development of motoneurons and primary sensory afferents in the thoracic and lumbar spinal cord of...
TRANSCRIPT

Development of Motoneurons andPrimary Sensory Afferents in theThoracic and Lumbar Spinal Cordof the South American Opossum
Monodelphis domestica
GRAHAM W. KNOTT,1* PETER D. KITCHENER,2 AND NORMAN R. SAUNDERS3
1Institut de Biologie Cellulaire et de Morphologie, Universite de Lausanne,Lausanne 1005, Switzerland
2Department of Anatomy and Cell Biology, The University of Melbourne, Parkville,Victoria, Australia 3052
3Department of Anatomy and Physiology, University of Tasmania, Hobart, Australia 7001
ABSTRACTThe postnatal development of the primary sensory afferent projection to the thoracic (T4)
and lumbar (L4) spinal cord of the marsupial species Monodelphis domestica was studied byusing anterograde and retrograde neuronal tracers. Large numbers of primary afferents andmotoneurons were labelled by application of the carbocyanine dye DiI into individual dorsalroot ganglia (DRG) afferents in short-term organ cultures. Dorsal root axons had entered thecord at birth, but most primary afferent innervation of the grey matter and the establishmentof cytoarchitectural lamination occurs postnatally. In addition to ipsilateral projections, someprimary afferents that projected to the dorsal horn extended across the midline into theequivalent contralateral regions of the grey matter. Similarly, motoneuron dendrites occasion-ally extended across midline and into the contralateral grey matter. The first fibresinnervating the spinal cord project to the ventral horn and formed increasingly complexterminal arbours in the motor columns between P1 and P7. After P5 many afferents were seenprojecting to the dorsal horn, with the superficial dorsal horn being the last region of thespinal grey to be innervated. Histochemical labelling with the lectin Griffonia simplicifoliaindicated that C fibre primary afferents had arborised in the superficial dorsal horn by P14.The sequence of primary afferent innervation is thus similar to that described in the rat, butthis sequence occurs over a period of several weeks in Monodelphis, compared with severaldays in the rat. J. Comp. Neurol. 414:423–436, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: carbocyanine dye; marsupial; dorsal horn; GSA
The morphological and functional development of spinalcord afferents and efferents has been described in anumber of mammalian and nonmammalian species. In allmammalian species described to date, which includes rat(Smith, 1983; Kudo and Yamada, 1987; Mirnics and Koer-ber, 1995), sheep (Rees et al., 1995), human (Konstan-tinidou et al., 1995), and mouse (Ozaki and Snyder, 1997),the projection of afferents to the spinal cord and theirarborisation in the spinal grey matter occurs in utero.Innervation of the spinal cord by primary afferents in thesheep begins midgestation and progresses to a maturepattern over the next several months in utero (Rees et al.,1995). The development of primary afferent innervation ofthe human spinal cord appears to be similar, in that it
begins around 3 months in utero and proceeds at arelatively slow pace. In rat and mouse, primary afferentinnervation of the spinal cord occurs late in utero over aperiod of just a few days, although the final pattern ofprojections are resolved over the first few postnatal weeks(Fitzgerald et al., 1994). In the developing chick, themature pattern of primary afferent projections to the
Grant sponsor: Motor Accident Insurance Board Foundation, Tasmania.*Correspondence to: Graham W. Knott, Institut de Biologie Cellulaire et
de Morphologie, Universite de Lausanne, Lausanne 1005, Switzerland.E-mail: [email protected]
Received 26 May 1998; Revised 6 July 1999; Accepted 22 July 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 414:423–436 (1999)
r 1999 WILEY-LISS, INC.

spinal cord is established relatively early in ovo (Davis etal., 1989; O’Donovan et al., 1992; Oppenheim et al., 1988).Marsupial species, in contrast, offer a potentially moreaccessible preparation with which to study primary affer-ent development because they are born at a very immaturestage of development, and much of neuronal developmentoccurs postnatally.
The present study has investigated the developmentand organisation of primary sensory and motor projectionsin the spinal cord of the South American grey short-tailedopossum, Monodelphis domestica. As with other marsupi-als, Monodelphis are born at a very immature stage ofdevelopment, with a gestation period of only 14 days. Thepostnatal developmental period is protracted, with eyeopening occurring during the 5th postnatal week. Despitetheir immaturity at birth, the young are not only able tobreathe and suck but they also make their way to the teat,although it is not clear whether this process involves adegree of help from the mother (Saunders et al., 1998). Theuse of Monodelphis domestica for developmental studieshas been exploited in recent studies of spinal cord regenera-tion after traumatic injury (Nicholls and Saunders, 1996;Varga et al., 1996; Wang et al., 1997; Saunders et al.,1998). It has been shown that after complete transection ofthe thoracic spinal cord by crushing, there is a substantialdegree of normal structural development in the crushedregion and near-normal motor behaviour when the ani-mals have grown to adulthood (Saunders et al., 1998). Theextent to which repair of the spinal cord is due to growth offibres that develop after the time of the crush (as opposedto regeneration of cut axons) is unknown. The interpreta-tion of these lesion studies requires the timetable ofdevelopment of spinal motor and sensory pathways to beconsidered.
The present study set out to describe the general patternof development of motoneurons and primary afferentingrowth and used a number of different methods. Theneuronal tracers DiI (see Honig and Hume, 1986; Gode-ment et al., 1987) and Neurobiotin (Kita and Armstrong,1991) were applied to cells of the dorsal root ganglia (DRG)in fixed and cultured nervous tissue, respectively. Toobtain a clear profile of the arrival of C-fibre afferents inthe dorsal horn, the plant lectin Griffonia Simplicifolia(GSA) was used. Lectins that bind to terminal a-D-galactose, such as GSA lectin and soy bean agglutinin,were shown to label this population of primary afferentsthat have small cell bodies and terminate in the superficiallayers of the dorsal horn of rats and cats (Plenderleith etal., 1989).
MATERIALS AND METHODS
Anaesthesia and tissue fixation
Monodelphis pups of various postnatal ages were ob-tained from the colony maintained within the Universityof Tasmania Animal House (for breeding and care seeFadem et al., 1982; Saunders, 1997; Saunders et al., 1989).The day of birth was defined as postnatal day 0 (P0). Allexperiments were conducted within the guidelines of theNational Health and Medical Research Council (Australia)and with the approval of the University of TasmaniaEthics (Animals) Committee. Young animals were killedwith an overdose of halothane inhalation anaesthetic andimmediately perfused transcardially with 0.1 M phosphate-buffered saline (PBS, 4°C, pH 7.4); once the animals were
completely exsanguinated, the perfusate was changed to4% paraformaldehyde in phosphate buffer (0.1 mM, pH 7.2at 4°C). After perfusion the spinal cord was exposeddorsally by carefully cutting away the vertebral archescovering the spinal cord, taking care not to damage thedorsal roots; preparations were then postfixed overnight inthe same fixative.
