covalent cross-linking of trnagly1 to the ribosomal p site via the dihydrouridine loop
TRANSCRIPT

Biochimica et Biophysica Acta 825 (1985) 161-168 161 Elsevier
BBA91471
Covalent cross-linking of tRNA~ ly to the ribosomal P site via the dihydrouridine loop
Jan-Kan Chen a, J~rgen H. Krauss b, Stephen S. Hixson b and Robert A. Zimmermann a,,
Departments of a Biochemistry and b Chemistry, University of Massachusetts, Amherst, MA 01003 (U.S.A.)
(Received February 8th, 1985)
Key words: tRNA modification; tRNA-rRNA cross-linking; Ribosome; Photoaffinity label; Protein synthesis; ( E. coli )
The dihydrouracil residue at position 20 of Escherichia coil tRNA °y has been replaced by the photoaffinity reagent, N-(4-azido-2-nitrophenyi)glycyl hydrazide (AGH). The location of the substituent was confirmed by the susceptibility of the modified tRNA to cleavage with aniline. When N-acetylglycyl-tRNA °y derivatized with AGH was bound noncovalentiy to the P site of E. coil 70 S ribosomes, 5-6% on average was photochemically cross-linked to the ribosomal particles in a reaction requiring poly(G,U), irradiation and the presence of the AGH label in the tRNA. Approximately two-thirds of the covalenfly attached tRNA was associated with 16 S RNA in the 30 S subunit. This material was judged to he in the P site by the criterion of puromycin reactivity. As partial RNAase digestion of the tRNA-16 S RNA complex produced labeled fragments from both 5' and 3' segments of the rRNA, there appeared to be more than one site of cross-linking in the 30 S subunit. The small amount of N-acetylglycyl-tRNA~ ~r associated with the 50 S subunit was also linked mainly to rRNA, but it was not puromycin-reactive.
Introduction
Although much has been learned about the size, shape and structure of the prokaryotic ribosome, few details are available about the organization of the protein and RNA constituents within individ- ual functional domains [1]. The binding sites for aminoacyl-tRNA and peptidyl-tRNA, known as the A and P sites, are of particular importance as they account for the correct positioning of the acceptor and donor groups during peptide bond
* To whom correspondence should be addressed. Abbreviations: Hepes, 4-(2-hydroxyethyl)-l-piperazineethane- sulfonic acid; Pipes, 1,4-piperazinediethanesulfonic acid; AGH, N-(4-azido-2-nitrophenyl)glycyl hydrazide; tRNA~ y, tRNA1 °ly reduced with NaBH4; tRNA°~H, tRNA~Y derivatized with AGH; AcGly, N-aeetylglycyl; D, dihydrouracil; acp3U, 3-(3- amino-3-carboxypropyl)uridine; DMSO, dimethyl sulfoxide.
formation. One technique for obtaining a more complete description of the A and P sites entails the establishment of covalent bonds between specific residues in aminoacyl- or peptidyl-tRNA and the ribosomal components at their sites of interaction. For this purpose, it is convenient to introduce affinity labels either at the aminoacyl moiety or at one of the easily modified minor bases frequently found in tRNA molecules. The ribosomal proteins or RNAs with which they react can then be identified and, in the most favorable cases, the precise amino acid or base involved in the cross-link can be determined. The results of such experiments, in combination with current knowledge of tRNA and ribosome structure, should ultimately yield a comprehensive portrait of ribosome architecture at the A and P sites.
Affinity labeling has already contributed im- portant information about the structure of tRNA
0167-4781/85/$03.30 © 1985 Elsevier Science Publishers B.V. (Biomedical Division)

162
binding sites in the Escherichia coli ribosome, but the data are still quite fragmentary. A variety of reagents attached to the aminoacyl group at the end of one arm of the L-shaped tRNA molecule have been found to label several proteins of the 50 S ribosomal subunit as well as the 23 S RNA [2,3]. Probes situated near the bend in the 'L' react mainly with proteins of both 30 S and 50 S sub- units [2,3], while the anticodon loop, at the end of the other arm of the tRNA, has been cross-linked to the 3' domain of 16 S RNA in the 30 S subunit [4-6]. In the present communication, we report the attachment of a photolabile substituent to a posi- tion not previously exploited in affinity labeling studies. Specifically, we have replaced the unique dihydrouracil of E. coli tRNA~lY--which occurs at position 20 * on the outer surface of the molecule just below the bend [7,8]--with the aryl azide reagent, N-(4-azido-2-nitrophenyl)glycyl hy- drazide. When derivatized N-acetylglycyl-tRNA~ ly is bound to the ribosomal P site and irradiated, this probe mediates photochemical cross-linking of the tRNA to the RNA components of both 30 S and 50 S subunits.
