characterization of a bacillus gene cluster required for ...20lundstr%f6m.pdf · lbpg luria broth...
TRANSCRIPT

F A C U L T Y O F S C I E N C E U N I V E R S I T Y O F C O P E N H A G E N D e p a r t m e n t o f B i o l o g y P h D S c h o o l o f S c i e n c e P r o j e c t f u n d e d b y E u r o p e a n C o m m i s s i o n u n d e r t h e 7 t h F r a m e w o r k P r o g r a m m e ( F P 7 ) , M a r i e - C u r i e I T N P r o j e c t 2 1 5 5 2 4 a n d N o v o z y m e s A / S
PhD thesis Sara Lundström
Characterization of a Bacillus licheniformis gene cluster required for functional expression of a bacteriocin
Academic advisor: Michael Askvad Sørensen
Submitted: 31/05/12

II
PREFACE Presented in this PhD thesis is the result of work carried out by me at the Department of Bacterial Gene Technology, Novozymes A/S, DK-2880 Bagsværd, Denmark. During this project I was enrolled as a PhD student at the University of Copenhagen, Faculty of Science. The PhD project was financed to equal parts by European Commission under the 7th Framework Programme (FP7), Marie-Curie ITN Project 215524 and Novozymes A/S. Academic advisor of this project was Professor Michael Askvad Sørensen at the Department of Biology, Biomolecular Sciences. This project was supervised by Jens Tønne Andersen with assistance by Michael Dolberg Rasmussen and Poul Erik Pedersen from Novozymes A/S, Department of Bacterial Gene Technology. The work carried out during my PhD is presented in five chapters (Chapters 4-8). These chapters are written in article format without an abstract, since Chapter 3 serves this purpose. Sara Lundström May 2012

III
ACKNOWLEDGEMENTS I thankfully acknowledge the award of a PhD fellowship by Marie Curie Initial Training Network, FP7 (Project 215524) and Novozymes A/S. I wish to express my deepest gratitude to my supervisors Jens Tønne Andersen, Michael Dolberg Rasmussen, and Poul Erik Pedersen at Novozymes A/S and my principal supervisor Michael Askvad Sørensen at the University of Copenhagen. Their guidance, knowledge, criticism, and encouragements have been a huge support during this research. I would also like to express gratitude to the TranSys Consortium for making this PhD fellowship such an inspiring and positive learning experience. The wonderful people involved have made these three years an amazing journey. Among the people involved in this study I would like to acknowledge the departments within Novozymes A/S whose work made this study possible. The department of Protein Technology at Novozymes A/S is recognized for their help with the N-terminal protein sequencing of formosin with a special thanks to Christian Isak Jørgensen and Clive Phipps Walter who carried out this work. Through their contribution the N-terminal of secreted formosin could be determined, subsequently leading to the determination of the N-terminal signal peptide cleavage site. The department of Protein Biochemistry is recognized for the purification of formosin and the Ci2a protein, with a special thanks to Peter Rahbek Østergaard for producing the purified samples of formosin. With this contribution the antibacterial effects of formosin could be evaluated and antibodies could be produced against both formosin and the Ci2a protein. For the production of antibodies against formosin and Ci2a the Toxicology department is recognized. Through their contribution western blotting analysis could be performed towards these two proteins. In addition to these departments at Novozymes A/S a special thanks is given to ImaGene-iT at Lunds University for their help in the localization study by providing guidance and expertise for the microscope imaging of the GFP fusions proteins. I also wish to thank my colleagues at the department of Bacterial Gene Technology for providing an inspiring scientific environment with amazing support and positive atmosphere. I am especially grateful for the wonderful support and time that Pia Andersen, Pernille Hvid Christensen, and Birthe Kate Lassen have given me during my time in this department. Furthermore, I am grateful for the advice that Anne Breüner and Brian Købmann at Novozymes A/S and Lisa Theorin at the University of Copenhagen gave me, when they took the time to proof-read parts of this PhD thesis. Finally, I want to thank my family and friends for their continuous encouragements and love which has given me the confidence to pursue my dreams. Dad and Marie, thank you for always being there for me. Fredrik, thank you for your love and for being my biggest (but perhaps skinniest) fan. Niclas, thank you for your big heart and for always having my back. Jimmy, thank you for always making me smile and being so kindhearted. Daniel, thank you for being my younger but “older” and wiser brother. Ola, thank you for your endless love and support.

IV
ABSTRACT The aim of this PhD project was to characterize the gene cluster required for functional expression of a
bacteriocin (UniProt ordered locus name BL00275) from Bacillus licheniformis ATCC 14580. This bacteriocin,
with the proposed name formosin (ForD) are encoded in a gene located in the chromosome of B. licheniformis
ATCC 14580 with three adjacent genes (UniProt ordered locus names BL00274-BL00272), proposed to be
named forE, forF, and forG, respectively. Genetic in silico analysis showed that these four genes are arranged in
an operon situated in a genomic island with host defensive properties. Structural and functional studies
demonstrated that ForE and ForG constitute an ABC transporter required in both secretion of and immunity to
formosin. ForF is an accessory protein to the ForEG ABC transporter containing an N-terminal transmembrane
domain. No function could be linked to ForF. Secretion analysis revealed formosin to have two secretion
signals; one N-terminal sec-dependent signal peptide and one C-terminal ABC transporter signal. However, only
when secreted through the ForEG ABC transporter could formosin be detected in the medium.
Characterization of formosin showed that it is a 9.6kDa heat-labile bacteriocin belonging to the lactococcin 972
family with an observed bacteriolytic effect on Bacillus subtilis. ForG is a structural homolog of a previously
described “immunity” protein associated with this protein family. Investigations of lactococcin 972-like protein
and adjacent genes, in addition to the results obtained within this project, concluded that the members of the
lactococcin 972 family are associated with ABC transporters and not transmembrane immunity proteins as
previously predicted.

V
ABBREVIATIONS 3Pcry Promoter system with three promoters and an mRNA stabilizing segment aa Amino acids ABC transporter ATP binding cassette transporter AmyQ Alpha-amylase from Bacillus amyloliquefaciens ATP Adenosine-5'-triphosphate ATCC American Type Culture Collection CFU Colony-forming unit Cm Chloramphenicol Ci2a Subtilisin-chymotrypsin inhibitor-2A (protease inhibitor) C-terminal Carboxyl-terminus of protein C-domain Carboxyl-terminus of signal peptide DNA Deoxyribonucleic acid D-loop Conserved motif in ATP binding domain of ABC transporters ENI motif Conserved motif in ATP binding domain of ABC exporters GC content Guanine-cytosine content GI Genomic Island GFP Green fluorescent protein HlyA Haemolysin A from Escherichia coli H-domain Hydrophobic domain of signal peptide H-loop Conserved motif in ATP binding domain of ABC transporters ICL Intracellular loops of membrane proteins kDa Molecular mass unit, kilo Dalton Km Kanamycin LBPG Luria broth with added glucose and potassium phosphate Man-PTS Mannose permease phosphotransferase system MBC Minimal bactericidal concentration MFP Membrane fusion protein MHB Müeller-Hinton II Broth MIC Minimal inhibition concentration mRNA Messenger ribonucleic acid NBD Nucleotide-binding domain N-domain Amine-terminus of signal peptide

VI
N-terminal Amine-terminus of protein OD600 Optical density measured at a wavelength of 600nm ORF Open reading frame PBS Phosphate buffered saline PCR Polymerase chain reaction PG Phosphatidylglycerol pH Potential Hydrogen pI Isoelectric point poly(U) Stretch of uracil residues RFP Red fluorescent protein rRNA Ribosomal ribonucleic acid SD Standard deviations Sec-pathway General secretory pathway SOE PCR Splicing by overlap extension polymerase chain reaction TA Teichoic acid Tat-pathway Twin-arginine translocation pathway TM Transmembrane TMD Transmembrane domain tRNA Transfer ribonucleic acid TY medium Medium containing tryptone and yeast extract UV Ultraviolet light Q-loop Conserved motif in ATP binding domain of ABC transporters Both one and three letter abbreviations was used for single amino acid residues

1
CONTENTS Preface . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . II Acknowledgements. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . III Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . IV Abbreviations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . V Contents . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 2. Review of bacteriocins in Gram positive bacteria. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 2.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 2.2 Antimicrobial peptides and bacteriocins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 2.3 Historical perspective. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 2.4 Expression . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5 2.5 Classification . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6 2.6 Mode of action . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 2.6.1 Pore-forming bacteriocins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 2.6.2 Inhibition of peptidoglycan biosynthesis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 2.6.3 Bacteriolytic enzymes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 2.6.4 Secondary mode of action of bacteriocins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 2.6.5 Other modes of action . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 2.7 Immunity and resistance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 2.7.1 Efflux ABC transporters. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 2.7.2 Immunity proteins. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12 2.7.3 Immunity to class III bacteriocins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12 2.7.4 Strain resistance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
2.8 Secretion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13 2.8.1 Sec-mediated secretion. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14 2.8.2 Secretion through ABC transporters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14 2.9 Applications . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15 3. Introduction to the practical part . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17

2
4. Genetic and functional characterization of a chromosomally encoded bacteriocin from Bacillus licheniformis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20 5. Secretion analysis of formosin, a bacteriocin from Bacillus licheniformis . . . . . . . . . . . . . . . . . . . . . . 31 6. Immunity associated with formosin production and an investigation of the cellular location of the ForEG ABC transporter. . . . . . . . . . . . . . . . . . . . . . . 45 7. In silico analysis of the ForEG ABC transporter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57 8. Characterization of the formosin accessory protein ForF found in association with the ForEG ABC transporter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63 9. Concluding remarks and future prospects . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75 References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80 Appendix
I. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 90 II . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91 III. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 92 IV . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94

3
CHAPTER 1 INTRODUCTION
The aim of this thesis was to characterize the gene cluster required for functional expression of a bacteriocin from Bacillus licheniformis. This bacteriocin was first discovered by Novozymes A/S in a project with the objective to find, identify, and remove genes of any unnecessary secreted proteins thereby decreasing the work required for purification. During this initial project to remove secreted proteins the gene of this bacteriocin was identified in the chromosome of Bacillus licheniformis ATCC 14580 and subsequently removed. This bacteriocin had a signal peptide similar to that of proteins secreted through the general secretory (Sec) pathway. Production required the expression of three genes in addition to the bacteriocin-gene. These four genes were located adjacent to each other on the chromosome of B. licheniformis. The bacteriocin was shown to be effective against closely related bacteria such as Bacillus subtilis. However, B. subtilis could be used as a heterologous expression host without any bactericidal effect if the four genes from B. licheniformis were expressed in this bacterium. It was suspected that this bacteriocin was secreted through the sec-pathway but it was unknown why three additional genes appeared to be vital for the production and producer immunity. Therefore, the aim of this project was to characterize these three genes with respect to their possible function in the production of and immunity to this bacteriocin. The investigated bacteriocin was encoded in a gene with the ordered locus name BL00275 in the UniProt database (Veith et al., 2004; The UniProt Consortium, 2012). However, it is proposed that this bacteriocin be given the name formosin with the gene being named forD. The three additional genes required for production and producer immunity had the ordered locus names BL00274, BL00273, and BL00272 and were accordingly proposed to be renamed forE, forF, and forG respectively. In this thesis, formosin and the three adjacently expressed proteins were characterized through in silico analysis. For functional studies B. subtilis was used as a heterologous expression host. Proteins involved in production or immunity were determined by the phenotypical properties of strains with different gene patterns of forDEFG. Secretion profile of formosin was established through signal peptide analysis such as N-terminal sequencing, exchange of the signal peptide, fusion to a reporter protein, and analysis of an internal signal able to promote secretion. Moreover, the subcellular locations of the ForE and ForG proteins were fused to a fluorescent protein and visualized through confocal microscopy. The thesis begins with an introductory chapter reviewing studies on bacteriocins of Gram positive bacteria. This is followed by an overview of the practical part which then continues with five chapters describing the methods used and results obtained in this project. The results presented in these five chapters pertain to the characteristics and genomic region of formosin, its secretion mechanism, producer immunity, as well as general characteristics of the ForE, ForF and ForG proteins. In the last chapter final conclusions are drawn and possible future prospects and implications of this thesis are presented.

4
CHAPTER 2 REVIEW OF BACTERIOCINS IN GRAM POSITIVE BACTERIA
2.1 INTRODUCTION This chapter is focused on bacteriocins in Gram positive bacteria where the best described bacteriocins are those produced by lactic acid bacteria. In some instances a few examples of bacteriocin systems in Gram negative bacteria are used as a comparison of the differences. The chapter gives an overview of what bacteriocins are and some main characteristics of the expression, secretion, mode of action, and immunity. 2.2 ANTIMICROBIAL PEPTIDES AND BACTERIOCINS Antimicrobial peptides are found in all domains of life (Bacteria, Archaea, and Eukaryote). In humans, they serve an important part in the innate immune system where they protect against bacteria, fungi, yeast, viruses, and cancer cells (Reddy et al., 2004). In plants, they serve as a natural defense and protect plants against various pathogens. In a microecological milieu, they serve as weapons in a microbiological war over limited resources. When an antimicrobial peptide or protein produced by bacteria kills other bacterial strains it is called a bacteriocin. These bacteriocins are ribosomally synthesized proteinaceous antibacterial compounds, produced and secreted by all major lineages of eubacteria and even found in archaebacteria such as Halophiles (Riley and Wertz, 2002; Torreblanca et al., 1994). These compounds typically exert their antimicrobial action on species closely related to the producer, subsequently killing competitors so that the producer may thrive (Tagg et al., 1976). Bacteriocins are a diverse group of proteins and peptides. Some are post-translationally modified such as the bacteriocin group lantibiotics which contains the non-proteinogenic amino acid called lanthionine. 2.3 HISTORICAL PERSPECTIVE Bacteriocins were originally called ‘colicins’ and were first described in 1925 by André Gratia. In his initial discovery Gratia described the antagonism action exerted by Escherichia coli V towards E. coli Ø (Gratia, 2000). The antagonistic effect was later shown to be caused by a bacteriocin today known as colicin V or microcin V (Cascales et al., 2007). In 1953 the term bacteriocin was proposed, which described the definition of colicin-type bacteriocins since these had thus far been the most widely studied. According to this, bacteriocin was defined as having a narrow bactericidal spectrum with lethal biosynthesis, intra-specific activity, and attachment to specific cell receptors (Tagg et al., 1976). While bacteriocin research had mainly been conducted in Gram negative bacteria, some research into Gram positive bacteria such as lactic acid bacteria had also been performed (Cotter et al, 2005; Rogers and Whittier,

5
1928). An important discovery was made in 1933 when the inhibitory effects of bacterially produced compounds present in milk were demonstrated and although not known at the time this was due to a bacteriocin (Whitehead, 1933). This bacteriocin, today known as nisin, was to become the most widely used bacteriocin. Nisin has thus far been approved as a food additive in over 50 countries since its initial approval in England almost 60 years ago (Cotter et al., 2005; Delves-Broughton, 2005). The introduction of Nisin to the market in 1953 and the concept of utilizing bacteriocins in food applications also shifted the bacteriocin research from Gram negative towards Gram positive bacteria (Heng et al., 2006b; Cotter et al., 2005). As research into bacteriocins in Gram positive bacteria became popular, problems with the original definition of bacteriocins became apparent. In Gram positive bacteria the biosynthesis was not lethal, did not show intracellular activity and some bacteriocins showed a wider bactericidal spectrum (Tagg et al., 1976). No bacteriocin has been introduced to the market on the same international scale as nisin, but bacteriocin research continues. New bacteriocins in various bacteria are continually being discovered and characterized. It has even been predicted that 99% of all bacteria produce at least one bacteriocin (Klaenhammer, 1988; Riley and Wertz, 2002). 2.4 EXPRESSION Genes encoding bacteriocins are typically located in mobile genetic elements such as plasmids, transposons, prophages, and genomic islands. Production of a particular bacteriocin involves the co-expression of other genes that encode proteins with functions such as immunity and when required also secretion, regulation, and biosynthesis of the bacteriocin (Nes et al., 1996; Jack et al., 1995). Bacteriocins and the associated genes are usually expressed in an operon, but may involve expression of up to four separate operons (Heng et al., 2006b). Bacteriocin production has in several instances been found to occur during different growth phases. Two examples of this are enterocin produced by Enterococcus faecium RZS C5 in the early exponential growth phase (Foulquié Moreno, 2003) and lactococcin 972 from Lactococcus lactis IPLA 972, the production of which starts in the late exponential growth phase (Martínez et al, 1999). Furthermore, depending on the transcription regulation system, expression may also be influenced by factors such as carbon source (de Vuyst and Vandamme, 1992; Drosinos et al., 2005; Bárcena et al., 1998), cell-density (Riley and Wertz, 2002), temperature (Diep et al., 2000), or presence of a bacteriocin-sensitive strain (Barefoot et al., 1994). In Gram negative bacteria the transcription regulations of the bacteriocins are typically part of cellular regulons such as the stress induced salt overly sensitive (SOS) regulon (Cascales et al., 2007). Bacteriocin transcription in Gram positive bacteria may be coupled to cellular regulons, but is in many cases regulated through specific bacteriocin expression systems. One such bacteriocin regulation system involves transcriptional repressors such as LtnR which represses the transcription of lacticin 3147 (Cotter et al., 2005; McAuliffe et al., 2001a). Another way to control bacteriocin expression is through quorum sensing. This involves a three-component signal-transduction system consisting of an induction factor, a transmembrane (TM) histidine kinase, and a DNA-binding response regulator (Cotter et al., 2005; Nes et al., 1996). Signal transduction occurs as the induction factor binds to the histidine kinase which activates the response regulator through phosphorylation. The activated response regulator can then e.g. activate transcription of the bacteriocin associated genes. Induction factors are peptide pheromones occasionally observed to be bacteriocin-like peptides, but the

6
bactericidal activity of an induction factor is quite insignificant compared to a real bacteriocin (Anderssen et al., 1998; Nes et al., 1996). Moreover, the induction factor might also be the bacteriocin itself like in the case of e.g. nisin A (Kuipers et al., 1995; Riley and Wertz, 2002). The expression profiles of different bacteriocins are quite diverse, but in general the production is regulated to occur under certain conditions. In Gram negative bacteria this involves a regulation coupled to e.g. a stress response and is typically lethal for the producing cell. In Gram positive bacteria the expression is coupled to e.g. cell-density and is not lethal for the producer. 2.5 CLASSIFICATION There have been various ways of classifying bacteriocins. This includes factors such as size, mode of action, modification, and activity against Listeria. The classification used today has been modified in several steps based on the initial classification scheme proposed by Klaenhammer in 1993. This scheme was composed of four major classes (I-IV) based on their probable structure and mode of action as it was predicted at that time. In this classification; class I was the lantibiotics, class II was small unmodified peptides, class III was larger heat-labile proteins, and class IV was complex bacteriocins with chemical motifs composed of lipids and carbohydrates (Klaenhammer, 1993). Class IV was later removed as no bacteriocin was fulfilling these criteria (Heng and Tagg, 2006a). In 2005 Cotter and colleagues redefined this classification scheme. They proposed a radical change resulting in only two classes, the lantibiotics (class I) and the non-lantibiotics (class II) as well as extensive subdivisions of these two classes. Moreover, they also suggested that class III should not be classified as bacteriocins but instead be named bacteriolysins. This new classification scheme lead to disagreement as seen in a correspondence between Cotter et al. (2006) and Heng and Tagg (2006a). Heng and Tagg (2006a) disapproved of statements made by Cotter et al. (2005) where bacteriocins were described as heat-stable, resulting in e.g. colicins no longer being considered bacteriocins. Another problem with this proposed classification was that it was partial towards bacteriocins produced by lactic acid bacteria that could be used in food applications (Cotter et al., 2005; Heng and Tagg, 2006a; Cotter et al., 2006). Heng and Tagg (2006a) also noted that not all class III bacteriocins are bacteriolytic and should therefore not be named as such.

7
Figure 1. Bacteriocin classification scheme as proposed by Heng et al. (2006b). This included a combination between the original Klaenhammer’s scheme (1993) and the scheme as proposed by Cotter et al. (2005). (Figure used from reference Heng et al., 2006b) In a book on bacteriocins written by Heng et al. (2006b), a more universal classification scheme was proposed. The new classification was a combination of the previous classifications by Klaenhammer (1993) and Cotter et al. (2005) and was designed to fit all bacteriocins produced by Gram positive bacteria (Heng et al., 2006b). This proposed classification (Figure 1) has now become generally accepted with the exception that there are still disagreements on the subdivion of class II and if class IV should be a separate class. Several diverse subdivisions of class II have been described (Klaenhammer, 1993; Cotter et al., 2005; Heng et al., 2006b; Nes et al., 2007; Nissen-Meyer et al., 2009). This is shown in the literature as the subdivisions vary between articles therefore causing confusion. What remains constant in all suggestions is that class IIa are the pediocin-like bacteriocins and IIb are the multi-component (also described as two-component) bacteriocins. The subdivision of class II has not been finally defined. If a bacteriocin is classified as class II, it is required to define which of the classification schemes was used when defining the subdivision to avoid confusion. 2.6 MODE OF ACTION Bacteriocins are usually regarded as having a narrow bactericidal spectrum, acting on bacteria closely related to the producer. However, this is a generalization in need of modification since e.g. nisin exerts its antimicrobial effect on a larger spectrum subsequently being effective against strains such as Listeria and Clostridium (Delves-Broughton, 2005). But the bactericidal spectrum remains relatively narrow due to the fact that bacteriocins produced by Gram positive strains cannot kill Gram negative strains under normal growth conditions (Jack et al., 1995; Cotter et al., 2005). The range of the bactericidal spectrum is dependent on the mode of action of the bacteriocin, presence of the cellular target in a strain, and specificity of the bacteriocin target binding domain. 2.6.1 Pore-forming bacteriocins Pore-forming bacteriocins are produced by both Gram positive and Gram negative bacteria. The size of the pores can vary but acts by disrupting the membrane potential and can in larger pores even cause UV absorbing material to leak out of the cells. The membrane association of these bacteriocins can generally be predicted through hydropathy plots and helical wheel projections.

8
In Gram positive bacteria, pore-forming bacteriocins are observed in class I and II bacteriocins. Pore-formation is either target specific or caused by association of cationic bacteriocins to the negatively charged phospholipids (Cotter et al., 2005). In both class I and II the pore-formations involve a multimeric state but the mechanisms of how this occurs differ. Pore-formation in class II was previously thought to be non-target specific. However, recent research has identified one cellular target of these bacteriocins to be the mannose permease phosphotransferase system (Man-PTS) (Kjos et al., 2011; Diep et al., 2007). Models have been proposed on how these class II bacteriocins interact with the membrane to create a pore (Figure 2). The barrel-stave, also called wormhole, (Figure 2A) and carpet model (Figure 2B) are the two most common models described, but it is predicted that different bacteriocins use different models to create pores. In a recently proposed model of the Man-PTS interacting bacteriocins it has been suggested that it is not the bacteriocins that make the pore. Instead it is proposed that the permease of Man-PTS opens irreversibly due to the binding of the bacteriocin, thereby creating a pore (Kjos et al., 2011).
Figure 2. Mechanism of action for pore-forming antimicrobial peptides. (A)The barrel-stave model, where the hydrophilic sides of the peptides line the pore. (B) The carpet model, where the membrane is enclosed in the peptides that then disrupts the membrane in a detergent-like manner. (C) The toroidal model is similar to the barrel-stave model but the heads of the phospholipids interact with hydrophilic portion of the peptides. (D) The molecular electroporation model, where cationic peptides create an electrical potential difference and as this potential reaches 0.2V a pore is created by molecular electroporation. (E) The sinking raft model, where a mass imbalance of the antimicrobial peptides between the inner and outer leaflet induces pore-formation. The peptides induce a transient pore by sinking into the membrane. This process is induced through self-association and creation of a curvature gradient along the membrane. (Figure adapted from reference Teixeira et al., 2012)
In class I not all peptides/proteins are pore-forming bacteriocins. The pore-formation occurs for the bacteriocins when their α-helical C-terminal dips into the membrane (Cotter et al., 2005; Jack et al., 1995). The pore-formation requires a multimeric state as observed for the class II bacteriocins. Using nisin as a model for the pore-formation event two mechanisms are proposed (Figure 3). One is the barrel-stave model (Figure 3C) and one is the wedge model (Figure 3B) which is similar to the toroidal model (Figure 2C), except that in the wedge model the bacteriocin remains on the surface of the phospholipids heads (Hoffmann et al., 2002). The

9
pore-formation is initiated via the interaction with the cell wall precursor lipid II. However, it has also been suggested that at higher concentrations of class Ia lantibiotics, these proteins are capable of forming non-target-specific wedge like pores (Héchard and Sahl, 2002).
Figure 3. Proposed mechanisms of pore-formation for nisin. (A) Nisin binds to lipid II thus inhibiting cell wall synthesis. (B) In the wedge model Nisin remains surface bound interacting with the phospholipid heads causing the membrane to distort and create a pore. (C) In the barrel-stave or wormhole model the hydrophobic side of the C-terminal α-helix of nisin interacts with the hydrophobic lipid tails to create a pore. (Figure from reference Hoffmann et al., 2002) In a Gram negative system such as for colicin A or B the bacteriocins must first be imported into the periplasm before they can interact with the cytoplasmic membrane leading to pore-formation. This involves a two step process which starts with the receptor binding to an outer membrane protein and is followed by the import process facilitated by either the Ton- or Tol-system (Braun et al., 1994; Kleanthous, 2010). When a pore-forming bacteriocin such as colicin A, interacts with the cellular membrane it undergoes a conformational change. While the exact mechanism of the pore-formation event is not yet fully understood it is known that each colocin molecule forms a pore (Cascales et al., 2007). 2.6.2 Inhibition of peptidoglycan biosynthesis Inhibition of cell wall biosynthesis is observed for both class Ia and Ib lantibiotics. The cell wall inhibition occurs as the lantibiotics interact with the lipid II cell wall precursor, thus preventing growth of the peptidoglycan chain. It is known that while some linear lantibiotics (class Ia) interact with both lipid I and lipid II cell wall precursors – globular lantibiotics (class Ib) only interact with lipid II (Bierbaum and Sahl, 2009). Moreover, the binding sites in lipid II are different in linear (Figure 3) and globular (Figure 4) lantibiotics (Héchard and Sahl, 2002).