General morphology
To study the general morphology and cytoarchitecture ofthe spinal cord at different ages, both thin paraffin (5–7µm) and thick vibratome (80–120 µm, VT1000; Leica,Deerfield, IL) sections were used. Vibratomed sectionswere counterstained with bisbenzimide, and paraffin sec-tions were counterstained with toluidine blue. To collectmaterial for paraffin histology, animals from various post-natal ages were killed and perfused (as described above)with Bouin’s fixative. Immediately after perfusion, tho-racic and lumbar regions of the entire spinal column wereremoved and fixed overnight in Bouin’s fixative.
DiI neuronal tracing
After postfixation in paraformaldehyde, single crystals(,0.1 mm diameter) of fluorescent carbocyanine dye ‘‘DiI’’:(1,18-dioctadecyl 3,3,3838-tetramethylindocarbocyanine per-chlorate, Molecular Probes, Inc., Eugene, OR) were in-serted into the DRG at T4 and L4 (see Godement et al.,1987 for details). In some preparations, the ventral rootswere severed to prevent labelling of ventral root axons andmotoneurons. After crystal insertion, the entire cord wasstored in fixative or in PBS containing 0.1% sodium azide(to prevent bacterial contamination) at room temperature(21°C) or at 37°C. The incubation periods varied between 1and 5 days, depending on the distance between the dyeplacement site and the spinal cord. After incubation, thespinal cords were embedded in 5% agar and cut intotransverse sections (80–120-µm-thickness) on a vibratome.In most cases sections were placed in a 2.5 µg/ml solutionof bisbenzimide (Sigma, St. Louis, MO) in PBS for 10–30minutes to provide a general counterstain. The sectionswere mounted in PBS on glass slides and coverslippedwith use of nail varnish (Australian Colours) to seal theedges. All sections were then examined under fluorescentillumination with a rhodamine filter set for DiI and aultraviolet (UV) filter set for bisbenzimide on an OlympusBX50 microscope. All sections exhibiting DiI fluorescencewere photographed to allow the rostrocaudal extent oflabelled projections to be determined. Some sections werephotoconverted by using diaminobenzidine (DAB, Sigma)to transform the DiI staining into a permanent label;sections mounted in 3.0 mg/ml DAB (in 0.1 M PBS) wereilluminated with rhodamine epifluorescence (310 objec-tive) for 30 minutes. After the sections were washed inPBS, they were then mounted on gelatin-coated slides,air-dried overnight, dehydrated in a graded alcohol series,cleared in xylene, and coverslipped with DPX.
Dorsal root ganglia injection of Neurobiotin
Young Monodelphis at various ages were anaesthetisedand killed as described above and then immersed immedi-ately in sterile Basal Medium Eagle’s (BME, Gibco, LifeTechnologies Ltd., Scotland, United Kingdom), which wasbubbled continuously with 5% CO2 in oxygen (for detailssee Saunders et al., 1992) at room temperature. The entirecentral nervous system (CNS) was then removed from the
424 G.W. KNOTT ET AL.

animal, taking care to leave the dorsal root gangliaattached to the spinal cord at T4 and L5. In the samemedium, organ culture preparations of the Monodelphisnervous system have survived for up to 7 days (Woodwardet al., 1993). Microelectrodes were filled with 3–4 µl of 6%Neurobiotin (Vector Laboratories Inc., Burlingame, CA)with a small amount of methyl green to visualise thesolution. The microelectrode tip was carefully broken backto a wider diameter so that under pressure a small amountof solution could be seen emerging from the tip. A singleinjection (,1 µl) was then made into the T4 and L5 DRGs.Preparations were then left in the culture medium for 4hours before fixation in 4% paraformaldehyde. Vibratomesections were cut as described above, and the Neurobiotintracer localised by using an avidin-biotin-peroxidase com-plex (ABC, Elite kit, Vector Laboratories) and then visual-ised by using DAB (Kita and Armstrong, 1991).
Lectin histochemistry
The binding of the lectin GSA (Vector Laboratories),which binds to terminal a-D-galactose residues, was exam-ined on transverse paraffin sections (5–7-µm-thickness).Sections were dewaxed in xylene, rehydrated in an alcoholseries (100% through to 50%), and washed three times in0.1 M PBS containing 0.2% Tween. A solution of GSA(1:250) and 0.05% gelatin blocker (Amersham, Essex,United Kingdom) was applied directly to sections andincubated overnight at 4°C. The lectin was then visualisedusing ABC as described above. After dehydration andclearing, sections were photographed under Normarskioptics.
Image preparation
Fluorescent images were photographed with Ilford HP5(ISO 400) black and white film and printed on Ilfordmultigrade paper. These prints were subsequently digi-tally scanned with an Agfa Duoscan and assembled on aMacintosh 8600 using Photoshop 4.0. Figures 7 and 9 werephotographed with Kodak Ektachrome 64T film andscanned digitally (Agfa Duoscan). Image brightness andcontrast were adjusted optimally on all scanned imagesand labelled within the Photoshop program. This softwarewas also used to collage together Figures 4 and 5.
RESULTS
Lumbar and thoracic DRG were labelled in a total of 58Monodelphis pups. DiI incubation times varied consider-ably, depending on the age of the pup (24–48 hours at P0and 7 days at P30) to obtain maximum labelling of theentire cell and its processes. However, there was somevariation in the amount of labelling seen within the spinalcord, presumably because of variation in the number ofcells and processes labelled and the distance from the DRGentry zone. This was particularly noticeable in the youngeranimals. At P0, DiI left for too long invariably resulted inexcessive labelling of cells and their processes. This madeit difficult to visualise primary afferents in the dorsal hornor the extent of their projections. Therefore, at theseyounger ages, after insertion of DiI crystals into the DRG,the spinal cords were incubated for no longer than 48hours at 35°C.