Materials and Methods
Materials E. coli tRNA~ ly with a stated amino acid accep-
tance of 1500 pmol/A260 unit was obtained from Subriden RNA. Unfractionated aminoacyl-tRNA synthetases and 70 S tight-couple ribosomes were prepared from E. coli MRE600 by standard pro- cedures [9,10]. [3H]Glycine (12.3 Ci/mmol), [14C]glycine (112 mCi/mmol), NaB3H4 (347 mCi/mmol), [y-aEp]ATP (2600 Ci/mmol) and Omnifluor were purchased from New England Nuclear, poly(G,U) from Miles Laboratories, ATP, Hepes and puromycin from Sigma, T4 poly- nucleotide kinase and RNAase Phy I from P-L Biochemicals, Pipes, RNAase T~ and RNAase U 2 from Calbiochem-Behring, and bacterial alkaline phosphatase and pancreatic RNAase A from Worthington. ITLG thin-layer chromatography sheets were supplied by Gelman. 14C-labeled 16 S RNA was isolated as described previously [11].
* Although D is actually residue 19 in E. cob tRNA~ ly, its position corresponds to that of residue 20 in the standard numbering system based on yeast tRNA Phe.
Synthesis of N-(4-azido-2-nitropher~vl)glycyl hv- drazide (A GH)
A solution containing 0.8 g (2.4 mmol) of the N-hydroxysuccinimide ester of N-(4-azido-2- nitrophenyl)glycine [12,13] in 60 ml of absolute ethanol was slowly mixed with 0.4 g (7.9 mmol) of hydrazine hydrate (64% hydrazine) in 40 ml of absolute ethanol. A further 50 ml of ethanol were added to homogenize the reaction mixture, which was then stirred overnight at 25°C. The dark red precipitate that formed was filtered and dried to give 0.42 g (71%) of AGH:
HzN-NH- C -CH2-NH-~.~-N 3
This product was characterized by a melting point of 157-158°C and an R F of 0.62 when subjected to thin-layer chromatography on ITLG sheets using a 10:1 (v/v) chloroform/ethanol mixture as eluent. An additional 0.20 g of impure AGH was obtained by concentrating the mother liquor from the initial precipitation. Spectral data for the pure AGH were: IR (KBr) 3490, 3210, 2115, 1680, 1650, 1635, 1565, 1530, 1410 and 1265 cm-l; NMR (DMSO-d6) 8 3.15 (m, 3H), 3.85 (m, 2H), 6.75 (d, 1H, J = 9Hz), 7.30 (dd, 1H, J = 3 and 9 Hz), and 7.60 (d, 1H, J = 3Hz); UV (MeOH) ~max 445 nm (c 5300). Use of the N-hydroxysuccini- mide ester of N-(4-azido-2-nitrophenyl)glycine labeled with 3H in the glycine moiety allowed the synthesis of [3H]AGH with a specific radioactivity of 75 mCi/mmol. In this case, approx. 25 mg of crude [3H]AGH obtained by evaporating the ethanol with a stream of nitrogen were dried briefly under reduced pressure and then chromato- graphed on a 0.35 cm i.d. × 120 cm long column packed with a slurry of 13 g of 100-200 mesh silica gel in chloroform. The column was washed successively with 11 ml of chloroform, 2.5 ml of a 20 : 1 (v/v) chloroform/methanol mixture, and 7.5 ml of a 10 : 1 (v/v) chloroform/methanol mixture, and the [3H]AGH was eluted with a 5:1 (v/v) chloroform/methanol mixture. An alternative pro- cedure for the synthesis of AGH has been reported [14].

163
Reduction of tRNA~ ty The reduction of dihydrouracil at position 20 of
tRNA~ ly to ureidopropanol ribonucleoside was carried out essentially as described by Yang and $611 [15]. Reaction mixtures contained approx. 15 A26 o units/ml of tRNA~ ly and 15 mg/ml of NaBH 4 in 200 mM Na2B40 7 (pH 9.8), 200 mM KC1, 10 mM MgCI 2. After incubation for 3 h at 20°C in the dark, an equal volume of 1 M HOAc was added to destroy excess borohydride and the reduced tRNA was recovered by precipitation with ethanol. To monitor the extent of reduction, 3H- labeled NaBH 4 was added to the reaction mixture and the incorporation of 3H into the ureidopropanol moiety was measured [16]. The release of 3H-labeled ureidopropanol was followed by incubation of reduced tRNA~ ly (2.5 A26 o units/ml) with a 200-fold molar excess of methyl- amine in 100 mM sodium formate (pH 3.0). re- duced tRNA~ ly will henceforth be referred to as tRNA~ y.
