complexity of early and middle successional stages in a rocky intertidal surfgrass community
TRANSCRIPT

Oecologia (Berlin) (1983) 60:56-65 Oecologia �9 Springer-Verlag 1983
Complexity of early and middle successional stages in a rocky intertidal surfgrass community Teresa Turner Department of Zoology, Oregon State University, Corvallis, OR 97331, USA
Summary. A variety of simple models have been proposed to describe ecological succession (e.g., Connell and Slatyer 1977), but these models do not address some agents that may increase complexity. To determine the complexity of a natural sequence, four null hypotheses were tested: (1) seasonality of growth, recruitment, and mortality does not influence succession; (2) the interspecific interactions that produce successional change are the same throughout the sequence; (3) consumers have no influence on succession, and (4) small scale spatial variation in establishment and mortality does not occur. These hypotheses were tested in a low zone rocky intertidal community normally dominated by the surfgrass Phyllospadix scouleri on the Oregon coast.
Succession experiments initiated in different seasons and an herbivore exclusion experiment falsified each of these hypotheses. At two sites (Boiler Bay and Squaw Island) experimental plots that were cleared in the spring were first colonized by Ulva sp., but those cleared in other seasons were first colonized by Phaeostrophion irregulare. Ulva ap- pears adapted to colonize space made available by winter storms, whereas Phaeostrophion takes advantage of space made available by sand movement in the fall. Another sea- sonal pattern was the sharp decline in total algal cover in the fall, when wave action increased. At Squaw Island, the presence of Phaeostrophion established in fall and winter significantly inhibited the summer establishment of Ulva, though Utva occupied some space epiphytically. In contrast, at Boiler Bay, a different, stronger type of interspecific in- teraction occurred: Phaeostrophion totally inhibited the es- tablishment of filamentous diatoms. The cover of Ulva in the summer was also influenced by herbivores, but Ulva cover declined in the fall in both herbivore exclusions and controls, suggesting that herbivores were not solely respon- sible for its replacement. Local variation was demonstrated, because after three years of succession, replicate plots often differed. The early colonists, Phaeostrophion and Ulva, con- tinued to dominate some plots; in other plots they had been replaced by middle successional species, including Cryptosiphonia woodii and Odonthaliafloccosa. In still other plots Rhodomela larix had replaced other species. Similarly complex successional sequences occur in many natural com- munities. Thus, the features that simple models do not ad- dress may add complexity to succession, and for some com- munities different approaches must be developed.
Introduction
Several simple models have been proposed to describe suc- cession. All of these models were designed to explain pre- dictable successional changes with little variation in the tim- ing of change or in the species composition of each stage. The oldest model (Clements 1916) postulates that succes- sional changes result from early space occupiers preparing the way (facilitation) for later species. This traditional mod- el has recently been questioned (Drury and Nisbet 1973; Connell and Slatyer 1977), and three alternative models have been proposed. (1) Successional changes may result solely from differences in life histories. Rapidly growing species dominate early in the successional sequence; slowly growing species dominate later: no interspecific interactions occur (Connell and Slatyer 1977). (2) Successional changes may result from natural enemies or physical conditions that kill early colonists, allowing later species to dominate. This view supposes that early species inhibit later ones. As a result, without this mortality late successional species would not replace initial colonizers or would do so more slowly (Connell and Slatyer 1977; Sousa 1979). (3) Successional changes may result from later species outcompeting early colonists. This view also supposes that early successional species inhibit later ones. If early space occupiers were ab- sent, later species would be able to grow faster, because they would not pay the energetic costs of competition (Horn 1981).
Despite the differing basic viewpoints of these models, they all fail to address certain features that may make suc- cession more complex. None of these models, for example, addresses seasonality of recruitment, growth, and mortality. And, although different types of interspecific interactions have been described, changes in the types of interactions during the course of succession have not been described. Only one of the models deals explicitly with the potential impact of consumers, and none of the models considers local variation in settlement and mortality. Of course, sim- ple models can never include the complexity of the natural world, but they can capture the salient features and have predictive value. For example, Sousa (1979) studied a suc- cessional sequence whose essential features were predicted by a simple model proposed by Connell and Slatyer (1977). In other cases, however, all these simple models fail, and more complex models must be constructed (Horn 1981).

57
Here I investigate whether these features add complexity to one system - early and middle successional stages in a rocky intertidal community normally dominated by surf- grass (the seagrass Phyllospadix scouleri). Marine rocky in- tertidal communities such as this one have a variety of fea- tures that lend them to investigations of these questions (Paine 1977). The principle natural disturbance, removal of organisms by waves, can be mimicked experimentally. Because naturally disturbed areas are relatively small, ex- periments can be replicated to detect spatial variation. Small marine herbivores can be manipulated to determine their effects. Also, successional changes are relatively fast. Other experimental studies conducted in marine communi- ties (e.g., Sousa 1979) provide a basis for comparison.
In this study I focus on four questions: (1) In which seasons do particular species recruit and
in which seasons are the different algal species most abun- dant ?
(2) Does the presence of established species inhibit or facilitate the recruitment of later colonizing species?
(3) Do herbivores influence early succession? (4) How different is the species composition of experi-
mental replicates and will these differences persist as alter- native equilibria?
Community and study sites
I investigated succession in surfgrass beds on the Oregon coast in low zone (+0.6-0.0 m) rocky intertidal horizontal benches. In areas with little disturbance, surfgrass can form a virtual monoculture (Turner 1982). It persists in all sea- sons for many years and prevents other species from invad- ing. Thus, surfgrass beds functionally resemble the persis- tent mussel beds described by Paine and Levin (1981). Phys- ical disturbances (waves or wave-borne logs) prevent mo- nopolization of space in both communities and produce a mosaic of patches in different successional stages. In this study I experimentally simulated these natural disturbances by removing surfgrass and other macroscopic organisms from the rock. I present data for 3 years on the resultant early and middle colonizing stages. Elsewhere I describe the undisturbed community (Turner ~ 982) and the initiation of the last successional stage, surfgrass recruitment (Turner 1983).
This study was carried out at two main sites and one secondary site (fully described in Turner 1982). The main sites differ primarily in substratum: Squaw Island (43~ 124~ has sandstone; Boiler Bay (44~ 124~ mudstone. Both sites experience moderate wave action and sand scour. I observed occasional pockets of sand up to 2 cm deep in experimental plots at both sites. An herbivore exclusion experiment was conducted at a third site 0.7 km south of Squaw Island. This site was similar both physically and biologically to the Squaw Island site.
