competitive and blocking enzyme-linked immunoassay for detection of fetal bovine serum antibodies to...
TRANSCRIPT

hmaf of Virofogicuf methods, 15 f 1987) 167-I 75 Elsevier
JVM 005%
167
Competitive and blocking enzyme-linked immunoassay for detection of fetal bovine serum
antibodies to bovine viral diarrhea virus
J.B. Katz and S.K. Hanson
Biofogics Virology Laboratqry, National Veterinary Servictx Laboraturies, Veterinary Services, Animnl and Plant Health Inspection Service, United States Department of Agriculture, Ames, Iowa,
U.S.A.
(Accepted 8 October 1986)
~._-..-- .l~ .-~~-
A competitive blocking enzyme-linked immunoassay (CELIA) was developed to detect bovine viral diarrhea virus (BVDV) antibodies in undiluted fetal bovine serum (FBS). The CELIA was based on competition of serum BVDV antibodies with b~otin-labelled anti-BVDV immunogiobn~~ns (Ig) for a hmited quantitiy of s&id-phase BVDV antigen. Antigen preparation was simple, FBS could be tested undiluted, and detergent-containing washes were unnecessary. A series of dilutions of postnatal bovine BVDV antiserum prepared in FBS and a set of 147 undiluted abhatoir FBS sampfes were tested by both CELIA and serum neutralization tests @NT). CELIA results on both sets of specimens corre- lated positively with SNT titers (r=D.99 and r=O.SS). Relative to the SNT, CELIA sensitivity was 100%; specificity was 76%. CELIA detected a level of BVDV antibody below the 1%titer threshold detect- able with the WT. Advantages, limitations, and theoretical differences between the CELIA and SNT are discussed. A similar comparison of CELIA with non-competitive enzyme-linked immunoassay ap- proaches to BVDV serodiagnosis is made. It is concluded that the CELIA is valuable in selecting only BVDV-seronegative FBS for use in vitologic cell culture media.
Enzyme immunoassay; BVD virus; Fetal bovine serum; Ceil cuhure medium --.l_l_----~ _I.II___~_~____~---~_,_
The bovine fetus may respond to challenge with bovine viral diarrhea virus (BVDV) in several ways (Van Oirschot, 1983). An active immune response may develop (Kniazeff and Rimer, 1967; Horner et al., 1973; Ohmann et al., 1982). Another potential response includes induction of persistent BVDV immunotoler- ante with viremia (Van Oirschot, 1983; McCiurkin et al., 1984; Bolin et al., 1985). In the former instance, fetal bovine serum (FBS) may contain BVDV antibodies (Coria and McClurkin, 1978; Ohmann et al., 1982); in the latter, FBS may contain BVDV- or virus-antibody complexes (Cutlip et al., 1980; Bolin et al., 1985).
Correspondence to: J.B. Katz, Biologics Virology Laboratory, National Veterinary Services Labora- tories, Veterinary Services, Animal and Piant Health Inspection Service, United States Department of Agricufture, P.O. Box 844, Ames, IA, U.S.A.
0166-093418711603.50 Q 1987 EIsevier Science Publishers B.V. (Biomedical Division)

168
FBS containing either BVDV, BVDV antibody, or both, is unsatisfactory as a component of cell culture media to be used for pcstivirus growth. Antibody may thwart viral isolation attempts for BVDV and two other serologically related to- gaviruses: hog cholera virus and border disease virus (Van Oirschot, 1983). Non- cytopathic BVDV may infect ceil cultures resulting in viral interference with cy- topathic BVDV used in the serum neutralization test (SNT) and in vaccine pro- duction (Gillespie et al., 1962).
