comparison of a chemically mediated and an immunologically mediated demyelinating lesion model
TRANSCRIPT

METHODS: A Companion to Methods in Enzymology 10, 440–452 (1996)
Article No. 0122
Comparison of a Chemically Mediated and anImmunologically Mediated DemyelinatingLesion ModelRichard Reynolds, Isabelle Cenci di Bello,Annette Meeson,* and Sara Piddlesden†Department of Anatomy, Charing Cross and Westminster Medical School, London W6 8RP, United Kingdom;*Department of Cell Biology, Skirball Institute, NYU Medical Center, New York, New York 10016; and†Department of Medical Biochemistry, University of Wales College of Medicine, Cardiff, United Kingdom
the environment eventually becomes inhibitory for suc-The production of two animal models for the central nervous cessful remyelination. Therefore, it is essential that
system degenerative condition multiple sclerosis is described attempts are made to understand the changes that takein detail. The first is a chemically mediated noninflammatory place in the cellular environment of areas of white mat-demyelinating lesion of the brain stem induced by the injection ter damage in both successfully remyelinating lesionsof a trypanocidal DNA binding dye, ethidium bromide, into the and chronic demyelinating lesions.cerebellomedullary cistern. The injection does not involve any In many animal models of demyelinating disease in-physical damage to the blood–brain barrier or the CSF–brain
volving chemical toxicity in the absence of inflamma-barrier and is simple to perform. The second lesion model is antion (1, 5, 13), remyelination is often highly efficient. Itimmunologically mediated demyelinating condition involving theis thought that remyelinating oligodendroglia in suchinjection of a T-cell line specific for myelin basic protein, followedlesions derive from immature oligodendroglial precur-by injection of a monoclonal antibody against the myelin surfacesors, although this has yet to be demonstrated. It is ofprotein, myelin/oligodendrocyte glycoprotein. We describe theinterest whether the absence of inflammatory cells inproduction of the antigen-specific T-cell line in detail. This modelthese lesions contributes to the efficient remyelination.is characterized by widespread inflammatory infiltrates accom-
panied by areas of demyelination. Both of these models are In contrast to the wealth of information available con-produced in the Lewis rat, allowing the direct comparison of cerning chemically mediated noninflammatory demye-mechanisms involved in demyelination and repair in the pres- linating lesions, very little is known concerning theence or absence of invading inflammatory cells. Despite the very efficiency of remyelination and the mechanisms in-different etiologies of the two lesion models, they are both acute volved in repair following autoimmune inflammatoryand result in efficient remyelination. q 1996 Academic Press, Inc. lesions. The majority of studies of inflammatory lesions
have used experimental allergic encephalomyelitis(EAE) in rats or mice, induced either by active inocula-tion with myelin basic protein (MBP; 7) or by passivetransfer of activated MBP-specific T lymphoblasts (8).Multiple sclerosis (MS) lesions are characterized byThis approach leads to primarily acute inflammatoryperivascular and periventricular inflammatory infil-foci around blood vessels and ventricles in the CNStrates containing predominantly T lymphocytes andwith demyelination absent or restricted to the dorsalmonocytes. These are accompanied by varying degreesroot entry and ventral root exit zones of the spinal cordof demyelination with preservation of axons. Recent(8). If injections of T lymphoblasts or MBP are followedstudies have provided evidence for limited remyelina-several days later by the injection of antibodies againsttion of these lesions (10, 11), but this appears to occurmyelin-associated glycoprotein (MOG), then the degreepredominantly during the early stages of the disease.of inflammation is increased and is now accompaniedAlthough oligodendroglial survival and proliferationby widespread demyelination (4). Antibody-augmentedhave been noted in early lesions (11), chronic MSdemyelinating EAE (ADEAE) thus combines the hu-plaques are characterized by a pronounced loss of oligo-moral and T-cell-mediated inflammatory events thatdendroglia. Thus it is likely that the cellular and hu-are thought to play a role in the development of MSmoral environment of the lesion site changes with in-
creasing frequency of inflammatory events, such that lesions.
440 1046-2023/96 $18.00Copyright q 1996 by Academic Press, Inc.
All rights of reproduction in any form reserved.
