development of vegetation patterns in early primary succession
TRANSCRIPT

Development of vegetation patterns in early primary succession
Bryndıs Marteinsdottir, Kristın Svavarsdottir & Thora Ellen Thorhallsdottir
AbstractQuestion: We investigated colonisation filters inearly plant community development on a glacialoutwash plain. We asked if these were related toseed limitation or to a lack of safe sites, if topogra-phical heterogeneity affected species patchiness andhow species life cycles influence successional trajec-tories.Location: An outwash plain (Skeijararsandur) insoutheast Iceland.Methods: We identified surface heterogeneity at twodifferent scales, ca. 10–15 cm (larger stones andestablished plants) and ca. 50m (shallow depres-sions representing dry river beds) at two study sites.We quantified species cover, flowering plant density,seed production, seed rain, seed bank density, seed-ling emergence and seedling survival from June 2005to June 2007 for the whole plant community, andmeasured seed production for five species.Results: Mean vegetation cover was o2.5% at thesites. Low emergence rates and high seedling mor-tality were the two main recruitment filters. Only1.4% of seedlings emerging in 2005 survived into the2007 growing season. Topographical heterogeneityhad little effect on plant colonisation. High annualvariation was recorded, and the two study sites (ca.2 km apart) differed in their colonisation success. Ofthe five species, establishment of Cerastium alpinumand Silene uniflorawas most limited by lack of seeds,whereas establishment of Luzula spicata, Poa glaucaand Rumex acetosella was most limited by safe sites.Conclusions: We conclude that colonisation pro-cesses and patterns in early primary succession on
Skeijararsandur were largely influenced by stochas-tic factors.
Keywords: Iceland; Microsite; Outwash plain; Re-cruitment; Seed bank; Seedling survival; Seed rain;Topography.
Nomenclature: Kristinsson (2001, 2009)
Introduction
Plant colonisation and vegetation developmentmay be directed and rate-limited by two differentprocesses. The first is related to the size and speciescomposition of the seed pool and probabilities ofdispersal and arrival onto the site. The second is themore deterministic environmental filtering reflectingsuitable microsites for germination and growth (e.g.Eriksson & Ehrlen 1992; del Moral & Bliss 1993).The absence of a species at a site or a seral stage maytherefore be due as much to its absence from theseed pool as to its inability to establish (Fastie 1995).Successful early establishment of a population invirgin terrain can have long lasting implications forsuccessional trajectories (del Moral & Lacher 2005).Those first colonists may act as nascent foci (sensuMoody &Mack 1988), and become a local source ofseeds. They may facilitate establishment of othercolonists by creating shelter (Castellanos et al. 1994;Franks 2003) and increasing subsurface biologicalactivity (Cargill & Chapin 1987; Blundon et al. 1993;Callaway 1995).
During the initial stages of primary succession,seed input comes mostly through long-distance dis-persal. With time, a seed bank builds up and localseed production is believed to gradually take over asthe major seed source (del Moral et al. 2005). Al-though the importance of an available seed pool inprimary succession has been emphasised in manystudies (Titus & del Moral 1998), few have at-tempted simultaneously to quantify the seed rainand seed bank and assess their spatial components(but see Wood & del Moral 2000; Bossuyt & Hermy2004).
In harsh, early successional environments, sur-face heterogeneity at various scales in the landscapecan create microsites that are differently favourablefor plant establishment. Depressions, large stones
Marteinsdottir, B. (corresponding author, Bryndis.
[email protected]): Institute of Biology,
University of Iceland, Sturlugotu 7, 101 Reykjavık,
Iceland. Present address: Department of Botany,
Stockholm University, SE-106 91 Stockholm, Sweden.
Svavarsdottir, K. ([email protected]): Soil
Conservation Service, Keldnaholt, 112 Reykjavık, Ice-
land
Thorhallsdottir, T.E. ([email protected]): Institute of Biol-
ogy, University of Iceland, Sturlugotu 7, 101
Reykjavık, Iceland.
Journal of Vegetation Science 21: 531–540, 2010DOI: 10.1111/j.1654-1103.2009.01161.x& 2010 International Association for Vegetation Science

and established plants can facilitate the colonisationprocesses by creating shelter, retaining moisture andtrapping seeds (Wood & del Moral 1987; Jumppo-nen et al. 1999; Franks 2003).
All the factors mentioned above are important forplant establishment, but have seldom been simulta-neously studied. In this paper, seed rain, soil seedbank, emergence of seedlings, seedling survival andvegetation cover were all considered and quantified.
We studied early primary succession on Skei-jararsandur, a vast (1000 km2), homogeneous gla-cial outwash plain in southeast Iceland. Our aim wasto determine which factors control plant colonisa-tion and development of vegetation patterns early insuccession. We asked: (1) Are the colonisation filterson Skeijararsandur related to seed limitation or to alack of suitable sites for establishment? (2) Does to-pographical heterogeneity affect species patchiness?(3) How do species life cycles influence successionaltrajectories?
Materials and Methods
Study sites
Two sites at a pioneering successional stagewere selected near the upper centre of Skeijar-arsandur, (site A: 631570N, 171090W, site B:631560N, 171120W), an outwash plain south of Skei-jararjokull, the largest outlet glacier of Vatnajokullicecap. The two study sites, about 2 km apart, weremostly level with a subtly undulating pattern of flatsand depressions (mean depth 0.1m, mean diameter26m), representing old flood pathways. These sitesappeared similar and were chosen as representativefor very large sparsely vegetated areas of Skeijar-arsandur. The soil in the area is classified as vitrisol,with mineralogy dominated by volcanic glass (Ar-nalds 2004). It is infertile, with mean nitrogen andcarbon concentrations of 0.01% and 0.05%, re-spectively. The soil was 74% sand, 20% gravel and6% silt/clay, and soil conditions were similar be-tween flats and depressions and between the twostudy sites, although site B had slightly finer soilgrain size than site A (Marteinsdottir 2007).
The climate on Skeijararsandur is maritime,with cool summers and mild winters. Temperatureand precipitation values during the study periodwere obtained from the weather station Fa-gurholsmyri (651530N, 161390W) approx. 26 kmfrom the study sites. The mean annual air tempera-tures were 5.11C and 5.81C in 2005 and 2006,respectively, and the mean summer (June-August)
temperature was 10.61C in both years. Mean annualand summer precipitation was 1593 and 469mmrespectively in 2005, and 2070 and 433mm respec-tively in 2006 (Icelandic Meteorological Office2007). During the study period the growing seasonat Skeijararsandur lasted from approximately mid-May to early September.
