chlorophyte dunaliella tertiolecta' - plant physiology
TRANSCRIPT

Plant Physiol. (1991) 97, 147-1530032-0889/91/97/01 47/07/$01 .00/0
Received for publication December 10, 1990Accepted May 7, 1991
Light Intensity-induced Changes in cab mRNA and LightHarvesting Complex 11 Apoprotein Levels in the Unicellular
Chlorophyte Dunaliella tertiolecta'
Julie LaRoche*, Anne Mortain-Bertrand2, and Paul G. Falkowski
Oceanographic and Atmospheric Sciences Division, Brookhaven National Laboratory, Upton, New York 11973
ABSTRACT
During a transition from high growth irradiance (700 micromolesquanta per square meter per second) to low growth irradiance(70 micromoles quanta per square meter per second), the unicel-lular marine chlorophyte Dunaliella tertiolecta Butcher increasesthe cellular pool size of the light-harvesting complex of photosys-tem 11 (LHC II). We showed that the increase in LHC II apoproteinsand in chlorophyll content per cell is preceded by an approxi-mately fourfold increase in cab mRNA. The increase in cab mRNAis detectable within 1.5 hours following a shift from high to lowlight intensity. An increase in the relative abundance of cab mRNAwas also found following a shift from high light to darkness andfrom high light to low light in the presence of gabaculine, achlorophyll synthesis inhibitor. However, the LHC II apoproteinsdid not accumulate in the latter experiments, suggesting that LHC11 apoprotein synthesis is coupled to chlorophyll synthesis at orbeyond translation. We propose that changes in energy balancebrought about by a change in light intensity may control a regu-latory factor acting to repress cab mRNA expression in high light.
Variations in light intensity often have profound effects onthe organization and function ofthe photosynthetic apparatusof both mature higher plants and algae (4, 8, 9, 19). Thephysiological response to variations in light intensity, var-iously called sun-shade-, light-shade-, or photoadaptation, isa reversible phenomenon that results in an adjustment of thepigment composition, absorption cross-section, and minimalturnover time for electron transport from water to C02, whichleads to an optimization of light harvesting and energy con-version capacity of the photosynthetic apparatus (3, 5).Compared with algae, higher plants have a reduced ability
to physiologically adapt to wide variations in irradiance. Mosthigher plants are genetically adapted (in a Darwinian sense)to HL' or LL. For example, Zea mays will not develop or
' This research was supported by the U.S. Department of Energy,Office of Basic Biological Energy Sciences, and by a Bourse Lavoisierfrom Ministere des Affaires Etrangeres to Dr. Anne Mortain-Bertrand.
2 Present address: Universite Bordeaux I, Laboratoire de Biologieet Physiologie Vegetale, Avenue des Facultes, 33405 Talence. cedexFrance.
3Abbreviations: HL, high light; LL, low light; LHC II, light har-vesting complex of photosystem II; gabaculine, 3-amino-2.3-dehydro-benzoic acid.
grow in the shade (2), whereas the tropical rain forest plant,Alocasia macrorhiza, cannot tolerate full sun (3). In contrast,most algae have evolved a physiological plasticity that allowsthem to photoadapt to wide variations in light intensity (5,8). Among the algae, the chlorophytes are the most physio-logically plastic in varying the Chl content per cell (5). Thesealgae have a photosynthetic apparatus similar to that foundin higher plants (30). The photoadaptation response of themarine chlorophyte Dunaliella tertiolecta has been character-ized at the biophysical (30), physiological (6, 7, 9, 30), andbiochemical levels (27, 30). However, the mechanism of pho-toadaptation is poorly understood at the molecular level.
In D. tertiolecta, the LHC II contains four polypeptides,each one different at the N-terminal end (18). The proteinsare encoded by a small cab gene family comprising three tofive members (18). In this alga, a 10-fold decrease in lightintensity typically results in a two- to threefold increase in thecellular abundance of light-harvesting Chl a/b binding pro-teins, accompanied by a corresponding increase in pigments(28, 29). Using a kinetic approach, Sukenik et al. (29) dem-onstrated that the increase in the LHC II complex in HLgrown cells transferred to LL follows first order kinetics witha half-time ranging between 16 and 24 h, and that the LHCII apoproteins are the first component of the photosyntheticapparatus to respond to a change to a lower light intensity.
In this paper, we examine changes in cab mRNA and LHCII apoprotein levels following a shift from high to low lightintensity. Our results reveal that the LL-induced increase inLHC II apoproteins followed a large initial increase in the cabmRNA. The mRNA increase can be detected within 1.5 hafter the shift to LL, and experiments with a nuclear transcrip-tion inhibitor suggest that transcription is required for the fulldevelopment of the photoadaptation response.
MATERIALS AND METHODS
Materials
Restriction endonucleases, random primers, and nick-translation kits were obtained from Bethesda Research Lab-oratories (Gaithersburg, MD). [a_-32P]dATP (3000 Ci/mmol),and [y_-32P]ATP (3000 Ci/mmol) were purchased from Du-Pont-New England Nuclear. Horseradish peroxidase immu-noblot detection kits were obtained from Bio-Rad.
