curcumin suppresses nadph-oxidase and mcp-1 …curcumin suppresses nadph-oxidase and mcp-1 mrna...
TRANSCRIPT

2010年度 修士論文
クルクミンはダウンヒル運動後のマウスの
骨格筋における NADPH-oxidase および
MCP-1 の遺伝子発現を抑制する
Curcumin suppresses NADPH-oxidase and
MCP-1 mRNA expression in skeletal muscle
of mice following downhill running
早稲田大学大学院 スポーツ科学研究科
スポーツ科学専攻 スポーツ医科学研究領域
5009A025-9
加藤 孝基
Kato, Koki
研究指導教員: 鈴木 克彦 准教授

目次
1) 緒 言 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1
2) 方 法 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 4
2-1) 実 験 動 物 と 飼 育 条 件
2-2) ダ ウ ン ヒ ル 走 行 運 動 負 荷
2-3) ク ル ク ミ ン 投 与
2-4) 採 血 お よ び 筋 組 織 採 取
2-5) ク レ ア チ ン キ ナ ー ゼ 活 性
2-6) real-time quantitative PCR
2-7) タ ン パ ク 質 濃 度 測 定
2-8) Hydrogen peroxide 濃 度 測 定
2-9) 統 計 処 理
3) 結 果 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 8
4) 考 察 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 21
5) 結 論 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 24
6) 謝 辞 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 25
7) 参 考 文 献 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 26
8) お わ り に - 今 後 の 展 望 ― ・ ・ ・ ・ ・ ・ ・ 31

1
1 ) 緒 言
生 体 には損 傷 に対 する防 御 機 構 が存 在 し 、組 織 の恒 常 性 を保 ってい
る。骨 格 筋 においても 、運 動 や圧 迫 ス トレス 、外 傷 に伴 う組 織 の損 傷 が引
き 起 こ さ れ る と 、 組 織 を 再 生 す る 機 構 が は た ら く 。 し か し な が ら 、 過 度 な 運
動 によ り 防 御 機 構 の バランスが崩 れる と 、組 織 再 生 の 遅 れ、機 能 障 害 な
どを引 き起 こす 。ダウンヒル運 動 などにみられる伸 張 性 筋 収 縮 は、筋 繊 維
の過 剰 な損 傷 や炎 症 反 応 を伴 うことが知 られている [ 1 , 2 ] 。これまでに実
験 動 物 を 用 いた 研 究 よ り 、 ダ ウン ヒ ル 運 動 は 炎 症 性 サ イ ト カ イン お よび 活
性 酸 素 の 産 生 を伴 う こ とが報 告 されて いる [ 3 , 4 ] 。 近 年 、 こ れ らの 炎 症
反 応 には好 中 球 やマクロファージなどの免 疫 細 胞 が関 与 していることが明
らかになった [ 5 ] 。
骨 格 筋 損 傷 後 の 炎 症 反 応 は 、 損 傷 し た 骨 格 筋 組 織 で の t u m o r
n e c r o s i s f a c t o r ( T N F ) - や i n t e r l e u k i n ( I L ) - 1 β 、 I L - 6 などの炎 症 性
サ イ ト カ イン の 産 生 に よ り 惹 起 さ れ る 。実 際 に 、マ ウス 前 脛 骨 筋 の カ ル デ ィ
オ トキシン誘 発 性 筋 損 傷 直 後 に、組 織 内 で T N F - α 、 と I L - 1 β の遺 伝 子
発 現 が増 加 するこ とが報 告 されている [ 6 ] 。 また 、ラ ッ ト を用 いた研 究 では、
ダウンヒル運 動 直 後 に外 側 広 筋 および腓 腹 筋 で T N F - α の遺 伝 子 発 現
が増 加 することが報 告 されている [ 7 ] 。 さ らに 、運 動 2 4 時 間 後 にマウス腓
腹 筋 の I L - 1 β タンパク質 濃 度 が増 加 することが報 告 されている [ 8 ] 。また、
骨 格 筋 損 傷 時 には、組 織 内 に好 中 球 、マクロファージなどの白 血 球 が浸
潤 することが知 られている [ 9 ] 。白 血 球 の浸 潤 は、 T N F - α や I L - 1 β によ り E
セ レ ク チ ン な ど の 細 胞 接 着 分 子 の 発 現 が 増 加 し 、 I L - 8 、 m o n o c y t e
c h e m o a t t r a c t a n t p r o t e i n ( M C P ) - 1 などのケモカインが損 傷 後 に増 加 す
ることで誘 導 される [ 6 , 1 0 , 1 1 ] 。最 も早 く組 織 に浸 潤 する白 血 球 は好 中
球 である。実 際 に、ダウンヒル誘 導 性 筋 損 傷 4 5 分 後 に損 傷 部 位 に浸 潤

2
するこ とが報 告 され ている [ 9 ] 。ケモカ インで誘 導 された好 中 球 は活 性 化
され、損 傷 組 織 に浸 潤 し貪 食 を行 う [ 1 2 ] 。 また 、筋 損 傷 時 には、 M C P - 1
の 作 用 に よ り マ ク ロ フ ァ ー ジ の 浸 潤 が 誘 導 さ れ る こ と が 知 ら れ て い る [ 1 3 ,
1 4 ] 。組 織 に浸 潤 したマク ロフ ァージは、損 傷 部 位 の貪 食 を行 うだけでな く 、
炎 症 の 鎮 静 化 と筋 サ テラ イ ト細 胞 を含 む 筋 幹 細 胞 の 活 性 化 も 引 き 起 こ
すこ とが考 え られている [ 1 5 ] 。一 方 で 、好 中 球 やマク ロフ ァージの形 質 膜
に存 在 し、酸 化 酵 素 としてはたら く N A D P H - o x i d a s e によ り活 性 酸 素 が産
生 される [ 1 6 , 1 7 ] 。 N A D P H - o x i d a s e は、好 中 球 やマクロファージが貪 食
を行 った 際 に 活 性 化 され 、電 子 供 与 体 として 酸 素 を 1 電 子 還 元 してスー
パーオキシド ( ・ O 2- ) を産 生 する [ 1 8 ] 。 また 、生 成 された ・ O 2
- は、過 酸 化
水 素 ( h y d r o g e n p e r o x i d e : H 2 O 2 ) に代 謝 される。実 際 に、 2時 間 のダウ
ンヒル運 動 後 、ラッ トの腓 腹 筋 およびヒラメ筋 の H 2 O 2 濃 度 が上 昇 すること
が報 告 されている [ 7 ] 。また、 H 2 O 2 は N F - κ B などのシグナル伝 達 経 路 を
活 性 化 し、それらの伝 達 経 路 は I L - 1 、 I L - 6 などの炎 症 性 サイトカインを
誘 導 するこ とが報 告 されている [ 1 9 ] 。生 体 には、運 動 および骨 格 筋 損 傷
によ り生 じた活 性 酸 素 を生 成 する系 と消 去 する系 が存 在 しバランスを保 っ
ているが、活 性 酸 素 生 成 系 が消 去 系 の能 力 を超 えると酸 化 ス ト レス状 態
とな り 、デオキシ リボ核 酸 ( D N A ) 、細 胞 膜 などの脂 質 およびタンパク質 を
傷 害 し 、 細 胞 死 を 引 き 起 こ す こ と が 考 え ら れ る [ 2 0 ] 。 ま た 、 過 剰 な 活 性
酸 素 によ り運 動 パフォーマンスが低 下 することも知 られている [ 2 1 ] 。
天 然 ウコンの主 成 分 であるクルク ミンは様 々な疾 患 モデルにおいて抗 炎
症 作 用 を示 すこ とが明 らかにされている [ 2 2 , 2 3 ] 。 クルク ミ ンの作 用 機 序
について、これまでに N F - B の活 性 を抑 制 することが報 告 されている [ 2 4 ,
2 5 ] 。 したがって、 N F - B シグナル伝 達 経 路 の活 性 抑 制 を介 して T N F - 、
I L - 1 β などの炎 症 性 サイトカインを抑 制 すると考 えられる。また、クルク ミンは

3
H 2 O 2 などの活 性 酸 素 の産 生 を阻 害 することが報 告 されている [ 2 6 ] 。クル
ク ミ ン は抗 炎 症 作 用 お よび 抗 酸 化 作 用 を示 す こ とか ら 、筋 損 傷 を伴 う 運
動 においても炎 症 状 態 を抑 制 する可 能 性 が考 えられる。実 際 に、マウスを
用 いた 動 物 実 験 で 、 ク ル ク ミ ン 投 与 に よ り ダ ウ ン ヒ ル 運 動 誘 発 性 筋 損 傷
後 の骨 格 筋 の I L - 1 β 、 I L - 6 、 T N F - α タンパク質 濃 度 が減 少 することが報
告 され て いる [ 8 ] 。 し か しな がら 、 クル ク ミ ン摂 取 に よ るダウン ヒル運 動 時 の
筋 組 織 での炎 症 反 応 抑 制 の作 用 機 序 に関 しては十 分 に解 明 されていな
い。クルク ミンは、 p h o r b o l m y r i s t a t e a c e t a t e ( P M A ) 誘 導 性 M C P - 1 の産
生 を 抑 制 す る こ と が 報 告 さ れ て い る [ 2 7 ] 。 し た が っ て 、 ク ル ク ミ ン に よ る 筋
損 傷 時 の炎 症 反 応 の改 善 には、マクロフ ァージ浸 潤 の抑 制 が関 与 してい
る可 能 性 が考 えられる。さ らに 、クルク ミンの抗 酸 化 作 用 から好 中 球 やマク
ロファージの活 性 酸 素 産 生 が抑 制 される可 能 性 も考 えられる。
そ こで 、本 研 究 で は 、高 吸 収 の クルク ミ ンを経 口 で 単 回 投 与 し 、マ ウス
のダウンヒル走 行 運 動 誘 導 性 筋 損 傷 後 の骨 格 筋 組 織 の炎 症 反 応 、酸
化 ス トレスおよび免 疫 細 胞 浸 潤 に及 ぼす影 響 を明 らかにするこ とを目 的 と
した。