10
Figure 4. Lipid II binding of globular lantibiotics (class Ib) causing inhibition of the cell-wall synthesis, a function also observed for class Ia. (Figure taken from reference Héchard and Sahl, 2002) Lactococcin 972 is another cell wall acting bacteriocin determined to specifically inhibit septum formation through interaction with lipid II (Martínez et al., 2000; 2008). This is a novel bacteriocin in the fact that it is the first non-lantibiotic bacteriocin demonstrated to bind lipid II (Martínez et al., 2008). Since lactococcin 972 inhibits septum formation it is only active against growing cells with no effects against non-dividing cells (Martínez et al., 2008). It should be noted that Martínez and colleagues did not draw a parallel between their observed septum inhibition and the fact that lipid II binding nisin and vancomycin are found to be more frequently attracted to the cell-division sites (Kramer et al., 2008). Moreover, it is also known that the septum is more frequently affected by the secondary mode of action (see section 2.6.4) where the cellular autolytic enzymes are activated (Jack et al., 1995; Sahl and Bierbaum, 1998). A comparison between the septum inhibition of lactococcin 972 and these mentioned facts would have made for an interesting discussion e.g. could this be an effect of a transient nature of the interaction between lipid II and lactococcin 972? 2.6.3 Bacteriolytic enzymes This is the mode of action observed in class IIIa bacteriocin such as lysostaphin. These are bacteriocins that function as zinc metalloproteases causing hydrolysis of the cell wall in sensitive strains (Kumar, 2008). They have two separate domains, an N-terminal peptidase domain and a C-terminal substrate binding domain. The N-terminal typically hydrolyses the pentaglycine interpeptide bridge in the cell wall, while the C-terminal determines the range of the bactericidal spectrum (Baba and Schneewind, 1996; Nilsen et al., 2003). 2.6.4 Secondary mode of action of bacteriocins For some bacteriocins, sensitive cells start lysing soon after exposure to the bacteriocin. This is an effect not caused by the disruption of the membrane potential, but rather the secondary mode of action of these bacteriocins (Jack et al., 1995). In order for the cells to start lysing the cell wall needs to be degraded. This is caused by the autolytic system of the bacteriocin-sensitive cells, not the bacteriocin. The autolytic system causes cell-lysis through two diverse mechanisms both of which have been shown to occur with different bacteriocins. In some instances the energy depletion caused by the pore-formation event leads to an imbalance between breaking down and rebuilding the cell wall in cultures of growing cells (Martínez-Cuesta et al., 2000). For cationic bacteriocins the autolytic enzymes are activated as these bacteriocins interact with the teichoic acids in the cell wall, thereby releasing the autolytic enzymes that are bound to these cell wall

11
components in their non-active state (Bierbaum and Sahl, 1987). This activation of the autolytic enzymes is observed to occur more frequently in the septum between two daughter cells (Jack et al., 1995). 2.6.5 Other modes of action In Gram positive bacteria the main modes of action are pore-formation and inhibition of cell wall synthesis as described above. Another previously mentioned mode of action is the enzymatical break down of the cell wall as observed for class IIIa bacteriocins. However, other modes of action have also been observed for bacteriocins in Gram positive bacteria which include quorum sensing and inhibition of spore outgrowth. In quorum sensing, e.g. nisin and subtilin act as pheromones promoting their own expression through a three-component signal-transduction system (Kleerebezem, 2004). These two bacteriocins also inhibit spore outgrowth through an uncommon didehydroalanine residue in position 5, which interacts with the spore-associated factor required for outgrowth (Sahl and Bierbaum, 1998). The mode of action for class IIIb bacteriocins is still unknown and their classification is based on their non-lytic mode of action compared to the bacteriolytic nature of class IIIa (Section 2.6.3). The mode of action of class IIIb is thought to involve interruption of the proton motive force, thus indicating membrane interaction (Heng et al., 2006b). However, in all known proteins of this class the function remains unknown. Many bacteriocins in Gram negative bacteria have an intracellular function which includes enzymatical modes of action such as DNase and RNase (Cascales et al., 2007). This has not been observed for bacteriocins in Gram positive bacteria. 2.7 IMMUNITY AND RESISTANCE Immunity is conferred through an ABC transporter and/or a specific immunity protein which are typically co-expressed with the bacteriocin in the producer (Bierbaum and Sahl, 2009). However, in the evolution of bacterial warfare the bacteriocin sensitive strains can become resistant due to the selective pressure of the bacteriocin presence. This resistance involves either structural or physiological changes in the strain depending on the mode of action for that particular bacteriocin. 2.7.1 Efflux ABC transporters ABC transporters responsible for conveying immunity are found to consist of one to three proteins. These transporters have a typical architecture consisting of four domains, two transmembrane domains and two nucleotide binding domains. They are quite diverse but can be recognized by characteristic features of the nucleotide binding domain (Wen and Tajkhorshid, 2011). In some bacteriocin systems an accessory protein is present, although the function of this protein is still unknown (Heng et al., 2006b). The immunity function is thought to be similar to that of the efflux observed in multidrug resistance ABC transporters except that they have more specific target recognition (Heng et al., 2006b). This mechanism of immunity is observed for both class I and class II bacteriocins where the ABC transporter causes efflux of membrane associated bacteriocin into the medium. These ABC transporters are in some cases found to work in synergy with the immunity protein described in the next section. One example of this is NisI and NisFEG where the immunity to nisin is increased as both immunity functions are present (McAuliffe et al., 2001b; Heng et al., 2006b).

12
2.7.2 Immunity proteins Immunity proteins are observed to be either membrane associated, secreted, or intracellular proteins (Heng et al., 2006b). The dedicated immunity proteins are generally encoded in the same region as the associated bacteriocins and in some cases they are even co-transcribed (Nes et al., 2007). Immunity proteins associated with lantibiotics (class I) are found to be either membrane associated or secreted. This membrane association can either be as membrane proteins with an N-terminal hydrophobic part e.g. PepI or as lipoproteins e.g. NisI (McAuliffe et al., 2001b; Heng et al., 2006b). Some of these lipoproteins are found in both a membrane attached state and a secreted state. This is the case of e.g. NisI, where only half of the secreted proteins undergo lipid modification subsequently resulting in only half being membrane attached (Koponen et al., 2004). The size of these proteins is quite diverse ranging from the small 69 amino acid long immunity protein (PepI) of Pep5 to the larger 245 amino acid long immunity protein (NisI) of NisA (Heng et al., 2006b). Cross-immunity has been demonstrated between immunity proteins associated with similar bacteriocins such as e.g. PepI and EciI or NisI and NsuI. However, in general these immunity proteins are considered to be bacteriocin-specific (McAuliffe et al., 2001b; Heng et al., 2006b). Class II is a quite diverse class of bacteriocins which is also reflected in the functions of the immunity proteins. In general these proteins share little homology, making them fairly specific to their individual bacteriocin (Drider et al., 2006; Nes et al., 2007). The specificity is conveyed by the C-terminal of the immunity protein (Johnsen et al., 2004). Their specific immunity function is quite elusive to this date and it has never been determined that any physical contact is made between the immunity protein and the associated bacteriocins (Kjos et al., 2011). Only one immunity function has been resolved thus far and it is only relevant to the class II bacteriocins known to interact with Man-PTS. The immunity proteins of these bacteriocins associate loosely to Man-PTS, but when a bacteriocin binds to Man-PTS the binding efficiency increases for the immunity protein which also increases binding efficiency of the bacteriocin, thereby locking the bacteriocin in place (Diep et al., 2007). However, it is still not known how the Man-PTS targeting bacteriocins exert their pore-formation and thus one cannot predict how this immunity protein functions to prevent it. Membrane bound proteases from the Abi protein family have also been observed to be associated with immunity to class II bacteriocins. In comparison to the other immunity proteins described they do exhibit cross-immunity against other bacteriocins. Their substrate has not yet been identified, but is not the bacteriocin itself. It has therefore been suggested that these Abi proteases either break down the cellular target of the bacteriocin or that they work through degradation of an anti-sigma factor thus changing the transcriptional profile of the Abi expressing cells in a way which conveys the immunity. (Kjos et al., 2010) 2.7.3 Immunity to class III bacteriocins The function of the immunity proteins associated with class IIIa is not fully understood. However, these immunity proteins are co-expressed in the plasmids encoding the bacteriocin. Since these bacteriocins are enzymes that break down the cell wall, it is suspected that the immunity proteins convey immunity by altering the cell wall structure in some way. This has been observed to be the mode of action of the immunity protein (Epr) associated with lysostaphin that changes the composition of the cell wall by addition of serine (Sugai et al., 1997; Heng et al., 2006b). Little is still known about the mode of action for class IIIb bacteriocins, so no predictions can been made on the functions of their producer immunity.

13
2.7.4 Strain resistance The mechanism of resistance (Table 1) can vary depending on the mode of action of the produced bacteriocin. The resistance could be a temporary shift involving cellular modifications such as changes in membrane fluidity, surface charge, and cell wall thickness. It may also be a more permanent resistance such as a mutation causing the cells to e.g. lack the ability to express a cellular target such as Man-PTS. In general, mutations are only an advantage under the selective pressure of a present bacteriocin. Under non-selective pressure the resistant strains most often lose their advantage and are quickly outcompeted. Cross-resistance is quite common since many bacteriocins are pore-forming peptides with cationic properties. Changes such as e.g. membrane fluidity, surface charge, and anionic capsule production will decrease the interaction efficiency with the membrane for any cationic, pore-forming bacteriocin thus causing cross-resistance. Resistance becomes a concern when the binding site of the cellular target is mutated to decrease binding efficiency of the bacteriocin or if the resistant strains acquire e.g. an immunity protein or an efflux ABC transporter. Since horizontal gene transfer occurs between various strains this may result in sensitive strains acquiring efflux ABC transporters, extracellular proteases, Abi-proteases, and immunity proteins. However, nisin has been used for over 50 years and no significant resistance has been observed thus far (Heng et al., 2006b). Table 1. Examples of mechanism of resistance against bacteriocins.
Mechanism of resistance Example
Down regulation of cellular target Man-PTS expression levels decrease or stop (Kjos et al., 2011)
Change of membrane fluidity More saturated PG1 (Limonet et al., 2004), More unsaturated PG (Vadyvaloo et al., 2002)
Decreased negative surface charge D-alanylation of TA2, Lysinylation of PG (Vadyvaloo et al., 2004)
Anionic capsule L. lactis produces exopolysaccharides which are anionic in nature due to the addition of phosphate groups (Looijesteijn et al., 1999)
Increased cell wall thickness Abnormal cell wall synthesis and autolysin inhibition (Maisnier-Patin and Richard, 1996)
Inhibition of cell-communication The secreted Sgc protein from Streptococcus gordonii interferes with quorum sensing in Streptococcus mutans required for bacteriocin production (Wang and Kuramitsu, 2005)
1 PG, Phosphatidylglycerol 2 TA, Teichoic acid 2.8 SECRETION In Gram negative bacteria, bacteriocin production has a lethal outcome for the producer. This is circumvented in Gram positive bacteria due to their producer immunity (section 2.7) and secretion mechanisms. Bacteriocins are typically secreted through dedicated transporters (ABC transporters), but some bacteriocins have been observed to be secreted through the general secretory pathway (Sec-pathway). The distinction of which translocation pathway is used is related to the properties of the N-terminal signal peptide (Tjalsma et al., 2004).

14
2.8.1 Sec-mediated secretion Sec-dependent signal peptides are distinguished by the presence of three domains and absence of a twin-arginine motif. The distinct domains consist of a positively charged N-domain, a hydrophobic H-domain which should be large enough to span the membrane, and finally a C-domain with the signal peptidase recognition site (Tjalsma et al., 2004). Some class II bacteriocins have been predicted, through the properties of their N-terminal signal peptide, to be secreted through the Sec-pathway. Moreover, enterocin P has been confirmed to be a sec-pathway secreted bacteriocin through signal peptide mutations and the use of the SecA inhibitor sodium azide (Herranz and Driessen, 2005). Examples of other bacteriocins predicted to be secreted through the sec-pathway are lactococcin 972 (Martínez et al., 1999), divergicin A (Worobo et al., 1995), and bacteriocin T8 (De Kwaadsteniet et al., 2006). 2.8.2 Secretion through ABC transporters Signal peptides directing secretion through ABC transporters are typically called leader peptides and are distinguished from Sec-dependent signal peptides by their lack of a hydrophobic H-domain (Tjalsma et al., 2004). These leader peptides are usually described as having a double-glycine motif (GG/GA/GS). However, other leader peptide types are known such as the FNLD-type or the quite different leader peptide of e.g. mersacidin. Recently some leader-less bacteriocins have been found which are thought to be secreted by ABC transporters. Leader peptides of the GG-type (Figure 5) are secreted and processed by a particular type of ABC transporter. These transporters are homodimers constructed by a protein with an N-terminal protease domain and a C-terminal ATP binding domain linked together via a middle transmembrane domain (Havarstein et al., 1995). The protease domain is a cystein protease and belongs to the peptidase C39 protein family (Nes et al., 2007). This is the most abundant ABC transporter leader peptide and is observed for most class II bacteriocins and some class I lantibioics (Nes et al., 1996). Some globular lantibotics such as cinnamycin and mersacidin have a different leader peptide which is longer and does not contain the GG-motif. However, these globular lantibiotics are still observed to be associated with the same type of ABC transporter as the GG-type leader peptides (Altena et al., 2000). Leader peptides of the FNLD-type (Figure 5) have a serine protease recognition motif and are found in class I lantibiotics such as nisin. This leader peptide is not processed by a protease domain within the ABC transporter as seen with the GG-type leader peptides. Instead this leader peptide is cleaved off after secretion by an extracellular serine protease that either is a special purpose one co-expressed with the lantibiotic (e.g. nisin) or a non-bacteriocin-specific protease from the producer strain (e.g. subtilisin). (Bierbaum and Sahl, 2009; Heng et al., 2006; McAuliffe et al., 2001)

15
Figure 5. Sequence alignment of leader peptides from bacteriocins known to belong to either GG-type or FNLD-type. Lactococcin A and Pediocin PA-1 are class II bacteriocins, while the other leader peptides belong to class I lantibiotics. The alignment was performed using ClustalW where the *(asterisk) represents a conserved amino-acid, a : (colon) indicates conservation between amino-acids of strongly similar properties, and a . (period) represents conservation between amino-acids of weakly similar properties. Conserved regions in these two types of leader peptides are indicated by blue boxes. Both the GG-type and the FNLD-type leader peptides contain negatively charged amino acids making the netcharge of the leader peptide neutral or slightly negative. The leader peptides are also shown to inhibit the activity on the bacteriocin, and are therefore thought to be a protection for the producer (Bierbaum and Sahl, 2009). Leader less bacteriocins that are secreted through ABC transporter systems have recently been discovered. These ABC transporters are sometimes also found to be involved in the immunity to the bacteriocin. The leader less peptides were first thought to be hemolytic peptides, but were later found to also be bactericidal. (Nes et al., 2007) Not much is known about their specific secretion mechanism, but in one instant a multidrug resistance ABC transporter is observed to function in the secretion and immunity (Gajic et al., 2003). 2.9 APPLICATIONS For more than 50 years nisin has been used as a food preservative. Nisin is produced by L. lactis, a bacterium generally regarded as safe (GRAS). This bacteriocin is effective against Gram positive bacteria and some spores, but could in a combination treatment with chelators (e.g. EDTA) be effective against Gram negative bacteria (Jack et al., 1995). Nisin (e.g. Nisaplin®) is thus far the only purified bacteriocin approved as a food additive on an international scale, although some variations might exist within different countries. However, there are three ways of using bacteriocins as food additives. First, one can add the purified bacteriocin as in the case of nisin (Delves-Broughton, 2005). Secondly, one can add fermentate from bacteriocin producers which also enhances flavor e.g. Microgard™, Alta™, and Perlac™ (Lemay et al., 2002). Lastly, one can add a bacteriocin producer such as various strains of L. lactis being used in production of yogurt, cheese, and other fermented foods. This is one of the major reasons why bacteriocin research still remains such a large subject in microbiology with GRAS bacteria being the key focus.

16
Bacteriocins are also of interest for medical applications. Since antibiotic-resistant strains are becoming more common, alternative treatments are being researched. Bacteriocins have in some cases been shown to be effective against these antibiotic-resistant bacteria. One example of this is the two component lacticin 3147 lantibiotic that has been found effective against pathogens such as methicillin-resistant Staphylococcus aureus (MRSA) and vancomycin-resistant Enterococcus faecalis (VRE) (Galvin et al., 1999). Bacteriocins used for a medical application must go through clinical trials which is very expensive. In November 2011 a phase I clinical trial of a bacteriocin referred to as NVB302 was initiated by a UK company called Novacta Biosystems (Novacta News, 2011). Bacteriocins are also used as probiotics. One such product is an oral probiotic, BLIS K12, which contains strains that produce e.g. the lantibiotic salivaricin A2 (Wescombe et al., 2006). This product was introduced to the market by one of the well-known scientists in bacteriocin research, John Tagg. Other probiotic products are found to contain fermentate of different strains and can even be marketed as bacteriocin-containing products e.g. Dr. Ohhira's, Probiotics, Essential Formulas (Essential Formulas Incorporated, 2011). Another large market for bacteriocins is as additives in animal feed. Addition of either fermentate from bacteriocin producers or bacteriocin producer strains to animal feed could promote a healthier gastric intestinal tract and fewer instances of food contamination by dangerous pathogens (e.g. Cole et al., 2006). There are advantages in using bacteriocins for food applications, since they are considered a natural product and are therefore described as natural preservatives (Cotter et al., 2005). If used in animal feed, bacteriocins in combination with prebiotics could reduce the large quantities of antibiotics given to farm animals today, which is a debated subject. In medical applications Man-PTS interacting bacteriocins could be utilized since this is a unique cellular target distinct from those of antibiotics. There are also disadvantages concerning the use of bacteriocins. One of these being the cost, as chemical preservatives are cheaper than bacteriocins. The developmental cost of bacteriocins for medical applications is very high compared to profits, unless clinical trials are sponsored by the government. Another concern is different adverse effects caused by the administered bacteriocins. This could include effects like disturbing the natural healthy flora but also more dangerous effects such as hemolysis of eukaryotic cells.

17
CHAPTER 3 INTRODUCTION TO THE PRACTICAL PART Introduction The aim of this thesis was to investigate the gene cluster required for functional expression of formosin, with a special regard to secretion and producer immunity. This was achieved by the characterization of genes adjacent to the formosin gene in a gene cluster from Bacillus licheniformis. The gene luster consists of four genes with the ordered locus name BL00275-BL00272 in the UniProt database (Veith et al., 2004; The UniProt Consortium, 2012). In this thesis the gene cluster was predicted to be an operon and the genes were recommended to be named forD-forG (Chapter 4). This for-operon (Figure 1) was found to be comprised of genes encoding a 14kDa pre-formosin (ForD), a 24.5kDa ATP binding protein (ForE), a 12.8kDa accessory protein (ForF), and a 71kDa transmembrane (TM) protein (ForG). ForE and ForG was in this thesis found to constitute an ABC transporter referred to as the ForEG ABC transporter (Chapter7).
Figure 1. The for-operon from Bacillus licheniformis consisting of the genes BL00275-BL00273 as assigned by UniProt, but have been given the gene names forD, forE, forF, and forG. This for-operon encodes for the bacteriocin called formosin, an ATP binding protein, an accessory protein, and a transmembrane protein. In the research performed in this thesis the for-operon was expressed in Bacillus subtilis for the purpose of isolating and identifying the genes required for formosin production and immunity. B. subtilis had prior to this thesis been observed to be sensitive to the antibacterial activity of formosin. When forD was inserted into B. subtilis, recombinants could be isolated but no formosin was produced. In contrast, when the entire for-operon was inserted into B. subtilis, formosin was produced and secreted in large amounts. In the following, a short summary will be given of the individual chapters in this thesis.

18
Chapter 4 Genetic and functional characterization of a chromosomally encoded bacteriocin from Bacillus licheniformis In this initial study, properties of formosin and the genomic region encoding the for-operon were investigated to gain insight into this bacteriocin system. Results of this study showed that formosin was expressed in a genetic region observed to be a genomic island with host defensive properties. Genes involved in formosin production were predicted to be arranged in an operon referred to as the for-operon. The antibacterial properties of formosin were demonstrated in B. subtilis, but did not result in a determination of the mode of action of formosin. However, it was determined that formosin had a secondary mode of action that caused cell lysis. The antibacterial activity of formosin was shown to be resistant to changes in pH, but was inactivated at temperatures of 50°C and over. Moreover, formosin was seen to belong to the lactococcin 972 family and results obtained in this thesis are therefore compared to lactococcin 972. Results from this study also suggest that the current model of the lactococcin 972 family might be incomplete. The current model proposes that this bacteriocin family is associated with a seven TM immunity protein (Martínez et al., 1999). However, in this study it is shown that this distinctive TM protein is part of an ABC transporter. This was demonstrated by a comparison of genes to adjacent lactococcin 972-like proteins and was further supported by the in silico analysis performed in Chapter 7. Chapter 5 Secretion analysis of formosin, a bacteriocin from Bacillus licheniformis The aim of the study presented in this chapter was to investigate the secretion profile of formosin. For this purpose different secretion analyses was performed in B.subtilis. These analyses included a determination of which genes in the for-operon were required for formosin secretion and an analysis of the “secretion promotion” of either the N-terminal signal peptide of formosin and formosin itself. Results showed that formosin contained two secretion signals. One signal peptide was situated in the N-terminal and had features similar to that of a sec-dependent signal peptide. The other signal peptide was situated in the C-terminal and promoted secretion through the ForEG ABC transporter. The N-terminal signal peptide was cleaved in both secretion pathways, but formosin could only be detected in the medium when secreted through the ForEG ABC transporter. Result that supports this is that the last 24 amino acids in the C-terminal of formosin were shown to promote secretion of a reporter protein in the presence of the ForEG ABC transporter. Further evidence to support the suggestion of a C-terminal signal peptide is reported in Chapter 8 where a homologue to the formosin system was found in Jonesia denitrificans. The formosin homologue in this system contained no N-terminal signal peptide, but showed 87% sequence similarity to the last 45 amino acids of the C-terminal of formosin. This is the first predicted C-terminal secretion signal observed in Gram positive bacteria. Chapter 6 Immunity associated with formosin production and an investigation of the cellular location of the ForEG ABC transporter In Chapter 4 it was demonstrated that B. subtilis is sensitive to the antibacterial actions of formosin. In contrast, expression of the for-operon in B. subtilis described in Chapter 5 resulted in a high production of formosin. Consequently, the aim of this study was to investigate the cause of this observed producer immunity. For this purpose, the genes in the for-operon were investigated for their immunity properties. Results showed that the ForEG ABC transported conveyed a small increase of immunity to formosin. This immunity was demonstrated in a competition study to be sufficient to outcompete a non-formosin-producing B. subtilis strain. The second aim of this of this study was to investigate the subcellular location of the ForEG ABC transporter. For this purpose, green fluorescent protein (GFP) was fused to the C-terminal of either the ForE or

19
the ForG protein so that their location within the cells could be determined in a confocal microscope. Results of this study indicated that the ForEG ABC transporter was localized in microdomains within the membrane similar to those that has been previously described for other protein translocation pathways (Buist et al., 2006). Chapter 7 In silico analysis of the ForEG ABC transporter In this study the ForEG ABC transporter was characterized in an in silico analysis. For this purpose, domains known to be conserved within the ATP binding part of ABC transporters were identified within the ForE protein. In contrast TM domains of ABC transporters do not contain conserved regions. Instead a topology model of ForG was predicted that could be compared to other TM proteins associated with ABC transporters. Furthermore, possible interactions sites between the ForE and ForG are discussed through the current knowledge of ATP binding proteins and conserved regions observed in the intracellular loops of the ForG protein. Chapter 8 Characterization of the formosin accessory protein ForF found in association with the ForEG ABC transporter In the previous chapters the functions and characteristics of formosin and the ForEG ABC transporter were investigated. However, the properties and function of the ForF accessory protein remained unknown. Hence, this last chapter was devoted to the characterization of this accessory protein. During this study the function of the accessory protein was tested with regards to secretion of formosin, but no apparent function could be observed. In Chapter 6 it was also observed that ForF had no apparent immunity properties by itself. In the characterization it was predicted that ForF was a single spanning membrane protein with an N-terminal TM region. The ForF protein was observed to be associated with the ABC transporter, as overexpression of this protein in the presence of ForE and ForG resulted in the ForF protein appearing in the medium. This was not predicted to be the result of secretion since the size of ForF seen in the medium indicated that the protein still contained the TM region. During the analysis of the ForF protein two homologues of this protein were found in J. denitrificans. These two ForF-homologues were both associated with the same ABC transporter, identified to be a homologue to the ForEG ABC transporter. Moreover, ForF was also compared to other accessory proteins adjacent lactococcin 972-like proteins with regards to size and predicted secondary structure. This comparison demonstrated the diversity of accessory protein associated with protein secreting ABC transporters in Gram positive bacteria.

20
CHAPTER 4 Genetic and functional characterization of a chromosomally encoded bacteriocin from Bacillus licheniformis
INTRODUCTION In microbiological warfare one strategy employed to facilitate niche competition is the secretion of small peptides or proteins that kill or inhibit the rival bacteria. These are called bacteriocins and are ribosomally synthesized proteins produced by both Gram positive and Gram negative bacteria (Tagg et al., 1976). Bacteriocins were first defined as bactericidal proteins effective against closely related bacteria (Tagg et al., 1976), but as new bacteriocins have been found, this definition has changed to include bacteriostatic, broad spectrum bacteriocins, and bacteriocin which undergo post-translational modifications (Cotter et al., 2005). Most bacteriocins are encoded within different types of mobile genetic elements that could be self-replicating or integrated into the bacterial chromosome (Jack et al., 1995). Production of a particular bacteriocin typically involves expression of several different genes that encodes proteins involved in the biosynthesis, secretion or immunity to the bacteriocin (Jack et al., 1995). Secretion of bacteriocins involves either a special purpose ABC transporter or the general secretory (sec) pathway (Drider et al., 2006). When secretion occurs through the sec pathway the N-terminal signal peptide has a typical sec-dependent signal peptide containing three domains consisting of a positively charged N-domain, a hydrophobic H-domain, and a C-terminal with the signal peptide cleavage recognition site (Tjalsma et al., 2004). In transport mediated by an ABC transporter the signal peptide, called leader peptide, lacks the H-domain and contains a double glycine motif in the C-terminal which mediates the signal peptide cleavage (Tjalsma et al., 2004). The present study reports on the characterization of formosin (ForD), a novel bacteriocin from Bacillus licheniformis ATCC 14580. This is a chromosomally encoded protein associated with three other genes present in the same locus as forD, named forE, forF, and forG. This study was aimed at describing the properties of formosin with regards to the genetics, effect against Bacillus subtilis, and homology to other known bacteriocins. The chromosomal region adjacent to the forD gene was investigated and an in silico model of the formosin associated gene cluster was made to predict if these four genes (forD-forG) were located in an operon. The antimicrobial property of formosin was demonstrated using B. subtilis. Furthermore, formosin was screened for sensitivity to changes in environmental factors such as pH and temperature. Results obtained in this study indicate that formosin is expressed in a polycistronic operon together with three other genes, two of which are coding for an ABC transporter and one being an unknown accessory protein. This operon was observed to be located in a chromosomal region defined as a genomic island with host defensive properties most likely obtained through horizontal gene transfer. Formosin was shown to be a pH tolerant but heat sensitive bacteriocin that has a fast acting bacteriolytic effect on B. subtilis. Formosin was observed to

21
have little sequence homology to any known and characterized proteins, but was observed to belong to the lactococcin 972 family. The current knowledge of the lactococcin 972 family has been defined by the research performed on lactococcin 972. Results from the study of formosin and the comparisons performed in this present study of different proteins belonging to the lactococcin 972 family revealed new insight into this bacteriocin family, including a new definition of proteins associated with the lactococcin 972 family. In the old model the bacteriocin is found to be associated with a seven transmembrane (TM) protein predicted to be an immunity protein (Martínez et al., 1999). In the new model defined by the results in this study it is instead proposed that the members of the lactococcin 972 family are associated with the expression of an ABC transporter with a characteristic topology of seven TM regions with two large extracellular domains. MATERIALS AND METHODS
Bacterial strain and growth conditions. The bacterial strain used in this study was B. subtilis 168, RUB 200 derivative JA1343 (amyE aprE nprE spoIIAC) which is a protease and sporulation deficient strain. Cells were grown at 37°C in liquid TY medium or Müeller-Hinton II Broth (MHB) and plated on LBPG agar plates (LB agar with 0.5% (w/v) glucose and 50mM potassium phosphate, pH 7.0). In the different tests purified formosin was added to the cells at various concentrations. Purified formosin was provided by the Protein Biochemistry department at Novozymes A/S. Growth experiment of B. subtilis 168 with ForD addition. B. subtilis 168 (JA1343) cells were grown at 37°C in liquid TY medium with a rotation speed of 250rpm. Growth tests were performed in 50mL cultures that were inoculated with exponentially growing JA1343 cells resulting in an OD600 of approximately 0.02. Formosin was added to the cultures when an OD600 of approximately 0.5-0.6 was reached. This addition was performed by adding 1mL of different dilutions of formosin to each culture, resulting in a final concentration of 0, 0.5, 1, 1.5, and 2 µg/mL of formosin. The progression of the cultures was then monitored for several hours through optical density measurements to observe the effect of the formosin addition. Sample preparation and western blotting analysis. During the growth experiment samples were taken from the cultures. They were prepared for SDS-PAGE by adding one part NuPAGE® LDS sample buffer (4x) to three parts sample and then incubated at 99°C for 3min and stored on ice. Samples used for SDS-PAGE were the samples taken from the culture that showed the highest inhibition that still recovered from the addition of formosin. From these samples 10µL was loaded to a precast NuPAGE® 4-12% Bis-Tris gel (Invitrogen). The gel was run with NuPAGE® MES SDS Running Buffer at 200V for 35min. To reduce background in the western blotting analysis, the gel was incubated 30min in MilliQ water. Blotting was performed for 7min using the iBlot® Dry Blotting system (Invitrogen) to a Nitrocellulose Transfer Stack designed for this system. Immunoblotting was made with rabbit primary antibodies against formosin (manufactured internally by Novozymes A/S) in combination with a Novex® WesternBreeze™ Chemoluminescent Anti-Rabbit kit (Invitrogen) using recommended procedures. Immunodetection was made by the provided reaction buffers from the WesternBreeze™ kit. Detection was made and photographed using BioSpectrum® Imaging System (UVP) in combination with VisionWorks®LS Analysis Software (UVP) using the exposure time of 20min. The images were processed in Adobe Photoshop® CS3 (Adobe Systems) by inverting the colors. Stability test of formosin to temperature and pH. To characterize the stability of formosin, effects of temperatures and pH were investigated. To determine the effect of different temperatures, samples of formosin were incubated for 30min at temperatures of 0°, 20°, 40°, 50°, 80°, and 100°C. To test the effect of

