chlorophyll organization in green photosynthetic bacteria
TRANSCRIPT

33
Biochimica et Biophysica A cta, 594 (1980) 33 -51 Elsevier/North-Holland Biomedical Press
BBA 86065
C H L O R O P H Y L L O R G A N I Z A T I O N IN G R E E N PHOTOSYNTHETIC BACTERIA *
JOHN M. OLSON
Biology Department, Brookhaven National Laboratory, Upton, NY 11973 (U.S.A.)
(Received June 3rd, 1980)
Contents
I. Classification and cell structure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
II. Photosynthetic unit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35 A. Chlorophylls . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
III. Cytoplasmic membrane (Chlorobiaceae) . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36 A. Unit-membrane vesicles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36 B. Reaction centers . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 C. Bacteriochlorophyll a-proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
IV. Chlorosome (Chlorobiaceae) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44 A. Baseplate structure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44 B. Rod elements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
V. Reaction-center complexes (Chlorobiaceae) . . . . . . . . . . . . . . . . . . . . . . . . . . 48 A. Intramembrane particles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
VI. Chloroflexacea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
VII. Summary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
Note added in proof . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
I. Classification and cell s t ruc ture
Green bacter ia include green sulfur bacter ia (Chlorobiaceae) and gliding f i l amentous
bacter ia (Chlorof lexaceae) [1] . Green sulfur bacter ia (e.g. Chlorobium and Prosthe- cochloris) are obligate p h o t o a u t o t r o p h s and str ict anaerobes that grow in d im light. Glid-
* A shorter version of this paper was presented at the 8th Annual Meeting of the American Society for Photobiology, 17-21 February 1980, in Colorado Springs, Colorado, U.S.A. Abbreviations: BChl, bacteriochlorophyll; BPh, bacteriopheophytin; CD, circular dichroism; EPR, electron paramagnetic resonance; SDS, sodium dodecyl sulfate. By acceptance of this article, the publisher and/or recipient acknowledges the U.S. Government's right to retain a nonexclusive, royalty-free license in and to any copyright covering this paper.
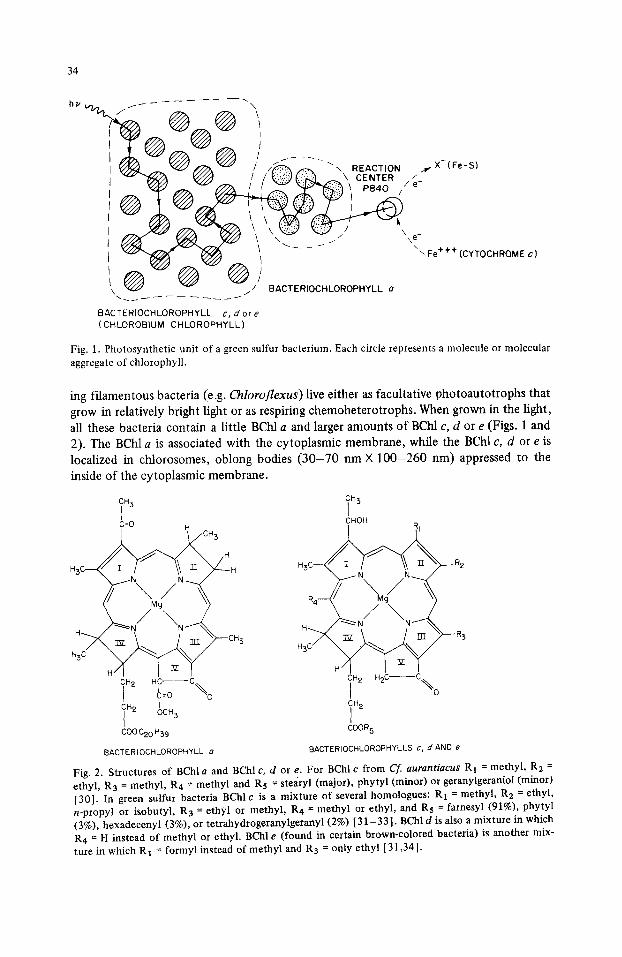
34
~ ; ~ - - ~ ( ~ - ~ / t ~ v'4¢/ / / ~ .<~. ~ \ REACTION ~e,X (Fe-S) "v-z-'~Z/j~ " I~yy/~ / / ~,!;',!~)(;~;~ ~ \ CENTER ./_
I / ~ ~1 P840 //e
x ~ . / BACTERIOCHLOROPHYLL a
BACTERIOCHLOROPHYLL c, dore (CHLOROBIUM CHLOROPHYLL)
Fig. 1. Photosynthetic unit of a green sulfur bacterium. Each circle represents a molecule or molecular aggregate of chlorophyll.
ing f d a m e n t o u s bac te r i a (e.g. Chloroflexus) live e i the r as facul ta t ive p h o t o a u t o t r o p h s t h a t
grow in relat ively b r igh t l ight or as respir ing c h e m o h e t e r o t r o p h s . When g rown in the l ight ,
all these bac te r i a c o n t a i n a l i t t le BChl a and larger a m o u n t s o f BChl c, d or e (Figs. 1 and
2). The BChl a is associa ted w i t h the cy t op l a s m i c m e m b r a n e , whi le the BChl c, d or e is
local ized in ch lo rosomes , o b l o n g bod ies ( 3 0 - 7 0 n m X 1 0 0 - 2 6 0 n m ) appressed to the
inside o f the cy top l a smic m e m b r a n e .
~H3 i H3
, ~ 0 "~//CH3 H CHOH RI
H C - - C. ill2 H2C
1 OCH 5 iH2 i H2 COO C20 H59 COORs
BACTERIOCHLOROPHYLL a BACTERIOCHLOROPHYLLS c, d AND e
Fig. 2. Structures of BChla and BChlc, d or e. For BChlc from Cf. aurantiacus R1 = methyl, R 2 = ethyl, R3 = methyl, R4 = methyl and Rs = stearyl (major), phytyl (minor) or geranylgeraniol (minor) [30]. In green sulfur bacteria BChl c is a mixture of several homologues: R1 = methyl, R 2 = ethyl, n-propyl or isobutyl, R3 = ethyl or methyl, R4 = methyl or ethyl, and Rs = farnesyl (91%), phytyl (3%), hexadecenyl (3%), or tetrahydrogeranylgeranyl (2%) [ 31-33] . BChl d is also a mixture in which R¢ = H instead of methyl or ethyl. BChl e (found in certain brown-colored bacteria) is another mix- ture in which RI = formyl instead of methyl and R 3 = only ethyl [31,34].

