chromosomes. 2 figure 28.1 genetic material organization: condensation nucleic acid: negative...
TRANSCRIPT

Chromosomes

2
Figure 28.1
Genetic material organization: condensationNucleic acid: negative chargedProteins: positive chargedNucleoproteins:DNA(RNA)-protein complex
Bacteria: nucleoid
Eukaryotic: 1-chromatin2-chromosomes
Packing ratio:DNA(RNA)-proteins
Packing ratio = the length of the DNA divided by the length of the unit that contains it.
Nucleic Acid Molecules are longer than their compartments

3
Viral Genomes Are Packaged into Their Coats
• The length of DNA/RNA that can be incorporated into a virus is limited by the structure of the head-shell.
Figure 28.2
How specific is the packaging of the viral genome?
Capsid (structural proteins and functional viral proteins)Nucleic acid (RNA)
Protein shell is formed by a two-layer disk and each layer contains 17 identical proteins. RNA interacts with each layer ofProteins.
DNA(RNA) /protein InteractionPrimary and tertiarystructures

4
• Nucleic acid within the head-shell is extremely condensed.
• Filamentous RNA viruses condense the RNA genome as they assemble the head-shell around it.
• Spherical DNA viruses insert the DNA into a preassembled protein shell.
Figure 28.3

5
Head-shell (protein) and DNA.
Insertion of DNAinto the phage head.Translocation (ATP-dependent process)Condensation(mechanism: protein-scaffolding- ?)
Viruses use protein coats for packingTranslocation (phage lambda)
1-step:cleavage at the cos sequence
2-step: Binding of terminase (2 subunits)
3-step: recruitment to the head
4-step: translocation
5-step: condensation
CohesiveComplementaryends
Cos sequences are ~200 base pairs long and essential for packagingCos N site: cleavage siteCos B site: It holds the terminaseCos Q site: It protects from degradation (cellular DNAses)

6
The Bacterial Genome Is a Nucleoid
• The bacterial nucleoid is ∼80% DNA by mass.– It can be unfolded by
agents that act on RNA or protein.
• The proteins that are responsible for condensing the DNA have not been identified.
• HU and H1 proteins
Figure 28.5
heat-unstable nucleoid proteinHistone-like protein H1 (H-NS)
High mobility group: Non-specific interaction

7
The Bacterial Genome Is Supercoiled
Figure 28.7
Loops
Organization of the genome into loops is associated with supercoiling of the DNA in vivo. Nicking the DNA relaxes the supercoils.
•The nucleoid has
∼100 independent
negatively supercoiled
domains.
•The average density
of supercoiling is
∼1 supercoil/100 bp.
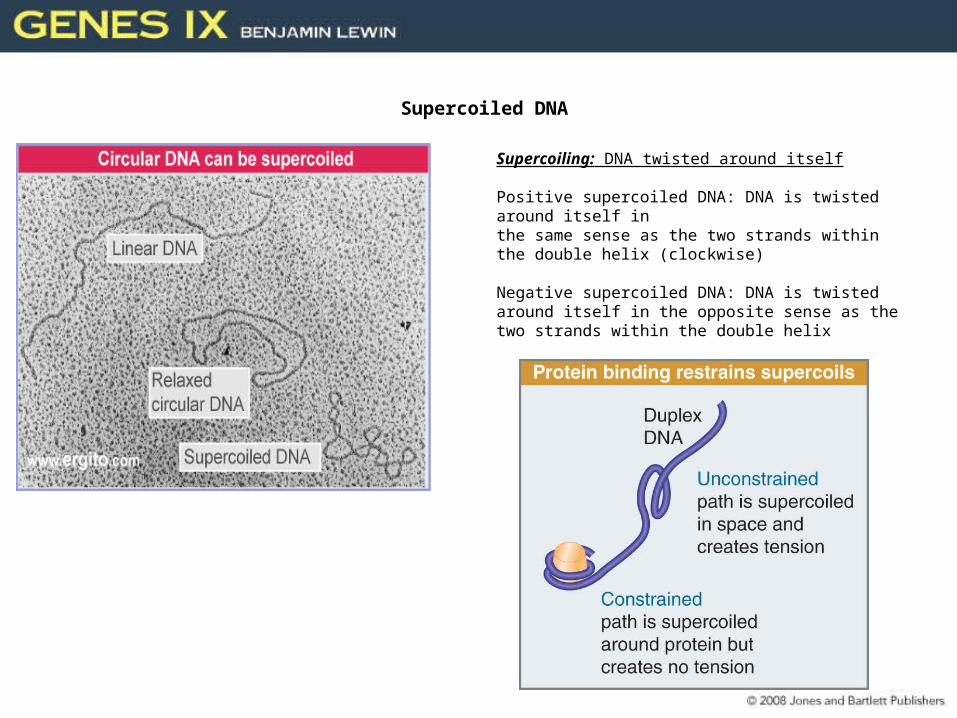
Supercoiled DNA
Supercoiling: DNA twisted around itself
Positive supercoiled DNA: DNA is twisted around itself in the same sense as the two strands within the double helix (clockwise)
Negative supercoiled DNA: DNA is twisted around itself in the opposite sense as the two strands within the double helix

