characterization of fn-c/h-v, a novel synthetic peptide from
TRANSCRIPT

Characterization of FN-C/H-V, a Novel Synthetic PeptideFrom Fibronectin That Promotes Rabbit Corneal EpithelialCell Adhesion, Spreading, and Motility
Daniel L. Mooradian,* James B. McCarthy* Amy P. N. Skubitz*]. Douglas Cameron,-\ and Leo T. Furcht*
Purpose. Fibronectin promotes corneal epithelial cell adhesion and motility in vitro and playsan important role in corneal re-epithelialization during corneal wound healing. Multiple do-mains contribute to the adhesion- and motility-promoting activity of fibronectin. The aim ofthis study was to identify amino acid sequences that contribute to the rabbit corneal epithelial(RCE) cell adhesion- and motility-promoting activity of the 33 and 66 kD carboxy-terminalheparin-binding fragments of fibronectin.
Methods. Synthetic peptides derived from the 33/66 kD fragments of fibronectin were testedfor their ability to directly promote RCE cell adhesion, spreading, and motility. To assess thecontribution of these peptides to the activity of fibroneclin and the 33/66 kD fragments offibronectin, synthetic peptides, and antibodies against these peptides were tested for theirability to block RCE cell adhesion, spreading, and motility.
Results. In this study, we identified a novel peptide sequence derived from the 33/66 kDfragments of fibronectin, FN-C/H-V (WQPPRARI), that directly promotes the adhesion,spreading, and migration of RCE cells in a concentration-dependent manner. A second pep-tide from the 33/66 kD fragments of fibronectin, FN-C/H-1V (SPPRRARVT), promoted RCEcell adhesion and spreading, but did not promote RCE cell migration. In contrast, two syn-thetic peptides from the 33/66 kD fragments of fibronectin that were previously shown topromote RCE cell adhesion (FN-C/H-I and FN-C/H-III) did not promote RCE cell spreadingor migration. Soluble FN-C/H-V inhibited RCE cell adhesion to surfaces coated with FN-C/H-V, the 33/66 kD fragments of fibronectin, and to fibronectin. In addition, polyclonal anti-FN-C/H-V IgG inhibited RCE cell adhesion to FN-C/H-V, the 33/66 kD fragments of fibronectin,and to fibronectin. Finally, polyclonal anti-FN-C/H-V IgG also inhibited RCE cell haptotacticmigration on the 33/66 kD fragments.
Conclusions. These data suggest that the amino acid sequence defined by peptide FN-C/H-Vcontributes to the adhesion-, spreading-, and motility-promoting activity of the 33/66 kDcarboxy-terminal heparin-binding fragments of fibronectin. Given the important role of fibro-nectin in corneal wound healing, these findings provide additional insight into the complexmolecular basis of corneal epithelial cell interactions with fibronectin and may be important inthe context of corneal wound healing. Invest Ophthalmol Vis Sci. 1993; 34:153-164.
From the ^Department of Laboratory Medicine ami Pathology/ A h e corneal epithelium provides a protective COVer-Biomedical Engineering Center, and ^Department of Ophthalmology, ing for the cornea and is essential for normal cornealUniversity of Minnesota Minneapolis Minnesota function. Failure of the cornea to re-epithelialize after/ his work was supported by National Institutes oj Health grants t Y- '06625 (LTF), AM-32660, and CA 43924 (JBM), and by an AmericanHeart Association—Minnesota Affiliate—Postdoctoral Fellowship(DLM). Reprint requests: Daniel L. Mooradian, University of Minnesota,Submitted for publication: March 13 1992; accepted July 16, 1992. Laboratory Medicine and Pathology, Box 609 UMHC, Minneapolis, MNProprietary Interest Category: N. 55455.
Investigative Ophthalmology & Visual Science, January 1993, Vol. 34, No. ICopyright © Association for Research in Vision and Ophthalmology 153
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018

154 Investigative Ophthalmology & Visual Science, January 1993, Vol. 34, No. 1
injury can result in persistent corneal defects and inulceration of the underlying corneal stroma.1 Fibro-nectin, a glycoprotein found in plasma and in extracel-lular matrices,2 plays a central role in corneal re-epi-thelialization. Fibronectin accumulates along with fi-brin at sites of corneal injury,3 providing a provisionalmatrix for epithelial cell adhesion and migration dur-ing wound healing. In addition, fibronectin promotesthe adhesion and migration of corneal epithelial cellsin vitro.4 Finally, the addition of exogenous fibronec-tin has been shown to promote corneal re-epitheliali-zation in vitro5 and in vivo6 and has been used clinicallyto treat persistent corneal epithelial defects.7-8
Plasma fibronectin is a large dimeric glycoproteincomposed of similar, but not identical polypeptide sub-units, termed the A and B chains, that are linked bydisulfide bonds.9 A number of functionally distinct do-mains of fibronectin have been identified (Fig. 1), in-cluding several domains that support cell adhe-sion.1011 The RGDS tetrapeptide sequence perhaps isthe most widely known.12 Rabbit corneal epithelial(RCE) cells adhere to a fragment of fibronectin thatcontains this sequence,4 and Nishida et al13 showedthat an RGDS-containing synthetic peptide can inhibitRCE cell adhesion to intact fibronectin. The RGDSsequence mediates cell adhesion via interactions withthe a5/31 integrin.14 Recent reports suggest that theactivity of the RGDS-containing central cell binding
domain of fibronectin may depend upon one or more"synergy" sites that contribute to full activity of thisdomain.15"17
The type III connecting segment, a region of al-ternative splicing,18 is a second cell-adhesion promot-ing domain in fibronectin,19 and two peptides withinthis domain also have been found to possess cell adhe-sion-promoting activity.20 Peptide CS-1, which con-tains a sequence (LDV) that promotes cell type-spe-cific adhesion,21 and peptide CS-5, which contains therelated sequence REDV,20 also promote cell adhesion.Cell adhesion to CS-1 and CS-5 is mediated by the«4j81 integrin.22'23
The 33/66 kD heparin-binding fragments fromthe carboxy-terminal third of the A chain and B chainof the fibronectin molecule also support cell adhe-sion.24 Previous studies have identified a number ofpeptide sequences within the carboxy-terminal hepa-rin-binding/cell adhesion-promoting domain thatpromote the adhesion of various cell types. Two ofthese peptides are FN-C/H-I25'26 and FN-C/H-II,26-28
which promote melanoma and neuroblastoma cell ad-hesion. Recent evidence suggests that melanoma cellsmay adhere to these sequences via a cell surface he-paran sulfate proteoglycan.29 In addition, FN-C/H-IIIrecently has been shown to support melanoma cell ad-hesion.30 Cell adhesion to these peptides is cell-typespecific and only two of these peptides (FN-C/H-I and
I II
27kD 46kD
III IV
75kD
(B cha in ) H2N
(A cha in ) H2N
V VI
66kD
CJ RGDS CJ
is *33kD 31 kD
33kD fragment(expanded)
type III(•12)
type type type II(CS)
1583—T(C)
FN-C/H-IIIFN-C/H-IV
|-2039
I I (T)
LCS-1I— FN-C/H-II— FN-C/H-I— FN-C/H-V
Ss s
COOH
FIGURE 1. Location of synthetic peptides within fibronectin. The 33/66 kD fragments offibronectin are generaied by trypsin (T) and cathepsin D (C) digestion of fibronectin. Hepa-rin binding domain (I). Collagen binding (noncovalent) domain (II). Free sulfhydryl (III).RGDS-containing central cell binding domain (IV). Carboxy-terminal heparin binding andcell adhesion-promoting domain (V). Free sulfhydryl (VI). The location of peptides FN-C/H-IV, FN-C/H-V, as well as the previously described peptides FN-C/H-I, FN-C/H-11, FN-C/H-III, and CS-1 within the 33 kD fragment also are shown.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018

Corneal Epithelial Cell Motility to Fibronectin 155
FN-C/H-III) have been shown to promote RCE celladhesion.31 In this study, we identify two additionalpeptide sequences derived from the 33/66 kD frag-ments of fibronectin—FN-C/H-IV (SPPRRARVT)and FN-C/H-V (WQPPRARI)—that promote RCEcell adhesion and spreading. In addition, we provideevidence that one of these peptides, FN-C/H-V, alsopromotes RCE cell haptotactic migration and contrib-utes to the adhesion- and motility-promoting activitiesof the 33/66 kD fragments of fibronectin.