35
I1. Photosynthetic unit
The photosynthetic units of green sulfur bacteria contain 1000-2000 BChl c, d or e molecules and about 100 BChl a molecules feeding excitation energy into a single reac- tion center [2]. (The smaller units found in gliding filamentous bacteria have less BChl c, d or e and are comparable in size to those of purple bacteria [1].) Fig. 1 illustrates the main functions of the chlorophylls in energy transfer in green bacteria. A photon absorbed by a molecule or molecular aggregate of BChl ¢, d or e raises that molecule or molecular aggregate to an excited singlet state. The excitation (lowest excited state) then migrates through other BChl c, d or e molecules or aggregates until it passes to a BChl a molecular aggregate. After further migration among various BChl a aggregates the excita- tion is localized on a specialized form of BChl a in the reaction center. In green sulfur bacteria this specialized BChl a (P-840) in the excited state is a powerful electron donor that transfers one electron to a low potential acceptor (probably an iron-sulfur center, E m = - 0 . 5 4 V) presumably by way of one or more intermediate carriers [3].
When green sulfur bacteria are transferred from high light intensity (20 0 0 0 - 1 0 0 000 lux) to low light intensity ( 2 0 - 1 0 0 lux) the specific BChl c or d content of whole cells at least doubles [1,4] and in some cases increases several-fold [1,5]. This increase is brought about by an increase in the number of chlorosomes per cell and by an increase in the size of the chlorosomes [1,4,5]. In Chloroflexus the BChl c content can easily be quadrupled by decreasing the intensity from 2000 lux to 400 lux [6].
IIA. Chlorophylls
The BChl a (Fig. 2) in green bacteria is identical to the BChl a in purple bacteria except for the substitution of geranylgeraniol for phytol by certain purple non-sulfur bac- teria (e.g. Rhodospirillum rubrum) [ 7 - 9 ] . BChl a accounts for 5 to 10% of the total chlorophyll in green sulfur bacteria and in vivo is responsible for the minor absorption band at 810 nm (Fig. 3). In the filamentous gliding bacterium Chloroflexus aurantT"acus BChl a accounts for the absorption bands at 805 and 865 nm shown in Fig. 4A.
0.9 l i I i I I I I i
730 0.8 458 r~ 0.7 446v /,?o
m <rY O. 5 , . j "~,\ 0 0.4 "
- , / \ \
N t 0.2 ' \ 0.1 - " "
O I I [ I I I I I L
400 500 600 700 800 WAVELENGTH (nm)
Fig. 3. Absorption spectra of cell free extracts of green sulfur bacteria containing BChl c (P. aestuarii strain 2K, solid curve) and BChl d (C. limicola f. thiosulfatophilum strain L, dashed curve).

36
0.8
0 .7
0.5
# o o.4 I
03
0.2~
74O
_ , o . . . . . . . . . . h , , ¢.r~n 700 800 900 700 800 900
WAVELENGTH (rim)
Fig. 4. A. Absorption spectrum of membrane fraction from Cf. aurantiacus grown under high intensity light (courtesy of S.G. Sprague and R.C. Fuller). B. Light-induced absorbance changes in cell free preparation of Cf aurantiacus in buffer. Reprinted by permission of Arch. Microbiol. from Pierson and Castenholz [91.
The main light-harvesting pigment of any given green bacterium is either BChl c, BChl d or BChl e. (BChl c and d used to be called chlorobium chlorophylls 660 and 650, respectively.) In vivo, BChl c has absorption bands at about 460 and 7 4 0 - 7 5 0 nm, whilst BChl d absorbs at about 450 and 730 nm (Figs. 3 and 4). In green sulfur bacteria these chlorophylls are esterified mainly by farnesol (C 1 sH26OH), but in C. aurantiacus BChl c is esterified mainly by stearyl alcohol (C 18H3 7OH) [ 10].
The most important difference between BChl a and BChl c, d or e is that ring II in BChl a is more reduced. This affects the n-rr* transitions so that the main absorption bands of BChl a in vivo lie below 400 nm and above 800 nm, whereas those of BChl c, d or e lie above 400 nm and below 800 nm (see Fig. 3). Another significant difference is that ring V is attached to a -COOCH3 in BChl a but to an -H in BChl c, d or e. Differences in the esterifying alcohol could be important in the binding of the various chlorophylls to specific proteins.
III. Cytoplasmic membrane (Chlorobiaceae)
IliA. Unit-membrane vesicles
When Chloroflexus is broken in a French pressure cell, the cytoplasmic membrane can be isolated mainly as flat pieces, but when sulfur bacteria are broken, the fragments are isolated (Fig. 5) as vesicles [ 11 ]. For Chlorobium limicola f. thiosulfatophilum the vesicle diameters range from 30 to 100 nm with a mean value of about 60 nm. Membrane thick- ness is between 7 and 8 nm [3]. In freeze-fracture electron micrographs (Fig. 6) many particles with a mean diameter of 8 nm can be seen on the inner face, but very few on the outer face. Since micrographs of intact ceils show that the particle density on the P (pro-

37
Whole cells in 10 mM sodium dithionite 3 X thru French pressure cell at 1400 atm (20 000 lb/inch 2)
Crude extract l Spin at 40 000 Xg for 90 min
Supernatant Spin at 105 Xg for 20 h on sucrose gradient (20-45%)
Membrane fraction (Complex I)
l Add 2 M NaCI and chromatograph
Complex I (Clean membranes)
Incubate in 2 M guanidine hydrochloride and chromatograph on Sepharose CL-6B
Complex II (membranes) and BChl a-protein
Fig. 5. Flow chart for the preparation of unit-membrane vesicles (Complexes I and II) from C limicola f. thiosulfatophilum.
toplasmic)-face of the cytoplasmic membrane is much higher than on the E (exoplasmic)- face [ 12], the inner face of the vesicle may correspond to the P-face of the cytoplasmic membrane.
The vesicle membranes are completely free of chlorosomes and contain essentially no BChl c. They do, however, contain BChl a (absorption maxima at 342, about 373, about 604, 810 and 835 nm), BPh c (674 nm and perhaps 560 nm), and carotenoid (471 and 506 nm) as shown in Fig. 7. They also contain photochemical reaction centers, evidenced by the light-induced absorbance changes at 610,830, 842 and 1157 nm reflecting the oxi- dation of P-840, and the absorbance change at 553 nm reflecting the oxidation of cyto- chrome c-553 (Fig. 8). Membrane preparations (Complex I ) f rom P. aestuarii [2] and 600-kdalton particles [13] prepared from these membranes with 1% Triton X-IO0 show similar absorption spectra and light-induced absorption changes. The best preparations from both Chlorobium and Prosthecochloris contain about 80 BChl a and 7 -8 BPh c molecules per reaction center (Swarthoff, T. and Amesz, J. (1980)personal communica- tion). At 100 K the absorption spectrum of BPh c in Complex I or particles from Pros- thecochloris shows three bands (665,670 and 675 nm) [4], and at 25°C the CD spectrum of BPh c in Complex I from Chlorobium shows a sizable negative band at 662 nm and an equivalent positive band at 676 nm (Olson, J.M. and Sutherland, J.C. (1980) unpublished results). These observations suggest an exciton interaction with components at 662-665 nm and 675-676 nm involving two or more BPh c molecules with some non-interacting BPh c (670 nm) also present.
About half the BChl a can be removed from the unit-membrane vesicles of Chlorobium by treatment with 2 M guanidine hydrochloride [3] or 2 M arginine hydrochloride [5]. Similar results can be obtained by treating the 600-kdalton particles from Prosthe- cochloris with 1 M guanidine hydrochloride [13]. The resulting 350-kdalton particles from Prosthecochloris retain much more photochemical activity than do the treated vesi- cles (Complex II) from Chlorobium, because the concentration of guanidine hydrochlo- ride is lower and the exposure time is shorter in the former case. The BChl a liberated by the guanidine hydrochloride is contained in water-soluble BChl a-proteins [6,7]. The