9
Eukaryotic DNA Has Loops and Domains Attached to a Scaffold
•DNA of interphase chromatin is negatively supercoiled into independent domains of ∼85 kb.•Metaphase chromosomes have a protein scaffold to which the loops of supercoiled DNA are attached.
Eukaryotic interphase DNA is organized similarly to prokaryotic DNA.
DNA loops are anchored in a protein scaffold that does not require DNA for formation.
DNA
ProteinScaffold

10
Specific Sequences Attach DNA to an Interphase Matrix
Figure 28.10
MAR sequences may have role in regulating supercoiling of loops.
S/MAR: it is mix.
DNA is attached to the nuclear matrix at specific sequences called MARs (matrix associated regions) or SARs (scaffold attachment regions).
The MARs are AT rich (>70%) but do not have any specific consensus sequence.

11
Chromatin Is Divided into Euchromatin and Heterochromatin
Chromatin is divided into euchromatin and heterochromatin
Chromosomes are compacted at Mitosis.Heterochromatin is moredensely packed thaneuchromatin
Individual chromosomes can be seen only during mitosis
During interphase, the general mass of chromatin is in the form
of euchromatin, which is less tightly packed than mitotic chromosomes.
Regions of heterochromatin remain densely packed throughout interphase.
Heterochromatin :-permanently condensed-consists in DNA sequence repeats (not transcribed) -reduced density of genes (inactivated)-replicates at late states of the S phase.-interacts with Histones

12
Chromosomes Have Banding Patterns• Certain staining techniques cause the chromosomes
to have the appearance of a series of striations.– They are called G-bands.
Figure 28.13
Giemsa staining allows visualization of chromosome binding patterns.
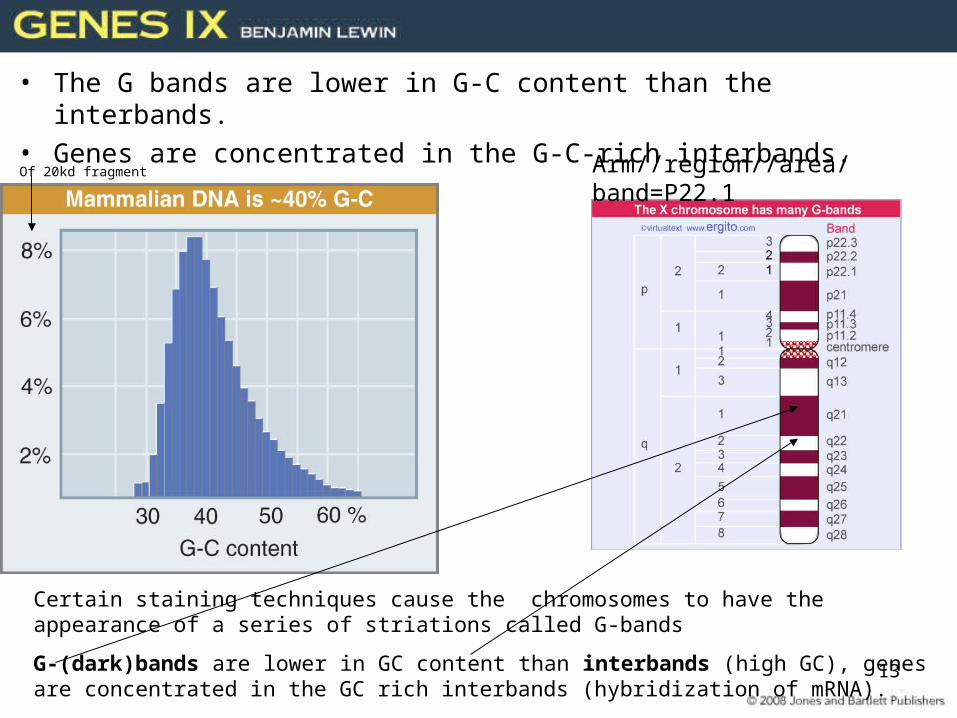
13
• The G bands are lower in G-C content than the interbands.• Genes are concentrated in the G-C-rich interbands.
Certain staining techniques cause the chromosomes to have the appearance of a series of striations called G-bands
G-(dark)bands are lower in GC content than interbands (high GC), genes are concentrated in the GC rich interbands (hybridization of mRNA).
Arm//region//area/band=P22.1Of 20kd fragment

