conference on practice improvement - stfm
TRANSCRIPT

The occurrence and significance of biogenic
opal in the regolith
Jonathan Clarke*
Department of Geology, CRC LEME, Australian National University, ACT 0200, Canberra, Australia
Received 23 November 2001; accepted 28 February 2002
Abstract
Biogenic opal produced by vascular plants, diatoms, and siliceous sponges have been found in soils and terrestrial sediments
of all continents except Antarctica since the middle of the 19th century. The opal particles range in size from fine silt to fine
sand. Almost all soils contain detectable opal up to levels of 2–3%, and a significant number contain values in excess of 5%.
Even higher values have been found from soils and sediments of all continents in a wide range of soil types. The most important
factor is poor soil drainage and seasonal to permanent water logging. This encourages the proliferation of silica producing
organisms. Such conditions have been found in the soils and aquatic sediments of the monsoonal tropics, tropical rain forests,
temperate forests, tropical savanna, tropical islands, semi-arid grasslands and savanna, and temperate woodland and grassland.
The presence of a volcanic substrate also appears favourable in some cases, but is not necessary. Biogenic opal preferentially
collects in the A horizon of soils and, to a lesser extent, in the B horizon. This preferential distribution facilitates identification
of palaeosols in stacked soil sequences. Biogenic opal is also a component of windblown dust, even in arid environments.
Biogenic opal is significant to regolith processes in a number of ways. Firstly, as in the case in marine environments, it is likely
to be important in silica cycling and storage because of its greater lability compared to quartz. Secondly, dissolution and
reprecipitation of opal A as opal CT or micro-quartz may play a role in cementation and silicification of regolith to form silica
hardpans and silcrete. Thirdly, the organisms that form biogenic opal can have considerable palaeoenvironmental significance
and be valuable in reconstructing regolith evolution. Finally, some forms of biogenic silica, in particular sponge spicules, can
present a health hazard. Their high abundance in some soils and sediments needs to be considered when assessing the health
implications of airborne dust.
D 2002 Elsevier Science B.V. All rights reserved.
Keywords: regolith; opal; phytolith; soil science; micropalaeontology
1. Introduction
There is an increasing awareness of the role of biota
in the regolith generally. Examples include the precip-
itation of iron and manganese (Skinner and Fitzpatrick,
1992) and interaction between bacteria and minerals
(McIntosh and Groat, 1997). Part of this role is the
deposition of biogenic silica in the form of opal.
Examples include phytoliths, diatoms, and siliceous
sponge spicules. Brewer et al. (1993) described phyto-
liths in Australian soils as ‘‘almost ubiquitous’’. In
contrast, the same authors describe diatoms as ‘‘have
been recorded’’ and sponge spicules as ‘‘rare’’ and
‘‘almost certainly inherited’’. Wilding et al. (1989)
0012-8252/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
PII: S0012 -8252 (02 )00092 -2
* Fax: +61-2-6125-5544.
E-mail address: [email protected] (J. Clarke).
www.elsevier.com/locate/earscirev
Earth-Science Reviews 60 (2003) 175–194

regarded most soils as containing up to 3% biogenic
opal and cited examples where up to 20% was present.
These values mean that opal comprises a significant
part of the silica component of many soils and can com-
monly exceed the abundances of potassium, calcium,
magnesium, sodium and phosphate. However, bio-
genic silica is normally regarded as a minor component
and the potential role of organisms in the large-scale
deposition of silica in the regolith is largely ignored.
Despite this, there is considerable evidence that
opal from phytoliths, diatoms, sponge spicules, and
other organisms are often abundant in the regolith in
many localities. Widespread and common accumula-
tions >2% by grain abundance are known from every
continent except Antarctica in environments as
diverse as coastal and inland swamps, forests, grass-
lands, and flood plains. They are present in the
tropical, temperate, and semi arid regions.
Assessing the actual abundance of biogenic silica
in regolith materials is difficult. Most studies concen-
trate on one component, such as phytoliths, and do not
discuss the presence of other forms. Some authors
(e.g., Brewer, 1955) give actual percentages (Table 1),
others list proportions (Table 2) (e.g., Schwandes and
Collins, 1994) in opal grains per gram, still others
(e.g., Hart, 1992) as percentage of a particular size
fraction, such as silt, or (Clark et al., 1992a) as cm2
cm3. None of the works, to date, appears to have used
the various chemical techniques used in the study of
biogenic opal in marine sediments (see Muller and
Schneider, 1993 and references therein).
This paper reviews occurrences of biogenic opal in
the regolith and their potential importance to regolith
processes. Far from being of minor importance, I argue
that biogenic opal is a key constituent of the regolith.
2. Status of terrestrial biogenic opal research
For more than 150 years, biogenic silica has been
known to be a component of the regolith. The earliest
mention of biogenic opal in soils was in 1841 by the
German microscopist Ehrenherg (Piperno, 1988). A
few years later, Gregory (1855) reported spicules and
diatoms as well as phytoliths in soils. Early studies on
spicules were listed by Smithson (1959), while Oehler
Table 1
Abundances of opaline material in selected soils (representative only)
% Opal Forms Setting Locality Source
19 Spicules Flooded forest Central Amazonia Chauvel et al. (1996)
3.2 Phytoliths, spicules,
diatoms
Coastal swamps Dalmore, Victoria,
Australia
Baker (1959b)
2.1 Phytoliths, diatoms Forested hills Mt. Gellibrand,
Victoria, Australia
Baker (1959b)
24 Spicules, diatoms,
phytoliths
Alluvial plain Duntroon, ACT,
Australia
Brewer (1955)
11 Spicules, diatoms,
phytoliths
Alluvial plain Deep Creek, NSW,
Australia
Brewer (1955)
2 Spicules, diatoms,
phytoliths
Alluvial plain Doughboy Creek,
NSW, Australia
Brewer (1955)
43 Phytoliths Bamboo forested
slope
Reunion Island Meunier et al. (1999)
0.9 Phytoliths Forested plain Amazonia Kondo and Iwasa
(1981)
4.5 Phytoliths Swamp Northern Sydney,
Australia
Hart (1992)
20 Phytoliths Grassland Oregon, USA Hart (1992)
100 Phytoliths Basalt flow East Africa Hart (1992)
30–60 Phytoliths Volcanic slopes Japan Hart (1992)
48 Phytoliths, spicules,
diatoms
Seasonal
wetlands
Magela Creek,
NT, Australia
Clark et al. (1992b)
100 Phytoliths, spicules,
diatoms
Seasonal
wetlands
Magela Creek,
NT, Australia
Clark et al. (1992b)
J. Clarke / Earth-Science Reviews 60 (2003) 175–194176

(1979) provided a review of the significance, origin,
and biogeochemical cycling of silica in the terrestrial
environment. Much has been documented on the
taxonomy of biogenic opal in soils, and some detailed
palaeoenvironmental and palaeoecological studies
have been carried, most on local scales (e.g., Gasse,
1987; Volkmer-Ribeiro, 1992). However, much of this
work has been fragmentary and focused on specific
applications, such as palaeoenvironmental and ar-
chaeological analysis. There has been little integration
of data from soil science, palaeoenvironmental, palae-
ontological, sedimentological, and regolith studies.
3. Sources of biogenic silica in soils
3.1. Plant opal
Many groups of plants produce silt-sized opal
grains known as phytoliths (Fig. 1A). These form
structural elements and provide a defense against
herbivores (Lewin and Reimann, 1969). They are
extremely abundant in grasses, which can contain up
to 10% silica by weight (Lovering, 1959) and horse-
tails (Equisetum) can contain up to 16% dry weight of
biogenic opal (Lewin and Reimann, 1969). However,
phytoliths also occur in a wide range of other plant
groups (see Piperno, 1988) and it would be a mistake
to assume that they are abundant only in grasses and
horsetails. Phytoliths are found in most soils, includ-
ing those from temperate forests of North America
(Wilding and Drees, 1971), the savanna landscapes of
Kenya (Runge, 1999) and tropical forests of Amazo-
nia (Kondo and Iwasa, 1981) and Africa (Runge,
1999). They locally reach abundances of 43% in the
B-horizon of some soils on Reunion Island (Meunier
et al., 1999). They offer considerable potential for
palaeoenvironmental reconstruction, owing to differ-
ent morphologies derived from different plant com-
munities (Runge, 1999; Wilding and Drees, 1971).
Phytoliths are the most common form of biogenic
silica in most terrestrial environments. A much rarer
occurrence is tabashir, massive bodies of opal found
in bamboo (Jones et al., 1966). Phytoliths may be
confused with sponge spicules, especially in older
literature (see review in Hart and Humphreys, 1997).
Both have a rod- or needle-shaped morphologies,
however sponge spicules typically have a central
canal, absent in phytoliths. Some spicules are highly
complex and irregular in shape, especially those of the
lithistid sponges, common in Eocene marine sedi-
ments in southern Australia, these, along with gem-
mule bodies and microscleres lack a central canal, and
may be confused with phytoliths.
Table 2
Other selected abundances of opaline material in soils (non-compatible units)
Amount Units Forms Setting Locality Source
0.5–6 % silt
fraction
Phytoliths Tropical
forest soils
D. R. Congo Runge (1999)
>0.1–2 % sand
fraction
Phytoliths Tropical
forest soils
D. R. Congo Runge (1999)
14 % silt
fraction
Phytoliths Tropical
forest soils
Central African Republic Runge (1999)
1.65 % sand
fraction
Phytoliths Tropical
forest soils
Central African Republic Runge (1999)
500,000 Grains/gram Spicules Histsol Florida Schwandes and Collins
(1994)
4600 Grains/gram Spicules Entisol Florida Schwandes and Collins
(1994)
48,000 Grains/gram Spicules Spodsol Florida Schwandes and Collins
(1994)
9500 Grains/gram Spicules Mollisol Florida Schwandes and Collins
(1994)
23,000 Grains/gram Spicules Ultisol Florida Schwandes and Collins
(1994)
f 10,000 Grains cm2 cm3 Phytoliths,
spicules, diatoms
Black soils Magela Creek, Northern
Territory
Clark et al. (1992a)
J. Clarke / Earth-Science Reviews 60 (2003) 175–194 177