Derivatization of tRNA~ ty From 0.5 to 1.5 A260 units of unlabeled tRNA~ y
in 50/~1 of 100 mM sodium formate (pH 3.0), 25% DMSO (Eastman, redistilled) were incubated in the dark with a 150-400-fold molar excess of 3H-labeled AGH for 2-3 h at 37°C. Unreacted AGH was extracted with ether and an aliquot of the tRNA was precipitated with trichloroacetic acid to assay the incorporation of radioactivity. Large batches of derivatized tRNA~ Iy, which will be referred to as tRNA~AI~n, were washed re- peatedly by precipitation with ethanol and dis- solved in 0.1% KOAc (pH 5.0). To verify the site of modification, 20-#g samples of tRNA~ ly and tRNA~AI~H were treated with alkaline phosphatase for 30 min at 37°C, extracted with phenol, labeled with 32p at the 5' terminus by incubation with [y-32p]ATP and polynucleotide kinase for 20 rain at 37°C, and purified electrophoreticaUy [17]. [5'- 32 P]tRNA~ly was subjected to partial digestion with RNAases "1"1,122, A and Phy I and with formamide [18,19]. Both [5'-32p]tRNA~IY and [5'- 32p]tRNA~IY n were also treated with 1 M aniline for 6 min at 60°C and dried under vacuum [20]. The products were separated by electrophoresis on gels containing 20% polyacrylamide in 90 mM Tris-90 mM H3BO 3 (pH 8.3), 2.5 mM Na2EDTA, 7 M urea.
Aminoacylation of tRNA~ & Aminoacylation was performed by incubating
tRNA~ Iy (1.6 A260 units/ml) with 3 mM ATP, 14 /~M 3H- or 14C-labeled glycine and an appropriate amount of aminoacyl-tRNA synthetase in 100 mM Tris-HC1 (pH 7.6), 14 mM MgC12 for 15 min at 37°C [21]. Gly-tRNA~ ly was N-acetylated accord- ing to Haenni and Chapeville [22] and the extent of acetylation assessed by the method of Schofield and Zamecnik [23].
Formation and analysis of AcGly-tRNA~n-ribo- some complexes
As the formation and analysis of covalent tRNA-ribosome complexes has been described [5], only a brief resume of the relevant procedures will be presented here. Noncovalent binding of AcGly-tRNA~l~n to 70 S ribosomes was carried out by incubating 150 pmol/ml of Ac[3H]GIy- tRNA~AlYH, 300 /~g/ml of poly(G,U) and 700 pmol/ml of 70 S tight couples in 50 mM Hepes-NaOH (pH 7.6), 100 mM NH4C1, 15 mM MgCl 2 for 15-20 min at 37°C. Binding was esti- mated by the retention of radioactivity on nitrocel- lulose filters in the presence of 20 mM Mg 2+ and the association of Ac[3HIGly-tRNAGAI~n with the ribosomal P site was inferred from the release of bound amino acid by 1 mM puromycin. Cross-lin- king of Ac[3H]Gly-tRNAGAI~H to the ribosome was accomplished by irradiating noncovalent 70 S complexes for 4 h with a Hanovia 450-W medium-pressure mercury lamp surrounded by a Pyrex filter. The irradiated complexes were pre- cipitated with ethanol, resuspended in 10 mM Pipes-NaOH (pH 6.3), 0.25 mM MgC12 and centrifuged through a 12-ml 5-20% (w/v) sucrose gradient in the same buffer with 100 mM NaC1 for 16 h at 25 000 rev/min in a Beckman SW41 rotor. The gradient effluent was monitored for ab- sorbance at 260 nm and the acid-insoluble radio- activity in each fraction was measured. The cross- linking efficiency was calculated as the fraction of noncovalently bound Ac[3H]GIy-tRNACAIYH that co-sedimented with the dissociated ribosomal sub- units at 0.25 mM Mg 2+. The subunits were then pooled, precipitated with ethanol, resuspended in 10 mM Pipes/NaOH (pH 6.3), 10 mM MgC12, 0.1% sodium dodecyl sulfate, and extracted with phenol. Ribosomal RNA was recovered from the
164
aqueous phase and analyzed by sucrose-gradient centrifugation before and after partial RNAase digestion.