AID Methods 343A / 6709$$$281 09-24-96 16:29:28 metha AP: Methods

441COMPARISON OF DEMYELINATING LESION MODELS
In this paper we describe, in detail, the methods for injection, if the needle is inserted too far it is likelyto damage the respiratory center of the brain stem,producing both ethidium bromide-induced demyelin-
ation (12) and anti-MOG-augmented demyelinating resulting in immediate death of the animal. The nee-dle is thus directed into the cisterna magna, or cere-EAE (6) in the Lewis rat. We briefly compare the two
types of lesions with respect to the timing of demyelin- bellomedullary cistern, an enlarged subarachnoidspace adjacent to the fourth ventricle (Fig. 1A). Theation and remyelination.ethidium bromide solution should be injected slowlyand the needle left in position for approximately 10 sfollowing delivery before removal. Very little leakage
MATERIALS AND METHODS should occur following this procedure. The injectionsite is then wiped with ethanol and the animal re-moved from the stereotaxic apparatus into a paper-A. Ethidium Bromide-Induced Demyelinationlined cage to allow recovery. If the procedure is cor-Young adult Lewis rats (male or female, 100–120 g)rectly carried out then the ethidium bromide is in-are anesthetized using gaseous halothane (for bothjected into the CSF circulation without any physicalinduction and maintenance of anesthesia). The ani-damage to the blood–brain barrier and without themals are mounted in a stereotaxic frame using stan-need for invasive surgery. No clinical symptomsdard ear bars to immobilize the head. The dorsal neckshould be seen in these animals, despite the oftenregion is then shaved and disinfected. The trunk oflarge areas of demyelination.the animal is supported at approximately 457 to allowB. Antibody-Augmented Demyelinating Experimentalthe needle to be inserted horizontally. Injections areAllergic Encephalomyelitisthen performed freehand using a Hamilton syringe
(100 ml, 710RN) fitted with a 357 angled polished 26- This section aims to describe the step-by-step meth-odology for the successful production of new T-cell linesgauge needle (3 cm long). Animals are injected with
20 ml of a 0.15% ethidium bromide solution in 0.9% and their injection into Lewis rats to produce EAE.Through successive cycles of activation and prolifera-saline (total dose 30 mg/animal). Control animals re-
ceive injections of saline only. The solutions are filter tion, a T-cell line will develop high specificity towardthe chosen antigen and increased encephalogenicity.sterilized before injection. The needle is inserted be-
tween and into the depression between the occipital Such activated cells, when injected in small numbers,will produce the animal model of experimental allergicbone and the neural spine of the atlas vertebrae. This
requires a little pressure, and it is usually possible (autoimmune) encephalomyelitis and result in symp-toms with a consistent clinical score prior to anti-MOGto feel the needle pass through the dura. Care must
be taken not to insert the needle too far; otherwise antibody being injected to produce antibody-aug-mented demyelinating EAE.needle tract damage will occur. Due to the site of the
1. Injection of MBP
Animals: 5–10 Lewis rats (we have used females of 160–170 g, although males are also adequate).Antigen: 100 mg GP MBP in a total volume of 100 ml PBS. Our best cell lines have been raised
against MBP isolated from GP but we have also immunized Lewis rats with a syntheticpeptide based on amino acids 67–84 of the guinea pig MBP sequence. Guinea pig MBPis isolated according to the method of Deibler et al. (2).
Adjuvant: Freund’s complete adjuvant supplemented with mycobacterium tuberculosis (Difco) to afinal concentration of 4 mg/ml.
Syringes: 2 1 1.0 ml glass Hamilton syringes with gas tight Teflon plunger connected by fine-gaugetubing (No. 25 or No. 27).
The Hamilton syringes are essential to allow very syringes kept on ice while mixing. The mixture shouldbe emulsified completely so that there is no separationthorough emulsification of the antigen with the adju-of the oily adjuvant from the PBS/antigen. Continuevant. Leakage past the plunger, while emulsifying themixing the solutions until you are ready to inject.solution, can be prevented by dipping the Teflon
plunger briefly into hot distilled water. Adjuvant with Animals should be well anesthetized as the footpadMBT is placed in one syringe (0.5 ml) and antigen (0.5 is extremely sensitive. Induction and maintenance ofml) diluted to the appropriate concentration in PBS in anesthesia is best carried out using gaseous halothane.
Lay the animal on its back with hind footpads facingthe other. Both solutions should be kept at 47C and
AID Methods 343A / 6709$$$282 09-24-96 16:29:28 metha AP: Methods

442 REYNOLDS ET AL.
the operator. Displace all the thoroughly emulsified into the upper part will still cause significant sorenessbut this will subside within 2–3 days. Discomfort ismixture into one syringe, disinfect the footpad with a
swab, insert the needle (gauge) 5–8 mm below the sur- minimized compared with injections into the walkingpart of the pad. The day of injection is counted as Dayface of the skin well into the upper part of the pad, and
carefully inject 50 ml into each of the pads. Injections 0. The animals are then left for 10 days before sacrifice.
2. Preparation of the T-Cell Linea. Media, antigens, and proliferation assay.
b-mercaptoethanol stock solution: 40 ml of b-mercaptoethanol in 10 ml DMEM mediumEagles Hepes (EH): 30 g Hepes per 5 liter of Eagle’s mediumComplete medium: DMEM supplemented with asparagine (0.18 g per 5 liter)
Penicillin (100 IU/ml), streptomycin (100 mg/ml)1% sodium pyruvate1% essential amino acids2 mM glutamineb-mercaptoethanol: 100 ml stock per 100 ml complete medium (10 mM)
Restimulation medium: Complete medium / 1% syngeneic (i.e., Lewis) rat serum. Make up justprior to use. Do not store.
Antigens: Make up in Eagles Hepes at the following concentrations:MBP: 200 mg/mlPPD: 100 mg/mlCon A: 25 mg/ml
Store aliquots at 0207C, thaw, and add to the proliferation assay ormedium at a dilution of 1:10
Tritiated thymidine: 2 Ci/mmol from AmershamPut 100 ml in 10 ml Eagles Hepes to make up stock solution
T-cell growth factor (TCGF): Use either MLA cell line supernatant or Balb/c splenocyte cultures as asource of IL-2. Incubate the spleen cell suspension in complete medium(10 ml/spleen) containing 0.1% mouse serum and 5 mg/ml concanavalinA for 24 h. Collect supernatant by centrifugation and filter sterilize.Store aliquots of 15–45 ml at 0207C.
Proliferation medium: Complete medium containing 15% heat-inactivated horse serum, 15%TCGF, or MLA supernatant as source of IL-2. Make up fresh.
