composition and temporal changes in macroinvertebrate communities of intermittent streams in...
TRANSCRIPT

This article was downloaded by: [Moraine Comm College]On: 10 December 2014, At: 00:44Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
New Zealand Journal of Marine andFreshwater ResearchPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnzm20
Composition and temporal changesin macroinvertebrate communities ofintermittent streams in Hawke's Bay,New ZealandRichard G. Storey a & John M. Quinn ba National Institute of Water and Atmospheric Research Limited , P.O. Box 11 115, Hamilton, New Zealand E-mail:b National Institute of Water and Atmospheric Research Limited ,P. O. Box 11 115, Hamilton, New ZealandPublished online: 19 Feb 2010.
To cite this article: Richard G. Storey & John M. Quinn (2008) Composition and temporal changes inmacroinvertebrate communities of intermittent streams in Hawke's Bay, New Zealand, New ZealandJournal of Marine and Freshwater Research, 42:1, 109-125, DOI: 10.1080/00288330809509941
To link to this article: http://dx.doi.org/10.1080/00288330809509941
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &

Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

New Zealand Journal of Marine and Freshwater Research, 2008, Vol. 42: 109-1250028-8330/08/4201-0109 © The Royal Society of New Zealand 2008
.109
Composition and temporal changes in macroinvertebratecommunities of intermittent streams in Hawke's Bay, New Zealand
RICHARD G. STOREYJOHN M. QUINN
National Institute of Water and AtmosphericResearch Limited
P. O. Box 11 115Hamilton, New Zealandemail: [email protected]
Abstract Macroinvertebrate communities in threestony-bottomed intermittent headwater streams incentral Hawke's Bay, New Zealand, were sampledonce per month (on average) for 11 months fromautumn 2005 to summer 2006, and compared withcommunities in three perennial streams and threeintermittent streams with perennial headwaters.Among the intermittent headwater sites, 95 taxa werefound, with Oligochaeta, Copepoda, Chironomidae,and Ostracoda occurring in greatest abundance. Eightmayfly, 6 stonefly and 16 caddisfly species werefound. Macroinvertebrate abundance at intermittentheadwater sites was equal to nearby perennialsites, but taxon richness per sample was lower.Community composition changed progressivelyduring the autumn-to-spring flow period (March-November 2005) at one intermittent headwater site,but at the other two sites a directional change wasnot evident, probably because stream flow stoppedbriefly several times during the flow period. Mostof the taxa collected in riffles during the flowperiod were also in disconnected pools after flowstopped in summer (January 2006), but some taxawere absent and the disconnected pools containedseveral additional species. The disconnected poolsin January had higher macroinvertebrate density andrichness than connected pools during the flow period.At intermittent sites with perennial headwaters,taxon richness and EFT (Ephemeroptera, Plecoptera,and Trichoptera) richness were intermediate between
M07040; Online publication date 13 March 2008Received 27 July 2007; accepted 20 November 2007
intermittent headwater sites and perennial streams.Two weeks after flow began, the intermittent siteswith perennial headwaters showed much higherEPT abundance and richness than the intermittentheadwater sites, but after 8 weeks the proportionaldifference was less.
Keywords benthic; temporary pools; colonisation;community composition
INTRODUCTION
Intermittent streams typically are identified with aridor semi-arid zones and Mediterranean climates, butthey can be abundant in humid temperate zones also(Williams & Hynes 1976; Dieterich & Anderson2000; Hansen 2001; Williams 2006). In NewZealand, previous studies (Pearson 1995; Parkynet al. 2006a,b) and anecdotal evidence suggest thatintermittent streams may be common in a number ofregions throughout the country. Despite this evidence,the fauna and ecology of intermittent streams havebeen little studied worldwide, or in New Zealand,where published studies of intermittent streamsnumber less than 10 (Pearson 1995; Death 2003;Fowler 2004; Datry et al. 2007; Robinson & Buser2007; Larnedetal. in press a,b). Limited knowledgeof the distribution of intermittent streams, poorrecognition of their biological values, and limitedunderstanding of the ecological processes sustainingtheir biological communities are currently hamperingefforts to conserve and manage intermittent streams,both overseas (Uys & O'Keefe 1997; Walker et al.1997; Sheldon et al. 2002; Maltchik & Medeiros2006) and in New Zealand (Parkyn et al. 2006a).Intermittent streams may become more commonin the future, as climate change scenarios predictgreater frequency and severity of drought in lowlandeastern parts of New Zealand (NIWA 2007), and asstreams are likely to come under increasing pressurefrom growing human demands for water.
Overseas research has highlighted importantdifferences in the macroinvertebrate community of
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