Methods
To determine the effect of temporal and spatial variation on successional sequences, I initiated succession experi- ments in four seasons at Squaw Island and in three seasons at Boiler Bay. Throughout this paper the term treatment refers to the season of initiation, and the replicates of each treatment are referred to as plots or clearings. In each sea- son at each site I removed surfgrass with a wrecking bar and putty knife from four replicate 0.25 m 2 areas, each
selected to have a surfgrass canopy cover >90%. Four unmanipulated plots nearby served as controls. At Squaw Island the plots are all within a 300 m 2 area; at Boiler Bay within a 900 m 2 area. Scraping with a putty knife does not remove all encrusting algae, but when natural distur- bances like waves remove surfgrass, encrusting algae also remain. The experimental plots were permanently marked with concrete nails and Sea Goin' Poxy Putty | I recorded patterns of space utilization by plants and densities of herbi- vores in these plots generally four times a year for 3 years. Surfgrass plants surrounding the experimental removals grew slightly into the plots, and a few surfgrass seeds re- cruited (Turner 1982), but this slow recovery did not appear to influence the other species.
To quantify patterns of space utilization within these experimental plots, I estimated percent cover of sessile in- vertebrates and plants using a flexible vinyl quadrat marked with 100 randomly placed dots: I counted the number of dots above each species (Menge 1976; Lubchenco and Menge 1978). Overlap caused some values to exceed 100% cover. That is, of one plant lay on top of another under a dot, each species had 1% cover. This aspect of the meth- ods differed from those of Menge (1976) but resembled those of Littler and Littler (1980). Those species present but not under any dots were recorded arbitrarily as 0.5% cover. Densities of limpets and the coiled snail Tegulafune- bralis were also recorded; some herbivores, including am- phipods, isopods, and the gastropod Lacuna marmorata, were too small to be accurately counted.
To normalize the data for parametric statistics, I made an arcsine transformation of the percent cover data as rec- ommended by Sokal and Rohlf (1969). In figures the mean percent covers and standard errors are plotted in degrees, but the ordinate axis is backtransformed for easier interpre- tation.
To determine the effect of herbivores on early succes- sion, stainless steel mesh fences were used to exclude them from four 25 cm x 25 cm clearings. The fences were 5 cm high with a 2 cm lip projecting outward at the top and had a 0.5 cm mesh size. The experiment ran from May 4, 1980, when the surfgrass (100% canopy cover) was re- moved, to October 23, 1980. During this period, I manually removed herbivores that entered the exclusions and cleaned the fences at least twice a month. Because the fences were not completely effective, herbivory was reduced, but not eliminated. The design included two control treatments. Unmanipulated controls consisted of four cleared plots lo- cated randomly at least 1 m from any exclusion fences and experienced natural successional changes. Fence controls consisted of four plots immediately seaward of the four fences (i.e., with a fence along one side). These plots were presumably accessible to herbivores but experienced some of the artifacts of the fences, such as changes in water move- ment and inoculation by the spores of drift algae entangled on the fences. I removed surfgrass from a 0.5 m buffer zone around each of the plots to eliminate the influence of shad- ing by adjacent surfgrass. Algal cover was estimated during this experiment with the same methods as above, except that only 25 randomly placed dots were used.
Results
The removal of surfgrass completely changes the commun- ity; within 3 months algal cover increases significantly

58
leo
50
IOC
5C n" LLI > O O I'-- 0
Z loo (...) r r LLI G_
50
IOO
50
COVER OF TWO EARLY COLONIZERS
SUMMER REMOVAL
RemovaE r
. . . . . . . . . . . . . .
FALL. REMOVAL
, i i , i i , JJ ,, J, , , ,, , , i , , , ,
WINTER REMOVAL
;PRING" REMOVAL I Ulvo
[-] Phaeosfroph/bn
Before 1 ~ ~ ~ Removol
.,. o , , 'Su' 'F ' 'W''Sp' 'Su' 'F ' 'W ' 'Sp ' 'Su' 'F ' W''Sp'
1978 J 1979 I f980 I t981
Fig. 1. Seasonal cover of two early colonizing species, Ulva sp. (dark bars) and Phaeostrophion irregulare (light bars), at Squaw Island following surfgrass removal in four seasons. I initiated four replicates in each season. Bars represent means, and lines represent standard errors, which are symmetrical about the means. Both epiphytic and epilithic cover are included. Ulva cover in summer 1979 is significantly higher in the spring removals, without Phaeos- trophion, than in the fall and winter removals, with Phaeostrophion (P < 0.05, Mann-Whitney U-test)
(Turner 1982) and many species invade (Appendix). In con- trol plots algal cover does not increase and no new species invade. In broad outline the successional sequence runs thus: at both sites, depending on the season, one of two species colonize abundantly within 3 months, the brown alga Phaeostrophion irregulare or the green alga Ulva sp. Members of the genus Ulva generally possess an annual blade with a perennial holdfast of small rhizoidal processes (Abbott and Hollenberg 1976). Phaeostrophion has an en- crusting perennial holdfast and annual blades. Additionally, at Boiler Bay, filamentous diatoms sometimes formed thick mats on bare substratum in the summer. In some plots Phaeostrophion and Ulva continued to dominate for the entire 3 years; in others they were replaced by a suite of middle successional algal species including mats of polysi- phonous red algae, the red blade-like algae Iridaea spp. and Gigartinapapillata, and the branching red algae Crypto- siphonia woodii and Odonthalia floecosa. In other plots the slowest growing middle successional species, Rhodomela
U/vo
70 ~ [ ] Phoeostroph/on
50 FALL REMOVAL
30 ~ l ~ ~ ~ ~ F r ~ / . ~ I0
.~ 1 50
> 3 0 C)
~ JO
I I / / - "
WINTER REMOVAL
/ / 50 SPRING REMOVAL -x*
30
,o
0 I w I s p l l s u l l F [ [ w l [
I 1979 I 1980 I 1981 Fig. 2. Seasonal cover of two early colonizing species, Ulva sp. and Phaeostrophion irregulare, at Boiler Bay following surfgrass removal in three seasons. I initiated four replicates in each season. Bars represent means, and lines represent standard errors, which are symmetrical about the mean. Both epiphytic and epilithic cover are included. Breaks in the axis indicate dates for which no data were taken. The asterisk indicates data were taken for only one replicate
larix, another branched red alga, was apparently in the process of replacing other middle successional species after 3 years. Rhodomela may eventually form nearly mono- specific stands. Observations and experiments indicate that it resembles surfgrass in persisting for years and in prevent- ing other species from invading (Turner 1982, 1983; D 'An- tonio 1982). Presumably, surfgrass will eventually replace all the middle succesional species.
