cytogenetic, fish and dna studies in 11 individuals from a family with two siblings with dup(21q)...
TRANSCRIPT

Hum Genet (1993) 92 : 127-132 human-- genetics �9 SpringerzVerlag 1993
Cytogenetic, FISH and DNA studies in 11 individuals from a family with two siblings with dup(21q) Down syndrome
O. Bartsch l, U. Krnig 2, M. B. Petersen 2, H. Poulsen 2, M. Mikkelsen 2, F. Palau 3, F. Prieto 3, E. Schwinger I
J Institut fur Humangenetik, Medizinische Universitfit, Ratzeburger Allee 160, D-23652 Ltibeck, Germany 2 Department of Medical Genetics, The John F. Kennedy Institute, DK-2600 Glostrup, Denmark 3 Unidad de Genrtica y Diagnrstico Prenatal, Hospital "La Fe", E-46009 Valencia, Spain
Received: 16 October 1992 / Revised: 21 January 1993
Abstract. A Spanish fami ly has p rev ious ly been de- scr ibed with two sibl ings with dup(21q) Down syndrome. The father has a normal karyotype . The mother has a mi- c rochromosome . Cytogenet ic , f luorescence in situ hy- br id iza t ion and D N A studies have now been carr ied out on the family. F ind ings include that the mother has three different ch romosome anomal ies , viz. (1) a ch romosome 22 with an unusual per icent romer ic region that contains a lphoid D N A from ch romosomes 21/13 and ch romosome 22, (2) an i soch romosome 21p in the frequent cell l ine and (3) an i soch romosome 21q in a rare second cell line. A poss ib le explana t ion is that the mother deve loped f rom a zygote with t r i somy 21 and that mitot ic error in ear ly de- ve lopment resul ted in the format ion o f two cell l ines with ka ryo types o f 47 ,XX,+i (21p) and 47,XX,+i(21q) , respec- tively. The unusual ch romosome 22 represents a hi therto undescr ibed ch romosome anomaly and one poss ib le ex- p lanat ion is a t rans locat ion of the short arms be tween c h r o m o s o m e s 21/13 and 22 in the ancest ry of the family. The re la t ionship be tween the unusual ch romosome 22 and the i soch romosome format ion in the mother is not known. However , all three ch romosome anomal ies involve the a lphoid D N A of ch romosome 21/13, indica t ing that this is not a chance finding.
Introduction
Prieto et al. (1981) descr ibed two sibl ings with dup(21q) Down syndrome whose mother had a b isa te l l i ted mi- c rochromosome . The cytogenet ic mechan i sms in the fam- i ly were diff icult to understand. Prieto et al. (1981) sug- ges ted that the m i c r o c h r o m o s o m e was a dup(21p) . As a contr ibut ion to the ongoing discuss ion whether the pres- ence o f b isa te l l i ted ex t r ach romosomes increase the spon- taneous meiot ic non-dis junct ion rate in man (e.g. Bartsch
Correspondence to: O. Bartsch, Institut for Klinische Genetik, Medizinische Akademie, Fetscherstrasse 74, D-01307 Dresden, Germany
et al. 1980; Ste inbach et al. 1983; A nne r rn et al. 1984), Prieto et al. (1981) asked whether the mic roc h ro mosome had induced the repeated "de novo" format ion of dup(21 q) ch romosomes in the maternal germline. To an- swer this quest ion, we studied 11 indiv iduals f rom the fami ly using ch romosome banding, f luorescence in situ hybr id iza t ion (FISH) and D N A analysis .
Materials and methods
Family OB02
Family OB02 (Fig. 1) is a three generation family previously de- scribed by Prieto et al. (1981). The siblings III4 and IIl5 have dup(21q) Down syndrome. With regard to the signs in the Jackson checklist for the clinical assessment of children with Down syn- drome (Jackson et al. 1986), IlI 4 had a permanently open mouth, a protruding tongue and a transverse palmar crease; III 5 had ab- normal teeth and a systolic murmur. Both sibs had oblique eye fis- sures, brushfield spots, a furrowed tongue, a short neck and in- curved 5th fingers. Additional findings included polydactyly in III 4; alopecia in III 5, and abnormal dermatoglyphics (palmar t" and Cummins index > 30), short stature, and an IQ of 40-55 (Wechsler and Santucci test) in both sibs. The other individuals from the family had no congenital anomalies.