Brain and spinal cord preparations, which had beencultured for a few hours immediately after the Neurobiotinhad been injected into the DRG, showed good tissue
preservation only in cases in which the entire vertebraeand dura had been removed from the spinal cord. Necrosisof the nervous tissue was evident within 2 hours if theoxygenated media was not allowed unrestricted access tothe spinal cord and DRGs. The use of Neurobiotin enabledlabelling of just a few fibres without any evidence oftranscellular transfer of this marker. DiI or Neurobiotinbacklabelled motoneurons were observed in all of thespinal cords with uncut ventral roots. Efferent axonsleaving the spinal cord must therefore lie in close proxim-ity to the DRG to absorb these markers. No backlabelledmotoneurons were visible in spinal cords whose ventralroots had been severed before labelling.
Afferent projections into thoracic andlumbar spinal cord at birth
Figure 1 shows toluidine blue-stained transverse paraf-fin sections through thoracic and lumbar spinal cordsegments at P0. Other than the region-specific features ofthe thoracic and lumbar spinal segments, no obviousdifferences appeared in the cytological differentiation be-tween these two levels at this age or at any later agestudied. P0 was the earliest age examined, and the charac-teristic three layers of the spinal cord were evident (neuro-epithelial layer, mantle layer, and marginal layer) in bothregions. A large central spinal canal with thin alar andbasal plates was found. The neuroepithelial layer con-tained a few mitotic figures indicating continued celldivision at the time of birth. Within the mantle layer theintermediolateral and ventral horn was easily distin-guished, containing well-differentiated motoneurons inthoracic and lumbar sections. No evidence of a presump-tive substantia gelatinosa or other laminations within thedorsal horn in either of these regions at P0 was found.
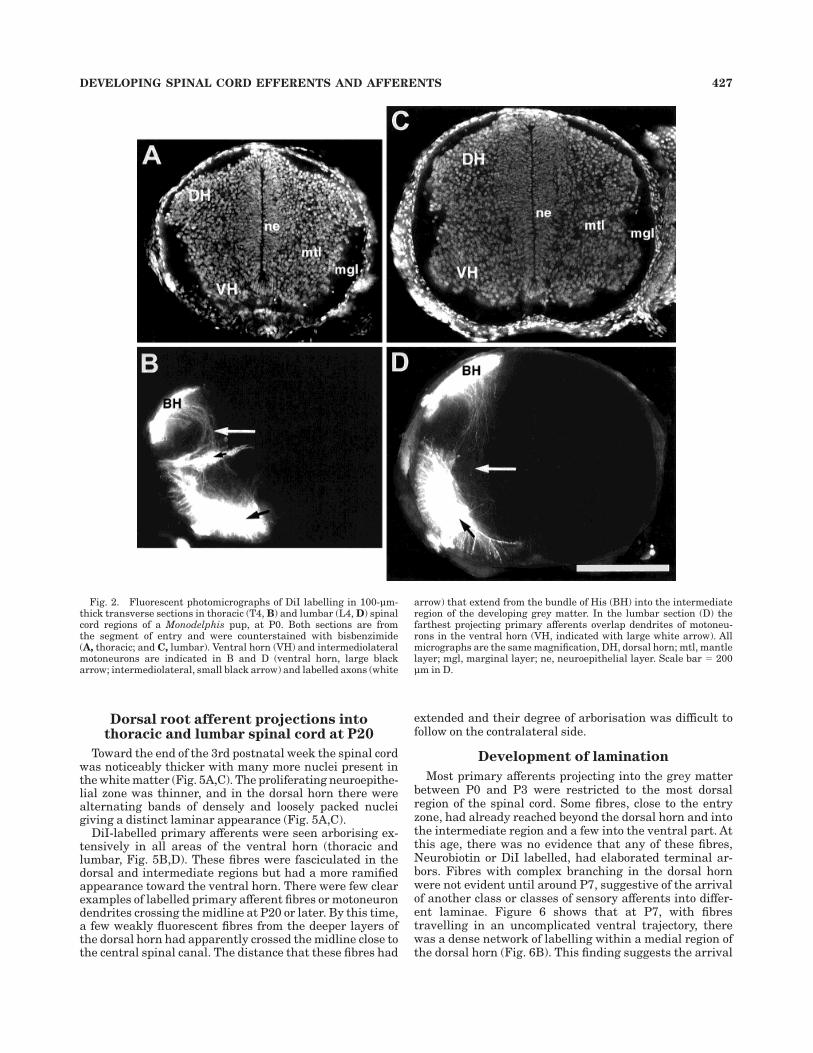
At P0, transverse sections of spinal cord labelled withDiI (Fig. 2) showed labelled fibres in a thin but broaddorsal root entry zone (Fig. 2B, D, and Fig. 3). This zoneextended as far medially as the underlying dorsal horn andlaterally to the most lateral aspect of the presumptivewhite matter. The mediolateral extent of the dorsal rootentry zone was most extensive in lumbar cord of youngeranimals (up to around P7). The penetration of primaryafferents into the developing spinal grey matter wasevident on the day of birth in lumbar and thoracic regionswhere a number of fibres, and small fascicles of fibres,were seen in the dorsal horn with a few projecting throughthe intermediate region toward the ventral horn. Thesefibres appeared to avoid the ventricular zone and projecteddirectly toward the ventral horn, where a significantnumber of motoneurons in the ventral and intermediolat-eral (thoracic) regions had been backlabelled. These groupsof cells occupied a significant area within the ventral hornand a large number of dendrites could be seen radiating tothe outer edges of the cord (Fig. 2B, D).
Injection of the neuronal tracer, Neurobiotin, into the T4and L4 DRG enabled a very small population of primaryafferent fibres to be labelled. At this early stage of develop-ment this method had some advantages over that of DiI;because only a few fibres were Neurobiotin positive, theirentire length could be seen in the transverse sections.Excessive fibre labelling often caused ‘‘flare’’ in the DiIpreparations at such young ages, making it difficult tolocalise single fibres and the exact extent of their growth.Figure 3 shows Neurobiotin-labelled primary afferent fi-bres in the spinal cord at P0. As suggested by the DiI-
DEVELOPING SPINAL CORD EFFERENTS AND AFFERENTS 425

labelled cords (Fig. 2), these afferents extended toward theventral region where fibres tipped with growth cones wereseen (Fig. 3C, D). Figure 3 also shows that in the lumbarsegment a number of motoneurons have been backlabelledin the ventral horn (Fig. 3A,B), and there was a degree ofoverlap between motoneuron dendrites and the few sen-sory afferents that were seen in this region.
