chimney building by male uca formosensis rathbun, … ratio of u. formosensispresent on the surface...
TRANSCRIPT

242
Zoological Studies 44(2): 242-251 (2005)
242
* To whom correspondence and reprint requests should be addressed.
Some crabs (and other crustaceans) buildstructures (pillars, hoods, chimneys, etc.) next totheir burrows (Powers and Bliss 1983). Some ofthese structures have been shown to function forsexual attraction, e.g., pillars (Uca: Christy 1988ab), hoods (Uca: Zucker 1974 1981, Christy et al.2002 2003), mudballs (Uca: Oliveira et al. 1998),and pyramids (Ocypode: Linsenmair 1967, Hughes1973); some are related to disturbance (fighting orburrow occupation) caused by neighbors, e.g.,hoods (Uca: Zucker 1974 and Cleistostoma:Clayton 1988), chimneys (Uca: Salmon 1987,Wada and Murata 2000), and barricades (Ilyoplax:Wada 1984); and some are related to environmen-tal regulation, e.g., hoods (Uca: Powers and Cole1976), chimneys (Uca: Crane 1975: 524, Thurman
1984), and igloos (Dotilla: Takeda et al. 1996).Crane (1975) reviewed these structures in fiddlercrabs of the world, and chimneys were found insome subgenera: Deltuca (U. dussumieri, U. arcu-ata, U. forcipata, U. coarctata, and U. urvillei),Thalassuca (U. tetragonon), Amphiuca (U.chlorophthalmus), Boboruca (U. thayeri), Minuca(U. vocator), and Celuca (U. cumulanta and U.stenodactylus). Other fiddler species were alsoreported to build chimneys, e.g., U. elegans of thesubgenus Australuca (George and Jones 1982)and U. subcylindrica of the subgenus Minuca(Thurman 1984) (reviewed in Table 1).
Uca formosensis Rathbun, 1921 is a speciesof fiddler crab endemic to Taiwan. This species ispoorly known except for its morphological charac-
Chimney Building by Male Uca formosensis Rathbun, 1921 (Crustacea:Decapoda: Ocypodidae) after Pairing: A New Hypothesis for ChimneyFunctionHsi-Te Shih1,2,*, Hin-Kiu Mok2, and Hsueh-Wen Chang3
1Department of Life Science, National Chung-Hsing University, Taichung, Taiwan 402, R.O.C.Tel/Fax: 886-4-22856496. E-mail: [email protected]
2Institute of Marine Biology, National Sun Yat-sen University, Kaohsiung, Taiwan 804, R.O.C.3Department of Biological Sciences, National Sun Yat-sen University, Kaohsiung, Taiwan 804, R.O.C.
(Accepted February 8, 2005)
Hsi-Te Shih, Hin-Kiu Mok, and Hsueh-Wen Chang (2005) Chimney building by male Uca formosensisRathbun, 1921 (Crustacea: Decapoda: Ocypodidae) after pairing: a new hypothesis for chimney function.Zoological Studies 44(2): 242-251. The construction of a chimney after pairing is first described for Uca for-mosensis Rathbun, 1921, a fiddler crab endemic to Taiwan. After attracting a female into his burrow, the malebuilds a chimney averaging 9.2 cm high and 7.7 cm in diameter at the base within a 4- or 5-d period before thenext neap tide. It is not related to sexual attraction because the chimney does not appear prior to or duringcourtship. To build the chimney, the male excavates the burrow which probably widens the shaft and, moreimportantly, deepens the burrow so that it reaches the water table, thus providing a moist chamber in which thefemale can incubate her eggs. The mudballs dug out from within the burrow are piled around the entrance andform the chimney, which herein is assumed to function as a way for the builder to hide from its enemies. Thesex ratio of U. formosensis present on the surface was also recorded and is discussed. http://www.sinica.edu.tw/zool/zoolstud/44.2/242.pdf
Key words: Fiddler crabs, Behavior, Underground mating, Mudflats, Taiwan.

Shih et al. -- Chimney Building by Uca formosensis 243
ters (see Shih et al. 1999). From preliminaryobservations, many large chimneys were periodi-cally found in the habitat of U. formosensis. Thechimneys of this species have not been describedin detail before (Shih et al. 1999). In this paper, wedescribe results based on successive observationsin Changhua, west-central Taiwan, and on datarecorded on mudflats along the western coast ofTaiwan. A chimney of U. formosensis is built bythe male of a pair after he has attracted a femaleinto his burrow for underground mating, and ittherefore is not a courtship signal. The function ofchimney building is proposed and discussed in thisstudy. The chimneys built by U. arcuata (de Haan,1835) are also recorded for comparison. In addi-tion, we also recorded and discuss the sex ratio ofU. formosensis present on the surface.