Sampling
Within each study site, 10 plots (10�10m) werelaid out, five plots in depressions and five on flats, re-presenting mesoscale environmental heterogeneity atthe sites. The plots were positioned in the closest de-pression/flat to a random point determined usingrandom compass bearings and distances. In July2005, 12 (0.5�0.5m) quadrats were randomly placedwithin each study plot (240 quadrats in total) formeasurements of seedling emergence and survival.Within quadrats, seedlings were counted, identified tospecies or group (i.e. graminoids and Galium spp.)and marked with a coloured wire ring, using differentcolours for each sampling date. Due to high seedlingdensity, smaller quadrat size (0.3�0.5m) was used for46 samples at site B. Seedling emergence and/or sur-vival was recorded six times during the study, 1 Julyand 22 August 2005, 12 June, 17 July and 20 August2006 and 20 June 2007.
For estimation of seed rain at the study sites, oneseed trap was randomly positioned in August 2005(N5 20) and two traps in August 2006 (N540),within each of the 20 plots. The traps were emptied inearly and late September in both years and the sam-ples combined before analysis. Seeds were countedand assigned to species or higher taxa, such as car-yophyllaceous species and graminoids. The seed trapswere 0.2�0.2m Astroturf grass mats anchored to theground with wire pins in each corner. This measure-ment only gives an abrupt estimate of the seed rain inthe area. Even though synthetic grass mats of this kindhave been used with good efficiency in various seedrain studies in arctic regions (e.g. Molau & Larsson2000), they can underestimate the seed rain of grami-noids and herbs (Larsson & Molau 2001). The seedrain was also only measured in late summer whenmost species in the area shed their seeds. Late summershedding of seeds has been shown to be much greaterthan winter shedding (Ryvarden 1971, 1975), but thismay be offset by higher probabilities of long-distancedispersal in winter. Despite limitations of the seed rainestimates in this study, our measurements indicate theextent and composition of the local seed rain and al-low us to compare seed rain between topographies,sites and years.
532 Marteinsdottir, Bryndıs et al.

In early August 2006, 10-m transects were laidout, adjacent to each study plot (in the same de-pressions and flats). Vegetation cover was deter-mined along transects using the point interceptmethod. Five quadrats (0.5�0.5m), with 100 reg-ularly distributed points, were laid out at regularintervals and the first hit recorded for each species ateach point. Vascular plants were identified to spe-cies, while bryophytes and lichens were eachrecorded as separate taxa. In addition, all vascularplant species were listed at both sites.
To analyse the seed bank, five 10-m transectswere laid out in depressions and five on flats at eachsite and seed bank data obtained by the seed germi-nation method (Baskin & Baskin 1998). Along eachtransect, five soil cores were collected at each ofthree microsites, representing the microscale topo-graphical variation at the sites: (1) the lee side of acushion plant (5–20 cm in diameter), (2) the lee sideof a stone (5–20 cm in diameter, max. height 10 cm)and (3) exposed controls, neither close to a stone nora cushion plant. The position of the cores was de-termined by choosing the closest appropriatemicrosite to a random point. A total of 300 soil coreswere collected in early May 2005 using an auger of5.3 cm in diameter down to 5-cm depth. The sampleswere spread as a maximum 2-mm thick layer ontrays filled with sterile peat (pH 5.5). The trays wererandomly placed in a greenhouse with additionalUV light during the day for the first two months.Soil moisture was maintained by regular watering.No seedlings appeared in control trays (sterile peat).Seedlings were assigned to a species or in case ofCaryophyllaceae, Galium spp. and graminoids, tohigher taxa as they could not be identified at thespecies level. Counts were conducted weekly duringthe first 10 weeks, but in alternate weeks thereafter.The study was terminated after 16 weeks, when nonew seedlings had emerged for six weeks.
For estimation on seed production, five com-mon perennial species, Luzula spicata, Cerastiumalpinum, Silene uniflora, Rumex acetosella and Poaglauca, were chosen to represent the community.These species are among the most common in thearea (collectively 49% of the total vegetation cover)and represent the two most important functionalgroups, graminoids and herbs. The above specieswill hereafter be referred to by their generic names.As abundance of flowering individuals was low, nodistinction was made in seed production betweentopographies or sites. For each species, 15 floweringindividuals, growing within or close to the samplingplots were selected from the sites. In mid-July 2006,after stigmas had withered, inflorescences of Luzula,
Cerastium and Silene were carefully bagged with1-mmmesh white nylon material. The bags were tiedto thin metal rods that arched at right angles overthe plant so the inflorescence did not bear the weightof the bag. This procedure was carried out on Ru-mex and Poa in early August. All seed bags werecollected in late September. Mature seeds, abortedseeds and unripe seeds were separated and counted.Pre-dispersal seed predation was estimated forLuzula, Poa, Rumex and Cerastium when pupae,larvae or holes after larvae were observed in fruits.For Silene, the procedure described by Pettersson(1994) was followed so only individuals with swollenovules were bagged, and seed predation recorded inthe field when a fruit was found to be completelyempty. The density of the bagged species and num-ber of flowers or inflorescence per individual wasrecorded in the 10 study plots at each site to obtainestimations of number of seeds perm� 2.
To evaluate the relative importance of local seedproduction in the area and the influence of differentfilters on the colonisation of those five species, a lifecycle diagram was constructed. Survival percentageswere estimated as follows: number of ovules perm� 2 was estimated from the mean number of ovulesper flower or inflorescence and the mean number offlowers or inflorescence perm� 2. Seed/ovule ratioswere obtained by comparing number of matureovules with the number of all ovules. Dispersed seedperm� 2 was obtained by multiplying the number ofmature ovules perm� 2 with the percentage of intactfruits. Germination percentage was calculated bydividing seedling density in 2005 by the number ofseeds in the spring seed bank in 2005. Survival wasthe percentage of seedlings emerging in 2005 thatwere alive at the end of the 2006 growing season.
