could be attainedby more sophisticated strategies. However, it was not clear whether this couldbe a finding specific to the social ecology of first-graders or whether it re-flected a more general developmental trend. Further research in the preschoolyears is needed to clarify this issue.
The studies described so far concur in indicating that aggressive behav-iors could be a proximal cause of peer rejection, at least in middle childhood.However, it is actually difficult to form a definitive conclusion for the pre-school period. Experimental play-group studies aimed at assessing differenttypes of aggressive behaviors of unacquainted young children may thus be agood strategy to determine whether specific aggressive behaviors antecedepeer rejection in the preschool years.
The fact that aggressive behaviors may be a proximal cause of peer expe-riences does not mean that the experience of peer rejection could not also be acause of aggressive behaviors. In other words, the relation between aggressivebehaviors and peer rejection could very well be bidirectional. In fact, there isat least some indirect evidence suggesting that rejection by peers yields aggres-sive interactions. For instance, when the same behavior is manifested by apopular child and a rejected child, the rejected child received a less positive an-swer (Dodge, 1983). In familiar groups, rejected boys are more often the tar-get of aggression than popular boys, whereas no difference is found innonfamiliar groups (Coie & Kupersmidt, 1983). These observations are inline with the two studies of preschoolers presented earlier (Masters &Furman, 1981; Boivin et al., 1990). They suggest that status acquisition inpeer group differs from status maintenance, in which reputation biases andscapegoat processes operate (Coie, 1990; Hymel, Wagner, & Butler, 1990).Thus, once established, peer rejection may be associated with negative peerexperiences, and aggressive behaviors could also be adopted as a response tothese experiences.
Could peer rejection and negative peer experiences during the preschoolyears contribute to subsequent adjustment problems, including aggressive be-haviors? The evidence with preschool children is scarce, but a series of studies
Peer Relationships and Aggressive Behavior 381

showed that cumulative peer rejection, starting in kindergarten, predictedgrowth in reactive aggression (rather than proactive aggression; Dodge et al.,2003). Interestingly, in one of these studies, peer rejection in kindergarten con-tributed uniquely to the prediction of aggressive behaviors in third grade, be-yond peer rejection in first grade and peer rejection in second grade. Further-more, early aggression also moderated the effect of rejection: Only childrenwho were above the median in aggression were likely to show increases in ag-gression linked to their cumulative experience of peer rejection over the years.These results strongly suggest that the experience of rejection in kindergartenmay exacerbate aggressive behavior development only among children ini-tially disposed toward aggression. The fact that it affected reactive aggression,rather than proactive aggressive behaviors, also supports the view that peer re-jection is a condition predisposing the child to aggressive behaviors.
In sum, the empirical evidence favors a bidirectional model of the associa-tion between peer rejection and aggressive behaviors. Aggressive behaviorsmay lead to rejection by peers, and the experience of being rejected by peers inchildhood may play an incremental role in the later development of aggressivebehaviors. Not only can this evidence be traced back to the preschool years,but the experience of rejection in kindergarten seems critical in that predic-tion. It would be highly relevant at this point to substantiate further the poten-tial contribution of peer rejection experiences in preschool to later forms ofaggression. In particular, there are specific lines of inquiry that would requireattention.
The first line of inquiry is process oriented. We need to move beyond thesimple description of the associations between aggressive behaviors and peerrelations, however specific they may be, to better understand the “dynamics”of these specific behavior–relationship associations during early childhood.Specifically, intraindividual sociocognitive and emotional processes (e.g., so-cial information processing deficits; Crick and Dodge, 1994) may underline,moderate, or mediate the links between peer relations and aggressive behav-iors. The early experiences of rejection could influence the child’s sociocogni-tive processes, leading to social-cognitive deficits and biases, and result inincreased aggressive behaviors. For example, retrospective reports of harshparenting has been associated with the child’s aggressive behaviors in kinder-garten, and specific social-cognitive deficiencies, including hostile biases in at-tributions, accounted for this relation (Dodge, Bates, & Pettit, 1990). Theseresults suggest that social-cognitive processes may mediate the relation be-tween specific aversive experiences and the early development of aggressivebehaviors. Mediation through social-cognitive process was also found byDodge et al. (2003): Negative peer status significantly predicted later aggres-sion, and a significant part of that prediction was accounted for by social-cognitive processes. What is needed is a closer examination of these processesas a function of different forms and functions of aggressive behaviors in pre-school.
The second line of inquiry is also process oriented, but aims at describing
382 D E T E R M I N A N T S O F A G G R E S S I O N

the nature of the peer experiences that are conducive to higher aggression. Re-search on peer victimization may be highly relevant to this objective in that itspecifically targets children who truly experience negative treatments by peers.Indeed, peer rejection and peer victimization are related phenomena, but theyshould not be confounded. Peer rejection is a measure of disliking and reflectsa negative attitude of the peer group toward a child, whereas peer victimiza-tion is a class of negative behaviors displayed toward a child (Boivin &Hymel, 1997; Boivin, Hymel, & Hodges, 2001). Peer victimization has mostlybeen documented in middle childhood, but there is evidence that such difficul-ties may exist in the preschool years (Alsaker & Valkanover, 2001; Kochen-derfer & Ladd, 1996), even as early as age 3 for specific forms of indirect orrelational victimization (Crick, Casas, & Ku, 1999). However, the main pointhere is that we need to document the nature of the peer interactions that areassociated with actual peer rejection in preschool. Asher, Rose, and Gabriel(2001) have recently identified more than 30 types of rejecting behaviors,which they classified in six different categories (from controlling and dominat-ing a child, to preventing a child from having access to a friend). Some of theseways of rejecting may be relevant for preschoolers. What is needed is a morecomplete and integrated view of how preschoolers reject another child.
Third, social peer interaction processes may also underlie, mediate, ormoderate the negative impact of aversive peer experiences, as well as the asso-ciation between peer relations and aggressive behaviors. For example, groupnorms (Boivin, Dodge, & Coie, 1995; Stormshak, Bierman, Bruschi, Dodge,& Coie, 1999) and specific relationships (e.g., affiliation with aggressive chil-dren; Boivin & Vitaro, 1995; having a protective friend; Hodges, Boivin,Vitaro, & Bukowski, 1997) have been shown to moderate the links betweenpeer experiences and aggressive behaviors among school-age children. Weneed to know whether and how these moderation effects operate in the pre-school period.
Two final considerations are warranted: First, it is important to note thatsome aggressive children may actually possess fairly high social status in thegroup (e.g., Prinstein & Cillessen, 2003; Rodkin, Farmer, Pearl, & Van Acker,2000), especially if the groups norms are supportive or neutral in regard to ag-gressive behaviors (Boivin et al., 1995). This is more likely the case among youngchildren, because specific forms of aggressive behaviors (i.e., the instrumental/proactive type) are sometimes positively related to popularity. When this is thecase, aggressive behaviors may be more likely to be valued and reinforced by, aswell as generalized within, the peer group. Consequently, high-status aggressivepreschoolers should be the focus of attention in future research.
Finally, despite the growing awareness that girls and boys may expressaggressive behavior differently (Bjorkqvist et al., 1992; Crick & Grotpeter,1995; Underwood, 2003), most of the research on peer relations and aggres-sion has been conducted with boys. Greater attention should be devoted tosex differences in the early study of the linkages between direct and indirect/relational aggression and peer rejection in the preschool years.
Peer Relationships and Aggressive Behavior 383

AN ECOLOGICAL PERSPECTIVE OF PEER INFLUENCEON AGGRESSIVE BEHAVIOR DURING EARLY CHILDHOOD
A full understanding of the role played by the child’s peer affiliates must takeinto account the fact that these peer relationships are embedded in other,larger social contexts (Dishion, French, & Patterson, 1995b). Accordingly,different levels of the young child’s social world are discussed in the followingsections of this chapter. The interactions between friends and clique membersare examined first. Typically, however, these interactions are embedded in alarger peer group and take place in settings such as children’s homes, neigh-borhoods, daycare centers, and schools. These settings vary greatly in terms ofthe amount of structure, adult supervision, and type of peers encountered, allconditions that may have a bearing on the development of aggressive behav-iors. Finally, in analyzing these different contextual “levels,” we examine theroles of parents and teachers in overseeing, supervising, and managing thechild’s relationships with peers during the preschool period (Ladd & Le Sieur1995; Parke & O’Neil, 1999).
Friends and Cliques
Although they are rejected by conventional peers, many aggressive preadoles-cents and early adolescents have friends, and most of them participate incliques (Boivin & Vitaro, 1995; Cairns et al., 1988). This seems to be true alsofor young children (Farver, 1996; Snyder, Horsch, & Childs, 1997). For in-stance, Farver (1996) showed that children at the center of teacher-identifiedpeer networks of 4-year-olds were also the most aggressive.
Only a few studies have examined whether homophily with respect toaggression (i.e., the tendency for aggressive children to affiliate with other ag-gressive children) exists among young children. For instance, members ofteacher-identified cliques of 4-year-olds were found to be moderately similarin aggression, as assessed by independent observers (Farver, 1996). Four- tofive-year-old children were more likely to select friends who were similar in re-spect to teacher-rated aggression; that is, 50% of aggressive children’s friendswere aggressive, as compared with 12% of nonaggressive children’s friends(Snyder et al., 1997). A further study is worth considering, in spite of the factit was conducted with grade school children; in this study, clique membershipin first grade was associated with similar characteristics in respect to aggres-sion (Estell, Cairns, Farmer, & Cairns, 2002). Thus, the few available studiessuggest that aggression-related homophily exists in early childhood at thedyadic and the clique levels, conditions that extend onward, and perhaps in-crease, into middle childhood and adolescence.
Many convergent processes may account for aggression homophily(Kandel, 1978). Aggressive children may associate because they are activelyrejected by conventional peers and gradually cast out of the group, leavingthem with no other choice than to affiliate with other aggressive children
384 D E T E R M I N A N T S O F A G G R E S S I O N

(Patterson, Reid, & Dishion, 1992). It may be that aggressive childrenproactively select other aggressive children, partly because they share an inter-est for rough play, but also because their mutual selection allows them todominate other peers and to have better access to resources in the peer group(Boivin & Vitaro, 1995; Poulin & Boivin, 2000). This latter view is in linewith the confluence model of Dishion, Patterson, and Griesler (1994) thatposits a gradual convergence of aggressive children into homogeneous groupsof aggressive and risk-taking adolescents. The confluence model implies an in-creasingly active selection of aggressive peers who share similar attitudestoward aggression. Once established, these deviant associations may also ac-centuate the behavioral similarity by having aggressive children mutuallyinfluencing each other, through deviancy training (Dishion et al., 1995a; seethe discussion of “mechanisms” below). To the extent that these mechanismshave cumulative effects over time, aggressive children could be progressivelytrapped in a social environment that is increasingly segregated from that oftheir better-adjusted peers. There is evidence that aggression homophily in-creases with age from middle childhood to late adolescence (Dion & Boivin,2003; Neckerman, 1992). However, whether these processes are operative inthe preschool years is not known.
Are (Deviant) Friends Significant for Later Adjustment in Early Childhood?
Very few studies have examined this question among preschool children.Snyder et al. (1997) reported a short-term effect of interacting with aggressivepeers. They found that the amount of time preschoolers spent with aggressivepeers predicted an increase in observed and teacher-rated aggressive behaviorover a 3-month interval. Conversely, children who spent minimal time (lessthan 15%) with aggressive peers showed a decrease in aggression over thesubsequent 3-month period. The findings of Snyder et al. (1997) indicate thataggressive friends may have a negative influence during early childhood.However, this effect was measured only over a 3-month period. Whether asimilar long-term effect exists at that age is still unknown.
What Are the Mechanisms That Could Account for the Possible Influenceof Aggressive Friends?
The answer to this question assumes that aggressive peers mutually influenceeach other through some form of socialization mechanisms and that this influ-ence is not spurious, that is, attributable to a third factor such as a personaldisposition or a family factor (see Vitaro, Tremblay, & Bukowski, 2001).Work by Dishion and colleagues on rule-breaking talk during dyadic interac-tions involving deviant adolescents indicates that positive verbal reinforce-ment by peers of past, actual, and future deviant behaviors becomes increas-ingly important in shaping social behavior (Dishion et al., 1995a). Thisprocess, labeled “deviancy training,” has received substantial empirical sup-
Peer Relationships and Aggressive Behavior 385

port. For example, the association between exposure to deviant peers by age10 and growth in arrests, substance use, and sexual intercourse from ages 10to 18 is mediated by deviancy training at age 14 (Patterson, Dishion, &Yoeger, 2000). However, very little is known about deviancy training in youngchildren. A precise examination of this process with young children is neededto confirm its existence in early childhood.
Direct training through conflicts and coercion among friends or cliquemembers may also play a role in explaining how and why affiliation with ag-gressive friends might instigate or increase aggressive behaviors. In their clas-sic observational study of preschoolers’ aggressive interactions, Patterson,Littman, and Bricker (1967) showed that the principles of operant condition-ing maintain aggressive behavior in the peer context. They found that theresponse to an aggressive behavior conditioned the likelihood of reenactedaggression in the future. When a child who was the victim of aggression re-acted by withdrawal and submissive behaviors, the aggressor was more likelyto subsequently attack the same target than when the victim retaliated. Nega-tive reinforcement in coercive peer interactions may thus shape young chil-dren’s behaviors in the same way it shapes this behavior during child–adultcoercive interchanges. Similarly, the use of verbal aggression is followed bytermination of a conflict episode for aggressive children, whereas a positiveverbal interchange or the absence of a response terminates the conflict episodefor nonaggressive children (Snyder & Brown, 1983). In other words, aggres-sive children are negatively reinforced for using coercive behaviors with theirpeers, whereas nonaggressive children are reinforced for using nonaversivestrategies. Hence, peer interactions during preschool years are sometimes oc-casions for coercive interchanges among peers which may, under some condi-tions (e.g., child’s submissiveness, adult and peer tolerance of aggression),serve as learning opportunities for aggression. If these conditions are present,then preschool and early elementary school settings may provide traininggrounds for further shaping of aggressive behaviors in children who mayalready have learned them at home. It is not clear, however, whether this typeof training is taking place within dyadic friendships or among members of aclique. Given the higher incidence of conflicts among preschool friendshipsinvolving at least one aggressive child, this could be the case (Snyder et al.,1997). However that may be, there are possible caveats to the long-term im-pact of deviancy training in preschool.
Caveats to Deviancy Training in Preschool
Why would affiliating with aggressive friends have little long-term effect onthe behavior trajectory of young children? One possible reason is that pre-school children do not systematically and exclusively affiliate with other ag-gressive children as much as preadolescents or adolescents do. Indeed, in thethree preschool studies reviewed earlier, aggression-related homophily was
386 D E T E R M I N A N T S O F A G G R E S S I O N

moderate. Hence, behavioral homophily may not yet be firmly established andspecialized during the preschool years. Young aggressive children may haveaccess to a variety of peers and friends, and these experiences may mitigate thenegative influence of deviant peers. For instance, to the extent that many ag-gressive children are not rejected by conventional peers during early child-hood, they would be less likely to be deprived of positive socialization experi-ences.
Another related possibility is that the associations among aggressivepreschoolers may be short-lived. There is indeed evidence showing thatchildhood friendships are not always stable and that clique compositionmay change substantially over a period of weeks, especially when involvingaggressive children (Cairns et al., 1995; Parker & Seal, 1996). However,Snyder et al. (1997) found no difference in the stability of strong mutual af-filiations of aggressive and nonaggressive children, although aggressive chil-dren were less likely than nonaggressive children to establish strong mutualaffiliations with classmates. Other researchers also reported that friendshipsduring the preschool years can be relatively stable and intimate (Howes,1987).
Furthermore, young children’s friendships may not carry the same weightas those of older children. Indeed, these early friendships are based on con-crete exchanges rather than shared values and interests, loyalty, and mutualsupport (Bigelow, 1977), qualities that may induce greater mutual influenceon behavior.
Finally, aggressive preschoolers may simply be more sensitive to adultrules than older aggressive children. Hence, parents and teachers may exert amoderating effect on aggressive friends’ influence by controlling children’s be-havior more closely and effectively. Parental supervision may also play a simi-lar role. In contrast to those of adolescence, the preschool peer environmentsare more closely structured and supervised by adults. However, even duringpreschool years the social settings may vary greatly in terms of adult structureand supervision, and this could make a difference with respect to exposure todeviant peers. For instance, the degree of parents’ monitoring of their kindergar-ten child has been associated with the child’s exposure to deviancy-producingexperiences and aggressive peers in a variety of contexts (Kilgore, Snyder, &Lentz, 2000).
Longitudinal studies that examine whether aggression-related homophilyincreases from the preschool years to late childhood are clearly needed. In ad-dition, it would be important to determine whether homophily is driven byspecific forms of aggressive behaviors (i.e., proactive aggression rather thanreactive aggression), as this seems to be the case with older children (Poulin &Boivin, 2000). Moreover, it would be relevant to learn about the qualitativefeatures of friendships during this period in order to understand why the earlyencounters with aggressive children may or may not leave traces in the longterm.
Peer Relationships and Aggressive Behavior 387

The Larger Peer System
By and large, our discussion of the peer influence on aggression in early child-hood has been limited so far to the peers with whom the child forms affectivebonds and spends more time interacting with, such as friends or clique mem-bers. However, the larger peer group to which the child is exposed may alsohave a unique impact on the level of aggression, especially in the case of stablepeer groups, such as daycare groups or kindergarten classrooms. As alreadysuggested, group norms with respect to aggression seem to be very salient dur-ing early childhood and may interact with initial dispositions for aggression.For example, it was found that a high level of aggression in a first-grade class-room uniquely contributed to an increase in aggression over a 5-year periodfor children who were already displaying high levels of aggression (Kellam,Ling, Merisca, Brown, and Ialongo, 1998). In other words, aggressive first-graders may become more aggressive when they are placed in a classroomwhere the level of aggression is high.
A recent observational study found evidence of a contagion effect indaycare: Children were significantly more likely to behave aggressively afteran aggressive act had occurred than when no aggressive act had occurred(Goldstein, Arnold, Rosenberg, Stowe, & Ortiz, 2001). Interestingly, conta-gion was stronger for aggressive behaviors directed toward another child thanfor aggressive behaviors directed at an object. Aggressive behaviors are notisolated acts in the classroom, but rather seem to be part of a larger group sys-tem, characterized by chain reactions. A next step would be to examinewhether such contagion effects are associated with changes in individual be-haviors. These findings suggest that the ecology of peer influence during earlychildhood is complex and that all levels of the peer system should be consid-ered.
The Behavior Settings
As early as 2 years of age, children start to spend more time with peers than withadults (Ellis, Rogoff, & Cromer, 1981). During the early years, these peer inter-actions are mainly confined to the home, where parents are more likely to or-chestrate and supervise social interactions with siblings and other children. In-deed, there are some indications that interactions with peers within the familymay account for individual differences in early developmental trajectories of ag-gression. In particular, siblings appear to be a predominant source of agonisticbehavior for preschoolers and may provide a training ground in aggressive be-haviors for younger siblings (Dunn & Kendrick, 1982). For instance, Tremblayet al. (2004) found that having a sibling of close age (i.e., less then 2 years in agedifference) was the best overall predictor of being described as highly physicallyaggressive between 17 and 42 months of age. These findings underscore the po-tential contributing role of sibling relationship for revealing, and perhaps foster-ing, aggressive behaviors very early in life.
388 D E T E R M I N A N T S O F A G G R E S S I O N

However, parental monitoring of these aggressive episodes are key to mit-igating this potential negative effect. There are different ways in which parentsmay influence their child’s peer interactions and relationships. For example,parents can provide opportunities for positive friendships while prohibitingdeviant ones. Parents may provide direct opportunities for peer interactionsby organizing social encounters between their child and selected other chil-dren. They can also provide indirect opportunities by coaching their childabout peer interactions (Ladd & LeSieur, 1995). In doing so, parents may im-prove their child’s ability to access more competent peers (Parke & Ladd,1992). Parental practices such as monitoring their children’s peer affiliations(Dishion & McMahon, 1998) and prohibiting their interactions with deviantpeers (Mounts, 2001) have been extensively studied during the late childhood/adolescence period. More research is needed on how parents manage youngchildren’s affiliation with aggressive peers.
As children get older, they interact with peers in a larger variety of set-tings (Sinclair, Pettit, Harrist, Dodge, & Bates, 1994). Other significant adults,such as classroom teachers, daycare staff, or after-school activity leaders, maythen also contribute in shaping a child’s peer environments (Searcy &Meadows, 1994). These settings are likely to vary with respect to the degree ofstructure and supervision provided by adults, as well as of the characteristicsof the peers (i.e., heterogeneity in age, level of peer aggression, etc.). Some set-tings may be more likely than others to provide opportunities for associationsamong aggressive children, as well as for deviancy training. Typically, theseare settings where the demands placed on the child and the degree of adultsupervision are low. Three behavioral settings are relevant to this discussion:(1) the neighborhood, (2) the childcare facility, and (3) the school.
The Neighborhood
During the preschool years, children gradually begin to form informal peergroups in their neighborhoods (Ladd & Golter, 1988). These peer groups aretypically mixed-aged groups (Ellis et al., 1981). The behavioral characteristicsof a child’s friends in this setting are likely to vary according to the socioeco-nomic status of the families living in the neighborhood. In socially disadvan-taged neighborhoods, such as public housing neighborhoods, the density ofaggressive children is higher, potentially increasing the child’s exposure to de-viant peers. In these neighborhoods, close parental supervision is crucial forlimiting the child’s access to deviant peers (Pettit, Bates, Dodge, & Meece,1999). Deviant friendships in a neighborhood have been studied mostly withadolescents (Dishion et al., 1995b), but this context is also relevant for youn-ger children because their friendship networks are mostly composed of peersfrom the same neighborhood (Ladd & Golter, 1988). According to Sinclair etal. (1994), the neighborhood is where 4–5-year-old children, especially chil-dren from low SES families, spend the greatest amount of time with peers.This is also where they are most frequently exposed to aggressive peers. Devi-
Peer Relationships and Aggressive Behavior 389

ancy training in these settings is likely, given the lack of structure and the lackof adult supervision. However, it is not clear whether these experiences predictan increase in children’s aggressive behaviors over time, above and beyondpersonal dispositions, family characteristics, and social interactions at thedyadic level.
Daycare and After-School Care Programs
In western societies, the amount of time young children spend in daycare hasbeen steadily increasing in the last decades (NICHD Early Child Care Re-search Network, 2003). In typical daycare settings, children spend most oftheir time interacting with peers. Preliminary evidence seems to indicate thattime spent in daycare may be associated with higher rates of aggression(Belsky, 2002; NICHD Early Child Care Research Network, 2003). Peer ef-fects may partly be responsible for this (Fabes, Hanish, & Martin, 2003). Atabout 2–3 years of age, a clear social structure involving specific patterns ofaffiliations and cliques emerges (Strayer & Santos, 1996). Children of thatage, especially boys (Hawley, 1999), often use aggressive means to reach highstatus in the social structure. Some of these cliques may thus contribute to theuse of aggression by modeling or reinforcement processes.
As already alluded to, the degree of adult supervision and structure mayplay a moderating role with respect to the possibility of a peer influence on ag-gression in daycare settings. Indeed, research on the impact of daycare on chil-dren’s adjustment constantly points to adult variables (e.g., child–staff ratio,staff qualifications) to explain the observed effects (Vandell & Shumow,1999). For example, Goldstein et al. (2001) found that the immediate pres-ence of the daycare teacher was associated with less contagion of aggression in36–72-month-old children. With close supervision of peer interactions, adultsmay simply prevent aggressive interactions from occurring at all.
Arrangements for child care are still necessary when children reach kin-dergarten and first grade. Indeed, because of the increasing rates of maternalemployment, the 3:00–5:00 P.M. period represents a new opportunity for addi-tional peer interactions (Vandell & Shumow, 1999). Some arrangements forthis period of time involve a low level of peer contact, such as staying with aparent or with a sitter, whereas others involve more peer contact, such asafter-school centers and out-of-school formal activities including lessons (mu-sic, dance), clubs (scouts), and organized sports. After-school centers varyconsiderably in respect to child–staff ratio, degree of structure, types of activi-ties, and heterogeneity in children’s age (Vandell & Shumow, 1999). Some ofthese settings may be a fertile ground for peer influence on aggression. Pierce,Hamm, and Vandell (1999) found that negative peer interactions in after-school care settings were associated with boys displaying high levels ofexternalizing behaviors at school, suggesting continuity in peer difficultiesfrom one setting to another. Similar findings concerning after-school care werealso reported (Vandell & Corasaniti, 1988).
390 D E T E R M I N A N T S O F A G G R E S S I O N

Deviant peer processes are less likely to take place in out-of-school formalactivities: First-graders involved in these activities tend to show fewer behav-ioral problems (Posner & Vandell, 1994). In these settings, aggressive behav-iors are not tolerated because they disrupt the activities. The levels of structureand adult supervision are usually high, and participants also tend to comefrom higher-income families.
The School
When they reach kindergarten, children spend several hours every day inter-acting with a stable group of same-age classmates. Most of the research onchildren’s friendship networks and aggression reviewed earlier has been con-ducted in such same-age classroom settings. However, the school structuremay also influence the nature of the peer relations of aggressive children. Asearly as first grade, aggressive children may be assigned to a segregated set-ting, such as a special classroom that involves other students with behaviorproblems. These special classrooms create an environment where aggressivechildren are given opportunities for developing friendships with each other(Searcy & Meadows, 1994). In addition to increasing aggression through themechanisms already described, these friendship groups may contribute to thecreation of an in-group/out-group type of social division between aggressivechildren and their mainstream nonaggressive peers.
CONCLUSION
Preschoolers’ peer relationships are complex. Preschool children may face avariety of peer experiences in a diversity of contexts, and it is important not toneglect the potential role of these experiences in the development of aggressivebehaviors. A review of the research suggests that starting in preschool, there isa bidirectional and differentiated association between peer relationship prob-lems and aggressive behaviors. Hostile/reactive aggressive behaviors, ratherthan instrumental/proactive forms of aggression, appear associated with, andperhaps augmented by, peer rejection. Homophily, in respect to aggression, isnoticeable during the preschool years. This pattern of association should be ofconcern.
Be that as it may, the empirical evidence linking peer relationships and ag-gressive behaviors in preschool is still thin. For instance, most of the researcheffort has been conducted with kindergarten children, and we still know verylittle about the association between early experiences with peers and aggres-sive behaviors in prekindergarten children. Nor is it clear whether these earlypreschool peer experiences have long-term consequences.
Longitudinal studies, starting early in life, are necessary to evaluate thepotential impact of peer experiences in the long term with respect to subtypesof aggressive behaviors. These future studies should measure the various
Peer Relationships and Aggressive Behavior 391

forms of peer relationships at different levels (e.g., dyads, cliques, groups) andin a variety of contexts (i.e., home, daycare, neighborhood). They should ex-amine the microprocesses involved in interactions between aggressive childrenand their friends, in addition to the mediating and moderating processes link-ing aggressive behaviors and peer experiences. Specifically, gender differences,as well as the direct and indirect role of parents, other adults, and siblings,should be documented more systematically.
Research may also take advantage of genetically informative and longitu-dinal designs to test specific models relative to the personal and family deter-minants of peer relationships, and to examine the specific role of peer rela-tionships in later adjustment. Of special interest is the possibility that peerexperiences may account for a significant part of nonshared environmentalinfluences on children’s later aggressive behaviors. Such studies could alsohelp unravel the developmental processes presiding over the development ofchildren’s peer relationships. Specifically, there is a possibility that gene–environment correlation may partly explain the nature of young children’speer experiences, for instance, rejection by peers of a child’s behavior, but alsohis or her selection of specific peers (see Deater-Deckard, 2001).
Finally, future research may also examine the feasibility and the impact ofexperimental interventions aimed at exposing young aggressive children tononaggressive peers and modifying the composition of their social networks.These interventions could serve as experimental manipulations in order to testmore decisively the putative causal role of these peer-related variables in re-spect to aggressive behaviors.
ACKNOWLEDGMENTS
Preparation of this chapter was supported in part by grants from the Canadian Insti-tute of Health Research and the Canada Research Chair Program awarded to MichelBoivin.
REFERENCES
Alsaker, F. D., & Valkanover, S. (2001). Early diagnosis and prevention of victimizationin kindergarten. In J. Juvonen & S. Graham (Eds.), Peer harassment in school: Theplight of the vulnerable and victimized (pp. 175–195). New York: Guilford Press.
Asher, S. R., & Coie, J. D. (1990). Peer rejection in childhood. New York: CambridgeUniversity Press.
Asher, S. R., Rose, A. J., & Gabriel, S. W. (2001). Peer rejection in everyday life. In M.Leary (Ed.), Interpersonal rejection (pp. 105–142). New York: Oxford UniversityPress.
Belsky, J. (2002). Quantity counts: Amount of child care and children’s socioemotionaldevelopment. Journal of Developmental and Behavioral Pediatrics, 23(3), 167–170.
392 D E T E R M I N A N T S O F A G G R E S S I O N

Bigelow, B. J. (1977). Children’s friendship expectations: A cognitive developmentalstudy. Child Development, 48, 246–253.
Bjorkqvist, K., Lagerspetz, K. M. J., & Kaukiainen, A. (1992). Do girls manipulate andboys fight? Developmental trends in regard to direct and indirect aggression. Ag-gressive Behavior, 18, 117–127.
Boivin, M., & Bégin, G. (1986). Temporal reliability and validity of three sociometric sta-tus assessments with young children. Canadian Journal of Behavioral Sciences, 18,167–172.
Boivin, M., Dodge, K. A., & Coie, J. D. (1995). Individual–group behavioral similarityand peer status in experimental play groups: The social misfit revisited. Journal ofPersonality and Social Psychology, 69(2), 269–279.
Boivin, M., Dorval, M., & Bégin, G. (1990). Statut auprès des pairs et comportement so-cial à la maternelle: Profils individuels et dyadiques. Revue Canadienne des Sciencesdu Comportement, 22, 121–139.
Boivin, M., & Hymel, S. (1997). Peer experiences and social self-perceptions: A two-stage mediational model. Developmental Psychology, 33, 1, 135–145.
Boivin, M., Hymel, S., & Hodges, E. V. E. (2001). Toward a process view of peer rejectionand harassment. In J. Juvonen & S. Graham (Eds.), Peer harassment in school: Theplight of the vulnerable and victimized (pp. 265–289). New York: Guilford Press.
Boivin, M., Tessier, O., & Strayer, F. F. (1985). La cohérence des choix sociométriques etl’évaluation de l’amitié chez les enfants d’âge pré-scolaire. Enfance, 329–343.
Boivin, M., & Vitaro, F. (1995). The impact of peer relationships on aggression in child-hood: Inhibition through coercion or promotion through peer support. In J.McCord (Ed.), Coercion and punishment in long-term perspectives (pp. 183–197).Cambridge, UK: Cambridge University Press.
Buhler, C. (1931). The social behavior of the child. In C. Murchison (Ed.), A handbook ofchild psychology (pp. 393–431). New York: Russell & Russell.
Cairns, R. B., Cairns, B. D., Neckerman, H. J., Gest, S. D., & Gariépy, J.-L. (1988). Socialnetworks and aggressive behavior: Peer support or peer rejection? DevelopmentalPsychology, 24, 815–823.
Cairns, R. B., Leung, M.-C., Buchanan, L., & Cairns, B. D. (1995). Friendships and so-cial networks in childhood and adolescence: Fluidity, reliability, and interrelations.Child Development, 66, 1330–1345.
Caplan, M., Vespo, J. E., Pedersen, J., & Hay, D. F. (1991). Conflict over resources insmall groups of 1- and 2-year-olds. Child Development, 62, 1513–1524.
Coie, J. D. (1990). Toward a theory of peer rejection. In S. R. Asher & J. D. Coie (Eds.),Peer rejection in childhood (pp. 365–402). New York: Cambridge University Press.
Coie, J. D., & Dodge, K. A. (1983). Continuities and changes in children’s social status: Afive-year longitudinal study. Merrill-Palmer Quarterly, 29, 261–281.
Coie, J. D., & Dodge, K. A. (1998). Aggression and antisocial behaviour. In W. Damon &N. Eisenberg (Eds.), Handbook of child psychology: Social emotional and personal-ity development (pp. 779–862). New York: Wiley.
Coie, J. D., & Kupersmidt, J. B. (1983). A behavioral analysis of emerging social status inboys’ groups. Child Development, 54, 1400–1416.
Crick, N. R., Casas, J. F., & Ku, H. C. (1999). Relational and physical forms of peer vic-timization in preschool. Developmental Psychology, 35(2), 376–385.
Crick, N. R., & Dodge, K. A. (1994). A review and reformulation of social information-processing mechanisms in children’s social adjustment. Psychological Bulletin,115(1), 74–101.
Peer Relationships and Aggressive Behavior 393