Seasonal patterns
Most of the species colonizing experimental plots after surf- grass removal had distinct seasonal patterns in recruitment, cover, or both. The early colonists at Squaw Island (Fig. 1) and Boiler Bay (Fig. 2) illustrate the trends. Only a few tiny blades of Ulva sp. were present in the late fall and winter. Over the summer, however, Ulva settled densely (or regrew from a holdfast) and grew very rapidly. The brown blade Phaeostrophion irregulare also had a striking seasonal pattern (Fig. 1). It settled (or regrew from a hold- fast) in the late fall, grew to occupy its maximum cover in winter, and declined as the blades became small and desiccated in the spring and summer.
Middle successional algae at Squaw Island also had sea- sonal patterns of abundance. Gigartina papillata, for exam- ple, colonized three replicates the first spring and reached peak cover that summer (Fig. 3). Its cover declined dramati- cally in the fall (a similar pattern to that observed by Slo- cure 1980), then increased in these same plots again in spring and summer 1980, and again in spring 1981. The fact that Gigartina continually grew in these plots and only

59
100 -
n~ LU > O O
D 5 0 -
<
LL O
o~
o
'w' Sp Su F W Sp Su F W Sp I I , 80 1,98,
Fig. 3. Seasonal cover of Gigartina papillata in the three replicates it invaded at Squaw Island. It did not invade Boiler Bay. For each replicate, I expressed the percent cover for each sampling date as a percentage of the maximum observed in that replicate. This allows comparisons of general trends in plots with different settlement densities. Maxima ranged from 17 to 52 percent cover. These percentages were arcsiue transformed, and the bars represent means, the lines standard errors, which are symmetrical about the means
Io0 - -
s: u > o o
D -~ 50- x
DATE OF REMOVAL [ ] Summer 1978
~1 [ ] W~n,.r ,9~-79
0 F I Iw I I Sp Su I I F I I W I I Sp I I SU I I F I I W I I Sp I Is /
1978 [ i979 l 1980 I 1981
Fig. 4. Seasonal cover of IHdaea spp. (L flaccida, L heterocarpa and L cordata combined) in the replicates it colonized at Squaw Island: four replicates of the treatments initiated in summer, fall, and spring, and three replicates initiated in winter. These species were not as abundant at Boiler Bay. For each replicate, I expressed the percent cover for each sampling date as a percentage of the maximum observed in that replicate. This allows comparisons of general trends in plots with different settlement densities. Maxima ranged from 5 to 81 percent cover. These percentages were arcsine transformed, and the bars represent means, the lines standard er- rors, which are symmetrical about the means
these plots suggests that Gigartina might regrow from pe- rennial cells or that its propagules do not settle far from parent plants. Iridaea spp. (L heterocarpa, L flaccida, and I. cordata), which have annual blades and a perennial crus- tose holdfast, exhibited a similar pat tern (Hruby 1975; Hansen and Doyle 1976; Hansen 1977; Fos ter 1982; Gaines 1983). They often colonized within 3 months of clearing but did not reach peak cover until summer (Fig. 4). Cover declined severely in fall and winter, but usually rebounded in the spring and summer.
A few species invaded plots every spring and summer at both sites. They usually did not occupy over 5% cover and often grew epiphytically. These included the red algae Halosaccion glandiforme and Porphyra sp., f i lamentous dia- toms, and the brown algae Leathesia difformis and Leathe- sia nana. Mainly because of these rarer species, erect algal species diversity (measured as number of species) peaked in summer (Turner 1982).
Algal cover as a whole also exhibited seasonal patterns. At Squaw Island, because the cover of most individual spe-
DATE OF REMOVAL [ ] Summer 1978
[ ] Fall 1978
�9 Winter 1978~79
[ ] Spring 1979
I ,979 I 1980 I ,981
Fig. 5. Seasonal cover of all upright algae, illustrating the conse- quences of four different seasons of initiation at Squaw Island. All values over 100% were considered as 100%. Bars represent means; lines standard errors
I 0 0 -
~> 50-
O- L I I
DATE OF REMOVAL
[] Fall 1978 �9 Winter 1978-79
[ ] Spring 1979
1979 l
//,
~2
1980
~5 .. J_
-L [ 1981
Fig. 6. Seasonal cover of all upright algae, in three seasons at Boiler Bay. All values over 100 % were considered as 100 %. Bars represent means; lines standard errors. Breaks in the axis indicate dates for which no data were taken. The asterisk indicates data were taken for only one replicate
cies peaked in the summer, total algal cover also peaked (Fig. 5) despite the opposi te trend in Phaeostrophion cover (Fig. 1). At Boiler Bay, algal cover peaked in the summer in most plots (Fig. 6), but clearings init iated in fall 1978 did not exhibit a fall decline until the second year.
Inhibition: variation in the interspecific interactions
Because some species recruited seasonally and space was cleared each season, comparisons between treatments in these experiments reveal the effect of established species on later colonizers (Dean and Hurd 1980). Only the first year 's da ta were analyzed, because after this time replicate plots were no longer similar. Other factors that might vary along with the presence of established species could con- found interpreta t ion of these observations. Time since initi- at ion is the most likely confounding factor. Plots exposed to propagules for a longer time may consequently have a greater cover of a species. Nonetheless, if cover is less in plots exposed longer the opposi te of the result expected from this confounding factor - then established species clearly inhibit later colonizers.
Less cover in plots exposed longer was exactly the result observed: thus, Phaeostrophion inhibited later colonizers at
60
both sites. At Squaw Island, Phaeostrophion inhibited the establishment of Ulva. In summer 1979 Ulva invaded all the treatments, but it occupied significantly more space in plots initiated in spring 1979 than in plots initiated in sum- mer or fall 1978 (Fig. 1; P<0.05, Mann-Whitney U-test). Although the winter colonist Phaeostrophion preempted some space, the inhibition was not strong, because Ulva occupied 64.6% cover in the presence of Phaeostrophion versus 98.5% in its absence. Ulva could invade despite a high Phaeostrophion cover, because it grew epiphytically on the small, desiccated Phaeostrophion blades.
At Boiler Bay both filamentous diatoms and Ulva in- vaded plots initiated in spring 1979. Diatoms generally in- vaded depressions, whereas Ulva invaded emergent areas. Differences in summer 1979 between plots initiated in fall 1978 and spring 1979 suggest Phaeostrophion completely preempts filamentous diatoms, which do not become epi- phytic on Phaeostrophion. Diatoms occupied a mean of 59.0% cover in plots without Phaeostrophion, but did not invade plots in which it was present. Reductions in Ulva cover caused by Phaeostrophion were not significant at Boi- ler Bay, suggesting that strength of interspecific interactions varies in space.