MATERIALS AND METHODS
Uca formosensis is distributed on the westcoast of Taiwan, including the Penghu Archipelago(the Pescadores) (Shih et al. 1999). From 7 to 21July 1996, successive observations on chimneybuilding were conducted in Shengang, ChanghuaCounty. Two plots (2 x 2 m) were selected inwhich to monitor the crabs
,behavior. From 08:00
to 18:00, numbers of males and females in eachplot and 3 temperature readings were recordedeach hour, except for the period when the habitatwas covered by water at high tide. The 3 tempera-tures recorded were the ground temperature insunshine (sun temperature), ground temperatureunder the shade of some object (shade tempera-ture), and soil temperature at a depth of about 12cm by inserting the probe of a Digital Thermo(Digital Taschen-Thermometer). The sex ratio,males/females, appearing on these plots was alsocalculated each hour. One hour
,s data in 21 July
are not included as sex ratio was 16:0.The process of chimney building and other
Table 1. A summary of chimneys built around burrow entrances by Uca fiddler crabs
Species Subgenus defined Materials Builder Referencesby Crane (1975)
U. formosensis ?a from within the burrow mostly male and this studyseldom female
U. arcuata Deltuca mostly from the surface both sexes Crane 1975, Yeh 1996,Wada and Murata 2000
U. coarctata Deltuca both sexes Crane 1975U. coarctata flammula Deltuca Crane 1975U. dussumieri Deltuca both sexes Crane 1975U. paradussumieri Deltuca Ng and Sivasothi 1999U. forcipata Deltuca from within the burrow female Crane 1975U. urvillei Deltuca from within the burrow male (?) and female Crane 1975U. elegans Australuca George and Jones 1982U. tetragonon Thalassuca from the surface female Crane 1975U. chlorophthalmus Amphiuca female Crane 1975
crassipesU. chlorophthalmus Amphiuca female Crane 1975U. thayeri Boboruca from the surface (?) female von Hagen 1970, Crane 1975,
Salmon 1987, Kellmeyer andSalmon 2001, Weaver andSalmon 2002
U. galapagensis Minuca male von Hagen 1968U. minax Minuca Montague 1980U. pugnax Minuca Montague 1980U. subcylindrica Minuca from within the burrow (?) Thurman 1984U. vocator Minuca both sexes Crane 1975U. cumulanta Celuca Crane 1975U. stenodactylus Celuca female (?) Crane 1975
aBased on Shih et al. (1999).

Zoological Studies 44(2): 242-251 (2005)244
related behaviors were recorded on video tape.Reproductive behaviors (e.g., courtship, surfacemating, underground mating, etc.) and otherbehaviors (e.g., fighting, feeding, etc.) wererecorded by ad libitum sampling (Martin andBateson 1993). If a certain behavior pattern wasobserved, it was recorded until it ceased. Someburrows with chimneys were dug out, and the sexand carapace width (CW) of the pair were mea-sured, as well as the reproductive condition of thefemale. The depth of the burrow and its relativedistance to the high tide mark were also recorded.Behaviors related to chimney building in Jhuwei(Taipei County), Jinshuei (Hsinchu City),Haishangu (Hsinchu City), Shengang (ChanghuaCo.), Dongshih (Chiayi Co.), Budai (Chiayi Co.),and Cigu (Tainan Co.) from Sept. 1994 to Apr.1997 were also recorded to supplement theresults.
Uca arcuata (de Haan) is another species inTaiwan that frequently builds chimneys. Its chim-ney building behavior was also recorded and isdescribed for comparison.
RESULTS
Chimneys of U. formosensis appeared on themudflats of the high intertidal zone during the peri-od of the neap tide in the warm season (Mar. toOct.) in Taiwan (Fig. 1A).
Chimney structure
After a male U. formosensis had attracted afemale to stay inside his burrow for undergroundmating, the male began to build a tower-like cylin-drical structure, a chimney (average height, 9.2(range, 4~14) cm; average diameter at the base,7.7 (range, 3~13 cm); n = 26) (Fig. 1C, D), aroundthe burrow entrance. The chimney was composedof large wet soil pellets (i.e., mudballs). The mate-rial for the chimney was carried up from within theburrow (Fig. 1G), not from the surface substratenear the burrow as with the chimney building of U.arcuata (Fig. 1K). Therefore, the color of the chim-ney and the substrate generally differed.Sometimes, both the entrance of the chimney (andalso the entrance of the burrow) would becomeplugged by mudballs (Fig. 1E, F; see below). Inaddition, because the builder did not trim the chim-ney like U. arcuata does (see“Discussion”), itssurface was rugged, and generally there were sev-eral large gaps among the mudballs.
Twelve females were observed to excavatemudballs from within the burrow and five of thesefemales were ovigerous. The female piled up themudballs loosely beside, but not around, the bur-row, and the resulting structure resembled a pyra-mid (Fig. 1H), not a cylinder like the male
,s chim-
ney.
Chimney building
We determined the process and timing ofchimney building from successive 15 d observa-tions conducted in Shengang, Changhua, Taiwan.
1. During the period of a neap tide, theamount of time that the tide covered the habitat ofU. formosensis (in the high intertidal zone)decreased each day. About 1 or 2 d before thetide did not cover its habitat, males began courtingwandering females. When a female stayed insidehis burrow, the male began to build a chimney.The male excavated mudballs from inside the bur-row, carrying them using its ambulatories on theminor side (Fig. 1G), and piled them on one side ofthe burrow entrance. The crab did not place mud-balls on the other side until the chimney was fin-ished. Therefore, the structure appeared hood-shaped while it was being built (Fig. 1B, G). Theoriginal entrance was plugged by mudballs to 3~5cm in depth (Fig. 1E), while the entrance of thechimney sometimes became plugged by 1 or 2mudballs (Fig. 1F). Meanwhile, the female stayedinside the burrow, and it was assumed that she didnot participate in the building process. A chimneycould be finished in 1 d.
