declaration and recommendation - egerton...
TRANSCRIPT

EFFECT OF EXTRACTS FROM SELECTED Aloe PLANT SPECIES ON THE
Anopheles gambiae sensu stricto AND Aedes aegypti MOSQUITOES
Chore Judith Kavulani
A Thesis Submitted to the Graduate School in Partial Fulfillment of the Requirements
for the Award of Master of Science Degree in Biochemistry of Egerton University
EGERTON UNIVERSITY
AUGUST 2012

DECLARATION AND RECOMMENDATION
DECLARATION
I hereby declare that this thesis is my original work and has not been submitted for award in
any institution of learning to the best of my knowledge.
Signed:……………………… Date:……..…………….
Chore, Judith Kavulani
SM14/1518/05
RECOMMENDATION
We confirm that this thesis has our approval to be presented for examination as per the
Egerton University regulations.
Dr. Paul Mireji
Department of Biochemistry and Molecular Biology,
Egerton University
Signed:………….................... Date:.…….........……….
Dr.Francis Wachira
Tea Research Foundation of Kenya (TRFK)
Signed:………….................... Date:.…….........……….
Prof. Euty Wathuta
Department of Biological Science,
Egerton University
Signed:………….................... Date:.…….........………
2

COPYRIGHT
© Judith Kavulani Chore, 2012
All rights reserved. No part of this project report should be reproduced or transmitted
in any form by means of mechanical including photocopying, recording or any information
storage or retrieval system without permission in writing from the author or Egerton
University on behalf of the author.
.
3

DEDICATION
This work is dedicated with love to my entire family members, my husband Henry
Chore; and my dear children Allan Amiani, Kelly Everia, Tony Kahi and Mercy Cheredi.
Your tireless support, endurance and belief in me carried the day for me.
4

ACKNOWLEDGEMENT
I give all due glory to God almighty without whose help I would not have come this
far. I am indeed indebted to my supportive supervisors; Prof. Francis Wachira and Dr. Paul
Mireji. The success of this study has been due to your diligent advice, conceptual discussions,
steady encouragement and constructive criticism. I would also like to thank my other
supervisor, Prof. Euty Wathuta for the advice on the subject matter of insect physiology. I am
grateful to Egerton University for providing an enabling environment and financial support
during this study. I would like to thank the entire staff of the Department of Biochemistry,
Egerton University, for their support during this study. I extend my gratitude to the entire
staff of Entomology Department, of the Pyrethrum Board of Kenya, Nakuru, for providing
me with resources, support and logistics. I am especially grateful to my entire family
members for their encouragement and support during this time of study. Lastly but not least I
would like to thank the entire staff of the Department of Chemistry, Egerton University, for
their assistance and support during the formative stage of this study. You all deserve the best
in your respective specialties as you promote scientific research.
5

ABSTRACT
Malaria control through insecticide application has significantly been compromised
by the advent of resistance to the insecticide in the Anopheles mosquito vector. These
developments have necessitated the need to bio prospect for and understand the mode of
action of novel insecticide introduction. Larvicidal potential in extracts from Aloe plants on
Anopheles gambiae sensu stricto and Aedes aegypti third instar (two days old) larvae were
evaluated. Extracts were obtained from Aloe turkanensis, Aloe ngongensis and Aloe fibrosa
plants using classical solvent extraction technique. Larvicidal potential of the extract were
evaluated through bioassay of the extract on third instar An. gambiae s.s and Ae. aegypti
larvae following 24 h exposure. The dose-response data obtained were analyzed by probit
analysis to establish the median lethal concentration (LC50) of the extract to the larvae. The
LC50 responses against third instar larvae of An. gambiae s. s. and Ae. aegypti were 0.08 mg /
ml and 0.11mg / ml, respectively at 95% intervals. All the A. ngongensis plant extracts had
larvicidal activity. Only the methanol, acetone and hexane plant extracts of A. fibrosa species
showed larvicidal activities at LC50 concentrations ranging between 0.66 and 3.90 mg / ml at
95% confidence interval. Phytochemical tests showed presence of flavonoids, tannins and
saponins in Aloe turkanensis plant extract. Apart from the hexane and chloroform plant
extracts of Aloe ngongensis, those of methanol, acetone and ethyl acetate showed presence of
flavonoids. All the Aloe fibrosa plant extracts of hexane, acetone and methanol showed
presence of flavonoids and tannins. However, only the acetone and methanol plant extracts
of Aloe fibrosa showed presence of saponins. None of the Aloe plant extracts gave positive
test for phlobatannins, terpenoids and steroids. The phenolic functional group was prevalent
in most of the plant extracts. The leased significant difference (LSD) of the means of the
developmental stages of the Aloe exposed Ae. aegypti were significantly different from those
of the negative controls at 5% level. The leased significant means of the egg deposits, egg
viability and mortality were significantly different in the Aloe exposed Ae. aegypti at 5%
level The findings of this study will contribute to current Anopheles vector control programs
by providing information on potential additional anti-Anopheles compound(s) and molecular
process (es) of subtractive hybridization with bioinformatics, that can be exploited in
development of novel insecticides.
TABLE OF CONTENTS
6

DECLARATION AND RECOMMENDATION.............................................................ii
COPYRIGHT.....................................................................................................................iii
DEDICATION....................................................................................................................iv
ACKNOWLEDGEMENT..................................................................................................v
ABSTRACT........................................................................................................................vi
TABLE OF CONTENTS..................................................................................................vii
LIST OF ABBREVIATIONS.............................................................................................x
LIST OF TABLES.............................................................................................................xi
LIST OF FIGURES..........................................................................................................xii
CHAPTER ONE: INTRODUCTION...............................................................................1
1.1 Background Information.................................................................................................1
1.2 Statement of the problem................................................................................................2
1.3 Objectives of the study....................................................................................................2
1.4 Hypotheses......................................................................................................................3
1.5 Justification.....................................................................................................................3
CHAPTER TWO: LITERATURE REVIEW..................................................................5
2.1 The mosquitoes...............................................................................................................5
2.2 Classification and Distribution of Anopheles gambiae complex..................................10
2.3. Life cycle of Anopheles gambiae and Aedes aegypti...................................................11
2.4 Control Measures for mosquitoes.................................................................................15
2.5 Use of plants in mosquito control.................................................................................17
2.6 Mode of Action of Insecticides.....................................................................................19
2.7 Vector Resistance to Insecticides..................................................................................21
2.8 Characteristics of Aloe species......................................................................................22
7

2.8 Aloe species Phytochemicals.........................................................................................24
2.9 Phytochemistry and plant pharmacology screened medicinal plants............................25
CHAPTER THREE: MATERIALS AND METHODS.................................................27
3.1Sampling of test Aloe species.........................................................................................27
3.2 Test Insect.....................................................................................................................27
3.3 Aloe leaf homogenization and subsequent solvent extraction of phytochemicals from plant
material.........................................................................................................................28
3.4 Larvicidal tests..............................................................................................................29
3.5 Physiological Responses...............................................................................................29
3.6 Phytochemical and functional group tests of the Aloe plant extracts............................30
3.7 Data analysis.................................................................................................................31
CHAPTER FOUR: RESULTS........................................................................................32
4.1 Larvicidal results...........................................................................................................32
4.2 Phytochemical qualitative analysis results of selected larvicidal Aloe plant extracts. .35
4.3 Analysis of chemical functional groups in Aloe extracts..............................................37
4.4 The effect of Aloe plant extracts on Ae. aegypti...........................................................39
CHAPTER FIVE: DISCUSSION, CONCLUSION AND RECOMMENDATION....42
5.1 Discussion.....................................................................................................................42
5.2 Conclusion.....................................................................................................................44
5.3 Recommendation...........................................................................................................44
REFERENCES....................................................................................................................45
APPENDICES.....................................................................................................................62
Appendix 1:F0 Post 24 hour exposure to Aloe extracts......................................................62
Appendix 2: Pupae emergence............................................................................................63
Appendix 3: Adult Male Emergence...................................................................................63
8

Appendix 4: Adult female emergence.................................................................................64
Appendix 5: Male: female sex ratio (Male/Total)...............................................................64
Appendix 6: Egg deposits (oviposition) studies on 30 males+30 females..........................65
Appendix 7: Age specific survivorship of adults for males ( F0) where; x=yx/y0.............65
Appendix 8: Age-specific survivorship of female adults (F0)............................................66
Appendix 9: Male Age specific mortality studies (F1).......................................................66
Appendix 10: F1 Egg Hatchability 48 hours post Aloe extracts exposure of 1500 eggs of
Aedes aegypti...............................................................................................................67
Appendix 11: Larvae emergence.........................................................................................67
Appendix 12: Pupae emergence..........................................................................................68
Appendix 13: Adult male emergence..................................................................................68
Appendix 14: Adult female emergence...............................................................................69
Appendix 15: Male and female sex ratio............................................................................69
Appendix 16: Aedes aegypti F1 oviposition.......................................................................70
Appendix 17: Qualitative analysis of terpenoids from selected larvicidal Aloe plant extract71
Appendix 18: Qualitative analysis of steroids of selected larvicidal Aloe plant extracts. . .72
Appendix 19: Qualitative analysis of flavanoids of selected larvicidal Aloe plant extracts73
Appendix 20: Qualitative analysis of tannins and polyphenols of larvicidal selected Aloe plant
extracts.........................................................................................................................74
Appendix 21: Phlobatannins analysis in selected Aloe plant extracts................................75
Appendix 22: Qualitative saponin test results for selected larvicidal Aloe plant extracts. .76
9

LIST OF ABBREVIATIONS
AFA Aloe fibrosa acetone extract
AFH Aloe fibrosa hexane
AFM Aloe fibrosa methanol extract
ANA Aloe ngongensis acetone extract
ANC Aloe ngongensis chloroform extract
ANE Aloe ngongensis ethyl acetate extract
ANH Aloe ngongensis hexane extract
ANM Aloe ngongensis methanol extract
ATE Aloe turkanensis ethyl acetate extract
DDT Dichloro diphenyl trichloroethane
EPI WHO Expanded Programme on Immunization
FAO Food for Africa Fund
Kdr Knockdown resistance
LC 50 Concentration required to kill 50% of the test organism
LC 90 Concentration required to kill 90% of the test organism
LC 99 Concentration required to kill 99% of the test organism
IPM Integrated pest management
IGRs Insect growth regulators
IRS Indoor residual spraying
MOH Ministry of Health
RBM Roll back malaria
UNICEF United Nations Childres’s Fund
10

LIST OF TABLES
Table 1: Probit analysis results for the concentrations (mg/ml) of selected Aloe plant extracts
against An. gambiae s. s. third instar larvae...............................................................33
Table 2: Probit analysis results for concentrations (mg/ml) of selected Aloe plant extracts
against Ae. aegypti third instar larvae........................................................................34
Table 3: Summary of the qualitative phytochemical constituents of the selected Aloe plant
extracts.......................................................................................................................36
Table 4: Aloe species functional groups..................................................................................38
Table 5: The effect of Aloe plant extracts on 1500 third instar larvae of Ae. aegypti parental
generation (F0) and first generation (F1) and the subsequent developmental stages
...................................................................................................................................40
LIST OF FIGURES
11

Figure 1: Female Aedes aegypti feeding on human flesh ( Wikipedia, 2012)........................7
Figure 2: Female Anopheles gambiae feeding on human flesh (Wikipedia 2012)....................8
Figure 3: Distinctive proboscis male mosquito (right) and female mosquito (left) (Constant
and Lam, 2012).......................................................................................................9
Figure 4: The hairy (plumose) male antennae and the thinly haired (pilose) antennae of
female Culicinae mosquito (Clements, 2002).........................................................9
Figure 5: The complete life cycle of a mosquito (Oklahoma University, 2012).....................14
Figure 6: Aloe turkanensis plant prevalent in North Western Kenya and the Karamajong
District of Uganda (Bosch, 2006).........................................................................23
Figure 7: The chemical formulae of the phytochemicals found to be prevalent in the Aloe
plant species (Dagne et al., 1996).........................................................................25
Figure 8: The responses of third instar larvae post 24 hour exposure to selected Aloe plant
extracts and the negative controls.........................................................................41
Figure 9: The effect of Aloe plant extracts on the pupae of Ae. aegypti 42
12

CHAPTER ONE
INTRODUCTION
1.1 Background Information
The mosquitoes are a family of small, midge-like flies: the Culicidae (Foot and Cook,
2005). Although a few species are harmless or even useful to humanity, most are a nuisance
because they consume blood from living vertebrates, including humans (Clements, 1992 and
Apperson et al., 2002). In feeding on blood, various species of mosquitoes transmit some of
the most harmful human and livestock diseases. Effective management of the malaria and
arbovirus vectors by current classes of mosquitocides is challenging, and necessitates
prospecting for novel insecticides with unique modes of action, different from the current
ones. Current adulticide classes (organochlorines, organophosphates, carbamates, pyrethroids
and DDT) are limited to three different modes of action (little target-site diversity), with two
different target sites (AchE or voltage gated sodium channel) and a single component (central
nervous system) of the vector. It is not therefore surprising that there is cross-resistance to the
insecticides in the vector to insecticides within the classes, phenomena currently widespread
across Sub-Sahara Africa (Chandre et al., 2000; Enayati and Hemingway, 2010). The
resistance to pyrethroids presents a real and immediate challenge to efficacy of otherwise
successful insecticide treated nets (ITN) based malaria control intervention against adult
vectors (Etang et al., 2004). Resistance development has also hampered larvae control
(Diabate et al., 2003), a viable alternative control of the vector populations. This initiative
has further been compounded by development of resistance in the larvae to new classes of
larvicides (IGRs, bacterial endotoxins and even Bacillus thuringiensis (Bti)) (Amy, et al.,
2005 and Wirth et al., 2005).
Recent phytochemical research has begun to reveal a variety of blend effects in the
bioactivities of plant natural products. Two principal blend effects have been demonstrated:
(1) enhanced biological activity resulting from synergistic or other additive effects of
moderately active or individually inactive compounds to give mixtures that are more active
than a linear summation of individual activities (Berenbaum and Zangeri, 1987; Isman et al.,
2008; Romeo et al., 1996; Bekele and Hassanali, 2001); and (2) mitigating effects of
structurally related or unrelated compounds against rapid resistance development that
characterizes most single-component bioactive compounds (Feng and Isman, 1995; Isman et
al., 1996) (characteristic of current mosquitocide classes). Most plants produce a variety of
1

secondary metabolites, which may or may not be structurally related, with multiplicity of
defense and non-defense functions against different pathogens and herbivores. This
phytochemical and functional diversity has undoubtedly arisen from sustained selective
forces in response to succession of attack by pathogens and herbivores and other selective
pressures over evolutionary time. Moreover, phytochemical blends rarely demonstrate acute
toxicity. They are often subtle and have longer-term growth-disrupting effects. This project
explored the larvicidal effect of Aloe plant extracts on Anopheles gambiae and Aedes aegypti
and the adverse influence on their parental (FO) and first generation (F1) developmental
stages.
1.2 Statement of the problem
Anopheles gambiae and Ae. aegypti are the principal Afro tropical vectors of malaria
and arbovirus respectively. These vectors have developed resistance to most conventional
insecticides, exposing most of the populations in Sub Saharan African and most of the tropics
at risk of contracting the diseases they vector. Adult mosquitoes are highly mobile flying
insects that can easily detect and avoid many intervention measures as compared with the
larvae that are confined within relatively small aquatic habitats. Malaria control needs
multiple initiatives given that plasmodium falciparum has developed resistance to most drugs,
including artemisinin-based combination therapy. Resurge in mosquito-borne diseases due to
inter-annual and inter-decadal climate variability calls for collaborative intervention measures
to contain epidemiology.
1.3 Objectives of the study
1.3.1 General Objective
To determine the effect of extracts from Aloe plant species on the immature stages of
Ae. aegypti (insecticidal) and the mortality of An. gambiae s. s. and Ae. aegypti mosquito
larvae.
1.3.2 Specific Objectives
1. To determine toxicity of organic extracts of A. turkanensis, A. ngongensis and A. fibrosa
plant species to An. gambiae s.s and Ae. aegypti larvae.
2. To determine phytochemical composition of A. turkanensis, A. ngongensis and A.
fibrosa plant extracts which are toxic to An. gambiae s.s and Ae. aegypti larvae.
2

3. To determine effect in An. gambiae s.s and Ae. aegypti larvae to A. turkanensis, A.
ngongensis and A. fibrosa plant extracts.
1.4 Hypotheses
1. Organic extracts of A. turkanensis, A. ngongensis or A. fibrosa plant species are not
significantly toxic to An. gambiae s.s and Ae. aegypti third instar larvae.
2. Aloe turkanensis, A. ngongensis and A. fibrosa plant extract phytochemical composition
do not significantly influence their toxicity to An. gambiae s.s and Aedes aegypti third
instar larvae.
3. There are no specific effects of Aloe plant extracts of A. turkanensis, A.fibrosa and A.
ngongensis on An. gambiae and Ae. aegypti third instar larvae.
1.5 Justification
Malaria control in the tropics and Africa South of the Sahara is based on insecticide-
treated nets (ITNs), indoor-residual spraying (IRS) with insecticides and prompt and effective
treatment of clinical malaria. Malaria control initiatives such as U.S. Presidents Malaria
Initiative, Roll Back Malaria, Global Strategic Framework for Integrated Vector Management
and other partnerships have been involved in mosquito-borne decease control campaigns.
Many believe integrated vector management (IVM), targeting both larval and adult
mosquitoes, is the future for malaria control. While ITNs are currently the priority strategy,
there is growing interest in attacking the aquatic stages of malaria vectors with microbial
larvicides, in conjunction with environmental management. Biological vector control makes
use of the predatory mosquito larvae (Toxorhychites spp.), the mermethid or mosquito-
attacking nematode, Romanomermis culcivorax and/or the mosquito-eating fish, Gambusa
affinis, killfish (fundulidae), minnows (cyprinidae), and/or tilapia and or guppies, e.g.,
Legister reliculatus wherever possible. Insect growth regulators (IGRs) such as methoprene
(altosid) are used to stop mosquito maturity to adult but insect resistance has been reported in
this class of larvicides, just as in the convectional insecticides.
Extracts from Aloe plant species have secondary metabolites with insecticidal
potential. If effective against An. gambiae and Ae. aegypti these extracts can contribute to the
management of these malaria, arboviruses and/or filarial worm vectors by integrating into the
current and existing control methods. Additionally, application of the phytochemicals in
vector control will also be more environmentally friendly than application of conventional
3

synthetic insecticides. The use of Aloe phytochemicals is potentially useful in the control of
the juvenile stages of the vector, particularly larvae, which has been overlooked in many
vector control programs. The use of Aloe plant extracts against larvae confined to water
habitats will be more efficient than the convectional use of insecticides against the more
elusive adult stage of the mosquitoes. It is therefore prudent to evaluate the larvicidal
potential of extracts (phytochemicals) from Aloe plant species on the larvae of the vectors and
identify physiological responses in the vector to the phytochemicals that can be exploited in
developing novel biolarvicides.
4