The SNT (Rossi and Kiesel, 1971) is widely used for BVDV serology. Non-com- petitive enzyme-linked imm~Inosorbent assays (ELISA) have been described for detection of BVDV antibodies in bovine serum (Howard et al.. 1985; Chu et at., 1985). We describe here a competitive blocking enzyme-linked immunoassay (CE- LIA) for the same purpose. Like the ELISA methods, the CELIA is rapid, eco- nomical, sensitive, and correlates well with the SNT. The CELIA will operate us- ing undiluted FBS and should theoretically detect anti-BVDV antibodies specifically, regardless of immunoglobulin (Ig) class and subclass. This contrasts with the non-competitive methods based on general detection of all bound Ig rec- ognized by an anti-Ig conjugate (Clark and EngvatI, 19% Attschuh and Van Re- genmortel. 1982; Kurstak, 1985).
Reference and test sera
A negative reference FBS (NR) was chosen on the basis of repeated negative SNT results. The NR was BVDV antibody seronegative by the alpha and beta SNT methods at a 1:2 dilution.
Serum from a calf hyperinlmuniz~d with NADL and NY-1 strains of BVDV was serially diluted in NR serum from I:16 through 1~4096. A f:12CW dilution of that serum was designated as a weak positive reference (WPR) serum and had a geo- metric mean titer of 1:3 by the SNT (n=S).
A group of 147 abbatoir FBS samples was tested by both the CELIA and the SNT.
Biotin-Eabelled lg preparation
BVDV h~erimmn~e serum was produced in a gnotobiotic calf immunized re- peatedly with NADL and NY-l BVDV strains grown in embryonic bovine kidney (EBK) cell culture. Prior to fractionation, the serum was absorbed with EBK cells to help reduce levels of antibodies which may have been generated against EBK cellular antigens.
The serum was fractionated on a protein A-Sepharose CL-4B column (Sigma Chemical, St. Louis, MO) as previously described (Schmerr et al,, 1985). The im- munoglobuli~-containing fractions were pooled and lab&led with aminohe~anoyl biotin at a 5:l (wiw) ratio according to the method of Guesdon et al. (1979).

169
SNT
A microtiter procedure utilizing day-old cultures of Madin-Darby bovine kid-
ney (MDBK) cells was employed (Rossi and Kiesel, 1971). The NADL strain of BVDV was used at Xl-150 median tissue culture infectious doses (TCID,,) per well. Tests were read by cytopathology at 96 h and stained for confirmatory immuno- fluorescence microscopy as needed. Titer was expressed as the reciprocal of the dilution producing the 50% neutralization end point.
CELIA microplate preparation
MDBK cell suspensions were inoculated with non-cytopathic NY-l BVDV at a 1:l multiplicity of infection, seeded into flasks and grown 72 h at 38°C under 5% CO,. Cells were then resuspended at 2 x lo4 cells per 0.2 ml. The stirred suspen- sion was dispensed (0.2 ml per well) into 96-well polystyrene tissue culture plates (Costar 3596, Data Packaging, Cambridge, MA). Wells in rows B-D, columns l-11, received infected cells and were designated antigen-bearing (AB) wells (Fig. 1). Uninfected cell suspensions prepared in an identical manner were dispensed into wells of rows E-G, columns l-11, of the same plates (Fig. 1). These wells were designated cell controls (CC). Rows A and H and column 12 were not utilized to avoid plate edge aberrations (Kurstak, 1985). Plates were incubated 8 h, washed once in phosphate buffered saline (PBS), fixed with 80% acetone-20% Hz0 for 45 min, dried, and stored at -80°C. Under these conditions, plates were stable in CELIA testing for at least 2 months.