2. On the day the tide first failed to cover thehabitat (e.g., during the day on 10 July, Fig. 2C),most chimneys had already been built. A few indi-viduals of U. formosensis without chimneys cameto the surface by the side of their burrow entrancesbut were generally inactive. On the following days,the ground temperature of the sand graduallyincreased, and no U. formosensis (or any othercrab species) was observed on the surface duringthis period (10~13 July, Fig. 2A). The mudflatsbecame cracked (Fig. 1I), and salt crystallized onthe surface (Fig. 1J). The highest sun, shade, andsoil temperatures measured were 45.9, 39.7, and41.1°C, respectively, on 13 July. This droughtperiod lasted for 4 or 5 d until the tide returned toits normal levels.
3. When the tide again covered the habitat(14 July), the chimneys were destroyed by thewater due to their loose structure. Then, increas-ing numbers of individuals began emerging from

Shih et al. -- Chimney Building by Uca formosensis 245
Fig. 1. Chimney building of Uca formosensis. (A) Numerous chimneys of U. formosensis protruding above the mudflat when the habi-tat was not covered by the tide, at which time it became extremely dry (Shengang, Changhua, Taiwan). (B) A hood-shaped chimney inthe middle stage of chimney building. (C and D) Two types of chimneys. (E) The burrow entrance also becoming plugged (arrow) bymudballs to about 3~5 cm in depth after removing the chimney. (F) The entrance of a chimney having become plugged (arrow) bymudballs. (G) A male building a chimney. (H) A chimney built by an ovigerous female. Note that it is loose and without a distinctshape. (I and J) Mudflats having become cracked, and salt having crystallized on the surface. (K) For contrast, a chimney built by U.arcuata with many scraping traces on its surface (arrow). Scale = 5 cm.
(A)
(C)
(G)
(I)
(D)
(B)
(E)
(H)
(K)
(F)
(J)

Zoological Studies 44(2): 242-251 (2005)246
their burrows (Fig. 2A), and they mainly fed andfought, but courtship was rare. The tides werehigh enough to periodically cover the habitat for 10to 11 d, during which time, chimneys were rare.When the next neap tide occurred, the chimneybuilding processes began again, synchronized withthe semilunar cycle.
Relationships between the number of crabson the surface and temperatures each hour areshown in Fig. 2A. The number of crabs was nega-tively correlated with sun temperature (R = - 0.37,p < 0.001) and shade temperature (R = - 0.28, p =0.005), but not soil temperature (R = 0, p = 0.99;d.f. = 98) on an hourly basis. However, R valueswere higher if the average number of crabs andtemperatures each day were used (sun tempera-ture: R = - 0.54, p = 0.11; shade temperature: R =- 0.70, p = 0.02; soil temperature: R = - 0.58, p =0.08; d.f. = 9). The most-significant negative cor-relation with the number of females (thick line inthe lower part of Fig. 2A) was with the sun temper-
ature (R = - 0.79, p = 0.006, d.f. = 9), and for thenumber of males (thin line in the lower part of Fig.2A) was with the shade temperature (R = - 0.64,p = 0.04).
When the tide covered the habitat after thedrought period, crabs emerged from their burrows.A greater number of males (with an average sexratio of males: females of 4.68, and a range of1.81~10.02, Fig. 2B) always appeared on the mud-flats. During the time when the habitat was cov-ered by the tide at least once a day (Fig. 2C),female numbers increased to a maximum (16July), followed by the maximum of male numberswhich occurred 3 d later (Fig. 2A). The sex ratio ofcrabs appearing on the mudflats also showed atendency to fluctuate with the tide. The ratioincreased to its highest value (of 10.02) 7 d afterthe tide began covering the habitat again (Fig. 2B).It is apparent that the increasing value of the sexratio was caused by an increase in the numbers ofmales, although it decreased slightly after 20 July,
Fig. 2. Relationships among temperatures, the number of Uca formosensis, sex ratio, and tide height recorded in the study areas inShengang, Changhua, Taiwan. (A) Sun, shade, and soil temperatures each hour shown in the upper part. The gray and white bars inthe lower part represent the number of males and females appearing on the surface, respectively. The average number of males (thinline) and females (thick line) each day is also shown. (B) The sex ratio (number of males/females) each hour (thin line) and its averagevalue each day (thick line with values shown above it). (C) Icons at the bottom indicating the moon phase. The gray bars are the tidalheight, days and nights, each day; the horizontal line is the height estimated for the study areas; the asterisk (*) above the gray bar indi-cates when the study area was covered by tide in the daytime; and icons in the upper part represent the processes of chimney building.See text for further explanations.
60
50
40
30
20
10
0
4.68 2.93
212019181716151413121110987
1.81 3.24 2.31 2.36 3.94 4.84 6.59 8.6910.02
25
30
35
40
45
50
55
60
65
70
20
10
0
480
460
440
420
400
Tem
pera
ture
(°C
)S
ex r
atio
Tid
e he
ight
(m
)
Cra
b nu
mbe
r
Sun temp.
Average male no.
Average female no.
Average sex ratio
July
Shade temp. Soil temp.(A)
(B)
(C)
Shih et al. -- Chimney Building by Uca formosensis 247
and there was also a decrease in the number offemales.
