characterization of thermobifida fusca cutinase-cbm...
TRANSCRIPT

1
1
2
Characterization of Thermobifida fusca Cutinase-CBM 3
Fusion Proteins and Their Potential Application in 4
Bioscouring 5
Yao Zhang1,
2, Sheng Chen
1, 2, Meng Xu
1, 2, Artur Cavoco-Paulo
3, Jing Wu
1, 2*
, 6
and Jian Chen1, 2*
7
1State Key Laboratory of Food Science and Technology,
2School of Biotechnology and 8
Key Laboratory of Industrial Biotechnology Ministry of Education, Jiangnan University, 9
Wuxi, 214122, China, 3
Department of Textile Engineering, University of Minho, 4800 10
Guimaraes, Portugal. 11
12
13
14
15
16
* Corresponding author. Mailing address: State Key Laboratory of Food Science and 17
Technology, Jiangnan University, Wuxi 214122, China. Jing Wu, Phone: 18
86-510-85327802. Fax: 86-510-85327802. E-mail: [email protected] or Jian 19
Chen, Phone: 86-510- 85329031. Fax: 86-510-85918309. Email: [email protected]
Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.00896-10 AEM Accepts, published online ahead of print on 20 August 2010
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2
ABSTRACT 21
Cutinase from Thermobifida fusca is thermally stable and has potential application 22
in the bioscouring of cotton in the textile industry. In the present study, the 23
carbohydrate-binding module (CBM) from T. fusca cellulase Cel6A (CBMCel6A) and C. 24
fimi cellulase CenA (CBMCenA) were fused, separately, to the carboxyl-terminus of T. 25
fusca cutinase. Both fusion enzymes, cutinase-CBMCel6A and cutinase-CBMCenA, were 26
expressed in E. coli and purified to homogeneity. Enzyme characterization showed that 27
both of them displayed similar catalytic properties and pH stability to T. fusca cuinase. 28
In addition, both fusion proteins displayed an activity half-life of 53 h at their optimal 29
temperature of 50 °C. Compared with T. fusca cutinase, in the absence of pectinase, the 30
binding activity on cotton fiber was enhanced by 2% for cutinase-CBMCel6A and 28% 31
for cutinase-CBMCenA; while in the presence of pectinase, the binding activity was 32
enhanced by 40% for the former and 45% for the latter. Notably, dramatic increase of up 33
to 3-fold was observed in the amount of released fatty acids from cotton fiber by both 34
cutinase-CBM fusion proteins when acting in concert with pectinase. This is the first 35
report of improving the scouring efficiency of cutinase by fusing it with CBM. The 36
improvement in activity and the strong synergistic effect between the fusion proteins 37
and pectinase suggests that they may have better applications in textile bioscouring than 38
the native cutinase.39
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

3
INTRODUCTION 40
Cotton fiber has a multilayered structure with its outermost surface being the 41
cuticle which is crosslinked to the primary cell wall of cotton fiber by esterified pectin 42
substances. The major component of the cuticle is cutin, an insoluble polyester 43
composed mainly of saturated C:16 and C:18 hydroxy and epoxy fatty acids (15, 17, 37, 44
48). During the process of scouring in the textile industry, the cuticle layer has to be 45
removed in order to improve the wettability of cotton fiber, which then facilitates 46
uniform dyeing and finishing. Traditionally, this process is performed by hot hydrolysis 47
in alkaline medium, which not only consumes large quantities of water and energy but 48
also causes severe pollution and fiber damage (21, 22, 43). Therefore, 49
environment-friendly scouring methods based on biocatalysts have been actively sought 50
(2, 41, 46). 51
Cutinase is a multi-functional esterase capable of degrading the cutin component of 52
the cuticle. Earlier reports showed that the fungal cutinase from Fusarium solani pisi 53
has potential use for cotton cuticle degradation and exhibits a good synergistic effect 54
with pectinase, an enzyme utilized to degrade pectin, in the scouring of cotton fiber (1, 8, 55
9, 15). Moreover, site-directed mutagenesis has been performed to substitute the 56
specific amino acid residues near the active site of cutinase (3) to improve its hydrolytic 57
activity towards polyesters. More recently, a cutinase from the thermophilic bacterium 58
Thermobifida fusca has been identified and overexpressed in E. coli in our laboratory 59
(11). The good thermal stability and alkali resistance of this recombinant T. fusca 60
cutinase make it potentially more amenable to textile bioscouring (11). 61
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

4
To further improve the applicability and/or catalytic efficiency of T. fusca cutinase, 62
the present study attempts to engineer a novel cutin-degrading enzyme, based on 63
analysis of the surface structure of cotton fiber. It has been observed that, in addition to 64
cutin, pectin, proteins and other components, there is also a large amount of cellulose on 65
the surface layer of cotton fiber (31). Thus it is tempting to hypothesize that if the 66
enzyme can be engineered to specifically bind to cellulose through a “gain of function” 67
modification, its concentration on the surface of cotton fiber could increase significantly. 68
Subsequently, its catalytic efficiency for cutin breakdown could be improved due to 69
proximity effect. In order to design such an enzyme, a fusion protein strategy in which a 70
cellulose binding protein/module will be attached to cutinase is considered. 71
It is well known that cellulase is capable of binding specifically to cellulose (35, 72
42). This enzyme has two separate modules, a catalytic module and a 73
carbohydrate-binding module (CBM) (12). The two modules are discrete structural and 74
functional units usually connected by a flexible linker (5, 18, 39). CBM has high 75
specific capacities for cellulose binding. Previously, it has been reported that CBM is 76
able to be fused to a chosen target protein by genetic manipulation (46), resulting in 77
enhanced binding of this fusion protein to cellulose (7, 40). For example, fusion 78
proteins were constructed by fusing CBM to β-glucosenucleotide enzyme (GUS) (14) or 79
β-glycosidase (BglA) (20), which facilitates biochemical analysis of scouring efficiency 80
for cotton fabrics. 81
In the present study, the CBM from T. fusca cellulase Cel6A (abbreviated as 82
CBMCel6A) and the CBM from C. fimi cellulase CenA (abbreviated as CBMCenA) were 83
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