Results
Derioatization of tRNA Gty at position 20 Prior to derivatization, the dihydrouracil re-
sidue at position 20 of tRNA~ b was converted to ureidopropanol by reduction with NaB3H4 [15,16]. Fig. la demonstrates that the reaction was quanti- tative, reaching a plateau when 2000 pmol of 3H had been incorporated per .4260 unit of tRNA. The specificity of the reductive step was confirmed by the reactivity of the 3H-labeled ureidopropanol moiety toward primary amines. As shown in Fig. lb, over 75% of the radioactivity was converted to acid-soluble form upon incubation of reduced tRNA Gly with methylamine.
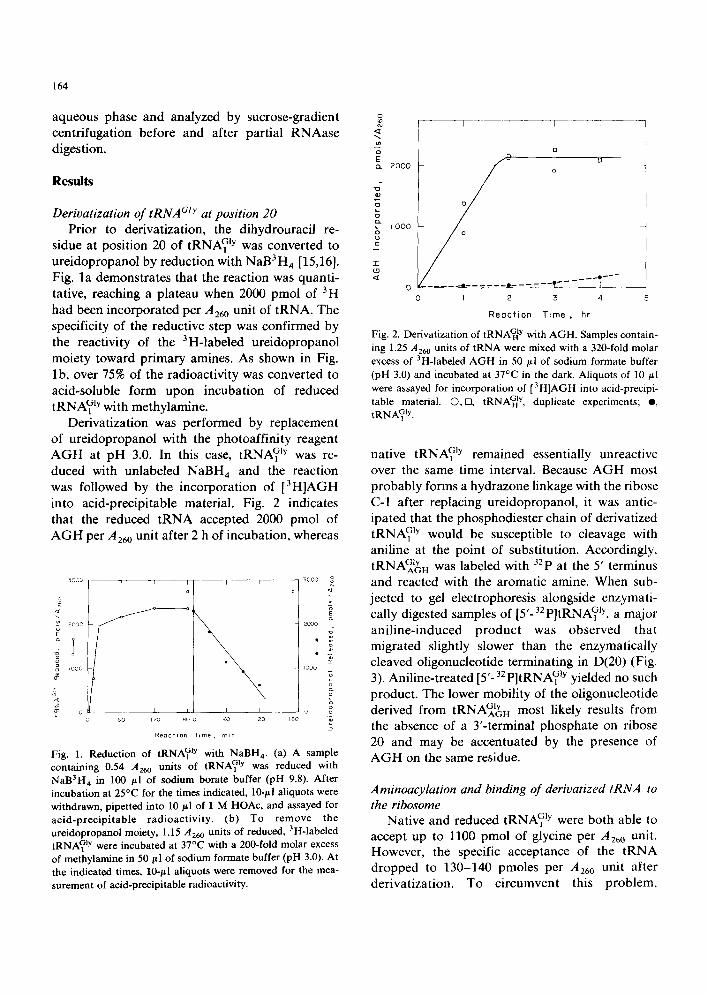
Derivatization was performed by replacement of ureidopropanol with the photoaffinity reagent AGH at pH 3.0. In this case, tRNA~ jy was re- duced with unlabeled NaBH 4 and the reaction was followed by the incorporation of [3H]AGH into acid-precipitable material. Fig. 2 indicates that the reduced tRNA accepted 2000 pmol of AGH per .4260 unit after 2 h of incubation, whereas
3000 - - i
o
~ , o o o
g
Z 0 - - I
6O
I t [
120 180 0 60 120
Reac t i on T ime , rn~n
3 0 0 0 o
c~ 2 o o 0
i ooo
c c c~ o
o. o
o ~
I SO
D
Fig. 1. Reduction of tRNA~ ly with NaBH 4. (a) A sample containing 0.54 A26 o units of tRNA~ ly was reduced with NaB3H4 in 100 #1 of sodium borate buffer (pH 9.8). After incubation at 25°C for the times indicated, 10-/~1 aliquots were withdrawn, pipetted into 10/~1 of 1 M HOAc, and assayed for acid-precipitable radioactivity. (b) To remove the ureidopropanol moiety, 1.15 A260 units of reduced, 3H-labeled tRNA~ Iy were incubated at 37°C with a 200-fold molar excess of methylamine in 50/~1 of sodium formate buffer (pH 3.0). At the indicated times, 10-/~1 aliquots were removed for the mea- surement of acid-precipitable radioactivity.