Afferent projections into thoracic andlumbar spinal cord after the 1st
postnatal week
After the 1st postnatal week greater numbers of axonsprojecting further ventrally into the cord were seen; thesefibres were closely associated with motoneurons in theventral horn. Actual numbers of primary afferents in thisregion were not possible to estimate because of theiroverlap with motoneuron dendrites; also, the number ofafferents labelled in any particular section depended onhow many DRG cells were labelled and how far from theentry zone the section was taken. Fibres were easilyidentifiable in sections more remote (rostrally and cau-dally) from the dense network of fluorescence in the dorsalhorn near the root entry zone. They could be seen project-ing medially, converging in the intermediate layer fromwhere they ramified ventrally. Another feature of the
labelling shown by these fibres was a certain amount offasciculation, particularly as the axons converged in theintermediate region before arborising in the ventral horn.The bisbenzimide counterstain (Fig. 4A,C) showed thatthe neuroepithelial layer had decreased in thickness,whereas both the alar and basal plates had thickened inthoracic and lumbar regions. At this age the dorsal andventral horns had increased in size along with an enlarge-ment of the presumptive white matter in the dorsal andlateral areas of the cord. DiI-labelled primary afferentfibres occupied a very broad band in the forming whitematter across the entire mediolateral extent of the dorsalregion. Some labelled fibres innervating the dorsal horn inthoracic sections were seen projecting horizontally to themidline and beyond to innervate the corresponding dorsalhorn regions on the contralateral side (Fig. 4B). Some ofthese fibres possessed varicosities suggestive of synapticspecialisations. Similarly, motoneuron dendrites in thecontralateral spinal cord exhibited terminal branching ofcomparable form and complexity to those branches on theipsilateral side. This pattern of crossing the midline wasnot present to any great extent in the lumbar segment.However, like thoracic motoneurons, those in the lumbarcord also showed dendrites extending a short distanceacross the midline.
Fig. 1. Photomicrographs of transverse, 5-µm-thick sections of thethoracic (T4, A) and lumbar (L4, B) spinal cord of Monodelphis, at P0,stained with toluidine blue. In both sections the thick neuroepitheliallayer (ne), mantle layer (mnl) containing large motoneurons in theventral horns (VH) and marginal layer (mgl) are evident (labelled in
B). The dorsal root ganglia (drg) are shown at the left hand edge ofeach micrograph and the central spinal canal (csc) at the right. Bothsections show the thick alar and basilar plates (labelled ap and bas,respectively). Both micrographs are the same magnification. DH,dorsal horn. Scale bar 5 100 µm in A.
426 G.W. KNOTT ET AL.
Dorsal root afferent projections intothoracic and lumbar spinal cord at P20
Toward the end of the 3rd postnatal week the spinal cordwas noticeably thicker with many more nuclei present inthe white matter (Fig. 5A,C). The proliferating neuroepithe-lial zone was thinner, and in the dorsal horn there werealternating bands of densely and loosely packed nucleigiving a distinct laminar appearance (Fig. 5A,C).
DiI-labelled primary afferents were seen arborising ex-tensively in all areas of the ventral horn (thoracic andlumbar, Fig. 5B,D). These fibres were fasciculated in thedorsal and intermediate regions but had a more ramifiedappearance toward the ventral horn. There were few clearexamples of labelled primary afferent fibres or motoneurondendrites crossing the midline at P20 or later. By this time,a few weakly fluorescent fibres from the deeper layers ofthe dorsal horn had apparently crossed the midline close tothe central spinal canal. The distance that these fibres had
extended and their degree of arborisation was difficult tofollow on the contralateral side.
Development of lamination
Most primary afferents projecting into the grey matterbetween P0 and P3 were restricted to the most dorsalregion of the spinal cord. Some fibres, close to the entryzone, had already reached beyond the dorsal horn and intothe intermediate region and a few into the ventral part. Atthis age, there was no evidence that any of these fibres,Neurobiotin or DiI labelled, had elaborated terminal ar-bors. Fibres with complex branching in the dorsal hornwere not evident until around P7, suggestive of the arrivalof another class or classes of sensory afferents into differ-ent laminae. Figure 6 shows that at P7, with fibrestravelling in an uncomplicated ventral trajectory, therewas a dense network of labelling within a medial region ofthe dorsal horn (Fig. 6B). This finding suggests the arrival
Fig. 2. Fluorescent photomicrographs of DiI labelling in 100-µm-thick transverse sections in thoracic (T4, B) and lumbar (L4, D) spinalcord regions of a Monodelphis pup, at P0. Both sections are fromthe segment of entry and were counterstained with bisbenzimide(A, thoracic; and C, lumbar). Ventral horn (VH) and intermediolateralmotoneurons are indicated in B and D (ventral horn, large blackarrow; intermediolateral, small black arrow) and labelled axons (white
arrow) that extend from the bundle of His (BH) into the intermediateregion of the developing grey matter. In the lumbar section (D) thefarthest projecting primary afferents overlap dendrites of motoneu-rons in the ventral horn (VH, indicated with large white arrow). Allmicrographs are the same magnification, DH, dorsal horn; mtl, mantlelayer; mgl, marginal layer; ne, neuroepithelial layer. Scale bar 5 200µm in D.
DEVELOPING SPINAL CORD EFFERENTS AND AFFERENTS 427

of primary afferents to a particular target field situateddeeper in the dorsal horn. This profile was similar in boththe lumbar and thoracic sections. After P7, the moresuperficial layers of the dorsal horn contained DiI-labelledfibres and endings (Fig. 6C,D,E), possibly correlating withthe known target field of small-diameter primary afferents(Fitzgerald, 1987). By the end of the 3rd postnatal week
the entire depth of the dorsal horn (thoracic and lumbar)appeared to be filled with complex arbors.
Labeling afferents in the superficialdorsal horn
The binding of the GSA lectin in the dorsal horn at P14and in adult spinal cord is shown in Figure 7 (Fig. 7A,B).
Fig. 3. Photomicrographs of primary afferent fibres labelled withNeurobiotin in the thoracic (T4, A and C) and lumbar (L4, B and D)regions of the P0 Monodelphis spinal cord. The sections are from thesegment of entry. Micrographs A and B show labelled axons in thedorsal horn (DH) and motoneurons in the ventral horn (VH) of bothsections. Small black arrows indicate, in A and B, endothelial cells of
blood vessels. Micrographs C and D are higher power magnifications ofregions arrowed in A and B (large black arrows) and show fibres tippedwith growth cones (small black arrows) that have extended into thedeveloping grey matter. Micrographs A and B are the same magnifica-tion. Scale bars 5 200 µm in A and 50 µm in C,D.