Fig. 1. Representative examples of opal secreting organisms showing the immense range in morphology. (A) Phytoliths (after Baker, 1959a). (B)
Marine and freshwater diatoms (after Braiser, 1980). (C) Marine and freshwater sponge spicules (after de Laubenfels, 1955). (D)
Silicoflagellates, (E) chrysomonads, (F) radiolaria, and (G) ebridians (all after Braiser, 1980). (H) Helizoans (after Moore, 1964).
J. Clarke / Earth-Science Reviews 60 (2003) 175–194178

Fig. 1 (continued).
J. Clarke / Earth-Science Reviews 60 (2003) 175–194 179

3.2. Diatoms
Diatoms (Fig. 1B) are found in lakes, rivers, and
soils. They also occur subaerially on plant and rock
surfaces in moist environments (Patrick, 1977). Dia-
toms comprise the bulk of the 24% of siliceous
remains reported by Brewer (1955) from the A2
horizons of soils in Canberra, Australia. Compara-
tively pure accumulations of diatoms (diatomites) are
known only from lake and swamp basins, however
such sediments may form part of the regolith, espe-
cially where they occur in stable continental environ-
ments. Modern Australian examples include the
Holocene diatomites of the Swan coast plain of West-
ern Australia (Gibson, 1976) and the Neogene diatom-
ites of Victoria (Kenley, 1976) which are intimately
associated with lakes and sediments formed by Qua-
ternary basaltic volcanism and associated drainage
diversion. Miocene Bonnie Doon Diatomite from
New South Wales is not, however, associated with
volcanism (Taylor et al., 1990). All these deposits
contain varying amounts of clastic material and sponge
spicules in addition to the diatom frustrules. Secondary
silica mobilisation is common in the Victorian depos-
its, as they have been exposed to percolating ground-
water, forming opaline bands. Diatoms are known
from all continents. The expansion and contraction
of large lakes in low relief continental environments,
such as the modern lake Chad, can result in deposition
of diatomaceous sediments over large areas and then
expose them to pedogenic processes (Thiry, 1999).
3.3. Sponges
Siliceous sponges (Fig. 1C) are normally only a
minor component of marine and terrestrial ecosys-
tems. Their presence in sediments and soils (Smith-
son, 1959) are similarly normally minor. Under
conditions not fully understood they can, however,
proliferate and even dominate. Late Eocene spicular
marine and marginal marine sediments, locally with
up to 100% sponge spicules (Clarke, 1994a), form the
land surface over large areas of southern Australia.
The reasons for this proliferation during a narrow time
interval are not fully known, but probably related to a
unique confluence of runoff, turbidity, and nutrient
conditions (Gammon et al., 2000).
Freshwater sponges produce much smaller spicules
than marine sponges, typically silt rather than sand-
sized. Thus, they are often missed by those looking
only at the larger fraction. They are known not only
from lakes (Harrison, 1988) and rivers (Chauvel et al.,
1996) but also bogs (Volkmer-Ribeiro, 1992) and
waterlogged soils (Schwandes and Collins, 1994).
They have been reported as composing more than
20% of the silt fraction of the soil in parts of Amazonia
(Chauvel et al., 1996) and in numbers of more than one
million spicules to the gram (Schwandes and Collins,
1994). They have been found to date in the soils of all
continents except Antarctica.
Even though the presence of such spicules has been
taken to indicate high levels of dissolved silica (see
Turner, 1985), this need not be the case. Spicule-rich
Fig. 1 (continued).
J. Clarke / Earth-Science Reviews 60 (2003) 175–194180

sediments were reported from rivers in Amazonia by
Chauvel et al. (1996) that have extremely low dissolved
silica levels of 2.1 ppb. In comparison, Aston (1983)
gives an average global silicon value for river water of
13.1 ppm, while Wollast and Mackenzie (1983) say
5.42 ppm.
3.4. Other organisms (Figs. 1 D–G)
Many groups of regolith bacteria are known to
dissolve silicates (see review of Silverman, 1979)
through enzyme and organic acid secretion. Some,
such as Proteus mirabilis, also store the silica within
their cells and in slime layers (Tesic and Todorovic,
1958; Lauwers and Heinen, 1974) as monomeric
silica. The fate of such silica in the regolith is not
known but it may provide both a source of silica for
uptake by higher plants and, potentially, as a means of
silica cementation should the silica in the slime layers
and dead bacterial cells between regolith grains
undergo polymerisation. The abundance of bacterial
remains in hydrothermal and marine chert deposits
suggests that bacteria do play a role in silica deposi-
tion (Ferris, 1997). Shaw et al. (1990) reported that
desiccation of formerly floating colonies of the fila-
mentous cyanobacteria Chloriflexus provided a tem-
plate for silica deposition in a silica-depositing
alkaline saline pan in Botswana. Widespread silicifi-
cation of bacterial cells has been reported (Ferris,
1997) and strongly supports a bacterial role in the
deposition of silica in the regolith.
Radiolaria are significant siliceous organisms in
the pelagic realm (Braiser, 1980). Not found in
terrestrial aquatic environments, they are likewise rare
in the coastal sediments likely to be incorporated into
the terrestrial regolith. However, the Australian rego-
lith includes epicontinental sediments of Cretaceous
age, and these can include radiolarian-rich deposits.
An example is the radiolarian-bearing Darwin Mem-
ber of the Bathurst Island Formation of the Northern
Territory (Nott, 1994) which may be the source of
silica in siliceous bands in the weathering profile.
Helizoans are Protozoa with a siliceous test similar to
radiolaria but restricted to freshwater environments.
They are rarely preserved because their fragility
normally results in the rapid disintegration of the test
after death (Braiser, 1980). Heliozoans however have
occasionally been reported from Quaternary sedi-
ments and are thought to indicate swampy to lacus-
trine environments (Piperno, 1988; Moore, 1964).
Another group of Protozoa are the testate amobae
known as rhizopods (Charman, 2000). While some
rhizopods form tests (xenosomic tests) of cemented
grains from the local environment, others have very
weak siliceous tests. These are known as idiosomic
tests and are formed by the parent amobae during
reproduction. Rhizopods are found in lakes and peat-
lands and in recent research has demonstrated their
potential for palaeoecological studies. Their larger
scale significance to silica cycling is unclear, as they
are only weakly mineralised.
Silicoflagellates are also important silica depositing
organisms in pelagic marine environments. They are
not, however known from terrestrial environments,
nor are they, unlike diatoms and sponges, known at
present to be locally common enough in coastal sedi-
ments to become significant components of the rego-
lith as a result of sea level changes (Braiser, 1980).
Whether they, as radiolaria do, occur in significant
numbers in marine epicontinental deposits such as the
Cretaceous of the Australian regolith, is not known.
Chrysomonads are mostly non-marine Chrysophyte
algae related to silicoflagellates (Braiser, 1980). They
appear to be of minor importance in siliceous non-
marine sediments.
Ebridians are a small and comparatively minor
group of siliceous marine plankton allied to dinofla-
gellates (Braiser, 1980) and are not presently known
to form significant accumulations on their own. They
are common in some diatomite deposits (Bohaty and
Harwood, 2000). Some Ebridians produce external
siliceous scales or spines while all produce siliceous
resting cysts termed statosphores (Smol, 1987).
4. Spatial distribution in the regolith
4.1. Opal in the landscape
No detailed study has been carried on the spatial
distribution of biogenic opal in soils. However, fol-
lowing Hart’s (1992) analysis of the data of Stace et
al. (1968), detectable opal is common in many
Australian soils profiles. Stace et al. (1968) compiled
147 representative soil profiles from 43 soil groups.
Micromorphological data was provided for 85 of
J. Clarke / Earth-Science Reviews 60 (2003) 175–194 181
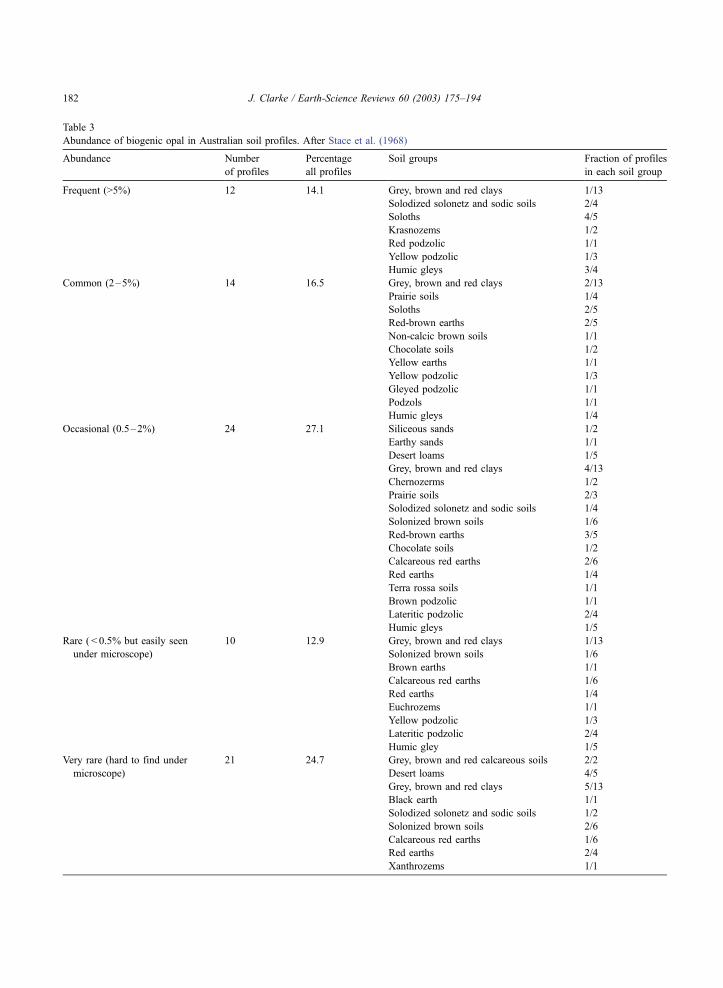
Table 3
Abundance of biogenic opal in Australian soil profiles. After Stace et al. (1968)
Abundance Number
of profiles
Percentage
all profiles
Soil groups Fraction of profiles
in each soil group
Frequent (>5%) 12 14.1 Grey, brown and red clays 1/13
Solodized solonetz and sodic soils 2/4
Soloths 4/5
Krasnozems 1/2
Red podzolic 1/1
Yellow podzolic 1/3
Humic gleys 3/4
Common (2–5%) 14 16.5 Grey, brown and red clays 2/13
Prairie soils 1/4
Soloths 2/5
Red-brown earths 2/5
Non-calcic brown soils 1/1
Chocolate soils 1/2
Yellow earths 1/1
Yellow podzolic 1/3
Gleyed podzolic 1/1
Podzols 1/1
Humic gleys 1/4
Occasional (0.5–2%) 24 27.1 Siliceous sands 1/2
Earthy sands 1/1
Desert loams 1/5
Grey, brown and red clays 4/13
Chernozerms 1/2
Prairie soils 2/3
Solodized solonetz and sodic soils 1/4
Solonized brown soils 1/6
Red-brown earths 3/5
Chocolate soils 1/2
Calcareous red earths 2/6
Red earths 1/4
Terra rossa soils 1/1
Brown podzolic 1/1
Lateritic podzolic 2/4
Humic gleys 1/5
Rare ( < 0.5% but easily seen 10 12.9 Grey, brown and red clays 1/13
under microscope) Solonized brown soils 1/6
Brown earths 1/1
Calcareous red earths 1/6
Red earths 1/4
Euchrozems 1/1
Yellow podzolic 1/3
Lateritic podzolic 2/4
Humic gley 1/5
Very rare (hard to find under 21 24.7 Grey, brown and red calcareous soils 2/2
microscope) Desert loams 4/5
Grey, brown and red clays 5/13
Black earth 1/1
Solodized solonetz and sodic soils 1/2
Solonized brown soils 2/6
Calcareous red earths 1/6
Red earths 2/4
Xanthrozems 1/1
J. Clarke / Earth-Science Reviews 60 (2003) 175–194182