MATERIALS AND METHODS
Corneal Epithelial Cell Culture
The experiments described in this study conformed tothe ARVO Resolution on the Use of Animals in Re-search and the University of Minnesota guidelines foranimal use. Corneal epithelial cell cultures were initi-ated as previously described.4 Briefly, 3-5 lb New Zea-land white rabbits (Birchwood Farms, Red Wing, MN)were killed and their eyes were enucleated, rinsed withHBSS (Hank's balanced salt solution), and transferredto HBSS containing 0.5 mg/ml gentamicin and 15mmol/1 HEPES. Corneas were excised and cut into1-2 mm2 blocks using a sterile razor blade. Theseblocks were incubated in dispase (1 U/ml, Grade II;Boehringer Mannheim, Indianapolis, IN) for 1 hr at25°C. The corneal epithelium was separated from theunderlying stroma with forceps and transferred to 60mm tissue culture dishes (Costar, Cambridge, MA)containing 0.5 ml of Dulbecco's modified Eagle's me-dium/Ham's nutrient mixture F-12 (DMEM/FIZ),10% fetal bovine serum (FBS), 5 Mg/ml insulin, 10 ng/ml epidermal growth factor, 0.5 Mg/ml hydrocorti-
sone, and 50 Mg/ml gentamicin. Epithelial explainswere allowed to attach for 24 hr at 37°C before addi-tional medium was added. Medium was changed everythird day. Epithelial cells migrated out of the cornealexplants and reached confluence within 10-14 days.Only primary cultures were used in these studies.
Isolation of Plasma Fibronectin and itsProteolytic Fragments
Human plasma fibronectin was purified as previouslydescribed.32 The 33/66 kD heparin-binding/cell adhe-sion-promoting fragments and the 27 kD ammo-ter-minal fragment (Fig. 1) were generated by proteolyticdigestion of intact fibronectin and purified as previ-ously described.25
Peptide Synthesis and Characterization
Synthetic peptides were synthesized in the Microche-mical Facility (University of Minnesota) with an Ap-plied Biosystems (Foster City, CA) peptide synthesizerusing previously described modifications of the Merri-field solid-phase method.33 Lyophilized crude pep-tides were purified by preparative reverse-phase highpressure liquid chromatography (HPLC) on a C-18column and were eluted with a linear gradient of aceto-nitrile (0-60%) containing 0.1% trifluoroacetic acid inwater. Peptide composition was verified by amino acidanalysis26 before use. Peptide purity was greater than95%, based on analytical HPLC. The sequences andselected properties of the synthetic peptides used inthis study are shown in Table 1 and Figure 1. Hydropa-thy indices were calculated using the method of Kyteand Doolittle.34
TABLE l. Synthetic Peptides From the 33/66 kD Fragmentsof FibronectinPeptideNomenclature Primary Sequence*
Hydropathy NetIndex\ ChargeX
FN-C/H-I YEKPGSPPREWPRPRPGV (residue nos. -24.3 +21906-1924)
FN-C/H-II KNNQKSEPLIGRKKT (residue nos. 1946-1960) -29.3 +4FN-C/H-III YRVRVTPKEKTGPMKE (residue nos. -23.7 +3
1721-1736)FN-C/H-IV SPPRRARVT (residue nos. 1784-1792) -12.2 +3FN-C/H-V WQPPRARI (residue nos. 1892-1899) -14.8 +2CS-1 DELPQLVTLPHPNLHGPEILDVPST -9.9 - 4
(residue nos. 1961-1985, 33 kD fragment only)
* The sequences shown use the single letter amino acid code: A, alanine; C, cysteine; D, aspartic acid; E,glutamic acid; F, phenylalanine; G, glycine; H, histidine; I, isoleucine; K, lysine; L, leucine; M, methio-nine; N, asparagine; P, proline; Q, glutamine; R, arginine; S, serine; T, threonine; V, valine; W, trypto-phan; Y, tyrosine.f Hydropathy indices were calculated by the method of Kyte and Doolittle.34 Using this method, hydro-philic regions of a protein have a negative hydropathy index.X The sum of all charged residues where lysine (K.) and arginine (R) residues are positively charged (+1)and glutamic acid (E) and aspartic acid (D) are negatively charged (—1) at neutral pH. Histidine (H) isassumed to be uncharged at neutral pH.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018

156 Investigative Ophthalmology & Visual Science, January 1993, Vol. 34, No. 1
Peptide Coupling to OvalbuminSynthetic peptides were conjugated to ovalbumin(OVA) using EDC (l-ethyl-3-(3-dimethylaminopro-pyl)-carbodiimide hydrochloride; Sigma ChemicalCo., St. Louis, MO) as previously described.26 Briefly,5 mg of each peptide was dissolved in 1 ml of water onice, and 5 mg of OVA was added. Fifty milligrams ofEDC in 150 ix\ of water was added to the peptide-OVAmixture, and the solution was mixed overnight at 4°Con a rotary shaker. The OVA-conjugated peptidesthen were dialyzed extensively against phosphate buff-ered saline (PBS) to remove excess EDC and any un-coupled peptide (Spectrapore 4, 14 kD cutoff; Spec-trum Medical Industries, Los Angeles, CA). The aver-age peptide coupling efficiency was 35% (10-15 molof peptide per mole of OVA) for each of the peptides,as determined by coupling I25I-labeled peptides ofknown specific activity to OVA and measuring the ra-dioactivity present in purified peptide-OVA conju-gates. Aliquots of the peptide-OVA conjugates werestored at -80°C.
Antibody Production, Purification, andCharacterizationPolyclonal antibodies were generated against FN-C/H-V or FN-C/H-II coupled to keyhole limpet hemo-cyanin (KLH; Sigma Chemical Co.) using EDC as previ-ously described.35 New Zealand white rabbits were im-munized on the back by multiple intradermalinjections of approximately 1 mg per rabbit of pep-tide/KLH conjugate in complete Freund's adjuvant.Subsequent biweekly boosts were given intramuscu-larly in incomplete Freund's adjuvant. Sera were col-lected 14 days after the fourth immunization andtested by ELISA for reactivity against uncoupled FN-C/H-V, fibronectin, and other proteins and pep-tides.35 Immunoglobulin G (IgG) was purified frompooled immune sera by precipitation with ammoniumsulfate followed by DEAE anion exchange chromatog-raphy, and the purity of the IgG was verified by SDS-PAGE.