CHAPTER TWO
LITERATURE REVIEW
2.1 The mosquitoes
Malaria remains one of the most devastating diseases occurring in the world today. It
is estimated that about 350 - 500 million clinical cases occur every year with approximately
1-3 million deaths in tropical Africa alone (WHO and UNICEF, 2003). This represents at
least one death in every thirty seconds (WHO, 1996). Majority of the cases occur in children
under five years and pregnant women are also especially vulnerable. Approximately 40% of
the world’s population lives in regions where malaria transmission is endemic (Agyepong,
2012). It has been established that 90% of the global malaria morbidity and mortality occurs
in sub-Saharan Africa (UNICEF, 2010). In Kenya, over 28 million Kenyans are at risk of
malaria infection and an estimated 34,000 children less than five years of age die annually
due to malaria (Mugo, 2012). It accounts for 30-50% of all outpatient attendance and 20% of
all admissions to health facilities. An estimated 170 million working days are lost to the
disease each year (MOH 2001). Malaria is also estimated to cause 20% of all deaths in
children under five (MOH 2006). The economic burden in Kenya due to malaria related cases
was 2.3 billion shillings (Kemri, 2012).
Malaria transmission in Kenya is by all the four species of human Plasmodium: P.
falciparum, P. malariae, P. ovale and P. vivax. P. falciparum which causes the severest form
of the disease accounts for 98 percent of all malaria infections. The major malaria vectors in
Kenya are members of An. gambiae complex and An.funestus (Lyimo et al 1992 and Keating
et al., 2005). Kenya has four malaria epidemiological zones: endemic, seasonal malaria
transmission, Malaria epidemic prone areas of western highlands of Kenya and low risk
malaria areas (Kemri, 2012). The malaria endemic zone lies at the altitudes ranging from 0 to
1300 meters around Lake Victoria in western Kenya and in the coastal regions. Rainfall,
temperature and humidity are the determinants of the perennial transmission of malaria. The
vector life cycle is usually short with high survival rate due to the suitable climatic conditions
(Jean-Michel et al., 2003). Transmission is intense throughout the year with annual
entomological inoculation rates between 30 and 100 (Kemri, 2012). Samples of blood taken
from mosquitoes across section of Kenyans have shown seroprevalence to the arbovirus in
both humans and mosquitoes (Luke et al., 2011 and Desiree et al., 2011). Kenya has had
multiple arbovirus outbreaks in the past two decades resulting in economic and public health
5

distress, including yellow fever in 1992 and 1995), chikungunya fever in 2004, and Rift
Valley fever (RVF) in 1997 and 2006 (Bird et al., 2008 WHO, 2011). Yellow fever is a viral
hemorrhagic fever which strikes an estimated 200 000 persons world-wide each year and
causes an estimated 30 000 deaths (WHO, 1992). Yellow fever virus is the prototype of the
family Flaviviridae, which currently contains over 70 viruses, of which most are arthropod-
borne, including the dengue viruses (Monath, 1991 and Kuniholm et al., 2006). Dengue fever
virus, the most common of the viruses, is transmitted by the bite of an Aedes mosquito
(Stacy, 2012). Infections can be spread among humans via the mosquito vector.
Approximately 2.5 billion people worldwide are at risk of infection with dengue fever in over
100 countries are at risk of infection and 20 million infections are reported annually (WHO,
2010). Patients present with fever, headache, eye, and back pain. The main vector of yellow
fever within village and urban settlements is female Aedes (Stegomyia) aegypti (only females
feed on blood to obtain protein for egg production) Gubler, (1988). The virus is transmitted
when a mosquito bites an infected human and then, after an extrinsic (in the mosquito)
incubation period of 12-21 days, bites a susceptible human. There is no specific treatment for
dengue/ severe dengue, but early detection and access to proper medical care lowers fatality
rates below 1% (WHO, 2012). Dengue prevention and control solely depends on effective
vector control measures. Ae. aegypti breeds readily in all types of domestic and peridomestic
collections of fresh water, including flower vases, water drums, tin cans, broken coconut
shells, old tyres and gutters (WHO, 1998). Apart from Brugia malayi and Wuchereria
bancrofti, Ae. aegypti also transmits the filarial worm. Lymphatic filariasis is estimated to
affect more than 120 million people worldwide, 98% of who live in the tropical and
subtropical regions of Africa and Asia (Michael et al., 2001 and Braga et al., 2003). There
are 72 countries with endemic lymphatic filariasis and a total population of more than 1.3
billion at risk for infection (Jamshaid, 2006). Approximately one third of infected individuals
have physical manifestations of elephantiasis. Physical disabilities due to elephantiasis and
other chronic organ damage result in the loss of nearly 6 million lives per year. The greatest
impact is in Asia followed by Africa (WHO, 2006).
Mosquitoes belong to the order Diptera, a group of insects that only have one pair of
wings located on the methothorax. There are about 3200 species of mosquitoes belonging to
the family Culicidae (Service and Ashford, 2001). Most species of medical importance fall
within the sub-families Anophelinae (of which the most important genus is Anopheles (Figure
2) and Culicinae (which comprises of Aedes (Figure 1), Culex and Mansonia) (Conn, 1997).
6

There are many species of non-biting gnats and midges, which resemble mosquitoes in
several features but do not have the long forwardly- projecting proboscis (Burgess and
Cowan, 1993). Both male and female mosquitoes feed on plant fluids and nectar (Carpenter
and Walter, 1974). However only the female mosquito typically requires a blood meal from a
warm –blooded animal before a viable batch of eggs can be laid. It is the female mosquito
only that is capable of sucking blood (Lehane, 2005). The internal male mouthparts are short
and only extend about a quarter of the length of the proboscis. In contrast, the female has
long, needle-like mouthparts, which are capable of piercing animal tissue (Kong and Wu,
2009). Male mosquitoes have a pair of long bushy (plumose) antennae, (Figures 3 and 4)
whereas the antennae of the female are sparsely haired or pilose (Burgess and Cowan, 1993).
Figure 1: Female Ae. aegypti feeding on human flesh (Wikipedia, 2012)
7

The female An. gambiae is distinguished from other mosquitoes by the palpi, which are as
long and straight as the proboscis, while palpi of female Culicinae are considerably shorter
(Figure 2). Unlike other mosquitoes, the Anopheles have discrete blocks of black and white
scale on their wings (Das et al., 2007) Adult Anopheles can also be identified by their typical
resting position: males and females rest with their abdomens sticking up in the air rather than
parallel to the surface on which they are resting. One important behavioral factor is the
degree to which an Anopheles species prefers to feed on humans (anthropophagy) or animals
such as cattle (zoophilic) Howell and Knols, 2009).
Figure 2: Female An. gambiae feeding on human flesh (Wikipedia 2012)
8

Generally the following features can be used to differentiate a male mosquito from a female
mosquito: male mosquitoes tend to hatch before the females, and also generally have shorter
lifespan.
Figure 3: Distinctive proboscis male mosquito (right) and female mosquito (left)
(Constant and Lam, 2012)
Figure 4: The hairy (plumose) male antennae and the thinly haired (pilose) antennae of
female Culicinae mosquito (Clements, 2002)
9

Both sexes have one pair of compound eyes (Gao et al., 2007). Mosquitoes undergo complete
life cycle: egg, larva and pupa before becoming adult (Schäfer and Lundström. 2006). The
immature stages are always associated with water which may occur in a ward range of
locations (Ulricke et al., 2004). The mosquito egg hatches into a small worm-like larva which
feeds on microorganisms in the water or on the water surface using paired mouth brushes on
the head (Lehane, 2005). Vision is rudimentary but larvae react rapidly to changes in light
intensity, moving actively with a wriggling of darting motion through water (Burgess and
Cowan, 1993). The bulky, thoracic part of the larva often has long bristles or hairs, which
assist in achieving balance. The larva passes through four stages or instars before molting to
the pupa stage (Nishiura et al., 2003).
The pupa is coma shaped; the head and thorax having fused to form cephalothorax,
with the abdomen hanging down from it. The pupa stage is actively mobile using a pair of
paddles located on the hind end of the abdomen to progress in a tumbling motion through the
water (Schäfer and Lundström, 2006). It does not feed but comes to water / air interface to
obtain oxygen through a pair of dorsal trumpets on the cephalothoraxes. The adult mosquito
can be seen developing through the papal skin. When the adult mosquito is fully developed,
the pupa comes out to the surface, and splits across the dorsum, the adult emerging to stand
on the water surface, while the exoskeleton hardens and dries. Males will typically emerge
first and swarm in the air over the breeding site (Leon and Steven, 2002). When the females
emerge mating will take place. The female mosquito stores sperm from a single mating in the
sperm theca and these will be used to fertilize eggs from alternate ovaries when required
(Burgess and Cowan, 1993). Thus the female mosquito needs to be fertilized once in a
lifetime but may lay up to ten or so batches of eggs (Clements, 1992). The female will
typically feed in subdued light, especially during the night, but some species will readily feed
during the day (Lehane, 2005). Depending on the species, female mosquitoes may rest
indoors or outside before or after feeding (WHO, 2006). Identification of the species is thus
essential in order that correct control measures are carried out.
2.2 Classification and Distribution of Anopheles gambiae complex
There are 460 species of the Anopheles mosquito, and about 100 of these are able to
transmit malaria and 30-40 are notorious vectors in many parts of the world (Lehrer, 2010).
Anopheles gambiae is one of the best known because of its predominant role in transmitting
of the most dangerous malaria parasite, Plasmodium falciparum (Snow et al., 2005). An.
10

gambiae mosquitoes belong to the family Culicidae, order Diptera in the phylum Arthropoda
of the animal Kingdom (Leunita and Ogbunugafor, 2008). They constitute a complex of
seven sibling species. An. gambiae s. s. is the prolific African malaria vector and has
expanded its niche into polluted habitats. Recent sdudies found An. gambiae larvae thriving
in a variety in a variety of anthropogenic urban water bodies, which contained pollution from
domestic and/or industrial sewage (Awolola et al., 2007). It comprises several chromosomal
forms like Bamako, Mopti and Savanna. Anopheles arabiensis and An. gambiae s s. are
isolated reproductively by mainly pre –mating barriers though both occur in sympatry in
tropical Africa. Their larval stages inhabit mainly fresh water habitats (Muirhead-Thompson,
1945). Anopheles melas Theobald and Anopheles merus Donitz are salt-water species found
on the West and East coasts of Africa respectively (Gillies and de Meillon, 1968). Anopheles
bwamba White is found in hot water springs in the Ugandan Bwamba County (White, 1985).
It breeds in brackish water .Anopheles quadriannulatus species B is found in Ethiopia and is
considered not as an important malaria vector due to its preference for cattle. Anopheles
quandriannulatus Theobald is a Plasmodium falcipurum vector in South Africa (Walker et
al., 2007). Anopheles gambiae sensu lato is the main vector transmitting P. falciparum south
of the Sahara. The sibling’s species belonging to An. gambiae complex are morphologically
indistinguishable. Differences between them have been detected using various techniques
including the use of banding patterns of polytene chromosomes, enzyme variations banding
sequences of mitotic sex chromosomes, DNA and RNA probes (Colluzi et al., 2002 and
Fanello et al., 2002).
2.3. Life cycle of Anopheles gambiae and Aedes aegypti
Mosquitoes act as vectors for many human diseases including malaria, West Nile,
yellow fever, encephalitis, and dengue (Service, 2001 and WHO, 2006). To understand a
disease like dengue fever it is essential to understand the life cycle of the mosquito host, the
human host, and the virus, and also to understand the various environmental factors that can
support disease transmission including potential epidemics and pandemics (Gubler, 1998).
Anophelines and Aedes mosquitoes go through complete metamorphosis of four stages in
their life cycle: egg, larva, pupa and adult (Service, 1993) (Figure 5). The first three stages
are aquatic and last 5-14 days, depending on the species and the ambient temperatures. Adult
female Anopheles mosquitoes act as vectors for the malaria parasites and filarial worms
(Gillies and De Meillon, 1968). The adult females can live up to a month (or more in
captivity) but most probably do not live more than 1-2 weeks in nature (Leunita and
11

Ogbunugafor, 2008). The adult female lays 50-200 eggs per oviposition. Eggs are laid singly
directly on water and are unique in having floats on either side. Each egg is protected by an
egg shell, which in many species is elaborately sculpted. Eggs are not resistant to drying and
hatch within 2-3 days, although hatching may take up to 2-3 weeks in colder climates
(Clements, 1992). Mosquito larvae have a well-developed head with mouth brushes used for
feeding, a large thorax and a segmented abdomen. In contrast to other mosquitoes, Anopheles
larvae lack a respiratory siphon and position themselves so that their body is parallel to the
surface of the water (Gillies and De Meillon, 1968). Larvae breathe through spiracles located
on the 8th abdominal segment and therefore must come to the surface frequently. The larvae
spend most of their time feeding on algae, bacteria, and other microorganisms in the surface
micro layer (Clements, 1992). They dive below the surface only when disturbed. Larvae
swim either by jerky movements of the entire body or through propulsion with the mouth
brushes. Larvae develop through four stages, or instars, after which they metamorphose into
pupae (Goma, 1959). At the end of each instar, the larvae molt, shedding their exoskeleton, or
skin, to allow for further growth. Larvae of mosquitoes occur in a wide range of habitats but
most species prefer clean, unpolluted water (Clements, 1999).
Larvae of Anopheles mosquitoes have been found in fresh or salt-water marshes,
mangrove swamps, rice fields, grassy ditches, the edges of streams and rivers, and small,
temporally rain pools (Gilbert and Calderone, 2007). The pupa is comma shaped when
viewed from the side. The head and surface of the cephalothorax splits and the adult
mosquito emerges (Clements, 1992). The whole process from egg to emergence of the adult
from the pupa takes not more than a week at tropical temperatures (Gillies and De Meillon,
1968). Adults of An. gambiae emerge during the late afternoon, and once mature, mate during
a twenty-minute period at dusk. When a female has matured a batch of eggs, she takes to the
wings and responds to stimuli from suitable oviposition sites. For most mosquitoes, the
oviposition site is a water body with particular characteristics: odor, taste, flow and shade; all
known to influence different species (Clements, 1999). Within an hour of completing one
gonotrophic cycle; a female may commence another. At warmer temperatures, like in the
tropics, Anopheles oviposits regularly every two or three days (Service, 1993). Many species
prefer habitats with vegetation. Others prefer habitats that have none. Some breed in open,
sun-lit pools while others are found only in shaded breeding sites in forests. A few species
breed in tree holes or the leaf axils of some plants (Blackwell and Johnson, 2000). The Aedes
aegypti mosquito can be recognized by white markings on legs (Womack, 1993) ( Figures 1
12

and 5). The mosquito originated in Africa but is now found in tropical and subtropical
regions throughout the world. The immature stages of the mosquitoes are always associated
with free water of some sort but the type and location vary considerably (Munga et al., 2005).
Some breeding places for Aedes aegypti are artificial water containers, such as the odd plastic
bucket, flowerpot "saucer", or discarded bottle or tires. Aedes females generally drop their
eggs singly, much as Anopheles do, but not as a rule into water (Wikipeadia, 2012). Instead,
they lay their eggs on damp mud or other surfaces near the water's edge. Such an oviposition
site commonly is the wall of a cavity such as a hollow stump or a container such as a bucket
or a discarded vehicle tire. The eggs generally do not hatch until they are flooded, and they
may have to withstand considerable desiccation before that happens (Nelson, 1984). They are
not resistant to desiccation straight after oviposition, but must develop to a suitable degree
first (Service, 1993). Once they have achieved that, however, they can enter diapause for
several months if they dry out. Clutches of eggs of the majority of mosquito species hatch as
soon as possible, and all the eggs in the clutch hatch at much the same time (Clements, 1992).
In contrast, a batch of Aedes eggs in diapause tends to hatch irregularly over an extended
period of time. This makes it much more difficult to control such a species than those
mosquitoes whose larvae can be killed all together as they hatch (Wikipedia, 2012). Some
Anopheles species do also behave in such a manner, though not to the same degree of
sophistication (Huang et al., 2006).
One method of classifying mosquitoes, which is important in the control of the larval
stage, is by the type of habitat in which the eggs are laid. Such species that lay eggs singly on
the moist soil usually near the edge of temporary pools of water are known as flood water
mosquitoes. These eggs only hatch after they have been flooded by water. Psorophora,
Aedes, and Ochlerotatus mosquitoes are floodwater mosquitoes which are most abundant
shortly after spring rainfall (Oklahoma, 2012). Those species that lay eggs on the surface of
the water, either clumped in rafts or as single floating eggs, are known as permanent water
mosquitoes. Anopheles, Culiseta, and Culex are permanent water mosquitoes. Eggs of Aedes
aegypti are long, smooth, ovoid shaped, and approximately one millimeter long. When first
laid, the eggs appear white, but within minutes they turn shiny- black. In a warm climate such
as the tropics, the eggs may develop in as little as two days, whereas in cooler temperatures
development can take up to one week (Foster and Walker, 2002). Anopheline and Culicine
larvae can be distinguished in the field by their different resting positions in the water
(Clements, 1992). Culicine larvae have siphons through which they obtain air from the
13

surface, enabling it to feed below the surface .Aedes aegypti larvae also have no palpate hairs.
The males of Aedes aegypti feed on nectar while the females almost exclusively feed on
blood (Wikipedia, 2012). This explains how the arboviruses are transmitted by the Aedes
aegypti mosquito as the female mosquito spreads the yellow fever, dengue fever and filarial
worms while in search of blood (Linquist et al., 2012). Aedes aegypti also becomes a vicious
vector of the arbovirus and filarial worms due to the female habit to take blood meals more
than once in a life time. The adult Aedes aegypti is a small to medium sized mosquito,
approximately 7 millimeters .Aedes adults have white scales on the dorsal (top) surface of
the that basal bands forming the shape of violin or lyre (Carpenter and La Casse, 1985).
Figure 5: The complete life cycle of a mosquito (Oklahoma University, 2012)
2.4 Control Measures for mosquitoes
Efforts to eradicate malaria have failed and parasitic resistance to the most commonly
used and affordable anti- malarial drugs is developing rapidly (Leunita and Ogbunugafor,
14

2008 and WHO, 2011). Insecticide resistance in the vector is also an evolving problem,
(Hargreaves et al, 2003). A malaria vaccine is the subject of most research but its testing is
incomplete and full development remains a distant goal (Francine et al., 2003). The use of
residue pyrethroids to treat bed nets became more fashionable as a means of controlling adult
stage malarial vectors for the following reasons: first, most Anophelines bite indoors late at
night and bed nets thus intercept mosquitoes as they approach sleepers in search of blood
(Clements, 1999). Secondly, in many countries there has been a tendency to replaster walls as
soon as they have been sprayed covering up insecticide deposit (WHO, 1996). While
insecticide treated nets (ITNs) are currently the priority strategy in containing the adult stage
of the mosquito, there is growing interest in attacking the aquatic stages of malaria vectors
with microbial larvicides in conjunction with environmental management (Utzinger et al.,
2002) This initiative that originally started in Tanzania, was meant to spread to other
countries under the Presidents’ Malaria Initiative (Utzinger et al., 2002). Global initiatives to
fight malaria and other vector borne diseases so as to manage and control them to tolerable
levels include: Roll Back Malaria (RBM), Medicines for Malaria Venture (MMV), Malaria
Vaccine Initiative (MVI), Mapping Malaria Risk in Africa (MARA), Multilateral Initiative
on Malaria (MIM), The Presidents Malaria Initiative (PMI) and other global partners (WHO,
2012). These initiatives are responsible for the distribution of insecticide treated bed nets, use
of treatment based on combination therapy (Mugo, 2012), concise delivery of insecticides
and drugs (RBM), development of new antimalarial drugs (MMV) the development of
chemical testing of malaria vaccines (MVI), the mapping of malaria risk (MARA) and the
promotion of malaria research and capacity building (MIM). A marked increase in malaria
has recently been noted in the African highlands, largely due to the rise of drug-resistant
strains of Plasmodium falciparum parasites (Bøcker et al., 2000). The ecological features of
the western highlands of Kenya support stable parasite transmission, and increasing
population pressure has led to the clearance of natural swamps, massive deforestation and
crop cultivation in the valley bottoms (Munga et al 2006). Because of these agricultural
changes, many water bodies are now exposed to the sun and provide ideal conditions for
vector proliferation and increased malaria transmission (Briet et al., 2003) .Malaria control in
these highlands is based on insecticide-treated nets (ITNs), indoor-residual spraying (IRS)
with insecticides and prompt and effective treatment of clinical malaria Many believe
integrated vector management (IVM), is the way forward in malaria control (Role Back
Malaria, 2005 and Nagera et al., 2011). Integration of larval source management into ongoing
15

programmes is likely to be most effective when transmission is moderate or low where
mosquito breeding sites are contained and well defined (Kitron et al., 1989). This will work
mainly in urban and peri urban areas. Larval control can be attained through environmental
management, large space coverage and community participation (Utzinger, 2001). This can
be achieved by use of chemical or biological control.
Mosquito larvae basically have the nature to breed everywhere in small amounts of water on
the surface of the ground and hence their control can be acceptable only under suitable
mapping and characterization of breeding sites. Vector control should target all stages of the
mosquito life cycle yet for the last 50 years it has focused on the adult mosquito control
(WHO, 1999). While ITNs have saved live in Africa, their use is limited as adult mosquitoes
feed outside houses and before sleeping hours. Hence larval source management targeting
both indoor and outdoor vector populations should contribute to greater reductions in
transmissions than ITNs alone (Kitron and Spielman, 1989).There has been reduced efficacy
of insecticide treated nets and indoor residue spraying for malaria control due to pyrethroids
resistance (Nʹ Guessan et al., 2007). Also there have been reports of emergence of resistance
in field populations of tropical Culex quiquefasciatus to the microbial agent Bacillus
sphaericus (Mulla et al., 2003). Integrated vector management (IVM) programme consists of
standard water management, open marsh-water management, biological control, larval and
adult mosquito control. These techniques are based on an understanding of the mosquito life
cycle. Water management exploits the fact that the larvae are vulnerable to removal of water
they need to survive on. Biological control uses fish and other predators to eat the larvae.
Water management and biological control are combined in open marsh water management.
Larval control targets mosquito’s larvae using highly specific materials such as bacterial
pesticides and insect growth regulators (Amy, 2006). Bacterial larvide (Bacillus thuringiensis
var.israelensis) is highly specific to mosquito larvae and is an environmentally friendly
product (WHO, 2005).
A new bacterial granular larvicide with the trade name Vectolex ®CG with live
Bacillus sphaericus as its active ingredient is used in larval control programme. This provides
a new form of cost effective long term control in areas that continually hold water and breed
mosquitoes such as drainage ditches and swamps (Gilbert and Calderone, 2007). A juvenile
growth hormone with the trade name Altosid prevents the mosquito from molting from the
larval stage to adult. Larvicidal effects of a neem (Azadiracta indica) oil formulation on the
malaria vector control has been explored and other phytochemicals explored for larvicidal
16

activity include extracts from Aloe species and cashew nut shell in their possible use in
malaria control (Amy, 2006). Protection against mosquito bites includes closing the doors
and windows in the evenings to prevent entry into human dwellings, using mosquito repellant
lotions, creams, mats or coils (WHO, 2006). Local infestations can be managed using hand
held or truck mounted sprayers. Adult control is accomplished using ground or aerial
applications of ultra- low-volume (ulv) aerosols of materials that rapidly degrade in the
environment (Gilbert and Calderone, 2007 and Nobert et al., 2010). Travelers to endemic
areas and high-risk individuals should be started on anti- malarial drugs to suppress malaria
(Kakkilaya 2006). WHO responds to dengue in the following ways: supports countries in the
confirmation of outbreaks through its collaborating network of laboratories; provides
technical support and guidance to countries for the effective management of dengue
outbreaks; provides training on clinical management, diagnosis and vector control at the
regional level with some of its collaborating centers; formulates evidence-based strategies
and policies; develops new tools, including insecticide products and application technologies;
gathers official records of dengue and severe dengue from over 100 Member States and
publishes guidelines and handbooks for dengue prevention and control for Member States
WHO, 2012). The Joint WHO/UNICEF Technical Group on Immunization in Africa
recommended in 1988 incorporation of yellow fever vaccine in routine child immunization
programmes of countries at risk for yellow fever, and the World Bank’s 1993 World
Development Report also strongly endorsed adding yellow fever vaccine to the EPI of the at-
risk countries (Lhuillier et al., 1989 and WHO, 1998).Measures of yellow fever immunization
monitoring and fast-tracking of positive viral load cases has to be adhered to at the country
level (Roger et al., 2009 and Kemri, 2012).
2.5 Use of plants in mosquito control
The use of plant and plant-derived products to control pests in the developing world is
well known and prior to the discovery of synthetic pesticides, plant or plant-based products
were the only pest-managing agents available to farmers around the world. There are repots
of medicinal plants with insecticidal including antifeedant, larvicidal, ovicidal and repellence
activities against mosquitoes. Such plants include those of Annonaceae, Papiloinaceae,
Meliaceae, Mimosaceae and Lamiaceae genera (Kalyana et al, 1985). The pool of plants
possessing insecticidal substances is enormous. These have generated extraordinary interest
in recent years, as potential sources of natural insect control agents. Today over 2000 species
of plants are known that possess some insecticidal activity (Jacobson, 1975, 1989).
17