profiles. Opal was detected in 81 of these (Table 3)
representing over 95% of soils studied. Just over
30% (26 examples) contained more than 2% opal,
and just over 56% (47 examples) contained more
than 0.5%.
Abundant (>2%) opal was found in soils from
western Australia, Victoria, New SouthWales, Queens-
land, South Australia, and the Australian Capital
Territory. All but three of these soils (88.5%) were
characterised by slow or impeded, drainage. In con-
trast, poor drainage was a feature of only 41.6%
profiles containing occasional opal grains. Profiles
with rare and very rare grains of biogenic opal were
poorly drained in only 30% and 33% of cases, respec-
tively.
These results indicate that there is a strong corre-
lation between the occurrence of high levels of bio-
genic opal in soils with poor drainage. Stace et al.
(1968) performed no soil petrography on alluvial or
organic-rich soils. Data on occurrences of exception-
ally high levels of biogenic opal indicate that they
often occur in such soils also. Examples include those
of the flood plains (Chauvel et al., 1996) and bogs
(Volkmer-Ribeiro, 1992) of Brazil, and the Magela
Creek flood plain of the Northern Territory of Aus-
tralia (Clark et al., 1992a,b) (Fig. 2). As post-organic-
rich soils, such as histosols are poorly drained, the
correlation with poor drainage and water logging,
noted by Brewer (1955), is thus probably much
stronger than these data indicate.
4.2. Opal in soil profiles
There have been many studies noting the pres-
ence of biogenic opal in soils, and the following
references provide an outline only. Biogenic opal is
normally preferentially concentrated in the A-hori-
zon of individual soil profiles (Oehler, 1979). It is
also present less commonly in the B-horizon, and
sometimes both (Simons et al., 2000). Accumulation
in the B-horizon is typically due to downward
movement resulting from bioturbation and percolat-
ing soil water (Hart and Humphreys, 1997; Boet-
tinger, 1994; Piperno, 1988). In some cases the
biogenic opal may be concentrated entirely in the
B-horizon (see Meunier et al., 1999; Schwandes and
Collins, 1994), but this is exceptional. Bobrova and
Bobrov (1997) and Bobrov and Bobrova (2001) re-
ported the concentration of phytoliths in illuvial and
eluvial horizons.
Because phytoliths are commonly perceived to be
concentrated in the A horizon of soils, increases in
their abundance within a profile have sometimes been
used as indicators of palaeosols in a succession (see
Dormaar and Lutwick, 1969). Care should be taken in
such interpretations, because of the downward move-
Abundance Number
of profiles
Percentage
all profiles
Soil groups Fraction of profiles
in each soil group
Krasnozems 1/2
Alpine humus soils 1/1
Absent 5 4.7 Siliceous sands 1/1
Solonized brown soils 2/6
Calcareous red earths 2/4
Undescribed Solonchalks
Alluvial soils
Lithosols
Red-brown hardpan soils
Redzinas
Wiesenboden
Calcareous sands
Grey-brown podzolic
Humus podzols
Peaty podzols
Neutral to alkaline peats
Acid peats
Table 3 (continued)
J. Clarke / Earth-Science Reviews 60 (2003) 175–194 183

ment in such circumstances noted above by Simons et
al. 2000) and should only be carried out with good
soil stratigraphic control.
5. Composition and chemistry
Biogenic silica is formed mainly as amorphous
opaline silica (opal A), but plant opal is also known
to contain opal CT (Wilding and Drees, 1974).
Diagenesis of the opal in soils results in further
production of opal CT and eventually its stablisation
as quartz (Wilding et al., 1989). Studies of the
subsequent precipitation of dissolved silica from
biogenic opal are rare, but amorphous opal, opal
CT and quartz are all possible secondary phases,
with crystalline forms becoming more likely with
time.
Plant opal can contain significant amounts of Al,
Fe, Ti, Mn, P, Cu, N, and C (Wilding et al., 1989).
The Al is known to play a key role in the surface
chemistry of plant opal, influencing dissolution and
interaction with organic acids (Bartoli, 1985). Much
or all of the Al is chemisorbed on the surface,
rendering the plant opal as reactive in the soil as iron
and aluminium hydroxides and allophanes. Nitrogen,
phosphorous, and carbon may be the result of inclu-
sion of lignin and cellulose during formation of the
phytoliths. Trace element composition of other forms
of biogenic opal, such as spicules and diatoms, is not
known.
Biogenic opal also shows high delta O18 values.
Diatoms typically show approximate values of + 29
to + 32 relative to SMOW, and phytoliths values of
+ 36 to + 39 (Wilding et al., 1989). This strongly
suggests that the isotopic value of regolith opal, at
Fig. 2. An opal factory (1): wetlands of Kakadu National Park, Australia contain up to 100% opal. Open water billabongs (A) and seasonal grass
and sedge wetlands (B) have the highest opal productivity. Paperbark wetlands (C) are less productive. Biogenic opal is visible as pale streaks in
the black soils of the seasonal wetlands (D). All photographs courtesy of R. Wasson.
J. Clarke / Earth-Science Reviews 60 (2003) 175–194184

