complement evasion by human pathogens - lambris · surface way, the complement cascade also...
TRANSCRIPT

Evading the manifold attacks of the immune system is a key determinant for the survival of pathogens within their hosts. It is not surprising, therefore, that the coex-istence and co-evolution of humans and microorganisms has produced a multitude of microbial mechanisms for attenuating or escaping these attacks. As a first line of defence against pathogenic intruders, and a mediator between the innate and adaptive immune responses, the complement system is a particular focus of these evasion strategies. Although this carefully regulated cascade of enzymes, protein complexes and receptors ensures the rapid recognition and elimination of foreign struc-tures, it also offers many sites of interference that can disrupt this balanced network of protein interactions. A detailed understanding of the individual processes and the underlying interactions on a molecular level is essential for describing the mechanisms of infectious diseases and the development of new therapies. Recent discoveries of complement-targeting proteins, the avail-ability of complete microbial genome sequences and advances in experimental methods have propelled this area of research, and provide fascinating insights into complement attack and evasion. Many pathogens seem to have developed parallel routes for escaping comple-ment, and several evasion principles are shared not only among members of the same genus but even among diverse organisms, such as bacteria, viruses, fungi and parasites.
In this Review, we will provide a comprehensive over-view and update of the exciting recent developments in this field. After a short introduction that will discuss the diverse role of the complement system in defence, dis-ease and infection, the emphasis will be on the functional
and structural aspects of the evasion strategies of human pathogens. Rather than separating them by organism, we classify distinct and common mechanisms for all patho-gens based on their mode of action. In light of recent findings, the unique evasion strategies of Staphylococcus aureus will be analysed in more detail. Finally, the potential impact of these developments on prospective antimicrobial and complement-specific therapeutics will be discussed.
The human complement systemThe complement system is a central component of the innate immune response and fulfils numerous func-tions, including the recognition of foreign cells, com-munication with and activation of adaptive immunity and the removal of cellular debris (reviewed in REFS 1–5). Complement consists of a well-balanced network of circulating and cell-surface-bound proteins, which serve as substrates, enzymes or modulators of a hier-archical series of extracellular proteolytic cascades. There are three established mechanisms of comple-ment activation; these are known as the classical, lectin and alternative pathways (FIG. 1a). The initial steps that trigger these activation processes differ considerably. The classical pathway is stimulated by the recogni-tion of antigen–antibody complexes on foreign-cell surfaces by the hexameric complement component C1q. Structurally similar pattern-recognition receptors, mannose-binding lectin (MBL) and ficolins, bind to carbohydrate ligands on microbial intruders and initiate the lectin pathway. Conversely, the alternative pathway is stimulated by the spontaneous hydrolysis of native C3 or the presence of foreign surface structures
*Department of Pathology and Laboratory Medicine, University of Pennsylvania School of Medicine, 422 Curie Boulevard, Philadelphia, Pennsylvania 19104, USA. ‡Division of Cell Biology and Biophysics, School of Biological Sciences, University of Missouri-Kansas City, 5100 Rockhill Road, Kansas City, Missouri 64110, USA. Correspondence to J.D.L. e-mail: [email protected]:10.1038/nrmicro1824
Pattern-recognition receptorA highly diverse group of soluble and surface-bound proteins that can detect specific molecular surface structures. These receptors are important for discriminating between self and non-self cells (for example, microorganisms) and are, therefore, found in various pathways of the immune system. Prominent examples in the complement system are C1q and mannose-binding lectin.
Complement evasion by human pathogensJohn D. Lambris*, Daniel Ricklin* and Brian V. Geisbrecht‡
Abstract | The human immune system has developed an elaborate network of cascades for dealing with microbial intruders. Owing to its ability to rapidly recognize and eliminate microorganisms, the complement system is an essential and efficient component of this machinery. However, many pathogenic organisms have found ways to escape the attack of complement through a range of different mechanisms. Recent discoveries in this field have provided important insights into these processes on a molecular level. These vital developments could augment our knowledge of the pathology and treatment of infectious and inflammatory diseases.
R E V I E W S
132 | FEBRUARY 2008 | VOLUME 6 www.nature.com/reviews/micro
©!2008!Nature Publishing Group!

C3aR
C3
C3a
C3bC4b
CRC5aR
C5C5a
C4C4a
C1q
Host cell
Nature Reviews | Microbiology
RCAmimicry
fI
C3b
C3aR
Cell lysis
Amplification
Opsonization
C3C3a
AP C3convertase
CP and AP C5convertases
CP and AP C5convertases
MAC
Lectinpathway
Classicalpathway
Alternative pathway
Initiation
CP C3convertase
AP C3convertase
CP C3convertase
C3b C3b iC3bC4b
CR
C5aR
AntigensCarbohydrates
Initial AP C3convertase
Initial AP C3convertase
Signalling
Antibodyinactivation
C5aRantagonism
C1 inhibition
C1-INH
RCA recruitment C3b degradation
MAC inhibition
iC3b
C3 inhibition
C5 inhibition
RCA recruitment and mimicryConvertase decay
ConvertaseinhibitionC2
inhibition
IncreasingC3b densityfB or fD
fB or fD
a Activation
b Evasion
Pathogen cell
Pathogen cell
RCA
C5b C6–C9
C2
C5C5a
C4C4a
P
P
C1qMBL or F
MBL or F
Host cellChemotaxisInflammation
Phagocytosis
Cleavage
Inhibition
Capturing
C1r/C1s(CP)
MASP (LP)
(FIG. 1a). Recent findings suggest that additional proc-esses, such as the C2-bypass6 and extrinsic protease7 pathways or properdin-mediated direct convertase assembly on microbial surfaces8, can also initiate complement activation.
All of the complement cascades culminate in the central cleavage of C3 and the generation of its active fragments C3a and C3b (FIG. 1a). Opsonization of foreign surfaces by covalently attached C3b fulfils three major functions: cell clearance by phagocytosis;
Figure 1 | Activation and evasion of complement. a | After activation of the complement system by antibody complexes (classical pathway (CP)), terminal mannose (lectin pathway (LP)) or by spontaneous and induced C3 hydrolysis (alternative pathway (AP)), the C3 convertases cleave C3 to its active fragments C3a and C3b. Covalent binding of C3b (opsonization) amplifies the cascade and mediates phagocytosis and adaptive immune responses by binding to complement receptors (CRs). Accumulation of deposited C3b also leads to the assembly of C5 convertases that activate C5 to C5a and C5b. Whereas C5b initiates the formation of the lytic membrane-attack complex (MAC), the anaphylatoxins C3a and C5a induce pro-inflammatory and chemotactic responses by binding to their receptors (C3aR and C5aR). On pathogenic surfaces, properdin (P) induces and stabilizes the AP C3 convertase, which leads to enhanced complement activity. b | Microorganisms have developed many ways to evade complement actions. Suppression of CP activation can be achieved by trapping endogenous C1 inhibitor (C1-INH) to the surface or by inactivating antibodies through the capture of their Fc regions. Whereas the recruitment of soluble regulators by capturing host proteins is a common strategy to impair downstream complement actions, certain viruses also produce structural mimics of these regulators. In addition, some microbial proteins have similar activities to CD59 in preventing MAC formation. Direct inhibition of C3, the C3 and C5 convertases, C5 or the C5a receptor (C5aR) is a prominent strategy of Staphylococcus aureus. Finally, a set of different microbial proteases can degrade many of the crucial components of the complement system. These proteases act directly or by capturing and activating a human protease. An extended list of complement evasion proteins can be found in Supplementary information S1 (table). Increased and decreased activity is represented by thick and thin arrows, respectively. F, ficolin; fB, factor B; fD, factor D; fI, factor I, MASP, MBL-associated serine protease; MBL, mannose-binding lectin; RCA, regulators of complement activation.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | FEBRUARY 2008 | 133
©!2008!Nature Publishing Group!

AnaphylatoxinA small protein fragment (approximately 10 kDa) that is generated during the activation of complement components. C3a and C5a trigger a range of inflammatory and immune-stimulating responses by binding to their receptors (C3aR and C5aR) on various effector cells. The chemotactic activity of C5a is 100-fold higher than that of C3a; no such activities or receptors have so far been described for C4a.
Toll-like receptorA pattern-recognition receptor that recognizes a range of surface structures on pathogens (for example, proteoglycans and lipopolysaccharides). Toll-like receptors are expressed on most immune cells and their activation and signalling induces numerous inflammatory, innate and adaptive immune responses.
Short consensus repeatThe structural building block of many complement regulators and receptors (for example, factor H and CR1). These -sheet-rich domains, which are
composed of approximately 60 residues, are also found in viral complement-evasion factors and other proteins (for example, selectins, clotting factor XIII B and GABA receptors).
amplification of complement activation by the forma-tion of a surface-bound C3 convertase; and assembly of the C5 convertases. Cleavage of C5 induces the forma-tion of a multiprotein pore complex (the membrane-attack complex (MAC)), which leads to cell lysis. Both the covalent attachment of C3b and the stabilization of the C3 convertase by the complement regulator properdin are greatly encouraged by hydroxyl-rich pathogen surfaces. A series of complement receptors mediate the recognition of opsonized cells by leuko-cytes, which results in phagocytosis and the stimula-tion of the adaptive immune system (for example, by B and T cells). Finally, the anaphylatoxins C3a and C5a are released during complement activation and trigger a range of chemotactic and pro-inflammatory responses (for example, the recruitment of inflammatory cells and an increase in microvasculature permeability). In this way, the complement cascade also supports and pro-motes the function of downstream mechanisms of the immune response.
Excessive complement activation on self tissue has severe effects and can lead to the development of various diseases2,9,10. In addition to a location- and time-based restriction to immediate sites of activation, a finely tuned set of soluble and membrane-bound regulators ensure that any action of complement on host cells is either prevented or actively inhibited. These structurally similar regula-tors of complement activation (RCA) comprise comple-ment receptor 1 (CR1), factor H, factor H-like protein-1 (FHL-1), C4-binding protein (C4BP), decay-accelerating factor (DAF) and membrane cofactor protein (MCP). RCA either destabilize the C3 convertase (decay-accelera-tion activity; carried out by CR1, factor H, FHL-1, C4BP and DAF) or promote the factor-I-mediated degradation of C3b to iC3b (cofactor activity; carried out by CR1, fac-tor H, C4BP, FHL-1 and MCP). CR1 also acts as a cofactor for an additional factor-I cleavage of iC3b to C3d. A struc-turally unrelated regulator protein, CD59, prevents forma-tion of the MAC. Finally, the glycoprotein C1-inhibitor controls the activity of several enzymes that are involved in the first steps of the classical and lectin pathways (C1r, C1s and MBL-associated serine protease).
This arsenal of proteases, active components and regulators is not only essential for the efficient elimina-tion of microbial intruders, but also modulates the func-tions of the adaptive immune system11. Complement also seems to be able to communicate with other important cascades and networks in the human body. Such connec-tions have, for example, been shown for the coagulation system7. Furthermore, recent data suggest that crosstalk between complement and Toll-like receptors might have synergistic effects in the immune response to infec-tions12,13. Despite all of its benefits, however, the many proteins that are involved in this powerful network also provide pathogenic organisms with numerous locations where anti-complement strategies can be executed.
Mechanisms of complement evasionThe ability to escape the elaborate machinery of the human immune system is a key determinant in the virulence of pathogens. In fact, immune-evasion strategies are often
focused on the complement system, which is a centre-piece of innate immunity and is generally regarded as the first line of defence against pathogenic microorganisms. Our knowledge of how these escape mechanisms func-tion on a molecular level has increased remarkably in recent years. Identification of the individual pathogenic proteins and their human targets has been one crucial step in this task and the list of complement-targeting proteins is constantly growing. Despite this plethora of complement-binding proteins (Supplementary infor-mation S1 (table)), their mechanisms of action can be condensed to a few successful strategies: the recruitment or mimicking of complement regulators; the modulation or inhibition of complement proteins by direct interac-tions; and inactivation by enzymatic degradation (FIG. 1b; TABLE 1). In addition to these strategies, many microor-ganisms also possess passive evasive features — a promi-nent example is the cell wall of Gram-positive bacteria, which prevents lysis by the MAC14. The following sec-tions focus on the active complement-evasion strategies and illustrate their common and unique features.
Making use of the host’s arsenal — acquiring regulators. The presence of RCA on cell surfaces is a key deter-minant for complement function on self and non-self cells. Although most pathogens do not express RCA on their surfaces, they have found ways to adapt by stably binding the RCA that circulate in human plasma15 (FIG. 1b; TABLE 1). In fact, of all the mecha-nisms of complement evasion that are used by various pathogens, trapping RCA is by far the most widely disseminated strategy for avoiding the complement response. RCA recruitment is common in bacteria16 (for example, Escherichia coli, Borrelia burgdorferi and streptococci), but has also been described for viruses17 (for example, HIV-1), fungi18,19 (for example, Candida albicans) and parasites20 (for example, Echinococcus spp.) (Supplementary information S1 (table)). The recruitment strategy has important advantages: RCA are the natural regulators of complement and are evo-lutionarily tuned to fulfil their functions; they are pro-duced by the host and are available in relatively high concentrations; and they share common structural features, called short consensus repeat (SCR) domains, that allow the same pathogen-derived binding protein to recruit different host RCA.
Although they are less potent than the cell-surface-bound CR1, the availability of C4BP, factor H and FHL-1 as soluble proteins means that these proteins are also primary targets. All three regulators show decay acceleration and cofactor activity. Owing to their size and multivalency, the captured regulators do not usually lose their overall activity. In many cases, the recruit-ment function can be attributed to a family of similar proteins in a bacterial genus. Typical examples are the complement-regulator-acquiring proteins in Borrelia spp., porins in Neisseria spp. and the M protein family in Streptococcus spp.16 The recruitment of regulators is not restricted to soluble RCA, however. The in vivo transfer of membrane-bound regulators (MCP, DAF and CD59) between human cells has been reported21,
R E V I E W S
134 | FEBRUARY 2008 | VOLUME 6 www.nature.com/reviews/micro
©!2008!Nature Publishing Group!

and the acquisition of CD59 by this mechanism has been suggested for E. coli22 and Helicobacter pylori23. Finally, some viruses camouflage themselves with RCA-containing host membrane during budding or stimulate RCA expression on the host cell24.
Certain viruses have found ways to produce soluble proteins that closely mimic the structure and function of host regulators (FIG. 1b; TABLE 1). The orthopoxvi-ruses variola (smallpox) and vaccinia express comple-ment-inhibitory proteins25,26 that consist of four SCR domains. These modules are the common building blocks and functional entities of all human RCA, and are also found in complement receptors (for example, CR2) and factors (for example, factor B). Structural analysis of the complete vaccinia virus complement-control protein (VCP)27 (FIG. 2a) clearly demonstrated its similarity to human RCA, such as DAF (FIG. 2b). Despite their small size compared with endogenous factor H (20 SCRs) and CR1 (30 SCRs), both VCP and the smallpox inhibitor of complement enzymes (SPICE) show decay acceleration as well as cofactor activity. Although these proteins vary in only 11 residues, the resulting changes in the electrostatic surface potential seem to have dra-matic effects on their regulatory potency; SPICE had a 1000-fold higher inhibitory activity for the alternative pathway than VCP28. Similar RCA-like proteins have also been described for monkeypox29 and cowpox30, two viruses that can infect humans. Interestingly, the monkeypox protein MOPICE (monkeypox inhibitor of complement proteins) contains only three SCR units and lacks decay-accelerating activity. Although a mem-brane-bound VCP homologue has been reported in
vaccinia virus (B5R)31, no complement-based functions could be detected for this protein (J.D.L., unpublished observations). Recent discoveries show that the same class of molecule is also produced in herpes viruses. A type 1 membrane protein that has four SCR repeats and dual complement-regulation activity has been isolated from Kaposi’s sarcoma-associated herpesvirus (HHV-8) and has been named Kaposica32.
The structural correspondence between these regu-lators and host RCA (FIG. 2) might reflect their ancient human origins, as the hijacking and implementation of sequences that encode beneficial host proteins has long been recognized as a hallmark of viral biology and evolution. However, viruses might not be the only microorganisms that follow this strategy. For example, complement-regulatory proteins have been described for various Ixodes spp. ticks. Both the Ixodes scapularis anti-complement protein (ISAC) and its counterpart from Ixodes ricinus (IRAC) exert decay-acceleration activity on the C3 convertases. However, any structural similari-ties between these proteins and host RCA has yet to be confirmed33,34.
Cutting through complement: pathogenic proteases. The degradation of complement components into smaller, non-functional fragments is the primary task of complement-active proteases. This class of eva-sion proteins has been reported almost exclusively in bacteria and targets a wide range of substrates. For example, the degradation of immunoglobulins (Igs) and C1q (for example, by Pseudomonas elastase (PaE) and alkaline protease (PaAP)) prevents activation
Table 1 | Examples of complement evasion proteins and their targets on host cells*
Complement evasion protein Host target
Antibody depletion
Staphylococcal protein A (SpA) IgG
Complement inhibition
Extracellular fibrinogen-binding protein (Efb) C3 and C3b-containing convertases
Staphylococcal superantigen-like protein-7 (SSL-7) C5
Staphylococcus complement inhibitor (SCIN) C3 convertases
Complement C2 receptor trispanning protein (CRIT) C2
Chemotaxis inhibitory protein of Staphylococcus aureus (CHIPS)
C5a receptor (C5aR)
Regulators of complement activation (RCA) recruitment
Complement-regulator-acquiring protein (CRASP)
Factor H, factor H-like protein-1 (FHL-1) and C4-binding protein (C4BP)
M protein family Factor H, FHL-1 and C4BP
RCA mimicry
Variola virus complement-control protein (VCP) C3b and C3 convertases
Smallpox protein of complement enzymes (SPICE) C3b and C3 convertases
Proteolytic degradation
Staphylokinase C3b and IgG (by activation of plasmin)
Pseudomonas elastase (PaE) C3
56 kDa protease C5a*See Supplementary information S1 (table) for an extended list of microbial evasion proteins.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | FEBRUARY 2008 | 135
©!2008!Nature Publishing Group!