14
DARK BANDS (G BANDS) PALE BANDS (CORRESPOND TO I BANDS)Stain strongly with dyes that bind preferentially to Stain weakly…..
AT-rich regions such as Giemsa and Quinicrine May be comparatively AT-rich Maybe comparatively GC-rich
DNase insensitive DNase sensitive
Condense early during the cell cycle but replicate late Condense late during cell cycle but replicate early
Gene poor. Genes may be large because exons are often Gene rich, Genes are comparatively small because of close
separated by very large introns clustering of exons
LINE rich but may be poor in Alu repeats LINE poor but may be enriched in Alu repeats

15
Special conditions for visualizing chromosomes and gene expression
Visualization of condensed chromosomes that express genes during an extended meiotic period in some amphibians (lampbrush chromosomes).
Visualization of amplified chromosomes (polytene chromosomes) in the larvae of Dipteran flies (Drosophila).

16
Lampbrush Chromosomes Are Extended
• Sites of gene expression on lampbrush chromosomes show loops that are extended from
the chromosomal axis.
Figure 28.16
When are chiasmata formed? DNA recombination
Why would the presence of ribonucleoprotein suggests gene expression? Nascent RNA chains

17
Polytene Chromosomes Form Bands
• Polytene chromosomes of dipterans have a series of bands that can be used as a cytological map.
Figure 28.18
(Drosophila)Dna probe(Labeled cDNAderived from a mRNA)
Are these genes transcribed?

18
Polytene Chromosomes Expand at Sites of Gene Expression
• Bands that are sites of gene expression on polytene chromosomes expand to give “puffs.”
Figure 28.21
Puff material site of transcription?
Characteristics of the puff materialDNA is in more relax conformationSynthesis of RNATranscriptionProteins required for transcription(RNA polymerase II and others)

19
The Eukaryotic Chromosome Is a Segregation Device
• A eukaryotic chromosome is held on the mitotic spindle by the attachment of microtubules to the kinetochore that forms in its centromeric region.
• Centromeres often have heterochromatin that is rich in satellite DNA sequences ( highly repetitive DNA sequence).
Figure 28.22

Centromeres are required for the correct segregation of chromosomes at mitosis and meiosis
Centromere or centromeric region: DNA: Proteins:Cohesins proteins (glue)Microtubules (MTOCs)Other proteins ?
Centromeric DNA:Is formed by the heterochromatin DNA ( very common but not a 100 % of co-localization)C-bands (specific staining- DAPI or 4',6-diamidino-2-phenylindole, 33258 hoechst )
Conserved DNA sequence(responsible for DNA-protein [microtubules] interaction)
Kinetochore- DNA and microtubules (part of the MTOC)
Centromere DNA sequences confer mitotic stability on pieces of dsDNA

21
Centromeres May Contain Repetitive DNA
• Centromeres in higher eukaryotic chromosomes contain large amounts of repetitive DNA.
• The function of the repetitive DNA is not known.
Saccharomyces cerevisiae ~120 bp
Schizosaccharomyces pombe ~40 to 100 kb
Drosophila ~200 to 600 kb
Arabidopsis >500 kb

22
Centromeres Have Short DNA Sequences in S. cerevisiae
• Centromeric DNA regions (CEN) elements are identified in S. cerevisiae by the ability to allow a plasmid to segregate accurately at mitosis.
• CEN elements consist of the short conserved sequences CDE-I and CDE-III.– They flank the A-T-rich region CDE-II.
Figure 28.25
CDE: cycle-dependent element