Cytogenetic studies
Prieto et al. (1981) analysed standard GTG-banded chromosomes from cultured blood lymphocytes from nine individuals. The sib- lings with Down syndrome (III4 and III5) had karyotypes of 46,XY, -21,+dup(21q). The grandparents I1 and I2, the father II6 and sibling III1 had normal karyotypes. The mother II5 and sib- lings III 2 and III 3 had karyotypes of 47,XX,+mar. The marker was a bisatellited microchromosome with positive satellites for sil- ver staining of tile nucleolus organizer region (AgNOR) at both ends. Prieto et al. (1981) studied different tissues from the mother (2 blood cultures, bone marrow and skin fibroblasts) and analysed a total of 349 GTG-banded metaphases, all of which had a 47,XX,+mar karyotype. In 1982, the mother requested steriliza- tion. Following gonadal biopsies and cell culture, 50 GTG-banded metaphases were analysed from each ovary. All analysed cells bad a karyotype of 47,XX,+mar, i.e. there was no evidence for cytoge- netic mosaicism.

128
I I ~ [ ~ 2 ~ [ ~ ( ~ y ~ ~ ~ "
I I I ( ~ ~ ) 2 + 3 ~ 4 i ~ , I , I
]~: = unusual chromosome 22 O = normal karyotype
( ~ ) = microchromosome ,,"r = Down syndrolne
�9 = isochromosome 21q [ ] = not studied
Fig. 1. Pedigree of the OB02 family and karyotypes after FISH and DNA studies
For this study, heparinized blood was obtained from all individ- uals shown in the pedigree (Fig. 1), except from 12 (deceased), II 1 and I17 (not available). Standard metaphase spreads were prepared from cultured blood lymphocytes, and chromosome heteromor- phisms were studied after quinacrine banding and AgNOR staining (Mikkelsen et al. 1980). For FISH, the protocol of Bartsch and Schwinger (1991) was used with the modification that the DNA probes were biotinylated and the detection was carried out with avidin-fluorescein-isothiocyanate (FITC) and biotinylated anti- avidin. DNA probes used were the chromosome 21/13 alphoid probe LI.26 (Devilee et al. 1986), the chromosome 22/14 alphoid probe p22-94 (JOrgensen et al. 1988) and the satellite III probe for the short arm of chromosome 15 pHSR (Wang et al. 1984). Hy- bridization stringency was high [2 x SSC (1 • SSC = 150 mM NaCl/15 mM sodium citrate, pH 7.0), 60% formamide, 37 ~ C]. The alphoid DNA sequences from chromosomes 13 and 21 are more than 99% homologous (JOrgensen et al. 1988). Therefore, FISH with the chromosome 21 / 13 alphoid probe does not distinguish be- tween the centromeres from chromosomes 21 and 13.
Molecular studies
EDTA blood for DNA analysis was obtained from individuals I 1, II 2, II 3, II 5, II 6, III 1, II13, III 4 and III 5. DNA was extracted and molecular studies were carried out using DNA polymorphisms from 15 loci covering the entire long arm of chromosomes 21 (Table 1). The polymorphisms studied were microsatellites ana- lysed by polymerase chain reaction amplification and polyacry- lamide gel electrophoresis (Petersen et al. 1992). The microsatel- lites were the dinucleotide repeats of loci D21S215 (Warren et al. 1993), D21S120 (Burmeister et al. 1990), D21S16 (Cruts et al. 1992a), D21SI3E (Stinissen and Van Broeckhoven 1991), D21S192 (Van Camp et al. 1991), D21S145 (Cruts et al. 1992b), D21S214, D21S210, D21S216 (Warren et al. 1991), D21S211 (Weber et al. 1991 ), D21S 167 (Guo et al. 1990), D21S 156 (Lewis et al. 1990), HMG 14 (Petersen et al. 1991 a), D21S 171 (Petersen et al. 1991b) and the tetranucleotide repeat within the IFNAR gene (Mclnnis et al. 1991).