Nature Reviews | Microbiology
a
bSCR1 SCR2 SCR3 SCR4
SCR1 SCR2 SCR3 SCR4
by the classical pathway35, whereas cleavage of C5a (by the 56 kDa protease from Serratia marcescens36 or the C5a peptidases from streptococci37) efficiently disa-bles C5a-mediated pro-inflammatory and chemotactic signalling. Streptococcal pyrogenic exotoxin B was found to degrade the complement regulator proper-din, which subsequently loses its ability to stabilize the convertases on pathogenic surfaces38. Inactivation of C3 by cleavage into non-functional fragments has been described for Pseudomonas spp. proteases (PaE and PaAP) and Porphyromonas spp. proteases (PrtH)35. A rare example of non-bacterial proteases that have complement activity has been observed in the para-sitic worm Schistosoma mansoni, which degrades iC3b using a 28-kDa protease, resulting in impaired binding to CR3 (REF. 39).
The acquisition and activation of host proteases is a more recently described alternative to the enzy-matic attenuation of complement. For example, the conversion of endogenous plasminogen into plasmin on bacterial surfaces can lead to the degradation of immunoglobulin G (IgG) and C3 fragments. Although a range of bacteria express plasminogen activators and/or receptors40, this mechanism is best described for S. aureus (discussed below)41.
Direct interventions — microbial complement inhibitors. A series of highly specific protein–protein interactions is the driving force behind the sophisticated activation and regulatory mechanisms of the complement system. Any interruption of the individual binding events can break this delicate balance and destabilize comple-ment efficiency. It is surprising, therefore, that only a
few direct complement inhibitors have been identified so far; most of these have been discovered in S. aureus and their molecular mechanisms are more thoroughly discussed in the following section. The CD59-like pro-tein from B. burgdorferi is one of the few examples of a direct complement inhibitor in other bacteria. This membrane-bound protein, which has affinity for C9 and C8b, is able to inhibit MAC formation. Similarly, streptococcal inhibitor of complement (SIC) has been found to prevent MAC formation by interfering with the C5b–C7 and C5b–C8 complexes. These functions of SIC seem to be paradoxical, as streptococci are already resistant to MAC-mediated cytolysis owing to their cell-wall structure14,42. This seemingly makes SIC ineffective, or at least redundant, in this respect. Although additional non-complement function-alities have been attributed to SIC, the reason for its MAC-inhibitory potential remains unsolved.
In herpes viruses, anti-complement activities have been attributed to several glycoproteins. The most ver-satile examples are the transmembrane gC1 and gC2 from herpes simplex virus type 1 and 2, respectively. Both glycoproteins were found to bind C3b and specifi-cally accelerate the decay of the alternative pathway C3 convertase43. Although gC2 has a tenfold stronger affin-ity for C3b44, only gC1 can also inhibit the interaction of C3b with C5 and properdin45. Other glycoproteins of the herpes virus family have Fc-receptor properties and can deplete antibody recognition and the activation of the classical pathway43,46. Ig-inactivating proteins have also been identified in bacteria, such as S. aureus and group G streptococci35.
Certain human parasites also express direct com-plement inhibitors. The complement C2 receptor trispanning protein has been detected in species of Schistosoma and Trypanosoma. This protein disrupts the interaction between C2 and C4 and, therefore, prevents formation of the classical pathway C3 con-vertase47. Furthermore, the Schistosoma spp. protein paramyosin prevents MAC formation by binding C8 and C9 (REF. 48), and has other immunomodulatory activities because of its binding affinity for C1q and IgG49. Finally, soft ticks of the genus Ornithodoros express at least one protein that keeps complement activation in check during their blood meal. The 17 kDa Ornithodoros moubata complement inhibitor has been shown to directly bind to C5 and prevent its cleavage, probably by inhibiting substrate binding to the C5 convertase50,51.
Finding the right mixture of activation and inhibition. Although complement inhibition and regulation is the predominant mechanism that is used to evade host immune defence by most pathogens, there is increas-ing evidence that some microorganisms actively induce complement activity for their own purposes. One of the most intriguing examples of fine-tuning antipodal actions that is used to ‘outwit’ host immune defence is that accomplished by HIV-1 — although this virus takes advantage of opsonization to enter human cells through complement receptors (BOX 1),
Figure 2 | Molecular mimicry of human complement regulators by viruses. Certain orthopox viruses and herpes viruses express proteins that closely mimic the structure and function of human regulators of complement activation. This structural similarity is illustrated for vaccinia virus complement-control protein (VCP; Protein Data Bank (PDB) code: 1G40 (REF. 27)) (a) and human decay-accelerating factor (DAF; PDB code: 1OJV102) (b), both of which have four short consensus repeat (SCR) domains. These SCRs (or complement control protein modules) are common structural motifs in complement components (for example, complement receptors and regulators) and other proteins (such as selectins).
R E V I E W S
136 | FEBRUARY 2008 | VOLUME 6 www.nature.com/reviews/micro
©!2008!Nature Publishing Group!

CR3CR2
C1q or mannose-binding lectin
Nature Reviews | Microbiology
RCA
Host cell
Cell entry Cell entry
EBV HIV-1
Complementactivation
Opsonization
RCA recruitment
Degradation
C3b
gp41gp120
gp350/220
iC3b
the terminal actions, such as MAC formation and phago-cytosis, must be avoided. Strikingly, the same glycopro-teins that activate complement through the classical and lectin pathways (gp41 and gp120, respectively) are also responsible for recruiting the regulators factor H and CD59 to the surface of the virus. In addition to inhibition of MAC formation by CD59, the degradation of C3b by factor I and factor H reduces amplification. However, the presence of iC3b still allows HIV-1 to hijack complement receptors to enter immune cells (BOX 1).
The diversity of the complement-targeting proteins (proteases, inhibitors and regulators) suggests that cer-tain bacteria might have developed a similarly elaborate
network of balanced complement activation and inhibi-tion. In this respect, S. aureus produces a nearly unparal-leled ensemble of immune-evasive molecules that target the complement system.
S. aureus — a master of complement evasionS. aureus is a prototypic opportunistic pathogen and a leading cause of nosocomial and community-acquired infections52. Perhaps more than any other bacterial path-ogen, S. aureus has evolved the ability to adapt to, and persist within, the diverse physiological microenviron-ments of its host (for example, skin and bones). Recent studies on this versatile bacterium have uncovered a formidable arsenal of virulence-facilitating proteins and structures that contribute to its pathogenesis.
Whereas the impact of S. aureus cell-surface-retained adhesins on host invasion and colonization has been firmly established53,54, the identities and functions of this bacterium’s immune-evading molecules have emerged only recently (reviewed in REFS 35,55–57). Several studies have revealed new mechanisms of immune evasion and modulation, and S. aureus has developed into a model system to study the fascinating interplay of host–pathogen interactions. Some strains synthe-size capsular polysaccharides58 that have been shown to impede antibody recognition59,60 and interfere with opsonization and C3b recognition by its receptors61. As the peptidoglycan-rich structure of its Gram-positive cell wall renders the bacterium resistant to MAC forma-tion42,55, most of the active evasion processes of S. aureus target the initial, amplifying and pro-inflammatory steps of complement activation.
Prevention of complement initiation. There are several mechanisms through which S. aureus impairs initiation of the classical pathway. Among these, the structure and function of staphylococcal protein A (SpA) (FIG. 3a) has been characterized most extensively. This surface-bound protein recognizes the Fc domain of Igs with high affinity62,63, which results in inverted tagging and block-age of the C1q (and Fc receptor) binding sites. It has been shown that SpA can also bind to the C1q receptor gC1qR/p33, which is highly expressed on the surface of activated platelets and endothelial cells64. However, this mechanism might be more closely associated with cell adhesion, through bridging to fibrinogen, than immune evasion65. Combined with the recent discovery that SpA can bind to von Willebrand factor and tumour-necrosis factor receptor 1 (REFS 66,67), these findings could reflect a far more versatile role for SpA than was initially real-ized. Interestingly, a second, less-characterized protein that has Ig-binding properties, called Sbi (S. aureus IgG-binding protein), has been described68. Whether Sbi shares the same physiological versatility as SpA remains to be established.
Distinct use of proteolytic enzymes. Even for pro-teases, S. aureus has developed an indirect but highly efficient way of counteracting complement. Although staphylokinase is not an enzyme itself, the forma-tion of a complex with plasminogen converts this
Box 1 | Hitchhiking with complement to enter host cells
Many human pathogens increase their virulence by avoiding confrontation with complement components or inactivating them through inhibition and degradation. However, a small group of pathogens, predominantly viruses, follow a completely different strategy by using complement receptors (CR) as an entry port into host cells. The general principle of binding to human target receptors using surface proteins, with subsequent internalization, is common to most viruses89, yet only a few target complement proteins3,17. For example, Epstein–Barr virus (EBV) expresses the glycoprotein gp350/220, which specifically interacts with CR2 on B cells and immature T cells (see the figure, left), ultimately causing infectious mononucleosis (Pfeiffer’s disease)90,91. In addition, surface-bound complement regulators serve as targets for the measles virus (haemagglutinin binds to membrane cofactor protein (MCP))92, human herpesvirus 6 (glycoprotein H binds to MCP)93 and some enteroviruses (coxsackievirus and echovirus bind to decay-accelerating factor (DAF))17. Similarly, surface-bound complement regulators, such as DAF and MCP, are recognized by other microbial proteins (for example, the Dr adhesin of Escherichia coli94) and have important roles in the infectivity and colonization of these pathogens.
Even without expressing complement-specific surface proteins, pathogens have found intriguing ways to use the hitchhiking principle — they voluntarily activate complement to become opsonized and take advantage of the deposited C3 fragments to interact with complement receptors on human cells. This formidable strategy has been observed in viruses (for example, HIV-1) (see the figure, right)3,95 and Mycobacterium tuberculosis, both of which use CR3 (REF. 96). However, the destructive potential of complement requires that the pathogen keeps complement activation in check by other mechanisms, such as the recruitment of host regulators of complement activity (RCA). Specific blockage of these activation and attachment steps could be a promising approach in the development of antimicrobial therapies.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | FEBRUARY 2008 | 137
©!2008!Nature Publishing Group!

Nature Reviews | Microbiology
a
b
SpA(B module)
Target: IgGsFunction: Attenuates classical
pathway
Efb(Carboxyl terminus)
SCIN
C3 (C3d domain)Inhibits C3 activation
C3 convertasesInhibits C3 activation
SSL-7 CHIPS
C5a receptorInhibits C5a signalling
Target: C5Function: Inhibits C5 activation
Human C5a
C5a receptorsReceptor activation
-defensinA cationic, cyclic, cysteine-rich peptide of 15–20 amino acids that belongs to a family of antimicrobial peptides. Whereas -defensins are mainly expressed in mammalian neutrophils, other members of this family have been described in various species within mammals, insects and plants. Although these peptides are thought to primarily disrupt microbial cell walls, they might also act as immunomodulators.
endogenous protein into the active serine protease plas-min. The broad substrate spectrum of plasmin includes C3 and human IgG. Even more strikingly, plasmin was recently shown to bind to the surface of S. aureus and remove deposited IgG, C3b and iC3b from its surface, thereby reversing the effect of opsonization41. Plasmin cleavage of IgG at its hinge region might also attenuate classical-pathway activation through C1q. In addition to plasmin activation, staphylokinase directly abolishes the bactericidal properties of -defensins, which makes it an important colonization factor69. Although the anti-opsonization activities of surface-activated plasmin have so far only been described for S. aureus, the pres-ence of plasminogen activators and receptors on various other bacteria40 suggests that this strategy could be more widely prevalent.
Suppression of complement amplification. Surprisingly, no RCA-recruiting proteins have been reported for S. aureus so far. However, recent reports have described the factor-I-mediated degradation of C3b to iC3b that is independent of factor H on the sur-face of certain S. aureus strains. The overall result of
this process is the decreased efficiency of bacterial killing by phagocytic neutrophils70,71. Although these findings are suggestive of a bacterially derived cofac-tor protein, current evidence suggests that S. aureus has evolved different pathways for counteracting the downstream actions of complement. By target-ing the central components C3 and C5, most of the opsonization, amplification and signalling steps can be efficiently suppressed. In fact, there are several unique S. aureus proteins that inhibit the conversion of native C3 to C3b. Extracellular fibrinogen-binding protein (Efb) was the first C3b-binding protein to be identified in S. aureus and was shown to inhibit opsonophagocytosis by granulocytes72. Biochemical and structural studies revealed that Efb binds to the thioester-containing C3d domain of C3 through its three-helix bundle carboxy-terminal domain (Efb-C)73,74 (FIG. 3a). Recently, an Efb-homologous protein (Ehp) from S. aureus was reported75 that shares a high level of structural similarity with Efb-C. Surprisingly, Ehp can bind two molecules of C3d and is a more potent inhibitor of C3 conversion through the alternative pathway75. Both proteins produce their effects by inducing conformational changes in C3 that render this central component unable to participate in downstream activation events74,75. A recent study indi-cates that Efb and Ehp (designated Ecb in this case76) can also bind to the C3b-containing convertases and, therefore, prevent the cleavage of both C3 (amplifica-tion) and C5 (generation of C5a)76. This convertase-centred inhibition is probably a manifestation of a conformational change in C3b that has occurred upon the binding of Efb74.
S. aureus also expresses another group of small, helical proteins that block complement activation. These staphylococcal complement inhibitors (SCINs) (FIG. 3a) seem to stabilize the C3 convertases in a non-functional state, thereby efficiently blocking all three pathways77. In contrast to Efb and Ehp, SCINs bind only to the assembled C3 convertases76–78 and not to C3 or its fragments. Both the Efb and SCIN families share a high degree of structural similarity with the IgG-binding module of SpA74,78 (FIG. 3a). However, a closer inspection of these structures reveals that the spatial arrangement of the helices is distinct. SCINs are more similar to SpA than Efb or Ehp, which may partly explain the differential binding specificities of these proteins.
Inhibition on the level of C5. In addition to the inter-ference at the stage of C3, S. aureus also produces two proteins that disrupt the downstream inflammatory responses that are carried out by C5 and its activation products. Staphylococcal superantigen-like protein-7 (SSL-7) (FIG. 3b) has recently been shown to bind native C5 with high affinity, resulting in the inhibi-tion of complement-mediated haemolysis and killing of E. coli in an in vitro setting79. SSL-7 also interacts with IgA in a C5-independent manner79. Separately, the chemotaxis inhibitory protein of S. aureus (CHIPS) (FIG. 3b) was found to impair neutrophil and
Figure 3 | Complement-targeting proteins in Staphylococcus aureus. A range of S. aureus proteins that interact with targets from the complement system have been structurally analysed. a | The immunoglobulin G (IgG)-binding module of staphylococcal protein A (SpA; Protein Data Bank (PDB) code: 1BDD63), the C3-binding domain of the extracellular fibrinogen-binding protein (Efb; PDB code: 2GOM74) and staphylococcal complement inhibitor (SCIN; PDB code: 2QFF78) share similar structural motifs, but show differential functionality. b | Both staphylococcal superantigen-like protein-7 (SSL-7; PDB code: 1V1O103) and the chemotaxis inhibitory protein of S. aureus (CHIPS; PDB code: 1XEE104) attenuate complement on the level of C5, but bind to unrelated targets (C5 protein and C5a receptor, respectively). The solution structure of human C5a (obtained by nuclear magnetic resonance at pH 5.2; PDB code: 1KJS105) is shown as a comparison. All protein-structure representations are coloured according to their secondary structure ( -helices in red and -sheets in yellow).
R E V I E W S
138 | FEBRUARY 2008 | VOLUME 6 www.nature.com/reviews/micro
©!2008!Nature Publishing Group!