CELIA procedure
NR and WPR sera were tested on every plate as negative and weak positive controls. All incubations were conducted on an orbital shaker at 37”C, except that substrate incubation was conducted at 21-23°C. All sera, including NR and WPR sera, were divided into two portions. Aliquots of the first portion (170 FVwell) were placed in 3(AB) wells and 3(CC) wells, i.e., each serum occupied one column (2-11) on a CELIA plate (Fig.1). The plate was incubated during this blocking step for 1 h. During this time, the biotin-labelled Ig preparation was diluted 1:24 in NR serum and added at 2% (v/v) to the second portion of each serum. After the first portions were discarded and the plate rinsed once with PBS, wells were refilled with 125-l~_l aliquots of the label-containing second portion of each serum. A l-h competitive incubation followed; the plate was then washed 4 times with PBS. A 1:2000 dilution of streptavidin-horseradish peroxidase conjugate in PBS (Zymed Laboratories, San Francisco, CA) was added (100 kliwell), incubated for 40 min, and the plate emptied and washed as before. Substrate (170 l-d/well) was added and the absorbance was minitored kinetically at 405 nm (OD,,,) with a mi- croplate reader-computer system (MR 580, Dynatech Laboratories, Alexandria, VA; Apple II Plus, Apple Computer, Cupertino, CA). Substrate consisted of 0.40 mM 2,2’-azino-di(3-ethylbenzthiazolinesulfonic acid) diammonium salt (ABTS,

170
Sigma Chemical) and 2.5 mM H,O, in 0.05 M citric acid buffer (pH 4.0). When the mean OD,,, of the 3 NR serum (AB) wells minus the mean OD,,, of the 3 NR serum (CC) wels reached 0.500 (45-60 min), the reactions on the entire plate were recorded for data processing. Conjugate/substrate background absorbance control wells in column 1 were treated identically to wells in other columns with the exception that both l-h serum incubations were conducted using only NR serum with no added biotinylated Ig (Fig. 1).
The mean OD,,, of the 3 (CC) wells was subtracted from the mean 0D405 of the 3 (AB) wells for each serum, yielding a corrected OD, (COD,). The COD,, values of the WPR serum and sera of unknown status were then compared to the COD value of the NR serum to determine the fractional inhibition of absorbance (FIA) relative to the NR serum:
FIA = [l - (COD,, of unknown serum)i(COD~~s NR serum)].
The NR serum FIA value by formula definition was 0. Sera with FIA values equal to or greater than the WPR serum FIA value were classified BVDV seropositive. Sera with FIA values ranging from 0 to 50% of the WPR serum FIA value were classified seronegative; those with FIA values between 50-100% of the WPR serum
Fig. 1. Microplate format and typical results of CELIA test. Cal. 6: NR serum; Cot. 7: WPR serum; Cots. 2, 3, 9, 11: BVDV seropositive FBS specimens; Cols. 4, 5, 8, 10: BVDV seronegative FBS spec- imens; Col. 1: conjugate-substrate control; Coi. 12 and Rows A, H: empty wells.

171
were designated suspect. In qualitative terms, a serum was considered positive if antigen-bearing wells exposed to that serum developed equal or less color than an- tigen-bearing wells exposed to the WPR serum. The inverse relationship between color intensity and seropositivity was a direct result of the competitive, blocking principle of the CELIA method.
Results
Assay standardization
As shown in Fig. 1, column 1, conjugate/substrate background OD,, was uni- formly negligible. Preliminary titrations were conducted to ensure that biotiny- lated ig, conjugate, and substrate concentrations were not limiting OD,,, during the first 90 min of substrate reaction. This was accomplished by reducing the num- bers of BVDV-infected and uninfected cells seeded into (AB) and (CC) wells until OD,, became dependent upon cell count per well. Antigen limitation of OD, commenced and increased as the well cell count was progressively reduced below 30000 cells/well. At the chosen concentration of 2 x lo4 cells/well, additional ti- trations verified that conjugate and substrate levels were not limiting OD,,s for at least 90 min after substrate addition. Biotinylated Ig concentration was adjusted to be present in only slight excess. This ensured that competing unlabelled anti- body in test sera would limit biotinylated Ig binding with resultant reduction in
OD,,,s.
lntraplate and plate-to-plate assay variability
Plate-to-plate and day-to-day assay variability were assessed by examining the FIA value of the WPR serum tested on every plate. In a series of 21 experiments, the mean WPR serum FIA was 0.24 2 0.07 (s + SD). If the WPR serum FIA deviated from 0.24 by more than 0.07, that experiment was repeated.