22
different pH, formosin was diluted to 2mg/mL in a solution of 0.01 % (w/v) acetic acid. Using sodium hydroxide or hydrochloric acid the pH was adjusted to the desired value ranging from pH 2-11. The solutions were sterilized by filtration and left at 4°C overnight and from the volume of each pH sample the end concentration was calculated. To test the bacteriocin activity microtiter plates were used containing MHB inoculated with 105cell/mL. The formosin solutions were added to a final concentration of 100µg/mL, which is more than ten times the concentration needed for a bactericidal effect (Chapter 6). The microtiter plates were incubated overnight at 37°C with a rotation speed of 300rpm. In silico analysis of the forD gene cluster. The DNA sequence encoding the forD gene cluster was analyzed with regards to transcription initiation and termination to predict if the genes in the gene cluster was a part of an operon. To find transcription terminators, programs such as FindTerm (Softberry), WebGeSTer (Mitra et al., 2011), TransTermHP (Kingsford et al., 2007), and ARNold (Macke et al., 2001; Gautheret and Lambert, 2001) were used. These programs predict stem-loop structures which are characteristic of rho independent terminators. From the resulting stem-loop structures the free energy, base pairing and presence of low energy base paring downstream of the stem-loop was evaluated and the probable terminators were presented. Finding potential promoters was made in two ways. First potential promoters were identified by searching for sigma factor A (σA) like sequences in the upstream region of the forD, forE, and forG. Secondly, more uncommon promoters was attempted to be identified through the search of the databases DBTBS (Ishii et al., 2001) and Virtual Footprint (Münch et al., 2005). Results from these databases were compared to each other and to the weight matrix of the aligned promoter sequences. In addition to the transcription initiation and termination the translation initiation was also evaluated for the four genes in the gene cluster by identifying possible Shine Dalgarno sequences (Shine and Dalgarno, 1975). Identification and characterization of homologues to formosin. A search for proteins similar to the formosin protein was performed in two ways. First a Basic Local Alignment Search Tool (BLAST) search was performed against the UniProtKB database (The UniProt Consortium, 2012) and secondly Pfam was used to identify if formosin contained any domains similar to a known protein family (Punta et al., 2012). Since the results showed that formosin belongs to the lactococcin 972 family, five different proteins belonging to this family derived from five different Gram positive bacteria were chosen for comparison. This included formosin, Lactococcin 972 and three other putative lactococcin 972-like proteins. Signal peptides were predicted using SignalP 4.0 (Petersen et al., 2011). The predicted mature proteins were characterized for relevant properties such as size, isoelectric point (pI), secondary structure, and presence of hydrophobic residues using the calculation program ProtParam (Gasteiger et al., 2005). Information of the pI will disclose if the protein is a cationic protein and the hydrophobic and aliphatic residues may give indications to the amphipathicity of the protein. The secondary structure of the mature protein was predicted using PSIPRED v3.0 (Jones, 1999) and the genes coded in the same region of the plasmid or chromosome as the lactococcin 972-like protein were identified and analyzed for function. Characterization of the genomic region adjacent to the forD gene. Genes in the region of 4140-4175kbp of the B. licheniformis chromosome were identified and characterized. The genes were identified by a BLAST search against the UniProtKB database (The UniProt Consortium, 2012) and further analyzed for known motifs or domains by Pfam (Punta et al., 2012) or InterProScan (Quevillon et al., 2005). To identify the five pseudogenes found in the region these were translated in all three reading frames and from the resulting translation the larger generated sequences were used in a protein BLAST. The identified protein coded by each pseudogene could then be analyzed as described for proteins coded by the other genes in the region. The functions of the genes in the investigated region indicated that this could be a genomic section that had been inserted from a
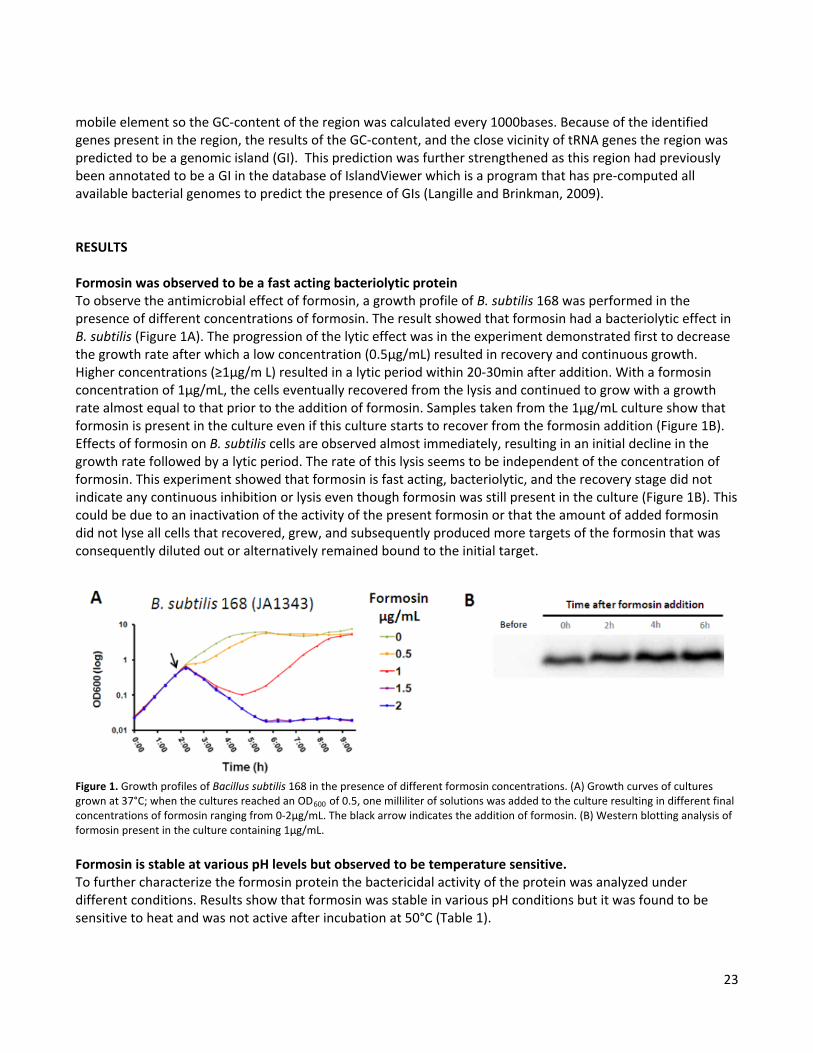
23
mobile element so the GC-content of the region was calculated every 1000bases. Because of the identified genes present in the region, the results of the GC-content, and the close vicinity of tRNA genes the region was predicted to be a genomic island (GI). This prediction was further strengthened as this region had previously been annotated to be a GI in the database of IslandViewer which is a program that has pre-computed all available bacterial genomes to predict the presence of GIs (Langille and Brinkman, 2009). RESULTS Formosin was observed to be a fast acting bacteriolytic protein To observe the antimicrobial effect of formosin, a growth profile of B. subtilis 168 was performed in the presence of different concentrations of formosin. The result showed that formosin had a bacteriolytic effect in B. subtilis (Figure 1A). The progression of the lytic effect was in the experiment demonstrated first to decrease the growth rate after which a low concentration (0.5µg/mL) resulted in recovery and continuous growth. Higher concentrations (≥1µg/m L) resulted in a lytic period within 20-30min after addition. With a formosin concentration of 1µg/mL, the cells eventually recovered from the lysis and continued to grow with a growth rate almost equal to that prior to the addition of formosin. Samples taken from the 1µg/mL culture show that formosin is present in the culture even if this culture starts to recover from the formosin addition (Figure 1B). Effects of formosin on B. subtilis cells are observed almost immediately, resulting in an initial decline in the growth rate followed by a lytic period. The rate of this lysis seems to be independent of the concentration of formosin. This experiment showed that formosin is fast acting, bacteriolytic, and the recovery stage did not indicate any continuous inhibition or lysis even though formosin was still present in the culture (Figure 1B). This could be due to an inactivation of the activity of the present formosin or that the amount of added formosin did not lyse all cells that recovered, grew, and subsequently produced more targets of the formosin that was consequently diluted out or alternatively remained bound to the initial target.
Figure 1. Growth profiles of Bacillus subtilis 168 in the presence of different formosin concentrations. (A) Growth curves of cultures grown at 37°C; when the cultures reached an OD600 of 0.5, one milliliter of solutions was added to the culture resulting in different final concentrations of formosin ranging from 0-2µg/mL. The black arrow indicates the addition of formosin. (B) Western blotting analysis of formosin present in the culture containing 1µg/mL. Formosin is stable at various pH levels but observed to be temperature sensitive. To further characterize the formosin protein the bactericidal activity of the protein was analyzed under different conditions. Results show that formosin was stable in various pH conditions but it was found to be sensitive to heat and was not active after incubation at 50°C (Table 1).

24
Table1. Bactericidal activity of formosin under various conditions.
Pretreatment of formosin
Bactericidal activity
pH (17h) 2 + 3 + 4 + 5 + 6 + 7 + 8 + 9 + 10 + 11 + Temperature (30min) 0°C + 20°C + 40°C + 50°C - 80°C - 100°C - The forD associated gene cluster The appearance of the forD associated gene cluster shows two large gaps in the gene cluster (Figure 2A). These gaps are usually indicative of genes being transcribed separately (Price et al., 2005). However, the presence of the gaps only suggests that this is not an operon. To verify this, the presence of an intrinsic terminator (also called rho independent terminator) and a promoter should be found in each gap. After evaluation of stem-loop structures found in the forD related gene cluster, three stem-loops were found (Figure 2D). Stem-loop 1 was predicted by FindTerm (Softberry) and located between the forD and forE genes (Figure 2D). Stem-loop 2 was predicted by WebGeSTer (Mitra et al., 2011) and located forty bases downstream of the forE gene (Figure 2D). Stem-loop 3 was predicted by WebGeSTer and found about fifty bases downstream of the stop codon of the forG gene (Figure 2D). For these three stem-loops the free energy of the stem-loop structure is observed to be high, but presences of a poly(U) tail is observed only in stem-loop 3 (Figure 2D) and in that case only three uracil is seen. These three uracil tail is shorter than the average length seen in B. subtilis or Escherichia coli (de Hoon et al., 2005). Stem-loop 1 was observed to be an intrinsic terminator on the complementary strand but not for the forD associated gene cluster. In the manual discrimination of potential promoters only one promoter was predicted which was observed to be located upstream of the forD gene (Figure 2C). When the databases DBTBS and Virtual footprint were used to find possible promoters it was discovered that the two databases were not generating equal results for the same predicted promoters. This was probably due to a low stringency of some the consensus sequences and the methods of which these predictions were made. Because of this only the sites that overlapped for the same sigma factor (σ) was used. This resulted in two predicted σH
sites (Figure 2C), one upstream of forD and one upstream of forE (Figure 2A). Translation initiation through the Shine Dalgarno (SD) site was also evaluated for these four genes (Figure 2B). They all contained a well defined SD sequence, but it was observed that upstream of the forD gene a large amount of purine residues was observed. The upstream region from the start codon of forD was observed to contain 18 purine residues in the

25
first 20 residues present. In addition to this, purine rich SD sequence of forD, it was also observed that two small open reading frames (ORFs) are located upstream of the forD gene. These are present in the regions 120-18nt and 114-18nt upstream of the start codon for the forD gene and could be affecting the translation initiation of forD if they are present in the mRNA.
Figure 2. Characteristics of the forD gene cluster from Bacillus licheniformis ATCC14580. (A) The genes in the forD gene cluster denoted to be forD (UniProt locus name: BL00275), forE (BL00274), forF (BL00273), and forG (BL00272) with a stem-loop that could cause transcription delay or termination marked as stem-loop 1, 2 and 3. Promoters (PA-PC) are also seen in the figure, where the black represents the manually predicted promoter and the grey is two promoters predicted by the matching result from Virtual footprint and DBTMS. (B) Translation initiation for the four genes where the Shine Dalgarno sequence (SD) and the start codon of each gene have been marked. (C) Potential promoter sequences seen upstream of the forD and forE genes where the putative -10 and -35 elements has been underlined. (D) Structure and Gibbs free energy (ΔG) of three stem-loops found in the region of the forD associated genes that could affect the transcription either by stalling or through rho independent termination. Hydrogen bonds are annotated as weak (blue) or strong (green) and the downstream region of the stem-loop are marked as red if a G or C is present as these reduce the chance of intrinsic termination.

26
Formosin is related to the lactococcin 972 family Formosin was not found to be similar to any proteins with known functions in a BLAST search (The UniProt Consortium, 2012) but was annotated by Pfam (Punta et al., 2012) to contain a domain similar to that of the lactococcin 972 family. Comparison of five example proteins annotated to belong to the lactococcin 972 family (Table 2) show that they all have a secondary structure consisting of an α-helical signal peptide and a mature protein of only β-strand structures. The mature proteins of all except Tfu_0814 are cationic and formosin is larger (9.6kDa) than the other proteins in this comparison (7.3-7.7kDa). The N-terminal signal peptides for all five proteins appear to be Sec-dependent due to the presence of an H-domain and lack of a twin-arginine motif (Tjalsma et al., 2004). The aliphatic index is most often used to predict stability of proteins (Ikai, 1980), but due to the predicted all beta secondary structure of these five proteins one can predict that the presence of aromatic, and not aliphatic, residues are more vital for protein stability. A high aliphatic and hydropathy index could be indicative of the probability of membrane association as seen in pore forming bacteriocin. Lactococcin 972 is not a pore forming bacteriocin (Martínez et al., 2000). The hydropathy index of formosin and lactococcin 972 is similar, while the aliphatic index is higher for formosin than for lactococcin 972. This could indicate that formosin is more prone to membrane association. However, since one cannot predict which of these hydrophobic or aliphatic residues are surface residues this prediction might be quite hypothetical. Table 2. Properties of mature proteins predicted to be in the lactococcin 972 family. Gene/ Locus
Origin Mass (kDa) pIA Net
charge Aliphatic
IndexB Hydropathy
IndexC 2nd
structureD Identity
ForD LclA forD Bacillus licheniformis
(ATCC 14580) 9.6 9.7 + 6 53.88 -0.915 All beta 100% 25%
lclA Lactococcus lactis subsp. lactis (Plasmid pBL1) 7.4 9.7 + 4 36.97 -0.982 All beta 25% 100%
Tfu_0814 Thermobifida fusca (YX) 7.3 6.98 ± 0 67.50 -0.157 All beta 20% 30% MWP015 Staphylococcus aureus
(Plasmid pMW2) 7.4 10.2 + 8 50.87 -0.525 All beta 15% 26%
SPAR17_1984
Streptococcus pneumoniae (GA08780) 7.7 9.48 + 4 38.45 -0.713 All beta 34% 40%
A pI, Isoelectric point B Aliphatic index calculated as Ikai, 1980 C Hydropathy index is the average hydropathicity of the amino acids (Kyte and Doolittle, 1982) calculated as the sum of the hydropathicity of all residues, divided by the number of residues. D Secondary structure predicted by PSIPRED v3.0 The lactococcin 972 family is predicted to be associated with two or three other proteins Lactococcin 972 has previously been predicted to be a Sec-translocated protein and been observed to be expressed in an operon containing lclA (lactococcin 972) and lclB (putative immunity gene)( Martínez et al., 1999). This has been the model for the lactococcin 972 family, but in this study a comparison of the five model proteins in table 2 showed that this model might be incomplete. Instead it is suggested that three to four genes are associated with each of the model proteins from the lactococcin 972 family (Figure 3). The genes associated with the lactococcin 972 family was in this study presented to be the genes coding for the bacteriocin, a transmembrane (TM) protein, an ATP binding protein and occasionally also a putative accessory protein. The five TM protein associated with the five model proteins are all predicted by Phobius (Käll et al., 2007) to have seven TM regions with two large extracellular domains, one in the N-terminal and a second between the TM4

27
and TM5 regions. Except for the forD gene cluster, the genes associated with the lactococcin 972-like protein were observed to be in close vicinity to each other indicating that these gene clusters might be transcribed on one single operon (Price et al., 2005).
Figure 3. Neighboring genes of each of the model proteins from the lactococcin 972 family coded by the genes: forD, lclA, TFU_0814, MWP015, and SPAR17_1984. Genes are colored according to their properties indicating the bacteriocin as red, a transmembrane protein as blue, an ATP binding protein as green, and a putative accessory protein as grey.
Figure 4. Chromosomal region (4140k-4175k bases) of Bacillus licheniformis ATCC 14580 showing the adjacent region of the forD gene. Genes coded on the minus strand are shown in blue and genes coded on the plus strand are shown in red. Grey genes are genes with framshifts (pseudogene) that are no longer coding a functional protein. Functions of gene or gene clusters are shown with a green background with the described protein function.
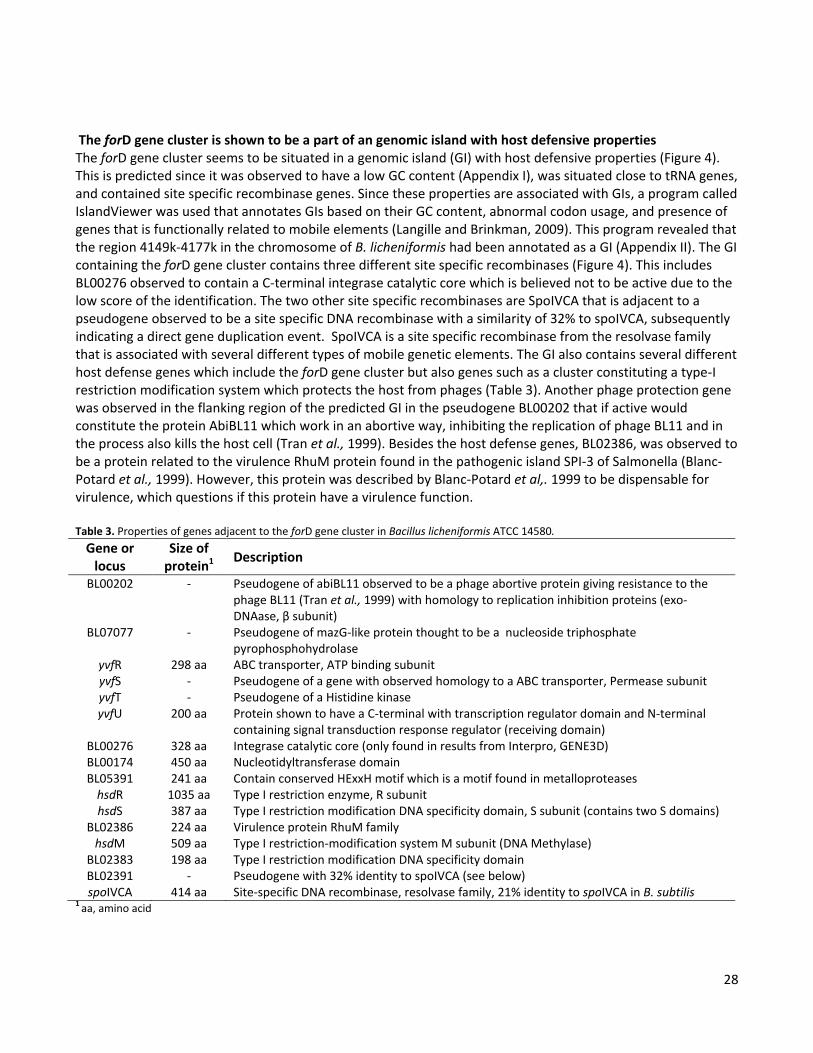
28
The forD gene cluster is shown to be a part of an genomic island with host defensive properties The forD gene cluster seems to be situated in a genomic island (GI) with host defensive properties (Figure 4). This is predicted since it was observed to have a low GC content (Appendix I), was situated close to tRNA genes, and contained site specific recombinase genes. Since these properties are associated with GIs, a program called IslandViewer was used that annotates GIs based on their GC content, abnormal codon usage, and presence of genes that is functionally related to mobile elements (Langille and Brinkman, 2009). This program revealed that the region 4149k-4177k in the chromosome of B. licheniformis had been annotated as a GI (Appendix II). The GI containing the forD gene cluster contains three different site specific recombinases (Figure 4). This includes BL00276 observed to contain a C-terminal integrase catalytic core which is believed not to be active due to the low score of the identification. The two other site specific recombinases are SpoIVCA that is adjacent to a pseudogene observed to be a site specific DNA recombinase with a similarity of 32% to spoIVCA, subsequently indicating a direct gene duplication event. SpoIVCA is a site specific recombinase from the resolvase family that is associated with several different types of mobile genetic elements. The GI also contains several different host defense genes which include the forD gene cluster but also genes such as a cluster constituting a type-I restriction modification system which protects the host from phages (Table 3). Another phage protection gene was observed in the flanking region of the predicted GI in the pseudogene BL00202 that if active would constitute the protein AbiBL11 which work in an abortive way, inhibiting the replication of phage BL11 and in the process also kills the host cell (Tran et al., 1999). Besides the host defense genes, BL02386, was observed to be a protein related to the virulence RhuM protein found in the pathogenic island SPI-3 of Salmonella (Blanc-Potard et al., 1999). However, this protein was described by Blanc-Potard et al,. 1999 to be dispensable for virulence, which questions if this protein have a virulence function. Table 3. Properties of genes adjacent to the forD gene cluster in Bacillus licheniformis ATCC 14580.
Gene or locus
Size of protein1 Description
BL00202 - Pseudogene of abiBL11 observed to be a phage abortive protein giving resistance to the phage BL11 (Tran et al., 1999) with homology to replication inhibition proteins (exo-DNAase, β subunit)
BL07077 - Pseudogene of mazG-like protein thought to be a nucleoside triphosphate pyrophosphohydrolase
yvfR 298 aa ABC transporter, ATP binding subunit yvfS - Pseudogene of a gene with observed homology to a ABC transporter, Permease subunit yvfT - Pseudogene of a Histidine kinase yvfU 200 aa Protein shown to have a C-terminal with transcription regulator domain and N-terminal
containing signal transduction response regulator (receiving domain) BL00276 328 aa Integrase catalytic core (only found in results from Interpro, GENE3D) BL00174 450 aa Nucleotidyltransferase domain BL05391 241 aa Contain conserved HExxH motif which is a motif found in metalloproteases
hsdR 1035 aa Type I restriction enzyme, R subunit hsdS 387 aa Type I restriction modification DNA specificity domain, S subunit (contains two S domains)
BL02386 224 aa Virulence protein RhuM family hsdM 509 aa Type I restriction-modification system M subunit (DNA Methylase)
BL02383 198 aa Type I restriction modification DNA specificity domain BL02391 - Pseudogene with 32% identity to spoIVCA (see below) spoIVCA 414 aa Site-specific DNA recombinase, resolvase family, 21% identity to spoIVCA in B. subtilis
1 aa, amino acid

29
DISCUSSION The reported data demonstrates that formosin has a quick acting bacteriolytic effect against B. subtilis. The protein appears to be stable in various pH conditions (pH2-11) but was observed to be inactivated by heat. Mode of action was never determined during the course of this study. However, from the appearance of the growth curve response to formosin addition a few suggestions could be made. When formosin was added to the culture the cells started to lyse. There are two types of quick acting bacteriolytic effects of bacteriocins, one being bacteriolytic enzymes which break down the cell wall (e.g. Lysostaphin) and the other being bacteriocin that has a secondary bacteriolytic effect (e.g. Nisin) (Cotter et al., 2005; Martínez-Cuesta et al., 2000). In a secondary bacteriolytic effect the bacteriocin usually has a primary pore-forming effect on the cells but in addition interacts with the cell in a way which causes autolytic enzymes to be activated or there is an imbalance between building and degrading the cellular wall during growth which subsequently leads to lysis (Jack et al., 1995; Martínez-Cuesta et al., 2000). If formosin was a bacteriolytic enzyme one might expect that the rate of lysis would be concentration dependent. In this study it was shown that the rate of lysis remained the same for the different concentrations indicating that formosin has a secondary bacteriolytic effect. The primary effect of formosin could not be revealed in this study but a comparison can be made to lactococcin 972 which belongs to the same bacteriocin family. Lactococcin 972 has been observed to be a non-pore forming bacteriocin that inhibits septum formation but does not cause lysis (Martínez et al., 1996). Inhibition of the septum formation is caused by an interaction of lactococcin 972 to the cell wall precursor lipid II (Martínez et al., 2008). The primary effect of formosin could be similar to lactococcin 972, but a pore formation could still be a possible primary effect since formosin has a higher aliphatic index than lactococcin 972. To determine the primary effect of formosin it would therefore be advisable to test both inhibition of septum formation (Martínez et al., 1996) and pore formation (McAuliffe et al., 1998). The gene cluster associated with formosin production was also investigated in this study. This gene cluster was observed to contain four genes coding for formosin, an ABC transporter consisting of two proteins, and a protein predicted to be an accessory protein. Three putative promoter sequences were found within the gene cluster through in silico investigations. Potential transcriptional terminators were also investigated and three stem-loop structures were found. Requirements for a rho independent terminator are a stem-loop structure with a downstream poly(U) tail (de Hoon et al., 2005). For the predicted stem-loop structures in or after the forD gene cluster only the last stem-loop is observed with a poly(U) tail and can be identified as a potential transcription terminator. The two stem-loop structures found between the genes in the gene cluster do not contain downstream poly(U) tails and thus do not qualify as rho independent terminator in the scope of this study. Instead these two stem-loop structures located between the genes could have another function such as transcription pausing (Farnham and Platt, 1981). Alternatively, these stem-loops could also be involved in either mRNA stability as some exonucleases are known to be inhibited by stem-loops or mRNA degradation as some endonucleases are observed to specifically degrade double stranded RNA structures (Belasco and Chen, 1988; Codon, 2003). Since no transcription terminator was predicted within the gene cluster it is in this study suggested that the gene cluster is named the for-operon. However, it would be advisable to confirm this experimentally by northern blotting analysis using probes toward the four genes in this putative for-operon. Such an analysis will show when in the growth cycle formosin is expressed as well as the size and gene content of the mRNA transcript(s). In the comparison of genes associated with the lactococcin 972-like proteins it was shown that the previous model for the lactococcin 972 family might be incomplete, since it was observed that the investigated

30
lactococcin 972-like protein all seem to be associated with an ABC transporter. Lactococcin 972 has previously been suggested to be associated with an adjacent gene predicted to code for a putative immunity protein with seven TM spanning regions (Martínez et al., 1999).This was concluded from results of a northern blotting analysis where a 2.5kb transcript was found to contain the lclA and lclB genes (Martínez et al., 1999). However, when genes associated with lactococcin 972-like protein were examined in this study it was observed that this conclusion was not correct. DNA sequence of the lactococcin 972 encoding plasmid revealed that an open reading frame (ORF) was located two nucleotides downstream of the stop codon of lclB. This ORF was found to code for an ATP binding protein subunit of an ABC transporter. But the predicted size of 2.5kb of the lclAB transcript does not fit the presence of this additional ORF. However, the estimation of a 2.5kb transcript was seen to be inaccurate since the northern blot results showed this transcript to be larger than the 23S rRNA in Lactococcus lactis (Martínez et al., 1999, Figure 4). According to the sequenced genome the 23S rRNA in L. lactis is approximately 2.9kb long, which would make the transcript in the initial Lactococcin 972 study larger than 2.9kb. This would fit the presence of the ATP binding subunit. Although this transcript was never confirmed to contain this third gene, the true length of the transcript and the close proximity of the lclB and the ORF indicate that the lclA-associated-operon contains three genes. From the present study it is therefore suggested that the model of the lactococcin 972 family is revised to say that this protein family is associated with an ABC transporter with seven transmembrane regions. The function of the ABC transporter associated with formosin will during the scope of this thesis be revealed which in turn could be a possible function for all ABC transporters associated with the lactococcin 972 family. The for-operon is located in a chromosomal part of the B. licheniformis chromosome that has been noted to be a genomic island (GI). It contains several genes which can be described as having host defensive properties, such as a restriction-modification system and a phage abortive protein. In addition, the presence of formosin will result in an advantage over other bacteria which can be killed by this bacteriocin. Three different recombinases were also found in the investigated region, one of which is SpoIVCA. SpoIVCA has in B. subtilis been observed to be involved in sporulation by excision of a DNA element termed skin that subsequently results in the creation of the sigma factor K (σK)(Kunkel et al., 1990). This skin element is observed to be derived from the PBSX prophage that has evolved to be an integral part of the sporulation process (Krogh et al., 1996). SpoIVCA from B. licheniformis and B. subtilis are both site-specific recombinases, but SpoIVCA in B. licheniformis does not have a function in sporulation. Instead it is suggested, based on the findings in this study, that the function is related to the mobility of the genomic island in which it is located.