428 G.W. KNOTT ET AL.

GSA labelling was evident at P14, appearing as sparselabelling in the very superficial and presumably presump-tive lamina II of both thoracic and lumbar sections. At noearlier ages was this labelling apparent in these regions.This lectin marker for C-afferents clearly labels the super-ficial dorsal horn of the mature spinal cord (Fig. 7C,D).Labelling the DRG cells with Neurobiotin, at P10 (Fig. 8),4 days before the first GSA binding in the dorsal hornfailed to show the presence of any fibre endings, orbranched axons, within this superficial region of the dorsalhorn. This finding suggests that 2 weeks postnatal wasabout the time that C-fibre afferents first arrive in the
superficial dorsal horn and is substantiated by the DiIlabelling seen at P15 (Fig. 6C) showing dense clusters thathad the morphology of terminal boutons in the superficialdorsal horn.
Development of motoneurondendritic arborisation
Lateral and medial motor column. Motoneurons inthe lateral and medial motor columns were backlabelledfrom the 1st postnatal day (P0) onward. Dendrites fromthese neurons were restricted to the ventral horn and the
Fig. 4. Fluorescent micrographs showing DiI labelling in 100-µm-thick transverse sections (B and D) counterstained with bisbenzimide(A,C) from the thoracic (T4) and lumbar (L4) spinal cord of Monodel-phis pups at P7. Sections are from the segment of entry. DiI-labelledventral horn (VH) and intermediolateral (IML) motoneurons (indi-cated with large white arrows and a small white arrow, respectively, inB and D) and axons extending dorsoventrally through the developinggrey matter on the ipsilateral side are present. Note the thin neuroepi-thelial layer and densely packed nuclei within the dorsal horns (DH,
upper micrographs). Labelled primary afferents can be seen across theentire mediolateral extent of the dorsal horn where there is anexuberance of labelling. Primary afferents ramify within the ventralhorn (VH) and are closely associated with dendrites of backlabelledmotoneurons. Labelled fibres, in the dorsal horn, can also be seencrossing the midline (indicated with stars) in the thoracic segment. Allmicrographs are the same magnification. Prints included in C and Dwere scanned digitally and collaged by using Photoshop (see Materialsand Methods). Scale bar 5 200 µm in D.
DEVELOPING SPINAL CORD EFFERENTS AND AFFERENTS 429

Figure 5

adjacent presumptive white matter during the first 3postnatal days. By P7, dendrites could be seen extendinginto the intermediate region, particularly in the lateraledge of the cord (Fig. 4B,D). These more extensive den-dritic patterns were difficult to examine in thoracic sec-tions because of the presence of backlabelled intermediolat-eral motoneurons and their processes.At this age, processesfrom these motoneurons were seen crossing the midline ina region of the ventral commissure. Occasionally, motoneu-rons in the ventral contralateral region were also backla-belled, although this was rare. This may be due to tran-sient and inappropriate axonal growth by the motoneuronitself because this phenomenon was never seen after P15in either the thoracic or lumbar region.
Throughout the time period studied in these experi-ments (P0–P30) motoneurons backlabelled from a singleDRG occupied a narrower region rostrocaudally than thefield covered by the afferent projections. It appeared thatmaximum numbers of motoneurons were backlabelled atthe level of entry of dorsal roots into the spinal cord. Thismotoneuron labelling decreased farther away from theentry point until, in the adjacent segments, only labelledprimary afferents could be seen in the ventral horn.Although it was apparent that later in development thedendrites from motoneurons became more exuberant, theirrostrocaudal projection fields have not been followed inthis study.
Intermediolateral column. In the thoracic region, in-termediolateral column motoneurons were backlabelledfrom the 1st postnatal day (P0). At this early age, den-drites of these neurons did not extend laterally beyond thegrey matter or medially past the boundary of the neuroepi-thelial zone and mantle region. At P7, dendrites were seenas far laterally as the extreme edge of the presumptivewhite matter and had fasciculated into a contralaterallyprojecting bundle that had grown dorsally and close to thecentral spinal canal. By P20 this region had adopted itsmature pattern as a series of horizontally arranged backla-belled cell bodies whose dendrites were lined up along themediolateral axis within the grey matter.
Rostrocaudal projection of primary afferentsearly in development
The projections of primary afferents in the grey matteralong the rostrocaudal axis at the time of birth (P0) did notappear to be confined to the segment of origin. In some
animals single crystals of DiI were inserted into the DRGat T4, and the ventral roots in this segment were cut toavoid backlabelling of any motoneurons (Fig. 9). Thelength of the T4 segment of spinal cord at this age (P0) wasapproximately 400 µm (Fig. 9A). However, labelled fibreswithin the grey matter were spread over a distance .700µm in three animals whose spinal cords in this region weresectioned transversely, therefore showing considerableoverlap with the neighbouring segments. These fibresappeared to be unbranched and projected into the dorsalhorn and beyond in a straight-line trajectory. The densityof fibres entering the dorsal horn decreased progressivelyat sites farther from the segment of entry.
In the L4 segment, the intersegmental distance wasapproximately 700 µm, and a single DiI crystal was placedin DRG-labelled primary afferents penetrating the greymatter rostrocaudally over a distance .1,400 µm. Like theafferents in the thoracic segments, fibres appeared to beunbranched and had only grown a short distance towardthe midline. This confirmed the labelling seen in thetransverse sections in which fibres were absent from theventricular zone. These measurements give an indicationof the rostrocaudal spread of primary afferent fibres atbirth, assuming that a reasonably representative portionof the sensory neurons was labelled with the single crystalplacement in the DRG of each preparation.
After the 1st postnatal week (Fig. 9B,D), in threeseparate cords sectioned horizontally, both thoracic andlumbar regions showed that labelled fibres had grown intothe dorsal horn of the adjacent segments and the segmentof entry. Therefore, the rostrocaudal extent of primaryafferent ingrowth, into the developing grey matter, over-lapped into the adjacent segments. However, unlike thenewborn spinal cord, these fibres often had small collater-als and more complex trajectories (Fig. 9C). Sections ofspinal cord farther from the segment of entry, similar toP0, showed fewer labelled fibres, but there was no indica-tion that the primary afferents lagged behind those closerto the entry region. At P20 the rostrocaudal extent ofprimary afferent projections in the thoracic segment hadoverlapped into the adjacent segments rostral and caudalto the segment of entry (intersegmental distance was,700 µm). However, in three spinal cords studied therewas a large variation in the degree of overlap, presumablya function of the number of DiI-labelled cells in the DRG.