Statistical analyses
Generalised linear models (GLM) with Poissonerror were used to test for soil seed bank differencesbetween sites, topography and microsites. Multiplecomparisons among microsites were done using at-test based on the best model, with Bonferroniadjusted significance values (P5 0.025). For dif-ferences in number of seedlings between sites, topo-graphies and years, a GLMwith quasi-Poisson error(due to over-dispersion) was used. The same ap-proach was used to test for seed rain differencesbetween site, year and topography. The seed rainand seedling emergence data had high variation be-tween years, so each year was analysed separately.GLMs with quasi-binomial error (due to over-dispersion) were used to analyse the effect of site and
Vegetation patterns in early primary succession 533
topography on first-year survival (from seedlingemerging to the first sampling date in the next sum-mer), total survival (from seedling emerging in 2005until the last sampling date in 2007) and for com-parison between years in first-year survival. Modelslooking at seedling emergence and survival includedcohorts as a covariate. The effect of site and topo-graphy on total vegetation cover was analysed in thesame way. In the GLMs, saturated models includedall variables as well as the interaction between them.To test for significance of each variable, the fits offull and reduced models were compared in a hier-archical manner using an F-test for models withquasi-Poisson or quasi-binomial error and a chi-square test for models with Poisson error (Crawley2002; Quinn & Keough 2002).
Statistical analyses were conducted using R2.4.1 for Windows (available from http://cran.r-project.org).
Results
Vegetation cover
A total of 28 vascular plant species were re-corded; 27 at site A and 19 at site B. Poa (30% of
total vascular plant cover), Rumex (20%), Arabi-dopsis petraea (13%), Festuca richardsonii (11%)and Thymus praecox (11%) were the most abundantspecies (Table 1). Moss and lichen cover was only0.09% and 0.01%, respectively. Vegetation coverwas significantly higher at site A (2.2%) than site B(1.2%) (F1, 18 5 4.74, P5 0.043), but did not differbetween flats and depressions (Table 2).
Seed rain
The total seed rain differed significantly betweenyears (F1,57567.87, Po0.001), with 393 versus80 seedm� 2 in 2005 and 2006, respectively.Rumexwasthemost abundant species, followed by graminoids andcaryophyllaceous species (Table 1). Seed rain did notdiffer between flats and depressions in either year. In2005, site B had a significantly higher total seed rainthan site A (F1,18527.89, Po0.001) (Table 2).
Soil seed bank
A total of 265 seeds germinated, equivalentto 400 seedsm� 2. The most abundant species/groups were Rumex (52% of total seedlings),graminoids (26%), T. praecox (7%) and A. petraea(7%) (Table 1). The seed bank density was
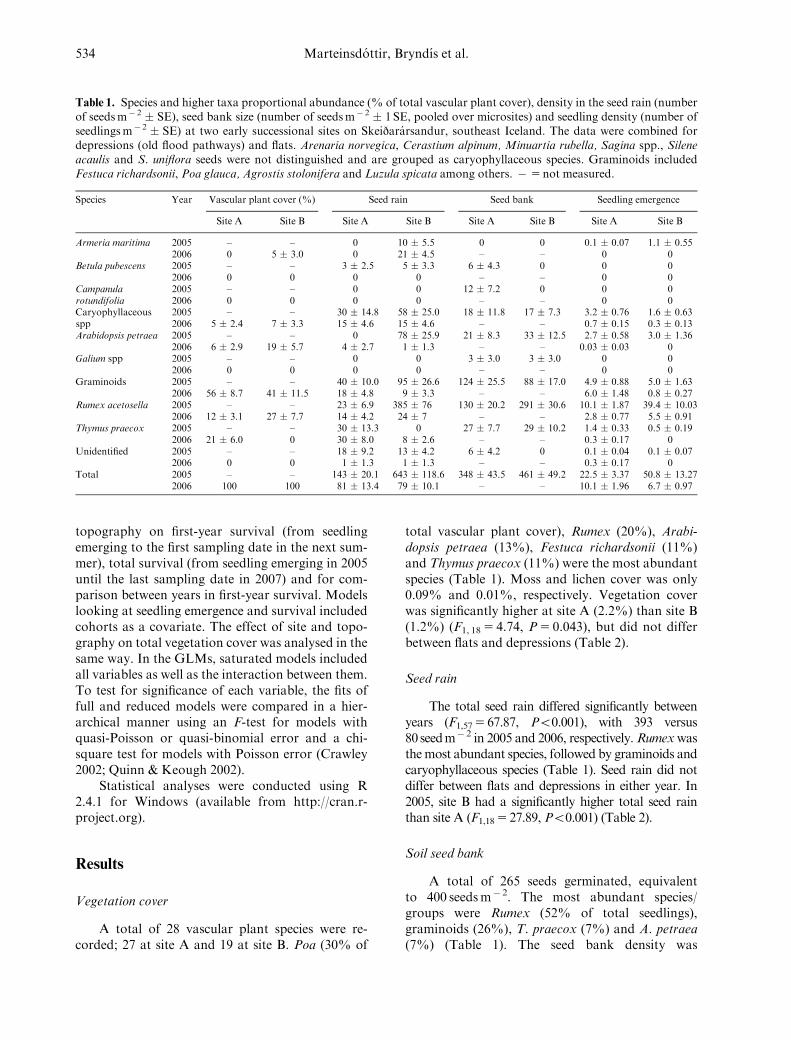
Table 1. Species and higher taxa proportional abundance (% of total vascular plant cover), density in the seed rain (numberof seedsm� 2 � SE), seed bank size (number of seedsm� 2 � 1 SE, pooled over microsites) and seedling density (number ofseedlingsm� 2 � SE) at two early successional sites on Skeijararsandur, southeast Iceland. The data were combined fordepressions (old flood pathways) and flats. Arenaria norvegica, Cerastium alpinum, Minuartia rubella, Sagina spp., Sileneacaulis and S. uniflora seeds were not distinguished and are grouped as caryophyllaceous species. Graminoids includedFestuca richardsonii, Poa glauca, Agrostis stolonifera and Luzula spicata among others. � 5 not measured.