31
CHAPTER 5 Secretion analysis of formosin, a bacteriocin from Bacillus licheniformis
INTRODUCTION Bacillus secretes numerous proteins into their extracellular environment and possesses a great capacity for secreting foreign enzymes (Simonen and Palva, 1993). This is why Bacillus serves as a good bacterial system for production in addition to studying both expression and secretion of different types of proteins. Because of the large variations of secreted proteins, different pathways have evolved to serve different needs. The main secretion pathway is the secretion (Sec) pathway that translocates a wide array of unfolded proteins. Many times resulting in a great yield of protein in the extracellular milieu. The second secretion pathway is the twin-arginine translocation (Tat) pathway that translocates proteins in their folded state. (Natale et al., 2008; Simonen and Palva, 1993; Pohl and Harwood, 2010; Tjalsma et al., 2004) In addition to these two general secretion pathways there are also special purpose secretion pathways such as the ABC transporters dedicated to secreting a specific protein into the medium at certain times of need (Pohl and Harwood, 2010; Simonen and Palva, 1993). The secretion pathway can in most cases be distinguished by the properties of the N-terminal signal peptide. The signal peptide for the Tat- and Sec-pathways consists of three different domains. The first domain is the N-domain which signals that the protein is destined for translocation because of the presence of positively charged amino acids. This is followed by a domain of hydrophobic amino acids called the H-domain and finally the C-domain that contains the signal required for the signal peptide cleavage. (Natale et al., 2008; Tjalsma et al., 2004) The signal peptide for ABC transporters, also called leader peptide, can in e.g. Bacillus subtilis generally be distinguished by the lack of a hydrophobic H-domain in the N-terminal signal peptide (Tjalsma et al., 2004). In this study the secretion of a novel bacteriocin, formosin, from Bacillus licheniformis was characterized through heterologous expression in B. subtilis. Formosin has been identified as a chromosomally encoded protein that has been assigned the ordered locus name BL00275 in the UniProt database (Veith et al., 2004; The UniProt Consortium, 2012). This is a 14 kDa protein with a cleavable N-terminal signal peptide resulting in a secreted protein with the final size of 9.6kDa. Different proteins from B. licheniformis have previously been expressed in B. subtilis with great success (Simonen and Palva, 1993), formosin however, could not initially be produced in B. subtilis. It was soon discovered that formosin was a part of a complex four-gene operon (Figure 1A). This for-operon contained the expression of formosin (ForD) followed by an ATP binding protein (ForE), an unknown protein referred to as an accessory protein (ForF), and ended with a transmembrane (TM) protein (ForG). Expression of the intact for-operon in B. subtilis resulted in high levels of secreted formosin. The additional genes in the for-operon seemed

32
to constitute a putative ABC transporter since the ATP binding protein was observed to contain motifs belonging to the ABC transporter family (Chapter 7). The signal peptide of formosin has properties more resembling that of a sec-dependent signal peptide, rather than that of a leader peptide for ABC transporters (Figure 1B). However, it is known that ABC transporters can be quite promiscuous in their substrate recognition (Kuchler and Thorner, 1992). The aim of the current study was to determine the secretion pathway and the process in which this secretion occurs with regards to signal peptide recognition.
Figure 1. (A) The for-operon required for production of formosin. The operon contains the genes BL00275-BL00273 as assigned by UniProt but has been given the gene names forD, forE, forF, and forG. (B) N-terminal signal peptide of formosin and its signal peptidase (SP) cleavage site predicted by results from N-terminal sequencing of secreted formosin. The three domains of the signal peptide are indicated as well as the positively charged amino-acids. To investigate the secretion, the for-operon was expressed in B. subtilis JA1343, which is a RUB200 derivative where genes of two extracellular proteases have been deleted (nprE and aprE) to increase yield. Secretion analysis was performed in several ways. (I) The genes required for secretion were determined by performing different deletions of the genes in the for-operon. (II) Secretion profiles of formosin, the N-terminal signal peptide, and different parts of the secreted 9.6kDa formosin protein were determined by fusions to a reporter protein. The reporter protein used in this study was a small protease inhibitor, Subtilisin-chymotrypsin inhibitor-2A (Ci2A), from Hordeum vulgare (Barley) (Halkier et al., 1993). (III) Secretion of formosin using a known Sec-dependent signal peptide was made by exchanging the endogenous N-terminal signal peptide of formosin to the signal peptide of α-amylase (AmyQ) from Bacillus amyloliquefaciens (Palva et al., 1981). Results from this study show that the ATP binding protein (ForE) and the TM protein (ForG) were required for secretion and were thought to constitute a putative ABC transporter for formosin. The N-terminal signal peptide was not the signal needed for translocation through the ABC transporter. Instead there seemed to be two secretion pathways for formosin. One promoted by the N-terminal signal peptide which subsequently lead to the degradation of formosin and a second internal signal peptide at the C-terminus of formosin that promoted secretion through the ABC transporter. It was also observed that only when secreted through the ABC transporter could formosin be detected in the medium. It was therefore believed that the secretion through the ABC transporter is a crucial step in the maturation. This is the first observed example of a protein with two different signal peptides and the first example of a C-terminal signal peptide found in Bacillus.

33
MATERIALS AND METHODS
Bacterial strains and growth conditions. Bacterial strains used for this study is listed in Table 1. B. subtilis cells were made competent by the method of Anagnostopoulos and Spizizen (Anagnostopoulos and Spizizen, 1961). Cells were grown in liquid TY media or Cal 18-2 media (Rasmussen et al., 2000) and plated on LBPG agar plates (LB agar with 0.5% (w/v) glucose and 50mM potassium phosphate, pH 7.0). When required chloramphenicol (6 µg/mL), Kanamycin (30 µg/mL) or D-alanine (200µg/mL) was supplemented. Incubation was made at 37°C for plates or TY medium and 30°C for Cal 18-2 medium with 250rpm for liquid cultures. All the produced strains were sequence verified using the Sanger-method (Sanger et al., 1977). DNA techniques. Chromosomal DNA from B. subtilis and B. licheniformis was extracted using the protocol for Gram positive bacteria by QIAamp® DNA Mini and Blood Mini Kit (Qiagen). Plasmids were extracted by QIAprep® Spin Miniprep Kit (Qiagen). PCR amplifications was preformed with Phusion® Hot Start High-Fidelity DNA polymerase (Finnzymes) using the recommended protocol. PCR products were purified using the QIAquick® Gel Extraction Kit (Qiagen). All gene constructs were put under the control of a triple promoter system (3Pcry) previously described to result in high yields (Widner et al., 2000). Construction of producing strain. The expression cassette was designed for insertion into the pel-region of B. subtilis by homologous recombination. The cassette was created through splicing by overlap extension PCR (SOE-PCR) as previously described (Horton et al., 1990). Briefly, the different segments in the expression cassette were amplified using primers with 5’-tails that are reverse complementary to the neighboring segment (Table 2). The for-operon of B. licheniformis was amplified by PCR using primers for-F and for-R. These genes were put under the control of a triple promoter system (3Pcry) previously described to result in high yields (Widner et al., 2000). The left side of the pel region used for homologous recombination together with this triple promoter was amplified from B. subtilis MB1510 using primers for-F and 3P-cry-R. The right side of the pel region together with the chloramphenicol resistance was amplified from the Escherichia coli plasmid pMB1508 using primers cam-F and pel-R. These three segments were then added in equal molar amounts to the SOE PCR where it was ligated through the overlapping regions of the primers and amplified by the pel-F and pel-R primers. The resulting segment was then transformed into B. subtilis JA1343 to create strain SLds1. Construction of deletions. Deletion of the different genes in the gene cluster was made through SOE PCR in the same manner as described above. Chromosomal DNA from the production strain SLds1 was used as a template for the amplifications. Primers were designed to flank the region of the deletion and contained a 5’-tail that were reverse complementary to one another so that they could be used to fuse the segments together in the SOE-PCR (Table 2). For the amplification of the two segments and as amplifying primers for the SOE the pel-F and pel-R was used. The segment was transformed into B. subtilis JA1343, resulting in strains SLds2-8. Construction of the fusion protein. To follow the secretion pattern of the different strains a reporter protein was fused to formosin. Subtilisin-chymotrypsin inhibitor-2A (Ci2A) from H. vulgare (Barley) was used as a reporter protein (amino acids: 14-84) and fused to the C-terminal of formosin. Two strains were created by SOE PCR using the primers pel-F and Fus-for-R or Fus-for-F and pel-R to create the left and right segments flanking the Ci2a insertion. As templates the strains SLds1 and SLds6 were used to create strains with and without the ABC transporter. The Ci2a gene was amplified from plasmid pCi2a using primers Ci2a-forD-F and Ci2a-forD-R. The three segments were then added to the SOE PCR in equal molar amounts and ligated to one segment that was amplified in the same reaction by the pel-F and pel-R primers. The resulting segment was then transformed into B. subtilis JA1343 to create strain SLds21 and 22. In addition to these two strains a control

34
strain was created that produced intracellular Ci2a. Using the strain SLds22, formosin was deleted in the same way as in the deletion strains using primers pel-F and Fus-Dfor-R or Fus-Dfor-F and pel-R for the deletion. This resulted in strain SLds53 that contained only the Ci2a protein without any secretion signal. Construct of the reporter protein fused to the signal peptide from formosin. These strains were created in the same manner as described for the fusion protein. However, in these strains only the signal peptide of formosin was fused to the Ci2a protein using different primers at that insertion site. Two primers were different reflecting the insertion site of the C-terminal end of formosin being in this case just the signal peptide. The primer Fus-for-R was exchanged to Fus-SP-R and Ci2a-ForD-F was exchanged for the Ci2a-SP-F primer. Two strains were created with (SLds80) or without the ABC transporter (SLds81) using the template of either SLds21 or SLds22. Creation of D-alanine deficient strain to be used for gene insertions. To utilize another insertion site in B. subtilis (JA1343) without the use of a selection marker a D-alanine deficient strain had previously been created by Novozymes by deleting the gene for alanine racemase, alr1. The strain SLds29 contained this deletion by insertion of a kanamycin selectable disruption cassette containing the triple promoter together with the expression of the comS gene to increase transformation rates (Liu et al., 1996). To create a strain that expressed the ABC transporter but not formosin, the forD gene was deleted from strain SLds1 by amplifying the flanking regions of the forD gene using SOE-primers. For the left segment primers d-forD-R and pel-F were used and for the right segment primers pel-R and d-forD-F were used. The two segments were then fused by an SOE PCR to one segment that could then be amplified in the same reaction by the pel-F and pel-R primers. The resulting segment was then transformed into B. subtilis JA1343 to create strain SLds11. The strain SLds11 was then made D-alanine deficient via horizontal gene transfer by adding chromosomal DNA from strain SLds29 to competent cells of SLds11. Selection of kanamycin and chloramphenicol resistance results in a D-alanine deficient strain that contains the ABC transporter (SLds60). Exchanging the signal peptide of formosin. To see the effect of an exchange of the signal peptide for formosin the endogenous signal peptide was exchanged for the signal peptide of α-amylase (AmyQ) from B. amyloliquefaciens (Palva et al., 1981). Through SOE PCR a construct was created to be inserted at the alr-region of the strains SLds29 and SLds60 to see the effect of secretion for the new signal peptide in the absence and presence of the ABC transporter. The construct was created by fusing together four segments in a SOE PCR. For the first segment primers dal-F and L-dal-R were used to amplify the left region of the alr and the promoters from strain SLds29. In the second segment primers SPamyQ-F and SPamyQ-R were used to amplify the signal peptide from amyQ using DSM 9553 as a template. For the third segment the sequence for mature formosin was amplified from strain SLds1 using primers forD-F and forD-R. In the fourth segment the alr gene was restored using JA1343 as a template with the primers R-dal-F and dal-R. These four segments were then used in an SOE PCR to ligate the segments together to the final construct that then could be amplified in the same reaction by the primers dal-F and dal-R. The resulting construct was inserted into competent cells from SLds29 and SLds60 to create one strain without the ABC transporter (SLds100) and one with the ABC transporter (SLds101).

35
Table 1. Strains and plasmids used in this study. Strain or plasmid Description and/or genotypeA Strain source Bacillus licheniformis ATCC14580 Natural producer of formosin (ForD) BGSC Bacillus subtilis 168 JA1343 RUB 200 derivative; amyE, aprE, nprE, spoIIAC Novozymes A/S, (Internal) MB1510 JA1343; Δpel::3Pcry Bjornvad et al., 2003 SLds29 JA1343; Δalr::3Pcry comS; KmR Novozymes A/S (Internal) SLds11 JA1343; Δpel::3Pcry forEFG; CmR This study SLds60 JA1343; Δalr::3Pcry comS; KmR + Δpel::3Pcry forEFG; CmR This study Bacillus amyloliquefaciens DSM 9553 Source of AmyQ signal peptide Novozymes A/S (Internal) Plasmid pMB1508 pUC19 derivative; contains CmR and used for insertion into pel-region Bjornvad et al., 2003 pCi2a Expression vector for Ci2a (used as PCR template) Novozymes A/S (Internal) Strains containing deletions SLds1 JA1343; Δpel::3Pcry forDEFG; CmR This study SLds2 JA1343; Δpel::3Pcry forDEF; CmR This study SLds3 JA1343; Δpel::3Pcry forDEG; CmR This study SLds4 JA1343; Δpel::3Pcry forDFG; CmR This study SLds5 JA1343; Δpel::3Pcry forDG; CmR This study SLds6 JA1343; Δpel::3Pcry forD; CmR This study SLds7 JA1343; Δpel::3Pcry forDE; CmR This study SLds8 JA1343; Δpel::3Pcry forDF; CmR This study Strains for different secretion analysis SLds21 JA1343; Δpel::3Pcry Φ(forD-ci2a) forEFG; CmR This study SLds22 JA1343; Δpel::3Pcry Φ(forD-ci2a); CmR This study SLds53 JA1343; Δpel::3Pcry ci2a; CmR This study SLds80 JA1343; Δpel::3Pcry Φ(SPforD-ci2a) forEFG; CmR This study SLds81 JA1343; Δpel::3Pcry Φ(SPforD-ci2a); CmR This study SLds100 JA1343; alr::3Pcry Φ(SPamyQ-forD) This study SLds101 JA1343; alr::3Pcry Φ(SPamyQ-forD) + Δpel::3Pcry forEFG; CmR This study Strains used for analysis of internal signal peptide SLds110 JA1343; alr::3Pcry Φ(forD42-73-ci2a) This study SLds111 JA1343; alr::3Pcry Φ(forD74-101-ci2a) This study SLds112 JA1343; alr::3Pcry Φ(forD102-126-ci2a) This study SLds113 JA1343; alr::3Pcry Φ(forD42-73-ci2a) + Δpel::3Pcry forEFG; CmR This study SLds114 JA1343; alr::3Pcry Φ(forD74-101-ci2a) + Δpel::3Pcry forEFG; CmR This study SLds115 JA1343; alr::3Pcry Φ(forD102-126-ci2a) + Δpel::3Pcry forEFG; CmR This study A 3Pcry is a promoter system with three promoters and the mRNA stabilization segment (Widner et al., 2000)
Creation of fusion between Ci2a and different parts of the mature formosin protein. Three different parts of formosin (ForD) were fused to the N-terminal of Ci2a. These strains were all created in the same manner using different primers. An example of this is the first strain that was created by four segments that were then ligated in an SOE PCR. The first segment was the left side of the alr insertion site and the promoters and it was

36
amplified from SLds29 using primers dal-F and preD1-R. The second segment was the first part of the mature ForD protein which was amplified from SLds1 using primers D1-F and D1-R. The third segment coded for Ci2a and was amplified by primers D1-ci2a-F and D-Ci2a-R from the strain SLds21. The fourth segment was the right site that contained the alr gene and this was amplified from strain JA1343 using primers Ci2a-dal-F and dal-R. After a SOE PCR using primers dal-F and dal-R the segment was transformed into either SLds29 or SLds60 to generate strains with/without the ABC transporter. In the same manner the two other constructs were created with the exception that the primers with the name D1 were changed to D2 or D3, depending on the section of the ForD protein that was fused to Ci2a. This resulted in the strains SLds110-115. Sample preparation. Secretion and production patterns of the different strains were determined by growing the cells for 48h (72h for B. licheniformis) in a special Cal 18-2 medium at 30°C. After this the cell-free medium was extracted by a two step centrifugation. Cells were removed by centrifuging the growth solution at 5000xg for 3min, after which the supernatant was taken and centrifuged at 20.000xg for 10min to remove any additional debris that might interfere with the SDS-PAGE. Samples were prepared for SDS-PAGE by adding two parts of supernatant, one part water and one part NuPAGE® LDS sample buffer (4x). For the cell-samples the cells were washed in a three step repetitive centrifugation in which the cells were centrifuged at 5000xg for 3min and the supernatant taken out and replaced with clean Cal 18-2 medium. After repeating the centrifugation three times, the resulting cell suspension was prepared for SDS-PAGE in the same manner as the medium samples. SDS-PAGE and Western blotting analysis. Samples were incubated at 99°C for 3min and put on ice until loading 10µL of the samples on precast NuPAGE® 4-12% Bis-Tris gel (Invitrogen). To estimate the size of the bands on the membrane 15µL of Novex® Sharp Pre-strained Protein standard was used for western blot size estimation. As a control 1µg of purified formosin (produced internally by Novozymes A/S) was used as a positive control. The gels were run with NuPAGE® MES SDS Running Buffer at 200V for 35min. To prevent background in the western blotting analysis the gels were incubated for 30min in distillated water. Blotting was performed for 7min using the iBlot® Dry Blotting system (Invitrogen) to a Nitrocellulose Transfer Stack designed for this system. Immunoblotting was performed using rabbit primary antibodies against either formosin or Ci2a (manufactured internally by Novozymes) in combination with a Novex® WesternBreeze™ Chemoluminescent Anti-Rabbit kit (Invitrogen) using recommended procedures and provided reaction buffers. Membranes were exposed and photographed using BioSpectrum® Imaging System (UVP) in combination with VisionWorks®LS Analysis Software (UVP) using the general exposure time of 15min with the exception for figure 3 where 25min was used to increase the signal. Images were processed in Adobe Photoshop® CS2 (Adobe Systems) by inverting the colors of the images. N-terminal sequencing. Sequencing of formosin was performed by Edman degradation on gel fragments cut out from the SDS-PAGE gels. Samples were sent to the Protein Technology group at Novozymes for analysis.
37
Table 2. Primers used in this study.
Primer ID Sequence (5’-3’)A Template Specificity Construction of production strain pel-F AGACGAGCTCCGCAATAAAA Varies Forward primer for pel-region 3Pcry-R CCACTCGGTTCCCTCCTCATTTTT MB1510 SOE primer promoter region for-F GGAGGGAACCGAGTGGGCGGTGACAAACTAAC ATCC14580 SOE primer for-operon for-R TCGACTATTACTGGGCTTCATCCGACATCTTA ATCC14580 SOE primer for-operon cam-F GATGAAGCCCCGTTAATCCGCGGATATATAGC pMB1508 SOE primer chloramphenicol pel-R ACCAGCCGTCCTTTCTACCT Varies Reverse primer pel-region Creation of deletions d-forD-R CGTTTTCCCTTTGTATTTCATTCCTCCTCTTTCATTCG SLds 1 SOE primer deletion of forD d-forD-F GGAATGAAATACAAAGGGAAAACGGTTGCT SLds 1 SOE primer deletion of forD d-forE-R TGGTTGATTTCATCTGTACCTCCCATCAAATTCTACG SLds 1 SOE primer deletion of forE d-forE-F GGAGGTACAGATGAAATCAACCACGAAGATCATT SLds 1 SOE primer deletion of forE d-forF-R TGTAGAAGTTGAGTTATTTCATGAGTTACCTCCTAAGTTGC SLds 1 SOE primer deletion of forF d-forF-F GGTAACTCATGAAATAACTCAACTTCTACACAATTCACTCG SLds 1 SOE primer deletion of forF d-forG-R CTTACCTCCGGTCACAATCCTCCAAGGGCTCATA SLds 1 SOE primer deletion of forG d-forG-F AGGATTGTGACCGGAGGTAAGATGTCG SLds 1 SOE primer deletion of forG d-forEF-R TGTAGAAGTTGAGTTACTGTACCTCCCATCAAATTCTACG SLds 1 SOE primer deletion of forEF d-forEF-F ATGGGAGGTACAGTAACTCAACTTCTACACAATTCACTCG SLds 1 SOE primer deletion of forEF d-forEFG-R CTTACCTCCGGTCACTGTACCTCCCATCAAATTCTACG SLds 1 SOE primer deletion of forEFG d-forEFG-F GAGGTACAGTGACCGGAGGTAAGATGTCG SLds 1 SOE primer deletion of forEFG Creation of forD-Ci2a fusion protein Fus-for-R CACCGCCACCACGCGTGTTGGTGCGATAGTAGTAACGACC SLds 1 or 6 SOE primer fusion of forD to Ci2a Ci2a-forD-F ACACGCGTGGTGGCGGTGCTGGTGACCGTCA pCi2a SOE primer amplification of Ci2a Ci2a-forD-R CCTTTAGATCTAGCCGACCCTGGGGAC pCi2a SOE primer amplification of Ci2a Fus-for-F TCCCCAGGGTCGGCTAGATCTAAAGGGAAAACGGTTGC SLds 1 or 6 SOE primer fusion of forD to Ci2a Fus-Dfor-F TCACCAGCACCCATTATTTCATTCCTCCTCTTTCA SLds22 SOE primer for deletion of ForD Fus-Dfor-R AGGAATGAAATAATGGGTGCTGGTGACCGTC SLds22 SOE primer for deletion of ForD Fusion of Ci2a to the signal peptide of ForD Fus-SP-R GGTCACCAGCACCTGCAAATGAAGGCACCGTA SLds21 or 22 SOE primer fusion Ci2a to signal peptide Ci2a-SP-F CCTTCATTTGCAGGTGCTGGTGACCGTCA SLds21 or 22 SOE primer fusion Ci2a to signal peptide Exchanging the signal peptide of ForD dal-F AATCATCGAAATAACCGCCAA SLds29 Forward primer for alr-region L-dal-R CGTTTTTGAATCATTCGGTTCCCTCCTCATTTTTATAG SLds29 SOE primer for left segment of alr-region SPamyQ-F AGGGAACCGAATGATTCAAAAACGAAAGCGGA DSM 9553 SOE primer for signal peptide from amyQ SPamyQ-R GGATTCACCCAGGCTGATGTTTTTGTAATCGG DSM 9553 SOE primer for signal peptide from amyQ forD-F TACAAAAACATCAGCCTGGGTGAATCCGGGTTATC SLds1 SOE primer for forD forD-R GCGATCTCTCCTCTCTTGCCATGCTAGCCCCGT SLds1 SOE primer for forD R-dal-F TAGCATGGCAAGAGAGGAGAGATCGCACTGATAATTG JA1343 SOE primer for right segment of alr-region dal-R AGAGAACCTTTTTTCGTGTGGC JA1343 Reverse primer for alr-region Fusion of Ci2a to different parts of the ForD protein preD1-R CGGATTCACCCACATTATTTCATTCCTCCTCTTTCATT SLds29 SOE primer for the 3’-region for part 1 D1-F GGAGGAATGAAATAATGTGGGTGAATCCGGGTTATC SLds1 SOE primer for part 1 of mature forD D1-R CACCAGCACCCGGGTGGTTGTACTCTGAACG SLds1 SOE primer for part 1 of mature forD D1-ci2a-F ACAACCACCCGGGTGCTGGTGACCGTCA SLds21 SOE primer to fuse Ci2a to part 1 of forD preD2-R GTGGACCTTTGTCATTATTTCATTCCTCCTCTTTCATT SLds29 SOE primer for the 3’-region for part 2 D2-F GAGGAATGAAATAATGACAAAGGTCCACGGCTC SLds1 SOE primer for part 2 of mature ForD D2-R CAGCACCCGCAGCCGTATCAATACTTCTATT SLds1 SOE primer for part 2 of mature forD D2-ci2a-F GATACGGCTGCGGGTGCTGGTGACCGTCA SLds21 SOE primer to fuse Ci2a to part 2 of forD preD3-R GGAATGAAATAATGGGCCGCTACTCTAATGCCTA SLds29 SOE primer for the 3’-region for part 3 D3-F CCAGCACCGTTGGTGCGATAGTAGTAACGACC SLds1 SOE primer for part 3 of mature forD D3-R CTATCGCACCAACGGTGCTGGTGACCGTCA SLds1 SOE primer for part 3 of mature forD D3-ci2a-F GGATTCACCCACATTATTTCATTCCTCCTCTTTCATTC SLds21 SOE primer to fuse Ci2a to part3 of forD D-Ci2a-R ATAAAATGTAATCAAAGTTACTAGCCGACCCTGGGGA SLds21 SOE primer for Ci2a
Ci2a-dal-F CCAGGGTCGGCTAGTAACTTTGATTACATTTTATAATTAATTTTAACAAAGTG
JA1343 SOE primer between Ci2a and alr
A Underlined sequences are the 5’tails used in the SOE-PCR.

38
RESULTS
N-terminal sequencing showed that the 14kDa formosin was cleaved to a 9.6kDa secreted protein. The N-terminal sequencing revealed the cleavage site for the signal peptide (Figure 1B). Formosin was found to be a 126-amino-acid long protein resulting in an 85 amino acid (aa) long protein with a size of 9.6kDa after secretion. The signal peptide was determined to be 41 aa long, containing an N-terminal domain with eight positive charges followed by a 15 aa long hydrophobic domain (H-domain). Properties of this signal peptide seemed to correspond to a typical sec-dependent signal peptide (Figure1B). However, previous results had indicated that the production was dependent on the expression of the entire for-operon. The additional genes in the for-operon are found to constitute an ABC transporter (Chapter 7) that was suspected of being involved in the secretion process.
Figure 2. Western blotting analysis for formosin present in the medium after two days (three days for B. licheniformis). In the far right lane 1µg of purified formosin was added for comparison. Strains containing different gene patterns inserted into the bacterial chromosome at the pel-locus. Letters D, E, F, and G represent the presence of the genes forD, forE, forF, and forG. Secretion of formosin is dependent on two other structural genes in the for-operon. To determine which genes of the operon were vital for the secretion, strains were created with different deletion patterns of the three other structural genes in the operon (Table 1). Results showed that the only deletion mutant still capable of formosin secretion was the deletion of the accessory protein (ForF) in strain SLds3 (Figure 2). Consequently, this shows that both the ATP binding protein (ForE) and the TM protein (ForG) were required for formosin secretion. Expression of formosin in the natural producer, B. licheniformis, required three days of growth to detect the protein in large amounts in the medium. When the accessory protein (ForF) was deleted, a reduction of the secretion could be observed. For this reason the accessory protein was kept as a part of the “ABC transporter construct” in later experiments.

39
Figure 3. Western blotting analysis for detection of formosin (ForD) within the cells after two days growth. Strains used in this analysis were the strains with different deletion patterns that were not observed to secrete formosin into the medium (Figure 2). According to the size of the two bands observed in two of the lanes the proteins found is the 14kDa uncleaved (strong band) and the ∼9kDa cleaved or degraded (weak band) formosin. Letters D, E, F, and G represent the presence of the genes forD, forE, forF, and forG. Uncleaved formosin and the TM protein interact within the cells. Western blotting analysis was made to determine the intracellular levels of formosin in the strains with different deletion patterns of the for-operon. The cell fractions of the strains that were not observed to secrete formosin were used for this analysis (Figure 2). Results showed that although the gene expression of forD should generate formosin production in all these strains, formosin could only be found in the cell fractions of the strains that contained the TM protein (ForG) (Figure 3). The highest amounts were observed for a band corresponding to uncleaved formosin (14kDa), but small amounts of the cleaved or degraded formosin (∼9kDa) were also detected in this analysis. These results indicate that an interaction between the TM protein (ForG) and formosin occurs within the cell.
Figure 4. Western blotting analysis of a formosin-Ci2a fusion protein (D-Ci2a) or intracellular produced Ci2a in the medium or cells after two days. The secretion was determined in the presence or absence of the three other structural genes in the for-operon, forEFG. Blotting was performed with rabbit antiserum for the Ci2a protein (A) and formosin (B). The expected size of the fusion protein after signal peptide cleavage is 17.9kDa.

