climate sensitive site index models for norway
TRANSCRIPT

Draft
Climate sensitive site index models for Norway
Journal: Canadian Journal of Forest Research
Manuscript ID cjfr-2015-0155.R3
Manuscript Type: Article
Date Submitted by the Author: 02-Feb-2016
Complete List of Authors: Antón-Fernández, Clara; Norwegian Institute of Bioeconomy Research Mola-Yudego, Blas; Norwegian Institute of Bioeconomy Research Dalsgaard, Lise; Norwegian Institute of Bioeconomy Research Astrup, Rasmus; Norwegian Institute of Bioeconomy Research
Keyword: climate change, Picea abies, Pinus sylvestris, boreal forest, productivity
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
1
Climate sensitive site index models for Norway
Clara Antón-Fernández, Blas Mola-Yudego, Lise Dalsgaard, and Rasmus
Astrup
1
Clara Antón-Fernández,1 Blas Mola-Yudego, Lise Dalsgaard, and Rasmus Astrup.Norwegian Institute of Bioeconomy Research
1Corresponding author (e-mail: [email protected]).
Can. J. For. Res. 99: 1–33 (2016) DOI: 10.1139/Zxx-xxx © 2016 NRC Canada
Page 2 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
2 Can. J. For. Res. Vol. 99, 2016
Abstract: The present study aims to develop biologically sound and parsimonious2
site index models for Norway to predict changes in site index under different climatic3
conditions. The models are constructed using data from the Norwegian national4
forest inventory (NNFI) and climate data from the Norwegian meteorological5
institute. Site index was modeled using the potential modifier funtional form, with6
a potential component (POT) depending on site quality classes, and two modifier7
components (MOD): temperature, and moisture. Each of these modifiers was based8
on a portfolio of candidate variables. The best model for spruce dominated stands9
included temperature as modifier (R2 = 0.56). In the case of pine and deciduous10
dominated stands, the best models included both modifiers (R2 of 0.40 and 0.54,11
for temperature and moisture respectively). We illustrate the use of the models by12
analyzing the possible shift in SI for year 2100 under one (RCP4.5) of the benchmark13
scenarios adopted by the IPCC for its fifth Assessment Report. The models presented14
can be valuable for evaluating the effect of climate change scenarios in Norwegian15
forest.16
Key words: Climate change, Picea abies, Pinus sylvestris, Norway spruce, Scots pine,17
deciduous, boreal forest, productivity.18
©2016 NRC Canada
Page 3 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 3
1. Introduction19
Quantification and prediction of stand productivity is a key question in forest research,20
with evident implications in forest planning. The productivity of a given area has traditionally21
been estimated by measuring the height of dominant trees at a given age. This value, known22
as site index (SI), is assumed to be a valid surrogate for the potential productivity of a23
particular site. Site index is widely used as a measure of site quality as it can be estimated24
from direct measurements of height and age, and it is a good predictor of volume growth and25
yield (Weiskittel et al., 2011). Site index is also a prevalent key predictor in most traditional26
growth and yield models.27
The site index reflects inherent site characteristics, such as soil and climate, which are28
directly related to forest productivity. Except in the case of intensively managed stands (e.g.29
plantations subject to irrigation or fertilization) site index is generally assumed to remain30
constant through time (Assmann, 1970), as average soil and climatic variables have been31
assumed to remain stable. Therefore, many approaches oriented towards the estimation of site32
index include topographic or locational variables (e.g. slope, latitude, altitude) that relate to33
climate and soil conditions, providing good accuracy in the estimates.34
However, this static approach does no longer hold in the present context of future climatic35
changes, as it is reasonable to expect changes in site index in parallel to changes in climate36
conditions (e.g. Kauppi et al., 2014). This climatic uncertainty context urges for the devel-37
opment of adequate tools for a more dynamic estimation of site index. This is particularly38
relevant in high latitudes, where current climate change scenarios predict the largest tempera-39
ture increases (Stocker et al., 2013). In these regions, temperature is the main variable driving40
forest productivity, and therefore the expected changes will strongly affect forest productivity41
(Peltola et al., 2010).42
Climatic projections for Norway forecast a generally warmer and wetter climate (Stocker43
et al., 2013). The forecasted conditions for Norway are outside the range of conditions cur-44
rently present in Norway, particularly in the southernmost part. Hence, the importance of45
having a site index model that gives biologically sound estimates outside the range of ob-46
served conditions. Several empirical approaches for predicting SI from biophysical predictors,47
©2016 NRC Canada
Page 4 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
4 Can. J. For. Res. Vol. 99, 2016
including climatic variables, have been developed. These approaches range from linear models48
(e.g., Nigh et al., 2004; Sharma et al., 2012) to nonparametric approaches (e.g., Sabatia and49
Burkhart, 2014; Weiskittel et al., 2011). Some nonparametric approaches seem to have gained50
some popularity, e.g. the random forest model (e.g., Crookston et al., 2010; Weiskittel et al.,51
2011), because of their high accuracy and lack of assumptions about how the independent52
variables relate to each other and the dependent variable. This approach has, however, an53
important drawback, that when used outside the range of the training data its predictions are54
unreliable (Sabatia and Burkhart, 2014). Linear regression models and similar approaches like55
generalized additive models, GAM, (e.g., Albert and Schmidt, 2010) or other semiparamet-56
ric additive smoothing approaches (e.g., Nothdurft et al., 2012) share, somehow, a similar57
drawback: since SI has biological constrains that are not directly considered within the linear58
model, linear models could predict unrealistic values when used outside the range of the fitting59
data.60
The present study aims to develop biologically sound and parsimonious models for pre-61
dicting changes in site index under different climatic conditions for the three main species in62
Norway. The models are constructed using data from the Norwegian national forest inventory63
and climate data from the Norwegian meteorological institute. We illustrate how the SI models64
can be used by estimating the SI shift in 2100 under one of the IPCC climate change scenarios.65
2. Material and methods66
2.1. Data sources67
The forest measurements for this study were based on data from the Norwegian National68
Forest Inventory (NNFI) measured during the period 2009-2013. The NNFI consists of perma-69
nent plots laid systematically on a 3x3 km grid, covering an extension from latitude 58.8N up70
to 70.8N. The grid covers the whole country except for the northernmost region (Finnmark).71
Plots with more than 20% of Sitka spruce (Picea sitchensis (Bong.) Carr.) were excluded, due72
to differences in measurement methods, and only plots located in productive forestland were73
considered, resulting in 8943 plots included in the calculations. The main species in Norway74
are, in this order, Norway spruce (Picea abies (L.) Karst.), Scots pine (Pinus sylvestris L.) and75
birch (Betula pendula Roth. and Betula pubecens Ehr.).76
©2016 NRC Canada
Page 5 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 5
Site index (SI) was defined as the average height of the 100 thickest trees per hectare at77
the reference age of 40 years, and it is estimated at each cycle of the NNFI. At each plot78
the SI was assessed for the main species by selecting at least one dominant tree within a 0.179
ha circle. The selection of the dominant tree was done by visual assessment. The heights of80
the selected trees were measured and their corresponding ages were estimated from increment81
cores. The resulting data was used to estimate the SI according to Tveite and Braastad (1981),82
which places SI in classes: 6 (5-6.5), 8 (6.5-9.5), 11 (9.5-12.5), 14 (12.5-15.5), 17 (15.5-18.5),83
20 (18.55-21.5), 23(21.5-24.5), and 26 (>24.5) m.84
In addition, records of soil depth and understory vegetation class were taken at each plot.85
The understory vegetation classes were grouped into 9 categories (Astrup et al., 2010), accord-86
ing to their position along a gradient in nutrients (poor, moderate, and rich) and moisture87
(wet, moderate, and dry). The soil depth was classified in 4 categories: soil depth below 2588
cm, between 25 and 50 cm, between 50 and 100 cm, and above 100 cm. The resulting com-89
bination of soil richness, moisture, and depth form a total of 36 potential combinations, most90
of which are only represented by a small number of plots. These combinations were the basis91
of a categorical variable that organized richness, moisture and soil depth into five biologically92
meaningful groups. The groups were defined to minimize the SI variability for each species93
within each group. The new site quality variable (SQ) had a value of 1 to indicate the poorest94
sites and 5 to indicate the best sites (Figure 1).95
The location of spruce, pine and deciduous dominated stands in Norway is therefore re-96
flected in their frequency in each of the SQ groups defined (Table 1). Pine dominated stands97
are mostly located on poor sites (SQ1) while they are rare on the best sites (SQ5), whereas98
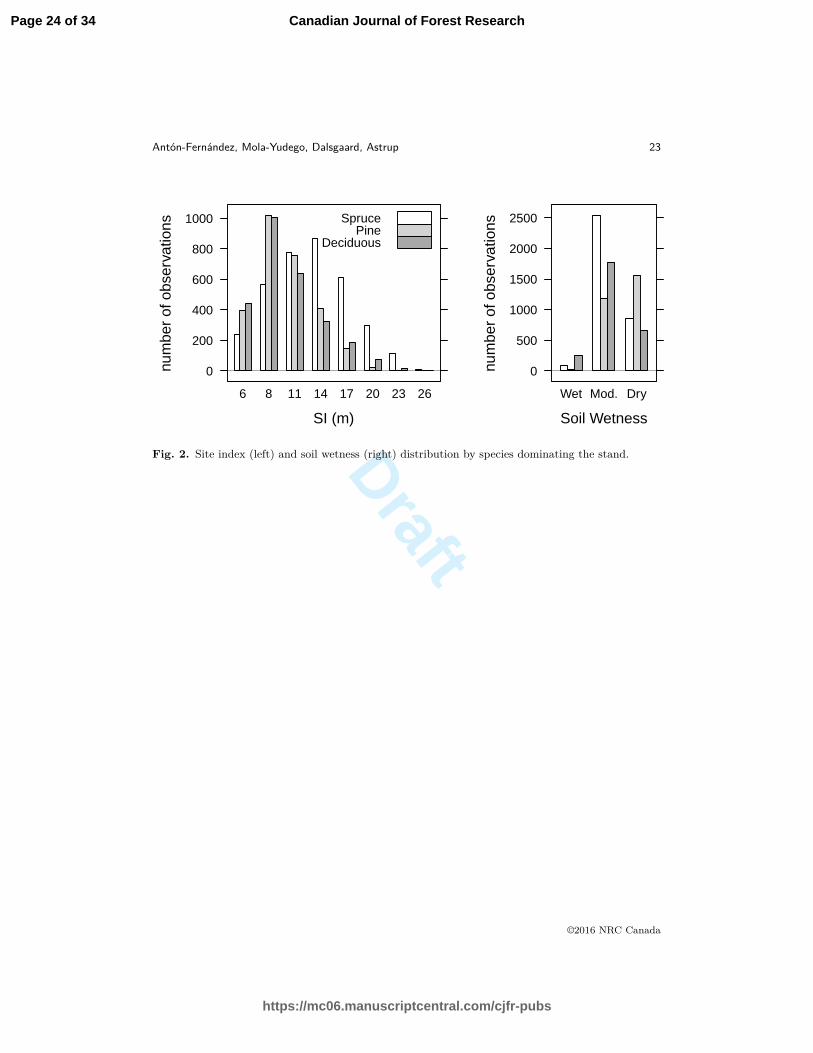
spruce dominated and deciduous dominated stands were mostly distributed in SQ3 and SQ4.99
The distribution of plots among SI classes is similar for deciduous and pine dominated stands100
(Figure 2). The lower SI classes are most commonly dominated by pine and deciduous species,101
while spruce dominated stands lead SI classes higher than 11m. The highest SI observed, 26m,102
is only found in few (12) spruce dominated stands. With respect to soil wetness, deciduous103
and spruce dominated stands are most common in moderate soils while pine are most common104
in dry soils (Figure 2). Most of the plots classified as wet are dominated by deciduous species,105
and only 18 of the plots classified as wet are dominated by pines.106
©2016 NRC Canada
Page 6 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
6 Can. J. For. Res. Vol. 99, 2016
Historical climate data (1900-2008) at the plot level were provided by the Norwegian mete-107
orological institute. Monthly precipitation and temperature were geographically interpolated108
from about 400 weather stations throughout Norway. 30-years averages were calculated us-109
ing data from 1979-2008 (MET7908). We used the multimodel (67) ensemble mean monthly110
temperature and precipitation outcomes from representative concentration pathway (RCPs)111
4.5 (Thomson et al., 2011) from the CMIP5 21st century experiments (Royal Meterological112
Institute of The Netherlands (KNMI), 2014), RCP4.5, for projecting SI changes in 2100 for113
NNFI plots currently on forestland. RCP4.5 is one of the benchmark scenarios adopted by the114
IPCC for its fifth Assessment Report (Moss et al., 2008).115
2.2. Modeling approach116
Since the aim of this study is to predict changes in site index under changing climatic117
conditions it is important that the selected modeling approach produces models that behave118
well beyond the range of current climatic conditions. We use a multiplicative potential (POT)119
modifier (MOD) functional form for the SI model (SI = POT ×MOD) where MOD is scaled to120
only have values between 0 and 1. This functional form has the advantage of being constrained121
and robust since the predicted SI will not exceed the potential even when approaching the122
limits of the empirical data used for model development. In this modelling approach both123
the potential and modifiers are estimated as parameters in a nonlinear regression model. The124
applied modelling approach is equivalent to that frequently applied in studies of tree diameter125
growth where an estimated potential maximum growth rate (estimated as part of a nonlinear126
regression model) is modified according to site factors or competition for light or belowground127
resources (e.g. Lilles and Astrup, 2012; Canham and Uriarte, 2006). The approach applied in128
this analysis is different than the more conventional potential modifier approach (as in Belcher129
