computer three-dimensional anatomical reconstruction of the human sinus node and a novel paranodal...
TRANSCRIPT

THE ANATOMICAL RECORD 294:970–979 (2011)
Computer Three-DimensionalAnatomical Reconstruction of theHuman Sinus Node and a Novel
Paranodal AreaNATALIE CHANDLER,1 OLEG ASLANIDI,2 DAVID BUCKLEY,3 SHIN INADA,1
STEVEN BIRCHALL,1 ANDREW ATKINSON,1 DANIELLE KIRK,1
OLIVER MONFREDI,1 PETER MOLENAAR,4 ROBERT ANDERSON,1
VINOD SHARMA,5 DANIEL SIGG,5 HENGGUI ZHANG,2 MARK BOYETT,1
AND HALINA DOBRZYNSKI1*1Cardiovascular Medicine, University of Manchester, Manchester, UK
2Biological Physics, University of Manchester, Manchester, UK3Division of Medical Physics, University of Leeds, Leeds, UK
4Queensland University of Technology and University of Queensland, Brisbane, Australia5Medtronic Inc, Minneapolis, MN, USA
ABSTRACTWe have previously shown in rabbit that the pacemaker of the heart
(the sinus node) is widespread and matches the wide distribution of theleading pacemaker site within the right atrium. There is, however, uncer-tainty about the precise location of the pacemaker in human heart, andits spatial relationships with the surrounding right atrial muscle. There-fore, the aim of the current study was to investigate the distribution ofthe sinus node tissue in a series of healthy human hearts and, for one ofthe hearts to construct a computer three-dimensional anatomical model ofthe sinus node, including the likely orientation of myocytes. A combina-tion of experimental techniques—diffusion tensor magnetic resonanceimaging (DT-MRI), histology, immunohistochemistry, image processingand computer modelling—was used. Our data show that the sinus nodewas larger than in previous studies and, most importantly, we identifieda previously unknown area running alongside the sinus node (the ‘‘para-nodal area"), which is even more extensive than the sinus node. This areapossesses properties of both nodal and atrial tissues and may have a rolein pacemaking. For example, it could explain the wide spread distributionof the leading pacemaker site in human right atrium, a phenomenonknown as the wandering pacemaker observed in clinics. In summary, a
Additional Supporting Information may be found in the onlineversion of this article.Abbreviations used: AM ¼ Atrial muscle; ANP ¼ Atrial natri-
uretic peptide; Ao ¼ Aorta; Cx40, Cx43 ¼ Connexin 40 and 43;3D ¼ Three-dimensional; DT-MRI ¼ Diffusion tensor magneticresonance imaging; HCN4 ¼ Ion channel responsible for hyper-polarization-activated or ‘funny’ current, If; IVC ¼ Inferior venacava; Kir2.1 ¼ Ion channel responsible for background inwardrectifier Kþ current, IK1; Kir6.2 ¼ Ion channel responsible forATP-sensitive Kþ current, IKATP; Kv1.4 ¼ Ion channel responsi-ble for transient outward Kþ current, Ito; LAA ¼ Left atrialappendage; MiRP2 ¼ Accessory protein for Kþ channels; Nav1.5, Nav�1 ¼ Naþ channel responsible for inward Naþ cur-rent, INa; PA ¼ Pulmonary artery; PV ¼ Pulmonary vein; RA ¼Right atrium; RAA ¼ Right atrial appendage; RV ¼ Right ven-tricle; SCV ¼ Superior caval vein; SK2 ¼ Ion channel responsi-
ble for Ca2þ-activated Kþ current, IKCa; TASK1 Twin-poredomain Kþ channel; Tbx3 ¼ Transcription factor; TC ¼ Termi-nal crest.
Natalie Chandler and Oleg Aslanidi equally contributed tothis work.Grant sponsor: Medtronic Inc.
*Correspondence to: Dr. Halina Dobrzynski, Faculty ofMedical and Human Sciences, University of Manchester, CoreTechnology Facility, 46 Grafton Street, Manchester M13 9NT,UK. E-mail: [email protected]
Received 19 June 2010; Accepted 27 January 2011
DOI 10.1002/ar.21379Published online 28 April 2011 in Wiley Online Library(wileyonlinelibrary.com).
VVC 2011 WILEY-LISS, INC.
novel 3D anatomical reconstruction presents a new picture of the distri-bution of nodal cells within the human right atrium. Anat Rec, 294:970–979, 2011. VVC 2011 Wiley-Liss, Inc.