-grasp domainA structural fold that consists of anti-parallel -strands that ‘grasp’ a single -helix. Initially described as the central structural element of ubiquitin, this fold was later identified in several other proteins. Despite its small size, it has a wide range of functions and is present in enzymatic, binding and signalling proteins. -grasp domains are central elements in the structures of a number of bacterial immune-evasion proteins.
monocyte responses to C5a signalling, primarily by interacting with and antagonizing the C5a recep-tor80–82. Although CHIPS does not share substantial homology in either sequence or secondary structure with the natural ligand C5a (FIG. 3b), the similar overall shape and size of these molecules might explain their competition for the same receptor. Interestingly, and despite their structurally and functionally unrelated targets (native C5 and C5a receptor, respectively), the structures of SSL-7 and CHIPS share some simi-larities55 (FIG. 3b). In this respect, the prominent fold that is observed in both proteins is related to the
-grasp domains that are found in several bacterial superantigens and superantigen-like proteins82,83, and are representative of a large family of bacterial immune-evasion proteins.
Therapeutic potentialOur increasing knowledge of individual microbial agents, their human targets and their mechanisms of action could not only facilitate our understanding of microbial infection but also pave the way towards new therapeutics. Currently, our arsenal of antibiotic drugs is becoming alarmingly ineffective against many severe infectious diseases and the pipeline of novel therapeutic concepts is thin. In the past, drug-screening initiatives focused mainly on bacterial metabolism targets rather than immune evasion84. Nevertheless, recent discover-ies of complement-attenuating proteins have revealed some promising, albeit indirect, targets for therapeutic intervention. One potential limitation of this approach is that microorganisms often have multiple, sometimes redundant, evasion mechanisms15, and not all of these are equally important in pathogenesis. For example, the acquisition of factor H is not essential for the mammalian infection of B. burgdorferi85. Although confirmation of individual proteins as virulence fac-tors by in vivo mutational studies should be a first and crucial step for selecting drug targets, it is clear that classifying promising targets by basic research will provide the foundation of any such development efforts. To this end, governmental initiatives, such as that implemented by the United States Food and Drug Administration (FDA) — for example, the Antibiotic Safety and Innovation amendment to the Revitalization Act and the Orphan Drug Act — may provide new incentives for antibiotic drug development in which evasion strategies are targeted86.
Whereas the development of new antibiotics might represent a speculative endeavour, the path towards the development of new therapeutics for complement-mediated diseases seems clearer. Increased activation or insufficient regulation of this attack system on human cells is known to induce an ever-growing list of autoimmune, inflammatory and ischaemic conditions9. In many cases, the mechanisms that are involved have been revealed and numerous attractive pharmacologi-cal targets within the complement network have been identified2. Nonetheless, the number of anti-complement therapeutics on the market remains surprisingly lim-ited10. In this respect, many microbial proteins are adept
at inhibiting the complement response, and these mol-ecules provide a broad structural and functional frame-work for complement-modulating agents. Although the therapeutic administration of soluble forms of human RCA has long been considered to be an ideal method for supporting the natural regulation of complement, their size (for example, soluble CR1) or restricted activ-ity (for example, soluble DAF and MCP) has limited this approach. In this context, the development of modified, non-immunogenic viral RCA mimics based on SPICE and VCP could overcome some of these limitations, as they have high decay-acceleration and cofactor activity but are comparatively small. Indeed, VCP has been tested in an in vitro model of Alzheimer’s disease and animal models of head trauma87,88. Furthermore, the structural information on these highly potent mimics27,28 could serve as a template for designing modified host RCA that have enhanced functional properties.
The versatile arsenal of direct complement inhibitors that is provided by S. aureus could be another promis-ing source of drug templates. As potent inhibitors of the alternative pathway, proteins such as Efb, Ehp and SCIN have clinical potential for pathological conditions in which the excessive or uncontrolled generation of C3b seems to result in disease (for example, age-related macular degeneration). Owing to its broad specificity for all C3 convertases, SCIN could also be effective for conditions in which activation of the classical pathway is involved (for example, systemic lupus erythematosus). Finally, impairing the signalling function of the C5a receptor using CHIPS could serve as a new approach for a range of inflammatory disorders. Although all these proteins are highly soluble and can be expressed in high quantities, their potential immunogenicity could severely limit their direct use as therapeutics, as has recently been shown for CHIPS82. However, the avail-ability of robust structural data for these proteins (in some cases, even as co-crystals with their human target (BOX 2)) should facilitate the rational design of smaller, synthetic compounds that mimic the potent functions of these microbial proteins.
PerspectivesThe coexistence of humans and microorganisms has generated a fascinating interplay of mechanisms that maintain the balance between infection, defence and evasion. For many infectious diseases, however, this bal-ance is changed in favour of the pathogenic intruders. Alongside many other factors, such as cell adhesion and entry or colonization efficiency, avoiding the attacks of the immune system is a key determinant of microbial survival inside an often hostile host environment. The diverse and elaborate evasion strategies that are discussed here illustrate the importance of the human complement system as a target for immune escape.
The new structural insights into complement evasion, combined with our increased knowledge of complement functions in microbial defence and human disease, provide an exciting platform for long-awaited progress in research. In this context, combinations of various techniques and more systemic approaches are
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | FEBRUARY 2008 | 139
©!2008!Nature Publishing Group!
Nature Reviews | Microbiology
Alignment with C3
Alignment with C3b
Efb-C
C3d
an important premise for coping with the complex nature of host–pathogen interactions. The continued availability of sequenced microbial genomes can only supplement our list of known complement-targeting proteins. With an eye to the future, continued studies
on such mechanisms of immune evasion and comple-ment inhibition should not only further our under-standing of these host–pathogen interactions, but also provide vital insights into better treatment strategies for many types of human diseases.
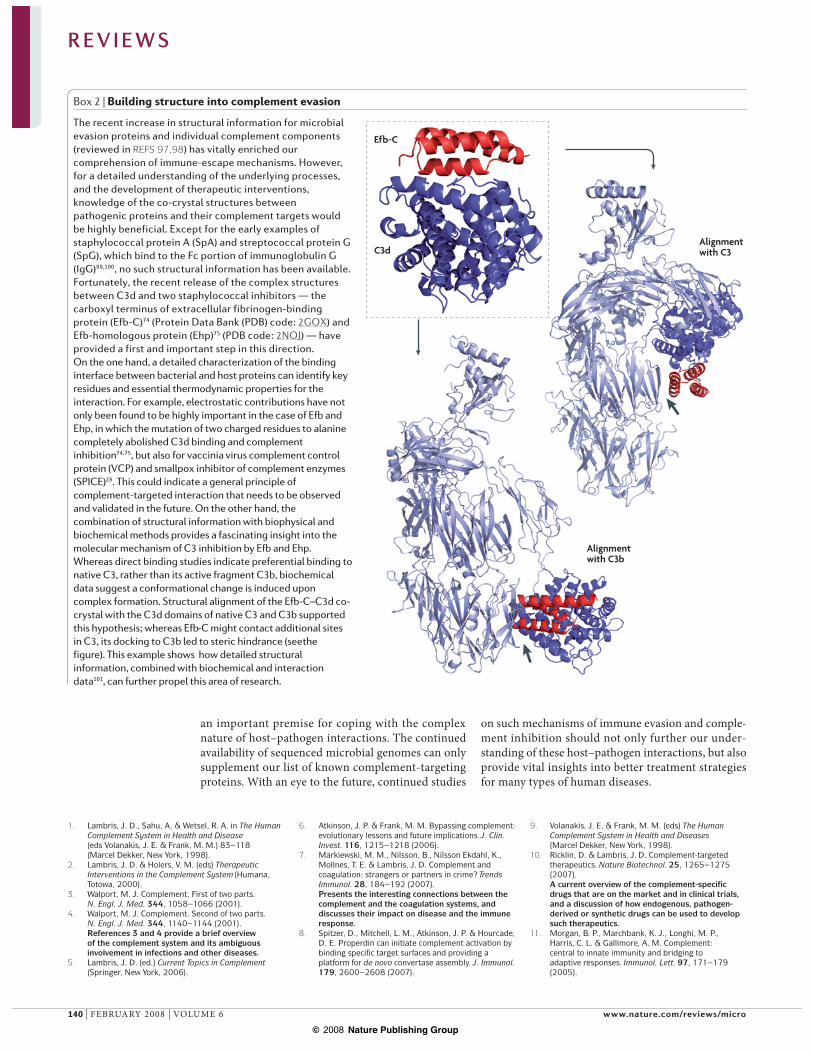
Box 2 | Building structure into complement evasion
The recent increase in structural information for microbial evasion proteins and individual complement components (reviewed in REFS 97,98) has vitally enriched our comprehension of immune-escape mechanisms. However, for a detailed understanding of the underlying processes, and the development of therapeutic interventions, knowledge of the co-crystal structures between pathogenic proteins and their complement targets would be highly beneficial. Except for the early examples of staphylococcal protein A (SpA) and streptococcal protein G (SpG), which bind to the Fc portion of immunoglobulin G (IgG)99,100, no such structural information has been available. Fortunately, the recent release of the complex structures between C3d and two staphylococcal inhibitors — the carboxyl terminus of extracellular fibrinogen-binding protein (Efb-C)74 (Protein Data Bank (PDB) code: 2GOX) and Efb-homologous protein (Ehp)75 (PDB code: 2NOJ) — have provided a first and important step in this direction. On the one hand, a detailed characterization of the binding interface between bacterial and host proteins can identify key residues and essential thermodynamic properties for the interaction. For example, electrostatic contributions have not only been found to be highly important in the case of Efb and Ehp, in which the mutation of two charged residues to alanine completely abolished C3d binding and complement inhibition74,75, but also for vaccinia virus complement control protein (VCP) and smallpox inhibitor of complement enzymes (SPICE)28. This could indicate a general principle of complement-targeted interaction that needs to be observed and validated in the future. On the other hand, the combination of structural information with biophysical and biochemical methods provides a fascinating insight into the molecular mechanism of C3 inhibition by Efb and Ehp. Whereas direct binding studies indicate preferential binding to native C3, rather than its active fragment C3b, biochemical data suggest a conformational change is induced upon complex formation. Structural alignment of the Efb-C–C3d co-crystal with the C3d domains of native C3 and C3b supported this hypothesis; whereas Efb-C might contact additional sites in C3, its docking to C3b led to steric hindrance (see the figure). This example shows how detailed structural information, combined with biochemical and interaction data101, can further propel this area of research.
1. Lambris, J. D., Sahu, A. & Wetsel, R. A. in The Human Complement System in Health and Disease (eds Volanakis, J. E. & Frank, M. M.) 83–118 (Marcel Dekker, New York, 1998).
2. Lambris, J. D. & Holers, V. M. (eds) Therapeutic Interventions in the Complement System (Humana, Totowa, 2000).
3. Walport, M. J. Complement. First of two parts. N. Engl. J. Med. 344, 1058–1066 (2001).
4. Walport, M. J. Complement. Second of two parts. N. Engl. J. Med. 344, 1140–1144 (2001).References 3 and 4 provide a brief overview of the complement system and its ambiguous involvement in infections and other diseases.
5. Lambris, J. D. (ed.) Current Topics in Complement (Springer, New York, 2006).
6. Atkinson, J. P. & Frank, M. M. Bypassing complement: evolutionary lessons and future implications. J. Clin. Invest. 116, 1215–1218 (2006).
7. Markiewski, M. M., Nilsson, B., Nilsson Ekdahl, K., Mollnes, T. E. & Lambris, J. D. Complement and coagulation: strangers or partners in crime? Trends Immunol. 28, 184–192 (2007).Presents the interesting connections between the complement and the coagulation systems, and discusses their impact on disease and the immune response.
8. Spitzer, D., Mitchell, L. M., Atkinson, J. P. & Hourcade, D. E. Properdin can initiate complement activation by binding specific target surfaces and providing a platform for de novo convertase assembly. J. Immunol. 179, 2600–2608 (2007).
9. Volanakis, J. E. & Frank, M. M. (eds) The Human Complement System in Health and Diseases (Marcel Dekker, New York, 1998).
10. Ricklin, D. & Lambris, J. D. Complement-targeted therapeutics. Nature Biotechnol. 25, 1265–1275 (2007).A current overview of the complement-specific drugs that are on the market and in clinical trials, and a discussion of how endogenous, pathogen-derived or synthetic drugs can be used to develop such therapeutics.
11. Morgan, B. P., Marchbank, K. J., Longhi, M. P., Harris, C. L. & Gallimore, A. M. Complement: central to innate immunity and bridging to adaptive responses. Immunol. Lett. 97, 171–179 (2005).
R E V I E W S
140 | FEBRUARY 2008 | VOLUME 6 www.nature.com/reviews/micro
©!2008!Nature Publishing Group!