110 New Zealand Journal of Marine and Freshwater Research, 2008, Vol. 42
intermittent streams in different parts of the world.Studies in United Kingdom (Smith et al. 2003),Switzerland (Ruegg & Robinson 2004), Germany(Meyer et al. 2003), Portugal (Aguiar et al. 2002),and Algeria (Arab et al. 2004), have shown thatintermittent streams are typically species-poorcompared with nearby perennial streams, whereasstudies in Australia (Boulton & Suter 1986) andUnited States (Dieterich & Anderson 2000) havefound that the species diversity in intermittent streamscan be as high as in perennial streams. The degree offaunal similarity between intermittent and perennialstreams also appears to differ between and withincountries. Studies in United States (New York,Delucchi 1988; Alabama, Feminella 1996), Kenya(Shivoga 2001) and Australia (Boulton & Lake 1992a)have found high faunal overlap, whereas studies inCanada (Muchow & Richardson 2000; Williams2006), United Kingdom (Smith & Wood 2002), andUnited States (Oregon, Dieterich & Anderson 2000)have found significant differences, including speciesabsent from nearby perennial streams and previously-undescribed species. The contrasting results amongthese studies may be explained partly by site-specificfactors such as connectivity to subsurface refuges orproximity to nearby perennial waters (Smith & Wood2002), but it is likely also that in different regionsdistinct intermittent stream communities have evolvedowing to different climatic conditions (McMahon& Finlayson 2003). Therefore, it is valuable tocontinue extending intermittent stream research tonew countries.
Intermittent streams are inherently dynamic,therefore recording the seasonal periodicity oftheir macroinvertebrate communities is essentialto adequately describe their fauna and understandtheir ecology. Several studies (e.g., Boulton & Lake1992b; Robinson et al. 2004; Acuna et al. 2005)have described shifts in species composition duringthe flowing water phase, first as early colonists aregradually replaced by later arrivals, and then asthe riffle habitat contracts towards the end of theflow period. These shifts reflect differences amongspecies in their over-summering strategies, lifehistory characteristics, source of colonists, toleranceof changing physico-chemical conditions andcompetitive or predatory abilities (Boulton & Lake1992b; Ruegg & Robinson 2004). Thus, documentingthe temporal sequence of species in intermittentstreams provides a basis for further examining theover-summering strategies of individual species andthe ecological interactions that maintain diversemacroinvertebrate communities. The pools remaining
after flow ceases represent an aquatic habitat distinctfrom the flowing water habitats (Williams 2006),and because of their unique physico-chemicalconditions, these disconnected pools may provide atemporal niche, allowing a different set of fauna to"time-share" the same habitat space with the loticfauna (Bogan & Lytle 2007). Therefore, a completedescription of the intermittent stream fauna shouldinclude the disconnected pools.
Ecologically two types of intermittent stream canbe distinguished—those that occur in the headwatersand those that occur in the mid-reaches of streamnetworks (Dieterich & Anderson 2000). Headwaterintermittent streams result from the longitudinalexpansion and contraction of stream networks inresponse to seasonally rising and lowering watertables near the tops of catchments. Intermittent mid-reaches occur where streams that originate in wetupland regions flow across fiat plains with watertables that are permanently or seasonally below theriver bed. Intermittent reaches are produced where theflow path expands and contracts seasonally accordingto rising and lowering of the local water table orchanges in the flow volume of the river (Larned etal. in press a). In New Zealand, both intermittentheadwater and intermittent mid-reach streams areknown to occur (Parkyn et al. 2006a; Larned et al.in press a). A key characteristic distinguishing thesetwo stream types ecologically is the presence ofa source of invertebrate colonists upstream of theintermittent reach (Dieterich & Anderson 2000).Invertebrate communities in intermittent mid-reachesare expected to contain fauna that has drifted fromupstream perennial waters as well as fauna thatsurvives drought in situ, thus they should resemblea perennial stream community more closely thanheadwater intermittent streams do (Fritz & Dodds2005). Other authors(e.g., Shivoga 2001) have notedthat intermittent stream communities tend to recovermore quickly after drought where there are perennialreaches upstream.
The aims of our study were: (1) to compare theaquatic macroinvertebrate community (abundance,richness and composition) of small headwaterintermittent streams and larger mid-reach intermittentstreams to nearby mid-reach perennial streams; (2)to compare the fauna in New Zealand intermittentstreams to the fauna in similar habitats overseas; (3) todescribe the temporal changes in macroinvertebratecommunity composition of both intermittent streamtypes during the flow period; and (4) to compare theflow period community to the community persistingin disconnected pools after flow has ceased.
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14
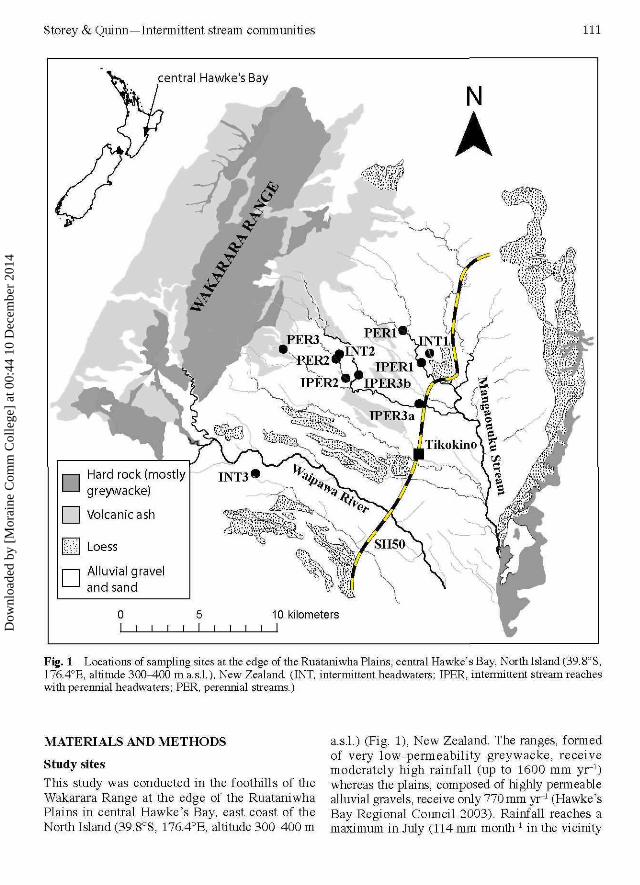
Storey & Quinn—Intermittent stream communities 111
Hard rock (mostlygreywacke^
Volcanic ash
Loess
Alluvial graveland sand
0 5 10 kilometers1 i i i i I i i i i
PERIINT2
IPER1IPER3b
Fig. 1 Locations of sampling sites at the edge of the Ruataniwha Plains, central Hawke's Bay, North Island (39.8°S,176.4°E, altitude 300^4-00 m a.s.l.), New Zealand (INT, intermittent headwaters; IPER, intermittent stream reacheswith perennial headwaters; PER, perennial streams.)
MATERIALS AND METHODS
Study sitesThis study was conducted in the foothills of theWakarara Range at the edge of the RuataniwhaPlains in central Hawke's Bay, east coast of theNorth Island (39.8°S, 176.4°E, altitude 300-400 m
a.s.l.) (Fig. 1), New Zealand. The ranges, formedof very low-permeability greywacke, receivemoderately high rainfall (up to 1600 mm yr"1)whereas the plains, composed of highly permeablealluvial gravels, receive only 770 mm yr"1 (Hawke'sBay Regional Council 2003). Rainfall reaches amaximum in July (114 mm month"1 in the vicinity
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

112 New Zealand Journal of Marine and Freshwater Research, 2008, Vol. 42
of the sampling sites) and a minimum in January(69 mm month"1), whereas evapotranspiration variesfrom 18 mm month"1 in July to 113 mm month"1
in January (Hawke's Bay Regional Council 2003).Maximum daily air temperatures on the Ruatani whaPlains range from 14°C in July to 24°C in January(Hawke's Bay Regional Council 2003). During thesummer months the water table beneath the plaindrops below the level of most stream beds, andstreams that originate in the Wakarara Range losetheir water vertically and dry up as they cross theplain (Hawke's Bay Regional Council 2003).
The study sites were situated on three types ofstream —intermittently-flowing streams that areintermittent to their headwaters (sites labelled INT),intermittently-flowing streams that have perennially-flowing headwaters (labelled IPER) and perennially-flowing streams (labelled PER). The first streamtype included three first-order tributaries (INT1-3) that originate at the edge of the plain (Fig. 1)and dry each summer for \-A months, accordingto local landowners (S. Macdonald & C. Hudsonpers. comm.). At the sample locations, these streamswere 1.5-14 cm deep and 0.5-1.1 m wide duringwinter basefiow(R. G. Storey pers.obs.). These threeintermittent streams flow into perennial streams,respectively 3 km, 300 m and >500 m downstreamof the study sites. Another three sites (IPER1-3)were located near the INT streams, also on alluvialgravels at the edge of the plains (Fig. 1). The IPERsites were on larger third-order streams that originatein the ranges and maintain perennial flow in theirheadwaters to points, respectively, >4, 1 and 10 kmupstream of the study sites. The IPERreaches dry outin most years for up to 3 months. During the summerbefore the study began, all of the INT and IPER siteshad been dry for several weeks (S. Macdonald & C.Hudson pers. comm.). At the sample locations, theIPER reaches were 6-9 cm deep and 2.8^.2 cmwide during winter basefiow (R G. Storey pers.obs.).Three perennial sites (PER1-3) were located as nearas possible to the INT sites, also in rolling hills at theedge of the plains and underlain by alluvial gravels.Two of the PER sites were on the same streams asIPER sites and the third on an adjacent stream. Atthe sample locations, the PER streams were 7-8cm deep and 2.3-3 m wide during winter basefiow(R. G. Storey pers.obs.). All of the study streams hada similar coarse substrate type dominated by pebblesand cobbles.
The main vegetation cover in the lowlands ispasture for sheep and cattle farming, but some pocketsof native forest remain, mostly in river gullies. All of
the study streams were surrounded by intact nativeforest at least 30 m wide on each bank and extendingat least 300 m upstream of the study site. Riparianvegetation cover over the stream channel was c. 80%at the INT sites and c. 60% at the IPER sites.
Data collectionBenthic macroinvertebrates were sampled from theintermittent (INT) sites throughout the flowing waterperiod and once during the disconnected pool phase.Sampling was conducted every 2 weeks from the endof March 2005 (autumn; 2 weeks after flowing waterappeared) until May 2005, and continued once permonth thereafter until mid-January (summer) 2006,except for December 2005 when no samples weretaken. Water flow ceased in these streams in earlyJanuary 2006, and disconnected pools remained forat least 1 month at all INT sites. Benthic sampleswere taken from IPER sites on the first samplingoccasion in March 2005 and again in mid-May2005, 8 weeks after flow began, by which timethe macroinvertebrate community was consideredto have a full complement of taxa (Fowler 2004).During May sampling, one IPER site (IPER3) waschanged as the new site better matched the other INTand IPER sites in terms of riparian forest cover (Fig.1). IPER3b was closer than IPER3a to a perennialreach (2.5 km cf. 10 km downstream), but similar toIPER3a in other physico-chemical parameters. PERsites were sampled once in mid-May to coincide withsampling at the IPER sites.
Benthic macroinvertebrates were collected using aSurber sampler with a 0.1 in2 base and a 250 /mi meshnet. Four samples were taken per site, located to includethe range of substrate sizes, water depths and currentvelocities found in riffles. Once in September 2005(during the flowing water period) and once in January2006 (when flow had ceased) pools were sampledinstead of riffles. To sample macroinvertebrates inpools, a hand net (250 /mi mesh) was used to sweep0.1 in2 of the pool bed and macrophytes, an areaequivalent to the Surber sampler.
Macroinvertebrate samples were preserved on sitewith 70% ethanol. In the laboratory, macroinvertebrateswere counted and identified under 50x magnificationusing the keys of Winterbourn et al. (2000) for insects,Towns & Peters (1996) for leptophlebiid mayflies,Smith & Ward (2007) for hydrobiosid caddisfiies,Fenwick & Duggan (2001) for amphipods, andWinterbourn (1973) for molluscs. All of the EPT(Ephemeroptera, Plecoptera and Trichoptera) andmollusc taxa were identified to genus, and most tospecies, half of the Coleoptera and Diptera taxa were
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