Keywords: sinus node; pacemaking; 3D anatomical model;arrhythmias
INTRODUCTION
The sinus node is the primary pacemaker of the heartand is responsible for the initiation of the heartbeat. Thehuman sinus node has previously been shown as a smallhighly localized and histologically distinct tissue area atthe junction of the superior caval vein with the rightatrium. The leading pacemaker site can, however, movespontaneously along the terminal crest of the right atrium,a phenomenon known as the wandering pacemaker. Inopen heart surgery, patients and in healthy individuals,the leading pacemaker site has been shown to be locatedanywhere along the terminal crest between the openings ofthe superior and inferior caval veins (Boineau et al., 1988;
Ramanathan et al., 2004; Ramanathan et al., 2006). Suchfindings are in agreement with studies on smaller mam-mals. For example, to stop sinus rhythm in the dog, anextensive lesion is required along the whole of the terminalcrest between the superior and inferior caval veins (Kal-man et al., 1995). A detailed computer three-dimensional(3D) model of the anatomy of the sinus node of the rabbit,based on serial sections processed using histology andimmunohistochemistry, shows the rabbit sinus node toextend from the superior to the inferior caval vein (Dobr-zynski et al., 2005), a finding consistent with electrophysio-logical data from the rabbit (Fedorov et al., 2006). It ispossible, therefore, that the human sinus node is a more
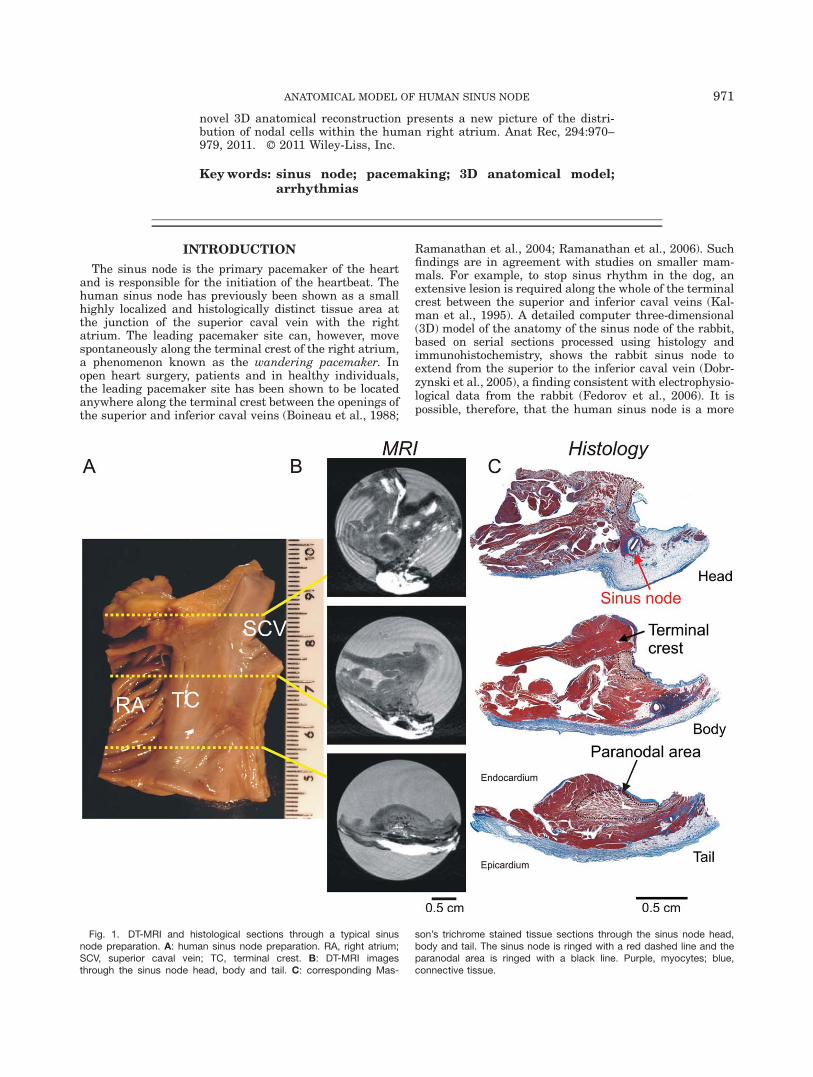
Fig. 1. DT-MRI and histological sections through a typical sinusnode preparation. A: human sinus node preparation. RA, right atrium;SCV, superior caval vein; TC, terminal crest. B: DT-MRI imagesthrough the sinus node head, body and tail. C: corresponding Mas-
son’s trichrome stained tissue sections through the sinus node head,body and tail. The sinus node is ringed with a red dashed line and theparanodal area is ringed with a black line. Purple, myocytes; blue,connective tissue.
ANATOMICAL MODEL OF HUMAN SINUS NODE 971

extensive structure than has been previously reported. Ina previous study of ion channel expression in the humansinus node (Chandler et al., 2009), we reported that run-ning alongside the sinus node is a discrete tissue with char-acteristics transitional between those of the sinus node andatrial muscle. Since this tissue had not been previouslyreported, we named it the ‘paranodal area’. In this study,we assess the structure and extent of the human sinusnode and the novel paranodal area. The 3D model of thehuman SAN, for the first time, (i) reveals the full extent ofthe SAN, which is much larger than shown before; (ii)shows spatial locations and shapes of the SAN and the newparanodal area within the human right atrium; (iii) showsorientations of cardiac fibres running through the SAN,paranodal area and atrial tissue. Spatial structure of theSAN and fibre orientations are the major factors definingthe patterns of AP generation in the SAN and spread intothe atrium. The DT-MRI results allowed, primarily, foreffective reconstruction of the fibre orientations, which wasimpossible to achieve using other methods.