Results
Cytogenetic and FISH studies
Individual III 1 had a normal banded karyo type of 46,XX; h o w e v e r , after Fish with the a lphoid 21/13 probe, the metaphases showed f luorescent segments on three G-
group chromosomes and two D-group ch romosomes (Fig. 2A). This indicates the presence of an unusual chromo- some 22 with a lphoid D N A from ch romosome 21 or 13. F ISH with the ch romosome 22/14 a lphoid D N A probe re- suited in the usual staining o f two G-group ch romosomes and two D-group ch romosomes (Fig .2B) . Thus the un- usual ch romosome 22 also carries a normal-s ized alphoid D N A segment from ch romosome 22. The f luorescent sig- nal with the a lphoid 21/13 probe on the unusual chromo- some 22 represents a reproducib le f inding that cannot be expla ined by incidental cross-hybr idizat ion. Med ium str ingency hybr id iza t ion condi t ions (i.e. approximate ly 2 x SSC, 50% formamide , 37~ with this probe usual ly re- suits in the addit ional staining of ch romosomes 18 and 20 (Kolvraa et al. 1991); when the s tr ingency is further de- creased, all ch romosomes with D N A from the a lphoid su- per fami ly 2 are stained (chromosomes 2, 4, 8, 9, 13, 14, 15, 18, 20, 21 and 22; Alexandrov et al. 1991).
The unusual ch romosome 22 was present in 114 (not shown), II 5 (Figs. 2C, D), III 1 (Fig. 2A) and III 5 (Fig. 2E), but not in I 1 (not shown). QFQ-banding and AgNOR- staining indicated that the ch romosome 22 anomaly segre- gated with the ch romosome 22 homologue " f" (Fig. 3), which was inheri ted from the grandfather 12.
The mic roch romosome was found in II 5, III 2 and III 3. It hybr id ized with the ch romosome 21/13 a lphoid D N A probe (Fig. 2C), but not with the a lphoid 22/14 probe and the ch romosome 15 short arm probe (not shown). Chro- mosomal he te romorphisms showed that both arms of the mic roch romosome were der ived from the ch romosome 21 homologue "c" f rom the g randmother I 1 (Fig. 3). It could
Fig.2 A-F. Five different metaphases after FISH. Arrowheads, Fluorescent segments on G-group chromosomes; small arrow, isochromosome 21 p; large arrows, isochromosomes 21 q. Fluores- cent segments on D-group chromosomes are not indicated. A, B Two metaphases from the normal sibling III I. A Following hy- bridization with the chromosome 21/13 alphoid DNA probe, fluo- rescent segments are on three G-group chromosomes (chromo- somes 21, unusual chromosome 22) and two D-group chromo- somes (chromosomes 13). B With the chromosome 22/14 alphoid DNA probe, fluorescent segments are found on two G-group chro- mosomes (chromosomes 22) and two D-group chromosomes (chromosomes 14). C, D Two metaphases from the mother II 5, hybridized with the chromosome 21/13 alphoid DNA probe. C The usual metaphase from II 5, in which fluorescent segments are found on the microchromosome (isochromosome 21p), three G- group chromosomes (chromosomes 21, unusual chromosome 22) and two D-group