DISCUSSION
This study has demonstrated that in the marsupialspecies Monodelphis domestica the overall pattern ofdevelopment of primary afferent projections during thefirst few weeks of postnatal life is broadly similar to that ineutherian species. However, this pattern of developmentoccurs within a different time frame with respect to birth,much of it occurring ex utero. On the day of birth, primaryafferents had already penetrated into the dorsal horn inboth the thoracic and lumbar segments, but few of theseafferents had reached beyond the intermediate region. Atbirth, Monodelphis is at a stage of development thatcorresponds approximately to a human embryo of earlierthan 8 weeks of gestation (Konstantinidou et al., 1995) andrat embryos between E16–E17 (Mirnics and Koerber,1995) and an E14–E15 mouse (Ozaki and Snider, 1997). Itwas previously shown that at P4 in Monodelphis, primaryafferents of the lumbosacral region are abundant in the
Fig. 5. Fluorescent photomicrographs showing DiI labelling (B,D)in 100-µm-thick transverse sections counterstained with bisbenzimide(A,C) in the thoracic (T4) and lumbar (L4) segments of the Monodel-phis spinal cord, at P20. The sections are from the segment of entry.DiI-labelled afferent fibres crowd the entire thickness of the dorsalhorn (DH) with some projecting into the ventral horn (VH). Backla-belled motoneurons are indicated in the ventral horns of both sections(large white arrows) and in the intermediolateral region (IML) in B(small white arrow). Ventral horn motoneurons have labelled den-drites, a few of which cross the midline into the contralateral side ofthe spinal cord (indicated with star in B and D). Micrograph D shows asuperficial layer (indicated with small white arrow) in the dorsal hornwith abundant fascicles of fibres projecting to deeper layers andcontaining few fine endings of primary afferents. Note also thedifferent densities of nuclei in the dorsal horn (A and C), indicatingthe formation of different laminae. All micrographs are the samemagnification. The prints in this figure were scanned digitally andcollaged by using Photoshop (see Materials and Methods). Scale bar 5200 µm in D.
DEVELOPING SPINAL CORD EFFERENTS AND AFFERENTS 431

Fig. 6. Fluorescent photomicrographs of DiI labelling in 100-µm-thick sections of the dorsal horn in the thoracic spinal cord ofMonodelphis at five different postnatal ages (A, P0; B, P7; C, P15;D, P22 and E, P30). All sections are from the segment of entry. Medialis to the right and lateral to the left in each micrograph. At P0 (A)labelled fibres present in the superficial region of the dorsal hornappear to have simple trajectories. At P7 (B) and later thin fascicles offibres traverse this superficial portion of the dorsal horn. At older ages
(P22, D and P30, E) the outer layer appears to be filled with a punctatelabelling (indicated with arrows in D and E). This type of fibrelabelling is sparse in the same region at P15 (C) and not evident atearlier ages. Fascicles of fibres projecting through the dorsal horn(seen from P7 onward and indicated with a white arrow in P30, E) areprominent more medially with a few fascicles seen at the lateral edge.Magnification is the same in all micrographs. Scale bar 5 100 µmin E.
432 G.W. KNOTT ET AL.

ventral horn, where they overlap with dorsally directedmotoneurons (Qin et al., 1993); this is confirmed in thepresent study (note that Qin et al. designate the day ofbirth as P1, making P4 equivalent to P3). In the opossumspecies Didelphis virginiana, ventral horn afferents couldalso be seen within the grey matter of the lumbar region,in close association with motoneuron dendrites, on the dayof birth (Wang et al., 1997).
By P7 primary afferents had reached the ventral horn,indicating that these fibres were probably class Ia fibresfrom muscle spindles (see Brown, 1981). As with thedemonstrated trajectories of ventral horn afferents in therat (Mirnics and Koerber, 1995) and mouse (Ozaki andSnider, 1997), they enter the dorsal horn toward the
medial aspect of the grey matter. This pattern was notobserved in the human embryo, where they enter thedorsal horn along its entire dorsolateral extent (Konstan-tinidou et al., 1995). The arrival and arborisation in thelumbar region of ventral horn afferents at P7 provides thebasic circuit for the development of a simple stretch reflex.In the cervical region in Monodelphis this pathway hasbeen shown to be present during the first postnatal week,by using electrophysiological analysis (Lepre et al., 1998).Whether the lumbar region would have the same degree ofconnectivity at this age remains unclear. Previous reportshave contributed some information on spinal circuit func-tional development in Monodelphis. Cassidy et al. (1994)examined the development of sensorimotor reflexes in thisspecies but not before P15. More recently, however, Saun-ders et al. (1998) observed vigorous ipsilateral and contra-lateral writhing movements from the hindlimbs at P7when squeezed with forceps; the nature of the afferentssubserving this response is not known. After the 1stpostnatal week many fibres were present and evidentlyterminating in the dorsal horn along the mediolateralextent of a region, presumably corresponding to layers IIIand IV. DiI labelling in this region gave this layer a denseand complex appearance, whereas Neurobiotin revealedthe detailed morphology of these fibres. These fibresterminating in the deeper dorsal horn possessed terminalpatterns typical of low-threshold afferents.
Visualisation of single small-calibre axons with DiI maybe difficult at later stages of development because of adecreased ability of DiI to diffuse along such fibres(Burkhalter et al., 1993). After placing DiI crystals on theentry zone of the dorsal roots, Konstantinidou et al. (1995)were unable to label any fibres in layers I and II of thehuman spinal cord. In the present study GSA was used toverify the DiI labelling, as was shown recently by Fernan-dez and Nicholls (1998), using the electron microscope,that even on the day of birth, in Monodelphis, groups ofsmall and large diameter axons are present in the centralstump of the dorsal root in the cervical region. The GSAstaining indicated the presence of C-fibre afferents withinthe presumptive layers I and II at P14. There was noevidence of GSA-labelled afferents in the grey matter atearlier ages examined. This suggests that toward the endof the 2nd postnatal week C-fibres began to arrive in thedorsal horn in Monodelphis. This also corresponds withthe appearance of DiI-labelled structures in this regionwith morphologies suggestive of synaptic terminals.