MATERIALS AND METHODS
Four preparations of the superior cavoatrial junctionfrom healthy human hearts (but unsuitable for trans-plantation) were obtained from the Prince Charles Hos-
pital, Australia. Patient details are given in SupportingInformation Table S1. The work was ethically approvedby the Prince Charles Hospital Ethics Committee (ethicsapproval, PCH EC2565) and the University of Manches-ter. One of the preparations was fixed in formalin over-night and was used to construct the computer model. Itwas scanned using DT-MRI to determine the anatomy ofthe preparation and myocyte orientation. The prepara-tion was then embedded in paraffin and serially sec-tioned. 58 sections at 0.5 mm intervals were stainedwith Masson’s trichrome to show the histology and a fur-ther 58 sections at the same levels were labeled usingthe immunoenzyme technique for connexin43 (Cx43) tohelp delineate the sinus node, paranodal area and atrialmuscle. The sections at the 58 levels were aligned basedon the DT-MRI. Next, based on the histology and Cx43labeling, the sections were ‘‘segmented,’’ that is, the dif-ferent tissues delineated. Amira 4.1 software (VisageImaging) was then used to construct a 3D anatomicalmodel as a mathematical array and the ‘‘isosurface"function was used to view it. Finally, the myocyte orien-tation computed from the DT-MRI was incorporated intothe model. The model is available as two text files inSupporting Information. The remaining three humansinus node preparations were used to confirm the mainfindings from the preparation used to construct the 3Danatomical model. The preparations were frozen in liq-uid N2 and cryosectioned; sections at different levelswere stained with Masson’s trichrome and variousmarker proteins were labeled using immunofluorescence.The 3D anatomical model was combined with mathemat-ical models of feasible human sinus node, paranodal andatrial action potentials (Chandler et al., 2009) – numeri-cal simulation was used to calculate the initiation andspread of the action potential through the tissue. Fulldetails of the methods used are available in SupportingInformation.
RESULTSDetermination of Tissue Geometry andIdentification of Different Cell Types
Figure 1A shows an endocardial view of the superiorcavoatrial junction preparation from a healthy humanheart used to construct the 3D anatomical model. Thepectinate muscles in the right atrial appendage, the ter-minal crest (a thick branch of atrial muscle) and thesmooth intercaval region can be seen (Fig. 1A). Thepreparation was scanned using DT-MRI, and Fig. 1Bshows examples of DT-MRI images taken from a planeperpendicular to the terminal crest as indicated by theyellow broken lines in Fig. 1A. Tissue sections were thencut from the preparation along the same plane. Fifty-eight sections were selected at 0.5 mm intervals andstained for histology using Masson’s trichrome, whichstains myocytes purple and connective tissue blue. TheMasson’s trichrome stained sections most closely corre-sponding to the DT-MRI images (Fig. 1B) are shown inFig. 1C. The images in Fig. 1B,C are selected to showthe head, body, and tail of the human sinus node.Another Masson’s trichrome stained section (at highermagnification) is shown in Fig. 2A. In the Masson’s tri-chrome stained sections, the sinus node could usually beeasily identified, because it was centred on the sinusnode artery and it appeared blue because of a high
Fig. 2. Masson’s trichrome staining (A) and immunoenzyme labelingof Cx43 (B) in adjacent sections through the sinus node, paranodalarea and atrial muscle of the terminal crest. In A: purple, myocytes;blue, connective tissue. In B: Cx43 labeling shown as small darkbrown spots. The sinus node is ringed with a red dashed line and theparanodal area is ringed with a green dashed line.
972 CHANDLER ET AL.

content of fibrous connective tissue. The atrial musclecould also be identified, because it was densely-packedand red in appearance. In addition, there was a thirdtissue, the paranodal area. In the Masson’s trichromestained sections, it could be identified because the myo-cytes were loosely packed. This was confirmed with highmagnification images of the Masson’s trichrome stainingin Fig. 3A. The arrangement of loosely packed cells dis-tinguished paranodal myocytes. Furthermore, theextracellular matrix in which the paranodal myocyteswere embedded was different in composition to theextracellular matrix of the sinus node, because itappeared white and not blue, that is, it was composed ofadipocytes rather than fibrous tissue. Another 58 sec-
tions at the same levels were labeled using immunoen-zyme for Cx43 to aid the identification of the threetissues. Cx43 is the principle gap junction protein in theheart. A representative example is shown in Fig. 2B.High magnification images of Cx43 labeling are shownin Fig. 3B. As we have shown previously (Chandleret al., 2009), Cx43 was highly expressed in the atrialmuscle, but was not detectable in the sinus node andwas expressed at an intermediate level in the paranodalarea. Although histology alone was sufficient to identifyand distinguish the sinus node, paranodal and atrialmuscle, the use of, and lack of Cx43 labeling was impor-tant in identifying the head and tail of the sinus nodetissue. HCN4 is commonly used as a positive marker of
Fig. 3. High magnification images of Masson’s trichrome staining(A), immunoenzyme labeling of Cx43 (B) and immunofluorescencelabeling of Cx43 (C) and Cx40 and caveolin3 (D) in the sinus node,paranodal area and atrial muscle. In A: purple, myocytes; blue, con-nective tissue. In B: Cx43 labeling shown as small dark brown spots;red arrows highlight Cx43-expressing myocytes and green arrows
highlight non-expressing myocytes. In C: Cx43 labeling shown assmall green spots; white arrows highlight Cx43-expressing myocytesand red arrows highlight non-expressing myocytes. In D: Cx40 label-ing shown as red spots and caveolin3 labeling shown in green; whitearrows highlight Cx40 expressing myocytes and red arrows highlightnon-expressing myocytes.