23
The Centromere Binds a Protein Complex
• A specialized protein complex that is an alternative to the usual chromatin structure is formed at CDE-II.
• The CBF3 protein complex that binds to CDE-III is essential for centromeric function.
• The proteins that connect these two complexes may provide the connection to microtubules.
Figure 28.26
Mutations in CDE affect normal Segregation of DNA

24
Telomeres Have Simple Repeating Sequences
• The telomere is required for the stability of the chromosome end.
• A telomere consists of a simple repeat where a C+A
-rich strand has the sequence C>1(A/T)1–4.
Figure 28.27

25
Telomeres Seal the Chromosome Ends
• The protein TRF2 catalyzes a reaction in which the 3′ repeating unit of the G+T-rich strand forms a D-T loop (5 to 10 kb).
– It displaces its homolog in an upstream region of the telomere.
Figure 28.30

26
Telomeres Are Synthesized by a Ribonucleoprotein Enzyme
• Telomerase (RT) uses the 3′–OH of the G+T telomeric strand to prime synthesis of tandem TTGGGG repeats.
• RNA as template
Figure 28.31


28
28.19 Telomeres Are Essential for Survival
Figure 28.32

29
Why are telomeres important?
The telomere can reach a length of 15,000 base pairs.
Telomeres function by preventing chromosomes from losing base pair sequences at their ends.
Telomeres also stop chromosomes from fusing to each other. However, each time a cell divides, some of the telomere is lost (usually 25-200 base pairs per division).
When the telomere becomes too short, the chromosome reaches a "critical length”==>apoptosis. • Telomeres allow cells to distinguish chromosome ends from broken DNA
◦ If DNA is broken there are two options after the cell cycle is stopped: Repair or Death▪ Repair can occur in two ways:
▪ Homologous Recombination (HR) -- ▪ Non-homologous end-joining (NHEJ) --
• Telomeres prevent chromosome fusions by NHEJ◦ Fusion-bridge-breakage cycles leads to genomic instability which in turn can
result in cell death or neoplastic transformation• Telomeres are specialized structures that are essential for protecting chromosome ends and
ensuring chromosome stability

Nucleosomes

31
The Nucleosome Is the Subunit of All Chromatin
• Micrococcal nuclease releases individual nucleosomes from chromatin as 11S particles.
Figure 29.2Endonucleases

32
• A nucleosome contains “Basic structure “
– ∼200 bp of DNA– two copies of each core histone – (H2A, H2B, H3, and H4)
Figure 29.3
Nucleosomes are the fundamental subunits of chromatin in eukaryotes.
The nucleosome is ~200 bp of DNA wrapped on the surface of a histone octamer (two H2AH2B dimers and one H3H4 tetramer). H1 is not required for the correct formation of the nucleosome, but is part of it.
Nucleosome organization:
Nucleosomes are the first level of organization (10 nm).
Nucleosome fiber ( coiling of the series of nucleosomes) is the second level of organization (30 nm).
The third level of organization is the packaging of the Nucleosome fiber itself.

33
• DNA is wrapped around the outside surface of the protein octamer.
Figure 29.5
~80 bp of DNA makes two turn around the histones core (H1 is no included)

34
DNA Is Coiled in Arrays of Nucleosomes
• >95% of the DNA is recovered in nucleosomes or multimers when micrococcal nuclease cleaves DNA of chromatin.
• The length of DNA per nucleosome varies for individual tissues in a range from 154 to 260 bp.
Figure 29.7/8
Chromatin
(Nucleosomes)n
*ExperimentalConditions“Micrococcal”
Buffer ~pH:8.0, 5mM CaCl2, 100mM NaCl.
Nuclease*

35
Nucleosomes Have a Common Structure
• Nucleosomal DNA is divided into the core DNA and linker DNA.– It depends on
the DNA’s susceptibility to micrococcal nuclease.
• The core DNA is the length of 146 bp that is found on the core particles produced by prolonged digestion with micrococcal nuclease.
Figure 29.9
Nucleosomes (~205bp)
Nucleosomes (~146bp)
Nuclease*
*Experimentalconditions

36
• Linker DNA~19 bp (constant ?) …..Linker DNA is the region of 8 to 114 bp that is susceptible to early cleavage by the enzyme.
• Changes in the length of linker DNA account for the variation in total length of nucleosomal DNA.
• H1 is associated with linker DNA and may lie at the point where DNA enters and leaves the nucleosome. Core DNA ~146 bp (no Histone 1)
Partially digested chromatin can be separated into mono, di, tri nucleosome fractions . Fully digested chromatin is in the mononucleosome and core particle form.
?