and Brand, 1982) where the potential is estimated in a separate analysis, and often using a130
different data set, than the one used for fitting the modifier. It is similar to the conventional131
potential modifier approach in the sense that the modifiers vary between 0 and 1 and modify132
how close to realization of the potential the estimated SI is.133
The SI was, therefore, modeled using the multiplicative potential modifier functional form.134
This functional form has two components: The potential component (POT), which can be135
©2016 NRC Canada
Page 7 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 7
interpreted as the mean SI at a given SQ when neither temperature or moisture are limiting,136
and a modifier component (MOD) which reduces the potential site index according to relevant137
variables, in the form SI = POT × MOD. The modifier component can consist of one or138
several multiplicative modifiers, each of them taking values between 0 and 1, e.g. MOD =139
MOD1 · MOD2, which results in MOD varying between 0 and 1, and the model predicting140
values between 0 and POT . The potential modifier functional form is commonly used to model141
growth, where the lower limit of zero makes sense. However, when applied to SI modeling a142
value of zero does not make biological sense. Therefore, to account for a lower SI limit of 6,143
which is the minimum SI measured in the NNFI, we redefined the basic form of the model as:144
6 + (POT − 6) × MOD.145
The candidate modifiers included a temperature effect, MODT , and a moisture surplus146
effect MODM . For each modifier, a simple portfolio of candidate variables was considered.147
The modifier MODT included variables related to temperature that may affect SI, namely the148
number of growing days (GD, number of days with average temperature above 5◦C), and com-149
binations of the 30-year monthly average temperatures, for instance the temperature sum for150
spring and summer months (TSUM). The modifier MODM included variables related to water151
availability that may restrict the SI, and included several combinations of the 30-year monthly152
average precipitation, the Palmer drought severity index (Palmer, 1965), PDSI, for the grow-153
ing season months, the topographic wetness index (Beven and Kirkby, 1979), TWI, and the154
monthly moisture surplus defined as the difference between 30-year monthly average precipita-155
tion and the average monthly potential evapotranspiration (MSJ). We followed Thornthwaite156
(1948) to calculate potential evapotranspiration (PET) using the 30-year monthly average157
temperature:158
PET = 16K
(
10T
I
)m
I =12∑
i=1
(
T
5
)1.514
; m = 6.75 × 10−7I3− 7.71 × 10−5I2 + 1.79 × 10−2 + 0.492
©2016 NRC Canada
Page 8 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
8 Can. J. For. Res. Vol. 99, 2016
K =(
N
12
)(
NDM
30
)
; N =(
24π
)
ωs
ωs = arccos (− tan φ tan δ) ; δ = 0.4093 sin(
2πJ
365− 1.405
)
159
Where T is the monthly-mean temperature (°C), I is a heat index, which is calculated as160
the sum of 12 monthly index values i, m is a coefficient depending on I, K is a correction161
coefficient computed as a function of the latitude and month, NDM is the number of days of162
the month, N is the maximum number of sun hours, ωs is the hourly angle of sun rising, φ is163
the latitude in radians, and δ is the solar declination in radians, J is the average Julian day of164
the month.165
The plots were grouped according to dominant species (spruce, pine, and deciduous plots)166
and equations were fit to each of these categories separately. All the candidate variables were167
preliminarily explored by examining graphically and analytically their explanatory power for168
site index, for each modifier and for each species. To explore the strength and shape of the169
relationship between site index and the candidate variables we fitted linear tail-restricted170
cubic splines (5 knots) functions to each candidate variable and species (Harrell, 2001). When171
analyzing the strenght of the association between site index and the candidate variables, cubic172
splines offer the advantage of allowing departures from the assumption of linearity in the173
association. In addition, correlation matrices were computed and examined, to determine the174
degree of intercorrelation among variables.175
The criteria for including the candidate variables in the final version of the models were:176
they had to be significant at the 0.05 level, and they had to provide the highest explanatory177
power of the candidate variables.178
Concerning the model shape of the modifiers, two basic forms were considered. Variables179
increasing with the value of the explanatory variable were modelled using the logistic functions180
1/ (1 + exp(−βX)), where X is a vector of explanatory variables, and β is the vector of param-181
eters to be estimated. Relationships reaching a peak at certain point and decreasing afterwards182
were modelled using the Weibull function γ/α · ((X − µ) /α)γ−1 exp (− ((X − µ) /α)γ), where183
X ≥ µ, and α>0, scaled to be bounded between 0 and 1. The Weibull function is a widely used184
©2016 NRC Canada
Page 9 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 9
and flexible function that have a shape parameter (γ), a scale parameter(α), and a location185
parameter (µ). To bound the Weibull function between 0 and 1, we added a scaling factor186
(1/ξ) which multiplied the whole function and was calculated as the maximum value of the187
function, and we redefined X as188
X =
X
µ
X ≥ µ
otherwise.
Thus, (X − µ) will never be negative, and values of X below µ will result in the modifier189
taking a value of 0.190
The basic form of the model was 6+(POT − 6)×MOD, where MOD = MODT ×MODM ,191
resulting in two models, named md1 and md2. Model md1 includes the temperature modifier192
MODT , and model md2 includes both the temperature and the moisture availability modifier193
MODM .194
Finally, the models were assessed in order to identify which modifiers, and in which com-
bination, resulted in a better model. The assessment was based in the Akaike information
criterion, AIC, (Akaike, 1974), and BIC (Schwarz, 1978), and the proportion of variance ex-
plained by the model relative to that explained by the simple mean of the data, adjusted by
the sample size (n) and the number of regressors (p):
R2
adj = 1 −
(
n − 1n − p − 1
)
(
1 − R2)
R2 = 1 −
(
∑
i (yi − fi)2
∑
i (yi − y)2
)
where y is the mean of the observed SI, fi is the ith model predicted SI, and yi is the ith195
observed SI.196
To avoid, as much as possible, problems with convergence we used a simulated annealing197
algorithm (Goffe et al., 1994) implemented in the likelihood package (Murphy, 2015) in R198
(R Core Team, 2015) to fit all models. The likelihood package allows to find the maximum199
likelihood estimates of statistical models using simulated annealing, a global optimization200
©2016 NRC Canada
Page 10 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
10 Can. J. For. Res. Vol. 99, 2016
algorithm, with or without bounded searches.201
2.3. Projections202
The temperature and precipitation outcomes from the multimodel ensemble for RCP4.5203
have a 0.5 degree spatial resolution. Since the source and resolution of these data are differ-204
ent from the ones used for fitting the models (MET7908), we estimated the 30-years monthly205
average temperature at each plot for the 2071-2100 period as the sum of MET7908 and the pro-206
jected change according to RCP4.5. We calculated the projected change according to RCP4.5207
as the difference between the average monthly temperature for the 1979-2008 period available208
from RCP4.5 and the 2071-2100 monthly average from RCP4.5. We used the estimates of the209
30-years monthly averages for the 2071-2100 period, calculated as the sum of MET7908 and210
the projected change according to RCP4.5, to calculate the variables required by the models,211
TSUM and MSJ.212
3. Results213
3.1. Model fitting214
Several candidate variables for MODT were available. The preliminary correlation matrices215
of the candidate variables showed that temperature and number of growing days were highly216
correlated and that the best temperature-based variable had higher correlation with SI than217
the number of growing days for all species. Among the best temperature variables there was218
little variation in their correlation with SI, and the correlation among them was high (>0.94).219
The graphical evaluation of the residuals of the linear tail-restricted cubic splines revealed220
that April, May, and June monthly temperatures performed best, but different months tended221
to perform better in different regions (e.g. northern region), while the sum of temperature in222
spring and early summer had a more consistent residual pattern. Therefore, we selected tem-223
perature in spring and early summer, defined as the sum of the 30-year average temperatures224
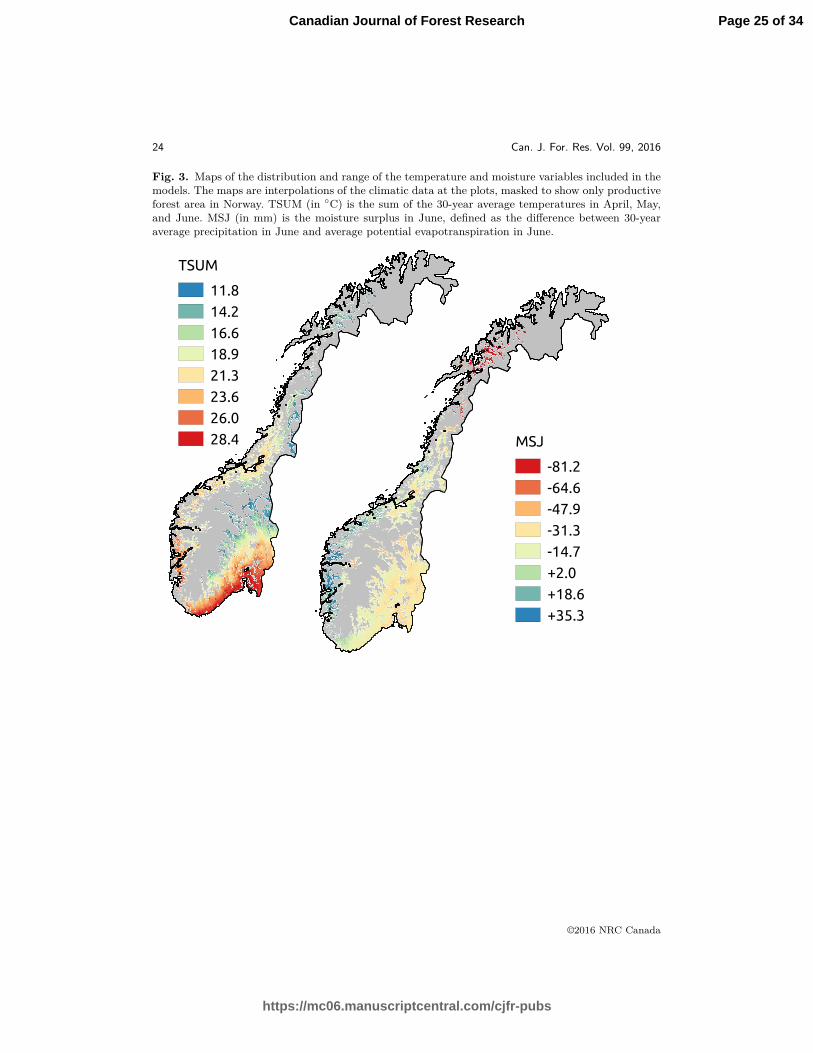
in April, May, and June (TSUM), for MODT . The range and distribution of these variables225
is shown in Figure 3.226
For MODM the correlations among candidate variables and SI were smaller than for227
MODT , but stronger for deciduous than for spruce or pine. Measures of goodness-of-fit (resid-228
©2016 NRC Canada
Page 11 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 11
ual standard error and coefficient of determination R2) of the linear tail-restricted cubic splines229
revealed that water surplus in June outperformed all variables. Therefore, we selected moisture230
surplus in June (MSJ), defined as the difference between 30-year average precipitation in June231
and average potential evapotranspiration in June.232
The preliminary analysis determined the parameters that varied with SQ and the most233
adequate functional form for each modifier and for each species. We assessed graphically the234
need to make each parameter dependent on the SQ group. Table 2 shows the two modifiers235
selected for each species. In general, the variability explained by the best models, as measured236
by R2, was highest for spruce dominated plots, followed by deciduous and pine dominated237
plots. For all species the largest variability was explained by temperature.238
Consistently with the limiting factor theory, the sensitivity analyses revealed that the effect239
of temperature on SI was smaller for poorer sites and that this effect increases with site quality240
for all species (Figures 4, 5, and 6)241
In the spruce dominated stands, a large part of the variability was already explained by the242
model including TSUM (md1) in addition to the SQ factor (Table 3). In pine dominated stands243
(Table 3), model md2, with two modifiers, performed best. The full model (md2) improved AIC244
and R2 while keeping the estimates of the parameters within the expected range. A decrease245
in moisture surplus at the wetter sites results in an increase in SI, but once the optimum246
was reached, a drier climate (lower moisture surplus) results in a decrease in SI. The effect of247
temperature was greater at optimal moisture surplus, and it was very limited at the lowest248
and highest moisture surplus limits.249
Thus, we regarded md2 (temperature as modifier), md2 (temperature and moisture surplus250
as modifiers) and md2 (temperature and moisture surplus as modifiers) as the best models251
for spruce, pine and deciduous dominated stands, respectively. The selected models show little252
apparent trend in plots of residual values against either predicted values (Figure 7) or any of253
the indendent variables (data not shown).254
3.2. Projections255
Climatic projections for Norway forecast a generally warmer and wetter climate, which256
results in a similar range of MSJ values, but a generally higher TSUM (Figure 8). Projections257
©2016 NRC Canada
Page 12 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
12 Can. J. For. Res. Vol. 99, 2016
for SI in 2100 for the NNFI plots used for fitting the models were calculated by adding the258
expected change to the current SI. That is, we used the best models for each species to estimate259
SI for current conditions and for conditions in 2100, we then calculated the difference between260
these two and applied it to the current SI on each plot. For all species the patterns of SI remain261
similar to the current patterns of SI. The projections show that in most plots there will be an262
increase or no change in SI, with pine being the species less affected by climate change. The263