Crick, N. R., & Grotpeter, J. K. (1995). Relational aggression, gender, and social-psycho-logical adjustment. Child Development, 66, 710–722.
Deater-Deckard, K. (2001). Annotation: Recent research examining the role of peer rela-tionships in the development of psychopathology. Journal of Child Psychology andPsychiatry, 42(5), 565–579.
Dion, É., & Boivin, M. (2003). Band apart: A developmental perspective on aggressiveboys’ peer affiliation. Manuscript submitted for publication.
Dishion, T. J., Andrews, D. M., & Crosby, L. (1995a). Antisocial boys and their friends inearly adolescence: Relationship characteristics, quality, and interactional process.Child Development, 66, 139–151.
Dishion, T. J., French, D. C., & Patterson, G. R. (1995b). The development and ecologyof antisocial behavior. In D. Cicchetti & D. Cohen (Eds.), Manual of developmentalpsychopathology (pp. 421–471). New York: Wiley.
Dishion, T. J., & McMahon, R. J. (1998). Parental monitoring and the prevention ofchild and adolescent problem behavior: A conceptual and empirical formulation.Clinical Child and Family Psychology, 1, 61–75.
Dishion, T. J., Patterson, G. R., & Griesler, P. C. (1994). Peer adaptations in the develop-ment of antisocial behavior: A confluence model. In L. R. Huesmann (Ed.), Aggres-sive behavior: Current perspectives (pp. 61–95). New York: Plenum Press.
Dodge, K. A. (1983). Behavioral antecedents of peer social status. Child Development,53, 1386–1399.
Dodge, K. A., Bates, J. E., & Pettit, G. S. (1990). Mechanisms in the cycle of violence. Sci-ence, 250, 1678–1683.
Dodge, K. A., Coie, J. D., Pettit, G., & Price, J. (1990). Peer status and aggression in boys’groups: Developmental and contextual analyses. Child Development, 61, 1289–1309.
Dodge, K. A., Lansford, J. E., Salzer Burks V., Bates, J. E., Pettit, G. S., Fontaine, R., &Price, J. M. (2003). Peer rejection and social information-processing factors in thedevelopment of aggressive behavior problems in children. Child Development,74(2), 374–393.
Dunn, J., & Kendrick, C. (1982). Siblings. Cambridge, MA: Harvard University Press.Ellis, S., Rogoff, B., & Cromer, C. C. (1981). Age segregation in children’s social interac-
tion. Developmental Psychology, 29, 63–73.Estell, D. B, Cairns, R. B., Farmer, T. W., & Cairns, B. D. (2002). Aggression in inner-city
early elementary classrooms: Individual and peer-group configurations. Merrill–Palmer Quarterly, 48, 52–76.
Fabes, R. A., Hanish, L. D., & Martin, C. L. (2003). Children at play: The role of peers inunderstanding the effects of child care. Child Development, 74(4), 1039–1043.
Farver, J. A. M. (1996). Aggressive behavior in preschoolers’ social networks: Do birds ofa feather flock together? Early Childhood Research Quarterly, 11, 333–350.
Goldstein, N. E., Arnold, D. H., Rosenberg, J. L., Stowe, R. M., & Ortiz, C. (2001). Con-tagion of aggression in day care classrooms as a function of peer and teacher re-sponses. Journal of Educational Psychology, 4, 708–719.
Goodenough, F. L. (1931). Anger in young children. Minneapolis: University of Minne-sota.
Hartup, W. W. (1983). Peer relations. In P. H. Mussen & E. M. Hetherington (Eds.),Handbook of child psychology: Vol. 4. Socialization, personality, and social devel-opment (4th ed., pp. 103–196). New York: Wiley.
394 D E T E R M I N A N T S O F A G G R E S S I O N

Hartup, W. W. (1996). The company they keep: Friendships and their developmental sig-nificance. Child Development, 67, 1–13.
Hartup, W. W., Glazer, J. A., & Charlesworth, R. (1967). Peer reinforcement andsociometric status. Child Development, 38, 1017–1024.
Hartup, W. W., & Laursen, B. (1991). Relationships as developmental contexts. In R. Co-hen, & A. W. Siegel (Eds.), Context and development (pp. 253–279). Hillsdale, NJ:Erlbaum.
Hawley, P. H. (1999). The ontogenesis of social dominance: A strategy-based evolution-ary perspective. Developmental Review, 19, 97–132.
Hodges, V. E., Boivin, M., Vitaro, F., & Bukowski, B. (1999). The power of friendship:Protecting against an escalating cycle of peer victimization. Developmental Psy-chology, 35(1), 94–101.
Howes, C. (1987). Peer interaction of young children. Monographs of the Society for Re-search in Child Development, 53(No. 217).
Hymel, S., Wagner, E., & Butler, L. (1990). Reputation bias: View from the peer group. InS. R. Asher & J. D. Coie (Eds.), Peer rejection in childhood (pp. 156–186). NewYork: Cambridge University Press.
Kandel, D. B. (1978). Homophily, selection, and socialization in adolescent friendships.American Journal of Sociology, 84, 427–436.
Kellam, S. G., Ling, X., Merisca, R., Brown, C. H., & Ialongo, N. (1998). The effect ofthe level of aggression in the first grade classroom on the course and malleability ofaggressive behavior into middle school. Development and Psychopathology, 10,165–185.
Kilgore, K., Snyder, J., & Lentz, C. (2000). The contribution of parental discipline, pa-rental monitoring, and school-based risk to early-onset conduct problems in Afri-can-American boys and girls. Developmental Psychology, 36, 1–11.
Kochenderfer, B. J., & Ladd, G. W. (1996). Peer victimization: Cause or consequence ofschool maladjustment? Child Development, 67, 1305–1317
Ladd, G. W., & Golter, B. S. (1988). Parents’ initiation and monitoring of children’s peercontacts : Predictive of children’s peer relations in nonschool and school settings?Developmental Psychology, 24, 109–117.
Ladd, G. W., & Le Sieur, K. D. (1995). Parents’ and children’s peer relationships. In M. H.Bornstein (Ed.), Handbook of parenting (pp. 377–409). Hillsdale, NJ: LEA.
Lafrenière, P. J., Strayer, F. F., & Gauthier, R. (1984). The emergence of same-sexaffiliative preferences among preschool peers: A developmental/ethological per-spective. Child Development, 55, 1958–1965.
Maccoby, E. E. (1988). The two sexes: Growing up apart, coming together. Cambridge,MA: Harvard University Press.
Martin, C. L., & Fabes, R. A. (2001). The stability and consequences of young children’ssame-sex peer interactions. Developmental Psychology, 37, 431–446.
Masters, J. C., & Furman, W. (1981). Popularity, individual friendship selection, and spe-cific peer interaction among children. Developmental Psychology, 3, 344–350.
McDougall, P., Hymel, S., Vaillancourt, T., & Mercer, L. (2001). The consequences ofearly peer rejection. In M. Leary (Ed.), Interpersonal rejection (pp. 213–247). NewYork: Oxford University Press.
Mounts, N. S. (2001). Young adolescents’ perceptions of parental management of peerrelationships. Journal of Early Adolescence, 21, 92–122.
Neckerman, H. J. (1992). A longitudinal investigation of the stability and fluidity of so-
Peer Relationships and Aggressive Behavior 395

cial networks and peer relationships of children and adolescents. Dissertation Ab-stracts International, 53, 9–B.
NICHD Early Child Care Research Network (2003). Does amount of time spent in childcare predict socioemotional adjustment during the transition to kindergarten?Child Development, 74(4), 976–1005.
Parke, R. D., & Ladd, G. W. (1992). Family–peer relationships: Modes of linkage.Hillsdale, NJ: Erlbaum.
Parke, R. D., & O’Neil, R. (1999). Social relationships across contexts: Family–peer link-ages. In W. A. Collins & B. Laursen (Eds.), Relationships as developmental contexts:Minnesota Symposia on Child Psychology (Vol. 30, 211–239). Hillsdale, NJ: LEA.
Parker, J. G., & Asher, S. R. (1987). Peer acceptance and later personal adjustment: Arelow-accepted children “at risk”? Psychological Bulletin, 102(3), 357–389.
Parker, J. G., & Seal, J. (1996). Forming, losing, renewing, and replacing friendships: Ap-plying temporal parameters to the assessment of children’s friendship experiences.Child Development, 67, 2248–2268.
Patterson, G. R., Dishion, T. J., & Yoerger, K. (2000). Adolescent growth in new forms ofproblem behavior: Macro- and micro-peer dynamics. Prevention Science, 1, 3–13.
Patterson, G. R., Littman, R. A., & Bricker, W. (1967). Assertive behavior in children: Astep toward a theory of aggression. Monographs of the Society for Research inChild Development, 32(No. 5).
Patterson, G. R., Reid, J. B., & Dishion, T. J. (1992). A social learning approach: Vol. 4.Antisocial boys. Eugene, OR: Castalia Press.
Pellegrini, A. D. (in press). Sexual segregation in childhood: A review of evidence for twohypotheses. Animal Behaviour.
Pettit, G. S., Bates, J. E., Dodge, K. A., & Meece, D. W. (1999). The impact of after-schoolpeer contact on early adolescent externalizing problems is moderated by parentalmonitoring, perceived neighbourhood safety, and prior adjustment. Child Develop-ment, 70, 768–778.
Pierce, K. M., Hamm, J. V., & Vandell, D. L. (1999). Experiences in after-school pro-grams and children’s adjustment in first-grade classrooms. Child Development, 70,756–767.
Posner, J. K., & Vandell, D. L. (1994). Low-income children’s after-school care: Are therebeneficial effects of after-school programs? Child Development, 65, 440–456.
Poulin, F., & Boivin, M. (2000). The role of proactive aggression and reactive aggressionin the formation and development of boys’ friendships. Developmental Psychology,36, 233–240.
Prinstein, M. J., & Cillessen, A. H. N. (2003). Forms and functions of adolescent peer ag-gression associated with high levels of peer status. Merrill–Palmer Quarterly, 49,310–342.
Rodkin, P. C., Farmer, T. W., Pearl, R., & Van Acker, R. (2000). Heterogeneity of popularboys: Antisocial and prosocial configurations. Developmental Psychology, 36, 14–24.
Ross, H. S., Lollis, S. P., & Elliot, C. (1982). Toddler–peer communication. In K. H. Ru-bin & H. S. Ross (Eds.), Peer relationships and social skills in childhood (pp. 73–98). New York: Springer-Verlag.
Rubin, K. H., Bukowski, W., & Parker, J. G. (1998). Peer interactions, relationships,and groups. In W. Damon & N. Eisenberg (Eds.), Handbook of child psychol-ogy: Social emotional and personality development (Vol. 3, pp. 619–700). NewYork: Wiley.
396 D E T E R M I N A N T S O F A G G R E S S I O N

Rubin, K. H., Watson, K., & Jambor, T. (1978). Free play behaviors in preschool and kin-dergarten children. Child Development, 49, 534–536.
Searcy, S., & Meadows, N. B. (1994). The impact of social structures on friendship devel-opment of children with behavior disorders. Education and Treatment of Children,17, 255–266.
Selman, R. L. (1980). The growth of interpersonal understanding. New York: AcademicPress.
Sinclair, J. J., Petit, G. S., Harrist, A. W., Dodge, K. A., & Bates, J. E. (1994). Encounterswith aggressive peers in early childhood: Frequency, age differences, and correlatesof risk for behaviour problems. International Journal of Behavioral Development,17, 675–696.
Snyder, J., & Brown, K. (1983). Oppositional behavior and noncompliance in preschoolchildren. Behavioral Assessment, 5, 333–348.
Snyder, J., Horsch, E., & Childs, J. (1997). Peer relationships of young children:Affiliative choices and the shaping of aggressive behavior. Journal of Clinical ChildPsychology, 26, 145–156.
Stormshak, E. A., Bierman, K. L., Bruschi, C., Dodge, K. A., & Coie, J. D. (1999). The re-lation between behavior problems and peer preference in different classroom con-texts. Child Development, 70, 169–182.
Strayer, F. F. (1989). Co-adaptation within the early peer group: A psychobiological studyof social competence. In B. H. Schneider, G. Attili, J. Nadel, & R. Weissberg (Eds.),Social competence in developmental perspective (pp. 145–174). Boston: KluwerAcademic.
Strayer, F. F., & Santos, A. J. (1996). Affiliative structures in preschool peer groups. SocialDevelopment, 5, 117–130.
Strayer, F. F., & Trudel, M. (1984). Developmental changes in the nature and function ofsocial dominance among young children. Ethology and Sociobiology, 5, 279–295.
Tremblay, R. E. (2000). The development of aggressive behavior during childhood: Whathave we learned in the past century? International Journal of Behavioral Develop-ment, 24, 129–141.
Tremblay, R. E., Nagin, D. S., Séguin, J. R., Zoccollillo, M., Zelazo, P., Boivin, M.,Pérusse, D., & Japel, C. (2004). Physical aggression during early childhood: Trajec-tories and predictors. Pediatrics, 114(1), e43–e50.
Underwood, M. K. (2003). Social aggression in girls. New York: Guilford Press.Vandell, D. L., & Corasaniti, M. A. (1988). The relations between third graders’ after-
school care and social, academic, and emotional functioning. Child Development,59, 868–875.
Vandell, D. L., & Shumow, L. (1999). After-school child care programs. The Future ofChildren, 9, 64–80.
Vandell, D. L., Wilson, K. S., & Buchanan, N. R. (1980). Peer interaction in the first yearof life: An examination of its structure, content, and sensitivity to toys. Child Devel-opment, 51, 481–488.
Vitaro, F., Tremblay, R. E., & Bukowski, W. M. (2001). Friends, friendships, and conductdisorders. In J. Hill & B. Maughan (Eds.), Conduct disorder in childhood (pp. 346–378). Cambridge, UK: Cambridge University Press.
Zahn-Waxler, C., Radke-Yarrow, M., & King, R. A. (1979). Child-rearing and children’sprosocial initiations toward victims of distress. Child Development, 50, 319–330.
Peer Relationships and Aggressive Behavior 397

DETERMINANTS OF AGGRESSIONSocial Capital and Physical Violence
19
Social Capital and Physical Violence
UBERTO GATTI and RICHARD E. TREMBLAY
Man is made fit for Society not by nature, but by training.—THOMAS HOBBES, On the Citizen (1647/1998, p. 25)
God makes all things good; man meddles with them and theybecome evil.
—ROUSSEAU, Émile (1762/1986, p. 5)
Although Rousseau and Hobbes disagreed on the nature of “natural” disposi-tions, they agreed on the fact that the social environment has an important im-pact on social development. It is generally accepted today, as it has been byphilosophers since Plato and Aristotle, that the social development of childrenis influenced by the environment in which they live, and in particular by therelationships they have with their own families and communities. The as-sumption is that “good” social environments will contribute to good psycho-social development, whereas, on the contrary, “bad” social environments willfacilitate social adjustment problems.
The aim of this chapter is to review the literature on the effects of socialenvironments on the development of physical aggression. We have chosen toconsider the social environment in terms of a relatively recent unifying con-cept, social capital. In recent years, this concept has increasingly come to thefore. Initially formulated to explain such phenomena as social stratificationand economic development, it has since been utilized to interpret various as-pects of life in society.
Social capital has been defined as interpersonal relationships that facili-tate action (Bourdieu, 1980). Social capital therefore represents an aspect ofsocial reality that individuals (or groups) utilize as a resource in order to fur-ther their own interests and to achieve goals that would otherwise be beyond
398

their reach (Coleman, 1990); that is to say, it is the investment and the instru-mental use of the resources inherent in social networks (Lin, 1999). Or, again,it has been seen as interpersonal trust (Fukuyama, 1995), or networks, norms,and trust that enable participants to act together more effectively to pursueshared objectives (Putnam, 1993). Social capital constitutes a resource that isoften compared to physical capital (tools, machinery, etc.) and human capital(skills acquired by the individual through education and socialization). Physi-cal capital is created through the transformation of raw materials; human cap-ital through the transformation of persons; social capital through changes inthe relationships between persons (Coleman, 1990).
These succinct definitions hint at two different (though not entirely inde-pendent) interpretations of the concept of social capital: The first, which ischiefly microsocial, stresses the importance of relationships, social networksor links that the individual can utilize; the second, which is mainly macro-social, attaches greater importance to the norms of reciprocity and civic en-gagement as the features of a given society. In general, however, it has to beadmitted that the definitions proposed for social capital are quite numerous;indeed, debate is currently under way with a view to clarifying and definingthe concept, the excessive extension of which may compromise its heuristicvalue (Portes, 1998).
The notion that social capital is linked to the development of children hasbeen argued especially by Coleman (1990), who draws a distinction betweensocial capital within the family and outside the family. In reality, however,leaving aside this dichotomy, it can be claimed that many diverse forms of so-cial capital exist and that these can be detected at different levels, rangingfrom the microsocial, which centers on the family, to the macrosocial, whichtakes the nation, region, or city as its unit of analysis; at an intermediate level,the district or neighborhood may be used as the unit of observation. More-over, social capital may exert different effects in different phases of develop-ment. It therefore seems appropriate to reflect on how the different types ofsocial capital might influence aggressive behavior at various ages. By settingthe developmental phases against the greater or lesser breadth of the contextin which the effect is felt, we created a classification for our analysis of theeffects of social capital. Table 19.1 illustrates the hypothesis that, in general,social capital at the microsocial level acts primarily in the early years of life,and at the macrosocial level its effects are chiefly felt during adulthood.
THE CHILD WELFARE SYSTEM
A form of social capital that may affect children’s adjustment, and thereforethe likelihood of aggressive behavior, is constituted by social and financial as-sistance to parents (generally mothers) in difficulty. In modern societies, wel-fare systems have been developed that loosen the bonds between the citizenand the labor market and compensate for financial disadvantages, thereby
Social Capital and Physical Violence 399

making health, social, and educational services available to people who wouldotherwise be unable to afford them. The relationships between such invest-ments and crime rates have generally been examined through synchronousdata. Messner and Rosenfeld (1997), for example, showed that investments inwelfare were able to reduce anomic pressure and, consequently, homiciderates in the various nations considered.
In an investigation involving several countries, Pampel and Gartner(1995) examined the effect of demographic changes—particularly increases inthe number of young people—on general homicide rates. These authors foundthat in countries where institutions for collective social protection were weak,an increase in the number of young people led to a rise in the homicide rate,whereas no such effect was recorded in countries where these institutions werestrong. In comparing homicide rates in various countries, Savolaien (2000)found that the effect of economic inequality on this type of crime was nonexis-tent in countries with high rates of investment in welfare.
DeFronzo conducted a series of studies on the relationship between the fi-nancial level of welfare aid and crime rates. In a study of 39 U.S. metropolitanareas, he found, after controlling for other social and economic factors, thatthe level of public assistance to poor families correlated negatively with therates of various crimes, including homicide, sexual assault, and burglary(DeFronzo, 1983). In a subsequent study (DeFronzo,1997) he examined theeffect of the mean level of social assistance, per person, in 141 U.S. cities, onhomicide, while controlling for a whole range of socioeconomic variables, andfound that the level of social assistance (Aid to Families with DependentChildren, AFDC) had a direct negative impact on homicide rates; moreover, itexerted an indirect negative effect on these rates through its association withfamily status (by reducing the number of families headed by a single mother).The high level of economic support therefore apparently reduced homiciderates through two different causal mechanisms. Note that all these studiesconcerned general crime rates, resulting from both juvenile and adult crime.
400 D E T E R M I N A N T S O F A G G R E S S I O N
TABLE 19.1. Action of Social Capital at Different Ages and in Different Contexts
0–2 years 3–5 years 6–11 years 12–18 years > 18 years
Child welfare system + + + +
Family + + + +
Daycare, preschool, school + + + +
Peers + + + +
Neighborhood + +
Town or region + +
Gang + +
Violent subcultureand organized crime
+ +

These results fall within the framework of the broader question of the ef-fects of welfare intervention that have been debated in recent decades. Indeed,such intervention is regarded by some as being counterproductive, fosteringdependence and passivity, stifling effort, and promoting marginalization; oth-ers, however, regard it as an indispensable tool for reducing the deprivation ofthose who belong to the marginal sectors of society, and, in the final analysis,for improving their social adjustment.
Awareness of the effects of welfare intervention acquired particular im-portance in the United States when, in 1996, a radical change in welfare policytook place. In that year the AFDC program was abolished and was replacedby the more restrictive Personal Responsibility and Work Opportunity Recon-ciliation Act (PRWORA), which limited the period of eligibility for aid andimposed prescriptions in the field of work. As a result, the number of familieson assistance, which had peaked at 5 million in 1994, had been halved byJune 1999 (Ku, 2001) and further declined to about 2 million in 2001. Ac-cording to The Economist (2001), the success of this reform went beyond themost optimistic forecasts, yet other commentators judged it to be a substantialfailure (Cancian, 2001). Still other experts, however, viewed the results of thereform as partly positive and partly negative. Zaslow, Tout, Smith, and Moore(1998), for instance, saw the results as positive in those cases in which moth-ers actually succeeded in finding and keeping a job, thus increasing family in-come, but as potentially negative when mothers were unable to find or keep ajob once the maximum permitted time on assistance had been reached. Morris(2002) analyzed the main evaluation studies and concluded that the new wel-fare structure may be advantageous for children but potentially harmful to ad-olescents; since the reform was implemented, smoking, alcohol use, small-timedelinquency, and maladjustment at school appear to have increased amongteenagers, who receive less supervision from their families. A particular nega-tive effect of the reform has been highlighted by Paxon and Waldfogel (2001),who claim that as a result of reduced aid to families in difficulty, cases of childabuse have increased.
The heated debate and the research efforts aimed at assessing the resultsof aid programs emphasize the importance of welfare systems in the politics ofa nation, especially in regard to their repercussions on families and child de-velopment. From a criminological point of view, it has been suggested (Vila,1994) that all of the factors that enhance child development, such as healthcare, education, and parenting, have a long-term effect and are able to reducecrime a generation later (the “nurturance” hypothesis). To test this hypothesis,Savage and Vila (1997) carried out analyses, with countries as units, aimed atascertaining whether indicators of child welfare were able to predict crime lev-els 20 years later. The nurturance hypothesis was not supported for crime ingeneral, but only for crimes of violence. In a further study, again carried outon a sample of countries, Savage and Vila (2002) examined the relationshipsbetween changes in child care services and changes in crime rates recorded in alater period. As covariates, the authors considered measures of economic
Social Capital and Physical Violence 401

growth and inequality, factors that reflected the opportunities to commitcrimes, and demographic factors. The results supported the nurturance hy-pothesis: (1) Improvements in primary school enrollments and in the numberof hospital beds available per capita predicted declines in homicide rates, (2)improvements in infant mortality rates, life expectancy at birth, and the num-ber of hospital beds predicted declines in the rates of violent crime, (3) and im-provements in life expectancy at birth were associated with lower rates oftheft.
Similar results were obtained by Gatti, Schadee, and Tremblay (2002a),who tested the hypothesis that better maternal and infant health care will bereflected in reduced crime rates a generation later. They analyzed the relation-ship between infant mortality and the lagged rates of homicide (20 years later)in the 95 Italian provinces, controlling for gross domestic product. Anotherform of social capital is constituted by programs that provide support forhigh-risk families. The Elmira Home Visitation Study (Olds et al., 1997) is oneof the best examples. This randomized clinical trial showed that home visita-tion by a nurse, starting during pregnancy and lasting until the child’s secondbirthday, significantly reduced child abuse by the parents, substance abuse andarrests among the mothers, and juvenile delinquency among the children.
The relationships between the quality and intensity of welfare interven-tion, on one hand, and children’s adjustment, on the other, may be variouswhen viewed from the standpoint of social capital. These interventions mayhelp weave a network of relationships that goes beyond the nucleus of thefamily and facilitates access to different types of resources (e.g., educationaland recreational). To some extent, proper welfare intervention should makeup for the deficiencies in the family’s social capital, even though, as mentionedearlier, the effects of the welfare system are not completely clear and are stilldebatable.
FAMILY SOCIAL CAPITAL
How social capital should be defined at the family level is controversial. Onone hand, some, such as Wright, Cullen, and Miller (2001), claim that a verybroad conception should be adopted—one that includes any type of relation-ship involving the child, both within and through the family, and which there-fore takes into account the time and effort that parents devote to their chil-dren, the intense and lasting affective links that are forged, and the prosocialdirectives that are proposed. On the other hand, according to Sampson,Morenoff, and Earls (1999), many researchers adopt an excessively broadconception of social capital, including aspects of life within the family, such asparental monitoring and expectations, which should not be included; in thisperspective, the notion of social capital should concern links with the localcommunity. In this review we adopt an intermediate stance—one that consid-ers those aspects of family life that imply a greater or lesser link with the com-
402 D E T E R M I N A N T S O F A G G R E S S I O N

munity. This is in line with the suggestions made by Coleman (1990), whostressed that some characteristics of the family, in particular single-parentstatus and the presence of a working mother, may weaken links with the com-munity.
In regard to the single-parent family, several studies revealed a correlationbetween this condition and antisocial behavior on the part of children (see,e.g., Rutter, Giller, & Hagell, 1998). Vaden-Kiernan, Ialongo, Pearson, andKellam (1995) investigated the relationships between the family structures andteacher-rated aggression of children, within the framework of a longitudinalstudy of 1,197 children from 19 public elementary schools in Baltimore,Maryland. The relationships between fourth-grade family structure and sixth-grade aggression were examined separately for boys and girls, using logisticregression analyses comparing mother-alone families with the other families,and controlling for income level, urban area, and fourth-grade aggression.The boys from mother-alone families were more likely to be rated as more ag-gressive by their teachers, whereas this was not the case for the girls. Pagani,Boulerice, and Tremblay (1997) conducted a similar study with a randomsample of the population of kindergarten children in the Canadian province ofQuébec (947 girls and 882 boys). They used logistic regressions to predict age-12 fighting from family status, controlling for age-6 fighting and other familycharacteristics (education, age, poverty). A short-term increase in fighting forboys and girls could be attributed to family transitions in both directions, thatis, divorce (becoming a single parent) and remarriage (ending single-parentstatus). With a large sample (n = 1,037) of kindergarten boys from low socio-economic areas of Montreal, Nagin and Tremblay (2001) studied the predic-tive value of family break-up before school entry for trajectories of physicalaggression from 6 to 15 years of age. After controlling for parent characteris-tics (age at birth of child, education, employment status) and child characteris-tics (IQ and behavior problems), they showed that family breakup beforeschool entry increased the odds of being on a high physical aggression trajec-tory by close to 50%. In the Dunedin study, the fact that a 13-year-old was thechild of a single parent predicted the likelihood of convictions for violentcrimes at 18 years (Henry, Caspi, Moffitt, & Silva, 2004). In London,Farrington (1989) found that parent–child separation before the age of 10years predicted self-reported violence in adolescence and adulthood, as well asthe likelihood of convictions for violent crimes. In a more recent study,Tremblay et al. (2004) found that children born in Canada in 1997–1998 tosingle mothers were more at risk of being on a high-level physical aggressiontrajectory between 17 and 42 months after birth than other children. These re-sults were obtained after controlling for a large number of parent and familycharacteristics.
Another aspect of family life that has long been investigated concernsthe relationship between the delinquency of parents and that of their chil-dren. This relationship has been variously interpreted in terms of geneticpredisposition, psychological influence, or environmental factors. From the
Social Capital and Physical Violence 403

social capital perspective, it may also be claimed that parental delinquencyreduces family social capital, in that involvement in illicit activities mightnot only restrict children’s access to socially valid interpersonal and commu-nity relationships, but also facilitate contact with groups involved in crime.Several longitudinal studies have revealed the link between parental delin-quency and violence on the part of children (Smith & Farrington, 2003),as recorded through self-reported surveys and official records of violent of-fenses committed either in adolescence or adulthood. Particularly significantin this field is the research conducted by Keenan and Shaw (1994), whomeasured the frequency and pervasiveness of aggression from videotapes ofthe behavior of 98 mother–child couples when the children (males and fe-males) were 18 and 24 months old. This is one of the very few studies thathave examined physical aggression at a very early age, and measured it notonly through parental perception but also through direct observation. Theseauthors found that parental delinquency was one of the strongest predictorsof aggression for boys at 24 months (but not at 18 months). They thereforeclaimed that the link between familial delinquency and boys’ aggression wasstronger when the children had reached a developmental stage at which theparents began to discipline their children.
Using the U.S. National Youth Study, Wright et al. (2001) evaluatedfamily social capital, using a composite index that included the time parentsspent with their children, the strength of the bonds between various familymembers, and the parents’ critical attitude toward delinquency. They found aninverse relationship between social capital and both short- and long-terminvolvement in delinquency, evaluated through answers given by the youthsabout their involvement in a range of crimes, such as theft, assault, robbery,and drug dealing. McCord (2002) obtained similar results with data from theCambridge–Somerville study. Social capital was measured by social workers’observations of 232 families with children between 10 and 16 years of age atthe start of the research. Outcome data, collected more than 40 years after thebeginning of the study, indicated that the social capital of parents reduced therisk of their children’s being convicted of violent crimes.
According to Coleman (1990), maternal employment also has an effecton social capital. He suggests that if a mother works, she will have less chanceto participate in the life of the community, to develop relationships withneighbors, and to share educational problems, and that this will have a nega-tive influence on children’s adjustment. This aspect of family social capitalassumes particular relevance in modern societies, in which women’s employ-ment outside the home has become the norm. Some have expressed concernregarding the well-being of children on the grounds that their developmentand adjustment may be hindered if their mothers work (Belsky, 2001). VanderVen, Cullen, Carrozza, and Wright (2001) used data from the National Longi-tudinal Survey of Youth and found that the features of the mothers’ jobs hadrelatively little or no direct influence on the delinquency of their children, butdid have a slight indirect effect through supervision. Lack of supervision had
404 D E T E R M I N A N T S O F A G G R E S S I O N

an effect regardless of whether early or current employment was considered. Itwas found that the effects of maternal employment might differ according tothe population considered. For instance, Han, Waldfogel, and Brooks-Gunn(2001) found that, in the general population, maternal employment in the firstyear of the child’s life had an effect that could be perceived in the sphere ofexternalizing problems when the child was 7–8 years old, but among AfricanAmerican children this effect did not emerge. However, research on the effectsof maternal employment must include research on the impact of the quality ofcare given to young children while their parents are working. We turn next tothis topic.
DAYCARE AND EARLY CHILDHOOD DEVELOPMENT CENTERS
In the last few decades, a heated debate has raged in regard to children’sdaycare, to the extent that the term daycare war has been coined to describethe widespread and fierce clash of views on such institutions (Karen, 1994).Some argue that early child care services are an important social resource thatmothers can exploit to improve their children’s socialization. Others arguethat daily separation from the mother during the first few years of the child’slife may put the child’s development at risk. We obviously need good data todecide which resources constitute a sound capital for children’s development.But few good studies have been carried out to date, and fewer than a handfuldeal with the development of aggression. There appears to be a consensus thatthe effects will depend on the quality of the daycare environment rather thanon the presence or absence of daycare (Tremblay, Barr, & Peters, 2003). Someearly studies on children’s disruptive behavior concluded that those who spentseveral hours of the day away from their mothers during the first year of lifewere more disobedient and more aggressive toward their peers between 3 and8 years of age (Haskins, 1985; Rubenstein & Howes, 1983). These studieswere criticized because they often failed to take into account the quality of thedaycare environment and selection effects, that is, family and child character-istics before the beginning of daycare. For example, a study of a representativesample of 3,431 Canadian children 2–3 years of age (Borge, Rutter, Côté, &Tremblay, 2004) found a higher level of aggression among children who werenot sent to daycare groups, but were looked after by their parents. This rela-tionship emerged only among children in high-risk families; among those inlow-risk families, who constituted the majority (87%), no difference in thelevel of aggression was recorded. Moreover, within the subgroup of childrenattending group daycare, placement instability was associated with a higherlevel of aggression after controlling for a family risk index (occupational level,maternal education, number of siblings, and family functioning).
The U.S. National Institute of Child Health and Human Developmentcreated the NICHD Early Child Care Research Network (2002, 2003) totackle the shortcomings of previous studies. It regularly assessed 1,300 chil-
Social Capital and Physical Violence 405

dren and their families starting with the children at 6 months of age. Childrenwho spent more time in child care during the first years of life displayedhigher levels of externalizing problems and problems with aggression at 24months and 54 months, as well as in kindergarten and first grade. These re-sults certainly fueled the public debate. However, we will need much more in-formation before the debate is settled. First, the differences were statisticallysignificant, but more time in daycare did not increase the risk of being consid-ered in the clinical range; second, developmental trajectories of aggressionwere not studied to determine which type of children are more likely to beaffected. Are the affected children those who tend to be highly aggressive? Forexample, Raine, Reynolds, Venables, Mednick, and Farrington (1998) re-ported that 3-year-olds who had a larger body size were more likely to use ag-gression at 11 years of age. One can imagine that being with peers for longerperiods of time, and dominating them, makes it less likely that a child willlearn not to use physical aggression. Alternately, could the affected children bethose who normally tend to learn rapidly not to use physical aggression?These children may need to keep their level of aggression high enough to sur-vive in a tough peer environment. Developmental trajectories during the pre-school years, coupled with data from the elementary school years, should helpin understanding who is affected and whether these effects are long lasting. Athird caveat is that we are looking at a longitudinal study. We will need exper-iments to determine which types of preschool environments help childrenlearn alternatives to aggression.
Experimental designs are indeed needed to answer questions concerningcause and effect. A number of randomized experiments have attempted to testthe effect of early preventive interventions, carried out in the preschool years.The Perry preschool project, also known as the High Scope project, is proba-bly the best known of these experiments (Weikart, 1998; Weikart & Schwein-hart, 1992). The main aim of this program was to prevent school failureamong poor African American children by intervening when they were 3 and4 years old. Children attended quality day centers and teachers visited moth-ers at home. The long-term results revealed not only that the treated subjectswere more successful at school than the control subjects, but also that theywere arrested less often both as adolescents and as adults (at ages 19 and 24the treated subjects were five times less likely to have been arrested). Theycommitted less serious crimes and had lower rates of recidivism. Significantreductions in antisocial behavior were also found in the Syracuse UniversityFamily Development Project, and a reduction in aggressive behavior was ob-served in both the Yale Child Welfare Project and the Houston Parent ChildDevelopment Center (for a review of the results, see Tremblay & Craig,1995). Administered during the preschool years, all these programs increasesocial capital, in that they improve relationships between children and par-ents, and between parents and the community. Unfortunately, none of thesestudies appear to have specifically studied their subjects’ development of ag-gression.
406 D E T E R M I N A N T S O F A G G R E S S I O N