o
,,J [ F
t2
2000 o 1
" o
o
o o . g i ooo
35
~.D
o ...... • ...... • ...... Y I
0 I 2 B 4
R e a c t i o n T i m e , h r
Fig. 2. Derivatization of tRNA~ y with AGH. Samples contain- ing 1.25 AE~ units of tRNA were mixed with a 320-fold molar excess of 3H-labeled AGH in 50 ~1 of sodium formate buffer (pH 3.0) and incubated at 37°C in the dark. Aliquots of 10 ~1 were assayed for incorporation of [3H]AGH into acid-precipi- table material. O,m, tRNAGH ly, duplicate experiments; e, tRNA1 Gly.
native tRNA Gb remained essentially unreactive over the same time interval. Because AGH most probably forms a hydrazone linkage with the ribose C-1 after replacing ureidopropanol, it was antic- ipated that the phosphodiester chain of derivatized tRNA1Gjy would be susceptible to cleavage with aniline at the point of substitution. Accordingly, tRNAGAbH was labeled with 32p at the 5' terminus and reacted with the aromatic amine. When sub- jected to gel electrophoresis alongside enzymati- cally digested samples of [5'-32 P]tRNAGly, a major aniline-induced product was observed that migrated slightly slower than the enzymatically cleaved oligonucleotide terminating in D(20) (Fig. 3). Aniline-treated [5'-32 P]tRNAO)y yielded no such product. The lower mobility of the oligonucleotide derived from tRNAGAIYH most likely results from the absence of a 3'-terminal phosphate on ribose 20 and may be accentuated by the presence of AGH on the same residue.
Aminoacylation and binding of derivatized tRNA to the ribosome
Native and reduced tRNA~ lu were both able to accept up to 1100 pmol of glycine per A2~ unit. However, the specific acceptance of the tRNA dropped to 130-140 pmoles per A260 unit after derivatization. To circumvent this problem,

T ~ L ~ A P h L Not Dry - + - +
A ~oG - - G A
U
u ~
~oG--
- - AGH~o
a b
Fig. 3. Location of the derivatized residue in tRNAGA~r~. (a) Aliquots of [5'-32p]tRNA~ ly (approx. 10000 cpm each) were partially digested with RNAase T 1 (G), U 2 (A), A (C,U), Phy I (G,A,U) and formamide (L), and subjected to electrophoresis in a 20% (w/v) polyacrylamide gel at pH 8.3 to establish the position of 1)(20). The sequence is numbered according to the standard system based on yeast tRNA phe. (b) A sample of tRNA GIs was derivatized to the extent of 986 pmol/A260 unit with AGH and labeled with 32p at the 5' end. Aliquots of [5'-32P]tRNA~ ly (Nat) and [5'-32P]tRNAGAI~H (Drv) (approx. 10000 cpm each) were then electrophoresed before ( - ) and after ( + ) aniline treatment under precisely the same conditions as in (a) in order to determine the location of the derivatized residue.
t R N A ~ y was generally charged and acetylated be- fore modification with AGH. Under the relatively harsh reaction conditions employed, 40-50% of
165
the AcGIy- tRNA~ y was deacylated during deriva- tization. Nonetheless, in double-label experiments carried out with [t4C]glycine and [3H]AGH, it was determined that the resulting AcGly-tRNA~Yri population typically contained 500-600 pmol of glycine and 1500-2000 pmol of A G H per A260 unit. Ac[3H]Gly-tRNA~AJYH prepared in the same way was found to bind to 70 S tight-couple ribo- somes just as well as unmodified Ac[3H]GIy - tRNA GIy in the presence of poly(G,U). At a molar tRNA : ribosome input ratio of 0.2 : 1, for instance, 75% of both species bound to the 70 S particles (data not shown). Furthermore, both complexes were sensitive to reaction with puromycin, demon- strating that the tRNAs were located in the P site.
