dendrochronological dating of historic log cabins on...
TRANSCRIPT

ORIGINAL ARTICLE
Dendrochronological dating of historic log cabins on the BelleMeade Plantation, Davidson County, Tennessee, USA
Lauren A. Stachowiak1 • Elizabeth A. Schneider2 • Maegen L. Rochner1 •
Savannah A. Collins1 • Henri D. Grissino-Mayer1
Received: 14 November 2014 / Accepted: 1 May 2015 / Published online: 10 May 2015
� Springer-Verlag Berlin Heidelberg 2015
Abstract The Harding Cabin is a one and one half story,
double pen log cabin located on the Belle Meade Plantation
in Davidson County, Tennessee, USA. Historical accounts
attribute the eastern cabin to Daniel Dunham who
originally settled in the area ca. 1780 and the western cabin
to John Harding who purchased the land from the Dunham
family in February 1807. We used dendrochronological
dating techniques to determine the years of harvest for trees
cut and used to build both cabins. We found that both
cabins were predominantly made from two genera not
commonly used to build log cabins in the southeastern
U.S., Fraxinus (ash) and Ulmus (elm). We obtained ring
width measurements from 30 cores from all walls on both
cabins and used graphical and statistical techniques to
crossdate the tree-ring series and create floating chronolo-
gies for both species. We crossdated these with reference
chronologies from the ITRDB. The Fraxinus chronology
was dated from 1657 to 1805 while the Ulmus chronology
was dated from 1583 to 1805. Cutting dates indicated that
most if not all trees used to build both cabins were cut in
late winter/early spring of 1807 before the trees had broken
dormancy. We propose that both cabins were built the
same year but by different families. We propose that the
Harding family built the eastern cabin while Harding’s
slaves built the western cabin, originally at a different lo-
cation on the property. Once the mansion was completed
by 1820, the slave cabin was relocated beside the Harding
cabin.
Keywords Dendroarchaeology � Tree rings �Construction history � Log cabin � Belle Meade Plantation �Southeastern U.S.
Introduction
Agencies that manage state and national historic sites
recognize the importance of the accuracy provided by
dendrochronological dating and have increased the demand
for tree-ring dating of historic structures. These agencies
are also recognizing that written records and oral histories
may have enhanced the historical significance of a structure
and may have added one to two generations to the age of a
structure (Grissino-Mayer 2009). Any wooden structure
can be analysed using tree-ring dating techniques provided
certain criteria are met and these criteria are carefully
evaluated prior to sampling for tree-ring analysis. For ex-
ample, the species of tree(s), number of rings, accessibility,
and preservation quality dictate whether tree-ring dating
will be successful. Furthermore, such empirical analyses of
tree-ring dates for historic structures provide valuable in-
formation for those wishing to achieve listings in the Na-
tional Register of Historic Places managed by the National
Park Service (Grissino-Mayer 2009). The use of tree rings
in archaeological research in the southeastern U.S. has
developed into a reliable and replicable science over the
past 35 years. Stahle (1979) spearheaded the use of
Communicated by L. A. Newsom.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00334-015-0535-x) contains supplementarymaterial, which is available to authorized users.
& Henri D. Grissino-Mayer
1 Laboratory of Tree-Ring Science, Department of Geography,
The University of Tennessee, Knoxville, TN 37996, USA
2 Department of Geography, Environment and Society, The
University of Minnesota, Minneapolis, MN 55455, USA
123
Veget Hist Archaeobot (2016) 25:105–115
DOI 10.1007/s00334-015-0535-x

dendrochronological methods in the dating of historic
structures throughout much of Arkansas, while Grissino-
Mayer and his students and colleagues have successfully
dated over 50 historic structures in Tennessee, Virginia,
North Carolina, Kentucky, Florida, Georgia and Michigan
(Grissino-Mayer and van de Gevel 2007; Henderson et al.
2009; Mann et al. 2009; Slayton et al. 2009; Garland et al.
2012; DeWeese et al. 2012; Grissino-Mayer et al. 2010;
Harley et al. 2011; Grissino-Mayer et al. 2013; Stachowiak
et al. 2014).
The Belle Meade Plantation, located in Davidson
County in central Tennessee, was added to the U.S. Na-
tional Register of Historic Places in December 1969. Long
before the picturesque plantation structures were built be-
ginning in the early 1800 s, the land was part of a trade
route used by Native American groups, and the forests
were used as primary hunting grounds for wild game. In
1780, Colonel John Donelson led a party of explorers and
settlers by river to central Tennessee and eventually
founded the city of Nashborough (later renamed Nash-
ville), Tennessee. Daniel Dunham (b. 1735, d. 1787) was a
member of this party. In a meander bend of Richland
Creek, south of today’s Nashville, Dunham settled the area
and moved his family to a 640 acre tract of land in 1784
(North Carolina Land Grant #173, 15 January 1784, issued
to Daniel Dunham, Grant Book 66, p. 191). That year, he
built a small fort or station (i.e., a post in a remote location)
at a main crossing to accommodate the high traffic of
travellers through the area at that time. The fort came to be
known as Dunham’s Station. While detailed written
records are scarce for the Dunham family and the region in
general during this period, several documents reveal
hardship for the Dunham family throughout their tenure on
the property. During a raid on the property in May 1787 by
Native Americans, Daniel Dunham and two of his children,
Joseph and William, were killed (Kilgore 1981; Wills
1991; Hoobler 2008). The station was later partially de-
stroyed in a fire in 1792 during another raid. Despite these
hardships, the remaining Dunham family members lived at
the station until the property was purchased by the Harding
family (Kilgore 1981; Hoobler 2008).
John Harding (b. 1777, d. 1865) moved with his family
to the area in 1798 and purchased the property from Daniel
A. Dunham (b. ca. 1784, d. ca. 1840, the son of Daniel
Dunham) on 21 February 1807. Scholars believe that the
original station structure and Dunham home currently oc-
cupies the eastern half of the current two-pen structure
(Kilgore 1981; Wills 1991) (Fig. 1). Henceforth, we call
this structure ‘‘Cabin 1.’’ Sometime later, the family of
John Harding joined a second log cabin (which we call
‘‘Cabin 2’’) on the western side of Cabin 1 (Wills 1991). By
1820, however, John Harding had built a large house on the
former ‘‘Richland Creek Plantation,’’ renaming the
mansion and the property ‘‘Belle Meade Plantation’’
(Fig. 2). He continued to purchase adjoining land such that
the plantation totalled 3,800 acres (1,530 ha) by the mid-
1820 s. Harding had income from the standing of stallions
and boarding of horses, and already owned several horses
he trained and raced. When his son William (b. 1808, d.