40
Absence of the ABC transporter results in secretion and degradation of formosin. To investigate the secretion pattern of formosin a fusion protein was made with a reporter protein. Ci2A was fused to the C-terminal of formosin (Table 1). Expression of the fusion protein was made in the presence and absence of the ABC-transporter. This resulted in two different secretion patterns for the two strains. When the ABC transporter is present (SLds21) the fusion protein and some degradation products with separate Ci2a or formosin could be detected in the medium (Figure 4). In the absence of the ABC transporter (SLds22) only the reporter protein Ci2a was detected in the medium (Figure 4A) while formosin or the fusion protein could not be detected (Figure 4B). To investigate if the Ci2a detected in strain SLds22 was a true secretion or if it was a result of lysed cells during growth resulting in free Ci2a, a strain (SLds53) producing only the reporter protein was created and tested for detection of the reporter protein in the medium (Figure 4A). Results showed that while the SLds53 strain had intracellular production of Ci2a, no significant amount of this protein could be detected in the medium. This demonstrates that the fusion protein is been secreted both in the absence and presence of the ABC transporter although in the absence of the ABC transporter formosin is been degraded leaving only the reporter protein Ci2a in the medium.
Figure 5. SDS-Page and western blotting analysis of the secretion of reporter protein, Ci2a, fused with the N-terminal signal peptide of formosin (SPForD). The secretion was tested in the presence or absence of the ABC transporter by co-expressing the three genes forE, forF, and forG. The N-terminal signal peptide of formosin promotes secretion independent of the ABC transporter. When formosin was fused to the Ci2a, secretion occurred in the absence of the ABC transporter. To investigate this secretion the reporter protein Ci2a was fused to the N-terminal signal peptide (Figure 1B) of formosin to examine the secretion pattern. The secretion then became independent of the ABC transporter (Figure 5). This shows that the N-terminal signal peptide is capable of propagating secretion through one of the pre-existing secretion systems in B. subtilis. Subsequently, these results show that the N-terminal signal peptide of formosin does not code for the secretion signal required for the ABC transporter since the presence of the ABC transporter did not alter the secretion pattern (Figure 5).

41
Figure 6. Western Blotting analysis of the secreted formosin in strains where the native N-terminal signal peptide (SPForD) has been exchanged for the sec-dependent signal peptide from AmyQ (SPAmyQ). The secretion is evaluated in strains with the presence or absence of the ABC transporter consisting of the three genes forE, forF, and forG. Changing the N-terminal signal peptide of formosin does not influence the secretion pattern. To test if formosin could be translocated through the sec-pathway, the endogenous signal peptide of formosin was exchanged for the AmyQ signal peptide. Two strains were created with and without the ABC transporter. Results showed that the secretion remained dependent of the presence of the ABC transporter regardless of the sec-type signal peptide from AmyQ (Figure 6). Consequently, these results show that the N-terminal signal peptide of formosin is not the signal required for secretion through the ABC transporter. The signal seems to be within formosin itself, since exchanging the N-terminal signal did not change the dependence on the ABC transporter. It should be noted that the presence of the N-terminal signal peptide was required for secretion. When an attempt was made to express only the mature part of formosin, the expression failed (results not shown). Results indicate that the secretion signal for the ABC transporter is present within the mature ForD protein, but the presence of the N-terminal part is required. Function of the N-terminal signal peptide could e.g. be to prevent folding prior to secretion.

42
Figure 7. Secretion propagation of different part of the mature formosin protein. (A) Demonstration of the different parts of the mature formosin protein (D1-D3) used as signal peptide in the N-terminal of the reporter protein Ci2a. (B) Western blotting analysis demonstrating the secretion of Ci2a after two days promoted by different parts of formosin. The secretion is evaluated in strains with the presence or absence of the ABC transporter consisting of the three genes forE, forF, and forG represented by the letters E, F, and G. The Ci2a protein was fused to different parts of formosin named D1, D2, or D3 A putative internal signal peptide for the ABC transporter is found in the C-terminal of formosin. Exchanging the endogenous signal peptide to a sec-dependent signal peptide did not alter the secretion pattern. Secretion remained dependent on the presence of the ABC transporter. To determine if there might be an internal signal peptide for the ABC transporter within formosin, different parts of the mature protein were fused to the N-terminal of the reporter protein Ci2a (Figure 7B). Secretion was tested in the presence or absence of the ABC transporter (Figure 7A). Results show that there is a small level of Ci2a coming out of the cells in all the strains where Ci2a is present. But when Ci2a is fused to the C-terminal part of formosin (SLds115) it is secreted in the presence of the ABC transporter in high enough levels to suspect specific secretion. The expected product of this fusion protein is 10.9kDa but the secreted product observed in the gel is 18-19kDa. The reason for this is unknown and the construct was confirmed by DNA sequencing to be correct. This band of 18-19kDa is only observed in the strain where the ABC transporter is present. This indicates that the ABC transporter is involved in the formation of this product since no such product is observed in the strain lacking the ABC transporter that contains the same fusion protein (SLds112).

43
DISCUSSION Results of this study suggest that formosin is secreted through a putative ABC transporter comprised of an ATP binding protein and a TM protein that is co-expressed with formosin in the for-operon. The N-terminal signal peptide of formosin is not the signal for this ABC transporter since it remained dependent on the ABC transporter when this endogenous signal peptide was exchanged to a sec-dependent signal peptide like the AmyQ signal peptide. It was also observed that the signal peptide from formosin could promote secretion of a reporter protein, Ci2a, independently of the ABC transporter. When a fusion protein was made between formosin and Ci2a, the reporter protein could be detected in the medium both in the presence and absence of the ABC transporter. However, only in the presence of the ABC transporter could formosin be detected. This suggest that this ABC transporter is involved in the maturation process of formosin since it seems that in the absence of the ABC transporter the fusion protein had been secreted, but that the formosin part of the fusion protein had been degraded leaving only Ci2a in the medium. Results from this part of the study also suggest that there is an alternative secretion pathway for formosin. In the absence of the ABC transporter this pathway would result in degradation of the formosin protein, leaving only Ci2a to be detected in the medium. Alternatively, formosin could be secreted through e.g. the sec-pathway. The ATP binding protein and the TM protein could instead constitute a chaperon that was required for formosin to be folded correctly into its mature form. This theory would explain why exchanging the N-terminal signal peptide does not alter the dependence of these two proteins and why it was capable of secreting the reporter protein both in the presence and absence of the three proteins in the for-operon. This alternate theory is challenged by the fact that the ATP binding protein contains an ABC transporter motif and that uncleaved formosin is only observed in the presence of the TM protein within the cells indicating interaction prior to secretion. Secondly, this alternate theory is also challenged by results showing that the C-terminal part of formosin is capable of promoting secretion of Ci2a in the presence of the ABC transporter. This suggests that the C-terminal part of formosin is the signal required for secretion through the ABC transporter. C-terminal signal peptide, also referred to as targeting signal, has been found to promote secretion through ABC transporters in Gram negative bacteria. Examples of this include an extracellular lipase, LipA, and a hemophore protein, HasA, from Serratia marcescens (Omori et al., 2001). The HlyA protein in Escherichia coli is another example and the secretion of this protein has been extensively studied since this secretion pathway promotes secretion through both cellular membranes by an ABC transporter in combination with the TolC protein (Nicaud et al., 1986; Koronakis et al., 1989). Secretion of other proteins through this specific secretion pathway has also been made by fusion of the HlyA targeting signal to different proteins that would otherwise be difficult to secrete into the medium (Gentschev et al., 1994; Kenny et al., 1991). Prior to this, C-terminal targeting signals have not been observed to promote secretion in Bacillus. Results in this study indicate such a potential C-terminal targeting signal. However, this secretion system differs from the examples in Gram negative bacteria since formosin contains a cleavable N-terminal signal peptide that was not observed in the other proteins in Gram negative bacteria with a confirmed C-terminal targeting signal. The N-terminal signal peptide has in this study been shown to promote secretion that results in degradation of formosin. The presence of these two separate secretion signals might be a way to regulate formosin, so that it becomes dependent on the presence of the ABC transporter and no intracellular build-up is created in the absence of the ABC transporter.

44
In this study of the C-terminal targeting signal the expression of the ABC transporter and the reporter protein was not optimized. This study only indicated the potential of the C-terminal to promote secretion through the ABC transporter. The C-terminal of formosin was fused to the N-terminal of the Ci2a protein which might decrease the efficiency of the secretion. In the example of the C-terminal target signal of the HlyA protein, the placement of the target signal to the protein to be secreted was found to be crucial in order for secretion to occur (Kenny et al., 1991). The results of this study showed a secreted protein with a much larger size (18-19kDa, Figure 7) when the C-terminal part of formosin (D3) was fused to the N-terminal of the Ci2a protein (10.9kDa). It would therefore be interesting to see the effect of the secretion when the C-terminal of formosin is fused the C-terminal of the Ci2a protein, but also to test the secretion potential of the C-terminal of formosin with other proteins. The ABC transporter is in this study shown to secrete large amounts of formosin. So in an optimized system the potential use of this secretion system for secretion of various substrates could be investigated by optimizing the expression of the two genes (forE and forG) in addition to a further investigation of the signal peptide requirements. The implications of these results also raise the question of how many of the proteins in Bacillus species are thought to be a sec-pathway secreted protein but has a primary secretion pathway through an ABC transporter by a C-terminal targeting signal or an unknown internal signal. In B. subtilis only a few protein are observed with a typical ABC transporter signal peptide even though 77 putative ABC transporters have been found in the genome (Tjalsma et al., 2000). The results of this study may also be relevant for the lactococcin 972 family since formosin belongs to this family (Chapter 4). As described in the previous chapter these proteins are found to be associated with an ABC transporter similar to the formosin ABC transporter and have an N-terminal signal peptide which seems to be a sec-dependent signal peptide (Chapter 4). Could the secretion of protein in the lactococcin 972 family all have a similar mechanism as the one indicated in this study?

45
CHAPTER 6 Immunity associated with formosin production and an investigation of the cellular location of the ForEG ABC transporter
INTRODUCTION Bacteriocins are ribosomally synthesized peptides or proteins produced by bacteria that have antimicrobial activity typically against species closely related to the producer (Belguesmia et al., 2011; Riley and Wertz, 2002). They could be encoded in the chromosome or in mobile genetic elements such as plasmids or transposons (Riley and Wertz, 2002). The production of a particular bacteriocin usually involves expression of an operon consisting of the bacteriocin gene which may include various elements such as a transporter and an immunity protein depending on the system (Nes et al., 1996). Secretion of a bacteriocin occurs in most cases through a special purpose transporter referred to as an ABC transporter (Cotter et al., 2005). The functions of these bacteriocin ABC transporters are quite diverse and some of these possess different types of dual functions. This secondary function could be a proteolytic activity (Havarstein et al., 1995; Paik et al., 1998) or a function in immunity to the secreted bacteriocin (Otto et al., 1998; Hille et al., 2001). One of the required features for a producer of bacteriocin is the need to have immunity to the bacteriocins inhibitory effects (Cotter et al., 2005). Since previous studies have shown that formosin could be produced in the formosin-sensitive strain Bacillus subtilis by expressing only the for-operon (Chapter 5) it appears that protein products derived from the for-operon are conveying immunity to the formosin-producing B. subtilis strain. Therefore the aim of this study is to investigate which of the additional genes in the for-operon might be conveying this resistance. The for-operon (Figure 1) encodes four proteins which are formosin (ForD), a 24.5kDa ATP binding protein (ForE), a 12.8kDa accessory protein (ForF), and a 71kDa transmembrane (TM) protein (ForG).
Figure 1. The for-operon required for the production of the ForD protein. The operon contains the genes with the locus names BL00275-BL00273 as assigned by UniProt, but have been given the gene names forD, forE, forF, and forG.

46
In addition to this immunity determination the cellular location of the ForE and ForG proteins was investigated by fusing the proteins to a green fluorescent protein (GFP). These two proteins constitute an ABC transporter which will be further demonstrated in Chapter 7. In this ABC transporter it is shown that ForE constitutes the nucleotide binding domain (NBD) while ForG is the TM domain (TMD). Results from this study demonstrated that the ForEG ABC transporter provides immunity to formosin. Since it has previously been shown that this ABC transporter is involved in secretion of formosin (Chapter 5), this means that the ForEG ABC transporter has a dual function, working both in the secretion of and immunity to formosin. The ForEG ABC transporter was also shown to localize in the membrane in microdomains similar to those previously described for other protein translocational pathways (Buist et al., 2006). No function could be linked to the accessory protein within the outcome of this study, but a characterization of this protein will be performed in a separate study (Chapter 8).
MATERIAL AND METHOD Bacterial strains and growth conditions. Bacterial strains used for this study are listed in Table 1. B. subtilis cells were made competent by the method of Anagnostopoulos and Spizizen (Anagnostopoulos and Spizizen, 1961). Cells were grown in liquid TY media or Cal 18-2 media (Rasmussen et al., 2000) and plated on LBPG agar plates (LB agar with 0.5% (w/v) glucose and 50mM potassium phosphate, pH 7.0). For MIC determination, cells were grown in cation adjusted Müeller-Hinton II Broth (Becton, Dickinson and Company, USA). When required Chloramphenicol (6 µg/mL), Kanamycin (30 µg/mL) or D-alanine (200µg/mL) was supplemented. Incubation was made at 37°C for plates or TY medium and 30°C for Cal 18-2 medium with 250rpm for liquid cultures. All the produced strains were sequence verified using the Sanger-method (primers not shown) (Sanger et al., 1977). DNA techniques. Chromosomal DNA from B. subtilis was extracted using the protocol for Gram positive bacteria by QIAamp® DNA Mini and Blood Mini Kit (Qiagen). PCR amplifications were preformed with Phusion® Hot Start High-Fidelity DNA polymerase (Finnzymes) using the recommended protocol. PCR products were purified using the QIAquick® Gel Extraction Kit (Qiagen). All gene constructs were put under the control of a triple promoter system (3Pcry) previously described to result in high yields (Widner et al., 2000). Creating strains for immunity screening. To test which genes were required for the immunity to formosin, deletion of the forD gene was made in the previously created SLds1-8 strains (Chapter 5). Deletion was made through splicing by overlap extension PCR (SOE-PCR) as previously described (Horton et al., 1990). Using this method, forD was deleted from strains SLds1-8 by amplifying the flanking regions of forD using chromosomal DNA from the different strains. For the left segment primers d-forD-R and pel-F were used and for the right segment primers pel-R and d-forD-F were used. Primers d-forD-F and d-forD-R were constructed with 5’-tails that were reverse complementary to each other (Table 2). These two segments were then added in equal molar amounts to the SOE PCR where the segments were ligated through the overlapping regions of the primers and amplified by the pel-F and pel-R primers. The resulting construct was then transformed into B. subtilis JA1343 cells to create strains SLds11-18 (Table 1). Growth experiment of B. subtilis 168 with ForD addition. B. subtilis 168 (JA1343) or (SLds11) cells were grown at 37°C in liquid TY medium with a rotation speed of 250rpm. Growth experiments were performed in 50mL cultures that were inoculated with exponentially growing cells resulting in an OD600 of 0.02. Formosin was

47
added to the cultures when an OD600 of approximately 0.5-0.6 was reached. This addition was performed by adding a solution of 1mL to each culture, resulting in a final concentration of 0, 0.5, 1, 1.5, and 2 µg/mL of formosin. The progression of the cultures was then monitored for several hours through optical density measurements to observe the effect of the formosin addition. Sample preparation and western blotting analysis from growth curves. During the growth experiment samples were taken from the cultures. They were prepared for SDS-PAGE by adding one part NuPAGE® LDS sample buffer (4x) to three parts sample and then incubated at 99°C for 3min and stored on ice. Samples used for the SDS-PAGE were the samples taken from the culture from either JA1343 or SLds11 that showed the highest inhibition but still recovered from the addition of formosin. From these SDS samples 10µL were loaded to a precast NuPAGE® 4-12% Bis-Tris gel (Invitrogen). The gel was run with NuPAGE® MES SDS Running Buffer at 200V for 35min. To reduce background in the western blotting analysis the gel was incubated for 30min in MilliQ water. Blotting was performed for 7min using the iBlot® Dry Blotting system (Invitrogen) to a Nitrocellulose Transfer Stack designed for this system. Immunoblotting was made with rabbit primary antibodies against formosin (manufactured internally by Novozymes A/S) in combination with a Novex® WesternBreeze™ Chemoluminescent Anti-Rabbit kit (Invitrogen) using recommended procedures. Immunodetection was made by the provided reaction buffers from the WesternBreeze™ kit. Detection was made and photographed using BioSpectrum® Imaging System (UVP) in combination with VisionWorks®LS Analysis Software (UVP) using the exposure time of 10 or 25min. The images were processed in Adobe Photoshop® CS3 (Adobe Systems) by inverting the colors. For the SLds11 strain samples the exposure time was too short, so in addition to inverting the colors an adjustment of the contrast was made. Antimicrobial susceptibility testing. Minimal inhibition concentration (MIC) and minimal bactericidal concentration (MBC) were determined by the micro dilution broth method according to National Committee for Clinical Laboratory Standards/ Clinical and Laboratory Standards Institute (NCCLS/CLSI) guidelines (The Clinical and Laboratory Standards Institute, 2011). Following these guidelines the test was performed in 96-well microtiter plates (one for each strain), where Müeller-Hinton II Broth (MHB) inoculated with an estimated 105cells/mL of exponentially growing cells had been added to each well. Cell concentration of each strain was confirmed by plating the remaining MHB, which was diluted to an expected amount of 10-100cells/plate on LBPG plates and grown overnight at 37°C. Purified formosin produced internally by Novozymes A/S was added in a two-fold dilution series with final concentrations ranging from 0.024-12.5 µg/mL. Each column had the same concentration and each row contained a new dilution step. As controls one row had pure MHB, while one row had an inoculated MHB without any formosin. The plates were put on a microplate shaker and left to grow for 16h at 37°C with a rotation speed of 300rpm. After incubation the LBPG plates were checked to verify that the MHB added had an acceptable concentration range of 2 to 8 x 105cells/mL. Growth of each well in the microtiter plate was checked and MIC for each strain was determined as the lowest concentration of formosin that had no detectable growth. To determine MBC, 10µL of the four concentrations that did not show visual growth (1 to 8 x MIC) were plated on LBPG plates and incubated overnight at 37°C. MBC is defined as the concentration of formosin that kills ≥99.9% of the bacteria and since the cells/mL was known, the MBC could be determined. After this experiment formosin could be evaluated as being a bactericidal or a bacteriostatic agent. A bacteriostatic agent is defined as having an MBC higher than 4xMIC otherwise it is considered bactericidal. Agar diffusion assay. Since formosin is a large protein in the context of agar diffusion this assay was preformed as previously described for nisin (Pongtharangkul and Demirci, 2004; Tramer and Fowler, 1964). Plates of LBPG agar were prepared with 0.75% agar supplemented with 1% (w/v) Tween20. After autoclaving the medium was

48
cooled down to 40°C and inoculated to an OD600 of 0.5 (approximately 108 CFU/mL). Equal amounts of the agar were poured into plates and left to solidify in room temperature. One sample well was created in the center of each plate through a vacuum punch, which is a small cylindrical metal tube with a 2mm diameter attached through a trap to a vacuum source. Purified formosin (20µL) at a concentration of 4mg/mL was added to the sample wells. To increase diffusion the plates were pre-incubated at 4°C for 24h prior to incubation at 37° for 16h. After incubation pictures were taken and the diameter of the zone of inhibition surrounding the sample wells was measured. Table 1. Strains used in this study.
Strain Description and/or genotype A Strain source Bacillus licheniformis ATCC14580 Natural producer of formosin BGSC Bacillus subtilis 168 JA1343 RUB 200 derivative; amyE, aprE, nprE, spoIIAC Novozymes A/S (Internal) NZgfp Strain with chromosomal GFP expression Novozymes A/S (Internal) NZrfp JA1343; alr::3Pcry RFP Novozymes A/S (Internal) SLds29 JA1343; Δalr::3Pcry comS; KmR Novozymes A/S (Internal) SLds22 JA1343; Δpel::3Pcry Φ(SPforD-ci2a); CmR Chapter 5 SLds1 JA1343; Δpel::3Pcry forDEFG; CmR Chapter 5 SLds2 JA1343; Δpel::3Pcry forDEF; CmR Chapter 5 SLds3 JA1343; Δpel::3Pcry forDEG; CmR Chapter 5 SLds4 JA1343; Δpel::3Pcry forDFG; CmR Chapter 5 SLds5 JA1343; Δpel::3Pcry forDG; CmR Chapter 5 SLds6 JA1343; Δpel::3Pcry forD; CmR Chapter 5 SLds7 JA1343; Δpel::3Pcry forDE; CmR Chapter 5 SLds8 JA1343; Δpel::3Pcry forDF; CmR Chapter 5 Strains created for immunity screening SLds11 JA1343; Δpel::3Pcry forEFG; CmR This study SLds12 JA1343; Δpel::3Pcry forEF; CmR This study SLds13 JA1343; Δpel::3Pcry forEG; CmR This study SLds14 JA1343; Δpel::3Pcry forFG; CmR This study SLds15 JA1343; Δpel::3Pcry forG; CmR This study SLds16 JA1343; Δpel::3Pcry CmR This study SLds17 JA1343; Δpel::3Pcry forE; CmR This study SLds18 JA1343; Δpel::3Pcry forF; CmR This study SLds70 JA1343; Δpel::3Pcry forDEFG; CmR Δalr::3Pcry comS; KmR This study SLds71 JA1343; Δpel::3Pcry forDEFG; CmR alr::3Pcry RFP This study Strains created for localization study B SLds30 JA1343; alr::3Pcry Φ(forG-PEPTlinker-GFP) This study SLds34 JA1343; alr::3Pcry Φ(forE-PEPTlinker-GFP) This study SLds38 JA1343; alr::3Pcry Φ(forG-PEPTlinker-GFP) +Δpel::3Pcry forDEF; CmR This study SLds41 JA1343; alr::3Pcry Φ(forE-PEPTlinker-GFP) +Δpel::3Pcry forDFG; CmR This study A 3Pcry is a promoter system with three promoters and the mRNA stabilization segment (Widner et al., 2000). B PEPTlinker is a four amino acid long linker between the two fused proteins comprised of Pro-Glu-Pro-Thr

49
Creating strains for competition study. To visualize the effect of the producers immunity to the effect of formosin, a culture mixed with a population of wildtype B. subtilis JA1343 (sensitive) and formosin producing B. subtilis SLds1 (resistant) was grown. To distinguish the two strains an intracellular production of red fluorescent protein (RFP) was expressed in SLds1, a strain known to secrete high amounts of formosin. This strain was produced by inserting the expression of RFP into the alr-region. This construct of the RFP expression in the alr-region had previously been made by Novozymes in the strain NZrfp (Table 1). In the alr-region the selection of insertion is made through the restoration of the alr gene so this region needed to be disrupted in SLds1. To create this disruption chromosomal DNA of the D-alanine deficient strain SLds29 was added to competent cells of SLds1. This created the strain SLds70 which was a kanamycin and chloramphenicol resistant as well as a D-alanine deficient strain. The expression of RFP was inserted by adding chromosomal DNA from the NZrfp strain to competent SLds70 cells and selecting for colonies where the alr gene had been restored. This created the strain SLds71 which was a formosin producer with an intracellular production of RFP. Competition growth study. The competition growth was performed by mixing a starting culture in liquid TY medium populated by two different strains, one being labeled by intracellular production of RFP. The growth of the entire culture was followed by measuring the optical density, and the composition of the population in the culture was determined by plating the culture on LBPG plates with a dilution resulting in 50-200colonies/plate. Two different competition growth studies were performed so a comparison could be made. In the first competition study the sensitive strain B. subtilis JA1343 was mixed with B. subtilis SLds71 which is a formosin secreting and resistant strain with intracellular expression of RFP. The second competition study was performed as a comparison using two strains both of which were formosin secreting and resistant strains, one with intracellular RFP expression (SLds71) and one without (SLds1). To start this mixed population culture, the strains were separately grown at 37°C with 250rpm to an optical density of ∼2 (OD600) and then diluted to yield an optical density of 1. The sensitive strain was added in a volume resulting in a calculated optical density of 0.006 and the resistant producer was added in a volume calculated to result in an optical density of 0.004. For the mixed population of the two resistant strains the culture was seeded by a volume calculated to result in an optical density of 0.005 for each strain. These two mixed cultures were then grown at 37°C with 250rpm. The progression of the growth was monitored for seven hours and every 30min a sample was taken out and the optical density measured (OD600) after which the sample was diluted and plated on LBPG plates. Plates were incubated overnight at 37°C after which the strain composition of the two different competition cultures was determined. The strain with the intracellular produced RFP was observed as pink colonies on the plates but to determine the composition with a low error rate the RFP-colonies were counted under UV light using red tinted glasses that block the blue spectrum light so that the RFP fluorescence of the colonies could be visualized. Creation of GFP-fusion proteins. Green fluorescent protein was fused to the C-terminal part of either the TMD protein (ForG) or the NBD protein (ForE). Strains were created using SOE PCR for the insertion into the alr-region of strain SLds29. GFP-fusion to the TM protein was created by four segments. For the first segment primers dal-F and ForG-dal-R were used to amplify the left side of the alr-region and the promoters from strain SLds29. In the second segment primers ForG-F and ForG-R amplified the forG gene using SLds1 as a template. For the third segment the GFP gene was amplified from strain NZgfp using primers GFP-F and GFP-R. Between the forG gene and GFP a linker (Pro-Glu-Pro-Thr) was inserted using the 5’-tail of the SOE primers ForG-R and GFP-F. In the fourth segment the alr gene was restored using B. subtilis JA1343 as a template with the primers GFP-dal-F and dal-R. These four segments were then ligated in an SOE PCR and the final construct was amplified by the primers dal-F and dal-R. The GFP fusion to the NBD protein (ForE) was created in the same manner except that three of the primers ForG-dal-R, ForG-F and ForG-R were exchanged for the primers ForE-dal-R, ForE-F and ForE-R, respectively. Resulting strains with the GFP-fusion proteins were named SLds30 for

50
the ForG-fusion and SLds34 for fusion to the ForE protein. These strains were complemented with the three other genes in the for-operon by transforming SLds30 with chromosomal DNA from SLds2 and transforming SLds34 with chromosomal DNA from SLds4. This resulted in strains containing the four genes with SLds38 having a GFP fusion to ForG and SLds41 where ForE was fused to GFP. The GFP used is the most commonly utilized derivative called enhanced GFP which contains the mutations S65T and F64L which make the fluorescence signal more stable, but the protein is in this study referred to as GFP. Confocal microscopy. To immobilize the bacteria, microscope slides were covered with a fine layer of 0.75% agar (in MilliQ H2O). Bacteria was grown overnight at 37°C on LBPG agar plates and diluted in 1xPBS buffer which was then streaked onto the agar covered microscope slides and a cover slip was put on the slides. The microscope images were taken with the assistance of ImaGene-iT at Lunds University using a Zeiss LSM 510 META confocal microscope combined with the software LSM 510. To excite and visualize the emission of GFP the dichroic mirror HFT 488 and longpass filter LP 505 were utilized. Table 2. Primers used in this study.
Primer ID Sequence (5’-3’)A Template Specificity Amplification primers for insertion sites pel-F AGACGAGCTCCGCAATAAAA Varies Forward primer for pel-region pel-R ACCAGCCGTCCTTTCTACCT Varies Reverse primer pel-region dal-F AATCATCGAAATAACCGCCAA Varies Forward primer for alr-region dal-R AGAGAACCTTTTTTCGTGTGGC Varies Reverse primer for alr-region Deletion of forD d-forD-R CGTTTTCCCTTTGTATTTCATTCCTCCTCTTTCATTCG SLds1-8 SOE primer deletion of forD d-forD-F GGAATGAAATACAAAGGGAAAACGGTTGCT SLds1-8 SOE primer deletion of forD GFP fusion proteins forG-dal-R GCTGGACCTCCAAGAACCATAATACATAATTTTCAAACTGATAAA
ATG SLds29 SOE primer left segment of alr-region
forG-F GAAAATTATGTATTATGGTTCTTGGAGGTCCAGCTT SLds1 SOE primer amplification of forG forG-R TCCTTTACTCATAGTCGGTTCCGGAATTCGCCGTTTGCTG SLds1 SOE primer to fuse GFP to forG with linker GFP-F CCGGAACCGACTATGAGTAAAGGAGAAGAACTTTTCA NZgfp SOE primer to fuse forG to GFP with linker GFP-R AAATGTAATCAAAGTTATTATTTGTATAGTTCATCCATGCCA NZgfp SOE primer amplification of GFP GFP-dal-F TGGATGAACTATACAAATAATAACTTTGATTACATTTTATAATT
AATTTTAACAAAGT SLds29 SOE primer right segment of alr-region
forE-dal-R TCTGTACCTCCCATCAAACATAATACATAATTTTCAAACTGATAAAATG
SLds29 SOE primer left segment of alr-region
forE-F GAAAATTATGTATTATGTTTGATGGGAGGTACAGATGT SLds1 SOE primer amplification of forE forE-R CTTTACTCATAGTCGGTTCCGGTGAGTTACCTCCTAAGTTGCC SLds1 SOE primer amplification of forE A Underlined sequences are the 5’tails used in the SOE-PCR.
Formosin secretion of GFP-fusion strains. Functionality of the GFP-fusion strains SLds 38 and SLds41 was tested by growing the cells for 48h in a special Cal 18-2 medium at 30°C. After this the cell-free medium was extracted by a two step centrifugation. Cells were removed by centrifuging the growth solution at 5000xg for 3min, after which the supernatant was taken and centrifuged at 20.000xg for 10min to remove any additional debris that might interfere with the SDS-PAGE. Samples were prepared for SDS-PAGE by adding two parts of supernatant, one part water and one part NuPAGE® LDS sample buffer (4x). Samples were incubated at 99°C for 3min and put on ice until loading 10µL of the samples on precast NuPAGE® 4-12% Bis-Tris gel (Invitrogen). The gels were run with NuPAGE® MES SDS Running Buffer at 200V for 35min. Gels were stained for 1h in InstantBlue™ (Gentaur) after which the gels where left overnight in MilliQ water to destain.