Species Year Vascular plant cover (%) Seed rain Seed bank Seedling emergence
Site A Site B Site A Site B Site A Site B Site A Site B
Armeria maritima 2005 – – 0 10 � 5.5 0 0 0.1 � 0.07 1.1 � 0.552006 0 5 � 3.0 0 21 � 4.5 – – 0 0
Betula pubescens 2005 – – 3 � 2.5 5 � 3.3 6 � 4.3 0 0 02006 0 0 0 0 – – 0 0
Campanularotundifolia
2005 – – 0 0 12 � 7.2 0 0 02006 0 0 0 0 – – 0 0
Caryophyllaceousspp
2005 – – 30 � 14.8 58 � 25.0 18 � 11.8 17 � 7.3 3.2 � 0.76 1.6 � 0.632006 5 � 2.4 7 � 3.3 15 � 4.6 15 � 4.6 – – 0.7 � 0.15 0.3 � 0.13
Arabidopsis petraea 2005 – – 0 78 � 25.9 21 � 8.3 33 � 12.5 2.7 � 0.58 3.0 � 1.362006 6 � 2.9 19 � 5.7 4 � 2.7 1 � 1.3 – – 0.03 � 0.03 0
Galium spp 2005 – – 0 0 3 � 3.0 3 � 3.0 0 02006 0 0 0 0 – – 0 0
Graminoids 2005 – – 40 � 10.0 95 � 26.6 124 � 25.5 88 � 17.0 4.9 � 0.88 5.0 � 1.632006 56 � 8.7 41 � 11.5 18 � 4.8 9 � 3.3 – – 6.0 � 1.48 0.8 � 0.27
Rumex acetosella 2005 – – 23 � 6.9 385 � 76 130 � 20.2 291 � 30.6 10.1 � 1.87 39.4 � 10.032006 12 � 3.1 27 � 7.7 14 � 4.2 24 � 7 – – 2.8 � 0.77 5.5 � 0.91
Thymus praecox 2005 – – 30 � 13.3 0 27 � 7.7 29 � 10.2 1.4 � 0.33 0.5 � 0.192006 21 � 6.0 0 30 � 8.0 8 � 2.6 – – 0.3 � 0.17 0
Unidentified 2005 – – 18 � 9.2 13 � 4.2 6 � 4.2 0 0.1 � 0.04 0.1 � 0.072006 0 0 1 � 1.3 1 � 1.3 – – 0.3 � 0.17 0
Total 2005 – – 143 � 20.1 643 � 118.6 348 � 43.5 461 � 49.2 22.5 � 3.37 50.8 � 13.272006 100 100 81 � 13.4 79 � 10.1 – – 10.1 � 1.96 6.7 � 0.97
534 Marteinsdottir, Bryndıs et al.
significantly higher at site B than site A (w1,562 5 4.89,
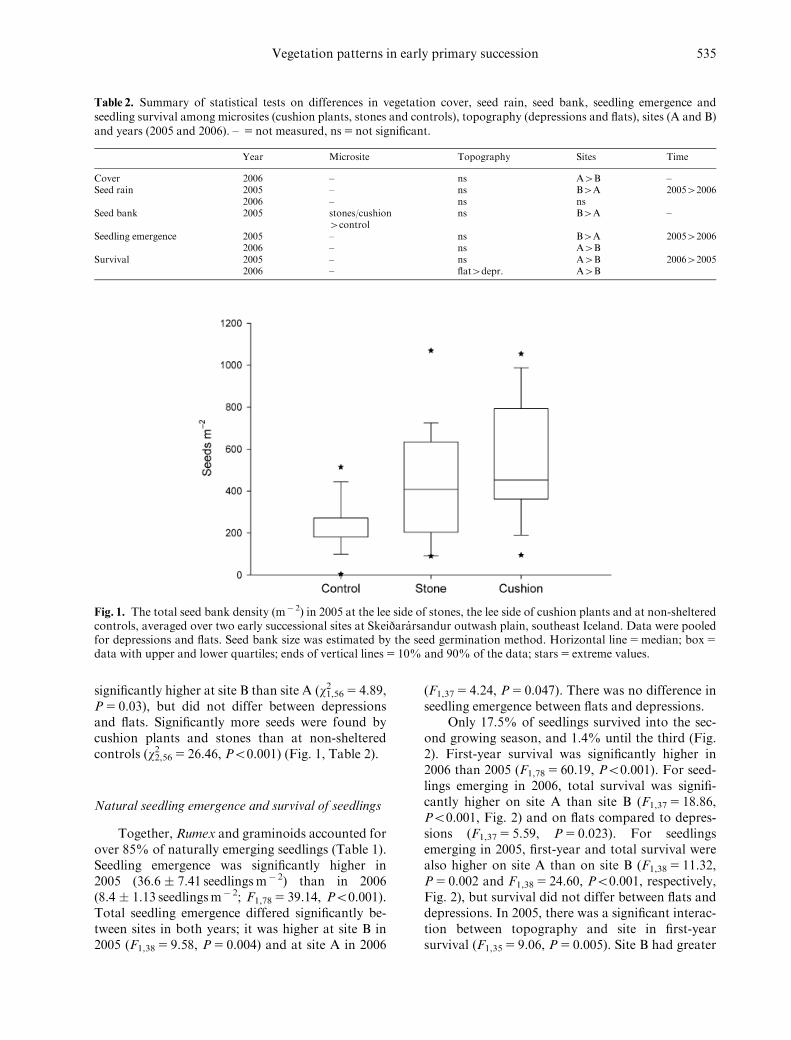
P5 0.03), but did not differ between depressionsand flats. Significantly more seeds were found bycushion plants and stones than at non-shelteredcontrols (w2,56
2 5 26.46, Po0.001) (Fig. 1, Table 2).
Natural seedling emergence and survival of seedlings
Together, Rumex and graminoids accounted forover 85% of naturally emerging seedlings (Table 1).Seedling emergence was significantly higher in2005 (36.6 � 7.41 seedlingsm� 2) than in 2006(8.4 � 1.13 seedlingsm� 2; F1,78 5 39.14, Po0.001).Total seedling emergence differed significantly be-tween sites in both years; it was higher at site B in2005 (F1,38 5 9.58, P5 0.004) and at site A in 2006
(F1,37 5 4.24, P5 0.047). There was no difference inseedling emergence between flats and depressions.