147
Dow
nloaded from https://academ
ic.oup.com/plphys/article/97/1/147/6087173 by guest on 21 D
ecember 2021

Plant Physiol. Vol. 97, 1991
Figure 1. Expression of cab genes in steady-state high light (700 IAmol quanta m-2 s-') andlow light (70 uamol quanta m-2 S-1). Cells weregrown to steady-state at a cell density of 5 x105 cells/mL and total RNA and proteins har-vested and measured as described in "Materialsand Methods." Chi a/b is a molar ratio, LHC IIrepresents the relative amount of LHC II apopro-teins determined by Western blots (see "Mate- I i 'rials and Methods"), and cab represents the cabmRNA levels determined by Northern blot analy- isis (see "Materials and Methods"). The standarderrors on individual measurements of Chi a/celland ChI a/b are less than 10%. For HL-growncells, the ChI a/cell and Chi a/b ratios averaged0.42 ± 0.04 pg/cell and 11.2 ± 1.2 respectively,in replicate experiments (n = 11). For LL-growncells, the Chi a/cell and ChI a/b averaged 1.2 ±0.2 pg/cell and 7.6 ± 0.9, respectively, in repli-cate experiments (n = 10). The differences be-tween HL and LL Chi a/cell and Chi a/b ratiosare statistically significant at P < 0.001.
Strain and Culture Conditions
The unicellular marine chlorophyte Dunaliella tertiolecta(Woods Hole clone DUN) was grown as previously described(9) in artificial seawater enriched with f/2 nutrients (13), at1 8TC. Continuous light was provided by banks of high outputfluorescent tubes. The cultures were maintained in a semicon-tinuous turbidostat mode at a cell density of 5 x I05 cells/mL. For kinetic studies, growth irradiance was shifted in asingle step from 700 (HL) to 70 (LL) rmol quanta m-2 s-'. Acontrol measurement was made 24 or 12 h before the start ofthe light shift, initial measurements were made just prior tothe shift in light intensity (time zero), and additional meas-urements were made up to 1 week after the light shift.
Pigment Determination and Cell Counts
Pigment determinations were performed as previously de-scribed (28). Culture aliquots were filtered onto a glass fiberfilter and homogenized in 90% acetone. Corrected spectrawere recorded on an Aminco DW2-C spectrophotometer, andChl a and b were calculated using the equation of Jeffrey andHumphrey (14). Cells were counted using a haemocytometer.The average count of eight fields was used to calculate thecellular pool size of Chl a. Standard errors of replicate Chl/cell measurements and Chl a/b ratios were always less than10 and 5% of the values cited in the text, respectively.
Protein Analysis
Total protein preparations were as previously described(28). Approximately 2.5 x 108 cells were concentrated bycentrifugation and resuspended in lysis buffer (100 mMNa2CO3, 100 mm DTT, and 200 ,M PMSF). The suspensionwas sonicated 2 min on ice using a Kontes microprobesonicator. SDS, glycerol, and bromothymol blue were addedto a final concentration of 0.5, 10, and 0.05%, respectively.The samples were heated for 2 min at 95"C. Protein concen-trations in the solubilized cell extracts were measured using a
lI jII ;,:
.4 !g | , i
BCA kit (Pierce Chemical Co., St. Louis). For SDS-PAGE,equal amounts ofprotein were loaded on 15% polyacrylamidegel, and proteins were transferred by electrophoresis to nitro-cellulose. The levels of LHC II apoproteins were determinedby immunodetection using anti-LHC II raised against pea(28). The relative quantity of LHC II apoprotein was esti-mated by densitometric scanning of the Western blots.
RNA Isolation and Northern Blot Hybridization
Two to five hundred milliliters of cell culture (approxi-mately 5 x 105 cells/mL) were harvested by centrifugationand the RNA isolated as described (18). Two to four micro-grams of RNA/lane were separated by electrophoresis on 1%agarose/formaldehyde gels and transferred to nitrocellulose(21). Hybridization solution and conditions were as in Man-iatis et al. (21). Restriction fragments containing a cDNAencoding a cab gene (pDTcabl) from D. tertiolecta (18) andan a-tubulin gene from Chlamydomonas reinhardtii werelabeled by nick-translation (21). The relative amounts of cabmRNA were estimated by densitometric scanning of autora-diograms. Peak areas from the densitometric scans were usedto calculate the relative abundance for each sample. Thedifferent intensities of cab mRNA at HL observed in thedifferent experiments are due to variations in the exposuresof the autoradiograms.