51
RESULTS
Immunity to formosin is conveyed by one or more of the three other genes in the for-operon. A previous study showed that B. subtilis was sensitive to the antibacterial action of formosin (Chapter 4) but at the same time it was observed that large amounts of formosin could be produced in B. subtilis by expressing the for-operon (Chapter 5). It seemed that one or more of the three genes in the for-operon were able to convey immunity to the accumulating concentration of formosin as it is produced in the B. subtilis production strain. To confirm this, growth profiles of a sensitive B. subtilis strain (JA1343) and a potentially resistant B. subtilis strain (SLds11) were made in the presence of different formosin concentrations. In SLds11 the three genes forE, forF, and forG were expressed while the forD gene had been deleted from the for-operon. Results show that the SLds11 strain was able to withstand a higher concentration of formosin which demonstrated that either the ABC transporter, the accessory protein, or both was conveying immunity in the formosin producer (Figure 2A and 2B). Strain JA1343 with 1µg/mL formosin and strain SLds11 with 1.5µg/mL formosin were shown to recover after a short lytic phase even though formosin was still present in the cultures (Figure 2C and 2D).
Figure 2. Growth profiles of Bacillus subtilis in the presence of different formosin concentrations. Cultures grown at 37°C; when the cultures reached an OD600 of 0.5, one milliliter of solution was added to the culture resulting in different final concentrations of formosin ranging from 0-2µg/mL. The black arrow indicates the addition of formosin. (A) Growth of B. subtilis (wt) with addition of different formosin concentrations. (B) Growth of a B. subtilis strain (SLds11) with the expression of the ABC transporter (ForE and ForG) and accessory protein (ForF), with addition of different formosin concentrations. (C) Western blotting analysis of formosin present in the B. subtilis (wt) culture containing 1µg/mL. (D) Western blotting analysis of formosin present in the B. subtilis (SLds11) culture containing 1.5µg/mL. The exposure time of the SLds11 samples during the western blotting analysis was 10min compared to 25min for the JA1343 strain, so the contrast was increased using Adobe Photoshop® CS3 (Adobe Systems) so intensity of (C) and (D) should not be compared to each other.

52
Table 3. Results of the antibacterial activity of ForD for different strains determined by two methods using a micro dilution broth method to demonstrate the MIC and MBC and an agar diffusion assay that show the growth inhibition.
Strain Genotype or description Micro dilution broth Agar diffusion
MIC (µg/mL) MBC Growth inhibition
± SD a (mm) B. licheniformis ATCC14580 Natural producer of ForD 3,13 4xMIC 17,6 ± 1,8
B. subtilis JA1343 Strain used to create strains SLds11-18 0,78 MIC 26,6 ± 0,5 B. subtilis SLds11 JA1343 Δpel::3Pcry forEFG; CmR 3,13 2xMIC 16,2 ± 0,8 B. subtilis SLds12 JA1343 Δpel::3Pcry forEF; CmR 0,78 MIC 25,4 ± 1,1 B. subtilis SLds13 JA1343 Δpel::3Pcry forEG; CmR 1,56 2xMIC 20,2 ± 0,4 B. subtilis SLds14 JA1343 Δpel::3Pcry forFG; CmR 0,78 MIC 25 ± 1 B. subtilis SLds15 JA1343 Δpel::3Pcry forG; CmR 0,78 MIC 25,6 ± 0,9 B. subtilis SLds16 JA1343 Δpel::3Pcry CmR 0,78 MIC 25,2 ± 0,8 B. subtilis SLds17 JA1343 Δpel::3Pcry forE; CmR 0,78 MIC 25,4 ± 0,9 B. subtilis SLds18 JA1343 Δpel::3Pcry forF; CmR 0,78 MIC 25,2 ± 0,8 a Calculated as mean value of the diameter for the observed growth inhibition of five plates with a standard deviation (SD)
The ABC transporter conveys immunity. To evaluate which genes in the for-operon convey immunity to formosin, strains with different gene patterns of the for-operon were screened for their sensitivity to formosin using two different methods. First a micro dilution broth method was used to determine the MIC and MBC of the different strains. Results demonstrate that the two proteins required for immunity to formosin were the NBD protein (ForE) and the TMD protein (ForG) (Table 3). This was observed in strain SLds13 containing these two proteins, where an increase in MIC was observed compared to B. subtilis JA1343. Higher immunity than that of the SLds13 strain was observed in SLds11, where proteins ForE and ForG are present in addition to the accessory protein (ForF). The SLds11 strain resulted in the same MIC as the B. licheniformis strain. The deletion of the accessory protein seemed to decrease immunity to formosin when one compared the MIC between SLds11 and SLds13, but no other immunity effect was observed in strains containing the accessory protein. MBC was equal to the MIC for all observed sensitive strains having a MIC of 0.78 µg/mL. However, in B. licheniformis the MBC was four times higher than the MIC and in the resistant B. subtilis strains SLds11 and SLds13 the MBC was two times higher than the MIC. Because of these small differences in MIC and MBC, formosin should be considered a bactericidal agent for all tested strains. For a second method an agar diffusion assay was performed and results from this assay correlated with the results from the micro dilution broth method. In sensitive strains a growth inhibition of 25-26mm was observed (Table 3, Figure 3). For B.licheniformis and SLds11 the growth inhibition was observed to be 16-17mm, while strain SLds13 showed a growth inhibition of 20mm. In all B. subtilis strains the zone of inhibition was observed to have a well-defined border but in B. licheniformis this border was more diffuse (Figure 3). In the MBC determination B. licheniformis was observed to have a four times higher MBC than MIC (Table 3) which could be a reason for this type of diffuse border.
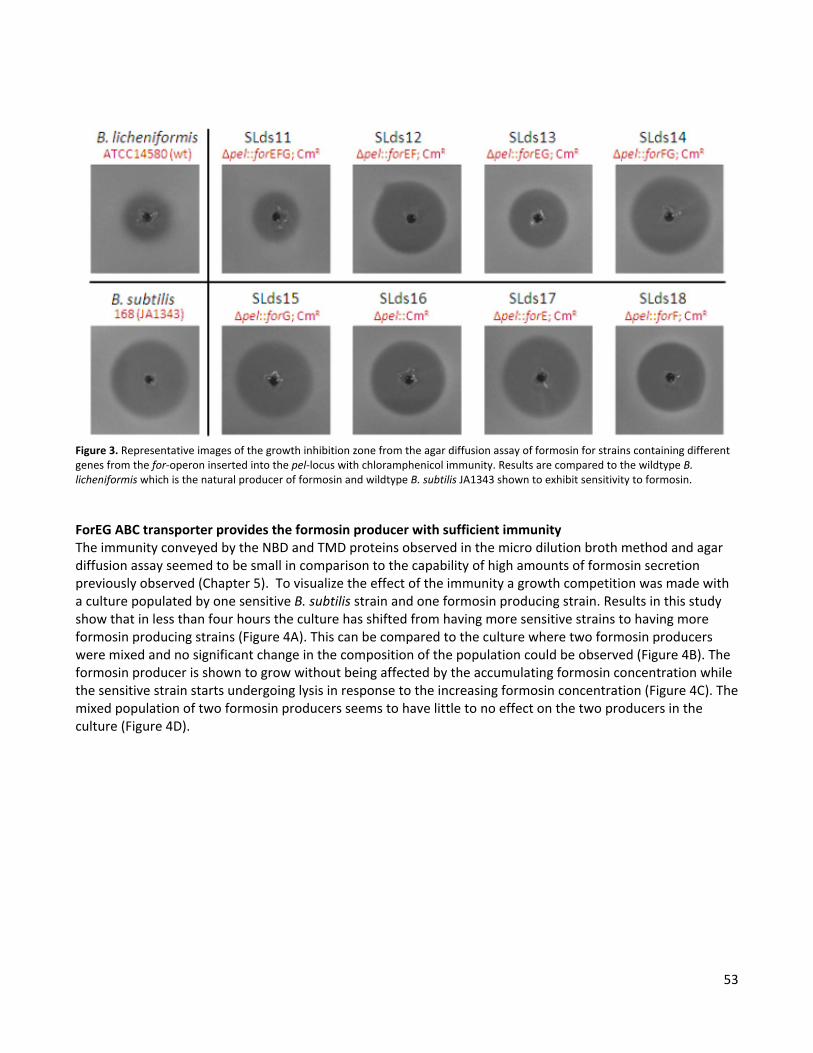
53
Figure 3. Representative images of the growth inhibition zone from the agar diffusion assay of formosin for strains containing different genes from the for-operon inserted into the pel-locus with chloramphenicol immunity. Results are compared to the wildtype B. licheniformis which is the natural producer of formosin and wildtype B. subtilis JA1343 shown to exhibit sensitivity to formosin.
ForEG ABC transporter provides the formosin producer with sufficient immunity The immunity conveyed by the NBD and TMD proteins observed in the micro dilution broth method and agar diffusion assay seemed to be small in comparison to the capability of high amounts of formosin secretion previously observed (Chapter 5). To visualize the effect of the immunity a growth competition was made with a culture populated by one sensitive B. subtilis strain and one formosin producing strain. Results in this study show that in less than four hours the culture has shifted from having more sensitive strains to having more formosin producing strains (Figure 4A). This can be compared to the culture where two formosin producers were mixed and no significant change in the composition of the population could be observed (Figure 4B). The formosin producer is shown to grow without being affected by the accumulating formosin concentration while the sensitive strain starts undergoing lysis in response to the increasing formosin concentration (Figure 4C). The mixed population of two formosin producers seems to have little to no effect on the two producers in the culture (Figure 4D).
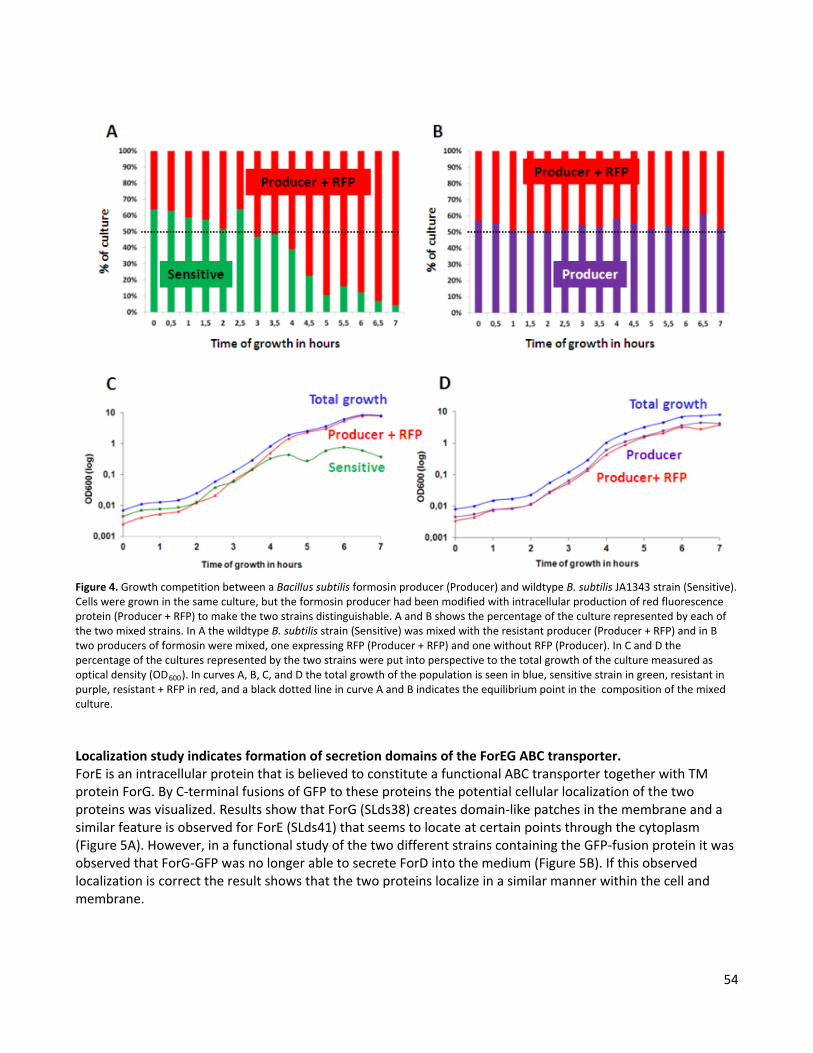
54
Figure 4. Growth competition between a Bacillus subtilis formosin producer (Producer) and wildtype B. subtilis JA1343 strain (Sensitive). Cells were grown in the same culture, but the formosin producer had been modified with intracellular production of red fluorescence protein (Producer + RFP) to make the two strains distinguishable. A and B shows the percentage of the culture represented by each of the two mixed strains. In A the wildtype B. subtilis strain (Sensitive) was mixed with the resistant producer (Producer + RFP) and in B two producers of formosin were mixed, one expressing RFP (Producer + RFP) and one without RFP (Producer). In C and D the percentage of the cultures represented by the two strains were put into perspective to the total growth of the culture measured as optical density (OD600). In curves A, B, C, and D the total growth of the population is seen in blue, sensitive strain in green, resistant in purple, resistant + RFP in red, and a black dotted line in curve A and B indicates the equilibrium point in the composition of the mixed culture. Localization study indicates formation of secretion domains of the ForEG ABC transporter. ForE is an intracellular protein that is believed to constitute a functional ABC transporter together with TM protein ForG. By C-terminal fusions of GFP to these proteins the potential cellular localization of the two proteins was visualized. Results show that ForG (SLds38) creates domain-like patches in the membrane and a similar feature is observed for ForE (SLds41) that seems to locate at certain points through the cytoplasm (Figure 5A). However, in a functional study of the two different strains containing the GFP-fusion protein it was observed that ForG-GFP was no longer able to secrete ForD into the medium (Figure 5B). If this observed localization is correct the result shows that the two proteins localize in a similar manner within the cell and membrane.

55
Figure 5. Results of the localization and formosin secretion for GFP-fusion proteins made with either ForE or ForG. (A) Observed localization of the ForG (SLds38) and ForE (SLds41) within the cells shown by GFP fusions to the C-terminal of the proteins. (B) Secretion of formosin in strain constraining the GFP fusion proteins after two days incubation at 30°C in Cal 18-2 medium. DISCUSSION Results from this study show that the ABC transporter constituted by the NBD protein (ForE) and the TMD protein (ForG) convey immunity to formosin. This illustrates that the ForEG ABC transporter has a dual function and functions both in the secretion of the formosin which was observed in Chapter 5 and in the immunity to formosin observed in this study. This immunity conveyed by the ForEG ABC transporter might seem small but the competition growth study demonstrated that this immunity is enough to protect the producer from the accumulating formosin concentration while non-producers are killed. During this study no function could be linked to the accessory protein. However, it was observed that the deletion of the accessory protein lowered the level of immunity. This could be because of the lack of the accessory protein resulting in a lower immunity or the lack of the gene in the for-operon resulting in a different expression pattern for the remaining genes. It would therefore be relevant to examine the effect of immunity to formosin if the accessory protein was complemented to the SLds13 where it had been deleted. If this complementation increases immunity it would indicate an immunity function of the accessory protein. This complementation was tested with regards to formosin secretion in Chapter 8, but regrettably never attempted with regards to a possible immunity function towards formosin. The location of the ForEG ABC transporter seemed to be arranged in microdomains throughout the cellular membrane. This subcellular localization was observed for both the TM protein ForG and the intracellular protein ForE. Subcellular sites for bacterial protein export have previously been observed in Gram positive bacteria (Buist et al., 2006). This includes localizations observed in B. subtilis for the two major secretion pathways such as the spiral-like structures of SecA and SecY in the sec-pathway (Campo et al., 2004) and the polar formations of TatC in the tat-pathway (Meile et al., 2006). The localization of the ForEG ABC transporter

56
seems to be creating a similar spiral-like pattern to that observed for the Sec-pathway in B. subtilis (Campo et al., 2004). However, this was never confirmed in this study since imaging stacks of different optical sections to produce 3D images was difficult due to bleaching of the GFP-signal. It was also shown that the ForG-GFP fusion protein could no longer secrete formosin which might question the observed localization. Moreover, it would also be interesting to determine if subcellular locations of the NBD (ForE) are present even in the absence of the TMD (ForG). Such a test would show if the NBD is co-localized with the TMD or if this observed localization is independent of the presence of the TMD. Since the function of the TMD (ForG) was disturbed by the fusion to GFP it would also be advisable to construct the fusion protein using a longer and more flexible linker-peptide between ForG and GFP. Besides the linker there was also an issue with a high degree of bleaching of GFP that needs to be resolved so that a 3D image of the localization can be made to provide additional information about the localization structures. Since formosin is related to the lactococcin 972 family (Chapter 4) results from this study could possibly be indicative of all ABC transporters associated with the lactococcin 972 family. Therefore, it would be of interest to test immunity functions of other ABC transporters associated with lactococcin 972-like proteins.

57
CHAPTER 7 In silico analysis of the ForEG ABC transporter
INTRODUCTION ATP binding cassette (ABC) transporters represent the largest and most diverse family of transporters (Higgins, 1992). These transporters utilize energy released in ATP hydrolysis to translocate a wide range of substrates across membranes working as importers or exporters, but never both (Rees et al., 2009). ABC transporters are recognized by characteristic features of the ATP binding cassette and have a typical architecture consisting of four domains, two transmembrane domains (TMDs) and two nucleotide binding domains (NBDs)( Wen and Tajkhorshid, 2011). While the NBDs possess conserved motifs involved in the ATP hydrolysis, the TMDs are quite variable in both sequence and structure, reflecting the diversity in translocated substrates (Rees et al., 2009). The ABC transporters are divided into three separate folds, two of them represented by the prokaryotic large or small ABC importers and the third being the ABC exporter found in both prokaryotes and eukaryotes (Rees et al., 2009; Wen and Tajkhorshid, 2011). In the previous chapters it was shown that the ForE and ForG proteins were vital for the formosin secretion and immunity (Chapter 5, Chapter 6). It was believed that these two proteins form an ABC transporter where ForE is the NBD and ForG is the TMD. In this study the putative ForEG ABC transporter was characterized in silico and a putative structure was predicted. In addition, the putative interaction site between ForE and ForG was investigated. In the NBD the Q-loop is known to interact with the TMD (Oswald et al., 2006; Wen and Tajkhorshid, 2011). In the TMD the interaction site for ABC importers occurs in two conserved coupling helices called the EAA motif (Wen and Tajkhorshid, 2011; Rees et al., 2009). In ABC exporters the interaction site is less defined and based on protein structures from ABC exporters that do not have a protein secreting function, so the site in ForG that interacts with ForE predicted in this study is highly speculative. In this study it was observed that ForE contained several of the conserved motifs of an NBD protein for ABC exporters. ForG was observed to be constructed of seven TM regions, and two putative sites on ForG were predicted as possible interaction sites for ForE or ForD in this study. MATERIAL AND METHOD Topology model of the ForG protein. A computer based topology model of the TMD protein (ForG) was created by evaluations of two different methods. The first method was a sequence based analysis comparing results from four different topology prediction programs. This included the programs OCTOPUS (Viklund and Elofsson, 2008b), SCAMPI-msa (Bernsel et al., 2008), HMMTOP (Tusnády and Simon, 2001), and TMHMM

58
(Krogh et al., 2001). In the second method a multiple sequence alignment was performed with selected results from protein sequence based BLAST (Altschul et al., 1990) that were aligned using ClustalW 2.1 (Larkin et al., 2007). This included four proteins from different Gram positive bacteria with sequence identity ranging from 38-18% to ForG. The topology was predicted for the aligned proteins using the TM topology prediction method MEMSAT3 (Nugent and Jones, 2009) and the signal peptide of the proteins was predicted using Phobius (Käll et al., 2004) since it is capable of distinguishing between a TM region and a signal peptide. Based on the consensus results from these five prediction programs a putative model of ForG was created. Results of the multiple sequence alignment could then be used to verify the correct number of TM regions. Characterization of the ForE. Initially the ForE protein was determined to be the ATP binding protein of an ABC transporter by a database search of protein patterns and families (Chapter 4). To verify this and to gain further understanding of ForE a multiple sequence alignment (Larkin et al., 2007) was performed as previously described for ForG. This alignment indicated conserved regions within ForE that were used to identify different functional domains from the current knowledge of ATP binding proteins in ABC transporters. Interaction sites in ForG. The multiple sequence alignment was also used to predict intracellular loops of ForG that could then be used to predict the interaction site with ForE. In addition, the secondary structure of the intracellular loops of ForG was predicted using PSIPRED v3.0 (Jones, 1999) to assist in the prediction of the interaction site. In the multiple sequences alignment only the three proteins with the highest homology (38-29% sequence identity) to ForG were used to reveal interaction sites. RESULTS AND DISCUSSION The predicted topology model of the ForG shows a seven TM protein. The topology, TM regions, and the signal peptide of the ForG protein were predicted by five different topology prediction programs (Appendix III, Figure 1) and a prediction of the signal peptide using Phobius (Käll et al., 2004). The results show a TM protein with an initial eight TM regions containing a signal peptide cleavage resulting in a protein consisting of seven TM regions (Figure 1). The prediction that ForG is a seven TM protein was further confirmed by a multiple sequence alignment (Appendix III, Figure 2) where proteins with homology to the ForG protein were observed to consist of the same number of TM regions. In addition to the predicted topology seen in Figure 1 one also has to consider the proline-induced disruption of a TM α-helix. This occurs when a proline residue is one to two helix turns (<3.6-7.2 residues) from the TM-region end (Nilsson et al., 1998). Four proline residues were found in the predicted TM regions of this topology model. The proline residues seen in TM4 and TM7 have by one or more prediction program been determined to disrupt the TM α-helix. In TM5 the proline residue could induce such a disruption of the TM region subsequently leading to a termination of the TM5 at the proline residue instead of the TM5 region predicted in the topology model (Figure 1). In TM8 the proline residue is found to be 7-8 residues from the predicted end for the TM region and thus it is unlikely that this residue induces a disruption. Instead, the presence of the proline in TM8 might indicate a helix kink observed to associate with a TM imbedded proline residue (Yohannan et al., 2004). In the multiple sequence alignment (Appendix III, Figure 2) it was observed that three of the five sequences used in the alignment contained a conserved proline residue in the last TM region (TM8). This imbedded proline might indicate an importance of a helix kink for the protein function or the conformational change that the protein undergoes during translocation. Although no conserved proline residue was found in two of the five aligned protein sequences (Appendix III, Figure 2), it is still possible that a kink is present in these two proteins since it is known that while proline creates a kink not all kinks are created by a proline (Yohannan et al., 2004).

59
Figure 1. Topology model of ForG showing the eight transmembrane (TM) regions as determined by five different prediction programs. The residues within each TM domain are all depicted to be in the membrane for all the prediction programs used. The residues with a grey background are residues where one or more prediction programs predicted them to be in the membrane. Residues in red at the N-terminal are the signal peptide predicted by Phobius. Proline residues predicted to be in a TM domain are highlighted since they may cause conflicts in the predicted topology model through a proline-induced disruption of a TM α-helix. Motifs found in ATP binding proteins of ABC exporters could be identified in ForE. Searches in databases able to recognize pattern, signature, and protein family, showed that ForE was predicted to be an NBD protein of an ABC transporter (Chapter 4). To confirm this, a multiple sequence alignment was used and several motifs known to be present in NBDs of ABC transporters were identified (Figure 2). The Walker A motif (Walker et al., 1982) was identified from the amino acid sequence GxxxxGK(T/S), where x is any type of amino acid. The Walker B motif (Hanson and Whiteheart, 2005) was identified from the amino acid sequence hhhhDE, where h is a hydrophobic amino acid. The ABC signature region (Davidson et al., 2008) was identified from the amino acid sequence known to be similar to LSGGQ. In addition three motifs were found through the observation of highly conserved amino acids or regions. The H-loop (Oswald et al., 2006) consisting of a conserved histidine residue in the C-terminal of ForE. The D-loop (Hanson and Whiteheart, 2005) was observed in connection to the Walker B motif consisting of the amino acid sequence (S/A)LD. The Q-loop (Oswald et al., 2006) was identified as a conserved glutamine residue in a region that was furthermore observed to contain three conserved negatively charged amino acids positioned six residues downstream of the glutamine residue. Also identified was the ENI motif which is known to be highly conserved among ABC exporters (Wen and Tajkhorshid, 2011).

60
Figure 2. Multiple sequence alignment of the ForE where structural domains have been identified. The seven proteins aligned in this figure are all ABC transporter associated proteins which are named according to their gene or ordered locus name. This includes (from the top): forE, an ATP binding protein from Bacillus licheniformis; GYMC61_1571, an ABC transporter related protein (60% identity) from Geobacillus sp. Y412MC61; lolD, an ABC superfamily ATP binding cassette transporter (40% identity) from Parascardovia denticolens; macB, an ABC superfamily ATP binding cassette transporter (40% identity) from Corynebacterium pseudogenitalium; Tfu_0812, an ABC-type antimicrobial peptide transport system ATPase component (40% identity) from Thermobifida fusca; smi_1970, an ABC transporter ATP-binding protein (35% identity) from Streptococcus mitis; SSGG_05546, an ABC transporter ATP-binding protein (40% identity) from Streptomyces roseosporus. The alignment was performed using ClustalW where the *(asterisk) represents a conserved amino acid, a : (colon) indicates conservation between amino acids of strongly similar properties, and a . (period) represents conservation between amino acids of weakly similar properties. Five conserved regions were observed in the alignment (colored background) that could then be identified as motifs in nucleotide binding domains (NBDs) of ABC transporters.

61
Figure 3. Multiple sequence alignment made by ClustalW of the three intracellular loops (ICL) of the transmembrane protein (ForG) indicating conserved regions and the secondary structure of ForG predicted by PSIPRED v3.0. Proteins aligned are named according to their gene or ordered locus name and are (from the top): forG from B. licheniformis; Jden_0042, an unknown protein (38% identity) from Jonesia denitrificans; Geoth_1312, an unknown protein (36% identity) from Geobacillus termoglucosidasius; BLIJ_0444, an unknown protein (29% identity) from Bifidobacterium longum. In the alignment the *(asterisk) represents a conserved amino-acid, a : (colon) indicates conservation between amino-acids of strongly similar properties, and a . (period) represents conservation between amino-acids of weakly similar properties. Conserved regions in the first and last intracellular loops of ForG might be possible interaction sites for ForE. In the region of the TMD protein (ForG) where it interacts with the NBD protein (ForE) or formosin (ForD), one might expect that there should be a degree of conservation between homologous proteins. For this purpose a multiple sequence alignment was performed for ForG and the three intracellular loops (ICL1-3) were screened for any conserved regions and their secondary structure. The N-terminal was not considered since it contained the signal peptide and the C-terminal was short with little conservation between the proteins in the multiple sequence alignment (Appendix III, Figure 2). Results show a conserved region in the ICL1 (R-RALRVL-G---RIQ) that was also predicted to have a β-strand configuration (Figure 3). This conserved region was also found to contain positively charged residues in an almost alternating pattern like “+a+a+aa+” where + is a positively charged residue and a is an uncharged residue. In ICL3 a second conserved region was observed (LQAKRDFPL) but no particular secondary structure was predicted in this region. Results from the predicted secondary structure also show that ICL2 and ICL3 were surrounded by predicted α-helices that were extensions of the membrane helices from the flanking TM regions. CONCLUSIONS In this study the analysis of the ForE protein showed that this protein contained conserved regions such as the signature region, ENI motif, and the Q-loop subsequently confirming that ForE is the NBD of an ABC transporter. The ENI motif has been observed to be highly conserved in ABC exporter systems and the Q-loop is in this analysis important since it is believed to be the site that interacts with the TMD (Wen and Tajkhorshid, 2011). In ABC transporters, NBDs are observed to be highly conserved in sequence and structure between different systems while TMDs exhibit a wide range of diversity in both sequence and numbers of membrane spanning regions (Rees et al., 2009). The topology model of ForG made in this study shows that this protein has seven membrane spanning regions and an additional membrane spanning region in the N-terminal which constitutes the signal peptide. A typical TMD usually contains 6 membrane spanning regions but a wide range of diversity has been observed in the number of membrane spanning regions (Rees et al., 2009). The seven TM regions and the two large extracellular domains in the TM protein associated with the lactococcin 972 family seem to be conserved (Data not shown). This indicated that while sequence homology between these TMD proteins might be low, the structure homology seems to be conserved.