1886) took over management of the plantation in 1839, he
continued the horse breeding and racing tradition his father
began, making Belle Meade one of the most renowned
breeders of thoroughbred horses in the U.S., a legacy that
lasted through the rest of the 19th century. After his death
in 1886, national financial crises, a withering economy, and
evangelical reform that disfavoured gambling slowly took
their toll on horse racing and breeding, and final dispersal
sales of the plantation’s horses took place in 1903 and
1904.
Fig. 1 View of the south walls of Cabin 1 (right) and Cabin 2 (left).
The dogtrot or breezeway is seen in the centre. Photo orientation is to
the north (photograph by H. D. Grissino-Mayer)
Fig. 2 The Tennessee Valley region with locations of the Belle
Meade Plantation and the four reference chronologies used in this
project (map created by L. A. Stachowiak)
106 Veget Hist Archaeobot (2016) 25:105–115
123

For over 200 years since the land was purchased by
John Harding, the two conjoined cabins remained on the
plantation, used as housing for slaves and later by freedmen
and other tenants during the 19th and 20th centuries. The
cabins are located approximately 200 metres from what is
now the main plantation house, and 100 metres from the
meander in Richland Creek. The cabins are separated by a
central breezeway (also called a dogtrot) in a style referred
to as a double pen cabin (Rehder 2012). The cabins feature
a second floor, or half story, accessible via steps in the
breezeway between the two cabins (seen in Fig. 1). Each
cabin has a limestone chimney located on either side of the
structure. A porch exists on both the north and south sides
of the cabins, and entrance doors to each pen are currently
located only through the breezeway. However, 19th cen-
tury photographs and evidence of cutting and replacement
of logs in each pen indicate that original doorways once
existed only on the south-facing walls.
Despite claims that Dunham was the builder of Cabin 1,
‘‘the true age of the cabin, alleged to be the original or
described as built on the site of Dunham’s Station, remains
conjecture’’ (Warden 1988). Because doubt exists which of
the two could have been built by Daniel Dunham, doubt
also exists about which cabin was built by John Harding.
The overarching goal of our project was to date precisely
the years of construction for both cabins and thereby help
evaluate the likely builder of each structure.
Materials and methods
Field methods
We extracted 12 mm diameter cores from both log struc-
tures using a specially designed, electrically powered hol-
low drill bit. Cores were specifically located and collected
from logs based on several factors. First, we collected cores
from the basal end of the log to ensure as many rings as
possible were included. Second, we collected cores only
from the underside of logs to ensure the holes left were
inconspicuous and to allow the sawdust to quickly evacuate
the drill bit. Third, we cored only along the rounded edges
of the log (and through bark, when possible) to capture the
outermost rings and thus determine the year(s) the trees
were harvested. The angle of the drill bit was carefully
monitored while coring by a second person who stood at
the log end and ensured alignment of the drill bit tip with
the pith. This allowed us to reach as close to the pith as
possible and therefore obtain the maximum number of
rings possible. Once extracted, the cores were immediately
mounted on core mounts with the cells vertically aligned.
All cores were then labelled according to cabin, compass
direction of wall, the log number, and core letter (if
needed), e.g., C1N005A = Cabin 1, north wall, log 5 (se-
quential from the bottom log), core ‘‘A.’’ Lastly, locations
from where all cores on all logs were extracted were
sketched for permanent archiving.
Sanding and measuring
Each core was sanded with a tabletop sander using progres-
sively finer sandpaper beginning with ANSI 80-grit
(177–210 lm) and endingwithANSI 400-grit (20.6–23.6 lm)
(Orvis and Grissino-Mayer 2002) to polish the wood to a fine
finish. Once sanded, tree rings on all cores were marked using
standard decadal dot notation (Stokes and Smiley 1996; Speer
2010) and then measured to 0.001 mm accuracy using a Vel-
mex measuring system coupled with MeasureJ2X software.
Measuring began with the innermost complete ring assigned to
relative year ‘‘1.’’ By convention, the outermost ring is not
measured because it may not be a complete ring if the tree was
harvested during the middle of the growing season.
Internal crossmatching
We used COFECHA to perform segmented time-series
correlation analysis to suggest a possible temporal placement
for all series in the data set (Holmes 1983; Grissino-Mayer
2001). We evaluated 40-year ring segments lagged by
10 years to provide more in-depth diagnostics that helped
ensure the proper temporal alignment had been achieved.
The program statistically compared each individual series
with a master chronology created from all other cores and
calculated the inter-series correlation coefficient, a statistical
metric of how well the ring-width patterns from the core
being tested matched the chronology created from all other
cores. A minimum value of 0.40 (usually p\ 0.001) was
desirable for results to be statistically convincing although
individual correlation values were often much higher (e.g.,
r = 0.55 with p\ 0.0001). A core was considered cross-
matchedwith all other cores when its coefficient was C 0.40
and COFECHA suggested a temporal adjustment that was
identical for all or most of the segments tested (e.g., ‘‘?41’’).
Suggested placements made by COFECHA were confirmed
visually by comparing line graphs of each series being tested
against line graphs of other series. The result of the internal
crossdating exercise was a set of tree-ring measurement
series that were properly aligned in time relative to each
other, but not absolutely dated.
External crossdating
Absolute (external) crossdating is possible because the
overarching regional climate signal imparts a common pat-
tern of wide and narrow rings in tree growth. Our reference
datasets consisted of three Quercus alba L. (white oak)
Veget Hist Archaeobot (2016) 25:105–115 107
123

chronologies and one Quercus stellataWangenh. (post oak)
chronology downloaded from the International Tree-Ring
Data Bank (ITRDB) (Grissino-Mayer and Fritts 1997), all
located in the surrounding region (Fig. 2). The reference
chronologies were created by Edward R. Cook of the La-
mont-Doherty Earth Observatory at Columbia University in
New York (KY004), Daniel N. Duvick (formerly) of Oak
Ridge National Laboratory in Tennessee (TN005 and
TN008), and Michael C. Stambaugh and Richard P. Guyette
of the University of Missouri (KY005):
1. Mammoth Cave Recollect, KY004, 37.1215�N,86.0504�W, 1649–1985, Q. alba;
2. Piney Creek Pocket Wilderness, TN005, 35.6484�N,84.8584�W 1651–1982, Q. alba;
3. Norris Dam State Park, TN008, 36.1734�N,84.1003�W, 1633–1980, Q. alba;
4. Land between the Lakes, KY005, 36.7473�N,88.0664�W, 1689–2005, Q. stellata.
The reference chronologies were selected because of
their close geographic proximity to the Belle Meade
Plantation, and because they exhibited the appropriate
depth back in time to ensure a successful temporal overlap
with the pattern of ring widths from the undated cores.