ANATOMICAL MODEL OF HUMAN SINUS NODE 973

the cardiac conduction system in small mammals (e.g.,mouse, rat, and rabbit) (Brioschi et al., 2010; Yanniet al., 2010). Unfortunately, HCN4 could not be used asa positive marker of the human sinus node due to a highbackground signal in the surrounding atrial muscle(data not shown). This made the anatomical boundary ofthe human sinus node more difficult to delineate. Cx43was, therefore, used a negative marker of the humansinus node.
The main findings from the sinus node preparationused to construct the anatomical model were confirmedin three other sinus node preparations. These prepara-tions were frozen and cryosections were cut. Cx43 wasagain labeled (using immunofluorescence rather thanimmunoenzyme). Four other marker proteins were alsoimmunolabeled, connexin40 (Cx40), atrial natriureticpeptide (ANP), caveolin3 and vimentin. Cx40 is a largeconductance connexin, ANP (a hormone) is onlyexpressed by atrial myocytes, caveolin3 is expressed inthe cell membrane of all cardiac myocytes, and vimentinis expressed by fibroblasts (one component of the extrac-ellular matrix). As expected, Cx43 labeling was absentfrom the sinus node, but present in the atrial muscle(Fig. 3C). In the paranodal area, Cx43 labeling was het-
erogeneous. The paranodal area contained a mixture ofareas some of which expressed Cx43 (like atrial muscle)and others that did not (like sinus node) (Fig. 3C). TheCx43 signal intensity was measured and found to be thelowest in the sinus node, the highest in the atrial mus-cle, and intermediate in the paranodal area (Fig. 4A).This suggests that approximately half of the paranodalarea expressed Cx43 (Fig. 4A). Double labeling for Cx40and caveolin3 revealed that Cx40 was distributed in asimilar manner to Cx43 (Fig. 3D). ANP was again dis-tributed in a similar manner to Cx43. It was absent inthe sinus node, present in some regions in the paranodalarea and present in the atrial muscle (Fig. 5A). This wasconfirmed by the ANP signal intensity (Fig. 4B). How-ever, double labeling of Cx43 and ANP revealed a morecomplex picture of the paranodal area. Not all regions inthe paranodal area that expressed Cx43 also expressedANP, and vice versa (Supporting Information Fig. S1).Caveolin3 labeling was present in all myocytes (Fig. 5B),and there was no change in the mean signal intensity ofcaveolin3 immunofluorescence in the sinus node, parano-dal, and right atrial myocyte regions (Fig. 4C). Immunolabeling of vimentin showed a much greater proportionof fibroblasts in the sinus node than in the atrial muscleand paranodal area (Figs. 4D, 5C). This supports the ob-servation from the Masson’s trichrome staining that theextracellular matrix was fibrous in the sinus node andfatty in the paranodal area. Because caveolin3 labelingis present in the cell membrane (Fig. 5B), it can be usedto measure the diameter of transversely cut myocytes(with a circular profile): the myocyte diameter in theparanodal area was intermediate between that in thesinus node and atrial muscle (Fig. 4E). In conclusion,these findings confirm the presence of three distinctmyocyte types in the tissue sections.
3D Anatomical Model of the Human Sinus Nodeand Paranodal Area
The 58 sections at �0.5 mm intervals from the prepa-ration used to construct the anatomical model were ‘‘seg-mented,’’ based on differences in the histology and theimmunoenzyme labeling of Cx43 (e.g., Fig. 2). The sec-tions were also outlined and aligned based on the DT-MRI (e.g., Fig. 1B). In Supporting Information, examplesof sections segmented in this way are shown in Fig. S2,and all sections are shown in Movie 1. Amira 4.1 soft-ware was used to construct a 3D anatomical model ofthe preparation from the 58 segmented sections. Figure6 shows endocardial, side, and epicardial views of themodel with the different tissues shown in different col-ours (see also Movie 2 Supporting Information). Themain features of the model were confirmed in the threeother sinus node preparations. The sinus node (red zone)was located beneath the epicardium of the terminal crestwith a layer of atrial muscle in between the sinus nodeand the endocardium (Fig. 6B). The sinus node was cres-cent shaped and entwined around the sinus node arteryand had a wide body and thin head and tail, which werelocated more toward the endocardial surface than thebody (Fig. 6). The sinus node measured 29.5 mm inlength, 1.8 mm in height, and 6.4 mm in width, which islarger than reported previously. For example, the dimen-sions of the human sinus node have previously beenmeasured using electron microscopy and reported to be
Fig. 4. Properties of the sinus node, paranodal area and atrial mus-cle. A–D: signal intensity for the expression of Cx43 (A), ANP (B), cav-eolin3 (C) and vimentin (D) as detected by immunofluorescence.Signal intensity was measured using Velocity software (Improvision)and is shown relative to the signal intensity in the atrial muscle. Means� SEM (n ¼ 3) shown. E: diameter of sinus node, paranodal and atrialmyocytes. Means � SEM (n ¼ 200) shown. *ysignificantly different(P < 0.05) from sinus node* or paranodal areay as determined byone-way ANOVA.