5
fused, separately, to the carboxyl terminus of T. fusca cutinase. The resulting fusion 84
enzymes were compared with the native cutinase in terms of their biochemical 85
properties as well as the catalytic efficiency in cutin breakdown on cotton fiber. This is 86
the first report of improving the scouring efficiency of cutinase by fusing it with CBM. 87
88
MATERIALS AND METHODS 89
Bacterial strains, plasmids, and culture conditions 90
The strain T. fusca, plasmid pET20b(+)-Tfu_0883, and pBSK-CBMCenA were lab 91
stocks (4,11). Plasmid pET20b(+)-Tfu_0883 was used as the gene source of T. fusca 92
cutinase. The strain T. fusca and plasmid pBSK-CBMCenA were used as the gene source 93
of CBMCel6A and CBMCenA, respectively. E. coli strains BL21 (DE3) was used as the 94
expression host, and pET20b(+) was used as the cloning and expression vector. Cells 95
were grown in Luria-Bertani medium at 37 °C and, if necessary, ampicillin in a final 96
concentration of 100 µg/ml was added to the medium. 97
Construction of cutinase-CBM fusion protein expression vectors 98
The gene encoding cutinase (NCBI accession number YP_288944) and CBMCenA 99
(NCBI accession number AAA23084.1) was amplified using plasmid 100
pET20b-Tfu_0883 and pBSK-CBMCenA as the template, respectively. The gene 101
encoding CBMCel6A and its linker (NCBI accession number YP_289135) was amplified 102
from T. fusca genomic DNA. Overlapping PCR was used to fuse the T. fusca CBMCel6A 103
or C. fimi CBMCenA to the C-terminus of T. fusca cutinase. Sequences of the primers are 104
given in Table 1. The procedure of fusion is shown in Figure 1. An NcoI restriction site 105
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

6
(boldface) was introduced at the 5' end of P1 while an EcoR1 restriction site (boldface) 106
was introduced at the 3' end of P4/P4'. The overlapping area was underlined. The 107
primers were synthesized by Shanghai Sangon Biological Engineering Technology & 108
Services Co. Ltd. 109
The PCR was performed using 35 successive cycles as following: denaturation at 110
94 °C for 0.5 min, annealing at 56 °C for 0.5 min, and primer extension at 72 °C for 1.5 111
min. PrimeSTAR HS DNA Polymerase was utilized during the process. The 112
amplification product was isolated and ligated into the vector pMD18T-simple. The 113
ligation mixture was used to transform chemically competent E. coli JM109. The 114
plasmid isolated from these transformants was verified by restriction analysis and the 115
gene sequence was confirmed by DNA sequencing. The plasmids with the correct 116
sequences of cutinase-CBMCel6A and cutinase-CBMCenA were named as 117
pMD18T/cutinase-CBMCel6A and pMD18T/cutinase-CBMCenA, respectively. The 118
resulting plasmids were digested with NcoI and EcoRI and ligated into the similarly 119
digested expression vector pET20b(+). The ligation mixture was used to transform 120
chemically competent E.coli JM109 cells. The plasmid isolated from these 121
transformants was verified by restriction analysis and the gene sequence was confirmed 122
again by DNA sequencing. The plasmids with the correct sequences of 123
cutinase-CBMCel6A and cutinase-CBMCenA were named as pET20b/cutinase-CBMCel6A 124
and pET20b/cutinase-CBMCenA, respectively. 125
Enzymes used for DNA manipulations were purchased from TakaRa 126
Biotechnology Co. Ltd. DNA sequencing was performed by Shanghai Generay 127
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

7
Biotechnology Co. Ltd. Genomic DNA extraction was performed following the method 128
of Sambrook et al. (1989). Plasmid DNA was extracted by using the Sangon EZ-10 Spin 129
Column Plasmid Mini-Preps kit. Plasmid and PCR products were recovered from 130
agarose gel using Sangon purification kit. 131
Expression and purification of the cutinase-CBM fusion proteins 132
E.coli BL21(DE3) cells harboring pET20b/cutinase-CBMCel6A or 133
pET20b/cutinase-CBMCenA were grown in TB medium containing ampicillin (100 134
µg/ml) at 37 °C. When the culture reached an A600 of 1.5 to 2.0, 135
isopropyl-1-thio-β-D-galactopyranoside (IPTG) was added to a final concentration of 136
0.5 mM. The culture after 18 h of induction was centrifuged (10,000 × g, 30 min, 4 137
°C) and the supernatant was collected. 138
The above supernatant was treated with 70% (w/v) saturated ammonium sulphate 139
solution and the solution was kept at 4 °C overnight. Precipitates were collected by 140
centrifugation and dissolved in 50 ml buffer A (20 mM sodium phosphate, 0.5 M NaCl, 141
and 20 mM imidazole, pH 7.4). The solutions were subsequently dialyzed against 2 142
liters of buffer A overnight and applied to a nickel affinity column pre-equilibrated with 143
buffer A. The samples were allowed to bind with Ni-NTA agarose at a flow rate of 1 144
ml/min, followed by washing with buffer A until the UV baseline was reached. Elution 145
was performed using a linear gradient from 0 to 500 mM imidazole in buffer A over 100 146
min. The fractions containing p-nitrophenyl butyrate (pNPB) hydrolase activity were 147
pooled and dialyzed against 2 liters of buffer B (20 mM Tris-HCl, pH 8.0) at 4 °C 148
overnight. The purified enzyme was concentrated by ultrafiltration and stored at -80°C. 149
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

8
Enzyme characterization of cutinase-CBMs 150
For enzyme characterization, all the values presented in graphs and tables are the 151
means of three replications. Esterase activity, cutinase activity, and lipase activity were 152
assayed as described previously (11), with pNPB, cutin and triolein as the substrates, 153
respectively. 154
Temperature optima of the fusion and native enzymes were measured at 155
temperatures ranging between 20-70 °C. The reaction was performed in a buffer 156
containing 20 mM Tris-HCl, 10 mM NaCl, and 50 mM sodium taurodeoxycholate at pH 157
8.0 using pNPB as substrates. Since the pH of Tris buffer is temperature dependent, the 158
buffers were adjusted to pH 8.0 at the desired temperature. Enzyme activity was 159
measured by pre-incubating the buffer at a desired temperature for 2 min to allow it to 160
reach the final pH of 8.0. The thermostability of the enzymes was determined by 161
incubating the enzymes in 20 mM Tris-HCl (pH 8.0) at 50 °C. At different intervals, 162
samples were taken and assayed for residual activity using pNPB as the substrate. 163
pH optima of the fusion and native enzymes was investigated between pH 6.0 to 164
9.0 using either potassium phospate buffer (pH 6.0-7.0) or Tris-HCl buffer (pH 7.0-9.0). 165
To determine the pH stability, 20 mM concentrations of the following buffers were used: 166
sodium acetate (pH 4.0-6.0), potassium phospate (pH 6.0-7.0), Tris-HCl (pH 7.0-9.0), 167
and glycine-NaOH (pH 9.0-11.0). The enzymes were pre-incubated in the various 168
buffers at 37 °C for 24 h, followed by the determination of residual activity using pNPB 169
as the substrate. 170
The Michaelis-Menten parameters, Vmax and Km, were determined from 171
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