These four chronologies were combined to create a single
regional reference chronology using the computer program
ARSTAN (Cook 1985).
We used COFECHA to statistically crossdate the undated
series by entering the reference Quercus chronology as the
dated dataset and the file containing the temporally adjusted
measurement series as the undated dataset. We again tested
40-year segments in the undated series lagged by 10 years to
maximize the number of segments tested and to develop
useful diagnostics for evaluating problematic segments or
cores. Crossdating was achieved when COFECHA suggested
a common temporal adjustment for all or most tested seg-
ments (e.g., ‘‘?1582’’) and these segments also displayed
statistically significant (usually p\0.001) correlations
against the reference chronology. Once crossdated, we used
program EDRM (Edit Ring Measurement) (Holmes 1992) to
manually adjust the rings to their exact calendar years and
created a chronology for each cabin using program ARSTAN.
We graphically verified the crossdating by overlaying the two
dated cabin chronologies along with the reference chronol-
ogy. Crossdating had to be convincing both graphically and
statistically (Grissino-Mayer 2001).
Determining cutting dates
Cutting dates for logs from a cabin indicate the years in
which the trees were harvested and are determined by
careful examination of the terminal rings on the cores un-
der high magnification. Only a few logs will have bark
attached to the outermost ring, making the year of the last
formed tree ring easily known. Most logs, however, do not
have intact bark and others may have the outermost rings
lost to decay or trimming during construction. Therefore, a
system has been established to determine the cutting dates
of each log and the likely year(s) of construction for the
cabin (Bannister et al. 1966; Nash 1999; Grissino-Mayer
et al. 2009).
B: Bark was present and attached to the last formed ring,
indicating the outermost ring is intact (a firm cutting date);
r: The outermost ring is intact around a smooth curved
surface of the log (considered a cutting date);
v: The date is within a few years of the cutting date,
based on the presence of sapwood (a near cutting date);
vv: A cutting date is not possible because we could not
determine how far we were from the outermost ring ever
formed on this tree (a non-cutting date);
??: A ring count was necessary on the outermost rings
because these were located in a detached sapwood portion of
the core, or the rings couldnot be crossdatedpast a certainpoint.
Once cutting dates for each sample were determined, we
noted any clustering of dates around certain calendar years
that could indicate the likely year(s) of tree harvesting for
cabin construction.
Results
Species used
Once we were able to view the sanded cores under the
microscope, we learned that the cabins were constructed
from multiple tree species (Table 1). Logs from Cabin 1
were cut from one of several Fraxinus (ash) species and
from one of several Ulmus (elm) species that grow in the
area (Fig. 3). Cabin 2 was made from four different tree
genera: Fraxinus, Ulmus, Quercus (oak), and Carya
(hickory) (Table 1). Each cabin was analysed by grouping
series from each genus together and testing the crossdating
of the floating series from the individual genus internally
against each other and externally against the reference
chronologies. Only Fraxinus and Ulmus were analysed
because we did not have enough cores that had enough
rings to test the Quercus and Carya series. We also com-
pared the final Fraxinus and Ulmus chronologies together
to evaluate crossdating between two hardwood tree genera,
and to test the crossdating of series between the two cabins.
Internal crossdating for Cabin 1
We were able to internally crossdate the tree rings from
seven cores representing all seven logs cut from Fraxinus
trees (ESMTable 1). The average interseries correlationwas
108 Veget Hist Archaeobot (2016) 25:105–115
123

0.53, which would be considered high for southeastern tree
species, while the average mean sensitivity was 0.23, a value
typical for trees with a moderate climate signal. COFECHA
flagged 15 of the 59 40-year segments tested as being
problematic (25 %), which is considerably higher than the
cut-off of 10 % we desire when crossdating tree rings in the
southeastern U.S. Of these 15 problem segments, eight oc-
curred on one core, C1N006A, which was the longest of the
seven Fraxinus cores in Cabin 1 (149 years) and also the
most challenging to crossdate. These eight segments had
correlations that were nonetheless all positive, indicating
some inherent climate signal, andwere bracketed by younger
and older segments that were not flagged (ESM Table 1).
Furthermore, this series, despite the problem segments, was
crossdated as having the same outermost ring (ring 149) as
five of the other six series, further confirming this series was
crossdated correctly.
The Ulmus cores, overall, contained tree rings that were
problematic to crossdate. Of the 11 logs in Cabin 1 that we
cored that were cut from Ulmus trees, we could crossdate
the tree-ring series from seven logs (ESM Table 2). We
found that Ulmus forms well-defined rings only when
young or middle-aged (Fig. 3a, d), but when some indi-
viduals reach maturity, the ring boundaries are not well-
defined, blending in with the distinct ‘‘ulmiform’’ bands in
the latewood to the point they cannot be distinguished. For
the seven that could be crossdated, Ulmus displayed a very
high average interseries correlation of 0.60 and an equally
impressive average mean sensitivity of 0.34 (ESM
Table 2). Equally important were the overall lengths of
individual series which averaged 145 years, with the
longest/oldest (C1W005A) being 210 years. Three series
had outermost rings with the same relative ring number
(210), suggesting these logs were cut in the same year; two
other logs had outermost ring numbers of 205. A final
notable feature of the crossdated Ulmus cores was the very
few (four) of the 84 40-year segments (5 %) flagged by
COFECHA. In general, when Ulmus can be crossdated, the
tree-ring series are exceptional.
Internal crossdating for Cabin 2
We were able to crossdate the tree rings from eight cores
representing all seven logs that were cut from Fraxinus
trees used to construct Cabin 2. The average interseries
correlation was 0.50 and the average mean sensitivity was
0.25 (ESM Table 3), both values comparable to Fraxinus
in Cabin 1. Of 69 40-year segments tested by COFECHA,
only four were flagged as being problematic (6 %). The
outermost ring number (102) was similar for six of the
seven logs, while the remaining log had a relative ring
number of 100. This finding indicates that these Fraxinus
trees were cut during the same year.