Fig.
6.
Fre
eze-
frac
ture
d pr
epar
atio
n o
f un
it-m
embr
ane
vesi
cles
(C
ompl
ex I
) fr
om C
. Km
icol
a f.
thio
sulf
atop
hilu
m.
Ves
icle
s w
ere
trea
ted
as d
escr
ibed
in [
12]
. B
efor
e fr
eezi
ng t
hey
wer
e su
spen
ded
in s
odiu
m d
ithi
onit
e (2
.5
mM
), s
odiu
m p
ho
sph
ate
(5 m
M)
and
glyc
erol
(30
% v
/v).
The
bar
rep
rese
nts
150
nm.
Cou
rtes
y o
f T
.H.
Gid
- di
ngs,
Jr.
and
L.A
. S
taeh
elin
.
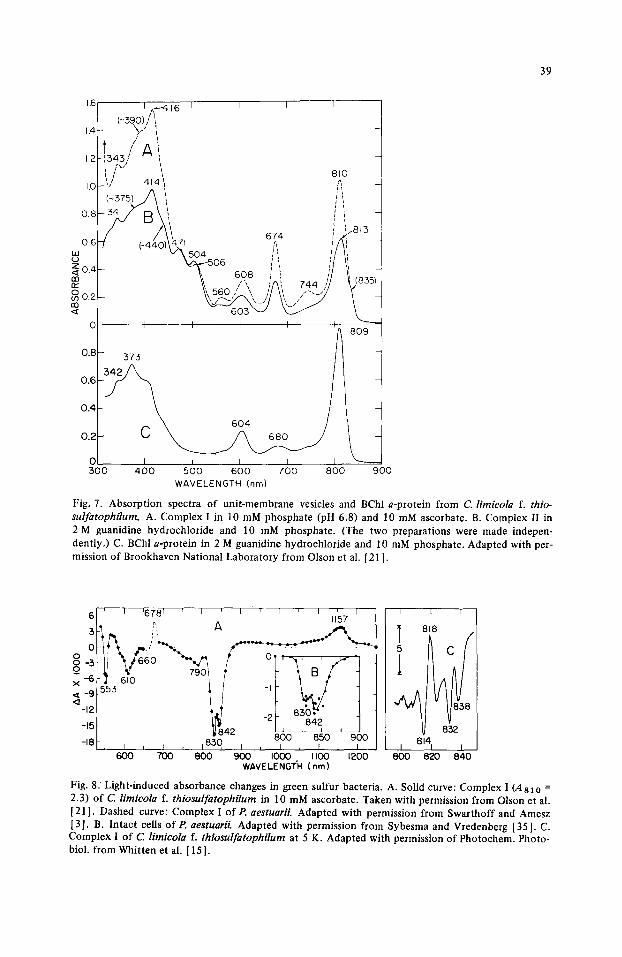
39
1.6 I . - 4 I ( ~ 3 9 0 ) / ~ 1 6 I I I I
,4~- ",,-' ', ~1343,'' A ~i i ! i
1.2 I f " h~ ' ~ 810
1.0 'J 414 '
0 .6 47 ,, /\, ',' ' ~ ~o4 :i / / ' , '-'~ o.4 k.--.~o6 i,, l/if
't, 6 0 8 ~ ,, , / /i 13B r r ~{, , " ]AL~ 7 4 4 /'/ \ ( 8 3 . \\ 560 / ~, / / \~, . - ._ . ' / [
I I I I i
0.8- 373 / ~ 809
/ 0.6 -
0.4 604
0.2 ~
O~ I J I I :500 4 0 0 5 0 0 6 0 0 7 0 0 8 0 0
WAVELENGTH (nm)
L_ 90O
Fig. 7. Absorption spectra of unit-membrane vesicles and BChl a-protein from C. lirnicola f. thio- sulfatophilum. A. Complex I in 10 mM phosphate (pH 6.8) and 10 mM ascorbate. B. Complex II in 2 M guanidine hydrochloride and 10 mM phosphate. (The two preparations were made indepen- dently.) C. BChl a-protein in 2 M guanidine hydrochloride and 10 mM phosphate. Adapted with per- mission of Brookhaven National Laboratory from Olson et al. [21 ].
6 ' I '678 [ ~ I ' I ' ' I ' I ' 7 I ° A , 5
~"6k l ' 6,0 -- / / -,L ~ ¢ / • ~ 9 553
'
- 1 8 ~ - , I I I ' 1 8 3 0 J , I , f ~ , "~ '~ 600 700 8 0 0 900 I000 I100 1200
WAVELENGTH (rim)
I I 1
T 818
832 I 8141 I
800 820 8 4 0
Fig. 8. Light-induced absorbance changes in green sulfur bacteria. A. Solid curve: Complex I (,4 a I 0 = 2.3) of C. limicola f. thiosul fatophilum in 10 mM ascorbate. Taken with permission from Olson et al. [21]. Dashed curve: Complex I of P. aestuarii. Adapted with permission from Swarthoff and Amesz [3]. B. Intact cells of P, aestuarii. Adapted with permission from Sybesma and Vredenberg [35]. C. Complex I of C limicola f. thiosul fatophilum at 5 K. Adapted with permission of Photochem. Photo- biol. from Whitten et al. [15].
40
absorption spectra of the Chlorobium BChl a-protein (Fig. 7C) and the Chlorobium vesi- cles before and after guanidine hydrochloride treatment (Fig. 7A and B) show that the shoulder at 835 nm in the vesicle spectra is missing from the BChl a-protein spectrum. This shoulder is ascribed to the reaction centers known to be present in the membrane preparations and in the 600- and 350-kdalton particles from Prosthecochloris.
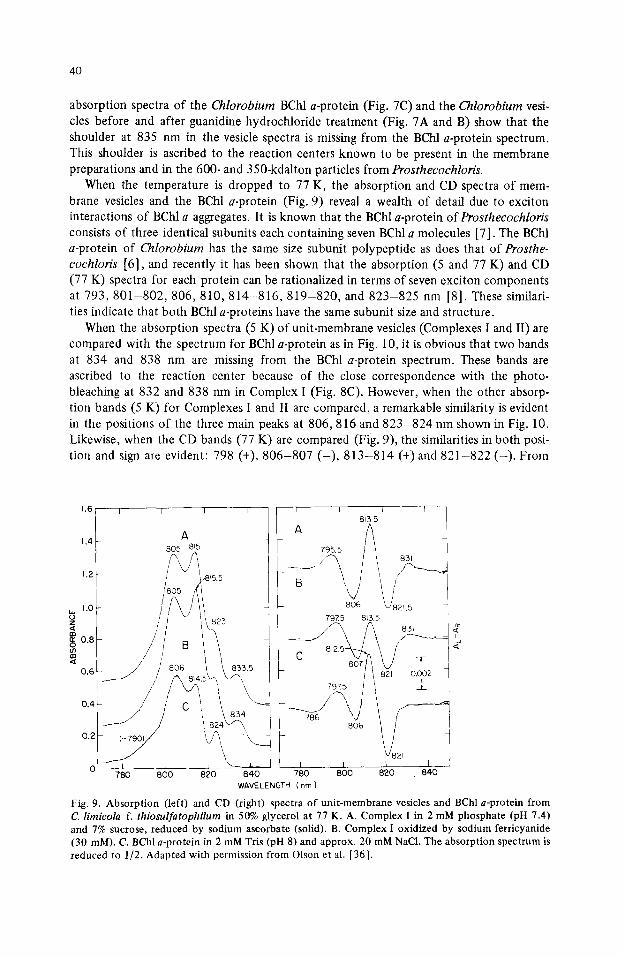
When the temperature is dropped to 77 K, the absorption and CD spectra of mem- brane vesicles and the BChl a-protein (Fig. 9) reveal a wealth of detail due to exciton interactions of BChl a aggregates. It is known that the BChl a-protein of Prosthecochloris consists of three identical subunits each containing seven BChl a molecules [7]. The BChl a-protein of Chlorobium has the same size subunit polypeptide as does that of Prosthe- cochloris [6], and recently it has been shown that the absorption (5 and 77 K) and CD (77 K) spectra for each protein can be rationalized in terms of seven exciton components at 7 9 3 , 8 0 1 - 8 0 2 , 8 0 6 , 8 1 0 , 8 1 4 - 8 1 6 , 8 1 9 - 8 2 0 , and 823-825 nm [8]. These similari- ties indicate that both BChl a-proteins have the same subunit size and structure.
When the absorption spectra (5 K) of unit-membrane vesicles (Complexes I and II) are compared with the spectrum for BChl a-protein as in Fig. 10, it is obvious that two bands at 834 and 838 nm are missing from the BChl a-protein spectrum. These bands are ascribed to the reaction center because of the close correspondence with the photo- bleaching at 832 and 838 nm in Complex I (Fig. 8C). However, when the other absorp- tion bands (5 K) for Complexes I and II are compared, a remarkable similarity is evident in the positions of the three main peaks at 806,816 and 823-824 nm shown in Fig. 10. Likewise, when the CD bands (77 K) are compared (Fig. 9), the similarities in both posi- tion and sign are evident: 798 (+), 806-807 ( - ) , 813-814 (+) and 821-822 ( - ) . From
1.6 I
1.4
1.2
w 1.0 o z
0.8 o m en
0.6
0.4
I I I
0.2
A 8O5 8q5
/ / 806 \\ 833.5
780 800 820 840
813.5
806 L/821 5 79X5 8~5.5 [
A 83, | e
/ / 8 2 ' o oo~ I
, , ys , , I
WAVELENGTH (nm)
Fig. 9. Absorption (left) and CD (right) spectra of unit-membrane vesicles and BChl a-protein from C. limicola f. thiosulfatophilum in 50% glycerol at 77 K. A. Complex I in 2 mM phosphate (pH 7.4) and 7% sucrose, reduced by sodium ascorbate (solid). B. Complex I oxidized by sodium ferricyanide (30 mM). C. BChl a-protein in 2 mM Tris (pH 8) and approx. 20 mM NaC1. The absorption spectrum is reduced to 1/2. Adapted with permission from Olson et al. [36].