Cell Adhesion AssayPrimary RCE cells were labeled with 35S-methionine(Tran35S-label; ICN Radiochemicals, Irvine CA) for 24hr before their use in adhesion assays. Purified pro-teins or OVA-conjugated peptides were diluted tovarious concentrations in PBS, pH 7.4, and 50 /il ali-quots were added to the wells of a 96-well Immulon Imicrotiter plate (Dynatech Laboratories, Inc., Chan-tilly, VA). Proteins and peptides were allowed to ad-sorb overnight at 37°C; then nonspecific binding siteswere blocked with 2 mg/ml OVA in PBS. Subcon-fluent RCE cells (8-10 days in culture) were harvestedusing trypsin:EDTA and resuspended in DMEM/F-12containing 10% FBS. The cells were centrifuged andresuspended two times in serum-free DMEM/F-12
containing 2 mg/ml OVA, then counted and resus-pended in serum-free DMEM/F-12 containing 2 mg/ml OVA at a final density of 5 X 104 cells/ml. Onehundred microliter aliquots of this cell suspensionwere added to protein/peptide-coated wells and incu-bated for 90 min at 37°C. Nonadherent cells wereremoved by washing three times with PBS containingOVA. Adherent cells were solubilized in 100 /A of 0.5N NaOH/1% sodium dodecyl sulfate, transferred toscintillation vials, and counted in a Beckman (Irvine,CA) 3801 scintillation counter.
To quantitate cell spreading, cells were fixed with3.6% formaldehyde in PBS after a 60 min adhesionassay. Adherent cells were stained for 30 min with 1%crystal violet in 10% ethanol. Plates then were washedtwice with 10% ethanol and air dried. A cell was consid-ered spread if cytoplasmic extensions (ie, pseudopo-dia, lamellipodia) were visible extending beyond thespherical cell profile noted after seeding. The percent-age of spread cells was calculated as the number ofspread cells divided by the number of adherent cellsmultiplied by 100. Results were based on cell counts infour randomly chosen medium power fields (XI00)for triplicate wells for each condition.
Inhibition of RCE Cell Adhesion by SolubleOVA-Coupled FN-C/H-VRCE cells were pre-incubated for 60 min at 37°C in100 Mg/ml of soluble OVA-coupled OVA, OVA-cou-pled FN-C/H-V or OVA-coupled FN-C/H-II. Theythen were added to Immulon I plates coated with 20Mg/ml of the 33/66 kD fragments of fibronectin, 6jug/ml of fibronectin, or 50 fig/ml of OVA-coupledFN-C/H-V. Adhesion assays were terminated after 60min, and cell adhesion was quantitated as describedabove.
Inhibition of RCE Cell Adhesion by PurifiedPolyclonal IgG Against FN-C/H-VImmulon I plates coated with 20 Mg/ml of the 33/66kD fragments of fibronectin, 6 Mg/ml of fibronectin,or 50 Mg/ml of OVA-coupled FN-C/H-V were pre-in-cubated for 30 min with 125 fx§/m\ of purified IgGfrom normal rabbits or rabbits immunized with FN-C/H-V or FN-C/H-II. RCE cells then were added to theplates and cell adhesion was quantitated after 60 min,as described above.
Cell Migration AssaySubconfluent corneal epithelial cells (8-10 days in cul-ture) were harvested using trypsin:EDTA and resus-pended in DMEM/F-12 containing 10% FBS. Thecells were counted and resuspended (two times) inserum-free DMEM/F-12 containing 2 mg/ml OVAand 50 Mg/ml gentamicin at a final density of 1 X 106
cells/ml. Polycarbonate filters (8 nm pore size; Cos-tar/Nucleopore, Cambridge, MA) were coated over-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018
Corneal Epithelial Cell Motility to Fibronectin 157
night (lower surface only) with purified proteins orOVA-conjugated peptides in PBS, pH 7.4. Thesefilters subsequently were rinsed in PBS and air-driedbefore their use. After assembly of the chemotaxischamber (Neuroprobe, Bethesda, MD) with a coatedpolycarbonate filter separating the upper and lowerwells, corneal epithelial cells were added to the upperwells of the chemotaxis chamber (5 X 104 cells/well).The chambers were incubated overnight (24 hr) at37°C in 5% CO2/95% O2; then the filters were re-moved, fixed, and stained. Cells that remained on theupper surface of the filters (ie, those cells that had notmigrated thorough the filter) were removed with acotton swab and the filters were mounted on glassslides for quantitation using a Zeiss (Thornwood, NY)photomicroscope equipped with an ocular grid. Mi-gration was expressed as the number of cells migratingper square millimeter based on cells counted in fourrandomly chosen high-powered fields (X250) for tripli-cate wells for each condition.
To measure the effect of purified polyclonal IgGagainst FN-C/H-V on RCE cell motility, experimentswere conducted as just described in the presence ofpurified IgG (32.5-125 /ig/ml) from normal rabbits orrabbits immunized with FN-C/H-V or FN-C/H-II.The upper and lower wells of the Boyden chambercontained equal concentrations of these antibodies toensure that a gradient of antibodies did not existacross the polycarbonate filter. RCE cell migrationwas quantitated as just described.
RESULTS
RCE Cell Adhesion to Synthetic PeptidesDerived From Fibronectin
We previously showed that the 33/66 kD carboxy-ter-minal heparin-binding/cell adhesion-promoting frag-ments of fibronectin promote the adhesion of RCEcells in a concentration-dependent manner.31 We alsoidentified two peptide sequences derived from the 33/66 kD fragments of fibronectin that promoted RCEcell adhesion and provided evidence that these peptidesequences contributed to the adhesion-promoting ac-tivity of the 33/66 kD fragments.31 However, in com-petition assays, these peptides only partially blockedRCE cell adhesion to the 33/66 kD fragments of fibro-nectin, suggesting that additional sites might contrib-ute to the adhesion-promoting activity of the 33/66kD fragments of fibronectin. To identify these addi-tional sites, we tested the ability of two other peptides(FN-C/H-IV and FN-C/H-V) derived from the pri-mary sequence of the 33/66 kD fragments of fibronec-tin (Fig. 1 and Table 1) to promote RCE cell adhesion.FN-C/H-IV corresponds to residue nos. 1784-1792in type III repeat no. 13 of plasma fibronectin and isfound in a region that recently was shown by Bober-Barkolow and Schwarzbauer35 to be important for the
heparin-binding activity of the 33 kD fragment. FN-C/H-V corresponds to residue nos. 1892-1899 intype III repeat no. 14 of plasma fibronectin. FN-C/H-IV and FN-C/H-V are similar to previously describedpeptides from this region of fibronectin (FN-C/H-I,FN-C/H-II, and FN-C/H-III) in that they are cationic,hydrophilic, and bind [3H]-heparin (data not shown).