Biochemical pesticides include plant-derived pesticides (botanicals) that can interfere with
the growth, feeding or reproduction of pests or insect pheromones applied for mating
disruption, monitoring or attract-and-kill strategies. Antioviposition and insecticidal activity
of Imperta cylindrical (Graminaceae) plant in Ethiopia has been documented Mohsen and et
al., 1995) Insect vector management is facing the economic and ecological challenge
worldwide due to the human and environmental hazards caused by majority of the synthetic
insecticide chemicals (FAO, 1992).
Identification of novel effective insecticidal compounds is essential to combat
increasing resistance rates that have been associated with synthetic convectional insecticides.
Plant extracts have been found to be advantageous for use in field mosquito programs (Abebe
and Ayehu, 1993). Secondary metabolites present in plants apparently function as defense
(toxic), which inhibits reproduction and other processes (Rattan, 2010). The botanical
insecticides are generally pest-specific and are relatively harmless to non-target organisms
including man. They are also biodegradable and harmless to the environment. Furthermore,
unlike conventional insecticides which are based on a single active ingredient, plant derived
insecticides comprise an array of chemical compounds which act concertedly on both
behavioral and physiological processes (Adeyemi, 2010). Thus the chances of insecticides
developing resistance to such bio insecticides are unlikely. Among the phytochemicals found
to have insecticidal potential are the triterpenoids, diterpenes, monoterpenes, alkaloids,
flavonoids and steroidal saponins (Isman and (Machial, 2006). Use of botanical involves the
development of the green technology using oil-in-water micro-emulsions as a nano-
insecticide delivery system to replace the traditional emulsifiable concentrated oil. This in
essence reduces organic solvents and increases the dispersity, wettability and penetration
properties of the droplets (Koul et al 2008). The antifeedant behavior of the plant
triterpenoids secondary metabolites prevent the insects from feeding but do not immediately
kill them hence the effective results can be accomplished by integrating them with more
creative strategies. Such a strategy can be incorporating them with insect growth regulators
(Adeyemi, 2010). Larvicidal activity of the leaf plant extracts of Paullina pinnata Linn
against An.gambiae has been attributed to the presence of alkaloids, tannins and saponins in it
Jaiyesimi et al., 2011. The plant extracts of Hemidesmus indicus, Gymnema sylvestre and
Eclipta prostrate have been found to have larvicidal potential against Culex quinquefasciatus
mosquito larvae (Gopiesh et al., 2007). Phytochemical constituents of Ethiopian medicinal
plants were screened and found to have larvicidal activity against Ae. aegypti, Ae. africanas
18

and Culex quinqufasciatus third and fourth instar larvae (Asfaw et al., 2007). Essential oils
from Striga hermothica, Hyptis spicigera and Ocimum basilicum leaf extracts have been
found to have mosquito repellency activity against An. gambiae and Culex quinquefastus
(Gabi et al 2012).The use of plant secondary metabolites in the fight against the diseases
vectored by mosquitoes will be more cost effective compared to the synthetic derived
chemical that has to be imported hence more uneconomical (Adeyemi, 2010).
2.6 Mode of Action of Insecticides
To understand the mode of action of insecticide, it is necessary to understand how the
targeted systems in pests normally function. This would help prevent development of
pesticide resistance in the target pests. Using pesticides with similar mode of action,
contributes to the problem of killing susceptible pests and leaving only those with resistance
to the entire class of pesticides. Development of pest resistance can be avoided or delayed by
rotating pest control chemicals that work through different modes of action (Amy, 2006).
Insecticides and miticides generally target the nervous system, growth and development or
energy production of the pest. Both human and insects have many different neurotransmitters
that work at different sites through the nervous system. Of the many neurotransmitters that
both insects and humans have, acetylcholine (ACh) and gamma-amino butyric acid (GABA)
are important targets of some insecticides. Acetylcholine can either excite or inhibit its target
neurons depending on the particular neuron and the receptors at the receptor sites.
Acetylcholine can cause particular neurons to “fire” continuing the nerve impulse
transmission, or it can cause the nerve impulse to stop at that particular site. In contrast,
GABA is an inhibitory neurotransmitter. When GABA is the neurotransmitter activated at the
synapse, the nerve impulse stops. Some insecticides interfere with the normal action of these
neurotransmitters. Organophosphate and carbamate insecticides are known as cholinesterase
inhibitors (Amy, 2006). They bind to the enzyme that is normally responsible for breaking
down acetylcholine after it has carried its message across the synapse. This causes over
stimulation of the nervous system and the insect dies. Although cholinesterase inhibition by
carbamates can be reversible, organophosphate poisoning is irreversible. Electrically charged
ions move along neurons in channels which include the sodium, calcium, potassium and
chloride channels. Many of the channels have gates that open or close in response to a certain
stimulus, which is an important mechanism through which some insecticides work.
Avermectins derived from a soil microorganism act by binding to the chloride channel
causing an inhibitory effect that causes the death of the pest. Organochlorine insecticides of
19

the cyclodiene type affect the chloride channel by inhibiting the GABA receptor, (Dong,
1997). Pyrethrins, which are naturally-occurring compounds derived from the
Chrysanthemum family are sodium channel modulators. They have a quick knockdown effect
against insects, but are unstable in the environment and therefore they do not last long enough
to kill the pest.
Pyrethroids are synthetic versions of pyrethrins and act in tiny channels through
which sodium is pumped to cause excitation of neurons, resulting in continued nerve impulse
transmission, tremors and eventually death (Amy et al; 2005). Bacillus thuringiensis (Bt)
which produces crystalline inclusions composed of endotoxins, referred to as crytoxin Bt
crystals, and must be ingested in the alkaline insect midgut, where they dissolve into
protoxins, which are cleaved by proteases into active toxins. When Bt is eaten by a larva, it
attacks the lining of the insect’s midgut and causes it to stop feeding and die. Some
insecticides such as organotin miticide inhibit oxidative phosphorylation directly, while
pyrroles work by uncoupling oxidative phosphorylation from electron transport. In either
case, the cell is unable to produce ATP as a source of its energy. Studies of the biophysics
and biochemistry of receptors and enzymes associated with insecticide targets elucidates
potential binding sites and illustrate that these proteins are dynamic molecules that interact in
various conformation with their antagonists and agonists (Amy,2006). Some insecticides
target the insect growth and development processes by interfering with hormones and others
by blocking the production of structural components of the exoskeleton. Juvenile hormone
(JH) analogue insecticides act as JH agonists and generally show the highest toxicity when
applied at the onset of metamorphosis (Thomas, 2004). A physiological basis for the toxicity
and morphogenetic effects of these insecticides has been suggested by linking their effects
with interference of the expression or action of certain genes especially the Broad- complex
(BR-C) transcript gene. Azadirachtin, derived from neem oil, interferes with the insect
development hormone, prothoracicotromic hormone. The active principle from the seed
kernel, the tetranortriterpenoid (limonoid), has the insecticidal effect (Tomlin, 2003). There
are numerous methods for studying gene expression which include the substractive reverse
transcriptase chain reaction (RT-PCR) of pests exposed to sub lethal doses of the insecticides.
Because of the potential effect of insecticides on the expression of certain genes, it is
important to establish the mode of action of insecticides by evaluating gene expression
profiles
20

2.7 Vector Resistance to Insecticides
A mechanism of pest resistance to insecticides is either through mutation within the
target site of the insecticides or alteration in the rate of insecticide detoxification. Three
enzyme systems: glutathione s-transferase, esterases and monooxygenases are involved in the
detoxification of the major insecticide classes. These enzymes act rapidly, metabolizing the
insecticides to non-toxic products or turning over the insecticide (sequestration). Gene
splicing, amplification and regulation plays a major role in mosquito insecticides (Chouaibou
et al., 2008) Replacement of alanine 302 in the RDL containing GABA receptor not only
interacts directly with the drug binding site within the ion channel pore but also allosterically
destabilizes the preferred desensitized state of the insecticide receptor. In the para gated
sodium channel, individual mutations in different channel polypeptides combine to cause
mutations similar to those found in the same polypeptide, suggesting that each mutation plays
a unique role in affecting channel function (Dong, 1997). In the para-voltage gated channel,
amino acid replacements are confined to only two positions (Guerrero et al., 1997). The first,
associated with the original kdr strain, is in S6 hydrophobic segment of homology domain II,
and the second is associated with another more resistant allele, termed-super-Kdr. This “kdr”-
like replacement is similar in the housefly, horn fly, cockroach the tobacco budworm
Helothis virescens, the aphid and the mosquito An. gambiae (Amy et al; 2005). Acetyl
cholinesterase encoding gene Ace, undergoes a limited subset of amino acid replacement
(Weill et al., 2000 and Weill et al., 2004). This causes insecticidal resistance despite the
widely differing sizes and structure of different organophosphate and carbamate insecticides
(Amy, 2006). Resistance to dieldrin and fibronil has been associated with chromosome
inversion 2La in An. gambiae. Pyrethroid insecticides’ resistance has also been reported in
Anopheles funestus in South Africa (Hargreaves et al., 2003).
Since the kdr mutation was first detected in An. gambiae from Africa in 1998, molecular
monitoring has been intensified world over. This mutation was found exclusively in the S
molecular form of the An. gambiae “group” in West and Central Africa (Weill, 2004). The
mutation was also found in Southern Benin, Cote-d’ Ivoire and Burkina Faso in the M
molecular form. A different Kdr type mutation (Leucine-phenylalanine) has been found in
An. gambiae in Western Kenya. Metabolic pyrethroid resistance has been found in Anopheles
sundaicus found in Southern Vietnam, a vector of local importance in this area (WHO, 2006).
A mutation confirming resistance to carbamates and organophosphates has been detected in
West Africa in An. gambiae. From the forgoing literature, management of resistance to
21

insecticides by insects relies on proper monitoring of the resistance. This activity has to be
integrated as a component of any malaria vector control program.
2.8 Characteristics of Aloe species
There has been a concerted effort worldwide to screen traditional medicinal plants for
their insecticidal potential (Asfaw et al., 2007). The active constituents have been found to be
antiprotozoal, antifungal, antibacterial, antinematode and effect on behavioral and
physiological responses on target organisms (Adeyemi, 2010). The Aloe species has active
phytochemical ingredients that need to be investigated for their potential use in biprospecting
for new botanicals in mosquito control measures (Isman and Machial, 2006). Aloe, in the
family of Aloaceae , is composed of herbaceous, shrubby or perennial, xerophytic, succulent
plants found in tropics and Southern African regions (Van Damme, 1991). The genus Aloe
has nearly 400 species confined to Africa. It is common in Kenya with about 60 taxa
recognized. Most of the taxa have a restricted distribution with Aloe secundiflora var
secundiflora being widespread in the country (Baillie et al., 2004). Aloe vera is one of the
species which originated from Africa and is a plant of dry sandy places and hilly sides. Its
fleshy cactus like appearance distinguishes from other herbs (Beentje and Smith; 2001). The
diversity pattern in Kenya indicates a high concentration of Aloe in three areas that are known
as Aloe hot spots and thus of high priority for conservation of the genus. These areas are; the
Kulal-Nyiro-Ndotos-Marsabit areas in the north, the Taita-Shimba Hills zone to the South-
East and the Naivasha-Baringo area in the Rift Valley. A. turkanensis occurs in north western
Kenya and in the Karamajong District of Uganda. Aloe turkanensis (Figure 6) grow on stony,
sandy soil or lava/ usually in the shades of shrubs in arid areas at 600-1,250m altitude
(Wabuyele et al., 2006). Aloe plant species have been used all over the world for their various
medicinal properties. Treatment of streptocin induced-diabetic rats with Aloe barbadensis (A.
vera), increases antioxidant enzymes like superoxide dismutase and it also decreases lipid
peroxidation product levels (Nwanjo, 2006). Cinnamoyl, P-coumaroyl, feruloyl, caffeoyl,
aloesin and related compounds have been isolated from Aloe species. The anti-inflammatory
and antioxidative activities of these compounds have been examined based on the structure-
activity relationship (Akira, 2008).
Investigation using the contact hypersensitivity response indicates a preventive effect
of aloesin on the UV-B-Induced immune suppression (Chaz, 2008). Aloesin inhibits tyrosine
hydroxylase and dihydrophenylalanine (DOPA) oxidase activities of tyrosinase from normal
human melanocyte cell lysates. This makes aloesin a positive pigment-altering agent for
22

cosmetic application (Yagi and Takeo, 2003). The fact that Aloe vera can alter skin
pigmentation points to the fact that its constituents could have physiological and physical
effect on target organisms, including mosquitoes (Adeyemi, 2010).
Figure 6: Aloe turkanensis plant prevalent in North Western Kenya and the
Karamajong District of Uganda (Bosch, 2006).
Neutral polysaccharides such as aloemannan and acemannan have shown anti tumor,
anti-inflammatory and immunosuppressive activities (Singh et al., 2000). Glycoprotein
fractions with body kinindegrading and cell proliferation-stimulating activities have been
identified from the nondialysate fraction of the gel of Aloe species (Tan and Vanitha 2004).
Aloesin has also been found to stimulate the proliferation of cultured human hepatoma SK-
Hep 1 cells, with other related and high molecular-weight materials such as verectin and
aloemannan could exert therapeutic properties for wounds, burns and inflammation
23

(Yakugaku, 2003). Aloe plant species have been considered to have a hypoglycemic effect
and many diabetic patients take the gel because of its hypoglycemic effect (Tiwari and Rao,
2002). Aloe vera, one of the most important species of Aloe is considered to have
hypotensive, hepatoprotective and blood purification property (Nwanjo, 2006). Chaz, (2008)
asserts that Aloe plant species have the property to boost the immune system, avert allergies,
sinusitis and bronchitis. The polysaccharides in Aloe species lowers down the body’s serum
lipids by lowering triglycerides, low-density lipoprotein levels and increasing high-density
lipoproteins (Yagi; Takeo, 2003). Several compounds from the Aloe –based alcoholic
beverages have been separated and identified (Tawfik et al., 2001). Among the noticeable
compounds include; aloesin, aloeresin A, hydroxyaloin, aloin A and B and aloinoside A and
B. Among other constituents isolated from Aloe vera, phenolic compounds containing
alkaloidal groups have insecticidal activities (Tan and Vanitha, 2004).
2.8 Aloe species Phytochemicals
The chemical composition of Aloe leave exudates has been investigated (Nauwinger,
2000). The compounds that have been identified can be classified into two main groups
namely chromones and anthraquinones. In some cases both types are present, and in others
only one. Many of the major constituents such as isoaloesin (1) aloesin (2) and aloin (3) of
Figure 7 occur in chemo-taxonomically distinct species. A. vera’s mucilaginous pulp
contains 93-96% water and the rest are solids that contain active compounds for its biological
activity. The mucilage of the leaves also contain various sugars and amino acids.
24

OO
CH3 O
CH3Glu
OHO
O
CH3 CH2
CH3
OH
Glu
1. Isoaloesin 2. Aloesin
Glu
OH
OH O OH3. Aloin
Figure 7: The chemical formulae of the phytochemicals found to be prevalent in the
Aloe plant species (Dagne et al., 1996)
2.9 Phytochemistry and plant pharmacology screened medicinal plants
Plants are nature’s chemical factories" providing the richest source of organic
chemicals on earth. The use of plants , whether herbs, shrubs or trees in parts or in whole
treatment management of diseases and disorders date back to pre-historic days (Egwaikhide,
2007). The detection of active principles in medicinal plants plays a strategic role in
phytochemical investigation of crude plant extracts and is very important in regards to their
potential pharmacological effects (Majaw and Moirangthem, 2009). The medicinal value of
these plants lies in some chemical substances that produce a definite physiological action on
the human body. The most important of these bioactive constituents of plants are alkaloids,
tannins, flavanoids, and phenolic compounds. (Edoega et al., 2005). In the recent years the
emphasis to control the mosquito populations has shifted from the use of convectional
chemicals towards more specific and environmentally friendly materials, which are generally
of botanical origin (Matasyoh et al., 2008). For this reason a lot of phytochemicals extracted
from various plant species have been tested for theirs larvicidal and repellent actions against
mosquitoes (Ciccia et al., 2000)
25

Terpenoids exert their effect as anti- bacteria, anti-fungi, anti-viral, anti- protozoan,
anti-allergens and anti-neoplastic agents (Maiyo et al., 2010). Membrane disruption may be
the mode of action employed by terpenoids on target organisms. Flavanoids are structural
derivatives of flavones. They contain conjugated aromatic systems often bound to sugar (s) as
glycosides. Flavanoids are water soluble. Flavanoids act as anti- microbials by complexing
extracellular proteins, soluble proteins and bacteria cell wall. Terpenoids form a large and
structurally diverse family of natural products derived C5 isoprene units. Typical structures
contain carbon skeletons represented by (C5)n and are classified as hemiterpenes (C5),
monoterpenes (C10), sesquiterpenes (C15), diterpenes (C20), sesterpenes (C25), triterpene (C30)
and tetrapenes (C40) (Rohmer M. 1999). Terpenoids exert their anti- bacteria, anti-fungi, anti-
viral, anti- protozoan, anti-allergens and anti-neoplastic agents (Maiyo et al., 2010).
Membrane disruption may be the mode of action employed by terpenoids on target organisms
and microbial cell membranes (Gadelha et al., 2005). Saponins are surface active agents.
They have soapy like properties. They can be detected by their ability to cause foaming and
to haemolyse red blood cells (Harboune, 1991). Steroidal saponins have been known to have
anti-protozoa activity by reacting with cholesterol in the protozoal cell membrane
(McAllister, 2001). Aloe plants from arid area of South Africa were found to contain
secondary phenol metabolites (Gutterman and Chausen, 2007). Phenolic compounds have
been found to have allelopathic potential on other vegetation (Anne et al., 2005). Tannins are
astringent, bitter plant polyphenols. They bind, precipitate or shrink proteins (Amy, 2006)
26