SCHOOL SOCIAL CAPITAL
Hagan and McCarthy (1997) have stressed that school is an important com-ponent of the social institutions network within which social capital is gener-ated. Indeed, in the light of the social capital paradigm, school is seen as acontext in which relationships that are important for the pupil’s adjustmentare developed—relationships that also involve teachers and parents.
A ready indication of the relationships between school social capital andpupil adjustment can be deduced from those studies that have correlated thesize of the school with the pupils’ behavior. Some studies revealed that stu-dents in larger schools experience a greater sense of alienation and frustrationthan those in small schools, and that the smaller schools are able to providetheir students with a better social environment. Moreover, it has been shownthat large schools report a far higher number of serious crimes and behaviorproblems (Ferris & West, 2002; National Center for Education Statistics,2002) than small schools; it may be supposed that the social relationshipswithin the smaller institutions are more closely knit, that the people knoweach other better and are more inclined to lend a hand in solving one another’sproblems.
Social capital at the school level particularly concerns the involvement ofparents in school activities. By using different samples of adolescents (in resi-dential treatment, ambulatory treatment, and untreated) Buysse (1997) foundthat a lack of social capital in the family and at school engendered a high riskof antisocial behavior on the part of the youth. Parcel and Dufur (2001) alsoanalyzed the influence of capital at home and at school and considered bothhuman and social capital. Using the National Longitudinal Survey of Youth(NLSY), to which they added indicators in the children’s schools (such as ameasure of school-wide parental involvement in school activities and scalesthat reflect school environment), they observed that family social capital andchild human capital had a large impact; but effects of school social capitalcould also be detected, and interactions between family and school social capi-tal were important as well.
In studying the relationships between school environment and the antiso-cial behavior of youth, Rutter, Maughan, Mortimore, Ouston, and Smitj(1979) identified a set of characteristics that apparently prevented deviant be-havior on the part of pupils. The school environment thus created was one inwhich teachers provided an example of good conduct, monitored and stimu-lated their pupils, and implemented firm and consistent disciplinary measures.More recently, Leblanc, Swisher, Vitaro, and Tremblay (2004) measured thesocial climate (discipline and academic pressure, in regard to teacher–pupilrelationships, and autonomy and job satisfaction, concerning relationshipswithin the teaching staff) of 217 high schools attended by 1,233 pupils whohad been followed since kindergarten. Students attending high schools whereteachers perceived fewer classroom behavior problems reported less physicallyaggressive antisocial behaviors. The analyses controlled for the students’ de-
Social Capital and Physical Violence 407

velopmental trajectory of aggression during elementary school, as well as forthe adverse family environment of the pupil and the sociodemographic factorsof the school, including school size.
Prevention experiments are, of course, better tools than correlationalstudies to test the causal effects of school social capital. Four experiments—the Seattle Social Development Project (Hawkins, Catalano, Kosterman,Abbott, & Hill, 1999), the Baltimore Prevention Study (Kellam, Rebok,Ialongo, & Mayer, 1994), the Montreal Longitudinal-Experimental Study(Tremblay, Kurtz, Mâsse, Vitaro, & Pihl, 1995), and the Fast Track preven-tion trial (Bierman et al., 2002)—measured the development of antisocial be-havior after attempting to improve relationships among pupils, parents andteachers, and the school environment in general. These four prevention exper-iments started in the first 2 years of elementary school and lasted between 2and 6 years. Both the Baltimore and Montreal experiments lasted 2 years andshowed a significant long-term reduction (4–8 years after the intervention) ofaggression in boys with the highest levels of physical aggression when theystarted school (Kellam et al., 1994; Lacourse, Nagin, Tremblay, Vitaro, &Claes, 2003). The Seattle experiment also showed a significant reduction of vi-olent delinquency during adolescence for boys and girls who participated inthe program from first to sixth grade, but not for those who participated onlyduring fifth and sixth grade. Fast Track is the latest of these four preventivetrials. Four years after the start of the intervention, the authors reported thatchildren in the control group, as compared with those in the interventiongroup, displayed less aggressive behavior, according to teacher and parent rat-ings (Bierman et al., 2002; Conduct Problems Prevention Research Group,2002). The results from these four experimental programs suggest that trajec-tories of physical aggression can be deflected by interventions that improverelationships between the pupil, his or her parents, peers, and teachers. Theyindicate, however, that the younger the children when the intervention starts,the more impact it will probably have.
PEER SOCIAL CAPITAL
Well-socialized peer groups are an important aspect of social capital—aresource that is available to children and may influence socialization in a posi-tive manner. As the child grows up, the peer group apparently becomes in-creasingly important, while the impact of the family environment declines. Inreference to the social capital paradigm, and therefore to resources in terms ofsocial relationships that can be utilized, we concentrate here on the effect ex-erted by the presence of prosocial peers, or the lack of this resource resultingfrom the phenomenon of peer rejection.
Various studies have reported that nondeviant friends are able to reduceinvolvement in antisocial behavior (Brown, Lohr, & McClenahan, 1986) andthat a prosocial group seems to lighten the impact of other risk factors
408 D E T E R M I N A N T S O F A G G R E S S I O N

(Quinton, Pickles, Maughan, & Rutter, 1993). It has also been observed thatpeer disapproval of delinquency reduces the likelihood that the individual willsubsequently commit violent crimes in general (Elliott, 1994), and sexualcrimes in particular (Ageton, 1983). With a sample of 868 boys selected fromkindergarten classrooms in poor areas of Montreal, Vitaro, Brendgen, andTremblay (2002) observed that, by early adolescence, those who were most in-fluenced by the aggressive–disruptive characteristics of their friends werethose who already had moderate behavior problems. These problems tendedto become milder if their friends were not aggressive–disruptive, but if theirfriends were aggressive–disruptive, then their own problems increased. It is in-teresting to note that boys who had serious problems, and boys who had noproblems, were not influenced by the characteristics of their friends. In a simi-lar study of 585 families in which there was a 5-year-old boy or girl, Criss,Pettit, Bates, Dodge, and Lapp (2002) found that peer acceptance and friend-ship acted as a moderator between the degree of family adversity and thechild’s externalizing behavioral problems.
In contrast, peer rejection can cause a child to miss out on a fundamentalexperience in the socialization process (Vitaro, Tremblay, & Bukowski,2001b) and can facilitate the emergence of adjustment problems and deviantbehavior. In a review of the literature on this subject, Parker and Asher (1987)concluded that there was a relationship between peer rejection and subsequentproblems of social adjustment and delinquency. However, they also raised thequestion of whether this rejection exerted an independent effect on the subse-quent behavioral disorders, or whether peer rejection was linked to the sub-ject’s aggressive behavior from the outset, and the subsequent disorders werechiefly linked to this early aggressiveness. Subsequent investigations enabledthis issue to be tackled and yielded some preliminary answers. On the basis ofbehavior observation, teacher and peer ratings, and peer interviews, Bierman,Smoot, and Aumiller (1993) subdivided a sample of 95 children examined atthree successive times (when they were 6–8, 8–10, and 10–12 years old) intofour subgroups: aggressive and rejected, aggressive and nonrejected, rejectedand nonaggressive, and neither aggressive nor rejected. Aggressive-rejectedchildren exhibited more diverse and severe conduct problems than aggressivechildren, in that the former displayed high levels of aggressiveness and disrup-tive behavior and a low level of prosocial activity, whereas the latter displayedonly a higher level of aggressive behavior. Moreover, 2 years later, the aggres-sive-rejected children continued to manifest a higher degree of aggressive anddisruptive behavior and a greater deficiency in prosocial activities than thechildren initially classified as aggressive-nonrejected. In a longitudinal study,which involved subjects from third grade to adolescence, Coie, Lochman,Terry, and Hyman (1992) observed that both aggressiveness and rejection, asevaluated in the third grade, independently predicted externalizing problemsin the sixth grade. The combination of aggressiveness and rejection predicted(only in males) acts of criminal assault and, in general, forms of antisocial be-havior that were more serious than those predicted by aggressiveness alone.
Social Capital and Physical Violence 409

Reactive aggression appears to be associated with peer rejection, whereas ac-tive aggression is enacted by children who are accepted by their peers (Dodge,Lochman, Harnish, Bates, & Pettit, 1997).
The effects of peer relationships on aggressive behavior have been alsodescribed in a relatively new field of research, concerning the consequences ofmutual antipathies (i.e., relationships characterized by reciprocal disliking) inchildhood and adolescence. In a study on 2,348 school-age children and 2,768adolescents, Abecassis et al. (2002) found that, controlling for peer rejection,same-sex antipathies were associated with aggressive behavior (perceived bypeers) and bullying others (self-reported), both in childhood and in adoles-cence, and to emotionality and lack of friendship support among adolescents;however, mixed-sex antipathies were related to antisocial and bullying behav-ior in boys but not in girls.
Finally, it should be noted that the influence of the peer group in reducingand controlling aggressive behavior can also be evaluated through experimen-tal interventions. Vitaro, Brendgen, Pagani, Tremblay, and McDuff (1999), forinstance, found that association with less deviant peers partly mediated the ef-fect on conduct disorders, assessed at the age of 13 years, of a multicomponentprevention program that targeted disruptive boys when they were ages 7–9. Ina subsequent study with the same sample, Vitaro, Brendgen, and Tremblay(2001a) showed that reduced disruptiveness and increased parental supervi-sion by age 11, as well as association with nondeviant peers by age 12, werepart of a chain of events that mediated the effect of the program on the initiallevel of delinquency at 13 years. Results of this kind have therefore confirmedthe notion that relationships with prosocial companions provide a form of so-cial capital that exerts a positive influence on the socialization process and onthe progressive control over disruptive behavior, including aggression.
NEIGHBORHOOD SOCIAL CAPITAL
At the beginning of the 1980s, Sampson and Groves (1989) used data from238 localities in Great Britain with a view to ascertaining whether some pa-rameters of community life, drawn from a national survey of 10,905 residents,were able to predict crime rates. They demonstrated that some features of thecommunity, which today would be identified as social capital (friendship net-works, participation in community activities, and supervision and control ofadolescent groups), were associated with lower crime rates, as measured boththrough self-reports and through victimization surveys. Sampson, Rauden-bush, and Earls (1997) subsequently utilized the concept of “collective effi-cacy” to interpret differences in the occurrence of violence in Chicago neigh-borhoods. Collective efficacy refers to social cohesion in a neighborhood,combined with the willingness of residents to act in favor of common inter-ests. It was measured by a scale of social control and a scale of social cohesionand trust. Controlling for race, age, and homicide rates in the previous years,
410 D E T E R M I N A N T S O F A G G R E S S I O N

collective efficacy at the neighborhood level correlated inversely with homi-cide, violence as perceived by citizens, and violence assessed through victim-ization surveys. Using the same indexes employed by Sampson et al. (1997),Browning (2002) found that collective efficacy was negatively associated withboth intimate homicide rates and nonlethal partner violence and that, further-more, in neighborhoods displaying higher levels of collective efficacy, womenwere more likely to disclose relationship conflicts to potential sources of sup-port.
In order to understand the origins of collective efficacy, Sampson et al.(1999) conducted a subsequent study in which they tried to identify, again atthe local community level, three aspects of social organization that influencechildren’s lives, development, and adjustment: intergenerational closure forchildren (when adults know the parents of their children’s friends, watch outfor the safety of children, etc.), reciprocal exchanges (the degree of interactionamong families regarding their children’s education and the swapping of ad-vice, material possessions, and information on childcare), and the degree of in-formal control combined with reciprocal support (expectations of actionwithin a community). They used the data from investigations conducted in1995 on a sample of 8,782 residents in 342 different Chicago neighborhoods.Residential stability and wealth were good predictors of the bond betweengenerations and of reciprocal exchanges. However, concentrated disadvantagecorrelated strongly with low expectations of shared social control over chil-dren. Spatial aspects proved to be very important, in that proximity to an areawith strong bonds between generations, frequent reciprocal exchanges, andgood informal control had a favorable impact above and beyond the struc-tural features of the neighborhood. Sampson et al. (1999) introduced these in-dicators on the grounds that some aspects of social capital, such as the exis-tence of groups, associations, and networks, might not, on their own, besufficient to favor child development and might even have a negative effect onsocial life if they operated in such a way as to exclude others. They thereforeclaimed that the objectives of local associations should be examined before de-ciding whether such groupings favor children’s collective needs.
Bellair (1997) used an interesting and innovative research method toinvestigate the effect of various types of social interaction on crime across 60urban neighborhoods. He found that getting together once a year or morewith neighbors had a strong negative association with burglary, motor vehicletheft, and robbery and that this form of interaction mediated a significant pro-portion of the effect of ecological characteristics on crime. Bellair suggestedthat even infrequent interaction and knowing one another could promptneighbors to engage in supervision and guardianship. This infrequent interac-tion corresponds to what Granovetter (1973) defined as “weak ties,” whichcan strengthen the community by creating linkages across networks. The pre-ventive effect of social bonds may not, however, be homogeneous. For in-stance, Warner and Rountree (1997) showed that strong social ties exerted apreventive effect in regard to assault rates in predominantly white neighbor-
Social Capital and Physical Violence 411

hoods, whereas in predominantly minority and mixed-race neighborhoodssuch ties were largely ineffective. A particular form of violent behavior, childmaltreatment, has been seen to be associated, at the neighborhood level, withthe lack of social resources, when income levels are equal (Vinson, Baldry, &Hargreaves, 1996).
Dubet (1987), through ethnographic studies, identified another relation-ship between violent behavior and scant social bonds. He ascribed the out-breaks of youth violence seen in recent years in the suburbs of some Frenchcities to the disappearance of the traditional working-class culture. In working-class neighborhoods there used to be a whole range of meeting places andspare-time activities and groups, which provided the young with role modelsand common objectives. In traditional working-class neighborhoods the fac-tory provided a nucleus for aggregation and socialization; children and ado-lescents therefore grew up within a network of relationships and expectationsthat fostered social and occupational development and provided models thatwere endorsed by the whole community. Although this setting also featuredsocial conflicts, the community was somewhat close-knit and rich in socialrelationships. De-industrialization and suburban transformation have demol-ished this social fabric; commerce is rare, meeting places nonexistent, build-ings dilapidated, and transport inadequate. What has emerged is a sort ofno-man’s-land, where the young try to survive, drug pushing abounds, crimeand violence are endemic, and a deep sense of insecurity prevails. In the lightof the social capital theories, what has disappeared from these neighborhoodsis a set of social networks, shared expectations, and community commitment.These are so essential to the social life and socialization of the young that theirdisappearance has facilitated the outbreaks of violence that have afflictedmany urban districts in Europe.
TOWN AND REGION
The relationship between regional social capital and violent crimes has re-cently been examined in a series of studies in the United States. Using datafrom the 50 U.S. states, Kennedy, Kawachi, Prothrow-Stith, Lochne, andGupta (1998) measured social capital by membership of groups and associa-tions and social trust, two items from the U.S. General Social Survey. Theyfound a correlation between a low level of social capital and marked inequal-ity on one hand, and violent crimes on the other. In a cross-sectional analysisthat also included the 50 U.S. states, Putnam (2000) found that an index ofsocial capital was inversely correlated with homicide rates and that this corre-lation was very high (r = –.80) for the period 1980–1995. Controlling for thevariable “fear of crime,” the (partial) correlation between social capital andhomicide remained high (r = –.53). Putnam concluded that the direction of therelationship could not possibly be from homicide to social capital, becauseany such relationship would have been mediated by the fear of crime. To ob-
412 D E T E R M I N A N T S O F A G G R E S S I O N

tain a clearer picture of the relationship that had emerged, he introduced arange of social and economic factors as independent variables in a multiple re-gression. He found that the model best able to predict homicide rates in thevarious states included four variables: social capital, mean poverty level, eth-nic makeup of the population (percentage of nonwhites), and the proportionof the population living in cities. In a similar study, Hemenway, Kennedy,Kawachi, and Putnam (2001) measured, at the state level, the degree of inter-personal trust (through the replies provided to the U.S. General Social Survey)and social commitment (assessed through participation in voluntary activities,religious observance, etc.). After controlling for the level of urbanization, pov-erty, and the percentage of nonwhite residents, they concluded that these indi-cators of social capital were inversely associated with the percentages of homi-cides and suicides committed with firearms.
Focusing their analysis on a more restricted territorial area, and thus in-creasing the sample size, Rosenfeld, Messner, and Baumer (2001) investigatedthe relationships between social capital and homicide in 99 geographical areasof the United States. They measured two aspects of social capital, civic com-mitment and social trust: The former was measured through voter turnoutand membership in voluntary associations, and the latter through respon-dents’ replies to the U.S. General Social Surveys of 1993, 1994, and 1996. Alow level of social capital was found to be associated with a high homiciderate, after controlling for a range of socioeconomic variables. Social capitalmediated the effect of the area’s being located in the southern states, but it didnot mediate the effect of economic deprivation. The statistical relationship be-tween social capital and homicide was not due to the influence of homicide onsocial capital. Subsequently, Messner, Baumer, and Rosenfeld (2002) broad-ened this focus to include robbery and assault, demonstrating that thesecrimes were also inversely related to social capital.
In one of the rare studies on social capital and violence outside the UnitedStates, we used Italian data and calculated an index of “civicness” (Putnam,1993) for each of the 20 administrative regions (Gatti, Tremblay, & Larocque,2003) and 95 provinces (Gatti, Tremblay, & Schadee, 2002b). The civicnessindex included three indicators: (1) voter turnouts in referendums or elections,(2) the percentage of citizens over the age of 13 years who read a newspaperevery day, and (3) the number of recreational, sporting, and cultural associa-tions per 100,000 inhabitants. The results revealed a negative correlationbetween civicness and the reported number of some types of serious crimes ofviolence, as recorded in the 1970s, 1980s, and 1990s, at the regional level.Data from the vast victimization survey conducted by the Istituto Nazionale diStatistica/National Institute of Statistics (ISTAT) confirmed the negative asso-ciation between civic sense and violent crimes. At the provincial level, multipleregression analyses were carried out, using as control variables unemploy-ment, family breakdown, and urbanization as predictors of homicide, in addi-tion to civic sense. Notwithstanding the introduction of these control vari-ables, civic sense continued to be negatively associated with homicide;
Social Capital and Physical Violence 413

however, the interactions among variables revealed that the protective effectof civic sense was exerted only in the southern provinces, where homiciderates are higher. The criminogenic effect of urbanization and unemploymentwas also observed only in the southern provinces, whereas family breakdownwas associated with homicide in the whole of Italy. Finally, the preventive ef-fect of civicness in regard to robbery was examined. This analysis revealedthat civic sense has a preventive effect on this type of crime, but only in themore urbanized provinces, where robbery rates are higher.
NEGATIVE SOCIAL CAPITAL
Although the majority of authors stress the positive aspects of social capital,some also underline its possible negative consequences (e.g., Portes, 1998). In-deed, social networks and the norms of reciprocal cooperation may be used toexclude those who do not belong to the group from access to resources (seeVaillancourt, Chapter 8, this volume). Community bonds may constitute apervasive social control tool, which hampers autonomy and privacy, andmembers of marginal minority groups may be tied down by bonds of solidar-ity that make it difficult for them to accept the dominant social norms and togain access to the world of traditional work. Finally, strong social bonds maybe used for patently illegal purposes within the sphere of organized crime oryouth gangs (Portes, 1998).
In regard to children’s development, the fact that youth gangs may consti-tute an environment in which aggressive behavior is facilitated has been welldocumented, especially for males. Numerous studies have revealed that youthswho belong to a gang commit more crimes than those who do not, and thatthis difference is greater with respect to serious crimes and crimes of violence.Longitudinal studies are especially suited to test three models, which Thorn-berry, Krohn, Lizotte, Smith, and Tobin (2002) have defined as the selectionmodel, the socialization model, and the enhancement, or mixed, model. Ac-cording to the selection model, the high level of delinquency and violenceamong gang members does not depend on the influence of the gang, but ratheris due to the fact that gangs attract and select those youths who already com-mit the most crimes. In contrast, the social facilitation model holds that it isthe gang that facilitates and triggers deviant behavior in youths who, beforejoining the gang, are no different from other young people. The enhancementmodel combines these two views: Youths who join gangs already display ahigher level of delinquency, and joining the gang exacerbates their deviant be-havior.
Using data from the Rochester Youth Development Study, Thornberry,Krohn, Lizotte, and Chard-Wierschem (1993) showed that, before joining agang, members did not display higher rates of delinquency or drug use thannonmembers. However, such behaviors increased upon entry into the gang,and delinquency declined once an individual left the gang (Thornberry et al.,
414 D E T E R M I N A N T S O F A G G R E S S I O N

1993). In a subsequent study, covering a longer period of time, Thornberry etal. (2002) found that, in addition to a strong facilitation effect, a weaker selec-tion effect was also present. Strong evidence for the social facilitation modeland partial support for selection was provided by Zhang, Welte, and Wiec-zorek (1999) through their analysis of the data from the Buffalo LongitudinalSurvey of Young Men. The enhancement model was confirmed by the DenverYouth Survey, a longitudinal study of families in high-risk neighborhoods(Esbensen & Huizinga, 1993), by a longitudinal study of schools in six Ameri-can cities (Esbensen, Peterson, Freng, & Taylor, 2002), and by the MontrealLongitudinal Experimental Study (Lacourse et al., 2003; Vitaro, Tremblay,Kerr, Pagani, & Bukowski, 1997). The latter study also revealed that thecriminogenic influence of the gang was independent of other risk factors, suchas previous delinquency, disruptiveness, and lack of parental supervision; gangmembership emerged as a significant predictor of the frequency of aggressivebehavior, above and beyond the effects of having delinquent friends (Gatti,Tremblay, Vitaro, & McDuff, in press). Data from the Seattle Social Develop-ment Project revealed that respondents who were gang members always hadthe highest frequency of delinquent behavior and drug use, and that compar-ing gang members, nongang youths who had delinquent friends, and nongangyouths who did not have delinquent friends, gang membership predicted self-reported and officially recorded delinquency beyond the effects of having de-linquent friends and prior delinquency (Battin-Pearson, Thornberry, Hawkins, &Krohn, 1998). The results from these studies demonstrate that the crimino-genic effect is not due simply to contact with other delinquent youths, but thatit is also linked to the influence of the gang itself.
Some ethnographic studies have revealed that the gang may constitute animportant form of social capital for youths—a resource that they can exploitin order to achieve their own objectives. Jankowsky (1991) obtained participantobservations over a 10-year period among 37 gangs in the metropolitan areasof Los Angeles, New York, and Boston. He concluded that in the Americansuburbs, gangs are not only an expression of social disorganization, but theyare also an organizational response aimed at enhancing the competitiveness oftheir members in acquiring scarce resources. Indeed, gang members enjoy cer-tain advantages in terms of money, status, and power, and the gang acceptsyouths as members if they are able to contribute to its prestige and efficiency.
Criminal organizations and subcultures may also be regarded as a formof social capital that facilitates violence. In particular, violent subcultures,which were first investigated by Wolfgang and Ferracuti (1967) and subse-quently analyzed by numerous criminologists, may, like the army or a boxingcoach, train the individual in using physical aggression. According to Wolf-gang and Ferracuti, a subculture is “a normative system of some sub-groupsmaller than the whole society,” which implies a process of learning sharedvalues. There are subcultures that value the use of physical violence, andcertain members (e.g., adult males) are required to use violence in certain cir-cumstances. In such situations, the individual is expected to act violently, and
Social Capital and Physical Violence 415

nonviolence may lead to ostracism or condemnation; in contrast, violence isrewarded rather than condemned and does not arouse guilt feelings in the in-dividual.
There are a few published biographies that shed light on the relationshipsbetween criminal organizations, subcultures, and aggressive behavior. A biog-raphy of Raffaele Cutolo (Marrazzo, 1984), a Neapolitan crime boss whoreigned over the Campania region for many years, and who is now in prison,provides us with the following illuminating episode. Raffaele’s father was anagricultural worker, who for years tilled a field as a sharecropper to supporthis family. One day, when Raffaele was still a child, the owner of the land toldhis father that the following year the field would be used for a different pur-pose, and that his services would therefore no longer be required. In despera-tion, Raffaele’s father turned to the local Mafia boss, whose word was law inthe village. The Mafia boss invited the Cutolo family to his home, and afterMass one Sunday, dressed in their best clothes, both parents and children re-spectfully presented themselves at the boss’s door. Raffaele’s father was ush-ered into another room by the boss, while the lady of the house offered cakesto Mrs. Cutolo and her children. When the two men reemerged, Mrs. Cutoloanxiously asked her husband if there was any news. She was told that every-thing had been settled. The following day, the landowner informed Mr.Cutolo that he had changed his mind and that his contract would be renewed.
It is easy to imagine the psychological impact that this event must havehad on all the members of the family, and especially on the young Raffaele,who had seen this family crisis solved by appealing to a local institution thatwas both powerful and efficient, although illegal. A few years later, however,when Raffaele was an adolescent, he was summoned by the Mafia boss, whoinstructed him to deliver a pistol secretly to a mafioso in another village.Other requests followed, and finally Raffaele Cutolo found himself involvedin serious crimes. From there, he went on to become the head of a powerfulorganized crime syndicate. It is clear from this account that the criminal fra-ternity constituted a form of social capital—a resource to be exploited in or-der to solve problems and achieve otherwise unreachable goals. But this socialcapital was also permeated by norms of reciprocity and expectations centeredon the use of violence, which ended up conditioning the social development ofthose living within this culture.
Some systematic studies have also evaluated the importance of violentsubcultures. Felson, Liska, South, and McNulty (1994), for example, studied2,123 boys in 87 public high schools. Respondents were asked a series ofquestions designed to measure whether they approved of nonaggressive re-sponses to some types of provocation. The questions consisted of a series ofpersonal values, and respondents were asked whether certain acts were “goodthing[s] for people to do.” Disagreement with these values was deemed to in-dicate a high score on the subculture of violence scale. The authors reachedthe conclusion that group-level values predicted violent behavior, regardless ofindividual-level values. Furthermore, this research demonstrated that subcul-
416 D E T E R M I N A N T S O F A G G R E S S I O N

ture differences in values were more frequently encountered within smallgroups involving interactions between their members, rather than withinbroad aggregations based on race, social class, or religion. Such examples il-lustrate the fact that social capital may also exist in illegal forms and that, insome cases, crime and violence constitute a shared resource that can be ex-ploited in order to achieve ambitions or, in certain circumstances, simply tosurvive.
CONCLUSIONS
We have seen that social capital can be defined in various ways and may oper-ate in more or less broad contexts and at different ages from early childhoodto adulthood. From the developmental standpoint, several empirical studieshave demonstrated that both familial and extrafamilial social capital have aneffect, even in the long term, on children’s adjustment, on academic resultsand socioeconomic success (Furstenberg & Hughes, 1995), and on health andwell-being (Morrow, 1999). In general, the social capital perspective hasproven useful in helping us to understand aggressive behavior and violence,though much more research is needed. Not all forms of social capital act inthe same way, and several studies have found that social capital seems to exerta preventive effect on aggressive behavior only in certain circumstances andonly in regard to some types of persons.
We have seen that the development of children and young people canbe influenced by the quality of relationships within the family, at school,and among peers, and by the characteristics of the communities and re-gional environments in which they live. For the most part, each of these lev-els of influence has been studied separately, though some researchers havebegun to consider different aspects of social capital simultaneously. This ap-pears to be an important advancement, in view of the multidimensional na-ture of social capital and its potential to link “the overly narrow purview ofpsychology and the overly broad purview of sociology” (Furstenberg andHughes, 1995).
In addition to its multidimensional character, the developmental aspect ofsocial capital should also be considered, by examining how the loss or acquisi-tion of a given form of social capital during the course of the individual’s lifemight influence engagement in aggressive behavior. The various types of socialcapital have a differential impact according to the age of the individual. Fam-ily social capital and the relationship resources offered by the child welfaresystem chiefly act during early childhood to middle childhood, when learningto regulate physical aggression appears to be at its peak. The influence ofpeers has been shown mainly during late childhood and adolescence; however,it is most likely present very early at home, through siblings, and in daycare.Violent subcultures and organized crime are more likely to exert a direct effecton adolescents and adults. Broadly speaking, it seems that social capital at the
Social Capital and Physical Violence 417

microlevel plays a stronger role during childhood, whereas the macrolevel actsespecially during adolescence and adult life.
Thus, the influence of social capital on the regulation of aggressive behav-ior should be studied from a developmental perspective. It can be claimed thatthe process of progressively waning physical aggression from early childhoodto adulthood (Sampson & Laub, 2003; Tremblay & Nagin, Chapter 5, thisvolume) is facilitated by the majority of the social relationships we have de-fined as social capital. All cultures, including nonhuman cultures, benefit fromhelping their young ones to control their ability to physically aggress others.This does not mean that they completely inhibit the power to physically ag-gress. Armies are, or should, always be ready for war. Individuals and sub-groups are at an advantage if they can defend themselves from an aggression.Thus, social capital that is advantageous for one group can be disadvanta-geous for another. This is why gangs and organized crime are consideredthreats to the mainstream, but constitute a resource for individuals who, fordifferent reasons, are not or have chosen not to be part of this mainstream.
In general, attempts should be made to improve social support (Colvin,Cullen, & Vander Ven, 2002) at all levels—through the family, welfare, andacademic institutions, formal and informal groups, neighborhoods and cities—to strengthen social networks, to increase community participation and socialcommitment, and to reduce individualism in favor of a community spirit.These objectives should be pursued both through specific intervention, for ex-ample, by providing support for mothers in difficulty, and through a broadercultural evolution toward greater cooperation and solidarity. Research on theeffects of these interventions should help us to understand which are the mosteffective strategies for a given culture, while testing our theories concerningthe origins and development of aggressive behavior. We will, of course, needto study to what extent different types of social capital have different effectson different types of aggressive behavior, at different periods of the life course.
REFERENCES
Abecassis, M., Hartup, W. W., Haselager, G. J. T., Ron, H. J., Scholte, R. H. J., & VanLieshout, C. F. M. (2002). Mutual antipathies and their significance in middle child-hood and adolescence. Child Development, 73(5), 1543–1556.
Ageton, S. S. (1983). Sexual assault among adolescents. Lexington, MA: LexingtonBooks.
Battin-Pearson, S. R., Thornberry, T. P., Hawkins, J. D., & Krohn, M. D. (1998). Gangmembership, delinquent peers, and delinquent behavior. Bulletin No. NCJ 171119.Youth Gang Series. Washington, DC: U.S. Department of Justice, Office of JusticePrograms, Office of Juvenile Justice and Delinquency Prevention.
Bellair, P. E. (1997). Social interaction and community crime: Examining the importanceof neighbor networks. Criminology, 35(4), 677–703.
Belsky, J. (2001). Developmental risks (still) associated with early child care. Journal ofChild Psychology and Psychiatry, 42, 845–860.
418 D E T E R M I N A N T S O F A G G R E S S I O N