9
Michaelis-Menten plots of specific activities at various substrate concentrations. Rates 172
were measured in triplicate using pNPB (100-2000 µM) as the substrate by continuous 173
spectrophotometric analysis. Initial reaction velocities were calculated from the linear 174
region (~60s) of the reaction progress curve and measured in triplicate by varying the 175
concentration of the substrate. Apparent kinetic constant Km was calculated using the 176
Graph Pad Prism program. 177
Protein quantification and SDS-PAGE analysis 178
Protein concentrations were determined using the Bio-Rad protein assay kit 179
(Bio-Rad), with purified bovine serum albumin (Promega) as the standard. Sodium 180
dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was performed in 10% 181
acrylamide gels and the proteins were visualized by staining with Coomassie brilliant 182
blue R-250. 183
Binding and catalytic activity of cutinase-CBMs towards cotton fiber 184
The adsorption property of the enzymes was tested with cotton fiber as described 185
previously with minor modifications (4). Prior to treatment, raw cotton fiber was boiled 186
for 1 min, and then dried at 40 °C overnight. 1 g of pretreated cotton fiber was mixed 187
with 0.1% penetrant and 0.5% bovine serum albumin (BSA) in Tris-HCl buffer (20 mM, 188
pH 8.0) at 25 °C. The mixture was incubated for 30 min to avoid non-specific binding. 189
Equal units of native cutinase or fusion enzyme (10 U/ml toward pNPB) was then added 190
to the above solution and incubated for another 1 h with shaking at 60 rpm. The reaction 191
mixtures were centrifuged at 3,000 ×g for 2 min and the amount of unbound enzyme 192
was estimated from the residual activity in the supernatant. The amount of cotton 193
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

10
fiber-bound enzyme was calculated from the difference between the initial enzyme 194
activity and unbound enzyme activity. Control assays were performed under the same 195
conditions, except in the absence of raw cotton fiber. Pectinase was added when needed. 196
The desorption property of the enzymes was performed according to a method 197
described previously (4). After 2 h of incubation, the above reaction solution was 198
diluted (1:20) with Tris-HCl buffer and the mixture was incubated for another 60 min 199
with shaking at 200 rpm at 25 °C. The amount of desorbed enzyme was estimated from 200
the enzymatic activity in the supernatant. All assays were performed in triplicate. 201
For the determination of released fatty acids catalyzed by cutinase or 202
cutinase-CBM, a 10 ml reaction mixture containing 1 g of raw cotton fiber and 50 µM 203
of native or fusion cutinase was incubated in 20 mM Tris-HCl buffer with or without 204
pectinase. The reaction mixture was shaken at 200 rpm for 18 h at 50 °C. At various 205
times, samples were removed and subjected to titration using 20 mM NaOH. 206
207
RESULTS 208
Construction and purification of cutinase-CBM fusion proteins 209
Cutinase-CBM fusion proteins were generated by fusing either the CBM of 210
cellulase Cel6A from T. fusca (CBMCel6A) or the CBM of cellulase CenA from C. fimi 211
(CBMCenA) to the C-terminus of T. fusca cutinase through overlapping PCR 212
amplification. The fused genes were subsequently inserted into the expression vector 213
pET-20b(+) which contains a C-terminal His6 tag and an N-terminal signal peptide PelB 214
to allow the expressed proteins to be secreted. The resulting constructs 215
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

11
pET20b/cutinase-CBMCel6A and pET20b/cutinase-CBMCenA were used for protein 216
expression in E.coli BL21(DE3). The pNPB hydrolyzing activity in the culture 217
supernatant was 94 U/ml for cutinase-CBMCel6A and 76 U/ml for cutinase-CBMCenA, 218
which was 230-fold and 190-fold to that of the control culture (in which E.coli cells 219
carried the vector pET-20b(+)), respectively. Both fusion enzymes were purified by 220
ammonium sulfate fraction and Ni-Sepharose affinity chromatography (Supplemental 221
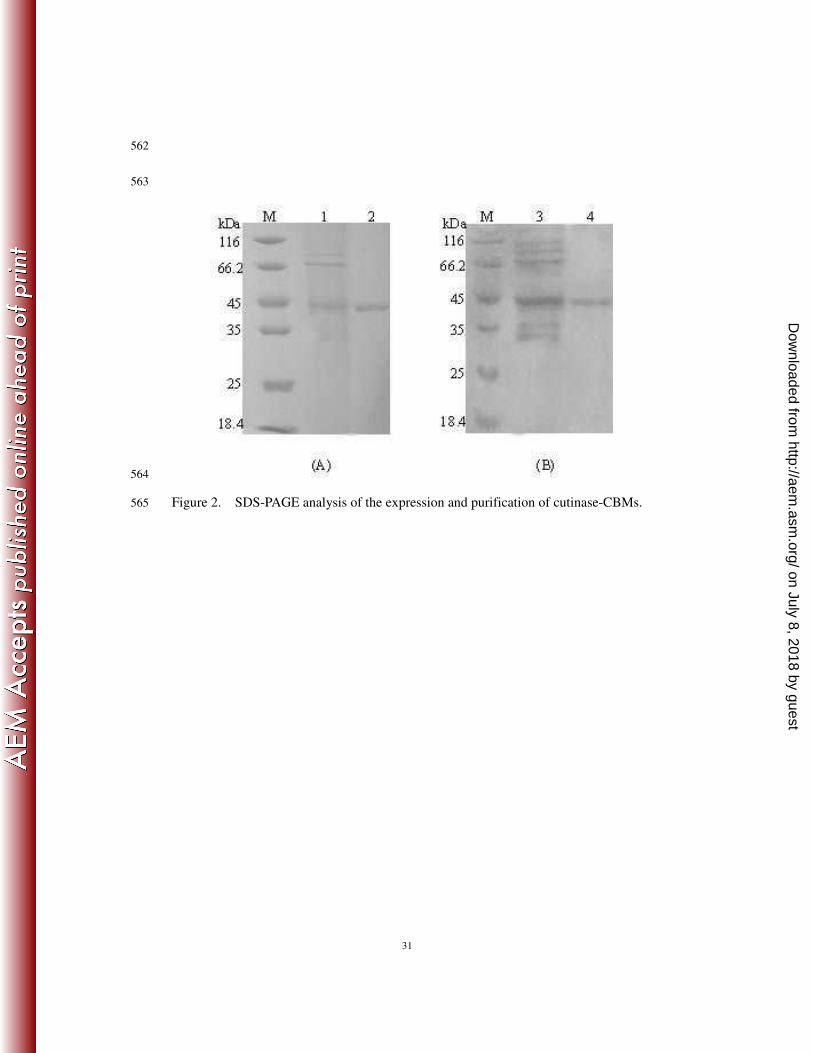
Table S1 and S2). SDS-PAGE results demonstrated that they were purified to 222
homogeneity with the same molecular mass of about 45 kDa (Figure 2). In addition, the 223
purified enzymes were active with a specific activity of 20000 U/µmol protein 224
(Supplemental Table S1) for cutinase-CBMCel6A and 15000 U/µmol protein 225
(Supplemental Table S2) for cutinase-CBMCenA. 226
Enzymatic properties of cutinase-CBM fusion proteins 227
The optimal temperature and pH of the fusion enzymes were determined at a 228
temperature range of 20-70 °C (Supplemental Figure S3A) and a pH range of 6-9 229
(Supplemental Figure S3C). For comparative purpose, T. fusca cutinase was also 230
subjected to similar analysis. The results showed that both cutinase-CBMCel6A and 231
cutinase-CBMCenA exhibited an optimal temperature at 50 °C, whereas the native 232
cutinase displayed an optimal temperature at 60 °C. Not surprisingly, all native and 233
fusion enzymes exhibited the same optimal pH of 8.0. Subsequently, the thermostability 234
was determined at the temperature of 50 °C (Supplemental Figure S3B), while the pH 235
stability was determined at pH values between 4 to 11 (Supplemental Figure S3D). The 236
half-life of the native cutinase was 70 h at 50 °C, while those of cutinase-CBM fusion 237
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