DNAase I and II make single-strand nicks (randomly).
DNA structure, periodicity and nucleosomes
Nuclear DNADNA-ase IRegular cutting ~10-11 bp
What is the interpretation of this regular cutting?
DNA is free in solution-no histones nuclear DNA
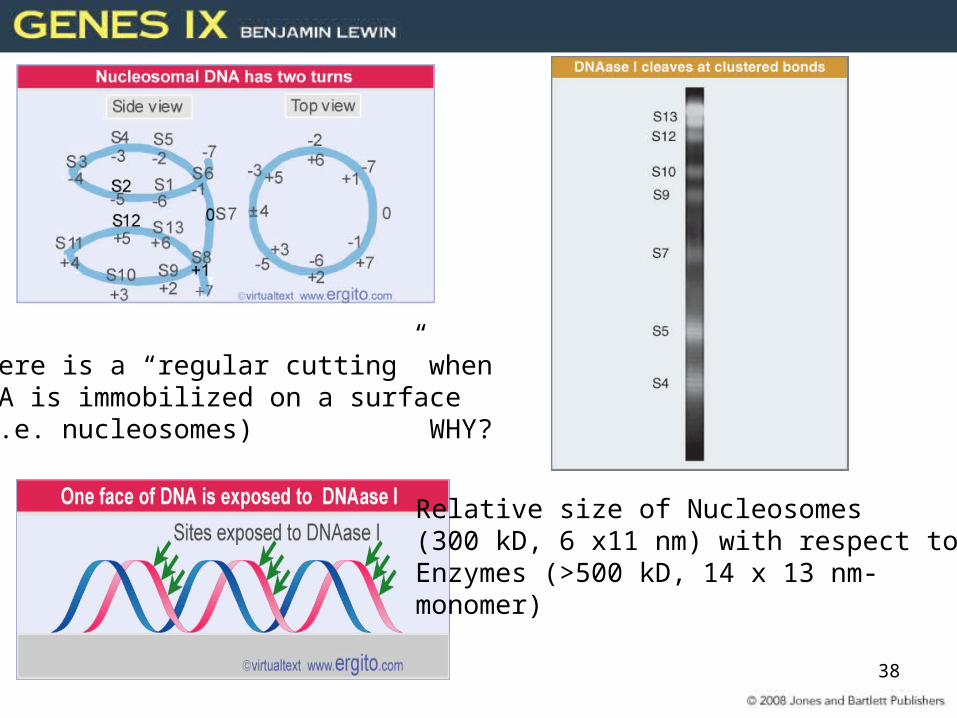
38
There is a “regular cutting” when DNA is immobilized on a surface (i.e. nucleosomes) WHY?
Relative size of Nucleosomes (300 kD, 6 x11 nm) with respect to Enzymes (>500 kD, 14 x 13 nm-monomer)

Supercoiling and the periodicity of DNA
SV40 (Simian Virus) has a small DNA genome that is useful for the study of nucleosomes.
The SV40 minichromosome is supercoiled and can be relaxed by altering the salt concentration (Topoisomerase-I).
Removal of the proteins (histones and nonhistones) adds supercoiling to the DNA.
Restoration of the histones reduces the supercoiling of the DNA.
The Periodicity of DNA Changes on the Nucleosome