Storey & Quinn—Intermittent stream communities 113.
identified beyond family, and the Oligochaeta wereidentified to family. Some samples of Ostracoda andCopepoda were identified to genus or species, butthe list of genera and species for these two groups islikely to be incomplete.
At the same time as benthic macroinvertebrateswere sampled, physico-chemical parameters ofthe stream water, stream bed and riparian areawere recorded. Dissolved oxygen (±0.1 mg litre"1),temperature (±0.1°C) and conductivity (±5 //S cm"1)weremeasuredusingYSI 2357andYSI 2354meters,and current velocity was estimated visually incategories of slow, medium, and fast. Physicaldimensions of the stream (wetted width, bankfullwidth, and water depth) were measured with tapemeasures, and the % cover of sediment particle sizes(Wentworth size classes; boulders >256 mm, cobbles64-256 mm, pebbles 4-64 mm, gravel 2 4 mm, sand1/16-2 mm, silt <1/16 mm), organic matter (wood,leaves, moss, filamentous algae and macrophytes)and overhead riparian vegetation were estimatedvisually. In January 2006, physical dimensions of eachpool were measured. Since water physico-chemicalparameters can fluctuate greatly in small disconnectedpools owing to photosynthesis, decomposition, andthe lack of water flushing and turbulence (Williams2006), spot measurements of physico-chemicalparameters were considered inadequate to characterisethe habitat. Therefore, during a period of fine weather,temperature (±0.1°C) and dissolved oxygen (±0.1 mglitre"1) of representative pools were logged for 54 hat intervals of 30 min.
From mid-May 2005, temperature loggers (OnsetTidbit®) were anchored on riffle areas of the streambed and on stream banks at the three INT sites todetect cessation of flow between sampling visits.Daily fluctuations in the streambed logger werecompared with those of the bank logger, and whenboth loggers showed similar fluctuations, the streambed was considered dry.
Data analysisMacroinvertebrate abundance data were summarisedusing measures of total abundance, EPT abundance,total taxon richness, and EPT richness. INT, PER,
and IPER sites were compared on the basis of theseunivariate measures using /-tests and analysis ofvariance (ANOVA) followed by Tukey's HSD (Zar1996), using SPSS vll . Abundance measures werelog(x+l)-transformed to reduce the correlationbetween variance and mean, satisfying an assumptionof ANOVA (Zar 1996).
The full taxonomic composition of the varioussites and sampling dates was compared with multi-dimensional scaling (MDS) ordinations on 4th root-transformed abundance data (Clarke & Warwick2001), using PRIMER v5 software (PRIMER-ELtd). Differences between sites were then testedusing analysis of similarities (ANOSIM; Clarke &Warwick 2001), and the taxa responsible for thesedifferences were determined using the SIMPERprocedure, with a 90% cut-off for low contributions(Clarke & Warwick 2001).
RESULTS
Flow persistenceStream flow began at all sites except INT 1 by mid-March 2005, and ceased by early January 2006.INT1 had surface pools but no flow on 30 March,however by 13 April flow had begun. During summer2005/06, aquifer levels were unusually high and poolsremained at INT sites throughout the 2005/06 dryperiod. Temperature logger measurements, rainfalldata and one direct observation suggested that flowwas continuous during the entire 2005 flow period atINT1, but at INT2 and INT3 surface flow in rifflesmay have stopped for a time between some samplingdates, e.g., for a week between 13 and 27 April, from13 to 16 May, 7 to 12 June, 9 to 18 September and25 September to 10 October 2005.
Physico-chemical characteristicsWater temperatures at the INT sites during the flowperiod varied between 7.1 and 15.2°C, dissolvedoxygen levels between 6.5 and 11.3 mg litre"1, andconductivity between 99 and 192 //S cm"1 (Table1). In May 2005, water temperatures at PER sites(between 8.0 and 9.1°C ) were equivalent to those
Table 1 Water physico-chemical characteristics of INT (intermittent headwater)sites during the flow period, March-November 2005.
March July/August November
Temperature (°Q 13.5-15.2 7.1-8.5 10.3-11.7Dissolved oxygen (mg litre"1) 6.5-9.3 9.2-11.3 7.1-9.8Conductivity (jiS cm"1) 124-192 102-151 99-168
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14
114 New Zealand Journal of Marine and Freshwater Research, 2008, Vol. 42
mossfilamentous
algaemacrophyte
leaf
small wood
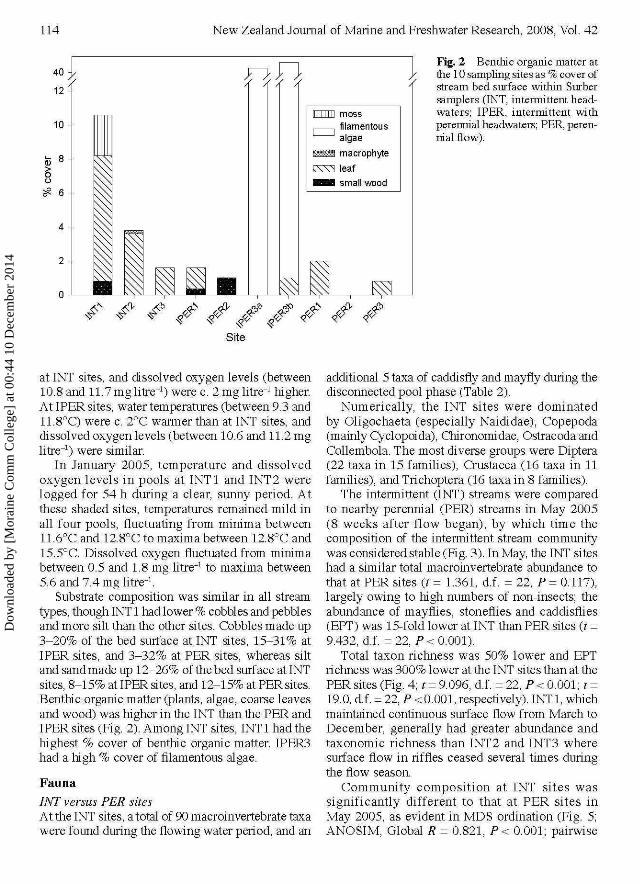
Fig. 2 Benthic organic matter atthe 10 sampling sites as % cover ofstream bed surface within Surbersamplers (INT, intermittent head-waters; IPER, intermittent withperennial headwaters; PER, peren-nial flow).
Site
at INT sites, and dissolved oxygen levels (between10.8 and 11.7 mg litre"1) were c. 2 mg litre"1 higher.At IPER sites, water temperatures (between 9.3 and11.8°C) were c. 2°C warmer than at INT sites, anddissolved oxygen levels (between 10.6 and 11.2 mglitre"1) were similar.
In January 2005, temperature and dissolvedoxygen levels in pools at INT1 and INT2 werelogged for 54 h during a clear, sunny period. Atthese shaded sites, temperatures remained mild inall four pools, fluctuating from minima between11.6°C and 12.8°C to maxima between 12.8°C and15.5°C. Dissolved oxygen fluctuated from minimabetween 0.5 and 1.8 mg litre"1 to maxima between5.6 and 7.4 mg litre"1.
Substrate composition was similar in all streamtypes, though INT 1 had lower % cobbles and pebblesand more silt than the other sites. Cobbles made up3-20% of the bed surface at INT sites, 15-31% atIPER sites, and 3-32% at PER sites, whereas siltand sand made up 12-26% of the bed surf ace at INTsites, 8-15% at IPER sites, and 12-15% at PER sites.Benthic organic matter (plants, algae, coarse leavesand wood) was higher in the INT than the PER andIPER sites (Fig. 2). Among INT sites, INT 1 had thehighest % cover of benthic organic matter. IPER3had a high % cover of filamentous algae.
FaunaINT versus PER sitesAt the INT sites, a total of 90 macroinvertebrate taxawere found during the flowing water period, and an
additional 5 taxa of caddisfiy and mayfly during thedisconnected pool phase (Table 2).
Numerically, the INT sites were dominatedby Oligochaeta (especially Naididae), Copepoda(mainly Cyclopoida), Chironomidae, Ostracoda andCollembola. The most diverse groups were Diptera(22 taxa in 15 families), Crustacea (16 taxa in 11families), and Trichoptera (16 taxa in 8 families).
The intermittent (INT) streams were comparedto nearby perennial (PER) streams in May 2005(8 weeks after flow began), by which time thecomposition of the intermittent stream communitywas considered stable (Fig. 3). In May, the INT siteshad a similar total macroinvertebrate abundance tothat at PER sites (t = 1.361, d.f. = 22, P = 0.117),largely owing to high numbers of non-insects; theabundance of mayflies, stonefiies and caddisfiies(EPT) was 15-fold lower at INT than PER sites (t =9.432, d.f. =22, P< 0.001).
Total taxon richness was 50% lower and EPTrichness was 300% lower at the INT sites than at thePER sites (Fig. 4; t = 9.096, d.f. = 22, P < 0.001; t =19.0, d.f. = 22, P < 0.001, respectively). INT 1, whichmaintained continuous surface flow from March toDecember, generally had greater abundance andtaxonomic richness than INT2 and INT3 wheresurface flow in riffles ceased several times duringthe flow season.
Community composition at INT sites wassignificantly different to that at PER sites inMay 2005, as evident in MDS ordination (Fig. 5;ANOSIM, Global R = 0.821, P< 0.001; pairwise
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