As with Ulmus in Cabin 1, we encountered considerable
difficulty in crossdating the Ulmus tree-ring series for
Cabin 2. We were only able to partially crossdate the tree
rings from eight cores representing five logs (ESM
Table 4). The tree-ring series could be crossdated only
during the earliest to middle sections on each core, such
that up to half the tree rings on any one core could not be
crossdated with confidence. We crossdated the Ulmus tree
rings as far as possible from innermost to outermost ring,
and then counted the rings past that point. At that point, the
thin ring boundaries blended into the ‘‘ulmiform’’ bands in
the latewood and could no longer be distinguished. We
report statistics only for the crossdated portions of the
Fig. 3 a Core taken from an Ulmus log (top) and a Fraxinus log
(bottom). b ‘‘Ulmiform’’ bands in the latewood that were diagnostic
of Ulmus cores. c Attached bark on a Fraxinus core. d Pith of a
Fraxinus core (photographs by L. A. Stachowiak)
Table 1 Tree genera used to build both cabins
Veget Hist Archaeobot (2016) 25:105–115 109
123

cores. The average interseries correlation was 0.48 and the
average mean sensitivity was 0.31, both values somewhat
lower than the values reported for Ulmus from Cabin 1.
Nine segments of the 69 40-year segments (13 %) that
could be crossdated were flagged by COFECHA as being
problematic but all segments were carefully re-inspected
and found to be correctly dated. The crossdated portions of
the cores resulted in a 169 year chronology, but ring counts
brought these cores out enough to create a 222-year long
dataset. Several cores had outermost rings that were within
a few years (e.g., ring numbers 218, 219, 220 and 222)
indicating that the ring counts for Ulmus could provide
some supporting information on possible cutting dates for
Cabin 2.
External (absolute) crossdating
We tested the correlations between the Fraxinus series
from both cabins and found excellent correspondence,
suggesting we could combine the two datasets into one
larger dataset. We created a floating chronology from this
dataset that spanned 149 years and tested this against the
composite reference chronology created from the four re-
gional Quercus chronologies using COFECHA. We found
the absolute crossdating when COFECHA suggested a
‘‘?1656’’ dating adjustment for all 12 40-year segments
tested (lagged 10 years) of the 149-year long dataset with
an average correlation of 0.43. We then added 1656 to all
rings in the Fraxinus measurement series and created an-
other chronology which we again tested against the master
reference chronology. We found a statistically significant
correlation (r = 0.39, n = 149, t = 5.14, p\ 0.0001) that
suggested that the Fraxinus series were now absolutely
dated, spanning from 1657 to 1805 (Fig. 4). The last ring
present on most cores was therefore the year 1806 which
could have been a partial or complete ring.
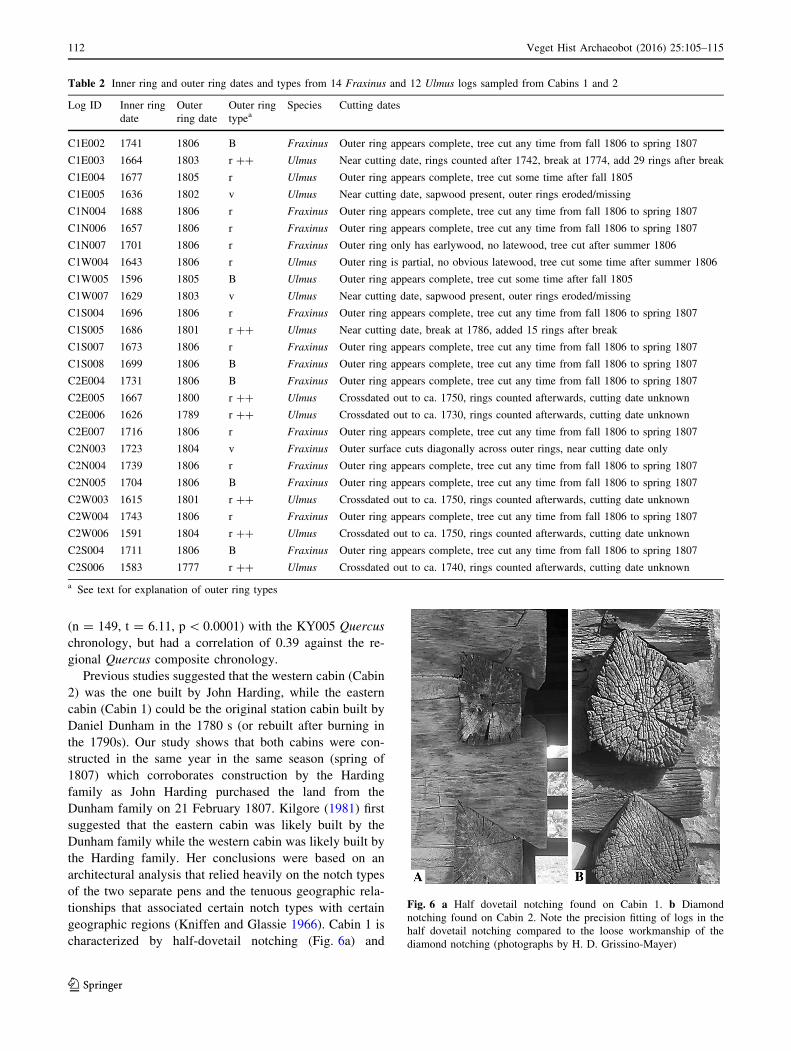
We tested the correlations between theUlmus series from
both cabins and again found excellent temporal correspon-
dence, suggesting we could combine the two datasets into
one larger dataset. We created a floating chronology from
this dataset that spanned 223 years and tested this against the
composite reference chronology created from the four re-
gional Quercus chronologies. COFECHA found a strong
dating adjustment of ‘‘?1582’’ for 12 of the 19 40-year
segments tested (lagged 10 years) in the 223-year long
dataset with an average correlation of 0.44. These correla-
tions and associated dating adjustments were present in the
early andmiddle portions of the floating chronology because
the outer portion of this dataset contained tree rings from
Cabin 2 that could not be crossdated. We added 1582 to the
calendar dates for all tree rings and created another chron-
ology to test against the master reference chronology. We
again found a statistically significant correlation (r = 0.36,
n = 173, t = 5.05, p\ 0.0001) that suggested the Ulmus
tree-ring data were now absolutely crossdated, spanning
1583 to 1805 (Fig. 5) (common period of overlap is 1633 to
1805 = 173 years). For the Ulmus cores collected from
Cabin 1, the date for the outermost unmeasured rings was
therefore 1806. No cutting dates could be obtained forUlmus
from Cabin 2 because they could not be crossdated com-
pletely. However, the outermost dates based on ring counts
(e.g., 1800 for C2E005, 1801 for C2W003, and 1804 FOR
C2W006) provide a terminus pro quem and support the
structure being built a few years after 1804.