Herbivores
An exclusion experiment suggests herbivores could influ- ence the cover of Ulva during the summer. I initiated the experiment May 4, 1980. By August 10, Ulva occupied 94.3% of the understory space in the exclusions but only 29.0% and 30.0% in the fence controls and unlnanipulated controls respectively, a highly significant difference ( P < 0.01, ANOVA). By October 23, Ulva still occupied more space in the exclusions than in the fence controls and un- manipulated controls (55.5% vs. 15.4% and 22.2% respec- tively). However, the difference was no longer significant (P> 0.05, ANOVA). Ulva may have decreased in cover be- cause of physical conditions in the fall. I terminated the experiment in October 1980, because the fences could not have withstood late fall and winter storms. I conclude that herbivores could depress the cover of Ulva early in succes- sion, but they apparently are not solely responsible for the decline of Ufva.
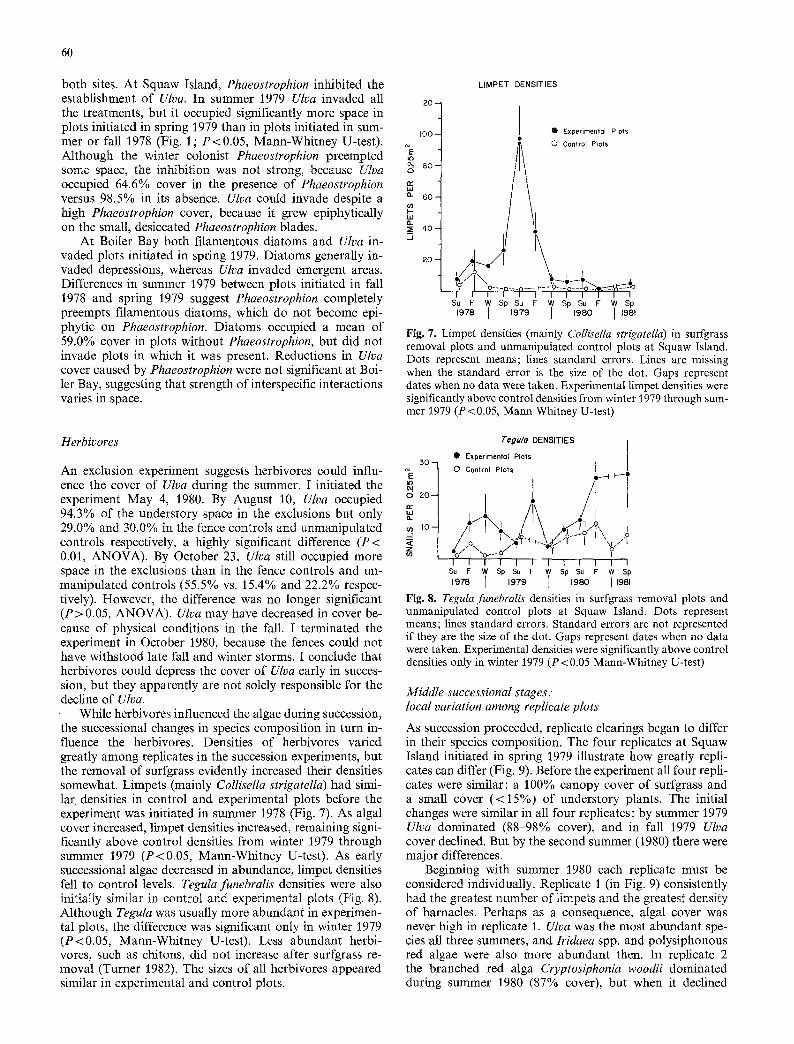
While herbivores influenced the algae during succession, the successional changes in species composition in turn in- fluence the herbivores. Densities of herbivores varied greatly among replicates in the succession experiments, but the removal of surfgrass evidently increased their densities somewhat. Limpets (mainly ColIisella strigatella) had simi- lar. densities in control and experimental plots before the experiment was initiated in summer 1978 (Fig. 7). As algal cover increased, limpet densities increased, remaining signi- ficantly above control densities from winter 1979 through summer 1979 (P<0.05, Mann-Whitney U-test). As early successional algae decreased in abundance, limpet densities fell to control levels. Teguta funebralis densities were also initially similar in control and experimental plots (Fig. 8). Atthough Teguta was usually more abundant in experimen- tal plots, the difference was significant only in winter 1979 (P<0.05, Mann-Whitney U-test). Less abundant herbi- vores, such as chitons, did not increase after surfgrass re- moval (Turner 1982). The sizes of all herbivores appeared similar in experimental and control plots.
120 -
IO0- "E -
8 0 -
,.=, o. 60-
== _~ 40- .-I
LIMPET DENSITIES
�9 Experimental Plots
/ 1 ! 0 COntrol plOts
r+" I " . . . . . . . . . . . . . . . , k ~ % T o _ ~ . . ~ . . § I I I I I I I I I I I I
Su F W Sp Su F W Sp Su F W Sp 1978 [ 1979 I 1980 11981
Fig. 7. Limpet densities (mainly Collisella strigatella) in surfgrass removal plots and unmanipulated control plots at Squaw Island. Dots represent means; lines standard errors. Lines are missing when the standard error is the size of the dot. Gaps represent dates when no data were taken. Experimental limpet densities were significantly above control densities from winter 1979 through sum- mer 1979 (P < 0.05, Mann-Whitney U-test)
5O
0 2 0
o_ m fO _.J
$
7egu/o DENSITIES
�9 Experimental Plots / I 0 Control Plots �9 |._~ p._/
! ,/ ( / -~ / , -oh" ~ /
I I I I I t I 1 I I I I Su F W Sp Su F W Sp Su F W Sp ,978 I ,979 I ,980 1,98,
Fig. 8. Tegula funebralis densities in surfgrass removal plots and unmanipulated control plots at Squaw Island. Dots represent means; lines standard errors. Standard errors are not represented if they are the size of the dot. Gaps represent dates when no data were taken. Experimental densities were significantly above control densities only in winter 1979 (P < 0.05 Mann-Whitney U-test)
Middle successional stages: local variation among replicate plots
As succession proceeded, replicate clearings began to differ in their species composition. The four replicates at Squaw Island initiated in spring 1979 illustrate how greatly repli- cates can differ (Fig. 9). Before the experiment all four repli- cates were similar: a 100% canopy cover of surfgrass and a small cover (<15%) of understory plants. The initial changes were similar in all four replicates: by summer 1979 Ulva dominated (88-98% cover), and in fall 1979 Ulva cover declined. But by the second summer (1980) there were major differences.