least in its particulate or dispersed form, can be used
to demonstrate biological origin. Webb and Long-
staffe (1997) argued that progressive depletion
reflected increasing values of evapotranspiration.
Bombin and Muehlenbachs (1980) earlier noted that
the oxygen isotope values varied according to both
temperature and humidity. This makes the use of these
isotopes as direct palaeoclimatic indicators problem-
atic, although they certainly appear to have potential
as evapotranspiration indicators. It is not known
whether these signatures persist through diagenesis
of the opal. Webb and Longstaffe also reported
extreme depletion in deuterium (� 125), the reasons
for which were not known at the date of publication.
This contrasts with the study of Fredlund (1993), who
found deuterium values consist with that of the waters
that supported plant growth, allowing for the usual
levels of biological fractionation. Carbon isotope
ratios of phytoliths were reported by Fredlund as
reflecting the C3/C4 ratio within the source vegeta-
tion. As is the case with oxygen isotopes, it is not yet
known whether these hydrogen and carbon survive
diagenesis.
As noted by Oehler (1979), non-crystalline forms
of silica are much more soluble than crystalline forms.
Wilding et al. (1989) said that amorphous silica was
more soluble than quartz by a factor of 10 or more.
Biogenic opal showed a great range in solubility.
Generally, those remains containing less organic car-
bon were more soluble than those that still contained
significant organic matter. In addition, some types of
biogenic opal are more soluble than others. Wilding
and Drees (1974) found that forest opal was 10–15
times more soluble than grass opal, owing to its
greater surface area. In sediments, biogenic opal is
more labile than quartz. Kosters and Bailey (1983)
identified sponge spicules as among the most chemi-
cally mobile silica sources in sediments from the
Mississippi Delta.
Once silica has been dissolved from biogenic opal
grains, it can then be re-precipitated lower in the soil
profile. Alexandre et al. (1997) described incipient
cementation of soil particles in the lower part of a soil
profile by opal remobilised from phytoliths and other
organisms. Dissolution and cementation of silica in
soils can occur quite rapidly under favourable con-
ditions, as shown by Breese (1960) in studies of
aeolian dust.
6. Biogeochemical cycling of silica in the regolith
Organisms play a vital role in both the dissolution
(Silverman, 1979) and deposition (Oehler, 1979) of
silica in the regolith. Ambivalent results were reported
in early studies cited by Jones and Handreck (1967)
on the effect of growing plants in silicon-free sub-
strates. Some studies showed no ill effect whereas
others did. One of the main functions of biogenic opal
in plants is in improving pest resistance, thus under
field conditions improved silicon availability to plants
is desirable. A more recent work cited in Epstein
(1999) shows that silicon-deprived plants are structur-
ally weaker, have various abnormalities, and are more
susceptible to biotic and abiotic stress. Results from
experimental studies in silicon-free media replicate
studies of plant pathologies found in silicon-poor
soils, especially lateritic and bauxitic soils of tropical
regions. In Epstein’s words, ‘‘the evidence is over-
whelming that silicon should be included among the
elements having a major bearing on plant life
(Epstein, 1999). Because of its greater chemical
mobility than crystalline silica phases, biogenic opal
plays a major role in the cycling of silica in soils
(Alexandre et al., 1997), just as it does in aquatic
environments (Konhauser et al., 1992).
As pointed out by Heinen and Oehler (1979), the
cycling of silica through the biosphere has evolved
through time. Silica bacteria may have been present
since the Precambrian. Marine sponges are known
since just before the end of the Neoproterozoic (Braiser
et al., 1997) and have formed strandline accumulations
since the Carboniferous (Carlson, 1994). Sponges have
been reported from freshwater environments of at least
Carboniferous age (De Laubenfels, 1955) and probably
form a significant source of silica in coals throughout
geological history (Davis et al., 1984). Equisetum, the
single modern representative of the Equisetales, a
dominant component of land vegetation from the
Devonian through to Permian, contains abundant silica
(Lewin and Reimann, 1969). If these values of silica
content reflect a characteristic of the taxon as a whole,
evolution of these plants would have significantly
increased the rates of silica cycling in the regolith.
Rates of silica cycling would have increased still
further with the evolution of grasses and their ecolog-
ical dominance from the Oligocene onwards (Braiser,
1980). Although diatoms may have appeared in the
J. Clarke / Earth-Science Reviews 60 (2003) 175–194 185

Jurassic, they are common in terrestrial environments
only from the Miocene (Braiser, 1980).
Thus, the pattern rate of silica cycling and the
potential for accumulations of biogenic silica in the
regolith has increased markedly with time. Major
increases would have occurred in the Mid to late
Palaeozoic and near to the Paleogene–Neogene boun-
dary.
Alexandre et al. (1997) reported that 92% of the
biogenic silica in the soil is recycled by plants and is
the main source of this nutrient. Only the remaining
8% accumulates in the soil. Konhauser et al. (1992),
in a study of the seasonally flooded forests of the
Amazon basin reported that diatoms played a key role
in silica exchange between the dissolved and precipi-
tated state, with the precipitated silica being stored as
gel coatings on wood. The significance of sponges in
silica cycling is not known, but the abundance of
siliceous spicules in some soils in the flooded forests
of the Amazon (19%) indicates that they, along with
diatoms, are likely to be significant in at least some
water logged environments.
7. Application to understanding and study of
regolith processes
7.1. Setting of exceptionally opal-rich environments
Although small values of opal are fairly ubiquitous
in many soils and sediments significant quantities
(e.g., >5%) occur less commonly. Apart from the
unusual stranded Eocene sediments of southern Aus-
tralia, examples are non-marine. Examples reviewed in
this paper include the Magela Creek flood plain,
(Northern Territory, Australia; Clark et al., 1992a,b),
Amazonia (Chauvel et al., 1996), Bungendore (NSW)
and Duntroon (ACT; both Brewer, 1955) (Fig. 3). The
Okavango Delta has abundant dissolution and precip-
itation of silica (Shaw et al., 1990; Shaw and Nash,
1998), and high levels of opal productivity. Peat
formed from grasses and sedges contain up to 30%
phytoliths, along with minor diatoms and sponge
spicules (McCarthy et al., 1989).
These opal-depositing environments differ in many
respects. With respect to vegetation, Amazonia con-
sists of flooded tropical rainforest, Bungendore and
Duntroon formerly temperate eucalypt woodland,
Magela Creek monsoon grassland, woodland, and
wetland, and the Okavango Delta seasonally to per-
manently flooded arid wetland. With respect to water
chemistry, Magela Creek and Amazonia waters are
acidic, the Okavango region strongly alkaline, and the
NSW and ACT examples near neutral. The Okavango
waters are also comparatively saline whereas the
others are dilute.
The common feature in all of these environments is
seasonal to permanent flooding. Under such condi-
tions the diatoms and sponge bloom in sufficient
quantities so as to make a significant component of
the sediment. The sponges and diatoms may be
epiphytic (Chauvel et al., 1996; Konhauser et al.,
1992; Clark et al., 1992b), or planktonic (Volkmer-
Ribeiro, 1992). Furthermore, in many wetlands the
plant taxa include those with high phytolith produc-
tion, such as grasses and sedges (Clark et al.,
1992a,b). This association of poor drainage and high
levels of opal reinforces the conclusions drawn above
from a review of soil micromorphology in Stace et al.
(1968) discussed above.
Under ideal conditions, these environments can
produce sediments as rich in biogenic silica as those
of the marine realm. Magela Creek forms part of the
East Alligator River drainage system in the Northern
Territory of Australia. The black alluvial soils, typi-
cally between 30 cm and 2 m thick, can average up to
35% biogenic opal over lateral distances of several km
and up to 48% in a single soil core. The mineral
fraction of individual beds within the black soils can
be composed entirely of biogenic opal (Clark et al.,
1992a,b).
Other unusually opal-rich environments are found
on volcanic soils in Africa, Japan (see references in
Hart, 1992) and Reunion (Meunier et al., 1999). The
high levels of opal in these soils may reflect unusually
rapid release and uptake of silica into plants as a result
of the fast weathering of soils derived from the
weathering of volcanic debris and rocks, especially
those low in silica.
Although high rainfall encourages water logging
and thus potentially high productivity of opal, it is not
necessary for it. Water logged soils and aquatic envi-
ronments occur in arid and semi arid environments,
provided there is sufficient water. More critical is
salinity, but provided it is not excessive, these environ-
ments are potentially highly productive, even in dry
J. Clarke / Earth-Science Reviews 60 (2003) 175–194186