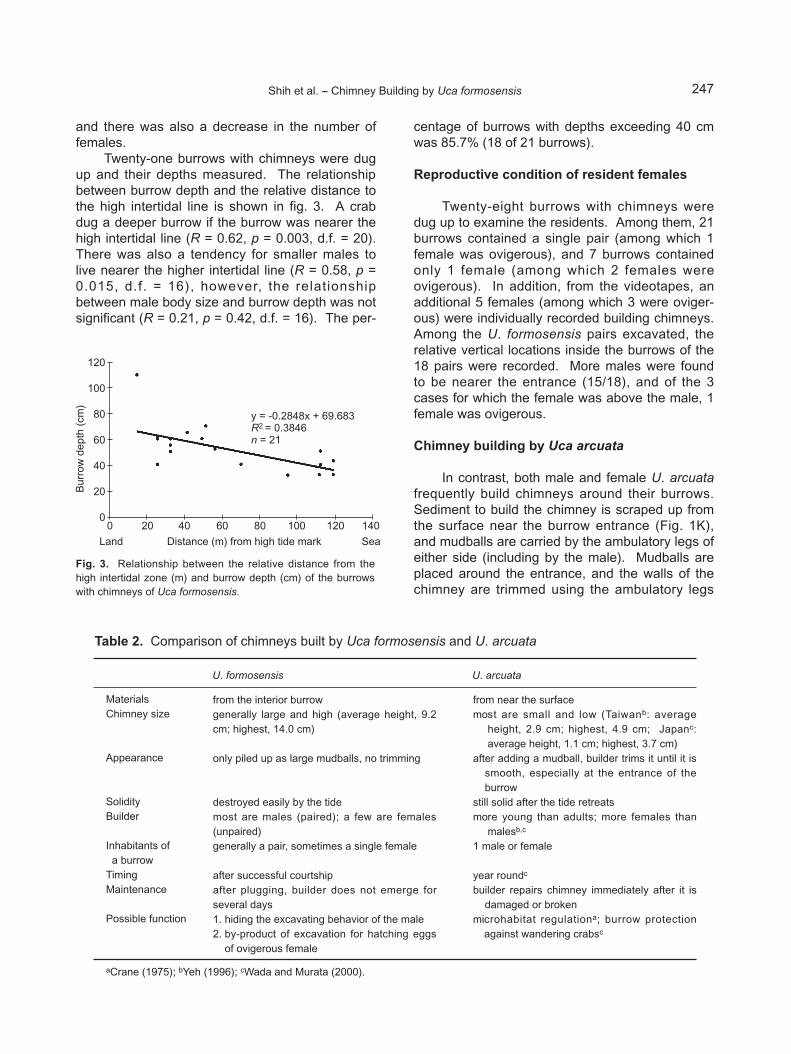
Twenty-one burrows with chimneys were dugup and their depths measured. The relationshipbetween burrow depth and the relative distance tothe high intertidal line is shown in fig. 3. A crabdug a deeper burrow if the burrow was nearer thehigh intertidal line (R = 0.62, p = 0.003, d.f. = 20).There was also a tendency for smaller males tolive nearer the higher intertidal line (R = 0.58, p =0.015, d.f. = 16), however, the relationshipbetween male body size and burrow depth was notsignificant (R = 0.21, p = 0.42, d.f. = 16). The per-
centage of burrows with depths exceeding 40 cmwas 85.7% (18 of 21 burrows).
Reproductive condition of resident females
Twenty-eight burrows with chimneys weredug up to examine the residents. Among them, 21burrows contained a single pair (among which 1female was ovigerous), and 7 burrows containedonly 1 female (among which 2 females wereovigerous). In addition, from the videotapes, anadditional 5 females (among which 3 were oviger-ous) were individually recorded building chimneys.Among the U. formosensis pairs excavated, therelative vertical locations inside the burrows of the18 pairs were recorded. More males were foundto be nearer the entrance (15/18), and of the 3cases for which the female was above the male, 1female was ovigerous.
Chimney building by Uca arcuata
In contrast, both male and female U. arcuatafrequently build chimneys around their burrows.Sediment to build the chimney is scraped up fromthe surface near the burrow entrance (Fig. 1K),and mudballs are carried by the ambulatory legs ofeither side (including by the male). Mudballs areplaced around the entrance, and the walls of thechimney are trimmed using the ambulatory legs
Table 2. Comparison of chimneys built by Uca formosensis and U. arcuata
U. formosensis U. arcuata
MaterialsChimney size
Appearance
SolidityBuilder
Inhabitants ofa burrow
TimingMaintenance
Possible function
aCrane (1975); bYeh (1996); cWada and Murata (2000).
120
100
80
60
40
20
0140120100806040200
y = -0.2848x + 69.683R2 = 0.3846n = 21
Land
Bur
row
dep
th (
cm)
SeaDistance (m) from high tide mark
Fig. 3. Relationship between the relative distance from thehigh intertidal zone (m) and burrow depth (cm) of the burrowswith chimneys of Uca formosensis.
from the interior burrowgenerally large and high (average height, 9.2cm; highest, 14.0 cm)
only piled up as large mudballs, no trimming
destroyed easily by the tidemost are males (paired); a few are females(unpaired)generally a pair, sometimes a single female
after successful courtshipafter plugging, builder does not emerge forseveral days1. hiding the excavating behavior of the male2. by-product of excavation for hatching eggs
of ovigerous female
from near the surfacemost are small and low (Taiwanb: average
height, 2.9 cm; highest, 4.9 cm; Japanc:average height, 1.1 cm; highest, 3.7 cm)
after adding a mudball, builder trims it until it issmooth, especially at the entrance of theburrow
still solid after the tide retreatsmore young than adults; more females than
malesb,c
1 male or female
year roundc
builder repairs chimney immediately after it isdamaged or broken
microhabitat regulationa; burrow protectionagainst wandering crabsc

Zoological Studies 44(2): 242-251 (2005)248
and both chelipeds. If a chimney becomes dam-aged or broken, the builder repairs it immediately.A new chimney can be finished in less than 30min, even if it had been badly damaged (see Table2).