RESULTS
Effect of Light Intensity on Steady-State Levels of theLHC 11 Complex
The amount of total Chl, the Chl a/b ratios, LHC IIapoproteins, and cab mRNA levels were measured in cellsgrown at steady-state in HL and LL (Fig. 1). Chl a/cell wasthreefold higher in the LL-grown culture, whereas the Chl a/b ratio was approximately 30% lower than that in the HLculture. LHC II apoproteins were approximately twofoldhigher in the LL cells. In addition, there was a change in the
148 LAROCHE ET AL.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/97/1/147/6087173 by guest on 21 D
ecember 2021
LIGHT-INTENSITY REGULATION OF cab mRNA IN DUNALIELLA TERTIOLECTA
a1.4
pg Chia/cell
0.012.0
Chia/Chlb(mol/mol)
2.0
Time (h) 0 9 18 27 36
MW(kbp)b
oe-tubulin
cab
LHCII
Time (h)
-1.6
-* * *1.2
(KDa)31
24.5O 9 18 27 36
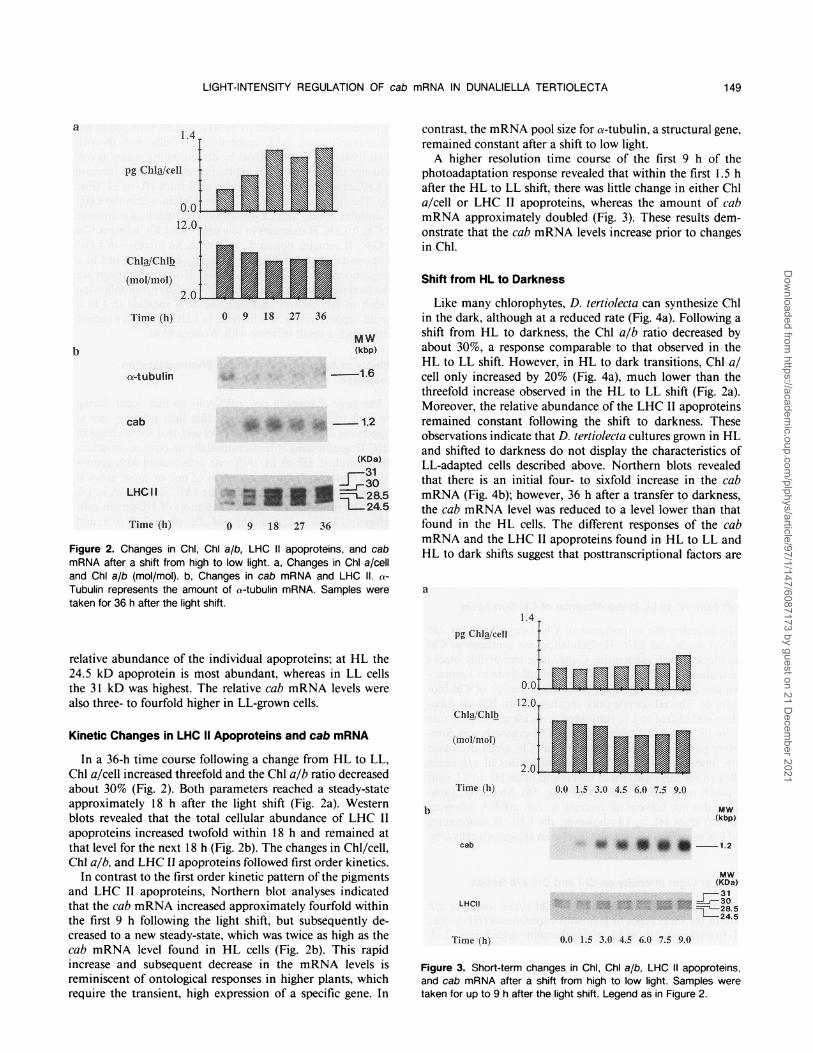
Figure 2. Changes in Chi, ChI a/b, LHC 11 apoproteins, and cabmRNA after a shift from high to low light. a, Changes in Chi a/celland ChI a/b (mol/mol). b, Changes in cab mRNA and LHC IL. a-Tubulin represents the amount of a-tubulin mRNA. Samples weretaken for 36 h after the light shift.
relative abundance of the individual apoproteins; at HL the24.5 kD apoprotein is most abundant, whereas in LL cellsthe 31 kD was highest. The relative cab mRNA levels werealso three- to fourfold higher in LL-grown cells.
Kinetic Changes in LHC 11 Apoproteins and cab mRNA
In a 36-h time course following a change from HL to LL,Chl a/cell increased threefold and the Chl a/b ratio decreasedabout 30% (Fig. 2). Both parameters reached a steady-stateapproximately 18 h after the light shift (Fig. 2a). Westernblots revealed that the total cellular abundance of LHC IIapoproteins increased twofold within 18 h and remained atthat level for the next 18 h (Fig. 2b). The changes in Chl/cell,Chl a/b, and LHC II apoproteins followed first order kinetics.