4
2 ) 方 法
1 . 実 験 動 物 と飼 育 条 件
本 実 験 では 1 0 週 齢 の C 5 7 B / 6 J 雄 マウス ( 紀 和 実 験 動 物 研 究 所 ,
W a k a y a m a , J a p a n ) を用 いた。室 温 2 1 ℃ 、湿 度 3 5 %、 2 1 : 0 0 - 9 : 0 0 を暗
期 に 9 : 0 0 - 2 1 : 0 0 を明 期 に設 定 した飼 育 室 において、ゲージ内 で飼 育 した。
M F ( オリエンタル酵 母 , T o k y o , J a p a n ) の飼 料 を使 用 し、飲 水 は水 道 水
を用 い、 と もに自 由 摂 取 とした 。マウスは 、無 作 為 に安 静 +クルク ミン非 投
与 群 ( n = 1 2 ) 、安 静 +クルク ミン投 与 群 ( n = 1 2 ) 、運 動 +クルク ミン非 投
与 群 ( n = 1 4 ) 、運 動 +クルク ミン投 与 群 ( n = 1 4 ) の 4 群 に分 けた。
な お 、 本 実 験 は 早 稲 田 大 学 動 物 実 験 委 員 会 の 承 認 を 得 て 行 っ た
( 1 0 K 0 0 1 ) 。
2 . ダウンヒル走 行 運 動 負 荷
ダウンヒル走 行 運 動 負 荷 には、小 動 物 用 運 動 負 荷 装 置 K N - 7 3 ト レ ッ
ド ・ ミ ル ( 夏 目 製 作 所 , T o k y o , J a p a n ) を 用 い た 。 ま ず 運 動 開 始 か ら
1 2 . 5 - 1 7 . 5 m / m i n , 傾 斜 - 1 4 ° の条 件 で 1 5 分 間 ウォーミングアップを負 荷
した。その後 、速 度 2 2 m / m i n , 傾 斜 - 1 4 ° の条 件 で 1 3 5 分 間 ダウンヒル走
行 運 動 を行 い、合 計 1 5 0 分 間 の走 行 運 動 を負 荷 した。
3 . クルク ミン投 与
運 動 +クルク ミン投 与 群 には運 動 終 了 直 後 に麻 酔 下 にてマウス 1匹 あ
た りクルク ミン成 分 3 m g を含 む P B S 溶 液 を 2 0 0 μ l 経 口 投 与 した。運 動 +
クルク ミン非 投 与 群 には運 動 終 了 直 後 に麻 酔 下 P B S を 2 0 0 μ l 経 口 投
与 し た 。 な お 、 経 口 投 与 に は マ ウ ス 用 ゾ ン デ ( 夏 目 製 作 所 , T o k y o ,
J a p a n ) を使 用 した 。 また 、安 静 群 も同 様 の方 法 で投 与 した。麻 酔 は実 験

5
小 動 物 用 ガス麻 酔 システム ( D S ファーマバイオメディカル , O s a k a , J a p a n )
を使 用 して、イソフルランフォーレン吸 入 麻 酔 液 ( アボッ トジャパン , T o k y o ,
J a p a n ) を用 いた。麻 酔 は、流 速 0 . 8 L / m i n 、濃 度 4 . 0 % に設 定 した。
4 . 採 血 および筋 組 織 採 取
クルク ミン投 与 2 4 時 間 後 に解 剖 を行 った。イソフルラン吸 入 麻 酔 下 で
開 腹 し 、 腹 部 大 動 脈 か ら 血 液 を ヘ パ リ ン 処 理 さ れ た 真 空 採 血 管
( T E R U M O , T o k y o , J a p a n ) に採 取 した 。プレ麻 酔 時 は、流 速 0 . 8 L / m i n 、
濃 度 4 . 0 % に設 定 し、維 持 麻 酔 時 は、流 速 0 . 8 L / m i n 、濃 度 1 . 0 %に設
定 した。採 取 した血 液 は、 2 , 6 0 0 x G 、 1 0 分 間 の条 件 で遠 心 を行 い、上 清
の血 漿 成 分 を分 離 し、 - 8 0 ℃ の冷 凍 庫 にて保 存 した。
ま た 、 両 脚 か ら 腓 腹 筋 を 摘 出 し た 。 摘 出 し た 腓 腹 筋 は 、 R N A l a t e r
( A p p l i e d B i o s y s t e m s , C a r l s b a d , C a l i f o r n i a , U S A ) に浸 して液 体 窒 素
で凍 結 させた後 、 - 8 0 ℃ の冷 凍 庫 にて保 存 した。
5 . クレアチンキナーゼ活 性
血 漿 中 の ク レ ア チ ン キナ ー ゼ 活 性 は 長 浜 ラ イフ サ ン エ ン ス ラ ボラ ト リ ーに
依 頼 して測 定 した。
6 . r e a l - t i m e q u a n t i t a t i v e P C R
腓 腹 筋 の T o t a l R N A は、 R N e a s y M i n i K i t ( Q I A G E N , V a k e n c i a ,
C a l i f o r n i a , U S A ) および R N a s e - F r e e D N a s e S e t ( Q I A G E N , V a k e n c i a ,
C a l i f o r n i a , U S A ) を用 いて抽 出 した。その後 N a n o D r o p s y s t e m ( N a n o
D r o p T e c h n o l o g i e s , W i l m i n g s t o n , D e l a w a r e , U S A ) を用 いて R N A 濃
度 を 測 定 し た 。 T o t a l m R N A は 、 H i g h C a p a c i t y c D N A R e v e r s e

6
T r a n s c r i p t i o n k i t w i t h R N a s e i n h i b i t o r ( A p p l i e d B i o s y s t e m s ,
C a r l s b a d , C a l i f o r n i a , U S A ) を用 いて、逆 転 写 反 応 によ り c D N A を作 製
し た 。 作 製 し た c D N A は 、 P o w e r S Y B R G r e e n P C R M a s t e r M i x
( A p p l i e d B i o s y s t e m s , C a r l s b a d , C a l i f o r n i a , U S A ) を 用 い て F a s t
r e a l - t i m e P C R 7 5 0 0 装 置 ( A p p l i e d B i o s y s t e m s , C a r l s b a d ,
C a l i f o r n i a , U S A ) によ り P C R を行 った。 P C R 条 件 は、 9 5 ℃ で 1 0 分 間
c D N A を変 性 させた後 に、 1 サイクル 9 5 ℃ で 1 5 秒 間 、 6 0 ℃ で 1 分 間 の
条 件 で 、 4 0 サ イ ク ル 繰 り 返 し た 。 構 成 的 に 発 現 す る 遺 伝 子 と し て 、
g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e ( G A P D H )を使 用 した 。
標 的 遺 伝 子 および内 因 性 遺 伝 子 として使 用 したプライマーは T a b l e 1 に
示 した。
7 . タンパク質 濃 度 測 定
腓 腹 筋 は T i s s u e P r o t e i n E x t r a c t i o n R e a g e n t ( T - P E R ) w i t h
P r o t e a s e i n h i b i t o r ( T h e r m o , R o c k f o r d , I l l i n o i s , U S A ) 溶 液 でホモジ
ナイズした後 、 1 0 , 0 0 0 x G 、 1 0 分 間 の条 件 で遠 心 操 作 を行 い上 清 のタン
パ ク 質 を 分 離 し た 。 タ ン パ ク 濃 度 は B C A P r o t e i n A s s a y ( T h e r m o ,
R o c k f o r d , I l l i n o i s , U S A ) を用 いて測 定 した。
腓 腹 筋 内 の T N F - α タ ン パ ク 質 濃 度 は M o u s e T N F - α E L I S A k i t
( R & D s y s t e m s , M c k i n l e y , M i n n e s o t a , U S A ) 、 I L - 6 タンパク質 濃 度 は
M o u s e I L - 6 E L I S A k i t ( R & D s y s t e m s , M c k i n l e y , M i n n e s o t a , U S A ) 、
M C P - 1 タンパク質 濃 度 は M o u s e M C P - 1 E L I S A k i t ( R & D s y s t e m s ,
M c k i n l e y , M i n n e s o t a , U S A ) を用 いて測 定 した。 M P O タンパク質 濃 度 は、
M o u s e M P O E L I S A k i t ( H y c u l t b i o t e c h , U d e n , N e t h e r l a n d ) を用 いて
測 定 した 。 な お 、吸 光 度 の 測 定 に は V E R S A M a x m i c r o p l a t e r e a d e r