projections show constant or increasing SI, with very rare occurrences of decreasing SI (Figure264
9). The largest changes in SI are for spruce dominated stands, where the projections show an265
increase in SI of at least 1 meter in most regions, with a higher increase in the central part of266
Norway and plots close to the tree line in the southern part of Norway. For pine dominated267
stands the increase in SI was more moderate, with most of the plots experiencing little change.268
For deciduous dominated stands the change in SI was bellow 3 meters in most plots, with little269
change in the northernmost regions, and only few plots experiencing increases of SI above 3270
meters. The range of MSJ on the original data (-115, 134) does not differ much from the range271
of MSJ on RCP4.5 (-119, 118), being the average change in MSJ -6.5 mm. The average change272
in TSUM in RCP4.5 with respect to MET7908 was 6.5 ◦C. The range of TSUM change from273
(0, 32.5) in MET7908 to (1.6, 38.5) in RCP4.5.274
4. Discussion275
The present paper explains the variations of SI in Norway according to climatic and soil276
conditions by using an empirical approach. The models are fit using an extensive data pool277
resulting from the NNFI and environmental variables from the Norwegian meteorological insti-278
tute. Thus, an application to recent climate conditions becomes straightforward. At the same279
time, the models are biologically sound, and efficiently represent the main limitations to forest280
growth in Norwegian conditions.281
However, there are limitations in the data used for the estimation of the SI: in general,282
the overall estimation of site index based on a reference height is a difficult task subject283
to measurement error and the site classes in Norway are defined in intervals, which reduces284
the precision of the predictions. Despite these limitations, the NNFI is the largest data set285
available for the region and covers systematically a broad geographical region with diverse286
©2016 NRC Canada
Page 13 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 13
climate conditions, thus providing a solid empirical basis for the models presented.287
The models presented precision levels similar to other recent studies of SI prediction using288
alternative statistical approaches (e.g. Seynave et al., 2005; Albert and Schmidt, 2010; Aertsen289
et al., 2010). In Norway, Sharma et al. (2012) presented SI models for Norway spruce and Scots290
pine with higher predictive power than the ones presented here. Sharma et al. (2012) used linear291
regression models and the year of stand establishment, the temperature sum (degree days above292
5ºC), location, topography, soil, and understory vegetation variables as explanatory variables.293
Although Sharma’s et al. R2 values may look promising (up to 0.85 for Norway spruce and294
0.72 for Scots pine), their models present several limitations when used for prognoses under295
climate change. They assume a linear relationship between temperature sum and SI, thus296
higher temperatures will always, regardless of precipitation, result in higher SI. The inclusion of297
the year of stand establishment results in a continuous trend of higher SI in newly established298
stands, which may result in illogical SI when used for prediction of long-term prognoses.299
Recent studies on SI prediction have focused on modeling the spatial SI trends. For example,300
Albert and Schmidt (2010) used generalized additive regression models with a two-dimensional301
location component to model the spatial SI trend. We chose not to include location components302
in our models because, in our case, location is correlated with the main variables of interest303
(the correlations between TSUM and latitude and longitude are 0.47, and 0.36 respectively,304
and between MMS and latitude and longitude are 0.47, and 0.72, respectively) and that would305
cause confounding of the effects of changes in the climatic variables. Other options to address306
spatial correlation is that of Nothdurft et al. (2012), who applied a simplified universal kriging307
method using B-spline basis functions to provide spatio-temporal predictions of site index308
for an area of Southwest Germany. The Nothdurft et al. (2012) approach requires that the309
mean function of the spatial site-index process, the one that depends on climatic variables, to310
be linear. Since the relationship between SI and most of the climatic variables is non-linear,311
the relationships are linearized through the B-spline functions. Nothdurft et al. (2012) chose312
the number and location of the B-spline knots heuristically to avoid illogical behavior when313
extrapolating beyond the range of the model data. The strength of the approach selected in the314
present paper is that we ensure biologically sound results beyond the current range of climatic315
conditions, and that behavior is defined by the data and the basic form of the models, avoiding316
©2016 NRC Canada
Page 14 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
14 Can. J. For. Res. Vol. 99, 2016
having to select heuristically some of the parameters. Other interesting approach to model the317
effects of biophysical factors on tree growth is to model the effect of biophysical factors on the318
potential tree height as it is done in SILVA (Pretzsch et al., 2002). This approach, however,319
requires data on dominant height, which is neither currently recorded by the NNFI nor is320
available from experimental plots in a representative scale for Norwegian conditions.321
Our results show that SI increases with temperature. Due to interaction effects between322
temperature and site quality, the increase becomes more pronounced with increasing site qual-323
ity. This results are congruent with Sigurdsson et al. (2013), who found that the effect of324
increase on temperature or CO2 on mature Norway spruce in the boreal zone was not evident325
unless nutrient availability was improved. Our results are also consistent with Pretzsch et al.326
(2014) findings that growth acceleration, likely due to climate change, for Norway spruce since327
1960 in Central Europe is stronger in more fertile sites.328
The effects of the variables included in the final version of the models were logical and329
in line with previous studies. In Norway, Andreassen et al. (2006) underlined the effect of330
temperature and precipitation on Norway spruce growth based on tree-ring series. Andreassen331
et al. (2006) concluded that, of the variables explored, June temperature and precipitation332
had the largest influence on tree ring increments of the variables explored, which is consistent333
with our results. For all three species, most of the variability was explained by MODT , which334
is also consistent with previous studies on the relationship between climatic factors and tree335
growth in Fennoscandia, which show that growth is mainly determined by summer temperature336
(e.g. Ge et al., 2011; Andreassen et al., 2006). Ge et al. (2011) used a process-based growth337
model to assess the impacts of climate change on stem wood growth. They found that most of338
the variability of annual growth in Norway spruce in Finland was explained by temperature,339
particularly June temperature, while the relationship between annual growth and monthly340
precipitation was more subtle. Andreassen et al. (2006) also found stronger correlations of341
growth with summer temperatures than with monthly precipitation for Norway spruce in342
Norway. The fact that moisture surplus is not included in the final model for spruce is not343
to say that that moisture surplus is not an important factor in spruce dominated stands344
productivity. Rather, it indicates that moisture surplus is not a limiting factor within the345
current range of spruce dominated stands.346
©2016 NRC Canada
Page 15 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 15
For pine dominated stands the optimal MSJ is lower than for deciduous dominated stands.347
This is not surprising, since pine dominated stands are already dominating the drier sites in348
Norway (Figure 2).349
One of our main objectives was to make the models biologically sound, which is a require-350
ment when the aim of the models is to forecast the productivity of large non-homogeneous351
areas. To achieve this objective we used empirical data and a modeling approach that bounds352
the response of the model between a maximum, the potential, and a minimum, defined as the353
minimum recorded SI (6m). As a result of this, even if the conditions on a plot for a certain354
species become unsuitable for the survival of the species or the plot becomes unproductive, the355
model will still give an estimate of the SI between the minimum and the maximum. Predict-356
ing the suitability of a site for the survival or establishment of a species should be answered357
separately, and it is not within the scope of this paper.358
There are other considerations that should be taken into account when applying climate359
sensitive SI models. One should be careful when applying these models to already established360
stands, as the climate-growth response of the stands already established might depend on361
the provenance of the trees, their age, and elevation, among other factors (Primicia et al.,362
2015). Productivity might also be affected by other factors that might change with time and363
that are not considered here, like CO2, nitrogen deposition (Sigurdsson et al., 2013), nutrient364
availability, and solar radiation.365
The potential component (POT) is the mean SI index at a given site quality (SQ) when366
neither temperature or moisture are limiting. Even under the best climatic conditions there367
are other factors that limit the growth of trees, such as light and nutrient availability, or me-368
chanical factors (e.g. wind, snow). Since the fit of a model is usually performed by minimizing369
weighted errors, the model will predict the average SI found under certain site conditions,370
temperature, and precipitation, which will never be the maximum SI observed because of the371
imperfect correlation between SI and the variables of the models, and the error implicit in372
the measurements of the variables. Thus, the models correctly predict trends due to a change373
in temperature or precipitation, but they will predict expected SI under given conditions. In374
order to predict future SI under climate change, we suggest that the models should be used375
to predict SI change (predicted SI in future climate - predicted SI in current climate) which376
©2016 NRC Canada
Page 16 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
16 Can. J. For. Res. Vol. 99, 2016
then is added to the currently observed SI. If no information about the current site index is377
available, the models can be used directly but with an expectation of a larger error than when378
current site index information is available. If there is information about current SI and the379
model is used to predict SI change, the predicted SI can exceed POT.380
Thanks to the broad array of conditions currently present in Norway we have been able to381
use the space-for-time substitution, an approach widely used in other fields such as biodiversity382
modeling (Blois et al., 2013), to predict climate change effects on SI for Norway. The space-for-383
time substitution assumes that spatial and temporal variation are equivalent. This approach is384
commonly used accross fields when long-term time-series data are not available. However, this385
approach has some drawbacks that should be considered. For example, as with any empirical386
model, caution should be used when extrapolating beyond the original range. Also, although387
SI is assessed at each cycle of the NNFI, the observed SI is the result of the stand history and388
climate throughout the life of the trees currently in the stand, and as so, it is an imperfect389
measurement of the current SI if a new stand were to be established. Since the observed SI is390
the result of the compound response over the life span of the trees, we chose to use the average391
of the climatic variables over the last 30-years, instead of using only more recent data, as the392
explanatory variable for our SI models.393
When the best models are applied using climate projections for 2100 (RCP4.5), the patterns394
of change observed in Figure 9 reflect the behavior of the models. The general trend for all plots395
considered and all species is an increase in SI for 2100. For more extreme climate scenarios (e.g.396