12
enzymes were both 53 h at 50 °C. Similar pH stability at a pH range of 6-9 was 238
observed for all three enzymes. 239
Previously, it has been shown that T. fusca cutinase has broad substrate specificity 240
against cutin, soluble esters and insoluble triglycerides (11). The activity of 241
cutinase-CBMs towards soluble ester had been confirmed in the above purification and 242
characterization analysis using pNPB as a substrate. For the insoluble triglycerides, both 243
cutinase-CBMCel6A and cutinase-CBMCenA were found to be capable of hydrolyzing 244
triolein with specific activities corresponding to 128% and 111% of the native cutinase, 245
respectively. Furthermore, their cutin hydrolyzing activities were evaluated under their 246
individual optimal temperature and pH. As shown in Table 2, the C16 and C18 family 247
fatty acid monomers released after enzymatic reaction were 61% for cutinase-CBMCel6A, 248
64% for cutinase-CBMCenA, and 59% for the native cutinase. The hydroxy fatty acids, 249
which are specific in cutin, were 3.6% for cutinase-CBMCel6A, 3.7% for 250
cutinase-CBMCenA, and 3.1% for the native cutinase. These results demonstrated that 251
both cutinase-CBMCel6A and cutinase-CBMCenA can hydrolyze cutin as efficiently as the 252
native cutinase. 253
The kinetics of the fusion enzymes was analyzed using pNPB as the substrate 254
(Table 3). Their Km values were similar to that of the native cutinase, while their 255
catalytic efficiency (Kcat/Km) was about 94% (cutinase-CBMCel6A) and 85% 256
(cutinase-CBMCenA) of the native cutinase. 257
Binding and hydrolytic activity of cutinase-CBMs towards cotton fiber 258
Adsorption of the enzyme on the surface of cotton fiber is the first step for cutinase 259
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

13
to perform its hydrolysis towards cutin (47). The binding experiments (Figure 3A) were 260
performed under the conditions with or without the presence of pectinase, an enzyme 261
utilized to remove pectin in bioscouring. The results showed that, compared with T. 262
fusca cutinase, the binding of cutinase-CBMCel6A was enhanced by 2% in the absence of 263
pectinase and 40% in the presence of pectinase, while the binding of cutinase-CBMCenA 264
was enhanced by 28% in the absence of pectinase and 45% in the presence of pectinase. 265
After dilution and re-equilibration, almost all of the bound enzymes were desorbed. 266
Thus, the binding of both cutinase and cutinase–CBMs with cotton fiber appeared to be 267
reversible. 268
In addition to binding, their hydrolytic efficiency towards cotton fiber was also 269
compared (Figure 3B). In the absence of pectinase, the amount of released fatty acids 270
was similar to that of the native cutinase for cutinase-CBMCel6A, but was 1.8-fold higher 271
for cutinase-CBMCenA. In the presence of pectinase, however, both fusion enzymes 272
released almost the same amount of fatty acids and exhibited a catalytic efficiency 273
3-fold higher than that of the native cutinase. This result is consistent with that of the 274
above binding experiment. The increased binding capability resulted in enhanced cutin 275
hydrolytic efficiency. 276
277
DISCUSSION 278
Previously, site-directed mutagenesis of the amino acid residues surrounding the 279
active site has been performed in order to obtain higher catalytic efficiency for cutinase 280
(3). In the present study, a fusion protein approach in which cutinase was fused with 281
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

14
CBM for improved affinity to cotton fiber was developed. By taking advantage of the 282
CBM’s specific binding to cellulose on the surface of cotton fiber where cutin also 283
exists, the concentration of fusion cutinase around cutin would be increased, which may 284
then result in enhanced enzyme catalytic efficiency due to proximity effect. Considering 285
the conditions of textile scouring process, the cutinase-CBM fusion protein has to meet 286
the following requirements: (1) no significantly decreased cutin hydrolyzing activity 287
when comparing with the native cutinase; (2) good thermal stability and alkali 288
resistance; (3) good binding affinity to cotton fiber; and (4) improved scouring effect for 289
cotton fiber. 290
In order to meet the above requirements, the choice of a suitable CBM is critical. 291
To date, characteristics of CBMs have been explored extensively (23, 45). They are 292
found mainly in carbohydrate degrading enzymes from fungi (e.g. Trichoderma reesei) 293
and bacteria (e.g. Cellulomonas fimi), including cellulase, xylanase, mannanase and a 294
number of non-hydrolytic proteins (33). Considering the source of the cutinase used in 295
the present study, naturally a CBM was first selected from the genome of T. fusca. 296
As identified by the CAZy ModO database (http://www.afmb.cnrs-mrs.fr/CAZY/), 297
the genome of T. fusca encodes a total of 17 hydrolytic enzymes that possess a CBM 298
(supplemental Table S3), with 6 of them experimentally characterized (underlined in 299
supplemental Table S3) (36). In addition, CBMs are divided into 59 different families 300
(http://www.afmb.cnrs-mrs.fr/CAZY/) and those from the second family were found to 301
be able to ease crystalline cellulose without significant fiber damage (10). Among the 302
six characterized CBMs from T. fusca, except for the CBM3 of Cel9A and the CBM4 of 303
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