Intraplate variation between the 3 (AB) wells used for each serum was assessed by considering the standard deviation of the 3 corresponding OD,, measure- ments. The standard deviation (pooled) of the ODdo for the 3 wells used to test NR serum on each plate was 0.07 (n=Zl). Triplicate wells used for WPR serum testing had an OD,,, p ooled standard deviation of 0.06 (n=21). For both serum types respectively, the 21 triplicate standard deviations did not differ significantly (P >O.lO) from one another, as revealed by Cochran’s test for variance homo- geneity. Similar triplicate well standard deviations were observed with FBS sam- ples of unknown serologic status.
Correlation of CELfA aad SNT
Dilutions of BVDV antiserum were tested in a randomized block design on CE- LIA plates and analyzed for the lowest level of significant non-zero response (Wil-
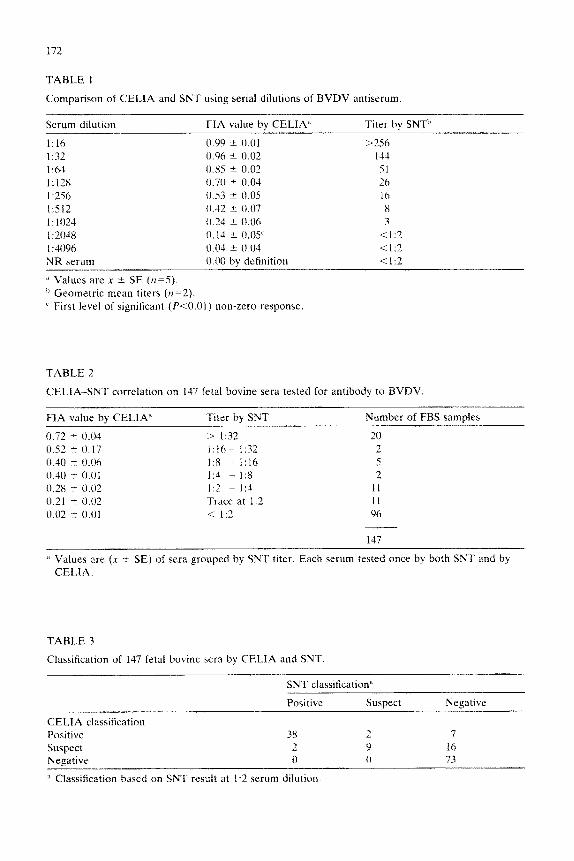
TABLE 1
Comparison of CELIA and SNT using serial dilutions of BVDV antiserum.
Serum dilution FIA value by CELIA’ Titer by SNT”
1:lh
1:x
1:64
1:12x 1:256
I:512
I.102
1:204x
I :3096
0.99 t 0.01 >
0.96 i- 0.02
0.85 ” 0.02 0.70 k 0.04 0.53 -+ 0.05 0.42 t 0.07 0.21 k 0.06 0. 14 rt 0.0s’
0.04 t 0.04
256
144
51
26
16
8
3
<I:2
4 I:2 NR serum
j’ Values are x it SE (r1=.5).
h Geometric mean titers (n=2).
L First level of significant (kO.01) non-zero response
0.00 by definition < 1:2
TABLE 2
CELIA-SNT correlation on 137 fetal bovine sera tested for antibody to BVDV
FIA value bv CELIA” Titer by SNT Number of FBS samples
0.72 2 0.04 > I:32 20 0.52 f 0.17 I:16 - 1:32 2 0.40 It- 0.06 I:8 - 1:16 5 0.40 t 0.01 I:4 - 1:8 2 0.28 t 0.02 1:3 - I:4 11 0.21 I?I 0.02 Trace at 1:2 11 0.02 -t 0.01 < I:2 96
147
d Values are (K rt SE) of sera grouped by SNT titer. Each serum tested once by both SNT and by
CELIA.