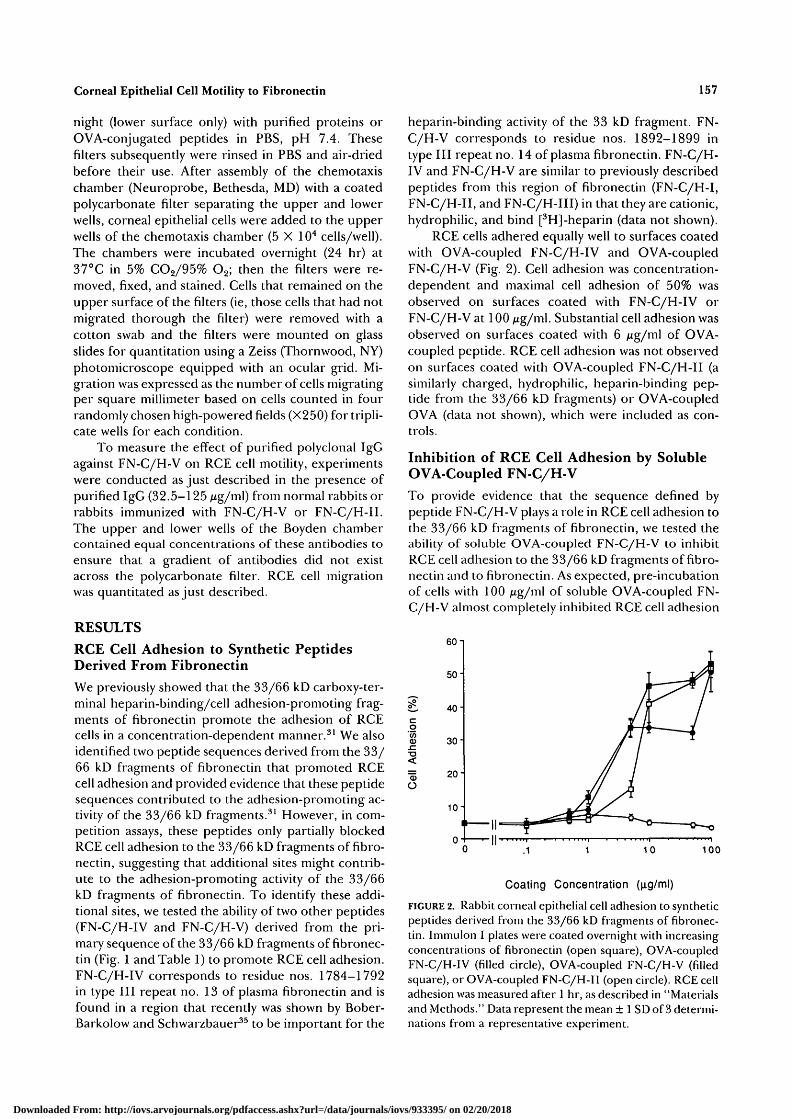
RCE cells adhered equally well to surfaces coatedwith OVA-coupled FN-C/H-IV and OVA-coupledFN-C/H-V (Fig. 2). Cell adhesion was concentration-dependent and maximal cell adhesion of 50% wasobserved on surfaces coated with FN-C/H-IV orFN-C/H-V at 100 /ug/ml. Substantial cell adhesion wasobserved on surfaces coated with 6 Mg/ml of OVA-coupled peptide. RCE cell adhesion was not observedon surfaces coated with OVA-coupled FN-C/H-II (asimilarly charged, hydrophilic, heparin-binding pep-tide from the 33/66 kD fragments) or OVA-coupledOVA (data not shown), which were included as con-trols.
Inhibition of RCE Cell Adhesion by SolubleOVA-Coupled FN-C/H-V
To provide evidence that the sequence defined bypeptide FN-C/H-V plays a role in RCE cell adhesion tothe 33/66 kD fragments of fibronectin, we tested theability of soluble OVA-coupled FN-C/H-V to inhibitRCE cell adhesion to the 33/66 kD fragments of fibro-nectin and to fibronectin. As expected, pre-incubationof cells with 100 ixg/m\ of soluble OVA-coupled FN-C/H-V almost completely inhibited RCE cell adhesion
60 i
50-
O
40-
30"
20"
10"
010 100
Coating Concentration (ng/ml)
FIGURE 2. Rabbit corneal epithelial cell adhesion to syntheticpeptides derived from the 33/66 kD fragments of fibronec-tin. Immulon I plates were coated overnight with increasingconcentrations of fibronectin (open square), OVA-coupledFN-C/H-IV (filled circle), OVA-coupled FN-C/H-V (filledsquare), or OVA-coupled FN-C/H-II (open circle). RCE celladhesion was measured after 1 hr, as described in "Materialsand Methods." Data represent the mean ± 1 SD of 3 determi-nations from a representative experiment.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018
158 Investigative Ophthalmology & Visual Science, January 1993, Vol. 34, No. 1
to surfaces coated with FN-C/H-V (Fig. 3). In addi-tion, soluble OVA-coupled FN-C/H-V also inhibitedRCE cell adhesion to the 33/66 kD fragments and tofibronectin-coated surfaces, by 25% and 20%, respec-tively. This inhibition was specific for peptide FN-C/H-V, because pre-incubation of RCE cells with 100Mg/ml of OVA-coupled FN-C/H-II or OVA-coupledOVA did not inhibit RCE cell adhesion to OVA-cou-pled FN-C/H-V, the 33/66 kD fragments of fibronec-tin, or to fibronectin (Fig. 3).
Inhibition of RCE Cell Adhesion by PolyclonalAntibodies Against FN-C/H-V
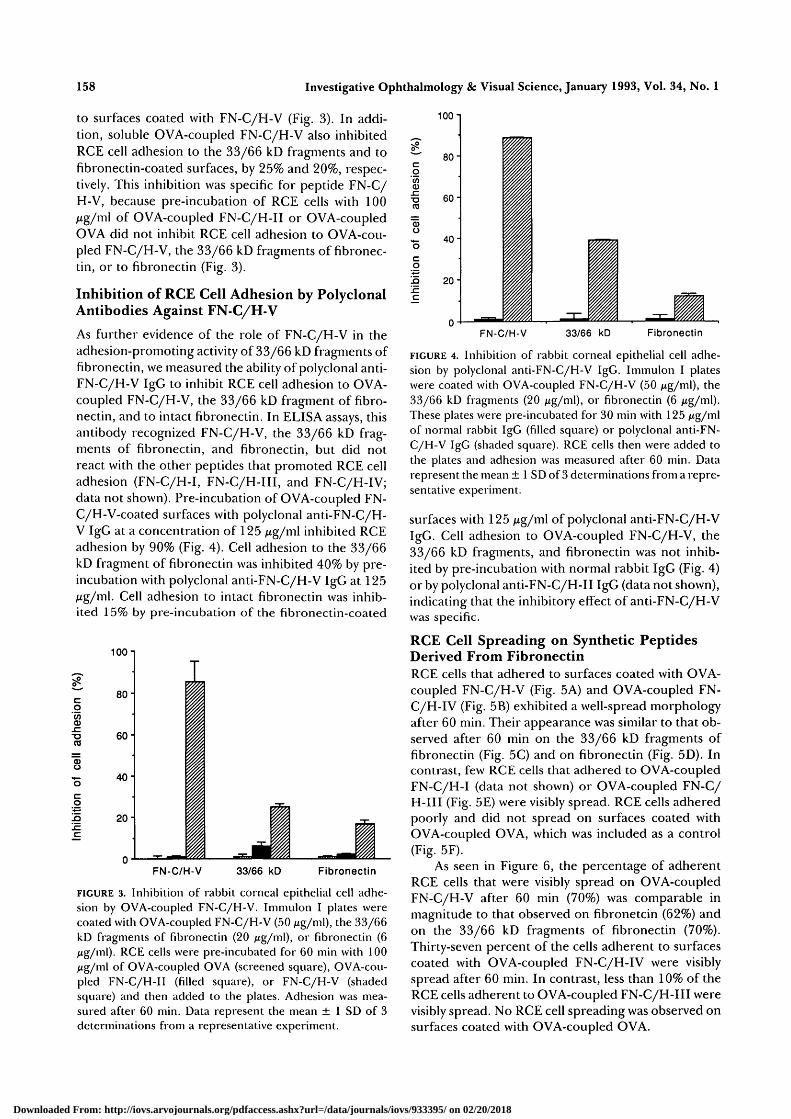
As further evidence of the role of FN-C/H-V in theadhesion-promoting activity of 33/66 kD fragments offibronectin, we measured the ability of polyclonal anti-FN-C/H-V IgG to inhibit RCE cell adhesion to OVA-coupled FN-C/H-V, the 33/66 kD fragment of fibro-nectin, and to intact fibronectin. In ELISA assays, thisantibody recognized FN-C/H-V, the 33/66 kD frag-ments of fibronectin, and fibronectin, but did notreact with the other peptides that promoted RCE celladhesion (FN-C/H-I, FN-C/H-III, and FN-C/H-IV;data not shown). Pre-incubation of OVA-coupled FN-C/H-V-coated surfaces with polyclonal anti-FN-C/H-V IgG at a concentration of 125 /ig/m\ inhibited RCEadhesion by 90% (Fig. 4). Cell adhesion to the 33/66kD fragment of fibronectin was inhibited 40% by pre-incubation with polyclonal anti-FN-C/H-V IgG at 125jug/ml. Cell adhesion to intact fibronectin was inhib-ited 15% by pre-incubation of the fibronectin-coated
100-1
co'55CD.c"O
O
J5
8 0 -
60"
40-
20-
o-1-FN-C/H-V 33/66 kD Fibronectin
FIGURE 3. Inhibition of rabbit corneal epithelial cell adhe-sion by OVA-coupled FN-C/H-V. Immulon I plates werecoated with OVA-coupled FN-C/H-V (50 Mg/ml), the 33/66kD fragments of fibronectin (20 ng/m\), or fibronectin (6/ig/ml). RCE cells were pre-incubated for 60 min with 100jig/ml of OVA-coupled OVA (screened square), OVA-cou-pled FN-C/H-II (filled square), or FN-C/H-V (shadedsquare) and then added to the plates. Adhesion was mea-sured after 60 min. Data represent the mean ± 1 SD of 3determinations from a representative experiment.
T3CO
JO
c
100 i
80"
60-
40"
20
FN-C/H-V 33/66 kD Fibronectin