15
Cel9B, all the others were identified to belong to the second family and thus were 304
candidates for fusing with cutinase. 305
As for the directionality of the fusion, the homology structural model of cutinase 306
showed that both the N- and C-termini of the enzyme were exposed to the solvent side 307
(11). Considering the presence of the N-terminal PelB signal sequence for 308
transmembrane localization, it appears that the CBM is better to be fused to the 309
C-terminus of cutinase. Another consideration is a possible linker sequence between the 310
cutinase and the CBM. For example, it has been reported that the linker between the 311
CBM and catalytic domain of a bacterial cellulase is composed entirely of the Pro-Thr 312
repetitive sequence (19). Such a linker in cellulases would possess certain flexibility and 313
avoid possible structural hindrance, which ensures the uniform movement of the two 314
domains on the fiber surface (19, 39, 44). Therefore, an appropriate linker between the 315
cutinase and the CBM is desired. 316
Putting together the above considerations, the CBM from T. fusca cellulase Cel6A, 317
which belongs to the second family of CBMs and has a 28-residue linker region, was 318
chosen to be fused to the C-terminus of cutinase. 319
Cutinase-CBMCel6A was well expressed and purified, and shown to be able to 320
hydrolyze not only cutin but also insoluble triglycerides (triolein) and soluble esters 321
(pNPB). In addition, it shares similar pH stability as cutinase and displayed optimal 322
temperature at 50°C and half-life of 53 h at 50 °C. When their binding and catalytic 323
efficiency towards cotton fiber were compared, cutinase-CBMCel6A did not appear to 324
have a significantly better performance than the native cutinase. However, in the 325
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

16
presence of pectinase, cutinase-CBMCel6A exhibited significant improvement in binding 326
and a dramatic 3-fold increase in catalytic efficiency. This sharp contrast is likely 327
because most of the cellulose on the surface of cotton fiber is not well exposed to the 328
solvent and is embedded in the epidermis full of pectins, proteins and other components, 329
thus limiting the binding of CBM to cotton fiber. When pectinase was added in the 330
reaction mixture, removal of pectin by this enzyme may lead to the exposure of 331
cellulose, resulting in increased adsorption of cutinase-CBMCel6A, which eventually led 332
to higher scouring efficiency of cotton fiber. 333
In addition to CBMCel6A, we also examined the possibility of using other CBMs 334
that have been experimentally characterized. The CBM of endoglucanase A (CenA) 335
from C. fimi, which also belongs to the second family of CBMs, was shown to have 336
high affinity to cellulose (13, 16, 49) and appears to be a suitable candidate. 337
Subsequently, CBMCenA was fused to the C-terminus of cutinase using the same linker 338
from T. fusca Cel6A. As expected, cutinase-CBMCenA displayed similar substrate 339
specificity and catalytic properties as the native cutinase. Interestingly, although the 340
CBMCenA is from a mesophilic bacterium, this fusion enzyme still retained decent 341
thermostability, which may be due to the presence of a disulfide bond in CBMCenA (16). 342
Notably, significant improvement in the binding and catalytic efficiency towards cotton 343
fiber was observed for cutinase-CBMCenA when compared to the native cutinase and 344
cutinase-CBMCel6A. In addition, similar to cutinase-CBMCel6A, strong synergistic effect 345
with pectinase was also observed with cutinase-CBMCenA. Thus, it appears that the 346
scouring effect of cutinase-CBMCenA is better than cutinase-CBMCel6A. 347
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

17
In conclusion, cutinase-CBM fusion proteins were successfully created by fusing a 348
CBM to the C-terminus of T. fusca cutinase. Compared with the native cutinase, both 349
fusion proteins, cutinase-CBMCel6A and cutinase-CBMCenA, share similar stability and 350
catalytic properties, but showed greatly enhanced binding and hydrolytic activity 351
towards cotton fiber. These improvements as well as the synergistic effect between the 352
fusion proteins and the pectinase suggests that they may have better application 353
potential in textile bioscouring. 354
355
ACKNOWLEDGMENT 356
This work was supported financially by the National High-tech Research and 357
Development Program of China (2009AA02Z204), the National Natural Science 358
Foundation of China (30970057), the National Outstanding Youth Foundation of China 359
(20625619), Research Program of State Key Laboratory of Food Science and 360
Technology (SKLF-MB-200802), the Key Program of National Natural Science 361
Foundation of China (20836003), Program of Innovation Team of Jiangnan University 362
(2008CXTD01) and the Self-determined Research Program of Jiangnan University (Yao 363
Zhang)364
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

18
REFERENCE 365
1. Agrawal*, P. B., V. A. Nierstrasz, G. H. Bouwhuis, and M. M. C. G. 366
Warmoeskerken. 2008. Cutinase and pectinase in cotton bioscouring: an 367
innovative and fast bioscouring process. Biocatalysis and Biotransformation 368
26:412-421. 369
2. Araujo, R., M. Casal, and A. Cavaco-Paulo. 2008. Application of enzymes 370
for textile fibres processing. Biocatal Biotransfor 26:332-349. 371
3. Araujo, R., C. Silva, A. O'Neill, N. Micaelo, G. Guebitz, C. M. Soares, M. 372
Casal, and A. Cavaco-Paulo. 2007. Tailoring cutinase activity towards 373
polyethylene terephthalate and polyamide 6,6 fibers. J Biotechnol 128:849-57. 374
4. Azevedo, H., D. Bishop, and A. Cavaco-Paulo. 2000. Effects of agitation 375
level on the adsorption, desorption, and activities on cotton fabrics of full 376
length and core domains of EGV (Humicola insolens) and CenA 377
(Cellulomonas fimi). Enzyme Microb Technol 27:325-329. 378
5. Black, G. W., J. E. Rixon, J. H. Clarke, G. P. Hazlewood, L. M. Ferreira, D. 379
N. Bolam, and H. J. Gilbert. 1997. Cellulose binding domains and linker 380
sequences potentiate the activity of hemicellulases against complex substrates. 381
J Biotechnol 57:59-69. 382
6. Blanco, J., J. J. Coque, J. Velasco, and J. F. Martin. 1997. Cloning, 383
expression in Streptomyces lividans and biochemical characterization of a 384
thermostable endo-beta-1,4-xylanase of Thermomonospora alba ULJB1 with 385
cellulose-binding ability. Appl Microbiol Biotechnol 48:208-17. 386
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