Beginning with summer 1980 each replicate must be considered individually. Replicate 1 (in Fig. 9) consistently had the greatest number of ]impets and the greatest density of barnacles. Perhaps as a consequence, algal cover was never high in replicate 1. Ulva was the most abundant spe- cies all three summers, and Iridaea spp. and polysiphonous red algae were also more abundant then. In replicate 2 the branched red alga Cryptosiphonia woodii dominated during summer 1980 (87% cover), but when it declined

61
lO0 -
5 0 -
0
~176 t 5 0 ~ o,
I.~ > o 0 0 mmo0o I
Z~: I00~ laJ 0 rr" i,i a.
50- Ri~
O
I 0 0 -
5 0
0
[. �9 , l , o
[ ] PhoaosYrophio~
[ ] Crypto~p~onio
l ~ ~ I I I I I
2.
3.
i I l i II I I 11 t l I i
4.
1979 ] 198( ] 1981
Fig. 9. Cover of four most abundant species, Ulva sp., Phaeostro- phion irregulare, Cryptosiphonia woodii, and Rhodomela larix, in each of four replicate plots numbered one to four (in the upper right hand corner) at Squaw Island initiated in spring 1979. Both epiphytie and epilithic cover are included. The algal cover before removal was beneath a 100% cover surfgrass canopy
in fall 1980, early successional species, Ulva an d Phaeostro- phion, again cap tured mos t o f the space. In contras t , in replicate 3 a l though Cryptosiphonia invaded, it never occu- pied as a high a cover as in replicate 2, possibly because ano the r b r anched red alga, Rhodomela larix, occupied 48% cover in summer 1980. This high cover of Rhodomela rela- tively early in succession might have been caused by re- growth f rom cells no t completely removed when the experi- men t was in i t ia ted (see below). In replicate 4 Crvptosiphonia replaced Ulva and d o m i n a t e d the next summer (1980). Rho- domela i nvaded replicate 4 in winter 1980, grew slowly, and replaced Cryptosiphonia as the mos t a b u n d a n t species by spr ing 1981. Thus , the four replicates experienced entire- ly different middle successional sequences.
Differences a m o n g repIicates occurred in several t reat- ments at bo th sites. A t Squaw Is land at the end of the exper iment (in spr ing 1981), six species had over 20% cover in at least one of the 16 plots (consider ing all the clearing dates in Table 1). The replicates ini t ia ted in summer 1978
Table 1. Abundant erect plant species in each replicate of surfgrass removal experiments from data taken in June 1981 at Squaw Island. Numbers preceding each species name in parentheses are percent covers, epilithic and epiphytic cover combined
Treatment Repli- Species with Other abundant (date care greatest % cover species succession # (>20% cover) initiated)
Summer 1 b (68) Ulva (55) Phaeostrophion 1978 2 u (66) Phaeostrophion none
3 b (48) Phaeostrophion (27) Ulva 4 a (43) Phaeostrophion none
Fall 1978 1 b (41) Phaeostrophion (32) Ulva, (24) polysiphonous
red algae 2 ~ (51) Rhodomela (27) Iridaeaflaccida 3 b (51) Phaeostrophion none 4 b (73) Ulva none
Winter 1 a (56) Phaeostrophion (38) Rhodomela 1979 2 b (29) Iridaeaflaecida (23) Phaeostrophion
3" (30) Rhodomela (24) Ulva 4 b (61) Phaeostrophion (41) Ulva
Spring 1 b (29) Ulva none 1979 c 2 b (53) Phaeostrophion (35) Cryptosiphonia
3 a (72) Rhodomela (35) Phaeostrophion 4 b (68) Rhodomela (34) Cryptosiphonia
Rhodomela present as an understory species in the surfgrass bed before the experiment was initiated
b RhodomeIa absent before the experiment was initiated c The replicates 1-4 in this treatment correspond to replicates 1-4
in Fig. 9
Table 2. Abundant erect plant species in each replicate of surfgrass removal experiments from data taken in April 1981 at Boiler Bay. Numbers preceding each species name in parentheses are percent covers, epilithic and epiphytic covers combined
Treatment Repli- Species with Other abundant (date cate greatest%cover species succession # (>20% cover) initiated)
Fa11 1978 I a (17) Rhodomela none 2 a (61) Rhodomela none 3 a (66) Rhodomela (23) Phaeostrophion 4 b (39) Rhodomela (29) Odonthalia
Winter 1 b (82) Cryptosiphonia none 1979 2" (42) Rhodomela (32) polysiphonous
red algae 3 a (3) Phaeostrophion none 4 b (34) Cryptosiphonia (30) Phaeostrophion
Spring 1 (30) polysiphonous none 1979 r red algae
2 (41) Phaeostrophion none 3 (57) Phaeostrophion (53) Rhodomela
(23) Odonthalia 4 (49) Odonthalia (33) polysiphonons
red algae
Rhodomela present as an understory species in the surfgrass bed before the experiment was initiated
b Rhodomela absent before the experiment was initiated ~ Data on the initial presence or absence of Rhodomela in this
treatment are unavailable

62
were fairly similar to each other. Phaeostrophion invaded these plots the first winter and maintained a high cover (Fig. 1). Epiphytic Ulva had a high cover in some of them. In contrast, replicates of the other three treatments (ini- tiated in fall, winter, and spring) did not resemble each other (Table 1). In many plots early successional species maintained a high cover, but in others Iridaea flaccida, Cryptosiphonia, and Rhodomela established a high cover. At Boiler Bay the replicates initiated in fall 1978 resembled each other fairly closely (Table 2). Rhodornela was the most abundant species in all four. In the other two treatments (initiated in winter and spring 1979), however, replicates differed greatly. A variety of species were abundant, includ- ing Phaeostrophion, Cryptosiphonia, polysiphonous red al- gae, and Odonthalia.