Having established that both Fraxinus and Ulmus cross-
date with Quercus tree-ring data, we next tested the corre-
lation between the Fraxinus andUlmus tree-ring series from
the two cabins. We found a statistically significant correla-
tion (r = 0.35, n = 149, t = 4.53, p\ 0.0001) that suggests
the two species do in fact crossdate against each other, but the
relationship is weaker than both relationships with the
Quercus tree-ring data.
Cutting dates
Most of the Fraxinus cores from both cabins clearly show
that the outermost measured tree ring dates to the year
1805. Careful inspection, however, revealed that most
Fraxinus cores have what could be a complete ring for
1806 (by convention, the outermost ring is never measured
because it could represent an incomplete ring), indicating
that the latest harvest of trees used to build both cabins
occurred sometime in the dormant season of 1806–1807
(Table 2). For most Fraxinus cores, the likely season for
harvest is late winter or early spring of 1807 because John
Harding purchased the property in late February of 1807
and likely began cutting trees immediately or soon after for
use in Cabin 1. Four Fraxinus cores have clear bark re-
tained, and these four cores provide convincing evidence
for tree harvest during the dormant season of 1806–1807.
Some Fraxinus cores suggest tree harvesting before the
dormant season of 1806–1807, but it is more likely that the
outermost ring or a partial outermost ring was trimmed
during construction or decayed off, despite these logs
having what appeared to be smooth surfaces.
The Ulmus cores provide supporting evidence for har-
vest during the dormant season of 1806-1807 although
most Ulmus cores have outermost ring dates that slightly
precede 1807 (Table 2). One Ulmus core (C1W005) had
bark but the outermost two rings were indistinct and we
could not tell conclusively if any wood had formed for
1806 yet. Based on these results, we can confirm that both
structures were built from Fraxinus and Ulmus trees that
were likely harvested in late winter or early spring of 1807,
before trees had come out of their winter dormant period.
110 Veget Hist Archaeobot (2016) 25:105–115
123
Discussion
The use of multiple species is uncommon for building log
structures in the southeastern U.S. as early settlers built
structures usually from a single tree species. Morgan
(1990) noted that 20 % of the log structures he studied in
east Tennessee were built from two species, usually pine
and tulip poplar. However, of nearly 50 historic structures
we have analysed over the last 15 years, the Harding
Cabins are the first we have encountered with two or more
species. The choice to use these species was likely based
on local availability and suggests Fraxinus and Ulmus were
locally abundant. Neither Morgan (1990) nor Rehder
(2012) mention the use of these two species to build log
structures, nor could we find any reference to using these
species as building materials in log structures in the
southeastern U.S.
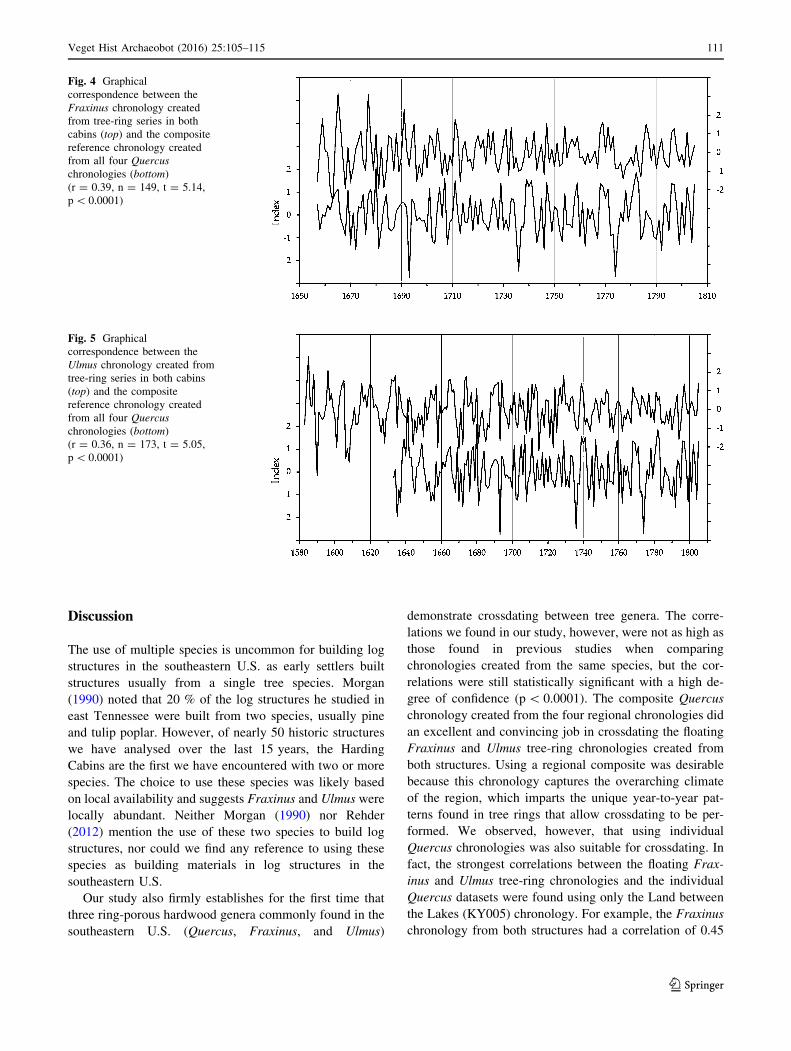
Our study also firmly establishes for the first time that
three ring-porous hardwood genera commonly found in the
southeastern U.S. (Quercus, Fraxinus, and Ulmus)
demonstrate crossdating between tree genera. The corre-
lations we found in our study, however, were not as high as
those found in previous studies when comparing
chronologies created from the same species, but the cor-
relations were still statistically significant with a high de-
gree of confidence (p\ 0.0001). The composite Quercus
chronology created from the four regional chronologies did
an excellent and convincing job in crossdating the floating
Fraxinus and Ulmus tree-ring chronologies created from
both structures. Using a regional composite was desirable
because this chronology captures the overarching climate
of the region, which imparts the unique year-to-year pat-
terns found in tree rings that allow crossdating to be per-
formed. We observed, however, that using individual
Quercus chronologies was also suitable for crossdating. In
fact, the strongest correlations between the floating Frax-
inus and Ulmus tree-ring chronologies and the individual
Quercus datasets were found using only the Land between
the Lakes (KY005) chronology. For example, the Fraxinus
chronology from both structures had a correlation of 0.45
Fig. 4 Graphical
correspondence between the
Fraxinus chronology created
from tree-ring series in both
cabins (top) and the composite
reference chronology created
from all four Quercus
chronologies (bottom)
(r = 0.39, n = 149, t = 5.14,
p\ 0.0001)
Fig. 5 Graphical
correspondence between the
Ulmus chronology created from
tree-ring series in both cabins
(top) and the composite
reference chronology created
from all four Quercus
chronologies (bottom)
(r = 0.36, n = 173, t = 5.05,
p\ 0.0001)
Veget Hist Archaeobot (2016) 25:105–115 111
123
(n = 149, t = 6.11, p\ 0.0001) with the KY005 Quercus
chronology, but had a correlation of 0.39 against the re-
gional Quercus composite chronology.