7
( M o l e c u l a r D e v i c e s , C a l i f o r n i a , U S A ) を使 用 した。
8 . H 2 O 2 濃 度 測 定
腓 腹 筋 内 の H 2 O 2 濃 度 は S e n s o L y t e A D H P h y d r o g e n p e r o x i d e
a s s a y k i t ( F r e m o u n t , C a l i f o r n i a , U S A ) を用 いて測 定 した。なお、蛍 光
強 度 の測 定 には F L U O s t a r O P T I M A ( B M G L A B T E C H , O f f e n b u r g ,
G e r m a n y ) を使 用 した。
9 . 統 計 処 理
結 果 は 、平 均 値 + 標 準 誤 差 で 表 した 。 運 動 お よび ク ル ク ミ ン投 与 を要
因 と した測 定 項 目 の 変 化 について は、二 元 配 置 分 散 分 析 を行 い、優 位
水 準 は 5 %未 満 とした。 P o s t h o c テス トは T u r k e y ’ s t e s t の検 定 を行 い、
優 位 水 準 は 5 %未 満 とした。
全 ての分 析 は S P S S V . 1 7 を用 いて行 った。

8
3 ) 結 果
ダウンヒル 運 動 お よびクルク ミ ン 投 与 に よる血 漿 中 のクレアチンキナーゼ 活
性 の変 化
血 漿 中 のクレアチン キナーゼ活 性 はダウンヒル運 動 による影 響 がみられ
た [ a n e f f e c t o f d o w n h i l l e x e r c i s e : F = 1 0 . 7 , P < 0 . 0 1 . , c u r c u m i n :
F = 0 . 0 0 7 , n . s . , a n d e x e r c i s e x c u r c u m i n i n t e r a c t i o n : F = 0 . 6 5 , n . s . ] 。
ま た 、 ク ル ク ミ ン 非 投 与 群 に お いて ク レ ア チン キナ ーゼ 活 性 は ダウン ヒ ル 運
動 によって増 加 する傾 向 がみられた ( P = 0 . 0 8 8 ) 。 しかしながら、ダウンヒル
運 動 群 に お いて 、 ク レ アチン キナ ーゼ 活 性 に ついて はク ル ク ミ ン 投 与 に よ る
有 意 な変 化 はみられなかった ( F i g . 1 ) 。
ダ ウン ヒ ル 運 動 お よび ク ル ク ミ ン 投 与 に よ る 骨 格 筋 組 織 の 炎 症 性 サ イ ト カ
インのタンパク質 濃 度 の変 化
筋 組 織 中 の T N F - のタンパク質 濃 度 はダウンヒル運 動 およびクルク ミン
投 与 に よ る影 響 はみ ら れ な かった [ a n e f f e c t o f d o w n h i l l e x e r c i s e :
F = 1 . 5 9 , n . s . , c u r c u m i n : F = 0 . 5 6 , n . s . , a n d e x e r c i s e x c u r c u m i n
i n t e r a c t i o n : F = 2 . 1 7 , n . s . ] 。 一 方 、 ク ル ク ミ ン 非 投 与 群 に お い て も 、
T N F - のタンパク質 濃 度 はダウンヒル運 動 による有 意 な変 化 はみられなか
った 。加 えて 、ダウン ヒル運 動 群 にお いて 、 T N F - の タンパク 質 濃 度 はクル
ク ミン投 与 による有 意 な変 化 はみられなかった ( F i g . 2 A ) 。
また 、筋 組 織 中 の I L - 6 のタンパク質 濃 度 はダウンヒル運 動 およびクルク
ミン投 与 による影 響 はみられなかった [ a n e f f e c t o f d o w n h i l l e x e r c i s e :
F = 0 . 0 1 n . s . , c u r c u m i n : F = 0 . 0 1 , n . s . , a n d e x e r c i s e x c u r c u m i n
i n t e r a c t i o n : F = 0 . 0 0 5 , n . s . ] 。また、クルク ミン非 投 与 群 において I L - 6 の
タンパク質 濃 度 はダウンヒル運 動 による有 意 な変 化 はみられなかった。加 え

9
て、ダウンヒル運 動 群 において 、 I L - 6 のタンパク質 濃 度 についてはクルク ミ
ン投 与 によって有 意 な変 化 はみられなかった ( F i g . 2 B ) 。
ダ ウン ヒ ル 運 動 お よび ク ル ク ミ ン 投 与 に よ る 骨 格 筋 組 織 の 炎 症 性 サ イ ト カ
インの遺 伝 子 発 現 の変 化
筋 組 織 中 の T N F - の遺 伝 子 発 現 はダウンヒル運 動 およびクルク ミン投
与 に よ る 影 響 は み ら れ な か っ た [ a n e f f e c t o f d o w n h i l l e x e r c i s e :
F = 0 . 2 9 , n . s . , c u r c u m i n : F = 0 . 2 8 , n . s . , a n d e x e r c i s e x c u r c u m i n
i n t e r a c t i o n : F = 0 . 0 8 , n . s . ] 。また 、クルク ミン非 投 与 群 において T N F - の
遺 伝 子 発 現 はダウンヒル運 動 による有 意 な変 化 はみられなかった。加 えて、
ダウン ヒル 運 動 群 にお いて 、 T N F - の 遺 伝 子 発 現 に ついて はク ルク ミ ン 投
与 による有 意 な変 化 はみられなかった ( F i g . 3 A ) 。
一 方 、筋 組 織 中 の I L - 6 の遺 伝 子 発 現 量 はダウンヒル運 動 およびクル
ク ミン投 与 の影 響 はみられなかった [ a n e f f e c t o f d o w n h i l l e x e r c i s e :
F = 2 . 2 8 , n . s . , c u r c u m i n : F = 4 . 1 6 , n . s . , a n d e x e r c i s e x c u r c u m i n
i n t e r a c t i o n : F = 0 . 0 8 , n . s . ] 。 また、クルク ミン非 投 与 群 において I L - 6 の遺
伝 子 発 現 はダウン ヒル 運 動 に よ る有 意 な変 化 はみ られな かった 。加 えて 、
ダウンヒル運 動 群 において、 I L - 6 の遺 伝 子 発 現 についてはクルク ミン投 与
による有 意 な変 化 はみられなかった ( F i g . 3 B ) 。
筋 組 織 中 の I L - 1 の遺 伝 子 発 現 はダウンヒル運 動 およびクルク ミン投
与 の影 響 はみられなかった [ a n e f f e c t o f d o w n h i l l e x e r c i s e : F = 0 . 0 1 ,
n . s . , c u r c u m i n : F = 0 . 4 4 , n . s . , a n d e x e r c i s e x c u r c u m i n i n t e r a c t i o n :
F = 2 . 4 7 , n . s . ] 。また、クルク ミン非 投 与 群 において I L - 1 の遺 伝 子 発 現 は
ダウン ヒ ル 運 動 に よ る有 意 な 影 響 はみ られな かった 。加 えて 、ダウン ヒ ル 運
動 群 に おいて 、 I L - 1 の遺 伝 子 発 現 について はクルク ミ ン 投 与 による有 意

10
な変 化 はみられなかった ( F i g . 3 C ) 。
加 えて、筋 組 織 中 の I L - 1 2 の遺 伝 子 発 現 はダウンヒル運 動 およびクル
ク ミ ン 投 与 に よ る 影 響 は み ら れ な か っ た [ a n e f f e c t o f d o w n h i l l
e x e r c i s e : F = 0 . 4 0 , n . s . , c u r c u m i n : F = 0 . 0 0 , n . s . , a n d e x e r c i s e x
c u r c u m i n i n t e r a c t i o n : F = 0 . 0 4 , n . s . ] 。また、クルク ミン非 投 与 群 におい
て I L - 1 2 の遺 伝 子 発 現 はダウンヒル運 動 による有 意 な変 化 はみられなかっ
た。加 えて、ダウンヒル運 動 群 において、 I L - 1 2 の遺 伝 子 発 現 についてはク
ルク ミン投 与 による有 意 な変 化 はみられなかった ( F i g . 3 D ) 。
ダウンヒル運 動 およびクルク ミン投 与 による骨 格 筋 組 織 の H 2 O 2 濃 度 およ
び N A D P H - o x i d a s e 遺 伝 子 発 現 の変 化
筋 組 織 中 の H 2 O 2 濃 度 はダウンヒル運 動 およびクルク ミン投 与 の影 響 は
み ら れ な か っ た [ a n e f f e c t o f d o w n h i l l e x e r c i s e : F = 3 . 5 6 , n . s . ,
c u r c u m i n : F = 2 . 7 4 , n . s . , a n d e x e r c i s e x c u r c u m i n i n t e r a c t i o n :
F = 2 . 4 7 , n . s . ] 。また 、クルク ミン非 投 与 群 において H 2 O 2 濃 度 はダウンヒル
運 動 によって有 意 に増 加 した ( P < 0 . 0 1 ) 。 しか しながら 、ダウン ヒル運 動 群
において H 2 O 2 濃 度 はクルク ミン投 与 によって有 意 に減 少 した ( P < 0 . 0 1 ,
F i g . 4 A ) 。
筋 組 織 中 の N A D P H - o x i d a s e の遺 伝 子 発 現 量 はダウンヒル運 動 によ
る影 響 がみられた [ a n e f f e c t o f d o w n h i l l e x e r c i s e : F = 8 . 7 4 , P < 0 . 0 1 ,
c u r c u m i n : F = 1 . 4 9 , n . s . , a n d e x e r c i s e x c u r c u m i n i n t e r a c t i o n :
F = 5 . 9 5 , P < 0 . 0 5 ] 。 また、クルク ミン非 投 与 群 において N A D P H - o x i d a s e の
遺 伝 子 発 現 量 はダウンヒル運 動 によって有 意 に増 加 した ( P < 0 . 0 1 ) 。 し
かしながら 、ダウンヒル運 動 群 において 、 N A D P H - o x i d a s e の遺 伝 子 発 現
量 はクルク ミン投 与 によって有 意 に減 少 した ( P < 0 . 0 5 , F i g . 4 B ) 。