Therefore, it would seem that the development of projec-tions from cells in the DRG occurs in a sequential mannerreaching targets deeper in the grey matter before the moresuperficial ones, a pattern that is similar to other mamma-lian species (Willis and Coggeshall, 1991). However, thetime course of this process is somewhat different indifferent species; in the rat, Ia afferents first reach themotor pool at E16 and the innervation of layers I and II bysmall-diameter fibres appears to be well advanced by E20(Fitzgerald, 1987). It would seem that this process inMonodelphis occurs in the first 2 weeks postnatally.
Comparison of thoracicand lumbar development
A number of studies using eutherian mammalian spe-cies have stated that a rostrocaudal ‘‘developmental gradi-ent’’ exists early in spinal cord formation when consideringthe spinal cord enlargements in the cervical and lumbar
Fig. 7. Photomicrographs showing staining of the GSA lectin in thedorsal horn (DH) of 5-µm-thick paraffin sections from the thoracic (Aand C) and lumbar (B and D) regions of the Monodelphis spinal cord.Medial is to the right and lateral to the left with the central spinalcanal (csc) indicated at the righthand edge in A and B. In the adult (Cand D) there is dense labelling in the superficial laminae of the dorsalhorn (indicated with black arrows). P14 (A and B) was the earliest agein which GSA labelling was evident with a fine punctate DAB stainingin the more superficial layers of the dorsal horn (black arrows).Photomicrographs were taken by using Normarski optics. Endothelialcells of capillaries are also labelled. Magnification is the same in allmicrographs. Scale bar 5 100 µm in A.
DEVELOPING SPINAL CORD EFFERENTS AND AFFERENTS 433

regions (Altman and Bayer, 1984). This finding is based onthe clear differences shown morphologically and electro-physiologically between these two levels. In the Monodel-phis there are clear differences in limb development(Cassidy and Cabana, 1993), spinal reflexes (Cassidy et al.,1994), and also in the myelination of the dorsal and ventralroots (Leblond and Cabana, 1997). It might be expected,therefore, that if there is a gradient in the maturation ofthe spinal cord, the innervation of the lumbar spinal cordby primary afferents would be delayed with respect to thethoracic region. Such a temporal difference in the patternof innervation was not observed. The present study hasnot, however, examined the cervical region, whose develop-ment may be more precocious in accord with the earlydevelopment of forelimbs compared with hindlimbs. Thepresent study examined general aspects of spinal corddevelopment and focused on the regions of the spinal cordrelevant to the experiments addressing the recovery oflocomotor function after thoracic lesions in neonatal Mono-delphis (Saunders et al., 1998).
Contralateral projectionsof primary afferents
Primary afferent fibres, which project to the contralat-eral side of the spinal cord, have been demonstrated atdifferent segmental levels in different stages of develop-ment in the rat (Smith, 1983; Smith, 1986). In the Monodel-
phis they appear to be more numerous in the thoracic thanin the lumbar segment, which is consistent with thelikelihood that a greater proportion of thoracic afferentsrelay information from the axial skin and muscle, there-fore maintaining somatotopic continuity across the dorsaland ventral midline, as shown in other species (e.g., cat,Ritz et al., 1985). At P30, the oldest age studied with DiI, itwas difficult to visualise these fibres, but in the two cordsexamined their number seemed to have decreased mark-edly in the thoracic segment and was similar to that in thelumbar segment. In other species, contralaterally project-ing fibres were less numerous in the adult (Smith, 1983;Smith, 1986). Whether the apparent decrease in thesefibres in the P30 Monodelphis is a reflection of a decreasetoward adulthood requires a larger sample than the twoanimals at this age in the present study.
In conclusion, the present study outlines the overallpattern of development of primary afferents and motoneu-rons within the spinal cord of the postnatal Monodelphisshowing that this species is similar to the rat becausethere is a sequence of ingrowth for the different majorclasses of primary afferents into the dorsal horn. Unlikeeutherian mammals, however, the greater part of theinnervation of the spinal cord by primary sensory afferentsoccurs postnatally, affirming the utility of Monodelphisdomestica for the study of spinal cord development andregeneration.
Fig. 8. Photomicrographs showing Neurobiotin labelled afferentsin the dorsal horn of the lumbar (L4) segment in spinal cord ofMonodelphis, at P10 (section thickness 5 100 µm). The section is fromthe segment of entry. Micrograph B is a higher power magnification ofthe region boxed in A. The photomicrographs have been taken byusing Normarski optics. Fibres appear to cross the superficial layers of
the dorsal horn (indicated with small black arrows in B) and branch inthe deeper regions. No labelled fibres branching or arborising in thesuperficial layers were present in any sections at this age. Photomicro-graph B is double the magnification of A and shows the superficialregion of the dorsal horn in A (enclosed in rectangle). Scale bar 5 100µm in A.
434 G.W. KNOTT ET AL.

Fig. 9. Darkfield photomicrographs of 150-µm-thick longitudinal-horizontal sections through thoracic spinal cord segments, T3, T4 andT5; DiI crystals were placed in the T4 DRG at two different ages (A, P0;B, P7). These sections show labelled primary afferents that have beenphotoconverted with DAB. The asterisks indicate the central spinalcanal (csc), and arrows mark the positions of dorsal root entry into thethree segments, T3, T4 and T5, from left to right. At P0 many labelledafferents are arranged longitudinally, and others are shown invadingthe grey matter at right angles, particularly in the segment of entry.
These ingrowing fibres overlap into the adjacent segments rostral andcaudal to this entry segment. After the 1st postnatal week (B) labelledafferents in the grey matter overlap considerably with the adjoiningsegments, and a few fibres have grown some distance into thecontralateral side (indicated with small white arrows in B). Thesecrossing fibres at P7 also show collaterals close to the midline (C).Micrographs A and B are the same magnification. Scale bar 5 200 µmin A and 50 µm in C.
DEVELOPING SPINAL CORD EFFERENTS AND AFFERENTS 435

LITERATURE CITED
Altman J, Bayer SA. 1984. The development of the rat spinal cord. Adv AnatEmbryol Cell Biol 85:1–165.
Brown AG. 1981. Organisation of the spinal cord. New York: Springer-Verlag.
Burkhalter A, Bernardo KL, Charles V. 1993. Developments of the localcircuits in human visual cortex. J Neurosci 13:1916–1931.
Cassidy G, Cabana T. 1993. The development of the long descendingpropriospinal projections in the opossum, Monodelphis domestica. DevBrain Res 72:291–299.