One cause of this local variation may be small variations in the initial surfgrass understory. For example, the mean cover of Rhodomela by the end of the experiment, in the plots that had had some initial cover of Rhodomela was 38.7%, but the mean cover in the plots that had had no initial cover was 10.7% (see footnotes in Tables 1 and 2). This highly significant difference (P<0.005; Mann-Whit- ney U-test) may have been caused by the thin crustose hold- fast of Rhodornela, which is very difficult to remove com- pletely (D'Antonio 1982). Upright branches regrow from the holdfast. Apparently, scraping the crust away with a putty knife did not completely remove the holdfasts, and the plants regrew. This regrowth is crucial, because Rhodo- mela rarely recruits from spores (D'Antonio 1982).
Discussion
Two different types of disturbance may select for the differ- ent seasonal patterns of growth and recruitment by Ulva and Phaeostrophion documented in this study. Along the Oregon coast space becomes available at two times: "winter" storms (November-April) remove organisms (e.g., Paine and Levin 1981) and the first storm in the fall removes the sand which has accumulated in the summer (e.g., Mark- ham 1973). The distribution of Phaeostrophion (limited to sand-influenced areas), the ability of its blades to regrow if they are removed by sand scour, and its fall and winter recruitment suggest that it takes advantage of space made available by sand movement (Mathieson 1965). On the other hand, Ulva, a thinner, more fragile blade, appears to take advantage of space made available by wave removal of organisms later in the year. At Squaw Island and Boiler Bay both processes make space available, and both algae occur.
The other distinct seasonal pattern documented in this study, dramatic fall declines in algal cover, could have a variety of causes. In Oregon the changes occurring in the fall include shorter days (= reduced time for photosynthe- sis), lower temperatures (night low tides mean algae may experience freezing temperatures), larger waves, larger freshwater input from heavy rains, and perhaps greater sand scour due to increased water movement. Hansen (1977) believed that large waves and short days, which re- duced growth, correlated best with declines in the cover of Iridaea cordata. But L cordata loses its blades at the same time the blades lose their cuticle, making them more susceptible to herbivores (Gerwick and Lang 1977; Gaines 1983). Thus, fall declines in algae might be caused by several factors.
Regardless of its cause, this seasonal decline in algal cover plays a central role in the community. The deaths of Ulva individuals in the fall might allow Cryptosiphonia to replace them, and the deaths of Cryptosiphonia individ- uals might allow Rhodomela or even Phaeostrophion to re- place them. Connell and Slatyer's (1977) simple model pre- dicts species replacement by physical conditions, but it does not predict backwards succession, such as from Cryptosi- phonia back to Phaeostrophion.
Variation in the types of interspecific interaction in- creases the complexity of succession in this community. One example is the difference in the ability of Phaeostrophion to inhibit Ulva and diatoms. Diatoms were completely in- hibited, but Ulva was only slightly inhibited, because it grew epiphytically on Phaeostrophion. The result is puzzling, however, because filamentous diatoms do grow epiphyti- cally on other species. Perhaps the host-epiphyte interaction is fairly specific. Without further experiments, the precise interaction between Uh, a and Phaeostrophion is unclear. Phaeostrophion may not be greatly harmed by Ulva, because its blades become small and desiccated in the summer in any case. Conversely, epiphytic growth may allow Ulva to live in undisturbed areas (Fig. 1 ; D'Antonio 1982).
Both inhibition and facilitation occur in this succes- sional sequence. Surfgrass has barbed seeds (Gibbs 1902), which must recruit by attachment to algae with a specific morphology to avoid dislodgement. Seeds fit on to branched algae with a central axis 1 mm in diameter, such as Rhodomela, Odonthalia, Cryptosiphonia, and some erect coralline algae. Therefore, this step is classified as obligate facilitation (Turner 1983).
The influence of herbivores on succession as a whole is unknown, but they appear to have only a subtle effect on the early stages. Herbivore exclusions produced a signifi- cant increase in Ulva, but in some places accessible to herbi- vores Ulva attained almost 100% cover (spring 1979 remov- als, Squaw Island, Fig. 1). The reduced Ulva cover pro- duced by herbivores did not result in a significant increase in other algal species. Fall declines in algal cover occurred regardless of whether herbivores were present.
The successional sequence of the plants influences the herbivores. The increased limpet densities early in succes- sion might have two causes. (1) Early successional plants are generally more attractive to marine herbivores than late successional plants (Lubchenco and Gaines 1981). (2) Early successional plants are smaller and therefore less likely to crowd out herbivores (e.g., Southward and Southward 1978; Underwood and Jernakoff 1981). The slightly greater Tegula densities in the successional plots might result from their preference for algae over surfgrass (Best 1964).
Spatial variation among replicates might have several causes. One cause suggested by the data is an historical effect, the presence or absence of Rhodomela in the initial surfgrass bed. Two other possible causes are spatial differ- ences in recruitment magnitude and in surrounding organ- isms. Rhodomela, for instance, has variable recruitment. The ability to capture space by vegetative growth can am- plify chance differences, because one propagule might grow to occupy a large patch. The organisms surrounding a suc- cessional plot could influence the sequence in four ways: (1) they could provide a source of propagules; (2) they could invade the plot themselves by vegetative growth, (3) they could produce variability in conditions near the edge of the plot by shading, whiplash, or allelopathy, or (4) they

63
could harbor different species of herbivores. Although few algal dispersal patterns are known, some species disperse only a few meters (Anderson and North 1966; Dayton 1973; Paine 1979; Deysher and Norton 1982). If this pat- tern is common, differences in surrounding plants could produce high local variability (Horn 1981). In my experi- ments, variation might also have been produced by vegeta- tive growth of Rhodomela plants at the borders of some plots. An alternate hypothesis, that the replicates are not true replicates but differ in their abiotic characteristics or in their microscopic biotic characteristics, also merits con- sideration. This possibility is worrisome and suggests some generalizations may be suspect.
Despite the variety of middle successional stages, all the replicates will probably have the same last successional stage dominated by surfgrass. Surfgrass plants surrounding the plots slowly invade by horizontal vegetative growth (Turner 1982). Even if the plots were not surrounded, they would probably still become surfgrass beds, though much more slowly, as plants recruited from seeds. The only condi- tion that might preclude surfgrass invasion is a persistent monoculture of blade-like species, such as Phaeostrophion, that cannot facilitate surfgrass (Turner 1983). Although an indefinitely persisting monoculture of any species seems im- probable, Phaeostrophion dominated several experimental plots for 3 years and forms large monocultures at Strawber- ry Hill and Seal Rock, sand-influenced areas on the Oregon coast (pers. obs.).