41
I I T I I I 0 1 I [ I I l I I I I I
A ~9~ B " '~
i t
,, , ~_ f I I / '~ / /
J / \/i ~- / ',, ,~ ~
770 7 9 0 8 1 0 8 3 0 8 5 0 7 7 0 7 9 0 8 1 0 8 3 0
W A V E L E N G T H ( n m }
I ~ I ] 1 I I tol I P I
rO ¢,J
(,D O0 0 ~o t~
8 5 0 7 7 0 7 9 0 8 1 0 8 3 0 8 5 0
Fig. 10. Absorption spectra (lower curves) and fourth derivative (upper) curves of unit-membrane vesi- cles and BChl a-protein from C. limicola f. thiosulfatophilum in 50% glycerol at 5 K. A. Complex I. B. Complex II. C. BChl a-protein. Adapted with permission of Photochem. Photobiol. from Whitten et al. [151.
these similarities in absorption and CD, Whitten et al. [15] concluded that the exciton interactions in the membrane-bound BChl a in Complexes I and II are probably quite sim- ilar to those in the water-soluble BChl a-protein. The membrane-bound BChl a molecules may well be organized in groups of seven inside protein subunits similar in configuration to the subunits of the water-soluble BChl a-protein.
IIIB. Reaction centers
The light-induced bleaching of P-840 (Fig. 8) in green sulfur bacteria, in unit-mem- brane vesicles (Complex I), or in 350- and 600-kdalton Triton particles is quite different from the bleaching ofP-865 (Fig. 4) in Chloroflexus [9] which is more like that o f P-870 and P-960 in purple bacteria. Two narrow troughs (half maximum width about 5 nm) appear at 830 and 842 nm (about 300 K) or at 832 and 838 nm (5 K). (These wavelength shifts with temperature ( - 2 nm and +4 nm respectively) are small compared to the shifts o f approx. - 2 0 nm observed in reaction centers of purple bacteria [3,20].) Also, there appears to be a red shift of an 814-nm band to 818 nm (5 K). In Chloroflexus and the purple bacteria a single broad band (half maximum width about 25 nm) at 8 6 0 - 8 9 0 nm (960 nm in BChl b organisms) is found. In contrast to the peculiarities in the optical properties of the reaction centers o f green sulfur bacteria, the light-induced EPR signal corresponding to oxidized P-840 is typical for many photosynthetic bacteria: g = 2.003, z3J/= 9.2 + 0.5 G [3,21]. Although P-840 may be a BChla dimer or special pair, as are P-870 and P-960, further study by electron nuclear double resonance is required to prove the point.

42
There are at least two possible interpretations of the two peaks in the absorption spec- trum of P-840 [5]. Each reaction center may have two peaks as a result of exciton inter- action between two or more BChl a molecules. In this interpretation, the small separation (12 nm at 300 K and 6 nm at 5 K) of the peaks would reflect a weak interaction between the BChl a molecules. Alternatively there might be two very similar types of reaction cen-
ter, one with P-830 and the other with P-842. (At 5 K each reaction center would have a band at 816 nm in addition to a single photobleachable band at either 832 or 838 nm.)
Further experiments are needed to show whether the bands at 830 and 842 nm (300 K)
can change independently of each other.
IIIC. Bacteriochlorophyll a-proteins
Figure 11 shows the subunit structure [17] of the BChl a-protein from P. aestuarii. Seven BChl a molecules with an average center-to-center distance of 1.2 nm are wrapped
C
N
Fig. 11. Subunit of the BChl a-protein from P. aestuarii. This diagram shows the arrangement of the polypeptide backbone and the BChl a core of one subunit. Circles denote a-carbon positions. Some connections within the chain are uncertain, and particularly ambiguous regions are indicated by broken lines. For clarity, magnesium atoms, BChl ring substituents, and phytyl chains are omitted. The direction of view is from the center of the trimer toward the exterior of the molecule. The three- fold axis of the trimer is horizontal in the plane of the figure. Reproduced with permission of J. Mol. Biol. from Matthews et al. [ 17 ].