12. Hawlisch, H. et al. C5a negatively regulates toll-like receptor 4-induced immune responses. Immunity 22, 415–426 (2005).
13. Zhang, X. et al. Regulation of Toll-like receptor-mediated inflammatory response by complement in vivo. Blood 110, 228–236 (2007).
14. Joiner, K., Brown, E., Hammer, C., Warren, K. & Frank, M. Studies on the mechanism of bacterial resistance to complement-mediated killing. III. C5b-9 deposits stably on rough and type 7 S. pneumoniae without causing bacterial killing. J. Immunol. 130, 845–849 (1983).
15. Zipfel, P. F., Wurzner, R. & Skerka, C. Complement evasion of pathogens: common strategies are shared by diverse organisms. Mol. Immunol. 44, 3850–3857 (2007).A supplementary description of the current topics in the field of complement evasion, with the focus on redundancy and multiplicity, the acquisition of host regulators and simultaneous binding to other host proteins.
16. Kraiczy, P. & Würzner, R. Complement escape of human pathogenic bacteria by acquisition of complement regulators. Mol. Immunol. 43, 31–44 (2006).
17. Bernet, J., Mullick, J., Singh, A. K. & Sahu, A. Viral mimicry of the complement system. J. Biosci. 28, 249–264 (2003).A broad overview of the use of complement proteins by viruses that discusses immune evasion and cell attachment and entry.
18. Meri, T. et al. The hyphal and yeast forms of Candida albicans bind the complement regulator C4b-binding protein. Infect. Immun. 72, 6633–6641 (2004).
19. Meri, T. et al. The yeast Candida albicans binds complement regulators factor H and FHL-1. Infect. Immun. 70, 5185–5192 (2002).
20. Inal, J. M. Parasite interaction with host complement: beyond attack regulation. Trends Parasitol. 20, 407–412 (2004).
21. Kooyman, D. L. et al. In vivo transfer of GPI-linked complement restriction factors from erythrocytes to the endothelium. Science 269, 89–92 (1995).
22. Rautemaa, R., Jarvis, G. A., Marnila, P. & Meri, S. Acquired resistance of Escherichia coli to complement lysis by binding of glycophosphoinositol-anchored protectin (CD59). Infect. Immun. 66, 1928–1933 (1998).
23. Rautemaa, R. et al. Survival of Helicobacter pylori from complement lysis by binding of GPI-anchored protectin (CD59). Gastroenterology 120, 470–479 (2001).
24. Tortorella, D., Gewurz, B. E., Furman, M. H., Schust, D. J. & Ploegh, H. L. Viral subversion of the immune system. Annu. Rev. Immunol. 18, 861–926 (2000).
25. McKenzie, R., Kotwal, G. J., Moss, B., Hammer, C. H. & Frank, M. M. Regulation of complement activity by vaccinia virus complement-control protein. J. Infect. Dis. 166, 1245–1250 (1992).
26. Rosengard, A. M., Liu, Y., Nie, Z. & Jimenez, R. Variola virus immune evasion design: expression of a highly efficient inhibitor of human complement. Proc. Natl Acad. Sci. USA 99, 8808–8813 (2002).
27. Murthy, K. H. et al. Crystal structure of a complement control protein that regulates both pathways of complement activation and binds heparan sulfate proteoglycans. Cell 104, 301–311 (2001).
28. Sfyroera, G., Katragadda, M., Morikis, D., Isaacs, S. N. & Lambris, J. D. Electrostatic modeling predicts the activities of orthopoxvirus complement control proteins. J. Immunol. 174, 2143–2151 (2005).
29. Liszewski, M. K. et al. Structure and regulatory profile of the monkeypox inhibitor of complement: comparison to homologs in vaccinia and variola and evidence for dimer formation. J. Immunol. 176, 3725–3734 (2006).
30. Miller, C. G., Shchelkunov, S. N. & Kotwal, G. J. The cowpox virus-encoded homolog of the vaccinia virus complement control protein is an inflammation modulatory protein. Virology 229, 126–133 (1997).
31. Engelstad, M., Howard, S. T. & Smith, G. L. A constitutively expressed vaccinia gene encodes a 42-kDa glycoprotein related to complement control factors that forms part of the extracellular virus envelope. Virology 188, 801–810 (1992).
32. Mullick, J., Bernet, J., Singh, A. K., Lambris, J. D. & Sahu, A. Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8) open reading frame 4 protein (kaposica) is a functional homolog of complement control proteins. J. Virol. 77, 3878–3881 (2003).
33. Daix, V. et al. Ixodes ticks belonging to the Ixodes ricinus complex encode a family of anticomplement proteins. Insect Mol. Biol. 16, 155–166 (2007).
34. Valenzuela, J. G., Charlab, R., Mather, T. N. & Ribeiro, J. M. Purification, cloning, and expression of a novel salivary anticomplement protein from the tick, Ixodes scapularis. J. Biol. Chem. 275, 18717–18723 (2000).
35. Rooijakkers, S. H. & van Strijp, J. A. Bacterial complement evasion. Mol. Immunol. 44, 23–32 (2007).
36. Oda, T. et al. Inactivation of chemotactic activity of C5a by the serratial 56-kilodalton protease. Infect. Immun. 58, 1269–1272 (1990).
37. Chmouryguina, I., Suvorov, A., Ferrieri, P. & Cleary, P. P. Conservation of the C5a peptidase genes in group A and B streptococci. Infect. Immun. 64, 2387–2390 (1996).
38. Tsao, N. et al. Streptococcal pyrogenic exotoxin B cleaves properdin and inhibits complement-mediated opsonophagocytosis. Biochem. Biophys. Res. Commun. 339, 779–784 (2006).
39. Ghendler, Y., Parizade, M., Arnon, R., McKerrow, J. H. & Fishelson, Z. Schistosoma mansoni: evidence for a 28-kDa membrane-anchored protease on schistosomula. Exp. Parasitol. 83, 73–82 (1996).
40. Lahteenmaki, K., Kuusela, P. & Korhonen, T. K. Bacterial plasminogen activators and receptors. FEMS Microbiol. Rev. 25, 531–552 (2001).
41. Rooijakkers, S. H., van Wamel, W. J., Ruyken, M., van Kessel, K. P. & van Strijp, J. A. Anti-opsonic properties of staphylokinase. Microbes Infect. 7, 476–484 (2005).
42. Frank, M. M. Annihilating host defense. Nature Med. 7, 1285–1286 (2001).
43. Lubinski, J., Nagashunmugam, T. & Friedman, H. M. Viral interference with antibody and complement. Semin. Cell Dev. Biol. 9, 329–337 (1998).
44. Rux, A. H. et al. Kinetic analysis of glycoprotein C of herpes simplex virus types 1 and 2 binding to heparin, heparan sulfate, and complement component C3b. Virology 294, 324–332 (2002).
45. Kostavasili, I. et al. Mechanism of complement inactivation by glycoprotein C of herpes simplex virus. J. Immunol. 158, 1763–1771 (1997).
46. Favoreel, H. W., Van de Walle, G. R., Nauwynck, H. J. & Pensaert, M. B. Virus complement evasion strategies. J. Gen. Virol. 84, 1–15 (2003).
47. Inal, J. M. & Sim, R. B. A Schistosoma protein, Sh-TOR, is a novel inhibitor of complement which binds human C2. FEBS Lett. 470, 131–134 (2000).
48. Deng, J., Gold, D., LoVerde, P. T. & Fishelson, Z. Inhibition of the complement membrane attack complex by Schistosoma mansoni paramyosin. Infect. Immun. 71, 6402–6410 (2003).
49. Gobert, G. N. & McManus, D. P. Update on paramyosin in parasitic worms. Parasitol. Int. 54, 101–107 (2005).
50. Nunn, M. A. et al. Complement inhibitor of C5 activation from the soft tick Ornithodoros moubata. J. Immunol. 174, 2084–2091 (2005).
51. Roversi, P. et al. The structure of OMCI, a novel lipocalin inhibitor of the complement system. J. Mol. Biol. 369, 784–793 (2007).
52. Lowy, F. D. Staphylococcus aureus infections. N. Engl. J. Med. 339, 520–532 (1998).
53. Patti, J. M., Allen, B. L., McGavin, M. J. & Hook, M. MSCRAMM-mediated adherence of microorganisms to host tissues. Annu. Rev. Microbiol. 48, 585–617 (1994).
54. Foster, T. J. & Hook, M. Surface protein adhesins of Staphylococcus aureus. Trends Microbiol. 6, 484–488 (1998).
55. Chavakis, T., Preissner, K. T. & Herrmann, M. The anti-inflammatory activities of Staphylococcus aureus. Trends Immunol. 28, 408–418 (2007).This comprehensive review discusses how S. aureus uses various target points (including complement) to escape the attack of the immune system.
56. Foster, T. J. Immune evasion by staphylococci. Nature Rev. Microbiol. 3, 948–958 (2005).
57. Rooijakkers, S. H., van Kessel, K. P. & van Strijp, J. A. Staphylococcal innate immune evasion. Trends Microbiol. 13, 596–601 (2005).
58. O’Riordan, K. & Lee, J. C. Staphylococcus aureus capsular polysaccharides. Clin. Microbiol. Rev. 17, 218–234 (2004).
59. Peterson, P. K. et al. Dichotomy between opsonization and serum complement activation by encapsulated staphylococci. Infect. Immun. 20, 770–775 (1978).
60. Cunnion, K. M., Lee, J. C. & Frank, M. M. Capsule production and growth phase influence binding of complement to Staphylococcus aureus. Infect. Immun. 69, 6796–6803 (2001).
61. Verbrugh, H. A., Peterson, P. K., Nguyen, B. Y., Sisson, S. P. & Kim, Y. Opsonization of encapsulated Staphylococcus aureus: the role of specific antibody and complement. J. Immunol. 129, 1681–1687 (1982).
62. Cedergren, L., Andersson, R., Jansson, B., Uhlen, M. & Nilsson, B. Mutational analysis of the interaction between staphylococcal protein A and human IgG1. Protein Eng. 6, 441–448 (1993).
63. Gouda, H. et al. Three-dimensional solution structure of the B domain of staphylococcal protein A: comparisons of the solution and crystal structures. Biochemistry 31, 9665–9672 (1992).
64. Nguyen, T., Ghebrehiwet, B. & Peerschke, E. I. Staphylococcus aureus protein A recognizes platelet gC1qR/p33: a novel mechanism for staphylococcal interactions with platelets. Infect. Immun. 68, 2061–2068 (2000).
65. Peerschke, E. I., Bayer, A. S., Ghebrehiwet, B. & Xiong, Y. Q. gC1qR/p33 blockade reduces Staphylococcus aureus colonization of target tissues in an animal model of infective endocarditis. Infect. Immun. 74, 4418–4423 (2006).
66. Hartleib, J. et al. Protein A is the von Willebrand factor binding protein on Staphylococcus aureus. Blood 96, 2149–2156 (2000).
67. Gómez, M. I. et al. Staphylococcus aureus protein A induces airway epithelial inflammatory responses by activating TNFR1. Nature Med. 10, 842–848 (2004).
68. Zhang, L., Jacobsson, K., Vasi, J., Lindberg, M. & Frykberg, L. A second IgG-binding protein in Staphylococcus aureus. Microbiology 144, 985–991 (1998).
69. Jin, T. et al. Staphylococcus aureus resists human defensins by production of staphylokinase, a novel bacterial evasion mechanism. J. Immunol. 172, 1169–1176 (2004).
70. Cunnion, K. M., Hair, P. S. & Buescher, E. S. Cleavage of complement C3b to iC3b on the surface of Staphylococcus aureus is mediated by serum complement factor I. Infect. Immun. 72, 2858–2863 (2004).
71. Cunnion, K. M., Buescher, E. S. & Hair, P. S. Serum complement factor I decreases Staphylococcus aureus phagocytosis. J. Lab. Clin. Med. 146, 279–286 (2005).
72. Lee, L. Y. et al. Inhibition of complement activation by a secreted Staphylococcus aureus protein. J. Infect. Dis. 190, 571–579 (2004).
73. Lee, L. Y., Liang, X., Hook, M. & Brown, E. L. Identification and characterization of the C3 binding domain of the Staphylococcus aureus extracellular fibrinogen-binding protein (Efb). J. Biol. Chem. 279, 50710–50716 (2004).
74. Hammel, M. et al. A structural basis for complement inhibition by Staphylococcus aureus. Nature Immunol. 8, 430–437 (2007).Describes the first co-crystal structure between a microbial complement-evasion protein and its host target domain; also elucidates the inhibition mechanism on a molecular level by combining biochemical and biophysical approaches.
75. Hammel, M. et al. Characterization of Ehp, a secreted complement inhibitory protein from Staphylococcus aureus. J. Biol. Chem. 282, 30051–30061 (2007).
76. Jongerius, I. et al. Staphylococcal complement evasion by various convertase-blocking molecules. J. Exp. Med. 204, 2461–2471 (2007).
77. Rooijakkers, S. H. M. et al. Immune evasion by a staphylococcal complement inhibitor that acts on C3 convertases. Nature Immunol. 6, 920–927 (2005).
78. Rooijakkers, S. H. M. et al. Staphylococcal complement inhibitor: structure and active sites. J. Immunol. 179, 2989–2998 (2007).The authors describe the crystal structure of a SCIN protein and identify its functional sites by analysing protein chimeras of SCIN and an inactive homologue.
79. Langley, R. et al. The staphylococcal superantigen-like protein 7 binds IgA and complement C5 and inhibits IgA-FcaRI binding and serum killing of bacteria. J. Immunol. 174, 2926–2933 (2005).
80. de Haas, C. J. et al. Chemotaxis inhibitory protein of Staphylococcus aureus, a bacterial antiinflammatory agent. J. Exp. Med. 199, 687–695 (2004).
81. Postma, B. et al. Chemotaxis inhibitory protein of Staphylococcus aureus binds specifically to the C5a
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | FEBRUARY 2008 | 141
©!2008!Nature Publishing Group!