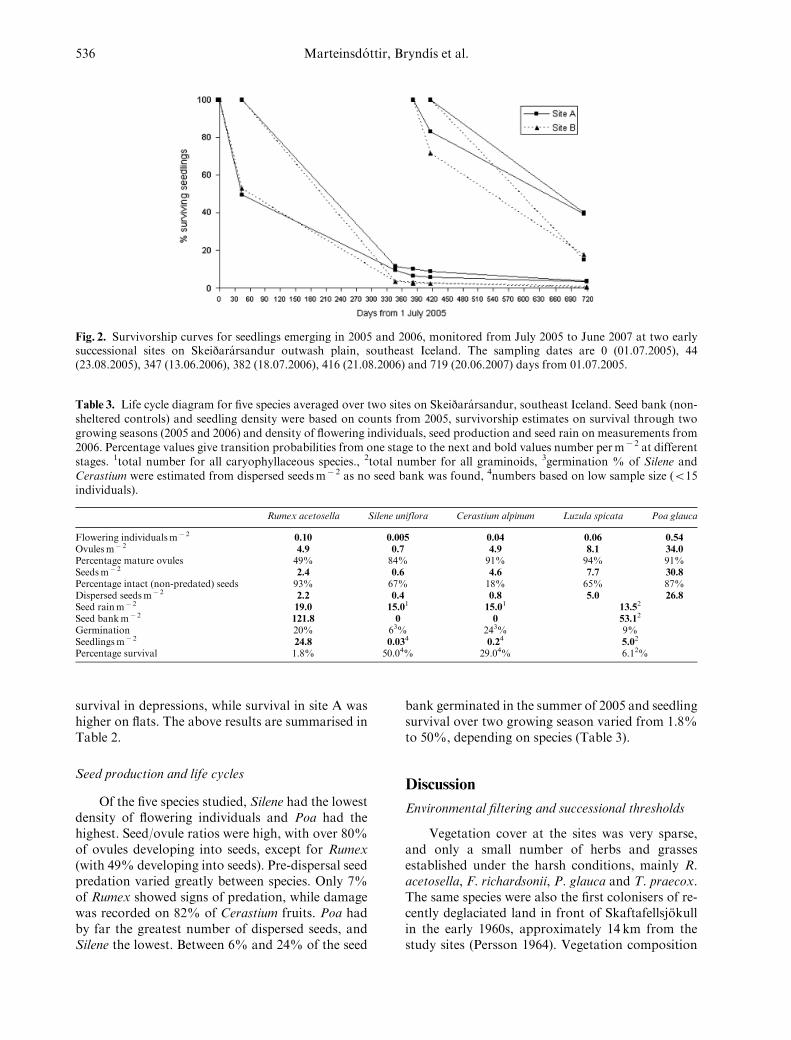
Only 17.5% of seedlings survived into the sec-ond growing season, and 1.4% until the third (Fig.2). First-year survival was significantly higher in2006 than 2005 (F1,78 5 60.19, Po0.001). For seed-lings emerging in 2006, total survival was signifi-cantly higher on site A than site B (F1,37 5 18.86,Po0.001, Fig. 2) and on flats compared to depres-sions (F1,37 5 5.59, P5 0.023). For seedlingsemerging in 2005, first-year and total survival werealso higher on site A than on site B (F1,38 5 11.32,P5 0.002 and F1,38 5 24.60, Po0.001, respectively,Fig. 2), but survival did not differ between flats anddepressions. In 2005, there was a significant interac-tion between topography and site in first-yearsurvival (F1,35 5 9.06, P5 0.005). Site B had greater
Table 2. Summary of statistical tests on differences in vegetation cover, seed rain, seed bank, seedling emergence andseedling survival among microsites (cushion plants, stones and controls), topography (depressions and flats), sites (A and B)and years (2005 and 2006). – 5 not measured, ns5 not significant.
Year Microsite Topography Sites Time
Cover 2006 – ns A4B –Seed rain 2005 – ns B4A 200542006
2006 – ns nsSeed bank 2005 stones/cushion ns B4A –
4controlSeedling emergence 2005 – ns B4A 200542006
2006 – ns A4BSurvival 2005 – ns A4B 200642005
2006 – flat4depr. A4B
Fig. 1. The total seed bank density (m� 2) in 2005 at the lee side of stones, the lee side of cushion plants and at non-shelteredcontrols, averaged over two early successional sites at Skeijararsandur outwash plain, southeast Iceland. Data were pooledfor depressions and flats. Seed bank size was estimated by the seed germination method. Horizontal line5median; box5
data with upper and lower quartiles; ends of vertical lines5 10% and 90% of the data; stars5 extreme values.
Vegetation patterns in early primary succession 535
survival in depressions, while survival in site A washigher on flats. The above results are summarised inTable 2.
Seed production and life cycles
Of the five species studied, Silene had the lowestdensity of flowering individuals and Poa had thehighest. Seed/ovule ratios were high, with over 80%of ovules developing into seeds, except for Rumex(with 49% developing into seeds). Pre-dispersal seedpredation varied greatly between species. Only 7%of Rumex showed signs of predation, while damagewas recorded on 82% of Cerastium fruits. Poa hadby far the greatest number of dispersed seeds, andSilene the lowest. Between 6% and 24% of the seed
bank germinated in the summer of 2005 and seedlingsurvival over two growing season varied from 1.8%to 50%, depending on species (Table 3).
Discussion
Environmental filtering and successional thresholds
Vegetation cover at the sites was very sparse,and only a small number of herbs and grassesestablished under the harsh conditions, mainly R.acetosella, F. richardsonii, P. glauca and T. praecox.The same species were also the first colonisers of re-cently deglaciated land in front of Skaftafellsjokullin the early 1960s, approximately 14 km from thestudy sites (Persson 1964). Vegetation composition
Fig. 2. Survivorship curves for seedlings emerging in 2005 and 2006, monitored from July 2005 to June 2007 at two earlysuccessional sites on Skeijararsandur outwash plain, southeast Iceland. The sampling dates are 0 (01.07.2005), 44(23.08.2005), 347 (13.06.2006), 382 (18.07.2006), 416 (21.08.2006) and 719 (20.06.2007) days from 01.07.2005.
Table 3. Life cycle diagram for five species averaged over two sites on Skeijararsandur, southeast Iceland. Seed bank (non-sheltered controls) and seedling density were based on counts from 2005, survivorship estimates on survival through twogrowing seasons (2005 and 2006) and density of flowering individuals, seed production and seed rain on measurements from2006. Percentage values give transition probabilities from one stage to the next and bold values number perm� 2 at differentstages. 1total number for all caryophyllaceous species., 2total number for all graminoids, 3germination % of Silene andCerastium were estimated from dispersed seedsm� 2 as no seed bank was found, 4numbers based on low sample size (o15individuals).