19
7. Bolam, D. N., A. Ciruela, S. McQueen-Mason, P. Simpson, M. P. 387
Williamson, J. E. Rixon, A. Boraston, G. P. Hazlewood, and H. J. Gilbert. 388
1998. Pseudomonas cellulose-binding domains mediate their effects by 389
increasing enzyme substrate proximity. Biochem J 331 ( Pt 3):775-81. 390
8. Carvalho, C. M., M. R. Aires-Barros, and J. M. Cabral. 1999. Cutinase: 391
from molecular level to bioprocess development. Biotechnol Bioeng 66:17-34. 392
9. Cavaco-Paulo, A. 1998. Processing textile fibers with enzymes: an overview. 393
ACS Symposium Series:180-189. 394
10. Cavaco-Paulo, A., J. Morgado, J. Andreaus, and D. Kilburn. 1999. 395
Interactions of cotton with CBD peptides. Enzyme and Microbial Technology 396
25:639-643. 397
11. Chen, S., X. Tong, R. W. Woodard, G. Du, J. Wu, and J. Chen. 2008. 398
Identification and characterization of bacterial cutinase. J Biol Chem 399
283:25854-62. 400
12. Ciolacu, D., J. Kovac, and V. Kokol. 2010. The effect of the 401
cellulose-binding domain from Clostridium cellulovorans on the 402
supramolecular structure of cellulose fibers. Carbohydr Res 345:621-630. 403
13. Damude, H. G., S. G. Withers, D. G. Kilburn, R. C. Miller, Jr., and R. A. 404
Warren. 1995. Site-directed mutation of the putative catalytic residues of 405
endoglucanase CenA from Cellulomonas fimi. Biochemistry 34:2220-4. 406
14. Degani, O., S. Gepstein, and C. G. Dosoretz. 2004. A new method for 407
measuring scouring efficiency of natural fibers based on the cellulose-binding 408
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

20
domain-beta-glucuronidase fused protein. J Biotechnol 107:265-73. 409
15. Degani, O., S. Gepstein, and C. G. Dosoretz. 2002. Potential use of cutinase 410
in enzymatic scouring of cotton fiber cuticle. Appl Biochem Biotechnol 411
102-103:277-89. 412
16. Din, N., I. J. Forsythe, L. D. Burtnick, N. R. Gilkes, R. C. Miller, Jr., R. A. 413
Warren, and D. G. Kilburn. 1994. The cellulose-binding domain of 414
endoglucanase A (CenA) from Cellulomonas fimi: evidence for the 415
involvement of tryptophan residues in binding. Mol Microbiol 11:747-55. 416
17. Fett, W. F., H. C. Gerard, R. A. Moreau, S. F. Osman, and L. E. Jones. 417
1992. Screening of Nonfilamentous Bacteria for Production of 418
Cutin-Degrading Enzymes. Appl Environ Microbiol 58:2123-2130. 419
18. Gilkes, N. R., B. Henrissat, D. G. Kilburn, R. C. Miller, Jr., and R. A. 420
Warren. 1991. Domains in microbial beta-1, 4-glycanases: sequence 421
conservation, function, and enzyme families. Microbiol Rev 55:303-15. 422
19. Gilkes, N. R., D. G. Kilburn, R. C. Miller, Jr., and R. A. Warren. 1989. 423
Structural and functional analysis of a bacterial cellulase by proteolysis. J Biol 424
Chem 264:17802-8. 425
20. Ha, J. S., Y. M. Lee, S. L. Choi, J. J. Song, C. S. Shin, J. H. Kim, and S. G. 426
Lee. 2008. Thermostable beta-glycosidase-CBD fusion protein for 427
biochemical analysis of cotton scouring efficiency. J Microbiol Biotechnol 428
18:443-8. 429
21. Hardin, I. R., Li, Y., Akin, D. 1998. Cotton wall structure and enzymatic 430
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

21
treatments. ACS Sysmposium Series 687:190-203. 431
22. Hartzell, M. M., Hsieh, Y.L. 1998. Enzymatic scouring to improve cotton 432
fabric wettability. J.Textile Res. 68:233-241. 433
23. Hashimoto, H. 2006. Recent structural studies of carbohydrate-binding 434
modules. Cell Mol Life Sci 63:2954-67. 435
24. Hilge, M., S. M. Gloor, W. Rypniewski, O. Sauer, T. D. Heightman, W. 436
Zimmermann, K. Winterhalter, and K. Piontek. 1998. High-resolution 437
native and complex structures of thermostable beta-mannanase from 438
Thermomonospora fusca - substrate specificity in glycosyl hydrolase family 5. 439
Structure 6:1433-44. 440
25. Irwin, D., E. D. Jung, and D. B. Wilson. 1994. Characterization and 441
sequence of a Thermomonospora fusca xylanase. Appl Environ Microbiol 442
60:763-70. 443
26. Irwin, D., D. H. Shin, S. Zhang, B. K. Barr, J. Sakon, P. A. Karplus, and D. 444
B. Wilson. 1998. Roles of the catalytic domain and two cellulose binding 445
domains of Thermomonospora fusca E4 in cellulose hydrolysis. J Bacteriol 446
180:1709-14. 447
27. Irwin, D. C., M. Cheng, B. Xiang, J. K. Rose, and D. B. Wilson. 2003. 448
Cloning, expression and characterization of a family-74 xyloglucanase from 449
Thermobifida fusca. Eur J Biochem 270:3083-91. 450
28. Irwin, D. C., S. Zhang, and D. B. Wilson. 2000. Cloning, expression and 451
characterization of a family 48 exocellulase, Cel48A, from Thermobifida fusca. 452
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