11
ダウンヒル運 動 およびクルク ミン投 与 による骨 格 筋 組 織 の M P O 濃 度 の変
化
筋 組 織 中 の M P O のタンパク質 濃 度 はクルク ミン投 与 による影 響 がみら
れ た [ a n e f f e c t o f d o w n h i l l e x e r c i s e : F = 0 . 1 8 , n . s . , c u r c u m i n :
F = 7 . 3 2 , P < 0 . 0 1 , a n d e x e r c i s e x c u r c u m i n i n t e r a c t i o n : F = 3 . 5 7 , n . s . ] 。
また、クルク ミン非 投 与 群 において M P O のタンパク質 濃 度 についてはダウン
ヒル運 動 によって有 意 な変 化 はみられなかった 。 しかしながら 、ダウンヒル運
動 群 において、 M P O のタンパク質 濃 度 はクルク ミン投 与 によって有 意 に増
加 した ( P < 0 . 0 1 , F i g . 5 ) 。
ダウンヒル運 動 およびクルク ミン投 与 による骨 格 筋 組 織 の M C P - 1 タンパク
質 濃 度 および M C P - 1 の遺 伝 子 発 現 の変 化
筋 組 織 中 の M C P - 1 のタンパク質 濃 度 はダウンヒル運 動 による影 響 がみ
ら れ た [ a n e f f e c t o f d o w n h i l l e x e r c i s e : F = 1 0 . 7 1 , P < 0 . 0 1 ,
c u r c u m i n : F = 0 . 7 9 , n . s . , a n d e x e r c i s e x c u r c u m i n i n t e r a c t i o n :
F = 0 . 8 5 , n . s . ] 。 また、クルク ミン非 投 与 群 において M C P - 1 のタンパク質 濃
度 はダウンヒル運 動 によって有 意 に増 加 した ( P < 0 . 0 1 ) 。 しかしながら 、ダ
ウンヒル運 動 群 において、 M C P - 1 のタンパク質 濃 度 についてはクルク ミン投
与 による有 意 な変 化 はみられなかった ( F i g . 6 A ) 。
また 、筋 組 織 中 の M C P - 1 の遺 伝 子 発 現 はダウンヒル運 動 およびクルク
ミ ン 投 与 に よ る 影 響 が み ら れ た [ a n e f f e c t o f d o w n h i l l e x e r c i s e :
F = 1 9 . 3 0 , P < 0 . 0 1 , c u r c u m i n : F = 4 . 6 0 , P < 0 . 0 5 , a n d e x e r c i s e x
c u r c u m i n i n t e r a c t i o n : F = 2 . 8 7 , n . s . ] 。一 方 、クルク ミン非 投 与 群 におい
て M C P - 1 の 遺 伝 子 発 現 は ダ ウ ン ヒ ル 運 動 に よ っ て 有 意 に 増 加 し た
( P < 0 . 0 1 ) 。 し か しな が ら 、ダウン ヒ ル 運 動 群 に お いて 、 M C P - 1 の 遺 伝 子

12
発 現 はクルク ミン投 与 によって有 意 に減 少 した ( P < 0 . 0 1 , F i g . 6 B ) 。
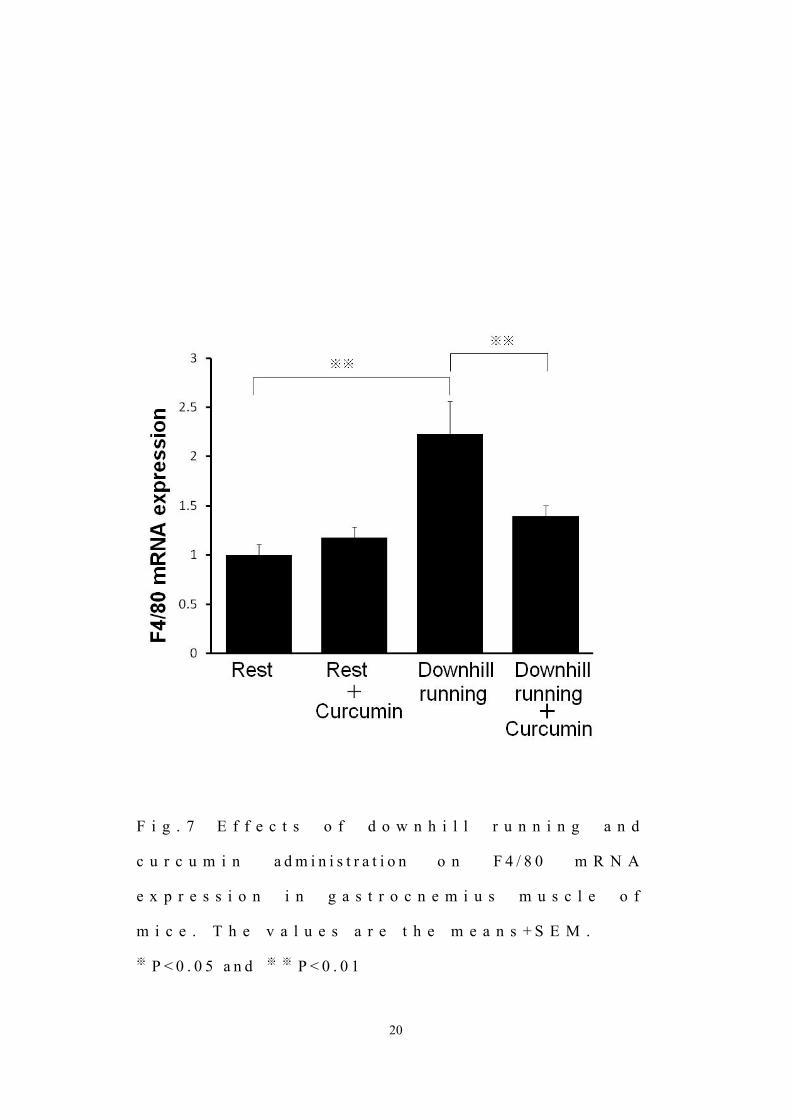
さ らに 、筋 組 織 中 の F 4 / 8 0 の遺 伝 子 発 現 はダウンヒル運 動 およびクルク
ミ ン 投 与 に よ る 影 響 が み ら れ た [ a n e f f e c t o f d o w n h i l l e x e r c i s e :
F = 1 2 . 1 5 , n . s . , c u r c u m i n : F = 3 . 1 5 , n . s . , a n d e x e r c i s e x c u r c u m i n
i n t e r a c t i o n : F = 5 . 1 7 , n . s . ] 。また 、クルク ミン非 投 与 群 において F 4 / 8 0 の
遺 伝 子 発 現 はダウンヒル運 動 によって 有 意 に増 加 した ( P < 0 . 0 1 ) 。 しか し
ながら、ダウンヒル運 動 群 において、 F 4 / 8 0 の遺 伝 子 発 現 はクルク ミン投 与
によって有 意 に減 少 した ( P < 0 . 0 1 , F i g . 7 A ) 。

13
gene forward reverse
GAPDH TGAAGCAGGCATCTGAGGG CGAAGGTGGAAGAGTGGGAG
TNF-a CCTCCCTCTCATCAGTTCTA ACTTGGTGGTTTGCTACGAC
IL-6 TAGTCCTTCCTACCCCAATTTCC TTGGTCCTTAGCCACTCCTTC
IL-1b CTGGAGAGTGTGGATCCCAAG GGAAGACACGGATTCCATGGTG
IL-12 GGAGCACGGCAGCAGAATA AACTTGAGGGAGAAGTAGGAATGG
NADPH oxidase TTGGGTCAGCACTGGCTCTG TGGCGGTGTGCAGTGCTATC
MCP-1 CTTCTGGGCCTGCTGTTCA CCAGCCTACTCATTGGGATCA
F4/80 CTTTGGCTATGGGCTTCCAGTC GCAAGGAGGACAGAGTTTATCGTG
T T a b l e 1 P r i m e r s e q u e n c e s f o r r e a l - t i m e R T - P C R
a n a l y s i s
G A P D H : G l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e
T N F t u m o r n e c r o s i s f a c t o r
I L : i n t e r l e u k i n
N A D P H o x i d a s e : n i c o t i n a m i d e a d e n i n e d i n u c l e o t i d e p h o s p h a t e
M C P - 1 : m o n o c y t e c h e m o t a c t i c p r o t e i n - 1

14
F i g . 1 E f f e c t s o f d o w n h i l l r u n n i n g a n d
c u r c u m i n a d m i n i s t r a t i o n o n p l a s m a c r e a t i n k i n a s e
a c t i v i t y . T h e v a l u e s a r e t h e m e a n s + S E M .
※ ※ P < 0 . 0 1

15
A
F i g . 2 E f f e c t s o f d o w n h i l l r u n n i n g a n d c u r c u m i n
a d m i n i s t r a t i o n o n T N F - ( A ) a n d I L - 6 ( B ) p r o t e i n
c o n c e n t r a t i o n s i n g a s t r o c n e m i u s m u s c l e o f m i c e . T h e
v a l u e s a r e t h e m e a n s + S E M .
B