974 CHANDLER ET AL.

13.5 (�2.5) mm in length, 1.2 (�0.3) mm in height, and5.6 (�1.4) mm in width (Sanchez-Quintana et al., 2005).This demonstrates the importance of using immunohis-tochemistry as well as histology to determine the limitsof the sinus node, which is narrow at the head and tail.At the resolution studied, there was no apparent connec-tion between the sinus node and the paranodal area,with an atrial overlay present between the two struc-tures (e.g., Fig. 2A). The paranodal area (yellow zone)was much more extensive than the sinus node andappeared to continue beyond the border of the tissuesample towards the inferior caval vein (Fig. 6).
Myocyte Orientation
DT-MRI was used to measure the speed (magnitude)and direction (angle) of diffusion of water moleculesthrough the tissue shown in Fig. 1A. In each voxel(250 lm � 250 lm � 500 lm; typical for such work)(Helm et al., 2005), we measured the primary eigenvec-tor (the angle of the preferred pathway of water diffu-sion) and the primary eigenvalue (the magnitude ofwater diffusion along the primary eigenvector) as wellas the secondary and tertiary eigenvectors and eigenval-ues (the angle and magnitude of water diffusion in thetwo orthogonal planes to the primary eigenvector). Asexpected, in all three tissues (sinus node, paranodal areaand atrial muscle), the secondary eigenvalue (the magni-tude of water diffusion along the secondary eigenvector)was significantly less than the primary eigenvalue (P <0.0001, paired t-test; Fig. 7A). The mean value of thesecondary eigenvalue was 49–73% of the mean value of
the primary eigenvalue. In ventricular muscle, the meanvalue of the secondary eigenvalue has been reported tobe 55% of the mean value of the primary eigenvalue(Helm et al., 2005). The angle of the primary eigenvectorhas been shown to correspond to the orientation (i.e.,angle) of myocytes in cardiac tissues, presumablybecause the intracellular compartment of the coupledmyocytes acts as a major pathway for water diffusion.The extracellular space is expected to act as anotherpathway for water diffusion, but perhaps water diffusionthrough the extracellular space is again constrained bythe myocytes. The angle of the primary eigenvector willcorrespond to the average angle of myocytes in a voxel.In the sinus node, the primary eigenvalue was less thanin the other tissues (Fig. 7A), perhaps because the sinusnode is made up of a low density (Fig. 3A) of poorly-coupled myocytes, as indicated by the absence of connex-ins (Fig. 3B–D). Figure 7B shows histograms of theangles (as viewed from the endocardial surface of thepreparation) in the three tissues. Although there was awide range of angles in atrial muscle, there was also apreferred angle of �70� (with respect to the horizontalplane of the figure). This suggests that many (but notall) atrial myocytes ran in the superior-inferior direction.This is again shown in Fig. 7C, which shows the angleof the primary eigenvector in each voxel in the sinusnode, paranodal area and atrial muscle (as viewed fromthe endocardial surface of the preparation). The anglesare shown by arrows (colour coded according to theangle). In the atrial muscle, there are many of the yel-low arrows, which are near vertical with an angle of�70�. This is consistent with images of tissue sectionscut perpendicular to the terminal crest in which manyatrial myocytes are cut transversely (e.g., in Fig. 7D thecut myocytes have a circular rather than an ellipsoidprofile). Careful inspection of the data shows that theatrial myocytes (as judged by the primary eigenvectors)ran longitudinally along the terminal crest and pectinatemuscles. This is best shown in Supporting InformationFig. S3. The myocytes in the paranodal area (as judgedby the primary eigenvectors) were orientated in a simi-lar manner to those in the terminal crest (Fig. 7B–D). Inthe sinus node, there was a wide range of angles, butthere was also a preferred angle of �50� (Fig. 7B). Thisis shown again in Fig. 7C, in which there are manygreen and blue arrows with an angle of �50�. Therefore,many sinus node myocytes were at an angle of �20�with respect to those in the paranodal area and atrialmuscle. This is consistent with images of tissue sectionscut perpendicular to the terminal crest in which manysinus node myocytes are cut transversely or obliquely(e.g., in Fig. 7D, the cut myocytes have a circular orshort elliptical profile rather than a elongated ellipticalprofile). An analysis of the angle of the primary eigen-vectors as viewed from the side of the preparation is pre-sented in Supporting Information Fig. S4. The angles ofthe primary eigenvectors are incorporated into the 3Danatomical model and are available in the SupportingInformation.