environments. Examples discussed in this paper
include Lake Chad and the Okavango Delta.
7.2. Dust
Most dust contains a small proportion of opal, this
being almost entirely of biogenic origin. Australian
windblown dusts (Baker, 1960) contain phytoliths,
sponge spicules, and diatoms. Drees et al. (1993)
reported that the main type of biogenic opal in dusts
from Niger is sponge spicules. These dusts have been
deflated from the Sahara or Sahel regions of Africa,
not the most obvious habitat for freshwater sponges.
Jones and Beavers (1963) and Wilding and Drees
(1968) used the presence of sponge spicules in soils
from ridge tops to indicate contributions from wind-
blown materials. Given the ubiquity of spicules and
spicule-like structures in many soils, this criterion
may prove to be of doubtful value. However, recog-
nising windblown components to soils is important to
the regolith geology of many areas (Greene, in press),
such as eastern Australia, where dryland soil salinity
is believed to be the result of accession of aeolian
material and potential tools in its recognition should
not be ignored. In addition, Wilding and Drees
(1974) suggested the clay-sized quartz particles in
some soil profiles that have been attributed to aeolian
accession may, in fact, be due to the recrystallisation
of phytoliths.
7.3. Palaeoenvironmental reconstruction
Opal-forming organisms are highly sensitive to
environmental variations and, properly interpreted,
Fig. 3. An opal factory (2): soils of the Canberra region contain significant amounts of opal. (A) Podzolic soils in gently undulating landscapes
north of Duntroon, Canberra, Australia contain up to 22% biogenic opal. (B) Alluvial soils at Barrack Flat, Queanbeyan, contain up to 7% opal
(Angela Harrison, unpublished data). (C) Soils at Deep Creek, east of Bungendore, ACT, Australia reportedly contain 11% biogenic opal (D).
Some 2% biogenic opal has been reported from Doughboy Creek, east of Bungendore.
J. Clarke / Earth-Science Reviews 60 (2003) 175–194 187

can be a useful guide in environmental reconstruction.
Both phytoliths and diatoms have been widely used in
palaeoenvironmental interpretation. Sponge spicules
have been used less because of difficulties in taxo-
nomic identification, but are good indicators of per-
manently waterlogged conditions and, where taxa are
recognisable, of marine influence.
Beavers and Stephen (1958) showed that phytoliths
in the soils of Illinois varied according to vegetation
type. Such patterns can be used to reconstruct ancient
vegetation patterns. One such example is the report of
Barboni et al. (1999), which provides an example of
phytoliths in the environmental reconstruction of
archaeologically significant regions in Ethiopia.
Because phytoliths are commonly perceived to be
concentrated in the A horizon of soils (Oehler,
1979), increases in their abundance within a profile
have sometimes been used as indicators of palaeosols
in a succession (Dormaar and Lutwick, 1969). As
noted above, care should be taken in such interpreta-
tions as exceptions are known. Piperno (1988) gave
an extensive review of the use of phytoliths in palae-
oenvironmental reconstruction and archaeology, with
the collection edited by Meunier and Colin (2001)
providing a summary of the state of the art.
The use of diatoms to reconstruct lacustrine sedi-
mentary environments is well known (Smol, 1987).
However, diatoms can be used even in arid and semi-
arid regions, as illustrated by the study of Gasse
(1987) from sub-Saharan Africa. In this example,
the diatoms live in a wide range of environments,
including former mega-Lake Chad, dilute swamps,
and small hypersaline lakes, fed by groundwater
discharge or ephemeral streams.
Harrison (1988) reviews the use of sponge spicules
from a Canadian lake. Sponge gemmules are also
valuable in reconstructing environments and have
been found in diverse environments such as bogs of
the Puget Lowland, Washington (Turner, 1985) and
the flood plain forests of Amazonia (Junk, 1984).
Piperno (1988) provides one of the few examples
of using a wide range of siliceous organisms in
environmental reconstruction. She was able to extract
a diverse assemblage of phytoliths, diatoms, sponge
spicules, and helizoans from a series of terrestrial
sediment cores from Panama. Piperno was able to
identify moist tropical forest, marine swamp, fresh-
water swamp and cropland vegetation and therefore
reconstruct an environmental and floral history of the
last 11,300 years. The record closely matched that
obtained from palynology and indicates the potential
utility of siliceous remains to document palaeoenvir-
onments in sediments that might not preserve organic
microfossils. Similarly, Clark et al. (1992a,b) used
palynology, diatom taxonomy, and distribution of
phytoliths and sponge spicules to determine the Hol-
ocene evolution of the Magela Creek floodplain in the
Northern Territory of Australia. These authors were
able to demonstrate how the environment evolved
from a mangrove swamp to seasonal freshwater wet-
land. Opal production was a clear indication of wet-
land development. However, very high levels of opal
productivity (>8% for bulk soil composites) were
associated with grass and sedge wetlands, rather than
wooded wetlands or mangroves. Sponges were the
main source of opal in the mangrove environment.
Despite these studies, the use of biogenic, opal,
apart from phytoliths, as a tool in regolith geology is
in its infancy. Most studies using siliceous remains
have concentrated on the Quaternary, rather than the
much longer time necessary in many regolith studies
as illustrated by Ollier and Pain (1996). Some work
has been done on siliceous remains in older sedi-
ments, such as that of Folk (1964) on Cainozoic
phytoliths. Marine sponge spicules have been impor-
tant in understanding Eocene environments of south-
ern Australia (Clarke, 1994a,b; Gammon et al., 2000).
Phytolith geochemistry is another possible but
poorly explored avenue for palaeoclimatic research.
Webb and Longstaffe (1997) indicated that increasing
enrichment in O18 reflected increasing degrees of
evapotranspiration. More needs to be known about
the range of O18 values of different taxa before this
can be routinely applied.
7.4. Silcretes siliceous hardpans, and hardsetting soils
Silcretes are silica-rich duricrusts found in many
parts of the globe (Thiry, 1999). They are enigmatic
features in that, unlike other duricrusts such as cal-
cretes, bauxites, and ferricretes, there are few known
modern analogues. Silcrete appears to form by a range
of different processes in a diverse range of landscape
contexts and eras (Thiry, 1999; Alley, 1996; Firman,
1994; Ollier, 1991), matched only by the diversity of
opinions as to its formative conditions. An important
J. Clarke / Earth-Science Reviews 60 (2003) 175–194188

series of observations, however is that it commonly
appears to have formed on former valley slopes and
floors, often, although not always, in quartz-rich
colluvium or alluvium. Importantly, to distinguish
silcretes from siliceous hardpans (see below), there
is also silicification of the host material, as well as
silica cementation. Once formed, such silcretes typi-
cally form inverted relief (Ollier and Pain, 1996).
Such silcretes need not have extended across the
entire landscape, but rather have formed in selected
localities by the lateral migration of silica-rich
groundwater.
The greater lability of biogenic opal compared to
other forms of silica means that it is a prime source of
silica for precipitation in the regolith. Several authors
have suggested a connection between silcretes and
biogenic opal. Gunn and Galloway (1978) raised the
possibility that biogenic silica leached from marine
organisms was an important component in some
Australian silcretes developed on Cretaceous marine
shales. Oehler (1979) similarly suggested that bio-
genic opal may play a role in silcrete formation,
postulating that dissolution of the siliceous remains
of terrestrial organisms may have contributed to the
formation of silcretes. Thiry (1999) noted silica dep-
osition by diatoms in the regolith during pluvial
highstands of Lake Chad (see also Gasse, 1987).
Few examples of recognisable biogenic silica remains
have been noted from silcretes, however. One exam-
ple is that of Clarke (1994b), which noted that almost
all the silcretes present in the Kambalda and Norse-
man regions of WA were developed on Eocene
spicular marine sediments, such as the Princess Royal
Spongolite and Pallinup Siltstone. The formation of
silcrete has largely obliterated the spicules and they
are preserved only as ghosts. These marine spicules
are quite large, typically 1–5 mm in length and 100
Am in diameter. It is likely that the smaller freshwater
spicules (100–300 Am in length and 3–10 Am across)
originally present in a silcreted sediment would be
completely destroyed or rendered unrecognisable, as
would the even more delicate diatoms and phytoliths.
Therefore, the remains of siliceous microfossils may
have been originally much more common in what are
now silcreted surficial sediments. A possible addi-
tional link between biogenic opal and silcretes is the
high levels of titanium found in many silcretes (Thiry,
1999) and the observation than phytoliths are com-
monly also enriched in this element (Wilding et al.,
1989). Much more work must be done to elucidate
this relationship, as detrital Ti from insoluble residules
or adsorbed Ti on clays and iron oxides are alter-
natives to a biological origin.
Clearly, the accumulation of biogenic silica in the
regolith alone is not the key to silcrete formation. As
the literature reviewed in this paper has shown, such
accumulations are not uncommon. Modern analogies
for silcrete are, however, difficult to find. What
remains unanswered is what conditions facilitate the
large-scale cementation and replacement by silica in
the regolith. If biogenic opal indeed plays a role in
this, it is through the stablisation of that opal as quartz.
It is possible however that accumulation of biogenic
silica, followed by major shifts in physical and chem-
ical hydrology associated with the Cainozoic climate
changes, may have been a factor.
Siliceous hardpans (Wright, 1983) are also com-
mon in many parts of inland Australia and also from
the Paris Basin (Thiry, 1999). They consist of silica-
cemented alluvium and colluvium. Other cementing
agents, including iron oxides, carbonate, and clay are
also present (Bettenay and Churchwood, 1974). Typ-
ically, they contain less silica and more iron and
carbonate than silcretes. Hardpans may form a con-
tinuum with silcretes, or represent an early stage in
silcrete development. Unlike silcretes, they are often
less indurated, lacking in silicification, and appear
more likely related to contemporary or near-contem-
porary landscape processes (Milnes et al., 1991) in
arid and semi-arid environments. Also, hardpans form
through siliceous cementation, rather than whole-scale
silicification. Despite this, their genesis and relation-
ship to silcrete is still obscure. Nor can they always be
related to overlying soil profiles (Wright, 1983).
However, the presence of common mobile silica from
biogenic opal would clearly be a favourable precursor
to their formation. As for silcretes, the fact that
biogenic opal is more common that hardpans suggests
that the key question is not the presence of opal in the
soil, but what factors encourage its stablisation in the
regolith as a quartz cement.
Hardsetting soils (Greene, in press) may be an early
precursor to hardpans. Hardsetting occurs through
cyclic wetting and drying of the soils. Slaking of soils
aggregates under conditions of rapid wetting, and/or
dispersion of the clay fraction alone. However, in some
J. Clarke / Earth-Science Reviews 60 (2003) 175–194 189