In contrast to the first order kinetic pattern of the pigmentsand LHC II apoproteins, Northern blot analyses indicatedthat the cab mRNA increased approximately fourfold withinthe first 9 h following the light shift, but subsequently de-creased to a new steady-state, which was twice as high as thecab mRNA level found in HL cells (Fig. 2b). This rapidincrease and subsequent decrease in the mRNA levels isreminiscent of ontological responses in higher plants, whichrequire the transient, high expression of a specific gene. In
contrast, the mRNA pool size for a-tubulin, a structural gene,remained constant after a shift to low light.A higher resolution time course of the first 9 h of the
photoadaptation response revealed that within the first 1.5 hafter the HL to LL shift, there was little change in either Chla/cell or LHC II apoproteins, whereas the amount of cabmRNA approximately doubled (Fig. 3). These results dem-onstrate that the cab mRNA levels increase prior to changesin Chl.
Shift from HL to Darkness
Like many chlorophytes, D. tertiolecta can synthesize Chlin the dark, although at a reduced rate (Fig. 4a). Following ashift from HL to darkness, the Chl a/b ratio decreased byabout 30%, a response comparable to that observed in theHL to LL shift. However, in HL to dark transitions, Chl a/cell only increased by 20% (Fig. 4a), much lower than thethreefold increase observed in the HL to LL shift (Fig. 2a).Moreover, the relative abundance of the LHC II apoproteinsremained constant following the shift to darkness. Theseobservations indicate that D. tertiolecta cultures grown in HLand shifted to darkness do not display the characteristics ofLL-adapted cells described above. Northern blots revealedthat there is an initial four- to sixfold increase in the cabmRNA (Fig. 4b); however, 36 h after a transfer to darkness,the cab mRNA level was reduced to a level lower than thatfound in the HL cells. The different responses of the cabmRNA and the LHC II apoproteins found in HL to LL andHL to dark shifts suggest that posttranscriptional factors are
a
1.4
pg Chla/cell
0.0 LiuiiXChlaJChlb
(mol/inol)
Time (h) 0.0 1.5 3.0 4.5 6.0 7.5 9.0
b MW(kbp)
cab
LHCII
Time (h)
_ _ w ~~1.2
MW(KDa)-31
0 1. 3.0 4.5 6.0s 7.5 9: <.0* , 30
-24.5
0.0 1.53.0 4.5 6.0 7.5 9.0
Figure 3. Short-term changes in Chi, ChI a/b, LHC 11 apoproteins,and cab mRNA after a shift from high to low light. Samples weretaken for up to 9 h after the light shift. Legend as in Figure 2.
149
Dow
nloaded from https://academ
ic.oup.com/plphys/article/97/1/147/6087173 by guest on 21 D
ecember 2021
Plant Physiol. Vol. 97, 1991
I, I &&'Ibc
t
.1
t
I-1
..)I
MW(kbp
rab
MW(KDal31
-30L-HCU3I. U 3z1~
24.5
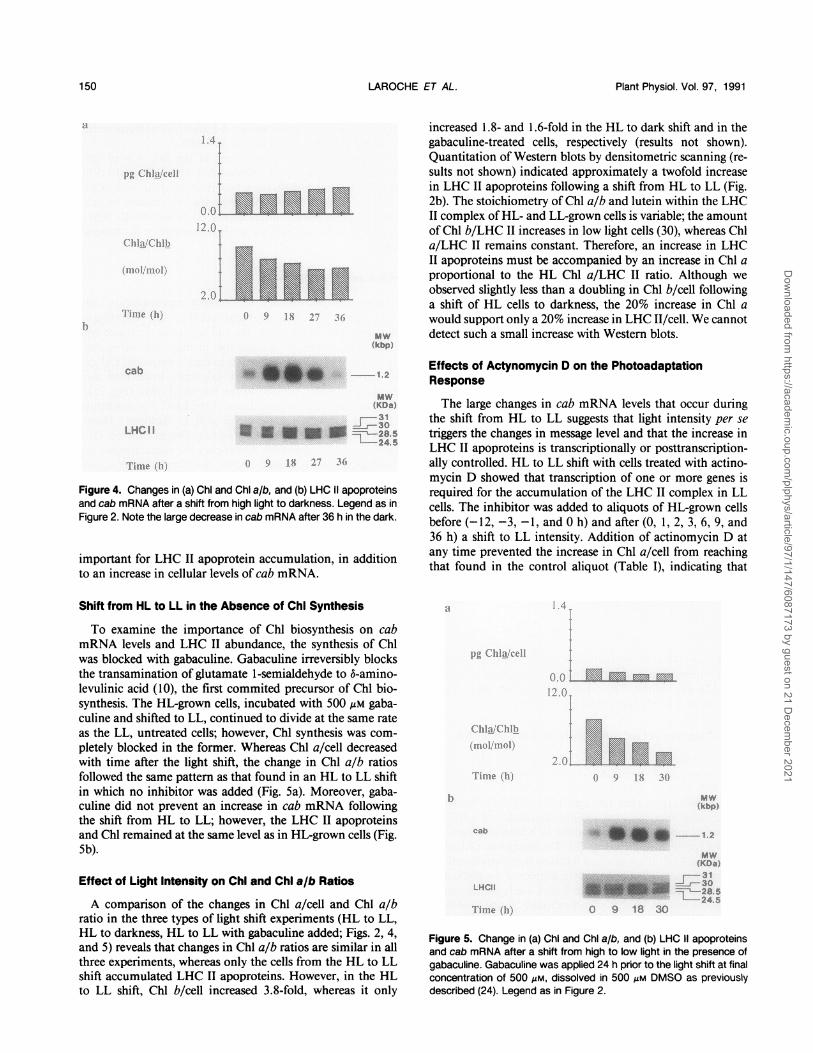
Figure 4. Changes in (a) ChI and Chi a/b, and (b) LHC II apoproteinsand cab mRNA after a shift from high light to darkness. Legend as inFigure 2. Note the large decrease in cab mRNA after 36 h in the dark.