16
F i g . 3 E f f e c t s o f d o w n h i l l r u n n i n g a n d
c u r c u m i n a d m i n i s t r a t i o n o n T N F - ( A ) , I L - 6
( B ) , I L - 1 ( C ) , a n d I L - 1 2 ( D ) m R N A e x p r e s s i o n s i n
g a s t r o c n e m i u s m u s c l e o f m i c e . T h e v a l u e s a r e
t h e m e a n s + S E M .
BA
C D

17
F i g 4 E f f e c t s o f d o w n h i l l r u n n i n g a n d c u r c u m i n
a d m i n i s t r a t i o n o n H 2 O 2 c o n c e n t r a t i o n ( A ) a n d
N A D P H - o x i d a s e m R N A e x p r e s s i o n ( B ) i n
g a s t r o c n e m i u s m u s c l e o f m i c e . T h e v a l u e s a r e t h e
m e a n s + S E M . ※ P < 0 . 0 5 a n d ※ ※ P < 0 . 0 1
A
B

18
F i g . 5 E f f e c t s o f d o w n h i l l r u n n i n g a n d
c u r c u m i n a d m i n i s t r a t i o n o n M P O p r o t e i n
c o n c e n t r a t i o n i n g a s t r o c n e m i u s m u s c l e o f
m i c e . T h e v a l u e s a r e t h e m e a n s + S E M . ※ ※ P < 0 . 0 5

19
F i g . 6 E f f e c t s o f d o w n h i l l r u n n i n g a n d
c u r c u m i n a d m i n i s t r a t i o n o n M C P - 1 p r o t e i n
c o n c e n t r a t i o n ( A ) a n d m R N A e x p r e s s i o n ( B ) i n
g a s t r o c n e m i u s m u s c l e o f m i c e . T h e v a l u e s a r e
t h e m e a n s + S E M . ※ ※ P < 0 . 0 5
B
A
20
F i g . 7 E f f e c t s o f d o w n h i l l r u n n i n g a n d
c u r c u m i n a d m i n i s t r a t i o n o n F 4 / 8 0 m R N A
e x p r e s s i o n i n g a s t r o c n e m i u s m u s c l e o f
m i c e . T h e v a l u e s a r e t h e m e a n s + S E M .
※ P < 0 . 0 5 a n d ※ ※ P < 0 . 0 1

21
4 ) 考 察
本 研 究 の 結 果 よ り 、 ダ ウン ヒ ル 運 動 に よ って 血 漿 ク レ ア チン キナ ーゼ 活
性 が上 昇 し、筋 損 傷 を誘 導 できたことが確 認 できた。しかしながら 、クルク ミ
ン投 与 によるダウンヒル運 動 後 の血 漿 クレアチンキナーゼ活 性 は、少 な く と
も 1 日 目 では変 化 しなかった 。また 、ダウンヒル運 動 後 のマウス腓 腹 筋 の
T N F - α 、 I L - 6 、 I L - 1 β 、 I L - 1 2 の遺 伝 子 発 現 量 、 T N F - α 、 I L - 6 のタンパク
質 濃 度 は 、 変 化 し な い こ と が 明 ら か に な っ た 。 し か し な が ら 、 筋 組 織 で の
H 2 O 2 e 濃 度 および N A D P H - o x i d a s e 遺 伝 子 発 現 はダウンヒル運 動 によ り
増 加 し 、 クルク ミ ン投 与 によ り減 少 するこ とが明 らかになった 。 また 、マク ロフ
ァージの浸 潤 を特 異 的 に誘 導 する M C P - 1 遺 伝 子 発 現 およびマクロファー
ジの浸 潤 を示 す F 4 / 8 0 遺 伝 子 発 現 がダウンヒル運 動 によ り増 加 し、クルク
ミン投 与 によ り減 少 することも明 らかになった 。一 方 で、好 中 球 の浸 潤 を示
すマーカーである M P O 濃 度 はダウンヒル運 動 の影 響 を受 けないことが示 唆
され た 。白 血 球 は活 性 酸 素 を産 生 す るこ とが知 られ て いる こ とか ら も 、 ク ル
ク ミン投 与 によるダウンヒル運 動 後 の H 2 O 2 濃 度 および N A D P H - o x i d a s e
遺 伝 子 発 現 抑 制 は、マクロファージの浸 潤 抑 制 が関 与 している可 能 性 が
考 えられる。今 回 の実 験 では運 動 直 後 にクルク ミンを投 与 し 1 日 目 にクレ
アチンキナーゼ活 性 を検 討 したため 、効 果 が生 じるのに作 用 時 間 が不 十
分 であった可 能 性 も考 えられ、運 動 負 荷 前 にクルク ミンを投 与 した場 合 の
筋 損 傷 予 防 効 果 について今 後 検 討 する必 要 がある。
これまでに 、激 しい運 動 後 のサイ トカ インの変 動 については、数 多 く報 告
がされている。ヒ トや実 験 動 物 を対 象 にした研 究 において、マラソンレースや
ダウンヒル運 動 後 に炎 症 性 サイトカインの遺 伝 子 発 現 が増 加 することが報
告 さ れ て い る [ 7 , 9 , 2 8 , 2 9 ] 。 一 方 で 、 ダ ウ ン ヒ ル 運 動 後 に T N F - α 、
I L - 1 β 、 I L - 1 2 の遺 伝 子 発 現 が変 化 しなかったことも報 告 されている [ 3 0 ] 。

22
本 研 究 では、ダウンヒル運 動 2 4 時 間 後 においてマウス骨 格 筋 の炎 症 性
サイ トカ インの遺 伝 子 発 現 は変 化 しなかった 。ダウンヒル運 動 後 の炎 症 性
サイ ト カインの変 動 は、動 物 の種 類 、運 動 条 件 、運 動 後 の時 間 経 過 によ
って結 果 に差 異 が生 じる可 能 性 が考 えられる。
活 性 酸 素 は、運 動 後 に筋 組 織 で増 加 することが知 られている。筋 損 傷
をともな う運 動 では、浸 潤 した免 疫 細 胞 による N A D P H - o x i d a s e の活 性 に
よ りスーパーオキシドが産 生 され、さらに H 2 O 2 が生 成 される [ 2 1 ] 。ラッ トを
用 いた研 究 では、 1 2 0 分 のダウンヒル運 動 2 4 時 間 後 に骨 格 筋 組 織 での
H 2 O 2 濃 度 が増 加 したことが報 告 されている [ 7 ] 。本 研 究 では、ダウンヒル
運 動 2 4 時 間 後 に マ ウ ス の 腓 腹 筋 に お い て H 2 O 2 濃 度 お よ び
N A D P H - o x i d a s e の遺 伝 子 発 現 が増 加 した。一 方 で、クルク ミン投 与 によ
ってダウンヒル運 動 後 の H 2 O 2 濃 度 および N A D P H - o x i d a s e 遺 伝 子 発 現
は有 意 に抑 制 された 。本 研 究 の結 果 から 、 クルク ミ ン投 与 によ りダウン ヒル
運 動 後 の酸 化 ストレス状 態 が軽 減 する可 能 性 が示 唆 された。 T N F - α は、
i n v i t r o において N A D P H - o x i d a s e 産 生 を促 進 することが明 らかにされて
いる [ 1 7 , 3 1 ] 。クルク ミンは N F - B の活 性 を抑 制 することから も 、クルク ミン
の示 す抗 炎 症 作 用 として 、 T N F - α などの炎 症 性 サイ トカインの発 現 抑 制
が関 与 すると考 えられる。しかしながら、本 研 究 では炎 症 性 サイトカインがダ
ウンヒル運 動 によ り有 意 に増 加 しなかった。したがって 、好 中 球 およびマクロ
ファージ由 来 の活 性 酸 素 を、細 胞 浸 潤 の抑 制 を介 して制 御 したもの と考
えられる。
M P O は、好 中 球 が活 性 化 すると 、食 胞 内 あるいは細 胞 外 に放 出 され
る。 M P O のタンパク質 濃 度 は、激 しい運 動 による好 中 球 の脱 顆 粒 に伴 い
増 加 することが知 られている [ 3 2 , 3 3 ] 。本 研 究 では、ダウンヒル運 動 2 4 時
間 後 のマウス骨 格 筋 の M P O 濃 度 は変 化 しないことが明 らかになった。好