CHAPTER THREE
MATERIALS AND METHODS
3.1Sampling of test Aloe species
Aloe plant species A. turkanensis, A. ngongensis and A. fibrosa were collected from
Turkana, Ngong and Kajiado regions of Kenya respectively. With the assistance of a
taxonomist, plant materials were identified and a voucher specimen was deposited at the
Department of Biological Sciences of Egerton University herbarium. The Aloe species were
designated labels and further identification of the samples subjected to the sequential organic
solvents was carried out:
Aloe species Voucher specimen
Identification Code
Solvent of Extraction Laboratory
Identification Code
A. turkanensis AT Ethyl acetate ATE
A. ngongensis AN Ethyl acetate ANE
A. ngongensis AN Hexane ANH
A. ngongensis AN Chloroform ANC
A. ngongensis AN Acetone ANA
A. ngongensis AN Methanol ANM
A. fibrosa AF Hexane AFH
A. fibrosa AF Acetone AFA
A. fibrosa AF Methanol AFM
3.2 Test Insect
Mosquito colony of An. gambiae s.s. was obtained from the Kenya Medical Research
Institute (KEMRI), Kisumu, Kenya. This colony was originally collected from Mbita field
station (00025'S, 34013'E), South Nyanza, Kenya in December, 2000 where An. gambiae s.s.
are abundant. At the time of this work, the colony was in the 35 th filial generation post field
sampling, away from any possible selection pressure by Aloe plant species extracts. A
laboratory mosquito colony of Ae. aegypti was also obtained from Pyrethrum Board of
Kenya, Nakuru that had not been under any possible selection pressure by Aloe plant species
27

3.2.1 Rearing and maintenance of mosquito colonies
The insectary strain of An. gambiae s.s. was reared in KEMRI insectary, Kisumu, in the
Republic of Kenya. The Ae. aegypti insectary strain was reared at the Pyrethrum Board of
Kenya insectary. The standard procedure for rearing Anopheles and Culicine mosquitoes was
followed (Ford and Green, 1972 and WHO, 2005). All life stages were reared in the insectary
(28 ± 2°C, 75 – 80 % Relative Humidity and 12L: 12D photoperiod) of Animal Rearing and
Quarantine Unit (ARQU) of KEMRI Kisumu, Kenya and the Pyrethrum board of Kenya,
Nakuru. From the day of emergence, adult mosquitoes were provided with 10% sugar
solutions soaked in cotton wools. The adult mosquitoes were sampled using an aspirator
connected to vacuum Three-day-old female mosquitoes were allowed to feed on
anaesthetized mice. The female mosquitoes were singled out due to their conspicuous thinly
haired antennae and forwardly projecting proboscis, as opposed to the males that have bushy
antennae. Approximately 2-3 days later, oviposition dishes were placed in the cages
containing gravid females. The eggs were placed on distilled water and were surrounded with
floating wax papers, which served to keep eggs from becoming stranded on the sides of the
hatching trays. Approximately 30 mg pulverized Tetramin fish food (Tetra GmbH, Melle,
Germany) per pan were sprinkled on the surface of the water twice daily. Pupae were
collected daily and transferred to breed cap bowls containing distilled water and placed inside
emergent cages covered with nets. A small moist filter paper wrapped in a conical shape is
put in the breed cap bowl containing distilled water and kept inside the cage overnight for the
mosquito to lay eggs. The bowls were placed in emergent cages for adult emergence.
3.3 Aloe leaf homogenization and subsequent solvent extraction of phytochemicals from
plant material
The leaf plant materials were cut into small pieces, and macerated in a blender. The
homogenates were extracted sequentially with solvents of increasing polarity as follows:
hexane, chloroform, ethyl acetate, acetone and methanol. The extracts were filtered through
active charcoal to remove the chlorophyll matter. The solvents were removed by roter
evaporation under pressure to give five extracts for each species. The yields were noted for
each extract.
3.4 Larvicidal tests
Bioassay was performed according to WHO procedure (WHO, 2005). Third instar larvae of
An. gambiae was sourced from Kenya Medical Research Institute laboratories (KEMRI)
Kisumu and bioassay in response to Aloe extracts carried out in their laboratories. The Ae.
28

aegypti were sourced from the Pyrethrum Board of Kenya where the bioassay to Aloe plant
extracts was conducted. Minimum and maximum ranges were determined by carrying out
bioassays at concentrations that could kill 10% and 95% of the third instar larvae
respectively. The Aloe plant extracts were solubilized in analytical grade dimethyl sulphoxide
(DMSO) and distilled water added to give serial dilution concentrations. Twenty larvae were
placed into each 50 ml disposable cups containing 15 ml of Aloe plant extract solution.
Larvae were fed with tetramin fish food during all testing. Serial dilutions of Aloe plant
extracts ranging from 0.05-2 mg/ ml were made. Three replicates were used for each
concentration level. Control test (negative control) was carried out in parallel using DMSO
and water for comparison. DMSO was kept at a concentration of not more than 1%. A
conventional insecticide (pyrethrum based larvicide, pylarvex) was used as a positive control
sample at 0.05 mg / ml. Larvae mortality in treated and control cups was recorded after 24
hours exposure. The dead larvae were counted and the median lethal concentrations (LC50)
determined by probit analysis (Finney 1971). The dead larvae in the four replicates were
pooled and expressed as percentage mortality for each concentration. Observation was made
on the behavior of the larvae to ascertain mortality by gently pricking them using end of a
pipette to provoke arousal.
3.5 Physiological Responses
3.5.1 Selection for Aloe species tolerance
Susceptible strains of third instar Ae. aegypti larvae were separately placed under selection
pressure with the extracts from Aloe plant species at concentrations that caused 50%
mortality (LC50). Aloe plant samples in which survivors failed to emerge as adults in
sufficient numbers were discarded. One thousand five hundred larvae were selected for the
parental and first generations and type of Aloe plant extract (n = 3) in three replicates each
consisting of 1500 larvae in 1500 ml water in polypropylene cylindrical pans with radius and
height of 10.5, 24.14 cm respectively. The susceptibility levels of Ae. aegypti to each Aloe
plant extract in successive generations were monitored by determining the LC50 values. A
control colony was reared simultaneously in a separate room and handled in the same manner
through all manipulations but was not exposed to Aloe plant extracts. The larvae were not fed
during the 24 h exposure. Survivors were normally propagated.
29

3.5.2 Physiological Resistance Diagnostic Test and Controls
Toxicity range tests (24 h) of Aloe plant extracts were conducted on each generation
using third instar An. gambiae s.s. and Ae. aegypti larvae. After determining the upper and
lower toxicity ranges of each Aloe plant extract, 24 h acute toxicity tests were conducted.
Three replicates (n = 25 per replicate) were exposed to five logarithmically separated Aloe
plant extract concentrations within the established toxicity response ranges (Finney 1971), in
400 ml of distilled water in the polypropylene cylindrical pans. Larval mortalities were
evaluated 24 h post exposure and LC50 determined by probit analysis (Finney 1971).
3.6 Phytochemical and functional group tests of the Aloe plant extracts
The Aloe plant extracts were assayed for the presence of the following phytochemical
groups: flavonoids, saponins, tannins and polyphenols, phlobatannins, steroids and
terpenoids. The Aloe plant extracts were also assayed for the presence of unsaturation,
alcohol groups, phenolic groups, aldehyde groups and carboxylic functional groups. The
phytochemical test for flavonoids was carried out by adding about 5 ml of dilute aqueous
ammonia was added to a portion of filtrate of the plant extract, followed by addition of
concentrated sulphuric acid. A positive test was confirmed by the formation of a yellow
coloration that disappeared instantly (Edeoga et al., 2005).Test for saponins was done by
adding three drops of olive oil was to 0.1 g of extract and shaking vigorously. Formation of
an emulsion indicated a positive test for saponins (Egwaikhide, 2007). Tannins and
polyphenols were tested for by adding few drops of 1% ferric chloride to 0.2 g of extract
previously dissolved in 10 ml of water. A brownish- green or blue- black coloration was
indicative of a positive test for either a polyphenol or tannin presence (Edeoga et al.,
2005).The test for phlobatannins was carried out by adding about 1ml of 1% ferric chloride to
0.1g of extract. Presence of depositions of a red precipitate was positive for phlobatannins
(Majaw and Moirangthem, 2009). To test for steroids, about 2 ml of acetic anhydride and 2ml
of sulphuric acid were added to 0.5g of extract. Change of color from violet to blue was
indicative of the presence of steroids (Egwaikhide, 2007). Lastly the terpenoids
phytochemical group was tested for by the Salkowski test where bout 3 ml of concentrated
sulphuric acid and 2 ml of chloroform were added to 5 ml of extract to form a layer. A red
coloration at the interface was indicative of the presence of terpenoids (Majaw and
Moirangthem, 2009). The functional groups in the Aloe plant extracts were assayed. Presence
of unsaturation was done by adding about 2ml of extract drop wise to 2ml of potassium
permanganate while shaking. Discoloration of the 1M potassium permanganate solution was
30

indicative of the presence of unsaturation in the extract. The dissolution of extract in a
solution of sodium hydroxide indicated presence of phenolic groups. The rate at which the
Aloe plant extracts reacted to form cloudiness with Luca’s reagent was indicative of the
presence of either the primary, secondary or tertiary alcohols. The presence of the aldehyde
functional group was tested by adding about 3-5drops of Tollen’s reagent to 2ml of plant
extract. Formation of a silver mirror indicated presence of aldehydes. (Harboune, 1991). The
presence of carboxylic acid group was assayed by addition of 3-5 drops of 1m sodium
hydrogen carbonate to 2 ml of sample extract. Solubility and effervescence of the sample was
confirmation of presence of carboxylic functional group (Harboune, 1991).
3.7 Data analysis
Acute mortality responses were corrected by Abbot’s formula (Busvine 1971) and
then transformed to Probits (Finney, 1971) for linear regression analyses and 50 % lethal
concentration (LC50) determination. Data sets with more than 10% control mortality were not
considered for analysis (Finney, 1971). Patterns of larval mortality, pupae formation and
mortality as well as male and female adult eclosion between the treatments were conducted
through cumulative frequency distribution. Probit analysis of concentration mortality data
was conducted to estimate the LC50, LC90 and LC99 values and associated 95 % confidence
limits. The effects of Aloe species selection on egg viability/hatchability, larval and pupa
mortalities, adult mortalities and female and male emergence were evaluated by one-way
analysis of variance (ANOVA) on three datasets. The 1% dimethyl sulphoxide (DMSO) and
distilled water were used as negative controls. Results that were significantly different were
analyzed using GENSTAT Version 10.s (Hemel Hempstead, GENSTAT software analysis
developer), (Jawara et al., 2009). The least significant difference was identified using Waller
– Duncan statistical procedures (Steel and Torrie, 1980).
31

CHAPTER FOUR
RESULTS
4.1 Larvicidal results
Larvicidal results for the preliminary assays of fifteen extracts from A. turkanensis, A.
ngongensis and A. fibrosa against third instar larvae of An. gambiae and Ae. aegypti showed
that only nine were active that could cause 60 % mortality at 2 mg/ml. Only the ethyl acetate
extract of Aloe turkanensis, hexane, ethyl acetate, acetone, chloroform and methanol extracts
of A. ngongensis and the hexane, acetone and methanol extracts of A.fibrosa showed activity.
The ethyl acetate soluble extract of A. turkanensis showed larvicidal activity where 99 %
mortality was achieved at a concentration of 0.25 mg/ ml within the lower and upper limits of
0.19-0.41 mg / ml respectively against third instar larvae of An. gambiae s.s (Table 1). In
comparison, A. turkanensis ethyl acetate plant extract exhibited 99 % mortality larvicidal
activity against third instar Ae. aegypti larvae, within the range of 0.25-0.56 mg / ml. upper
and lower limits with a median value of 0.33mg / ml (Table 2). The LC50 responses against
third instar larvae of An. gambiae s.s. and Ae. aegypti using the ethyl acetate extract of Aloe
turkanensis were 0.08 mg / ml and 0.11mg / ml, respectively. The A. ngongensis plant
extracts in acetone, ethyl acetate, hexane, and methanol and chloroform all showed larvicidal
activity against third instar larvae of the Anopheles and the Aedes mosquitoes species,
exhibiting LC50 values tabulated in tables 1 and 2. Among the assayed A. ngongensis plant
extracts, the hexane extract comparably showed larvicidal activity of LC50 concentration of
0.11 mg /ml against third instar larvae of Ae. aegypti mosquito species. Only the methanol,
acetone and hexane plant extracts of A. fibrosa species showed larvicidal activities at LC50
concentrations ranging between 0.66-3.90 mg / ml against third instar larvae of An. gambiae
and Ae. aegypti mosquito species.
32

Table 1
Probit analysis results for the concentrations (mg/ml) of selected Aloe plant extracts against An. gambiae s. s. third instar larvae
Aloe species Extraction solvent LC50 (95CL) LC90 (95CL) LC99 (LC95) Slope (β ± SE) ; χ2≠
A.turkanensis Ethyl acetate 0.08 (0.07-0.1) 0.15 (0.13-0.21) 0.25 (0.19-0.41) 4.89 ±0.78 1.24
A.ngongensis Ethyl acetate 0.66(0.52-0.84) 2.11(1.49-3.90) 5.45(3.15-15.24) 2.5±0.41 0.31
A.ngongensis Hexane 0.52(0.41-0.65) 1.54(1.11-2.72) 3.75(2.24-9.79) 2.7±0.44 0.36
A.ngongensis Chloroform 0.81(0.65-1.03) 3.51(2.27-8.33) 6.43(3.59-21.07) 2.5±0.47 1.21
A.ngongensis Acetone 1.21(0.98-1.55) 3.65(2.53-7.41) 8.91(5.01-28.90) 2.68±0.48 0.95
A.ngongensis Methanol 2.67(2.40-3.17) 4.92(3.90-8.18) 8.04(5.59-18.37) 4.8±0.10 3.44
A. fibrosa Methanol 2.74(1.9-5.53) 14.64(6.68-128.69) 57.32(17.04-1787.74) 1.76±0.44 0.23
A.fibrosa Acetone 2.70(2.41-3.15) 4.99(3.97-8.18) 8.23(5.74-18.58) 4.80±0.97 0.80
A.fibrosa Hexane 2.13(1.84-2.57) 4.62(3.50-8.21) 8.66(5.60-22.34) 2.82±0.73 0.42
95 CI = 95% confidence interval;
SE = standard error,
LC 50=lethal concentration of Aloe plant extract that kills 50% of the An. gambiae larvae
LC 90=lethal concentration of Aloe plant extract that kills 90% of the An. gambiae larvae
LC 99=lethal concentration of Aloe plant that kills 99% of the An. gambiae larvae
χ2≠ = Greek small letter Chi squared
β ± is the Greek sign beta-plus or minus.
33

Table 2
Probit analysis results for concentrations (mg/ml) of selected Aloe plant extracts against Ae. aegypti third instar larvae
Aloe species Extraction solvent LC50 (95CL) LC90 (95CL) LC99 (LC95) Slope (β ± SE) ; χ2≠
A.turkanensis Ethyl acetate 0.11(0.09-0.12) 0.20(0.16-0.27) 0.33(0.25 0.56) 4.7±0.76 0.42
A.ngongensis Ethyl acetate 0.15(0.13-0.17) 0.32(0.25-0.5) 0.62(0.41-1.43) 3.7±0.70 1.21
A.ngongensis Hexane 0.11(0.08-0.15) 0.48(0.29-1.24) 1.67(0.76-7.94) 1.0±.0.34 1.03
A.ngongensis Chloroform 0.33(0.28-0.37) 0.62(0.52-0.86) 1.05(0.79-1.84) 4.58±0.76 3.02
A.ngongensis Acetone 0.77(0.67-0.89) 1.57(1.26-2.42) 2.82(1.97-5.82)4 4.11±0.73 0.09
A.ngongensis Methanol 0.39(0.34-0.45) 0.79(0.63-1.22) 1.42(0.99-2.95) 4.11±0.73 2.70
A. fibrosa Methanol 3.90(3.37-4.47) 7.84(6.36-11.54) 9.56(7.43-15.48) 4.23±0.73 1.95
A.fibrosa Acetone 0.66(0.53-1.02) 1.76(1.1-6.41) 3.94(1.91-29.60) 2.99±0.73 0.06
A.fibrosa Hexane 2.39(2.07-2.89) 5.15(4.60-13.06) 9.64(6.21-25.12) 3.84±0.75 1.18
95 CI = 95% confidence interval;
SE = standard error,
LC 50=lethal concentration of Aloe plant extract that kills 50% of the Ae. aegypti larvae
LC 90=lethal concentration of Aloe plant extract that kills 90% of the Ae. aegypti larvae
LC 99=lethal concentration of Aloe plant that kills 99% of the Ae. aegypti larvae
χ2≠ = Greek small letter Chi squared
β ± is the Greek sign beta-plus or minus.
34

4.2 Phytochemical qualitative analysis results of selected larvicidal Aloe plant extracts
The qualitative analysis of the ethyl acetate extract of A. turkanensis gave positive test
results for saponins, tannin and polyphenol test and also the flavanoids test (Appendices 19,
20 and 22). Apart from the hexane and chloroform extracts of A. ngongensis plant extract, the
others of methanol; acetone and ethyl acetate gave positive flavanoids results (Appendix 19).
All the A. fibrosa plant extracts exhibited presence of flavonoids (Table 3). Tannins were
present in all the analyzed Aloe plant extracts except those of the hexane, ethyl acetate and
the chloroform extracts of the A. ngongensis plant extract (Appendix 20). Apart from the
ethyl acetate and the chloroform extracts of A. ngongensis and also the hexane extract of A.
fibrosa all the other assayed Aloe plant extracts showed the presence of saponins (Appendix
22) .All the analyzed plant extracts exhibited absence of steroid, phlobatannins and terpenoid
phytochemicals ( Appendices 17, 18 and 21).
35

Table 3
Summary of the qualitative phytochemical constituents of the selected Aloe plant extracts
Aloe species
Solvent of extraction Terpenoid test Steroid test Flavonoid test Saponin test Tannin/Polyphenol test
Phlobatannin test
ATE Ethyl acetate - - + + + -
ANE Ethyl acetate - - + - - -
ANA Acetone - - + + + -
ANC Chloroform - - - - - -
ANH Hexane - - - + - -
ANM Methanol - - + + + -
AFA Acetone - - + + + -
AFM Methanol - - + + + -
AFH Hexane - - + - + -
(+) Positive for test reaction(-) Negative for test reactionATE Aloe turkanensis ethyl acetate plant extractANE Aloe ngongensis ethyl acetate plant extractANA Aloe ngongensis acetone plant extractANC Aloe ngongensis chloroform plant extractANH Aloe ngongensis hexane plant extractANM Aloe ngongensis methanol plant extractAFA Aloe fibrosa acetone plant extractAFM Aloe fibrosa methanol plant extractAFH Aloe fibrosa hexane plant extract
36

4.3 Analysis of chemical functional groups in Aloe extracts
All the Aloe plant extracts analyzed exhibited absence of unsaturated bonds in their
structures .Only the ethyl acetate plant extract of A. ngongensis showed presence of tertiary
alcohols while the A. fibrosa acetone extract showed presence of secondary alcohol
functional group. The other Aloe plant extracts showed possibility of presence of primary
alcohols. Apart from the chloroform plant extract of A. ngongensis, all the analyzed plant
extracts showed presence of the phenolic radical. Also predominantly present in all the
assayed Aloe plant extracts was the aldehyde functional group whose presence was detected
by the reduction of ionized silver to elemental silvery mirror. All larvicidal Aloe plant
extracts showed presence of carboxylic acid functional group apart from the chloroform
extract of A. ngongensis plant. The analysis of the functional groups in the assayed Aloe plant
extracts is illustrated in Table 4.
37

Table 4
Aloe species functional groups
Aloe plant extract Solvent of extraction
Unsaturation test
Carboxylic group test
Alcohol group test Aldehyde group test
Phenol group test
ATE Ethyl acetate - + - + +
ANE Ethyl acetate - + Tertiary alcohol + +
ANA Acetone - + - + +
ANC Chloroform - - - + -
ANH Hexane - + - + +
ANM Methanol - + + +
AFA Acetone - + Secondary alcohol + +
AFM - - + - + +
AFH - - + - + +
(+ ) Positive for reaction test(-) Negative for reaction testATE Aloe turkanensis ethyl acetate plant extractANE Aloe ngongensis ethyl acetate plant extractANA Aloe ngongensis acetone plant extractANC Aloe ngongensis chloroform plant extractANH Aloe ngongensis hexane plant extractANM Aloe ngongensis methanol plant extractAFA Aloe fibrosa acetone plant extractAFM Aloe fibrosa methanol plant extractAFH Aloe fibrosa hexane plant extract
38