Storey & Quinn—Intermittent stream communities 115
Fig. 3 Density of benthic mac-roinvertebrates in May 2005 (INT,intermittent headwaters; IPER, in-termittent with perennial headwa-ters; PER, perennial flow).
Minor taKaOligochaetaMolluscaCrustaceaOther arthropodsDipteraColeopteraTnchopteraPlecopteraEphemeroptera
Fig. 4 Taxonomic richness ofbenthic invertebrates (total persite) in May 2005 (INT, intermit-tent headwaters; IPER, intermit-tent with perennial headwaters;PER, perennial flow).
Minor taxaOligochaetaMolluscaCrustaceaOther arthropodsDipteraColeopteraTnchopteraPlecopteraEphemeroptera
# # # $r <F <FSite
Table 2 Taxonomic composition of INT (intermittent headwater) and IPER (intermittent, but connected to perennialheadwaters) sites. INT data are total values for the flow season (March-November 2005); IPER data are total valuesof March and May 2005 sampling dates. Where the pool phase community added extra taxa, the total of pool andflowing phase is included in brackets.
Species (or next
EphemeropteraPlecopteraTrichopteraColeopteraDipteraOther insectaAcariCrustaceaMolluscaOligochaetaOther phylaTotal
lowest taxon)
6(8)6
13 (16)8
2271
16344
90 (95)
INT sites
Families (or nextlowest taxon)
1(1)3
7(8)7157111
$44
63 (64)
Av. abundanceper sample
101221109542:
221854918261688
Species (or nextlowest taxon)
71
124113151
•3452
IPER sites
Families (or nextlowest taxon)
31646315134
37
Av. abundanceper sample
36119513-53298
252
926
370
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

116 New Zealand Journal of Marine and Freshwater Research, 2008, Vol. 42
Stress: 0.13
PER3;PER3
Fig. 5 Multi-dimensional scaling(MDS) ordination of macroinver-tebrate community compositionat INT (intermittent headwater),IPER (intermittent with perennialheadwater) and PER (perennialflow) sites in May 2005.
Table 3 Community composition of INT (intermittent headwater) sites and comparison to PER (perennial flow)sites in May 2005. EPT refers to Ephemeroptera, Plecoptera, and Trichoptera. Taxa with the highest % contribution towithin-group similarity in SIMPER analysis are those occurring most abundantly and consistently among INT sites.
Taxa with highestcontribution to within-groupsimilarity among INT sites
NaididaeCyclopoidaCollembolaAcariOstracodaHydrobiosisChironominiNematodaEriopterini(F. Tipulidae)Orthocladiinae
14.309.028.137.946.826.395.504.55
4.253.70
EPT taxa occurringat all INT sites on atleast one occasion
DeleatidiumZephlebiaNeozephlebiaAcroperlaZelandobiusPsilochoremaOxyethiraPolyplectropus
Taxa at INT sitesbut not PER
ZelandobiusAcroperlaOrthopsycheOeconesusTarapsyche olisNesoperla fulvescensKempynusParadixa
PsychodidaeStratiomyidaeZygoptera
Taxa abundant atPER sites, butabsent from INT
IchthybotusColoburiscusHelicopsychePycnocentriaOlingaPycnocentrodesAoteapsycheArchichauliodes
testfor INT-PER, R = 1.0, significant atP = 0.001).Taxa most strongly characterising INT sites includedNaidid oligochaetes, cyclopoid copepods, andCollembola (Table 3). Several EPT taxa occurredat all INT sites, including Deleatidium, Zephlebia,Acroperla, and Psilochorema. No undescribedspecies were found in this study, but several species,e.g., Zelandobius, Acroperla, Tarapsyche olis, andNesoperla fulvescens were found in the intermittentstreams but not in the neighbouring perennial ones(Table 3).
IPER versus INT and PER sites
At IPER sites, 52 macroinvertebrate taxa were foundin March and May 2005 samples combined. Thefauna at IPER sites was dominated numerically byOligochaeta and Trichoptera (Table 2). Diptera andEphemeroptera were also abundant, but Crustaceaaccounted for only a modest proportion of thetotal abundance. The most diverse groups wereTrichoptera (12 taxa in 6 families) and Diptera (11taxa in 6 families).
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