Comparisons of low zone rocky intertidal communities in different areas provide further insights on succession. For example, two California workers, Sousa (1979) in Santa Barbara and Northcraft (1948) in Monterey, found that Ulva sp. colonized first throughout the year. Ulva may re- cruit in only one season in Oregon, because the physical environment there exhibits greater seasonal change. Day length, temperature, and wave action (B. Menge pers. com- mun.) vary less in Santa Barbara. Whether this difference represents a latitudinal difference or a difference between sites unrelated to latitude is not clear.
Sousa's (1979) results also differ from the present study in the role of herbivores. When Sousa excluded herbivores, Ulva persisted through the winter, and middle successional red algae did not invade. In the presence of herbivores, however, Ulva was removed, its inhibition broken, and red algae invaded. By contrast, in Oregon Ulva declines in the fall even in herbivore exclusions. Two explanations could account for this difference. First, Oregon appears to experi- ence greater changes in the physical environment that could remove Ulva. Second, the large herbivores most important in California, the crab Pachygrapsus crassipes and the sea hare Aplysia californiea, are absent from my study sites. In California, limpets temporarily reduced the abundance of Ulva but did not effect its replacement. Since limpets and other grazing molluscs were the main herbivores ex- cluded by my experiments, these animals appear to play similar roles in the two communities.
Although simple models describe some successional se- quences (e.g., Sousa 1979), seasonal variation, variation in interspecific interactions, and spatial variation are not un- usual features in natural communities. Examples of sea- sonal variation are legion (Keever 1950, 1979; Sutherland 1974; Foster 1975; Kain 1975; Neushul et al. 1976; Paine 1977; Emerson and Zedler 1978; Hay and South 1979; Sousa et al. 1981). In marine fouling communities abilities
of different species to invade space and to resist invasion vary greatly (Sutherland and Karlson 1977; Dean and Hurd 1980). Stochastic spatial variation in the intensity of recruit- ment influences both marine and terrestrial communities (e.g., Sutherland 1974, 1980; Robles and Cubit 1981; Greene and Schoener 1982; Hils and Vankat 1982). Even backwards succession, analogous to the replacement of Cryptosiphonia by Ulva, occurs in other communities (Drury and Nisbet 1973).
Historical effects may also occur in many communities, because most natural disturbances do not remove all parts of all organisms. Algae (e.g., Chondrus crispus; Lubchenco 1980) or simple colonial organisms like sponges may be able to recover from just a few cells, and terrestrial plants can resprout or grow from a seed bank, which may reflect the history of the site (Marks 1974; Harper 1977; Van Hulst 1980; Shugart et al. 1981). Surrounding organisms influ- ence succession in a fouling community (Kay and Keough 1981), where sponges rapidly invade newly available space adjacent to them. Thus, in many situations simple models can not predict successional change. This complexity could be incorporated into ecological theory by developing a vari- ety of models and applying each of them when they fit the natural history of the organisms.
Acknowledgments. This paper is a portion of a Ph.D. dissertation presented to the Zoology Department of Oregon State University. My advisors, Drs. B. Menge and J. Lubchenco, and my committee members, Drs. C. Bayne and P. McEvoy, helped me throughout my research. Another steadfast helper was J. Lucas, who provided advice, recorded data in the field, and drafted figures. All of my field helpers deserve thanks, especially J. Seger, C. Turner, and S. Turner. L. Ashkenas, C. D'Antonio, S. Gaines, J. Myers, and A. Olson have also helped by reading other versions of this paper. I am grateful as well to the United States Coast Guard for allowing me access to the Squaw Island site. S. Sargent typed the manu- script. I received financial support from Sigma Xi, the Scientific Research Society, the Zoology Department at Oregon State Uni- versity, National Science Foundation grants to Drs. B. Menge and J. Lubchenco, and a National Science Foundation doctoral im- provement grant.
Appendix. Species that colonized experimental plots but are not listed in text. Species designations follow Abbott and Hollenberg (1976). Unless noted, species occurred at both sites
Rhodophyta
Bossiella plumosa Callithamnion pikeanum a Ceramium sp. Corallina officinalis" Corallina vancouveriensis coralline crusts Cryptopleura violacea Endocladia muricata ~ Farlowia mollis Laurencia spectabilis Microcladia borealis Odonthalia lyalli ~ Odonthalia washingtoniensis b Plocamium cartilagineum Plocamium tenue Ptilota filicina b
Phaeophyta
Analipus japonicus Costaria costata b Egregia menziesii Fucus distichus" Haplogloia andersonii b Hedophyllum sessile Petalonia fascia Ralfsia sp. Scytosiphon sp.
Chlorophyta
Bryopsis sp. a Cladophora sp. Monostroma oxyspermum Rhizoclonium riparium a Spongomorpha sp. a

64
Animalia
Anthopleura elegantissirna Balanus cariosus Balanus glandula Bryozoa a Chthamalus dalli Mytilus sp. a
a occurred only at Squaw Island b occurred only at Boiler Bay
References Abbott IA, Hollenberg GJ (1976) Marine algae of California. Stan-
ford University Press, Stanford Anderson EK, North WJ (1966) In situ studies of spore production
and dispersal in the giant kelp Macrocystis. In: Young EG, McLachlan JL, (eds) Proc Int Seaweed Symposium 5, Halifax, 73-85, Pergamon Press, New York
Best B (1964) Feeding activities of Tegula funebralis. Veliger 6 (Suppl): 42-45
Clements FE (1916) Plant succession: An analysis of the develop- ment of vegetation. Carnegie Inst Wash Publ 242:1-512
Connell JH, Slatyer RO (1977) Mechanisms of succession in natu- ral communities and their role in community stability and orga- nization. Am Nat 111:1119-1144
D'Antonio CM (1982) Population and community ecology of the red alga, Rhodomela larix (Turner) C. Agardh, on the central Oregon coast. MS Thesis, Oregon State University, Corvallis, Oregon
Dayton PK (1973) Dispersion, dispersal, and persistence of the annual intertidal alga, Postelsia palmaeformis Ruprecht. Ecolo- gy 54:433-438
Dean TA, Hurd LE (1980) Development in an estuarine fouling community: The influence of early colonists on later arrivals. Oecologia (Berl) 46: 295-301