and formylated peptide receptor. J. Immunol. 172, 6994–7001 (2004).
82. Wright, A. J. et al. Characterisation of receptor binding by the chemotaxis inhibitory protein of Staphylococcus aureus and the effects of the host immune response. Mol. Immunol. 44, 2507–2517 (2007).
83. Geisbrecht, B. V., Hamaoka, B. Y., Perman, B., Zemla, A. & Leahy, D. J. The crystal structures of EAP domains from Staphylococcus aureus reveal an unexpected homology to bacterial superantigens. J. Biol. Chem. 280, 17243–17250 (2005).
84. Payne, D. J., Gwynn, M. N., Holmes, D. J. & Pompliano, D. L. Drugs for bad bugs: confronting the challenges of antibacterial discovery. Nature Rev. Drug Discov. 6, 29–40 (2007).Although not directly related to complement evasion, this insider view on new screening initiatives for antibiotic drugs by pharmaceutical companies provides noteworthy information on targets and challenges for this endeavour.
85. Woodman, M. E. et al. Borrelia burgdorferi binding of host complement regulator factor H is not required for efficient mammalian infection. Infect. Immun. 75, 3131–3139 (2007).
86. A step in the right direction. Nature Rev. Drug Discov. 6, 419 (2007).
87. Daly, J. & Kotwal, G. J. Pro-inflammatory complement activation by the A peptide of Alzheimer’s disease is biologically significant and can be blocked by vaccinia virus complement control protein. Neurobiol. Aging 19, 619–627 (1998).
88. Pillay, N. S., Kellaway, L. A. & Kotwal, G. J. Vaccinia virus complement control protein significantly improves sensorimotor function recovery after severe head trauma. Brain Res. 1153, 158–165 (2007).
89. Marsh, M. & Helenius, A. Virus entry: open sesame. Cell 124, 729–740 (2006).
90. Nemerow, G. R., Mold, C., Schwend, V. K., Tollefson, V. & Cooper, N. R. Identification of gp350 as the viral glycoprotein mediating attachment of Epstein–Barr virus (EBV) to the EBV/C3d receptor of B cells: sequence homology of gp350 and C3 complement fragment C3d. J. Virol. 61, 1416–1420 (1987).
91. Tsoukas, C. D. & Lambris, J. D. Expression of EBV/C3d receptors on T cells: biological significance. Immunol. Today 14, 56–59 (1993).
92. Naniche, D. et al. Human membrane cofactor protein (CD46) acts as a cellular receptor for measles virus. J. Virol. 67, 6025–6032 (1993).
93. Santoro, F. et al. Interaction of glycoprotein H of human herpesvirus 6 with the cellular receptor CD46. J. Biol. Chem. 278, 25964–25969 (2003).
94. Nowicki, B., Hart, A., Coyne, K. E., Lublin, D. M. & Nowicki, S. Short consensus repeat-3 domain of recombinant decay-accelerating factor is recognized by Escherichia coli recombinant Dr adhesin in a model of a cell–cell interaction. J. Exp. Med. 178, 2115–2121 (1993).
95. Datta, P. K. & Rappaport, J. HIV and complement: hijacking an immune defense. Biomed. Pharmacother. 60, 561–568 (2006).
96. Schorey, J. S., Carroll, M. C. & Brown, E. J. A macrophage invasion mechanism of pathogenic mycobacteria. Science 277, 1091–1093 (1997).
97. Janssen, B. J. & Gros, P. Structural insights into the central complement component C3. Mol. Immunol. 44, 3–10 (2007).
98. Lambris, J. D. & Morikis, D. (eds) Structural Biology of the Complement System (CRC, Boca Raton, 2005).
99. Deisenhofer, J., Jones, T. A., Huber, R., Sjodahl, J. & Sjoquist, J. Crystallization, crystal structure analysis and atomic model of the complex formed by a human Fc fragment and fragment B of protein A from Staphylococcus aureus. Hoppe-Seyler’s Z. Physiol. Chem. 359, 975–985 (1978).
100. Sauer-Eriksson, A. E., Kleywegt, G. J., Uhlen, M. & Jones, T. A. Crystal structure of the C2 fragment of streptococcal protein G in complex with the Fc domain of human IgG. Structure 3, 265–278 (1995).
101. Ricklin, D. & Lambris, J. D. Exploring the complement interaction network using surface plasmon resonance. Adv. Exp. Med. Biol. 598, 260–278 (2007).
102. Lukacik, P. et al. Complement regulation at the molecular level: the structure of decay-accelerating factor. Proc. Natl Acad. Sci. USA 101, 1279–1284 (2004).
103. Al-Shangiti, A. M. et al. Structural relationships and cellular tropism of staphylococcal superantigen-like proteins. Infect. Immun. 72, 4261–4270 (2004).
104. Haas, P. J. et al. The structure of the C5a receptor-blocking domain of chemotaxis inhibitory protein of Staphylococcus aureus is related to a group of immune evasive molecules. J. Mol. Biol. 353, 859–872 (2005).
105. Zhang, X., Boyar, W., Toth, M. J., Wennogle, L. & Gonnella, N. C. Structural definition of the C5a C terminus by two-dimensional nuclear magnetic resonance spectroscopy. Proteins 28, 261–267 (1997).
Acknowledgements This work was supported by National Institutes of Health grants GM-069736, GM-62134, AI-30040, AI-072106 and AI-068730, and by the research incentive funds of the School of Biological Sciences at the University of Missouri-Kansas City.
Competing interest statementThe authors declare competing financial interests: see web version for details.
DATABASESEntrez Genome: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=genomecowpox | HHV-8 | HIV-1 | monkeypox | vaccinia | variolaEntrez Genome Project: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=genomeprjBorrelia burgdorferi | Candida albicans | Escherichia coli | Helicobacter pylori | Schistosoma mansoni | Serratia marcescens | Staphylococcus aureusProtein Data Bank: http://www.rcsb.org/pdb/home/home.dohuman C5a | CHIPS | DAF | Efb | SCIN | SpA | SSL-7 | VCP
FURTHER INFORMATIONJohn D. Lambris’s homepage: http://www.lambris.net
SUPPLEMENTARY INFORMATIONSee online article: S1 (table)
ALL LINKS ARE ACTIVE IN THE ONLINE PDF
R E V I E W S
142 | FEBRUARY 2008 | VOLUME 6 www.nature.com/reviews/micro
©!2008!Nature Publishing Group!
Review
Nature Reviews Microbiology 6, 132-142 (February 2008) |
doi:10.1038/nrmicro1824
Complement evasion by human pathogens
John D. Lambris1 (#a1) , Daniel Ricklin1 (#a1) & Brian V. Geisbrecht2 (#a2)
Summary
The human complement system has a pivotal role in the recognition, opsonization
and elimination of microbial intruders. This functionality is maintained by a well-
balanced interaction network of serum proteins and cell-surface receptors.
Over thousands of years of co-evolution, many microorganisms have developed
specific complement-evasion strategies to escape the attack of the immune system.
Although some of these strategies are highly specific for a single species, others are
shared more broadly among bacteria, viruses, fungi and parasites.
The most prevalent complement-evasion mechanism seems to be the capture of
soluble host complement regulators on the microbial surface or the expression of
their structural mimics. However, the inactivation of complement components by
proteolytic degradation or specific inhibition of essential functional sites is also
frequently observed.
Staphylococcus aureus has a particularly wide and diverse arsenal of complement-
evasion proteins, many of which have been discovered only recently. These
numerous evasion strategies could contribute to the high virulence of this
bacterium.
Structural biology is an indispensable tool for characterizing the structure and
function of complement-evasion proteins. Recent publications of the co-crystal
structures of evasion proteins with their human targets have allowed an even deeper
insight into the molecular basis of these escape mechanisms.
The rapid increase in the structural and functional understanding of complement
evasion could serve as an important starting point for the development of
antimicrobial or complement-targeting therapeutics.

About the authors
JOHN D. LAMBRIS
John D. Lambris is the Dr Ralph and Sallie Weaver Professor of Research Medicine at
the department of Pathology and Laboratory Medicine of the University of
Pennsylvania, Philadelphia, USA. After earning his Ph.D. in biochemistry from the
University of Patras, Greece, he dedicated his research to various aspects of the
complement system. In his laboratory at the University of Pennsylvania, he applies
ideas and methods that are embodied in engineering, computer science, physics,
chemistry and other fields to address today's challenges in complement research. His
current research efforts include the development of small-size complement inhibitors,
the crosstalk of complement with other pathways and complement evasion by
bacteria and viruses. He has been the president of the International Complement
Society and is the founder and executive director of the Aegean Conferences.
DANIEL RICKLIN
Daniel Ricklin is a postdoctoral researcher at the University of Pennsylvania,
Philadelphia, USA, in the research group of John D. Lambris. He earned his M.Sc. in
pharmaceutical sciences from the Swiss Federal Institute of Technology in Zurich,
Switzerland, and his Ph.D. in pharmaceutical chemistry from the University of Basel,
Switzerland. His current research is centred on the structure, function and interaction
network of complement proteins and the therapeutic intervention of complement-
related diseases.
BRIAN V. GEISBRECHT
Brian V. Geisbrecht is an assistant professor of cell biology and biophysics at the
University of Missouri–Kansas City (UMKC), Missouri, USA. He earned a Ph.D. in
biological chemistry from The Johns Hopkins University School of Medicine,
Maryland, USA, which was followed by a postdoctoral appointment in biophysics and
biophysical chemistry at the same institution. Since joining UMKC in 2004, his
laboratory has focused on understanding the molecular basis of host–pathogen
interactions in infectious diseases by using various complementary approaches from
structural biology (X-ray crystallography, nuclear magnetic resonance and X-ray
scattering), biophysics and biochemistry.

SU
PPLE
MEN
TAR
Y IN
FOR
MAT
ION
In
fo
rmat
pro
vid
ed b
y L
amb
ris
et
al.
(F
EB
RU
AR
Y 2
00
8)
NA
TU
RE
RE
VIE
WS
| MICRO
BIOLO
GY
www.nature.com/reviews/micro
Su
pp
lem
en
tary
in
form
ati
on
S1
(ta
ble
): M
icro
bia
l co
mp
lem
en
t-ta
rget
ing p
rote
ins
P
ath
og
en
Pro
tein
T
arg
et
Ac
tio
n
Re
f
Ba
cte
ria
Actinobacillus
sp
p.
Om
p1
00
Ou
ter
me
mb
ran
e p
rote
in 1
00
fH
Re
cru
itm
en
t o
f re
gu
lato
rs
1
Bordetella
sp
p.
FH
A
Fila
me
nto
us h
em
ag
glu
tin
in
C4
BP
, (f
H,
FH
L-1
) R
ecru
itm
en
t o
f re
gu
lato
rs
2
Borrelia
sp
p.
CR
AS
P
Co
mp
lem
en
t re
gu
lato
r-a
cq
uir
ing
su
rfa
ce
pro
tein
s
fH, F
HL
-1
Re
cru
itm
en
t o
f re
gu
lato
rs
3
Erp
O
sp
E/F
-re
late
d p
rote
ins
fH
Re
cru
itm
en
t o
f re
gu
lato
rs
4,
5
n/a
C
D5
9-lik
e p
rote
in
C8
, C
9
Pre
ve
nts
MA
C f
orm
atio
n
6
Escherichia
sp
p.
Om
pA
Ou
ter
me
mb
ran
e p
rote
in A
C
4B
P
Re
cru
itm
en
t o
f re
gu
lato
rs
7
Stc
E
Se
cre
ted
pro
tea
se
of
C1
este
rase
in
hib
ito
r C
1-I
NH
R
ecru
itm
en
t o
f C
1-I
NH
8
Tra
T
Tra
T o
ute
r m
em
bra
ne
pro
tein
C
5b
6
Pre
ve
nts
MA
C f
orm
atio
n
9
Fusobacterium
sp
p.
n/a
U
nkn
ow
n fa
cto
r fH
R
ecru
itm
en
t o
f re
gu
lato
rs
10
Haemophilus
sp
p.
n/a
U
nkn
ow
n fa
cto
r C
4B
P, fH
R
ecru
itm
en
t o
f re
gu
lato
rs
11,
12
Moraxella
sp
p.
Usp
A1
/2
Ub
iqu
ito
us s
urf
ace
pro
tein
A1
/ A
2
C4
BP
Re
cru
itm
en
t o
f re
gu
lato
rs
13
Neisseria
sp
p.
LO
S
Lip
oo
ligo
sa
cch
arid
e
fH, F
HL
-1
Re
cru
itm
en
t o
f re
gu
lato
rs
GN
A1
87
0
Ge
no
me
-de
rive
d n
eis
se
ria
l a
ntig
en
18
70
fH, F
HL
-1
Re
cru
itm
en
t o
f re
gu
lato
rs
14
Po
r O
ute
r m
em
bra
ne
po
rin
s
C4
BP
, fH
, F
HL
-1
Re
cru
itm
en
t o
f re
gu
lato
rs
15,
16
n/a
T
yp
e IV
pili
MC
P
Atta
ch
me
nt
to e
pith
elia
l ce
lls
17
Porphyromonas
sp
p.
prt
H
prt
H p
rote
ase
C3
, Ig
G
De
gra
de
s C
3 a
nd
Ig
G
18
©!2008!N
atu
re P
ub
lis
hin
g G
rou
p

SU
PPLE
MEN
TAR
Y IN
FOR
MAT
ION
In
fo
rmat
pro
vid
ed b
y L
amb
ris
et
al.
(F
EB
RU
AR
Y 2
00
8)
NA
TU
RE
RE
VIE
WS
| MICRO
BIOLO
GY
www.nature.com/reviews/micro
Ps
eu
do
mo
na
s s
pp
.
Pa
E
Pse
ud
om
on
as e
lasta
se
C1
q, C
3
De
gra
de
s C
1q
an
d C
3
19
Pa
AP
P
se
ud
om
on
as a
lka
line
pro
tea
se
C
1q
, C
3
De
gra
de
s C
1q
an
d C
3
19
Tu
f E
lon
ga
tio
n fa
cto
r fH
, F
HL
-1
Re
cru
itm
en
t o
f re
gu
lato
rs
20
Se
rra
tia
sp
p.
n/a
5
6 k
Da
pro
tea
se
C
5a
, C
1-I
NH
D
eg
rad
es C
5a
, d
eg
rad
es C
1-I
NH
21,
22
Sta
ph
ylo
co
cc
us
sp
p.
CH
IPS
C
he
mo
taxis
in
hib
ito
ry p
rote
in o
f S
. a
ure
us
C5
aR
A
nta
go
niz
es C
5a
23
Efb
E
xtr
ace
llula
r fib
rin
og
en
-bin
din
g p
rote
in
C3
/C3
b/C
3d
In
hib
itio
n o
f C
3 a
nd
C3
b-c
on
tain
ing
co
nve
rta
se
s
24-2
6
Eh
p a
E
fb-h
om
olo
go
us p
rote
in
C3
/C3
b/C
3d
In
hib
itio
n o
f C
3 a
nd
C3
b-c
on
tain
ing
co
nve
rta
se
s
26,
27
SA
K
Sta
ph
ylo
kin
ase
C
3b
, Ig
G
(via
Pla
sm
in)
Cle
ave
s c
om
ple
me
nt p
rote
ins,
re
mo
ve
s C
3b
fro
m s
urf
ace
28
Sb
i S
. a
ure
us Ig
G-b
ind
ing
pro
tein
Ig
G
Inh
ibits I
g in
tera
ctio
n w
ith
C1
q
29
SC
IN
Sta
ph
ylo
co
cca
l co
mp
lem
en
t in
hib
ito
r C
3 C
on
ve
rta
se
s
Inh
ibits C
3 a
ctiva
tio
n t
o C
3a
/C3
b
26,
30
Sp
A
S. a
ure
us p
rote
in A
Ig
's, g
C1
q-R
In
hib
its I
g in
tera
ctio
n w
ith
C1
q
31
SS
L-7
S
tap
hylo
co
cca
l su
pe
ran
tig
en
-lik
e p
rote
in 7
C
5
Pre
ve
nts
C5
cle
ava
ge
32
Str
ep
toc
oc
cu
s s
pp
.
Ba
c
-Pro
tein
Ig
A,
fH
Re
cru
itm
en
t o
f re
gu
lato
rs
33
Fb
a
Fib
ron
ectin
-bin
din
g p
rote
in
fH, F
HL
-1
Re
cru
itm
en
t o
f re
gu
lato
rs
34
Hic
b
Fa
cto
r H
-bin
din
g in
hib
ito
r o
f co
mp
lem
en
t fH
R
ecru
itm
en
t o
f re
gu
lato
rs
35
Ide
S
IgG
-de
gra
din
g E
nzym
e o
f S
. p
yro
ge
ne
s
IgG
C
lea
ve
s Ig
G, n
o in
tera
ctio
n w
ith
C1
q
36
M b
S
urf
ace
pro
tein
s M
fa
mily
(A
rp, S
ir,
etc
.)
fH, C
4B
P, F
HL
, F
HR
, M
CP
R
ecru
itm
en
t o
f re
gu
lato
rs
37
PL
Y
Pn
eu
mo
lysin
Ig
G, C
1q
C
om
ple
me
nt a
ctiva
tio
n /
de
ple
tio
n
38,
39
Psp
A
Pn
eu
mo
co
cca
l su
rfa
ce
pro
tein
A
un
kn
ow
n
Po
ten
tia
l im
pa
irin
g o
f A
P a
nd
co
mp
lem
en
t re
ce
pto
rs
40
Psp
C c
P
ne
um
oco
cca
l su
rfa
ce
pro
tein
C
fH
(C
3,
IgA
) R
ecru
itm
en
t o
f re
gu
lato
rs,
Po
ten
tia
l d
eg
rad
atio
n o
f C
3/C
3b
41-4
4
scp
A/B
S
tre
pto
co
cca
l C
5a
pe
ptid
ase
C
5a
D
eg
rad
es C
5a
, D
isru
pts
sig
na
ling
45-4
7
SIC
S
tre
pto
co
cca
l in
hib
ito
r o
f co
mp
lem
en
t C
5b
-7, C
5b
-8
Pre
ve
ntio
n o
f M
AC
fo
rma
tio
n
48,
49
SP
E B
S
tre
pto
co
cca
l p
yro
ge
nic
exo
toxin
B
Pro
pe
rdin
, Ig
's
De
gra
de
s P
rop
erd
in,
Ig's
36,
50
Sp
G
Str
ep
toco
ccu
s p
rote
in G
Ig
's
Inh
ibits I
g in
tera
ctio
n w
ith
C1
q
51
©!2008!N
atu
re P
ub
lis
hin
g G
rou
p