Bierman, K. L., Coie, J. D., Dodge, K. A., Greenberg, M. T., Lochman, J. E., McMahon,R. J., & Pinderhughes, E. E. (2002). Using the Fast Track randomized preventiontrial to test the early-starter model of the development of serious conduct problems.Development and Psychopathology, 14(4), 925–943.
Bierman, K. L., Smoot, D. L., & Aumiller, K. (1993). Characteristics of aggressive-re-jected, aggressive (non-rejected) and rejected (non-aggressive) boys. Child Develop-ment, 64, 139–151.
Borge, A., Rutter, M., Côté, S., & Tremblay, R. E. (2004). Early childcare and physicalaggression: Differentiating social selection and social causation. Journal of ChildPsychology and Psychiatry, 45(2), 367–376.
Bourdieu, P. (1980). Le capital social: Notes provisoires, Actes de la Recherche en Sci-ences Sociales, 31, 2–3.
Brown, B. B., Lohr, M. J., & McClenahan, E. L. (1986). Early adolescents’ perceptions ofpeer pressure. Journal of Early Adolescence, 6, 139–154.
Browning, C. R. (2002). The span of collective efficacy: Extending social disorganizationtheory to partner violence. Journal of Marriage and Family, 64(4), 833–850.
Buysse, W. H. (1997). Behaviour problems and relationship with family and peers duringadolescence. Journal of Adolescence, 20(6), 645–659.
Cancian, M. (2001.) Rhetoric and reality of work-based welfare reform. Social Work,46(4), 309–314.
Coie, J. D., Lochman, J. E., Terry, R., & Hyman, C. (1992). Predicting early adolescentdisorder from childhood aggression and peer rejection. Journal of Consulting andClinical Psychology, 64, 53–63.
Coleman, J. S. (1990). Foundation of social theory. Cambridge, MA: Harvard UniversityPress.
Colvin, M., Cullen, F. T., & Vander Ven, T. (2002). Coercion, social support, and crime:An emerging theoretical consensus. Criminology, 40(1), 19–42.
Conduct Problems Prevention Research Group. (2002). Evaluation of the first 3 years ofthe Fast Track prevention trial with children at high risk for adolescent conductproblems. Journal of Abnormal Child Psychology, 30, 19–35.
Criss, M. M., Pettit, G. S., Bates, J. E., Dodge, K. A., & Lapp, A. L. (2002). Family adver-sity, positive peer relationship, and children’s externalizing behavior: A longitudinalperspective on risk and resilience. Child Development, 73(4), 1220–1237.
DeFronzo, J. (1983). Economic assistance to impoverished Americans. Relationship toincidence of crime. Criminology, 21(1), 119–136.
DeFronzo, J. (1997). Welfare and homicide. Journal of Research on Crime and Delin-quency, 34(3), 395–406.
Dodge, K. A., Lochman, J. E., Harnish, J. D., Bates, J. E., & Pettit, G. S. (1997). Reactiveand proactive aggression in school children and psychiatrically impaired chroni-cally assaultive youth. Journal of Abnormal Psychology, 106, 37–51.
Dubet, F. (1987). La galère: Les jeunes en survie. Paris: Fayard.The Economist. (2001, August 25). Welfare reform: America’s great achievement. The
Economist, 25–27.Elliott, D. S. (1994). Serious violent offenders: Onset, developmental course, and termi-
nation—The American Society of Criminology 1993 presidential address. Crimi-nology, 32, 1–21.
Esbensen, F., & Huizinga, D. (1993). Gangs, drugs, and delinquency in a survey of urbanyouth. Criminology, 31, 565–589.
Esbensen, F., Peterson, D., Freng, A., & Taylor, T. J. (2002). Initiation of drug use, drug
Social Capital and Physical Violence 419

sales, and violent offending among a sample of gang and non-gang youth. In C.R.Huff (Ed.), Gangs in America (3rd ed., pp. 37–50). Beverly Hills, CA: Sage.
Farrington, D. P. (1989). Early predictors of adolescent aggression and adult violence. Vi-olence and Victims, 4, 79–100.
Felson, R. B., Liska, A. E., South, S. J., & McNulty, T. L. (1994). The subculture of vio-lence and delinquency: Individual vs. school context effects. Social Forces, 73(1),155–173.
Ferris, J. S., & West, E. G. (2002). Economies of scale, school violence, and the optimalsize of schools. Carleton Economic Papers, No. 02–01. Ottawa, QC: Carleton Uni-versity.
Fukuyama, F. (1995). Trust: The social virtues and the creation of prosperity. New York:Free Press.
Furstenberg, F. F. Jr., & Hughes, M. E. (1995). Social capital and successful developmentamong at-risk youth. Journal of Marriage and the Family, 57(3), 580–592.
Gatti, U., Schadee, M. A. H., & Tremblay, R. E. (2002a, September). Lagged effects ofnurturance on crime in Italian provinces. Paper presented at the Second Conferenceof the European Society of Criminology, Toledo, Spain.
Gatti, U., Tremblay, R. E., & Larocque, D. (2003). Civic community and juvenile delin-quency: A study of the regions of Italy. British Journal of Criminology, 43, 22–40.
Gatti, U., Tremblay, R. E., & Schadee, H. (2002b, July). Social capital and violent crimein Italian provinces. Paper presented at the 15th World Meeting of the InternationalSociety for Research on Aggression, Montreal, Canada.
Gatti, U., Tremblay, R. E., Vitaro, F., & McDuff, P. (in press). Youth gangs, delinquencyand drug use: A test of the selection, facilitation, and enhancement hypotheses.Journal of Child Psychology and Psychiatry.
Granovetter, M. (1973). The strength of weak ties. American Journal of Sociology, 78,1370–1380.
Hagan, J., & McCarthy, B. (1997). Mean streets: Youth crime and homelessness. Cam-bridge, MA: Cambridge University Press.
Han, W., Waldfogel, J., & Brooks-Gunn, J. (2001) The effects of early maternal employ-ment on later cognitive and behavioral outcomes. Journal of Marriage and Family,63(2), 336–354.
Haskins, R. (1985). Public school aggression among children with varying day-care ex-perience. Child Development, 56, 689–703.
Hawkins, J. D., Catalano, R. F., Kosterman, R., Abbott, R., & Hill, K. G. (1999). Pre-venting adolescent health-risk behaviors by strengthening protection during child-hood. Archives of Pediatrics and Adolescent Medicine, 153, 226–234.
Hemenway, D., Kennedy, B. P., Kawachi I., & Putnam, R. D. (2001). Firearm prevalenceand social capital. Annals of Epidemiology, 11, 484–490.
Henry, B., Caspi, A., Moffit, T. E., & Silva, P. A. (1996). Temperamental and familial pre-dictors of violent and nonviolent criminal convictions: Age 3 to age 18. Develop-mental Psychology, 32, 614–623.
Hobbes, T. (1647/1998). De Cive. New York: Cambridge University Press.Jankowsky, M. S. (1991). Islands in the street: Gangs in American urban society. Berke-
ley, CA: University of California Press.Karen, R. (1994). Becoming attached. New York: Warner Books.Keenan, K., & Shaw, D. S. (1994). The development of aggression in toddlers: A study of
low-income families. Journal of Abnormal Child Psychology, 22(1), 53–77.Kellam, S. G., Rebok, G. W., Ialongo, N., & Mayer, L. S. (1994). The course and mallea-
420 D E T E R M I N A N T S O F A G G R E S S I O N

bility of aggressive behavior from early first grade into middle school: Results of adevelopmental epidemiologically-based preventive trial. Journal of Child Psychol-ogy and Psychiatry, 35, 259–281.
Kennedy, B. P., Kawachi, I., Prothrow-Stith, D., Lochne, K., & Gupta, V. (1998). Socialcapital, income inequality, and firearm violent crime. Social Science and Medicine,1, 7–17.
Ku, I. (2001). The effect of welfare on children’s education. Social Service Review, 75(2),245–270.
Lacourse, E., Nagin, D. S., Tremblay, R. E., Vitaro, F., & Claes, M. (2003). Developmen-tal trajectories of boys’ delinquent group membership and facilitation of violent be-haviors during adolescence. Development and Psychopathology, 15, 183–197.
LeBlanc, L., Swisher, R., Vitaro, F., & Tremblay, R. E. (2004). Antisocial behavior andhigh school social climate: A 10 year longitudinal and multilevel study. Manuscriptsubmitted for publication.
Lin, N. (1999). Social networks and status attainment. Annual Review of Sociology, 25,467–487.
Marrazzo, G. (1984). Il camorrista. Napoli: Tullio Pironti.McCord, J. (2002, July). Social capital, family socialization, and aggressive behavior. Pa-
per presented at the 15th World Meeting of the International Society for Researchon Aggression, Montreal, Canada.
Messner, S. F., Baumer, E., & Rosenfeld, R. (2002, July). Social capital and violent crimerates across U. S. communities. Paper presented at the 15th World Meeting of the In-ternational Society for Research on Aggression, Montreal, Canada.
Messner, S. F., & Rosenfeld, R. (1997). Political restraint of the market and levels of crim-inal homicide: A cross-national application of institutional-anomie theory. SocialForces, 75(4), 1393–1416.
Morris, P. (2002). The effect of welfare reform policies on children. Social Policy Report,16(1), 4–19.
Morrow, V. (1999). Conceptualizing social capital in relation to the well-being of chil-dren and young people: A critical review. The Sociological Review, 47(4), 744–765.
Nagin, D., & Tremblay, R. E. (2001). Parental and early childhood predictors of persis-tent physical aggression in boys from kindergarten to high school. Archives of Gen-eral Psychiatry, 58, 389–394.
National Center for Education Statistics. (2002). Digest of education statistics. Washing-ton DC: U.S. Department of Education.
NICHD Early Child Care Research Network. (2002). Child-care structure → process →outcome: Direct and indirect effects of child-care quality on young children’s devel-opment. Psychological Science,13(3), 199–206.
NICHD Early Child Care Research Network. (2003). Does amount of time in child carepredict socioemotional adjustment? Child Development, 74(4), 976–1005.
Olds, D. L., Eckenrode, J., Henderson, C. R., Jr., Kitzman, H., Powers, J., Cole, R.,Sidora, K., Morris, P., Pettitt, L. M., & Luckey, D. (1997). Long-term effects ofhome visitation on maternal life course, and child abuse and neglect. Journal of theAmerican Medical Association, 278, 637–643.
Pagani, L., Boulerice, B., & Tremblay, R. E. (1997). The influence of poverty upon chil-dren’s classroom placement and behavior problems during elementary school: Achange model approach. In G. Duncan & J. Brooks-Gunn (Eds.), Consequences ofgrowing up poor (pp. 311–339). New York: Sage.
Social Capital and Physical Violence 421

Pampel, F., & Gartner, R. (1995). Age-structure, socio-political institutions, and nationalhomicide rates. European Sociological Review, 11, 243–260.
Parcel, T. L., & Dufur, M. J. (2001). Capital at home and at school: Effects on child socialadjustment. Journal of Marriage and Family, 63(1), 32–47.
Parker, J. G., & Asher, S. R. (1987). Peer relations and later personal adjustment: Arelow-accepted children at risk? Psychological Bulletin, 102, 357–389.
Paxon, C., & Waldfogel, J. (2001, November). Welfare reforms, family resources andchild maltreatment. Paper presented at the 23rd annual research conference of theAssociation for Public Policy Analysis and Management, Washington, DC.
Portes, A. (1998). Social capital: Its origins and applications in modern society. AnnualReview of Sociology, 24, 1–24.
Putnam, R. D. (1993). Making democracy work: Civic tradition in modern Italy. Prince-ton, NJ: Princeton University Press.
Putnam, R. D. (2000). Bowling alone. The collapse and revival of American community.New York: Simon & Schuster.
Quinton, D., Pickles, A., Maughan, B., & Rutter, M. (1993). Partners, peers and path-ways: Assortative pairing and continuities in conduct disorder. Development andPsychopathology, 5, 763–783.
Raine, A., Reynolds, C., Venables, P. H., Mednick, S. A., & Farrington, D. P. (1998).Fearlessness, stimulation-seeking, and large body size at age 3 years as early predis-positions to childhood aggression at age 11 years. Archives of General Psychiatry,55(8), 745–751.
Rosenfeld, R., Messner, S. F., & Baumer, E. P. (2001). Social capital and homicide. SocialForces, 80(1), 283–310.
Rousseau, J. J. (1762/1986). Émile (B. Foxley, Trans.). London: Dent.Rubenstein, J., & Howes, C. (1983). Social-emotional development of toddlers in day
care: The role of peers and individual differences. In S. Kilmer (Ed.), Advances inearly education and day care (Vol. 3, pp. 13–45). Greenwich, CT: JAI Press.
Rutter, M., Giller, H., & Hagell, A. (1998). Antisocial behavior by young people. NewYork: Cambridge University Press.
Rutter, M., Maughan, B., Mortimore, P., Ouston, J., & Smitj, A. (1979). Fifteen thousandhours: Secondary schools and their effects on children. Cambridge, MA: HarvardUniversity Press.
Sampson, R. J., & Groves, B. W. (1989). Community structure and crime: Testing social-disorganization theory. American Journal of Sociology, 94, 774–802.
Sampson, R. J., & Laub, J. H. (2003). Life-course desisters? Trajectories of crime amongdelinquent boys followed to age 70. Criminology, 41(3), 555–592.
Sampson, R. J., Morenoff, J. D., & Earls, F. (1999). Beyond social capital: Spatial dynam-ics of collective efficacy for children. American Sociological Review, 54(5), 633–660.
Sampson, R. J., Raudenbush, S. W., & Earls, F. (1997). Neighborhood and violent crime:A multilevel study of collective efficacy. Science, 277, 918–924.
Savage, J., & Vila, B. (1997). Lagged effects of nurturance on crime: A cross-nationalcomparison. Studies on Crime and Crime Prevention, 6(1), 101–120.
Savage, J., & Vila, B. (2002). Changes in child welfare and subsequent crime rate trends:A cross-national test of the lagged nurturance hypothesis. Applied DevelopmentalPsychology, 23, 51–82.
Savolaien, J. (2000). Inequality, welfare state, and homicide: Further support for the in-stitutional anomie theory. Criminology, 38(4), 1021–1039.
422 D E T E R M I N A N T S O F A G G R E S S I O N

Smith, C. A., & Farrington, D. P. (2003). Continuities in antisocial behavior andparenting across three generations. Journal of Child Psychology and Psychiatry, 44,1–18.
Thornberry, T. P., Krohn, M. D., Lizotte, A. J., & Chard-Wierschem, D. (1993). The roleof juvenile gangs in facilitating delinquent behavior. Journal of Research in Crimeand Delinquency, 30, 55–87.
Thornberry, T. P., Krohn, M. D., Lizotte, A. D., Smith, C. A., & Tobin, K. (2002). Gangsand delinquency in developmental perspective. Cambridge, UK: Cambridge Univer-sity Press.
Tremblay, R. E., Barr, R. G., Peters, R. D. (Eds.). (2003). Encyclopedia on Early Child-hood Development [Online]. Montreal, QC: Centre of Excellence for Early Child-hood Development. Available at http://www.excellence-earlychildhood.ca/docu-ments/TremblayANGxp.pdf.
Tremblay, R. E., & Craig, W. (1995). Developmental crime prevention. In M. Tonry & D.Farrington (Eds.), Building a safer society: Strategic approach to crime prevention.Chicago: The University of Chicago Press.
Tremblay, R. E., Kurtz, L., Mâsse, L. C., Vitaro, F., & Pihl, R. O. (1995). A bimodal pre-ventive intervention for disruptive kindergarten boys: Its impact through mid-ado-lescence. Journal of Consulting and Clinical Psychology, 63(4), 560–568.
Tremblay, R. E., Nagin, D. S., Séguin, J. R., Zoccolillo, M., Zelazo, P., Boivin, M.,Pérusse, D., & Japel, C. (2004). Physical aggression during early childhood: Trajec-tories and predictors. Pediatrics, 114, e43–e50.
Vaden-Kiernan, N., Ialongo, N. S., Pearson, J., & Kellam, S. (1995). Household familystructure and children’s aggressive behavior: A longitudinal study of urban elemen-tary school children. Journal of Abnormal Child Psychology, 23(5), 553–568.
Vander Ven, T. M., Cullen, F. T., Carrozza, M. A., & Wright, J. P. (2001). Home alone:The impact of maternal employment on delinquency. Social Problems, 48(2), 236–257.
Vila, B. (1994). A general paradigm for understanding criminal behavior: Extending evo-lutionary ecological theory. Criminology, 32, 501–549.
Vinson, T., Baldry, B., & Hargreaves, J. (1996). Neighbourhoods, networks and childabuse. British Journal of Social Work, 26, 523–542.
Vitaro, F., Brendgen, M., Pagani, L., Tremblay, R. E., & McDuff, P. (1999). Disrup-tive behavior, peer association, and conduct disorder: Testing the developmentallinks through early intervention. Development and Psychopathology, 11, 287–304.
Vitaro, F., Brendgen, M., & Tremblay, R. E (2001a). Preventive intervention: Assessingits effects on the trajectories of delinquency and testing for mediational processes.Applied Developmental Science, 5(4), 201–213.
Vitaro, F., Brendgen, M., & Tremblay, R. E (2002). Reactively and proactively aggressivechildren: Antecedent and subsequent characteristics. Journal of Child Psychologyand Psychiatry, 43(4), 495–505.
Vitaro, F., Tremblay, R. E., & Bukowski, W. M. (2001b). Friends, friendship and conductdisorders. In J. Hill & B. Maughan (Eds.), Conduct disorders in childhood and ado-lescence (pp. 346–378). Cambridge, UK: Cambridge University Press.
Vitaro, F., Tremblay, R. E., Kerr, M., Pagani, L., & Bukowski, W. M. (1997). Disruptive-ness, friends’ characteristics, and delinquency in early adolescence: A test of twocompeting models of development. Child Development, 68, 676–689.
Warner, B., & Rountree, P. W. (1997). Local social ties in a community and crime model:
Social Capital and Physical Violence 423

Questioning the systemic nature of informal social control. Social Problems, 44,520–536.
Weikart, D. P. (1998). Changing early childhood development through educational inter-vention. Preventive Medicine, 27(2), 233–237.
Weikart, D. P., & Schweinhart, L. J. (1992). High/Scope preschool program outcomes. InJ. McCord & R. E. Tremblay (Eds.), Preventing antisocial behavior: Interventionsfrom birth to adolescence (pp. 67–86). New York: Guilford Press.
Wolfgang, M. E., & Ferracuti, F. (1967). The subculture of violence: Towards an inte-grated theory in criminology. London: Tavistock Publications.
Wright, J. P., Cullen, F. T., & Miller, J. T. (2001). Family social capital and delinquent in-volvement. Journal of Criminal Justice, 29, 1–9.
Zaslow, M., Tout, K., Smith, S., & Moore, K. (1998). Implications of the 1996 welfarelegislation for children: A research perspective. Social Policy Report. Society for Re-search in Child Development, 12(3), 1–35.
Zhang, L., Welte, J. W., & Wieczorek, W. F. (1999). Youth gangs, drug use, and delin-quency. Journal of Criminal Justice, 27, 101–109.
424 D E T E R M I N A N T S O F A G G R E S S I O N

DETERMINANTS OF AGGRESSIONSex Differences in Aggressive Behavior
20
Sex Differences in Aggressive BehaviorA Developmental and Evolutionary Perspective
JOHN ARCHER and SYLVANA CÔTÉ
INTRODUCTION: EVOLUTIONARY AND DEVELOPMENTAL EXPLANATIONS
Explanations of Behavior
Tinbergen (1963) set out four types of explanation of behavior. Two con-cerned the evolution of behavior, its historical antecedents and its survivalvalue, and two concerned causes within the individual, its development andimmediate causation. A full explanation of any form of behavior that humansshare with animals involves all four explanations. Aggression is such a form ofbehavior, having its origins in evolutionary history. Psychologists studyinghuman aggression typically concentrate on individual-level explanations, im-mediate causation and development, referred to as proximal explanations.Evolutionary psychologists and ethologists are also concerned with the twoevolutionary-based explanations, referred to as distal explanations. In thischapter we consider the development of sex differences in human aggressionin relation to evolutionary explanations.
Evolutionary History
The evolutionary history of aggression can be appreciated by noting its wide-spread occurrence throughout the animal kingdom, from animals with verysimple nervous systems, such as sea anemones, to humans (Archer, 1988). It istherefore not something that can realistically be viewed only as a consequenceof social influences on human development, although there are social environ-ments that greatly increase an individual’s level of aggression, and individuals
425

who are more aggression-prone than others whatever their social environ-ment.
Evolutionary Function
Fighting is widespread throughout the Animal kingdom. It occurs in two maincontexts, in warding off danger and as a consequence of competition for re-sources (Archer, 1988). Competition is a wider term than physical aggressionand results from a simple ecological principle. Organisms typically producemore offspring than their environment can sustain; resources are thereforescarce, and some will obtain more than others. Occasionally, this is the resultof chance, but animals are typically selected for features that enable them tooutcompete and outbreed others of their kind. This process often occurs overa period of time and is not immediately obvious. For example, one animal’sdigestive system may be more efficient than another’s, or one animal may bebetter at finding food than another.
Aggressive behavior is a form of competition that has evolved where or-ganisms have the ability to displace one another quickly, and there are re-sources worth fighting over. Thus, animals that can remove others by forcewill seek to do so. Resources that are located in one place and can be obtainedby actively displacing a rival will be fought over, whereas those that are evenlyand abundantly distributed over a wide area will not. Thus, hyenas fight overa carcass whereas cows do not fight over grass. Broadly speaking, the occur-rence of aggression in animals can be understood in terms of the costs andbenefits of fighting (Archer, 1988), and this principle can be extended to hu-man aggression.
Sexual Selection
A more specific application of the cost–benefit analysis is incorporated intothe principle of sexual selection (Darwin, 1871), which involves intermalecompetition and female choice. With a few exceptions, males have more togain than females have through fighting one another (or more to lose throughnot fighting). Trivers (1972) explained why this is the case. The specializationof germ cells leads to unequal parental investment. Females show greater ini-tial investment in terms of the energy and time costs of producing egg cells,and therefore a greater cost is sustained if they choose a low-fitness mate.Males show less initial investment, and therefore sustain a lower cost throughchoosing a low-fitness mate. These factors result in females being more selec-tive in their mate choice. They also lead to males being in competition withother males for both access to females and the resources that make them moreacceptable to females.
These considerations enable us to understand why greater physical ag-gression by males than females is observed in many animal species and is com-
426 D E T E R M I N A N T S O F A G G R E S S I O N

mon throughout the mammals. Trivers’s (1972) theory of parental investmentpredicts that sex differences in aggression (and in associated features such assize and strength) are most pronounced when male parental investment is ab-sent and competition between males is at its most intense. This is observed inspecies such as the elephant seal (Le Boeuf, 1974), when one male defends agroup of as many as 40 females against other males. In species in which themale contributes considerably to subsequent parental investment, sex differ-ences are minimal. In those relatively unusual cases in which male parentalinvestment exceeds that of females, it is the females who are larger and moreaggressive.
Human Sex Differences in Evolutionary Perspective
Comparison of sex differences in body size with those in other primates sug-gests that there has, in evolutionary history, been a tendency toward mildpolygyny in humans (Plavcan & van Schaik, 1997). A range of studies sug-gests that the sexual natures of men and women, and sex differences in aggres-sion, fit this pattern (Archer & Lloyd, 2002), although there is also consider-able variation between males (Archer & Mehdikhani, 2003). A range ofevidence on direct physical aggression shows that it is greater among malesthan females—at least toward adults of the same sex. There are large sex dif-ferences in measures of physical aggression (Archer, 2004), and in recordedviolent crime (Courtwright, 1996; Daly & Wilson, 1990), for same-sex en-counters.
Development from an Evolutionary Perspective
Given this evolutionary background, we can understand why human aggres-sion occurs and why there are sex differences in direct forms of human aggres-sion. However, how aggression develops and when the sex differences firstappear in development is another question, not readily answered by these evo-lutionary considerations alone. To appreciate some of the complexities thatare involved when we consider development from an evolutionary perspective,it is worth noting a number of general principles that have arisen from re-search on development from an evolutionary perspective (Archer, 1992a;Bjorklund & Pellegrini, 2002; Stamps, 2003). In each case, we indicate brieflyhow the principle might apply to the development of aggression and sex dif-ferences in aggression.
The first principle is that behavior serving an evolutionary function mayarise from different developmental processes in different animals. Thus,greater aggressiveness by males is achieved in different ways in different spe-cies. In some birds and mammals, testosterone directly enhances male aggres-siveness, but as we show in this chapter, it is likely that different processesoperate to produce this same end point in humans.
Sex Differences in Aggressive Behavior 427

Second, some types of behavior that develop during childhood serve thefunction of preparing the individual for a particular role in adulthood. Sex dif-ferences in aggression and rough-and-tumble play are likely examples of thisin humans (Bjorklund & Pellegrini, 2002). In other cases, behavior may servean adaptive function during childhood itself. We show that physical aggres-sion develops early in childhood, and that the objective of obtaining and re-taining resources may be one reason why this occurs.
Third, children are born not with a tabula rasa, but with a program fordevelopment that has evolved in human ancestral environments to serve anadaptive outcome. However, the potential range of outcomes, or phenotypes,that can be produced from this program is wide. This is termed the norm ofreaction, and it is a particularly important principle for considering develop-ment in modern human environments, which are very different from those ofour prehistoric ancestors. Thus, the modern school environment may lead toonce-adaptive behavior, such as high levels of aggression and rough-and-tumbleplay, being viewed as maladaptive.
Fourth, some evolved forms of behavior are little affected by variability inthe environments in which the individuals develop, whereas others are muchaffected by it. The onset of physical aggression early in life may not be readilysusceptible to environmental modification, whereas its subsequent develop-ment is likely to be influenced to a much greater extent by the type of socialand physical environment in which the individual develops.
Fifth, individuals select and modify the environments in which they de-velop. For example, as a consequence of their own preferences, boys andgirls form sex-segregated groups early in their social development, and theselast through to the teenage years (Maccoby, 1998). Such groups constitutedifferent social environments, in which boys and girls come to develop dif-ferent patterns of behavior including aggression, boys showing more directphysical forms, and girls more indirect forms involving manipulation of re-lationships.
A sixth principle is that selection may favor phenotypic diversity ratherthan a single adaptive type. There are various reasons for this, such as thatdifferent phenotypes are adapted for different environments, or that a rarephenotype has an advantage simply because it is unusual. Males of manyanimal species show what are termed alternative reproductive strategies, forexample, specialized either for fighting other males or for sneaking up onfemales when other males are distracted. In humans, it has been suggestedthat males vary from the more aggressive risk-prone male who favors ashort-term promiscuous reproductive strategy to the less aggressive, morerisk-averse male who favors a longer-term parental reproductive strategy(Archer & Mehdikhani, 2003). It follows that a variety of interlinked char-acteristics, or trait syndromes, are likely to develop together in the same indi-vidual. Thus, an aggression-prone individual may show a number of otherhigh-risk and impulsive types of behavior that in our society are typically la-beled as antisocial or criminal.
428 D E T E R M I N A N T S O F A G G R E S S I O N

THE DEVELOPMENT OF PHYSICAL AGGRESSION
The social learning perspective has provided an influential account of the de-velopment of social behavior, including aggression, in terms of principles orig-inally derived from learning theory (Bandura, 1973; Loeber & Stouthamer-Loeber, 1998; Reiss & Roth, 1993). Aggressive and violent behaviors areviewed as being learned during childhood and adolescence through exposureto social influences such as violent television, aggressive role models, or devi-ant peers (Johnson, Cohen, Smailes, Kasen, & Brook, 2002; Patterson,Dishion, & Bank, 1984; Thornberry, 1998). From this perspective alone, wewould expect children to become more aggressive and violent as they becomeolder.
The social learning perspective provides a stark contrast with a view ofdevelopment informed by evolutionary principles. First, there is no recogni-tion of an evolved program for the development of aggression to serve the im-mediate function of resource competition during childhood or as a prepara-tion for conflicts during adulthood. Evolutionary approaches to developmentrecognize that social learning is an important influence in behavior (Stamps,2003). However, it is one among a wider range of genetic and environmentalinfluences and is likely to extend beyond the facilitation of aggression by so-cial learning that has been the focus of most investigations from this perspec-tive.
Recent longitudinal studies indicate that children become less, not more,physically aggressive and violent over time (Broidy et al., 2003; Cairns,Cairns, Neckerman, Fergusson, & Gariepy, 1989; Nagin & Tremblay, 1999;Tremblay et al., 1999). Studies have found that most children followed declin-ing trajectories of physical aggression between kindergarten and grade 6(Broidy et al., 2003; Nagin & Tremblay, 1999). If the development of aggres-sion were dependent solely on the cumulative effects of social influences tend-ing to promote aggressive behavior, we would expect a significant group ofchildren to begin their use of physical aggression during the elementary schoolyears. Such a group has not been not identified (Kingston & Prior, 1995;Loeber & Stouthamer-Loeber, 1998; Tremblay et al., 1991). The majority ofthose at risk for later violent behavior were already on high trajectories ofphysical aggression in kindergarten (Broidy et al., 2003; Nagin & Tremblay,1999).
The possibility that aggression in middle childhood simply reflects thecontinuation of a behavior pattern already present during the first years of lifeis supported by research on the development of disruptive behavior disordersduring the preschool years. Physically aggressive behavior is common amonginfants and toddlers (Goodenough, 1931; Hay, Castle, & Davies, 2000;Tremblay et al., 1999).
However, most studies have examined the development of aggressionduring limited periods of development. An exception is the National Longitu-dinal Study of Canadian Youth (NLSCY), in which the developmental trajec-
Sex Differences in Aggressive Behavior 429

tories of physical aggression between infancy and preadolescence were esti-mated, with longitudinal data collected on a population sample (n = 8,208).The developmental patterns identified with mother reports indicated thataggression was part of most (65.5%) preschoolers’ behavioral repertoire, butthat the majority of children had successfully ceased to use physical aggressionby the end of the elementary school years.
As shown in Figure 20.1, four distinct trajectory groups were identified inthe NLSCY. The first comprised children with stable low levels of aggression.The second group—the rapid-desister group—included children with moder-ate levels of physical aggression in toddlerhood, who desisted rapidly andmaintained low levels from school entry to preadolescence. The third groupwas labeled the “moderate desisters”: it included children with slow but con-stant declines in physical aggression between 2 and 11 years. Finally, children
430 D E T E R M I N A N T S O F A G G R E S S I O N
FIGURE 20.1. Trajectories of physical aggression for boys and girls (n = 8,208).