FIGURE 4. Inhibition of rabbit corneal epithelial cell adhe-sion by polyclonal anti-FN-C/H-V IgG. Immulon I plateswere coated with OVA-coupled FN-C/H-V (50 iig/m\), the33/66 kD fragments (20 jig/ml), or fibronectin (6 /tig/ml).These plates were pre-incubated for 30 min with 125 Mg/mlof normal rabbit IgG (filled square) or polyclonal anti-FN-C/H-V IgG (shaded square). RCE cells then were added tothe plates and adhesion was measured after 60 min. Datarepresent the mean ± 1 SD of 3 determinations from a repre-sentative experiment.
surfaces with 125 Mg/ml of polyclonal anti-FN-C/H-VIgG. Cell adhesion to OVA-coupled FN-C/H-V, the33/66 kD fragments, and fibronectin was not inhib-ited by pre-incubation with normal rabbit IgG (Fig. 4)or by polyclonal anti-FN-C/H-II IgG (data not shown),indicating that the inhibitory effect of anti-FN-C/H-Vwas specific.
RCE Cell Spreading on Synthetic PeptidesDerived From FibronectinRCE cells that adhered to surfaces coated with OVA-coupled FN-C/H-V (Fig. 5A) and OVA-coupled FN-C/H-IV (Fig. 5B) exhibited a well-spread morphologyafter 60 min. Their appearance was similar to that ob-served after 60 min on the 33/66 kD fragments offibronectin (Fig. 5C) and on fibronectin (Fig. 5D). Incontrast, few RCE cells that adhered to OVA-coupledFN-C/H-I (data not shown) or OVA-coupled FN-C/H-III (Fig. 5E) were visibly spread. RCE cells adheredpoorly and did not spread on surfaces coated withOVA-coupled OVA, which was included as a control(Fig. 5F).
As seen in Figure 6, the percentage of adherentRCE cells that were visibly spread on OVA-coupledFN-C/H-V after 60 min (70%) was comparable inmagnitude to that observed on fibronetcin (62%) andon the 33/66 kD fragments of fibronectin (70%).Thirty-seven percent of the cells adherent to surfacescoated with OVA-coupled FN-C/H-IV were visiblyspread after 60 min. In contrast, less than 10% of theRCE cells adherent to OVA-coupled FN-C/H-III werevisibly spread. No RCE cell spreading was observed onsurfaces coated with OVA-coupled OVA.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018

Corneal Epithelial Cell Motility to Fibronectin
F >
FIGURE 5. Rabbit corneal epithelial cell spreading on fibronectin, the 33/66 kD fragments offibronectin, and synthetic peptides. lmmulon I plates were coated overnight with 6 Mg/m' offibronectin, 20 Mg/m' of the 33/66 kD fragments of fibronectin, or 50 ^g/ml of OVA-cou-pled FN-C/H-III, OVA-coupled FN-C/H-IV, OVA-coupled FN-C/H-V, or OVA-coupledOVA. The plates then were blocked with OVA, and RCE cells were added. Adherent cellswere fixed and stained after 1 hr, as described in the "Materials and Methods." RCE cellspreading on OVA-coupled FN-C/H-V (A), OVA-coupled FN-C/H-IV (B), fibronectin (C),the 33/66 kD fragments of fibronectin (D), OVA-coupled FN-C/H-III (E), and OVA-cou-pled OVA (F). (Bar = 30 mm.)
RCE Cell Haptotactic Migration onFibronectin and Fibronectin Fragments
The 33/66 kD fragments promoted the haptotacticmigration of RCE cells in a concentration-dependent
manner (Fig. 7). Significant haptotactic migration wasobserved on polycarbonate filters coated with 1 pM ofthe 33/66 kD fragments of fibronectin; at a concen-tration of 500 pM, the 33/66 kD fragments promotedRCE cell migration nearly as effectively as did fibronec-tin. The 27 kD fragment of fibronectin, which was in-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018

160 Investigative Ophthalmology & Visual Science, January 1993, Vol. 34, No. 1
OVALBUMIN
FN-C/H-V
FN-C/H-IV
FN-C/H-III
33/66 kD
FIBRONECTIN
1000"!
Cell Spreading (%)
FIGURE 6. Quantitation of RCE cell spreading on fibronec-tin, the 33/66 kD fragments of fibronectin, and syntheticpeptides. Immulon I plates were coated overnight with 10ixg/m\ of fibronectin, the 33/66 kD fragments of fibronec-tin, OVA-coupled FN-C/H-III, OVA-coupled FN-C/H-IV,OVA-coupled FN-C/H-V, or OVA-coupled OVA. Theplates then were blocked with OVA, and RCE cells wereadded. RCE cell spreading was measured after 1 hr, as de-scribed in "Materials and Methods." Data represent themean ± 1 SD of 3 determinations from a representative ex-periment.
eluded as a control, did not promote the haptotacticmigration of RCE cells at any concentration tested.