Rumex acetosella Silene uniflora Cerastium alpinum Luzula spicata Poa glauca
Flowering individualsm� 20.10 0.005 0.04 0.06 0.54
Ovulesm� 24.9 0.7 4.9 8.1 34.0
Percentage mature ovules 49% 84% 91% 94% 91%Seedsm� 2 2.4 0.6 4.6 7.7 30.8
Percentage intact (non-predated) seeds 93% 67% 18% 65% 87%Dispersed seedsm� 2
2.2 0.4 0.8 5.0 26.8
Seed rainm� 2 19.0 15.01 15.01 13.52
Seed bankm� 2 121.8 0 0 53.12
Germination 20% 63% 243% 9%Seedlingsm� 2
24.8 0.034
0.24
5.02
Percentage survival 1.8% 50.04% 29.04% 6.12%
536 Marteinsdottir, Bryndıs et al.

was similar to the species composition of the in-coming seed rain and the seed bank. On glacierforeland in the central European Alps, high corre-lations were also found between the seed bank, seedrain and vegetation (Erschbamer et al. 2001), and inprimary succession on Mount St. Helens, Wa-shington, the common species in the seed rain werealso the most common species in the vegetation(Wood & del Moral 2000).
Selection pressures change throughout a plant’scourse of development, and site qualities may bediscordant with respect to life cycle stages, i.e. it maypresent a favourable environment at one stage in theplant life cycle but difficult at another (del Moral etal. 1995; Schupp 1995). In the current study, site Bhad higher average seed rain and seedling densitythan site A, which had higher species richness andvegetation cover, possibly reflecting the higherseedling survival recorded there. Finer soil grain sizefound at site B could possibly explain some of thatdifference, as finer soil is more prone to be blown bythe wind and such sand drift may cause seedlingmortality. Sudden seedling losses in primary succes-sion have been ascribed to unstable substrates, e.g.on pumice plains near Mount St. Helens (Tsuyuzaki& Titus 1996).
Marked annual fluctuations were observed inseedling density, seed rain and in seedling survival.Seed rain and seedling densities were higher in 2005,while seedling survival was higher in 2006, recallingthe life cycle stage discordance mentioned above,although here it is temporal rather than spatial. An-nual fluctuations are well documented in primarysuccession and have often been linked to stochasticfactors, e.g. in emergence of seedlings in primarysuccession onMount St. Helens (Wood & del Moral1987; Titus & del Moral 1998) and seed rain on thefloodplains of the Tanana River (Walker et al.1986). Overall, a spatio-temporal variation was ex-perienced at a large scale (between sites), but not at asmaller scale (within sites).
Only a few studies have attempted to quantifyseed rain for a range of species in early successionalhabitats. Our figures are low, e.g. only about one-fifteenth of the annual seed rain recorded at barrenmid-elevation and early successional site on MountSt. Helens, but comparable to barren sub-alpine sites(Wood & del Moral 2000). However, we only mon-itored the seed rain for one month (during which weassumed most dispersal occurs) and may thereforehave underestimated the total annual seed rain.
Seed rain density and seed bank size of exposedcontrols were of the same order of magnitude. Onlya very small part of the available seed pool emerged
in 2005, implying that plant establishment on Skei-jararsandur was not immediately limited by lack ofseeds. Seedling recruitment was low (36.6 seedlingsm� 2 in 2005 and 8.4 seedlingsm� 2 in 2006), evenwhen compared to other sparsely vegetated areas inIceland (o10% vegetation cover, e.g. Magnusson1974; Elmarsdottir et al. 2003). In barren, early suc-cessional sites near Mount St. Helens, seedlingdensity was estimated at 41.6m� 2 (Tsuyuzaki et al.1997). The very low values on Skeijararsandurstrongly implicate seedling establishment as a majorfilter of plant establishment on these sites.
On average, 17.5% of seedlings survived intothe second growing season and 1.4% until the thirdgrowing season, making juvenile survival anothermajor filter in the colonisation process. Drought isgenerally considered a major limiting factor forplant establishment on barren sites (Noble et al.1979; Wood & del Moral 1987) but in Iceland frostheave has been observed to be one of the main cau-ses of winter mortality of young birch (Aradottir1991) and willow seedlings (Aradottir et al. 2006).No signs of death caused by frost heave were ob-served here, but desiccation clearly contributed toseedling losses. Frequent sand drifts that buried theseedlings were another major cause of mortalityduring the study, and many wire rings used to markseedlings were found buried in sand, especially inearly spring.
Topography and plant establishment
Depressions can trap seeds, create shelter andretain moisture (Harper et al. 1965; Jumpponen etal. 1999; Marone et al. 2004), thus facilitating plantestablishment. Plant preferences for depressionshave frequently been reported at primary succes-sional sites, e.g. for seedling establishment on glacierforeland (Jones & del Moral 2005), pumice barrens(del Moral 1999) and river floodplains (Walker et al.2006). On recently deglaciated terrain, more flower-ing individuals were found in concave rather than onconvex surfaces (Jumpponen et al. 1999). The onlysignificant topographically related difference inplant establishment found here was higher seedlingsurvival on flats than in depressions. A positive re-lationship between depressions and vegetation coverhas been observed in the uppermost, more stable,but still poorly vegetated part, of Skeijararsandur,where mosses were predominant in the vegetation(Geissler 2005; Martin 2007). This begs the questionwhy such a relationship was not found at our studysites. We propose two possible explanations. First,during the frequent and intense storms, sand is more
Vegetation patterns in early primary succession 537

likely to collect in depressions and on the lee side ofshallow slopes. Sand scouring and thick sand driftsmay be significant in limiting plant establishmentand growth in depressions, overriding other positivequalities associated with them. A second explana-tion relates to the role of mosses in facilitating theestablishment of vascular plants. In the better vege-tated parts of Skeijararsandur, bryophytes aregenerally more abundant in depressions than onflats (Martin 2007), but bryophyte cover was negli-gible at our study sites. If bryophytes preferdepressions at more stable sites, they could facilitateestablishment of vascular plants by enhancing mi-crobial activities (Bardgett & Walker 2004) andincreasing fertility, water-holding capacity and or-ganic matter of the soil (Sohlberg & Bliss 1984). Apositive association of vascular plant cover with de-pressions might therefore be conditional on theprior establishment of mosses.
Microsite patterns were only assessed for seedbank density. Cushion plants, and to a lesser extentsmall (5–20 cm in diameter) stones, acted as seedtraps on the flat surface of Skeijararsandur. Micro-sites that trap seeds do not necessarily increase plantestablishment (Schupp 1995). This pattern was seenat site A, whereas stones and cushion plants did notenhance the survival of transplanted seedlings(Marteinsdottir 2007). If probabilities of seed ger-mination and seedling survival are independent ofmicrosite, successful recruitment is more likely tooccur at microsites that trap more seeds.