Covalent cross-linking of AcGIy-tRNAGa~n to the P site
Noncovalent complexes containing Ac[3H]GIy - tRNA~AI~H, poly(G,U) and 70 S ribosomes were irradiated with a medium-pressure mercury lamp. The 30 S and 50 S subunits were then dissociated and fractionated by centrifugation on sucrose gradients at low Mg 2+ concentration to determine the distribution of radioactivity. Of the AcGIy- tRNA~Af~H initially bound, 2-8% (average 5.5%, eight experiments) became covalently attached to the ribosomal subparticles. From 65 to 80% (aver- age 71%) of the cross-link was directed to the 30 S subunit, while 20-35% (average 29%) was directed to the 50 S subunit (Fig. 4a). Cross-linking did not occur in the absence of irradiation or polynucleo- tide template (Fig. 4b, c), nor when AcGIy- tRNA~ ly or AcGIy- tRNA~ y was substituted for AcGIy-tRNA°AI~H (data not shown). Over 70% of the 3H radioactivity associated with the 30 S sub- unit was released by puromycin, indicating that the tRNA was cross-linked to the P site in this case (Fig. 4d). By contrast, less than 35% of the Ac[3H]GIy-tRNA~AI~H that co-sedimented with the 50 S subunit was puromycin-reactive, suggesting that at least some of these molecules were attached to the ribosome in a nonspecific fashion.
Phenol extraction of each subunit fraction per- mitted further investigation of the cross-linked products, as tRNA-rRNA complexes partition to the aqueous phase while tRNA-protein complexes are sequestered in the phenol phase [24]. The aqueous phases were subsequently analyzed by

166
I I I
0.8 ffl
0.6
O
~ 0 .4
0.2
~o r,J
0 0
50S
I-!
I I !
I / l
/
2"
I
o 3? /
i/l. I I
I 0 20
I I
- h ~ '
tO
S =
I
20 I0
F r a c t i o n N u m b e r
:[ .:M
1
2 0 Io
2~, 10 4 _
I ,',. I O '~
2000
I 0 0 0
0
20 ~-' L-I
(2
Fig. 4. Covalent cross-linking of AcGly-tRNAG~H to E. coli ribosomes. Noncovalent Ac[3H]Gly- tRNA~H-poly(G,U) .70 S ribosome complexes were irradiated for 4 h and sedimented through 5-20% (w/v) sucrose gradients containing 0.25 mM Mg 2 +. The acid-precipitable radioactivity in each fraction was measured. (a) Complete system. (b) Unirradiated. (c) Poly(G,U) omitted. (d) Complete system incubated with 1 mM puromycin for 15 rain at 37°C after irradiation.
sucrose gradient centrifugation to verify that the tRNA co-sedimented with the 16 S and 23 S RNAs (see Fig. 5a). Data reported in Table I
demonstrate that an average of 67% of the cross- linked AcGIy-tRNA~AJYU was attached to 16 S RNA. Roughly one-fourth of the tRNA was asso-
o I I
c
E
,.a
<I
n (..)
2 0 0
I O 0
S ' - - - - -
I I o
Con t ro l
';i \
i,
I0 20
F r o c l i o n
I
+ RNose Tj
]6S 12S 8S
1
io Number
0 ] f 400 ~_~ 2 0 0
E
lO0
Z
50
I a
0
2o ~
Fig. 5. Partial RNAase T l digestion of AcGIy-tRNA~t~H-16 S RNA complex. Ac[aH]OIy-tRNACA]~H-16 S RNA (95 #g) was mixed with 14C-labeled 16 S RNA (1.5 #g) in 400 #1 of 10mM potassium phosphate (pH 6.2), 20 mM MgCI2, 350 mM KC1 containing 20 #g of bovine serum albumin, incubated for 10 rain at 40°C, and chilled on ice. Samples were then treated with (a) no addition or (b) RNAase T 1 at an enzyme : substrate ratio of 1:400 (w/w) for 15 rain at 0°C and fractionated by centri- fugation through 3-15% (w/v) sucrose gradients in the same buffer for 17 h at 28000 revs/min in a Beckman SW41 rotor. Each tube was assayed for acid-precipitable radioactivity.
TABLE I
CROSS-LINKING OF DERIVATIZED AcGIy-tRNAG~H TO RIBOSOMAL COMPONENTS
Covalent tRNA-subunit cross-linking was estimated from ra- dioactivity co-sedimenting with 30 S and 50 S peaks after irradiation of noncovalent Ac[3H]Gly-tRNACA~H.poly(G,U). 70 S ribosome complexes (see Fig. 4). Each subunit fraction was extracted with phenol, t R N A - r R N A cross-linking was estimated from radioactivity that remained in the aqueous phase and co-migrated with rRNA upon subsequent sucrose gradient centrifugation, tRNA-protein cross-linking was esti- mated from radioactivity that partitioned into the non-aqueous phase. Data represent averages of several experiments.