TABLE 3
Classification of 137 fetal bovine sera by CELIA and SNT.
SNT classification”
Positive Suspect
CELIA classification
Positive 38 2
Suspect 2 9 Negative tl 0
’ Classification based on SNT result at I:2 serum dilution
Negative
7
16 73

173
liams, 1971). Table 1 presents the CELIA-SNT correlation on these sera. The correlation coefficient (r) between the 2 procedures was 0.99. The CELIA re- sponded significantly (P ~0.01) to a 2-fold serum dilution below the limit of SNT detection. At this serum dilution, the FIA was 0.14 ? 0.05 (x 1 SE, n=5). The WPR serum further demonstrated the high sensitivity of the CELIA: this serum had an SNT titer of 1:3, and produced a mean 24% decrease in OD,,,j relative to the NR serum. Table 2 presents the CELIA-SNT correlation on 147 FBS samples. The r value between the two methods was 0.85 for this group of sera.
A summary classification of the FBS samples by both procedures is shown in Table 3. These data indicated that, relative to the SNT, the CELIA was 100% sensitive and 76% specific.
Discussion
CELIA and SNT results correlated highly using both undiluted FBS samples and dilutions of postnatal BVDV antiserum in FBS. Recent reports of non-competitive ELISA procedures for deteeting bovine antibodies to BVDV have demonstrated that those procedures also correlate well with the SNT (Chu et al., 198.5; Howard et al., 1985). Like the other procedures. the CELIA detects antibody levels at and below the sensitivity level of an SNT performed at the 1:2 serum dilution. The CELIA also shares the advantages of rapidity and economy. An additional ad- vantage is that ongoing cell culture capability is not required.
The theory of competitive and blocking inhibition underlying the CELIA pre- dicts that further reduction of well antigen content and biotinylated Ig concentra- tion may further increase test sensitivity. This would permit detection of lower lev- els of BVDV antibody than was currently demonstrated (Clark and Engvall, 1980; Kurstak, 1985). To ensure high sensitivity, it is important to verify that well an- tigen content does limit OD,,,S and that biotinylated Ig is as dilute as possible with- out being OD,,,S limiting. The 76% specificity of the CELIA compared with the SNT may reflect the high sensitivity of the CELIA. The reduced specificity may also result from detection of non-neutralizing BVDV antibodies which would be inapparent in the SNT (Bolton et al., 1981; Lehtohen and Eerola, 1982), but re- sult in a seropositive classi~cation by CELIA.
Serum from persistently infected fetuses may be antibody negative but contain non-cytopathic BVDV (McClurkin et al., 1984: Bolin et al., 1985). Viremic sera from these fetuses might appear falsely seropositive in the SNT as a consequence of noll-cytopathic interference (Gillespie et al., 1962; Bolin et al., 198.5). These sera might also appear as false-positives upon CELIA, but only if enough virus was present to measurably inhibit the binding of labelled Ig to the viral antigen fixed in the (AB) wells.
Various patterns of Ig class and subclass response to BVDV challenge have been documented (Fernelius, 1966; Ohmann et al., 1982; Howard et al., 1985). The class(es) of Ig produced by a challenged fetus may depend upon fetal age and the length of time between challenge and serum collection. Some ELISA methods de-

174
pend upon class and/or subclass specific anti-Ig conjugates (Chu et al., 1985). These methods may not detect antibodies of other Ig classes and subclasses, Unlike these methods, the CELIA does not depend upon direct detection of all bound Ig, but rather upon the interference of biotinylated Ig binding by functionally specific un- labelled serum antibodies. However, fetal serum antibodies of lower avidity than those in the labelled Ig preparation may not compete well with labelied Ig for fixed antigen. The absence of false-negatives in the CELIA testing of the 147 FBS sam- ples tends to discount this as a practical problem.