4.4 The effect of Aloe plant extracts on Ae. aegypti
The effects of the Aloe species plant extracts on the egg, larvae, pupa and adult stages
of the mosquito Ae. aegypti are summarized in Table 5 and Appendices 1 to 16. During this
study, both the parental (F0) and the first generation (F1) Aloe exposed groups were
adversely affected in their developmental stages (Figures 7 and 8). While the mean larval
mortality in the control groups in F0 was 2.67% and 9.33% in distilled water and DMSO
respectively, the Aloe plant exposed larvae exhibited mortalities of 48.60%, 51.55% and
51.55% with LSD of 62.5 and coefficient of variation of 7.1% with F value of ˂ 0.01.The
mean larval mortality in the F1 A. ngongensis ethyl acetate plant extract exposed larvae was
14.38%compared to 2.9% in the untreated control(Appendix 1) While the pupae mortality in
the controls was between 1.03% and 2.13%, that in the Aloe exposed group ranged between
12.12% and 13.10% in F0 generation (Appendix 2). The ability of the Aloe exposed
mosquitoes to lay eggs was equally hampered as evidenced in table 3 (there were strong
negative correlations with egg deposits of 4000 per treatment and fecundity of 133
(Appendices 6 and 16)). In comparison the water and DMSO control mosquitoes deposited
7333 and 6030 eggs respectively (hence fecundity of 244. and 201).
The egg viability in the F1 generation group exposed to Aloe plant extracts was
adversely affected ranging between 19.8% and 49.3% of the possible 1500 exposed eggs.
This is in contrast to the control group which had more eggs hatching of up to 94.6% and
96.7% out of the batch of 1500.eggs (Appendix 10). The age specific studies on the 28th day
revealed there were between 14 and 16 males in the control group as opposed to greatly
reduced numbers in the Aloe plant exposed mosquitoes (averagely 2-6 (Appendices 7, 8 and
9). On the same day the females in the control group were between 24 and 25 in contrast with
the Aloe exposed female mosquitoes that were between 7 and 22 out of the original number
of 30. Consequently the ratio of surviving males and females on the 28 th day was 0.47 and
0.44 in the water and DMSO control environment mosquitoes. This is opposed to the lower
ratios of 0.35, 0.27 and 0.20 in the respective Aloe exposed mosquito batches (Appendix 8).
Also profoundly affected were the rates of both the female and male mosquito emergence
(53-224). This is in contrast to the Aloe unexposed mosquitoes that showed male and female
emergence of between 647 and 628 from the possible exposed 1500 egg batch (Appendices 3
and 4).
39

Table 5
The effect of Aloe plant extracts on 1500 third instar larvae of Ae. aegypti parental generation (F0) and first generation (F1) and the subsequent developmental stages
ATTRIBUTES CONTROL 1%DMSO AFA ANE ATE LSD F %CV
F0 larvae mortality 40.0a 140.0b 729.0c 773.3c 779.7c 62.5 <.001 7.1
F1 larvae mortality 44.0a 143.0b 183.7c 215.7d 145.0b 21.3 <.001 7.7
F0 Pupae mortality 15.3a 26.7a 88.7b 92.0b 101.3b 28.4 <.001 23.3
F1 Pupae mortality 28.3a 25.3a 79.3b 61.3c 24.3a 13.3 <.001 16.1
F0 egg deposits 7394.0a 6030.0b 4357.0c 4210.0c 4181.0c 775. <.001 5.5
F1 Egg deposits 7033.0a 7615.0b 7083.0c 4948.0d 4597.0d 416.4 <.001 3.5
F1 Egg viability 1453.0a 1419.0a 749.0b 700.0b 297.0c 79.3 <.001 4.6
F0 ♂ Age specific 16.7a 14.7a 6.33b 3.33b 2.33b 5.2 <.001 11.6
F0 ♀ Age specific 25.7a 24.3a 21.7a 10.8b 7.8b 5.4 <.011 15.8
Survivorship ♂ and ♀ 0.47a 0.44a 0.35b 0.27c 0.20d 0.06 <.001 82
FI ♂ emergence 647.0a 628.3b 224.3c 193.7d 62.0e 12.2 <.001 1.8
F1♀emergence 624.3a 607.7b 220.0c 186.7d 53.3e 13.2 <.001 2.1LSD = Least Significant Difference of means (5% level)CV = coefficient of variationDMSO = 1% solution of dimethyl sulphoxideATE = Ethyl acetate plant extract of Aloe turkanensis ANE = Ethyl acetate plant extract of Aloe ngongensisAFA = Acetone plant extract of Aloe fibrosaF0 and F1 = parental and first filial generations respectivelyThe symbols ♂ and♀ mean the male and the female mosquitoes respectivelyThe means with the same letter superscript on the same row are not significantly different ((P<.0.05)
40

Control 1% DMSO 1% Ethyl acetate
AFA ANE ATE0
10
20
30
40
50
60
70
80
90
100
DeadAlive
Figure 8: The responses of third instar larvae post 24 hour exposure to selected Aloe
plant extracts and the negative controls
(a) Control= Distilled water(b) 1% DMSO= 1% Dimethyl sulphoxide solution(c) 1% ethyl acetate =1% ethyl acetate solution(d) ANE= LC50 Aloe ngongensis ethyl acetate plant extract(e) ATE= LC50 Aloe turkanensis ethyl acetate plant extract(f) AFA=LC50 Aloe fibrosa plant extract(g) Dead larvae(h) Surviving larvae
41

Control1% DMSO
ANEATE
AFA
0.00%
10.00%
20.00%
30.00%
40.00%
50.00%
60.00%
70.00%
80.00%
90.00%
100.00%
Mortality Survivorship
Figure 9: The effect of Aloe plant extracts on the pupae of Ae. aegypti
(a) Control= Distilled water(b) 1% DMSO= 1% Dimethyl sulphoxide solution(c) ANE= LC50 Aloe ngongensis ethyl acetate plant extract(d) ATE= LC50 Aloe turkanensis ethyl acetate plant extract(e) AFA=LC50 Aloe fibrosa plant extract(f) Dead pupae(g) Surviving pupae
42

CHAPTER FIVE
DISCUSSION, CONCLUSION AND RECOMMENDATION
5.1 Discussion
This study has confirmed that plant extracts can be used as larvicides and deterrents
against mosquitoes. The A. turkanensis leaf extract effected 99% mortality against An.
gambiae at 0.25mg/ml within the lower and upper ranges of 0.19mg/ml and 0.41mg/ml
respectively. This is comparable to the larvicidal effect of Lantana camara Linn against
mosquito species Ae. aegypti and Culex quinquefasciatus where 100% mortality was attained
at 1.0mg/ml and 3.0mg/ml respectively (Sathish and Maneemegalai, 2008) .There are reports
of medicinal plants with insecticidal activity including ovicidal, antifeedant, larvicidal and
repellent activities (Asfaw et al., 2007). Mosquito larvicidal and cytotoxic activities of three
Annona species against third instar Culex quinquefasciatus and brine shrimp larvae have been
reported (Magdula et al., 2009). Also the developmental stages of the bio assayed Ae. aegypti
were markedly affected by exposure to Aloe plant extracts. This observation correlates with
that of Imperta cylindrical (Graminaceae) which showed antioviposition and insecticidal
effects against mosquitoes (Mohsen et al., 1995). In this study flavonoids were present in
most of the assayed Aloe species.
Flavonoids have been found to have insecticidal activity against wooly apple aphid,
Eriosoma lanigerum (Mazen, 2012). Sathish and Maneemegalai (2008) found saponins,
flavonoids, terpenoids and cardiac glycosides as secondary metabolites present in plant
extracts with insecticidal potential. It is therefore hypothesized that larvicidal activity could
be due to the disruption of the larval cell membrane (Mayo et al., 2010). This is because the
mode action of flavonoids is by disrupting the cell membrane. That all the assayed Aloe
larvicidal extracts gave a positive Tollen,s test suggests presence of potentially free aldehyde
groups. This is collaborated by the fact that flavanoids are structural derivatives of flavones
containing conjugated aromatic systems often bound to sugar(s) as glycosides (Berenbaum
and Zangeri, 1987; Harboune, 1991). The toxicity of benzaldehyde, an aromatic aldehyde,
against the immature and adult stages of Ae. aegypti and Culex quinquefasciatus has been
reported (Paulraj et al., 2011). In this study the A. turkanensis ethyl acetate plant extract
exhibited the highest larvicidal activity against third instar larvae of Ae. aegypti and An.
gambiae. This extract exhibited presence of flavonoids, saponins and the tannins and
polyphenol group. Polyphenols have previously been isolated from A. vera leaf plant extracts
43

(Dagne et al., 1998).Screening of some Ethiopian medicinal plants with larvidal activity
revealed constituent presence of saponins, polyphenols, alkaloids and glycosides
All the three species of analyzed larvicidal Aloe plant extracts depicted absence of
terpenoids and steroids thus probably suggesting that the larvicidal metabolites not derived
from the isoprene units. That larvicidal activity in assayed Aloe plant extracts is not attributed
to terpenoids concurs with Phytochemistry analysis of the Meliaceae plant in which very
minimal insecticidal activity was reported (Isman et al 1996). Also profoundly absent in all
the larvicidal plant extracts of Aloes under study were the phlobatannins. Most of the
phytochemicals extracted from various plant species have indeed been tested and found to
exhibit larvicidal and repellent action against mosquitoes (Ciccia et al., 2001; Innocent et al.,
2010). The larvicidal activity of Aloe plant extracts assayed in this study against An. gambiae
and Ae. aegypti larvae corroborates earlier results achieved for plant extracts as alternative
larvicides for mosquito control (Berenbaum et al., 1991, Matasyoh et al., 2008). The
physiological responses of all the developmental stages of Ae. aegypti were profoundly
affected by the Aloe plant extracts. The larval mortality in the Aloe exposed treatments was
higher than in the control experiments. This was also depicted in survivorship studies where
the untreated mosquitoes outlived those exposed to Aloe plant extracts. The same trend was
observed in the egg deposits whereby the Aloe plant extract exposed mosquitoes had lower
fecundity as compared to the control group. The egg viability in the control groups was
higher in comparison to the Aloe exposed groups .The females in this study tended to outlive
their male counterparts. This concurs with the established norm of the mosquito life span
(Hurd et al 1995). That plant extracts affect the physiology of mosquitoes is also evidenced
by the repellent effect of constituents of essential oils of Suregada zanzibariensis
(Agiospermae Euphobiaceae) leaves to the mosquito An. gambiae s.s (Innocent et al., 2010).
It has been established that there is insecticidal, repellent and oviposition deterrent activity of
selected essential oils against Anopheles stephensi, Ae. aegypti and Cu. quinquefasciatus
(Veena et al., 2011). Despite the above observation it is generally held that the efficacy of
plant derived phytochemicals is a function of the environmental conditions (Berenbaum and
Zangeri, 1987).
While vector control is one of the options employed to contain malaria epidemics,
there is continuous increase in resistance of mosquitoes to convectional insecticides (Chandre
et al., 1999. Diabate et al., 2003 and Hunt, et al., 2010). There have also been reports of
chemical insecticide residuals deposited in food (FAO, 1992). This has necessitated the need
44

to bio prospect for secondary metabolites of botanical origin (Adeyemi 2010), that could be
environmentally friendly. The plant- derived natural products as larvicides have the
advantage of being harmless to beneficial non-target organisms and the environment
(Pitasawat et al., 2007). Hence the larvicidal effect of the bioactive Aloe plant extracts shall
be used alongside other convectional vector control methods to break Ae. aegypti and An.
gambiae mosquito breeding cycles. None of the extracts exhibited the presence of steroids.
Hence possibly phytosteroids may not be the active agents responsible for their larvicidal
effects. That flavanoids in this plant extracts could possibly be larvicidal is supported by
work done using essential oils of chloroxylon Swietenia DC against Ae. aegypti and An.
stephensi (Kiran et al 2006). The major constituents in the members of Aloeceae are typically
chromones and anthraquinones, or anthrones, and so possibly phenolic group could be
responsible for the larvicidal effect on mosquitoes (Matasyoh et al., 2008).
Tannins have been known to have histopathological effects on the midgut epithelium
of Papilio polyexenes and Papilio glaucus (family of white and black swallow tail butterflies)
(Berenbaum and Steinly, 1986). Hence it is hypothesized that the prevalence of the phenolic
group in the assayed Aloe plant extracts could play a role in physiological and larvicidal
responses against the mosquito. This study confirmed that Ae. aegypti survivorship, fecundity
and egg viability were significantly reduced due to exposure to the selected Aloe plant
extracts. Effect on the oviposition and embryo toxicity of plant extract of Indigofera
suffruticosa on early development of Ae. aegypti has been reported (Cardoso et al., 2011).
The fact that natural products can have inhibitory effect on an organism has been established
(Flavi et al., 2005). Vector borne diseases are among the major causes of illness and death in
many developing countries affecting substantial portion of the productive force. Medicinal
plants with larvicidal properties have paramount importance for the local control of the
mosquito. With the advent of drug resistance to the most commonly used drugs against the
malaria parasite; integrated vector control approach with global initiatives is the way to
contain vector-borne diseases (Adeyemi, 2010).
5.2 Conclusion
(1)The organic extracts of A. turkanensis, A. ngongensis and A. fibrosa species are
toxic to third instar larvae of Ae. aegypti and An. gambiae and hence their larvicidal activity
can be used in mosquito control.
(2) The phytochemical composition A. turkanensis, A.ngongensis and A. fibrosa
influence their toxicity to third instar larvae of An. gambiae and Ae. aegypti.
45

(3) The extracts of A. turkanensis, A. fibrosa and A. ngongensis have deterrent effect
on the developmental stages of An. gambiae and Ae. aegypti mosquitoes. hence they can be
integrated in existing vector control programmes to control the mosquito vector.
5.3 Recommendations
(1) Extracts of A. turkanensis, A. ngongensis and A. fibrosa with larvicidal activity
should be used as alternative larvicides against mosquitoes.
(2) The respective larvicide-extract responsive genes should be established through
comparative analysis of gene transcripts from sub lethal dose (LC50) survivors of the extract
exposure and non-exposed control mosquito larvae, through subtractive hybridization
molecular approaches coupled to bioinformatics analysis.
(3) Chemical purification of the extracts should be carried out to aid in definite
identification of the constituent larvicides in the Aloe plant larvicides.
(4) An attempt should be made to establish the mode of action of the larvicides for
future incorporation in formulations to counter vector resistance to insecticides.
46
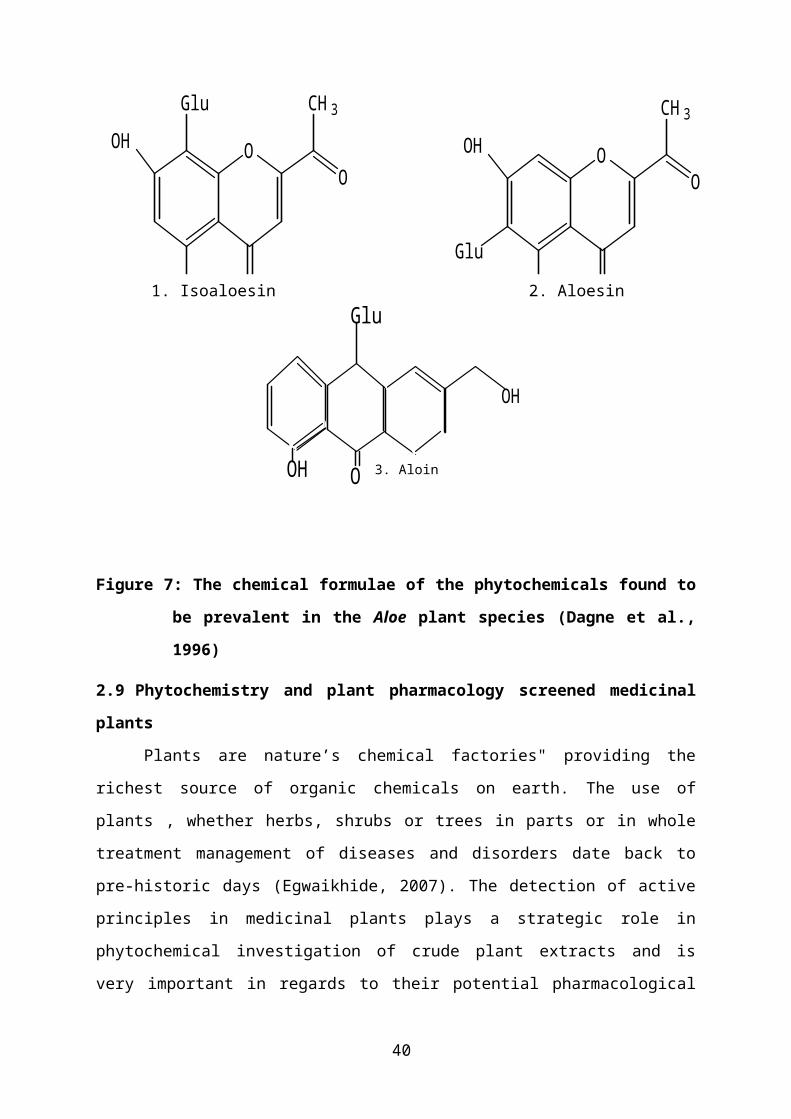
REFERENCES
Abebe D. and Ayehu A. (1993). Medicinal plants and enigmatic health practices of Northern
Ethiopia, Addis Ababa BSPE, pp. 511
Adeyemi M. M. (2010). The potential of secondary metabolites in plant material as deterrents
against insect pests. African Journal review of Pure and Applied Chemistry. 4:243-246.
Agyepong N., Mak-Mensah E. E. and Brown C. A. (2012). Prevalence of Anopheles gambiae
s. s and their pyrethroids knock down resistance pattern in communities in Kumasi
metropolis using polymerase chain reaction (PCR). European Journal of experimental
Biology. 2:304-310.
Asfaw D., Aseyed T., Abebe D., Kissi M., Daniel M. and Girum T. (2007). Screening of some
Ethiopian medicinal plants of for mosquito larvicidal effects and phytochemical
constituents. Journal of Pharmacology online. 3:231-243
Akira Yagi (2008). The chemistry of low molecular weight chromones and glycoproteins from
Aloe vera. Accessed online 01/08/2008 at file://E: \chemistry of Aloe
Amy L., Ranson H., McCall P.J., Randile N.P., Black C. W., Walker L. D. and Donlley M. J.
2005). A simplified high through method for pyrethroids knock down resistance (Kdr)
detection in Anopheles gambiae. Journal of Malaria 4: 1475-2875.
Amy E.B, (2006). Mode of action of insecticides and related pest control chemicals for
production agriculture, ornamentals and turf. Pesticides information Leaflet No.43. http://
pesticides. UMD.EDUC. Accessed 22/09/2008.
Anne B., Sophie L., Christine R., Stephane G., Sylvie D. and Catherine F. (2005). Allelopathic
potential of Medicago arborea, a Mediterranean invasive shrub. Journal of Ecology, 15:
193-198.
Apperson C, S., Harrison B.A., Unnasch T. R., Hassan H. K., Irby W.S., Savage H.M., Watson
D. W., Rueda L. M., Engber B. R. and Nasci R. S. (2002). Host-feeding habits of Culex
and other mosquitoes (Diptera: Culicidae) in the Borough of Queens in New York City,
with ccharacters and techniques for identification of Culex mosquitoes. Journal of Medical
Entomology. 39: 777-785.
Awolola T. S., Oduola A.O., Obansa J. B., Chukwurar N. J and Unyimundu J. P.(2007).
Anopheles gambiae s.s breeding in polluted water in urban Lagos, southwestern Nigeria.
Journal of Vector Borne Diseases. 44:241-244.
Baillie J. E. M. B., Hilton-Taylor C. and Stuart S.N. editors. (Eds). 2004) (. IUCN Red List of
Threatened Species: A global Species Assessment IUCN. Gland & Cambridge.
47