43
in a protein cocoon with the seven phytyl tails (not shown) forming the inner core. The seven porphine rings, although not parallel, lie close to an average plane. All Mg atoms are five-coordinated. Histidine residues serve as ligands to the Mg atoms (not shown) in BChls 1 ,3 , 4, 6 and 7. A carbonyl oxygen is the ligand to BChl 5, and a water molecule is the ligand to BChl 2. Each BChl a molecule is anchored to the protein through extensive H-bonding, liganding to Mg, and hydrophobic interactions through the phytyl tail. This basic arrangement is thought to apply to the firmly bound BChl a in membranes (i.e., that BChl a not solubilized by guanidine HC1) as well as to the water-soluble BChl a-protein in both P. aestuarii and C. limicola.
The Prosthecochloris BChl a-protein exists as a trimer (Fig. 12) in solution [6] and in crystals [7,22], and the Chlorobium protein is assumed to be the same. In the trimer the BChl a aggregates in each subunit form a triangular funnel around the three-fold axis. The closest distance between BChl a molecules in adjacent subunits is 2.4 nm compared with 1.2 nm within one subunit. Prosthecochloris BChl a-protein crystallizes in two modes, hexagonal (P63) and trigonal (P31) [3]. In trigonal crystals the trimers are close packed, and the closest distance between BChl a molecules in adjacent trimers is about 3 nm.
Fig. 12. BChl a-protein trimer with BChl a molecules omitted. The polypeptide backbone is projected down the trimer axis (central triangle). Reproduced with permission of J. Mol. Biol. from Matthews et al. [17].

44
In a recent attempt to understand theoretically the low temperature absorption and CD spectra in the region of the 809-nm band, Pearlstein and Hemenger [23] found it im- possible to fit the experimental data with the conventional assignment of the lowest sin- glet transition of BChl a as y-polarized. This has led some to doubt the adequacy of present-day exciton theory.
IV. Chlorosome (Chlorobiaceae)
IVA. Baseplate structure
Staehelin et al. [ 12] in a beautiful freeze-fracture study have shown that between each chlorosome and the cytoplasmic membrane in C. limicola f. thiosulfatophilum there is a baseplate, 5 - 6 nm thick, which appears to be a two.dimensional crystalline array of BChl a-protein (Fig. 13). The electron micrographs show a 6-nm spacing between parallel ridges, which lie at an angle between 40 ° and 60 ° with the long axis of the chlorosome.
With the assumption that the Chlorobium BChl a-protein can crystallize in essentially the same trigonal mode [22] as does the Prosthecochloris BChl a-protein, it is possible to understand the baseplate as a two-dimensional crystal in the trigonal space group P31 as shown in Fig. 14. In the baseplate, the 31 screw axis makes an angle of 25 ° with the plane of the cytoplasmic membrane as shown in the side view (Fig. 14). The total thickness of the baseplate according to this interpretation is 8.7 nm, o f which 3 - 4 nm is embedded in the membrane. The spacing (normal to the membrane) between rows of trimers is 6.0 nm.
rod elements (~lOnm in
o,,o°,o ..... .o.,.° envelope layer (qalactolipid m o n o . f ' ~ ~ ~ ~C" ~ fj~N, ~ ,jJl~ ~ ~ ~/ /
iayerP)
c~,t (hpld bllayer ~5 nm) crystalline Ixiseplate
large intramembrane particles (periodicity ~ 6nm; Bchl o-protein?) (mostly 10-14nm in diometer~ reaction center - non -crystalline LH-Bchl o complexes?)
Fig. 13. Model of a chlorosome and its associated cytoplasmic membrane based on freeze-fracture studies of C. limicola f. thiosulfatophilum. The hydrophobic core of the chlorosome contains 10-30 rod elements surrounded by matrix material. The 2-4 nm wide envelope layer adjacent to the cyto- plasm is thought to consist of a monolayer of galactolipid molecules. A crystalline base plate, 5-6 nm thick, connects the chlorosome core to about 30 large intramembrane particles inside the cytoplas- mic membrane. See text for details. Reproduced with permission from Staehelin et al. [ 12].

8.7 n m
PERSPECTIVE VIEW 65 ~ ~- 6.0 nm-~
SIDE VIEW
45
X/ Fig. 14. Proposed arrangement of BChl a-protein trimers in the baseplate of C. limicola f. thiosulfato. philum. Perspective view: Each subunit is represented by a 4.8-nm sphere, and each trimer is repre- sented by three spheres connected by an overlying 'triangle' with vertices at the poles of the spheres. The trimers are arranged in a two-dimensional crystal (trigonal P31) in which subunit d touches sub- unit c and subunit f touches subunit h. Side view: The 31 screw axis of the crystal makes an angle of 25 ° with the plane of the cytoplasmic membrane (8.0 nm thick). A reaction-center complex in the membrane is represented in cross section by a 4.8-rim reaction-center sphere (RC) and two 4.8-nm BChl a-containing subunits (B). Top view: The uppermost subunits (stippled) form channels in the top surface of the baseplate. The channels spaced 6.8 nm apart run at an angle of about 55 ° to the chan- nels spaced 6.0 nm apart. The rod elements in the chlorosome may be aligned parallel to the 6.8-nm channels.
, k ,
The trimers are spaced 8.3 nm apart in each row as shown in the top view (Fig. 14). In
the upper surfaces of the baseplates there are grooves at about 55 ° to the direction of the rows of trimers. These grooves are formed by the uppermost subunit in each trimer (sub- units a and d i nF ig . 14). If the overlying rod elements (see Fig. 13) tend to line up with the grooves in the baseplate, the long axis of each chlorosome would be expected to lie about 55 ° away from the rows of trimers in the baseplate. Since the forces holding the trimers together in the baseplate are the same as those holding crystals together, the base- plate must be a self-assembling structure.
IVB. R o d elements
Staehelin et al. [12] showed the existence of 1 0 - 3 0 rod elements approx. 10 nm in diameter inside each chlorosome in C. limicola f. thiosulfatophilum (Fig, 13). Previously Cruden and Stanier [1,24] had noted in several chlorosomes 10-25 ring-like structures, 9 - 1 0 nm wide with a small central hole about 3 nm in diameter. In glutaraldehydeofixed, sections the ring-like structures appear as five or six subunits arranged around the central hole and remain intact after the extraction of BChl with methanol. This resistance to methanol extraction suggests that the ring-like structures may be proteinaceous. The