62
Interaction sites of the TMD to the NBD have been observed in small and large ABC importers to occur in two conserved coupling helices called the EAA motif (Wen and Tajkhorshid, 2011; Rees et al., 2009). However, in the ABC exporter the EAA is absent and the interaction site is less defined (Wen and Tajkhorshid, 2011). The interaction occurs on the intracellular part of the TM protein, so in this study the ICL was screened for conserved regions in an attempt to find the site in ForG that could interact with the ForE protein. Two conserved regions were found in the ICL, one in ICL1 and another in ICL3. In ICL1 a pattern of alternating positively charged residue was found in the conserved region that was predicted to be in a β-strand configuration. Since a β-strand position neighboring residues on opposite sides of the backbone these alternating positive charges will create a positively charged side of the predicted β-strand. This is interesting since it was observed that there was a cluster of negatively charged residues downstream of the Q-loop in ForE. In ICL3 the conserved region itself was not predicted to contain any defined secondary structure but the region was observed to have flanking α-helices that connect to the helices in the TM regions, which in turn could convey conformational change in the TM helices induced by the ATP hydrolysis in the ForE protein. For the substrate targeting of ForD it could be observed in Chapter 5 (Figure 3) that uncleaved ForD was associated with the ForG protein. Because of this result it is suspected that the specificity of the targeting of ForD secretion occurs in ForG. Since results of the secretion analysis (Chapter 5) also indicated that the C-terminal was the translocation signal in ForD it is expected that this is the part of ForD interacting with ForG in the ICL or the intracellular phase of the TM regions. However, it is difficult to predict any type of interaction site between the ForD and ForG protein since it has been observed in the C-terminal target signal of e.g. HlyA that it is not sequence specific. Instead, it is believed that the targeting is an independently folded target motif (Kenny et al., 1991). But this still leaves the possibility that some of the conserved regions of the ICLs could be interaction sites with ForD rather than the interaction site between the ForE and ForG. The described interaction sites to ForE observed in ICL1 and ICL3 are highly speculative interaction sites but they may serve as good targets for mutational studies to experimentally determine the sites on ForG that interact with ForE. Since the sites also could be targets for ForD, one needs to distinguish between the loss of interaction to ForE or ForD. Both interactions are assumed to cause a disruption of the ForD secretion, but if the interaction is disrupted to ForE, one might still expect that uncleaved ForD will be associated with ForG within the cells as observed in Chapter 5 (Figure 3).

63
CHAPTER 8 Characterization of the formosin accessory protein ForF found in association with the ForEG ABC transporter INTRODUCTION
The term accessory protein is a broad description of a most often uncharacterized protein that is associated with another protein of known function. The term is used for proteins involved in e.g. protein folding, cell signaling, and secretion (Fink, 1999; Ortiz de Orué Lucana and Groves, 2009; Varcamonti et al., 2001). Secretion related accessory proteins are associated with ATP-binding cassette (ABC) transporters involved in protein secretion in both Gram positive and Gram negative bacteria (Young and Holland, 1999). In addition, these accessory proteins have also been associated with ABC transporters involved in antibiotic resistance in Gram negative bacteria (Nikaido and Zgurskaya, 2001). In this study the accessory protein ForF is characterized. This is an accessory protein associated with the ForEG ABC transporter which is observed to be involved in both secretion of and immunity to the chromosomally encoded bacteriocin from Bacillus licheniformis called formosin (Chapter 5, Chapter 6). Genes associated with formosin are encoded in the for-operon consisting of formosin (ForD), a nucleotide binding domain (NBD) of an ABC transporter (ForE), an accessory protein (ForF), and a transmembrane domain (TMD) of an ABC transporter (ForG) (Chapter 7). The function of ForF with regards to formosin secretion is examined through complementation test and different forF-deletion strains. To characterize the ForF accessory protein the sequence of this protein is analyzed in silico to find possible homologues, predict a secondary structure, and analyze properties of the N-terminal. The broad definition of the name accessory proteins makes the description of these types of proteins difficult. To avoid confusion the criteria for being characterized as an accessory proteins in this study are therefore specified, although other studies might have a different definition. Accessory proteins are proteins not part of the fundamental ABC transporter model consisting of two NBDs and two TMDs. They are proteins encoded in genes adjacent to the ABC transporter genes. They are single spanning membrane proteins with an N-terminal transmembrane (TM) region. However, they could also be secreted proteins, provided that their function can be directly linked to the function of the ABC transporter and that they are not a substrate of the ABC transporter. In addition, these proteins should not be predicted to have functions unrelated to the ABC transporter system. In Gram negative bacteria, accessory proteins are sometimes called membrane fusion proteins (MFP). These are proteins with a characteristic N-terminal TM spanning region and average size of ∼450 amino acids, which are found to facilitate secretion in ABC transporter systems through the inner and outer membrane (Sablon et al., 2000). In Gram positive bacteria this feature is not required since these bacteria only have one membrane, but a few MFP homologues have been identified in Gram positive bacteria (Harley et al., 2002; Chung et al.,

64
1992). The function of accessory proteins in Gram positive bacteria is unknown and there are very few examples where a function has been defined for any of these accessory proteins. The only example found for this study is the accessory protein LncD that has been observed to be essential for the secretion of Lactococcin A (Franke et al., 1996; Varcamonti et al., 2001) and has been characterized as an MFP homologue (Harley et al., 2002). However, defining accessory proteins from Gram positive bacteria based on their homology to proteins from Gram negative bacteria, known to have specific functions important for secretion over two cellular membranes, limits the perspective view of these proteins. As part of the characterization of ForF, other accessory proteins associated with ABC transporters of the Lactococcin 972 family were also identified and characterized. The identified accessory proteins associated with this type of ABC transporter could then be compared to ForF. These accessory proteins were observed to be diverse, as four different classes of accessory proteins could be distinguished. Characterization of the ForF accessory protein revealed that the protein had no noticeable function in the secretion of formosin. However, ForF was observed to be associated to the ForEG ABC transporter as overexpression of this accessory protein resulted in the detection of this protein in medium when the ABC transporter was present. The detection of ForF in the medium was interesting since the predictions performed in this study showed that this was a single spanning membrane protein with an N-terminal TM region. This is made even more intriguing since the size of the protein observed in the medium indicates that the N-terminal TM region is still present in the protein. Homologues to ForF identified during this study also indicated that the protein might be functional in a homomultimeric state. Furthermore, these homologues also showed that a non-lactococcin 972-like protein could potentially be using a very similar ABC transporter system to the lactococcin 972 family by having homology to the C-terminal part of the formosin protein. MATERIALS AND METHODS
Bacterial strains and growth conditions. Bacterial strains used for this study are listed in Table 1. B. subtilis cells were made competent by the method of Anagnostopoulos and Spizizen (Anagnostopoulos and Spizizen, 1961). Cells were grown in liquid TY media or Cal 18-2 media (Rasmussen et al., 2000) and plated on LBPG agar plates (LB agar with 0.5% (w/v) glucose and 50mM potassium phosphate, pH 7.0). When required chloramphenicol (6 µg/mL) or Kanamycin (30 µg/mL) was supplemented. Incubation was made at 37°C for plates or TY medium and 30°C for Cal 18-2 medium with 250rpm for liquid cultures. All the produced strains were sequence verified using the Sanger-method (primers not shown) (Sanger et al., 1977). DNA techniques. Chromosomal DNA from B. subtilis was extracted using the protocol for Gram positive bacteria by QIAamp® DNA Mini and Blood Mini Kit (Qiagen). PCR amplifications were preformed with Phusion® Hot Start High-Fidelity DNA polymerase (Finnzymes) using the recommended protocol. PCR products were purified using the QIAquick® Gel Extraction Kit (Qiagen). Expression cassettes or gene deletions were made through splicing by overlap extension PCR (SOE-PCR) as previously described (Horton et al., 1990). Primers used are listed in Table 2. All gene constructs were put under the control of a triple promoter system (3Pcry) previously described to result in high yields (Widner et al., 2000). Insertion of forF into the amyE locus. In the B. subtilis strain used (JA1343) there is a small deletion of the amyE gene. However, the site can still be utilized as an insertion point using the flanking regions of amyE. This is exploited to create a strain that contains the forF gene expressed in trans so the effect of the ForF protein can be observed by inserting this construct into different strains. The strain is created through SOE PCR fusing

65
together four segments. The left side of the amyE region was amplified from B. subtilis JA1343 using the primers amy-F and L-amy-R. From the strain SLds18 the promoter system (3Pcry) and forF was amplified using primers 3P-forF-F and 3P-forF-R. The selection used was kanamycin resistance which was amplified from SLds29 using primers Km-F and Km-R. The right side of the amyE region was amplified from B. subtilis JA1343 using the primers R-amy-F and amy-R. The four segments were purified and added in equal molar amounts to the SOE PCR where these segments were ligated through the overlapping regions of the primers and amplified by the amy-F and amy-R primers. The resulting construct was transformed into B. subtilis JA1343 to create strain SLds20. By adding chromosomal DNA from SLds20 to competent cells of SLds3, SLds11, SLds22, and SLds1 the strains SLds19, Slds170, SLds171, and SLds172 were created, respectively. In strain SLds22 and SLds171 formosin is fused to a reporter protein called Ci2a used to determine the secretion pattern of the formosin in a previous study (Chapter 5). This protein is a small protease inhibitor, Subtilisin-chymotrypsin inhibitor-2A (Ci2A), from Hordeum vulgare (Barley) (Halkier et al., 1993). Table 1. Bacillus subtilis 168 strains used in this study.
Strain Description and/or genotypeA Strain source JA1343 RUB 200 derivative; amyE, aprE, nprE, spoIIAC Novozymes A/S (Internal) SLds29 JA1343; Δalr::3Pcry comS; KmR Novozymes A/S (Internal) Slds22 JA1343; Δpel::3Pcry Φ(SPforD-ci2a); CmR Chapter 5 SLds1 JA1343; Δpel::3Pcry forDEFG; CmR Chapter 5 SLds11 JA1343; Δpel::3Pcry forEFG; CmR Chapter 6 SLds18 JA1343; Δpel::3Pcry forF; CmR Chapter 6 SLds3 JA1343; Δpel::3Pcry forDEG; CmR Chapter 5 SLds3new JA1343; Δpel::3Pcry forDEFG; CmR This study SLds3mut JA1343; Δpel::3Pcry forDE-forG(V226L); CmR This study SLds20 JA1343; ΔamyE::3Pcry forF; KmR This study SLds19 JA1343; Δpel::3Pcry forDEG; CmR + ΔamyE::3Pcry forF; KmR This study SLds170 JA1343; Δpel::3Pcry forEFG; CmR + ΔamyE::3Pcry forF; KmR This study SLds171 JA1343; Δpel::3Pcry Φ(SPforD-ci2a); CmR + ΔamyE::3Pcry forF; KmR This study SLds172 JA1343; Δpel::3Pcry forDEFG; CmR + ΔamyE::3Pcry forF; KmR This study A 3Pcry is a promoter system with three promoters and the mRNA stabilization segment (Widner et al., 2000).
Creation of alternate deletion of ForF. Strain SLds3 was previously created to screen for the genes required for secretion of formosin (Chapter 5). In this strain the forF gene was deleted from the for-operon which lowered the secretion level. Since the forE and forF genes overlap with four nucleotides the deletion in SLds3 strain was created leaving two amino acids from ForF followed by an inframe stopcodon so not to disrupt the forE gene. To test the effect of this initial deletion, a second deletion of forF was made. This time leaving the first 13 amino acids of ForF, followed by an inframe stopcodon. This strain was created using SOE PCR as previously described for the deletion of forF (Chapter 5) with the exception that instead of using the primers d-forF-F and d-forF-R the primers d-forF2-F and d-forF2-R were used. The resulting construct was then transformed into B. subtilis JA1343 to create strain SLds3new. Besides the SLds3 and SLds3new strains a third strain was discovered during the DNA sequencing of SLds3 (Chapter 5). This strain was found to contain a missense mutation in the TMD protein (ForG) where Valine226 had been changed to Leucine226. The strain was given the name SLds3mut and tested to see any effect of the mutation with regards to the secretion of formosin.
Sample preparation and western blotting analysis. Secretion patterns of the different strains were determined by growing the cells for 48h in a special Cal 18-2 medium at 30°C. After this the cell-free medium was extracted by a two step centrifugation. The cells were removed by centrifuging the growth solution at 5000xg for 3min,

66
after which the supernatant was taken and centrifuged at 20.000xg for 10min to remove any additional debris that might interfere with the SDS-PAGE. Samples were prepared for SDS-PAGE by adding two parts of supernatant, one part water and one part NuPAGE® LDS sample buffer (4x). These samples were incubated at 99°C for 3min and put on ice until loading 10µL of the samples on precast NuPAGE® 4-12% Bis-Tris gel (Invitrogen). To estimate the size of the bands on the western blotting membrane 15µL of Novex® Sharp Pre-strained Protein standard was loaded in one of the lanes. The gel was run with NuPAGE® MES SDS Running Buffer at 200V for 35min. To reduce background in the western blotting analysis the gel was incubated for 30min in MilliQ water. Blotting was performed for 7min using the iBlot® Dry Blotting system (Invitrogen) to a Nitrocellulose Transfer Stack designed for this system. Immunoblotting was made using rabbit primary antibodies against either formosin or Ci2a (manufactured internally by Novozymes A/S) in combination with a Novex® WesternBreeze™ Chemoluminescent Anti-Rabbit kit (Invitrogen) using recommended procedures. Immunodetection was made using the provided reaction buffers from the WesternBreeze™ kit. Detection was made and photographed using BioSpectrum® Imaging System (UVP) in combination with VisionWorks®LS Analysis Software (UVP) using the exposure time of 15min. Images were processed in Adobe Photoshop® CS3 (Adobe Systems) by inverting the colors of the images. For SDS-gel images gels were stained for 1h in InstantBlue™ (Gentaur) after which the gels where left overnight in MilliQ water to destain. Table 2. Primers used in this study. Primer ID Sequence (5’-3’)A Template Specificity Amplification primers for insertion sites pel-F AGACGAGCTCCGCAATAAAA SLds1 Forward primer for pel-region pel-R ACCAGCCGTCCTTTCTACCT SLds1 Reverse primer pel-region amy-F GTCAAGTGGCTGCGGTTTAT JA1343 Forward primer for amyE-region amy-R CCGTCCAAATATGTGCTGAC JA1343 Reverse primer for amyE-region Partial deletion of forF d-forF2-R ATTGTGTAGAAGTTGAGTTATATGACGACACCGATAATGATCTT SLds1 SOE primer deletion for part of forF d-forF2-F TCGGTGTCGTCATATAACTCAACTTCTACACAATTCACTCG SLds1 SOE primer deletion for part of forF Insertion of forF into the amyE locus L-amy-R GTTCGCTTCGCTTCGTTTTGCAAACATTCTTGA JA1343 SOE primer left segment of amyE-region 3P-forF-F TTGCAAAACGAAGCGAAGCGAACACTTGATT SLds18 SOE primer amplification of 3Pcry and forF 3P-forF-R TCCTAAGCATCCTTCACAATCCTCCAAGGGCTCATA SLds18 SOE primer amplification of 3Pcry and forF Km-F CTTGGAGGATTGTGAAGGATGCTTAGGAAGACG SLds29 SOE primer amplification of Km Km-R AGAACCGCTTAAGCCCCGATTTAAGCACACCCTTT SLds29 SOE primer amplification of Km R-amy-F GCTTAAATCGGGGCTTAAGCGGTTCTCTTCC JA1343 SOE primer right segment of amyE-region A Underlined sequences are the 5’tails used in the SOE-PCR.
Predicting signal peptide or TM region in the N-terminal of ForF. To distinguish if the N-terminal of the ForF accessory protein contains a signal peptide or if ForF is a single-spanning membrane protein, different predictions were used. To predict if ForF is a membrane protein, topology and TM prediction programs such as MEMSAT3 (Nugent and Jones, 2009), OCTOPUS (Viklund and Elofsson, 2008b), SCAMPI-msa (Bernsel et al., 2008), and TMHMM (Krogh et al., 2001) were used. To predict the presence of a signal peptide, prediction programs such as SignalP-HMM (Nielsen and Krogh, 1998), SPEPlip (Fariselli et al., 2003), PrediSi (Hiller et al., 2004), and SignalP 4.0 (Petersen et al., 2011) were used. Besides the specialized programs of TM and signal peptide predictions, three other programs were used. These were Phobius (Käll et al., 2007), SOSUIsignal (Gomi et al., 2004), and SPOCTOPUS (Viklund et al., 2008a) which are programs able to detect and distinguish between TM regions and signal peptides. Identifying proteins similar to ForF. Homologues of the ForF accessory proteins were identified by a protein BLAST (Basic Local Alignment Search Tool) program used to search the UniProtKB database (The UniProt Consortium, 2012). From these results two homologues were aligned with ForF using ClustalW 2.1 (Larkin et

67
al., 2007) and this alignment was compared to the secondary structure of the ForF protein predicted using PSIPRED v3.0 (Jones, 1999). Comparison of putative accessory proteins associated with the lactococcin 972 family. The genomic regions of genes annotated to code for a putative protein belonging to the lactococcin 972 family were searched to identify possible accessory proteins. These accessory proteins were identified by the criteria that they had to be expressed adjacent to the ABC transporter, contain an N-terminal TM region, and have no unrelated function. The N-terminal TM region or signal peptide was predicted by the same method as described for the ForF protein and possible functional domains were identified by InterProScan (Quevillon et al., 2005). Prediction of the secondary structures was performed using PSIPRED v3.0 (Jones, 1999). After the different accessory proteins had been characterized they were placed into different groups depending on their predicted secondary structure and size. Groups that had more than one protein were aligned using ClustalW (Larkin et al., 2007) to determine the sequence identity within the group. RESULTS
ForF was detected in the medium but no function of this protein could be determined. The accessory protein (ForF) from the for-operon was expressed in trans within different strains to observe the effect of the formosin secretion. First the ForF protein was complemented in trans to the strain where it had been deleted in the for-operon, resulting in strain Slds19. Result showed that the complementation of ForF did not restore the high level secretion of formosin (ForD) observed in strain SLds1 which expresses the intact for-operon (Figure 1A). In strain SLds172, ForF was complemented in a strain SLds1, a strain already expressing the intact for-operon thus having a high level of secretion of formosin. This resulted in a lower formosin secretion in strain SLds172 compared to SLds1 (Figure 1A). In strain SLds171, ForF was complemented into strain SLds22 to see if ForF could be involved in the stabilization of formosin. SLds22 contained a fusion protein of ForD-Ci2a where the fusion protein was secreted but formosin was degraded and only Ci2a could be detected in the medium (Chapter 5). Results show that complementation made in strain SLds171 did not stabilize formosin which still was undetectable in the medium (Figure 1A). In strain SLds170 ForF was complemented in trans to a strain that already contained ForE, ForF and ForG to observe the effect of ForF in the presence of the ABC transporter. In this strain a band of approximately 12-13kDa could be observed in the SDS-PAGE (Figure 1A). This band was observed in all the complementation strains containing the ForEG ABC transporter. The 12-13kDa band observed in the SDS-PAGE (Figure 1A) corresponds in size with the entire length of the ForF protein. If a signal peptide cleavage would occur (see later results) this is predicted to result in a 10kDa band which is the approximate size of formosin. In strain SLds170, formosin is not expressed and no detectable band was observed at 10kDa that would be indicative of a signal peptide cleavage of ForF. N-terminal sequencing of the 12-13kDa band from the SDS-PAGE was attempted using Edman degradation, but results were inconclusive due to the impurity of the sample.

68
Figure 1. SDS-PAGE and Western blotting analysis of the growth medium after two days of incubation of different Bacillus subtilis strains. Letters D, E, F, and G represent the presence of the genes forD, forE, forF, and forG. (A) Secretion analysis from ForF expressing strains where secretion profiles of formosin (ForD) and Ci2a is used to analyze the effect of ForF. When ForF was expressed in strains with the ForEG ABC transporter a band of 12-13kDa appears in the medium. (B) Secretion of formosin in strains containing different deletion sites of the ForF protein and a missense mutation in the ForG protein. Deletion site of ForF and a missense mutation in ForG change the levels of secreted formosin. In the for-operon the translation of the NBD protein (ForE) and the accessory protein (ForF) overlap with four nucleotides. Because of this the initial deletion of ForF made in strain SLds3 left two amino acids of the ForF protein followed by a stop codon. This deletion resulted in a decreased level of secreted formosin (ForD) compared to SLds1 containing the intact for-operon (Figure 1B). To test the effect of the deletion site of ForF, a new strain called SLds3new was created that still contained the first 13 amino acids of the ForF protein followed by a stopcodon. In SLds3new no formosin secretion could be detected after two days growth and it seemed that this deletion site completely disrupted the secretion of formosin (Figure 1B). In addition to these two strains, a third strain called SLds3mut was isolated during the initial creation of the SLds3 strain. This strain contained the same DNA sequence as the SLds3 strain with the exception of a missense mutant of the ForG protein V226L. Secretion of formosin in strain SLds3mut resulted in levels equal to that of the SLds1 strain which contained the intact for-operon (Figure 1B). The expression cassette in the pel-locus of SLds3mut and SLds3 was moved to new strains of B.subtilis JA1343, but the secretion level remained the same (results not shown) indicating that the increased secretion of formosin observed in SLds3mut was a result of the missense mutation. It is not known if this is because of an effect in mRNA stability or a functional improvement of ForG. However, the increase in formosin secretion observed for SLds3mut strain shows that ForF is not vital for the secretion of formosin.

69
Figure 2. N-terminal sequence of ForF with the prediction of the region being either a signal peptide (SP) or a membrane protein (MP), using different prediction programs. Prediction of a transmembrane (TM) region (h) was made using the prediction programs MEMSAT3, OCTOPUS, SCAMPI-msa, and THHMM. If topology prediction was available the result is shown in the figure as amino acids being either internal (i) or external (o). Programs used to predict the signal peptide(s) and the cleavage site were SPEPlip, SignalP-HMM, SignalP 4.0, and PrediSi. In addition programs such as SPOCTOPUS, SOSUIsignal, and Phobius that could distinguish between a signal peptide and a TM region were also used. ForF is predicted to be a single-pass membrane protein. Since the properties of a signal peptide and an N-terminal TM region is difficult to distinguish, a comparison of signal peptide and TM predictions is made. In all TM prediction programs ForF is predicted to be a membrane protein (Figure 2). In the signal peptide prediction three of the four programs predicted ForF to contain a signal peptide. However, scores of the signal peptide prediction were not considered high. In SignalP 4.0 the average calculated score was 0.33 and since the cutoff is 0.450 it was not predicted to be a signal peptide. In SignalP-HMM a different calculation method was used to predict presence of a signal peptide based on the presence of e.g. a hydrophobic region. SignalP-HMM predicted a signal peptide with a probability of 0.983, but had difficulty determining a site for the signal peptide cleavage and the site with the highest probability showing a probability of 0.333. The programs capable of distinguishing a TM region from a signal peptide all concluded that the N-terminal of ForF was a TM region and not a signal peptide. Results indicate that ForF has a single spanning membrane region, since programs capable of distinguishing between a signal peptide and a TM region predicted a TM protein. It was also observed that there was difficulty determining the signal peptide cleavage site, which might indicate that no cleavage site is present.

70
Figure 3. Comparison of genetic organization of the for-operon in B. licheniformis and the homologue region found in J. denitrificans. Genes are colored according to their properties indicating formosin or formosin-like protein as pink, transmembrane protein (TMD) as blue, ATP binding protein (NBD) as green, and accessory protein as grey. Homologue to the ForF accessory proteins indicates multimeric state of this accessory protein. Only a few homologues of ForF were identified and they were found in Bacillus, Geobacillus, and Jonesia bacteria. Bacillus homologues of ForF were located adjacent ABC transporter systems similar to ForEG ABC transporter. Geobacillus homologues were associated either with pseudogenes and ABC transporter related genes or in a larger cluster of ABC transporter genes of two NBD and three TMD-like proteins which probably constitute one or two complex heteromeric ABC transporters. These Geobacillus homologues showed that a ForF-like protein could be associated with other ABC transporter systems, but was not investigated further. However, two interesting homologues to the ForF accessory protein were identified in Jonesia denitrificans. These two homologues were observed to be neighboring genes adjacent to a partial formosin homologue and an ABC transporter (Figure 3). The first ForF homologue Jden_0039 was found to have 63% similarity to ForF, and the second ForF homologue Jden_0040 was found to have 76% similarity to ForF. These two ForF homologues were identified to be associated with the same ABC transporter. This could be indicating that ForF are active in a homomultimeric state. However, this is a prediction and no result from this study indicates that. Comparison of all three accessory proteins revealed that the N-terminal with its predicted TM region showed little sequence homology while several regions of the predicted cytoplasmic parts of the proteins were observed to be similar (Figure 4). Homologues to the other proteins encoded in the for-operon were also found in the region adjacent the ForF homologues. A partial 45 amino acid long formosin homologue Jden_0041 was identified and calculated to have 87% similarity to the last 45 amino acids of formosins C-terminal part. The Jden_0042 had 71% similarity to the TMD in the for-operon and the Jden_0043 had 81% similarity to the NBD protein in the for-operon (Appendix IV). The genes in the for-operon and the homologues observed in J. denitrificans all seem to have a high degree of conservation although the gene locations are quite different.