Cassidy G, Boudrias D, Pflieger J-F, Cabana T. 1994. The development ofsensorimotor reflexes in the Brazilian opossum Monodelphis domestica.Brain Behav Evol 43:244–253.
Davis BM, Frank E, Johnson FA, Scott SA. 1989. Development of centralprojections of lumbosacral sensory neurons in the chick. J Comp Neurol279:556–566.
Fadem BH, Trupin GL, Maliniak E, VandeBerg JL, Hayssen V. 1982. Careand breeding of the gray, short-tailed opossum (Monodelphis domes-tica). Lab Anim Sci 32:405–409.
Fernanadez J, Nicholls JG. 1998. Fine structure and development of dorsalroot ganglion neurons and Schwann cells in the newborn opossumMonodelphis domestica. J Comp Neurol 396:338–350.
Fitzgerald M. 1987. Prenatal growth of fine diameter primary afferents intothe rat spinal cord: a transganglionic tracer study. J Comp Neurol261:98–104.
Fitzgerald M, Butcher T, Shortland P. 1994. Developmental changes in thelaminar termination of A fibre cutaneous afferents in the rat spinaldorsal horn. J Comp Neurol 348:225–233.
Godement P, Vanselow J, Thanos S, Bonhoeffer F. 1987. A study indeveloping visual systems with a new method of staining neurones andtheir processes in fixed tissue. Development 101:697–713.
Honig MG, Hume RI. 1986. Fluorescent carbocyanine dyes allow livingneurons of identified origin to be studied in long-term cultures. J CellBiol 103:171–187.
Kita H, Armstrong W. 1991. A biotin containing compound N-(2-amino-ethyl) biotinamide for intracellular labelling and neuronal tracingstudies: comparison with biocytin. J Neurosci Methods 37:141–150.
Konstantinidou AD, Silos-Santiago I, Flaris N, Snider WD. 1995. Develop-ment of the primary afferent projection in the human spinal cord. JComp Neurol 354:11–22.
Kudo N, Yamada T. 1987. Morphological and physiological studies ofdevelopment of the monosynaptic reflex pathway in the rat lumbarspinal cord. J Physiol 389:441–459.
Leblond H, Cabana T. 1997. Myelination of the ventral and dorsal roots ofthe C8 and L4 segments of the spinal cord at different stages ofdevelopment in the gray opossum, Monodelphis domestica. J CompNeurol 386:203–216.
Lepre M, Fernandez J, Nicholls JG. 1998. Re-establishment of directsynaptic connections between sensory axons and motoneurons afterlesions of neonatal opossum CNS (Monodelphis domestica) in culture.Suppl Eur J Neurosci 10:2500–2510.
Mirnics K, Koerber R. 1995. Prenatal development of rat primary afferentfibres: central projections. J Comp Neurol 355:601–614.
Nicholls JG, Saunders NR. 1996. Regeneration of immature mammalianspinal cord after injury. Trends Neurosci 19:229–234.
O’Donovan M, Sernagor E, Sholomenko G, Ho S, Antal M, Yee W. 1992.Development of spinal motor networks in the chick embryo. J Exp Zool261:261–273.
Oppenheim RM, Schneiderman A, Shimizu I, Yaginuma H. 1988. Onset anddevelopment of intersegmental projections in the chick embryo spinalcord. J Comp Neurol 275:159–180.
Ozaki S, Snider WD. 1997. Initial trajectories of sensory axons towardlaminar targets in the developing mouse spinal cord. J Comp Neurol380:215–229.
Plenderleith MB, Cameron AA, Key B, Snow PJ. 1989. Soybean agglutininbinds to a subpopulation of small diameter primary sensory neurones inthe rat and cat. Neuroscience 31:683–695.
Qin YQ, Wang XM, Martin GF. 1993. The early development of majorprojections from caudal levels of the spinal cord to the brainstem andcerebellum in the gray short-tailed Brazilian opossum, Monodelphiadomestica. Dev Brain Res 75:75–90.
Rees S, Nitsos I, Rawson J. 1995. Prenatal development of somatosensoryprimary afferent connections in the sheep. Reprod Fertil Dev 7:427–430.
Ritz LA, Culberson JL, Brown PB. 1985. Somatotopic organisation in thecat spinal cord segments with fused dorsal horns: caudal and thoraciclevels. J Neurophysiol 54:1167–1177.
Saunders NR, Balkwill P, Knott GW, Habgood MD, Møllgård K, TreherneJM, Nicholls JG. 1992. Growth of axons through a lesion in the intactCNS of fetal rat maintained in long-term culture. Proc R Soc Lond B250:171–180.
Saunders NR. 1997. Marsupials as models for studies of development andregeneration of the central nervous system. In: Saunders N, Hinds L,editors. Marsupial biology: recent research, new perspectives. Sydney:University of New South Wales Press.
Saunders NR, Adams E, Reader M, Møllgård K. 1989. Monodelphisdomestica (gray short-tailed opossum): an accessible model for studiesof early neocortical development. Anat Embryol 180:227–236.
Saunders NR, Kitchener PD, Knott GW, Nicholls JG, Potter A, Walls TJ.1998. Development of walking, swimming and neuronal connectionsafter spinal cord transection in the neonatal opossum, Monodelphisdomestica. J Neurosci 18:339–355.
Smith CL. 1983. The development and postnatal organisation of primaryafferent projection in the thoracic spinal cord of the rat. J Comp Neurol269:96–108.
Smith CL. 1986. Sensory neurons supplying the touch domes near the bodymidlines project bilaterally in the thoracic spinal cord of rats. J CompNeurol 245:541–552.
Varga ZM, Fernandez J, Blackshaw S, Muller KJ, Adams WB, Nicholls JG.1996. Neurite outgrowth through lesions of neonatal opossum spinalcord in culture. J Comp Neurol 366:1–13.
Wang XM, Terman JR, Martin GF. 1997. Evidence that regeneration of cutaxons contributes to growth of descending axons through the lesionafter transection of the midthoracic spinal cord in the developingopossum, Didelphis virginiana. Soc Neurosci Abstr 23:1727.
Willis WD, Coggeshall RE. 1991. Sensory mechanisms of the spinal cord.2nd edition. New York: Plenum Press.
Woodward SKA, Treherne JM, Knott GW, Fernandez J, Varga ZM, NichollsJG. 1993. Development of connections by axons growing throughinjured spinal cord of neonatal opossum in culture. J Exp Biol 176:77–88.
436 G.W. KNOTT ET AL.