22
Eur J Biochem 267:4988-97. 453
29. Jung, E. D., G. Lao, D. Irwin, B. K. Barr, A. Benjamin, and D. B. Wilson. 454
1993. DNA sequences and expression in Streptomyces lividans of an 455
exoglucanase gene and an endoglucanase gene from Thermomonospora fusca. 456
Appl Environ Microbiol 59:3032-43. 457
30. Jung, H., D. B. Wilson, and L. P. Walker. 2003. Binding and reversibility of 458
Thermobifida fusca Cel5A, Cel6B, and Cel48A and their respective catalytic 459
domains to bacterial microcrystalline cellulose. Biotechnol Bioeng 84:151-9. 460
31. Krakhmalev, V. A., Paiziev,A.A. 2006. Spiral structures of cotton fiber. 461
Cellulose 13:45-52. 462
32. Lao, G., G. S. Ghangas, E. D. Jung, and D. B. Wilson. 1991. DNA 463
sequences of three beta-1,4-endoglucanase genes from Thermomonospora 464
fusca. J Bacteriol 173:3397-407. 465
33. Levy, I., and O. Shoseyov. 2002. Cellulose-binding domains: 466
biotechnological applications. Biotechnol Adv 20:191-213. 467
34. Li, Y., D. C. Irwin, and D. B. Wilson. 2007. Processivity, substrate binding, 468
and mechanism of cellulose hydrolysis by Thermobifida fusca Cel9A. Appl 469
Environ Microbiol 73:3165-72. 470
35. Linder, M., J. Winiecka-Krusnell, and E. Linder. 2002. Use of recombinant 471
cellulose-binding domains of Trichoderma reesei cellulase as a selective 472
immunocytochemical marker for cellulose in protozoa. Appl Environ 473
Microbiol 68:2503-8. 474
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

23
36. Lykidis, A., K. Mavromatis, N. Ivanova, I. Anderson, M. Land, G. 475
DiBartolo, M. Martinez, A. Lapidus, S. Lucas, A. Copeland, P. Richardson, 476
D. B. Wilson, and N. Kyrpides. 2007. Genome sequence and analysis of the 477
soil cellulolytic actinomycete Thermobifida fusca YX. J Bacteriol 478
189:2477-86. 479
37. Nini, L., L. Sarda, L. C. Comeau, E. Boitard, J. P. Dubes, and H. 480
Chahinian. 2001. Lipase-catalysed hydrolysis of short-chain substrates in 481
solution and in emulsion: a kinetic study. Biochim Biophys Acta 1534:34-44. 482
38. Posta, K., E. Beki, D. B. Wilson, J. Kukolya, and L. Hornok. 2004. Cloning, 483
characterization and phylogenetic relationships of cel5B, a new endoglucanase 484
encoding gene from Thermobifida fusca. J Basic Microbiol 44:383-99. 485
39. Quentin, M., M. Ebbelaar, J. Derksen, C. Mariani, and H. van Der Valk. 486
2002. Description of a cellulose-binding domain and a linker sequence from 487
Aspergillus fungi. Appl Microbiol Biotechnol 58:658-62. 488
40. Richins, R. D., A. Mulchandani, and W. Chen. 2000. Expression, 489
immobilization, and enzymatic characterization of cellulose-binding 490
domain-organophosphorus hydrolase fusion enzymes. Biotechnol Bioeng 491
69:591-6. 492
41. Sae-be, P., U. Sangwatanaroj, and H. Punnapayak. 2007. Analysis of the 493
products from enzymatic scouring of cotton. Biotechnol J 2:316-25. 494
42. Sakka, K., G. Takada, S. Karita, and K. Ohmiya. 1996. Identification and 495
characterization of cellulose-binding domains in xylanase A of Clostridium 496
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

24
stercorarium. Ann N Y Acad Sci 782:241-51. 497
43. Sawada, K., and M. Ueda. 2001. Enzyme processing of textiles in reverse 498
micellar solution. J Biotechnol 89:263-9. 499
44. Shen, H., M. Schmuck, I. Pilz, N. R. Gilkes, D. G. Kilburn, R. C. Miller, 500
Jr., and R. A. Warren. 1991. Deletion of the linker connecting the catalytic 501
and cellulose-binding domains of endoglucanase A (CenA) of Cellulomonas 502
fimi alters its conformation and catalytic activity. J Biol Chem 266:11335-40. 503
45. Shoseyov, O., Z. Shani, and I. Levy. 2006. Carbohydrate binding modules: 504
biochemical properties and novel applications. Microbiol Mol Biol Rev 505
70:283-95. 506
46. Tomme, P., A. Boraston, B. McLean, J. Kormos, A. L. Creagh, K. Sturch, 507
N. R. Gilkes, C. A. Haynes, R. A. Warren, and D. G. Kilburn. 1998. 508
Characterization and affinity applications of cellulose-binding domains. J 509
Chromatogr B Biomed Sci Appl 715:283-96. 510
47. Tomme, P., D. P. Driver, E. A. Amandoron, R. C. Miller, Jr., R. Antony, J. 511
Warren, and D. G. Kilburn. 1995. Comparison of a fungal (family I) and 512
bacterial (family II) cellulose-binding domain. J Bacteriol 177:4356-63. 513
48. Walton, T. J., and P. E. Kolattukudy. 1972. Determination of the structures 514
of cutin monomers by a novel depolymerization procedure and combined gas 515
chromatography and mass spectrometry. Biochemistry 11:1885-96. 516
49. Wong, W. K., B. Gerhard, Z. M. Guo, D. G. Kilburn, A. J. Warren, and R. 517
C. Miller, Jr. 1986. Characterization and structure of an endoglucanase gene 518
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

25
cenA of Cellulomonas fimi. Gene 44:315-24. 519
50. Zhang, S., G. Lao, and D. B. Wilson. 1995. Characterization of a 520
Thermomonospora fusca exocellulase. Biochemistry 34:3386-95. 521
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

26
Figure Legends 522
523
Figure 1. Construction of cutinase-CBM fusion proteins. (A) T. fusca cutinase was 524
joined with the CBM from T. fusca Cel6A using overlapping extension PCR. The 525
fusion of T. fusca cutinase with CBM from C. fimi CenA followed the similar strategy. 526
(B) Amino acid composition of the fusion proteins. 527
528
Figure 2. SDS-PAGE analysis of the expression and purification of cutinase-CBMs. 529
(A) cutinase-CBMCel6A. lane M, molecular mass markers; lane 1, culture supernatant 530
of cutinase-CBMCel6A; lane 2, purified cutinase-CBMCel6A. (B) cutinase-CBMCenA. 531
lane M, molecular mass markers; lane 1, culture supernatant of cutinase-CBMCenA; 532
lane 2, purified cutinase-CBMCenA. 533
534
Figure 3. Scouring effects of cutinase-CBMs and cutinase on cotton fiber. (A) 535
Adsorption on cotton fiber. Purified cutinase or cutinase-CBM fusion protein was 536
incubated in 20 mM Tris-HCl buffer (pH 8.0, 25 °C). without pectinase, with 537
pectinase. (B) Analysis of products released from cotton fiber. 50µM purified cutinase 538
or cutinase-CBM fusion protein was incubated in 20 mM Tris-HCl buffer (pH 8.0, 50 539
°C) within/without pectinase,and the amount of released fatty acids was measured by 540
titration with 0.02 N NaOH. ◆ cutinase-CBMCel6A alone, ◇ cutinase-CBMCel6A and 541
pectinase, ▲ cutinase-CBMCenA alone, △ cutinase-CBMCenA and pectinase, ■ cutinase 542
alone, □ cutinase and pectinase.543
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