Use of the 3D Anatomical Model
Figure 8A shows the leading pacemaker site (red area)and the activation sequence of the atria of a patient(from Boineau et al., 1988). In this patient, the leading
Fig. 5. Expression of ANP (red signal; A), caveolin3 (red signal; B)and vimentin (blue signal; C) as detected by immunofluorescence inthe sinus node, paranodal area and atrial muscle. ANP is present in in-tracellular vesicles, caveolin3 is present in the cell membrane andvimentin is present in fibroblasts. Scale bar for all images shown atthe bottom right. Blue arrows highlight ANPexpressing myocytes;green arrows highlight non-expressing myocytes.
ANATOMICAL MODEL OF HUMAN SINUS NODE 975

pacemaker site was at the junction of the superior cavalvein with the right atrium. For comparison, Fig. 8Bshows the model of the sinus node and paranodal areasuperimposed on a schematic diagram of the heart.Because the model is a mathematical array with accu-rate anatomy, segmentation of different tissue types andmyocyte orientation, it can be used in mathematical sim-ulations. Based on ion channel expression, we have for-mulated a mathematical model of the action potential inthe human sinus node (Chandler et al., 2009). We haveused this model of the human sinus node action poten-tial together with a model of the human atrial potential(Courtemanche et al., 1988). We assumed that the para-nodal area is made of 50% sinus node myocytes and 50%atrial myocytes (Fig. 3C) to compute the initiation andconduction of the action potential in the 3D anatomicalmodel of the human sinus node and paranodal area. Thecomputed ‘heart rate’ was �1.1 Hz or 67 beats/minute.As shown in Fig. 8B, the action potential was first initi-ated at a discrete site at the head of the sinus node andit was then conducted to the remainder of the tissue viathe paranodal area. The activation sequence in Fig. 8Bis similar to that in Fig. 8A.
DISCUSSION
This study presents the first computer 3D reconstruc-tion of the human sinus node and novel paranodal area.Anatomical models of the human sinus node have beenpresented previously. James (1961) described the humansinus node as a small nodule at the junction of the supe-rior caval vein with the right atrium and ‘Purkinjetracts’ leading away from it, which is now known not tobe the case. Truex et al. (1967) produced a similar pic-
ture of the human sinus node (but without ‘Purkinjetracts’) and they created a 3D model of the sinus node inwax. Since the 1960s, more accurate representations ofthe human sinus node have been produced. For example,Sanchez-Quintana et al. (2005) showed that the humansinus node is perhaps larger than earlier reported andhas a crescent shape, centred on the sinus node arteryand it extends down the terminal crest. In theserespects, the description of Sanchez-Quintana et al.(2005) is consistent with the present study. However,Sanchez-Quintana et al. (2005) identified the sinus nodebased on histological criteria only and in our experienceit is only possible to identify the extremities of thehuman sinus node based on immuno labeling of markerproteins as well as histology. It may be significant thatin the present study the sinus node was longer (29.5mm) than in the study of Quintana et al. (2005) (13.5 �2.5 mm). Our model is also different from the earliermodels in that it includes the novel paranodal area andmyocyte orientation.
Mismatch Between Known Structure andFunction of Sinus Node
It has been reported by others that there is a mis-match between the known structure of the sinus nodeand pacemaking function (Kalman et al., 1995; Schuess-ler et al., 1996). Typically, the human sinus node isdepicted as a small nodule at the junction of the superiorcaval vein with the right atrium and the action potentialcan be initiated at any site along the terminal crest fromthe superior to the inferior caval vein (Boineau et al.,1988). The present study shows that the sinus nodeextends from the superior caval vein along the terminal
Fig. 6. 3D anatomical model of the human sinus node. Endocardial, side and epicardial views shown.Different tissues shown in different colours: red, sinus node; yellow, paranodal area; green, atrial muscle;blue, adipose tissue. PM, pectinate muscles; TC, terminal crest.
976 CHANDLER ET AL.

crest further towards the inferior caval vein than hasbeen previously recognized. However, as shown in Fig.8B the human sinus node only extends about a third ofthe distance between the superior and inferior caval
veins and, therefore, there is still a mismatch betweenstructure and function. A possible explanation for themismatch is the paranodal area. The sinus node prepa-rations available to us (Fig. 1A) did not extend to the
Fig. 7. Myocyte orientation. A: histogram of the primary (red) andsecondary (blue) eigenvalues in the sinus node, paranodal area andatrial muscle. B: histogram of fibre angles (angles of the primaryeigenvector) in the three tissues. C: myocyte orientation in the three
tissues superimposed on endocardial views of the preparation. Orien-tation is indicated by the direction as well as the colour of the arrows.D: high magnification views of caveolin3 immuno labeling in the threetissues in sections cut perpendicular to the terminal crest.
ANATOMICAL MODEL OF HUMAN SINUS NODE 977

inferior caval vein and the paranodal area was large atthe inferior edge of the preparation. It is possible, there-fore, that the paranodal area extends to the inferiorcaval vein, but this needs to be clarified in future stud-ies. In addition, it is well known that the terminal crestcan be the focus of atrial tachycardias, the so-called‘‘cristal tachycardias’’ (Kalman et al., 1998). Approxi-mately two-thirds of focal right atrial tachycardiasoccurring in the absence of structural heart disease arisealong the terminal crest, with a gradation in frequencyfrom the superior to the inferior terminal crest (Kalmanet al., 1998). Once again, the only structure with a pro-pensity for pacemaking extending the length of the ter-minal crest is the paranodal area and, therefore, this isa candidate substrate for the cristal tachycardias.Whether the paranodal area is capable of pacemakingunder physiological or pathophysiological conditions isconsidered below.