Cell freezing solution: 45% complete medium, 45% horse serum, 10% DMSO.
b. Isolation of T cells from popliteal lymph nodes 10 c. First activation cycle (without antigen-presentingcells). Resuspend the cells in restimulation mediumdays postsensitization. Animals should be sacrificed
by cervical dislocation and dipped in 70% alcohol to at 107 cells/ml. Remove 2 ml of the cell suspension for aproliferation assay and dilute 1 in 2 with restimulationminimize contamination at a later stage. The popliteal
lymph nodes are dissected out in a laminar flow bench medium. Add MBP to the remainder of the cell suspen-sion at a final concentration of 20 mg/ml. If you areand placed in EH medium. Fat should be trimmed off
and the nodes homogenized in 20–30 ml EH using a using Petri dishes, plate 5 ml/60-mm dish or use ap-proximately 10–12 ml per 75-cm2 tissue culture flask.glass hand homogenizer. The cell suspension is then
centrifuged at 1500 rpm for 10 min (47C) and washed Incubate the cell suspension at 377C and 10% CO2 for72 h. Count the start of the culture period as Day 1three times by suspension and centrifugation. Follow-
ing the last spin the viable cells in the suspension are and 72 h as Day 4. The stimulating period of 72 h isquite critical. Less than 72 h is insufficient. Initially,counted using trypan blue (0.5% in PBS). Expect to
obtain 7–10 1 107 viable T cells per animal. If a sub- few lymphoblasts (large rounded T cells) are present.Within 2–4 h most cells adopt a less rounded shape,stantial amount of connective fatty tissue remains after
the third wash, resuspend the cell pellet in a small becoming elongated and forming numerous clumps con-taining hundreds of cells by Day 2. During this activa-volume (approx. 10 ml) of stimulation medium and pass
the entire suspension through a 40-mm nylon cell tion phase, they also increase in numbers (1.5- to 2-fold).strainer (Falcon product code 2340).
AID Methods 343A / 6709$$$282 09-24-96 16:29:28 metha AP: Methods

443COMPARISON OF DEMYELINATING LESION MODELS
d. Proliferation assay. Plate out 12 wells of 200 ml h in culture add 20 ml (1 mCi) per well of [3H]thymidinefrom the stock solution. After 72 h in culture the cellscell suspension (5 1 106/ml) into a flat-bottomed 96-
well ELISA plate and a further 12 wells with 100 ml are harvested into 1.5-ml microtubes and centrifugedat 14,000 rpm for 2 min. The pellet is washed withcell suspension / 100 ml restimulation medium (some-
times it is easier to assess cell proliferation when fewer 1 ml PBS, centrifuged again, and resuspended in 200ml PBS. The suspension is added to 5 ml scintillationcells are present). To the first triplicate group of control
wells, add only Eagles Hepes (10 ml). To other triplicate fluid and counted. The stimulation index (mean anti-gen count divided by mean control count) should in-groups of wells, add antigens at a 1 in 10 dilution from
stock so that the final concentrations will be 10 mg/ml crease significantly for MBP relative to PPD with in-creasing cycles of activation, indicating that the cellPPD, 20 mg/ml MBP, and 2.5 mg/ml ConA. Incubate
the plate for a total of 72 h (377C, 10% CO2). After 60 line is becoming more specific for MBP.
FIG. 1. A diagram of the rat brain stem indicating the site of the ethidium bromide injection (A) and the possible extent of the demyelinatinglesion (A and B). The shaded areas represent the approximate areas in which the lesions may be found, although they never cover thewhole of these areas. 4V, fourth ventricle; STN, spinal tract of the trigeminal nerve; ICP, inferior cerebellar peduncle.
AID Methods 343A / 6709$$$282 09-24-96 16:29:28 metha AP: Methods

444 REYNOLDS ET AL.
e. First proliferation cycle. After 72 h in culture tap When thawing cells allow vials to warm at room tem-perature for 1–2 min and then finish thawing at 377Cthe flask gently or pipette the contents of dish or flaskfor a couple of minutes until the cell suspension is com-to remove any attached cells and harvest by centrifuga-pletely thawed. Immediately place the contents of thetion at 1500 rpm for 10 min. Wash the cells with EHvial into a centrifuge tube containing about 20 ml ofmedium (207C). Count the T cells after the first washwarm proliferation medium. Centrifuge at 1500 rpm(elliptical or large rounded but not the smaller popula-for 10 min. Resuspend the pellet in about 4–5 ml warmtion of rounded cells). Resuspend the cells in prolifera-proliferation medium, incubate in a Petri dish for 1 htion medium, initially at approximately 5–101 106 peras before, and slowly dilute to the appropriate density,milliliter, and allow them to settle in the incubator atas described under section 2e.377C (10% CO2) for 30–60 min. During this time the
blast cells become particularly proliferative and form3. Cycles of Activation and Proliferationlarge clumps containing hundreds of cells. The mor-