cases, cementation of the subsoil can also occur as a
result release of soluble silica during wetting and
reprecipitation as the soil dries (Chartres et al.,
1990). The presence of potentially labile silica in the
form of biogenic opal may facilitate this process.
Another possible link is the relationship sometimes
noted between some silcretes and volcanic rocks,
especially basalts (see Ollier, 1991). In the past this
was normally attributed to silica released during
hydrothermal alteration or rapid weathering. While a
hydrothermal origin is ruled out by modern workers
(Taylor and Eggleton, 2001), the exceptionally high
levels of biogenic opal found in soils developed on
some volcanic rocks (Meunier et al., 1999, and
references in Hart, 1992) suggest that it is the organ-
isms that are responsible for this association.
7.5. Health
The size, geometry and composition of biogenic
silica bodies in the regolith is a potential health issue.
The large marine spicules of the Eocene of WA can
cause contact dermatitis among geologists logging
percussion and air core holes in the palaeovalley fills
of the area. The irritation caused by the spicules is
increased by the hypersaline waters which often satu-
rates these deposits. The numbers of open pits through
the Eocene cover is also increasing, although no
effects among mine waters are as yet reported. Sponge
spicules in soils of southwestern Western Australia,
probably also Eocene in age, have been linked to hoof
disease in horses (Carroll, 1932). Stone et al. (1970)
mentioned similar contact dermatitis amongst agricul-
tural workers in areas of freshwater spicule-rich soil of
New York, as did miners of freshwater spiculites in
Brazil (Volkmer-Ribeiro, 1992).
Although respirable dust (must with a median
aerodynamic diameter of 10 Am and less) is normally
regarded as an urban problem, it is also a major health
issue in rural areas (Clausnitzer and Singer, 1996). The
lower size limit of freshwater sponge spicules ( < 5 Amacross and some less than 3 Am) places them within the
definition of hazardous mineral fibre (Skinner et al.,
1988). Some of the cellular damage caused by fibrous
materials is from fibre penetration. Many sponge
spicules have sharper terminations than non-biogenic
mineral fibres, which greatly increases their penetra-
tion potential. Other cellular damage is caused by
mineral reactivity, however the biological reactivity
of opal is unknown. The presence of spicules in
windblown dust raises the possibility that they may
present a respirable silica hazard in dust-affected areas.
Silicified root hair cells (Piperno, 1988) also have a
high aspect ratio, are sharp, and potentially fall into the
hazardous size range. Most are probably too large to
pose a cancer risk, although respiratory irritation has
been reportedly caused by them (Parry et al., 1984).
8. Conclusions
Biogenic opal in soils was first recognised in the
1840s. The potential importance of silica-secreting
plants in the formation of silcretes was suggested in
the 1950s. Despite periodic reviews in the 1970s and
1980s, and considerable research of aspects of bio-
genic silica in soils and terrestrial sediments since
then, regolith science is not much closer to under-
standing the larger scale significance of biogenic opal
than we were 50 years ago.
What is known is that juvenile biogenic opal is
present in most soils in at least trace amounts and
common (up to 2–3% in the A horizons many soils on
all continents except Antarctica. Phytoliths, sponges,
and diatoms largely produce the opal. Furthermore, in
a significant number of cases, it can be present in large
amounts comprising more than 10% of the mineral
fraction, and in some examples 100%. There is a
strong correlation between poorly drained or at least
seasonally water logged conditions, and abundant
biogenic opal. In some cases, volcanic substrates are
also conducive to abundant soil opal. Biogenic opal
may play an important role in the cycling of silica in
soils and aquatic sediments, in the genesis of the
siliceous cements of hardpans, and silcretes, and may
be significant in environmental health.
A number of approaches for future research appear
promising. The first is taxonomy; what opal-secreting
organisms are present in the regolith, in what numbers,
and how can they be recognised from their remains.
The second is the ecological limits and roles repre-
sented by each organism, which requires both drainage
basin and catena studies. Thirdly, we need to know the
stratigraphic distribution of these remains especially
through the Cainozoic. These studies will enable a
better understanding of the both the regolith signifi-
J. Clarke / Earth-Science Reviews 60 (2003) 175–194190

cance of biogenic opal in a wide range of environments
and also constrain the conditions under which they
may become the dominant part. Fourthly, we need to
identify the possible geochemical signatures of bio-
genic opal in mature regolith profiles. Fifthly, research
is needed into the physical, chemical, and biological
conditions necessary to convert labile opal into stable
quartz, leading to the formation of silica cements,
hardpans, and silcretes. Sixthly, the role of opal in
nutrient cycling needs better documentation. Finally,
the potential health hazards of biogenic opal in respi-
rable dusts would also be a fruitful research area.
Study of the importance of opal producing organ-
isms in the regolith is clearly a multidisciplinary
process. After more than 160 years, the importance
of biogenic opal in the regolith is surely an idea whose
time has come.
Acknowledgements
I would like to thank Doreen Bowdery, Robin
Clark, Tony Eggleton, John Field, Dianne Hart, Megan
Kilby, Ian Roach, and Bob Wasson for their help and
suggestions in preparing this paper. Graham Taylor
and J. Meunier made several helpful and encouraging
comments in their reviews of the manuscript. The
paper is published with the permission of CRC LEME.
References
Alexandre, A., Meunier, J.D., Colin, F., Koud, J.M., 1997. Plant
impact on the biogeochemical cycle of silicon and related pro-
cesses. Geochim. Cosmochim. Acta 61 (3), 677–682.
Alley, N.F., 1996. Cainozoic stratigraphy, palaeoenvironments, and
geological evolution of the Lake Eyre Basin. Palaeogeogr. Pa-
laeoclimatol. Palaeoecol. 144, 239–263.
Aston, S.R., 1983. Natural water and atmospheric chemistry of
silicon. In: Aston, S.R. (Ed.), Silica Geochemistry and Bio-
chemistry. Academic Press, New York, pp. 39–76.
Baker, G., 1959a. Opal phytoliths in some Victorian soils and ‘red
rain’ residues. Aust. J. Bot. 7, 64–87.
Baker, G., 1959b. A contrast in the opal phytolith assemblies of two
Victorian soils. Aust. J. Bot. 7, 88–96.
Baker, G., 1960. Phytoliths in some Australian dusts. Proc. R. Soc.
Vic. 72, 21–40.
Barboni, D., Bonnefille, R., Alexandre, A., Meunier, J.D., 1999.
Phytoliths as paleoenvironmental indicators, west side middle
Awash valley, Ethiopia. Palaeogeogr. Palaeoclimatol. Palaeoe-
col. 152, 87–100.
Bartoli, F., 1985. Crystallography and surface properties of biogenic
opal. J. Soil Sci. 36, 335–350.
Beavers, A.H., Stephen, I., 1958. Some features of the distribution
of plant opal in Illinois soils. Soil Sci. 86, 1–5.
Bettenay, E., Churchwood, H.M., 1974. Morphology and strati-
graphic relationships of the Wiluna hardpan in arid Western
Australia. J. Geol. Soc. Aust. 21 (1), 73–80.
Bobrov, A.A., Bobrova, E.K., 2001. Phytolith assemblages in pod-
zol and podzolic soils of the south-west coast of the Ohotsk Sea.
In: Meunier, J.D., Colin, F. (Eds.), Phytoliths; Applications in
Earth Sciences and Human History. A.A. Balkema, Rotterdam,
pp. 365–370.
Bobrova, E., Bobrov, A., 1997. Phytoliths in soils: species compo-
sition, distribution along a soil profile, and value as environmen-
tal indicators. In: Pinilla, A., Juan-Tresserras, J., Machado, M.A.
(Eds.), First European Meeting on Phytolith Research. Centro de
Ciencias Medioambientales del Consejo Superior de Investiga-
ciones Cientificas Serrano, 115B. 2800, Madrid, pp. 5–13.
Boettinger, J.L., 1994. Biogenic opal as an indicator of mixing in an
alfisol/vertsol landscape. Proceedings of the 9th International
Working Meeting on Soil Micromorphology, 17–26.
Bohaty, S.M., Harwood, D.M., 2000. Ebridian and silicoflagellate
bisotratigraphy from Eocene McMurdo erratics and the South-
ern Ocean. In: Stilwell, J.D., Feldmann, R.M. (Eds.), Paleobi-
ology and Paleoenvironments of Eocene rocks, McMurdo
Sound, East Antarctica. Antarct. Res. Ser., vol. 76. American
Geophysical Union, Washington, pp. 99–160.
Bombin, M., Muehlenbachs, K., 1980. Potential of 18O/16O ratios
in opaline plant silica as a continental paleoclimatic tool. Amer-
ican Quaternary Association Abstracts and Program, Sixth Bi-
ennial Meeting at Orono, Maine, 43–44.
Braiser, M.D., 1980. Microfossils George Allen and Sons, London.
Braiser, M.D., Green, O., Sheilds, G., 1997. Ediacarian sponge
spicule clusters from southwestern Mongolia and the origins
of the Cambrian fauna. Geology 25 (4), 351–354.
Breese, G.F., 1960. Quartz overgrowths as evidence of silica depo-
sition. Aust. J. Sci. 23, 18–20.
Brewer, R., 1955. Diatom skeletons and sponge spicules in the soils
of New South Wales. Aust. J. Sci. 17, 177–179.
Brewer, R., Sleeman, J.R., Foster, R.C., 1993. The fabric of Aus-
tralian soils. Soils: An Australian Viewpoint. Division of Soils,
CSIRO, pp. 439–476.
Carlson, E.H., 1994. Paleoshoreline patterns in the transgressive–
regressive sequences of Pennsylvanian rocks in the northern
Appalachian Basin, USA. Sediment. Geol. 93, 209–222.
Carroll, D., 1932. Mineralogy of fine sand fractions of some Aus-
tralian soils. J. R. Soc. West. Aust. 18, 125–144.
Charman, D.J., 2000. Biostratigraphic and palaeoenvironmental ap-
plications of testate amobae. Quat. Sci. Rev. 20, 1753–1764.
Chartres, C.J., Kirby, J.M., Raupach, M., 1990. Poorly ordered
silica and aluminosilicates as temporary cementing agents in
setting soils. Soil Sci. Am. J. 54, 1060–1067.
Chauvel, A., Walker, I., Lucas, Y., 1996. Sedimentation and pedo-
genesis in a central Amazonian black water basin. Biogeochem-
istry 33, 77–95.
Clark, R.L., East, T.J., Guppy, J., Mahon, D., Mcbride, P., Wasson,
R.J., 1992a. Late Quaternary stratigraphy of the Magela Plain.
J. Clarke / Earth-Science Reviews 60 (2003) 175–194 191