in the high physical aggression group were also desisting. However, their de-clines were of such small magnitude that in preadolescence (age 11) they stillexhibited notably higher levels of physical aggression than did their peers.
It is important to note that no significant group of children with an onsetof physical aggression during this period (between 2 and 11 years) was identi-fied. According to a retrospective study with younger children, most infantsstart to use physical aggression between the first and the second years afterbirth (Tremblay et al., 1999). Its onset is therefore likely to be earlier and to berelatively stable, rather than being dependent on specific environmental influ-ences that may lead to variable onset (see Tremblay & Nagin, Chapter 5, thisvolume). This suggests that physical aggression is an evolved pattern of behav-ior that develops early in life, irrespective of the sort of social influences high-lighted by social learning theory.
From the evidence reviewed in this section, it is clear that human aggres-sion appears early in life and is part of social interactions right from their be-ginning. Social learning theory has perhaps laid too much emphasis on thelearning of specific forms of aggressive behavior as a general mechanism forexplaining the developmental progression of aggression. Although such spe-cific aggressive actions can undoubtedly be learned from peers, parents, andmedia representations, they do not account for broad changes with age. How-ever, social factors such as coercive parenting, violent models, or violent mediamay be related to the failure, for a minority of children, to inhibit and regulatetheir aggression.
Why do high levels of physical aggression appear at such young ages? Wehave already highlighted the importance of competition for resources in func-tional explanations of aggression. Among young children, we would predictthat aggression arises when more than one child claims the same resources. Inmodern affluent societies, children do not have to fight for their lives, as oc-curs occasionally in other species (Frank, 1997; Stinson, 1979), or over re-sources necessary for survival, such as food, as occurs in the young of manybirds and mammals (e.g., Bekoff, 1981; Geist, 1978). They may, however,fight over other resources, such as a preferred food, a toy, or a play location.Campbell (1999) argued that most conflicts at preschool ages involve compet-ing for limited resources, such as guarding a toy or a territory.
THE DEVELOPMENT OF SEX DIFFERENCES IN PHYSICAL AGGRESSION
Sex differences have been found from the onset of physical aggression itself, inchildren as young as 17 months, according to mothers’ reports of specific actsof physical aggression (Baillargeon, 2002). In this study, boys were twice aslikely as girls to hit another child frequently. Observational studies show dif-ferences of fairly substantial magnitudes at around 3 years of age. Hay et al.(2000) found an effect size of d = 0.64, for a (relatively large) sample of 66children between 18 and 30 months of age. Campbell, Shirley, and Caygill
Sex Differences in Aggressive Behavior 431

(2002) observed considerably more negative interactions with peers by boysthan girls in children who were 27 months old. Negative interactions includeda child’s grabbing another child’s toy (d = 1.42) and a child’s resisting an-other’s attempt to grab his or her toy (d = 0.86).1 Observational studies ofchildren 4 years of age have also yielded large effect sizes (e.g., Sears, Rau, &Alpert, 1965: d = 1.27; McGrew, 1972: d = 1.29).
The conclusion that sex differences in physical aggression are presentearly in life is counter to earlier accounts based on the assumption that theyarise as a result of the differential socialization of boys and girls (e.g.,Tieger, 1980). In a review of sex differences in several types of childhoodbehavior problems, Keenan and Shaw (1997) put forward a similar view.They reviewed findings from five observational studies (n = 20–90) con-cerned with the development of aggression during the preschool years. Theyreported that boys were more aggressive than girls in only one of thesestudies (Gunnar, Senior, & Hartup, 1984) and concluded that there is nosex difference in aggression for toddlers, although there is an increasing di-vergence in girls’ and boys’ behavior during the preschool period. This wasbased on two longitudinal studies indicating that girls exhibited less aggres-sion by age 5.
Keenan and Shaw’s conclusion that there are no sex differences for tod-dlers is not supported by closer examination of the evidence they cite. Theirown study of children ages 18 and 24 months interacting with their mothersshowed sex differences in global aggression at both ages (18 months: d = 0.49;confidence interval [CI] = 0.07–0.91; p = .02; 24 months: d = 0.39; CI = 0.02–0.80; p = .06), those at the younger age being greater (figures calculated fromTable 2 in Shaw, Keenan, & Vondra, 1994). Jacklin and Maccoby (1978) ob-served dyads of 33-month-old children: Comparing aggression among maledyads with that among female dyads (i.e., taking each dyad as a unit) pro-duced a small effect size in the male direction (d = 0.18). Cummings, Iannotti,and Zahn-Waxler (1989) observed children playing with their best availablefriends at 2 and at 5 years. For the two ages combined (the only calculationpossible), effect sizes for sex differences in aggression were d = 0.26 in themale direction. Caplan, Vespo, Pedersen, and Hay (1991) observed 1- and 2-year-olds in mixed-sex groups of three for 25 minutes. The main comparisonwas between the level of conflict in groups with one male and groups with onefemale. A footnote indicated that there was no significant sex difference at anindividual level, but no direction was reported (for this sample size, the dvalue could have been anywhere between -0.57 and 0.57). The remainingstudy (Gunnar et al., 1984) found significantly longer duration of conflictsamong male than female dyads (d = 1.04).
A meta-analysis of the four studies that enabled effect sizes to be calcu-lated produced a weighted mean of d = 0.44 (CI = 0.15–0.74; p = .003), show-ing that there was a moderate sex difference in the male direction. This con-trasts with Keenan and Shaw’s conclusion, based on a narrative review, thatthere are no sex differences at young ages. It supports the conclusion from the
432 D E T E R M I N A N T S O F A G G R E S S I O N

other studies described above, that the sex difference occurs as early as it ispossible to measure children’s aggressive behavior.
Keenan and Shaw’s (1997) review also raised the question of how sex dif-ferences in physical aggression change throughout childhood, as they claimedthat it increased from about 2 to 5 years of age, based on two longitudinalstudies. A meta-analysis of 40 observational studies carried out between 1965and 2000 (Archer, 2004) showed no significant difference in the effect size forthe sex difference in aggression (overall d = 0.53) between three age catego-ries, under 6 years, 6–11 years, and over 11 years. However, this involvedcross-sectional comparisons of different studies, and there are limitations tothe inferences that can be drawn from such evidence. Large-scale representa-tive samples provide a way of overcoming the limitations, although at youngerages they involve less direct assessments of behavior, inasmuch as they gener-ally rely on mothers’ reports.
In the NLSCY (see the preceding section), sex differences in physical ag-gression were examined by comparing the proportion of boys and girls as-signed to the different trajectory groups. The results showed that boys weremore likely to be on the highest physical aggression trajectories (odds ratio[OR] = 1.56; 95% CI = 1.41–1.71) and less likely to be on the lowest trajecto-ries (OR = 0.85; 95% CI = 0.82–0.89). This suggests that boys were alreadymore physically aggressive than girls at 2 years of age, a difference that per-sisted throughout childhood. These sex differences are consistent with thefindings from the individual observational studies presented earlier.
The NLSCY study also found that the initial sex difference became morepronounced during the course of childhood, as boys were less likely to be as-signed to the rapid-desister trajectory group (OR= 0.88; 95% CI = 0.82–0.94). These findings support Keenan and Shaw’s (1997) conclusion that sexdifferences in physical aggression increase during the preschool years and thatthis is attributable to girls’ faster rate of decline between 2 and 5 years. Thus,girls may learn more quickly than boys to inhibit or control (physically) ag-gressive behaviors (Kochanska, Murray, & Harlan, 2000). Finally, there wereno significant sex differences for the moderate desister group (OR = 1.05;95% CI = 1.00–1.09).
One further point about the sex differences found in the NLSCY study isthat they are far from absolute. Many boys are found in the low groups andthe rapid-desister groups, and many girls are found in the high-trajectorygroup. Such variation may be worth investigating in relation to the evolution-ary hypothesis of alternative reproductive strategies, which would predict con-sistent ways of responding, with an early developmental onset and a consis-tent outcome.
A meta-analysis of sex differences in self-reports of physical aggression(Archer, 2004) showed similar effect sizes for the age categories 6–11 years,12–3 years, and 14–17 years. The lack of any further widening in the sex dif-ference at puberty provides indirect evidence that pubertal testosterone doesnot increase male aggressiveness, as it does in some other mammals (e.g.,
Sex Differences in Aggressive Behavior 433

Beeman, 1947; Edwards, 1969). More direct support for this conclusioncomes from a longitudinal study of boys going through puberty (Halpern,Udry, Campbell, & Suchindran, 1994), which found no influence of rising tes-tosterone levels on physical aggression.
The meta-analysis of self-reported physical aggression (Archer, 2004) didfind larger sex differences in the age categories 18–21 and 22–30 years than atyounger (14–17 years) or older ages (31–55 years). Studies of violent crimeand of homicides show that there is a pronounced peak in males betweenabout 20 and 29 years of age. This was found for homicides in data sets fromfour locations with different homicide rates, England and Wales, Canada, Chi-cago, and Detroit (Daly & Wilson, 1990), and for violent crimes in the mod-ern United States, 19th-century England and Wales, and early 19th-centuryEurope (Courtwright, 1996; Quetelet, 1833/1984). Although these studies dif-fer from those we have considered so far in that they involve much rarer vio-lent acts, defined by their damaging consequences rather than the type of be-havior, they show a striking parallel in their pattern of age and sexdistribution.
The evidence reviewed in this section indicates that there is a large sexdifference in physical aggression, beginning early in life, suggesting that it isnot the product of a gradual process such as socialization (Tremblay et al.,1999). However, there is also evidence from a longitudinal study that the sexdifference in physical aggression, such as hitting, biting, and attacking others,may become larger between toddlerhood and kindergarten. There is no evi-dence that the sex difference widens at puberty as a consequence of the actionof the male hormone testosterone. However, studies of self-reported physicalaggression and of violent acts converge to suggest a widening of the sex differ-ence between 18 and 30 years of age. These are peak years for the develop-ment of a constellation of characteristics that enable a young man to success-fully compete with other males.
THE DEVELOPMENT OF SEX DIFFERENCESIN PHYSICAL AGGRESSION: AN EVOLUTIONARY VIEW
We noted that resource competition provides an evolutionary reason for theappearance of physical aggression early in life. However, it is less clear whythere would be sex differences in this aggression early in life. We might expect,in the absence of reproductive competition, boys and girls to behave similarly,inasmuch as they would be competing for similar resources. Campbell (1999)articulated this view, arguing that, as toddlers, children’s conflicts are mainlyrelated to competing for limited resources, such as guarding a toy or a terri-tory. She also argued that there are no sex differences in this type ofresource-oriented aggression among toddlers, whereas there is a sex differ-ence in status-oriented aggression by 4 years of age. The first conclusion wasbased on a narrative review of a very small number of studies that sought to
434 D E T E R M I N A N T S O F A G G R E S S I O N

distinguish these different categories of aggression. It is inconsistent withother findings (e.g., Gunnar et al., 1984; Jacklin & Maccoby, 1978), and witha later study by Campbell et al. (2002), which reported very large sex differencesat 27 months in negative interactions (including a child’s grabbing anotherchild’s toy and a child’s resisting another’s attempt to grab his or her toy).
Campbell is more likely to be correct in her identification of status dis-putes as an important area in which the sexes diverge in the early years. It isclear that as children become older there is increasing sex segregation and dif-ferent types of social relations in boys’ and girls’ groups, based initially onself-selection of playmates (Archer, 1992b; Maccoby, 1998). Boys are moreconcerned with status and dominance—for example, who is tough and who isthe boss (Weisfeld, 1994)—and they operate in larger play groups. Girls formmore intense relations with one or two close friends (Archer, 1992b). Thesecontrasting patterns have wide-ranging implications for later sex differences inseveral areas of social behavior, including other forms of aggression (Maccoby,1998). Because of the greater emphasis on physically based dominance inchildhood, boys’ exchanges are more likely to develop into physical aggres-sion, whereas girls develop strategies of indirect aggression that become prom-inent in the teenage years. Nevertheless, as we noted earlier, the expectationthat there will be a greater contrast between the frequencies of physical ag-gression in boys’ and girls’ groups as they become older has, as yet, only lim-ited empirical support.
Sex differences in physical aggression prior to the ages of reproductivecompetition are found in the young of many mammals, together with sex dif-ferences in rough-and-tumble play. In addition to the advantages during child-hood of gaining tangible rewards, such as food and other valued resources,success in physical fights, whether over status or resources, may have twoother longer-term consequences. The first involves the learning of techniquesof fighting that may prove valuable in young adulthood, and the learning ofcues associated with when it is preferable not to fight (assessment strategies;Archer, 1994). The second involves the longer-term advantage, conferred by aphysically based dominance acquired during childhood, for dominance inyoung adulthood. This may have been considerable in the ancestral environ-ment, and may still be so where physical prowess forms the basis of domi-nance orders in adulthood, as occurs in many masculine subcultures.
If the sex difference in physical aggression found in childhood arisesmainly from the concern of boys with dominance relations and the learning ofways of fighting in preparation for adulthood, we would expect it to belargely manifest in relation to same-sex disputes. If instead it arises from re-source competition (as suggested for early ages), we would expect the aggres-sion to be directed to any child who has a preferred toy, irrespective of his orher sex.
There are relatively few studies that have separated the sex of the oppo-nent when considering children’s aggression. Where they have, the evidencesupports the view that sex differences in physical aggression concern aggres-
Sex Differences in Aggressive Behavior 435

sion to same-sex opponents. A different pattern (in the female direction) isfound whenever the (less frequent) between-sex physical aggression is studied.For example, an observational study by Barrett (1979) of 6- to 7-year-old chil-dren found a substantial sex difference in physical aggression in the maledirection for same-sex opponents (d = 1.05)2, and a difference in the femaledirection for opposite-sex opponents (d = –0.53). Pellegrini and Long (2002)followed up a sample of children at 6-monthly intervals from 12.8 years ofage and found at all ages except the youngest one, a sex difference in the maledirection for same-sex aggression and in the female direction for opposite-sexaggression. Effect sizes were in the moderate range, from 0.29 to 0.46. Cairnsand Cairns (1994) reported similar findings using questionnaires at 11 and 14years of age. A questionnaire study involving an older age (16.8 years) found asimilar pattern, with d = 1.03 for same-sex aggression and –0.95 for opposite-sex encounters (Hilton, Harris, & Rice, 2000). Gergen (1990) and Harris(1992) found similar results using self-reports at 19 years of age.
SEX DIFFERENCES IN VERBAL AGGRESSION
The evolutionary origins of verbal aggression are seen in the vocal threat dis-plays of animals. Some aspects of human aggression are directly comparable,such as shouting in a loud and intimidating way at someone, accompanied bythreatening gestures. These forms are found as early as children show socialinteractions, and accompany physical aggression. Other types of verbal ag-gression are uniquely human, for example, making verbal threats that specifyintended violent actions, such as “I’ll smash your head in.” Insults are alsouniquely human and generally involve remarks or accusations that challengeimportant aspects of a person’s identity, denigrating the person in some way.For example, a teenage boy may be called names that imply that he is cow-ardly, such as “chicken” or “yellow,” whereas a teenage girl may be callednames that imply that she is sexually promiscuous, such as “slag” or “slut”(Campbell, 1995). These insults challenge characteristics that are especiallyvalued in the particular sex. Traditionally, insults were viewed as attacks on aperson’s honor or reputation and, in circumstances where there was no effec-tive rule of law, had to be avenged to prevent loss of face and status for boththe person and his or her family (Nisbett & Cohen, 1996; Ruff, 2001). Thus,insults such as “son of a bitch” or “bastard,” which have largely lost their po-tency in modern society, were once regarded as denigrating statements about aperson’s whole family. More subtle forms of verbal aggression develop later inchildhood, for example, making someone the butt of remarks that are deni-grating to that person, but humorous to others. Being able to use language inthis way is particularly important in groups of teenage boys and young men(Benson & Archer, 2002). There is therefore considerable diversity within thecategory “verbal aggression.”
Yet verbal aggression by both sexes has generally been considered as a
436 D E T E R M I N A N T S O F A G G R E S S I O N

single overall category, so that the use of particular forms of verbal aggres-sion by the two sexes has seldom been studied. Meta-analytic summaries ofsex differences in aggression can only provide a broad view of changes withage. The sex difference for self-reported verbal aggression is in the same di-rection as that for physical aggression, but is generally smaller in size (Ar-cher, 2004). The difference was smaller at 6–11 years (d = 0.19) than at 11to 17 years (d = 0.36). Studies involving observations showed little differ-ence across age categories for verbal aggression. However, there are individ-ual studies showing sex differences in verbal aggression at young ages.Fagot and Hagan (1985) found differences in the male direction of d = 0.31between 2 and 3 years of life. McGrew (1972) and Sears et al. (1965) foundlarger differences (d = 0.53 and 1.05) at 4 years of age. As in the case ofphysical aggression, these sex differences are restricted to same-sex encoun-ters, when the sex of the opponent is considered (Barrett, 1979; Tucker,1989; Hilton et al., 2000).
THE DEVELOPMENT OF SEX DIFFERENCES IN INDIRECT AGGRESSION
Indirect aggression refers to covert forms of aggression, as opposed to director face-to-face forms. It may be physical, such as in destroying someone’sproperty, or verbal, through deliberately attacking a person’s social standingor via malicious gossip or social ostracism (Archer, 2001). Most studies haveconcentrated on verbal forms, such as getting others to dislike a person, be-coming friends with another as a form of revenge, telling a person’s secrets toanother, saying bad things about people behind their backs, and telling othersto avoid someone. Lagerspetz, Björkqvist, and Peltonen (1988) were the firstto carry out a systematic study of indirect aggression using peer ratings, whichare particularly suited to a study of this form of covert aggression. They docu-mented large sex differences in the female direction, which were found in laterstudies (Björkqvist, Lagerspetz, & Kaukiainen, 1992a). Sex differences havealso been found in a very similar form of aggression, termed relational (Crick& Grotpeter, 1995). It is a matter of dispute whether relational aggression isthe same as indirect aggression (Björkqvist, 2001) or closely overlaps with it(Archer, 2001).
Björkqvist, Osterman, and Kaukiainen (1992b) proposed that the expres-sion of aggression changes over the life course. Normatively, as children ma-ture, aggression changes from mainly physical to verbal, and then from verbalto indirect. In their cross-sectional analysis, levels of particular forms of indi-rect aggression, such as gossip, were much lower in both sexes at 8 years ofage, but were higher at 11, 15, and 18 years, and sex differences were particu-larly pronounced at these three older ages. A meta-analysis of studies usingpeer reports to investigate indirect aggression (Archer, 2004) showed that ef-fect sizes increased with age over the range of 6–17 years, so that by the teen-age years they were substantial. This coincides with the larger sex differences
Sex Differences in Aggressive Behavior 437

found from 11 years onward in Björkqvist’s analysis, and with evidence fromthe NLSCY, presented in the following discussion.
Vaillancourt (Chapter 8, this volume) presents in detail theories of the de-velopment of indirect aggression, as well as developmental trajectories (forages 4 and 11 years) estimated from the NLSCY (see also Vaillancourt et al.,2004). Briefly, the results indicate that the development of indirect aggressionfollows a pattern opposite to that for physical aggression in the same sample:While physical aggression decreases during childhood, indirect aggression in-creases.
The development of physical and indirect aggression found in the NLSCYsupport the notion that boys and girls use different types of aggressive behav-ior over the course of childhood. Indeed, the results show not only a strongertendency for girls to cease using physical aggression, they also show a strongertendency for girls to increasingly use indirect aggression over time (i.e., girlsare more likely than boys to follow rising trajectories of indirect aggression).It is noteworthy that the NLSCY shows that both sexes use both forms of ag-gression, physical and indirect. Early in childhood (from age 2), girls alreadyuse less physical aggression and more indirect (or relational) aggression thanboys do. These sex differences become even more pronounced over time, withboys being less likely to exhibit declining levels of physical aggression prior tokindergarten, and girls being more likely to follow rising trajectories of indi-rect and relational aggression during middle childhood. Even so, boys andgirls were represented on all types of trajectories, so that we again have tocaution that these generalizations hide considerable individual variation. Forinstance, a substantial proportion of girls (33.1%) followed a high and mostlystable trajectory of physical aggression, and a substantial proportion of boys(40.3%) followed a high and rising trajectory of indirect aggression. Thus, thesex differences were related to the proportion of boys and girls on trajectoriesreflecting high levels or changing levels of physical and indirect aggression.
These longitudinal data broadly agree with the cross-sectional analysisof Björkqvist and his colleagues, and with the meta-analysis of age differ-ences. An analysis of the same data found distinct factors for physical andindirect aggression, which were stable over time (Vaillancourt, Brendgen,Boivin, & Tremblay, 2003). Sex differences at younger ages (4 to 7 years)were in the female direction, but small in size, across four cohorts. Gen-erally, they became larger from 8 to 11 years, –0.26 and –0.28 being thelargest values.3
THE EVOLUTIONARY SIGNIFICANCE OF SEXDIFFERENCES IN INDIRECT AGGRESSION
Campbell (1999) proposed that females tend to use forms of aggression thatcarry low risks for physical injury, such as indirect aggression (see alsoBjörkqvist, 1994). She related this, in evolutionary terms, to the greater value
438 D E T E R M I N A N T S O F A G G R E S S I O N

attached by women to protecting their own lives, because the survival of theirinfants is more dependent on the mothers’ care and defense than the fathers’.In this context, the cost of direct and physical aggression, the forms that repre-sent risk for one’s physical safety, is greater for women than men. The greateruse of indirect and relational aggression among girls, and of physical aggres-sion among boys, is also relevant for competition with peers in the socialdomain. In an earlier section, we referred to the importance of same-sex peergroups during childhood, and the values adopted in these groups, for explain-ing sex differences in physical aggression. This can be extended to an under-standing of indirect aggression. Specifically, Maccoby (1998) and Crick andGrotpeter (1995) suggested that the structure of female friendships, whichtend to be based on intimacy and cohesiveness, facilitates the use of indirectand relational forms of aggression among girls. A cohesive social network en-ables this form of covert aggression to be used more effectively (Green, Rich-ardson, & Lago, 1996).
CONCLUSIONS
In this chapter we have sought to place the development of human sex differ-ences in aggression into an evolutionary context. The evolution of aggressioncan be understood in terms of its origins in the natural world, and its benefitsand costs in particular situations. Sex differences in aggression can be under-stood in similar terms, by considering the imbalance in parental investment inthe two sexes. Aggression begins early in postnatal life in many animals, anddata from small-scale observational studies, and from a large-scale longitudi-nal study, indicate that this is the case for humans. Such aggression consistslargely of physical forms, with accompanying vocalizations. Higher levels arefound in boys than girls from an early age, and these encounters arise fromboth property-related and dominance disputes. Following the trajectories ofaggression in a large representative sample showed a gradual decline in physi-cal aggression overall, suggesting that rather than having to be socialized intobeing aggressive, children gradually learn to inhibit their direct forms of ag-gression. More boys than girls were found on the developmental trajectoriescontaining the highest rates of physical aggression, and more girls than boyswere found on the lowest trajectories. Indirect or relational aggression ap-pears later in development, and girls were more likely than boys to follow ris-ing trajectories in this form of aggression, coinciding with findings that the sexdifference in indirect aggression is particularly pronounced in the teenageyears. These contrasting developmental pathways in girls’ and boys’ aggres-sion are not absolute differences, but show an overlap between the sexes. Nev-ertheless, they can be understood in terms of the different forms of social rela-tions that are typical of boys’ and girls’ social groups, as well as in terms ofthe different selection pressures on males and females resulting from sexualselection.
Sex Differences in Aggressive Behavior 439

NOTES
1. All these effect sizes were calculated from the published data using DSTAT (Johnson, 1989).2. Effect sizes were calculated by the first author, using D-STAT (Johnson, 1989).3. Values calculated from the authors’ data.
REFERENCES
Archer, J. (1988). The behavioural biology of aggression. Cambridge, UK: CambridgeUniversity Press.
Archer, J. (1992a). Ethology and human development. Savage, MD: Barnes & Noble.Archer, J. (1992b). Childhood gender roles: Social context and organization. In H.
McGurk (Ed.), Childhood social development: Contemporary perspectives (pp.31–61). Hove, UK, & Hillsdale, NJ: Erlbaum.
Archer, J. (1994). Violence between men. In J.Archer (Ed.), Male violence (pp. 121–140).London and New York: Routledge.
Archer, J. (2001). A strategic approach to aggression. Social Development, 10, 267–271.Archer, J. (2004). Sex differences in aggression in real-world settings: A meta-analytic re-
view. Review of General Psychology, 8, 291–322.Archer, J., & Lloyd, B. B. (2002). Sex and gender (2nd ed.). Cambridge, UK: Cambridge
University Press.Archer, J., & Mehdikhani, M. (2003). Variability among males in sexually-selected at-
tributes. Review of General Psychology, 7, 219–236.Baillargeon, R. (2002). Gender differences in physical aggression at 17 months of age:
Results from the longitudinal study of child development in Quebec. In S. Côté(Chair), Sex differences and similarities in aggression: Correlates and consequencesfrom infancy to mid-life. Symposium conducted at the 15th world meeting of the In-ternational Society for Research on Aggression, Montreal, Canada.
Bandura, A. (1973). Aggression: A social learning analysis. Englewood Cliffs, NJ:Prentice-Hall.
Barrett, D. E. (1979). A naturalistic study of sex differences in children’s aggression.Merrill–Palmer Quarterly, 25, 193–203.
Beeman, A. E. (1947). The effect of the male hormone on aggressive behavior in mice.Physiological Zoology, 20, 373–405.
Bekoff, M. (1981). Development of agonistic behaviour: Ethological and ecological as-pects. In P. F. Brain & D. Benton (Eds.), Multidisciplinary approaches to aggressionresearch (pp. 161–178). Amsterdam: Elsevier/North Holland.
Benson, D. A., & Archer, J. (2002). An ethnographic study of sources of conflict betweenyoung men in the context of the night out. Psychology, Evolution and Gender, 4, 3–30.
Bjorklund, D. J., & Pellegrini, A. D. (2002). The origins of human nature: Evolutionarydevelopmental psychology. Washington, DC: APA.
Björkqvist, K. (1994). Sex differences in physical, verbal and indirect aggression: A re-view of recent research. Sex Roles, 30, 177–188.
Björkqvist, K. (2001). Different names, same issue. Social Development, 10, 272–275.Björkqvist, K., Lagerspetz, K. M. J., & Kaukiainen, A. (1992a). Do girls manipulate and
boys fight? Developmental trends in regard to direct and indirect aggression. Ag-gressive Behavior, 18, 117–127.
440 D E T E R M I N A N T S O F A G G R E S S I O N