important for LHC II apoprotein accumulation, in additionto an increase in cellular levels of cab mRNA.
increased 1.8- and 1.6-fold in the HL to dark shift and in thegabaculine-treated cells, respectively (results not shown).Quantitation of Western blots by densitometric scanning (re-sults not shown) indicated approximately a twofold increasein LHC II apoproteins following a shift from HL to LL (Fig.2b). The stoichiometry of Chl a/b and lutein within the LHCII complex ofHL- and LL-grown cells is variable; the amountof Chl b/LHC II increases in low light cells (30), whereas Chla/LHC II remains constant. Therefore, an increase in LHCII apoproteins must be accompanied by an increase in Chl aproportional to the HL Chl a/LHC II ratio. Although weobserved slightly less than a doubling in Chl b/cell followinga shift of HL cells to darkness, the 20% increase in Chl awould support only a 20% increase in LHC II/cell. We cannotdetect such a small increase with Western blots.
Effects of Actynomycin D on the PhotoadaptationResponse
The large changes in cab mRNA levels that occur duringthe shift from HL to LL suggests that light intensity per setriggers the changes in message level and that the increase inLHC II apoproteins is transcriptionally or posttranscription-ally controlled. HL to LL shift with cells treated with actino-mycin D showed that transcription of one or more genes isrequired for the accumulation of the LHC II complex in LLcells. The inhibitor was added to aliquots of HL-grown cellsbefore (-12, -3, -1, and 0 h) and after (0, 1, 2, 3, 6, 9, and36 h) a shift to LL intensity. Addition of actinomycin D atany time prevented the increase in Chl a/cell from reachingthat found in the control aliquot (Table I), indicating that
Shift from HL to LL in the Absence of Chi Synthesis
To examine the importance of Chl biosynthesis on cabmRNA levels and LHC II abundance, the synthesis of Chlwas blocked with gabaculine. Gabaculine irreversibly blocksthe transamination of glutamate 1-semialdehyde to 6-amino-levulinic acid (10), the first commited precursor of Chl bio-synthesis. The HL-grown cells, incubated with 500 ,uM gaba-culine and shifted to LL, continued to divide at the same rateas the LL, untreated cells; however, Chl synthesis was com-pletely blocked in the former. Whereas Chl a/cell decreasedwith time after the light shift, the change in Chl a/b ratiosfollowed the same pattern as that found in an HL to LL shiftin which no inhibitor was added (Fig. 5a). Moreover, gaba-culine did not prevent an increase in cab mRNA followingthe shift from HL to LL; however, the LHC II apoproteinsand Chl remained at the same level as in HL-grown cells (Fig.Sb).
Effect of Light Intensity on Chi and Chi a/b Ratios
A comparison of the changes in Chl a/cell and Chl a/bratio in the three types of light shift experiments (HL to LL,HL to darkness, HL to LL with gabaculine added; Figs. 2, 4,and 5) reveals that changes in Chl a/b ratios are similar in allthree experiments, whereas only the cells from the HL to LLshift accumulated LHC II apoproteins. However, in the HLto LL shift, Chl b/cell increased 3.8-fold, whereas it only
;hia cvc.
ia ( tlkI
kAb
.H C
,lItle !h
,~ ~~m
3C
Figure 5. Change in (a) Chi and ChI a/b, and (b) LHC II apoproteinsand cab mRNA after a shift from high to low light in the presence ofgabaculine. Gabaculine was applied 24 h prior to the light shift at finalconcentration of 500 AM, dissolved in 500 AM DMSO as previouslydescribed (24). Legend as in Figure 2.
150 LAROCHE ET AL.
t t t)
ON"
Dow
nloaded from https://academ
ic.oup.com/plphys/article/97/1/147/6087173 by guest on 21 D
ecember 2021

LIGHT-INTENSITY REGULATION OF cab mRNA IN DUNALIELLA TERTIOLECTA
Table I. Effect of Actinomycin D on the Change in Chl a/cell and inChl a/b Ratio Following a Shift from HL to LL
All measurements were made after 36 h incubation in LL exceptfor the HL initial measurements, which were performed immediatelyprior to the light shift. The number in brackets in the treatment columncorresponds to the time (in h, relative to the shift to low light at timezero) at which actinomycin D (25 gg/mL final concentration) wasadded to the aliquots. Each measurement is followed by the per-centage change relative to the difference between the initial HL andthe final LL samples, untreated with actinomycin D.