chromosomes (chromosomes 13). D Unique metaphase from I15; the fluorescent segments lie on one metacen- tric chromosome (possibly an isochromosome 21q), one G-group chromosome (possibly the unusual chromosome 22), and two D- group chromosomes (chromosomes 13). E Metaphase from sibling II1 5 with dup(21q) Down syndrome, after hybridization with the chromosome 21/13 alphoid DNA probe: signals are lk)und on the isochromosome 21 q, two G-group chromosomes (chromosome 21, unusual chromosome 22), and two D-group chromosomes (chro- mosomes 13). F Same metaphase as in D but photographed with a Leitz N2 filter combination (excitation 530-560 nm) to enable better visualization of chromosome contours. Note the similar ap- pearance of the possible isochromosome 21q in D, F and the i(21q) in E. The metaphase in D, F was thoroughly examined under the microscope and contained no microchromosome

129
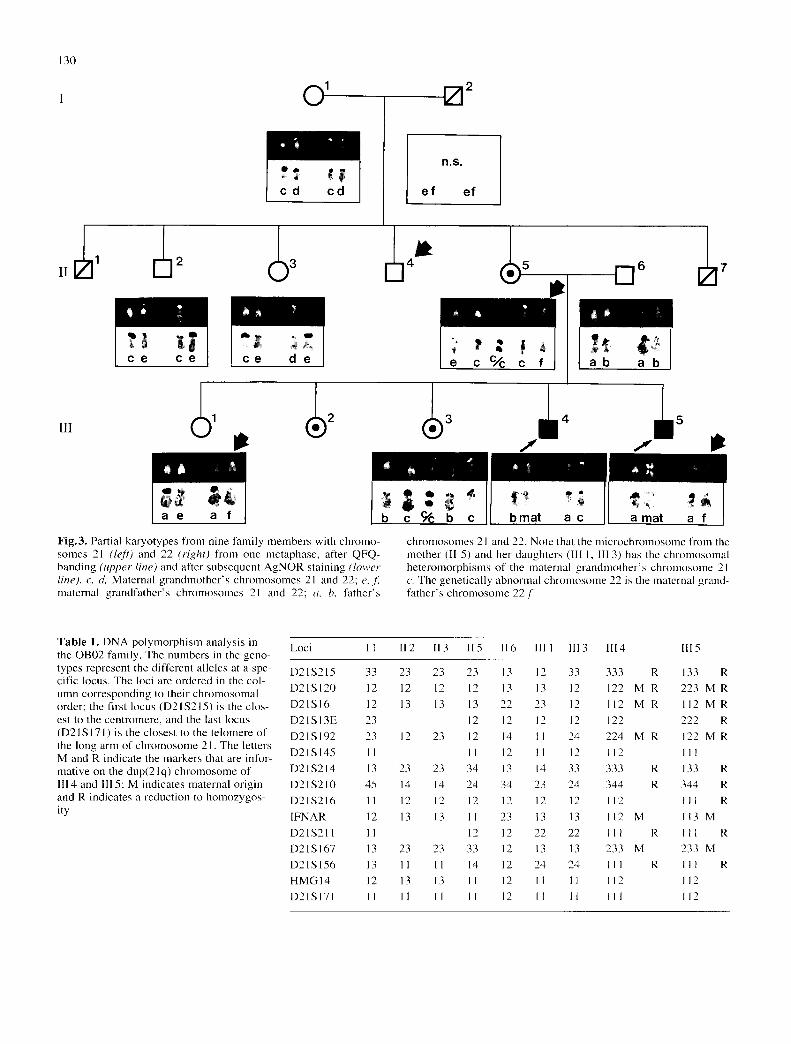
130
01 [7] 2
n.s, I el ef
ii~1 ~2 ~3 ~4"
III
Fig.3. Partial karyotypes from nine family members with chromo- somes 21 (left) and 22 (right) from one metaphase, after QFQ- banding (upper line) and after subsequent AgNOR staining (lower line), c, d, Maternal grandmother's chromosomes 21 and 22; e, f maternal grandfather's chromosomes 21 and 22; a, b, father's
chromosomes 21 and 22. Note that the microchromosmne from the mother (II 5) and her daughters (Ill 1, Ill 3) has the chromosomal heteromorphisms of the maternal grandmother's chromosome 21 c. The genetically abnormal chromosmne 22 is the maternal grand- father's chromosmne 22.[
Table 1. DNA polymorphism analysis in the OB02 family. The numbers in the geno- types represent the different alleles at a spe- cific locus. The loci are ordered in the col- umn corresponding to their chromosomal order; the first locus (D21S215) is the clos- est to the centromere, and the last locus (D21S171) is the closest to the telomere of the long arm of chromosome 21. The letters M and R indicate the markers that are infor- mative on the dup(21q) chromosome of lIl 4 and III 5; M indicates maternal origin and R indicates a reduction to homozygos- ity
Loci I 1 I12 II3 II5 II6 I111 1113 1114 I115
D21S215 33 23 23 23 13 12 33 333 R
D21SI20 12 12 12 12 13 13 12 122 M R
D21S16 12 13 13 13 22 23 12 112 M R
D21SI3E 23 12 12 12 12 122
D21S192 23 12 23 12 14 11 24 224 M R
D21S145 11 11 12 11 12 112
D21S214 13 23 23 34 13 14 33 333 R
D21S210 45 14 14 24 34 23 24 344 R
D21S216 11 12 12 12 12 12 12 112
IFNAR 12 13 13 II 23 13 13 112 M
D21S211 11 12 12 22 22 111 R
D21S167 13 23 23 33 12 13 13 233 M
D21S156 13 11 11 14 12 24 24 I11 R
HMGI4 12 13 13 11 12 I1 11 112
D21S171 11 11 I1 11 12 I1 11 111
133 R
223 M R
l l 2 M R
222 R
122 M R
l l l
133 R
344 R
I l l R 113 M
I11 R
233 M
111 R
112
112

be excluded that they were derived from a chromosome 13 (not shown). This indicates that the microchromosome is an isochromosome 21p.