Losses at different life cycle stages
Comparisons between the five species showedthat the main filters acted during different life cyclestages. Excepting Rumex, most ovules developedinto mature seeds, and there was no indication thatthe growing season was too short for seed matura-tion. Local seed dispersal was limited by the lowdensity of flowering individuals for all species butother factors, notably pre-dispersal seed predation,were important as well.
The results suggest that the Rumex seed maymostly represent an external seed source as the highseed rain density out-weighed local seed production.Rumex seeds are relatively large but they are oftendispersed together with the membranous perianth,which may enhance dispersal. Effective long-dis-tance dispersal across the flat sandur surface mayoccur during frequent strong winds and storms(Martin 2007). The two graminoids are likely to behighly dependent on local seed production, as theirseed production in 2006 was higher than the seed
rain for all graminoids combined in the same year.The same probably applied to Silene and Cerastium.Rumex and graminoids were the only groups thatcollected sizable seed reserves in the soil, which givesthem an advantage, as establishment is not whollydependent on new seeds arriving each year. Thesegroups also had the highest vegetation cover.
Our results suggest that even during the firststages of succession when vegetation cover is stillvery low, local seed production may be an importantsource of seed. They also indicate that establishmentof Rumex, Luzula and Poa was mostly limited bylack of suitable microsites for seed germination andseedling survival, while the lack of seeds may havebeen most limiting for Silene and Cerastium.
Conclusions
This study demonstrates the importance of bothstochastic factors and environmental filters in plantestablishment in early primary succession. Most ofSkeijararsandur has extremely sparse vegetationcover. Only a few species have been able to establishand survive in its harsh environment despite a rela-tively rich regional species pool. Strong environ-mental filters are implicated as only a fraction ofseeds arriving at the sites become adult plants.
Slight undulations and stones and cushionplants produce the most noticeable spatial patternon the otherwise homogeneous outwash plain. Ex-cept for slight correlations with survival and seedbank density, we were unable to detect a relation-ship of this spatial pattern with plant establishment.Hence, we were unable to identify particular safesites in the sandur environment.
Strong correlations were established betweenthe seed rain, seed bank and species composition.Considering the temporal and spatial variation ob-served in this study, and the impact of sand drifts onseedling survival, we conclude that spatial vegeta-tion patterns in early primary succession onSkeijararsandur are largely determined by stochas-tic factors. This may result in different trajectoriesand rate of succession, even in areas with similarconditions, leading to the development of a vegeta-tion mosaic. The importance of stochastic eventsalso indicates a low predictability in primary suc-cession.
Acknowledgements. We are grateful to all who helped in
the field, in particular Jamie Ann Martin. We thank Rey-
kjavik Botanic Gardens for their support in lending us
538 Marteinsdottir, Bryndıs et al.

greenhouse space for the seed germination experiment.
This study was funded by The Research Fund of Rannıs
(The Icelandic Centre for Research; Grant 040263031)
and a Graduate Scholarship (B.M.) from The Graduate
Research Fund of Rannıs (Grant 050210005). Finally, we
thank Roger del Moral, Ove Eriksson, Hans Henrik
Bruun and two anonymous reviewers for commenting on
the manuscript.
References
Aradottir, A.L. 1991. Population biology and stand
development of birch (Betula pubescens Ehrh.) on
disturbed sites in Iceland. PhD thesis, Texas A&M
University, College Station, Texas, US.
Aradottir, A.L., Svavarsdottir, K. & Magnusson, S.H.
2006. Landnam vıjis og arangur vıjisaninga
(Establishment of willows and evaluation of seeding
experiments). In: Svavarsdottir, K. (ed.) Innlendar
vıjitegundir. Lıffræji og notkunarmoguleikar ı
landgræjslu (Native willows in Iceland: Biology and
potential use for reclamation). pp. 59–72. Landgræjsla
Islands, Gunnarsholt, IS (In Icelandic).
Arnalds, O. 2004. Volcanic soils of Iceland. Catena 56:
3–20.
Anon (Icelandic Meteorological Office). 2007. Data from
the synoptic weather station on Fagurholsmyri.
Bardgett, R.D. & Walker, L.R. 2004. Impact of coloniser
plant species on the development of decomposer
microbial communities following deglaciation. Soil
Biology & Biochemistry 36: 555–559.
Baskin, C.C. & Baskin, J.M. 1998. Seeds. Ecology,
biogeography, and evolution of dormancy and
germination. Academic Press, San Diego, CA, US.
Blundon, D.J., Macisaac, D.A. & Dale, M.R.T. 1993.
Nucleation during primary succession in the Canadian
Rockies. Canadian Journal of Botany 71: 1093–1096.
Bossuyt, B. & Hermy, M. 2004. Seed bank assembly
follows vegetation succession in dune slacks. Journal
of Vegetation Science 15: 449–456.
Callaway, R.M. 1995. Positive interactions among plants.
Botanical Review 61: 306–349.
Cargill, S.M. & Chapin, F.S. 1987. Application of
successional theory to tundra restoration - a review.
Arctic and Alpine Research 19: 366–372.
Castellanos, E.M., Figueroa, M.E. & Davy, A.J. 1994.
Nucleation and facilitation in salt-marsh succession:
interactions between Spartina maritima and
Arthrocnemum perenne. Journal of Ecology 82: 239–
248.
Crawley, M.J. 2002. Statistical computing. An introduction
to data analysis using S-plus. Wiley, London, UK.
del Moral, R. 1999. Plant succession on pumice at Mount
St. Helens, Washington. American Midland Naturalist
141: 101–114.
del Moral, R. & Bliss, L.C. 1993. Mechanisms of primary
succession: insights resulting from the eruption of
Mount St. Helens. Advances in Ecological Research
24: 1–66.
del Moral, R. & Lacher, I.L. 2005. Vegetation patterns 25
years after the eruption of Mount St. Helens
Washington, USA. American Journal of Botany 92:
1948–1956.
del Moral, R., Titus, J.H. & Cook, A.M. 1995. Early
primary succession onMount St. Helens, Washington,
USA. Journal of Vegetation Science 6: 107–120.
del Moral, R., Wood, D.M. & Titus, J.H. 2005. Proximity,
microsites, and biotic interactions during early
succession. In: Dale, V.H., Swanson, F. & Crisafulli,
C. (eds.) Ecological responses to the 1980 Eruption of
Mount St. Helens. pp. 93–110. Springer-Verlag, Berlin,
DE.