27
544
545
Table 1. Primers used in the construction of the fusion genes 546
P1 : 5'-GGAATACCATATGTCCATGGCCAACCCCTACGAGCGCGG-3' (NcoI)
P2 : 5'-CGCGGCGATCGCCATGAACGGGCAGGTGGA-3'
P3 : 5'-TCCACCTGCCCGTTCATGGCGATCGCCGCG-3'
P4 : 5'-CATCTCGAGAGAATTCGGGCAGGTAAGGGTCGGAACAG-3' (EcoR1)
P2': 5'-GCGGCAGCCGGGAGCGGGAGGCGGCGTGGG-3'
P3': 5'-CCCACGCCGCCTCCCGCTCCCGGCTGCCGC-3'
P4': 5'-CATCTCGAGAGAATTCGGGGTGCCCGTGCAGGTGGTGC-3' (EcoR1)
Restriction enzyme sites are in bold and italicized. 15 base pairs of the overlapping regions are 547
underlined.548
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

28
549
Table 2. Monomeric products released from cutin hydrolysis by cutinase-CBMs and 550
cutinase. 551
Cutin hydrolysis by cutinase-CBM and cutinase was carried out in 25 mM potassium phosphate 552
buffer (pH 8.0) at 50 °C, for 18 h. Each value represents the mean of three independent 553
measurements, and the variation about the mean is below 5%. 554
555
Cutin hydrolysis products cutinase
hydrolysis
Area (%)
cutinase-CBMCel6A
hydrolysis
Area (%)
cutinase-CBMCenA
hydrolysis
Area (%)
Hexadecanoic acid 27.6 30.6 31.4
Octadecenoic acid 26.7 25.2 27.5
9-Octadecenoic acid 0.50 0.43 0.36
9,12-Octadecadienoic acid 1.08 1.02 1.01
16-Hydroxyhexadecanoic acid 0.54 0.54 0.58
18-Hydroxyoctadeca-9-enoic acid 1.01 0.87 0.91
18-Hydroxyoctadeca-9,12-dienoic acid 1.12 1.76 1.70
9,10,18-Trihydroxyoctadecanoic acid 0.47 0.47 0.51
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

29
556
557
558
Table 3. Kinetic parameters of cutinase-CBMs and cutinase 559
Kinetic parameters cutinase cutinase-CBMCel6A cutinase-CBMCenA
KmpNPB
(µM) 640 ± 40 620 ± 40 620 ± 30
Kcat(s-1
) 220 ± 10 200 ± 10 180 ± 10
Kcat/Km(s-1
) 0.34 0.32 0.29
560
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

30
561
cutinase-CBM /pET-20b
4.9kb
pelB
cutinase
NcoI
EcoR1His tagT7 terminator
f1 origin
Amp
ColE1 pBR322 origin
Linker
CBMCel6A
Cel6A
cutinase
P1
P2
cutinase
P1
P2
CD Linker CBMCel6A
P3
P4
Plasmid pET20b-Tfu_0883 Thermobifida fusca total DNA
cutinase
P1
Linker CBMCel6A
P4
PCR PCR
Overlapping PCR
pET20b(+) -NcoI/EcoRI
-Ligation
(A)
Linker CBMCel6Acutinase Linker CBMCel6Acutinasecutinase
(cutinase-CBMCel6A) ANPY…EVEEYRSTCP--------MAIA…PTPTPPP---------GSSG…NSNSVPTLTC
cutinase: 1~260aa Linker of CBMCel6A: 261-288aa CBMCel6A: 289~387aa
(cutinase-CBMCenA) ANPY…EVEEYRSTCP--------MAIA…PTPTPPP---------APGC…SLNGTTCTGT
cutinase: 1~260aa Linker of CBMCel6A: 261-288aa CBMCenA: 289~395aa
(B)
(cutinase-CBMCel6A) ANPY…EVEEYRSTCP--------MAIA…PTPTPPP---------GSSG…NSNSVPTLTC
cutinase: 1~260aa Linker of CBMCel6A: 261-288aa CBMCel6A: 289~387aa
(cutinase-CBMCenA) ANPY…EVEEYRSTCP--------MAIA…PTPTPPP---------APGC…SLNGTTCTGT
cutinase: 1~260aa Linker of CBMCel6A: 261-288aa CBMCenA: 289~395aa
(B)
Figure 1. Construction of cutinase-CBM fusion proteins.
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
31
562
563
564
Figure 2. SDS-PAGE analysis of the expression and purification of cutinase-CBMs.565
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

32
566
567
Figure 3. Scouring effects of cutinase-CBMs and cutinase on cotton fiber. 568
569
(A)
0
10
20
30
40
50
60 without pectinase
with pectinase
Bin
din
g r
atio
(%
)
cutinase cutinase-CBMCel6A
cutinase-CBMCenA
(B)
0 2 4 6 8 10 12 14 16 180
20
40
60
80
100
120
cutinase cutinase+pectinase
cutinase-CBMCel6A
cutinase-CBMCel6A
+pectinase
cutinase-CBMCenA
cutinase-CBMCenA
+pectinase
Fre
e f
atty
aci
ds
(µm
ol)
Time (h)
on July 8, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 2010, p. 7896 Vol. 76, No. 230099-2240/10/$12.00 doi:10.1128/AEM.02348-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
ERRATUM
Characterization of Thermobifida fusca Cutinase–Carbohydrate-Binding ModuleFusion Proteins and Their Potential Application in Bioscouring
Yao Zhang,1,2 Sheng Chen,1,2 Meng Xu,1,2 Artur Cavaco-Paulo,3 Jing Wu,1,2* and Jian Chen1,2*State Key Laboratory of Food Science and Technology1 and School of Biotechnology and Key Laboratory of
Industrial Biotechnology,2 Ministry of Education, Jiangnan University, Wuxi 214122, China, andDepartment of Textile Engineering, University of Minho, 4800 Guimaraes, Portugal3
Volume 76, no. 20, pages 6870–6876, 2010. Page 6870: The article byline should read as shown above.
7896