In: Wasson, R.J. (Ed.), Modern Sedimentation and Evolution of
the Magela Creek Plain. Res. Rep. Supervising Scientist for the
Alligator Rivers Region, vol. 6. pp. 28–80.
Clark, R.L., East, T.J., Guppy, J., Mahon, D., Mcbride, P., Wasson,
R.J., 1992b. Late Quaternary evolution of the Magela Plain. In:
Wasson, R.J. (Ed.), Modern Sedimentation and Evolution of the
Magela Creek Plain. Res. Rep. Supervising Scientist for the
Alligator Rivers Region, vol. 6. AGPS, Canberra, pp. 81–157.
Clarke, J.D.A., 1994a. Evolution of the Lefroy and Cowan Palae-
odrainages, Western Australia. Aust. J. Earth Sci. 41, 55–68.
Clarke, J.D.A., 1994b. Geomorphology of the Kambalda region.
Aust. J. Earth Sci. 41 (3), 229–240.
Clausnitzer, H., Singer, M.J., 1996. Respirable-dust production
from agricultural operations in the Sacramento Valley, Califor-
nia. J. Environ. Qual. 25, 877–884.
Davis, A., Suzanne, J.R., Rimmer, S.M., Yeakel, J.D., 1984. Some
genetic implications of silica and aluminosilicates in peat and
coal. Int. J. Coal Geol. 3, 293–314.
De Laubenfels, M.W., 1955. Porifera. Treatise on Invertebrate Pale-
ontology Part E. Geological Society of America, pp. E21–E112.
Dormaar, J.F., Lutwick, L.E., 1969. Infrared spectra of humic acids
and opal phytoliths as indicators of paleosols. Can. J. Sci. 49,
29–37.
Drees, L.R., Manu, A., Wilding, L.P., 1993. Characteristics of aeo-
lian dusts in Niger, West Africa. Geoderma 59, 213–223.
Epstein, E., 1999. Silicon. Annu. Rev. Plant Physiol. Plant Mol.
Biol. 50, 641–664.
Ferris, F.G., 1997. Formation of authigenic minerals by bacteria.
In: Mcintosh, J.M., Groat, L.A. (Eds.), Biological–Mineralog-
ical Interactions. Mineral. Assoc. Can. Short Course, vol. 25,
pp. 187–208.
Firman, J.B., 1994. Paleosols in laterite and silcrete profiles. Evi-
dence from the south east margin of the Australian Precambrian
shield. Earth Sci. Rev. 36, 149–179.
Fredlund, G.G., 1993. Paleoenvironmental interpretation of stable
carbon, hydrogen, and oxygen isotopes from opal phytoliths,
Eustis Ash Pit, Nebraska. In: Pearsall, D.M., Piperno, D.R.
(Eds.), Current Research in Phytolith Analysis: Applications
in Archaeology and Paleoecology. MASCA Res. Pap. Sci. Ar-
chaeol., vol. 10, pp. 37–46.
Folk, R.L., 1964. Note on the occurrence of phytoliths in some
Cenozoic sedimentary rocks. J. Paleontol. 38, 773–775.
Gammon, P.R., James, N.P., Bone, Y., Clarke, J.D.A., 2000. Sed-
imentology and lithostratigraphy of a Late Eocene sponge-do-
minated sequence, southern Western Australia. Aust. J. Earth
Sci. 47 (6), 1087–1103.
Gasse, F., 1987. Diatoms for reconstructing palaeoenvironments
and palaeohydrology in tropical semi-arid zones. Hydrobiologia
154, 127–163.
Gibson, A.A., 1976. Diatomite—Western Australia. In: Knight, C.L.
(Ed.), Economic Geology of Australia and Papua new Guinea.
Australas. Inst. Min. Metall. Monogr., vol. 8, pp. 111–112.
Greene, R.S.B., 2002. Hardsetting soils. In: Lal, R. (Ed.), Encyclo-
pedia of Soil Science. Marcel Dekker, NewYork, USA (in press).
Gregory, W., 1855. On the presence of diatomacaeae, phytolitharia,
and sponge spicules in soils which support vegetation. Proc.
Bot. Soc., Edinb., 69–72.
Gunn, R.H., Galloway, R.W., 1978. Silcretes in south-central
Queensland. In: Langford-Smith, T. (Ed.), Silcrete in Australia.
Department of Geography, University of New England, Armi-
dale, pp. 51–71.
Harrison, F.W., 1988. Methods in Quaternary ecology #4. Fresh-
water sponges. Geosci. Can. 15 (1), 193–198.
Hart, D.M., 1992. A field appraisal of the role of plant opal in the
Australian environment. PhD thesis, Department of Earth Sci-
ences, Macquarie University, Australia (unpublished).
Hart, D.M., Humphreys, G.S., 1997. The mobility of phytoliths in
soils: pedological considerations. In: Pinilla, A., Juan-Tresser-
ras, J., Machado, M.A. (Eds.), First European Meeting on Phy-
tolith Research. Centro de Ciencias Medioambientales del Con
sejo Superior de Investigaciones Cientificas Serrano, 115B,
2800, Madrid, pp. 93–100.
Heinen, W., Oehler, J.H., 1979. Evolutionary aspects of biological
involvement in the cycling of silica. In: Trudinger, P.A., Swain,
D.J. (Eds.), The Biogeochemical Cycling of Mineral-Forming
Elements. Elsevier, Amsterdam, pp. 431–443.
Jones, R.L., Beavers, A.H., 1963. Sponge spicules in Illinois soils.
Proc. Soil Sci. Soc. Am. 27, 438–440.
Jones, L.H.P., Handreck, K.A., 1967. Silica in soils, plants, and
animals. Adv. Agron. 19, 107–149.
Jones, L.P.H., Milne, A.A., Saunders, J.V., 1966. Tabashir: an opal
of plant origin. Science 151, 464–466.
Junk, W.J., 1984. Ecology of the varzea, floodplain of Amazonian
whitewater rivers. In: Sioli, H. (Ed.), The Amazon. Monogr.
Biol., vol. 56. Kluwer, Dordrecht, pp. 214–244.
Kenley, P.R., 1976. Diatomite—Victoria. In: Knight, C.L. (Ed.),
Economic Geology of Australia and Papua New Guinea. Aus-
tralas. Inst. Min. Metall. Monogr., vol. 8, pp. 111–112.
Kondo, R., Iwasa, Y., 1981. Opal phytoliths in humic yellow later-
osols and yellow laterosols in the Amazon region. Res. Bull.
Obihiro Univ. 12, 231–239.
Konhauser, K.O., Mann, H., Fyfe, W.S., 1992. Prolific organic SiO2
precipitation in a solute deficient river, Rio Negro, Brazil. Geol-
ogy 20, 227–230.
Kosters, E.C., Bailey, A., 1983. Characteristics of peat deposits in
the Mississippi River Delta Plain. Trans. Gulf Coast Assoc.
Geol. Soc. 33, 311–325.
Lauwers, A.M., Heinen, W., 1974. Biodegradation and utilization
of silica and quartz. Arch. Microbiol. 95, 67–78.
Lewin, J.C., Reimann, B.E.F., 1969. Silicon and plant growth. An-
nu. Rev. Plant Physiol. 20, 289–304.
Lovering, T.S., 1959. Significance of accumulator plants in rock
weathering. Bull. Geol. Soc. Am. 70, 781–800.
McCarthy, T.S., McIver, J.R., Caincross, B., Ellery, W.N., Ellery,
K., 1989. The inorganic chemistry of peat from the Maunachira
channel swamp system, Okavango Delta, Botswana. Geochim.
Cosmochim. Acta 53, 1077–1089.
McIntosh, J.M., Groat, L.A. (Eds.), 1997. Biological–Mineralogi-
cal Interactions. Mineral. Assoc. Can. Short Course, vol. 25.
Meunier, J.D., Colin, F. (Eds.), 2001. Phytoliths; Applications in
Earth Sciences and Human History. A.A. Balkema, Dordrecht,
378 pp.
Meunier, J.D., Colin, F., Alarcon, C., 1999. Biogenic silica storage
in soils. Geology 27, 835–838.
J. Clarke / Earth-Science Reviews 60 (2003) 175–194192