Component Percent of covalently bound AcGly-tRNA~AI~H in
subunits subunit components
30 S subunit 16 S RNA protein
50 S subunit 23 S RNA protein
71
29
67 4
24 5

ciated with 23 S RNA. Cross-linking to protein components of the 30 S and 50 S subunits was 5% or less in both cases. The Ac[3H]GIy-tRNACAI~H-16 S RNA complexes were next isolated and sub- jected to partial hydrolysis with RNAase T 1 in order to identify the portion of the rRNA to which the tRNA was attached. Under the conditions employed, the 16 S RNA is cleaved to 12 S and 8 S fragments that represent the 5' 60% and 3' 40% of the intact molecule, respectively [11]. As a sub- stantial fraction of the covalent Ac[3H]GIy - tRNAGAlYH co-migrated with each of the two rRNA fragments (Fig. 5), there appears to be more than one site of cross-linking in the 16 S RNA.
Discussion
E. coli tRNA~ Ir has been derivatized with the photoaffinity reagent AGH at position 20 in the D loop. As this modification represents a replace- ment of dihydrouracil by AGH, the probe may fit into the space originally occupied by the base. In any event, the photolabile group is located no farther than 12 A from the phosphodiester back- bone and therefore may not extend much beyond the boundaries of the unmodified tRNA molecule. When AcGIy-tRNA~ ly derivatized with AGH was bound to the ribosomal P site in presence of poly(G,U) and irradiated with visible light, up to 8% of the tRNA became cross-linked to the 70 S particles. Of the covalently bound AcGly-tRNA~ ly, approx. 70% was associated with the 30 S, and 30% with the 50 S, ribosomal subunits. Most of the tRNA linked to the 30 S subunit was judged to be in the P site because puromycin treatment of the irradiated 70 S complexes resulted in the removal of radioactive glycine from the 30 S fraction. The site to which the tRNA was attached in the 50 S particle is unclear, however, as the bulk of the glycine associated with the large subunit was not released by puromycin. In both subunits, the tRNA was cross-linked primarily to ribosomal RNA; in neither case did cross-linking to proteins represent more than a small fraction of the total. A pre- liminary attempt to localize the site of tRNA attachment indicated that more than one segment of the rRNA molecule had been labeled. This may reflect the close proximity of portions of the nucleic
167
acid chain that are widely separated in the nucleo- tide sequence, as suggested by current models of 16 S RNA structure [25].
Ribosomal components of the peptidyl-tRNA binding site include protein and RNA molecules of both subunits. Peptidyl-tRNA analogs formed by attaching reactive substituents to amino groups within the aminoacyl moiety label a region of the 50 S subunit that contains proteins L2, Ll l , L15, L18 and L27 as well as a segment of the 23 S RNA [2,3,26,27]. At the opposite end of the tRNA molecule, naturally modified uridine residues oc- curring in the 5' position of the anticodon can be photochemically cross-linked to 16 S RNA in the P site [4,5,28]. In the case of tRNA TM, 5-carboxy- methoxyuridine at position 34 has been shown to form a cyclobutane dimer with C(1400) in the rRNA [6,29]. In addition, E. coli tRNA Arg sub- stituted with a series of aryl azides at the 2-thio- cytidine residue in position 32 of the anticodon loop inserts into 30 S subunit proteins as well as 16 S RNA [30].
Information on the cross-linking of tRNA to the P site through its central region, where the two main helical segments of the molecule come to- gether, is rather sparse. Thus, it has been reported that a photoaffinity reagent attached to the 3-(3- amino-3-carboxypropyl)uridine residue at position 47 of E. coli tRNA Phe labels 23 S RNA and proteins of both 30 S and 50 S subunits, although the sites of covalent attachment have not yet been identified [3]. The present results demonstrate that 16 S RNA is also located near to the central region of the tRNA, but in this instance the probe faces outward from a point just below the vertex of the bend in the L-shaped molecule, whereas the sub- stituent attached to acp3U(47) faces inward.
Overall, data on the cross-linking of tRNA to the P site confirm the view that functional do- mains within the ribosome are composed of both protein and RNA in close array. Definition of the precise relationships between the tRNA and its neighboring ribosomal components at the pepti- dyl-tRNA binding site awaits a more exact analy- sis of the complexes already described as well as efforts to establish cross-links from additional positions in the tRNA molecule.

168
Acknowledgement
T h i s w o r k was s u p p o r t e d b y R e s e a r c h G r a n t
G M 2 2 8 0 7 f r o m t h e N a t i o n a l I n s t i t u t e s o f H e a l t h .