RCE Cell Haptotactic Migration on SyntheticPeptides Derived From Fibronectin
OVA-coupled FN-C/H-V promoted the haptotacticmigration of RCE cells in a concentration-dependentmanner (Fig. 8). RCE cell migration was observed onfilters coated with as little as 5 jitM of OVA-coupledFN-C/H-V, whereas maximal cell migration was seenon filters coated with 100 /xM of OVA-coupled FN-C/H-V. A slight haptotactic response was observed athigh coating concentrations of FN-C/H-IV (50 pM),but this response was not clearly dose-dependent.RCE cells did not migrate on filters coated with FN-C/H-I, FN-C/H-III (data not shown), or OVA-coupledOVA (Fig. 7). These findings emphasize that cell adhe-sion is required but not sufficient for cell motility, be-cause at least three peptides from the 33/66 kD frag-ments that promote RCE cell adhesion (FN-C/H-I,FN-C/H-III, and FN-C/H-IV) do not promote RCEcell motility.
Inhibition of RCE Cell Haptotactic Migrationby Polyclonal Antibodies Against FN-C/H-V
To provide evidence for a contribution by FN-C/H-Vto the motility-promoting activity of the 33/66 kDfragments of fibronectin, the ability of polyclonal anti-FN-C/H-V IgG to inhibit RCE haptotactic migrationon these fragments was studied. RCE cell haptotacticmigration on the 33/66 kD fragment of fibronectin
o
100
Coating concentration (M x 10 )
FIGURE 7. RCE cell haptotactic migration on fibronectin andfragments of fibronectin. Polycarbonate filters were coatedovernight with intact fibronectin (open circle), the 33/66 kDfragments (filled circle), or the 27 kD fragment of fibronec-tin (filled square). Haptotactic migration was measured after24 hr, as described in "Materials and Methods." Data repre-sent the mean ± 1 SD of 12 determinations from a represen-tative experiment.
was inhibited in a concentration-dependent manner inthe presence of anti-FN-C/H-V IgG (Fig. 9). Maximalinhibition of 80% was observed at an IgG concentra-tion of 125 ng/m\. In contrast, RCE cell haptotacticmigration was not inhibited in the presence of normalrabbit IgG at concentrations up to 125 fig/ml, con-firming the specificity of the anti-FN-C/H-V inhibi-tory effect (Fig. 9). Likewise, polyclonal anti-FN-C/H-II did not inhibit RCE cell haptotactic migration
250 n
200
150
100
50-
100
Coating Concentration ( M x 1 0 " 6 )
FIGURE 8. RCE cell haptotactic migration on OVA-coupledFN-C/H-IV and FN-C/H-V. Polycarbonate filters werecoated overnight with OVA-coupled FN-C/H-IV (filled cir-cle), OVA-coupled FN-C/H-V (open circle), or OVA-cou-pled OVA (filled square). Haptotactic migration was mea-sured after 24 hr, as described in "Materials and Methods."Data represent the mean ± 1 SD of 12 determinations froma representative experiment.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018

Corneal Epithelial Cell Motility to Fibronectin 161
100
a.I
60 80 100 120 140
IgG (jig/ml)
FIGURE 9. Inhibition of RCE cell haptotactic migration tothe 33/66 kD fragments by polyclonal anti-FN-C/H-V IgG.Polycarbonate filters were coated overnight with the 33/66kD fragments (20 /xg/ml) and placed in modified Boydenchambers. RCE cells were harvested and added to thesechambers with increasing concentrations of polyclonal anti-FN-C/H-V IgG (filled circle) or normal rabbit IgG (opencircle). The antibodies were present in the upper and lowerwells of the Boyden chamber. Haptotactic migration wasmeasured after 24 hr, as described in "Materials and Meth-ods." Data represent the mean ± 1 SD of 12 determinationsfrom a representative experiment.
(data not shown). The dramatic inhibition of RCE cellmotility (80%) by anti-FN-C/H-V IgG at a concentra-tion (125 /ug/mO that produced a much smaller de-crease in cell adhesion (40%) provides evidence thatFN-C/H-V contributes to the motility-promoting activ-ity of the 33/66 kD fragments of fibronectin.
DISCUSSION
Fibronectin plays an important role in corneal epithe-lial cell adhesion and motility during corneal woundhealing. Multiple domains contribute to the cell adhe-sion-promoting activity of fibronectin.1011 The RGDStetrapeptide sequence perhaps is the most wellknown.12 RCE cells adhere to a fragment of fibronec-tin that contains this sequence,4 and Nishida et al13
showed that an RGDS-containing synthetic peptidecan inhibit RCE cell adhesion to intact fibronectin.Previous studies have shown that the 33/66 kD hepa-rin-binding fragments of fibronectin also promote theadhesion of RCE cells; these studies have identifiedtwo synthetic peptides that contribute to the adhesion-promoting activity of these fragments.31 In this report,we demonstrated that the 33/66 kD fragments of fi-bronectin also promote RCE cell haptotactic migra-tion, and we identified two peptides derived from the33/66 kD fragments of fibronectin (FN-C/H-IV and
FN-C/H-V) that promote RCE cell adhesion andspreading, one of which (FN-C/H-V) also promotesRCE cell migration.
FN-C/H-IV and FN-C/H-V are chemically synthe-sized peptides from the 33/66 kD heparin-binding,cell adhesion-promoting fragments of fibronectin.These peptides promoted the adhesion of RCE cells ina concentration-dependent manner. RCE cell adhe-sion to FN-C/H-IV and FN-C/H-V was peptide-spe-cific because FN-C/H-II, also a hydrophilic and cat-ionic heparin-binding peptide derived from the 33/66kD fragments of fibronectin, did not promote RCEcell adhesion.31 Importantly, FN-C/H-II does pro-mote the adhesion of a variety of other cell types,2627
supporting the argument that cell adhesion to pep-tides from the 33/66 kD fragments of fibronectin alsois cell-type specific and that the activity of these pep-tides is not simply the result of net charge or net hy-dropathy indices.
Bober-Barkalow and Schwarzbauer36 showed thatmanipulation of the amino-terminal portion of typeIII repeat no. 13 in the 33 kD fragment that containsresidues within the FN-C/H-IV sequence results in adramatic loss of the heparin-binding activity of thisfragment. Our results suggest this sequence (FN-C/H-IV) may contribute to RCE cell adhesion and spread-ing on the 33 kD fragment as well. Our results alsosuggest that FN-C/H-V, a sequence found in type IIIrepeat no. 14, is an important RCE cell adhesion-,spreading-, and motility-promoting sequence in the33/66 kD fragments of fibronectin.
This conclusion is based on several lines of evi-dence. First, we have demonstrated that FN-C/H-Vdirectly promotes the adhesion, spreading, and hapto-tactic migration of RCE cells and therefore mimics the33/66 kD fragments of fibronectin in this regard. Sec-ond, soluble FN-C/H-V partially inhibited the adhe-sion of RCE cells to the 33/66 kD fragments of fibro-nectin and to fibronectin, as would be expected if FN-C/H-V and the 33/66 kD fragments are ligandscompeting for a common RCE cell receptor. Third,polyclonal anti-FN-C/H-V IgG partially inhibitedRCE cell adhesion to the 33/66 kD fragments of fibro-nectin and to fibronectin. This is consistent with amodel in which the sequence defined by peptide FN-C/H-V is an important cell adhesion-promoting do-main within the 33/66 kD fragments of fibronectin.