Milnes, A.R., Wright, M.J., Thiry, M., 1991. Silica accumulations
in saprolites and soils of South Australia. In: Nettleton, W.D.
(Ed.), Occurrence, Characteristics, and Genesis of Carbonate,
Gypsum and Silica Accumulations in Soils. Soil Sci. Soc.
Am. Spec. Publ., vol. 26, pp. 121–149.
Moore, R.C., 1964. Protista 3: Protozoa (chiefly Radiolaria and
Tintinnina). Treatise on Invertebrate Paleontology Part D. Geo-
logical Society of America, Lawrence, KS.
Muller, P.J., Schneider, R., 1993. An automated method for the
determination of opal in sediments and articulate matter.
Deep-Sea Res., Part I 40, 425–446.
Nott, J., 1994. Long-term landscape evolution in the Darwin
region and its implications for the origin of landsurfaces in
the north of the Northern Territory. Aust. J. Earth Sci. 41,
407–415.
Oehler, J.H., 1979. Deposition and diagenesis of biogenic silica. In:
Trudinger, P.A., Swain, D.J. (Eds.), The Biogeochemical Cy-
cling of Mineral-Forming Elements. Elsevier, Amsterdam, pp.
467–479.
Ollier, C.D., 1991. Aspects of silcrete formation in Australia. Z.
Geomorphol. 35 (2), 151–163.
Ollier, C., Pain, C., 1996. Regolith, Soils, and Landforms Wiley,
Chichester.
Parry, D.W., Hodson, M.J., Sangster, A.G., 1984. Some recent
advances in the study of silicon in higher plants. Philos. Trans.
R. Soc. London, Ser. B 304, 537–549.
Patrick, R., 1977. Ecology of freshwater diatoms–diatom commun-
ities. In: Wener, D. (Ed.), The Biology of Diatoms. Bot.
Monogr., vol. 13. Blackwell, London, pp. 284–332.
Piperno, D.R., 1988. Phytolith Analysis: An Archaeological and
Geological Perspective. Academic Press, San Diego.
Runge, F., 1999. The opal phytolith inventory of soils in central
Africa—quantities, shapes, classification, and spectra. Rev. Pa-
laeobot. Palynol. 107, 23–53.
Schwandes, I.P., Collins, M.E., 1994. Distribution and significance
of freshwater sponge spicules in selected Florida soils. Trans.
Am. Microsc. Soc. 113 (3), 242–257.
Shaw, P.A., Nash, D.J., 1998. Dual mechanisms for the formation of
fluvial silcretes in the distal reaches of the Okavango delta fan,
Botswana. Earth Surf. Processes Landf. 23, 705–714.
Shaw, P.A., Cooke, H.J., Perry, C.C., 1990. Microbialitic silcretes in
highly alkaline environments: some observations from Sua Pan,
Botswana. S. Afr. J. Geol. 93 (5/6), 803–808.
Silverman, M.P., 1979. Biological and organic chemical decompo-
sition of silicates. In: Trudinger, P.A., Swain, D.J. (Eds.), The
Biogeochemical Cycling of Mineral-Forming Elements. Elsev-
ier, Amsterdam, pp. 445–465.
Simons, N.A., Hart, D.M., Humphreys, G.S., 2000. Phytolith depth
functions in a podzol and a texture contrast soil, Sydney. In:
Adams, J.A., Metherell, A.K. (Eds.), Soil 2000: New Horizons
for a New Century. Aust. and NZ Second Joint Soils Conference,
vol. 3. New Zealand Society of Soil Science, Canterbury, pp.
193–194.
Skinner, H.C.W., Fitzpatrick, R.W. (Eds.), 1992. Biomineralization:
processes of iron and manganese—modern and ancient environ-
ments. Catena. (Supplement 21).
Skinner, H.C.W., Ross, M., Frondel, C., 1988. Asbestos and Other
Fibrous Materials. Oxford Univ. Press, New York.
Smithson, F., 1959. Opal sponge spicules in soils. J. Soil Sci. 10 (1),
105–109.
Smol, J.P., 1987. Methods in freshwater ecology #1. Freshwater
algae. Geosci. Can. 14 (4), 206–215.
Stace, H.C.T., Hubble, G.D., Brewer, R., Northcoate, K.H., Slee-
man, J.R., Mucahy, M.J., Hallsworth, E.G., 1968. A Handbook
of Australian Soils. Rellim Technical Publications, Glenside,
South Australia.
Stone, E.L., Dawson, J.E., Zeeuw, C.D., 1970. Dermatitis caused by
sponge spicules. Soil Sci. Soc. Am. Proc. 34, 151–152.
Taylor, G.T., Eggleton, R.A., 2001. Regolith Geology and Geo-
morphology. Wiley, Chichester.
Taylor, G., Gasse, F., Walker, P.H., Morgan, P.J., 1990. The palaeo-
ecological and palaeoclimatic significance of Miocene fresh-
water diatomite deposits from southern New South Wales,
Australia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 77 (2),
127–143.
Tesic, Z.P., Todorovic, M.S., 1958. Contribution to knowledge of
the specific properties of silicate bacteria. Zemljiste, 233–240.
Thiry, M., 1999. Diversity of continental silicification features:
examples from the Cenozoic deposits of the Paris Basin and
neighbouring basement. Spec. Publ. Int. Assoc. Sedimentol. 27,
87–127.
Turner, J., 1985. Sponge gemmules from lake sediments in the
Puget Lowland, Washington. Quat. Res. 24, 240–243.
Volkmer-Ribeiro, C., 1992. The freshwater sponges in some peat-
bog ponds in Brazil. Amazonia 12 (2), 317–335.
Webb, E.A., Longstaffe, F.J., 1997. Oxygen isotope geochemistry
of silica phytoliths. In: Pinilla, A., Juan-Tresserras, J., Machado,
M.A. (Eds.), First European Meeting on Phytolith Research.
Centro de Ciencias Medioambientales del Consejo Superior de
Investigaciones Cientificas Serrano, 115B. 2800, Madrid, pp.
233–243.
Wilding, L.P., Drees, L.R., 1968. Distribution and implications of
sponge spicules in surficial deposits in Ohio. Ohio J. Sci. 68,
92–99.
Wilding, L.P., Drees, L.R., 1971. Biogenic opal in Ohio soils. Soil
Sci. Soc. Am. Proc. 35, 1004–1010.
Wilding, L.P., Drees, L.R., 1974. Contributions of forest opal and
associated crystalline phases to fine silt and clay fractions of
soils. Clay Clay Miner. 22, 295–306.
Wilding, L.P., Smeck, N.E., Drees, L.R., 1989. Silica in soils:
quartz, crystabolite, tridymite and disordered silica polymorphs.
In: Dixon, J.B., Weed, S.B. (Eds.), Minerals in Soil Environ-
ments, 2nd edn. Soil Sci. Soc. Am. Book, vol. 1. pp. 913–974,
Madison, WI.
Wollast, R., Mackenzie, F.T., 1983. The global cycle of silica. In:
Aston, S.R. (Ed.), Silica Geochemistry and Biochemistry. Aca-
demic Press, New York, pp. 39–76.
Wright, M.J., 1983. Red-brown hardpans and associated soils
in Australia. Trans. R. Soc. South Aust. 107, 252–254.
J. Clarke / Earth-Science Reviews 60 (2003) 175–194 193

Jonathan Clarke graduated with a BSc
(Hons.) from the University of Tasmania
and completed his PhD degree at Flinders
University in South Australia. He worked
for 10 years at WMC Resources Ltd. in
mineral exploration and research before
joining the geology department at the
Centre for Landscape, Environment, and
Mineral Exploration at the Australian
National University. His past research inter-
ests have included Cambrian reef, platform,
and slope carbonates, cool water carbonate deposition, coal geology,
modern sediments on the Australian shelf, evolution of the Great
Australian Bight, and regolith geology in Western Australia. Other
current research interests include the history of aridity of the
Atacama Desert, Eocene carbonate, clastic, and biosiliceous sed-
imentation in the Eucla Basin, and terrestrial Mars analogues.
J. Clarke / Earth-Science Reviews 60 (2003) 175–194194