DISCUSSION
Chimney building by Uca formosensis wasobserved daily during 15 d periods. However,chimneys were destroyed by the tide, so subse-quent observations of the behavior of chimneybuilders and their mates were impossible.According to Nakasone and Murai (1998), the pair-ing time (the time which a male and female staytogether within a burrow after successful courtship)of narrow-fronted fiddlers (e.g., 5.7 (range, 3~9) dfor U. rosea, Murai et al. 1996; and 5.2 (range,4~8) d for U. tetragonon, Goshima et al. 1996) ismuch longer than that of broad-fronted species(e.g., 1.8 (range, 1~3) d for U. pugilator, Christy1978; and 2.0 (range, 1~5) d for U. lactea ,Goshima and Murai, 1988). At our study site, thehabitat of U. formosensis was not covered by thetide for 4 to 5 d (Fig. 2C), and the male was seento leave its burrow once the chimney had beendestroyed by the tide (pers. obs.). The amount oftime that the narrow-fronted U. formosensis spentin mate-guarding was presumed to match thisdrought period. It is possible that the female U.formosensis stays inside the burrow until the eggshatch after the male has left. The incubation dura-tion in most female fiddler crabs ranges from 12 to15 d (Zucker 1978, Christy and Salmon 1984,Salmon 1987, Salmon and Zucker 1988), exceptfor the incubation duration of U. tetragonon whichis 28 d (Goshima et al. 1996). Females of severalUca species that live in the upper intertidal zonerelease larvae during the largest-amplitude noctur-nal high tide of the lunar month (Morgan andChristy 1995). During this study, only 1 ovigerousfemale was seen feeding on the surface, and otherovigerous females were seen piling up mudballs(Fig. 1H). Therefore, when the chimney wasdestroyed by the tide, we presumed that thefemale U. formosensis stayed underground for afurther 16 to 18 d until the night of the next springtide.
The number of crabs appearing on the mud-flats each day was negatively correlated with thesun, shade, and soil temperatures (Fig. 2A).When the temperature was too high, U. formosen-sis did not appear on the surface. However, tem-
perature, sediment dryness, and food availabilityare likely interrelated, and without further study, itis unclear which factors most affect crab activity.
Johnson (2003) suggested that the male-biased sex ratios in U. pugilator may be due to dif-ferential mortality between the sexes. During thepresent study, individuals of U. lactea and U. arcu-ata were seen being eaten by birds, and discardedchelae of the 2 species were frequently found, butnot of individuals of U. formosensis (Shih 1997).However, individuals of U. formosensis wereobserved being attacked by other crabs during thestudy. Both cases occurred at dusk when the lightwas weak. In one case, an adult male U. for-mosensis was preyed upon, and his viscera wereeaten by a Helice formosensis. In another case, ayoung male U. formosensis was attacked by ayoung H. wuana, but the fiddler autotomized hismajor cheliped and escaped after his chelipedplunged into the legs of the attacker (Shih 1997).While the differential mortality caused by thedefensive chela weapon may be a possible factorfor explaining the male-biased sex ratio of U. for-mosensis, other factors related to the mortality offemales cannot be excluded. The sex ratio of U.formosensis on the surface tended to fluctuate withthe tide (ranging from 1.81 to 10.02, Fig. 2B).When the number of females began decreasing,the number of males began to dramaticallyincrease (decreasing slightly after 20 July) (Fig.2A). One day after the tide had risen to periodical-ly cover the habitat, female numbers increased toa maximum which may have been because thefemales were maximizing their food intake inpreparation for breeding after a period of inactivity.When the sun temperature increased, the femalesspent less time on the surface possibly to escapelethal temperatures.
deRivera and Vehrencamp (2001) suggestedthat the degree of synchrony of receptive females(to the tidal height of the habitat) and femalegametic investment (clutch size) can explain thesex ratio. An upper intertidal fiddler speciesbreeds more synchronously than lower intertidalspecies (Morgan and Christy 1995), and the sexratio approaches 1 (deRivera and Vehrencamp2001). Females of fiddler species with large clutchsizes produce a male-biased sex ratio because thefemale
,s cycle length of reproduction is longer (i.e.,
females spend more time underground during thehatching period, and fewer females appear on thesurface) (Christy and Salmon 1984). However,these 2 parameters are interrelated in a way thatcancels out their influences, because smaller fid-

Shih et al. -- Chimney Building by Uca formosensis 249
dler species tend to have larger clutch sizes andinhabit areas higher in the tidal zone. Uca for-mosensis is a large-sized species (CW > 30 mm,Shih et al. 1999) and inhabits the high intertidalzones, but the sex ratio did not approach equalitybased on the 2 parameters, indicating that thefemale gametic investment as incubation daysinstead of clutch size better explains the highmale-bias sex ratio of U. formosensis.