phology of the cells will also change from a rounded to If the cells are not to be frozen down after the firsta more elliptical grain-like shape, with this shape being activation then they should be left in proliferation me-visible even within the clumps. Cell density at this dium for the full 5 days and be restimulated on thestage is crucial. Once these clumps have formed then 5th day. Cultures should be fed/split daily during thisdilute cells with proliferation medium down to approxi- phase, as required. Wherever possible, avoid handlingmately 3–8 1 106 per milliliter and incubate at the large numbers of flasks all at once. Freeze all cells innew density for another hour or two. Then dilute as one batch and attempt to build up stock of cells atbefore to a lower density of Ç1–2 1 106 per milliliter, all stages of activation and proliferation that can beincubate for a further hour or two, dilute again down rescued should contamination occur.to 6–8 1 105 cells per milliliter, and leave 2–3 h or a. Preparation of syngeneic thymocytes as antigen-overnight. At each stage, the number of blasts (single presenting cells. Thymocytes must be prepared on theor in clumps) increases dramatically; 20–50 clumps same day that the restimulation of T cells with antigenshould be visible per field (if using 20X objective). The is scheduled. Lewis rats of 6–8 weeks of age (femalefollowing day, 12–18 h later, the flasks should be dense or male) are used. The numbers of rats required forwith numerous clumps. At this stage dilute to the final thymocyte preparation is dependent on the number ofrecommended density of 4–5 1 105 cells per milliliter. T cells needing restimulation. An approximate ratio forCells should be left in proliferation medium for at least T cell:thymocyte of 1:30 is required. Therefore 108 T5 days prior to restimulation (maximum 8 days). Cells cells will require 3 1 108 thymocytes. Approximatelyshould be checked daily and split as necessary. If the 109 thymocytes are usually obtained per thymus.cells change from an elliptical to a rounded shape they Animals are sacrificed by cervical dislocation, andmust be restimulated. the thymuses are removed aseptically and washed as
described for lymph nodes. Homogenize the thymusesf. Cryopreservation. Cells can be frozen at any time,in EH medium using a glass hand homogenizer andexcept immediately at the end of any activation cycle.wash the cell suspension 31 in EH by centrifugation.After the activation phase the cells should be placed inCount the cells following the last wash. Centrifuge theproliferation medium for at least a few hours beforecells, put 1.0 ml of EH medium on the pellet, andfreezing, as their numbers will increase two- to three-gamma irradiate (to abolish cell division) with 5000fold in that time. It is better to freeze cells down at therads from a cobalt or cesium source. Irradiation appa-early stage of proliferation. If frozen down on Days 4ratuses are utilized in hematology departments of mostor 5, many cells die on thawing. Therefore, if the cellshospitals and used for virus inactivation of blood prod-are frozen down after 1 day in proliferation medium,ucts. Time for irradiation is about 6–14 min. It is wisethey will need 4 further days in proliferation mediumto count the cells again afterward, as some may die. Ifon thawing before being restimulated with antigen.an irradiation source is not available then use mitomy-Freeze cells at a density of 1–2 1 107 cells per vialcin-C (20 mg/ml). If mitomycin-C is used, then the T-(1.5-ml vials). On freezing, the viability of the cellscell line will not be able to go through as many cyclesshould be 90% or greater. Use cold freezing solutionof activation and proliferation before dying due to cyto-and freeze the cells in an appropriate freezing appara-toxicity of residual mitomycin-C to the T cells.tus that uses a constant rate of freezing. The apparatus
available in the UK is called Planer P.T.C. 200 (Planer b. Harvesting of T cells for the second activation cycle.Products Ltd., Sunbury on Thames, England). With The procedure is the same for this and any subsequentsuch an apparatus cells should be frozen down at an activation. Centrifuge the T cells at 1500 rpm for 10initial rate of 017C per minute to 01007C, then 057C min, wash once with restimulating medium, and count.per minute to 01507C. Vials are then stored in liquid Wash the irradiated thymocytes once with restimulat-
ing medium and resuspend at a density of 108 thymo-nitrogen.
AID Methods 343A / 6709$$$282 09-24-96 16:29:28 metha AP: Methods

445COMPARISON OF DEMYELINATING LESION MODELS
cytes per 5 ml restimulating medium. Add the thymo- clumps, and an alteration in the morphology from theusual grain-like to a rounded appearance within 2–cyte suspension to the pellet of T cells to give a ratio
of approximately 1 T cell:30 thymocytes. Remove 2 ml 3 days in proliferation medium. This emphasizes theimportance of freezing as many vials as possible in theof T-cell thymocyte mixture for the proliferation assay
and carry out the assay as described previously (see early cycles, following first, second, and third activa-tion. Only a fraction of the total number of cells thatsection 2d). Add the stimulating antigen, MBP (20 mg/
ml, 1/10 dilution from stock), to the remainder of the will be needed for testing the effectiveness of the lineshould be subjected to successive cycles of activationmixture and plate in Petri dishes or flasks. Incubate
these cultures for 72 h as described before (2c). and proliferation. The other vials should be placed inlong-term storage and thawed, expanded, and freshlyc. Harvesting of T cells at the end of the second activa-activated for induction of EAE when required.tion cycle and isolation of freshly activated T cells by