23
中 球 は損 傷 後 1 時 間 以 内 に組 織 に浸 潤 し始 めることが知 られている。実
際 に、筋 損 傷 を伴 う運 動 後 の M P O 濃 度 を検 討 した研 究 は、多 くが運 動
直 後 および 1 時 間 後 に上 昇 したことを示 している 。本 研 究 で測 定 したダウ
ンヒル運 動 2 4 時 間 後 には、好 中 球 の次 に浸 潤 するマクロファージによって
貪 食 さ れ た 可 能 性 が 考 え ら れ る 。 これ に つ い て は 、 運 動 直 後 か ら 数 時 間
後 の M P O の変 動 や組 織 学 的 検 討 を行 い明 らかにする必 要 がある。また 、
クルク ミン投 与 によ りダウンヒル運 動 2 4 時 間 後 の M P O の濃 度 が増 加 した
が、その要 因 としてマクロファージとの関 連 が考 えられる。マクロファージは好
中 球 を貪 食 することが知 られている。したがって 、クルク ミン投 与 によ りマクロ
フ ァージの 浸 潤 が抑 制 された ため 、貪 食 されずに組 織 に 残 る好 中 球 が増
加 した可 能 性 が考 えられる。
マクロファージは、筋 損 傷 を伴 う運 動 後 1 日 以 内 に、損 傷 組 織 に浸 潤
することが知 られている [ 5 ] 。組 織 へのマクロファージの浸 潤 は、 N F - B シ
グナル伝 達 経 路 を介 した M C P - 1 などのケモカインの産 生 によ り誘 導 される
[ 1 3 , 1 4 ] 。また、浸 潤 したマクロファージは、 T N F - α 、 I L - 1 β などのサイ トカイ
ンを産 生 する こ とが知 られている [ 3 4 ] 。 ダウン ヒル運 動 を用 いた研 究 で は 、
9 0 分 間 の走 行 運 動 2 4 時 間 後 および 4 8 時 間 後 において、マウス腓 腹 筋
のマク ロフ ァージの浸 潤 が増 加 するこ とが報 告 されている [ 3 5 ] 。本 研 究 よ
り 、ダウンヒル運 動 2 4 時 間 後 にマクロファージが筋 組 織 に浸 潤 する可 能
性 が示 唆 された。一 方 で、クルク ミンはダウンヒル運 動 後 のマクロファージの
浸 潤 を抑 制 する可 能 性 が示 唆 された 。これまでに 、糖 尿 病 ラッ ト を用 いた
研 究 で、 M C P - 1 の血 中 濃 度 がクルク ミン投 与 によ り抑 制 されることが報 告
されている [ 3 6 ] 。本 研 究 においても、クルク ミンは F 4 / 8 0 発 現 同 様 に、筋
組 織 の M C P - 1 発 現 を減 少 させることが示 された。したがって、クルク ミンに
よるマクロファージ浸 潤 の抑 制 には骨 格 筋 の M C P - 1 産 生 の抑 制 が関 与

24
する可 能 性 が示 唆 された 。浸 潤 した 好 中 球 やマク ロフ ァージは活 性 酸 素
を産 生 することが知 られている [ 3 7 ] 。 N A D P H - o x i d a s e は好 中 球 やマクロ
フ ァージの形 質 膜 に存 在 し 、好 中 球 やマク ロフ ァージが貪 食 を行 った際 に
活 性 化 さ れ る 。 本 研 究 に お いて マ ク ロ フ ァ ージが 筋 組 織 に 浸 潤 す る 可 能
性 が示 唆 された。したがって、ダウンヒル運 動 による N A D P H - o x i d a s e の遺
伝 子 発 現 の増 加 は、マク ロファージの浸 潤 による可 能 性 が示 唆 される 。 ま
た、クルク ミンは F 4 / 8 0 発 現 同 様 に、ダウンヒル運 動 後 の H 2 O 2 濃 度 および
N A D P H - o x i d a s e 遺 伝 子 発 現 を減 少 させた 。 したがって 、クルク ミ ン投 与
によるダウンヒル運 動 後 の H 2 O 2 濃 度 および N A D P H - o x i d a s e 遺 伝 子 発
現 の減 少 は、マクロファージの浸 潤 抑 制 が関 与 している可 能 性 が考 えられ
る。
5 ) 結 論
ク ル ク ミ ン 投 与 は ダ ウ ン ヒ ル 運 動 後 の H 2 O 2 濃 度 お よ び
N A D P H - o x i d a s e 遺 伝 子 発 現 を抑 制 した。また、クルク ミン投 与 はダウンヒ
ル運 動 後 の M C P - 1 および F 4 / 8 0 遺 伝 子 発 現 を抑 制 した。したがって、ク
ル ク ミ ン 投 与 はマ ク ロ フ ァ ージ の 浸 潤 抑 制 を 介 し て 、 活 性 酸 素 産 生 を 抑
制 する可 能 性 が示 唆 された。

25
6 ) 謝 辞
本 稿 の作 成 にあたり 、御 懇 篤 な御 指 導 ならびに激 励 を賜 りました本 研
究 科 の鈴 木 克 彦 准 教 授 に深 甚 なる謝 意 を表 します。本 研 究 科 入 学 当
初 から本 稿 提 出 に至 るまで、免 疫 学 を学 んで間 もない浅 学 非 才 な私 に、
多 くの助 言 や暖 かい御 指 導 をしていただきました。ここに心 から御 礼 申 し上
げます。
また、副 査 として御 指 導 いただきました坂 本 静 男 先 生 、福 林 徹 先 生 に
も厚 く御 礼 申 し上 げます。
実 験 計 画 、実 験 手 法 、論 文 作 成 に至 るまで懇 切 丁 寧 に御 指 導 して
頂 きました予 防 医 学 研 究 室 博 士 課 程 の川 西 範 明 氏 に心 より御 礼 申 し
上 げます。また、本 稿 作 成 や大 学 院 生 活 を様 々な面 から支 えて下 さった
予 防 医 学 研 究 室 の皆 様 に深 く感 謝 致 します。
セラバリューズ株 式 会 社 の大 塚 喜 彦 様 には、クルク ミンの提 供 から研 究
における助 言 など様 々な面 で御 協 力 して頂 きましたこと厚 く御 礼 申 し上 げ
ます。
最 後 に、本 稿 作 成 のための大 学 院 生 活 を物 心 両 面 から支 えて くださっ
た両 親 に心 より感 謝 致 します。

26
7 ) 参 考 文 献
1 ) N o s a k a K , N e w t o n M . C o n c e n t r i c o r e c c e n t r i c t r a i n i n g e f f e c t o n
e c c e n t r i c e x e r c i s e - i n d u c e d m u s c l e d a m a g e . M e d S c i S p o r t s
E x e r c . 3 4 ( 1 ) : 6 3 - 6 9 . 2 0 0 2 .
2 ) P e a k e J , N o s a k a K , S u z u k i K . C h a r a c t e r i z a t i o n o f i n f l a m m a t o r y
r e s p o n s e s t o e c c e n t r i c e x e r c i s e i n h u m a n s . E x e r c I m m u n o l
R e v . 1 1 : 6 4 - 8 5 . 2 0 0 5 .
3 ) T h o m p s o n D , B a i l e y D M , H i l l J , H u r s t T , P o w e l l J R , W i l l i a m s C .
P r o l o n g e d v i t a m i n C s u p p l e m e n t a t i o n a n d r e c o v e r y f r o m e c -
c e n t r i c e x e r c i s e . E u r J A p p l P h y s i o l . J u n ; 9 2 ( 1 - 2 ) : 1 3 3 - 1 3 8 .
2 0 0 4 .
4 ) P e a k e J M . E x e r c i s e - i n d u c e d a l t e r a t i o n s i n n e u t r o p h i l d e g r a -
n u l a t i o n a n d r e s p i r a t o r y b u r s t a c t i v i t y : p o s s i b l e m e c h a n i s m s o f
a c t i o n . E x e r c I m m u n o l R e v . 8 : 4 9 - 1 0 0 . 2 0 0 2 .
5 ) T i d b a l l J G , W e h l i n g - H e n r i c k s M . M a c r o p h a g e s p r o m o t e m u s c l e
m e m b r a n e r e p a i r a n d m u s c l e f i b r e g r o w t h a n d r e g e n e r a t i o n
d u r i n g m o d i f i e d m u s c l e l o a d i n g i n m i c e i n v i v o . J P h y s i o l .
5 7 8 ( P t 1 ) : 3 2 7 - 3 3 6 . 2 0 0 7 .
6 ) P e l o s i L , G i a c i n t i C , N a r d i s C , B o r s e l l i n o G , R i z z u t o E , N i -
c o l e t t i C , W a n n e n e s F , B a t t i s t i n i L , R o s e n t h a l N , M o l i n a r o M ,
M u s a r ò A . L o c a l e x p r e s s i o n o f I G F - 1 a c c e l e r a t e s m u s c l e r e -
g e n e r a t i o n b y r a p i d l y m o d u l a t i n g i n f l a m m a t o r y c y t o k i n e s a n d
c h e m o k i n e s . FA S E B J . 2 1 ( 7 ) : 1 3 9 3 - 1 4 0 2 . 2 0 0 7 .
7 ) L i a o P , Z h o u J , J i L L , Z h a n g Y. E c c e n t r i c c o n t r a c t i o n i n d u c e s
i n f l a m m a t o r y r e s p o n s e s i n r a t s k e l e t a l m u s c l e : r o l e o f t u m o r
n e c r o s i s f a c t o r - a l p h a . A m J P h y s i o l R e g u l I n t e g r C o m p P h y s i o l .
2 9 8 ( 3 ) : R 5 9 9 - 6 0 7 2 0 1 0 .
8 ) D a v i s J M , M u r p h y E A , C a r m i c h a e l M D , Z i e l i n s k i M R ,
G r o s c h w i t z C M , B r o w n A S , G a n g e m i J D , G h a f f a r A , M a y e r E P.