Out of 400 metaphases analysed from II 5 with the alphoid 21/13 probe, 399 had a karyotype of 47,XX,+mi- crochromosome. One metaphase contained fluorescent segments on an E-group-sized metacentric chromosome, one G-group chromosome (out of two G-group chromo- somes present) and no microchromosome (Fig. 2D); how- ever, a full chromosome count from the metaphase was not possible. The stained metacentric chromosome closely resembled the dup(2 l q) from III 5 (Fig. 2E). This supports the hypothesis that II 5 has a rare second cell line with dup(21q).
Normal findings were obtained with the chromosome 21/13 alphoid DNA probe in I 1, I2, II3 and II6, and with the chromosome 22/14 alphoid DNA probe and the chro- mosome 15 short arm DNA probe in all individuals studies.
Molecular studies
DNA results are summarized in Table 1. The dup(21q) chromosomes from the two siblings with Down syndrome (III4 and III5) have identical alleles at all 15 loci. Five DNA polymorphisms clearly indicate the maternal origin of the dup(21q). Reduction to homozygosity was found for eight and ten DNA polymorphisms, respectively, indi- cating that the dup(21q) is an isochromosome 21q. The grandmother (I 1) has all alleles of the i(21q), which agrees with the cytogenetic finding that the i(21p) is deriv- ed from a grandmaternal chromosome 21 (homologue "c").
Discussion
We found three different chromosome anomalies in the family. They must have originated from at least two dif- ferent events. The unusual chromosome 22 is the oldest chromosome anomaly in the family. It was probably in- herited from I2, as indicated by its presence in II 4 and II 5, its absence in I 1, the quinacrine fluorescence hetero- morphisms and the AgNOR staining. The findings after FISH suggest that two different alphoid DNA domains are present on the unusual chromosome 22, namely, from chromosome 21 (or 13) and chromosome 22. Similar findings have not been described previously. We studied 104 unrelated individuals with the chromosome 21/13 alphoid DNA probe [including four probands with dup(21q) Down syndrome] and found no other unusual chromosome 22. Thus, this must be a rare finding. A pos- sible explanation is a reciprocal translocation of short arms and parts of the pericentric heterochromatin between a chromosome 21 or 13 and a chromosome 22 in the an- cestry of the family. DNA sequences from the short arm of chromosome 15 are sometimes found on chromosome 14 (Smeets et al. 1991). This can be explained by a chromo- somal translocation, but also by other means, e.g. the se- lective amplification of DNA sequences common to 15p and 14p (Stergianou et al. 1992). However, the findings of Smeets et al. (1991) and Stergianou et al. (1992) differ from our findings in that they used a short arm satellite III DNA probe where we used an alphoid DNA probe. The
131
alphoid DNA represents a part of the centromere with sig- nificant functional importance (Haaf et al. 1992).