Beentje H. and Smith S. (2001). FTEA and after. In Robbrecht E; Degreef J. and Friis I editors.
(Eds). Journal of Plant Systematics and Phytogeography for the Understanding of African
Biodiversity. Proceedings of the XVIth AETFAT Congress Systematics and Geography of
Plants. 71:2265–2290
Bekele J. and Hassanali A. (2001). Blend effects in the toxicity of the essential constituents of
Ocimum kilimands-charium and Ocimum kenyense (Lambiatae) on two post harvest insect
pests. Journal of Phytochemistry. 57: 385-391.
Berenbaum R. and Steinly N. (1986). Histopathological effects of tannins on the midgut
epithelium of Papilio polyexenes and Papilio glaucus. Journal of Entomology and
experimental Applications.39: 3-9.
Berenbaum. and Zangeri A. R. (1987). Furanocoumarins in Wild Parsnip: effect of
photosynthetically active radiation, ultraviolet light and nutrients. Journal of Ecology. 68:
516-570.
Berenbaum R., Nitao J. K. and Zangeri A. R. (1991). Adaptive significance of furanocoumarins
diversity in Pastinaca sativa (Apiaceae). Journal of Chemical Ecology. 20: 207-215.
Bird B. H., Githinji J. W., Macharia J. M., Kasiiti J .L., Muriithi R. M and Gacheru S.G. (2008)
Multiple virus lineages sharing recent common ancestry were associated with a large Rift
Valley fever outbreak among livestock in Kenya during 2006–2007. Journal of
Virology.82:11152–66.
Blackwell A. and Johnson S.N. (2000). Electrophysiological investigation of larval water and
potential chemo-attractants for Anopheles gambiae s.s Journal of Annals of Tropical
Medicine and Parasitology. 94: 389-398.
Bødker R., Kisinza W., Malima R., Msangeni H. A. and Lindsay S.W. (2000). Resurgence of
malaria in the Usambara Mountains, Tanzania is due to an epidemic of drug-resistant
parasite. Journal of Global Change. 43:1-134.
Bosch C.H. (2006). Aloe turkanensis Christian [Internet]. Record from Protabase. Schmeizer G.
H. and Curib-Fakim A. (Editors). Prota (Plant Resourses of Tropical
Africa/Resourseshttp;//database.prota.org/search.htm>ACC
Braga C., Dourado M. I., Ximenes R. A., Alves L., Brayner F., Rocha A. and Alexander N.
(2003). Field evaluation of the whole blood immunochromatographic test for rapid
bancrofti filariasis diagnosis in the northeast of Brazil. Journal Review of Institute Medical
Tropics. 45:125–129.
48

Briet O.J., Dossou-Yovo J., Akodo E., van de Giesen N. and Teuscher T.M. (2003). The
relationship between Anopheles gambiae density and rice cultivation in the savannah zone
and forest zone of Cote d’Ivoire .Journal of Tropical Medicine in International Health.8:
43-48.
Busvine J. R. (1971). A critical Review of the Techniques for testing Insecticides, CAB,
London, pp.: 263-288.
Burgess N. R.H. and Cowan G. O. (1993). Color Atlas of Medical Entomology. pp. 10.
Carpenter S. J. (1955). Mosquitoes of North America (North Mexico). University of California
Press, Berkeley, CA. pp. 360.
Carpenter S. J. and Walter J. L. (1974). Mosquitoes of North America (north of Mexico) p. 1
Chaz M. (2008).’ Get an immune boost with Aloe vera’ “Ezzine Articles” 20 April 2008, 02
May 2008.<http// e-zine articles.com Get-an-immune-system-boost-with-Aloe-vera.
Cardoso V. J.R., Roberto M. P.L., Daniela do Amaral F. N., Everson M. B. and Sonia P. L.
(2011). Oviposition and embryo toxicity of Indigofera suffruticosa on early development of
Aedes aegypti (Diptera: Culicidae). Academic editor Diethand Wahner-Roedler volume
2012 pp. 1-5.
Chandre F., Darriet F., Manguin S., Brengues C., Carnevale P. and Guillet P. (1999).
Pyrethroids cross-resistance spectrum among populations of Anopheles gambiae s.s. from
Cote D, Ivoire. Journal of the American Mosquito Control Association. 15: 53-59.
Chandre F., Darret F., Duchon S. (2000). Modifications of pyrethroids effects associated with
kdr mutation in Anopheles gambiae. Journal of Medical and Veterinary Entomology. 8:63-
72.
Chouaibou M., Etang J., Brevault T., Nwane P., Hinzoumbe C. K., Mimpfoundi R. and Simard
F. (2008). Dynamics of insecticide resistance in the malaria vector Anopheles gambiae s. l.
from an area of extensive cotton cultivation in Northern Cameron. Journal of Tropical.
Medical International Health. 13:476-486
Ciccia G., Coussio J. and Mongelli E. (2001). Insecticidal repellent activity against Aedes
aegypti larvae of some medicinal South American plants. Journal of Ethno pharmacology
and Pharmacy. 71:267.
Clements A.N. (1992). The Biology of Mosquitoes Development, Nutrition and Reproduction,
Chapman and Hall, London.pp.536
Clements A. N. (1999). The Biology of Mosquitoes Sensory Reception and Behavior, CABI,
UK.pp.756.
49
Clements A.N. (2002). The Biology of Mosquitoes. Volume 3 Chapman and Hall. London. pp.
1
Colluzi M., Sabatia A., Della Tore A., Deco M.A. and Petracca U. (2002). A polytene
chromosome analysis of the Anopheles gambiae species complex. Journal of Science.
298:1415-1417.
Conn J. E. (1997). Systematics of Mosquito Disease Vectors (Diptera, Culicidae): Impact of
Molecular Biology and Cladistics Analysis .Journal of Annual Review of Entomology. 42:
351-369
Constant B. and Lam R. (2012). Mosquito life cycle. Accessed online 23/7/2021
Dagne E., Wyk B., Stephenson D., and Steglich W. (1996). Three oxanthrones from Aloe
littoralis .Journal of Phytochemistry. 42: 1683-1687.
Dagne E., Bisrat D., Codina C. and Bastida J. (1998). A. C. O-diglucosylated oxanthrone from
Aloe littoralis. Journal of Phytochemistry 48: 903-905.
Das S., Garver L. and Dimopoulos G. 2007). Protocol for mosquito rearing (An. gambiae).
Journal of Visions Experiments. 5: 221.
Desiree A. L., Sutherland L.J., Muiruri S., Muchiri E, M., Gray L. R., Zimmerman P. A., Hise
A. G. and Charles King H .(2011). Arbovirus Prevalence in mosquitoes, Kenya. ELD
Journal Volume 17:2.
Diabate A., Baldet T., Chandre C., Dabire K. R., Kengne P., Gulguemde T. R., Simmard F.,
Guillet P., Hemingway J. and Hougard J. M. (2003). Kdr mutation, a Genetic Marker to
Assess events of introgression between the molecular M and S forms of Anopheles
gambiae (Diptera: Culicidae) in tropical Savannah area of West Africa. Journal of Medical
Entomology. 40:1905-1906.
Dong K. (1997). A single amino acid change in the para sodium channel protein is associated
with knockdown resistance (kdr) to pyrethroids resistance in German cockroaches Journal
of Insect Biochemistry and Molecular Biology 27:93-100.
Edoega H.O., Okwu D. E. and Mbaebie B. O. (2005). Phytochemical constituents of some
Nigerian plants. African Journal of Biotechnology. 4: 685-688.
Egwaikhide P. A. (2007). Analysis of the phytochemical content and activity of plectranthus
glandulosis whole plant. Journal of Middle East Scientific Research 2: 135-138.
Enayati A. and Hemingway J. (2010). Malaria management past, present and future. Journal of
Annual Review and Entomolology.55:569-591
50

Etang J., Manga L., Jean-Claude T., Gullet P., Fondjo.E. and Chandre F. (2004). Spectrum of
metabolic based resistance to DDT and pyrethroids in Anopheles gambiae s.l. populations
from Cameroon. Journal of Vector Ecology. 32: 1.
Fanello C., Santolamazza F. Della Torre A. (2002). "Simultaneous identification of species and
molecular forms of the Anopheles gambiae complex by PCR-RFLP". Journal of Medical
and Veterinary Entomology 16: 461.
FAO (1992). Pesticide residues in food. Report, 116-146.
Feng R. and Isman M. B. (1995). Selection for resistance to Azadiracta in the green peach
aphid, Myzus persicae. Journal of Cell Molecular Life Science.51:831-833.
Finney D.J. (1971). Probit analysis-a statistical treatment of the sigmoid response curve, 3rd
edition. Cambridge, United Kingdom.
Flavia C., Maskia L., Tamara R.C., Wagner V. and Iracilda Z.C. (2005). Inhibition of hydrogen
peroxide, nitric oxide and TNF for production in peritoneal macrophages by ethyl acetate
fraction from Alchornea glandulosa. Journal of Biological and Pharmaceuticals Bulletin.
28: 1726
Ford H. R. and Green E. (1972). Laboratory rearing of Anopheles albimanus. Journal of
Mosquito News. 32:509-513.
Foster W. A. and Walker C. B. (2002). Mosquitoes (Culicidae). Journal of Medical and
Veterinary Entomology 597: 203-262.
Foote R. H. and Cook R. D. (2005) Mosquitoes of Medical Importance. pp. 168.
Fraincine H., Abdoulage A., Djimde W.M. and Thomas C. (2003). The importance and future
of malaria in Africa. Accessed 1/8/2008.
Gabi B., Lawal A. O. and Hauwa B. S. (2012). Mosquito Repellent activity and phytochemical
characterization of essential oils from Striga hermonthica, Hyptis spicigera and Ocimum
basilicum leaf extracts. British Journal of Pharmacology and Toxicology 3:43-48.
Gadelha A.P.R., Vidal F., Castro T.M., Lopez C.S., Alberella N., Coelho N., Figueiredos. F.L.
and Monteiro-leal L.H. (2005). Susceptibility of Giandia lamblia to Hoveria dukis
extracts. Journal of Parasitology Research. 97:399-407.
Gao X., Yan X., Yao X., Xu1 L., Zhang K., Zhang J., Yang B. and Jiang L. (2007). The dry-
style antifogging properties of mosquito compound eyes and artificial analogues prepared
by soft lithography. Journal of Advanced Materials. 19:2213-2227.
Gilbert A. and Calderone, (2007). Mosquito control. Suffolk county Government. Accessed
3/9/[email protected]/health/suffolk.vectorplan/ pdf/final/appendix
51

Gillies M.T. and De Meillon. (1968). The Anophelinae of Africa South of the Sahara.
(Ethiopian zoogeographical region) South African Journal. Institute for Medical Research,
Johannesburg, 343
Goma L. K. H. (1959). Periodic pupation in Anopheles gambiae Giles. Journal of
Entomological Society of South Africa. 22:275-276
Gospiesh V. K. and Kannabiran K. (2007). Larvicidal effect of Hemidesmus indicus, Gymnema
sylvestre and Eclipta prostrate against Culex quinquefasciatus mosquito larvae .African
Journal of Biotechnology. 6: 307-311.
Gubler D. J. (1998). Resurgent vector –borne diseases as a global health problem. Journal of
Emergent Infectious Diseases. 4: 442-450
Guerrero F. D., Jamroz. R. C., Kammaiah D. and Kunz S.E. (1997). Toxicological and
molecular characterization of pyrethroids-resistant Haematobia irritants: Identification of
kdr and super-kdr point mutations. Journal of Insect Biochemistry and Molecular Biology.
27:745-755.
Gutterman Y. and Chausen V. (2007). Secondary phenol metabolites (SPHMS) distribution
and content of some Aloe species originated from arid zones of South Africa: A review.
Journal of Food Technology. 2: 555-569.
Harboune J. B. (1991). Phytochemical Methods. A guide to Modern Techniques of Plant
Analysis. Chapman and Hall. New York, Tokyo. Melbourne, Madras. Second edition.
Textbook, pp. 120.
Hargreaves K., Hunt R.H., Brooke B.D., Muthembu J., Weeto M. M., Awolola T.S. and
Coetzee M. (2003). Anopheles arabiensis and An. quadriannulatus resistance to DDT in
South Africa. Journal of Medical and Veterinary Entomology.17:417-422.
Huang J. Walker, Edward D, Vulule J. Miller J. R. (2006) Daily temperature profiles in and
around Western Kenyan larval habitats of Anopheles gambiae as related to egg mortality.
Malaria Journal. 5:87.
Hunt R. H., Edwardes M.and Coetzee M. (2010). Pyrethroids resistance in Southern African
Anopheles funestus extends to Likoma Island in Lake Malawi. Journal Parasites and
Vectors 3: 122
Hurd H., Hogg C. J. and Renshaw M. (1995). Interactions between blood feeding, fecundity
and infection in mosquitoes. Journal of Parasitology Today. 11:411-416.
52

Innocent E., Cosam C. J. and Gikonyo N. K. (2010). Constituents of the essential oil of
Suregada zanzibariensis leave are repellent to the mosquito Anopheles gambiae s.s
Journal of Insect Science. 10: 1-8.
Isman M.B., Matsuura H., Mackinnon S., Durst T., Towers G.H.N. and Arnason J. T. ( 1996).
Phytochemistry of the Meliaceae: so many terpenoids, so few insecticides. In
Phytochemical diversity and redundancy in ecological interactions, Plenum Press, New
York, London pp. 155-178.
Isman M. BB. and Machial C. M. (2006). Pesticides based on plant essential oils: from
traditional practice to commercialization. In Rai M. and Carpinella M. C. (Eds.). Naturally
occurring Bioactive Compounds. Journal of Elsevier, BV.11: 29-44
Isman M. B., Akhtar Y. and Yeoung Y. K. (2008). Comparative bioactivity of selected extracts
from Meliaceae and some commercial botanical insecticides against two noctuid
caterpillars, Trichoplasia ni and Pseudaletia unipunota. Journal of Phytochemistry
Review. 7: 77-78.
Jacobson M. C. (1975). Insecticides from plants: A review of the literature, 1954-1971:
Agricultural Handbook, US Department of Agriculture, Washington D. C., pp. 462:138.
Jacobson M. C. (1989). Botanical pesticides, past, present and future. Insecticides of plant
origin (Ed. Arnason, J. T.). Proceedings of the Chemical Society, Washington. D. C., pp. 1
-10.
Jaiyesimi A. A. and Anthony O. (2011). Larvicidal properties of Paullina pinnata Linn leaf
against Anopheles gambiae. Journal of Pharmacognosy Communication.1:1
Jamshaid I. (2006). Determination of the prevalence of lymphatic filariasis among migrant
workers in Kuwait by detecting circulating filarial antigen. Journal of Medical Microbiology.
55: 401-405
Jawara M., Smallegange R. C., Jeffries D., Nwakurima D. C., Awolola T. S. Bart G., Knols W.
T, and Conway D. J. (2009). Optimizing Odor Baited Traps Methods for collecting
mosquitoes during the malaria season in the Gambia. Journal of PLoS ONE, 4: 8167.
Jean-Michel L. P. T., Jean-Bernard D., Laurence M., Patrick R., Gilbert L. G., Voahirana G.
and Vincent R. (2003). Distribution of the species of the Anopheles gambiae complex and
first evidence of Anopheles merus as a malaria vector in Madagascar. Malaria Journal.
2:33.
Kakkilaya B.S. (2006). Control of malaria: scandalously scarce resource Journal of Nature.
3:417-419.
53

Kalyanasundaram M. and Das P. K. (1985). Larvicidal and synergist activity of plant extracts
for mosquito control. Indian Journal of Medical Resources. 82:19-23
Keating J., Mbogo C.M., Mwangangi J., N Zovu J. G., Guiyun Y., Githure J. I. and Beier J.
(2005). Anopheles gambiae s.l. and Anopheles funestus mosquito Distributions at 30
villages along the Kenyan coast. Journal of Medical Entomology. 43: 241-246.
Kemri. (2012). Kenya Malaria Fact Sheet. Malaria in Kenya at a glance. Accessed online on
23/7/2012 at http//www.org/inclee.php/aboutkemri/board-of-management
Kiran S.R., Bhawan K., Devis P. S., Rajeswara R. and Reddy J. K. (2006). Composition and
larvicidal activity of leaves and stem essential oils of chloroxylon Sweetenia DC against
Aedes aegypti and Anopheles stephensi. Journal of Bioscience Technology, 97: 2481-2484.
Howell P.I.and Knols B. G J. (2009). The Biology of mosquito mating. Malaria Journal. 2:8
Kitron U. and Spielman A. (1989). Suppression of transmission of malaria though source
reduction: antianophelline measures applied in Israel, The United States and Italy. Journal
Review of Infectious Diseases.11:391-406.
Kohl O., Walia S. and Dhaliwal G. S. (2008). Essential oils as green pesticides: Potential
constraints. Journal of Biopestic International. 4: 63-84.
Kuniholm M. H., Wolfe N.D., Huang C. Y., Mpoudi-Ngole E., Tamoufe U., Le Breton M.,
Burke D.S. and Gubler D. J. (2006). Seroprevalence and distribution of Flaviridae,
Togaviridae, and Bunyaviridae arboviral infections in rural Cameroonian adults. American
Journal of Tropical Medicine and Hygiene. 75: 371
Kong X. Q. and Wu C. W. (2009). Measurement and Prediction of Insertion Force for the
Mosquito Fascicle Penetrating into Human Skin. Journal of Bionic Engineering. 6: 143-
152.
Lehane M. J. (2005). The biology of blood-sucking in insects. Cambridge University Press. pp.
58
Lehrer S. (2010). Anopheles / mosquito transmission of brain tumor. Journal of Medical
Hypotheses. 74:167-168.
Leiriao P., Rodrigues C.D., Sonia S.A. and Mota M.M. (2004). Survival of protozoan intra
cellular parasites in host cells. Journal of EMBO reports 5:1142-1147
Leon B. and Steven S. (2002). Why Study Ecology in Temporary Pools? Journal Israel of
Zoology 47: 4
Leunita A, S. and Ogbunugafor C. B. (2008). Behavioral evidence for the existence of a region-
specific oviposition cue in Anopheles gambiae s.s Journal. of Vector Ecology. 33:321-324.
54

Lhuillier M., Mazzariol M. J., Zadi S., Le-Cam N., Benteja M. C., Adamowiez L., Marie F. N.
and Fritzel B.(1989). Study of combined vaccination against yellow fever and measles in
infants from six to nine months. Journal of Biology standing. 17:9-15.
Luke E. M., Rodney L C., Lillian A M., Trish P., Fredrick O., Victor O. O., Randal J S., Cindy
A R. and Nicholas A. (2011). Seroprevalence and distribution of arboviral infections
among rural Kenyan adults: A cross-sectional study. .Journal of Virology 8:371.
Linquist M., Kramer Laura and Dennis L. (2012). Mosquito reference manual. Accessed online
on 25/7/2012
Lyimo E.O., Taakken W. and Koella J.C. (1992). Effect of rearing temperature and larval
density on larval survival, age at pupation and adult size of Anopheles gambiae. Journal of
Experimental Entomology. 63:265-271.
Magdula J. J., Innocent E. and Otieno J.N. (2009). Mosquito larvicidal and cytotoxic activities
of 3 Annona species and isolation of active principles. Journal of Medicinal Plants
Research.3:674-680.
Maiyo Z.C., Ngure R.M., Matasyoh J.C. and Chepkorir R. (2010). Phytochemical constituents
and antimicrobial activity of leaf exudates of three Amaranthus plant species. African
Journal of Biotechnology. 9: 3178-3182.
Majaw S. and Moirangthem E., (2009). Qualitative and quantitative Analysis of Clerodendron
colebrookiarium Walp. Leaves and Zingibe cassumunar Roxb. Rhizomes. Journal of
Ethno botanical Leaflets, 13: 578-589.
Matasyoh J.C., Wathuta E. U., Kariuki S. T., Chepkorir R. and Kavulani J. (2008). Aloe plant
extracts as alternative larvicides for mosquito control. African Journal of Biotechnology. 7:
912-915.
Mazen A. (2012). Impact of flavonoids against woolly apple aphid, Eriosoma lanigerum
(Hausman) and its sole parasitoid Aphelinus mali (Haid). Journal of Agricultural Science.
4:2.
Michael E., Simonsen P.E., Malecela M., Jaoko W. G., Pedersen E. M., Mukoko D.,
Rwegoshora R. T. and Meyrowitsch D. W. (2001). Transmission intensity and the
immunoepidemiology of bancrofti filariasis in East Africa. Journal of Parasite
Immunology. 23: 373-388.
Mcllister T.M., Anneh C.B., Cockwill CL., Olsorim E., Wang Y. and Cheeke P.R. (2001).
Studies on use of Yucca schidigera to control giardiasis. Journal of Veterinary
Parasitology. 97: 85-89.
55