References
1 Garrett, R.A. (1979) in International Review of Biochem- istry, Vol. 25 (Offord, R.E., ed.), pp. 123-177, University Park Press, Baltimore
2 Kuechler, E. and Ofengand, J. (1979) in Transfer RNA: Structure, Properties, and Recognition (Schimmel, P.R., $611, D. and Abelson, J.N., eds.), pp. 413-444, Cold Spring Harbor Laboratory, New York
3 Ofengand, J. (1980) in Ribosomes: Structure, Function, and Genetics (Chambliss, G., Craven, G.R., Davies, J., Davis, K., Kahan, L. and Nomura, M., eds.), pp. 497-529, Univer- sity Park Press, Baltimore
4 Zimmermann, R.A., Gates, S.M., Schwartz, I. and Ofen- gand, J. (1979) Biochemistry 18, 4333-4339
5 Prince, J.B., Hixson, S.S. and Zimmermann, R.A. (1979) J. Biol. Chem. 254, 4745-4749
6 Prince, J.B., Taylor, B.H., Thurlow, D.L., Ofengand, J. and Zimmermann, R.A. (1982) Proc. Natl. Acad. Sci. USA 79, 5450-5454
7 Hill, C.W., Combriato, G., Steinhart, W., Riddle, D.L. and Carbon, J. (1973) J. Biol. Chem. 248, 4252-4262
8 Kim, S.-H. (1979) in Transfer RNA: Structure, Properties, and Recognition (Schimmel, P.R., $611, D. and Abelson, J.N., eds.), pp. 83-100, Cold Spring Harbor Laboratory, New York
9 Muench, K. and Berg, P. (1966) in Procedures in Nucleic Acid Research, Vol. 1 (Cantoni, G.L. and Davies, D.R., eds.), pp. 375-383, Harper and Row, New York.
10 Debey, P., Hoa, G.H.B., Douzou, P., Godefroy-Colburn, T., Graffe, M. and Grunberg-Manago, M. (1975) Biochemistry 14, 1553-1559.
11 Muto, A., Ehresmann, C., Fellner, P. and Zimmermann, R.A. (1974) J. Mol. Biol. 86, 411-432
12 Hsiung, N. and Cantor, C.R. (1974) Nucleic Acids Res. 1, 1753-1762.
13 Ofengand, J., Schwartz, I., Chinali, G., Hixson, S.S. and Hixson, S.H. (1977) Methods Enzymol. 46, 683-702
14 Chicheportiche, R., Balerna, M., Lombet, A., Romey, G. and Lazdunski, M. (1979) J. Biol. Chem. 254, 1552 1557
15 Yang, C.-H. and $611, D. (1974) Proc. Natl. Acad. Sci. USA 71, 2838-2842
16 Wintermeyer, W., Schleich, H.-G. and Zachau, H.G. (1979) Methods Enzymol. 59, 110-121
17 Silberklang, M., Gillum, A.M. and RajBhandary, U.L. (1979) Methods Enzymol. 59, 58-109
18 Donis-Keller, H., Maxam, A.M. and Gilbert, W. (1977) Nucleic Acids Res. 4, 2527-2538
19 Simoncsits, A., Brownlee, G.G., Brown, R.S., Rubin, J.R. and Guilley, H. (1977) Nature 269, 833-836
20 Peattie, D.A. (1979) Proc. Natl. Acad. Sci. USA 76, 1760-1764
21 Cedergren, R.J., Beauchemin, N. and Toupin, J. (1973) Biochemistry 12, 4566-4570
22 Haenni, A.-L. and Chapeville, F. (1966) Biochim. Biophys. Acta 114, 135-148
23 Schofield, P. and Zamecnik, P.C. (1968) Biochim. Biophys. Acta 155, 410-416
24 Schwartz, I., Gordon, E. and Ofengand, J. (1975) Biochem- istry 14, 2907-2914
25 Noller, H.F. (1984) Annu. Rev. Biochem. 53, 119-162 26 Johnson, A.E. and Cantor, C.R. (1980) J. Mol. Biol. 138,
273-297 27 Barta, A., Steiner, G., Brosius, J., Noller, H.F. and Kuechler,
E. (1984) Proc. Natl. Acad. Sci. USA 81, 3607-3611 28 Ofengand, J., Liou, R., Kohut, J., Ill, Schwartz, I. and
Zimmermann, R.A. (1979) Biochemistry 18, 4322-4332 29 Ofengand, J. and Liou, R. (1980) Biochemistry 19,
4814-4822 30 Chen, J.-K., Franke, L.A., Hixson, S.S. and Zimmerman,
R.A. (1985) Biochemistry, in the press