27
C u r c u m i n e f f e c t s o n i n f l a m m a t i o n a n d p e r f o r m a n c e r e c o v e r y
f o l l o w i n g e c c e n t r i c e x e r c i s e - i n d u c e d m u s c l e d a m a g e . A m J
P h y s i o l R e g u l I n t e g r C o m p P h y s i o l 2 9 2 ( 6 ) : R 2 1 6 8 - 2 1 7 3 . 2 0 0 7 .
9 ) F i e l d i n g R A , M a n f r e d i T J , D i n g W, F i a t a r o n e M A , E v a n s W J ,
C a n n o n J G . A c u t e p h a s e r e s p o n s e i n e x e r c i s e . I I I . N e u t r o p h i l
a n d I L - 1 a c c u m u l a t i o n i n s k e l e t a l m u s c l e . A m J P h y s i o l R e g u l
I n t e g r C o m p P h y s i o l . 2 6 5 : R 1 6 6 – 1 7 2 . 1 9 9 3 .
1 0 ) S h e p h a r d R J . A d h e s i o n m o l e c u l e s , c a t e c h o l a m i n e s a n d l e u c o -
c y t e r e d i s t r i b u t i o n d u r i n g a n d f o l l o w i n g e x e r c i s e . S p o r t s M e d
3 3 ( 4 ) : 2 6 1 - 2 8 4 . 2 0 0 3 .
1 1 ) B e l c a s t r o A N , A r t h u r G D , A l b i s s e r T A , R a j D A . H e a r t , l i v e r ,
a n d s k e l e t a l m u s c l e m y e l o p e r o x i d a s e a c t i v i t y d u r i n g e x e r c i s e .
J A p p l P h y s i o l . 8 0 : 1 3 3 1 – 1 3 3 5 . 1 9 9 6 .
1 2 ) L o w e D A , W a r r e n G L , I n g a l l s C P, B o o r s t e i n D B , A r m s t r o n g R B .
M u s c l e f u n c t i o n a n d p r o t e i n m e t a b o l i s m a f t e r i n i t i a t i o n o f
e c c e n t r i c c o n t r a c t i o n - i n d u c e d i n j u r y . J A p p l P h y s i o l .
7 9 ( 4 ) : 1 2 6 0 - 1 2 7 0 . 1 9 9 5 .
1 3 ) W a r r e n G L , H u l d e r m a n T , M i s h r a D , G a o X , M i l l e c c h i a L ,
O ' F a r r e l l L , K u z i e l WA , S i m e o n o v a P P. C h e m o k i n e r e c e p t o r
C C R 2 i n v o l v e m e n t i n s k e l e t a l m u s c l e r e g e n e r a t i o n . FA S E B J .
1 9 ( 3 ) : 4 1 3 - 4 1 5 . 2 0 0 5 .
1 4 ) S h i n W S , S z u b a A , R o c k s o n S G . T h e r o l e o f c h e m o k i n e s i n
h u m a n c a r d i o v a s c u l a r p a t h o l o g y : e n h a n c e d b i o l o g i c a l i n s i g h t s .
A t h e r o s c l e r o s i s . 1 6 0 ( 1 ) : 9 1 - 1 0 2 . 2 0 0 2 .
1 5 ) S e g a w a M , F u k a d a S , Ya m a m o t o Y, Ya h a g i H , K a n e m a t s u M ,
S a t o M , I t o T , U e z u m i A , H a y a s h i S , M i y a g o e - S u z u k i Y, T a k e d a
S , T s u j i k a w a K , Ya m a m o t o H . S u p p r e s s i o n o f m a c r o p h a g e
f u n c t i o n s i m p a i r s s k e l e t a l m u s c l e r e g e n e r a t i o n w i t h s e v e r e
f i b r o s i s . E x p C e l l R e s . 3 1 4 ( 1 7 ) : 3 2 3 2 - 3 2 4 4 . 2 0 0 8 .

28
1 6 ) B e d a r d K , K r a u s e K H . T h e N O X f a m i l y o f R O S - g e n e r a t i n g
N A D P H o x i d a s e s : p h y s i o l o g y a n d p a t h o p h y s i o l o g y . P h y s i o l R e v .
8 7 ( 1 ) : 2 4 5 - 3 1 3 . 2 0 0 7 .
1 7 ) L i J M , F a n L M , C h r i s t i e M R , S h a h A M . A c u t e t u m o r n e c r o s i s
f a c t o r a l p h a s i g n a l i n g v i a N A D P H o x i d a s e i n m i c r o v a s c u l a r
e n d o t h e l i a l c e l l s : r o l e o f p 4 7 p h o x p h o s p h o r y l a t i o n a n d b i n d i n g
t o T R A F 4 . M o l C e l l B i o l . 2 5 ( 6 ) : 2 3 2 0 - 2 3 3 0 . 2 0 0 5 .
1 8 ) D r ö g e W. F r e e r a d i c a l s i n t h e p h y s i o l o g i c a l c o n t r o l o f c e l l
f u n c t i o n . P h y s i o l R e v . 8 2 ( 1 ) : 4 7 - 9 5 . 2 0 0 2 .
1 9 ) P a h l H L . A c t i v a t o r s a n d t a r g e t g e n e s o f R e l / N F - k a p p a B t r a n -
s c r i p t i o n f a c t o r s . O n c o g e n e . 1 8 ( 4 9 ) : 6 8 5 3 - 6 8 6 6 . 1 9 9 9 .
2 0 ) A l e s s i o H M . E x e r c i s e - i n d u c e d o x i d a t i v e s t r e s s . M e d S c i S p o r t s
E x e r c . 2 5 ( 2 ) : 2 1 8 - 2 2 4 . 1 9 9 3 .
2 1 ) P o w e r s S K , J a c k s o n M J . E x e r c i s e - i n d u c e d o x i d a t i v e s t r e s s :
c e l l u l a r m e c h a n i s m s a n d i m p a c t o n m u s c l e f o r c e p r o d u c t i o n .
P h y s i o l R e v 8 8 : 1 2 4 3 - 1 2 7 6 ; 2 0 0 8 .
2 2 ) S i k o r a E , S c a p a g n i n i G , B a r b a g a l l o M . C u r c u m i n , i n f l a m m a t i o n ,
a g e i n g a n d a g e - r e l a t e d d i s e a s e s . I m m u n A g e i n g . J a n 1 7 ; 7 ( 1 ) : 1 .
2 0 1 0
2 3 ) M e n o n V P, S u d h e e r A R . A n t i o x i d a n t a n d a n t i - i n f l a m m a t o r y
p r o p e r t i e s o f c u r c u m i n . A d v E x p M e d B i o l . 5 9 5 : 1 0 5 - 2 5 . R e v i e w .
2 0 0 7 .
2 4 ) S i n g h S , A g g a r w a l B B . A c t i v a t i o n o f t r a n s c r i p t i o n f a c t o r
N F - k a p p a B i s s u p p r e s s e d b y c u r c u m i n ( d i f e r u l o y l m e t h a n e )
[ c o r r e c t e d ] . J B i o l C h e m 2 7 0 : 2 4 9 9 5 – 2 5 0 0 0 , 1 9 9 5 .
2 5 ) H a n S S , K e u m Y S , S e o H J , S u r h Y J . C u r c u m i n s u p p r e s s e s a c -
t i v a t i o n o f N F - k a p p a B a n d A P - 1 i n d u c e d b y p h o r b o l e s t e r i n
c u l t u r e d h u m a n p r o m y e l o c y t i c l e u k e m i a c e l l s . J B i o c h e m M o l
B i o l 3 5 : 3 3 7 – 3 4 2 , 2 0 0 2 .

29
2 6 ) B i s w a s S K , M c C l u r e D , J i m e n e z L A , M e g s o n I L , R a h m a n I .
C u r c u m i n i n d u c e s g l u t a t h i o n e b i o s y n t h e s i s a n d i n h i b i t s
N F - k a p p a B a c t i v a t i o n a n d i n t e r l e u k i n - 8 r e l e a s e i n a l v e o l a r
e p i t h e l i a l c e l l s : m e c h a n i s m o f f r e e r a d i c a l s c a v e n g i n g a c t i v i t y .
A n t i o x i d R e d o x S i g n a l . 7 ( 1 - 2 ) : 3 2 - 4 1 . 7 : 3 2 – 4 1 , 2 0 0 5 .
2 7 ) L i m J H , K w o n T K . C u r c u m i n i n h i b i t s p h o r b o l m y r i s t a t e a c e t a t e
( P M A ) - i n d u c e d M C P - 1 e x p r e s s i o n b y i n h i b i t i n g E R K a n d
N F - k a p p a B t r a n s c r i p t i o n a l a c t i v i t y . F o o d C h e m T o x i c o l .
4 8 ( 1 ) : 4 7 - 5 2 . 2 0 1 0 .
2 8 ) P e a k e J M , S u z u k i K , H o r d e r n M , W i l s o n G , N o s a k a K , C o o m b e s
J S . P l a s m a c y t o k i n e c h a n g e s i n r e l a t i o n t o e x e r c i s e i n t e n s i t y
a n d m u s c l e d a m a g e . E u r J A p p l P h y s i o l . 9 5 ( 5 - 6 ) : 5 1 4 - 5 2 1 . 2 0 0 5 .
2 9 ) H a m a d a K , Va n n i e r E , S a c h e c k J M , W i t s e l l A L , R o u b e n o f f R .
S e n e s c e n c e o f h u m a n s k e l e t a l m u s c l e i m p a i r s t h e l o c a l i n -
f l a m m a t o r y c y t o k i n e r e s p o n s e t o a c u t e e c c e n t r i c e x e r c i s e .
FA S E B J . 1 9 ( 2 ) : 2 6 4 - 2 6 6 . 2 0 0 5 .
3 0 ) B u f o r d T W , C o o k e M B , S h e l m a d i n e B D , H u d s o n G M , R e d d L ,
W i l l o u g h b y D S . E f f e c t s o f e c c e n t r i c t r e a d m i l l e x e r c i s e o n i n -
f l a m m a t o r y g e n e e x p r e s s i o n i n h u m a n s k e l e t a l m u s c l e . A p p l
P h y s i o l N u t r M e t a b . 3 4 ( 4 ) : 7 4 5 - 7 5 3 . 2 0 0 9 .
3 1 ) F r e y R S , R a h m a n A , K e f e r J C , M i n s h a l l R D , M a l i k A B . P K C z e t a
r e g u l a t e s T N F - a l p h a - i n d u c e d a c t i v a t i o n o f N A D P H o x i d a s e i n
e n d o t h e l i a l c e l l s . C i r c R e s . 1 7 ; 9 0 ( 9 ) : 1 0 1 2 - 1 0 1 9 . 2 0 0 2 .
3 2 ) K o k o t K , S c h a e f e r R M , T e s c h n e r M , G i l g e U , P l a s s R , H e i d l a n d
A . A c t i v a t i o n o f l e u k o c y t e s d u r i n g p r o l o n g e d p h y s i c a l e x e r c i s e .
A d v E x p M e d B i o l . 2 4 0 : 5 7 - 6 3 . 1 9 8 8 .
3 3 ) M o r o z o v V I , T s y p l e n k o v P V, G o l b e r g N D , K a l i n s k i M I . T h e
e f f e c t s o f h i g h - i n t e n s i t y e x e r c i s e o n s k e l e t a l m u s c l e n e u t r o p h i l
m y e l o p e r o x i d a s e i n u n t r a i n e d a n d t r a i n e d r a t s . E u r J A p p l