SU
PPLE
MEN
TAR
Y IN
FOR
MAT
ION
In
fo
rmat
pro
vid
ed b
y L
amb
ris
et
al.
(F
EB
RU
AR
Y 2
00
8)
NA
TU
RE
RE
VIE
WS
| MICRO
BIOLO
GY
www.nature.com/reviews/micro
Ye
rsin
ia s
pp
.
Ya
dA
Y
ers
inia
ad
he
sin
A
fH
Re
cru
itm
en
t o
f re
gu
lato
rs
52
Vir
us
es
He
rpe
s v
iru
se
s
gC
1/2
T
ran
sm
em
bra
ne
gly
co
pro
tein
s C
1,C
2 (
HS
V)
C3
b
Bin
ds to
C3
b,
de
ca
y a
cce
lera
tio
n (
on
ly A
P),
in
hib
its b
ind
ing
of
pro
pe
rdin
& C
5
53
gE
+g
I G
lyco
pro
tein
s E
+I
(HS
V)
IgG
F
c-r
ece
pto
r, le
ss a
ctiva
tio
n
54,
55
gp
34
,68
G
lyco
pro
tein
s 3
4, 6
8 (
HC
MV
) Ig
G
Fc-r
ece
pto
rs,
less a
ctiva
tio
n
56,
57
gp
I+g
pIV
G
lyco
pro
tein
s I+
IV (
VZ
V)
IgG
F
c-r
ece
pto
r, le
ss a
ctiva
tio
n
58
KC
P d
Ka
po
si's
sa
rko
ma
-asso
cia
ted
co
mp
lem
en
t co
ntr
ol
pro
tein
(K
SH
V)
C3
b
Mim
ics r
eg
ula
tors
(c
ofa
cto
r/d
eca
y a
cce
lera
tio
n
59
Re
tro
vir
us
es
gp
41
E
nve
lop
e g
lyco
pro
tein
41
(H
IV)
C1
q, fH
, C
D5
9
Dire
ct C
P a
ctiva
tio
n, R
ecru
itm
en
t o
f re
gu
lato
rs,
de
cre
ase
CD
59
e
xp
ressio
n
60-6
2
gp
12
0
En
ve
lop
e g
lyco
pro
tein
12
0 (
HIV
) M
BL
, fH
D
ire
ct L
P a
ctiva
tio
n,
Re
cru
itm
en
t o
f re
gu
lato
rs
62,
63
Ta
t T
ran
sa
ctiva
tor
of
tra
nscrip
tio
n (
HIV
) C
1-I
NH
In
du
ce
s C
1-I
NH
exp
ressio
n
64
Po
xv
iru
se
s
IMP
C
ow
po
x c
on
tro
l in
fla
mm
atio
n m
od
ula
tory
pro
tein
(C
ow
po
x V
iru
s)
C3
b, C
on
ve
rta
se
s
Mim
ics r
eg
ula
tors
(c
ofa
cto
r/d
eca
y a
cce
lera
tio
n)
65
MO
PIC
E
Mo
nke
yp
ox in
hib
ito
r o
f co
mp
lem
en
t e
nzym
es
(mo
nke
yp
ox v
iru
s)
C3
b
Mim
ics r
eg
ula
tors
(o
nly
co
facto
r a
ctivity)
66
SP
ICE
S
ma
llpo
x in
hib
ito
r o
f co
mp
lem
en
t e
nzym
es (
va
rio
la
viru
s)
C3
b, C
on
ve
rta
se
s
Mim
ics r
eg
ula
tors
(co
facto
r/d
eca
y a
cce
lera
tio
n)
67
VC
P
Va
ccin
ia v
iru
s c
om
ple
me
nt co
ntr
ol p
rote
in (
va
ccin
ia
viru
s)
C3
b, C
on
ve
rta
se
s
Mim
ics r
eg
ula
tors
(c
ofa
cto
r/d
eca
y a
cce
lera
tio
n)
68
Filo
vir
us
es
NS
1
No
n-s
tru
ctu
ral p
rote
in 1
(W
est
Nile
viru
s)
fH
Re
cru
itm
en
t o
f re
gu
lato
rs
69
Fu
ng
i
As
pe
rgillu
s f
um
iga
tus
n/a
U
nkn
ow
n fa
cto
r fH
, F
HL
-1, C
4B
P
RC
A r
ecru
itm
en
t 83
©!2008!N
atu
re P
ub
lis
hin
g G
rou
p

SU
PPLE
MEN
TAR
Y IN
FOR
MAT
ION
In
fo
rmat
pro
vid
ed b
y L
amb
ris
et
al.
(F
EB
RU
AR
Y 2
00
8)
NA
TU
RE
RE
VIE
WS
| MICRO
BIOLO
GY
www.nature.com/reviews/micro
Ca
nd
ida
alb
ica
ns
CR
AS
P-1
C
om
ple
me
nt re
gu
lato
r-a
cq
uir
ing
su
rfa
ce
pro
tein
1
fH, F
HL
-1, C
4B
P
Re
cru
itm
en
t o
f re
gu
lato
rs
84,
85
Gp
m1
p
Ph
osp
ho
gly
ce
rate
mu
tase
fH
, F
HL
-1
Re
cru
itm
en
t o
f re
gu
lato
rs
86
Pa
ras
ite
s
Ec
hin
oc
oc
cu
s s
pp
.
n/a
H
yd
atid
cyst w
all
fH
Re
cru
itm
en
t o
f re
gu
lato
rs
70
Ixo
de
s s
pp
.
IRA
C
Ixo
de
s r
icin
us a
nti-c
om
ple
me
nt p
rote
in
AP
co
nve
rta
se
D
eca
y a
cce
lera
tio
n
71
ISA
C
Ixo
de
s s
ca
pu
laris a
nti-c
om
ple
me
nt
pro
tein
A
P c
on
ve
rta
se
D
eca
y a
cce
lera
tio
n
72
On
ch
oc
erc
a s
pp
.
mf
Mic
rofila
ria
e
fH
Re
cru
itm
en
t o
f re
gu
lato
rs
73
Orn
ith
od
oro
s s
pp
.
Om
CI
Orn
ith
od
oro
s m
ou
ba
ta c
om
ple
me
nt
inh
ibito
r C
5
Bin
ds to
C5
(p
ote
ntia
lly b
locks b
ind
ing
to
C5
co
nve
rta
se
) 74
Sc
his
tos
om
a s
pp
.
CR
IT
Co
mp
lem
en
t C
2 r
ece
pto
r tr
isp
an
nin
g
C2
In
hib
its C
P c
on
ve
rta
se
fo
rma
tio
n
75
m2
8
28
kD
a m
em
bra
ne
se
rin
e p
rote
ase
iC
3b
C
lea
ve
s iC
3b
, re
str
icts
CR
3 b
ind
ing
76,
77
Pm
y e
P
ara
myo
sin
C8
, C
9,
C1
q,
IgG
P
reve
nts
MA
C f
orm
atio
n
De
cre
ase
s A
P a
ctiva
tio
n
78-8
1
Try
pa
no
so
ma
sp
p.
CR
IT
Co
mp
lem
en
t C
2 r
ece
pto
r tr
isp
an
nin
g
C2
In
hib
its C
P c
on
ve
rta
se
fo
rma
tio
n
75
T-D
AF
T
ryp
an
oso
ma
de
ca
y-a
cce
lera
tin
g fa
cto
r C
on
ve
rta
se
D
esta
bili
ze
s c
on
ve
rta
se
82
aE
hp h
as a
lso b
ee
n t
erm
ed e
xtr
acellu
lar
co
mple
ment-
bin
din
g p
rote
in (
Ecb).
bA
s H
ic is n
ot
a m
em
ber
of
the c
lassic
al P
sp
C f
am
iliy b
ut
a
Psp
C-l
ike p
rote
in,
it is lis
ted a
s s
epara
te p
rote
in.
cF
orm
er
na
mes:
cho
lin-b
indin
g p
rote
in (
Cbp
A),
Str
epto
coccus p
neu
mo
nia
e s
ecre
tory
Ig
A-
bin
din
g p
rote
in (
SpsA
) an
d p
neu
mococcal C
3-b
indin
g p
rote
in A
(P
bcA
). d
KP
C h
as a
lso b
ee
n t
erm
ed k
ap
osic
a.
e P
ara
myosin
has p
revio
usly
bee
n d
escrib
ed a
s S
chis
toso
me c
om
ple
ment
inhib
itor
pro
tein
1 (
SC
IP-1
). A
P,
altern
ative p
ath
way;
C1-I
NH
, C
1 e
ste
rase inhib
itor;
C4
BP
, C
4-
bin
din
g p
rote
in;
C5a
R,
C5
a r
ecepto
r; C
P,
cla
ssic
al path
way;
DA
F,
decay a
ccele
rating f
acto
r; f
H,
facto
r H
; F
HL,
facto
r H
-lik
e p
rote
in;
Ig,
imm
uno
glo
bulin
; M
BL,
man
nose-b
indin
g lectin;
MC
P,
me
mb
rane c
ofa
cto
r pro
tein
.
©!2008!N
atu
re P
ub
lis
hin
g G
rou
p

SU
PPLE
MEN
TAR
Y IN
FOR
MAT
ION
In
fo
rmat
pro
vid
ed b
y L
amb
ris
et
al.
(F
EB
RU
AR
Y 2
00
8)
NA
TU
RE
RE
VIE
WS
| MICRO
BIOLO
GY
www.nature.com/reviews/micro
Su
pp
lem
en
tary
in
form
ati
on
S1
(ta
ble
): R
efe
ren
ce
s
1.
Asa
ka
wa
, R
. e
t a
l. O
ute
r m
em
bra
ne
pro
tein
1
00
, a
ve
rsa
tile
viru
len
ce
fa
cto
r o
f A
ctin
ob
acill
us a
ctin
om
yce
tem
co
mita
ns.
Mo
l.
Mic
rob
iol. 5
0,
11
25
-11
39
(2
00
3).
2
. B
erg
ga
rd, K
., J
oh
nsso
n, E
., M
oo
i, F
.R. &
Lin
da
hl, G
. B
ord
ete
lla p
ert
ussis
bin
ds th
e
hu
ma
n c
om
ple
me
nt re
gu
lato
r C
4B
P: ro
le o
f fila
me
nto
us h
em
ag
glu
tin
in.
Infe
ct.
Im
mu
n.
65
, 3
63
8-3
64
3 (
19
97
).
3.
Kra
iczy, P
., S
ke
rka
, C
., K
irsch
fin
k, M
., B
rad
e,
V. &
Zip
fel, P
.F. Im
mu
ne
eva
sio
n o
f B
orr
elia
b
urg
do
rfe
ri b
y a
cq
uis
itio
n o
f h
um
an
co
mp
lem
en
t re
gu
lato
rs F
HL
-1/r
eco
ne
ctin
an
d
Fa
cto
r H
. E
ur.
J.
Imm
un
ol. 3
1,
16
74
-16
84
(2
00
1).
4
. H
ellw
ag
e, J.
et a
l. T
he
co
mp
lem
en
t re
gu
lato
r fa
cto
r H
bin
ds to
th
e s
urf
ace
pro
tein
Osp
E o
f B
orr
elia
bu
rgd
orf
eri.
J. B
iol. C
he
m.
27
6, 8
42
7-
84
35
(2
00
1).
5
. S
teve
nso
n, B
., E
l-H
ag
e,
N.,
Hin
es,
M.A
., M
ille
r,
J.C
. &
Ba
bb
, K
. D
iffe
ren
tia
l b
ind
ing
of h
ost
co
mp
lem
en
t in
hib
ito
r fa
cto
r H
by B
orr
elia
b
urg
do
rfe
ri E
rp s
urf
ace
pro
tein
s: a
po
ssib
le
me
ch
an
ism
un
de
rlyin
g th
e e
xp
an
siv
e h
ost
ran
ge
of L
ym
e d
ise
ase
sp
iro
ch
ete
s.
Infe
ct.
Im
mu
n.
70
, 4
91
-49
7 (
20
02
).
6.
Pa
usa
, M
. e
t a
l. S
eru
m-r
esis
tan
t str
ain
s o
f B
orr
elia
bu
rgd
orf
eri e
va
de
co
mp
lem
en
t-m
ed
iate
d k
illin
g b
y e
xp
ressin
g a
CD
59
-lik
e
co
mp
lem
en
t in
hib
ito
ry m
ole
cu
le. J.
Imm
un
ol.
17
0, 3
21
4-3
22
2 (
20
03
).
7.
Pra
sa
da
rao
, N
.V., B
lom
, A
.M.,
Vill
ou
tre
ix, B
.O.
& L
insa
ng
an
, L
.C. A
no
ve
l in
tera
ctio
n o
f o
ute
r m
em
bra
ne
pro
tein
A w
ith
C4
b b
ind
ing
pro
tein
m
ed
iate
s s
eru
m r
esis
tan
ce
of E
sch
erich
ia c
oli
K1
. J.
Imm
un
ol. 1
69
, 6
35
2-6
36
0 (
20
02
).
8.
La
the
m, W
.W., B
erg
sb
ake
n, T
. &
We
lch
, R
.A.
Po
ten
tia
tio
n o
f C
1 e
ste
rase
in
hib
ito
r b
y S
tcE
, a
m
eta
llop
rote
ase
se
cre
ted
by E
sch
erich
ia c
oli
O1
57
:H7
. T
he
Jo
urn
al o
f e
xp
erim
en
tal
me
dic
ine
19
9, 1
07
7-1
08
7 (
20
04
).
9.
Pra
mo
on
jag
o, P
. e
t a
l. R
ole
of
Tra
T p
rote
in,
an
a
ntico
mp
lem
en
tary
pro
tein
pro
du
ce
d in
E
sch
erich
ia c
oli
by R
10
0 fa
cto
r, in
se
rum
re
sis
tan
ce
. J.
Imm
un
ol. 1
48
, 8
27
-83
6 (
19
92
).
10
. F
rib
erg
, N
., C
arl
so
n, P
., M
eri
, S
. &
Ja
rva
, H
. F
uso
ba
cte
riu
m n
ecro
ph
oru
m,
the
ca
use
of
Le
mie
rre
's s
yn
dro
me
, b
ind
s fa
cto
r H
. M
ol.
Imm
un
ol. 4
4, 3
97
6 (
20
07
).
11
. H
alls
tro
m, T
., J
arv
a,
H.,
Rie
sb
eck, K
. &
Blo
m,
A.M
. In
tera
ctio
n w
ith
C4
b-b
ind
ing
pro
tein
co
ntr
ibu
tes to
no
nty
pe
ab
le H
ae
mo
ph
ilus
influ
en
za
e s
eru
m r
esis
tan
ce
. J.
Imm
un
ol. 1
78
, 6
35
9-6
36
6 (
20
07
).
12
. H
alls
tro
m, T
., Z
ipfe
l, P
.F., B
lom
, A
.M.,
F
ors
gre
n, A
. &
Rie
sb
eck, K
. H
ae
mo
ph
ilus
influ
en
za
e in
tera
cts
with
th
e h
um
an
co
mp
lem
en
t in
hib
ito
r fa
cto
r H
. M
ole
cu
lar
Imm
un
olo
gy 4
4, 3
92
6 (
20
07
).
13
. N
ord
str
om
, T
., B
lom
, A
.M.,
Fo
rsg
ren
, A
. &
R
iesb
eck, K
. T
he
em
erg
ing
pa
tho
ge
n M
ora
xe
lla
ca
tarr
ha
lis in
tera
cts
with
co
mp
lem
en
t in
hib
ito
r C
4b
bin
din
g p
rote
in th
rou
gh
ub
iqu
ito
us s
urf
ace
p
rote
ins A
1 a
nd
A2
. J.
Imm
un
ol. 1
73
, 4
59
8-
46
06
(2
00
4).
14
. M
ad
ico
, G
. e
t a
l. T
he
me
nin
go
co
cca
l va
ccin
e
ca
nd
ida
te G
NA
18
70
bin
ds th
e c
om
ple
me
nt
reg
ula
tory
pro
tein
fa
cto
r H
an
d e
nh
an
ce
s s
eru
m
resis
tan
ce
. J.
Imm
un
ol. 1
77
, 5
01
-51
0 (
20
06
).
15
. R
am
, S
. e
t a
l. B
ind
ing
of
C4
b-b
ind
ing
pro
tein
to
p
orin
: a
mo
lecu
lar
me
ch
an
ism
of se
rum
re
sis
tan
ce
of N
eis
se
ria
go
no
rrh
oe
ae
. T
he
Jo
urn
al o
f e
xp
erim
en
tal m
ed
icin
e 1
93
, 2
81
-29
5
(20
01
).
16
. R
am
, S
. e
t a
l. T
he
co
ntr
astin
g m
ech
an
ism
s o
f se
rum
re
sis
tan
ce
of
Ne
isse
ria
go
no
rrh
oe
ae
an
d
gro
up
B N
eis
se
ria
me
nin
gitid
is.
Mo
l. Im
mu
no
l.
36
, 9
15
-92
8 (
19
99
).
17
. K
alls
tro
m, H
., L
isze
wski, M
.K.,
Atk
inso
n, J.P
. &
Jo
nsso
n, A
.B. M
em
bra
ne
co
facto
r p
rote
in
(MC
P o
r C
D4
6)
is a
ce
llula
r p
ilus r
ece
pto
r fo
r p
ath
og
en
ic N
eis
se
ria
. M
ol. M
icro
bio
l. 2
5, 6
39
-6
47
(1
99
7).
1
8.
Sch
en
ke
in, H
.A.,
Fle
tch
er,
H.M
., B
od
na
r, M
. &
M
acrin
a, F
.L.
Incre
ase
d o
pso
niz
atio
n o
f a
prt
H-
de
fective
mu
tan
t o
f P
orp
hyro
mo
na
s g
ing
iva
lis
W8
3 is c
au
se
d b
y r
ed
uce
d d
eg
rad
atio
n o
f co
mp
lem
en
t-d
erive
d o
pso
nin
s. J.
Imm
un
ol.
15
4, 5
33
1-5
33
7 (
19
95
).
19
. H
on
g, Y
.Q.
& G
he
bre
hiw
et,
B.
Effe
ct
of
Pse
ud
om
on
as a
eru
gin
osa
ela
sta
se
an
d
alk
alin
e p
rote
ase
on
se
rum
co
mp
lem
en
t a
nd
is
ola
ted
co
mp
on
en
ts C
1q
an
d C
3. C
lin.
Imm
un
ol. Im
mu
no
pa
tho
l. 6
2,
13
3-1
38
(1
99
2).
2
0.
Ku
ne
rt, A
. e
t a
l. I
mm
un
e E
va
sio
n o
f th
e H
um
an
P
ath
og
en
Pse
ud
om
on
as a
eru
gin
osa
: E
lon
ga
tio
n F
acto
r T
uf
Is a
Fa
cto
r H
an
d
Pla
sm
ino
ge
n B
ind
ing
Pro
tein
. J.
Imm
un
ol. 1
79
, 2
97
9-2
98
8 (
20
07
).
21
. O
da
, T
. e
t a
l. I
na
ctiva
tio
n o
f ch
em
ota
ctic a
ctivity
of C
5a
by th
e s
err
atia
l 5
6-k
ilod
alto
n p
rote
ase
.
Infe
ct.
Im
mu
n. 5
8,
12
69
-12
72
(1
99
0).
2
2.
Mo
lla, A
., A
ka
ike
, T
. &
Ma
ed
a,
H.
Ina
ctiva
tio
n o
f va
rio
us p
rote
ina
se
in
hib
ito
rs a
nd
th
e
co
mp
lem
en
t syste
m in
hu
ma
n p
lasm
a b
y th
e
56
-kilo
da
lto
n p
rote
ina
se
fro
m S
err
atia
m
arc
esce
ns. In
fect.
Im
mu
n. 5
7,
18
68
-18
71
(1
98
9).
©!2008!N
atu
re P
ub
lis
hin
g G
rou
p