RCP8.5) the results may be considerably different than under RCP4.5. The moisture indicator,397
MSJ, does not change dramatically between MET7908 and RCP4.5, which is why there are only398
very few plots with a decrease in SI greater than 1 meter (Figure 9). Contrary to the presented399
results, studies from other parts of the boreal forest indicate reduced forest productivity. For400
example, projections for Canada under climate change indicate a likely decrease in productivity401
in areas with relative high water stress and low water-holding capacity, but an increase on forest402
productivity on sites with high water-holding capacity during the 21st century (Johnston et al.,403
2009). In northern Finland, where soil moisture seldom limits forest growth, stem wood growth404
are expected to be higher under climate change, while in sourthern Finnland stem wood growth405
is, on average, lower under climate change due to limited water availability compared with the406
©2016 NRC Canada
Page 17 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 17
current climate (Ge et al., 2011). Climatic projections for Norway forecast a generally warmer407
and wetter climate (Stocker et al., 2013), and at least for RCP4.5, it seems that the higher408
precipitation will, in general, compensate the higher water requirements and result in higher409
productivity for most areas in Norway and mainly for spruce and deciduous dominated stands.410
Finally, despite the restrictions of the modelling approach and the obvious limitations411
in forest growth prognoses, the models presented in this paper present a robust and easily412
applicable basis for SI prediction in boreal conditions.413
5. References414
References415
Aertsen, W., Kint, V., van Orshoven, J., Özkan, K., Muys, B., 2010. Comparison and ranking416
of different modelling techniques for prediction of site index in Mediterranean mountain417
forests. Ecological Modelling 221, 1119–1130.418
Akaike, H., 1974. A new look at the statistical model identification. IEEE transactions on419
automatic control 19, 716–723.420
Albert, M., Schmidt, M., 2010. Climate-sensitive modelling of site-productivity relationships421
for Norway spruce (Picea abies L. Karst.) and common beech (Fagus sylvatica L.). Forest422
Ecology and Management 259, 739–749.423
Andreassen, K., Solberg, S., Tveito, O.E., Lystad, S.L., 2006. Regional differences in climatic424
responses of Norway spruce (Picea Abies L. Karst) growth in Norway. Forest Ecology and425
Management 222, 211–221.426
Assmann, E., 1970. Principles of forest yield study. Pergamon Press Ltd., Oxford.427
Astrup, R., Dalsgaard, L., Eriksen, R., Hylen, G., 2010. Utviklingsscenarioer for karbonbind-428
ing i Norges skoger [Development scenarios for carbon sequestration in Norwegian forest].429
Oppdragsrapport fra Skog og landskap 16/10. Norwegian Forest and Landscape Institute.430
Ås, Norway. In Norwegian with English summary.431
Belcher, D.M., Brand, G.J., 1982. A description of STEMS- the stand and tree evaluation and432
modeling system. Gen. Tech. Rep.433
©2016 NRC Canada
Page 18 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
18 Can. J. For. Res. Vol. 99, 2016
Beven, K.J., Kirkby, M.J., 1979. A physically based, variable contributing area model of basin434
hydrology. Hydrological Sciences Bulletin 24, 43–69.435
Blois, J.L., Williams, J.W., Fitzpatrick, M.C., Jackson, S.T., Ferrier, S., 2013. Space can436
substitute for time in predicting climate-change effects on biodiversity. Proc. Natl. Acad.437
Sci. U.S.A. 110, 9374–9379.438
Canham, C.D., Uriarte, M., 2006. Analysis of neighborhood dynamics of forest ecosystems439
using likelihood methods and modeling. Ecological Applications 16, 62–73.440
Crookston, N.L., Rehfeldt, G.E., Dixon, G.E., Weiskittel, A.R., 2010. Addressing climate441
change in the forest vegetation simulator to assess impacts on landscape forest dynamics.442
Forest Ecology and Management 260, 1198–1211.443
Ge, Z.M., Kellomäki, S., Peltola, H., Zhou, X., Wang, K.Y., Väisänen, H., 2011. Impacts of444
changing climate on the productivity of Norway spruce dominant stands with a mixture of445
Scots pine and birch in relation to water availability in southern and northern Finland. Tree446
Physiol. .447
Goffe, W.L., Ferrier, G.D., Rogers, J., 1994. Global optimization of statistical functions with448
simulated annealing. Journal of Econometrics 60, 65–99.449
Harrell, F.E., 2001. Regression modelling strategies with applications to linear models, logistic450
regression, and survival analysis. Springer, New York, NY.451
Johnston, M.H., Campagna, M., Gray, P.A., Kope, H.H., Loo, J.A., Ogden, A.E., O’Neill, G.,452
Price, D.T., Williamson, T.B., 2009. Vulnerability of Canada’s tree species to climate change453
and management options for adaptation: an overview for policy makers and practitioners.454
Res. Coun. Pub. 12416-1E10. Canadian Council of Forest Ministers.455
Kauppi, P.E., Posch, M., Pirinen, P., 2014. Large Impacts of Climatic Warming on Growth of456
Boreal Forests since 1960. PLoS ONE 9, e111340.457
Lilles, E.B., Astrup, R., 2012. Multiple resource limitation and ontogeny combined: a growth458
rate comparison of three co-occurring conifers. Can. J. For. Res. 42, 99–110.459
©2016 NRC Canada
Page 19 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 19
Moss, R.H., Babiker, M., Brinkman, S., Calvo, E., Carter, T., Edmonds, J.A., Elgizouli, I.,460
Emori, S., Lin, E., Hibbard, K., others, 2008. Towards new scenarios for analysis of emis-461
sions, climate change, impacts, and response strategies. Technical Report. Pacific Northwest462
National Laboratory (PNNL), Richland, WA (US).463
Murphy, L., 2015. likelihood: Methods for maximum likelihood estimation, R package version464
1.7.465
Nigh, G., Ying, C.C., Qian, H., 2004. Climate and productivity of major conifer species in the466
interior of British Columbia, Canada. Forest Science 50, 659–671.467
Nothdurft, A., Wolf, T., Ringeler, A., Böhner, J., Saborowski, J., 2012. Spatio-temporal pre-468
diction of site index based on forest inventories and climate change scenarios. Forest Ecology469
and Management 279, 97–111.470
Palmer, W., 1965. Meteorological drought. Res. Pap. 45 45. U.S. Weather Bureau.471
Peltola, H., Ikonen, V.P., Gregow, H., Strandman, H., Kilpeläinen, A., Venäläinen, A., Kel-472
lomäki, S., 2010. Impacts of climate change on timber production and regional risks of473
wind-induced damage to forests in Finland. Forest Ecology and Management 260, 833–845.474
Pretzsch, H., Biber, P., Schütze, G., Uhl, E., Rötzer, T., 2014. Forest stand growth dynamics475
in Central Europe have accelerated since 1870. Nature Communications 5.476
Pretzsch, H., Biber, P., Ďurský, J., 2002. The single tree-based stand simulator SILVA: con-477
struction, application and evaluation. Forest Ecology and Management 162, 3–21.478
Primicia, I., Camarero, J.J., Janda, P., Čada, V., Morrissey, R.C., Trotsiuk, V., Bače, R.,479
Teodosiu, M., Svoboda, M., 2015. Age, competition, disturbance and elevation effects on480
tree and stand growth response of primary Picea abies forest to climate. Forest Ecology and481
Management 354, 77–86.482
R Core Team, 2015. R: A language and environment for statistical computing. R Foundation483
for Statistical Computing. Vienna, Austria.484
©2016 NRC Canada
Page 20 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
20 Can. J. For. Res. Vol. 99, 2016
Royal Meterological Institute of The Netherlands (KNMI), 2014. CMIP5 Scenario Runs485
- Surface Variables [online]. http://climexp.knmi.nl/selectfield\_cmip5.cgi\?id=486
someone@somewhere. [accessed 14 jun 2014].487
Sabatia, C.O., Burkhart, H.E., 2014. Predicting site index of plantation loblolly pine from488
biophysical variables. Forest Ecology and Management 326, 142–156.489
Schwarz, G., 1978. Estimating the dimension of a model. Ann. Stat. 6, 461–464.490
Seynave, I., Gégout, J.C., Hervé, J.C., Dhôte, J.F., Drapier, J., Bruno, E., Dumé, G., 2005.491
Picea abies site index prediction by environmental factors and understorey vegetation: a492
two-scale approach based on survey databases. Canadian Journal of Forest Research 35,493
1669–1678.494
Sharma, R.P., Brunner, A., Eid, T., 2012. Site index prediction from site and climate variables495
for Norway spruce and Scots pine in Norway. Scandinavian Journal of Forest Research 27,496
619–636.497
Sigurdsson, B.D., Medhurst, J.L., Wallin, G., Eggertsson, O., Linder, S., 2013. Growth of498
mature boreal Norway spruce was not affected by elevated [CO2] and/or air temperature499
unless nutrient availability was improved. Tree Physiology 33, 1192–1205.500
Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia,501
Y., Bex, V., Midgley, P.M. (Eds.), 2013. Climate change 2013: The physical science basis.502
Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental503
Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom and504
New York, NY, USA.505
Thomson, A.M., Calvin, K.V., Smith, S.J., Kyle, G.P., Volke, A., Patel, P., Delgado-Arias, S.,506
Bond-Lamberty, B., Wise, M.A., Clarke, L.E., Edmonds, J.A., 2011. RCP4.5: a pathway for507
stabilization of radiative forcing by 2100. Climatic Change 109, 77–94.508
Thornthwaite, C.W., 1948. An approach toward a rational classification of climate. Geograph-509
ical review 38, 55–94.510
Tveite, B., Braastad, H., 1981. Bonitering for gran, furu og bjork. Norsk Skogbruk 27, 17–22.511
©2016 NRC Canada
Page 21 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 21
Weiskittel, A.R., Crookston, N.L., Radtke, P.J., 2011. Linking climate, gross primary produc-512
tivity, and site index across forests of the western United States. Canadian Journal of Forest513
Research 41, 1710–1721.514
©2016 NRC Canada
Page 22 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Page 23 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research
Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 23
SI (m)
num
ber
of o
bser
vatio
ns
0
200
400
600
800
1000
6 8 11 14 17 20 23 26
SprucePine
Deciduous
Soil Wetness
num
ber
of o
bser
vatio
ns
0
500
1000
1500
2000
2500
Wet Mod. Dry
Fig. 2. Site index (left) and soil wetness (right) distribution by species dominating the stand.
©2016 NRC Canada
Page 24 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research
Draft
24 Can. J. For. Res. Vol. 99, 2016
Fig. 3. Maps of the distribution and range of the temperature and moisture variables included in themodels. The maps are interpolations of the climatic data at the plots, masked to show only productiveforest area in Norway. TSUM (in ◦C) is the sum of the 30-year average temperatures in April, May,and June. MSJ (in mm) is the moisture surplus in June, defined as the difference between 30-yearaverage precipitation in June and average potential evapotranspiration in June.
����
����
���
��
����
���
��
�
��� ���
����
���
�����
�����
�����
��
����
�����
©2016 NRC Canada
Page 25 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 25
Fig. 4. Sensitivity graphs for the spruce model (md2).
TSUM (ºC)
SI (
m)
10
15
20
25
10 20 30 40
SQ2
SQ3
SQ4
SQ5
©2016 NRC Canada
Page 26 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
26 Can. J. For. Res. Vol. 99, 2016
Fig. 5. Contour plots showing the behavior of md2 for pine dominated stands. Grey rectanglesindicate combinations of MSJ, TSUM and SQ that existed in the fitting dataset.
TSUM (ºC)
MS
J (m
m)
−50
0
50
10 20 30
78
9
SQ1
7
89
1011
1213
SQ2
78
910
1112
1314
SQ3
10 20 30
−50
0
507
8 910
1112
1314
1516
17
SQ4
©2016 NRC Canada
Page 27 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research
Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 27
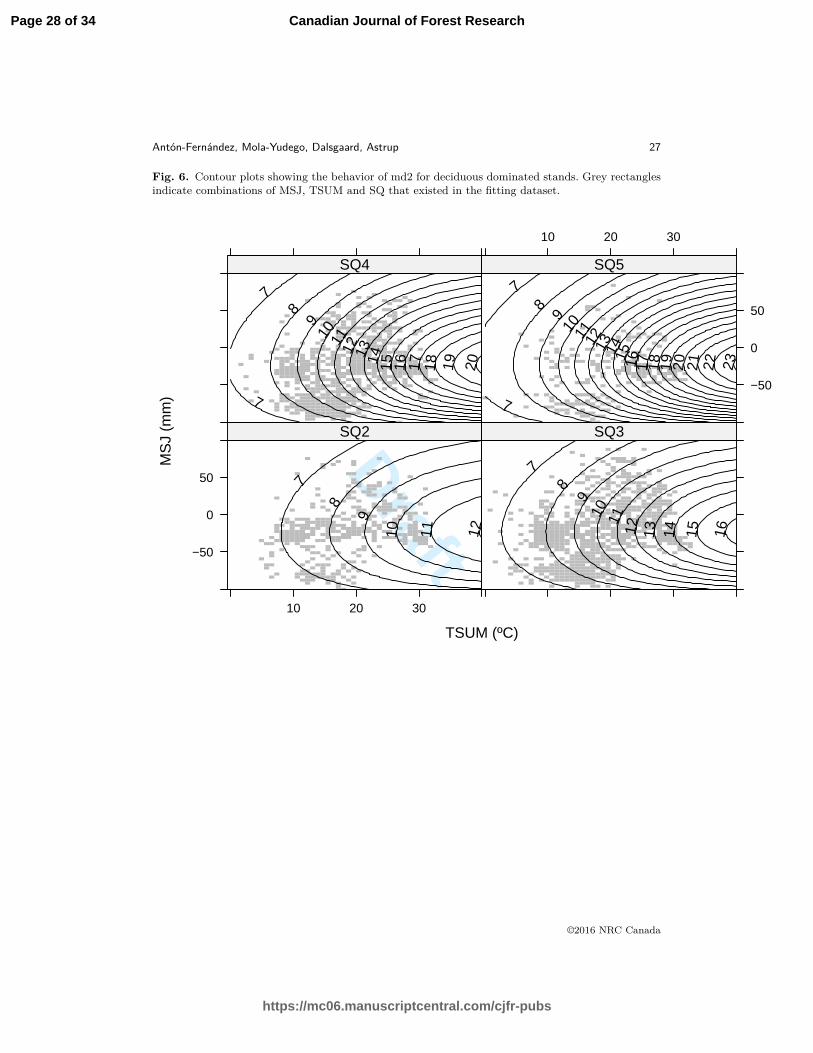
Fig. 6. Contour plots showing the behavior of md2 for deciduous dominated stands. Grey rectanglesindicate combinations of MSJ, TSUM and SQ that existed in the fitting dataset.
TSUM (ºC)
MS
J (m
m)
−50
0
50
10 20 30
7
8
9
10 11 12
SQ2
7
89
10 1112 13 14 15 16
SQ3
7
78
910 11 12 13 14 15 16 17 18 19 20
SQ4
10 20 30
−50
0
50
7
7
89
10 111213 14 15 16 17 18 19 20 21 22 23
SQ5
©2016 NRC Canada
Page 28 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
28 Can. J. For. Res. Vol. 99, 2016
Fig. 7. Residuals of the best model for each species against the predicted SI. A loess smooth functionwith span smoothing parameter = 2/3 and degree of local polynomial = 1 is shown in black.
Predicted SI (m)
Res
idua
ls (
m)
−10
−5
0
5
10
10 15 20
spruce−10
−5
0
5
10
pine−10
−5
0
5
10
deciduous
©2016 NRC Canada
Page 29 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 29
Fig. 8. Distribution of TSUM and MSJ under current climate (C) and RCPs 4.5 projections (F).