Storey & Quinn—Intermittent stream communities 117
Overall abundance at IPER sites (Fig. 3) was 26%lower than at INT sites (non-significant) and 63%lower than at PER sites (F = 3.888, d.f. = 2,33, P =0.030; Tukey's HSD P = 0.024). EPT abundancewas intermediate between INT and PER sites, andthe differences with both other stream types werestatistically significant (F = 25.066, d.f. = 2,33,P< 0.001; Tukey's HSD P = 0.030 and/>< 0.001respectively).
Total taxon richness and EPT richness at IPERsites (Fig. 4) were also intermediate between INTand PER sites, and for both of these measures, thedifferences among the three stream types werestatistically significant (for taxon richness, F =31.315, d.f. = 2,33, P< 0.001, Tukey's HSD P <0.001, P< 0.001; for EPT richness, F= 136.986,d.f. = 2,33, P < 0.001, Tukey's HSD P = 0.038, P <0.001).
As well as abundance and richness, communitycomposition of IPER sites was intermediate betweenthat of INT and PER sites (Fig. 5) and significantlydifferent from both (Global R = 0.821, P< 0.001;pairwise tests: INT-IPER, R = 0.609; IPER-PER,R = 0.768; both significant at P = 0.001). Taxamost strongly separating IPER sites from PER sitesincluded Olinga, Coloburiscus and Pycnocentrod.es,whereas those most strongly separating IPER fromINT sites included Deleatidium, Ostracoda andCollembola (Table 4).
Temporal changesin intermittent stream communitiesINT sites
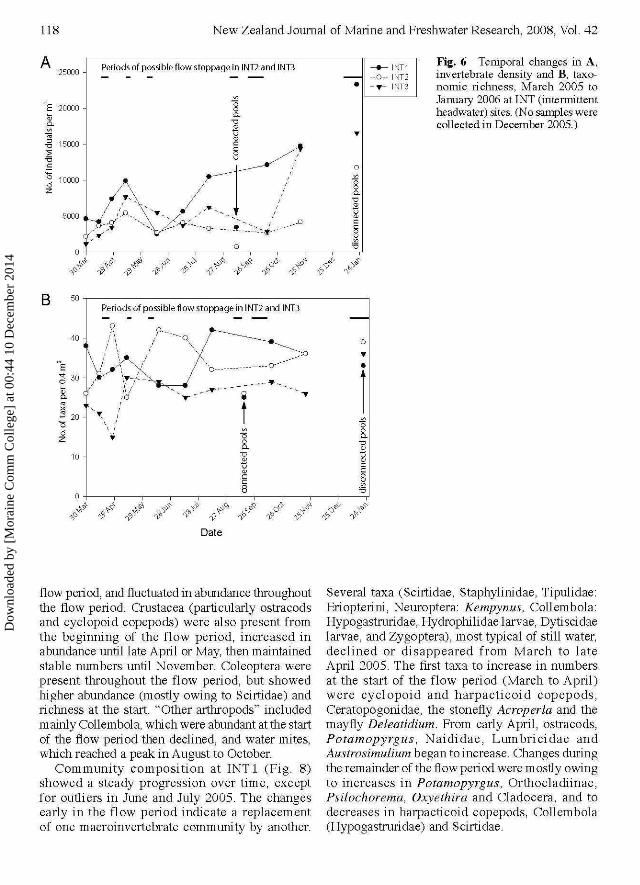
Total macroinvertebrate abundance and taxonomicrichness at INT sites (Fig. 6) showed considerablevariability overtime. Nevertheless, macroinvertebrateabundance at all sites showed a strong increase overthe first 8 weeks of flow, and also as the streamflow reduced between October and November 2005.Taxon richness showed less clear trends over timeand less consistency among sites. Overall, therewas no clear increase in richness, but different sitesreached peaks of richness at different times. At INT 1and INT3, richness declined for the first 2-A weeksand then increased.
Because INT1 was the only INT site thatmaintained continuous flow throughout the flowperiod, the remainder of the analysis focused onINT1, and patterns in the other two streams werenoted when they differed markedly from INT 1.
The abundance of EPT taxa at INT1 was verylow at the first appearance of surface water in March(25m~2), and reached a peak of 800 m~2 in May (Fig.7). Caddisfiy numbers increased later in the yearthan stonefiies or mayflies. Stonefiies declined from410 m~2 in August to 35 m~2 in October, whereasmayflies maintained a high abundance of c. 200m~2 to November. Diptera (especially Chironomidaeand Tipulidae) were present from the start of the
Table 4 Taxa accounting for separation in ordination space between IPER (intermittent, but connected to perennialheadwaters) sites, INT (intermittent headwater) sites, and PER (perennial) sites in May 2005. Taxa separating eachpair of sites are ranked on the basis of % contribution to the separation of sites in SIMPER analysis, with averagedissimilarity/SD of dissimilarities indicated for each taxon. (*absent from IPER; fabsent from INT; X absent fromIPER.)
Top 10 taxaseparating IPERfrom PER sites
Lower abundanceOlingaColoburiscus*Pycnocentrode sAoteapsyche*NeozephlebiaHelicopsycheHydra enidaeAustroperla cyrene*Hydora
Higher abundanceCyclopoida
%contribution
5.974.904.714.604.084.003.663.213.15
3.35
Av. dissim/SD
3.555.001.464.391.722.872.021.961.31
2.23
Top 10 taxaseparating IPERfrom INT sites
Higher abundanceDeleatidiumHydrobiosisTanytarsinifHydora~\A rchichauliodes\Zelandobius\
Lower abundanceOstracodaCollembolaChironominiAcroperlax
%contribution
5.073.663.573.563.553.52
4.143.883.533.43
Av. dissim/SD
1.901.211.971.553.331.34
1.251.401.691.13
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14
118 New Zealand Journal of Marine and Freshwater Research, 2008, Vol. 42
A Periods of possible flow stoppage in INT2 and INT3 - • - I N T 1--O-- INT2- T - INT3
Periods of possible flow stoppage in INT2 and INT3
Fig. 6 Temporal changes in A,invertebrate density and B, taxo-nomic richness, March 2005 toJanuary 2006 at INT (intermittentheadwater) sites. (No samples werecollected in December 2005.)
Date
flow period, and fluctuated in abundance throughoutthe flow period. Crustacea (particularly ostracodsand cyclopoid copepods) were also present fromthe beginning of the flow period, increased inabundance until late April or May, then maintainedstable numbers until November. Coleoptera werepresent throughout the flow period, but showedhigher abundance (mostly owing to Scirtidae) andrichness at the start. "Other arthropods" includedmainly Collembola, which were abundant at the startof the flow period then declined, and water mites,which reached a peak in August to October.
Community composition at INT1 (Fig. 8)showed a steady progression over time, exceptfor outliers in June and July 2005. The changesearly in the flow period indicate a replacementof one macroinvertebrate community by another.
Several taxa (Scirtidae, Staphylinidae, Tipulidae:Eriopterini, Neuroptera: Kempynus, Collembola:Hypogastruridae, Hydrophilidae larvae, Dytiscidaelarvae, andZygoptera), most typical of still water,declined or disappeared from March to lateApril 2005. The first taxa to increase in numbersat the start of the flow period (March to April)were cyclopoid and harpacticoid copepods,Ceratopogonidae, the stonefly Acroperla and themayfly Deleatidium. From early April, ostracods,Potamopyrgus, Naididae, Lumbricidae andAustrosimulium began to increase. Changes duringthe remainder of the flow period were mostly owingto increases in Potamopyrgus, Orthocladiinae,Psilochorema, Oxyethira and Cladocera, and todecreases in harpacticoid copepods, Collembola(Hypogastruridae) and Scirtidae.
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

Storey & Quinn—Intermittent stream communities 119
Fig. 7 Temporal changes inben-thic macroinvertebrate density atintermittent headwater site INT1,March 2005 to January 2006. (Nosamples were collected in Decem-ber 2005.)
connectedpools
Minor taKaOligochaetaMolluscaCrustaceaOther arthropodsDipteraColeopteraTnchopteraPlecopteraEphemeroptera
Fig. 8 Multi-dimensional scaling(MDS) ordination of macroinver-tebrate community composition atintermittent headwater site INT 1:temporal changes from Marchto November 2005 and faunaof connected pools (September2005) and remnant pools (Janu-ary 2006).
Mar
Stress: 0.15
Jan (pools)
Nov
Sept (pools)
Community composition at INT2 and INT3 didnot show a clear temporal progression as at INT1.However, ANOSIM showed that for each site thefirst samples in March were separated from the laterdates (INT2: R = 0.624, P = 0.001; INT3: ./? = 0.567,P = 0.001).
IPER sites
At the IPER sites, macroinvertebrate abundance was890-1950 individuals m~2 in March and increasedby 100-200% from March to May (Fig. 6A). Boththe initial abundance and % increase were the sameorder of magnitude as those at the INT sites. EPTabundance at the IPER sites (6.5-53.5 m~2) was,on average, 18 times higher at the IPER than at theINT sites (0.0-2.5 individuals m~2), and increased byonly 1.50- to 1.75-fold from March to May 2005,
compared with 15-to 30-fold at the INT sites. Therewere also large differences in EPT abundance amongthe three IPER sites. In March 2005, EPT abundanceat IPER2 (53.5 individuals m~2) was six times higherthan at IPER3 and IPER1 (6.5 and 9.5 individualsm~2, respectively).
Taxonomic richness changed by -20 to +45%from March to May among IPER sites, comparedwith -8 to +30% among the INT sites over the sameperiod (Fig. 6B). The only IPER site that did notincrease in richness over this period was IPER2,which was the closest of the three sites to a perennialreach, and had high richness from March.
Community composition at IPER sites (Fig. 9)was distinctly different from that at the INT sites,and this distinction persisted over time. However,temporal changes were similar in the two stream
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