40
The Nucleosome Is the Subunit of All Chromatin
• Micrococcal nuclease releases individual nucleosomes from chromatin as 11S particles.
Figure 29.2Endonucleases
IntermediateStructures?
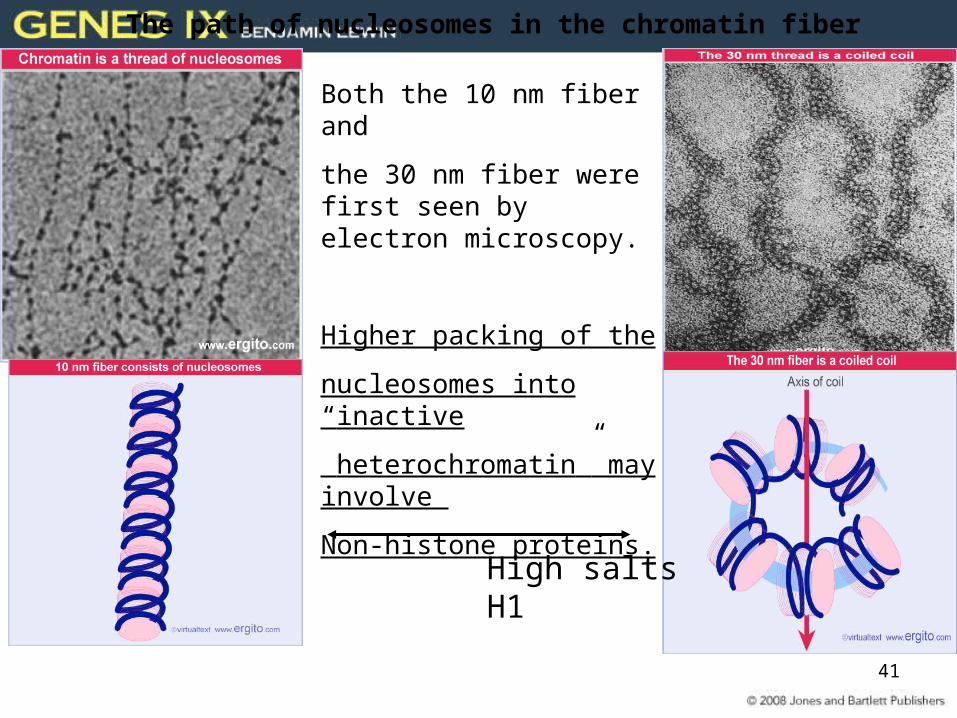
41
Both the 10 nm fiber and
the 30 nm fiber were first seen by electron microscopy.
Higher packing of the
nucleosomes into “inactive
heterochromatin” may involve
Non-histone proteins.
The path of nucleosomes in the chromatin fiber
High saltsH1

42
The Path of Nucleosomes in the Chromatin Fiber
• 10 nm chromatin fibers:– are unfolded from 30
nm fibers – consist of a string of
nucleosomes
Figure 29.23

43
• 30 nm fibers have six nucleosomes/turn.– They are organized
into a solenoid.
• Histone H1 is required for formation of the 30 nm fiber.
Figure 29.25
Which one of these two nucleo. forms are found in the nucleus?

44
Organization of the Histone Octamer
• The histone octamer has a kernel of an H32-H42 tetramer associated with two H2A-H2B dimers.
Figure 29.17

45
• Each histone is extensively interdigitated with its partner.
• All core histones have the structural motif of the histone fold (alpha helices). – N-terminal tails extend out
of the nucleosome.
Figure 29.20
Crystal structure of the histone core and DNA has been resolved

46
What is the relevance of this orientation?
Regulation of the histone protein function Acetylation, Methylation and Phosphorylation-Transient reaction ( i.e. on and off).

47
Reproduction of chromatin requires assembly of nucleosomes
Replication of DNA-Separation of Strands of DNA-Large protein complex binding(DNA Polymerase)
Nucleosomes --disruption--formation

48
Labeling (proteins) experiments suggest that nucleosomes are reformed from pre-formed subunits.
Tetramer (H4-H3) and dimer ( H2AH2B)
CAF-1: chromatin assembly factor
Why ?…..we have these two models
1-Do the histones “preform”(holohistone) a protein octamer around which the DNA is subsequently wrapped?
2-Does the histone octamer assemble on DNA from free histones?
3-Are other proteins required/involved during DNA assembly?

49
Reproduction of Chromatin Requires Assembly of Nucleosomes
• Histone octamers are not conserved during replication.– H2A-H2B
dimers and H32-H42 tetramers are conserved.
Figure 29.28
1H and 2H (D) aminoacids

50
• There are different pathways for the assembly of nucleosomes:– during replication– independently of replication (DNA damage)
• Accessory proteins are required to assist the assembly of nucleosomes.
• Chromatin assembly factor-1( CAF-1)

51
• CAF-1 is an assembly protein that is:– required for deposition
of H32-H42 tetramers following replication
• A different assembly protein and a variant of histone H3 may be used for replication-independent assembly (DNA damage).
PCNA: proliferating cell nuclear factor (sliding clamp)

52
Nucleosome Displacement and Reassembly Require Special Factors
• Ancillary factors are required both:– for RNA polymerase to displace
octamers (FACT) during transcription
– for the histones to reassemble into nucleosomes after transcription (CAT)
Figure 29.41
FCAT: Facilitates Chromatin transcription
CAT