SU
PPLE
MEN
TAR
Y IN
FOR
MAT
ION
In
fo
rmat
pro
vid
ed b
y L
amb
ris
et
al.
(F
EB
RU
AR
Y 2
00
8)
NA
TU
RE
RE
VIE
WS
| MICRO
BIOLO
GY
www.nature.com/reviews/micro
23
. d
e H
aa
s, C
.J.
et a
l. C
he
mo
taxis
in
hib
ito
ry
pro
tein
of
Sta
ph
ylo
co
ccu
s a
ure
us, a
ba
cte
ria
l a
ntiin
fla
mm
ato
ry a
ge
nt. T
he
Jo
urn
al o
f e
xp
erim
en
tal m
ed
icin
e 1
99
, 6
87
-69
5 (
20
04
).
24
. H
am
me
l, M
. e
t a
l. A
str
uctu
ral b
asis
fo
r
co
mp
lem
en
t in
hib
itio
n b
y S
tap
hylo
co
ccu
s
au
reu
s. N
at.
Im
mu
no
l. 8
, 4
30
-43
7 (
20
07
).
25
. L
ee
, L
.Y.
et a
l. I
nh
ibitio
n o
f co
mp
lem
en
t a
ctiva
tio
n b
y a
se
cre
ted
Sta
ph
ylo
co
ccu
s a
ure
us
pro
tein
. J. In
fect.
Dis
. 1
90
, 5
71
-57
9 (
20
04
).
26
. Jo
ng
eriu
s, I.
et a
l. S
tap
hylo
co
cca
l co
mp
lem
en
t e
va
sio
n b
y v
ario
us c
on
ve
rta
se
-blo
ckin
g
mo
lecu
les. T
he
Jo
urn
al o
f e
xp
erim
en
tal
me
dic
ine
20
4, 2
46
1-2
47
1 (
20
07
).
27
. H
am
me
l, M
. e
t a
l. C
ha
racte
riza
tio
n o
f E
hp
: A
se
cre
ted
co
mp
lem
en
t in
hib
ito
ry p
rote
in f
rom
S
tap
hylo
co
ccu
s a
ure
us. J. B
iol. C
he
m.
(20
07
).
28
. R
oo
ijakke
rs, S
.H., v
an
Wa
me
l, W
.J.,
Ru
yke
n,
M.,
va
n K
esse
l, K
.P. &
va
n S
trijp
, J.A
. A
nti-
op
so
nic
pro
pe
rtie
s o
f sta
ph
ylo
kin
ase
. M
icro
be
s
Infe
ct.
7, 4
76
-48
4 (
20
05
).
29
. Z
ha
ng
, L
., J
aco
bsso
n, K
., V
asi, J
., L
ind
be
rg, M
. &
Fry
kb
erg
, L
. A
se
co
nd
Ig
G-b
ind
ing
pro
tein
in
S
tap
hylo
co
ccu
s a
ure
us. M
icro
bio
log
y 1
44
( P
t 4
), 9
85
-99
1 (
19
98
).
30
. R
oo
ijakke
rs, S
.H. e
t a
l. Im
mu
ne
eva
sio
n b
y a
sta
ph
ylo
co
cca
l co
mp
lem
en
t in
hib
ito
r th
at
acts
o
n C
3 c
on
ve
rta
se
s. N
at.
Im
mu
no
l. 6
, 9
20
-92
7
(20
05
).
31
. K
ron
va
ll, G
. &
Ge
wu
rz,
H.
Activa
tio
n a
nd
in
hib
itio
n o
f Ig
G m
ed
iate
d c
om
ple
me
nt
fixa
tio
n
by s
tap
hylo
co
cca
l p
rote
in A
. C
lin.
Exp
. Im
mu
no
l. 7
, 2
11
-22
0 (
19
70
).
32
. L
an
gle
y, R
. e
t a
l. T
he
sta
ph
ylo
co
cca
l su
pe
ran
tig
en
-lik
e p
rote
in 7
bin
ds I
gA
an
d
co
mp
lem
en
t C
5 a
nd
in
hib
its I
gA
-Fc a
lph
a R
I b
ind
ing
an
d s
eru
m k
illin
g o
f b
acte
ria
. J.
Imm
un
ol. 1
74
, 2
92
6-2
93
3 (
20
05
).
33
. A
resch
ou
g, T
., S
talh
am
ma
r-C
arl
em
alm
, M
.,
Ka
rlsso
n, I.
& L
ind
ah
l, G
. S
tre
pto
co
cca
l b
eta
pro
tein
ha
s s
ep
ara
te b
ind
ing
site
s f
or
hu
ma
n
facto
r H
an
d Ig
A-F
c.
J. B
iol. C
he
m.
27
7, 1
26
42
-1
26
48
(2
00
2).
3
4.
Pa
nd
irip
ally
, V
., W
ei, L
., S
ke
rka
, C
., Z
ipfe
l, P
.F.
& C
ue
, D
. R
ecru
itm
en
t o
f co
mp
lem
en
t fa
cto
r H
-
like
pro
tein
1 p
rom
ote
s in
tra
ce
llula
r in
va
sio
n b
y
gro
up
A s
tre
pto
co
cci. In
fect.
Im
mu
n.
71
, 7
11
9-
71
28
(2
00
3).
3
5.
Ja
nu
lczyk, R
., Ia
nn
elli
, F
., S
joh
olm
, A
.G.,
Po
zzi,
G. &
Bjo
rck, L
. H
ic, a
no
ve
l su
rfa
ce
pro
tein
of
Str
ep
toco
ccu
s p
ne
um
on
iae
th
at in
terf
ere
s w
ith
co
mp
lem
en
t fu
nctio
n.
J. B
iol. C
he
m.
27
5,
37
25
7-3
72
63
(2
00
0).
36
. vo
n P
aw
el-R
am
min
ge
n,
U. &
Bjo
rck,
L.
Ide
S
an
d S
pe
B: im
mu
no
glo
bu
lin-d
eg
rad
ing
cyste
ine
p
rote
ina
se
s o
f S
tre
pto
co
ccu
s p
yo
ge
ne
s. C
urr
. O
pin
. M
icro
bio
l. 6
, 5
0-5
5 (
20
03
).
37
. Ja
rva
, H
., J
okira
nta
, T
.S., W
urz
ne
r, R
. &
Me
ri,
S. C
om
ple
me
nt
resis
tan
ce
me
ch
an
ism
s o
f str
ep
toco
cci. M
ol. I
mm
un
ol. 4
0,
95
-10
7 (
20
03
).
38
. M
itch
ell,
T.J
., A
nd
rew
, P
.W.,
Sa
un
de
rs,
F.K
.,
Sm
ith
, A
.N. &
Bo
uln
ois
, G
.J. C
om
ple
me
nt
activa
tio
n a
nd
an
tib
od
y b
ind
ing
by p
ne
um
oly
sin
via
a r
eg
ion
of th
e to
xin
ho
mo
log
ou
s to
a
hu
ma
n a
cu
te-p
ha
se
pro
tein
. M
ol. M
icro
bio
l. 5
, 1
88
3-1
88
8 (
19
91
).
39
. P
ato
n, J.C
., R
ow
an
-Ke
lly,
B. &
Fe
rra
nte
, A
. A
ctiva
tio
n o
f h
um
an
co
mp
lem
en
t b
y t
he
p
ne
um
oco
cca
l to
xin
pn
eu
mo
lysin
. In
fect.
Imm
un
. 4
3, 1
08
5-1
08
7 (
19
84
).
40
. R
en
, B
. e
t a
l. T
he
vir
ule
nce
fu
nctio
n o
f S
tre
pto
co
ccu
s p
ne
um
on
iae
su
rfa
ce
pro
tein
A
invo
lve
s in
hib
itio
n o
f co
mp
lem
en
t a
ctiva
tio
n a
nd
im
pa
irm
en
t o
f co
mp
lem
en
t re
ce
pto
r-m
ed
iate
d
pro
tectio
n. J. Im
mu
no
l. 1
73
, 7
50
6-7
51
2 (
20
04
).
41
. D
ave
, S
., B
roo
ks-W
alte
r, A
., P
an
gb
urn
, M
.K. &
M
cD
an
iel, L
.S. P
sp
C,
a p
ne
um
oco
cca
l su
rfa
ce
p
rote
in, b
ind
s h
um
an
fa
cto
r H
. In
fect.
Im
mu
n.
69
, 3
43
5-3
43
7 (
20
01
).
42
. K
err
, A
.R. e
t a
l. T
he
co
ntr
ibu
tio
n o
f P
sp
C to
p
ne
um
oco
cca
l viru
len
ce
va
rie
s b
etw
ee
n s
tra
ins
an
d is a
cco
mp
lish
ed
by b
oth
co
mp
lem
en
t e
va
sio
n a
nd
co
mp
lem
en
t-in
de
pe
nd
en
t m
ech
an
ism
s. In
fect.
Im
mu
n. 7
4, 5
31
9-5
32
4
(20
06
).
43
. A
ng
el, C
.S., R
uze
k, M
. &
Ho
ste
tte
r, M
.K.
De
gra
da
tio
n o
f C
3 b
y S
tre
pto
co
ccu
s
pn
eu
mo
nia
e. J. In
fect. D
is. 1
70
, 6
00
-60
8
(19
94
).
44
. C
he
ng
, Q
., F
inke
l, D
. &
Ho
ste
tte
r, M
.K. N
ove
l p
urifica
tio
n s
ch
em
e a
nd
fu
nctio
ns fo
r a
C3
-b
ind
ing
pro
tein
fro
m S
tre
pto
co
ccu
s
pn
eu
mo
nia
e. B
ioch
em
istr
y 3
9,
54
50
-54
57
(2
00
0).
4
5.
Ch
mo
ury
gu
ina
, I.
, S
uvo
rov,
A.,
Fe
rrie
ri,
P. &
C
lea
ry, P
.P.
Co
nse
rva
tio
n o
f th
e C
5a
pe
ptid
ase
g
en
es in
gro
up
A a
nd
B s
tre
pto
co
cci. I
nfe
ct.
Im
mu
n.
64
, 2
38
7-2
39
0 (
19
96
).
46
. W
exle
r, D
.E.,
Ch
en
ow
eth
, D
.E. &
Cle
ary
, P
.P.
Me
ch
an
ism
of a
ctio
n o
f th
e g
rou
p A
str
ep
toco
cca
l C
5a
in
activa
tor.
Pro
c. N
atl.
Aca
d.
Sci. U
.S.A
. 8
2, 8
14
4-8
14
8 (
19
85
).
47
. W
exle
r, D
.E.,
Ne
lso
n, R
.D. &
Cle
ary
, P
.P.
Hu
ma
n n
eu
tro
ph
il ch
em
ota
ctic r
esp
on
se
to
g
rou
p A
str
ep
toco
cci: b
acte
ria
-me
dia
ted
in
terf
ere
nce
with
co
mp
lem
en
t-d
eri
ve
d
ch
em
ota
ctic fa
cto
rs.
Infe
ct. I
mm
un
. 3
9, 2
39
-24
6
(19
83
).
48
. A
ke
sso
n, P
., S
joh
olm
, A
.G.
& B
jorc
k, L
. P
rote
in
SIC
, a
no
ve
l e
xtr
ace
llula
r p
rote
in o
f S
tre
pto
co
ccu
s p
yo
ge
ne
s in
terf
erin
g w
ith
co
mp
lem
en
t fu
nctio
n.
J. B
iol. C
he
m.
27
1, 1
08
1-
10
88
(1
99
6).
4
9.
Fe
rnie
-Kin
g,
B.A
. e
t a
l. S
tre
pto
co
cca
l in
hib
ito
r o
f co
mp
lem
en
t (S
IC)
inh
ibits t
he
me
mb
ran
e
atta
ck c
om
ple
x b
y p
reve
ntin
g u
pta
ke
of
C5
67
o
nto
ce
ll m
em
bra
ne
s. Im
mu
no
log
y 1
03
, 3
90
-
39
8 (
20
01
).
©!2008!N
atu
re P
ub
lis
hin
g G
rou
p