TSUM
MS
J
−100
−50
0
50
100
0 10 20 30 40
FFFFF
F FF
FF FFF
FFFF FFF
FF FFF
FF FFFF
F FFFFFF
F FF FFF FFF
FFF FFFF
FFFF FFFF
FFFF F
FFFFFFF F
F FF FFFF FFF
FFFFFFF
F
F
FF F
F
FF
FF
FFF
FFF
FF
FFF F
F FF FFF
FF
FFF FFF F
FFF
F FFFF
F
FFFFF
F FFFFF FFFFFFFF F
F F FF FFF F
F
F
FF
FFFF
F F
F
FF
FF
F
FF
FF
FFF
FFFFFFF
F
F FF
FF FFF
FFFFFF FFFF
F FFF
F
FFF
F F
FF
FFF
FF FF FF
FFF F F
FFFF
FFF
FFF
FF
F
FF
FFF FF
FF F
F
FF
FF FFF F
FF
FF
FF FFFF
FF
FF
F FFFFF
FFFFFFF F F
F FF
FFFF
F
FF
FF
F
F
FF
F
FF
F F FFF FF
FF
F
F FF
FFF
F
F
F FF
F
FFF
F
FFF
FF
FFF FF FF
FF F
F
FF
FF
F
F
FF
FF
F
F
F
FF F FFF
FFF
FF FFFF
FFFF FFF F F FF
FF
FFFF FF
FF
F
FFF
FF FF F
F
FF FFF
FFF
F
F
FF
F
FF FF F
FF
F
FF
FF
F F
F
F
FFF
F
FF
F
F
FFF
FF FFF F
F
FFF
FF
FFFF
FFFFF
F
F
FFF
FFF
FFF
F
FF
F
FFFF
FF
F F
FF F
FF
F
FF FFF F
FFF
F
FFF
F F
FF FF
FFF
F
FF FFF
F FFFFF F
FF F
FF FF
FFF
F
F
FFF F
F F
F FF
F
FF
F
F
FF
F FFFF
FF
F
F
FF FF
FF
FF
FF F
FF
F FFF
F
F
F
FF
FF
F F
F
FF F
F
F
F
F
F
FFF
F
F
FFFF F
FFF
F
F FF
FF
FF
F
FF
FFF
FF
F
F
F
F
F
F
F
F
FF
FF
F F FFF FFFF
F F
FF
F
FFF
FFFF F
FF
FF
FFFF F F
F
FF
FF
F FF F F F FF
FFF
F F
FF
F
F
F
FF
F FF
FF F
FF
FF F
FF F
FFF
FF
FFFFFF
FFFF
F FFF FF F
F
FFFF
FF
FF
F FFF FF FF
FF F
FF
F
F
F
F
FF
FF
FF
FF
FF F
FF
FF
F
FFFF
F FF
F
FF
FF
FF
F
F FF
F FF
FFF
FF F
F FF
FF
FF
FF FFFF
FFFF
FF FFF F
F
FFF F
F F
F
F
FF
FFFF
FF F
F FFFF
FFF
FF
FF
FF
FF
F
F
FF
F
F
F
F
F
FF
F
FF
FF
FF FF F FF
FF
FF
F FFF
F F
F
FF
F
FF
F FF
FF
FFF
FFF
FFFF
FF
FFF
FFFF F
FFF
F
F
FF
FF
F
FFF
FFFF FF
F FF
FFF
FF
FFF
FFF
FFFF
F
F
FF
F
F
F
FF
FF
FFF F
F
F F F
F FFFF
FFFFF
FFFF
FFF
FFFFFFFF
FFFFF
FFFFFFFF FFF
F FFFFFF
FF FFFFFFFFFF FFFF
FF FFF F FFFFF FFF FFF F
FFFFFFF FFF F
FFFF
FFF FF FFFF
FF
FFF
FFFFFFFFFF
F F
F FFFFF FFF FFF FFFFFF FFFFF FF FFF FFFFF
FF FFF FFFFFFFFF FF FFFFF FFF FFF F
FFF FF
FFFF FF FF
F
F FF FF FFF FFFFFF FF F FFFFFFFFF FFFF FFF
FFFF
FFFFF
F
FFFFFFF
FF FFF FFF FFF
F
F
FFF
FFF FF FFFF FFF F FFFFFF FFFF
FF FFF FFF FFF FFFFF F FFFF FFFFFFF
FFFFFF FFFF
FFFFFF F
FFF
FF FFFF
FFFF
FFFFFFF
FFFFFF
F FFF FFFFFF
FFF FFFFF FFFFFF FFFFFF FF
FFF
FF FFFFFF FFFFFFFFFFFFFF
FFFF
FFF
FFFFF FFFFFFFFFFFF FF FFFFFFFFF FF
FFFFFF FFFFFFFFFF FF FF FFFFFFF
FF
FFF
FF
FF
FF
F FFFF
FF
FFF FFFF
F FFFF
FF FFF
FFF FFFF
FF F
FF F FFFFFF
FFFFFF
F FF
FFF
FF FF FFF FFF F
FF FF FFFFFFFFF FF FF FF FFFFFFF FFFFFF FFF FFFFFF FF FFFFFFFFFF FF FFF FF FF FFF FFFF FFFF
FF
F F FFFFF FFF FFF FF FF
F F F FF FF
FFFFF
FFFFFFF FF F FFFFFF
F FFF FFFF FFFF
FF FFFFFFFFFFFFF FFFFFF FFF FFF FFF FFFFFFF
FFFFFFF FFFFF FFF FFFF
FFFF
FFFFFF
FFFF F F
FFFF
FF FF FFFF
FFF FFF
FFFF FF
FF
FF
FFFFFF
FFF
FFF
FF FFF FF F F
FFF
F FFF FFFFFFF
FFFFF F
FFFFFFFF
FF FFF
FF
FF FFF F
FF FFF FFF F F
F FF FFFFFFFF
FFFF
FF FFF
F FFF
FFF
FFFFFF
FF
FF FFFF
FFFFFFFF F FFFF F FFFF
FF FF FF F
FF
FFF
FFFFF
FFFF F
FF FF FFFF
FFF
FF FFF FF
FF F FF
FFF FFF F FF
FFF
FFF FFFFF F FFF FF FFFF FF
FFF FF FFFFF
FF
FF
FF FF F F
FFF
FFFF
F
F FFF F
FF
FFF F
FF FFF
FF FF
FFF
FF FF
FFFFF
FFFFF F FFFFF
F
F
FFF F
FF
FFFF FFFFFFF
F FFFF F
FFF
FFFF F F
FF
FF
F
FFF F FF
FFF
FF F
F
FFFF
FF FF FF
F FFF
FF FFF FFFFF
FFFF F FFF
FFFF
FF FFFF FF
F FFFFFF
FF F F
FFFF FFFFFF FF
FFFFF
F FFFFF FF FF
FFFF
FFF
F FFFFFF F
FF F
F FFF FF
FF
FF
F F FFFFFF FFF F FF
FFF
FF F FFF
F F FF FF
FFFF FF FF FF F
FF FF F FF FFFFFFFF FF FFFFFF FFF FFFFFF F F FFFFF F FF F
FFFFFFF F F FFF F FFFFFF
FF FF
FFFFFF
F F FFF
FF FFFF FF
FFFFFFF FFF FFF
F FFF
FFFF FFFFF F
F
F FF FF
F FFF FFF
FFF
FFFF F FF FFF
FF
FF
FF
F
FF FFFF FFF
FF FFFFFF
FF
FFFFFFFFFFFFF
FFF
FFFF FF
F
F FF F
FFFFF
F F FFFFFFF FFFFF FF FFF F
FF FF
F FF FF F
FF
F
F F FF FFF FF FFFFF FFF FFFFFF F
FFFF
FF FF
FFFFF F F FFFFFF
FFFFF
F
FF
FFFF FF FFFF
FFF FFF FFFFFF
FFF FFFF F FF
FF F
FFFF FF
FF
FFF F FF FFFFFF FFFFFFFFF
FF
F FFFFF F
FFFF F
FFFF
F
F
FF
FFF FFF
FFF
FFFF
FFFF
FFFFFFF
FF FF
FF F F FFF F
FFF
FF
FFF
FFFFF FF F
F
F
F
F
F
F
F
FF F FFF
FF
FFF
FFF
FF
F
F
F F FF FFF FFFF
FFFFF
FFFF
FFF FFFF FF
FFF FFFFF
FFFFFF
FFFF
F FF F
FF F
FF FF F FF FF
FF
FFF F FFFFF
FF
FF FF
FF
FFFF
FF
FF
FF
F
FF
F FFFF
FF FFFF
FFF
F FFF FFFF FFF
F
FFF F
F
FFFFF
F FFF
F FF
FFF
F F
FF
FF F
FFF
FF
FF
FFF
F
F FF
FFF
FF
FFFF
F
FFFF
FFF
FFF F
FFFFFF
F FFF
FF FF
F
FFF
FF F
FF
F FFF FFF
FF
F
F
FFF
FF
FF
FF
FFF
FF
FF
FFFF FF F
FFFFF
FFFFFFF FFF
FFF
FF FF FFF F FF FF
FF
FFFFFFFFFF
F FF
FF
F FF FFF F
F
FF
F
FF
F FF
F F F FFFF FFF
F
FF
FF FF
FFFFFFF
F
FF
F FF FFFFF
FF
FF
F
FFFFFFF
F FFF FF FFFF
F
FF
FF
FFF
F
F
F
FF
FFFF
FF
FFF
FFFFF
FFFFFFFF FFFFFFFFF
F
FFFFF
FFFF FFFFFFFFF FF
F
FFF FFF FFFFFFF FFFFFF FFFFFFF FF
FF
F FFF FFFF FF
FF
FF FF
F FF FFF
FF
FF
FF FFFFFFFFFFFF
FFFFFFFFFFFFFFFF
F
FFFF
FFFFFFF
FFFFFFFFFFF
F FFFFFFFFFFFFFFFF
FFFFFFFFFFFFFFF FFFFFFFFFFFFFFFFFF FFFFFFFFFF FFFF
FFFFFFFFFF
FF F
FFFFF
FFF
FF F
F
F FF F FFFFF
FF
FFFFF FFF F F FFFF
FFF FF
FF
FF F
FFF F FFFF FF
FFFF FFFF
F F FFF F F FF FFFFF
FF
FFF FFF FFF FF FF FFFF
FF FF
FFF
FFFF FF FFF FF FFF
FF
F
FF
F
FFFFFF
FFFFFF FFF FF FFFFF FF
FFFFF FFF F F F
FF FF FF FFFF F FFFFFFFFFF
FFFFFFFFFFFF
FFFFFF
FFFFFFFFF FFFF FFFF FFFFFFFFF
FFFFFFFFFFFFFFFFFFFFF
FFFFFFFFF
FFFFFFFF FFFFFFFFFFFFFFFFFFFFFFF
FFFFFFFFFFF FFFFFFFF
FFFF FFFFFFFFFFFFFFFFFF FFFFF FFFFFFFFFFFFFFFFF FFFF FFFFF FF F F
F F F F FFFFFFFFF
F F FFFFFFF
F FF
F FFFFF FFFF
F
FF FF F
FF
FF FF F
FFF
F FFF FFFFF
FFFFFFF FF FFF F
FFFFF FF FFF
F FF
FFF FFF FFFFFFF
FF F
FFF
FF FF
FF
F
F FF FF FF FF F
FFFF FF
FFFFF
F
FF FF
F FFFFFFF FFF FF
FFF FFFF
FFF
FFF
FFFFFF
FF F
F
F FFF
FF
FF F
FFFFFFFFF
FF FFFFFFFFFFFFFFFFFF
FFFFF
F FFFFFF
FFFFFF FFF FFFFFFFFFFFF FFFFFF FFFFFFF FFFFF FFFFFFF
FFFFF FFF
FFFFFFF
FF
FF
FFF
FFFFFF
FFF
F FFF
FFFF
FFF F
FFFFF FFFF FFF FFF FFFFFFFF FFF FFFFFFFFFFFF F FFFFFFFFFFFFF
FFFFFFF
F FFFFFFF FFFFFFFFF
FF FF FFF FFFFFF FFFFFFFFF
FFF F
FFFF FFF FFF FF FFF FF
FFFFFFF F
FF
F F F FF FF FF FF
FFF FFF FFF F
F FFFFF FFFF
FF
F
FFF
FF FFF
F
F
FFFF
FF
FFFFF
FFFFF FFF
FF FFFFF F FFFF FFFFF
FF
FF FF F
FF FFFFF
F FFFFF
FFFFFFF F
FFF FF
FFFF
FFF FF FF
FF
FFF
FFFFFFFFF FF
FF
F FFF FF
FFFFFFF FFF
F FF FFF FFFFFF FF F FFFF FFFF
FFF
FFFFFFFFFFFF
FFFFFFFF F FF FF
FFFFFFFFF F FFFF FFFFF F
FFFFFFF F
FFF FF FFF FFFF F FFFFFF FFFF F FFF
FFFF
F
FF
FFF FFF
FFFFFFFFF FF F
F F FFFF
FFF F F
F
F FF
FF
FFFFFF
FFF FFF
FF FFFFFFF F FFFFF FFF
FFF FFFFFFFF F
FFFFFF
FF FFFFFFF FF FFFF FFFFF FFF FFF F FFF
FFFF FFF FFFFF FFFFFF FF FFFFFFF
FF FF
FFF FF FFFF FFF
FFF
FF FF FFF F FFF
FFF FF
FF F
FFF
FFFFFF
FF
F FFFFFFFF FFF F
FFF F FF FF
FFFF
FFF
FF FFF FF FFFFFF FFFF FFF FFFFFF FFFFF FF
FFFF F FFFFF FFF FF
FF FF FF FFF FFFFFF
F FFFFFF FFFF
FFFF FFFF F
F FFF FF FFFFFF F
FFFF FFFF FFFF FF FF FF F
F
FF
F FF
FFFFFF FFFFF FFFFFFF F FF FFFF F
FFFFFF F