46
/
Fig. 15. Proposed model of a rod element in C. limicola f. thiosulfatophilurn. Protein subunits (15 kdalton), represented by 3.3-nm spheres, are arranged in rings of six with 3.3-nm holes in the centers. Rod elements are composed of close-packed rings spaced 2.9 nm apart. Each subunit is associated with 12-14 BChl c molecules (not shown). The BChl c in a highly aggregated, quasi-crystalline state occu- pies the central hole in the rod element and the spaces on the outer surface without increasing the overall diameter beyond 10 rim.
model shown in Fig. 15 combines a large number of close-packed ring-like structures (six subunits each) into a single rod element. For simplicity each subunit is represented as a sphere, 3.3 nm in diameter with an estimated weight of 15 kdalton on the assumption that the subunits are proteins.
The chlorosomes observed by Staehlin et al. [12] ranged in size from 40 × 70 nm to 100 × 260 nm and contained between 10 and 30 rod elements. From the chromosome model shown in Fig. 13, it appears that the width of the baseplate is about 10 nm less than the width of the chlorosome. The ratio of number of rod elements to width of base- plate is therefore about 1 per 3 nm for both small and large chlorosomes. Since each trimer in the baseplate occupies 50 nm 2, the number of trimers in a baseplate 100 × 30 nm is 60 (1260 BChl a) and the number o f rod elements is ten. According to the model in Fig. 15, each rod element (assumed to be 100 nm long)should contain 35 rings and 210 subunits. The total number of subunits in ten rod elements would be 2100, and the total number of BChl a molecules both in the baseplate and embedded in the membrane would be about 2500. Since the ratio o f BChl c to BChl a is known to be 10-20 , the number of BChl c molecules associated with each subunit should be 12-24 . If each subunit is a pro- tein, and the BChl c exists in a highly aggregated form (individual molecules 0 .2-0 .3 nm apart), the BChl c can be accommodated in the core and around the outside of each rod element. The volume occupied by the BChl c is estimated by assuming that the volume occupied by one BChl c molecule is 1.4 nm 3, the volume occupied by one chlorophyll a molecule in a crystal [25]. Since the volume of a subunit is about 19 nm 3 and that of a ring 114 nm 3, the total volume of one ring and its associated BChl c ( 1 2 - 2 4 molecules per subunit) would be 2 1 5 - 3 1 6 nm 3. In the model (Fig. 15) the rings are close-packed 2.9 nm apart. Each ring with its associated BChl c may be considered to be a 2.9-nm thick circular section of the rod element. The diameter of such a circular section (and the entire rod element) containing 12 BChl c molecules per subunit would be x/(4/rr)(215/2.9) = 9.7 rim. For 14 and 24 molecules per subunit, the calculated diameters are 10.1 and 11.8 nm, respectively. 1 2 - 1 4 BChl c molecules per subunit seems to be a reasonable estimate con- sistent with the data, the assumption of quasi-crystalline BChl c, and the subunit model

47
TABLE 1
AMINO ACID COMPOSITION OF CHLOROSOME PROTEIN [26] AND BChl a-PROTEIN [16] FROM C. LIMICOLA F. THIOSULFATOPHILUM STRAIN TASSAJARA
Residues Chlorosome protein BChl a-protein Difference (mol%) (tool%) (%)
Polar 41.7 45.5 -8 Acidic 20.2 19.6 +3
Asp 12.5 11.1 +13 Glu 7.7 8.5 -9
Basic 9.0 13.5 -33 Lys 4.1 5.1 -20 His 1.2 2.1 -43 Arg 3.7 6.3 -41
Hydroxyl 12.5 12.4 +1 Thr 5.6 3.3 +70 Ser 6.9 9.1 -24
Nonpolar 58.2 54.5 +7 Aliphatic 50.1 45.4 + 10
Gly 10. l 10.5 -4 Ala 13.0 6.6 +97 Val 7.8 10.7 -27 lie 5.7 4.0 +42 Leu 7.9 5.9 +34 Pro 4.4 5.9 -25 Met 1.1 1.2 +8 Cys 0.1 * 0.6 -
Aromatic 8.1 9.1 - 1 1 Tyr 2.3 2.8 -18 Phe 4.2 5.0 -16 Trp. 1.6 1.3 +23
Total 99.9 100.0
* Not treated with perchloric acid.
for the rod element. The calculated protein to BChl c ratio is 1.3 -+ 0.1, which is consis- tent with the experimental values o f 1.2 [24] and 1.7 [11] found for whole chlorosomes from C. limicola f. thiosulfatophilum. However, the earlier value of 0.5 [6] cannot easily be reconciled With the model or with the more recent experimental values.
The amino acid composit ion of the chlorosome protein [26] is compared with that o f the water-soluble BChl a-protein [fi] in Table I. The major differences are the higher con- tents o f threonine and alanine in the chlorosome protein. The content of non-polar amino acids residues is 7% higher in the chlorosome protein, but it is surprising that the differ- ence is not greater in view of the more hydrophobic environment of the chlorosome pro- tein and the known aqueous solubility of the BChl a-protein. Since the ratio of protein to BChl c in a specific complex is probably less than 1.7, the number of histidine residues per BChl c would be less than 0.35. This eliminates the possibility that a major way of binding BChl c to the protein is via histidine liganding to Mg atoms. (In the BChl a-pro- tein from Prosthecochloris, five of the seven BChl a molecules in each subunit are liganded to histidine residues.)
Bystrova et al. [7] have suggested that highly aggregated forms of BChl c could be due

48
to the formation of intermolecular bonds in which the keto group of ring V (Fig. 2) on one BChl c serves as a ligand to the Mg of another BChl c.
Since chlorosomes are rich in glycolipids [11,28] and phospholipids [11], it seems probable that both are necessary for the organization of BChl c and/or stabilization of the rod elements.
V. Reaction-center complexes (Chlorobiaceae)
The 600- and 350-kdalton particles from P. aestuarii contain about 75 and 35 BChl a molecules per reaction center, respectively [3]. These particles contain approx. 42- and 45-kdalton polypeptides (SDS-polyacrylamide gel electrophoresis) which together account for most of the protein (Swarthoff, T., personal communication). The 42-kdal- ton polypeptide, which cannot be distinguished from the denatured subunit of the water- soluble BChl a-protein [6], is present in a higher proportion in the 600-kdalton particle than in the 350-kdalton particle. Since the water-soluble BChl a-protein is released from the 600-kdalton particle upon treatment with guanidine hydrochloride, it seems clear that two trimers (42 BChl a molecules and 280 kdalton) are removed to yield the 350-kdalton particle. Most of the remaining BChl a molecules in the 350-kdalton particle are thought to be associated in aggregates of seven with 42-kdalton polypeptides to form five approx. 50-kdalton subunits similar in size to the subunits of the water-soluble BChl a-protein. The 35 BChla molcules in the 350-kdalton particle correspond to the tightly bound BChl a in the cytoplasmic membrane, i.e. that BHcl a not solubitized by guanidine hy- drochloride. The reaction center with its associated carotenoids would then account for the remaining 100-kdalton mass of the particle. The 45-kdalton polypeptide may be asso- ciated with the reaction-center part of the particles from Prosthecoehloris. In Chlorobium unit-membrane vesicles [3] 21- and 3 l-kdalton polypeptides may be associated with the reaction centers, and in Chloroflexus membranes [6] three polypeptides, 24, 27 and 29 kdalton, may be associated with the reaction center as in purple bacteria.
VA. Intramembrane particles
Staehelin et al. [ 12] observed in Chlorobium that the cytoplasmic membrane contains between 20 and 45 intramembrane particles (diameter 13-16 n m ) a t each chlorosome attachment site, and they suggested that these particles may correspond to reaction- center complexes containing BChl a not associated with the water-soluble BChl a-protein of the baseplate. If the large particles in Chlorobium are assumed to be equivalent to the 350-kdalton particles from Prosthecochloris, then the reaction-center subunits (RC) and the BChl a-containing subunits (B) of each reaction-center complex may be represented as 4.8-nm spheres arranged as shown in the side view of Fig. 14. The diameter of such a complex, viewed normal to the membrane, would appear to be between 12 and 15 nm.
The number of reaction centers for a given area of baseplate can be estimated. For a small chlorosome the baseplate is approx. 2100 nm 2 (30 × 70) and for a large chloro- some, approx. 23 000 nm 2 (90 × 260). This suggests that the baseplate area per reaction center might be as low as 100 nm 2 or as high as 500 nm 2. These two area values corre- spond to 2 and 10 BChl a-protein trimers, respectively, for each reaction-center particle. The photosynthetic unit for a small chlorosome-baseplate-membrane combination would then include at least 37 BChl a molecules in the reaction-center particle, 42 BChl a mole- cules in the two trimers, and 900 +- 100 BChl c molecules in 70 rod-element subunits. For