53
Do Nucleosomes Lie at Specific Positions?
• Nucleosomes may form at specific positions as the result either of:– the local structure of DNA – proteins that interact with
specific sequences
• The most common cause of nucleosome positioning is when proteins binding to DNA establish a boundary.
Nucleosome Positioning
Rest. EnzMicrococcal nuclease

Nucleosome positioning
Intrinsic positioning depends on the physical properties of certain DNA sequences (like rotational positioning) because the DNA has a particular bend to it --AT vs GC regions).
Extrinsic positioning depends on border regions with no nucleosomes that start the positioning (like translational positioning). (i.e. length of the DNA linker)
(i.e. the position of the histone octamers on DNA depend on the sequence of the DNA)
Why is the nucleosome positioning important?
Micrococcal Nuclease(?)
Access to DNA.
is not random
Why?

Me
Nucleosome depleted regions (NDR)
preinitiation complex (PIC)
56
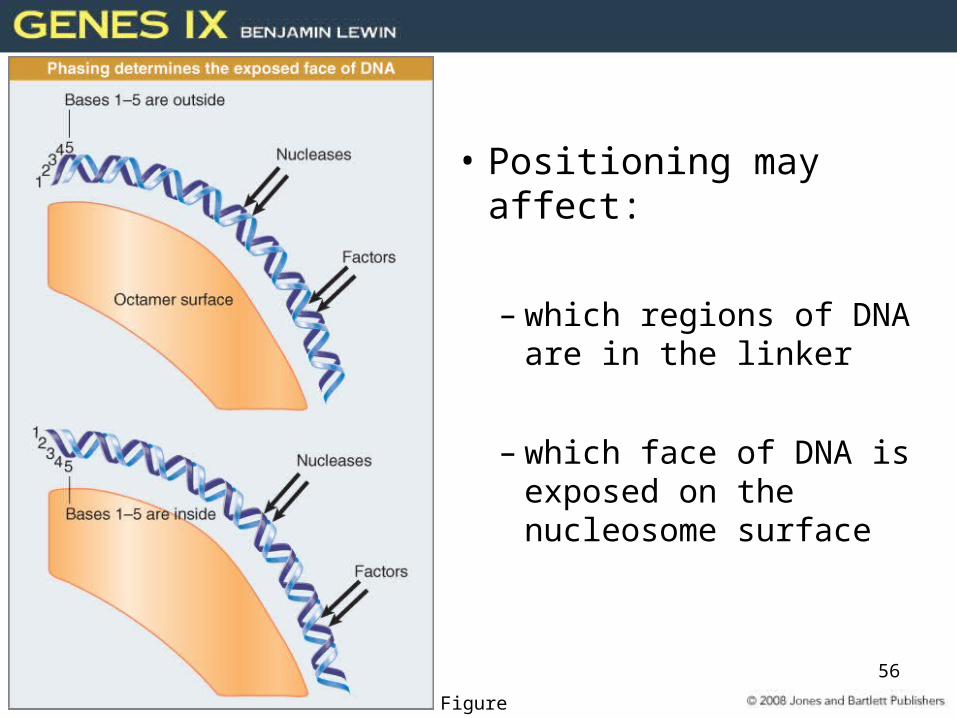
• Positioning may affect:
– which regions of DNA are in the linker
– which face of DNA is exposed on the nucleosome surface
Figure 29.34

57
Histone Octamers Are Displaced by Transcription
• RNA polymerase displaces histone octamers during transcription in a model system.– Octamers re-associate with
DNA as soon as the polymerase has passed.
• Nucleosomes are reorganized when transcription passes through a gene.
Figure 29.38
Size matter?RNA polymerase vs. nucleosome

RNA polymerase disturbs the extrinsic positioning of nucleosomes during transcription.
……..BUT, disturbed positioning can be readjusted when transcription is repressed.
RNA polymerase displaces DNA from histones
nuclease
- + -
?

59
DNAase Hypersensitive Sites Reflect Changes in Chromatin Structure
Figure 29.51
Nucleosome gaps (border regions in extrinsic positioning) allow polymerases, transcription factors and repressors to bind to the promoter region of genes.
Transcribed genes are much more sensitive to DNAase than other parts of the chromosome.

60
Domains Define Regions That Contain Active Genes
• A domain containing a transcribed gene is defined by increased sensitivity to degradation by DNAase I.
Figure 29.52
Chromosomal domain: is a region of altered structure of DNA( no-nucleosomal structure) including at least one active transcriptional units.