120 New Zealand Journal of Marine and Freshwater Research, 2008, Vol. 42
March Stress: 0.23
March
May
Fig. 9 Multi-dimensional scal-ing (MDS) ordination of changesin macroinvertebrate communitycomposition at INT (intermittentheadwater) and IPER (intermittentwith perennial headwater) sitesbetween March and May 2005.
types. Among the 10 main taxa driving the temporalchange in both stream types, were Naididae(SIMPER, 7.9% and 5.8% contribution in INT andIPER sites, respectively), Hydrobiosis (5.3 and 5.6%,respectively), Ostracoda(5.6 and 3.7%), cyclopoidcopepods (5.4 and 3.5%) and Collembola (4.6 and3.4%). The similarity between INT and IPER sites,was 42% in March 2005 and 51% in May 2005. Thusthere was a slight convergence in composition asthe communities matured between March and May2005.
Disconnected pools
At INT sites, the disconnected pools in January hadhigher taxonomic richness and considerable highermacroinvertebrate density than the connected poolsin September (Fig. 6). They also had slightly higherrichness and density than the riffles during the flowperiod. Community composition in the January poolswas distinct from that of the September pools andsomewhat distinct from that of the riffle community(e.g., Fig. 7 and 9). However, the difference betweenJanuary and previous months was mostly owingto different proportions among taxa, rather than achange in taxa; most of the taxa found in the riffleswere able to maintain at least a small populationin the disconnected pools. The disconnected poolshad much higher abundances of caddisflies (owingto Polyplectropus, which reached between 475 and2455 individuals m~2 at the three INT sites), higherabundances of crustaceans (2400-16 000 individualsm~2; mostly cyclopoid copepods and ostracods),and lower abundances of mayflies and stonefiies(20-1265 and 0-18 individuals m~2, respectively)
than riffles. At INT1, the disconnected pools alsocontained high numbers of beetles (1100 individualsm~2; all Scirtidae) and at INT2 and 3, Diptera weremuch more abundant in the disconnected pools(4900 and 9800 individuals nr2, respectively; mostlyCulicidae, Chironomidae, andPsychodidae)thaninriffles. Some taxa found in riffles during the flowperiod were not found in the disconnected pools,including Acroperla (no stonefiies were found atINT1 andINT3), Hydrobiosis, andCeratopogonidae.There were also a few taxa in the disconnectedpools, Triplectides cephalotes, Pseudoeconesus andPsilochorema donaldsoni, that were not found inriffles or connected pools. Overall, the communityin the disconnected pools in January 2005 wassomewhat different to that in the connected poolsin September 2005 (SIMPER average similarity 50%and 37% for INT 1 and INT2, respectively), mostlyowing to the absence of taxa from the connectedpools. The similarity was greater between theJanuary pool community and the March-Novemberriffle community (SIMPERaverage similarity 55%,43% and 62% for INT1, 2 and 3, respectively).
DISCUSSION
Intermittent headwater sites comparedwith perennial sites and overseas studiesAs expected, the headwater intermittent streamshad lower richness than nearby perennial streams.Community composition also differed significantly,but total macroinvertebrate abundance in the
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

Storey & Quinn—Intermittent stream communities 121
headwater intermittent streams was comparable tonearby perennial streams owing to high numbers ofmicro-crustaceans and oligochaetes.
Despite being taxon-poor compared with theperennial streams, the headwater intermittent streamswere more diverse than we expected in all majortaxonomic groups. Our sites were considerablyricher than those in several New Zealand intermittentstream studies (e.g., Parkyn et al. 2006b; Datry etal. 2007; Larned et al. in press a), but similar tothose in Death (2003), after adjusting faunal lists inthose studies to account for differences in taxonomicresolution and the habitat type sampled.
Compared with sites in overseas studies, oursites had moderate to high taxon richness. Rifflehabitats in Canada (Williams & Hynes 1976) andGermany (Meyer et al. 2003) yielded totals of 41and 34 taxa, respectively; two intermittent streamsin southeast Australia yielded a total similar toours (94 taxa; Boulton & Lake 1992c), thoughAustralian intermittent streams are consideredto have particularly diverse intermittent streamfauna (Boulton & Suter 1986). In our study, thedisconnected-pool fauna added only a few taxa,thus the overall richness for the year (95 taxa) wasconsiderably less than in the Australian study (166taxa; Boulton & Lake 1992c), but still comparableto three intermittent streams in United States (92taxa; Dieterich & Anderson 2000) and greater thanthe Canadian study (63 taxa) (Williams & Hynes1976). The EPT richness in our study (30 taxa overthe year) was intermediate between the EPT richnessreported in the Australian (42) and North American(55) studies on one hand, and the Canadian (11) andGerman (10) studies on the other.
We did not expect the intermittent stream faunain New Zealand to be as diverse as that overseas.Although New Zealand perennial streams havesimilar taxon richness to comparable streams inNorth America, Europe and Asia (Thompson &Townsend 2000), we expected that New Zealand'sturbulent history of glaciation, tectonic uplift andinundation would not have allowed species toevolve adaptations to an intermittent habitat, as isthought to have occurred in Australia (McMahon &Finlayson 2003) and the Mediterranean (Gasith &Resh 1999). However, in many intermittent streamstudies the fauna consists largely of opportunisticspecies that also occur widely in perennial streams(Progar & Moldenke 2002; Williams 2006). NewZealand's stream fauna includes many species withopportunistic characteristics such as generalistfeedine, flexible life histories, and extended flight
and egg-hatching periods (Winterbourn et al. 1981),and therefore may be well predisposed to takeadvantage of intermittent habitats.
Whereas several overseas studies have shownintermittent streams to have a distinct speciesassemblage and little overlap with the fauna of nearbyperennial streams (e.g., Dieterich & Anderson 2000;Williams 2006), a similar comparison from our datais unlikely to yield this result. In our study, 38% ofthe taxa in the headwater intermittent (INT) streamswere not found in the higher-order perennial (PER)streams. However, most of those taxa, e.g., Kempynus(Order Neuroptera), Paradixa, Psychodidae,Stratiomyidae (Order Diptera) and Zygoptera, arecommonly found in small perennial headwaters,stream margins, slow-flowing backwaters, or matsof decaying vegetation in perennial streams. Othertaxa, e.g., Zelandobius, Acroperla, Orthopsyche,and Oeconesus, are common stream insects thatmay have been missed by chance in the perennialstreams. In contrast to some overseas studies (e.g.,Johansson &Nilsson 1994; Feminella 1996; Dieterich& Anderson 2000) we found no new species atintermittent sites, though two species (N.fulvescensand T. olis) that are rare in perennial streams(Winterbourn et al. 2000) occurred several timesat INT sites, suggesting they may be well-adaptedto intermittent conditions. Overall, in our studyintermittent stream communities appeared distinctfrom perennial stream communities by the presenceof taxa that have some semi-aquatic larval stages (e.g.,N. fulvescens, Acroperla trivacuata, Zelandobiusfucillatus-group, Kempynus sp.) and absence ofcertain genera (Coloburiscus, Archichauliodes andAoteapsyche) that are widespread and abundantin most New Zealand streams (Quinn & Hickey
Intermittent streams with perennial headwatersIPER sites were expected to contain a mixture of taxathat survived the dry period in situ and those thatdrifted from upstream. Drift should result in IPERsites having higher EPT abundance and higher taxonrichness than intermittent headwater streams. IPERsites did have higher EPT abundance and taxonomicrichness, though total macroinvertebrate abundancewas similar in both intermittent stream types.
Temporal changesin intermittent stream communitiesWe expected to see a steady increase in macro-invertebrate abundance and taxonomic richness asaquatic taxa recolonised the previously dry habitat.
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