The failure of soluble peptide FN-C/H-V and anti-bodies against FN-C/H-V to completely block RCEcell adhesion to the 33/66 kD fragments of fibronec-tin is consistent with previous reports that describemultiple active cell adhesion-promoting domainswithin the 33/66 kD fragments.25~28>30>31 Furthermore,the finding that soluble FN-C/H-V and anti-FN-C/H-V IgG were less effective as inhibitors of RCE adhesionto fibronectin is not surprising, because not only areadditional cell adhesion-promoting sites available
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018

162 Investigative Ophthalmology & Visual Science, January 1993, Vol. 34, No. 1
within the 33/66 kD fragments, but the RGDS adhe-sion-promoting sequence in fibronectin is also pres-ent12 and has been shown to promote RCE cell adhe-
4,13sion.
Finally, polyclonal anti-FN-C/H-V IgG almostcompletely inhibited RCE cell motility on the 33/66kD fragments of fibronectin. These findings are con-sistent with experimental results that demonstrate FN-C/H-V is one of at least four peptide sequences withinthe 33/66 kD fragments of fibronectin that contributeto RCE cell adhesion (FN-C/H-I, FN-C/H-III, FN-C/H-IV, and FN-C/H-V), but is the only sequence fromthe 33/66 kD fragments of fibronectin that has beenshown to promote RCE cell motility. Although we can-not rule out contributions by as yet unidentified se-quences within the 33/66 kD fragments, the magni-tude of the inhibition observed in the presence of anti-FN-C/H-V IgG suggests a particularly important rolefor the sequence defined by FN-C/H-V in the motility-promoting activity of the 33/66 kD fragments.
An unresolved aspect of these studies is the rela-tively low potency of peptide FN-C/H-V when com-pared to intact fibronectin or to the 33/66 kD frag-ments of fibronectin. It has been shown by others thatcell adhesion-promoting synthetic peptides frequentlyare less active than the intact proteins from which theyare derived.37 A synthetic peptide, for example, maylack important secondary structural features that con-tribute to receptor recognition37 and therefore mayexhibit reduced activity when compared to its parentmolecule. In addition, a synthetic peptide may repre-sent a small part of a larger contiguous or noncon-tiguous binding site and therefore may only partiallyreproduce the activity of the parent molecule.15 A use-ful approach in this regard has been to develop mu-tated versions of larger functional domains using site-directed mutagenesis. Obara et al15 and others1617
have used this approach to identify "synergy" sitesthat may contribute to the adhesion-promoting activ-ity of the RGDS-containing central cell binding do-main. Similar studies may be needed to address theissue of FN-C/H-V and its activity relative to fibro-nectin.
The cellular receptor or receptors for FN-C/H-Vhas not been identified. Although members of the in-tegrin family of cell adhesion receptors may be in-volved,37-38 as has been suggested for FN-C/H-II,29 theheparin-binding properties of the 33/66 kD frag-ments of fibronectin and of FN-C/H-V itself stronglyfavor a role for cell surface proteoglycans in RCE celladhesion to peptide FN-C/H-V. A number of reportshave shown that cell surface proteoglycans can play arole in cell adhesion to extracellular matrix glycopro-teins.39"43 Cell surface proteoglycans have been impli-cated in cell adhesion to several peptides from the 33/66 kD fragments of fibronectin.24~26-2930 Identifyingthe cell surface receptor or receptors that mediate ad-
hesion to FN-C/H-V, as well as to FN-C/H-I, FN-C/H-III, and FN-C/H-IV, may provide an explanationfor the distinct RCE cell responses to these peptides.
The mechanisms that underlie the motility-pro-moting effects of FN-C/H-V likewise are unknown,although it seems likely that some form of transmem-brane signaling occurs during this process, becausecytoskeletal reorganization, cell spreading, and cellmotility are active processes and occur in this systemonly in response to specific peptides. Studies are nowunderway to identify the signal transduction pathwaysinvolved in FN-C/H-V-mediated RCE cell adhesion,spreading, and motility.
Given the important role of fibronectin in cornealwound healing in vivo, these findings provide addi-tional insight into the complex molecular basis of cor-neal epithelial cell interactions with fibronectin andmay be important in the context of corneal woundhealing. For example, peptide FN-C/H-V may be auseful alternative to fibronectin as a topical stimulantof corneal re-epithelialization and wound healing invivo. Alternatively, FN-C/H-V (or other FN-C/H pep-tides) may enhance re-epithelialization of intracornealimplants when covalently coupled to implant biomate-rial surfaces.
Key Wordsadhesion, cornea, epithelium, fibronectin, motility.
Acknowledgments
The authors thank Eric Kjellesvig, Xiaoling Wang, JudyKahm, Dan Mickelson, and Bina Vachhani for their excel-lent technical assistance during these studies.
References
1. Berman M, Manseau E, Law M. Ulceration is corre-lated with degradation of fibrin and fibronectin at thecorneal surface. Invest Ophthalmol Vis Sci. 1983;24:1358-1366.
2. Yamada KM, Olden K. Fibronectin: Adhesive glyco-proteins of cell surface and blood. Nature. 1978;175:179-184.
3. Suda T, Nishida T, Ohashi T, Nakagawa S, Manabe R.Fibronectin appears at the site of corneal stromalwound in rabbits. Curr Eye Res. 1981; 1:553-556.
4. Cameron JD, Hagen ST, Waterneld RR, Furcht LT.Effects of matrix proteins on rabbit corneal epithelialcell adhesion and migration. Curr Eye Res. 1988;7:293-301.
5. Nishida T, Nakagawa S, Awata T, Ohashi Y, WatanabeK, Manabe R. Fibronectin promotes epithelial migra-tion of cultured rabbit cornea in situ. J Cell Biol.1983;97:1653-1657.
6. Nishida T, Makagawa S, Nishibayashi C, Tanaka H,Manabe R. Fibronectin enhancement of corneal epi-thelial wound healing of rabbits in vivo. Arch Ophthal-mol. 1984; 102:455-456.
7. Nishida T, Ohashi Y, Awata T, Manabe R. Fibronec-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018

Corneal Epithelial Cell Motility to Fibronectin 163
tin: A new therapy for corneal trophic ulcer. Arch Oph-Lhalmol. 1983; 101:1046-1048.
8. Phan T-M, Foster CS, Boruchoff SA, Zagachin LM,Colvin RB. Topical fibronectin in the treatment ofpersistent corneal epithelial defects and trophiculcers. Am] Ophthalmol. 1987; 104:494-501.
9. Sekiguchi K, Fukuda M, Hakomori S-J. Domain struc-ture of hamster plasma fibronectin. J Biol Chem.1981;256:6452-6462.
10. Hynes RO. Fibronectins. New York: Springer-Verlag;1990.
11. Mosher DF. Fibronectin. New York: Academic Press;1989.
12. Pierschbacher MD, Ruoslahti E. The cell attachmentactivity of fibronectin can be duplicated by small syn-thetic fragments of the molecule. Nature. 1984;309:30-33.
13. Nishida T, Nakagawa S, Watanabe K, Yamada KM,Otari T, Berman MB. A peptide from fibronectin cell-binding domain inhibits attachment of epithelial cells.Invest Ophthalmol Vis Sci. 1988; 29:1820-1825.