Prieto et al. (1981) suggested that the mother's mi- crochromosome may be an isochromosome 21 of short arms. Our observations support their assumption. The DNA results indicate that the dup(21q) chromosomes from the siblings with Down syndrome are also isochromo- somes 21, but of the long arms. This agrees with the find- ing of Antonarakis et al. (1990) that most "de novo" dup(21 q) chromosomes from patients with Down syndrome seem to be isochromosomes and not translocations. We have observed one possible i(21q) cell from the mother's blood (Fig. 2D, F). This supports the hypothesis that the mother has a rare second cell line with i(21q), although of course, mosaicism with 1/400 i(21q) cells is not convinc- ing evidence for real mosaicism. However, the DNA find- ings that the isochromosomes 21q from the two siblings with Down syndrome are identical and of maternal origin add support to the conclusion that II 5 is a cytogenetic mosaic. In the literature, parental mosaicism for dup(21q) was reported in six out of 11 families with two siblings with dup(21q) Down syndrome (Steinberg et al. 1984; Hall t985; Priest et al. 1988). A low level of cytogenetic mosaicism was recently described in a 27-year-old woman of normal intelligence with one dup(21q) Down syndrome child (Croci and Franchi 1991); in this case, the dup(21q) was present in 11 out of 2659 metaphases.
It seems probable that the mother has two cell lines with i(21p) and i(21q), respectively, resulting from one mitotic error. This error may have taken place in the mother's zygote, or shortly thereafter. It can be excluded that the mother developed from a 46,XX zygote [which maldivided into two daughter cells with karyotypes of 47,XX,+i(21p) and 45,XX,-21,-21,+i(21q)], because in that case, both daughter cell lines would have to be isodi- somic for 21q; this disagrees with the DNA findings. Moreover, this developmental possibility would have re- quired two different mitotic errors in one cell division, namely non-disjunction of one chromosome 21 and isochromosome formation of the other chromosome 21.
The findings suggest that the mother's zygote had a 47,XX,+21 karyotype after a meiosis II error in the grand- mother's gametogenesis, and that it contained the chro- mosome 21 homologues "c", "c", and "e". Isochromo- some formation from one chromosome 21 "c" at the zy- gote stage or shortly thereafter would then result in daughter cells with karyotypes of 47,XX,+i(21p) and 47,XX,+i(21q). In this case, the cells with i(21p) will be near-normal, whereas the cells with i(21q) will contain four copies of 21q. This may explain the paucity of i(21q) cells in the mother's blood and skin.
Of course, we cannot know whether the isochromo- some formation and the unusual chromosome 22 are re- lated to each other. It could be a coincidence. However, the presence of an underlying mechanism must be consid- ered, because the chance occurrence of two rare cytoge- netic anomalies in one family is per s e a highly unusual event and, moreover, both anomalies involve the alphoid 21/13 DNA. The unusual chromosome 22 was transmitted to the mother from the grandfather, and thus pre-existed in the cell in which the isochromosomes 21p and 21q were

132
f o r m e d . N e v e r t h e l e s s , it s e e m s u n l i k e l y tha t the t wo di f - f e r e n t a l p h o i d D N A d o m a i n s o n the u n u s u a l c h r o m o s o m e 22 w e r e a c a u s a t i v e f a c t o r in the p r o c e s s o f i s o c h r o m o - s o m e f o r m a t i o n . O t h e r o b s e r v a t i o n s are n e e d e d to de te r - m i n e the s i g n i f i c a n c e o f n o r m a l - a p p e a r i n g c h r o m o s o m e s
w i t h " c h i m e r i c " c e n t r o m e r e s , i.e. c e n t r o m e r e s c o n t a i n i n g D N A d o m a i n s f r o m t w o d i f f e r e n t c h r o m o s o m e s .
Acknowledgements. We wish to thank Peter Devilee for providing probe L 1.26, Arne Leth Bak for probe p22-94, J. L. Hamerton for probe pHSR, and Andreas Gal for helpful comments. The work in the laboratory of M. B. P. and M. M. was funded by grants from the Danish Human Genome Research Programme and the E. C. Human Genome Analysis Programme. The activities of E. P. were supported by the Deutscher Akademischer Austauschdienst, "Ac- ciones lntegradas Hispano-Alemanas".