Properties and Possible Function of theParanodal Area
This study has shown that the paranodal area is ananatomically distinct area of the myocardium. The para-nodal area can be recognized in previous histologicalstudies of the human sinus node, albeit not identified assuch by previous investigators (e.g., Sanchez-Quintanaet al., 2005). The paranodal area runs in parallel is inclose proximity to the sinus node, but does not appear toconnect directly with it as determined at the resolutionobtained used in the current study (Fig. 2A). The para-nodal tissue is more extensive than the sinus node andcontinues beyond the inferior margin of our preparationand therefore a further study is needed to investigatethe full extent of this tissue. In the paranodal area, the
myocytes are diffusely organized in an extracellular ma-trix that is different from that of the sinus node (Fig.3A). The extracellular matrix of the paranodal area con-tains few fibroblasts, as indicated by vimentin expres-sion unlike in the sinus node (Fig. 4D) and instead itcontains adipocytes (Fig. 3A). On average, the myocytesin the paranodal area are intermediate in diameter com-pared to sinus node and atrial myocytes (Fig. 5E). Theparanodal area is heterogeneous in that it contains bothatrial-like (expressing Cx40, Cx43 and ANP) and sinusnode-like (not expressing Cx40, Cx43 and ANP) areas(Figs. 3C, D and 5A). In our earlier study on ion channelexpression in the human sinus node, we observed thatthe expression pattern of some ion channels in the para-nodal area is intermediate between that of the sinusnode and atrial muscle. For example, expression ofKir2.1, Nav1.5 and Navb1 is high in the atrial muscleand low in the sinus node (Chandler et al., 2009). Thisexplains why the atrial muscle has a stable restingpotential, whereas the sinus node does not, and theatrial muscle has an action potential with a fastupstroke, whereas the sinus node has an action potentialwith a slow upstroke. In the paranodal area, expressionof Kir2.1, Nav1.5 and Navb1 is intermediate (Chandleret al., 2009). Tbx3 is a transcription factor responsiblefor the nodal phenotype of the sinus node. Ectopicexpression of Tbx3 in atrial muscle represses the atrialphenotype and imposes a nodal phenotype (Hoogaarset al., 2007). In the human, as expected, Tbx3 is highlyexpressed in the sinus node, but poorly expressed in theatrial muscle (Chandler et al., 2009). Tbx3 expression inthe paranodal area is significantly greater than in theatrial muscle but is less than in the sinus node (Chan-dler et al., 2009). However, the paranodal area is notalways transitional as it shows greater expression of
Fig. 8. Comparaison of the activation sequence of the sinus nodein a patient and in the 3D anatomical model. A: dorsal view of the atriaof a patient during open heart surgery showing the leading pacemakersite (red) and the activation sequence of the atria; the activationsequence is shown as a colour scale and by the isochrones in ms.From Boineau et al., 1988. B: model of the sinus node (red) and para-nodal area (yellow) superimposed on a schematic diagram of the heart
(dorsal view). Ao, aorta; IVC, inferior caval vein; PA, pulmonary artery;PV, pulmonary veins; RA, right atrium; RV, right ventricle; SVC, supe-rior caval vein. The activation sequence is shown by the isochrones inms; the leading pacemaker site is shown by the white spot. Simulatedaction potentials from the sinus node (SN), paranodal area (PA) andatrial muscle (AM) are shown.
978 CHANDLER ET AL.

Kv4.2, Kir6.1, TASK1, SK2 and MiRP2 compared to boththe sinus node and atrial muscle (Chandler et al., 2009).
What is the possible function of the paranodal area?The poor expression of Cx40 and Cx43 in the paranodalarea (Fig. 3B-D) means that the paranodal myocytes arepoorly coupled. The poor expression of Nav1.5 and Navb1(Chandler et al., 2009) means that the myocytes willhave an action potential with a slow upstroke; and, mostimportantly, the poor expression of Kir2.1 (Chandleret al., 2009) may mean that the myocytes lack a stableresting potential. All of these are important attributes ofnodal tissue but not atrial muscle. The lack of Kir2.1, inparticular, may mean that the myocytes in the parano-dal area are capable of pacemaking under physiologicalor pathophysiological conditions. However, the paranodalarea does not show discernible expression of HCN4, theprincipal ion channel responsible for the pacemaker cur-rent, If. The paranodal area may have another function.Theoretically, it is difficult for the sinus node to drivethe more hyperpolarized atrial muscle that is for theaction potential to exit from the sinus node into theatrial muscle (Joyner and van Capelle, 1986). In thehuman, at the junction between the two tissues, anatrial overlay is sandwiched between the sinus node andparanodal area (Fig. 2A). Because of the lack of Kir2.1,the paranodal area may be depolarized in comparisonwith atrial muscle. The paranodal area may depolarizethe atrial overlay and in this way facilitate the exit ofthe action potential from the sinus node into the atrialmuscle. Further work is required to determine functionsof the paranodal area, as well as ANS innervations.