Previous studies suggested that the western cabin (Cabin
2) was the one built by John Harding, while the eastern
cabin (Cabin 1) could be the original station cabin built by
Daniel Dunham in the 1780 s (or rebuilt after burning in
the 1790s). Our study shows that both cabins were con-
structed in the same year in the same season (spring of
1807) which corroborates construction by the Harding
family as John Harding purchased the land from the
Dunham family on 21 February 1807. Kilgore (1981) first
suggested that the eastern cabin was likely built by the
Dunham family while the western cabin was likely built by
the Harding family. Her conclusions were based on an
architectural analysis that relied heavily on the notch types
of the two separate pens and the tenuous geographic rela-
tionships that associated certain notch types with certain
geographic regions (Kniffen and Glassie 1966). Cabin 1 is
characterized by half-dovetail notching (Fig. 6a) and
Table 2 Inner ring and outer ring dates and types from 14 Fraxinus and 12 Ulmus logs sampled from Cabins 1 and 2
Log ID Inner ring
date
Outer
ring date
Outer ring
typeaSpecies Cutting dates
C1E002 1741 1806 B Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C1E003 1664 1803 r ?? Ulmus Near cutting date, rings counted after 1742, break at 1774, add 29 rings after break
C1E004 1677 1805 r Ulmus Outer ring appears complete, tree cut some time after fall 1805
C1E005 1636 1802 v Ulmus Near cutting date, sapwood present, outer rings eroded/missing
C1N004 1688 1806 r Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C1N006 1657 1806 r Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C1N007 1701 1806 r Fraxinus Outer ring only has earlywood, no latewood, tree cut after summer 1806
C1W004 1643 1806 r Ulmus Outer ring is partial, no obvious latewood, tree cut some time after summer 1806
C1W005 1596 1805 B Ulmus Outer ring appears complete, tree cut some time after fall 1805
C1W007 1629 1803 v Ulmus Near cutting date, sapwood present, outer rings eroded/missing
C1S004 1696 1806 r Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C1S005 1686 1801 r ?? Ulmus Near cutting date, break at 1786, added 15 rings after break
C1S007 1673 1806 r Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C1S008 1699 1806 B Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C2E004 1731 1806 B Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C2E005 1667 1800 r ?? Ulmus Crossdated out to ca. 1750, rings counted afterwards, cutting date unknown
C2E006 1626 1789 r ?? Ulmus Crossdated out to ca. 1730, rings counted afterwards, cutting date unknown
C2E007 1716 1806 r Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C2N003 1723 1804 v Fraxinus Outer surface cuts diagonally across outer rings, near cutting date only
C2N004 1739 1806 r Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C2N005 1704 1806 B Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C2W003 1615 1801 r ?? Ulmus Crossdated out to ca. 1750, rings counted afterwards, cutting date unknown
C2W004 1743 1806 r Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C2W006 1591 1804 r ?? Ulmus Crossdated out to ca. 1750, rings counted afterwards, cutting date unknown
C2S004 1711 1806 B Fraxinus Outer ring appears complete, tree cut any time from fall 1806 to spring 1807
C2S006 1583 1777 r ?? Ulmus Crossdated out to ca. 1740, rings counted afterwards, cutting date unknown
a See text for explanation of outer ring types
Fig. 6 a Half dovetail notching found on Cabin 1. b Diamond
notching found on Cabin 2. Note the precision fitting of logs in the
half dovetail notching compared to the loose workmanship of the
diamond notching (photographs by H. D. Grissino-Mayer)
112 Veget Hist Archaeobot (2016) 25:105–115
123

Kilgore (1981) noted that the half-dovetail notching was
common in North Carolina and eastern Tennessee, the re-
gion from where Daniel Dunham and his family emigrated.
In contrast, Cabin 2 is characterized by the very uncommon
diamond notching where both upper and lower sides of the
end section of the log are chamfered to produce the crown
(Gavin 1997) (Fig. 6b). This notch type can be traced to
settlers who emigrated from near the Virginia/North
Carolina border (Gavin 1997). Although John Harding
originated from Goochland County in central Virginia,
Wills (1991) noted that Harding likely learned the diamond
notching technique ‘‘in Virginia, where that method was
prevalent’’.
A major historical inconsistency exists, however: if both
cabins were constructed by the Harding family contem-
poraneously, why then is one cabin (Cabin 1) well-pre-
served while the other (Cabin 2) is in a poor state of
preservation? And why use two different notch types? The
two cabins stand in stark visual contrast to each other.
Cabin 1 is characterized by logs that are in an excellent
state of preservation. Furthermore, Cabin 1 demonstrates
unusual precision with the log notches fitting perfectly into
each other, top and bottom (Fig. 6a), suggesting a master
builder was responsible for building the cabin. The wall
construction follows a basal- to distal-end overlaying
technique, and each log was clearly chosen specifically for
each successive layer. The logs on all four walls were
meticulously hewn, producing straight sides of unusual
precision. Cabin 2 is characterized by logs that are in a
poor state of preservation, with logs fitting loosely to-
gether. The uncommon diamond notching does not require
precision fitting and could be created by someone unskilled
in cabin building or notching. Many wide gaps exist in the
notching between the logs, and the sequential size order of
each ascending log is not well organized. Any person
carefully inspecting the logs in both structures would
clearly conclude that both structures were built during
different periods.
To help resolve this puzzle, we first evaluated whether
the Fraxinus and Ulmus trees used to build both structures
could have come from the same Fraxinus and Ulmus
populations. We observed that the Fraxinus trees used to
construct Cabin 1 were considerably older (maximum
150 years, average 112 years) compared to the Fraxinus
trees used to construct Cabin 2 (maximum 102 years, av-
erage 79 years). Furthermore, the Fraxinus trees used to
build Cabin 1 were clearly larger in diameter (aver-
age = 31 cm) than the Fraxinus trees used to build Cabin
2 (average = 21 cm), based on extrapolations of the pith to
outer edge distance of all the Fraxinus cores we sampled.
The Fraxinus trees also crossdated well against each other.