References
Alexandrov IA, Mashkova TD, Akopian TA, Medvedev LI, Kissilev LL, Mitkevich SP, Yurov YB (1991) Chromosome- specific alpha satellites: two distinct families on human chro- mosome 18. Genomics 11 : 15-23
Annerdn G, Wahlstr6m J, Tommerup N (1984) Marker chromo- somes in parents to children with Down's syndrome. Clin Genet 25 :140-147
Antonarakis SE, Adelsberger PA, Petersen MB, Binkert F, Schinzel AA (1990) Analysis of DNA polymorphisms suggests that most de novo dup(21q) chromosomes in patients with Down syndrome are isochromosomes and not translocations. Am J Hum Genet 47 : 968-972
Bartsch O, Schwinger E (1991) A simplified protocol for fluores- cence in situ hybridization with repetitive DNA probes and its use in clinical cytogenetics. Clin Genet 40 :47 -56
Bartsch HD, Azevedo Moreira LM de, ROhrborn G, Claussen U, Gebauer HJ (1980) Possible origin of a small bisatellited addi- tional chromosome. Hum Genet 54 : 319-322
Burmeister M, Cox DR, Myers RM (1990) Dinucleotide repeat polymorphism located at D21S 120. Nucleic Acids Res 18:4969
Croci G, Franchi F (1991) Parental mosaicism in de novo translo- cation (21 q21 q) Down's syndrome. J Med Genet 28:502
Cruts M, Backhovens H, Van Broeckhoven C (1992a) Dinu- cleotide repeat polymorphism at the D21S16 locus. Nucleic Acids Res 20:1159
Cruts M, Backhovens H, Van Broeckhoven C (1992b) Dinu- cleotide repeat polymorphism at the D21S145 locus. Nucleic Acids Res 20:1159
Devilee P, Slagboom P, Cornelisse CJ, Pearson PL (1986) Se- quence heterogeneity within the human alphoid repetitive DNA family. Nucleic Acids Res 14:2059-2074
Guo Z, Sharma V, Patterson D, Litt M (1990) TG repeat polymor- phism at the D21S 167 locus. Nucleic Acids Res 18 : 4967
Haaf T, Warburton PE, Willard HF (1992) Integration of human alpha-satellite DNA into simian chromosomes: centromere pro- tein binding and disruption of normal chromosome segregation. Cell 70 : 681-696
Hall BD (1985) Recurrence risk in de novo 21q21q translocation Down syndrome. Am J Med Genet 22 : 417-4 18
Jackson JF, North ER III, Thomas JG (1976) Clinical diagnosis of Down's syndrome. Clin Genet 9 :483-487
JCwgensen AL, Kolvraa S, Jones C, Bak AL (1988) A subfamily of alphoid repetitive DNA shared by the NOR-bearing human chromosomes 14 and 22. Genomics 3 :100-109
Kolvraa S, Koch J, Gregersen N, Jensen PKA, JOrgensen AL, Pe- tersen KB, Rasmussen K, Bolund L ( 1991 ) Application of fluo- rescence in situ hybridization techniques in clinical genetics: use of two alphoid repeat probes detecting the centromeres of
chromosomes 13 and 21 or chromosomes 14 and 22, respec- tively. Clin Genet 39 :278-286
Lewis JG, Weber J L Petersen MB, Slaugenhaupt SA, Kwitek A, May PE, Warren AC, Chakravarti A, Antonarakis SE (1990) Linkage mapping of the highly informative DNA marker D21S156 to human chromosome 21 using a polymorphic GT dinucleotide repeat. Genomics 8 :400-402
Mclnnis MG, Lufalla G, Slaugenhaupt SA, Petersen MB, Uze G. Chakravarti A, Antonarakis SE (1991) Linkage mapping of high- ly informative DNA polymorphisms within the human interferon- alpha receptor gene on chromosome 21. Genomics 11:573 576