Mohsen Z. H., Jawad A. M. al Saad M. and Al Naib A. (1995). Antioviposition and insecticidal
activity of Imperta cylindrical (Graminaceae).Journal of Medical Veterinary
Entomology.9: 441-442
MOH (2001) Kenya malaria Fact Sheet. Malaria in Kenya at a glance. Kemri report. Accessed
online 22/7/ 2012.
MOH (2006). Kenya malaria Fact Sheet. Malaria in Kenya at a glance. Accessed
online22/7/2012.
Monath T. P. (1991) Yellow Fever: Victor, Victoria? Conqueror, Conquest Epidemics and
Research in the Last Forty Years and Prospects for the Future. American Journal of
Tropical. Medicine and Hygiene: 45:1-43.
Mugo B. (2012).The Impact of the Global Fund: “The will to win: how the global fund has
changed millions of lives in Kenya”. Remarks by Hon. Beth Mugo E.G.H., M.P, during
the Global Fund Session at the European Parliament, Brussels, Belgium. Malaria News
11TH April, 2012. Accessed online on 22/ 7/2012.
Mulla M. S., Thavara V., Tawatsin A., Chopsri J. and Su T. (2003). Emergence of resistance
management in field populations of tropical Culex quiquefasciatus to the microbial agent
Bacillus sphaericus. Journal of American Mosquito Control Association. 19:39-49.
Muirhead–Thompson, R.C.N. (1945). Studies on breeding places and control of Anopheles
gambiae in the Coastal district of Sierra Leone. Journal of Bulletin of Entomological
Research, 36:185-252.
Munga S., Minakawa N., Zhou G. B., Okeyo-Owuor J., Githeko A. K. and Yan G. (2005).
Oviposition Site Preference and Egg Hatchability of Anopheles gambiae: Journal of
Effects of Land Cover Types. 42: 993-997.
Munga S., Minakawa N., Zhou G., Mushinzimana E., Barrack O. O and Githeko A. K. (2006).
Association between land cover and habitat productivity of malaria vectors in Western.
American Journal of Tropical Medicine and Health. 74:75-69.
Najera J. A., Kouznetsov R, L. and Delacollette C. (2011). Malaria control forecasting and
prevention.Accessed27/1/2012@http//www.rollbackmalaria.org/docs/najeraepedemics/
najll.htm
Nauwinger H. D. (2000). African traditional medicines: a dictionary of plant use and
application. Med Pharm Scientific; Stuttgart, Germany. pp. 589.
Nelson M. J. (1986). Aedes mosquitoes: Biology and Ecology. Pan American Health
Organization, Washington, D. C.
56

NʹGuessan R., Corbel V., Akogbeto M. and Rowland M. (2007). Reduced efficacy of
insecticide treated nets and indoor residue spraying for malaria control in pyrethroids
resistance area, Benin. Journal of Emerging Infectious diseases. 13:199-206
Nishiura J., Polly T. and Ray K. (2003). Methoprene Interferes with Mosquito Midgut
Remodeling During Metamorphosis Journal of Medical Entomology. 40:498-507.
Nobert B., Dusan P. and Marija Z. 2010. Mosquitoes and their control. European mosquito
Bulletin. Springer Publishers. pp. 25.
Nwanjo H. U (2006). Antioxidant activity of the exudates forms Aloe barbadensis leaves in
diabetic rats. Journal of Bouline International. 18: 77-81.
Oklahoma University (2012). Entomology and Plant Pathology. Media News. Fax
405.744.6039. Accessed online 21/7/2012.
Paulraj M.G., Reagan A. D. and Ignacimuthu S. (2011). Toxicity of benzaldehyde and
propionic acid against immature and adult stages Ae .aegypti (Linn) and Culex
quinquefasciatus (Say) (Diptera: Culicidae). Journal of Entomology 8: 539-547.
Pitasawat B., Chapakaew D., Choochote W., Jitpakdi A., Chaithong U., Kanjanapothi R.,
Tippawangkosol P., Riyong D., Tuetun B. and Chaiyasit D. (2007). Aromatic plant
derived essential oil: An alternative larvicide for mosquito control. Journal of Fitoterapia
78: 205 - 210.
Rattan R.S.(2010). Mechanism of action of insecticidal secondary metabolites of plant origin:
Crop protection.29:913-920.
Roger W. (2009). Communicable disease epidemiology and control: A global perspective.CA
BI, pp. 178.
Rohmer M. (1999). The discovery of the mevalonate-independent pathway for isoprenoid
biosynthesis in bacteria, algae and higher plants. Journal. Natural Products Report. 16.
565-574
Roll Back Malaria Partnership. Roll Back Malaria: global strategic plan. 2005-2015. Geneva:
RBM; 2005.
Romeo J. T., Saunders J.A. and Barbosa P. (1996). (Eds) Recent advances in Phytochemistry,
Vol. 30. Phytochemical diversity and redundancy in ecological interactions Plenum press,
New York, London pp. 155-178.
Royston M. R., Gilbert J. C., Lynn B. R. and Allan S. W. (1974). An introduction to Modern
Experimental Organic Chemistry. Second edition, pp. 440
57

Sathish K. and Maneemegalai. (2008). Evaluation of larvicidal effect of Lantana camara Linn
against Aedes aegypti and Culex quiquefasciatus. Journal of Advances in Biological
Research.2:39-43.
Schäfer M.L. and Lundström J.O. (2006). Different responses of two flood - water mosquito
species, Aedes vexans and Ochlerotatus sticticus (Diptera: Culicidae), to larval habitat
drying. Journal of Vector Ecology. 31:123-128.
Service, M. W. (1993). Mosquito Ecology: field sampling methods, Chapman and Hall,
London. pp. 988.
Service M. W. and Ashford R. W. (2001). Encyclopedia of Arthropod Infections of Man and
Domesticated Animals. Chapman and Hall, London pp. 319.
Singh M., Dhanalakshmi E. and Rao M. (2000). Chemo modulatory action of Aloe vera on the
profiles of enzymes associated with carcinogen metabolism and antioxidant status
regulation in mice .Journal of Phytomedicine 28:317-324.
Snow R.W., Guerra C. A., Noor A. M., Myint H. Y. and Hay I.S. (2005). The global
distribution of clinical episodes of Plasmodium falciparum malaria. Journal of Nature
434: 214-217
Steel R. G. D. and Torri J. H. (1980). Principles and procedures of statistics. A Biometrical
approach. Second edition. pp. 170-176
Stacy A. G.(2012) 33-Year-Old Liver Transplant Recipient Returns From Hunting Trip in
Sudan With Intermittent Fevers and Gross Hematuria. Journal of Laboratory Medicine:
43:3-6
Tan B.K.H. and Vanitha J. (2004). Immunomodulatory and antimicrobial effects of some
traditional Chinese herbs: A review. Journal of .Current Medicinal Chemistry, 11: 1423-
1430
Tawfik K. M. Sheteawi S. A. and El-Gawad Z. A. (2001). Growth and aloin production of Aloe
vera exudates under different ecological conditions. Ethiopian Journal of Biology
(Botany). 3: 149-159.
Tiwari A.K. and Rao M. (2002). Diabetes mellitus and multiple therapeutic approaches of
phytochemicals: present status and future prospects. Journal of Current Science 83: 30-38.
Thomas A.W. (2004). The molecular site of action of juvenile hormone and juvenile
metamorphosis: How these compounds kill insects. Journal of Insect Physiology. 50:111-
121.
58

Tomlin C.D. S. (2003). The pesticide manual. British crop protection council Publication 13
edition Pg. 37
UNICEF (2010). Fact of the week-90: Of all malaria deaths worldwide, currently 90 per cent
occur in Sub- Sahara Africa. Millennium Development Goal 4: Reduce Child
Mortality.Updated:11 October 2010. http:// www.chilinfo.org./malaria.htm1
Ulrike F., George S.., Gerry F., Killeen B. G. J., Knolls G.J. and Norbert B. (2004). The
practical importance of permanent and semi permanent habitats for controlling aquatic
stages of Anopheles gambiae sensu lato mosquitoes: operational observations from a rural
town in western Kenya. Journal of Tropical Medicine and International Health. 9 :1274–
1289
Utzinger A., Tozah T. and Singer B.H. (2001). Tropical medicine and international health
Blackwell Synergy. Efficacy and cost effectiveness of environmental management for
malaria control. Journal of Medical International Health. 6:677-687
Utzinger J., Tanner M., Kammen D, M., Killeen G. F. and Singer B. H. (2002).Integrated
programme is key to malaria control. Journal of Nature 4: 419-431.
Van Damme P. (1991). Plant Ecology of the Nabib desert. Journal of Africa focus, 7: 355-400.
Varavikova E. and Theodore H.T. (1998). The new public health. Journal of Emergent
Infectious Diseases. 4:442-450
Veena P., Tripathi A.K., Aggarwal K.K. and Khanuja S. P. S. (2011). Insecticidal, repellent
and oviposition deterrent activity of selected essential oils against Anopheles stephensi,
Aedes aegypti and Culex quinquefasciatus. Journal of Bioresource Technology, 96: 1749-
1757
Wabuyele E., Bjorå C. S., Nordal I. and Leonard E. N. (2006). Distribution, Diversity and
conservation of the genus Aloe in Kenya. Journal of East African Natural History. 95:213-
225.
Walker E. D., Thibault A.R., Thelen A. P., Bullard B. A., Huang J., Odiere M. R., Bayoh N.M.,
Wilkins E. E. and Vulule J. M. (2007). A Review Distribution of African malaria
mosquitoes belonging to the Anopheles gambiae complex. Journal of Malaria 27: 60: 23.
Weill, M., Chandre F., Brengues C., Manguin S., Akogbreo M., Pasteur N., Guillet, P. and
Raymond, M. (2000). The unique mutation in ace-1 giving high insecticide resistance is
easily detected in mosquito vectors. .Journal of Insect Molecular Biology. 9: 451-455.
59
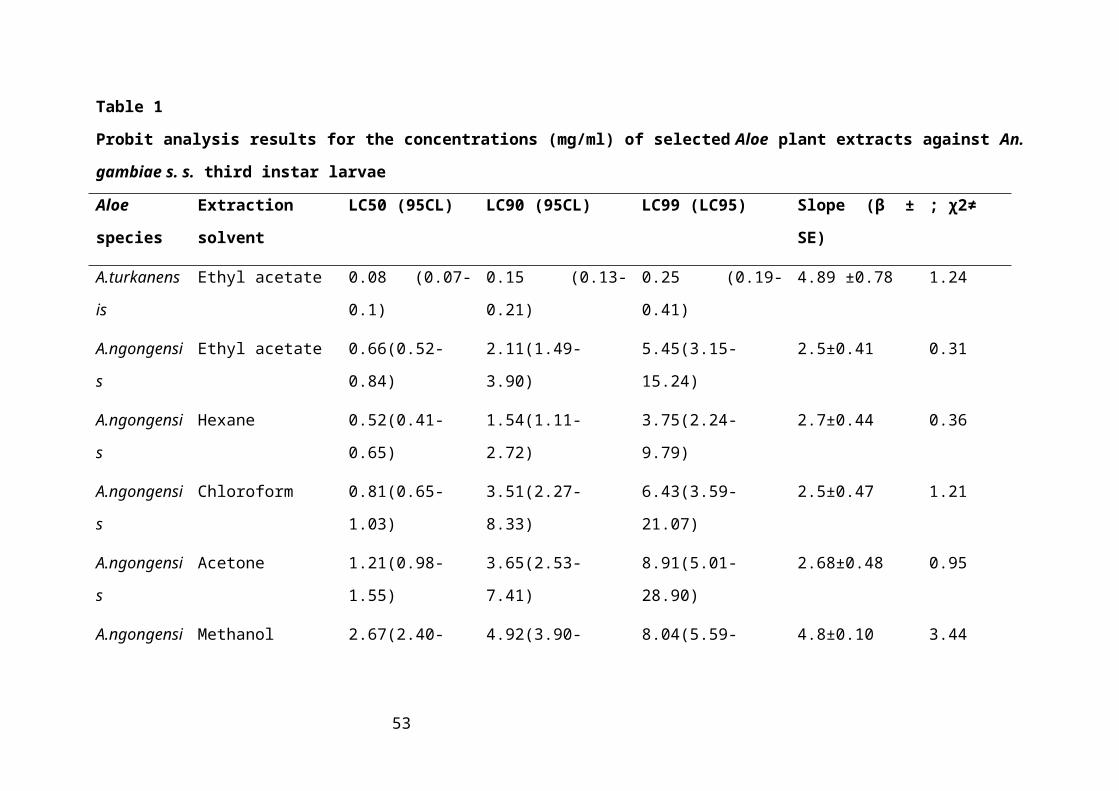
Weill M., Malcolm C., Chandre F., Mogensen K., Bertholomiew A. S., Marquina B. and
Raymond M. (2004). The unique mutation in ace-1 giving high insecticide resistance is
easily detected in mosquito vectors. Journal of insect Molecular Biology. 3: 1-7.
Wikipedia, the free Encyclopedia (2012). Mosquito. Accessed online 20/7/2012 at
http://en.wikipedia.org/wiki/mosquito#mw-head
White G. B.(1985) .Anopheles bwambae sp., a malaria vector in the Semliki Valley, Uganda,
and its relationships with other sibling species of the An.gambiae complex (Diptera:
Culicidae). Journal of Systematic Entomology. 10:501-522.
Wirth M. C., Hyun-Woo P., Walton W. E. and Federiri B. A. (2005). Cyt1A of Bacillus
thuringiensis delays evolution of resistance to Cry11A in the mosquito Culex
quinquefasciatus Journal of Applied Environmental Microbiology. 71: 1185-1189.
WHO, 1992. Division of Epidemiological Surveillance and Health Situation and Trend
Assessment. Global Health Situation and Projections: Estimates. Geneva, Switzerland.
WHO, (1998). Division of emerging and other communicable diseases surveillance and control
global programme for vaccines and immunization expanded programme on immunization.
WHO. (1996). Investing in health research for development. Report of the Ad Hoc Committee
on Health Research Relating to Future Intervention Options; 1996 Report No.
TDR/Gen/96.1. Geneva: The organization
WHO and UNICEF (2003). The Africa malaria Report WHO/CDS/MAL/2003.1093. Retrieved
27th April 2003 from http://mosquito.who.int/amd2003/
WHO (2005). Guidelines for laboratory and field testing of mosquito larvicides. World Health
Organization communicable disease control, prevention and eradication. WHO pesticide
evaluation scheme.
WHO. (2006). Malaria vector control and personal protection. Report of WHO study Group
WHO Technical Report Series 936.
WHO,(2009). Integrated malaria vector control with microbial larvicides and insecticide
treated nets in Western Kenya: a controlled trial. Bulletin of the World Health
Organization. 87: 645-732
WHO. (2010). First WHO Report on neglected diseases 2010: working to overcome the global
impact of neglected tropical diseases.
WHO (2011). Urgent action essential to protect malaria therapies says WHO
WHO (2011) .Global plan for artemisinin resistance containment. pp 1-87.
60
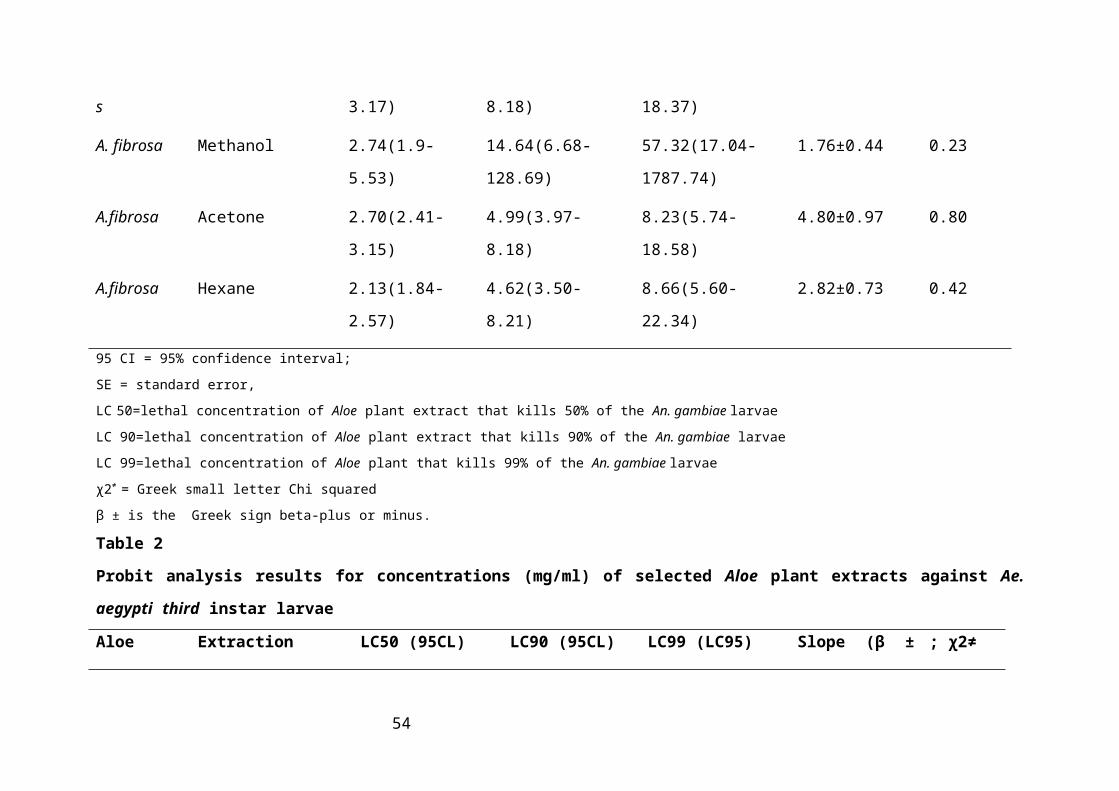
WHO (2011). Global Alert and Response (GAR). Yellow fever: a current threat. Accessed 27 th
January 2011 @ http://www.who.int/csr/disease/yellowfev/impactl/en/index.html
WHO (2012). Dengue and the severe dengue. Fact sheet No. 117.
Womack M. (1993). "The yellow fever mosquito, Aedes aegypti. Journal of Wing Beats 5: 4
Yagi A. and Takeo S. (2003). Anti-inflammatory constituent’s aloesin and aloemannan in Aloe
species and effects of tanshinon VI in Salvia miltiorrhiza on heart accessed online
08/08/2012. yagi @fupharm.fukuyama-u.ac.jp.
Yakugaku Z. (2003). Ant-inflammatory constituents of aloesin and aloemannan in Aloe
species. Journal of Public Medicine. 123:517-532.
61

APPENDICES
Appendix 1
F0 Post 24 hour exposure to Aloe extracts
Treatment Rep. 1 Rep. 2 Rep. 3
Mean
Mortality
%
Mortality
Mean
Survivorship
%
Survivorship
Dead Alive Dead Alive Dead Alive
Control 25 1475 55 1445 40 1460 40 2.67 1460 97.33
1%Dimethyl Sulphoxide 150 1350 115 1385 155 1345 140 9.33 1360 90.67
1% ethyl acetate 495 1005 390 1110 440 1060 442 29.44 1058 70.56
LC50 A. ngongensis 835 665 760 740 725 775 773.33 51.56 726.67 48.44
LC50.A. turkanensis 759 741 775 725 805 695 779.67 51.98 720.33 48.02
LC50 A.fibrosa 730 770 720 780 737 763 729 48.6 771 51.4
62

Appendix 2
Pupae emergence
Treatment Rep. 1 Rep. 2 Rep. 3Mean
Mortality
% Mortalit
yMean
Survivorship%
Survivorship
Dead Alive Dead Alive Dead Alive
Control 15 1445 5 1455 26 1434 15.336 1.05 1444.67 98.95
1%Dimethyl Sulphoxide 10 1350 30 1330 40 1320 26.67 1.96 1333.33 98.04
LC50 A. ngongensis 107 620 77 650 92 635 92 12.65 635.00 87.35
LC50.A. turkanensis 101 619 112 608 91 629 101.33 14.07 618.67 85.93
LC50 A.fibrosa 104 667 90 681 72 699 88.67 11.5 682.33 88.50
Appendix 3
Adult Male Emergence
63
Treatment Rep. 1 Rep. 2 Rep. 3 Mean %
Control 671 668 663 667.33 88.98
DMSO 659 663 669 663.67 88.49
LC50 A. ngongensis 323 319 326 322.67 43.02
LC50 A. turkanensis 309 312 299 306.67 40.89
LC50 A. fibrosa