The expansion rate is minimal during the first roundgradient centrifugation. All steps should be carriedof activation and proliferation. If the cultures areout at room temperature. Harvest the cells by centrifu-healthy, the increase in cell numbers during the secondgation and wash with EH (section 2e). Count the cellsand subsequent stimulations will be between 2- and 4-and resuspend in 3.0 ml EH at a cell density not ex-fold and 5- to 10-fold or more during proliferationceeding 107/ml. To separate the activated T cells fromphases. The flow diagram in Fig. 2 illustrates the pro-thymocytes, carefully layer the 3-ml cell suspensiongression of the cell line through successive cycles ofonto 5 ml Histopaque (Histopaque 1077; Sigma oractivation and proliferation. It emphasizes the essen-Lymphopaque, Pharmacia). Centrifuge for 30 min attial stages when cells are to be cryopreserved and the1200 rpm (note: no brake on centrifuge). Collect theimportance of expanding small cell numbers throughlayer of activated T cells that forms at the interface,the subsequent third and fourth activation to specifi-wash 21 in proliferation medium (not EH, as it causescally test the encephalitogenic response in vivo, ratherclumping), resuspend in proliferation medium, andthan rely only on the antigen and proliferative re-count and plate at decreasing cell densities as describedsponses of the cell line in vitro.previously (section 2e). Expand the cultures in prolifer-
ation medium, feeding and/or splitting cultures as re- 4. Testing the Encephalitogenicity of the T-Cell Linequired, remembering to freeze an appropriate number
a. Injection of T cells. Freshly activated T lympho-of vials in the early days of proliferation (1–3 days) orcytes, after the third (or subsequent) activation cy-to proceed to the third activation phase on Day 5 incle, are isolated by gradient centrifugation, washedproliferation medium.21, and finally resuspended in medium or PBS at
d. Further cycles of activation and proliferation. It an appropriate range of densities for intravenous in-is unlikely that there will be sufficient cells at this jection into the tail vein of female Lewis rats (usesecond activation stage to inject different doses to test three female Lewis rats of 160–170 g per dose ofthe encephalitogenicity of the line in Lewis rats in vivo. cells). The following doses should be tested: 105, 5 1However, cells that are freshly activated on their third 105, 106, 5 1 106, and 107 cells per ml. Use 1 1 107
cycle (and any subsequent cycle) should be isolated by only if you have plenty of cells in reserve. It is bestgradient centrifugation, as described above, and the to use three animals at each dose point as there is aisolated cells washed 21 in DMEM or RPMI medium certain amount of variability in the clinical symp-in the absence of additives and resuspended in either toms seen between animals. Count cells prior to andmedium or phosphate-buffered saline for injections (see after diluting T cells to ensure that precise doses aredetails under section 4). injected.
Cycles of proliferation and activation can be re- Weigh and label the individual animals and placepeated, most commonly up to four times. With each three to a cage. This can either be done a few hoursconsecutive activation, both antigen specificity and en- before or while the animal is still anesthetized fol-cephalitogenicity of the cell line are expected to in- lowing the injections. The same person should al-crease. The antigen specificity is determined from the ways handle the animals to minimize stress. Lewisprofile of [3H]thymidine incorporation and the encepha- rats are gentle creatures and do not bite, and atlitogenicity assessed by observing the extent of clinical about 6–8 weeks and a weight of 160 g are quitesymptoms resulting from injections of freshly activated small and easy to handle. Anesthetize the animalcells following the second, third and/or fourth stimula- well with halothane. Lay the animal on its side andtion cycle. We have also succeeded in inducing clinical follow the path of the vein from the base of the tailEAE after a fifth round of T-cell activation. However, (on both sides) through to the end. It is best to injectfrequently, after the fourth cycle of activation, the cell toward the tip of the tail so that, on the first attempt,line will collapse, as observed by a decrease in the pro- if the cell suspension fails to enter properly, you can
move toward the base of the tail without leakage ofliferation response, a failure of the T cells to form
AID Methods 343A / 6709$$$283 09-24-96 16:29:28 metha AP: Methods

446 REYNOLDS ET AL.
cells injected lower down occurring. If you have an (withdraw and expel in and out of syringe) and with-draw 0.5 ml into the syringe ready for injection. Theextra pair of hands available, use them! The other
person can anesthetize the animal and weigh and vein is not always visible even though it is quitesuperficial. Withdraw a little blood into the syringelabel it while you thoroughly mix the cell suspension
FIG. 2. A schematic flow diagram of the experimental procedures for the production of an MBP-specific T-cell line.
AID Methods 343A / 6709$$$283 09-24-96 16:29:28 metha AP: Methods

447COMPARISON OF DEMYELINATING LESION MODELS
to ensure that you are in the vein and then slowly corded daily. On Day 4 after injection of the T cells,when the only apparent signs of disease may be weightinject the cell suspension over approximately 2 or 3
min. If you are located properly there is no resistance loss, the animals are injected intraperitoneally, underhalothane anesthesia, with 3.0 mg per gram bodyto injecting and the cell suspension can be seen to
enter the circulation, with the vein changing to a weight of purified anti-MOG antibody. The appropriatelevel of antibodies will need to be determined to observewhitish color.maximum demyelination without producing too severeb. Monitoring of clinical symptoms of EAE. Clinicala clinical disease (see Ref. 3). The above level is a guideseverity resulting from injection of MBP-specific T lym-only, as the effective dose will depend on the methodsphocytes should correlate well with the T-cell dosesused to purify the antibody and the particular cloneused. Ideally animals should be checked daily for clini-of antibody used. Because the injection of anti-MOGcal symptoms as these can change rapidly. The firstantibodies augments the clinical symptoms observedindication of a response is weight loss, which can occurwith T-cell only injection, it is expected that the clinicalas early as Day 4 but may not occur until Day 8. Thecourse of disease following anti-MOG injection will beloss is minimal in the first days (4–7 g) but increasesseverely debilitating.to 10–20% of body weight by Days 10–12. Together