Here we propose and discuss some possiblehypotheses to explain the functions of the chim-neys built by U. formosensis. The 1st is the sexualattraction hypothesis. Because the chimney of U.formosensis is built after a male has successfullyattracted a female into his burrow, all possiblefunctions, direct or indirect, of construction for sex-ual attraction, e.g., the hood of U. musica terpsi-chores (Zucker 1974, Christy et al. 2002) and thepillar of U. beebei (Christy 1988a b), are excluded.The 2nd is the sunshade effect hypothesis. Itseems likely that the large chimney of U. for-mosensis provides a sunshade effect for loweringthe temperature of the burrow underneath.However, since the bottom of the burrow is notdirectly under its entrance, and nearly 86% of bur-rows were more than 40 cm deep (Fig. 3); the sun-shade effect of lowering burrow temperatures isprobably negligible. The 3rd is the ventilationhypothesis. The chimney of U. formosensis isgenerally loosely constructed from mudballs, andmany large gaps remain in the walls of the chim-ney; thus, the wind can easily ventilate the burrowthrough these gaps. However, after the chimney isfinished, the burrow entrances can becomeplugged (Fig. 1F), a behavior also seen in otherspecies of Uca (Salmon and Zucker 1988), andthus a ventilatory function of the chimney is highlyunlikely. The 4th is the hypothesis of warding offintruders. McCann (1938) proposed that the func-tion of the castle built by the terrestrial (freshwater)crab, Paratelphusa (Barytelphusa) guerini (H.Milne Edwards) on Salsette I., India, was to keepintruders away during estivation, because it actedas a bluff which is mistaken for a clod of earth.Silas and Sankarankutty (1960) also citedMcCann
,s opinion to explain the castle built by the
land crab, Cardisoma carnifex (Herbst). The bur-row with paired fiddler crabs was found to beusurped by other males (e.g., U. rosea, Murai et al.1996, and U. lactea perplexa, Nakasone and Murai1998). Wada and Mutata (2000) also consideredthat the function of the chimney of U. arcuata wasto protect the burrow from usurpation by wander-ing crabs. Therefore, the function of the chimney
of the mating burrow may possibly be to keepintruders away to prevent usurpation. However, inthe case of the chimney of U. formosensis, afterthe chimney was finished, the environment on themudflats greatly deteriorated (no tide covered itand temperatures increased, see Fig. 2), and nocrab was seen on the surface during that period. Itis unlikely that a pair inside a burrow would be dis-turbed by other crabs. Therefore, the chimney ofU. formosensis does not seem to play a defensiverole. The last possible hypothesis is that the chim-ney hides the burrower. There are 2 reasons whymales need to excavate their burrows after attract-ing a mate. First, a male
,s burrow is for a single
crab before pairing (Fig. 4A). After pairing, thediameter of the burrow shaft, at least the spacenear the bottom, must be enlarged to accommo-date 2 individuals (Fig. 4B(1)). Second, it is impor-tant to dig a deeper burrow to reach the watertable for the female, because the neap tide isapproaching (with lower water levels at that time),and moisture or water in the burrow is necessaryfor incubation (Kellmeyer and Salmon 2001,Weaver and Salmon 2002; Fig. 4B(2)). Generally2 types of chimneys, with either a wide or slenderdiameter (Fig. 1C, D), were constructed by U. for-mosensis. The burrow depth was correlated withthe tidal level (Fig. 3), but was not related to male
Fig. 4. A possible mechanism of chimney building by Uca for-mosensis. (A) Diagrammatic representation of a typical burrowof a male without a chimney before pairing. (B) After pairing.The shaft of the burrow must be enlarged (1), the bottom of theburrow is dug deeper until it reaches the water table (2), andthe male piles the mudballs around the burrow entrance to hidehimself while excavating (3). (C) After the excavation behavioris completed. The male may plug the entrances of both thechimney and burrow. See“Discussion”for further explana-tions.
(A) (B) (C)
(1)
(3) x
x
(2)♂
♂
♂♂
♀
♀
♀*

Zoological Studies 44(2): 242-251 (2005)250
body size (p = 0.42). Therefore, we predict thatthe width of a chimney is positively related to thesize of the female and/or male while the height ofthe chimney is positively related to the vertical dis-tance to the water table.
This is a new hypothesis for chimney buildingby U. formosensis, and it also has implications formore-terrestrial species that have to burrow to thewater table on dry beaches. When the male U.formosensis carries the mudballs to the surface,he is faced with the danger of predation, orusurpation of the burrow and/or female by othermales. One possible solution for him is to pilemudballs around the burrow entrance using hismajor cheliped, while the ambulatories of the minorside clasp onto the entrance, thus hiding himselfbehind the wall of the chimney so that he canrapidly withdraw into the burrow if necessary (Fig.4B(3)). In U. rosea, after a pair enters a burrow,the male also repeatedly carries mud from the bur-row and drops it onto the surface a short distancefrom the entrance until he plugs the entrance(Murai et al. 1996). It is possible that U. formosen-sis has evolved chimney building to hide the malefrom predators or other crabs that may attempt tousurp the burrow and female. The chimney struc-ture here is hypothesized to be a by-product ofburrowing. After the excavating behavior is com-pleted, the male may plug the entrances to boththe chimney and burrow (Figs. 1E, F, 4C) to pre-vent any enemy from entering at that moment andalso to maintain the moist environment within theburrow during the 4 to 5 d period of drought.