Björkqvist, K., Osterman, K., & Kaukiainen, A. (1992b). The development of direct andindirect aggressive strategies in males and females. In K. Björkqvist & P. Niemela(Eds.), Of mice and women: Aspects of female aggression (pp. 51–64). San Diego,CA: Academic Press.
Broidy, L. M., Nagin, D. S., Tremblay, R. E., Bates, J. E., Brame, B., Dodge, K., Fergusson,D., Horwood, J., Loeber, R., Laird, R., Lynam, D., Moffitt, T. E., Pettit, G. S., &Vitaro, F. (2003). Developmental trajectories of childhood disruptive behaviors andadolescent delinquency: A six site, cross-national study. Developmental Psychol-ogy, 39, 222–245.
Cairns, R. B., & Cairns, B. D. (1994). Lifelines and risks: Pathways of youth in our time.New York and London: Harvester Wheatsheaf.
Cairns, R. B., Cairns, B. D., Neckerman, H. J., Ferguson, L. L, & Gariepy, J. L. (1989).Growth and aggression: I. Childhood to early adolescence. Developmental Psychol-ogy, 25, 320–330.
Campbell, A. (1995). A few good men: Evolutionary psychology and female adolescentaggression. Ethology and Sociobiology, 16, 99–123.
Campbell, A. (1999). Staying alive: Evolution, culture, and women’s intrasexual aggres-sion. Behavioral and Brain Sciences, 2, 203–252.
Campbell, A., Shirley, L., & Caygill, L. (2002). Sex-typed preferences in three domains:Do two-year-olds need cognitive variables? British Journal of Psychology, 93, 203–217.
Caplan, M., Vespo, J., Pedersen, J., & Hay, D. F. (1991). Conflict and its resolutionin small groups of one- and two-year-olds. Child Development, 62, 1513–1524.
Courtwright, D. T. (1996). Violent land: Single men and social disorder from the frontierto the inner city. Cambridge, MA: Harvard University Press.
Crick, N. R., & Grotpeter, J. K. (1995) Relational aggression, gender, and social-psycho-logical adjustment. Child Development, 66, 710–722.
Cummings, E. M., Iannotti, R. J., & Zahn-Waxler, C. (1989). Aggression between peersin early childhood: Individual continuity and developmental change. Child Devel-opment, 60, 887–895.
Daly, M., & Wilson, M. (1990). Killing the competition: Female/female and male/malehomicide. Human Nature, 1, 81–107.
Darwin, C. (1871). The descent of man and selection in relation to sex. London: Murray.Edwards, D. A. (1969). Early androgen stimulation and aggressive behavior in male and
female mice. Physiology and Behavior, 40, 333–338.Fagot, B. I., & Hagan, R. (1985). Aggression in toddlers: Responses to the assertive acts
of boys and girls. Sex Roles, 12, 341–351.Frank, L. G. (1997). Evolution of genital masculinization: Why do female hyaenas have
such a large “penis”? Trends in Ecology and Evolution, 12, 58–62.Geist, V. (1978). On weapons, combat and ecology. In L. Krames, P. Pliner, & T. Alloway
(Eds.), Advances in the study of communication and affect: Vol. 4. Aggression,dominance and individual spacing (pp. 1–30). New York: Plenum Press.
Gergen, M. (1990). Beyond the evil empire: Horseplay and aggression. Aggressive Be-havior 16, 381–398.
Goodenough, F. L. (1931). Anger in young children. Westport, CT: Greenwood Press.Green, L. R., Richardson, D. R., & Lago, T. (1996). How do friendship, indirect, and di-
rect aggression relate? Aggressive Behavior, 22, 81–86.Gunnar, M. R., Senior, K., & Hartup, W. W. (1984). Peer presence and the exploratory
Sex Differences in Aggressive Behavior 441

behavior of eighteen- and thirty-month-old children. Child Development, 55,1103–1109.
Halpern, C. T., Udry, J. R., Campbell, B., and Suchindran, C. (1994). Relationships be-tween aggression and pubertal increases in testosterone: A panel analysis of adoles-cent males. Social Biology, 40, 8–24.
Harris, M. B. (1992). Sex and ethnic differences in past aggressive behaviors. Journal ofFamily Violence, 7, 85–102.
Hay, D. F., Castle, J., & Davies, L. (2000). Toddlers’ use of force against familiar peers: Aprecursor of serious aggression? Child Development, 71, 457–467.
Hilton, Z., Harris, G. T., & Rice, M. E. (2000). The functions of aggression by male teen-agers. Journal of Personality and Social Psychology, 79, 988–994.
Jacklin, C. N., & Maccoby, E. E. (1978). Social behavior at 33 months in same-sex andmixed-sex dyads. Child Development, 49, 557–569.
Johnson, B. T. (1989). Software for the meta-analytic review of research literatures.Hillsdale, NJ: Erlbaum.
Johnson, J. G., Cohen, P., Smailes, E. M., Kasen, S., & Brook, J. S. (2002) Televisionviewing and aggressive behavior during adolescence and adulthood. Science, 295,2468–2471.
Keenan, K., & Shaw, D. (1997). Developmental and social influences on young girls’early problem behavior. Psychological Bulletin, 121, 95–113.
Kingston, L., & Prior, M. (1995). The development of patterns of stable, transient, andschool-age onset aggressive behavior in young children. Journal of American Acad-emy of Child and Adolescent Psychiatry, 34, 348–358.
Kochanska, G., Murray, K. T., & Harlan, E. T. (2000). Effortful control in early child-hood: Continuity and change, antecedents, and implications for social develop-ment. Development and Psychopathology, 36, 220–232.
Lagerspetz, K. M. J., Björkqvist, K., Peltonen, T. (1988). Is indirect aggression typical offemales? Gender differences in aggressiveness in 11 to 12–year-old children. Aggres-sive Behavior, 14, 403–414.
Le Boeuf, B. J. (1974). Male-male competition and reproductive success in elephant seals.American Zoologist, 14, 163–176.
Loeber, R., & Stouthamer-Loeber, M. (1998). Development of juvenile aggression and vi-olence. Some common misconceptions and controversies. American Psychologist,53, 242–259.
Maccoby E. E. (1998). The two sexes: Growing up apart, coming together. Cambridge,MA: Belknap Press/Harvard University Press.
McGrew, W. C. (1972). An ethological study of children’s behavior. New York: AcademicPress.
Nagin, D. S., & Tremblay, R. E. (1999). Trajectories of boys’ physical aggression, opposi-tion, and hyperactivity on the path to physically violent and non violent juvenile de-linquency. Child Development, 70, 1181–1196.
Nisbett, R. E., & Cohen, D. (1996). Culture of honor: The psychology of violence in theSouth. Boulder, CO: Westview Press.
Patterson, G. R., Dishion, T. J., & Bank, L. (1984). Family interaction: A process modelof deviancy training. Aggressive Behavior, 10, 253–267.
Pellegrini, A. D., & Long, J. D. (2002). A longitudinal study of bullying, dominance, andvictimization during the transition from primary school through secondary school.British Journal of Developmental Psychology, 20, 259–280.
Plavcan, J. M., & Van Schhaik, C. P. (1997). Intrasexual competition and body weight di-
442 D E T E R M I N A N T S O F A G G R E S S I O N

morphism in anthropoid primates. American Journal of Physical Anthropology,103, 27–68.
Quetelet, A. (1984). Research on the propensity for crime at different ages. (S. F.Sylvester, Trans.). Cincinnati, OH: Anderson. (Original work published 1833)
Reiss, A. J., & Roth, J. A. (1993). Understanding and preventing violence. Washington,DC: National Academy Press.
Ruff, J. R. (2001). Violence in early modern Europe 1500–1800. Cambridge, UK: Cam-bridge University Press.
Sears, R. R., Rau, L., & Alpert, R. (1965). Identification and childrearing. Stanford, CA:Stanford University Press.
Shaw, D. S., Keenan, K., & Vondra, J. I. (1994). Developmental precursors of external-izing behavior: Ages 1 to 3. Developmental Psychology, 30, 355–364.
Stamps, J. (2003). Behavioural processes affecting development: Tinbergen’s fourthquestion comes of age. Animal Behaviour, 66, 1–13.
Stinson, C. H. (1979). On the selective advantage of fratricide in raptors. Evolution, 33,1219–1225.
Thornberry, T. P. (1998). Membership in youth gangs and involvement in serious and vio-lent offending. In R. Loeber & D. P. Farrington (Eds.), Serious and violent juvenileoffenders: Risk factors and successful interventions (pp. 147–166). ThousandOaks, CA: Sage.
Tieger, T. (1980). On the biological basis of sex differences. Child Development, 51, 943–963.
Tinbergen, N. (1963). On the aims and methods of ethology. Zeitschrift fur Tierpsy-chologie, 20, 410–413.
Tremblay, R. E., Japel, C., Perusse, D., Boivin, M., Zoccolillo, M., Montplaisir, J., &McDuff, P. (1999). The search for the age of “onset” of physical aggression: Rous-seau and Bandura revisted. Criminal Behavior and Mental Health, 9, 8–23.
Tremblay, R. E., Loeber, R., Gagnon, C., Charlebois, P., Larivée, S., & LeBlanc, M.(1991). Disruptive boys with stable and unstable high fighting behavior patternsduring junior elementary school. Journal of Abnormal Child Psychology, 19, 285–300.
Trivers, R. (1972). Parental investment and sexual selection. In B. B. Campbell (Ed.), Sex-ual selection and the descent of man (pp. 136–179). Chicago: Aldine.
Tucker, M. L. (1989). Anger and aggression in early adolescence: The influence of hor-mones and social context. Dissertation Abstracts International, 50(2), 761B. (UMINo. 8821665)
Vaillancourt, T., Brendgen, M., Boivin, M., & Tremblay, R. E. (2003). Longitudinal con-firmatory factor analysis of indirect and physical aggression: Evidence of two fac-tors over time? Child Development, 74, 1628–1638.
Vaillancourt, T., Côté, S., Farhat, A., Leblanc, J. C., Boivin, M., & Tremblay, R. E.(2004). The development of indirect aggression among Canadian children. Manu-script submitted for publication.
Weisfeld, G. (1994). Aggression and dominance in the social world of boys and girls. In J.Archer (Ed.), Male violence (pp. 42–69). London and New York: Routledge.
Sex Differences in Aggressive Behavior 443

This page intentionally left blank

Part IV
CHALLENGES FORTHE FUTURE

This page intentionally left blank

CHALLENGES FOR THE FUTUREWhere Are We Going?
21
The Developmental Originsof AggressionWhere Are We Going?
RICHARD E. TREMBLAY and SYLVANA CÔTÉ
The aim of this book was to review the state of knowledge on the develop-mental origins of aggression. Contributions were divided in two categories:Chapters 2–10 describe different types of aggressive behavior and the changesthat occur as individuals increase in age; Chapters 11–20 review factors oftenconsidered to be proximal and distal determinants of aggressive behavior. Likeall attempts to carve nature at its joints, separating the development of aggres-sive behavior from putative determinants is a perilous enterprise, especiallywhen nature has not yet been precisely described. The aim of this final chapteris to summarize where we come from, where we stand, and where we appearto be going with reference to selected key issues.
As we planned this book, it became clear that we were following a pathtraced by another book published 30 years earlier (De Wit & Hartup, 1974).In Chapter 1 of this volume, Willard W. Hartup, an editor of the previousbook, highlights three important changes that have occurred over these pastthree decades. First, investigators shifted their attention from the aggressiveact to the aggressive individual. Second, the research became truly develop-mental; that is, longitudinal data were collected, enabling investigators to lookat changes over time within individuals. Third, investigators increasingly ag-gregated the concepts of aggression and antisocial behavior. In this chapter, weadd four more changes that we believe are important to future research on ag-gression: More studies are focused on aggression during the first 3 years afterbirth; there is an explosion of studies on the biological bases of human aggres-
447

sion; we have started to systematically study gene–environment interactions;and preventive interventions are being used as experiments to test causal fac-tors. Six of these seven changes should mark the next few decades of research,because they are essential for the unraveling of the developmental origins ofaggression. However, one of the seven changes (aggregating aggression andantisocial behavior) may be a serious handicap in achieving this task.
CHANGES IN AGGRESSIVE ACTS OVER TIME WITHIN INDIVIDUALS
Two of the three changes Hartup highlights in Chapter 1 go hand in hand. Ifwe are studying aggressive behavior, the aggressive act must obviously remainthe basic phenomenon of study; however, understanding development impliesa focus on changes over time within individuals. Here are examples of devel-opmental questions that target the aggressive acts of individuals over time:When, during an individual’s life course, do different types of aggressive actsstart? Does the frequency of these acts change over the life course? Do the vic-tims of these acts change during the life course? Do the determinants of theseacts change during the life course? Do the consequences of these acts changeduring the life course? How and when, during the life course, can we changethese developmental trajectories?
The traditional approach to answering these questions was a cross-sectional design, that is, comparing samples of individuals of different ages. Inthe De Wit and Hartup (1974) book there are two very good examples of this(Hapkiewicz, 1974; Hoving, LaForme, & Wallace, 1974). Note that Hap-kiewicz’s chapter has a perfect developmental title: “Developmental Patternsof Aggression.” But there are many more recent examples, such as the follow-ing chapters in other books: “The Development of Direct and Indirect Aggres-sive Strategies in Males and Females” (Björkqvist, Österman, & Kaukiainen,1992) and “Do Children in Canada Become More Aggressive as They Ap-proach Adolescence?” (Tremblay et al., 1996).
We now have longitudinal data to study intra-individual changes in ag-gression, and these studies are reviewed in the first 10 chapters of this volume.However, many, if not most of the studies on the determinants of aggression(Chapters 11–20) provide little developmental information on aggression, be-cause the aggressive acts (the dependent variable) to be explained by one ormany determinants (independent variables) tend to be assessed at only onepoint in time. Many longitudinal studies are used to conduct cross-sectionalanalyses, or predictive analyses, that is, assessing the strength of the correla-tion between independent variables and aggression either at the same point orat two different points in time. For example, see Chapter 11 on genetics andaggression (Pérusse & Gendreau), and count how many of the cited studiesused the within-individual development of aggressive behavior as the pheno-type to be explained by genes and environment. Studies on intra-individual
448 C H A L L E N G E S F O R T H E F U T U R E

change in playful aggression (Chapter 7, by Peterson & Flanders), indirectaggression (Chapter 8, by Vaillancourt) and proactive–reactive aggression(Chapter 9, by Vitaro & Brendgen) are either nonexistent or extremely rare.Most of the studies attempt to find causes of a phenotype measured at onepoint in time, and hence not the development of a behavior over time. Themaintenance of a cross-sectional approach is most evident in the data avail-able for the meta-analyses of sex differences in aggression presented in Chap-ter 20 by Archer and Côté.
It is tempting to conclude that studying the development of a phenome-non does not come naturally to humans. This is best exemplified by the behav-ior of many of the young, and not so young, investigators who come to workon our longitudinal data sets. They say they are attracted by the richness ofthe longitudinal data that we have collected. They spend weeks, and oftenmonths, reviewing the literature, examining the data that was collected, andcome up with questions such as, “Do obstetrical complications predict violentbehavior in late adolescence?” “Does behavior in kindergarten predict violentbehavior in early adulthood?” “Do different types of aggression in early ado-lescence predict equally well spouse aggression?” Each of these questions hassome theoretical and practical value. Each necessitates longitudinal data thatspan decades. But they are not asking one of the developmental questionsasked at the start of this section.
The preoccupation is with prediction from time X to time Y, rather thandescribing the development of a phenotype and identifying the mechanismsthat unfold over time. Humans are in a hurry to know—“can’t wait for 20years to confirm a theory.” From a developmental science perspective, show-ing that characteristics at time X predict characteristics at time Y is as simplis-tic as showing that one, or an array of genes predict behavior at a given pointin time. Selecting two points in time from a data bank that includes repeatedassessments over long periods of time is similar to dichotomizing a continuousmeasurement scale; there is a huge loss of information. By the same analogy,the important developmental questions are concerned with the pattern of thedistribution of the scores, not by the creation of two groups that are some-what different. Taking a developmental perspective means, first, to under-stand the development (change over time) of the phenomenon we are trying topredict, and second, to describe the causal chain of events over time that ex-plain the development. However, a premium is generally given to studies thataim at testing causal theories (that is: testing the strength of the correlation be-tween a “determinant” and an “outcome”). Although there is a need for stud-ies with such objectives, the need for studies that describe the development ofa phenomenon over time is obviously a prerequisite for understanding thecause of the development. Those who tend to value theory testing more thandescriptions should look back to the revolutions created by the simple descrip-tions of Copernicus, Galileo, and Mendelyev (see also Tinbergen, 1963).
Galileo certainly did not find it easy to convince his colleagues that Co-
Where Are We Going? 449

pernicus’s description of the solar planet trajectories was closer to the truththan the trajectories their theories led them to imagine, notwithstanding whoor what caused the phenomenon. As the longitudinal data become availableand the next generation of investigators come into their prime, the traditionsshould change. The research efforts aimed at explaining behavior develop-ment should be accompanied by efforts aimed at describing that development,ideally over the life course. Unfortunately, the best designed and longest run-ning longitudinal studies with birth cohorts apparently did not obtain re-peated assessments of aggressive behaviors between birth and school entry(e.g., Fergusson & Horwood, 1995; Moffitt, Caspi, Dickson, Silva, &Stanton, 1996; Power, Manor, & Fox, 1991; Raine et al., 2001). Thus, to de-scribe the life course development of different forms of aggressive behavior,we will have to count on new birth cohorts. This means that complete lifecourse data will become available in not less than 90 years! Someone shouldstart soon, concentrate on collecting as much data as possible over the earlyyears, be persistent, and be prepared to train many generations of investiga-tors. This is the price we have to pay to make serious claims of conducting“life course” research on human development.
CLASSIFICATION: THE AGGRESSIVE AND ANTISOCIAL BEHAVIOR CASE
Chapter 2 by Gendreau and Archer shows the complexity of creating a classi-fication of aggressive behaviors, the basic building block of a science. This isnot a new problem (see e.g., De Wit & Hartup, 1974), and it is a problem thatall sciences have to handle with care (Appel, 1987; Burkhardt & Smith,1988). From that perspective, we strongly believe that the tendency over thepast 30 years to aggregate aggressive and antisocial behaviors (see Hartup,Chapter 1, this volume) is a serious mistake.
First, from an evolutionary perspective, aggression is an adaptive behav-ior. It is a tool that individuals learn to control during their development. Forexample, playful aggression is a natural behavior of nonhuman (e.g., juvenilerats, cats, dogs, monkeys) and human animals (see Pellis, Pellis, & Foroud,Chapter 3, and Petersen & Flanders, Chapter 7, this volume). Adult humanscontinue to play fight through physically aggressive games such as boxing,wrestling, rugby, American football, and ice hockey. Humans also produceand consume a huge amount of fictive violence in books, plays, electronicgames, and movies. Many humans learn to master different complex forms ofphysical aggression for defensive purposes, for example, by training in martialarts, by training in the art of boxing, or by learning to use weapons such asguns and rifles. The practice of aggregating aggression and antisocial behaviorthat appeared in the research over the past 30 years (see Hartup, Chapter 1,this volume; Coie & Dodge, 1998; Tremblay, 2000) probably reflects the am-bivalent relationship humans have with aggression. For example, there havebeen strong movements in the past 30 years to prevent children from playing
450 C H A L L E N G E S F O R T H E F U T U R E

aggressive games (e.g., parents should not buy toy guns) and to protect chil-dren from aggressive contents in books, movies, and video games. It is arguedthat these activities train them to become “antisocial” (e.g., Anderson et al.,2003; Jones, 2002). Meanwhile, violent movies and violent video games havethe biggest commercial success, games such as hockey are said to have becomemore violent than ever, thousands of young men are trained for preemptivewars, and we see, every day on television, the thousands of people violentlykilled during these wars. These are certainly not the most noble of human be-haviors; however, they should not be aggregated with “antisocial behaviors”that are defined as law-breaking behaviors by criminologists and as mental ill-nesses by psychiatrists. Similarly, indirect aggressions (see Vaillancourt, Chap-ter 8, this volume) manifested by elementary school girls or university profes-sors are aggressions, but they should not be confused with the “antisocialbehaviors” that are of interest to criminologists, psychiatrists, and clinicalpsychologists.
Second, if not all aggressions are antisocial, it is also true that not all anti-social behaviors are aggressions. For example, in their review of the state ofknowledge on aggression, Coie and Dodge (1998) include behaviors such assubstance abuse and risky sexual behavior in their definition of antisocial be-havior. It seems a far stretch to label these “aggressive.” Many of the chaptersin this book had to rely on studies of the general “antisocial behavior” con-cept, because there were not enough studies that specifically targeted aggres-sive behavior. One would expect that age of onset and determinants of verbaland indirect aggression trajectories would be different from onset and deter-minants of antisocial behaviors described as smoking marijuana (substanceabuse) and not wearing a condom during sexual intercourse (a risky sexualbehavior) (Coie & Dodge, 1998).
We believe that instead of aggregating aggressive behaviors with antiso-cial behaviors, we need to go in the opposite direction if we are to understandthe developmental origins of aggression. We need to disaggregate aggressivebehaviors. Many studies over the past few years have targeted subcategoriesof aggression, such as physical aggression (see Tremblay & Nagin, Chapter 5,this volume), indirect aggression (see Vaillancourt, Chapter 8, this volume),playful aggression (see Pellis, Pellis, & Foroud, Chapter 3, and Petersen &Flanders, Chapter 7, this volume), and proactive and reactive aggression (seeVitaro & Brendgen, Chapter 9, this volume). We predict that in the long run,the “splitters,” those who disaggregate the components of aggressive behav-iors, will make a more important contribution to understanding the develop-mental origins of aggression, than the “lumpers,” those who aggregate manyforms of aggression and a large variety of antisocial behavior. The success ofthe physical and biological sciences has been strongly linked to the “splitters”approach. For instance, from measuring the circumference of the skull in thenineteenth century (Lombroso, 1896), investigators moved on to identifyingthe large components of the brain (MacLean, 1949) in the middle of the 20thcentury. We are now able to precisely describe the lifespan development of
Where Are We Going? 451

every part of an individual’s brain without using a scalpel (see Paus, Chapter12, this volume), and we are discovering the importance of prions (Si et al.,2003). The work of Eric Kandel, the senior collaborator of the last cited arti-cle, is probably one of the best examples of the scientific value of “reduction-ism” (Kandel, 2000). The young psychoanalyst decided that the human andnonhuman primate brains were too complex to understand. He chose to studyhow a sea slug’s brain (Aplysia) learns. His reductionist work over four de-cades revealed how memory is created in the brain of a mollusk, a rat, and ahuman. Without doubt, Sigmund Freud, the young biologist, would be proudof the accomplishment of his Vienna-born colleague. The development of dif-ferent forms of aggressive behavior appears much less complex than the devel-opment of the human brain. We simply need more investigators who willpatiently observe and describe this development.
AGGRESSION DURING EARLY CHILDHOOD
Aristotle starts the second chapter of Book I of Politics with the following sen-tence: “He who considers things in their first growth and origin, whether astate or anything else, will obtain the clearest view of them” (Aristotle, 1943,p. 248). How obvious! It has been repeated many times since then, once byHamburg and van Lawick-Goodall in a chapter in De Wit and Hartup (1974):“There is a great need for direct studies, utilizing systematic observationaltechniques, of aggressive behavior and its precursors in human infancy andearly childhood” (p. 72). Out of 45 chapters in that book, only three providedsome information on preschool children. There were a number of importantstudies on preschool children in the 1920s and 1930s (see Hay, Chapter 6, andTremblay & Nagin, Chapter 5, this volume). There was also a wave of thesestudies in the 1970s and 1980s, apparently stimulated by the ethological workwith primates (e.g., Blurton Jones, 1972; McGrew, 1972; Restoin et al., 1985;Smith, 1974; Strayer & Trudel, 1984), but they were cross-sectional. Thereare still occasional observational studies on aggression in early childhood (seeHay, Chapter 6, this volume). But the vast majority of studies on the develop-ment of aggression in humans were, and still are carried out with individualsin either elementary or secondary schools.
One reason why few longitudinal preschool studies have been conducted iscertainly because they are exceptionally difficult to do. The tradition has been touse direct observations—methods of data collection that are time-consumingboth at the data collection and the data analysis levels, even for very small sam-ples. Yet, there are good methodological reasons for choosing to study school-age children: (1) They are easy to find, because they all have to go to school, (2)representative samples of populations can be easily created, and (3) reports ofaggressive behaviors can be obtained from different sources—teachers, parents,peers, self-reports. However, the methodological challenges of studying pre-schoolers should not be a sufficient cause for their paucity. Recent epidemiologi-
452 C H A L L E N G E S F O R T H E F U T U R E

cal studies with very large representative cohorts of newborns have shown thatthe methods used with older children (i.e., parent and educator reports on chil-dren’s behavior) can provide useful information on the developmental origins ofaggression during early childhood (e.g., Côté, Vaillancourt, LeBlanc, Nagin, &Tremblay, 2004; Dionne, Tremblay, Boivin, Laplante, & Pérusse, 2003; King-ston & Prior, 1995; Tremblay et al., 2004b).
A second reason for neglecting early childhood aggression over the past30 years may have been the popularity of the social learning theory (Bandura,1973). It is probably not an exaggeration to say that most of the research onthe development of human aggression in the last few decades of the 20th cen-tury was strongly inspired by the social learning paradigm. Although Bandurahad studied aggressive behavior in nursery schoolchildren, most of the longi-tudinal studies initiated with a social learning perspective were attempting tofind how schoolchildren learn to aggress from their environment. Parents,peers, movies, and television were the obvious models (e.g., Cairns & Cairns,1994; Farrington, 2003; Huesmann & Miller, 1994; Lefkowitz, Eron, Walder,& Huesmann, 1977; Loeber, Farrington, Stouthamer-Loeber, & Van Kammen,1998; Patterson, 1982; Thornberry, 1998; Tremblay, Vitaro, Nagin, Pagani,& Séguin, 2003).
A third reason for neglecting early childhood aggression is possibly re-lated to funding. Research priorities have been and will remain dictated bypolicy issues. Each time an adolescent commits a homicide, the pressure in-creases to understand the causes of violence during adolescence and to findways of helping or restraining at-risk adolescents. It is not easy to convinceagencies and peers that a study of infants’ aggression is the answer to under-standing, and preventing, violence during adolescence. Thus, most funding ofresearch on violence is channeled toward studies of individuals who are bigenough to hurt others seriously when they aggress.
The increasing evidence that control over physically violent behavior ismostly learned by humans during early childhood (see Tremblay & Nagin,Chapter 5, this volume) should help convince policymakers and funding agen-cies that research on early childhood aggression could help our understandingof the developmental origins of aggression and eventually prevent the violentbehaviors of adolescents who scare the population. However, if there were asudden increase in funds to study aggression during early childhood, therewould be a receptor problem, inasmuch as there are very few investigatorswith the necessary training. Fortunately, there are examples indicating that in-vestigators working on violence during adolescence or adulthood can becomeinterested in studying aggression in the early years. In fact, the investigators ofaggression needed for the future are individuals who focus on the develop-ment of aggression throughout the lifespan. The development of human be-havior cannot be understood if it is cut into investigator-convenient age slices.
The problem with direct observation, discussed earlier, will probably besolved in the near future by multidisciplinary teams that will include this time-consuming but important approach within a global strategy. For example,
Where Are We Going? 453

large population cohorts of children can be studied using parent ratings, whilesubsamples are strategically selected for intensive direct observations of socialinteractions, paired with key biological data collections such as saliva samplesand brain scans (see Paus, Chapter 12, Pihl & Benkelfat, Chapter 13, and VanGoozen, Chapter 14, this volume).
BIOPSYCHOSOCIAL STUDIES OF AGGRESSION
The great divide between social and biological approaches to human behavioris probably nowhere more evident than in the research on aggression over thepast three decades. The De Wit & Hartup (1974) book showed a preoccupa-tion with considering both sides of the question. But each article was con-cerned with either one or the other perspective. Numerous books on aggres-sion and violence during the past decades omitted biological issues, and bookson the biological bases of aggression did not include psychosocial factors.Most investigators preoccupied with the social “causes” find it difficult tolearn the biological jargon. In fact, many resist considering the hypothesis thatviolence could in some ways be influenced by biological factors. For example,in the not so distant past, during the planning phase of one of the most expen-sive longitudinal studies of child development and antisocial behavior in theUnited States, plans for biological assessment apparently had to go under-ground (for the original plan, see Chapter 15 and Appendix II in Tonry et al.,1991). The National Consortium on Violence Research, another very largeenterprise of the 1990s, funded by the U.S. National Science Foundation, wasessentially focused on the social factors. Similarly, those who focused on thebiological aspects of aggression generally did not take into account social fac-tors.
Considering these parallel trajectories, the changes that are occurring areimpressive. The 1996 NATO Advanced Study Institute on the Biosocial Basesof Violence represented an important step in that direction (Raine, Farrington,Brennan, & Mednick, 1997). Compare the number of biological studies onhumans reviewed in this book (especially in Chapter 12, by Paus; Chapter 11,by Pérusse & Gendreau; Chapter 13 by Pihl & Benkelfat; and Chapter 14, byVan Goozen) with those in the De Wit and Hartup (1974) book. The ideaswere present in 1974. An example that strongly influenced our work comesfrom Hamburg and van Lawick-Goodall (1974): “We are inclined to believethat the hormonal changes of puberty, especially in males, may facilitate thelearning of aggressive behavior” (p. 76) (see Van Goozen, Chapter 14, fortests of this hypothesis). The increasing number of biopsychosocial studies ap-pears largely due to the development of biological measurement techniquesthat are relatively easy to use with humans. This is the case with molecular ge-netics, brain scans, and radio immunoassays of saliva. This methodologicalprogress will continue, but biopsychosocial studies are still a small part of thenumerous studies on human aggressive-antisocial behaviors.
454 C H A L L E N G E S F O R T H E F U T U R E

The best incentive for behavioral and social scientists to include the bio-logical components is probably the increasingly clear demonstration that theenvironment, both physical and social, has an important impact on the devel-oping organism and that, in turn, these biological consequences have a long-term impact on social behavior (Francis, Diorio, Plotsky, & Meaney, 2002;Keating & Hertzman, 1999; Meaney, 2001; Weaver et al., 2004). Smokingduring pregnancy is a very good example. Children of mothers who smokedduring pregnancy are at higher risk of numerous problems, including lowbirth weight, hypertension, hyperactivity, inattentiveness, impulsiveness, andaggression (Tremblay, Barr, & Peters, 2004a; Pausova, Paus, Sedova, &Bérubé, 2003; Tremblay et al., 2004b; Wakschlag, Pickett, Cook, Benowitz, &Leventhal, 2002). At first sight, this is the domain of biologists, however, asecond look shows that women who smoke during pregnancy tend to havelow education, to be poor, to have a history of problem behaviors, to have amate who has similar problems, or to be a single parent already during preg-nancy. To what extent are the effects of smoking during pregnancy on aggres-sion explained by these social characteristics or, vice versa, the effects of thesocial characteristics explained by the effect of the nicotine on the developingcentral nervous system? There is no way of knowing if the social scientist con-ducts a study without measuring smoking while the biologist is doing a paral-lel study and does not measure the history of antisocial behavior of the par-ents. The best alternative for the advancement of knowledge is for thebiologist and the social scientist to use the same subjects and to do analysesthat take into account the biological and social variables. This is actually hap-pening, and the teams that take these multidisciplinary approaches are flour-ishing.
The next step is to realize that if smoking during pregnancy is injuringbrain development, a biological phenomenon, the preventive solutions aremostly in the domain of the psychosocial specialists. How do we get womenwith a history of problem behaviors to stop smoking during pregnancy? Thisleads back to our comments earlier concerning the importance of early child-hood in understanding the development of aggressive behavior. If early in de-velopment the social environment has an important impact on biological de-velopment, and this biological change has a long-term impact on socialbehavior development, it becomes still more obvious that we need to followAristotle’s advice: Start at the beginning if you want to understand the devel-opmental origins of aggression.
GENE–ENVIRONMENT INTERACTIONS
The gene–environment interactions (G×E) issue is but a sub-section of the pre-vious discussion. In the De Wit & Hartup (1974) book there were two chap-ters on genetics. One was a summary of an impressive master’s degree thesison the criminality of adopted children in Denmark (Hutchings, 1974). The
Where Are We Going? 455

second was a comparative study of agonistic behavior in different strains ofmice (Stewart, 1974). There were no twin studies and no molecular geneticstudies. The review by Pérusse and Gendreau in Chapter 11 of this volumeshows the impact of the genetic revolution over the past two decades. A newgeneration of social scientists can hear the word genes associated with humanbehavior without becoming inflamed and referring to eugenics. Many “psy-chosocial” investigators are recognizing that genetic information can be usedas a control variable to study more adequately environmental effects. Duringthe 1980s and 1990s, David Rowe made almost yearly presentations at theannual meeting of the American Society of Criminology, aimed at convincingcriminologists that controlling for genetic factors through twin, sibling, andadoption designs was important for identifying environment effects, especiallyfamily effects, on antisocial behavior (see Chapter 15 by Rowe in Tonry et al.,1991).
An important number of social scientists have moved much beyond whatRowe would have expected before his untimely death. It is obvious that mostinvestigators with rich longitudinal data sets on human development need tocollect genetic information on their subjects to add to the huge amount of datathey have collected on behavior and the environment. This will lead to an im-portant increase in G×E papers. It should eventually become next to impossi-ble to publish a paper on human development in a top scientific journal with-out having a wide range of information on genes and environment. In thesame way that we routinely control for socioeconomic status (SES), control-ling for specific genetic polymorphism will most probably become standardpractice. This will obviously not simplify our task. But it will lead to sounderknowledge, once we have learned to deal with the idea of G×E in our hypothe-ses and our analyses.
Concerning the developmental origins of aggression, we must repeat thatthe first problem we need to solve is not the measurement of genes, nor themeasurement of the environment, but the measurement of the phenotype. It isunlikely that there is a simple G×E leading to any expression of aggression atany point in time, or even multiple points in time, during the life course. Thestudies reviewed by Pérusse and Gendreau in Chapter 11 of this volumeclearly used a hotchpotch of assessments at various ages, with various popula-tions, to identify the “aggression” phenotype. We are far from a G×E scienceof the development of aggressive behavior in humans, but we are much closerthan we were 10 years ago. Fortunately, we can count on animal studies, ascan be seen in Chapter 4 of this volume, by Suomi.
TOWARD PSYCHOSOCIAL EPIGENOMICS
One of the important changes of the past three decades has been the increas-ing awareness of the need for experimental preventive interventions (e.g.,Dishion, McCord, & Poulin, 1999; Farrington, Ohlin, & Wilson, 1986;
456 C H A L L E N G E S F O R T H E F U T U R E