with weight loss, the following progressive symptoms 6. Analysis of the Lesions by Immunohistochemistrycan be observed and should be scored as follows:
a. Tissue preparation. The Lewis rats are anesthe-tized with sodium pentobarbitone and perfused
0.5 Partial loss of tail tone through the left ventricle with phosphate-buffered sa-1.0 Full loss of tail tone line (PBS) followed by 4% paraformaldehyde. Tissue2.0 Hindlimb weakness blocks are removed and postfixed in 4% paraformalde-3.0 Hindlimb paralysis hyde at 47C for 4–5 h. Tissue blocks are then cryopro-4.0 Hind and forelimb paralysis tected in 30% sucrose at 47C overnight. Five- to seven-5.0 Moribund micrometer-thick sections are cut and mounted on6.0 Dead glass slides precoated with gelatin/chrome alum and
poly-L-lysine. Cut sections are kept in humid conditionsfor at least 1 h before immunostaining. For histologicalGood animal husbandry is essential. Animals that havestaining cut sections were either used immediately orhind- and forelimb paralysis do not necessarily prog-stored at 0207C.ress to moribund. If they are regularly checked and
b. Immunostaining of cryostat sections. Immuno-kept warm, severely disabled animals may recover astaining of cryostat sections is carried out as describedday or two later. However, moribund animals shouldpreviously (12). Sections are incubated in sodium boro-be sacrificed. The ability of cells to induce EAE shouldhydride 0.5 mg/ml (2 1 5 min) to reduce nonspecificbe retested after each stimulation, as the appropriatebinding, washed thoroughly with PBS, and incubatedlevel of cells to inject to induce clinical disease willwith methanol (10 min, 47C). Sections are then incu-change with increasing number of activation cycles. It bated with affinity-purified rabbit anti-MBP antibodyis therefore important, before starting a large series of (diluted 1:500) overnight at room temperature. Sec-
experiments, to ensure that a sufficient stock of cells tions are then washed thoroughly with PBS and incu-at the same stage in the cycle are available. bated with tetramethylrhodamine-conjugated goat
anti-rabbit IgG antibody for 1 h. After immunostaining5. Injection of Anti-MOG Antibodies to Induce all sections are washed thoroughly with PBS and then
Demyelinating EAE mounted in PBS:glycerol 9:1.Anti-MOG monoclonal antibody must first be puri- c. Histology. Cryostat sections or epon-embedded
fied using protein A affinity chromatography to yield semi-thin sections are stained with hematoxylin andan IgG fraction. This is best achieved from ascitic fluid eosin or toluidine blue, then dehydrated through a se-using proprietary protein A columns with which appro- ries of alcohols, and finally mounted in DPX (Agar Sci-priate purification protocols are provided. Two differ- entific).ent monoclonal antibodies have been used for this pur-pose: clone Z-12, an IgG2a (9), and clone 8-18C5, anIgG1 (4). Purified antibody should be diluted in PBS to RESULTSa convenient concentration (1.0 mg/ml) to enable theinjection of approximately 0.5 ml per animal.
Ethidium Bromide LesionsExperimental animals should first be injected witha level of T cells that will induce clinical disease with Clinical symptoms have never been observed in our
experiments following ethidium bromide-induced lesionsa score of about 1.0–2.0, and symptoms should be re-
AID Methods 343A / 6709$$$283 09-24-96 16:29:28 metha AP: Methods

448 REYNOLDS ET AL.
as described in this paper. Injection of ethidium bromide ADEAE Lesionsinto the cerebellomedullary cistern of young adult rats Clinical Symptomsreproducibly gives rise to areas of demyelination in the
Onset of the clinical symptoms may be apparent asbrain stem. The lesions are usually bilateral, but notearly as 5 DPI, with disease progression often reachingalways, and generally affect the inferior cerebellar pedun-a score of 2.0–3.0 at 6–7 DPI. Animals with a diseasecle and to a lesser extent the neighboring spinal tract ofscore above 3.0 at 6 DPI are unlikely to survive. At thisthe trigeminal nerve and the spinocerebellar and olivoc-stage animals should be checked twice daily, as theerebellar tracts. The location can vary a little and is gen-symptoms can rapidly change. Mortality should remainerally found in the areas shown in Fig. 1. The lesions arelow in all recipients injected with anti-MOG antibodieslarge enough to be seen on toluidine blue-stained cryostatprovided that the clinical score resulting from injectionsections and so can be easily identified during the sec-of activated T cells alone did not exceed 2.0. The heighttioning procedure. If the injections of ethidium bromideof clinical symptoms is seen at 8–9 DPI in animals in-are inaccurate then either no demyelination or significantjected with T cells and antibody compared with 5–6 DPIneedle tract damage is seen, although this is commonlyin animals injected only with activated T cells. Clinicalfatal in this area. The dose of ethidium bromide used hasstatus has usually returned to normal by 12–14 DPI.proved to be critical. If lower doses are used then very