This hypothesis can also explain the excavat-ing behavior by the ovigerous (or assuming it willsoon be ovigerous) female of U. formosensis. Afemale may excavate mudballs from within a bur-row because the male of the pair has disappeared(e.g., killed by a predator). A more-plausibleexplanation is that the female had already beenfertilized in surface mating, and she needs to dig adeeper burrow by herself in order to reach thewater table for incubation, which is similar to themale
,s work in underground mating. However, the
female was unable to build a structure like themale
,s chimney (Fig. 1H).
When burrows of U. formosensis were dugout, most males were found to be above thefemale at the bottom in paired burrows (15 of 18burrows). This phenomenon had previously beennoted, and of all pairs excavated, males werenearer the entrance in those observations (vonHagen 1962, Nakasone et al. 1983, Yamaguchi1998). Yamaguchi (1998) suggested that the rela-
tive position of U. lactea has to do with the matingbehavior process. In this study, there was general-ly a large space inside the burrow of U. formosen-sis , and a pair could meet each other andexchange their positions inside the burrow (pers.obs.). Therefore, it is possible that we were mis-guided by the relative positions of the crabs whenthe burrow was dug out. However, the femaleshould generally be in the lower position near thebottom, because an ovigerous female needs amoist, even water-containing, environment. Asanother explanation, some predators or othercrabs may enter the burrow, and so the male maygenerally maintain an upper position for defensivepurposes (Nakasone and Murai 1998).
Acknowledgments: We wish to express ourthanks to Prof. Heinrich-Otto von Hagen(Department of Biology-Zoology, Phil ipps-University of Marburg, Marburg, Germany) for dis-cussions about chimney functions. We also wishto thank 2 anonymous reviewers for greatlyimproving this manuscript.
REFERENCES
Christy JH. 1978. Adaptive significance of reproductive cyclesin the fiddler crab Uca pugilator: a hypothesis. Science199: 453-455.
Christy JH. 1988a. Pillar function in the fiddler crab Uca bee-bei (I): effects on male spacing and aggression. Ethology78: 53-71.
Christy JH. 1988b. Pillar function in the fiddler crab Uca bee-bei (II): competitive courtship signaling. Ethology 78: 113-128.
Christy JH, PRY Backwell, S Goshima, T Kreuter. 2002.Sexual selection for structure building by courting malefiddler crabs: an experimental study of behavioral mecha-nisms. Behav. Ecol. 13: 366-374.
Christy JH, JK Baum, PRY Backwell. 2003. Attractiveness ofsand hoods built by courting male fiddler crabs, Uca musi-ca: test of a sensory trap hypothesis. Anim. Behav. 66:89-94.
Christy JH, M Salmon. 1984. Ecology and evolution of matingsystems of fiddler crabs (genus Uca). Biol. Rev. 59: 483-509.
Clayton DA. 1988. Hood construction as a spacing mecha-nism in Cleistostoma kuwaitense (Crustacea:Ocypodidae). Mar. Biol. 99: 57-61.
Crane J. 1975. Fiddler crabs of the world (Ocypodidae: genusUca). Princeton, NJ: Princeton Univ. Press.
deRivera CE, SL Vehrencamp. 2001. Male versus femalemate searching in fiddler crabs: a comparative analysis.Behav. Ecol. 12: 182-191.
George RW, DS Jones. 1982. A revision of the fiddler crabs ofAustralia (Ocypodinae: Uca). Rec. West. Aust. Mus.Supplement 14: 5-99.
Goshima S, T Koga, M Murai. 1996. Mate acceptance andguarding by male fiddler crabs Uca tetragonon (Herbst).

Shih et al. -- Chimney Building by Uca formosensis 251
J. Exp. Mar. Biol. Ecol. 196: 131-143.Goshima S, M Murai. 1988. Mating investment of male fiddler
crabs, Uca lactea. Anim. Behav. 36: 1249-1251.Hughes DA. 1973. On mating and the“copulation burrows”
of crabs of the genus Ocypode (Decapoda, Brachyura).Crustaceana 24: 72-76.
Johnson PTJ. 2003. Biased sex ratios in fiddler crabs(Brachyura, Ocypodidae): a review and evaluation of theinfluence of sampling method, size class, and sex-specificmortality. Crustaceana 76: 559-580.
Kellmeyer K, M Salmon. 2001. Hatching rhythms of Uca thay-eri Rathbun: timing in semidiurnal and mixed tidalregimes. J. Exp. Mar. Biol. Ecol. 260: 169-183.
Linsenmair KE. 1967. Konstruktion und Signalfunktion derSandpyramide der Reiterkrabbe Ocypode saratan Forsk.(Decapoda Brachyura Ocypodidae). Z. Tierpsychol. 24:403-456.
Martin P, P Bateson. 1993. Measuring behaviour: an introduc-tory guide. 2nd ed. Cambridge, UK: Cambridge Univ.Press.
McCann C. 1938. Notes on the common land crabParatelphusa (Barytelphusa) guerini (M.-Eds.) of SalsetteIsland. J. Bombay Nat. Hist. Soc. 39: 531-542.
Montague CL. 1980. A natural history of temperate WesternAtlantic fiddler crabs (genus Uca) with reference to theirimpact on the salt marsh. Contrib. Mar. Sci. 23: 25-55.
Morgan SG, JH Christy. 1995. Adaptive significance of the tim-ing of larval release by crabs. Am. Nat. 145: 457-479.