McCord, & Tremblay, 1992; Tremblay et al., 1999a; Tremblay, 2003). De Wit andHartup (1974) did not include any experimental, or even quasi-experimental,intervention studies. Two studies made use of juvenile delinquents as subjects(Parke, 1974; Leyens & Camino, 1974), and Patterson (1974) presented ananalysis of a boy referred to treatment for a high rate of “noxious” behavior.However, in their concluding chapter, De Wit and Hartup (1974) made an in-teresting set of comments on how children learn to regulate aggression. Theysuggested that two processes are involved: “a) moment-to-moment control ofthe ongoing actions of the child, and b) training the child for long-term self-control.” These processes can be linked to research domains covered in thisvolume: Chapter 17 on family factors (Zoccolillo et al.) and Chapters 12(Paus), 16 (Dionne), and 15 (Séguin & Zelazo), respectively, on brain develop-ment, language development, and development of executive functions.
Research on parent training and children’s social skills training blos-somed during the past three decades; however, judging from the reviews inthis book, these efforts did not generate clear knowledge on the developmen-tal origins of aggression. There appear to be four main reasons for this situa-tion. First, children who were the targets of the interventions were generallyschool-age children, well past the “origins” phase. The study by Gross et al.(2003) is a notable exception. Second, rather than targeting aggression, inter-vention programs targeted the general concepts of aversive, externalizing, orobnoxious behavior (e.g., Patterson, 1982; Wahler, 1987; Webster-Stratton,1998). Third, interventions showing long-term effects (more than a few years)were multimodal, precluding the attribution of the effect to a specific compo-nent (e.g., Lacourse et al., 2002; Olds et al., 1998). Fourth, the vast majorityof these studies were not designed to test developmental hypotheses concern-ing aggression and did not control for most of the other potential causal ef-fects. Chapter 17, by Zoccolillo et al., in this book describes the many com-peting explanations for a trajectory of chronic physical aggression: forexample, genetic factors, parents’ history of problem behaviors, parents’ edu-cation, smoking during pregnancy, parental responsiveness to the child, coer-cive parenting, abuse and neglect. Whether parenting skills are determinantstill needs to be demonstrated by controlling for alternative hypotheses, in-cluding genetic and perinatal effects (see Harris, 1998; Raine, 2002; Rowe,1994).
In regard to the gene–environment interaction, Caspi et al. (2002) re-ported a significant G×E for violent behavior, using assessments of child mal-treatment and a MAOA genetic polymorphism. This is one good indicationthat studies that do not take into account both genetic and environmental fac-tors are unlikely to provide adequate information on causal factors. However,we often forget that G×E studies, even with longitudinal data, are simplycorrelational studies if there is no manipulation of genes or environments.Correlational studies cannot prove causal effects. Hence, G×E studies of thedevelopmental origins of aggression with large-population birth cohorts willsimply give indications of possible causal factors. The ultimate proofs of cau-
Where Are We Going? 457

sality must come from experiments in which potential causal factors are ma-nipulated.
Most probably, we are far from genetic manipulations studies to test theireffects on the developmental trajectories of human aggression. Manipulationsof environments, however, are the daily business of thousands of people paidto help families and individuals with behavior problems or who are at highrisk of having these problems. Governments are investing billions in thesesocial programs, without much evidence that they are having the intended ef-fects (e.g., McCord, 2003; Timmins, 2001; Tremblay, LeMarquand, & Vitaro,1999b). The next decade should see a substantial increase in experimental ma-nipulations of environments to test for G×E.
This may sound far-fetched to some, but it is not much different fromgenotyping subjects in a longitudinal study. One of the new fields riding onthe crest of the genetic revolution is pharmacogenomics. Chemists create med-ications that compensate for genetic deficits or take into account genetic dif-ferences. These chemical products act at the protein level, but they are envi-ronmental products and thus constitute an environmental manipulation. So isnutrition; special diets are given to children born with a multitude of geneticdefects (e.g., Scriver & Kaufman, 2001). The aim of these diets is either togive the children proteins they are lacking because of the genetic defect or toprevent them from eating nutrients that could harm their development be-cause of a genetic deficit. Such a diet may be needed throughout the individ-ual’s life, or for only a limited time during a critical period for the growth of aphysiological structure.
We can generalize this approach by thinking about parenting as a “nutri-ent.” The results of the longitudinal birth cohort study in New Zealand re-ported by Caspi et al. (2002; see also, Pérusse & Gendreau, Chapter 11, thisvolume) indicates that boys maltreated by their parents (inadequate nutrient)are more or less at risk of becoming chronically aggressive, depending onwhich of two MAOA genotypes (high or low activity) they inherited. A practi-cal implication of these results is that, because maltreated boys with the high-activity MAOA polymorphism appear “genetically” protected from chronicphysical aggression, preventive interventions should specifically target boyswithout that protective factor. The parents of these at-risk boys should receiveparent training, as early as possible, to prevent maltreatment. The Elmirastudy (Olds et al., 1998) has shown that a nurse home visitation programcould prevent both child abuse by parents and delinquency when the childrenbecome adolescents. Such effects should be observed mainly for the maleswithout the MAOA protective polymorphism. Thus, an experimental preven-tive intervention that targets quality of parenting, for boys who lack the pro-tective genetic factor, would not only demonstrate the cost-effectiveness ofparenting interventions based on genetic screening, but would also provide anexperimental demonstration of the causal nature of the statistical interactionshown in correlational evidence. With psychosocial interventions, we can con-
458 C H A L L E N G E S F O R T H E F U T U R E

duct psycho-socio-genomic experiments, but the most exciting challenge ispsycho-socio-epigenomics. There is increasing evidence that gene expression isturned on and off by environmental factors (Szyf, 2003). For example, a re-cent study of maternal behavior with rat pups shows that increased lickingand grooming, as well as arched-back nursing altered rat pups’ DNAmethylation (Weaver et at, 2004). These results suggest that environment ma-nipulation not only can compensate for genetic defects but can also turn on oroff gene expression at different points in time during the developmental pro-cess.
In the not too distant future we should see numerous preventive experi-ments that will collect genetic information to test the cost-effectiveness of theintervention and, at the same time, test G×E hypotheses. However, we can im-mediately go back to subjects in successful interventions, such as the Elmirastudy, and genotype them to verify whether indeed the subjects who mostbenefited from that intervention had the low-activity MAOA genotype or anyother genetic risk. Unfortunately, such analyses would probably not provideuseful information on the developmental origin of early aggression, inasmuchas the published work from the Elmira study indicates that early aggressiondata were not collected. This leads us to underscore the fact that preventionexperiments with a G×E design need to specifically target the period frompregnancy to school entry, if we are to understand the developmental originsof aggression. These early studies will also be extremely useful to test the bio-logical embedding hypothesis (Keating & Hertzman, 1999) by studying themediating role of endophenotypes such as brain development (see Paus, Chap-ter 12, this volume) and HPA activity (see Van Goozen, Chapter 14, this vol-ume). Similar studies can also be undertaken to test other putative environ-mental causes of chronic aggression, such as smoking during pregnancy (seeWakschlag et al., 2002), influences of peers (see Boivin, Vitaro, & Poulin,Chapter 18, this volume), and television (see Johnson, Cohen, Smailes, Kasen,& Brook, 2002). In fact, we should be testing within the same studies the dif-ferential impact of parents, peers, and fictive aggression (on television, inmovies, and in video games).
CONCLUSION
Planning what is needed to take us from where we are to where we wouldlike to be forces us, first, to take stock of what has been done; second, toidentify what has not been done; third, to figure out what else needs to bedone; and finally, to deal with the whirl of new ideas stimulated by theplanning process. As we glance at this feast of new research within ourreach, we can imagine how excited a clone of Charley Darwin would be tocontribute to the next 50 years of research on the developmental origins ofaggression.
Where Are We Going? 459

12 August 1849
To M. A. T. Whitby
My dear Madam,
I cannot express too strongly my thanks for the extraordinary troublewhich you have taken in the interesting experiment, of which you send methe results.—I had given up all hopes of knowing whether peculiarities in thecaterpillar state were hereditary, but now the point is amply proved: there isindeed a wide difference between a probability, however high & such an ex-periment as you have made.—
. . . I dare not do more than hint my curiosity to know whether theFrales would prove hereditary,—i.e., whether it would be possible to make abreed with cocoons destitute of silk.—In the eyes of all silk-growers, thisassuredly would appear the most useless of experiments ever tryed.—(Darwin, 1832/1985, p. 248)
REFERENCES
Anderson, C. A., Berkowitz, L., Donnerstein, E., Huesmann, L. R., Johnson, J. D., Linz,D., Malamuth, N. M., & Wartella, E. (2003). The influence of media violence onyouth. Psychological Science in the Public Interest, 4(3), 81–110.
Appel, T. (1987). The Cuvier–Geoffroy debate: French biology in the decades beforeDarwin. New York: Oxford University Press.
Aristotle. (1943). On man and the universe. New York: Walter J. Black-Roslyn.Bandura, A. (1973). Aggression: A social learning analysis. New York: Holt.Björkqvist, K., Österman, K., & Kaukiainen, A. (1992). The development of direct and
indirect aggressive strategies in males and females. In K. Björkqvist & P. Niemelä(Eds.), Of mice and women: Aspects of female aggression (pp. 51–64). Toronto: Ac-ademic Press.
Blurton Jones, N. (1972). Characteristics of ethological studies of human behaviour. InN. Blurton Jones (Ed.), Ethological studies of child behaviour (pp. 3–33). NewYork: Cambridge University Press.
Cairns, R. B., & Cairns, B. D. (1994). Life lines and risks: Pathways of youth in our time.New York: Cambridge University Press.
Caspi, A., McClay, J., Moffitt, T., Mill, J., Martin, J., Craig, I. W., Taylor, A., & Poulton,R. (2002). Role of genotype in the cycle of violence in maltreated children. Science,297, 851–854.
Coie, J. D., & Dodge, K. A. (1998). Aggression and antisocial behavior. In W. Damon &N. Eisenberg (Eds.), Handbook of child psychology: Social, emotional, and person-ality development. (Vol. 3, pp. 779–862). Toronto: Wiley.
Côté, S., Vaillancourt, T., LeBlanc, J., Nagin, D. S., & Tremblay, R. E. (2004). The devel-opment of physical aggression during childhood: A nation wide longitudinal studyof Canadian children. Manuscript submitted for publication.
Darwin, C. (1832/1985). The correspondence of Charles Darwin, Vol. 4. London: Cam-bridge University Press.
De Wit, J., & Hartup, W. W. (Eds.). (1974). Determinants and origins of aggressive be-havior. The Hague, Netherlands: Mouton.
460 C H A L L E N G E S F O R T H E F U T U R E

Dionne, G., Tremblay, R. E., Boivin, M., Laplante, D., & Pérusse, D. (2003). Physical ag-gression and expressive vocabulary in 19-month-old twins. Developmental Psy-chology, 39(2), 261–273.
Dishion, T. J., McCord, J., & Poulin, F. (1999). Iatrogenic effects in early adolescent in-terventions that aggregate peers. American Psychologist, 54(9), 755–764.
Farrington, D. (2003). Key results from the first forty years of the Cambridge study in de-linquent development. In T. P. Thornberry & M. D. Krohn (Eds.), Taking stock ofdelinquency: An overview of findings from contemporary longitudinal studies (pp.137–183). New York: Kluwer Academic/Plenum Press.
Farrington, D. P., Ohlin, L. E., & Wilson, J. Q. (1986). Understanding and controllingcrime: Towards a new research strategy. New York: Springer-Verlag.
Fergusson, D. M., & Horwood, L. J. (1995). Early disruptive behavior, IQ, and laterschool achievement and delinquent behavior. Journal of Abnormal Child Psychol-ogy, 23(2), 183–199.
Francis, D. D., Diorio, J., Plotsky, P. M., & Meaney, M. J. (2002). Environmental enrich-ment reverses the effects of maternal separation on stress reactivity. Journal of Neu-roscience, 22, 7840–7843.
Gross, D., Fogg, L., Webster-Stratton, C., Garvey, C., Julion, W., & Grady, J. (2003). Par-ent training of toddlers in day care in low-income urban communities. Journal ofConsulting and Clinical Psychology, 71(2), 261–278.
Hamburg, D. A., & van Lawick-Goodall, J. (1974). Factors facilitating development ofaggressive behavior in chimpanzees and humans. In J. De Wit & W. W. Hartup(Eds.), Determinants and origins of aggressive behavior (pp. 59–86). The Hague,Netherlands: Mouton.
Hapkiewicz, W. G. (1974). Developmental patterns of aggression. In J. De Wit & W. W.Hartup (Eds.), Determinants and origins of aggressive behavior (pp. 201–207). TheHague, Netherlands: Mouton.
Harris, J. R. (1998). The nurture assumption: Why children turn out the way they do.New York: Free Press.
Hoving, K. L., LaForme, G. L., & Wallace, J. R. (1974). The development of children’s in-terpersonal aggression in a competitive task. In J. De Wit & W. W. Hartup (Eds.),Determinants and origins of aggressive behavior (pp. 193–200). The Hague, Neth-erlands: Mouton.
Huesmann, L. R., & Miller, L. S. (1994). Long-term effects of repeated exposure to mediaviolence in childhood. In L. R. Huesmann (Ed.), Aggressive behavior: Current per-spectives (pp. 153–186). New York: Plenum Press.
Hutchings, B. (1974). Genetic factors in criminality. In J. De Wit & W. W. Hartup (Eds.),Determinants and origins of aggressive behavior (pp. 255–266). The Hague, Neth-erlands: Mouton.
Johnson, J. G., Cohen, P., Smailes, E. M., Kasen, S., & Brook, J. S. (2002). Televisionviewing and aggressive behavior during adolescence and adulthood. Science, 295,2468–2471.
Jones, G. (2002). Killing monsters: Why children need fantasy, super heroes, and make-believe violence. New York: Basic Books.
Kandel, E. R. (2000, December 8). The molecular biology of memory storage: A dialogbetween genes and synapses. Nobel lecture. Stockholm: Karolinska Instituet.
Keating, D. P., & Hertzman, C. (1999). Developmental health and the wealth of nations.New York: Guilford Press.
Kingston, L., & Prior, M. (1995). The development of patterns of stable, transient, and
Where Are We Going? 461

school-age onset aggressive behavior in young children. Journal of the AmericanAcademy of Child and Adolescent Psychiatry, 34, 348–358.
Lacourse, E., Côté, S., Nagin, D. S., Vitaro, F., Brendgen, M., & Tremblay, R. E. (2002).A longitudinal-experimental approach to testing theories of antisocial behavior de-velopment. Development and Psychopathology, 14, 909–924.
Lefkowitz, M. M., Eron, L. D., Walder, L. O., & Huesmann, L. R. (1977). Growing up tobe violent. A longitudinal study of the development of aggression. New York:Pergamon Press.
Leyens, J., & Camino, L. (1974). The effects of repeated exposure to film violence on ag-gression and social structures. In J. De Wit & W. W. Hartup (Eds.), Determinantsand origins of aggressive behavior (pp. 509–516). The Hague, Netherlands: Mou-ton.
Loeber, R., Farrington, D. P., Stouthamer-Loeber, M., & Van Kammen, W. B. (1998). An-tisocial behavior and mental health problems: Explanatory factors in childhoodand adolescence. Mahwah, NJ: Erlbaum.
Lombroso, C. (1896). L’umo delinquente (5th ed., Vols. 1, 2, & 3). Torino: Broca.MacLean, P. (1949). Psychosomatic disease and the “visceral brain”: Recent develop-
ments bearing on the Papez theory of emotion. Psychosomatic Medicine, 11, 338–353.
McCord, J. (2003). Cures that harm: Unanticipated outcomes of crime prevention pro-grams. Annals of the American Academy of Political and Social Science, 587, 16–30.
McCord, J., & Tremblay, R. E. (Eds.). (1992). Preventing antisocial behavior from birththrough adolescence. Experimental approaches. New York: Guilford Press.
McGrew, W. C. (1972). An ethological study of children’s behavior. New York: AcademicPress.
Meaney, M. J. (2001). Maternal care, gene expression, and the transmission of individualdifferences in stress reactivity across generations. Annual Review of Neuroscience,24, 1161–1192.
Moffitt, T. E., Caspi, A., Dickson, N., Silva, P. S., & Stanton, W. (1996). Childhood-onsetversus adolescent-onset antisocial conduct problems in males: Natural history fromages 3 to 18 years. Development and Psychopathology, 8(2), 399–424.
Olds, D., Henderson, C. R., Cole, R., Eckenrode, J., Kitzman, H., Luckey, D., Pettitt, L.,Sidora, K., Morris, P., & Powers, J. (1998). Long-term effects of nurse home visita-tion on children’s criminal and antisocial behavior: Fifteen-year follow-up of a ran-domized controlled trial. Journal of the American Medical Association, 280(14),1238–1244.
Parke, R. D. (1974). Some methodological problems and some future trends. In J. De Wit& W. W. Hartup (Eds.), Determinants and origins of aggressive behavior (pp. 499–508). The Hague, Netherlands: Mouton.
Patterson, G. R. (1974). Stimulus control in natural settings. In J. De Wit & W. W.Hartup (Eds.), Determinants and origins of aggressive behavior (pp. 391–400). TheHague, Netherlands: Mouton.
Patterson, G. R. (1982). A social learning approach to family intervention: III. Coercivefamily process. Eugene, OR: Castalia.
Pausova, Z., Paus, T., Sedova, L., & Bérubé, J. (2003). Prenatal exposure to nicotinemodifies kidney weight and blood pressure in genetically susceptible rats: A case ofgene–environment interaction. Kidney International, 64(3), 829–835.
Power, C., Manor, O., & Fox, J. (1991). Health and class: The early years. London:Chapman & Hall.
462 C H A L L E N G E S F O R T H E F U T U R E

Raine, A. (2002). Annotation: The role of prefrontal deficits, low autonomic arousal,and early health factors in the development of antisocial and aggressive behavior inchildren. Journal of Child Psychology and Psychiatry, 43, 417–434.
Raine, A., Farrington, D. P., Brennan, P., & Mednick, S. A. (Eds.). (1997). Biosocial basesof violence. New York: Plenum Press.
Raine, A., Venables, P. H., Dalais, C., Mellingen, K., Reynolds, C., & Mednick, S. A.(2001). Early educational and health enrichment at age 3–5 years is associated withincreased autonomic and central nervous system arousal and orienting at age 11years: Evidence from the Mauritius Child Health Project. Psychophysiology, 38(2),254–266.
Restoin, A., Montagner, H., Rodriguez, D., Girardot, J. J., Laurent, D., Kontar, F.,Ullmann, V., Casagrande, C., & Talpain, B. (1985). Chronologie des comporte-ments de communication et profils de comportement chez le jeune enfant. In R. E.Tremblay, M. A. Provost, & F. F. Strayer (Eds.), Ethologie et développement del’enfant (pp. 93–130). Paris: Editions Stock/Laurence Pernoud.
Rowe, D. C. (1994). The limits of family influence. Genes, experience, and behavior.New York: Guilford Press.
Scriver, C. R., & Kaufman, S. (2001). Hyperphenylalaninemia: Phenylalanine hydroxy-lase deficiency. In C. R. Scriver, A. L. Beaudet, W. S. Sly, & D. Valle (Eds.), The meta-bolic and molecular bases of inherited disease (pp. 1667–1724). New York:McGraw-Hill.
Si, K., Giustetto, M., Etkin, A., Hsu, R., Janisiewicz, A. M., Miniaci, M. C., Kim, J. H.,Zhu, H., & Kandel, E. R. (2003). A neuronal isoform of CPEB regulates local pro-tein synthesis and stabilizes synapse-specific long-term facilitation in aplysia. Cell,115(7), 893–904.
Smith, P. K. (1974). Aggression in a preschool playgroup: Effects of varying physical re-sources. In J. De Wit & W. W. Hartup (Eds.), Determinants and origins of aggres-sive behavior (pp. 97–105). The Hague, Netherlands: Mouton.
Stewart, J. M. (1974). Genetic and ontogenetic determinants of agonistic behavior in ani-mal societies: Prototypic experiments with mice. In J. De Wit & W. W. Hartup(Eds.), Determinants and origins of aggressive behavior (pp. 267–276). The Hague,Netherlands: Mouton.
Strayer, F. F., & Trudel, M. (1984). Developmental changes in the nature and function ofsocial dominance among young children. Ethology and Sociobiology, 5, 279–295.
Szyf, M. (2003). DNA methylation: Enzymology.In D. Cooper (Ed.), Nature encyclope-dia of the human genome (pp. 1–8). New York: Nature Publishing Group.
Thornberry, T. (1998). Membership in youth gangs and involvement in serious and vio-lent offending. In R. Loeber & D. P. Farrington (Eds.), Serious and violent juvenileoffenders: Risk factors and successful interventions (pp. 147–166). ThousandOaks, CA: Sage.
Timmins, N. (2001, August 27). Fact, not fashion in testing time for social policy: Gov-ernments are taking an interest in a pioneering think-tank’s “what works” approachto policy making. Financial Times, Comment and Analysis.
Tinbergen, N. (1963). On aims and methods of ethology. Zeitschrift fur Tierpsychologie,20, 410–433.
Tonry, M., Ohlin, L. E., Farrington, D. P., Adams, K., Earls, F., Rowe, D. C., Sampson, R.J., & Tremblay, R. E. (1991). Human development and criminal behavior: Newways of advancing knowledge. New York: Springer-Verlag.
Tremblay, R. E. (2000). The development of aggressive behaviour during childhood:
Where Are We Going? 463

What have we learned in the past century? International Journal of Behavioral De-velopment, 24(2), 129–141.
Tremblay, R. E. (2003). Why socialization fails: The case of chronic physical aggression.In B. B. Lahey, T. E. Moffitt, & A. Caspi (Eds.), Causes of conduct disorder and juve-nile delinquency (pp. 182–224). New York: Guilford Press.
Tremblay, R. E., Barr, R., & Peters, R. de V. (Eds.). (2004a). Tobacco and pregnancy. Ency-clopedia on early childhood development. Centre of Excellence for Early ChildhoodDevelopment. Retrieved October 2003, from http://www.excellence-earlychildhood.ca/theme.asp?id=1&lang=EN
Tremblay, R. E., Boulerice, B., Harden, P. W., McDuff, P., Pérusse, D., Pihl, R. O., &Zoccolillo, M. (1996). Do children in Canada become more aggressive as theyapproach adolescence? In Human Resources Development Canada and StatisticsCanada (Eds.), Growing up in Canada: National Longitudinal Survey of Childrenand Youth (pp. 127–137). Ottawa: Statistics Canada.
Tremblay, R. E., Japel, C., Pérusse, D., McDuff, P., Boivin, M., Zoccolillo, M., &Montplaisir, J. (1999a). The search for the age of “onset” of physical aggression:Rousseau and Bandura revisited. Criminal Behavior and Mental Health, 9, 8–23.
Tremblay, R. E., LeMarquand, D., & Vitaro, F. (1999b). The prevention of ODD andCD. In H. C. Quay & A. E. Hogan (Eds.), Handbook of disruptive behavior disor-ders (pp. 525–555). New York: Kluwer Academic/Plenum Press.
Tremblay, R. E., Nagin, D. S., Séguin, J. R., Zoccolillo, M., Zelazo, P., Boivin, M.,Pérusse, D., & Japel, C. (2004b). Physical aggression during early childhood: Tra-jectories and predictors. Pediatrics 114(1), e43–e50.
Tremblay, R. E., Vitaro, F., Nagin, D. S., Pagani, L., & Séguin, J. R. (2003). The MontrealLongitudinal and Experimental study: Rediscovering the power of descriptions. InT. Thornberry (Ed.), Taking stock of delinquency: An overview of findings fromcontemporary longitudinal studies (pp. 205–254). New York: Kluwer Academic/Plenum Press.
Wahler, R. G. (1987). Contingency management with oppositional children: Some criti-cal teaching issues for parents. In J. Wilson & G. C. Loury (Eds.), From children tocitizens: Vol. 3. Families, schools, and delinquency prevention (pp. 112–131). NewYork: Springer-Verlag.
Wakschlag, L., Pickett, K. E., Cook, E., Benowitz, N. L., & Leventhal, B. (2002). Mater-nal smoking during pregnancy and severe antisocial behavior in offspring: A review.American Journal of Public Health, 92(6), 966–974.
Weaver, I. C. G., Cervoni, N., Champagne, F. A., D’Alessio, A. C., Sharma, S., Shecke, J.R., Dymov, S., Szyf, M., & Meaney, M. J. (2004). Epigenetic programming by ma-ternal behavior. Nature Neuroscience, doi: 10.1038/nn1276.
Webster-Stratton, C. (1998). Preventing conduct problems in Head Start children. Jour-nal of Consulting and Clinical Psychology, 66(5), 715–730.
464 C H A L L E N G E S F O R T H E F U T U R E

IndexIndex
Index
“f” following a page number indicates a figure; “t” following a page number indicatesa table
Abstract play, 142–151Abuse. See MaltreatmentAcademic achievement
predictors of violence and, 210proactive and reactive aggression and,
187Active defense, 111–112Adjustment problems
peer rejection and, 380proactive and reactive aggression and,
186–188schools and, 407
Adolescence. See also Developmentage–violent crime curve and, 85–87taggression prior to, 87–90, 89fgender differences and, 285homicide and, 209indirect aggression and, 160–161, 163language development and, 338–340,
340–341neuromodulators and, 262–263, 271–
273overview, 98–101, 99fproactive and reactive aggression and,
185subtyping aggression and, 38testosterone and, 286–287violence during, 202
Adoption studies, 231–232Adrenocorticotropic (ACTH) hormone,
296. See also HormonesAdrogenic gonadal hormones, 284. See
also TestosteroneAdrogens, 288–290. See also Hormones
Adulthood, play fighting in rats during,50–51
Affective aggression, 37–38Affective disorders. See Mood disordersAfter-school care, 390–391. See also
DaycareAge differences, 160–161Age-graded theory, 209Age–violent crime curve
aggression prior to adolescence and, 87–90, 89f
overview, 85–87fAggression
aggressive competence, 125aggressive performance, 125anger as a cause of, 94antisocial behavior vs. prosocial aggres-
sion, 5–6association with language, 343–347brain mechanisms influences on, 36–38child characteristics and causes of, 14,
93during childhood, 87–90, 89f, 93–98,
97f, 430f–431, 452–454childhood-onset persistent, 353–370,
362t, 363t, 364tcognitive antecedent of, 31–33consequences of, 31–33defensive, 38, 111–112defining, 5, 108–110, 353–354development of, influences on, 38–39development of prior to age 6, 90–93f,
92fdevelopmental trajectory of, 7–8
465

Aggression (continued)direct, 34environment and causes of, 95, 96evolution and causes of, 95expression of, 30f, 33–34, 38–39friends as determinant of, 15–16, 17–18frustration–aggression model, 178–179game, 38genetic–developmental model and, 224–
227, 228–234, 230fgenetics and causes of, 12–13, 96–97,
100heritability of, 355–368, 362t, 363t,
364thostile, 31–33, 109, 183–184impulsive, 264indirect, 34, 158–171, 383, 437–439,
451individual differences and causes of, 96during infancy, 107, 108–110, 124–126instrumental, 31–33, 38, 109, 184, 381integrative model of, 39–41fintermale, 37–38language–aggression hypothesis, 331–
332language development and causes of, 96,
340–341motivational system and causes of, 94neocortex, influences on, 37neural circuitries, influences on, 37nonviolent, 5–6offensive, 37parental, 28parenting as determinant of, 14peer relationships and causes of, 15, 96personally motivated, 32–33physical, 83–85, 308–309pleasure-motivated, 32predicting, 449–450as a predictor for antisocial behavior, 5prefrontal cortex, influences on, 37pregnancy and causes of, 96in preschool age group, 90–93f, 92f,
430f–431, 452–454proactive, 40–41f, 179–197prosocial, 5–6, 33punishment as, 33rage-like, 37reactive, 36, 40–41f, 178–197, 381regulation of, 142–143relational, 6, 34, 159–160, 383, 437relief-motivated, 32risk factors for early development of,
121–124
serotonin metabolism and causes of,96
siblings and causes of, 97smoking and causes of, 97social aggression, 34, 159–160social learning hypothesis and causes of,
94social systems and causes of, 100socially motivated, 32–33studies of, 250–255, 252f, 254f, 448–
450subtypes of, 26–29, 30f, 31–42, 41f,
450–452taxonomies of, 27–29, 39–41fteasing, 38testosterone and causes of, 96during toddler years, 124–126, 430f–
431, 452–454verbal, 108, 436–437war as, 33, 85weaning, 28
Aggressive competence, 125Aggressive performance, 125Alcohol use. See also Substance use
molecular genetics and, 232, 233neuromodulators and, 262during pregnancy, 210in rhesus monkeys, 76
Allelic variations, 13Alternative reproductive strategies,
428Androgens, 285–286Anger. See also Reactive aggression
as a cause of aggression, 94defining aggression and, 354during infancy, 110overview, 179, 282peer relationships and, 381proactive and reactive aggression and,
188–189, 195in young children, 91–93f, 92f
Antecedents of aggressionintegrative model and, 40polythetic analysis and, 30fof proactive and reactive aggression,
194–196subtyping aggression based on, 34–
39Anthropomorphism, 26–27Antidepressants, 263Antisocial behavior. See also Homicide;
Violenceadoption studies on, 231–232aggression as a predictor for, 5
466 Index

cognitive skills and, 314–315cortisol and, 292–293daycare and, 406developmental pathway model of, 205f–
206evolutionary perspective of, 450–452executive function and, 310–312as a familial disorder, 356friendships and, 16future research on, 368–370intergenerational transmission of, 355–
368, 362t, 363t, 364tlanguage development and, 333, 346maltreatment and, 359–360in parents, 360–368, 362t, 363t,
364tpeer relationships and, 409prefrontal damage and, 148–149vs. prosocial aggression, 5–6school environment and, 407
Antisocial personality disorder, 37Anxiety
indirect aggression and, 167language development and, 341molecular genetics and, 233during pregnancy, 123proactive and reactive aggression and,
187, 188Appearance, 167Arousal, 268Attachment system
childhood-onset persistent aggression(CPA) and, 358–359
as a risk factor, 123–124Attention-deficit/hyperactivity disorder
(ADHD)DHEA/DHEAS and, 289–290executive function and, 311–312, 317–
318genetic factors and, 122language development and, 343–344neuromodulators and, 270physical aggression and, 308proactive and reactive aggression and,
187testosterone and, 289
Attentional control, 273–275Attribution biases pathway, 346Authority conflict pathway, 205f–206. See
also Developmental pathwayAutism, abstract play and, 148Autonomy, during infancy, 111–112Avoidance of conflict, 116–117. See also
Conflict
B
Behaviorchanges in, 12–19, 35–36evolutionary perspective of, 427–
428executive function and, 310–312, 320–
321, 322explanations of, 425language development and, 333, 336–
337, 339–340physical aggression and, 308–309relationship to brain mechanisms, 242–
243Behavioral control, 261Bilateral view of socialization, 13–18. See
also SocializationBiological indicators. See also Hormones
contextualism and, 18–19overview, 13, 298–299, 454–455proactive and reactive aggression and,
188–190Biopsychosocial research, 454–455Birth complications, 96. See also Preg-
nancyBoxing stance, across species, 26–27Boys. See also Gender differences
developmental trajectory of aggressionin, 7–8
in play fighting in rats, 49–50proactive and reactive aggression and,
196use of indirect aggression by, 161–
162Brain mapping. See also Brain mechanisms
overview, 242–243, 255–256principles of, 243–246, 244f, 245f,
246fstudies of aggression using, 250–255,
252f, 254fBrain mechanisms. See also Brain map-
ping; Prefrontal cortexabstract play and, 144–146influences of on aggression, 36–38MRI studies of brain development and,
246–250neuromodulators and, 262–263relationship to behavior, 242–243smoking during pregnancy and, 455
Bullyinghistory of, 84peer relationships and, 381relationships and, 17as a research construct, 5–6
Index 467