122 New Zealand Journal of Marine and Freshwater Research, 2008, Vol. 42
Abundance and taxonomic richness were variableover the sampling period, partly owing to slightdifferences among field staff in positioning the Surbersamplers and to low and high flow events precedingthe sampling dates. Nevertheless, macroinvertebrateabundance at all sites showed a strong increaseover the first 8 weeks of flow, suggesting thatrecolonisation was a dominant process during thisperiod. Taxon richness at INT1 and INT3 declinedfor the first 2-A weeks and then increased, suggestingthat an initial community was replaced by a laterone.
We expected to see a progressive development inintermittent stream communities as early colonistswere replaced by later arrivals. Such a gradual changewas seen at INT 1 (the outliers in June and July maybe explained partly by different field staff placingthe Surber sampler in slightly finer substrate); atINT2 andINT3 development of the community wasnot directional, probably because of interruptions inwater flow, but the community present in the first 2weeks of flow was different from that in subsequentsamples. The temporal changes in INT1 showedsome similarities and some differences to overseasstudies. Typically a suite of early colonisers, includingChironomidae, Simuliidae and Ceratopogonidae(Boulton 2003; Lake 2003; Robinson et al. 2004;Acuna et al. 2005), is gradually replaced by laterarrivals, including EPT taxa and elmid beetles(Boulton 2003; Robinson et al. 2004). In our study,the early flow-period community was dominated bystill-water and semi-aquatic taxa, which appearedto have quickly colonised pools formed by thefirst rains. Chironomids were present from the firstappearance of surface water, but copepods, mayfliesand stonefiies increased in numbers more rapidlythan chironomids.
We expected that IPER sites would show a morediverse and "typical" stream community than INTsites early in the flow period, and that over timethe community at INT sites would become moresimilar to the IPER community. This convergencein the communities is because the IPER sites maybe colonised by drift, which is rapid and shouldcarry a variety of typical stream taxa, whereas mosttaxa at the INT sites must survive the dry periodin situ, and recolonise through slower pathwayssuch as egg hatching. Our results largely confirmedthese expectations. From March to May there was astrong convergence in EPT abundance and a slightconvergence in community composition betweenINT and IPER sites. Therefore, our results confirmthe findings of other studies that drift from perennial
upstream sources significantly increases the recoveryrate of intermittent stream communities after drought(Shivoga 2001; Fritz & Dodds 2005). We also foundevidence that distance from a perennial source affectsthe importance of drift as a recolonisation pathwayand the rate of community recovery after drought(Fritz & Dodds 2005; Aguiar et al. 2002; Larned etal. in press a). Since drift plays an important role inthe composition and ecology of intermittent streamcommunities, the effect of distance from source is anarea worthy of further research in New Zealand.
Disconnected poolsMost intermittent stream studies have noted largedifferences in the macroinvertebrate communitybetween flowing water habitats (riffles and pools)and the disconnected pools that remain after flowstops (e.g., Williams & Hynes 1976; Boulton 2003;Bogan & Lytle 2007). We found much greatermacroinvertebrate density and somewhat highertaxonomic richness in the disconnected Januarypools than in the connected September pools,which may be a result of the reduction in availablehabitat as riffles dried in summer (e.g., Boulton& Lake 1992b). We did not see a large differencein taxon richness between the disconnected poolsand the flowing water riffle community, but, apartfrom the still-water caddisfiy Polyplectropus, therewere very few EPT individuals in pools, as alsoobserved by Bonada et al. (2006) and Williams(1996). Some low-oxygen tolerant taxa, such aschironomids and gastropods, were more abundantin the disconnected pools than in riffles. However,in contrast to other studies, only a few lentic taxaappeared in summer pools. Therefore, the summerpool habitat did not increase greatly the overallbiodiversity of the intermittent stream system atthese sites as reported by Bogan & Lytle (2007). Ourresult may be because INT sites in our study werelocated in dense native bush (two in steep gullies),thus they lacked the physical habitat provided bymacrophytes and they may not be as accessible toaerially colonising Hemiptera and Coleoptera as themore open sites in other studies. The summer poolcommunity in nearby pasture streams had a muchhigher richness in these two taxa (Storey & Quinnunpubl. data).
Our results show that intermittent headwaterstreams can support diverse macroinvertebratecommunities, including some rare taxa, thereforethey have natural values that deserve protection.Adequate protection must begin with surveys toassess the abundance and distribution of intermittent
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

Storey & Quinn—Intermittent stream communities .123.
streams. Our results also showed that intermittentstream communities are likely to be less diversethan those in perennial streams, therefore wewould expect loss of species if perennial streamsbecome intermittent through climate change or waterabstraction. Finally, intermittent streams connectedwith upstream perennial reaches have morediverse aquatic communities than those without.Therefore, to maintain macroinvertebrate diversityin intermittent mid-reach streams, their connectivityto upstream perennial reaches must be maintainedand protected from on-line structures such as damsand reservoirs.
ACKNOWLEDGMENTS
We thank Graham Sevicke-Jones, Brett Stansfield, VickieHansen and Andrew Lamason for help with field work;Spencer and Kate Macdonald, Carola and Mike Hudsonfor site access and help with field work; Ian Andrews,Ian Duggan, Ilaria Mazzini, Ian McLellan, and BrianSmith for help with identifications; and Holly Ferguson,Claire Taylor, and Nathan Wagstaff for help with samplesorting.
REFERENCES
Acuna V, Munoz I, Giorgi A, Omella M, Sabater F,Sabater S 2005. Drought and postdrought recoverycycles in an intermittent Mediterranean stream:structural and functional aspects. Journal of theNorth American Benthological Society 24(4):919-933.
Aguiar FC, Ferreira MT, Pinto P 2002. Relative influenceof environmental variables on macroinvertebrateassemblages from an Iberian basin. Journal ofthe North American Benthological Society 21(1):43-53.
Arab A, Lek S, Lounaci A, Park Y 2004. Spatialand temporal patterns of benthic invertebratecommunities in an intermittent river (NorthAfrica). Annales de Limnologie 40(4): 317-327.
Bogan MT, Lytle DA 2007. Seasonal flow variationallows 'time-sharing' by disparate aquaticinsect communities in montane desert streams.Freshwater Biology 52(2): 290-304.
Boulton AJ 2003. Parallels and contrasts in the effects ofdrought on stream macroinvertebrate assemblages.Freshwater Biology 48: 1173-1185.
Boulton AJ, Lake PS 1992a. The ecology of twointermittent streams in Victoria, Australia. II.Comparisons of faunal composition betweenhabitats, rivers and years. Freshwater Biology27: 99-121.
Boulton AJ, Lake PS 1992b. The ecology of twointermittent streams in Victoria, Australia.III. Temporal changes in faunal composition.Freshwater Biology 27: 123-138.
Boulton AJ, Lake PS 1992c. The macroinvertebrateassemblages inpools and riffles in two intermittentstreams (Werribee and Lerderderg Rivers, southerncentral Victoria). Occasional Papers from theMuseum of Victoria 5: 55-71.
Boulton AJ, Suter PJ 1986. Ecology of temporary streams— an Australian perspective. In: de DeckkerP, Williams WD ed. Limnology in Australia.Dordrecht, Netherlands, Dr. W. Junk Publishers.Pp. 313-327.
Clarke KR, Warwick RM 2001. Change in marinecommunities: an approach to statistical analysisand interpretation 2nd ed. Plymouth, PRIMER-E. 156 p.
Datry T, Larned ST, Scarsbrook MR 2007. Responsesof hyporheic invertebrate assemblages to large-scale variation in flow permanence and surface-subsurface exchange. Freshwater Biology 52(8):1452-1462.
Death RG 2003. Spatial patterns in lotic invertebratecommunity composition: is substrate disturbanceactually important? Canadian Journal of FisheriesandAquatic Sciences 60(5): 603-611.
Delucchi CM 1988. Comparison of community structureamong streams with different temporal flowregimes. Canadian Journal of Zoology 66:579-586.
Dieterich M, Anderson NH 2000. The invertebrate faunaof summer-dry streams in western Oregon Archivfur Hydrobiologie 147(3): 273-295.
Feminella JW 1996. Comparison of benthicmacroinvertebrate assemblages in small streamsalong a gradient of flow permanence. Journal ofthe North American Benthological Society 15(4):651-669.
Fenwick G, Duggan I 2001. Biodiversity identificationworkshop: freshwater amphipods and commonplanktonic crustaceans. National Institute ofWater and Atmospheric Research Ltd, PalmerstonNorth
Fowler RT 2004. The recovery of benthic invertebratecommunities following dewatering in two braidedrivers. Hydrobiologia 523: 17-28.
Fritz KM, Dodds WK 2005. Harshness: characterisationof intermittent stream habitat over space and time.Marine and Freshwater Research 56(1): 13-23.
Gasith A, Resh VH 1999. Streams in Mediterraneanclimate regions: abiotic influences and bioticresponses to predictable seasonal events. AnnualReview of Ecology and Systematics 30: 51-81.
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