Deysher L, Norton TA (1982) Dispersal and colonization in Sar- gassum muticum (Yendo) Fensholt. J Exp Mar Biol Ecol 56:179-195
Drury WH, Nisbet ICT (1973) Succession. J Arnold Arbor 54:331-368
Emerson SE, Zedler JB (1978) Recolonization of intertidal algae: An experimental study. Mar Biol 44:315-324
Foster MS (1975) Algal succession in a Macrocystispyrifera forest. Mar Biol 32:313-329
Foster MS (1982) Factors controlling the intertidal zonation of Iridaeaflaccida. J Phycot 18 : 285-294
Gaines SD (1983) Herbivory and between-habitat diversity: The differential effectiveness of a marine plant defense. Ecology in press
Gerwick WH, Lang NJ (1977) Structural, chemical, and ecological studies on iridescence in Iridaea (Rhodophyta). J Phycol 13:121-127
Gibbs RE (1902) Phyllospadix as a beach-builder. Am Nat 36 : 101-109
Green CH, Schoener A (1982) Succession on marine hard sub- strata: A fixed lottery. Oecologia (Berl) 55:289-297
Hansen JE (1977) Ecology and natural history of Iridaea cordata (Gigartinales, Rhodophyta) growth. J Phycol ~ 3:395-402
Hansen JE, Doyle WT (1976) Ecology and natural history of Iri- daea cordata (Rhodophyta: Gigartinaceae): Population struc- ture. J Phycol 12:273-278
Harper JL (1977) Population biology of plants. Academic Press, New York
Hay CH, South GR (1979) Experimental ecology with particular reference to proposed commercial harvesting of Durvillaea (Phaeophyta, Durvilleales) in New Zealand. Bot Mar 22:431-436
Hils MH, Vankat JL (1982) Species removals from a first-year old-field plant community. Ecology 63:705-711
Horn HS (1981) Some causes of variety in patterns of secondary succession. In: West DC, Shugart HH, Botkin DB (eds) Forest succession, 24-35, Springer Verlag, New York
Hruby T (1975) Seasonal changes in two algal populations from the coastal waters of Washington state. J Ecol 63 : 881-890
Kain JM (1975) Algal recolonization of some cleared subtidal ar- eas. J Ecol 63:739-765
Kay AM, Keough MJ (1981) Occupation of patches in the epi- faunal communities on pier pilings and the bivalve Pinna bicolor at Edithburgh, South Australia. Oecologia (Berl) 48:123-130
Keever C (1950) Causes of succession on old fields of the Piedmont, North Carolina. Ecol Monogr 20:229~50
Keever C (1979) Mechanisms of plant succession on old fields of Lancaster County, Pennsylvania. Bull Torrey Bot Club 106:299-308
Littler MM, Littler DS (1980) The evolution of thallus form and survival strategies in benthic marine macroalgae: Field and lab- oratory tests of a functional form model. Am Nat 116:25-44
Lubchenco J (1980) Algal zonation in the New England rocky intertidal community: An experimental analysis. Ecology 61 : 333-344
Lubchenco J, Gaines SD (1981) A unified approach to marine- plant herbivore interactions. I. Populations and communities. Ann Rev Ecol Syst 12:405-437
Lubchenco J, Menge BA (1978) Community development and per- sistence in a low rocky intertidal zone. Ecol Monogr 48 : 67-94
Markham JW (1973) Observations on the ecology of Laminaria sinclairii on three northern Oregon beaches. J Phycol 9:336.341
Marks PL (1974) The role of pin cherry (Prunus pensylvanica L.) in the maintenance of stability in northern hardwood ecosys- tems. Ecol Monogr 44 : 73-88
Mathieson AC (1965) Contributions to the life history and ecology of the marine brown alga Phaeostrophion irregulare S. et G. Ph.D. dissertation, University of British Columbia, Vancouver, Canada
Menge BA (1976) Organization of the New England rocky intertid- al community: Role of predation, competition and environmen- tal hetrogeneity. Ecol Monogr 46:355-393
Neushul M, Foster MS, Coon DA, Woessner JW, Harger BWW (1976) An in situ study of recruitment, growth and survival of subtidal marine algae: techniques and preliminary results. J Phycol 12: 397-408
Northcraft RD (1948) Marine algal colonization on the Monterey Peninsula, California. Am J Bot 35 : 396-404
Paine RT (1977) Controlled manipulations in the marine intertidal zone, and their contributions to ecological theory. In: The changing scenes in natural sciences 1776-1976. Academy of Natural Sciences, Special Pnbl 12:245 270
Paine RT (1979) Disaster, catastrophe, and local persistence of the sea palm Postelsia palmaeformis. Science 205 : 685-687
Paine RT, Levin SA (1981) Intertidal landscapes: disturbance and the dynamics of pattern. Ecol Monogr 51:145-178
Robles CD, Cubit J (1981) Influence of biotic factors in an upper intertidal community: Dipteran larvae grazing on algae. Ecolo- gy 62:1536-1547
Shugart HH, West DC, Emanuel WR (1981) Patterns and dynam- ics of forests: An application of simulation models. In: West DC, Shugart HH, Botkin DB (eds) Forest succession, 74-94, Springer Verlag, New York
Slocmn CJ (1980) Differential susceptibility to grazers in two phases of an intertidal alga: Advantages of heteromorphic gen- erations. J Exp Mar Biol Ecol 46:99-110
Sokal RR, Rohlf FJ (1969) Biometry. WH Freeman and Co, San Francisco
Sousa WP (1979) Experimental investigations of disturbance and ecological succession in a rocky intertidal algal community. Ecol Mongr 49 : 227254
Sousa WP, Schroeter SC, Gaines SD (1981) Latitudinal variation in intertidal algal community structure: The influence of graz- ing and vegetative propagation. Oecologia (Berl) 48 : 297-307
Southward AJ, Southward ED (1978) Recolonization of rocky

65
shores in Cornwall after use of toxic dispersants to clean up the Torrey Canyon spill. J Fish Res Bd Can 35:682-706
Sutherland JP (1974) Multiple stable points in natural communi- ties. Am Nat 108:859-873
Sutherland JP (1980) Dynamics of the epibenthic community on roots of the mangrove Rhizophora mangle, at Bahia de Buche, Venezuela. Mar Biol 58 : 75-84
Sutherland JP, Karlson RH (1977) Development and stability of the fouling community at Beaufort, North Carolina. Ecol Monogr 47 : 425-446
Turner T (1982) Community organization and succession in rocky intertidal surfgrass beds. Ph.D. dissertation, Oregon State Uni- versity, Corvallis, Oregon
Turner T (1983) Facilitation as a successional mechanism in a rocky intertidal community. Am Nat 121:729-738
Underwood A J, Jernakoff P (1981) Effects of interactions between algae and grazing gastropods on the structure of a low-shore intertidal algal community. Oecologia (Berl) 48:221 233
Van Hulst R (1980) Vegetation dynamics or ecosystem dynamics: Dynamic sufficiency in succession theory. Vegetatio 43 : 147-151
Received December 20, 1982