The Ulmus trees cut to build both structures were generally
quite old with little difference in maximum ages (Cabin
1 = maximum 210 years, while Cabin 2 = maximum
222 years), but the Ulmus trees used in Cabin 1 were
younger (average = 145 years) compared to the Ulmus
trees in Cabin 2 (average = 168 years). However, Ulmus
specimens used to build Cabin 1 were much larger, with an
average diameter of 28.4 cm compared to the smaller di-
ameter Ulmus used to build Cabin 2 (average = 20.4 cm).
Like the Quercus, the Ulmus trees from both structures
crossdated well with each other. This also might help ex-
plain why the Ulmus tree rings in Cabin 2 were more
difficult to crossdate: the Ulmus trees cut to build Cabin 2
were both older but narrower in diameter, meaning more
rings per unit measurement. This could, however, be an
artifact of the location we chose to sample on the indi-
vidual Ulmus logs.
These findings suggest that the Fraxinus and Ulmus
trees used in Cabin 1 came from the same Fraxinus and
Ulmus tree populations used to build Cabin 2, with only
one major difference. The trees used in Cabin 1 were larger
than the trees used to build Cabin 2, a finding easily con-
firmed by visual inspection of the two structures them-
selves. We suggest that the larger trees in the original, local
Fraxinus and Ulmus stands were selectively cut first in
early 1807 and used to build Cabin 1, while the remaining
smaller diameter Fraxinus and Ulmus trees were then
harvested, also in early 1807, to build Cabin 2. If larger
diameter trees had been available, the builder of Cabin 2
would surely have used these rather than the smaller di-
ameter trees because using larger diameter trees would
mean fewer trees needed to be cut, hewn, and notched. An
alternative explanation suggests that the builder of Cabin 2
was specifically instructed to, or preferred to harvest, only
the smaller diameter trees.
Based on this mounting visible, physical, and den-
drochronological evidence, we propose a radical interpre-
tation of the construction of both cabins: the cabins were
not built by the same person or by same family. If John
Harding and his family had purposely built two cabins at
the same time in spring 1807, both cabins would likely
show similar structural features, such as similar log sizes,
only one notch type used, clear arrangement of the logs
(distal and basal log ends alternating on each side of the
cabin), and precision fitting of the logs one on top of the
other. Instead, we propose two very different groups of
people built the two cabins. We propose that Cabin 1 was
built by the John Harding family using large diameter trees
that had been harvested first from the local stands of trees,
before the trees broke dormancy in the late winter/early
spring of 1807. The cabin was meticulously built with great
attention to detail. Cabin 2 was built by another group of
people, and we suggest that the cabin was likely built by
the slaves owned by John Harding, Ben and Dicey, ‘‘who
helped [John Harding] turn his woods and fields into a
Veget Hist Archaeobot (2016) 25:105–115 113
123

working farm’’ (Wills 1991). The trees were cut after the
Harding family had harvested the larger ones to use in their
cabin, and the cabin was built by individuals who likely
were less skilled in the craft of cabin construction. These
findings would repudiate the suggestion that the second
cabin was built by John Harding sometime around 1812 as
his family grew to include three children: Amanda (b.
October 1807), William (b. September 1808), and El-
izabeth (b. April 1812) (Wills 1991).
Furthermore, we propose the two cabins were built at
different locations based primarily on the major differences
in physical appearances of the two cabins. If both cabins
had been built at the same time at the same location by the
same builders, the two cabins would likely have looked
very similar. We propose that Cabin 2 likely was built
elsewhere on the property in 1807 and perhaps served as
living quarters for the Harding family slaves. In 1819, John
Harding began making significant purchases of building
materials and internal furnishings and the main mansion of
the plantation was completed by late 1820 (Wills 1991).
We suggest that, once the Harding family relocated to the
larger mansion, Harding then gave the original cabin to a
slave family that lived and worked on the plantation. To
centralize the living arrangement, and to accommodate the
growth of extended families, the original cabin built by the
slaves was relocated to the original location of the Harding
Cabin. This suggests that the breezeway and added front
and back porches were constructed by the slave families.
The two cabins continued to serve as living quarters for the
families even after emancipation. Photographs of the two
cabins dated ca. 1880 and ca. 1890 clearly show many
individuals of an African-American extended family or
families (Fig. 7).
Conclusions
The two cabins that make up the Harding Cabin provided
a unique opportunity for the advancement of dendroar-
chaeology in the Southeast. The two cabins that collec-
tively make up the Harding Cabin on the Belle Meade
Plantation were built primarily from two tree genera,
Fraxinus and Ulmus. These are the first structures known
to be made from these two genera in the southeastern
U.S., and each cabin represents one of the few historic
structures known to be built from two or more tree spe-
cies. The individual Fraxinus and Ulmus tree-ring series
from each structure crossdated well within each species.
The Fraxinus and Ulmus tree-ring series also crossdated
well against the regional composite Quercus chronology
which establishes that, for the first time, crossdating exists
between hardwood genera in the southeastern U.S. These
two cabin chronologies represent the first developed from
these two species in the southeastern U.S. Once abso-
lutely crossdated, the Fraxinus chronology from both
cabins was anchored in time from 1657 to 1805 while the
Ulmus chronology was anchored in time from 1583 to
1805. The Ulmus chronology, however, is accurate only
in the early to middle portions of the chronology, because
the rings from many Ulmus cores could not be crossdated
past a certain year. Nonetheless, the Ulmus chronology is
now the oldest in Tennessee and one of the oldest overall
in the southeastern U.S. Because the Ulmus chronology
crossdated with Quercus and Fraxinus, future studies can
use Ulmus tree-ring data to date historic structures should
no nearby Quercus chronologies exist (e.g., in western
Tennessee). The high level of crossdating between tree
rings from both cabins suggests that the Fraxinus and
Ulmus trees used to build both cabins came from the
same local forest stand.
Careful inspection of the outermost rings suggests that
the Fraxinus and Ulmus trees were cut in the dormant
season of 1806/1807, most likely in late winter/early spring
of 1807 because John Harding purchased the land on 21
February 1807. This refutes the assertion that Cabin 1
possibly represented the original cabin built by Daniel
Dunham or built later after his death. Differences in visual
appearance, level of workmanship, preservation quality,
notch types, tree size, and tree ages suggest that the two
cabins likely were built by two different families. We
propose that John Harding and his family built Cabin 1
while Cabin 2 was likely built by the slaves that lived on
the property. We further propose that Cabin 2 was perhaps
built elsewhere on the property, then relocated and con-
solidated with Cabin 1 once the main mansion was com-
pleted in 1820, after which the two cabins served as living
quarters for slaves and freedmen after emancipation.