Appendix 4
Adult female emergence
Appendix 5
Male: female sex ratio (Male/Total)
64
Treatment Rep. 1 Rep. 2 Rep. 3 Mean %
Control 641 646 635 640.67 85.42
DMSO 633 640 637 636.67 84.89
LC50 A. ngongensis 310 315 312 312.33 41.89
LC50 A. turkanensis 309 305 286 300 40
LC50 A. fibrosa 346 352 351 349.67 46.62
Treatment Sex ratio
Control 0.51
1%Dithyl Sulphoxide 0.51
LC50 A. ngongensis 0.51
LC50 A. turkanensis 0.51
LC50 A. fibrosa 0.50
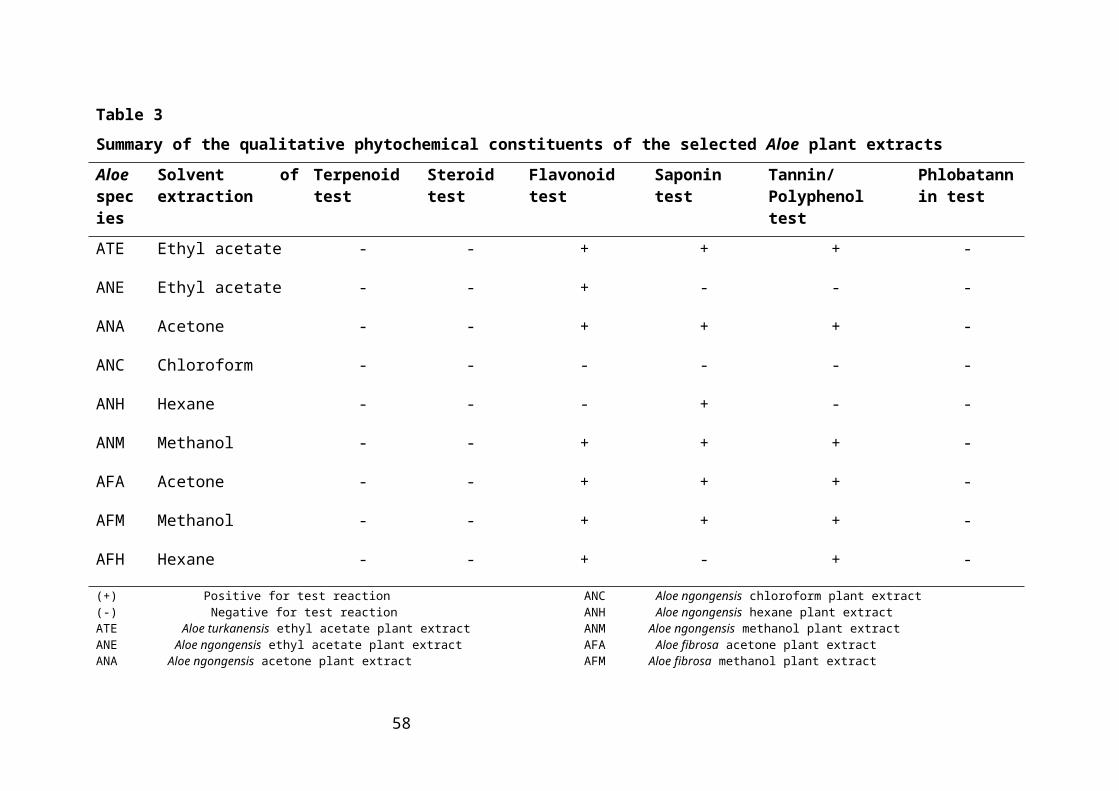
Appendix 6
Egg deposits (oviposition) studies on 30 males+30 females
Appendix 7:
Age specific survivorship of adults for males ( F0) where; x=yx/y0
65
Treatment Rep. 1 Rep. 2 Rep. 3 Mean ED Fecundity
Control (dist. Water) 6461 8121 7600 7394.00 246.67
1%Dimethyl Sulphoxide 5947 6300 5844 6030.33 201.01
LC50 A. ngongensis 4100 4230 4300 4210.60 140.33
LC50 A. turkanensis 4428 3996 4120 4181.33 139.37
LC50 A. fibrosa 4500 4670 4800 4656.67 155.22
Treatment Rep. 1 Rep. 2 Rep. 3 Mean x=yx/y0 (28th day)
Control 42 46 40 42.66 0.71
1% Dimethyl sulphoxide 39 37 42 39.33 0.67
LC50 A. ngongensis 14 17 12 14.33 0.23
LC50 A. turkanensis 10 13 9 10.67 0.18
LC50 A. fibrosa 25 29 31 28.33 0.47

Appendix 8
Age-specific survivorship of female adults (F0)
Appendix 9:
Male Age specific mortality studies (F1)
66
Treatment Rep. 1 Rep. 2 Rep. 3 Mean x=yx/y0 (28th day)
Control 23 29 25 25.67 0.85
1% Dimethyl sulphoxide 25 28 20 24.33 0.81
LC50 A. ngongensis 10 9 13 10.67 0.36
LC50 A. turkanensis 8 6 9 7.67 0.26
LC50 A. fibrosa 19 24 22 21.67 0.72
Treatment Rep. 1 Rep. 2 Rep. 3 Mean x=yx/y0 (28th day)
Control 19 14 17 16.67 0.56
1% Dimethyl sulphoxide 14 11 19 14.67 0.49
LC50 A. ngongensis 4 5 1 3.33 0.11
LC50 A. turkanensis 2 4 1 2.33 0.08
LC50 A. fibrosa 9 4 6 6.33 0.21

Appendix 10:
F1 Egg Hatchability 48 hours post Aloe extracts exposure of 1500 eggs of Aedes aegypti
Appendix 11
Larvae emergence
Treatment Rep 1 Rep. 2 Rep. 3 Mean Mortality % Dead Mean Alive % Alive
Dead Alive Dead Alive Dead Alive
Control 43 1410 50 1403 39 1414 44 3.03 1409 96.97
1%DMSO 152 1267 141 1278 136 1283 143 10.8 1276 89.92
LC50 A. ngongensis 217 483 200 500 230 470 216 30.86 484 69.14
LC50 A. turkanensis 134 163 147 150 154 143 145 48.82 153 51.52
LC50 A. fibrosa 172 577 180 569 199 550 184 24.56 565 75.43
67
Treatment Rep. 1 Rep. 2 Rep. 3 Mean Viability % viability
Control 1452 1437 1470 1453 96.87
1% Dimethyl sulphoxide 1398 1460 1400 1419 94.6
LC50 A. ngongensis 750 630 721 700 46.67
LC50 A. turkanensis 300 250 340 297 19.8
LC50 A. fibrosa 800 690 757 749 49.93

Appendix 12
Pupae emergence
Treatment Rep. 1 Rep. 2 Rep. 3Mean
Mortality%
MortalityMean
Survivorship%
Survivorship
Dead Alive Dead Alive Dead Alive
Control 32 1377 29 1380 24 1385 28.33 2.01 1380.67 97.98
1%Dimethyl Sulphoxide 26 1250 34 1242 16 1260 25.33 1.98 1250.67 98.01
LC50 A. ngongensis 63 421 54 430 67 417 61.33 12.67 422.66 87.32
LC50.A. turkanensis 21 132 24 129 28 125 24.33 15.9 128.67 84.09
LC50 A.fibrosa 80 485 72 493 86 479 79.33 14.04 485.67 85.96
Appendix 13
Adult male emergence
Treatment Rep 1 Rep. 2 Rep. 3 Mean %
Control 648.00 654.00 639.00 647.00 86.27
1%DMSO 627.00 635.00 623.00 628.33 83.77
LC50 A. ngongensis 192.00 189.00 200.00 193.66 25.82
LC50 A. turkanensis 57.00 60.00 69.00 62.00 8.27
LC50 A. fibrosa 218.00 230.00 225.00 224.33 29.91
68

Appendix 14
Adult female emergence
Treatment Rep 1 Rep. 2 Rep. 3 Mean %
Control 628.00 628.00 617.00 624.33 83.24
1%DMSO 603.00 622.00 598.00 607.67 81.02
LC50 A. ngongensis 186.00 182.00 192.00 186.67 24.88
LC50 A. turkanensis 55.00 57.00 48.00 53.33 7.11
LC50 A. fibrosa 215.00 224.00 221.00 220.00
Appendix 15
Male and female sex ratio
69
Treatment Sex ratio
Control 0.51
1%Dithyl Sulphoxide 0.51
LC50 A. ngongensis 0.51
LC50 A. turkanensis 0.51
LC50 A. fibrosa 0.50

Appendix 16
Aedes aegypti F1 oviposition
70
Treatment Rep. 1 Rep. 2 Rep. 3 Mean oviposition Fecundity
Control (dist. Water) 7300.00 7000.00 6800.00 7033.00 234.43
1%Dimethyl Sulphoxide 7342.00 7742.00 7760.00 7614.66 253.82
LC50 A. ngongensis 4847.00 4997.00 5000.00 4948.00 164.93
LC50 A. turkanensis 4500.00 4390.00 4900.00 4596.66 153.22
LC50 A. fibrosa 6934.00 7200.00 7114.00 7082.66 236.08

Appendix 17
Qualitative analysis of terpenoids from selected larvicidal Aloe plant extracts
71
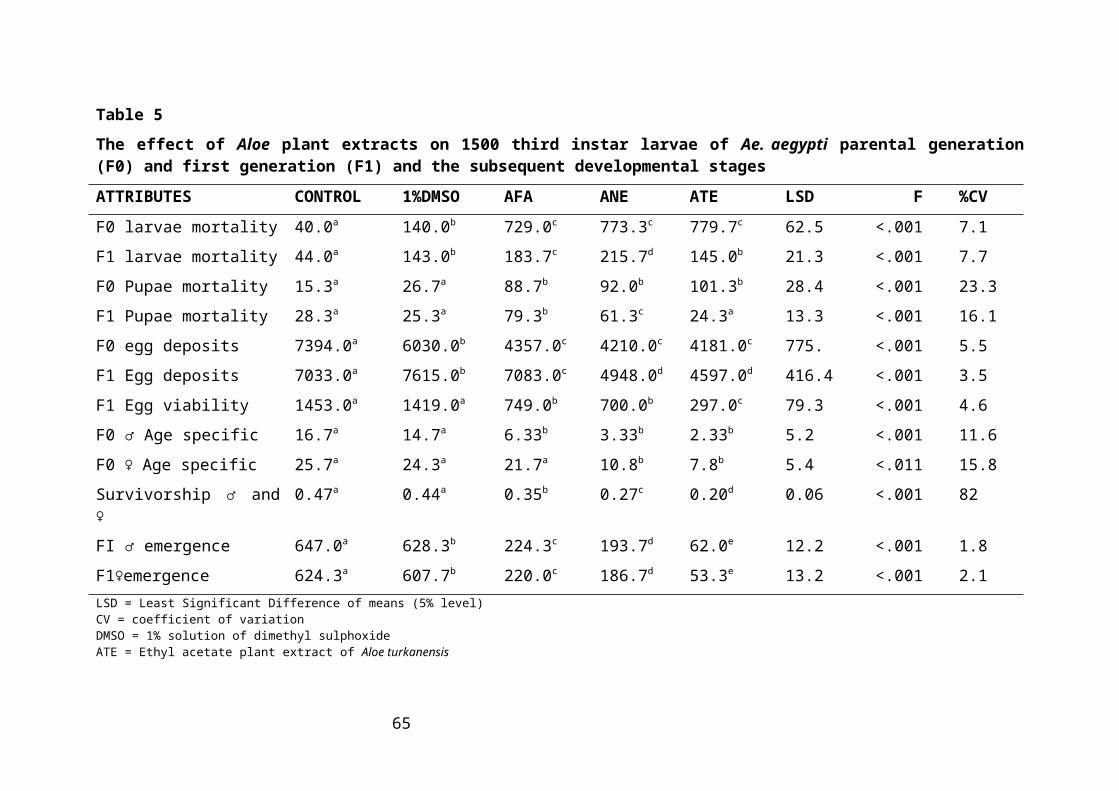
Appen
dix 18
Qualit
ative
analysis of steroids of selected larvicidal Aloe plant extracts
EXTRACT Observation Inference
ATE Formation of a brownish yellow solution was observed Absence of steroids
ANH A brownish colored solution was formed that was stable on standing Absence of steroids
ANE A yellow colored solution was formed Absence of steroids
ANC A brown colored solution was formed Absence of steroids
72
Extract Observation Inference
ATE A dark brown coloration at the interface Absence of terpenoids
ANH A dark brown coloration at the interface Absence of terpenoids
ANE A light brown coloration of interface Absence of terpenoids
ANC A brown coloration of interface Absence of terpenoids
ANA A brown of interface Absence of terpenoids
ANM A dark brown coloration of interface Absence of terpenoids
AFH A dark brown coloration of interface Absence of terpenoids
AFA A dark brown coloration of interface Absence of terpenoids
AFM A reddish brown coloration at interface Presence of terpenoids

ANA A brown colored solution was formed Absence of steroids
ANM A light brown colored solution was formed Absence of steroidsAFH A reddish brown colored solution was formed Absence of steroids
AFA A reddish brown colored solution was formed Absence of steroids
AFM A dark brown solution formed that is stable on standing Absence of steroids
73
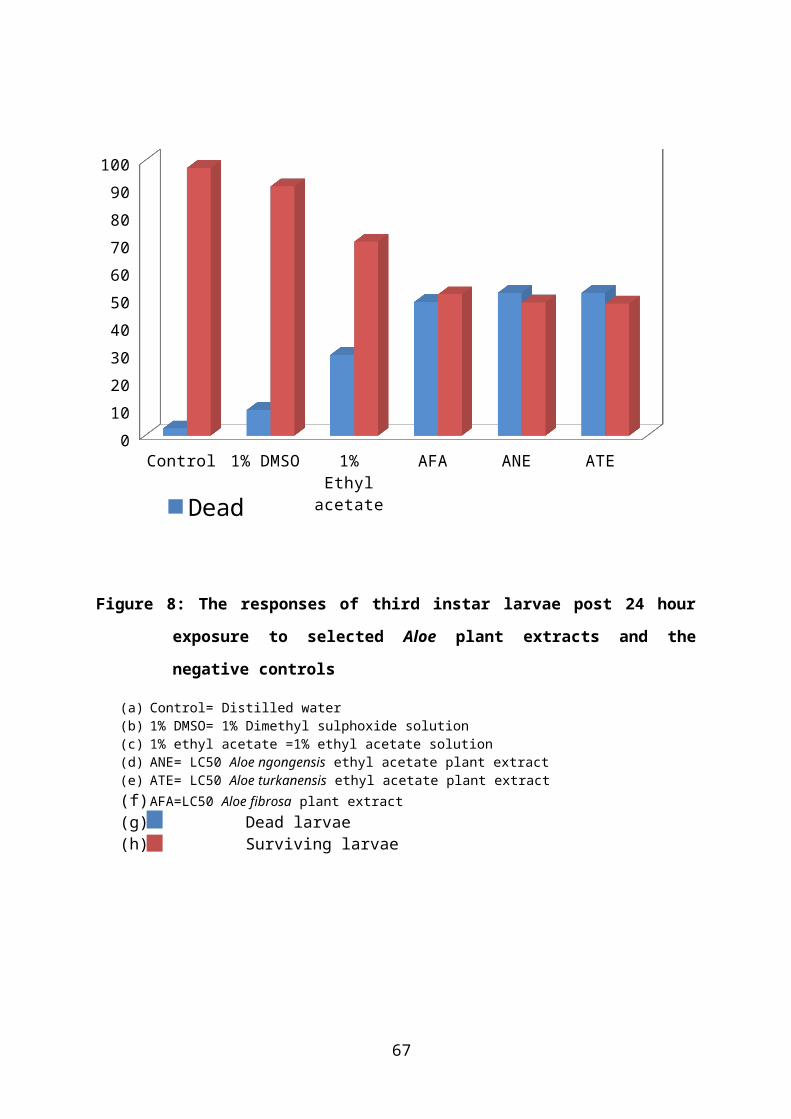
Appendix 19
Qualitative analysis of flavanoids of selected larvicidal Aloe plant extracts
74
Extract Observation Inference
ATE A yellow coloration was formed Presence of flavanoids
ANH A clear solution was formed Absence of flavanoids
ANE A yellow coloration was formed Presence of flavanoids
ANC A colorless solution Absence of flavanoids
ANA A yellow coloration was formed Presence of flavanoids
ANM A faint yellow coloration was observed Presence of flavanoids
AFH A faint yellow coloration was observed Presence of flavanoids
AFA A yellow coloration was formed Presence of flavanoids
AFM A yellow coloration was formed Presence of flavanoids
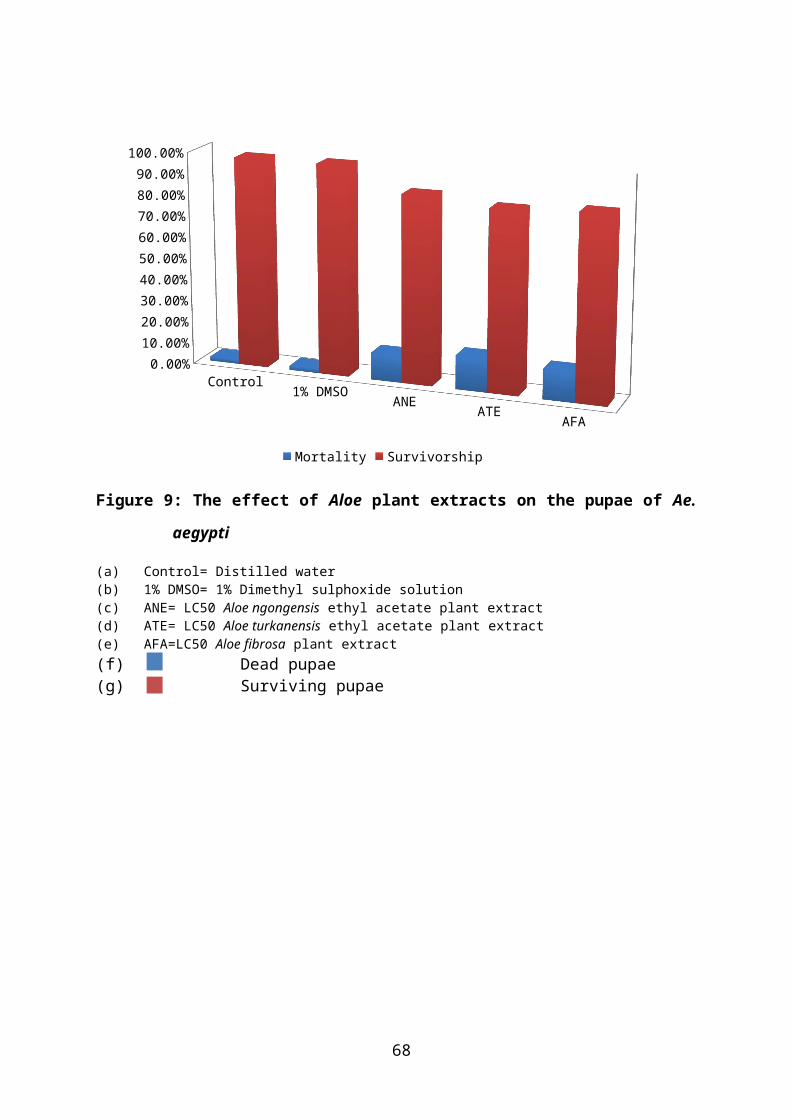
Appendix 20
Qualitative analysis of tannins and polyphenols of larvicidal selected Aloe plant extracts
Extract Observation Inference
ATE Formation of a brownish green solution Presence of tannins
ANH Formation of an orange solution Absence of tannins
ANE Formation of an orange solution Absence of tannins
ANC Formation of a light orange solution Absence of tannins
ANA Formation of a brown green solution Presence of tannins
ANM Formation of a blue black solution Presence of tannins
AFH Formation of a blue black solution Presence of tannins
AFA Formation of blue black solution Presence of tannins
AFM Formation of blue black solution Presence of tannins
75

Appendix 21
Phlobatannins analysis in selected Aloe plant extracts
Extract Observation Inference
ATE A clear solution forms Absence of phlobatannins
ANH A clear solution forms Absence of phlobatannins
ANE A clear solution forms Absence of phlobatannins
ANC A clear solution forms Absence of phlobatannins
ANA A clear solution forms Absence of phlobatannins
ANM A clear solution forms Absence of phlobatannins
AFH A clear solution forms Absence of phlobatannins
AFA A clear solution forms Absence of phlobatannins
AFM A clear solution forms Absence of phlobatannins
76

Appendix 22
Qualitative saponin test results for selected larvicidal Aloe plant extracts
77
Extract Observation Inference
ATE Formation of an emulsion Presence of saponins
ANH Formation of an emulsion Presence of saponins
ANE Formation of an emulsion Presence of saponins
ANC Formation of two immiscible layers Absence of saponins
ANA An emulsion was observed Presence of saponins
ANM Formation of two immiscible layers Absence of saponins
AFH Two immiscible layers were formed Absence of saponins
AFA Formation of an emulsion Presence of saponins
AFM Formation of an emulsion Presence of saponins

78