In summary, we have produced a detailed 3D anatomi-cal model (including myocyte orientation) of the humansinus node and running in parallel with the sinus nodeis an extensive paranodal area. This area possesses prop-erties of both nodal and atrial tissues and may have a rolein pacemaking and could explain the wide spread distri-bution of the leading pacemaker site (wandering pace-maker) in the human right atrium as observed in clinics.
LITERATURE CITED
Boineau JP, Canavan TE, Schuessler RB, Cain ME, Corr PB, CoxJL. 1988. Demonstration of a widely distributed atrial pacemakercomplex in the human heart. Circulation 77:1221–1237.
Brioschi C, Micheloni S, Tellez JO, Pisoni G, Longhi R, Moroni P,Billeter R, Barbuti A, Dobrzynski H, Boyett MR, DiFrancesco D,Baruscotti M. 2010. Distribution of the pacemaker HCN4 channelmRNA and protein in the rabbit sinoatrial node. J Mol Cell Car-diol 47:221–227.
Chandler NJ, Greener ID, Tellez JO, Inada S, Musa H, Molenaar P,DiFrancesco D, Baruscotti M, Longhi R, Anderson RH, Billeter R,Sharma V, Sigg DC, Boyett MR, Dobrzynski H. 2009. Moleculararchitecture of the human sinus node: insights into the functionof the cardiac pacemaker. Circulation 119:1562–1575.
Courtemanche M, Ramirez RJ, Nattel S. 1998. Ionic mechanismsunderlying human atrial action potential properties: insightsfrom a mathematical model. Am J Physiol 275:H301–H321.
Dobrzynski H, Li J, Tellez J, Greener ID, Nikolski VP, Wright SE,Parson SH, Jones SA, Lancaster MK, Yamamoto M, Honjo H,Takagishi Y, Kodama I, Efimov IR, Billeter R, Boyett MR. 2005.Computer three-dimensional reconstruction of the sinoatrial node.Circulation 111:846–854.
Fedorov VV, Hucker WJ, Dobrzynski H, Rosenshtraukh LV, EfimovIR. 2006. Postganglionic nerve stimulation induces temporal inhi-bition of excitability in the rabbit sinoatrial node. Am J Physiol291:H612–H623.
Helm PA, Tseng HJ, Younes L, McVeigh ER, Winslow RL. 2005. Exvivo 3D diffusion tensor imaging and quantification of cardiaclaminar structure. Magn Reson Med 54:850–859.
Hoogaars WM, Engel A, Brons JF, Verkerk AO, de Lange FJ, WongLY, Bakker ML, Clout DE, Wakker V, Barnett P, Ravesloot JH,Moorman AF, Verheijck EE, Christoffels VM. 2007. Tbx3 controlsthe sinoatrial node gene program and imposes pacemaker func-tion on the atria. Genes Dev 21:1098–1112.
James TN. 1961. Anatomy of the human sinus node. Anatom Rec141:109–139.
Joyner RW, van Capelle FJL. 1986. Propagation through electricallycoupled cells: how a small SA node drives a large atrium. BiophysJ 50:1157–1164.
Kalman JM, Lee RJ, Fisher WG, Chin MC, Ursell P, Stillson CA,Lesh MD, Scheinman MM. 1995. Radiofrequency catheter modifi-cation of sinus pacemaker function guided by intracardiac echo-cardiography. Circulation 92:3070–3081.
Kalman JM, Olgin JE, Karch MR, Hamdan M, Lee RJ, Lesh MD.1998. Cristal tachycardias: origin of right atrial tachycardias fromthe crista terminalis identified by intracardiac echocardiography.J Am Coll Cardiol 31:451–459.
Ramanathan C, Ghanem RN, Jia P, Ryu K, Rudy Y. 2004. Noninva-sive electrocardiographic imaging for cardiac electrophysiologyand arrhythmia. Nat Med 10:422–428.
Ramanathan C, Jia P, Ghanem R, Ryu K, Rudy Y. 2006. Activa-tion and repolarization of the normal human heart under com-plete physiological conditions. Proc Natl Acad Sci USA 103:6309–6314.
Sanchez-Quintana D, Cabrera JA, Farre J, Climent V, AndersonRH, Ho SY. 2005. Sinus node revisited in the era of electroana-tomical mapping and catheter ablation. Heart 91:189–194.
Schuessler RB, Boineau JP, Bromberg BI. 1996. Origin of the sinusimpulse. J Cardiovasc Electrophysiol 7:263–274.
Truex RC, Smythe MQ, Taylor MJ. 1967. Reconstruction of thehuman sinoatrial node. Anatom Rec 159:371–378.
Yanni J, Boyett MR, Anderson RH, Dobrzynski H. 2010. The extentof the specialized atrioventricular ring tissues. Heart Rhythm6:672–680.
ANATOMICAL MODEL OF HUMAN SINUS NODE 979