Elmarsdottir, A., Aradottir, A.L. & Trlica, M.J. 2003.
Microsite availability and establishment of native
species on degraded and reclaimed sites. Journal of
Applied Ecology 40: 815–823.
Eriksson, O. & Ehrlen, J. 1992. Seed and microsite
limitation of recruitment in plant-populations.
Oecologia 91: 360–364.
Erschbamer, B., Kneringer, E. & Schlag, R.N. 2001. Seed
rain, soil seed bank, seedling recruitment, and survival
of seedlings on a glacier foreland in the Central Alps.
Flora 196: 304–312.
Fastie, C.L. 1995. Causes and ecosystem consequences of
multiple pathways of primary succession at Glacier
Bay, Alaska. Ecology 76: 1899–1916.
Franks, S.J. 2003. Facilitation in multiple life-history
stages: evidence for nucleated succession in coastal
dunes. Plant Ecology 168: 1–11.
Geissler, J. 2005. Small-scale vegetation patterns in early
successional stage at Skeidararsand, SE-Iceland. MSc
Thesis, WestfalischeWilliams Universitat, Munster, DE.
Harper, J.L., Williams, J.T. & Sager, G.R. 1965. The
behaviour of seed in soil. I. The heterogeneity of soil
surfaces and its role in determining the establishment
of plants from seed. Journal of Ecology 53: 273–386.
Jones, C.C. & del Moral, R. 2005. Effects of microsite
conditions on seedling establishment on the foreland
of Coleman Glacier, Washington. Journal of
Vegetation Science 16: 293–300.
Jumpponen, A., Vare, H., Mattson, K.G., Ohtonen, R. &
Trappe, J.M. 1999. Characterization of ‘safe sites’ for
pioneers in primary succession on recently deglaciated
terrain. Journal of Ecology 87: 98–105.
Kristinsson, H. 2001. A guide to the flowering plants and
ferns of Iceland. Mal og menning, Reykjavık, IS.
Kristinsson, H. 2009. Flora of Iceland. Available at: http://
www.floraislands.is/engflora.htm,Accessed 1March 2009.
Larsson, E.L. & Molau, U. 2001. Snowbeds trapping seed
rain – a comparison of methods. Nordic Journal of
Botany 21: 385–392.
Magnusson, S.H. 1974. Plant colonization of eroded areas
in Iceland. PhD thesis, Lund University, SE.
Marone, L., Cueto, V.R., Milesi, F.A. & de Casenave, J.L.
2004. Soil seed bank composition over desert
Vegetation patterns in early primary succession 539

microhabitats: patterns and plausible mechanisms.
Canadian Journal of Botany 82: 1809–1816.
Marteinsdottir, B. 2007. Small scale spatial patterns and
colonization proccesses in early successional
environment. MSc thesis, University of Iceland,
Reykjavık, IS.
Martin, J.A. 2007. The ecology of kettleholes in
successional environment: Skeijararsandur, Iceland.
MSc thesis, University of Iceland, Reykjavık, IS.
Molau, U. & Larsson, E.L. 2000. Seed rain and seed bank
along an alpine altitudinal gradient in Swedish
Lapland. Canadian Journal of Botany 78: 728–747.
Moody, M.E. &Mack, R.N. 1988. Controlling the spread
of plant invasions – the importance of nascent foci.
Journal of Applied Ecology 25: 1009–1021.
Noble, J.C., Bell, A.D. & Harper, J.L. 1979. Population
biology of plants with clonal growth. 1. Morphology
and structural demography of Carex arenaria. Journal
of Ecology 67: 983–1008.
Persson, A. 1964. The vegetation at the margin of the
receding glacier Skaftafellsjokull, southeastern
Iceland. Botaniska Notiser 117: 323–354.
Pettersson, M.W. 1994. Large plant size counteracts early
seed predation during the extended flowering season
of a Silene uniflora (Caryophyllaceae) population.
Ecography 17: 264–271.
Quinn, G.P. & Keough, M.J. 2002. Experimental design
and data analysis for biologists. Cambridge University
Press, Cambridge, UK.
Ryvarden, L. 1971. Studies in seed dispersal 1. Trapping
of diaspores in the alpine zone at Finse, Norway.
Norwegian Journal of Botany 18: 215–226.
Ryvarden, L. 1975. Studies in seed dispersal 2. Winter-
dispersed species at Finse, Norway.Norwegian Journal
of Botany 22: 21–24.
Schupp, E.W. 1995. Seed–seedling conflicts, habitat
choice, and patterns of plant recruitment. American
Journal of Botany 82: 399–409.
Sohlberg, E.H. & Bliss, L.C. 1984. Microscale pattern of
vascular plant distribution in two high arctic plant
communities.Canadian Journal of Botany 62: 2033–2042.
Titus, J.H. & del Moral, R. 1998. Seedling establishment
in different microsites on Mount St. Helens,
Washington, USA. Plant Ecology 134: 13–26.
Tsuyuzaki, S. & Titus, J.H. 1996. Vegetation development
patterns in erosive areas on the Pumice Plains of
Mount St. Helens. American Midland Naturalist 135:
172–177.
Tsuyuzaki, S., Titus, J.H. & del Moral, R. 1997. Seedling
establishment patterns on the Pumice Plain, Mount St.
Helens, Washington. Journal of Vegetation Science 8:
727–734.
Walker, L.R., Zasada, J.C. & Chapin, F.S. 1986. The role
of life-history processes in primary succession on an
Alaskan floodplain. Ecology 67: 1243–1253.
Walker, L.R., Bellingham, P.J. & Peltzer, D.A. 2006.
Plant characteristics are poor predictors of microsite
colonization during the first two years of primary
succession. Journal of Vegetation Science 17: 397–406.
Wood, D.M. & del Moral, R. 1987. Mechanisms of early
primary succession in sub-alpine habitats on Mount
St. Helens. Ecology 68: 780–790.
Wood, D.M. & del Moral, R. 2000. Seed rain during early
primary succession onMount St. Helens, Washington.
Madronos 47: 1–9.
Received 24 April 2009;
Accepted 30 November 2009.
Co-ordinating Editor: Dr. Hans Henrik Bruun.
540 Marteinsdottir, Bryndıs et al.