C
Cats, gamma-aminobutyric acid (GABA)and, 269
Cause of aggressionduring early childhood, 93–98, 97flanguage–aggression association and,
344–347of proactive and reactive aggression,
194–196Cerebral cortex, 250–255, 252f, 254f. See
also Brain mechanismsChild care
contagion effect in, 388peer relationships and, 377, 390–391as a risk factor, 124social capital and, 405–406
Child characteristicscauses of aggression and, 93as a determinant of aggression, 14
Child development perspectives, 10–12.See also Development
Childhood. See also Development; Pre-school age group
aggression during, 87–90, 89f, 430f–431, 452–454
causes of aggression during, 93–98, 97fempathy during, 136executive function and, 309–310, 312–
318gender differences and, 432–434hormones and, 288–290language development and, 334–335,
338–340morality in, 117–118overview, 98–101, 99frisk factors and, 121–124violence during, 202
Childhood-onset persistent aggression(CPA). See also Childhood
causes of, 354–355as a familial disorder, 355–368, 362t,
363t, 364tfuture research on, 368–370overview, 353
Chimpanzees, 135–137. See also MonkeysCingulate cortex, 251, 252. See also Brain
mechanismsCingulate motor areas, 251. See also Brain
mechanismsClassification of aggression. See Subtypes
of aggressionCliques, 384–387. See also PopularityCoercive control, 25
Coercive parenting, 359. See alsoParenting
Cognitive antecedent of aggression, 31–33Cognitive development, 96. See also Cog-
nitive skills; DevelopmentCognitive skills
global measures of, 313–315during infancy, 112neuromodulators and, 273–275peer relationships and, 379play fighting and, 55–57, 58
Cold executive function, 320–321, 322.See also Executive function
Collective efficacy, 410–411Competition
abstract play and, 143reproductive objectives and, 167for resources, 435
Complex social play, 144. See also PlayConduct disorder
cortisol and, 189, 293, 294DHEA/DHEAS and, 290executive function and, 311–312hormones and, 296language development and, 333, 337lithium and, 275physical aggression and, 308smoking and, 357–358, 360subtyping aggression and, 39testosterone and, 288–289
Conflictinfancy and, 108–109normative course of, 110–118, 114f,
115fduring the toddler years, 116–117
Consequences of aggressionpolythetic analysis and, 30fsubtyping aggression based on, 31–33
Constructs in aggression literature, 4–6Contextualism, 18–19Cooperation
lack of, 149–150rough-and-tumble play and, 138–
142Corpus collosum. See also Brain mecha-
nismsMRI studies of brain development and,
247–249overview, 251
Cortisol. See also Hormones; Hypotha-lamic–pituitary–adrenal (HPA) axis
overview, 282–283, 291–296proactive and reactive aggression and,
189–190
468 Index

Covert pathway, 205f–206. See also Devel-opmental pathway
Criminal behaviorage–violent crime curve and, 85–87fchildhood-onset persistent aggression
(CPA) and, 355daycare and, 406developmental trajectory of, 206–207genetic studies on, 228–234, 230flanguage development and, 333, 341maltreatment and, 359–360neighborhoods and, 411–412peer rejection and, 409–410proactive and reactive aggression and,
187–188social capital and, 413testosterone and, 287welfare system and, 400–402
CSF 5–HIAAinteraction with environment, 74–76fparenting and, 74in rhesus monkeys, 67–69, 69–70, 70–
72, 71fCulture
proactive and reactive aggression and,196
role of, 133–134
D
Daycarecontagion effect in, 388peer relationships and, 377, 390–391as a risk factor, 124social capital and, 405–406
Defensive aggressiondevelopment and, 38during infancy, 111–112
Defining aggressionduring infancy, 108–110problem of, 353–354for research purposes, 5
Delinquencychildhood-onset persistent aggression
(CPA) and, 354developmental aspects of, 207–209developmental trajectory of, 206–207friendships and, 16genetic studies on, 228–234, 230flanguage development and, 333maltreatment and, 359–360in the parents, 403–404peer relationships and, 409predictors of, 209–211
proactive and reactive aggression and,187–188, 195
social capital and, 414–415Depression
indirect aggression and, 167maternal, 123–124, 140serotonin and, 264–265
Development. See also Adolescence; Brainmechanisms; Childhood; Infancy;Language development; Preschoolage group; Toddler years
abstract play and, 149aggression prior to age six, 90–93f, 92fbrain, 246–250causes of aggression and, 93–98, 97fchild development vs. lifespan perspec-
tives of, 10–12contextualism and, 18–19daycare and, 405–406determinants of change in, 12–19evolutionary perspective of, 427–428,
429–431, 430fof executive function, 309–310friendships and, 16gender differences and, 439genetic factors and, 223–224, 456of homicide, 204–207, 205f, 207–209influences of on aggression, 38–39neuromodulators and, 270–273normative change and, 6–10f, 7f, 9foverview, 98–101, 99fof physical aggression, 83–85, 309of rhesus monkeys, 65, 67–69, 68f, 77–
78fsocial capital and, 399, 400t, 414, 417of social understanding, 138studies of aggression using, 448–450testosterone and, 284–287welfare system and, 401–402
Developmental model. See also Develop-ment
overview, 448–450of proactive and reactive aggression,
190–193Developmental pathway. See also Develop-
mentof homicide, 204–206, 205fprevention and, 214
Developmental trajectories. See also Devel-opment
of homicide, 206–207overview, 7–8sociometric status and, 10violence and, 211–214t, 212f, 213f
Index 469

Deviancy trainingoverview, 385–386in preschool children, 386–387
DHEA/DHEAS. See also Hormonesin childhood, 289–290future research on, 296–298
Direct aggression, 34Dominance, 138–142Dominance hierarchy, 135–136Dopamine. See also Neuromodulators
development and, 270–273executive control and, 273–275inhibition and, 261–262, 276overview, 265–268play fighting in rats and, 57
Dorsal premotor cortex, 251. See alsoBrain mechanisms
Dorsolateral prefrontal cortex. See alsoBrain mechanisms
executive control and, 273functions of, 144–145
Drug usedopamine and, 270–271gang involvement and, 414–415
E
Ecological perspective, 384–391Elephant seals, 236Emotion regulation, 346Emotional antecedents, 34–36Emotions, 188–190, 316Empathy, 133–138Empiricism, vs. nativism, 12–13Employment, maternal, 404–405. See also
Daycare; Socioeconomic statusEnemies, 16–17. See also RelationshipsEngagement, 268Enhancement model, 414–415Environment. See also Social learning
hypothesisbiological components and, 455causes of aggression and, 95, 96childhood-onset persistent aggression
(CPA) and, 355, 356, 356–360contextualism and, 18–19evolutionary perspective and, 100, 428genetic factors and, 228–234, 230f,
455–456hormones and, 295–296interaction with genetics, 74–76flanguage–aggression association and,
343–344maltreatment and, 457–459
neuromodulators and, 262norepinephrine and, 268predictors of violence and, 210–211proactive and reactive aggression and,
191as a risk factor, 123–124role of, 398rough-and-tumble play and, 139school, 407–408
Escalation theoryoverview, 208–209prevention and, 214
Ethnicity, 196Etiology pathways, 343–344Evolution of aggression
causes of aggression and, 95gender differences and, 434–436, 437–
439indirect aggression, 158–159, 165–170overview, 84–85, 425–428, 429–431, 430fsubtyping aggression and, 450–451
Executive control, 273–275Executive function
development of, 309–310in early childhood, 312–318hot and cold, 320–321overview, 307–308, 321–322problem-solving framework and, 318–
320relationship with externalizing or antiso-
cial disorders, 310–312Explanatory models, 32Exploratory activity, 139Expression of aggression
development and, 38–39polythetic analysis and, 30fsubtyping aggression based on, 33–34
F
Family. See also Family relationships;Parenting
childhood-onset persistent aggression(CPA) and, 355–368, 362t, 363t,364t, 368–370
social capital and, 402–405Family history, 185Family relationships. See also Relation-
shipschild development vs. lifespan perspec-
tives and, 11developmental trajectory of aggression
and, 8as a risk factor, 123–124
470 Index

Fantasy, 138Fast Track prevention trial, 408. See also
PreventionFear
responses to, 35–36rough-and-tumble play and, 139
Fear system, 37–38Females. See also Gender differences
aggressive competence and, 125developmental trajectory of aggression
in, 8in play fighting in rats, 49–50proactive and reactive aggression and,
196use of indirect aggression by, 161–162,
163–17050:50 rule. See also Play fighting
adult play fighting in rats and, 51juvenile play fighting in rats and, 55–
56overview, 48, 58
5–HIAA concentration levelsinteraction with environment, 74–76fparenting and, 74in rhesus monkeys, 67–69, 69–70, 70–
72, 71f5–HT. See also Neuromodulators
dopamine and, 265, 267gamma-aminobutyric acid (GABA) and,
266–267inhibition and, 261–262, 274–275overview, 262, 263–265
5–HTTinteraction with environment, 74–76finteraction with rearing conditions, 13
fMRI, 245f, 246f, 255–256. See also Brainmapping
Forceduring infancy, 109, 112–115f, 114ftoddlers’ use of, 125
Friends. See also Peer relationships; Rela-tionships
as a determinant of aggression, 15–16,17–18
preschool age children and, 384–387Frontal cortex
adolescence and, 272–273brain mapping studies on, 250–255,
252f, 254fFrontal lobes, 250–255, 252f, 254f. See
also Brain mechanismsFrontopolar cortex, 251. See also Brain
mechanismsFruit flies, boxing stance in, 26–27
Frustration–aggression model, 178–179.See also Reactive aggression
Functional MRI, 245f, 246f, 255–256. Seealso Brain mapping
Funding of research, 453
G
Game aggression, 38Game theory, 51Gamma-aminobutyric acid (GABA). See
also Neuromodulatorsinhibition and, 261–262, 274–275, 276overview, 262, 269–270serotonin and, 266–267
Gang involvement, 414–416Gender differences
aggressive competence and, 125childhood-onset persistent aggression
(CPA) and, 356development of, 431–436dopamine and, 267evolutionary perspective of, 427interaction with environment, 74–76flanguage development and, 339, 341,
348neuromodulators and, 262normative change and, 8overview, 5, 439peer relationships and, 379in play fighting in rats, 49–50preschool age children and, 383in proactive and reactive aggression, 196serotonin, 264–265sexual selection and, 426–427single-parent families and, 403testosterone and, 283–284, 284–286in toddlers’ use of force, 118–119, 120,
120–121in use of indirect aggression, 161–162,
163–165, 170–171, 437–439in verbal aggression, 436–437
Genetic–developmental model. See alsoGenetics
of aggression in animals, 224–227of aggression in humans, 228–234, 230foverview, 234–236
Geneticscauses of aggression and, 96–97, 100childhood-onset persistent aggression
(CPA) and, 355–368, 356, 362t,363t, 364t
as a determinant of aggression, 12–13environment and, 455–456
Index 471

Genetics (continued)hormones and, 295–296human studies and, 228–234, 230flanguage–aggression association and,
343–344, 344–345maltreatment and, 457–459mouse studies on, 224–227neuromodulators and, 262overview, 223–224, 234–236rhesus monkeys and, 69–70as a risk factor, 122
Genotype driven approaches, 223–224Girls. See also Gender differences
aggressive competence and, 125developmental trajectory of aggression
in, 8in play fighting in rats, 49–50proactive and reactive aggression and,
196use of indirect aggression by, 161–162,
163–170Grey matter, 252–254. See also Brain
mechanisms
H
Harm, subtyping aggression based on,31
Health care, 402Heritability of aggression. See also
Geneticschildhood-onset persistent aggression
(CPA) and, 355–368, 362t, 363t,364t
as a risk factor, 122High Scope project, 406. See also DaycareHippocampus, 144. See also Brain mecha-
nismsHomicide. See also Antisocial behavior;
Violencedevelopmental aspects of, 207–209heterogeneity of, 203–207, 205foverview, 202–203predictors of, 209–211prevention of, 214–215social capital and, 413violence trajectories and, 211–214t,
212f, 213fwelfare system and, 400
Hormones. See also Cortisol; Testosteroneas an explanation for violence, 86–87tfuture research on, 296–298genetic factors and, 295–296overview, 281–283, 298–299, 454–455
Hostile aggressionduring infancy, 109overview, 31–33, 183–184
Hot executive function, 320–321. See alsoExecutive function
Hunger, 139Hunting, 28Hyenas, empathy in, 136Hypothalamic–pituitary–adrenal (HPA)
axis. See also Brain mechanismscortisol and, 291–296future research on, 296–298genetic factors and, 295–296proactive and reactive aggression and,
189–190rough-and-tumble play and, 141
I
Impulsive aggression, 264Indirect aggression. See also Relational
aggression; Social aggressionage differences in, 160–161correlates of, 162–163gender differences in, 161–162, 437–
439overview, 34, 158–159, 159–160, 170–
171, 451in preschool children, 383use of by females, 163–170
Individual differencescauses of aggression and, 96changes in aggression and, 448–450compared to normative change, 6–10f,
7f, 9f5–HIAA concentration levels in rhesus
monkeys and, 68–69genetic factors and, 235homicide and, 209play fighting in rats and, 57in rhesus monkeys, 66–67in toddlers’ use of force, 118–121
Infancy. See also Developmentaggression during, 107, 124–126defining aggression during, 108–
110empathy during, 136executive function and, 310individual differences and, 118–121innate features of, 137–138language development and, 334–335,
336–337normative course of conflict and aggres-
sion in, 110–118, 114f, 115f
472 Index

proactive and reactive aggression and,191–192
risk factors and, 121–124Inhibition
lack of, 149neuromodulators and, 262–263, 273–
274overview, 261, 276proactive and reactive aggression and,
188Inhibition theories, 142–143Initiative, 111–112Injury, subtyping aggression based on,
31Innate features of aggression, 133–138Instrumental aggression. See also Proactive
aggressiondevelopment and, 38during infancy, 109overview, 31–33, 184peer relationships and, 381
Instrumental force, 109Insults. See Verbal aggressionIntegrative model of aggression, 39–41fIntelligence
executive function and, 312, 313–314language development and, 332, 338–
339, 341, 348neuromodulators and, 273
Intent, subtyping aggression based on, 31–33
Intergenerational transmissionof CPA
future research on, 368–370overview, 353, 355–368, 362t, 363t,
364tIntermale aggression, 37–38Interventions
child development vs. lifespan perspec-tives and, 11
childhood-onset persistent aggression(CPA) and, 369
daycare and, 406language–aggression association and,
345need for, 456–459peer relationships and, 392proactive and reactive aggression and,
195–196schools and, 408social, 11state dependence theory and, 209
Isolation, 52–53, 56
L
Labeling, 209Language–aggression hypothesis, 331–332.
See also Language developmentLanguage development. See also Develop-
mentcauses of aggression and, 96history of, 331–332overview, 330–331, 347–348peer relationships and, 378, 379prediction of future aggression and,
340–341in the preschool age group, 314prevalence throughout, 333–343theoretical models of association to
aggression, 343–347Learning, 95Lexical development, 332–333. See also
Language developmentLifespan perspectives, 10–12Limbic structures, 146Linnoila, Markku, 63. See also Rhesus
monkeysLithium, 275
MMacaques, 56–57. See also MonkeysMacro-social pathway, 343–344Males. See also Gender differences
developmental trajectory of aggressionin, 7–8
in play fighting in rats, 49–50proactive and reactive aggression and, 196use of indirect aggression by, 161–162
Maltreatmentchildhood-onset persistent aggression
(CPA) and, 359–360genetic factors and, 457–459hormones and, 295–296monoamine oxidase and, 13neighborhoods and, 412play and, 150proactive and reactive aggression and,
186Maternal employment, 404–405. See also
DaycareMaternal responsiveness, 358–359Maturation status, 169Mediofrontal cortex, 252. See also Brain
mechanismsMemory. See also Executive function
neuromodulators and, 273–275overview, 321
Index 473

Mesial frontal cortex, 251. See also Brainmechanisms
Micro-social pathway, 345–346Mid-dorsolateral frontal cortex, 251, 252.
See also Brain mechanismsMid-ventrolateral frontal cortex, 251, 252.
See also Brain mechanismsMolecular genetics, 232–234. See also
GeneticsMonkeys. See also Rhesus monkeys
indirect aggression among, 167neuromodulators and, 275overview, 135–137rough-and-tumble play in, 56–57, 284
Monoamine oxidasegenetic factors and, 232–234maltreatment and, 360, 458–459parental maltreatment and, 13
Mood, testosterone and, 287Mood disorders
language development and, 341serotonin, 264–265
Moralistic aggression, 28Morality
in early childhood, 117–118play and, 143rough-and-tumble play and, 141–142
Motivational systemabstract play and, 144–146brain mechanism research on, 37causes of aggression and, 94overview, 137, 151–152predation and, 28subtyping aggression based on, 31–33
Mouse models. See also Rodentsdopamine and, 270gamma-aminobutyric acid (GABA) and,
270gender differences in, 285genetic–developmental model, 224–227norepinephrine and, 269
MRI. See also Brain mappingfunctional, 245f, 246foverview, 255–256structural, 243–244fstudies of aggression using, 250–255,
252f, 254fstudies of brain development, 246–250
N
Name calling. See Verbal aggressionNativism, vs. empiricism, 12–13Natural selection, 166
Neglect. See MaltreatmentNeighborhoods
peer relationships and, 389–390social capital and, 410–412
Neocortexinfluences of on aggression, 37integrative model and, 40
Neural circuitriesabstract play and, 144–146influences of on aggression, 37
Neuroendocrinological status, 37Neuroimaging. See Brain mappingNeuromodulators
development and, 270–273executive control and, 273–275inhibition and, 261–262overview, 262–263, 276
Neurotransmitters. See alsoNeuromodulators
dopamine, 265–268gamma-aminobutyric acid (GABA), 269–
270inhibition and, 261–262norepinephrine, 268–269serotonin, 263–265
Nonviolent aggression, 5–6Noradrenaline, 268Norepinephrine. See also Neuromodulators
inhibition and, 261–262overview, 268–269
Norm of reaction, 428Normative change, 6–10f, 7f, 9f
O
Object-oriented nature of aggression, 38Offensive aggression, 37. See also
Proactive aggressionOntogenetic organization
comparing different levels of, 26–29overview, 39
Oppositional–defiant disorder (ODD)cortisol and, 294DHEA/DHEAS and, 289–290executive function and, 312, 317–
318hormones and, 296physical aggression and, 308testosterone and, 288–289
Orbitofrontal cortex, 251, 252. See alsoBrain mechanisms
Orbitofrontal–prefrontal cortex, 144Organized crime, 416, 417–418. See also
Gang involvement
474 Index

Ostracism, 380–383Overt pathway, 205f–206. See also Devel-
opmental pathway
P
Pain, 35–36Parallel development model, 191Parent–child relationship, 14–15. See also
Family relationships; Parenting;Relationships
Parental aggression, 28Parenting
childhood-onset persistent aggression(CPA) and, 357, 358–360, 368–369
cognitive skills and, 313–314as a determinant of aggression, 14gene–environment interactions and, 75–
76fhormones and, 298interventions and, 457–459language–aggression association and,
343–344peer relationships and, 389predictors of violence and, 210proactive and reactive aggression and,
185, 191in rhesus monkeys, 73–74as a risk factor, 123–124rough-and-tumble play and, 140serotonin transporter gene (5–HTT)’s
interaction with, 13social-cognitive processes and, 382socialization and, 14–15use of force during infancy and, 112–
115f, 114fParents
antisocial behavior of, 360–368, 362t,363t, 364t
social capital and, 403–404Peer rejection. See also Peer relationships
during the preschool years, 380–383social capital and, 409–410
Peer relationships. See also Friends; Rela-tionships
causes of aggression and, 96as a determinant of aggression, 15in early childhood, 378–380, 384–391gene–environment interactions and, 75–
76findirect aggression and, 162–163during infancy, 110, 112overview, 376–378predictors of violence and, 210
during the preschool years, 380–383,391–392
proactive and reactive aggression and,185–186, 188, 191–192
rejection from, 380–383, 409–410in rhesus monkeys, 73–74as a risk factor, 124social capital and, 408–410during the toddler years, 116–117use of force during infancy and, 113
Performance theory, 14. See also Socializa-tion
Perry preschool project, 406. See alsoDaycare
Person-oriented nature of aggression, 38Personal Responsibility and Work Oppor-
tunity Reconciliation Act(PRWORA), 401. See also Welfaresystem
Personality disorderslanguage development and, 331molecular genetics and, 232–233proactive and reactive aggression and,
187Personally directed force
during infancy, 109toddlers’ use of, 119
Personally motivated aggression, 32–33Phenotype driven approaches, 224, 235–
236Phylogenetic organization, 26–29, 40Play. See also Play fighting; Rough-and-
tumble playabstract, 142–151overview, 152, 451social understanding and, 138
Play fighting. See also Rough-and-tumbleplay
in adult rats, 50–51in juvenile rats, 52–55, 54f, 55–57overview, 47–48, 58, 138–142in rats, 48–50, 49f
Pleasure-motivated aggression, 32Polythetic analysis, 29–30f. See also Sub-
types of aggressionPopularity, 163, 165, 169Population heterogeneity theories
overview, 208prevention and, 214–215
Postpartum period, 298Predatory aggression. See Proactive
aggressionPredicting aggression, 449–450
Index 475

Prefrontal cortex. See also Brainmechanisms
abstract play and, 145, 146–147brain mapping studies on, 250–255,
252f, 254fdamage to, 148–149executive control and, 273influences of on aggression, 37inhibition and, 276integrative model and, 40overview, 251
Pregnancycauses of aggression and, 96childhood-onset persistent aggression
(CPA) and, 357–358dopamine and, 270–271hormones and, 285–286, 297–298predictors of violence and, 210risk factors and, 123smoking during, 123, 210, 455
Premotor cortex, 251. See also Brainmechanisms
Prenatal malnutrition, 271. See also Preg-nancy
Preschool age group. See also Childhood;Development
aggression during, 90–93f, 92f, 430f–431, 452–454
executive function and, 310, 312–318gender differences and, 433language development and, 334–335,
336–337peer rejection and ostracism during,
380–383peer relationships and, 378–380, 384–
391, 391–392Prevention. See also Interventions
of homicide and violence, 214–215need for, 456–459schools and, 408state dependence theory and, 209
Primary motor cortex, 251. See also Brainmechanisms
Primates. See also Rhesus monkeysindirect aggression among, 167neuromodulators and, 275overview, 135–137rough-and-tumble play in, 56–57, 284
Proactive aggression. See also Instrumentalaggression
causality of, 194–196correlates of, 184–190developmental model of, 190–193gender, ethnicity, and culture and, 196
integrative model and, 40–41fmeasurement and validity issues regard-
ing, 180–184overview, 179–180, 197
Proactive use of forceduring infancy, 109during the toddler years, 119
Problem solving, 195Problem-solving framework
executive function and, 318–320overview, 321–322
Procedural morality, 141–142. See alsoMorality
Prosocial aggressionvs. antisocial aggression, 5–6overview, 33
Protest, 111Proximal antecedents
integrative model and, 40overview, 34–36
Psychoanalytic view, 134–135Punishment
as aggression, 33childhood-onset persistent aggression
(CPA) and, 359physical, 84
QQuantitative genetics, 228–232, 230f. See
also Genetics
RRage-like aggression, 37Rage system, 37–38Random effect growth modeling, 206–
207Rats. See also Rodents
boxing stance in, 26–27brain mechanism research on, 37dopamine and, 267, 270–271empathy in, 136neuromodulators and, 271, 275norepinephrine and, 268play fighting in, 48–50, 49f, 55–57, 58,
139, 142play fighting in adults, 50–51play fighting in juveniles, 52–55, 54f
Reactive aggressioncausality of, 194–196correlates of, 184–190developmental model of, 190–193gender, ethnicity, and culture and, 196integrative model and, 40–41f
476 Index

measurement and validity issues regard-ing, 180–184
overview, 36, 178–180, 197peer relationships and, 381
Reactive use of force, 109Regions, 412–414Regulation of aggression, 142–143Rejection, peer. See also Peer relationships
during the preschool years, 380–383social capital and, 409–410
Relational aggression. See also Indirectaggression
compared to indirect aggression, 159–160
gender differences in, 6, 437overview, 34in preschool children, 383
Relationships. See also Family relation-ships; Peer relationships
in rhesus monkeys, 64–66socialization and, 13–18violence in, 188
Relief-motivated aggression, 32Reproductive objectives
gender differences in, 428, 435indirect aggression and, 166–170
Repulsion hypothesis, 17Resistance, 111Resolution of conflict, 116–117. See also
ConflictRetaliation, 111Revenge, 150Rhesus monkeys. See also Monkeys
development of, 67–69, 68feffects of early social experiences on,
72–74empathy in, 136field studies of, 70–72, 71fgene–environment interactions and, 74–
76fgenetic factors and, 69–70individual differences and, 66–67overview of research with, 77–78frough-and-tumble play among, 139–140social aspects of, 64–66
Risk factorschildhood-onset persistent aggression
(CPA) and, 355, 355–368, 362t,363t, 364t
early development of aggression, 121–124
homicide and, 209–211population heterogeneity theory and,
208
Rodentsboxing stance in, 26–27brain mechanism research on, 37dopamine and, 267, 270, 270–271empathy in, 136gamma-aminobutyric acid (GABA) and,
270gender differences in, 285genetic–developmental model, 224–227neuromodulators and, 271, 275norepinephrine and, 268, 269play fighting in, 48–50, 49f, 55–57, 58play fighting in adults, 50–51play fighting in juveniles, 52–55, 54frough-and-tumble play among, 139, 142testosterone and, 284
Role play, 138Rough-and-tumble play. See also Play
fightinggender differences in, 284, 428, 435overview, 138–142, 152peer relationships and, 381in rhesus monkeys, 65–66social understanding and, 138
S
Schizophrenia, 274School achievement
predictors of violence and, 210proactive and reactive aggression and,
187School departure, 167Schools
environment of, 428peer relationships and, 391social capital and, 407–408
Selection model, 414–415Selective breeding, 225–227. See also
GeneticsSelf-esteem, 167Self-regulation
language–aggression association and,346
overview, 307–308Self-selection model, 209Semantic development, 332–333. See also
Language developmentSerotonin. See also Neuromodulators
dopamine and, 265, 267gamma-aminobutyric acid (GABA) and,
266–267inhibition and, 261–262, 274–275overview, 262, 263–265
Index 477

Serotonin metabolismcauses of aggression and, 96in rhesus monkeys, 66–67, 69–70
Serotonin receptors, 232–234Serotonin transporter gene (5–HTT)
interaction with environment, 74–76finteraction with rearing conditions, 13
Sex differences. See Gender differencesSex segregation, 435. See also Gender dif-
ferencesSexual encounters
overview, 451play fighting and, 48, 49–50, 52, 55
Sexual selection, 166, 426–427Shared etiology pathways, 343–344Shared genetic liability pathway, 343–344Siblings. See also Family relationships
causes of aggression and, 97conflicts with, 117preschool age children and, 388–389
Single-parent family, 403. See also FamilySmoking. See also Substance use
causes of aggression and, 97childhood-onset persistent aggression
(CPA) and, 357–358, 362t–363tconduct disorder and, 360during pregnancy, 123, 210, 455
Social aggression. See also Indirect aggres-sion; Relational aggression
compared to indirect aggression, 159–160
overview, 34Social bonding, 49Social capital
daycare and, 405–406at the family level, 402–405negative, 414–417neighborhoods and, 410–412overview, 398–399, 417–418peer relationships and, 408–410schools and, 407–408town and region and, 412–414
Social cognitionslanguage–aggression association and,
346parenting and, 382proactive and reactive aggression and,
183Social development, in rhesus monkeys,
64–66. See also Rhesus monkeysSocial–development perspective, 38–39Social experience, in rhesus monkeys, 72–
74Social facilitation model, 415
Social forces, 137Social-interactionist perspective, 32Social interventions, 11. See also Interven-
tionsSocial learning hypothesis. See also Envi-
ronmentcauses of aggression and, 94overview, 125, 429, 453proactive aggression and, 179
Social organizationinnateness of, 133–138in rhesus monkeys, 64–66
Social skillsabstract play and, 147–149indirect aggression and, 162–163language development and, 342peer relationships and, 378, 379play fighting and, 50, 52–53, 55–57,
58proactive and reactive aggression and,
195in rhesus monkeys, 73–74training in, 457
Social status, 162–163Social systems
causes of aggression and, 100contextualism and, 18–19
Social testing, 49Socialization
gender differences and, 432language–aggression association and,
343–344model of, 414–415overview, 95responses to, 151–152in rhesus monkeys, 64–66from unilateral to bilateral views of, 13–
18Socially motivated aggression, 32–33Socioeconomic status. See also Welfare
systemage–violent crime curve and, 89cognitive skills and, 313–314indirect aggression and, 169–170language–aggression association and,
343–344language development and, 341peer relationships and, 389–390physical aggression and, 308predictors of violence and, 210–211proactive and reactive aggression and,
187social capital and, 413
Sociometric status, 10
478 Index

Specific language impairments (SLI), 332.See also Language development
Speech and language difficulties (SLD),332. See also Language development
Splenium, 249. See also Brain mechanismsState dependence theory
overview, 208–209prevention and, 214
Strain differences, 69–70Stress system
cortisol and, 291–292hormones and, 297–298
Structural MRI, 243–244f. See also Brainmapping; MRI
Subcortical structures, 37Substance use. See also Alcohol use;
Smokinglack of behavioral control and, 261overview, 451proactive and reactive aggression and,
187, 195serotonin and, 264
Subtypes of aggression. See also Taxon-omies of aggression
based on its antecedents, 34–39based on its expression, 33–34based on the consequences, 31–33integrative model and, 39–41fmultilevel analysis of, 29–30foverview, 41–42, 450–452phylogenetic and ontogenetic organiza-
tion of, 26–29Suicide
indirect aggression and, 167molecular genetics and, 233serotonin and, 264
Supervision, 404–405Supplementary motor area, 251. See also
Brain mechanisms
T
Tantrums, 100Taxonomies of aggression. See also Sub-
types of aggressionearly, 27–29integrative model and, 39–41foverview, 39
Teasing aggression, 38. See also Verbalaggression
Temperamentproactive and reactive aggression and,
184–185, 191as a risk factor, 122
Terrorism, 33Testosterone. See also Hormones
causes of aggression and, 96as an explanation for violence, 86–87tfuture research on, 296–298overview, 282–283, 283–291
Theory of mindabstract play and, 149–150executive function and, 316, 320–321language–aggression association and,
346–347Threat, 35–36. See also Reactive aggres-
sionToddler years. See also Development;
Infancyaggression during, 124–126, 430f–431,
452–454anger during, 110executive function and, 310gender differences and, 432individual differences and, 118–121language development and, 334–335peer relationships and, 378–380risk factors and, 121–124use of force during, 114–115f
Towns, 412–414Transsexuals, 287Tryptophan hydoxylase
genetic factors and, 233overview, 264
Type A personality, 178–179
UUnilateral view of socialization, 13–18. See
also Socialization
VVentral premotor cortex. See also Brain
mechanismsabstract play and, 145overview, 251
Ventrolateral–medial hypothalamus, 37–38Ventromedial cortex, 251. See also Brain
mechanismsVerbal abilities, 315Verbal aggression
gender differences in, 436–437overview, 108
Verbal organization, 273–275Victimization
in preschool children, 383relationships and, 17as a research construct, 5–6
Index 479

Video games, 150Vigilance, 268Violence. See also Antisocial behavior;
Homicidedevelopment of, 83–85, 207–209genetic studies on, 228–234,
230flanguage development and, 333neighborhoods and, 412vs. nonviolent aggression, 5–6overview, 202–203play and, 149, 150predictors of, 209–211prevention of, 214–215proactive and reactive aggression and,
187–188, 195serotonin and, 264testosterone and, 287trajectories of, 211–214t, 212f,
213fVisuospatial abilities, 315Vocabulary. See Language development
WWar
as aggression, 33evolution of aggression and, 85
Weaning, 52Weaning aggression, 28Welfare system. See also Socioeconomic
statusoverview, 399–402social capital and, 418
White matter, 248–250. See also Brainmechanisms
Wisconsin Card Sorting Test (WCST), 310,311. See also Executive function
World wars, 85
YYoung male syndrome, 169
ZZoomorphism, 26–27
480 Index