Increases in the number of T cells injected simplylittle demyelination is seen. When higher doses wereleads to an increased clinical score and therefore in-tested we began to see some hemorrhagic lesions withcreased mortality. However, due to differences in theintense inflammation. When older animals were used weencephalitogenicity of T-cell lines raised against MBPgenerally saw smaller lesions. Lesions can be inducedas described above, the correct T-cell dose will have toequally reproducibly in outbred Sprague–Dawley rats.be determined for each new cell line and after eachcycle of activation. Increased doses of anti-MOG anti-Demyelinationbody generally lead to an increase in the clinical scoreA detailed analysis of the cellular changes seen inand the degree of inflammation. Higher doses of anti-the above lesions has been published by us previouslybody do not tend to lead to larger areas of demyelin-(12), and only a brief description follows. Initial signsation, but rather to more numerous smaller areas.of a lesion are seen at about 4 days postinjection (4Therefore, both T-cell and antibody dose need to beDPI) and are characterized by splitting of the myelincarefully titrated.lamellae followed by status spongiosus in the affected
area (Fig. 3A). These areas are initially hypocellular, CNS Inflammationsuggesting the death of both oligodendrocytes and as-Histological analysis of ADEAE animals at early stagestrocytes, but rapidly become populated by macrophages
reveals widespread inflammatory cell invasion of thefrom 6 DPI onward. Numbers of macrophages continueCNS. Perivascular cuffs of inflammatory cells are ob-to increase up to 10 DPI, when a large number of my-served predominantly in the pons/medulla and at all lev-elin debris and lipid laden cells can be found betweenels of the spinal cord (Figs. 3C and 3D). Perivascularbundles of demyelinated axons (Fig. 3B). Axons incuffing is also seen to a lesser extent in the cerebellumthese demyelinated lesions appear normal. Maximumand in the hypothalamus and thalamus, close to the thirddemyelination, as assessed by MBP immunostaining,ventricle. Areas of heavy periventricular infiltration areis observed at about 10 DPI (Figs. 4A and 4B). Thereseen around the fourth ventricle and the central canal ofdo not appear to be any inflammatory infiltrates in thethe spinal cord. Subpial inflammation is also a featurevicinity of the lesions, suggesting that the majority ofof the reaction in the spinal cord and to a lesser extent inmacrophages are derived from resident microglia.the pons/medulla. The degree of inflammation increasescaudally along the spinal cord, where large confluentRemyelinationareas of inflammatory cell invasion were seen at the tho-The first signs of remyelination are seen between 11racic, lumbar, and sacral levels (for details see Ref. 6).and 14 DPI and are characterized by abnormally thinMaximum inflammation is generally seen at around 6–myelin sheaths (Fig. 5A). Remyelination is only seen9 DPI and rapidly diminishes thereafter, until it becomesonce the myelin debris has been cleared from the le-absent by 14–16 DPI. The time course of clinical symp-sions. The majority of the remyelination is carried outtoms correlates well with the degree of inflammation butby oligodendroglia, with only small pockets of Schwannis delayed by 1–2 days.cell-derived myelin seen close to the pial surface. By 15
DPI remyelination is well underway and is essentiallyDemyelinationcomplete by 25–30 DPI. A small number of poorly re-
myelinated areas are sometimes seen after long recov- Areas of demyelination, detected by a lack of positivestaining with antibodies to MBP in areas of normallyery times, characterized by the presence of abnormal
myelin figures and cystic areas. dense myelin, have been detected only in animals with
AID Methods 343A / 6709$$$284 09-24-96 16:29:28 metha AP: Methods

449COMPARISON OF DEMYELINATING LESION MODELS
antibody augmented EAE (Figs. 4C and 4D). No demye- size of the areas of demyelination increases to reach amaximum size by 12–14 DPI. Demyelinated lesionslination is detected in Lewis rats injected with MBP-
specific T cells alone or T cells and nonspecific IgG. show the same widespread distribution as the inflam-matory infiltrates, being largest in the pons/medulla,Axons appeared spared, as determined by positive im-
munostaining for neurofilament proteins. Pockets of close to the fourth ventricle, and at the lower levels ofthe spinal cord. Demyelination is seen in both the graydemyelination are detected as early as 6 DPI in the
pons/medulla and at all levels of the spinal cord. The and the white matter of the spinal cord. The large num-
FIG. 3. Toluidine blue-stained semi-thin plastic sections of the inferior cerebellar peduncle of rats injected with ethidium bromide at 6DPI (A) and 10 DPI (B) and hematoxylin/eosin-stained cryosections of the cervical spinal cord of ADEAE animals at 10 DPI (C and D). Thesplitting of myelin lamellae and the removal of myelin debris following ethidium bromide-induced damage is clearly illustrated in A andB, respectively. The perivascular nature of the inflammatory infiltrates is illustrated in C and D. Scale bars represent 15 mm (A and B),90 mm (C), and 45 mm (D).
AID Methods 343A / 6709$$$284 09-24-96 16:29:28 metha AP: Methods
450 REYNOLDS ET AL.
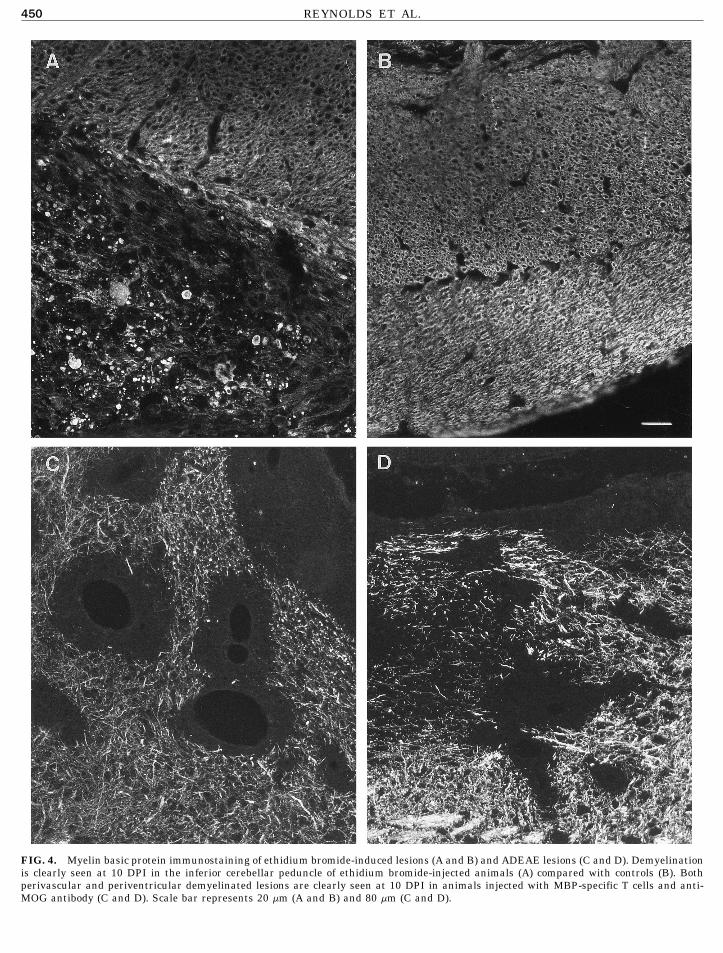
FIG. 4. Myelin basic protein immunostaining of ethidium bromide-induced lesions (A and B) and ADEAE lesions (C and D). Demyelinationis clearly seen at 10 DPI in the inferior cerebellar peduncle of ethidium bromide-injected animals (A) compared with controls (B). Bothperivascular and periventricular demyelinated lesions are clearly seen at 10 DPI in animals injected with MBP-specific T cells and anti-MOG antibody (C and D). Scale bar represents 20 mm (A and B) and 80 mm (C and D).