49
a large chlorosome combination the photosynthetic unit would include at least 37 BChl a molecules in the reaction-center particle, 210 BChla molecules in the ten trimers, and 4500 -+ 400 BChl c molecules in 350 rod-element subunits. It appears that the ratio of BChl c to BChl a (baseplate) is kept approximately constant, but that the ratio of BChl a (baseplate) plus BChl c (rod elements) to reaction-center complexes increases as chloro- some size increases. This means that the ratio BChl c : BChl a : reaction center can vary roughly from 900 : 80 : 1 (small chlorosome) to 4500 : 250 : 1 (large chlorosome).
In the baseplate of a small chlorosome there are two BChl a-protein trimers for each reaction-center complex, the same as in Chlorobium unit-membrane vesicles (Complex I) and in Prothecochloris 600-kdalton Triton particles. Since no baseplates are visible in freeze-fracture micrographs (Fig. 6) of Complex I, the baseplates appear to be destroyed along with the chlorosomes during cell breakage. The BChl a-protein trimers associated with reaction-center complexes in the membrane appear to remain attached to the unit- membrane vesicles (Complex I) and to the 600-kdalton Triton particles. The other BChl a-protein trinaers of the baseplate apparently are removed from the membrane during cell breakage. Guanidine hydrochloride seems to be required to remove the two trimers bound to the reaction-center complex.
VI. Chloroflexacea
Although reaction-center BChl a is associated with the cytoplasmic membrane in all green bacteria [3,28], the gliding filamentous bacterium Cf. aurantiacus differs from the green sulfur bacteria in several respects with regard to chlorophyll organization. In addi- tion to the reaction-center BChl a peaks at 805 and 865 nm in Chloroflexis there is another small peak at 790 nm [6,11,28]. The 790-nm BChl a is associated with the chlo- rosome baseplate and remains with the chlorosome when it is separated from the cyto- plasmic membrane [11]. The association between the BChl c of the chlorosome and the BChl a of the baseplate is so close that light absorbed by BChl c is emitted as fluorescence by BChl a (807 nm) as well as by BChl c (755 nm) in the ratio of 1.5 : 1 (Sprague, S.G., personal communciation). (The 810-nm BChl a-protein in the baseplate of sulfur bacteria remains with the cytoplasmic membrane when the chlorosomes are removed.) The base- plate in Chloroflexus appears only about 2 nm thick and shows ridges at 90 ° to the chlo- rosome axis [28,29], whereas the baseplate in sulfur bacteria appears to be 5 - 6 nm thick with striations at 4 0 - 6 0 ° to the chlorosome axis [12].
Gregg [28] found by SDS-polyacrylamide gel electrophoresis that purified ctvlorosomes (including baseplates) from strain J-lO-fl contained four major polypeptides (12, 14, 28 and 60 kdalton). More recently, Schmidt et al. [6] found 10-, 15-, 40- and 57-kdalton polypeptides in chlorosomes from strain OK-70-fl. The four peptides of Gregg are prob- ably the same as the peptides of Schmidt et al., even though the discrepancy between 28 and 40 kdalton is rather large. The 14-15-kdalton peptide accounts for most of the total protein and is considered to be a good candidate for the protein part of a specific BChl c-protein complex [28]. The 28 to 40-kdalton polypeptide might be associated with the 790-nm BChl a-in the baseplate [6]. Since no 790-rim BChl a complex has yet been iso- lated from Chloroflexus, it is not possible to say whether the 790-nm BChl a is really asso- ciated with protein in a subunit or trimer analogous to the subunits or trimers of BChl a-protein in green sulfur bacteria.
The rod elements in the chlorosomes of Chloroflexus are about 5.2 nm in diameter with a periodicity in the long direction of approx. 6 nm [29]. It is difficult to imagine

50
with the available information a subunit for the Chloroflexus rod element which is similar to the subunit proposed for the rod elements of Chlorobium and Prosthecochloris.
VII. Summary
Light-harvesting BChl c, d or e is thought to be located inside the rod elements which fill the chlorosome appressed to the inside of the cytoplasmic membrane of green photo- synthetic bacteria. BChl a is known to be part of a BChl a-protein which forms a crystal- line baseplate between the rod elements in the chlorosome and the inside of the cyto- plasmic membrane. Reaction-center complexes are most probably buried under the base- plate inside the membrane. Energy transfer is from BChl c, d or e in the rod elements to BChl a in the baseplate and then to BChl a in the reaction-center complexes.
The rod elements in green sulfur bacteria are thought to be composed of approx. 15- kdalton protein subunits, each associated with 12-14 BChl c, d or e molecules. Six sub- units would be required to form a lO-nm ring, and about 35 rings would be necessary to form a 100-nm rod element. The baseplate appears to be a two-dimensional crystal (tri- gonal space group P31) of BChl a-pr0tein trimers with the 31 screw axis tilted 25 ° out of the plane of the membrane. The reaction-center complex is thought to be made up of a 100-kdalton carotenoid reaction-center core and five 50-kdalton subunits, each contain- ing seven BChl a molecules. Each reaction-center complex is apparently linked directly to two BChl a-protein trimers in the baseplate.
The reaction centers in green sulfur bacteria may be of one type (containing P-840), or of two types (containing P-830 or P-842). In filamentous gliding bacteria the reaction centers appear to contain only P-865. The number of BChl a molecules in these reaction centers is not known, but is assumed to be at least two.
Note added in proof (Received October 13th, 1980)
Upon further reflection, the author believes that it is impossible to decide whether the unit-membrane vesicles of Chlorobium are inside4n or inside~ut, based on the freeze- fracture electron micrograph shown in Fig. 6. Independent evidence for inside-out vesicles is the fact that guanidine hydrochloride liberates BChl a-protein molecules from Complex I into the medium.
From linear dichroism measurements of intact cells, Complex I, 600-kdalton particles, 350-kdalton particles, and BChl a-protein molecules from P. aestuarii, Swarthoff et al. [14] have concluded that on the average the 3-fold axes of the BChl a-protein trimers in the cytoplasmic membrane of intact cells make angles between 55 ° and 90 ° with the mem- brane instead of 25 ° as shown in Fig. 14 (side view). This conclusion is clearly inconsis- tent with the baseplate arrangement proposed in the present paper.
Evidence for energy transfer from BPh c to BChl a in the reaction center is found in the fluorescence emission spectra of Complex I and BChl a-protein from P. aestuarii [37]. This energy transfer suggests that one or more BPh c molecules are close to or part of the reaction center pigment complex.
Acknowledgements
I thank Ms. Sallie Sprague and Prof. R. Clinton Fuller (University of Massachusetts) for permission to use the unpublished absorption spectrum of Chloroflexus membranes in