Treatment Cell x 1 06/mL % pg Chl a/cell % Chl a/b %
HL initial 0.52 0 0.36 0 13.0 0LL (-12) 0.58 8 0.59 20 11.3 25LL (-3) 0.84 42 0.60 20 12.3 10LL (-1) 1.02 65 0.47 10 12.1 13LL (0) 0.95 57 0.65 25 10.4 38LL (1) 0.97 59 0.58 19 10.1 43LL (2) 1.01 64 0.65 24 9.8 47LL (3) 0.93 54 0.73 32 8.4 68LL (6) 1.03 67 0.88 44 8.8 62LL (9) 1.09 75 0.96 51 8.0 74LL (36) 1.28 100 1.53 100 6.2 100LL final 1.28 100 1.52 100 6.2 100
transcription is required for at least 9 h after the light shift. Itshould be noted that addition of actinomycin D for up to 12h before the light shift supported an increase in Chl a/cell of20%, similar to the increase observed in cells shifted todarkness (Fig. 4a). However, the 20% increases in Chl a/cellobserved in the HL to dark shift and the HL to LL shift inthe presence of actinomycin D are much smaller than thethreefold increase in Chl a/cell that is found between HL-and LL-acclimated cells.
DISCUSSION
In D. tertiolecta, a shift from HL to LL results in an
immediate, rapid reduction in growth (29), and a relativeincrease in the synthesis and accumulation of LHC II. Theaccumulation of the LHC II complex is dependent on theincreased activity of two biosynthetic pathways relative to thecellular growth rate, namely Chl and LHC II apoproteinbiosyntheses. There is evidence suggesting that these twopathways are interrelated (12, 23, 24), but the control pointsbetween them remain obscure.The comparison of cab mRNA and LHC II apoproteins in
HL or LL steady-state grown cells indicates that the cab genesare expressed at a higher level in cells grown in steady-stateLL (Fig. 1). Following a shift from HL to LL, cab mRNAlevels reached a transient maximum, which coincided with a
period of rapid LHC II apoprotein accumulation. Measure-ments of Chl, LHC II apoproteins, and cab mRNA withinthe first 9 h after a shift from HL to LL indicated that theincrease in cab mRNA precedes, and is initially independentfrom, an increase in Chl synthesis. Our results demonstratethat in D. tertiolecta the LL-induced increase in the LHC II
apoproteins followed an increase in cab mRNA levels andthat Chl synthesis is not required for this initial increase. Theincrease in cab mRNA relative to other mRNA species during
photoadaptation to LL is very specific. In addition to a-tubulin mRNA (Fig. 2B), the levels of several other mRNAspecies encoding photosynthetic proteins, including psaB,psbA, rbcL, were measured and showed either no variationor a slight decrease in the first 36 h after a shift from HL toLL (J. LaRoche, unpublished results).The cab gene probe used in this study hybridizes to several
members of the cab gene family of D. tertiolecta (18). Al-though we cannot assess to what extent individual cab genesare differentially expressed after a shift to low light intensity,it is clear that the overall response results in an increase incab transcripts.The cells failed to photoadapt when transferred to LL in
the presence of actinomycin D (Table I), suggesting thatnuclear gene transcription is required to initiate the photoad-aptive response. Addition of actinomycin D to cell aliquotsat intervals following the shift to low light indicated thattranscription of at least one, and possibly more, gene isrequired for up to 9 h during the course of photoadaptation.However, it is not possible to determine from the results ofthis experiment whether the relative increase in the cabmRNA levels result specifically from an increase in transcrip-tion of the cab genes or from a change in the stability of thismessage relative to others.
In D. tertiolecta cells synchronized to a light/dark cycle,the cab mRNA levels are slightly higher in the light than inthe dark (J. La Roche, unpublished results). Therefore, it isimportant to note that the cab mRNA response to a light/dark cycle is opposite to that found in an asynchronousculture shifted to darkness (Fig. 4). These observations suggestthat the fluctuations in cab mRNA levels measured in cellssynchronized to a light/dark cycle and HL-grown cells shiftedto LL or darkness are controlled by different factors.The Chl a/b ratio of D. tertiolecta decreases following a
shift from HL to LL. The light-harvesting complexes of algae,and higher plants in general, are rich in Chl b or Chl c, whichpreferentially bind to LHC apoproteins. Moreover, Sukeniket al. (30) demonstrated that in D. tertiolecta, the Chl a/bratio of the ensemble of LHC II complexes from LL-growncells is actually half that of HL-grown cells. During photoad-aptation, two independent responses can explain the changein Chl a/b ratio: a change in the rate of Chl a synthesis, and/or a change in the conversion rate of Chl a to Chl b. Thisstudy and others (20, 25) demonstrate that a change in Chla/b occurs when chlorophytes are transferred to darkness, orwhen Chl synthesis is blocked by gabaculine, as well as aftera change in light intensity. The change in pigment composi-tion is possibly a byproduct of changes in the pigment syn-thesis rate and pool size generated by a wide range of stressstimuli.