30
P h y s i o l . 9 7 ( 6 ) : 7 1 6 - 7 2 2 . 2 0 0 6 .
3 4 ) T i d b a l l J G . I n t e r a c t i o n s b e t w e e n m u s c l e a n d t h e i m m u n e s y s t e m
d u r i n g m o d i f i e d m u s c u l o s k e l e t a l l o a d i n g . C l i n O r t h o p R e l a t
R e s . ( 4 0 3 S u p p l ) : S 1 0 0 - 1 0 9 . 2 0 0 2 .
3 5 ) T s i v i t s e S K , M c L o u g h l i n T J , P e t e r s o n J M , M y l o n a E , M c G r e g o r
S J , P i z z a F X . D o w n h i l l r u n n i n g i n r a t s : i n f l u e n c e o n n e u t r o -
p h i l s , m a c r o p h a g e s , a n d M y o D + c e l l s i n s k e l e t a l m u s c l e . E u r J
A p p l P h y s i o l . 9 0 ( 5 - 6 ) : 6 3 3 - 6 3 8 . 2 0 0 3 .
3 6 ) J a i n S K , R a i n s J , C r o a d J , L a r s o n B , J o n e s K . C u r c u m i n s u p -
p l e m e n t a t i o n l o w e r s T N F - a l p h a , I L - 6 , I L - 8 , a n d M C P - 1 s e c r e -
t i o n i n h i g h g l u c o s e - t r e a t e d c u l t u r e d m o n o c y t e s a n d b l o o d
l e v e l s o f T N F - a l p h a , I L - 6 , M C P - 1 , g l u c o s e , a n d g l y c o s y l a t e d
h e m o g l o b i n i n d i a b e t i c r a t s . A n t i o x i d R e d o x S i g n a l .
1 1 ( 2 ) : 2 4 1 - 2 4 9 . 2 0 0 9 .
3 7 ) F o r m a n H J , T o r r e s M . R e a c t i v e o x y g e n s p e c i e s a n d c e l l s i g -
n a l i n g : r e s p i r a t o r y b u r s t i n m a c r o p h a g e s i g n a l i n g . A m J R e s p i r
C r i t C a r e M e d . 1 6 6 ( 1 2 P t 2 ) : S 4 - 8 . 2 0 0 2 .

31
8 ) おわりに―今 後 の展 望 -
本 研 究 では、クルク ミンの及 ぼす影 響 を炎 症 性 サイトカイン 、酸 化 ス トレ
ス、免 疫 細 胞 浸 潤 の観 点 から検 討 した。しかしながら 、クルク ミン投 与 によ
るダウンヒル運 動 後 の筋 組 織 の分 解 および再 生 機 構 への影 響 、さらには
パフォーマンスへの影 響 に関 しての検 討 は行 っていない。
近 年 、クルク ミンは筋 委 縮 や加 齢 性 筋 肉 減 弱 症 (サルコペニア )の発
症 を抑 制 することが示 唆 された ( A l m d a r i N e t a l . 2 0 0 9 ) 。筋 委 縮 および
サルコペニアの発 症 には、 T N F - α などの炎 症 性 サイ トカインが重 要 な役 割
を示 す可 能 性 が示 唆 されている ( G o o d m a n M N e t a l . 1 9 9 1 ) 。 T N F - α は
筋 のタンパク質 分 解 に関 与 していることが知 られてお り 、 T N F - α 抗 体 の投
与 によ り筋 委 縮 を抑 制 したこと ( C o s t e l l i P e t a l . 1 9 9 3 ) からも 、 T N F - α が
筋 のタンパク質 分 解 を促 進 するこ とが考 えられる 。実 際 に 、 T N F - α は、筋
タンパク質 分 解 作 用 を有 しているユビキチン ・プロテアソーム経 路 の活 性 に
関 与 していることが明 らかになった ( L i Y P e t a l . 1 9 9 9 ) 。ユビキチン ・プロ
テアソーム系 路 は、筋 損 傷 を伴 う激 しい運 動 後 においても筋 タンパク質 分
解 を 引 き 起 こ す た め 、 筋 修 復 を 遅 延 す る 要 因 と さ れ る ( L i Y P e t a l .
2 0 0 3 ) 。 したがって 、 クルク ミンは激 しい運 動 後 の筋 タンパク質 の分 解 にも 、
N F - B の 抑 制 を 介 し て 関 与 し て い る 可 能 性 が 考 え ら れ る 。 今 後 は 、
T N F - α などの炎 症 性 サイ トカ インが増 加 するダウンヒル運 動 モデルを用 い
て 、 ク ル ク ミ ン 摂 取 によ る運 動 後 の 筋 修 復 へ の 関 与 を検 討 す る必 要 があ
る。
また 、これまでに活 性 酸 素 と運 動 パフォーマンスの関 係 は数 多 くの研 究
によ り検 討 されている ( P o w e r s S K e t a l . 2 0 0 8 ) 。適 度 な筋 肉 中 の活 性
酸 素 は 運 動 パ フ ォ ー マ ン ス を 向 上 さ せ る が 、 激 運 動 時 の 活 性 酸 素 の 過
剰 な増 加 はパフォーマンス低 下 をもたらすことが知 られている ( R e i d M B e t

32
a l . 1 9 9 3 ) 。実 際 に、筋 組 織 での過 剰 な H 2 O 2 によ り 、筋 収 縮 能 力 が低 下
することや ( O b a T e t a l . 1 9 9 6 ) 、活 性 酸 素 は運 動 誘 発 性 の筋 疲 労 に関
与 することも報 告 されている ( G a n d e v i a S C e t a l . 2 0 0 8 ) 。 したがって、抗
酸 化 作 用 を示 す薬 物 およびサプリメン トの摂 取 によ り 、激 運 動 時 の運 動 パ
フォ ーマ ンスが改 善 す る可 能 性 が考 えられ る 。 これま でに 、抗 酸 化 作 用 を
示 す N - a c e t y l c y s t e i n e 、ビタ ミン C およびビタミン E などの薬 物 、サプリ メ
ン ト投 与 に よる活 性 酸 素 産 生 抑 制 を介 したパフォーマンスへ の影 響 が検
討 されて きた 。 しか しながら 、実 際 に は 、ス ポーツ 選 手 へのサプ リ メ ン ト補 給
による活 性 酸 素 産 生 の影 響 は明 確 にされていない。 6 週 間 のビタミン E の
投 与 は、ウル ト ラマラソンのランナーでは筋 損 傷 への影 響 が無 いことが示 さ
れ ( M a s t a l o u d i s A e t a l . 2 0 0 6 ) 、生 体 に害 を及 ぼすとされる脂 質 膜 の
過 酸 化 に も 影 響 しな いこ とが報 告 さ れ ている ( L e e u w e n g u r g h C e t a l .
2 0 0 1 ) 。 また、ビタ ミン C 投 与 による激 運 動 後 の筋 損 傷 の抑 制 についても
数 多 く検 討 されているが ( P e a k J M e t a l . 2 0 0 3 ) 、明 確 な効 果 はみられず、
1 0 0 0 m g 以 上 の投 与 では逆 にその回 復 を阻 害 するという結 果 が報 告 され
ている ( C l o s e G L e t a l . 2 0 0 5 ) 。
近 年 、 D a v i s J M らによ り 、クルク ミン投 与 がマウスのダウンヒル運 動 のパ
フォーマンスを向 上 させることが報 告 された ( D a v i s J M e t a l . 2 0 0 7 ) 。その
要 因 と して 、 本 研 究 で 得 ら れ た 、 ク ル ク ミ ン の ダ ウン ヒ ル 運 動 後 の 抗 酸 化
作 用 が関 与 している可 能 性 が考 えられる 。 クルク ミ ン投 与 に よ るダウンヒル
運 動 後 の筋 組 織 の 酸 化 ス ト レス状 態 とパフォーマンス との関 係 について 、
更 なる検 討 が必 要 である。