SU
PPLE
MEN
TAR
Y IN
FOR
MAT
ION
In
fo
rmat
pro
vid
ed b
y L
amb
ris
et
al.
(F
EB
RU
AR
Y 2
00
8)
NA
TU
RE
RE
VIE
WS
| MICRO
BIOLO
GY
www.nature.com/reviews/micro
50
. T
sa
o, N
. e
t a
l. S
tre
pto
co
cca
l p
yro
ge
nic
exo
toxin
B
cle
ave
s p
rop
erd
in a
nd
in
hib
its c
om
ple
me
nt-
me
dia
ted
op
so
no
ph
ag
ocyto
sis
. B
ioch
em
. B
iop
hys. R
es. C
om
mu
n. 339
, 7
79
-78
4 (
20
06
).
51
. B
jorc
k, L
. &
Kro
nva
ll, G
. P
uri
fica
tio
n a
nd
so
me
pro
pe
rtie
s o
f str
ep
toco
cca
l p
rote
in G
, a
no
ve
l Ig
G-b
ind
ing
re
ag
en
t. J
. Im
mu
no
l. 133
, 9
69
-97
4
(19
84
).
52
. C
hin
a, B
., S
ory
, M
.P.,
N'G
uye
n, B
.T.,
De
B
ruye
re, M
. &
Co
rne
lis, G
.R.
Ro
le o
f th
e Y
ad
A
pro
tein
in
pre
ve
ntio
n o
f o
pso
niz
atio
n o
f Y
ers
inia
e
nte
roco
litic
a b
y C
3b
mo
lecu
les.
Infe
ct.
Im
mu
n.
61
, 3
12
9-3
13
6 (
19
93
).
53
. K
osta
va
sili
, I.
et a
l. M
ech
an
ism
of
co
mp
lem
en
t in
activa
tio
n b
y g
lyco
pro
tein
C o
f h
erp
es s
imp
lex
viru
s. J.
Imm
un
ol. 158
, 1
76
3-1
77
1 (
19
97
).
54
. Jo
hn
so
n, D
.C. &
Fe
en
str
a, V
. Id
en
tifica
tio
n o
f a
n
ove
l h
erp
es s
imp
lex v
iru
s t
yp
e 1
-in
du
ce
d
gly
co
pro
tein
wh
ich
co
mp
lexe
s w
ith
gE
an
d
bin
ds im
mu
no
glo
bu
lin.
J. V
iro
l. 61
, 2
20
8-2
21
6
(19
87
).
55
. P
ara
, M
.F.,
Ba
ucke
, R
.B. &
Sp
ea
r, P
.G.
Gly
co
pro
tein
gE
of
he
rpe
s s
imp
lex v
iru
s t
yp
e 1
: e
ffe
cts
of
an
ti-g
E o
n v
irio
n in
fectivity a
nd
on
viru
s-in
du
ce
d fc-b
ind
ing
re
ce
pto
rs.
J. V
iro
l. 41
, 1
29
-13
6 (
19
82
).
56
. A
tala
y, R
. e
t a
l. Id
en
tifica
tio
n a
nd
exp
ressio
n o
f h
um
an
cyto
me
ga
loviru
s tra
nscrip
tio
n u
nits
co
din
g fo
r tw
o d
istin
ct F
cg
am
ma
re
ce
pto
r
ho
mo
log
s. J. V
iro
l. 76
, 8
59
6-8
60
8 (
20
02
).
57
. L
ille
y, B
.N.,
Plo
eg
h,
H.L
. &
Tir
ab
assi, R
.S.
Hu
ma
n c
yto
me
ga
loviru
s o
pe
n r
ea
din
g f
ram
e
TR
L1
1/IR
L1
1 e
nco
de
s a
n im
mu
no
glo
bu
lin G
F
c-b
ind
ing
pro
tein
. J.
Viro
l. 75
, 1
12
18
-11
22
1
(20
01
).
58
. L
itw
in,
V.,
Ja
ckso
n, W
. &
Gro
se
, C
. R
ece
pto
r p
rop
ert
ies o
f tw
o v
arice
lla-z
oste
r vir
us
gly
co
pro
tein
s, g
pI a
nd
gp
IV, h
om
olo
go
us to
he
rpe
s s
imp
lex v
iru
s g
E a
nd
gI.
J. V
iro
l. 66
, 3
64
3-3
65
1 (
19
92
).
59
. S
pill
er,
O.B
. e
t a
l. C
om
ple
me
nt
reg
ula
tio
n b
y
Ka
po
si's
sa
rco
ma
-asso
cia
ted
he
rpe
sviru
s
OR
F4
pro
tein
. J. V
iro
l. 77
, 5
92
-59
9 (
20
03
).
60
. C
ho
ng
, Y
.H.
& L
ee
, M
.J.
Exp
ressio
n o
f co
mp
lem
en
t in
hib
ito
r p
rote
in C
D5
9 in
hu
ma
n
ne
uro
na
l a
nd
glia
l ce
ll lin
es t
rea
ted
with
HIV
-1
gp
41
pe
ptid
es. J. N
eu
rovir
ol. 6
, 5
1-6
0 (
20
00
).
61
. E
be
nb
ich
ler,
C.F
. e
t a
l. H
um
an
im
mu
no
de
ficie
ncy v
iru
s t
yp
e 1
activa
tes t
he
cla
ssic
al p
ath
wa
y o
f co
mp
lem
en
t b
y d
ire
ct
C1
b
ind
ing
th
rou
gh
sp
ecific
site
s in
th
e
tra
nsm
em
bra
ne
gly
co
pro
tein
gp
41
. T
he
Jo
urn
al
of e
xp
erim
en
tal m
ed
icin
e 174
, 1
41
7-1
42
4
(19
91
).
62
. S
toib
er,
H.,
Sch
ne
ide
r, R
., J
an
ato
va
, J.
&
Die
rich
, M
.P. H
um
an
co
mp
lem
en
t p
rote
ins C
3b
, C
4b
, fa
cto
r H
an
d p
rop
erd
in r
ea
ct w
ith
sp
ecific
site
s in
gp
12
0 a
nd
gp
41
, th
e e
nve
lop
e p
rote
ins
of H
IV-1
. Im
mu
no
bio
log
y 193
, 9
8-1
13
(1
99
5).
6
3.
Th
iele
ns, N
.M.,
Ta
cn
et-
De
lorm
e, P
. &
Arla
ud
, G
.J.
Inte
ractio
n o
f C
1q
an
d m
an
na
n-b
ind
ing
lectin
with
vir
use
s. Im
mu
no
bio
log
y 205
, 5
63
-5
74
(2
00
2).
6
4.
Izm
ailo
va
, E
. e
t a
l. H
IV-1
Ta
t re
pro
gra
ms
imm
atu
re d
en
dritic c
ells
to
exp
ress
ch
em
oa
ttra
cta
nts
fo
r a
ctiva
ted
T c
ells
an
d
ma
cro
ph
ag
es. N
at. M
ed
. 9
, 1
91
-19
7 (
20
03
).
65
. M
ille
r, C
.G.,
Sh
ch
elk
un
ov, S
.N. &
Ko
twa
l, G
.J.
Th
e c
ow
po
x v
iru
s-e
nco
de
d h
om
olo
g o
f th
e
va
ccin
ia v
iru
s c
om
ple
me
nt co
ntr
ol p
rote
in is a
n
infla
mm
atio
n m
od
ula
tory
pro
tein
. V
iro
log
y 229
, 1
26
-13
3 (
19
97
).
66
. L
isze
wski, M
.K.
et a
l. S
tru
ctu
re a
nd
re
gu
lato
ry
pro
file
of
the
mo
nke
yp
ox in
hib
ito
r o
f co
mp
lem
en
t: c
om
pa
riso
n t
o h
om
olo
gs in
va
ccin
ia a
nd
va
rio
la a
nd
evid
en
ce
fo
r d
ime
r fo
rma
tio
n. J.
Imm
un
ol. 176
, 3
72
5-3
73
4 (
20
06
).
67
. R
ose
ng
ard
, A
.M.,
Liu
, Y
., N
ie, Z
. &
Jim
en
ez, R
.
Va
rio
la v
iru
s im
mu
ne
eva
sio
n d
esig
n:
exp
ressio
n o
f a
hig
hly
eff
icie
nt
inh
ibito
r o
f
hu
ma
n c
om
ple
me
nt. P
roc. N
atl.
Aca
d. S
ci.
U.S
.A. 99
, 8
80
8-8
81
3 (
20
02
).
68
. M
cK
en
zie
, R
., K
otw
al, G
.J.,
Mo
ss, B
., H
am
me
r,
C.H
. &
Fra
nk,
M.M
. R
eg
ula
tio
n o
f co
mp
lem
en
t a
ctivity b
y v
accin
ia v
iru
s c
om
ple
me
nt-
co
ntr
ol
pro
tein
. J. In
fect.
Dis
. 166
, 1
24
5-1
25
0 (
19
92
).
69
. C
hu
ng
, K
.M.
et
al. W
est N
ile v
iru
s n
on
str
uctu
ral
pro
tein
NS
1 in
hib
its c
om
ple
me
nt a
ctiva
tio
n b
y
bin
din
g th
e r
eg
ula
tory
pro
tein
fa
cto
r H
. P
roc.
Na
tl.
Aca
d. S
ci. U
.S.A
. 103
, 1
91
11
-19
11
6
(20
06
).
70
. D
iaz, A
., F
err
eira
, A
. &
Sim
, R
.B. C
om
ple
me
nt
eva
sio
n b
y E
ch
ino
co
ccu
s g
ran
ulo
su
s:
se
qu
estr
atio
n o
f h
ost
facto
r H
in
th
e h
yd
atid
cyst w
all.
J.
Imm
un
ol. 158
, 3
77
9-3
78
6 (
19
97
).
71
. D
aix
, V
. e
t a
l. I
xo
de
s tic
ks b
elo
ng
ing
to
th
e
Ixo
de
s r
icin
us c
om
ple
x e
nco
de
a fa
mily
of
an
tico
mp
lem
en
t p
rote
ins.
Inse
ct. M
ol. B
iol. 16
, 1
55
-16
6 (
20
07
).
72
. V
ale
nzu
ela
, J.G
., C
ha
rla
b, R
., M
ath
er,
T.N
. &
R
ibe
iro
, J.M
. P
uri
fica
tio
n, clo
nin
g,
an
d
exp
ressio
n o
f a
no
ve
l sa
liva
ry a
ntico
mp
lem
en
t p
rote
in f
rom
th
e t
ick,
Ixo
de
s s
ca
pu
laris. J. B
iol.
Ch
em
. 275
, 1
87
17
-18
72
3 (
20
00
).
73
. M
eri,
T. e
t a
l. O
nch
oce
rca
vo
lvu
lus m
icro
fila
ria
e
avo
id c
om
ple
me
nt a
tta
ck b
y d
ire
ct b
ind
ing
of
facto
r H
. J.
Infe
ct.
Dis
. 185
, 1
78
6-1
79
3 (
20
02
).
74
. N
un
n, M
.A. e
t a
l. C
om
ple
me
nt
inh
ibito
r o
f C
5
activa
tio
n f
rom
th
e s
oft
tic
k O
rnith
od
oro
s
mo
ub
ata
. J.
Imm
un
ol. 174
, 2
08
4-2
09
1 (
20
05
).
75
. In
al, J
.M.
& S
im, R
.B.
A S
ch
isto
so
ma
pro
tein
, S
h-T
OR
, is
a n
ove
l in
hib
ito
r o
f co
mp
lem
en
t w
hic
h b
ind
s h
um
an
C2
. F
EB
S L
ett. 470
, 1
31
-1
34
(2
00
0).
7
6.
Gh
en
dle
r, Y
., P
ariza
de
, M
., A
rno
n,
R.,
M
cK
err
ow
, J.H
. &
Fis
he
lso
n, Z
. S
ch
isto
so
ma
m
an
so
ni: e
vid
en
ce
fo
r a
28
-kD
a m
em
bra
ne
-a
nch
ore
d p
rote
ase
on
sch
isto
so
mu
la. E
xp
.
Pa
rasito
l. 83
, 7
3-8
2 (
19
96
).
©!2008!N
atu
re P
ub
lis
hin
g G
rou
p

SU
PPLE
MEN
TAR
Y IN
FOR
MAT
ION
In
fo
rmat
pro
vid
ed b
y L
amb
ris
et
al.
(F
EB
RU
AR
Y 2
00
8)
NA
TU
RE
RE
VIE
WS
| MICRO
BIOLO
GY
www.nature.com/reviews/micro
77
. M
ariko
vsky, M
., A
rno
n,
R.
& F
ish
els
on
, Z
. S
ch
isto
so
ma
ma
nso
ni: lo
ca
liza
tio
n o
f th
e 2
8
kD
a s
ecre
ted
pro
tea
se
in
ce
rca
ria
. P
ara
site
Imm
un
ol. 12
, 3
89
-40
1 (
19
90
).
78
. D
en
g, J., G
old
, D
., L
oV
erd
e, P
.T.
& F
ish
els
on
,
Z. In
hib
itio
n o
f th
e c
om
ple
me
nt m
em
bra
ne
a
tta
ck c
om
ple
x b
y S
ch
isto
so
ma
ma
nso
ni
pa
ram
yo
sin
. In
fect.
Im
mu
n. 71
, 6
40
2-6
41
0
(20
03
).
79
. L
acle
tte
, J.P
. e
t a
l. P
ara
myo
sin
in
hib
its
co
mp
lem
en
t C
1. J. Im
mu
no
l. 148
, 1
24
-12
8
(19
92
).
80
. M
cIn
tosh
, R
.S.,
Jo
ne
s, F
.M., D
un
ne
, D
.W.,
McK
err
ow
, J.H
. &
Ple
ass,
R.J
. C
ha
racte
riza
tio
n
of im
mu
no
glo
bu
lin b
ind
ing
by s
ch
isto
so
me
s.
Pa
rasite
Im
mu
no
l. 28
, 4
07
-41
9 (
20
06
).
81
. P
ariza
de
, M
., A
rno
n, R
., L
ach
ma
nn
, P
.J. &
F
ish
els
on
, Z
. F
un
ctio
na
l a
nd
an
tig
en
ic
sim
ilari
tie
s b
etw
ee
n a
94
-kD
pro
tein
of
Sch
isto
so
ma
ma
nso
ni (S
CIP
-1)
an
d h
um
an
C
D5
9. T
he
Jo
urn
al o
f e
xp
erim
en
tal m
ed
icin
e
179
, 1
62
5-1
63
6 (
19
94
).
82
. T
am
bo
urg
i, D
.V. e
t a
l. A
pa
rtia
l cD
NA
clo
ne
of
tryp
om
astig
ote
de
ca
y-a
cce
lera
tin
g fa
cto
r (T
-D
AF
), a
de
ve
lop
me
nta
lly r
eg
ula
ted
co
mp
lem
en
t in
hib
ito
r o
f T
ryp
an
oso
ma
cru
zi, h
as g
en
etic a
nd
fu
nctio
na
l sim
ilari
tie
s t
o th
e h
um
an
co
mp
lem
en
t
inh
ibito
r D
AF
. In
fect.
Im
mu
n. 61
, 3
65
6-3
66
3
(19
93
).
83
. V
og
l, G
. e
t a
l. Im
mu
ne
eva
sio
n b
y a
cq
uis
itio
n o
f co
mp
lem
en
t in
hib
ito
rs:
Th
e m
ou
ld A
sp
erg
illu
s
bin
ds b
oth
fa
cto
r H
an
d C
4b
bin
din
g p
rote
in.
Mo
l. I
mm
un
ol. (
20
07
).
84
. M
eri,
T. e
t a
l. T
he
ye
ast C
an
did
a a
lbic
an
s b
ind
s
co
mp
lem
en
t re
gu
lato
rs fa
cto
r H
an
d F
HL
-1.
Infe
ct.
Im
mu
n. 70
, 5
18
5-5
19
2 (
20
02
).
85
. M
eri,
T. e
t a
l. T
he
hyp
ha
l a
nd
ye
ast
form
s o
f C
an
did
a a
lbic
an
s b
ind
th
e c
om
ple
me
nt
reg
ula
tor
C4
b-b
ind
ing
pro
tein
. In
fect.
Im
mu
n.
72
, 6
63
3-6
64
1 (
20
04
).
86
. P
olte
rma
nn
, S
. e
t a
l. G
pm
1p
is a
fa
cto
r H
, F
HL
-1
an
d p
lasm
ino
ge
n-b
ind
ing
su
rfa
ce
pro
tein
of
Ca
nd
ida
alb
ica
ns. J. B
iol. C
he
m.
(20
07
).
©!2008!N
atu
re P
ub
lis
hin
g G
rou
p