Mechanism of Light Intensity Regulation
In greening seedlings and Chlamydomonas, Chl synthesisis required to stabilize nascent LHC II apoproteins. Mortain-Bertrand et al. (24) demonstrated that the light intensity-induced increase in the LHC II apoproteins requires Chlbiosynthesis. However, it does not appear to be regulated byposttranslational stabilization of the nascent apoprotein byChl molecules, but is the result of an increase in synthesis of
151
Dow
nloaded from https://academ
ic.oup.com/plphys/article/97/1/147/6087173 by guest on 21 D
ecember 2021

Plant Physiol. Vol. 97, 1991
the specific apoproteins (24). An increase in cab mRNAfollowing a shift from HL to LL in the presence of gabaculine,or after a shift to darkness, did not lead to an increase in LHCII apoproteins (Fig. 5). Thus, our results suggest that Chlsynthesis is not required for the initial increase in cab mRNAlevels and that LHC II apoprotein synthesis is coupled to Chlsynthesis at or beyond the translational level.Gabaculine does not inhibit the cab mRNA accumulation,
suggesting that any Chl biosynthetic intermediate beyond 5-aminolevulinic acid is unlikely to regulate the increase in cabmRNA. However, our observations do not preclude the pos-
sibility that the accumulation of a Chl intermediate may exertnegative feedback on the cab mRNA levels (15).We suggest that energy availability and its effect on linear
electron transport play an important role in controlling thechange in cab mRNA during photoadaptation. Variations ofthis hypothesis have been suggested independently by severalworkers (1, 5, 11, 22). In Chlorella and cyanobacteria, sub-micromolar additions of DMCU induced a photoadaptiveresponse in cells maintained in HL (1, 17). The effect ofDCMU at HL strongly suggests that light intensity does nothave a direct effect on the regulatory factor per se. A similarinterpretation can be derived from the work of Shugarmanand Appleman (27) and Kindle (16), who showed in twodifferent species of chlorophytes that supplementing a culturewith an organic carbon source has the same effect as raisingthe light intensity; that is, it reduces the amount of Chl/cell.These results can be explained in terms of a balance betweenphotosynthetic energy and photosynthate accumulationwithin the cell. Sheen (26) has provided strong evidence ofmetabolic repression of higher plants' photosynthetic genes
by several sugars. However, the mechanism for such repres-
sion remains elusive.We propose that, in D. tertiolecta, cab gene expression is
repressed by a factor present only at HL, which is, in turn,regulated via changes in noncyclic photosynthetic electrontransport. Our results reveal that cab mRNA shows the sameinitial increase upon removal of high light growth conditions,regardless of whether the cells are shifted to darkness or toLL. These observations suggest that cab genes are suppressedin HL rather than activated by LL.
ACKNOWLEDGMENTS
We are indebted to Dr. Donald Weeks for his gift of the a-tubulinprobe from Chlamvdomonas, and to Ms. Barbara Randolf Andersonfor the gift of all other Chiamydomonas photosynthetic gene probes.We also thank Dr. John Bennett for his advice and comments duringthis work and we are grateful to K. Wyman for technical assistance.
LITERATURE CITED
1. Beale SI, Appleman D (1971) Regulation by degree of lightlimitation of growth. Plant Physiol 59: 230-235
2. Bennett J, Schwender JR, Shaw EK, Tempel N, Ledbetter M,Williams RS (1987) Failure of corn leaves to acclimate to lowirradiance. Role of protochlorophyllide reductase in regulatinglevels of five chlorophyll-binding proteins. Biochim BiophysActa 892: 118-129
3. Boardman NK (1977) Comparative photosynthesis of sun andshade plants. Annu Rev Plant Physiol 28: 355-377
4. Eskins K, Westhoff P, Beremand PD (1989) Light quality andirradiance level interaction in the control of expression of light-
harvesting complex of photosystem II. Plant Physiol 91: 163-169
5. Falkowski PG (1980) Light-shade adaptation in marine phyto-plankton. In PG Falkowski, ed, Primary Productivity in theSea. Plenum Press, New York, pp 99-119
6. Falkowski PG (1984) Kinetics of adaptation to irradiance inDunaliella tertiolecta. Photosynthetica 18: 62-68
7. Falkowski PG, Dubinsky Z, Wyman K (1985) Growth-irradiancerelationships in phytoplankton. Limnol Oceanogr 30: 311-321
8. Falkowski PG, LaRoche J (1991) Acclimation to spectral irra-diance in algae. J Phycol 26: 8-14