Murai M, S Goshima, K Kawai, HS Yong. 1996. Pair formationin the burrows of the fiddler crab Uca rosea (Decapoda:Ocypodidae). J. Crustacean Biol. 16: 522-528.
Nakasone Y, M Murai. 1998. Mating behavior of Uca lacteaperplexa (Decapoda: Ocypodidae). J. Crustacean Biol.18: 70-77.
Ng PKL, N Sivasothi, eds. 1999. A guide to the mangroves ofSingapore II: animal diversity. Singapore: SingaporeScience Centre.
Oliveira RF, PK McGregor, FRL Burford, MR Custódio, CLatruffe. 1998. Functions of mudballing behaviour in theEuropean fiddler crab Uca tangeri. Anim. Behav. 55:1299-1309.
Powers LW, DE Bliss. 1983. Terrestrial adaptations. In FJVernberg, WB Vernberg, eds. The biology of Crustacea.Vol. 8, Environmental adaptations. New York: AcademicPress, pp. 271-333.
Powers LW, JF Cole. 1976. Temperature variation in fiddlercrab microhabitats. J. Exp. Mar. Biol. Ecol. 21: 141-157.
Salmon M. 1987. On the reproductive behavior of the fiddlercrab Uca thayeri, with comparisons to U. pugilator and U.vocans: evidence for behavioral convergence. J.Crustacean Biol. 7: 25-44.
Salmon M, N Zucker. 1988. Interpreting differences in the
reproductive behaviour of fiddler crabs (genus Uca). In GChelazzi, M Vannini, eds. Behavioral adaptation to inter-tidal life. New York: Plenum Press, pp. 387-407.
Shih HT. 1997. The fiddler crab that belongs to Formosa, Ucaformosensis. Where should they go? - the present condi-tion of an endemic fiddler crab of Taiwan. Taiwan Nat.Sci. 54: 68-80. (in Chinese)
Shih HT, HK Mok, HW Chang, SC Lee. 1999. Morphology ofUca formosensisa Rathbun, 1921 (Crustacea: Decapoda:Ocypodidae), an endemic fiddler crab from Taiwan, withnotes on its ecology. Zool. Stud. 38: 164-177.
Silas EG, C Sankarankutty. 1960. On the castle building habitof the crab Cardisoma carnifex (Herbst) (familyGeocarcinidae), of the Andaman Islands. J. Mar. Biol.Assoc. India. 2: 237-240.
Takeda S, M Matsumasa, HS Yong, M Murai. 1996.“Igloo”construction by the ocypodid crab, Dotilla myctiroides(Milne-Edwards) (Crustacea; Brachyura): the role of an airchamber when burrowing in a saturated sandy substra-tum. J. Exp. Mar. Biol. Ecol. 198: 237-247.
Thurman CL II. 1984. Ecological notes on fiddler crabs ofsouth Texas, with special reference to Uca subcylindrica.J. Crustacean Biol. 4: 665-681.
von Hagen HO. 1962. Freilandstudien zur Sexual- undFortpflanzungs- Biologie von Uca tangeri in Andalusien.Z. Morph. Ökol. Tiere 51: 611-725.
von Hagen HO. 1968. Studien an peruanischenWinkerkrabben (Uca). Zool. Jahrb. Syst. 95: 395-468.
von Hagen HO. 1970. Zur Deutung langstieliger und gehörnterAugen bei Ocypodiden (Decapoda, Brachyura). FormaFunctio 2: 13-57.
Wada K. 1984. Barricade building in Ilyoplax pusillus (DeHaan) (Crustacea: Brachyura). J. Exp. Mar. Biol. Ecol.83: 73-88.
Wada K, I Murata. 2000. Chimney building in the fiddler crabUca arcuata. J. Crustacean Biol. 20: 507-511.
Weaver A, M Salmon. 2002. Hatching rhythms of Uca thayeri:evidence for phenotypic plasticity. J. Crustacean Biol. 22:429-438.
Yeh CL. 1996. Observations of chimney function and territorialbehavior in the fiddler crab Uca arcuata de Haan, 1835.Master
,s thesis, Fu Jen Catholic Univ., Taipei. (in
Chinese)Zucker N. 1974. Shelter building as a means of reducing terri-
tory size in the fiddler crab, Uca terpsichores (Crustacea:Ocypodidae). Am. Midl. Nat. 91: 224-236.
Zucker N. 1978. Monthly reproductive cycles in three sym-patric hood-building tropical fiddler crabs (genus Uca).Biol. Bull. 155: 410-424.
Zucker N. 1981. The role of hood-building in defining territo-ries and limiting combat in fiddler crabs. Anim. Behav. 29:387-395.

(A)
(C)
(G)
(D) (E)
(B)
(I)
(H)
(J)
(F)
(K)
Fig. 1

****
******
*
*
7 8 9 10 11 12 13 14 15 16 17 18 19 20 21400
420
440
460
480
July
0
10
204.68 2.93 1.81 3.24 2.31 2.36 3.94 4.84 6.59 8.69
0
10
20
30
40
50
60
Cra
b nu
mbe
r
25303540455055606570
10.02
Sun temp. Shade temp. Soil temp.
Tide
hei
ght (
m)
Tem
pera
ture
(°C
)Se
x ra
tio
(A)
(B)
(C)
Average female no.
Average male no.
Average sex ratio
Fig. 2

(1)
(2)
(3) ×
×
*(A) (B) (C)
Fig. 4