14. Pytela R, Pierschbacher MD, Ruoslahti E. Identifica-tion and isolation of a 140kd cell surface glycoproteinwith properties expected of a fibronectin receptor.Cell. 1985;40:191-198.
15. Obara M, Kang MS, Yamada KM. Site-directed muta-genesis of the cell-binding domain of human fibronec-tin. Separable, synergistic sites mediated adhesivefunction. Cell. 1988;53:649-657.
16. Aota S, Nagai T, Yamada KM. Characterization of re-gions of fibronectin besides the arginine-glycine-aspartic acid sequence required for adhesive functionof the cell binding domain using site directed muta-genesis./flio/ Chem. 1991; 266:15938-15943.
17. Nagai T, Yamakawa N, Aota S, Yamada SS, Olden K,Yamada KM. Monoclonal antibody characterizationof two distinct sites required for function of the cen-tral cell-binding domain of fibronectin in cell adhe-sion, cell migration and matrix assembly. J Cell Biol.1991;114:1295-1305.
18. Kornblihtt AR, Umezawa K, Vibe-Pedersen K, BaralleFE. Primary structure of human fibronectin: Differ-ential splicing may generate at least 10 polypeptidesfrom a single gene. EMBOJ. 1985;4:1755-1759.
19. Humphries MJ, Akiyama SK, Komoriya A, Olden K,Yamada KM. Identification of an alternatively splicedsite in human plasma fibronectin that mediates celltype-specific adhesion./ Cell Biol. 1986; 103:2637-2647.
20. Humphries MJ, Akiyama SK, Komoriya A, Olden K,Yamada KM. Identification of two distinct regions ofthe type III connecting segment of human plasma fi-bronectin that promote cell type-specific adhesion. /Biol Chem. 1987;262:6886-6892.
21. Komoriya A, Green LJ, Mervic M, Yamada SS, Ya-mada KM, Humphries MJ. The minimal essential se-quence for a major cell type-specific adhesion site(CS1) within the alternatively spliced type III connect-ing segment domain of fibronectin is leucine-asparticacid-valine./Zfco/ Chem. 1991;266:15075-15079.
22. Wayner EA, Garcia-Pardo A, Humphries MJ, McDon-ald JA, Carter WG. Identification and characteriza-
tion of the T lymphocyte adhesion receptor for analternative cell attachment domain (CS-1) in plasmafibronectin. J Cell Biol. 1989; 109:1321-1330.
23. Mould AP, Komoriya A, Wayner EA, Yamada KM,Humphries MJ. The CS5 peptide is a second site inIIICS region of fibronectin recognized by the integrina4j81. Inhibition of a4/31 function by RGD peptidehomologues./ZfcoZ Chem. 1991; 266:3579-3585.
24. McCarthyJB, Hagen ST, Furcht LT. Human fibronec-tin contains distinct adhesion- and motility-promotingdomains for metastatic melanoma cells. J Cell Biol.1986; 102:179-188.
25. McCarthyJB, Skubitz APN, Zhao Q, Yi X-Y, Mickel-son DJ, Klein DJ, Furcht LT. RGD-independent celladhesion to the carboxy-terminal heparin-bindingfragment of fibronectin involves heparin-dependentand -independent activities. / Cell Biol. 1990;110:777-787.
26. Haugen PK, McCarthyJB, Skubitz APN, Furcht LT,Letourneau PC. Recognition of the A chain carboxy-terminal heparin binding region of fibronectin in-volves multiple sites: Two contiguous sequences actindependently to promote neural cell adhesion./ CellBiol. 1990; 111:2733-2745.
27. Wilke MS, Skubitz APN, Furcht LT, McCarthy JB.Human keratinocytes adhere to two distinct heparin-binding synthetic peptides derived from fibronectin.Invest Dermatol. 1991; 97:573-579.
28. McCarthy JB, Chelberg MK, Mickelson DJ, FurchtLT. Localization and chemical synthesis of fibronectinpeptides with melanoma adhesion and heparin bind-ing activities. Biochemistry. 1988; 27:1380-1388.
29. Drake SL, Klein DJ, Mickelson DJ, Oegema TR,Furcht LT, McCarthyJB. Cell surface phosphatidylin-ositol-anchored heparan sulfate proteoglycan initiatesmouse melanoma cell adhesion to a fibronectin-de-rived, heparin-binding synthetic peptide. / Cell Biol.1992;117:1331-1341.
30. Iida J, Skubitz APN, Furcht LT, Wayner EA,McCarthy JB. Coordinate role for cell surface chon-droitin sulfate proteoglycan and a4/31 integrin in me-diating melanoma cell adhesion to fibronectin./ CellBiol. 1992; 118:431-444.
31. Mooradian DL, McCarthyJB, Cameron JD, SkubitzAPN, Furcht LT. Rabbit corneal epithelial cells ad-here to two distinct heparin-binding synthetic pep-tides derived from fibronectin. Invest Ophthalmol VisSci. 1992;33:3034-3040.
32. Mooradian DL, Weatherbee J, Lucas R, Furcht LT.Transforming growth factor-/3l binds to immobilizedfibronectin./ Cell Biochem. 1990;41:189-200.
33. Koliakos GG, Kouzi-Koliakos K, Furcht LT, RegerLA, Tsilibary EC. The binding of heparin to type IVcollagen: Domain specificity with identification ofpeptide sequences from the al(IV) and «2(IV) chainwhich preferentially bind heparin. / Biol Chem.1989;264:2313-2323.
34. Kyte J, Doolittle RF. A simple method for displayingthe hydropathic character of a protein. / Mol Biol.1982;157:105-132.
35. Skubitz APN, Charonis AS, Tsilibary EC, Furcht LT.Localization of a tumor cell adhesion domain of lam-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018

164 Investigative Ophthalmology & Visual Science, January 1993, Vol. 34, No. 1
inin by a monoclonal antibody. Exp Cell Res.1987; 173:349-360.
36. Bober-Barkalow FJ, Schwarzbauer JE. Localization ofthe major heparin-binding site in fibronectin. J BiolChem. 1991; 266:7812-7818.
37. Ruoslahti E, Pierschbacher MD. New perspectives incell adhesion: RGD and integrins. Science. 1987; 233:491-497.
38. Hynes RO. Integrins, a family of cell surface recep-tors. Cell. 1987; 48:549-555.
39. Rapraeger AC, Bernfield M. Heparan sulfate proteo-glycans from mouse mammary epithelial cells: A puta-tive membrane proteoglycan associates quantitativelywith lipid vesicles. J Biol Chem. 1983; 258:3632-3636.
40. Woods A, Couchman JR, Hook M. Heparan sulfateproteoglycans of rat embryo fibroblasts. J Biol Chem.1985;260:10872-10879.
41. Mugnai G, Lewandowska K, Choi HU, Rosenberg LC,Culp LA. Ganglioside-dependent adhesion events ofhuman neuroblastoma cells regulated by the RGDS-dependent fibronectin receptor and proteoglycans.Exp Cell Res. 1988; 175:229-247.
42. Culp LA, Mugnai G, Lewandoska K, Vallen EA, KosirMA, Houmiel KL. Heparan sulfate proteoglycans ofras-transformed 3T3 or neuroblastoma cells. Ann NYAcadSci. 1989;556:194-216.
43. Hook M, Kjellen L, Johansson S, Robinson J. Cell-sur-face glycosaminoglycans. Annu Rev Biochem. 1984;53:847-869.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933395/ on 02/20/2018