FF FFFF
FFF
FFFF FF FFFFFF FF
FFFFFF
FF
FFFF FFF
FFFF FF
F FFFFF
FF FFFFFFFFFF FFFFFFFF FFFF FFFFFFFFFFFFF FFF
FFF
FFFFFF
FFFF F
FFFFFFFFF FFFFFFF FFFF FF FF FFF F
FFF FF F FFFFFFFFFFFFFFFFFF FF FFF FFF
FFFFFFF FFFFFFFF FFF FF FFFFF FFFFF
FFF FFF FFF
FFFFF
FFF FFFFFF FFFFFFFF FF FFFF FFFF FFF FFF FF
F FF F
FFF
FF FFFFFFFF FF
FF FF FF
FFF
FF FFF
FFF FFFFF FFFFFF
F FF FFF FFFFFFF FFFF F F
F FF FFFFFF FFF
FFFFFF
FFFFF FF
FFFFFF
FF
FF
FFFF
FFF
FF
FFFF FFFF
FF FF FFFF F
FFF
FF F FF
FFF
FFF FFF FFFFFFFFFF F FFFF F FFFF FFF
FFF
F FF FFFFFF FF
FFFFFFFF FFF
FF FF
FFFFF FFFFF F F
F F FFFFFFF FF
FFF FFFF
FFF FF
FFF FF FF
FFF
FFFFF
FFFF
FFF
FFFFFF FFF F
F FF FFFF FFF FFFFFFFFFFFFFF
FF FF FFFFFF
FFF FFFFF
FFFFFFFFFFFF
FF
FFFFFFF FFF
FFFFFF F FFF FFF FFF F
FFFFFF
FFFFFFFF
FFF
F FFFF F
FF
F FFF
FFF
FF
FFF FFFF
FFF
FFF
F F
F
FFF
F
F
FF
F
FFF
FF
FF
F
FFFFFFFF
FF
FF
FFFFFF FF
FF
FF
FF FF F
FF FFFFFF F
F FF
FFF
F FF
FFFF
FF
FF
FFFF FFFF
FFF F
FF
FFF
FF
FF F FFF FFFFFF F FF
FFFF FF FF
FF
FF
FFFFF
FFF
FFFF FF
FFF FFFFFFFF
FF FF
FFFF FFFF FF
FF FF FF
F FF
F FFFF F FFF FFF FFFF FF
FF
FF
F FFF FFFFF FFFFFFFFFF F F
F FFFF FFF FFFF FFFF FF FF FF FFF FF
FFF
FFF
F
F
FF FFFFF F F FF F
FFFFF
FFFF
FF FF F FFFFFF F
FFFFFFF
FFF FF F FFFF FF F FF FF
FFFFFF F
FFFF FFFF FFFFFFF F FF
FFFFF F FF F FFFFFFF
FFFFFF
FF FF
FFF
FF
FFFFF F
F FFFFF
FFF
F FFF FF FF
FF F FF
FFFFFF FFFF FFFFF F FFF F FFFFF F FFFF FFFFFFFFF FF
FFFFFFFFFF FFFFFF
FFFF
FFFFFF FF FF FFFFF FF FFFFFF F FFF FFF FF FF FFF FFF FFFFFFF FFFF F
F F FFF FF
F FFF
FF FF F FF
FFF
F
FF F FF F
F F FFFFF F F F FFF FFFFFFFF FFF FF FFF F
FF FFFFFF FF FFFFFFFF FFF
F FFF FFFFFFF
FF FFF
FF FFF
FF FF F F F
F FFF
FFF FF F
F FFF FFFFF FF F FFFFFF FFFF FF FFFF
FFFFFF FFFF FFF
FFF FFFFFF F
FFFFFFF
F FFF
FFF
FFFF
F FF FF
FFFFFFFFFFFF
FFFF FFFFF
FF
FFFFFF
FFFFFF FFFF FFFFF
F
F
F
F
F FFFF FFFFFF FFF FFFF
FFFFFFF F FF FF FF FFF
FF
FFFFFFFFFF
FFF FF
FFF
FF F
FF FF FFFFFF FF
FF
FFF
FFFFF
FF FFF F FF F
FFF
F
F FF FF FF FF
FF FF FFFFFF
FFF FFFFF
FF FF FFF F FFF FF FFFFF FFFFFF
F FF FFF F FF FFFF FFF
FFFFF FFF F
FFFF FFFF FFF FF
FFFF
F FF FFF
FF FFFF
FFF FFFFFF
F F FF
FFFFFFF FFFF
FFFF
FF
FF FF F
FF
F
F
FFF
F
FF FF
FFFF FFF
FFFF
FF FF
FF
FFFF
FF
FF
FFFF FFF
FFFFFF
FF
FFFF
FFFFFFFFFFFF FFFF
FFFF FFFFF F
FFF FFFFF
FF
FF
FFF FFFF
FFFF
FF
FFFF FFF
FFFF FF
FF F
F FFF
FFF
FFFFFFF F
F F
FF
FFF
FFFF FFFF
FF
FFFF
F FF
F FFFF
FFFF
F
F
F
FF
FFF
F
FF
FFFF F
FFF FFF
FFF FF FFF
FFFFF
FFF
FFFFFF FF
FFF
F
F
FF
FF
FFF
FF
FFFFFFFFF
F
F
FF
FFFFFFF FF
FFF
FFFF
F FF FFF F
FFF
F FF FFFF
FF
FFF
FFFF
FFF
FF FFF FFF
F
FF
F
FF
FFFF
F
FF
F
FFFFF
FF F
FFFF FF
FF F
FFFF
FF
FF FFF
FF
FFFFFFF
FF FFFF FFFFF FFFFFF FF FFFFFFF
FFFF FFFF FFFFFFFF
FF FFFFF FF FF FFF FFF F FF
FFFFF
FF FFFFFFF FF FFFFFF
FFFFF
FFFFF
F
FFFF FF
FF
FF
F
F
FF
FF
FF
FFF FFFFF
F FF
FFF
F
FFF
FFF
F
FFFFFFFFFFFFFF FF FFFFFFFFF FFFF FFFFFFFF FFFFF FF FFF FFFFFFF FFFFFFF F
FFFFFFF FFFF FFFFFF FF FFFFFFFFFF
FFFFFFF FFFF FF
FFFFFFF
F FFF FFFFFFFFF
FFFFFF FFFF FF FFF
FF FFFFFF F
FF
FFFFFFF FFFF FFF
FFF
FFFFFFFFFFFFFFFF
FFF
FFFFFF
FFFF
F
FFFFF
FFFF FFFF FFF F FFFF FFFFFFF
FFFFFFFFFFFFFFF F FFFFF F
FF FFF FFFF FFFFFFFFFFF
FFF FFFF FFFFFFFF FFF F
FF FF FFF FFFF F FF FF FF FFFF FFFFFF FFFFFF FF FFFFFFFFFF F FF F
FF
FFF
FFF F FF F FF F F
F FFFFFFFF
FFF FF FFF FFFFFFFF FFFFF FFFFFFF FFFF FFFFFFFFFF F FFFFFFFFFFF FFF FF FF FFFFFFFFFF FFFFFF FFFFF F FFF FFFFFF
FFF FFF FFF FFFF F FFFFFFFF FF FFFFFFFFFF FF FFFFF FF FFFFF
FF FF
FF FFF F FFF FFFFF
FF FF
FFFF FF FF
FF F
F
FF
F
FF
FF
FFF FFFFF FFFF
F
FFF FFFF F
FFF
FFFFFFFF
FFF
FF FF FF
FFFF FFF FF
FF
FFFFFFF
F FF FFFF
FF
FF FF FFFFFFF FFFF F
FFFFFFFF
F FFFFFFFF
FFF
F FF FFFFFF FFFFFFF FFF F
FFFFF FF
FFFF F FFF
FFFF
FFF FF FFFFF
FF FFF
FFFF FFFFF FFFFF
FFF FFFF F FFF F FFF FFF
FFFFFFF
F
FF
F
F
F
FFFFF
FFF
F
FFF FF FF FF FFFFF
F
FF
FF
F
FFF
F
FFFF
FF FFF
F
F
F
FFFFF
F
FF
FF
F FF FF
FFFF FF FFFFF FF
FFFF FF
F
FFFFF
FFF
FFFFFFFFF
F FF
FF
F FF
F
FF
FFF
FF
F
FF
FFFF FFF
FFFF FF
F
F
F
FF
FF
FF
F
F
F
F
FFF
FF
FF
F
FF
FF
F
FFFF F
F F FF
FF
FF
FFF FF
F
FF FF F
FF
FF
F
F F
FFF
F
F
FFF
FFF
FFF FFFFF
FFFF FF FFF
F FF FF
FF FF FFF FF
FF
FF
F F FF
FFF
FF F
FFF FF FFFF
FF F FF
FFFF F
F
FF
FFFF
FFF
FFF
F
FF
FFF FFFFF
F
F
FF
FF
F
F
F
FFFF
FFFFF FF
F FFFFFF
F
F
FF
F FFFF
F F
FFF
FF
FF
F
FFF
FF
FFFF
F
F
F FFF
FF
F
FFFFF
F
FF FF
F
FFF
FF F
F
F
FF
F F
F
F
FF
F
F
FFFFFF
FFFFF F FFFF FFFF
FFFFFF FFF FFFF F
FF
F
F
F
F
FF
FF
F FFFFF
FF
FF FFFF
F
F FFF FF
FF
FFFF
F FF
FFFFFF
FF
FFFF F
FFFF F
FFF
F
FF
F
FF F
FF
FFFF
F FF
F FFF FF F F
F
F
F
F
F
F
F
FFFFF
FFFF FFFF FF
FFF
FFFF F F
F F
F
FF
F
F
FF F
F
F
FF FF
FF
F
FF
F FFF F F F
FFFF
F FF FF
F FFF F
F
FFF
F F F
FF
FFF
F
F FFFFF
FF F
FF
FF FFF FF
FFFFF
F FFF
FFFF FFF F
FF
FFF
F
FFF
FFFF FF FF
FFFFF FFF FF
F FF FFF
FFFFFFF
FFFFFFF FFFF
FFF F
FF F
F FFFF
FF
F FFFFFF
F
F
F
FFF
FF
FFFFF
FFF
FF F
F FF FF
FFFF F
F FFF FFF
FF
FF
FF F
F
F
F
FF
FF FFFF F
F FF
FF
F
F FF
F
FF F FFFF
FFF
FF
FF
FF
FFFFF FFFF FF
FF FFF FF
FFF
F FF F F
FFFFFF F
FF
FFF
F FFF FFF F
F
FF
FFF
FFFF
FFFF
FFFF FFFF
FF
F
F FF FF
FF FF FFFFFF
FF
F FFF
F FFFFFF FFF FFF F
FF FF
FF
FF F
F
FFFF FF
F FFF F
FFFFF FF F FF
F FF FFF FFF
FF F FF FFFF FF
FF
FFF
F
FFF
FF F FFFF FFF
FFF
FF
F FF
FFFF F
F F FFF
FF
FFF F
FF
F
FFF
FF
F
FFF
FFFFF
F
FFFFFF
F
FF
FF FFF
FF
FFF
F
F FFFFFFFF
FF
FF
FF FFFF
F F FFF FFFF FF F FF FF FFFFFFFF
FFF FFFFF
F FFF
FF
F
FF
FF
FFF
FF F
FFFFFFF
FFF F
FF
FFF FFFF FFF
F FFFFF
FF
FF FF
FF FF
FF
FFF FFFF FF
F F FFFF FFF F FFFFF FF FFFFFFFF
F FFFFFF FF FFF FFF FFF
F FFFFFF
F F F FF
FF FFFFFF FFF FF
FFFFF
FFF F
F
FFFF
FF
FF
F F
F
F
F
FF FFF
FF
FF
FFF FF
FF
FFF F FFF
FF
FF
FFFF
F
FF
FFF FFF
F
FFFFFF FF
FFFFF FF F
FF
FFFF F
FF
FF FF FFF F F FFFF
F F FFF
FF
FF FFF F
F FFF
F F
FF FFFFF
FFFF FFF F FFFFF
CCCCC
C CC
CC
CC
CCCC
C CCC
CC CCCCC CCCC
C CCCCCC
C CCCCC CCC
CCC CCCCCCCC CCC
CCCCC C
CCCCCCCCC C
CCCCC CCCC
CCCC
CC
C
C
CC C
C
CC
CC
CCC
CCC
CC
CCC C
C CCCC
CCC
CCC C
CC C
CCC
C CCCC
C
CCCCC
C CCCCC CCCCCCCC C
C C CC CCC C
C
C
CC
C CCC
C C
C
CC
CC
C
CC
CC
CCC
CCCCCCC
C
C CC
CC CCCCCCCC
CCCCC
CCC
CC
CC CC C
C
C
CCC
CC CC CC
CCC C C
CCC
C
CCC
CCC
CC
C
CC
CCC C
CC
C C
C
CC
CC CCC C
CC
CC
CC C
CCCC
CC
CC CC
CCC
CCCCC
CC C C
C CC
CCCC
C
CC
CC
C
C
CC
C
C C
C C CCC CC
CC
C
CCC
CC C
C
C
CC
CC
CCC
C
CCC
CC
CCCCC CCCC C
C
CC
CC
C
C
CCC
CC
C
C
CCC CCC
CCC CC CCCC
CCCC CCC C C CC
CC
C CCC CC
CC
C
C CCCC CC C
C
CCCCC
CCC
CC
CC
C
CC CC C
CC
C
CC
CC
C C
C
C
CCCC
CC
CC
CCC
CC CCC C
CCCC
CCCC
CC
CCCCC
C
C
CCC
CC
CCCC
C
CC
C
CCCC
CC
C C
CC C
CC
C
CCCCC C
CCC
C
CCC
CC
CCCCC CC C
CC CCC C CCC
CC C
CC C
CC CC
CCC
C
C
CCC C
C C
CCC
C
CC
C
C
CC
C CCCC
CC
C
C
CC CC
CC
CC
CC C
CC
C CCC