9. Falkowski PG, Owens TG (1980) Light-shade adaptation. Twostrategies in marine phytoplankton. Plant Physiol 66: 632-635
10. Flint DH (1984) Gabaculine inhibits b-ALA synthesis in chloro-plasts. (abstract No. 965). Plant Physiol 75: S-170
1 1. Fujita Y, Iwama Y, Ohki K, Murakami A, Hagiwara N (1989)Regulation of the size of light-harvesting antennae in responseto light intensity in the green alga Chlorella pyrenoidosa. PlantCell Physiol 30: 1029-1037
12. Genge SD, Polger D, Hiller RG (1974) The relationship betweenchlorophyll b and pigment-protein complex II. Biochim Bio-phys Acta 347: 22-30
13. Guillard RRL, Ryther JH (1962) Studies of marine planktonicdiatoms. I. Cvclotella nana (Hustedt) and Detonula confervacea(Cleve). Can J Microbiol 8: 229-239
14. Jeffrey SW, Humphrey GW (1975) New spectrophotometricequations for determining chlorophylls a, b, cl, c2 in higherplants, algae and natural phytoplankton. Biochem PhysiolPflanz 167: 191-194
15. Johanningmeier U (1989) Possible control of transcript levels bychlorophyll precursors in Chlamvdomonas. Eur J Biochem177: 4 17-424
16. Kindle KL (1987) Expression of a gene for a light-harvestingchlorophyll a/b-binding protein in Chlamvdomonas reinhard-Iii. effect of light and acetate. Plant Mol Biol 9: 547-583
17. Koenig F (1990) Shade adaptation in cyanobacteria. Furthercharacterization of Anacvstis shade phenotype as induced bysublethal concentrations of DCMU-type inhibitors in stronglight. Photosynth Res 26: 29-37
18. LaRoche J, Bennett J, Falkowski PG (1990) Characterization ofa cDNA encoding for the 28.5 kDa LHCII apoprotein fromthe unicellular marine chlorophyte, Duinaliella tertiolecta.Gene 95: 165-171
19. Leong T-Y, Goodchild DJ, Anderson JM (1985) Effects of lightquality on the composition, function, and structure of photo-synthetic thylakoid membranes of Asplenium aiustralasicuim(Sm.) Hook. Plant Physiol 78: 561-567
20. Maloney M, Hooberk JK, Marks DB (1989) Kinetics of chlo-rophyll accumulation and formation of chlorophyll-proteincomplexes during greening of Chlamvdomonas reinhardtii y-1at 38'C. Plant Physiol 91: 1100-1106
21. ManiatisT, Fritsch EF, Sambrook J (1982) Molecular Cloning.A Laboratory Manual. Cold Spring Harbor Laboratory. ColdSpring Harbor, NY
22. Melis A, Manodori A, Glick RE, Ghirardi ML, McCauley SW,Neale PJ (1985) The mechanism of photosynthetic membraneadaptation to environmental stress conditions: a hypothesis onthe role of electron-transport capacity and of ATP/NADPHpool in the regulation of thylakoid membrane organizationand function. Physiol Veg 23: 757-765
23. Michel H, Tellenbach MH, Boschetti A (1983) A chlorophyllh-less mutant of Chlam.vdomonas reinhardiii lacking in the light-harvesting chlorophyll a/b complex but not in its apoproteins.Biochim Biophys Acta 724: 417-424
24. Mortain-Bertrand A, Bennett J, Falkowski PG (1990) Photore-gulation of the light-harvesting chlorophyll protein complexassociated with photosystem11 in Diunaliellatertioleuta. PlantPhysiol 94: 304-311
25. Pick U, Gounaris K, Barber J (1987) Dynamics of photosystem11 and its light harvesting system in response to light changesin the halotolerant alga Dunaliella salina. Plant Physiol 85:194- 198
152 LAROCHE ET AL.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/97/1/147/6087173 by guest on 21 D
ecember 2021

LIGHT-INTENSITY REGULATION OF cab mRNA IN DUNALIELLA TERTIOLECTA
26. Sheen J (1990) Metabolic repression of transcription in higherplants. Plant Cell 2: 1027-1038
27. Shugarman PM, Appleman D (1966) Chlorophyll synthesis in(li/orella. 11. Effect of glucose and light intensity on the lagphase. Plant Physiol 41: 1701-1708
28. Sukenik A, Bennett J, Falkowski P (1988) Changes in theabundance of individual apoproteins of light-harvesting chlo-rophyll a/h-protein complexes of photosystem I and II with
growth irradiance in the marine chlorophyte Duinaliella terti-olecta. Biochim Biophys Acta 932: 206-215
29. Sukenik A, Bennett J, Mortain-Bertrand A, Falkowski PG (1990)Adaptation of the photosynthetic apparatus to irradiance inDunaliella tertiolecia. Plant Physiol 92: 891-898
30. Sukenik A, Wyman KD, Bennett J, Falkowski PG (1987) Anovel mechanism for regulating the excitation of photosystemII in a green alga. Nature 327: 704-707
153
Dow
nloaded from https://academ
ic.oup.com/plphys/article/97/1/147/6087173 by guest on 21 D
ecember 2021