51
Fig. 4A. 1 am also grateful to Dr. Thomas Giddings, Jr., and Prof. Andrew Staehelin (Uni-
versity of Colorado) for permission to use the unpublished electron micrograph shown in Fig. 6. Dr. Jan Amesz (Leiden) and Dr. Karin Schmidt (G6ttingen) kindly sent me copies of their papers [6,14] prior to publication. Several colleagues (J. Amesz, M. Dienes, B.W.
Matthews, R.M. Pearlstein, S.G. Sprague, L.A. Staehelin, J.P. Thornber, and W.B. Whir- ten) made helpful comments on the first draft of this paper.
References
1 Pierson, B.K. and Castenholz, R.W. (1978) in The Photosynthetic Bacteria (Clayton, R.K. and Sis- trom, W.R., eds.), pp. 179-197, Plenum Press, New York
2 Fowler, C.E., Nugent, N.A. and Fuller, R.C. (1971) Proc. Natl. Acad. Sci. U.S.A. 68, 2278-2282 30lson, J.M. and Thornber, J.P. (1979) in Membrane Proteins in Energy Transduction (Capaldi,
R.A., ed.), pp. 279-340, Marcel Dekker, New York 4 Broch-Due, M., Ormerod, J.G. and Fjerdingen, B.S. (1978) Arch. Microbiol. 116,269-274 5 Holt, S.C., Conti, S.F. and Fuller, R.C. (1966) J. Bacteriol. 91,349-355 6 Schmidt, K., Maarzahl, M. and Mayer, F. (1980) Arch. Microbiol. 127, 87-98 7 Katz, J.J., Strain, H.H., Harkness, A.L., Studier, M.H., Svec, W.A., Janson, T.R. and Cope, B.T.
(1972) J. Am. Chem. Soc. 94, 7938-7939 8 Kiinzler, A. and Pfennig, N. (1973) Arch. Mikrobiol. 91, 83-86 9 Gloe, A. and Pfennig, N. (1974) Arch. Microbiol. 96, 93-101
10 Gloe, A. and Risch, N. (1978) Arch. Microbiol. 118, 153-156 11 Schmidt, K. (1980) Arch. Microbiol. 124, 21-31 12 Staehlin, L.A., Golecki, J.R. and Drews, G. (1980) Biochim. Biopbys. Acta 589, 30-45 13 Swarthoff, T. and Amesz, J. (1979) Biochim. Biophys. Acta 548,427-432 14 Swarthoff, T., de Grooth, B.G., Meiburg, R.F., Rijgersberg, C.P. and Amesz, J. (1980) Biochim.
Biophys. Acta 593, 51-59 15 Whitten, W.B., Pearlstein, R.M. and Olson, J.M. (1979) Photochem. Photobiol. 29,823-828 16 Olson, J.M. (1978) in The Photosynthetic Bacteria (Clayton, R.K. and Sistrom, W.R., eds.), pp.
161-197, Plenum Press, New York 17 Matthews, B.W., Fenna, R.E., Bolognesi, M.C., Schmid, M.F. and Olson, J.M. (1979)J. Mol. Biol.
131,259-285 18 Whitten, W.B., Olson, J.M. and Pearlstein, R.M. (1980) Biochim. Biophys. Acta 591,203-207 19 Pierson, B.K. and Castenholz, R.W. (1974) Arch. Microbiol. 100, 283-305 20 Clayton, R.K. and Yamamoto, T. (1976) Photochem. Photobiol. 24, 67-70 21 Olson, J.M., Prince, R.C. and Brune, D.C. (1977) Brookhaven Symp. Biol. 28,238-44 22 Fenna, R.E., Matthews, B.W., Olson, J.M. and Shaw, E.K. (1974) J. Mol. Biol. 84,231-240 23 Pearlstein, R.M. and Hemenger, R.P. (1978) Proc. Natl. Acad. Sci. U.S.A. 75, 4920-4924 24 Cruden, D.L. and Stanier, R.Y. (1970) Arch. Mikrobiol. 72, 115-134 25 Donnay, G. (1959) Arch. Biochem. Biophys. 80, 80-85 26 Schmitz, R. (1967) Arch. Mikrobiol. 56,238-247 27 Bystrova, M.I., Mal'gosheva, I.N. and Krasnovskii, A.A. (1979)Mol. Biol. 13,440-451 (translated
from Molekulyarnaya Biologiya 13,582-594) 28 Gregg, T.I. (1978) Ph.D. Thesis, University of Massachusetts, Amherst 29 Staehelin, L.A., Golecki, J.R., Fuller, R.C. and Drews, G. (1978) Arch. Microbiol. 119, 269-277 30 Risch, N., Brockmann, H., Jr. and Gloe, A. (1979) Liebigs Ann. Chem. 1979, 408-418 31 Jones, O.T.G. (1978) in The Photosynthetic Bacteria (Clayton, R.K. and Sistrom, W.R., eds.), pp.
751-777, Plenum Press, New York 32 Caple, M.B., Chow, H. and Strouse, C.E. (1978) J. Biol. Chem. 253, 6730-6737 33 Smith, K.M., Bushell, M.J., Rimmer, J.. and Unsworth, J.F. (1980) J. Am. Chem. Soc. 102, 2437-
2448 34 Brockmann, H., Jr. (1976) Phil. Trans. R. Soc. Lond. B. 273,277-285 35 Sybesma, C. and Vredenberg, W.J. (1963) Biochim. Biophys. Acta 75,439-441 36 Olson, J.M., Ke, B. and Thompson, K.H. (1976) Biochim. Biophys. Acta 430, 524-537 37 Karapetyan, N.V., Swarthoff, T., Rijgersberg, C.P. and Amesz, J. (1980) Biochim. Biophys. Acta
593,254-260