C
C
C
CC
CC
C C
C
CC C
C
C
C
C
C
CCC
C
C
CCCC C
CCC
C
C CC
CC
CC
C
CC
CCC
CC
C
C
C
C
C
C
C
C
CC
CC
C C CCC CCCC
C C
CC
C
CCC
CCC
CC
CCCC
CCCCC C
C
CC
CC
C CC CC CCC
CCC
CC
CC
C
C
C
CC
C CC
CC C
CC C
C CCC C
CCCCC
C
CCCCC
CCCC
C CCC CC C
C
CCCC
CC
CC
C CCC CC C
CC
CC
CC
C
C
C
C
C C
CC
CC
CC
CC C
CC
CC
C
CCC CC CC
C
CC
C
C
CC
C
C CC C CC
CCC
C
C CC C
C
CC
CC
CC C
CCC
CCCC
CCCCC C
C
CCC C
C C
C
C
CC
CCC
CC
C CC CCCC
CCC
CC
CC
CC CC
C
C
CC
C
C
C
C
C
CC
C
CC
CC
CCC
C C CC
CC
CC
C CCC
CCC
CC
C
CC
C CC
CC
C CCCCC
CCCC
CC
CCC
CCC
C C
CCC
C
C
C
CCC
C
CCC
CCCC CCC C
C
CCC
CC
CC CC
CCCCC
CC
C
CC
C
C
C
C
C
C
CCC
C CC
C C C
CC
CCCCCC
CCCCCC
CCCCCCCCCC
CCCCCC
CCCCCCCC C CCCCCCCCC
CC CCCCCCCCCCCCCC
CC CCC C CCCCC CCCCCCC
CCCCCCC CCC C
CCCC
CCC CC CCCCC CC
CC CCCCCCCCCC
C C
C CCCCC CCC CCC CCCCCC CCCCC CC CCCCCCCCCC CCC CCC
CCCCCC CC CCCCCCCC CCC CCC
C CCCC
CC CC CCC
C CC CC CC
C CCCCCC CC C CCCCCC CCC CCCC CC
CCCC CC
CCCCC
CCCCCCC
CC CCC CCC CCC
C
C
CCC CCC
CCCCCC CCC C CCCCCC CCCCCC CCC C
CC CCC CCCCC C CCCC CCCCCCC
CCCCCC CCCC CCCC
CCCC
CCCC CCCC
CCCC
CCCCCC
CCCCCC
CC CCCCCCCC
CC
CC CCCCC CCCCCC CCCCCCCC
CCC
CC CCCCCC CCCCCCCCCCCCCC
CCCC
CCCCCCCC CC
CCCCCCCCCC CC CCCCCCCCC CC
CCCCCC CCCCCCCCCC CC CC CCCCCCCCC
CCCC
C CC
CC C CCCCC
CCCC CCC
C C CC CCCC CCC
CCC CC
CC
CC CCC C CC
CCCCCC
CCCCC CC
CCC
CCCC CCC CCC CCC CC CCCCCCCCC CC CC CC CCCCCCC CCCCCC CCC CCCCCCCC CCCCCCCCCC CC CCC CC CC CCC CCCC C
CCCC
CC C CCCCCCCC CCC CC C
CC C C CC C
CC
CCCCCCCCCCC CC C CCCCCC
CCCC CCCCCCCC
CCCCCCCCCCCCCCC CCCCCC CCC CCCCCCCCCCCCC
CCCCCCC CCCCCCCCCCCC
CCCC
CCCC
CCC
CCC C C CCCC
CC CCCCCC CC
C CCCC
CCC CC
CC
CC CCC CCC
CCC
CCCC
C CCC CC C C
CCC
C C CC CC CCCCCCCCC
C CCC CCCCCCC
C CCCC
CCC CCC C
CC CCC CCC C C
C CCCCC CCC
CC
CCCCCC CC
C
C CCC
CCCC
CCCCCCC CCCCCC
CCCCCCCC C CCCC C CCCCC
C CC CCCC
CC
CCCC
CCCCCCC C
CC CC CCCC C CC CC CCC CC
CC C CC
CCC CCC C CC
CCC
CCC CCCCC C CCCCC CCCCCC
CCC CC CCC
CCC
CCC
CC C
C C CC
CC
CCCC
C
C CCC C
CCCC
C C
CC CCC
CCCC
CCC
CC CC
CCCCCCC
CCC C CCCCCC
CCCC C
C CCCCC CCCC
CCCC CC
CC CC CC
CCCC C C
CC
CC
C
CCC C CC
CCC
CCC
C
CCC
CCC CC CC
C CCC
CC CCC CCCCCCC
CC C CCCCCC
C CC CC CC CCCCCCCCC
CC C CCCCC CCC
CCC CCC
CCCCC C CCCC CC CC
CCCCCCC
C C CCCCC C CC CC CCC CC
CC
CC
C C CCCCCC CC
C C CCCC
C CC C CCC
C C CC CC
CCCC CC CC CCC CCCC CCC CCCCCCCC CC CCCCCC CCC CCCCCC C C CCCCC C CC C
CCCCCCC C C CCC C CCCCCCCC C
CCCCCC
CC C C
CCCC CCCC CC
CCCCCCC CCCCCC
C CCC
CCCC CCCCC C
CC CC C
CC CCC CC
CCCC
CCCC C CCCCC
CC
CC
CC
C
CC CCCC CCC
CC CCCCCC
CC
CCCCCCCCCCCCC
CCC
CCCCCC
CC C
C CCCCC
CC C C
CCCCCC CCCCCCCCCC C
CC CC
C CCCC C
CC
CC C CC CCC CC CCCCC CCC CCCCCCC
CCCC CC CC CCCCC C C CCCCCC
CCCCC
C
CC CCCC CC CCCC
CCC CCC CCCCCC
CCC CCCC C CC C
C CCCCC C
CC
CCCC C CC CCCCC
C CCCCCCCCC
CC
CCC CC C CC CCC C
CCCC
C
C
CC
C CC CCCC
C
C
CCCC
CCCC
CCCCCCC
CC CC
CC C CCCC C
CCC
CC
CCC
CCCCC CC
C
C
CC
C
C
C
C
CC C CCC
CCCCC
CCC
CC
C
C
C C CC CCC CCCCCC
CCCCCCC
CCC CCCC CCCCC CCC
CC CCCCCC
CCCC
C CC C
CC C
CCCC C CC CC C
C CCC C CCCC CC
CC
C CC
CC
CCCC
CC
CC
CC
C
CC
CC
CCC
CC CCCC
CCC
C CCC CCCC C
CCC
C
CC CC
CCC
CC
C CCC
C CC
C
CC
C C
CC
CC C
CCC
C
C
CC
CCC
C
C CC
CCC
CC
CCCC
C
CC
CC
CCC
CCC C
CCCCCC
C CC
CC
C CC
C
CC
C
CC C
CC
C CCC CC C
CC
C
C
CCC
CC
C
CC
C
CCC
C CC
CCCC
C CC C
CCCCC
CCCC CCC CCC
CCC
CC CC CCC C CC
CCC
CC
CCCCCCCCC
C CC
CC
CCC CC
C CC
CC
C
CC
C CCC C C CCCC CC
C
C
CC
CC CC
CCCCCCCC
CC
C CC CCCCC
CC
CC
C
CCCCCCC
C CCC CC
CCCCC
CC
CC
CC
CC
C
C
CC
CC
CCCC
CCC
CCCCCC
CCCCCCC CC
CCCCCCC
C
CCCCC
CCCC CCCCCCCCCCC
C
CCC CCC CCCCCCC CCCCCC CCCCCCC CCCC
C CCC CCCC CCCC
CC CC
C CC CC
CCC
CC
CCCCCCCCCCCCCC
CCCCCCCCCCCCCCCC
CCC
CCCCCCC
CC
CCCCCCCCCCC
C CCCCCCCCCCCCCCC
CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCC CCCCCCCCCC CCCCCCCCCCCC
CCC
C CCCCC
CCC
CC
C CC
C CCCCCCCC
CC
CCCCC CCC C C CCC
CCCCC
C
CC
CC C
CCC CCCCC C
CC
CCC C
CCCCC CCCC C C
C CCCCCCC
CCC CCC CCC CCCC CCCC
CC CC
CCC
CCCC CC CCC CC CCC
CC
CCC
CC
CCCCCC
CCCCC CCC CC CCCCCCC
CCC
CC CCC C C C
CC CC CC CCCC C CCCCCCCCCCCCCCCCCCCCCC
CCCCCCCCCCCCCCC CCCC CCCC CCCCCC
CCC
CCCCCCCCCCCCCCCCCCCCC
CCCCCCCCC
CCCCCCCCCCCCCCCCCCCCCC
CCCCCC
CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCC
CCCCCCCCCC CCCCCCCCCCCCCCCCCCCCCCCCCC CCCCCCC C CC C C CCCCCCCCCC C CCC
CCCC CC C
CC CCC
CC CCCC C
CC CC C
CC
CC CC CCC
CC CCCCCC
CCC
CCCCCC CCCCC C
CCCCC CCCCC
CCC
CCC CCC CCCCCCC
CC C
CCC
CC CC
CC
C
C CC CC CC CC CCCCC C
CCCCCC
C
CC
CC
CCCCC CCC CCCCC CCC CC
CCCC
C
CC
C
CCCCCCCC C
C
C CCCC
CC
C CCCCCC
CCCC
CC CCCCCCCCCCCCCCCCCCC CCCCC C
CCCCCCCCCCC CCC CCCCCCCCCCCC CCCCCC CCCCCCC CCCCC C
CCCCCC
CCCCCC
CCCC
CCCCCC
CCC
CCC
CCCCCC
CCC
CCC C
CCCC
CC
C CCCCCC CCCC CCC CCC CCCCCCCC CCC CCCCCCCCCCCCCCCC
CCCCCCCCCCCCC
CCCCCCCCCCCC CCCCCCCCCCC CC CCCCCCCCC CCCCCCCCCCC CC
CCCC CCC CCC CCCCCCC CCC CCCC CCC
C CC CC CC CC CCCCC C
CC CCC CC CCCCC CCC
CC
C CC
CCCC C
CCC
C
CCCC
CC
CCCCC
CCCCC CC
CCC CCCCC C CCCC CCCCC
CC
CCCC C
CCCCCCCC CCC
CCC CCCCCC CCCC CC
CCCC
CCCCC CC CC CC
CCCCCCCCCCC CCC
C CCCC CC CCCCCC CCC
C CC CCC CCCCCC CC C CCCC CCCC
CCC
CCCCCCCCCCCC
CCCCCCCC C CCC
CCCCCCCCCC CCCCCC
CCCC CCCCCCCC C
CCC CC CCC CCCC CCCCCCCCCCC C CCC
CCCC
C
CC
CCC CC
CCCCCCCCCC CC C
C C CCCC
CCC C C
C
C CC
CC
CCCCCC
CCC CC CCC CCCCCCC C CCCCC CCC
CCCCCCCCCCC C
CCCCCCCC CCCCCCC CC CCCC CCCCC CCCCCC C CCCCCCC CCC CCCCC CCCCCC CC CCCCCCCCC CC
C CC CC CCCC CCCCC
C
CC CC CCC C CCC
CCC CC
CC C
CCC
CCCCCC
CCC CCCCC CCC CCC C
CCC C CC CCCC
CCC
CC
CC CCC CC CCCCCC CCCC CCC CCCCCCCCCCC CC
CCCC C CCC CCCCC CCC
C CC CCCCCCCCCCCCCCCCCC CC
CCCCCC CCCC CCCCCCC CC
CCCC CCCCC CCCC CCCC CC CCCCCC
C CC C C CCCCCC CCCCC CCCCCCC CCC CC
CC CCCCCC
C CCC CCCC
CCC
CCCCCC CCCCCC CC
CCCCCC
CC
CCCC CCCCC
CC CCCCCCC C CC CCCCCCCCCCCCCCCCCC C
CCCCCCCCCCCCCCCC CCC
CCCCCC
CCCCC
CC CCCCCCCC
CCCCCCCCC CCCC CC CC CCC CCCC CCC CCCC
CCCCCCCCCCCCCC CC CCCCCCCCC
CCCCCCCCCCCC CCC CC CCCCC CCCCCC CC CCC CCC
CC CCCCCC CCCCCC CCCCCCCC CC CCCC CCCC CCC CCC C C
C CC CCCC
CC CCCCCCCC CC CC CC C
CCCC
CC CCC CCC CCCCC CCCCCC
C CC CCC CCCCCC
C CCCC C CC CC CCCCC C CCC
CCCCCC
CCCCC CCCCC
CCCC
C CC
CCC CCC
CC
CCC
CC CCCC CCCC CCCC C
CCC CC C C
C
CCC
CCC CCC CCCCCCCCCC C CCCC C CCCC CCCC CC
C CC CCCCCC CC
CCCCCCCC CCC
CC CCCCCCC CCC
CC C CC C CCC
CCCC CC
CCCCCCC
CCC CC
CCC CC CC
CCC
CCCCC
CCCC
CCC
CCCCC
C CCC CC CC CCCCCCCCCCCCCCCCCCCCC CC CC CCCCC
CCCC CCCCC
CCCCCCCCCCCC
CC
CCCCCCC CCC
CCCCCC C CCC CCCCCC C
CCCCCCCCCC
CCCC
CCC
C CCCC CCC
CCCC
CCC
CCCCC CCC
CC
CCC CC
CC
C
C CCC
CC
CC
CCC
CC
CC
C
CCCCCC CC
CCC
C
CCCCCC CC
CC
CC
CC CC CCC CCCCCC C
C CC
CC
CCC
C
CCC CC
C CC
CCCC CCCC
CCC CCC
CCC C
CCC C C
CC CCCCCC C CCCCCC CC CC
CC
CC