Fig. 7 Photograph dated ca. 1890 showing the joined cabins which
housed an African-American family (photograph courtesy of the Belle
Meade Plantation)
114 Veget Hist Archaeobot (2016) 25:105–115
123

Acknowledgments We thank personnel at Belle Meade Plantation
who first expressed interest in our carrying out this interesting project,
especially Board Member Paul Clements, Executive Director Alton
Kelley, and Curator John Lamb. Historic pictures, photographs, and
documents were provided by the Belle Meade Plantation for better
understanding the historical context of both cabins.
References
Bannister B, Gell EA, Hannah JW (1966) Tree-ring dates from
Arizona N-Q: Verde-Show Low-St. Johns Area. Laboratory of
Tree-Ring Research, University of Arizona, Tucson
Cook ER (1985) A Time Series Analysis Approach to Tree-Ring
Standardization. Dissertation, University of Arizona, Tucson
DeWeese GG, Bishop WJ, Grissino-Mayer HD, Parrish BK, Edwards
SM (2012) Dendroarchaeological dating of the Chief John Ross
House, Rossville, Georgia. Southeast Arch 31:221–230
Garland NA, Grissino-Mayer HD, Deagan K, Harley GL, Waters G
(2012) Dendrochronological dating of wood from the Fountain
of Youth Park Archaeological Site (8SJ31), St. Augustine,
Florida, U.S.A. Tree-Ring Res 68:69–78. doi:10.3959/2010-11.1
Gavin M (1997) The diamond notch in Middle Tennessee. Mater Cult
29:13–23
Grissino-Mayer HD (2001) Evaluating crossdating accuracy: a
manual and tutorial for the computer program COFECHA.
Tree-Ring Res 57:205–221
Grissino-Mayer HD (2009) An introduction to dendroarcheology in
the southeastern United States. Tree-Ring Res 65:4–10. doi:10.
3959/2008-19.1
Grissino-Mayer HD, Fritts HC (1997) The International Tree-Ring
Data Bank: an enhanced global database serving the global
scientific community. Holocene 7:235–238. doi:10.1177/
095968369700700212
Grissino-Mayer HD, van de Gevel SL (2007) Tell-tale trees: historical
dendroarcheology of log structures at Rocky Mount, Piney Flats,
Tennessee. Hist Archaeol 41:32–49
Grissino-Mayer HD, LaForest LB, van de Gevel SL (2009)
Construction history of the Rocky Mount Historic Site
(40SL386), Piney Flats, Tennessee from tree-ring and documen-
tary evidence. Southeast Arch 28:64–77
Grissino-Mayer HD, Kobziar LN, Harley GL, Russell KP, LaForest
LB, Oppermann JK (2010) The historical dendroarchaeology of
the Ximenez-Fatio House, St. Augustine, Florida, U.S.A. Tree-
Ring Res 66:61–73. doi:10.3959/2009-8.1
Grissino-Mayer HD, Maxwell JT, Harley GL, Garland NA, Holt DH,
Absher C, Beale BJ, Boehm MS, De Graauw KA, Rautio AM,
Dye AW (2013) Dendrochronology reveals the construction
history of an early 19th century farm settlement, southwestern
Virginia, USA. J Archaeol Sci 40:481–489. doi:10.1016/j.jas.
2012.05.038
Harley GL, Grissino-Mayer HD, LaForest LB, McCauley P (2011)
Dendrochronological dating of the Lund-Spathelf House, Ann
Arbor, Michigan, USA. Tree-Ring Res 67:117–121. doi:10.
3959/2010-2.1
Henderson JP, Grissino-Mayer HD, van de Gevel SL, Hart JL (2009)
The historical dendroarcheology of the Hoskins House, Tannen-
baum Historic Park, Greensboro, North Carolina, U.S.A. Tree-
Ring Res 65:37–45. doi:10.3959/2007-11.1
Holmes RL (1983) Computer-assisted quality control in tree-ring
dating and measurement. Tree-Ring Bull 43:69–78
Holmes RL (1992) User’s manual for program EDRM. Laboratory of
Tree-Ring Research, University of Arizona, Tucson
Hoobler JA (2008) A guide to historic Nashville Tennessee. History
Press, Charleston
Kilgore SJ (1981) Brief history of the Association for the Preservation
of Tennessee Antiquities and its Belle Meade Mansion Historic
Site Museum. Thesis, Middle Tennessee State University,
Murfreesboro
Kniffen FB, Glassie H (1966) Building in wood in the eastern United
States: a time-place perspective. Geogr Rev 56:40–66
Mann DF, Grissino-Mayer HD, Faulkner CH, Rehder JB (2009) From
blockhouse to hog house: the historical dendroarcheology of the
Swaggerty Blockhouse, Cocke County, Tennesse, U.S.A. Tree-
Ring Res 65:57–67. doi:10.3959/2007-4.1
Morgan J (1990) The log house in east Tennessee. University of
Tennessee Press, Knoxville
Nash SE (1999) Time, trees, and prehistory: tree-ring dating and the
development of North American archaeology, 1914–1950.
University of Utah Press, Salt Lake City
Orvis KH, Grissino-Mayer HD (2002) Standardizing the reporting of
abrasive papers used to surface tree-ring samples. Tree-Ring Res
58:47–50
Rehder JB (2012) Tennessee log buildings: a folk tradition. Univer-
sity of Tennessee Press, Knoxville
Slayton JD, Stevens MR, Grissino-Mayer HD, Faulkner CH (2009)
The historical dendroarcheology of two log structures at the
Marble Springs Historic Site, Knox County, Tennessee, U.S.A.
Tree-Ring Res 65:23–39. doi:10.3959/2007-5.1
Speer JH (2010) Fundamentals of tree-ring research. University of
Arizona Press, Tucson
Stachowiak LA, Schneider EA, Rochner ML, Collins SA, Swiney CP,
Grissino-Mayer HD, McKenzie TG (2014) Dendrochronological
dating of the historic McKenzie Home, Meigs County, Ten-
nessee, USA. Tree-Ring Res 70:31–39. doi:10.3959/1536-1098-
70.1.31
Stahle DW (1979) Tree-ring dating of historic buildings in Arkansas.
Tree-Ring Bull 39:1–28
Stokes MA, Smiley TL (1996) An introduction to tree-ring dating.
University of Arizona Press, Tucson
Warden ML (1988) The Belle Meade Plantation. Association for the
Preservation of Tennessee Antiquities, Nashville
Wills R II (1991) The history of Belle Meade: mansion, plantation,
and stud. Vanderbilt University Press, Nashville
Veget Hist Archaeobot (2016) 25:105–115 115
123