AID Methods 343A / 6709$$343a 09-24-96 16:29:28 metha AP: Methods

451COMPARISON OF DEMYELINATING LESION MODELS
bers of myelin debris containing macrophages seen in so that by 17–20 DPI it appears almost complete, withall areas appearing to be repaired by 30 DPI. The onsetthe ethidium bromide lesions are not observed in the
ADEAE lesions, suggesting myelin removal by strip- of remyelination appears similar in all areas containinglesions.ping of the myelin layers in the latter lesions.
RemyelinationRemyelination, indicated by the presence of thin my- CONCLUDING REMARKS
elin, is seen as small areas at the edge of the lesionsfrom approximately 15 DPI (Fig. 5B). The extent of
A knowledge of the cellular and molecular eventsremyelination increases rapidly over the next few daysthat lead to successful remyelination of experimentallyinduced lesions of various etiologies should provide vi-tal information toward the goal of understanding andreversing the lack of repair that occurs in the vast ma-jority of MS lesions. We have described here two modelsof demyelinating disease in the rat with very differentetiologies, but which both lead to extensive repair de-spite the extensive white matter pathology.
We have described a simple and reproducible chemi-cally mediated acute demyelinating lesion of the ratbrain stem that does not require extensive surgery orinjection of the toxic agent directly into the brain pa-renchyma. Such a lesion allows the investigation of thebasic cellular and molecular mechanisms involved inthe regeneration of oligodendrocytes, and therefore theprocess of remyelination, without the added complica-tion of extensive immune system involvement.
We have also described the production of a demyelin-ating lesion model that more faithfully reproduces thepathological characteristics of MS lesions. The exten-sive distribution and the perivascular, periventricular,and subpial localization of the lesions found in the hu-moral and cell-mediated pathology of ADEAE are simi-lar to those seen in MS. Although this model does notreproduce the relapsing/remitting course of MS, it doesallow us to study the effects of immune system involve-ment on the process of repair of demyelinated lesions.
ACKNOWLEDGMENTS
This work was supported by The Wellcome Trust and The MultipleSclerosis Society of Great Britain and Northern Ireland.
REFERENCES
1. Blakemore, W. F. (1978) Neuropathol. Appl. Neurobiol. 4, 47–59.
2. Deibler, G. E., Martenson, R. E., and Kies, M. W. (1972) Prep.Biochem. 2, 139–165.
3. Lassmann, H., Brunner, C., Bradl, M., and Linington, C. (1988)FIG. 5. Remyelination of both ethidium bromide-induced (A) andActa Neuropathol. 75, 566–576.T-cell-induced lesions (B). MBP immunostaining clearly demon-
4. Linington, C., Lassmann, H., Brunner, C., and Vass, K. (1988)strates the thin myelin sheaths characteristic of remyelinationAm. J. Pathol. 130, 443–454.against the bright background of normal undamaged myelin. Scale
bar represents 30 mm (A) and 40 mm (B). 5. Ludwin, S. K. (1978) Lab. Invest. 39, 597–612.
AID Methods 343A / 6709$$$284 09-24-96 16:29:28 metha AP: Methods

452 REYNOLDS ET AL.
6. Meeson, A. P., Piddlesden, S., Morgan, B. P., and Reynolds, R. 10. Prineas, J. W., Kwon, E. E., Goldenberg, P. Z., Ilyas, A. A., Qua-ries, R. H., Benjamins, J. A., and Sprinkle, T. J. (1989) Lab.(1994) Exp. Neurol. 129, 299–310.Invest. 61, 489–503.7. Pender, M. P. (1987) J. Neuroimmunol. 15, 11–24.
11. Raine, C. S., Scheinberg, L. C., and Waltz, J. M. (1981) Lab.8. Pender, M. P., Nguyen, K. B., and Willenborg, D. O. (1989) J. Invest. 45, 534–546.
Neuroimmunol. 25, 125–142. 12. Reynolds, R., and Wilkin, G. P. (1993) J. Neurosci. Res. 36, 417–434.9. Piddlesden, S., Lassmann, H., Zimprich, F., Morgan, B. P., and
Linington, C. (1993) Am. J. Pathol. 143, 555–564. 13. Yajima, K., and Suzuki, K. (1979) Lab. Invest. 41, 385–392.
AID Methods 343A / 6709$$$285 09-24-96 16:29:28 metha AP: Methods