CCCCC
CCC
CCCC CC
CCC CCCCCCCCCC CC CC
CC CCCC CCCC C
C CCC C
CC CCCC C C
CC CCC CCCC CCC
CC
CC CCC CCCCC CCCCCCCC
CC C CC CCCC CCC CCCCCCCC CC CC CC CCC CC CCC
CCC
C
C
CC CCCCC C C CCCCC
CCC
CCCC
CCCCC CCCCCC C
CCCCCCCCCC CC CCCCC CCC CC C
CC
CCCCCCC
CCC CCCCCC CCCCC CCC CCCCC C CC C CCC
CCCCC CCCCCC
C CC CCC
CC
CCCCC C C CCCC
CCC
CCCCC CC C
CC
C C CC
CCCCCC CCCC CCCCC C CCC C CCCCC C CCCC CCCCCCCCC CC CCCCCCCCCC CCCCCC
CCCC CCCCCC CC CC CCCCCCC CCCCCC CCCC CCC C
C CCCCC CCCCCCCCCC CCCCCC
C CCC CCC CCC
CC CC C CC
CCC
C
CC C CC CC C CCC
CC C C C CCC CCCCCCCC CCC CC CCC CCC CCCCCC CC CCCCCCCCCCC
C CCC CCCC
CCC
CC CCC
CC CCC
CC CC C C C
C CCC
CCC CC C
CCCC CCCCC CC CCCCCCC C
CCC CC CCCCC CCCCC CCCCCCC
CCC CCCCCC C
CCCCCCC
C CCC
CCC CC
CCC
CC CC
CCCCCCCCCCCC
CCCC CCCCC
CC CC CCCC CCCCCC CCCC CC
CCC
C
C
C
CC CCCC CCCCCC CCC CCCC
CCCCCCCC CC CCCC CCC
CC
CCCCCCCCCC
CCC CC
CC CC
C C
CC CC CCCCCC CC
CC
CCC
CCCCC
CC CCC C CC C
CCC
C
C CC CC CC CC
CC CC CCCCC
CCCC CC
CCCCC CC CCCC CC
C CC CCCCC CCCCCCC CC CC
CC CC CCCC CC
CCCCCC CC
C CC
CCC CCCC CCC CCC
CCC
C CC CCC
CC CCCC
CCC CCCC
CC
CC CC
CCCCC
CC CCCC
CCCC
CC
CC CC C
CC
C
C
CCC
C
CC CC
CCCC CCC
CCC
CCC CC
CCCCC
CC
C
CC
CCCC CCC
CCCCCC
CC
CCCC
CCCCCC
CCCCCC CCCC
CCCC CCCCC C
CCC CCCCC
CC
CC
CCC CCCC
CCCC
CC
CCCC CCC
CCCC CC
CC CC CCC
CCC
CCCCCCCC
C C
CC
CC
C
CCCC CCCC
CC
CC
CCC C
C
C CCCC
CCCC
C
C
C
CC
CCC
C
CC
CCCC C
CCC CCC
CCC C
C CCC
CCCCC
CCC
CCCCCC CC
CCC
C
C
CC
CC
CCC
CC
CCCCCCCCC
C
C
CC
CCCCCCC C
C CCC
CCCC
C CC CCC C
CCC
C CC CCCC
CC
CCC
CCCC
CCC
CC CCC CC
C
C
CC
C
CC
CCCC
C
CC
C
CCCCCC
C CC
CCC CC
CCC
C
CCC
CC
CC CCC
CCCCC
CCCCCC CCCC CCC
CC CCCCCCCC CCCCCCC
CCCCCCCC CCCC
CCCC
CC CC
CCC CC CC CCC CCC C CCCC CCC
CC CCCCCCC
CC CCCCCC
CCCCC
CC
CCC
C
CCCC CC
CC
CC
C
C
CC
CC
CC
CCC CCCCC
C CC
CCC
C
CCC
CCC
C
CCCCCCCCCCCCCC CCCCCCCCCCCCCCCCCCCCCCC CCCCC CCCCC CCCCCCCCCCCCCC C
CCCCCCC CCCC CCCCCC CC CCCCCCCCCCCCCCCCCCCCC CCCCC
CCCCC CCC CCC
CCCCCCCCCCCC CCC
C CC CCCC
CCCCCCC CC CCCC
CCCC CCCC CCCCCC
CCCCCCCC CCCCCCCC
CCC
CCCCCCCC CCC
CC CCC CCCC CCCC CCC C CCCCCCCCCCC
CCCCCCCCCCCCCCC CCCCCC C
CC CCCCCCC CCCCCCCCCCC
CCC CC
CC CCCCCCCC CCC CCC CC CCC CCCC C CC CC CC CCCC CCCCCC CCCCCC CC CCCCCCCCCC CCC C
CC
CCCCCC CCC C C
C C CC CCCC
CCCCCCC CC CCCCCCCCCCC CCCCC CCCCCCC CCCCCCCCCCCCCCC CCCCCCCCCCCCCC CC CC CCCCCCCCCC CCCCCC CCC
CC C CCC CCCCCC
CCC CC
C CCC CCCC C CCCCCCCC CC CCCCCCCCCCCCCCCCC CCCCCCCCC C
CCC
CCC
C CCC CCCCCCC C
CCCCCC
CCCCCC
C
CC
C
CC
CC
CCC CCCCC CCCC
C
CCC CCCC C C
CC CCCCCCCC
CCC
CC CCCC
CCCC CCC CCCC
CCCCCCC
C CC CCCC
CC
CC C
C CCCCCCC CCCC CC
C
CCCCCC
C CCCCCCCCCC CC CC CCCC CC CCCCCCC CCC C
CC CCC CCCCCC C CCC
CCCCCCC CC CCCCC
CCCCC
CCC
CCCCCC CCCCC
CCC CCCC C CCC C CCC CCC
CCCCCCC
C
CC
C
C
C
CCCCC
CCC
C
CCCCC CC CC CCCCC
C
CC
CC
C
CCC
C
CCCC
CC CCC
C
C
C
CCCCC
C
C
C
CC
CC
C CC
CCCC CC CC
CCC CC
CCCC CC
C
CCCCC
CCC
CCCCC
CCC
CC C
C
CC
CC
C
C
CC
CCC
CC
C
CC
CCCC CCC
CCCC CC
C
C
CCC
CC
CC
C
C
C
C
CCC
CC
CC
C
CC
CC
C
CCCC
CCC CC
CC
CC
CCC C
C
C
CC CC C
CC
CC
C
C C
CCCC
C
CCC
CCC
CCCCCCCC CCCC CCCC
CC CC C C
CC CC CCC CCC C C
CC C C
CC
CC CC C
CCC CC CCCCCC C CC
CCCC C
C
CC
CCCC
CCC
CCC
C
CC
CCC CCCCC
C
C
CC
CC
C
CC
CCCC
CCCCC CC
C CCCCCC
C
C
CC
C CCCC
C C
CCC
CC
C
C
C
CCC
C
C
C CCC
C
C
C CCC
CC
C
CCCCC
C
CC C
C
C
CCCCC C
C
CCCC C
C
C
CC
C
C
CCCCCC CCCCC C CCCC CC
CC CCCCCC CCC CCCC C
CC
C
C
C
C
CC
CC
CCC
CCC
CCCC CCCC
C
C CCC CC
CC
CCCC
C CC
CCCCCC
CC
CCCC C
CCCC C
CCC
C
CC
CC
C C
CC
CCCC
CCCCCCC CC C C
C
C
C
C
C
C
C
CCCCC
CCCC CCCC CC
CCC
CCCC
C CC C
C
CC
CC
CC C
C
C
CC CCC
C C
CC
C CCC C C CCC
CC
C CC CC
C CCC C
C
CCC
C C C
CCCCC
C
C CCCCC
CC C
CC
CC CCC CC
CCCCCC C
CCCCCC CCC C
CC
CCC
CCCCCCCC C
C CCCCC
CC CCC CCC C
C CCCCCCCC
CC
CCCCCCC CCC CCC
C CCC CC CCCC
CCC CCC
CCC
C
CC
CCC
CC
CCCC
C CC CCC C
C CC CC
CCCC C
C CCC CCC
CC
CC
CC C
C
C
CC
CCC C
CCC CC C CC
CC
C CC
C
CC C CCC C
CCC
CC
CC
CC
CCCCC CCCCCC CC C
CCCCC
CCC CC C C
CCCCCC C
CC
CCC
C CC
C CCC CC
CC
CCC
CCCC
CCCC
CCC C CCCC CC
C
C CC CCCC CC CCCC
CCCC
C CCC
C CCCCCC CCC CCC C
CC
CC CCC
CCC
CCCC CC
CCCCC
CCCCC CC CCCC CC CCC C
CC
CC C CC CCCCCCC
CCCC
C
CC CCC
CC
CCC CCC
CCCC C
C CC
CCCC CC C CC
CC
C
C CC CC
C
C
CCC
CC
C
CCC
CCCCC
C
CCCCCC
C
CCCCC
CC
CCCC
CC
C CCCCCCCC
CC
CC
CC CCCC
C C CCC CCCC CC C CC CC CCCCCCCC CCC CCCCC
C CCC
CC
C
CC
CCC
CCC
CCC CC CCCC
CCC CCC
CCC CCCCCC
CCCCCC
CC
C
CCC
CCC C
CC
CCCCCCCC C
CC C CCCC CCC C CC
CCC CCCCCCCCCCC CCCCCC CC CCCCCC C
CCC C
CCCCCC C C CC
CC CCCCCCCCC CC
CCCCC
CCC C
CCCCC
C CCCC C
C
C
C
CC CCCC
C CC CCC CCC
CCCC C
CCC
CC
C
C CCCC
CCC
CCCCCCC
CC CCCC CCC
CCCC CC CC
CCC
CC CCCCC CC CCC CC CCCC
C C CCCCC CC CCC C
C C CCC C CC CCC CC
CC CC CCC C CCCCC
©2016 NRC Canada
Page 30 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Page 31 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 31
Table 1. Number of observations by sitequality and species dominating the stand.The + indicates categories aggregatedwith the next closest category.
Spruce Pine Deciduous
SQ1 + 1183 +SQ2 712 927 433SQ3 1697 540 1061SQ4 915 110 928SQ5 172 + 265Total 3496 2760 2687
©2016 NRC Canada
Page 32 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
32 Can. J. For. Res. Vol. 99, 2016
Table 2. Definition of the components for each species group. [SQ] indicates when the parametersvary with the site quality parameter.
spruce dominatedP OT π0[SQ]
MODT (1 + exp (−τ1 [SQ] · T SUM + τ0 [SQ]))−1
MODM (1 + exp (−µ1 · MSJ + µ0))−1
pine dominatedP OT π0[SQ]
MODT (1 + exp (−τ1 · T SUM + τ0 [SQ]))−1
MODM ξ−1· µ1/α · ((MSJ − µ0) /α)µ1−1 exp (− ((MSJ − µ0) /α)µ1 )
deciduousP OT π0[SQ]
MODT (1 + exp (−τ1 · T SUM + τ0))−1
MODM ξ−1· µ1/α · ((MSJ − µ0) /α)µ1−1 exp (− ((MSJ − µ0) /α)µ1 )
©2016 NRC Canada
Page 33 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Antón-Fernández, Mola-Yudego, Dalsgaard, Astrup 33
Table 3. Parameter estimates and measures of goodness of fit for the twocandidate models for the three species.
spruce pine pine deciduous deciduousParameters md1 md1 md2 md1 md2
π0[SQ1] 9.0797 9.469π0[SQ2] 15.4929 12.7145 13.3917 14.4688 13.041π0[SQ3] 21.7727 13.3343 14.7684 20.1651 18.0119π0[SQ4] 22.9493 16.7999 17.8809 25.3659 22.5634π0[SQ5] 26.1775 29.5949 25.9418τ1 0.1576 0.2068 0.1126 0.1125τ0 3.1924 2.7009τ0[SQ1] 1.8948 2.4028τ0[SQ2] 2.3125 2.9211τ0[SQ3] 1.7933 2.4689τ0[SQ4] 2.5339 3.0053ξ 0.0076 0.0060µ1 1.5042 1.8939µ0 -103.5199 -115.2986α 98.0557 137.4867AIC 17427 13121 12844 12742 12539BIC 17508 13180 12921 12784 12598R2
adj 0.56 0.33 0.40 0.50 0.54RMSE 2.91 2.60 2.47 2.58 2.49
©2016 NRC Canada
Page 34 of 34
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research