In contrast to the non-competitive and non-blocking ELISA tests the CELIA will operate using undiluted sera, without the need for highly purified antigen, or the use of detergents during washes. The ability to use undiluted samples may per- mit detection of those sera with antibody levels otherwise undetectable after di- lution.
Immunoassay techniques including the CELIA have inherent weaknesses. Sin- gle dilution assays may err due to prozone inhibition, and low avidity antibodies may not bind or compete well for fixed antigen (De Savigny and Voller, 1980; Lehtonen and Eerola, 1982). Avidin-biotin based detection systems may err due to elevated binding of the avidin-enzyme conjugate. This problem may be non- specific or the result of high background biotin levels in test sera (Kurstak, 1985). Viral inhibitory factors of a non-antibody nature may be overlooked by immu- noassay techniques but detectable by the SNT (Rossi and Kiesel, 1974). These po- tential problems did not appear to adversely affect CELIA sensitivity in the pres- ent study.
The CELIA procedure is a simple, economical, and sensitive method of ex- amining FBS for BVDV antibody. The ability of BVDV to produce apparently immunotolerant viremic fetuses implies that serology alone cannot assure freedom of cell culture systems from BVDV-related problems. Examination of FBS by both serologic and virologic means is required to accomplish that end.
References
Altschuh, D. and Van Regenmortel. M.H.V. (1982) J. Immunol. Methods 50, 99. Bolin, S.R., McClurkin, A.W. and Coria, M.F. (1985) Am. J. Vet. Res. 46, 2385.
Boiton, D.C., Chu. H.J., Ardans. A.A., Kelley, B. and Zee, Y.C. (1981) Vet. Microbial. 6, 256.
Chu, H.J., Zee, Y.C., Ardans, A.A. and Dai, K. (1985) Vet. Microbial. 10, 325.
Clark, B.R. and Engvall, E. (1980) In: Enzyme-Immunoassay (Maggio, E.T., ed.), p. 167. CRC Press, Boca Raton, FL.
Corm, M.F. and McClurkin, A.W. (1978) Can. J. Comp. Med. 42, 239.
Cutlip. R.C., Mc~lurkin. A.W. and Corm, M.F. (1980) Am. J. Vet. Res. 11, 1938. De Savigny, D. and Voller. A.J. (1980) J. Immunoassay 1, 105. Fernelius, A.L. (1966) J. Immunoi. 96, 488.
Gillespie, J.H., Madin, S.H. and Darby, N.B. (1962) Proc. Sot. Exp. Biol. Med. 110, 248.
Guesdon, J.L., Ternynck, T. and Avrameas, S. (1979) J. Histochem. Cytochem. 27, 1131. Horner, G.W., Johnson, R.H., Dennett, D.P. and Lane, W.R. (1973) Aust. Vet. J. 49, 325. Howard, C.J., Clarke: M.C. and Brownlie. J. (1985) Vet. Microbial. 10, 359. Kniazeff, A.J. and Rimer, V. (1967) Nature (London) 214, 805. Kurstak. E. (1985) Bull. W.H.O. 63. 793.

175
Lehtonen, O.P. and Eerola. E. (1982) J. Immunol. Methods 54, 233. McCIurkin, A.W., Littledike, E.T., Frank, G.H.. Coria, M.F. and Bolin, S.R. (1984) Can. J. Comp.
Med. 48, 156. Ohmann. H.B., Jensen, M.H., Sorensen, K.J. and Dalsgaard, K. (1982) Can. J. Comp. Med. 46.357. Rossi, C.R. and Kiesel, G.K. (1971) Appl. Microbial. 22. 32. Rossi. C.R. and Kiesel, G.K. (1974) Appl. Microbial. 27. 114. Schmerr, M.J.F., Patterson, J.M., Van Der Maaten, M.J. and Miller, J.M. (198s) Mol. Immunol. 22.
613. Van Oirschot. J.T. (1983) Vet. Microbial. 8, 321. Williams, D.A. (1971) Biometrics 27. 103.