Mikkelsen M, Poulsen H, Grinsted J, Lange A (1980) Non-dis- junction in trisomy 21: study of chromosomal heteromorphisms in 110 families. Ann Hum Genet 44:17 28
Petersen MB, Schinzel AA, Binkert F, Tranebjaerg L, Mikkelsen M, Collins FA, Economou EP, Antonarakis SE (1991a) Use of short sequence repeat DNA polymorphisms after PCR amplifi- cation to detect the parental origin of the additional chromo- some 21 in Down syndrome. Am J Hum Genet 48:65 71
Petersen MB, Weber JL, Slaugenhaupt SA, Kwitek AE, Mchmis MG, Chakravarti A, Antonarakis SE (1991b) Linkage mapping of D21S171 to the distal long arm of human chromosome 21 using a polymorphic (AC)n dinucleotide repeat. Hum Genet 87:401-404
Petersen MB, Frantzen M, Antonarakis SE, Warren AC, Van Broeckhoven C, Chakravarti A, Cox TK, Lund C, Olsen B. Poulsen H, Sand A, Tommerup N, Mikkelsen M (1992) Com- parative study of microsatellite and cytogenetic markers for de- tecting the origin of the nondisjoined chromosome 21 in Down syndrome. Am J Hum Genet 51 :516 525
Priest JH, Blackston RD, Pearse LA, Warren ST (1988) Molecular evidence for true isochromosome 21. Hum Genet 81 : 1 3
Prieto F, Badia L, Ribes C, Medina VH (1981) Trisomie 21 par translocation 21/21 chez deux fils d 'une m6re avec un mi- crochromosome surnum6raire. Ann Gdndt (Paris) 24:117 119
Smeets DFCM, Merckx GFM, Hopman AHM (1991) Frequent oc- currence of translocations of the short arm of chromosome 15 to other D-group chromosomes. Hum Genet 87 :45-48
Steinbach P, Djalali M, Hansmann I, Kattner E, Meisel-Stosiek M, Probeck HD, Schmidt A, Wolf M (1983) The genetic signifi- cance of accessory bisatellited marker chromosomes. Hum Genet 65 :155-164
Steinberg C, Zackai EH, Eupnu DL, Mennuti MT, Emanuel BS (1984) Recurrence risk fl)r de nov() 21q21q translocation Down syndrome: a study of 112 families. Am J Med Genet 17: 523-530
Stergianou K, Gould CP, Waters JJ, Hultdn M (1992) High popu- lation incidence of the 15p marker D I5ZI mapping to the short arm of one homologue 14. Hum Genet 88 : 364
Stinissen P, Van Broeckhoven C (1991) A new (CA)n repeat poly- morphism at the D21S 13E locus. Nucleic Acids Res 19 : 5089
Van Camp G, Bakchovens H, Cruts M, Wehnert A, Van Hul W, Stinissen P, Van Broeckhoven C (1991) Identification of chro- mosome 21 DNA polymorphisms for genetic studies in AIz- helmet ' s disease and Down syndrome. Hum Genet 87:649-653
Wang HS, Riddell DC, Donald LJ, Cameron EC, Tonogai J, Holden JJA, Higgins M, Stromas I, White BN, Hamerton JL (1984) Mapping of a repetitive 1.8 kb Kpnl sequence to the short ann of chromosome 15. Cytogenet Cell Genet 37 : 601- 602
Warren AC, Mclnnis MG, Kalaitsidaki M, Adelsberger P, Lewis JG, Antonarakis SE (1991) Dinucleotide repeat polymorphisms as linkage markers for chromosome 21. Cytogenet Cell Genet 58 : 2042
Warren AC, Petersen MB, Van Hul W, Mclnnis MG, Van Broeck- hoven C, Cox TK, Chakravarti A, Antonarakis SE (1993) D21S215 is a (GT) n polymorphic marker close to centromeric alphoid sequences on chromosome 21. Genomics 13:1365-1367
Weber JL, Kwitek AE, May PE, Wilkie PJ, Decker RA ( 1991 ) Mi- crosatellite DNA polymorphisms. Cytogenet Cell Genet 58: 1864