124 New Zealand Journal of Marine and Freshwater Research, 2008, Vol. 42
Hansen WF 2001. Identifying stream types andmanagementimplications. Forest Ecology and Management143(1-3): 39-46.
Hawke's Bay Regional Council 2003. Ruataniwha Plainswater resources investigation. EnvironmentalMonitoring Group Technical Report, June 2003.
Lake PS 2003. Ecological effects of perturbation bydrought in flowing waters. Freshwater Biology48: 1161-1172.
Lamed ST, Hicks DM, Schmidt J, Davey AJH, Dey K,Scarsbrook MR, Arscott DB, Woods RA in pressa. The Selwyn River of New Zealand: a benchmarksystem for alluvial plain rivers. River Researchand Applications.
Larned ST, Datry T, Robinson CT in press b. Invertebrateand microbial responses to inundation in anephemeral river reach in New Zealand: effects ofpreceding dry periods. Aquatic Sciences.
Maltchik L, Medeiros ESF2006. Conservation importanceof semi-arid streams in north-eastern Brazil:implications of hydrological disturhance andspecies diversity. Aquatic Conservation-Marineand Freshwater Ecosystems 16(7): 665-677.
McMahon TA, Finlayson BL 2003. Droughts and anti-droughts: the low flow hydrology of Australianrivers. Freshwater Biology 48(7): 1147-1160.
Meyer A, Meyer El, Meyer C 2003. Lotic communities oftwo small temporary karstic stream systems (EastWestphalia, Germany) along a longitudinal gradientof hydrological intermittency. Limnologica 33(4):271-279.
Muchow C, Richardson J 2000. Unexplored diversity:macroinvertebrates in coastal British Columbiaheadwater streams. Proceedings of a conferenceon the biology and management of species andhabitats at risk. 503 p.
NIWA 2007. Climate change impacts: New Zealandand the South Pacific. IPCC fourth assessmentreport. 4 p.
Parkyn S, Wilding TK, Croker G 2006. Small headwaterstreams of the Auckland Region Volume 4:Natural values. NIWA Client Report HAM2006-134. National Institute of Water andAtmosphericResearch Ltd, Hamilton. 52 p.
Parkyn S, Phillips N, Smith BJ 2006. Aquatic invertebratebiodiversity and spatial characteristion of non-perennial streams in native forest in the Waikato.NIWA Client Report HAM2006-041. NationalInstitute of Water and Atmospheric Research Ltd,Hamilton. 28 p.
Pearson CP 1995. Regional frequency analysis of lowflows in New Zealand rivers. Journal of Hydrology(NZ) 33(2): 94-122.
Pires AM, Cowx IG, Coelho MM 2000. Benthicmacroinvertebrate communities of intermittentstreams in the middle reaches of the GuadianaBasin (Portugal). Hydrobiologia 435: 167-175.
Progar RA, Moldenke AR 2002. Insect production fromtemporary and perennially flowing headwaterstreams in western Oregon. Journal of FreshwaterEcology 17(3): 391-407.
Quinn JM, Hickey CW 1990. Characterisation andclassification of benthic invertebrate communitiesin 88 New Zealand rivers in relation toenvironmental factors. New Zealand Journal ofMarine and Freshwater Research 24: 387-409.
Robinson CT, Buser T 2007. Density-dependent life historydifferences in a stream mayfly (Deleatidium)inhabiting permanent and intermittent streamreaches. New Zealand Journal of Marine andFreshwater Research 41: 265-271.
Robinson CT, Tockner K, Burgherr P 2004 Drift benthosrelationships in the seasonal colonization dynamicsof alpine streams. Archiv fur Hydrobiologie160(4): 447-470.
Ruegg J, Robinson CT 2004. Comparison ofmacroinvertebrate assemblages of permanentand temporary streams in an Alpine flood plain,Switzerland Archiv fur Hydrobiologie 161(4):489-510.
Sheldon F, Boulton AJ, Puckridge JT 2002. Conservationvalue of variable connectivity: aquatic invertebrateassemblages of channel and floodplain habitats ofa central Australian arid-zone river, Cooper Creek.Biological Conservation 103: 13-31.
Shivoga WA 2001. The influence of hydrology on thestructure of invertebrate communities in twostreams flowing into Lake Nakuru, Kenya.Hydrobiologia 458(1-3): 121-130.
Smith BJ, Ward JB 2007. A guide to the New ZealandHydrobiosidae caddisflies. National Institute ofWater andAtmospheric Research Ltd, Hamilton
Smith H, Wood PJ 2002. Flow permanence andmacroinvertebrate community variability inlimestone spring systems. Hydrobiologia 487(1):45-58.
Smith H, Wood PJ, Gunn J 2003. The influence ofhabitat structure and flow permanence oninvertebrate communities inkarst spring systems.Hydrobiologia 510(1-3): 53-66.
Thompson R, Townsend CR 2000. New Zealand's streaminvertebrates: an international perspective. In:Collier KJ, Winterbourn MJ ed New Zealandstream invertebrates: ecology and implicationsfor management. Christchurch, New ZealandLimnological Society. Pp. 415.
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14

Storey & Quinn—Intermittent stream communities 125
Towns DR, Peters WL 1996. Fauna of New Zealand.No. 36: Leptophlebiidae. Duval CT ed Lincoln,Manaaki Whenua Press. 143 p.
Uys MC, O'Keefe JH 1997. Simple words and fuzzyzones: early directions for temporary river researchin South Africa. Environmental Management 21:517-531.
Walker KF, Puckridge JT, Blanch SJ 1997. Irrigationdevelopment on Cooper Creek, central Australia—prospects for a regulated economy in a boom-and-bust ecology. Aquatic Conservation-Marineand Freshwater Ecosystems 7(1): 63-73.
Williams DD 1996. Environmental constraints intermporary fresh waters and their consequencesfor the insect fauna. Journal of the North AmericanBenthological Society 15(4): 634-650.
Williams DD 2006. The Biology of Temporary Waters. 1sted. Oxford, Oxford University Press. 337 p.
Williams DD, Hynes HBN 1976. The ecology of temporarystreams I. The faunas of two Canadian streams.Internationale Revue gesamten Hydrobiologie61(6): 761-787.
Winterbourn MJ 1973. A guide to the freshwater molluscaof New Zealand. Tuatara 20(3): 141-159.
Winterbourn MJ, Rounick JS, Cowie B 1981. Are NewZealand stream ecosystems really different?New Zealand Journal of Marine and FreshwaterResearch 15: 321-328
Winterbourn MJ, Gregson KLD, Dolphin CH 2000.Guide to the Aquatic Insects of New Zealand2nd ed. Auckland, Entomological Society of NewZealand 102 p.
Zar J 1996. Biostatistical Analysis. 3rd ed Upper SaddleRiver, N.J., Prentice Hall.
Dow
nloa
ded
by [
Mor
aine
Com
m C
olle
ge]
at 0
0:44
10
Dec
embe
r 20
14