71
Figure 4. Multiple sequence alignment of the ForF protein from Bacillus licheniformis and the two homologues Jden_0040 and Jden_0039 from Jonesia denitrificans. The alignment was performed using ClustalW 2.1. Sequence similarities seen in blue are indicated as *(asterisk) representing a conserved amino-acid, a : (colon) representing a conservation between amino-acids of strongly similar properties, and a . (period) representing a conservation between amino-acids of weakly similar properties. The secondary structure of ForF was predicted using PSIPRED v3.0 and shown below the sequence alignment. The red area indicates the predicted transmembrane region of the three proteins. Four groups of accessory proteins are associated with the lactococcin 972 family. Different accessory proteins associated with the lactococcin 972 family was found and characterized using criteria described in the material and methods section. Many ABC transporters associated with the lactococcin 972 family did not have an accessory protein but in some cases accessory proteins were present. Accessory proteins that could be located were grouped according to size and secondary structure (Table 3). Four groups could be identified that were found to be small (<120 amino acids) or large (>200 amino acids) with a secondary structure of either β (Group 3 and 4) or a mixture of α+β (Group 1 and 2). Proteins found in group 2 had 98% identity but they are associated with proteins with lower sequence identity although the accessory protein showed high levels of identity. No homologues in either known proteins, protein families, or domains could be identified for group 1 (formosin accessory protein) or group 4. Group 2 were identified as bacterial extracellular solute-binding proteins, family 3, which are proteins associated with ABC transporter that aids in the transport selection towards polar residues (Tam and Saier, 1993). Group 3 were found to be similar to the YxeA protein from B. subtilis, which is a protein induced in the presence of a pore forming antimicrobial peptide, LL-37, and co-transcribed with an ABC transporter (Pietiäinen et al., 2005; Joseph et al., 2002). The YxeA protein family is small proteins (115 resides) associated with ABC transporters that are found exclusively in Gram positive bacteria (InterPro, accession number IPR006542). There was an observed diversity in Group 3 since three of the proteins had a small α-helical region in the C-terminal, while two of the proteins consisted of an extracellular structure of only β-strands. However, all five proteins were identified to belong to the YxeA family. Common features of these four groups are that they all are expressed within or in close proximity of the ABC transporter genes and all have an N-terminal TM region anchoring them to the cellular membrane. The distinction between an N-terminal TM region or signal peptide in group 3 was less obvious than for the other groups, but a comparison of the results concluded that it most likely was a TM region which contradicts previous prediction for YxeA (Joseph et al., 2004). However, it should be noted that they did this prediction
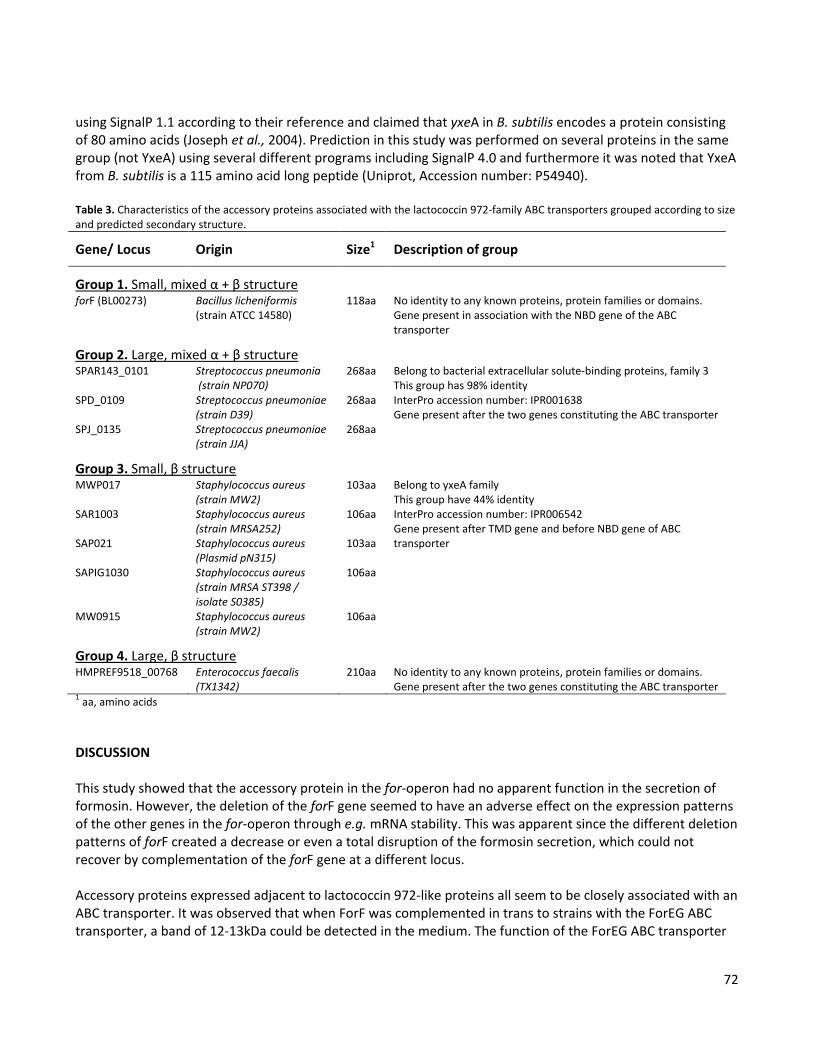
72
using SignalP 1.1 according to their reference and claimed that yxeA in B. subtilis encodes a protein consisting of 80 amino acids (Joseph et al., 2004). Prediction in this study was performed on several proteins in the same group (not YxeA) using several different programs including SignalP 4.0 and furthermore it was noted that YxeA from B. subtilis is a 115 amino acid long peptide (Uniprot, Accession number: P54940). Table 3. Characteristics of the accessory proteins associated with the lactococcin 972-family ABC transporters grouped according to size and predicted secondary structure.
Gene/ Locus Origin Size1 Description of group
Group 1. Small, mixed α + β structure forF (BL00273) Bacillus licheniformis
(strain ATCC 14580) 118aa No identity to any known proteins, protein families or domains.
Gene present in association with the NBD gene of the ABC transporter
Group 2. Large, mixed α + β structure SPAR143_0101 Streptococcus pneumonia
(strain NP070) 268aa Belong to bacterial extracellular solute-binding proteins, family 3
This group has 98% identity InterPro accession number: IPR001638 Gene present after the two genes constituting the ABC transporter
SPD_0109 Streptococcus pneumoniae (strain D39)
268aa
SPJ_0135 Streptococcus pneumoniae (strain JJA)
268aa
Group 3. Small, β structure MWP017 Staphylococcus aureus
(strain MW2) 103aa Belong to yxeA family
This group have 44% identity InterPro accession number: IPR006542 Gene present after TMD gene and before NBD gene of ABC transporter
SAR1003 Staphylococcus aureus (strain MRSA252)
106aa
SAP021 Staphylococcus aureus (Plasmid pN315)
103aa
SAPIG1030 Staphylococcus aureus (strain MRSA ST398 / isolate S0385)
106aa
MW0915 Staphylococcus aureus (strain MW2)
106aa
Group 4. Large, β structure HMPREF9518_00768 Enterococcus faecalis
(TX1342) 210aa No identity to any known proteins, protein families or domains.
Gene present after the two genes constituting the ABC transporter 1 aa, amino acids
DISCUSSION This study showed that the accessory protein in the for-operon had no apparent function in the secretion of formosin. However, the deletion of the forF gene seemed to have an adverse effect on the expression patterns of the other genes in the for-operon through e.g. mRNA stability. This was apparent since the different deletion patterns of forF created a decrease or even a total disruption of the formosin secretion, which could not recover by complementation of the forF gene at a different locus. Accessory proteins expressed adjacent to lactococcin 972-like proteins all seem to be closely associated with an ABC transporter. It was observed that when ForF was complemented in trans to strains with the ForEG ABC transporter, a band of 12-13kDa could be detected in the medium. The function of the ForEG ABC transporter

73
has been shown to be involved in the secretion of, and immunity to formosin (Chapter 5, Chapter 6). Due to these previous results the presence of the ForF protein in the medium is probably not caused by a secretion via the ForEG ABC transporter. Instead it could be speculated that the ForEG ABC transporter has a stabilizing effect for the ForF accessory protein. The size of ForF protein observed in the medium corresponds to the entire amino acid sequence of ForF subsequently indicating that there has been no signal peptide cleavage and that the TM region is still present. ForF observed in the medium could be an effect of an overexpression of ForF in the presence of the ForEG ABC transporter. However, it might also be hypothesized that this is an indication of the two states of the protein, one being in the medium and one where it is bound to the membrane. This has previously been observed for e.g. the immunity protein (NisI) associated with nisin where half of the expressed proteins are membrane bound and half is present in the medium (Koponen et al., 2004). However, NisI is an immunity protein attached to the membrane as a lipoprotein and not an accessory protein with an N-terminal TM region. In this study a comparison of different accessory proteins associated with the lactococcin 972 family was performed. Most ABC transporters did not have any accessory protein, and for the accessory protein found it was observed that there was a diversity regarding both secondary structure and size. The large α + β proteins were found to belong to the bacterial extracellular solute-binding proteins, family 3. These are described as proteins aiding in the import through ABC transporters with the affinity towards polar residues (Tam and Saier, 1993). The group of small β proteins was found to belong to the YxeA-family, but little is known about the function of this protein family. However, the YxeA-family of proteins might hypothetically be of great importance as YxeA have been implied in protection against cationic antimicrobial peptides not just from bacterial origin (Pietiäinen et al., 2005; Joseph et al., 2004). For the small α + β protein (ForF) and the large β protein no domains or similar proteins with known function could be detected. However, identification of two homologues to ForF indicates that the protein could be functioning as a homomultimeric protein. Due to the close proximity of the accessory protein genes to the ABC transporter genes, the functions of the accessory proteins seem to be linked to the ABC transporter. The function of the accessory proteins are exerted in the extracellular environment. The family 3 bacterial extracellular solute-binding proteins seem to have affinity towards polar amino acids aiding in import (Tam and Saier, 1993). The YxdLM ABC transporter and its accessory protein YxeA from B. subtilis are induced in the presence of the human antimicrobial peptide LL-37 and predicted to convey immunity to cationic antimicrobial peptides (Pietiäinen et al., 2005; Joseph et al., 2004). This might suggest that accessory proteins have a function in immunity by increase affinity to the bacteriocin for the ABC transporter. Alternatively, they might be involved in the secretion process having functions such as activation, maturation, and/or folding of the secreted bacteriocin. This theory is not supported by the results presented in this study of ForF, but another accessory protein (LncD) has been observed to be vital in the secretion of Lactococcin A (Franke et al., 1996; Varcamonti et al., 2001; Stoddard et al., 1992). Since there is an observed diversity in secondary structure it may be that the accessory proteins have different functions so it might be speculated that some work in the secretion process while others work in the immunity process associated ABC transporters. Moreover, some of the accessory proteins identified in this study might also be independently working immunity proteins that have been wrongfully annotated as accessory proteins. This is not the case for ForF since previous studies did not suggest an immunity property of ForF alone (Chapter 6). During this study it was difficult to ascertain if the results were due to a transcriptional effect or a protein effect. One example of this is the complementation of the accessory protein to the strain containing the intact for-operon in strain SLds172. This lowered the formosin secretion normally observed when expressing the

74
intact for-operon is strain SLds1. This could be a direct effect of the high concentrations of the ForF protein, but it might also be the effect of the presence of two identical, constitutive and strong promoters in the cell which could lower transcription initiation frequency or saturate the translation machinery. A second example is the difference in the forF deletion sites in SLds3 and SLds3new. The Slds3 were still active in the formosin secretion while SLds3new was not. This could be a due to an adverse effect on the mRNA. A third example is the increased formosin secretion of the ForG missence mutant in SLds3mut where a hydrophobic residue in a TM region was exchanged to a different hydrophobic residue (Valine226 to Leucine226). The absence of the formosin accessory protein (ForF) usually results in loss or decrease of formosin secretion, but this ForG missense mutant had a formosin secretion comparable to SLds1 which expressed the intact for-operon. While it seems more likely that these three examples are effects occurring on the transcriptional level it would be advisable to confirm this hypothesis by measuring the different levels of transcriptional units in the different strains through northern blotting analysis. Unfortunately this could not be done in the time span of this study. Alternatively, one might consider redesigning the strains and separately expressing the genes at different locus. These new strains would also provide the opportunity to use inducible or constitutive promoters in different combinations with the four different genes. While ABC transporters in general have been widely studied, the function of the accessory proteins is a research topic that needs further investigation. This is especially true with regards to the accessory proteins found in Gram positive bacteria. In these bacteria they are associated with protein or peptide interacting ABC transporters such as e.g. bacteriocin secreting ABC transporters (Young and Holland, 1999). They are also observed in association with ABC transporters linked to non-bacterially-produced antimicrobial peptides. One such system is the previously mentioned YxdLM ABC transporter and its accessory protein YxeA from B. subtilis which are predicted to convey immunity to cationic antimicrobial peptides (Pietiäinen et al., 2005; Joseph et al., 2004). Antimicrobial peptides are an important part of the innate immune system in humans while bacteriocins and peptides involved in quorum sensing have importance in the food and medical industry since e.g. these are regulating the microecological milieu. Because of this the research of the ABC transporters involved with these proteins or peptides is of medical and economical value. To fully understand and characterize these ABC transporters it is important to identify the function and significance of the diversities of the associated accessory proteins seen in Gram positive bacteria. Results and predictions made in this study have demonstrated the diversity and lack of knowledge surrounding the accessory proteins in Gram positive bacteria. Hopefully, this has signified the importance of future research into these accessory proteins such as determining their functions and the possible significance of the diversity in secondary structure.

75
CHAPTER 9 CONCLUDING REMARKS AND FUTURE PROSPECTS Introduction During the studies presented in this thesis, characteristics of the gene cluster required for functional expression of formosin were investigated, with a special regard to secretion and producer immunity. To better understand this process the thesis also included a partial characterization of formosin, although this was not the main aim of this PhD project. Presented here are the concluding remarks of this PhD thesis and the future prospects of these findings. Formosin This is a chromosomally encoded bacteriocin from Bacillus licheniformis ATCC 14580 situated in a genomic island with host defensive properties. The formosin gene forD is predicted to be arranged in an operon with three other genes, named forE, forF, and forG. This operon, referred to as the for-operon, contains the expression of formosin (ForD), an ATP binding protein (ForE), an unknown protein referred to as an accessory protein (ForF), and a transmembrane (TM) protein (ForG). Three promoters were predicted for the for-operon in this thesis but their function was never confirmed and should perhaps be viewed more as a guideline for further analysis than a result. (Chapter 4) Formosin is a pH tolerant but heat sensitive 9.6kDa bacteriocin that has a bacteriolytic effect on Bacillus subtilis (Chapter 4, Chapter 5). Formosin is predicted to have a sec-dependent signal peptide and a secondary structure consisting of β-strands (Chapter 4). This β-strand configuration could be associated with antimicrobial peptides such as the pore-forming defensins (Ganz, 2003). However, formosin is related to lactococcin 972 which is a non-pore-forming bacteriocin that inhibits septum formation in Lactococci through an interaction with lipid II (Martínez et al., 2008). No primary mode of action of formosin was determined in this thesis but it was observed that formosin has a secondary mode of action causing cell-lysis (Chapter 4). This secondary mode of action is not observed for lactococcin 972 (Martínez et al., 1999). Bacteriocins are of interest in different fields such as food and medical applications. In food applications they can be used in food preservation, start cultures for fermented food, or as an additive to animal feeds to promote a healthy gut flora. In medical applications they can be used as an alternative to antibiotics with a particular interest towards treatment of multidrug-resistant bacteria. Concerns about antibiotic-resistance and food safety has limited approval of new bacteriocins on the market while previously approved bacteriocins such as nisin can still be used. Possible applications of formosin on the market today are limited by these constrictions, but might have great prospects due to formosins sensitivity to heat. This heat sensitivity could be utilized to inactivate the bacteriocin, subsequently reducing the risk for developing antibiotic resistance and any other adverse effects of having an active antimicrobial protein present. However, prior to these

76
applications a more detailed study of formosin is needed such as mode of action, cell targets (e.g. lipid II), interference with antibiotic resistance, and cytotoxicity effects. Lactococcin 972 family In the current model of this protein family the bacteriocin is associated with an immunity protein with seven TM regions (InterPro accession number: IPR006540 and IPR006541). This model is based on the result of an expression study where a 2.5kb mRNA transcript containing the lactococcin 972 gene (lclA) and the immunity gene (lclB) were observed (Martínez et al., 1999). During this thesis an alternative model was proposed for this protein family which states that they are associated with an ABC transporter that has a characteristic topology of seven TM regions and two large extracellular domains. This new model is proposed since several lactococcin 972-like proteins, including lactococcin 972, were found to be associated with an ATP binding protein of an ABC transporter, in addition to the previously described TM immunity protein (Chapter 4). This proposed model contradicts results from the initial analysis of the lactococcin 972 gene expressions, since only genes encoding a TM protein (LclB) and lactococcin 972 (LclA) were observed in the mRNA transcript. However, in the investigation of lactococcin 972 performed in chapter 4, it was observed that an open reading frame predicted to be an ATP binding protein of an ABC transporter was situated two nucleotides downstream of the lclB gene. Moreover, it was also noted that the mRNA transcript analyzed by Martínez and colleagues was not 2.5kb as stated, instead it had the size of >2.9kb since it was observed to be larger than the 23S rRNA in Lactococcus lactis. (Chapter 4; Martínez et al., 1999) The ABC transporters associated with the lactococcin 972 protein family seemed to have a particular topology consisting of seven TM regions and two large extracellular domains, one in the N-terminal and one between the TM4 and TM5 regions (Chapter 7). For some of the lactococcin 972-like proteins an accessory protein was also associated with the ABC transporter. These accessory proteins seem quite diverse in structure, even though the ABC transporter is structurally conserved and the associated bacteriocin belongs to the same protein family (Chapter 8). Secretion mechanism of formosin During the study presented in Chapter 5 of this thesis it was shown that formosin contained two signal peptides. The first signal peptide is located in the N-terminal of the protein, which promotes secretion through one of the pre-existing secretory pathways in B. subtilis. The second signal peptide is located in the C-terminal of the protein, which promotes the secretion through the ForEG ABC transporter. The N-terminal signal peptide of formosin was demonstrated to be vital and cleaved off in both secretion pathways. However, only in the presence of the ForEG ABC transporter could formosin be detected in the medium. The N-terminal was predicted to be a sec-dependent signal peptide since it had an H-domain which is not present in signal peptides of ABC transporters. Moreover, it did not contain a lipobox as seen in lipoproteins, or a twin-arginine motif as seen in Tat-signal peptides (Tjalsma et al., 2004). This predicted sec-dependent signal peptide could be used to promote secretion of a reporter protein in high amounts without the presence of the ForEG ABC transporter. In fact, one could consider the N-terminal signal peptide of formosin to be a sec-dependent signal peptide with high efficiency due to the level of reporter protein secreted when this signal peptide is used. (Chapter 4, Chapter 5) During the test of the N-terminal signal peptide of formosin it became apparent that this was not the signal required for the secretion through the ForEG ABC transporter. Instead a second signal peptide was discovered in the C-terminal of formosin. This C-terminal signal peptide was demonstrated to promote secretion of a

77
reporter protein, but only in the presence of the ForEG ABC transporter. This supports the idea that there is a C-terminal signal peptide present in formosin. However, the secreted product observed in the western blotting analysis is 19-18kDa in size while the expected product should be 11kDa. This still indicates that there has been a secretion, although the band should be purified and identified through mass spectrometry since something has created a covalent bond to the secreted product. Furthermore, it would be prudent to attempt to secrete other proteins to confirm these results. (Chapter 5) Evidence of a C-terminal signal peptide was further validated as homologues to the ForEG transporter and the associated ForF accessory protein were found in Jonesia denitrificans. The ABC transporter and the accessory proteins of this homologous system were very similar to ForEFG, but the formosin homologue was different. This was a small 45 amino acid long protein that did not have an N-terminal signal, instead the formosin homologue had 87% similarity to the C-terminal part of formosin (Appendix IV). This lack of an N-terminal signal peptide and the homology to the C-terminal of formosin might be indicative of a C-terminal signal peptide within this formosin homologue. To further confirm the results of this study it would be of interest to test the secretion mechanism of this formosin homologue. (Chapter 8) Why are there two signal peptides in the same protein? Hypothetically this could serve as a defense mechanism since the secondary signal peptide in the N-terminal will result in degradation of formosin as observed in Chapter 5. If formosin was to become active in the absence of the ABC transporter it would start killing the producer as demonstrated in Chapter 6. Alternatively, the N-terminal signal peptide with its TM region could serve a purpose in the secretion process. It might ensure that the protein is not folding prior to secretion since folded substrates might inhibit secretion like observed for e.g. HasA (Debarbieux and Wandersman, 2001). But it might also be that the ABC transporter is using the TM region of the N-terminal signal peptide to insert formosin into the membrane where it is later processed by a signal peptidase. This would explain why the N-terminal signal peptide is cleaved off during secretion. Moreover, secretion through the ForEG ABC transporter was vital for formosin presence in the medium since secretion through the sec-pathway resulted in degradation of formosin (Chapter 5). This shows that the secretion through the ForEG ABC transporter was vital for maturation, which could be due to the directionality of the protein as it undergoes translocation. In the sec-pathway the translocation occurs N-terminal first, while translocation through the ForEG ABC transporter might occur C-terminal first. C-terminal signal peptides have previously been described to promote secretion through ABC transporters in Gram negative bacteria e.g. secretion of HlyA in Escherichia coli (Nicaud et al., 1986; Koronakis et al., 1989). The C-terminal signal peptide of formosin described in this thesis is the first C-terminal secretion signal observed in Gram positive bacteria. Formosin is observed to be a lactococcin 972-like bacteriocin with an N-terminal sec-dependent signal peptide and a C-terminal signal peptide. As a future prospect in understanding signal peptide differentiation and protein secreting ABC transporters in Gram positive bacteria, it would be interesting to find additional transport systems using C-terminal signal peptides. Even though a homologue to formosin was found without an N-terminal signal peptide one might still expect that proteins that have a C-terminal signal peptide have an N-terminal signal peptide as well. To find proteins with a potential C-terminal signal peptide one might look at proteins adjacent similar ABC transporter systems as the one observed in formosin, starting with the lactococcin 972 family. Lactococcin 972-like proteins are currently believed to have a sec-dependent signal peptide but are in this study found to be associated with an ABC transporter system which is a structural homolog to the formosin ABC transporter.

78
The ForEG ABC transporter ABC transporters have a typical architecture consisting of four domains, two transmembrane domains (TMDs) and two nucleotide binding domains (NBDs) (Wen and Tajkhorshid, 2011). This is believed to be the structure of the ForEG ABC transporter where the TMD (ForG) works as a homodimer together with two NBD (ForE) which provide the energy required for the function of this transporter. The ForE NBD was found to have conserved domains predicting this to be the NBD of an ABC exporter (Chapter 7). The topology of the TMD was predicted to consist of seven TM region and two extracellular domains (Chapter 7) which seemed to be structurally conserved for TMD associated with the lactococcin 972 family (Chapter 4; Chapter7). The ForEG ABC transporter was observed to have a dual function, being involved both in secretion of and immunity to formosin (Chapter 5; Chapter 6).The TMD (ForG) was found to be associated with uncleaved formosin within cells, predicting ForG to be the interaction partner to formosin in the secretion process (Chapter 5). The secretion through the ForEG ABC transporter is not promoted by the N-terminal signal peptide of formosin. Instead results from this thesis indicate that the secretion is promoted by the C-terminal part of formosin (Chapter 5). The ForEG ABC transporter was demonstrated to provide a two to four fold increase in immunity to formosin compared to a sensitive strain (Chapter 6). Even though this immunity increase seemed small, this was demonstrated to be enough to protect a formosin producer while a sensitive strain was lysed (Chapter 6). Functions of the ForEG ABC transporter demonstrated in this thesis and the conserved topology to other ABC transporters associated with lactococcin 972-like proteins makes this an interesting subject for future research. One topic of interest might be to investigate if this conserved topology relates to a C-terminal secretion mechanism among Gram positive bacteria. Another interesting research topic could be to investigate the mechanism behind the dual functions in secretion and immunity to see if e.g. the two large extracellular domains in the TMD can be coupled to one of these functions. Moreover, it might also be interesting to see how specific the immunity provided by these ABC transporters is, since these were observed to be structural homologues. The ForF accessory protein Characterization of the ForF accessory protein described in Chapter 8 predicted that this protein had an N-terminal TM domain. Most of the protein was situated outside of the cell with an α+β secondary structure. ForF has no apparent function in the secretion of formosin, but deletion of this protein has adverse effects on the expression of the other genes in the for-operon. When ForF is expressed in the presence of the ForEG ABC transporter this accessory protein is detected in the medium. This was predicted to be a result of a stabilization effect of the ForEG ABC transporter and a result of an overexpression. There are two studies that might aid in understanding ForF. One is to test if ForF increases immunity to formosin for the ForEG ABC transporter. Secondly, the subcellular location of ForF should be tested. Accessory proteins associated with bacteriocin ABC transporters in Gram positive bacteria have never been properly characterized to the same extent as they have been in other ABC transporter systems. To better characterize the ForF protein the study also included comparisons of other potential accessory proteins of ABC transporters associated with lactococcin 972-like proteins (Chapter 8). Although the ABC transporters of this bacteriocin family have been found to be structurally similar, an assortment of different accessory proteins were found. These different accessory proteins were assigned to four different groups based on size and secondary structure. Two of the four groups were identified to different protein families. One belonging to the YxeA family, where YxeA is a protein believed to provide immunity against cationic antimicrobial peptides

79
(Joseph et al., 2002). The other belonging to the bacterial extracellular solute-binding protein family 3, which is a protein family involved in the import of peptides with a selection towards polar residues (Tam and Saier, 1993). Identification of these two protein families seems to indicate an immunity function of these accessory proteins, where one group interacts on cationic bacteriocins and the other group interacts with polar residues. Accessory proteins of protein secreting or interacting ABC transporters in Gram positive bacteria are a subject where more research is needed. In similar ABC transporters they might be absent or present, and found in an assortment of different sizes and secondary structures as demonstrated in this thesis. Compared to the study of new bacteriocins or research into the protein interacting ABC transporters the accessory protein might seem less fascinating. However, research into new bacteriocins is usually done for possible applications or to understand the dynamics of the microflora in general. To understand the dynamics of the microflora or to produce this for applications in food or medical industries one needs to understand the secretion and immunity processes for either the producer or target bacteria. Both of which might be potential functions for the accessory proteins. Alternatively, research into these accessory proteins might be important in medical research into antimicrobial peptides involved in the innate immune system. Little is known about these accessory proteins in Gram positive bacteria and it is not known if they provide immunity and if so how specific this immunity is. One such example is the YxeA like proteins, since YxeA is believed to function in the immunity towards cationic peptides such as e.g. the human antimicrobial peptide LL-37 (Joseph et al., 2002).

80
REFERENCES Altena, K., Guder, A., Cramer, C., Bierbaum, G. 2000. Biosynthesis of the lantibiotic mersacidin: organization of a type B lantibiotic gene cluster. Appl. Environ. Microbiol. 66: 2565-2571 Altschul, S.F., Gish, W., Miller, W., Myers, E.W. & Lipman, D.J. 1990. Basic local alignment search tool. J. Mol. Biol. 215: 403-410 Anagnostopoulos, C., and J. Spizizen. 1961. Requirements for transformation of Bacillus subtilis. J Bacteriol. 81: 741–746. Anderssen, E.L., Diep, D.B., Nes, I.F., Eijsink, V.G.H., Nissen-Meyer, J. 1998. Antagonistic Activity of Lactobacillus plantarum C11: Two New Two-Peptide Bacteriocins, Plantaricins EF and JK, and the Induction Factor Plantaricin A. Appl. Environ. Microbiol. 64: 2269-2272 Baba, T., Schneewind, O. 1996. Target cell specificity of a bacteriocin molecule: a C-terminal signal directs lysostaphin to the cell wall of Staphylococcus aureus. EMBO J. 15: 4789–4797 Bárcena, J.M.B., Siñeriz, F., González de Llano, D., Rodríguez, A., Suárez, J.E. 1998. Chemostat Production of Plantaricin C by Lactobacillus plantarum LL441. Appl. Environ. Microbiol. 64: 3512-3514 Barefoot, S.F., Chen, Y-R., Hughes, T.A., Bodine, A.B., Shearer, M.Y., Hughes, M.D. 1994. Identification and Purification of a Protein That Induces Production of the Lactobacillus acidophilus Bacteriocin Lactacin B. Appl. Environ. Microbiol. 60: 3522-3528
Belasco, J.G., Chen, C-Y. A. 1988. Mechanism of puf mRNA degradation: the role of an intercistronic stem-loop structure. Gene. 72: 109-117 Belguesmia, Y., Madi, A., Sperandio, D., Merieau, A., Feuilloley, M., Prévost, H., Drider, D., Connil, N. 2011. Growing insights into the safety of bacteriocins: the case of enterocin S37. Res Microbiol.162: 159-163 Bernsel, A., Viklund, H., Falk, J., Lindahl, E., von Heijne, G.,Elofsson, A. 2008. Prediction of membrane-protein topology from first principles. Proc Nat Acad Sci USA 105:7177–7181 Bierbaum, G., Sahl, H-G. 1987. Autolytic system of Staphylococci simulans 22: influence of cationic peptides on activity of N-acetylmuramoyl-L-alanine amidase. J. Bacteriol. 169:5452–5458. Bierbaum, G., Sahl, H-G. 2009. Lantibiotics: mode of action, biosynthesis and bioengineering. Curr. Pharm. Biotechnol. 10: 2–18 Bjornvad, M. E., Jorgensen, P. L., Hansen, P. K. 7 May 2003. Homologous recombination into bacterium for the generation of polynucleotide libraries. World Intellectual Property Organization. WO 03/095658
Blanc-Potard, A.B., Solomon, F., Kayser, J., Groisman, E.A. 1999. The SPI-3 pathogenicity island of Salmonella enterica. J. Bacteriol. 3: 998-1004 Bradford, M.M. 1976. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72: 248–254

81
Braun, V., Pilsl, H., Groß, P. 1994. Colicins: structures, modes of action, transfer through membranes, and evolution. Arch. Microbiol. 161: 199-206
Buist, G., Ridder, A. N. J. A., Kok, J., Kuipers, O. P. 2006. Different subcellular locations of secretome components of Gram-positive bacteria. Microbiology. 152: 2867-2874 Campo, N., Tjalsma, H., Buist, G., Stepniak, D., Meijer, M., Veenhuis, M., Westermann, M., Müller, J. P., Bron, S., Jongbloed, J. D. H. 2004. Subcellular sites for bacterial protein export. Mol. Microbiol. 53: 1583-1599 Cascales, E., Buchanan, S.K., Duché, D., Kleanthous, C., Lloubès, R., Postle, K., Riley, M., Slatin, S., Cavard, D. 2007. Colicin biology. Microbiol. Mol. Biol. Rev. 71:158-229 Chung, Y.J., Steen, M.T., Hansen, J.N. 1992. The subtilin gene of Bacillus subtilis ATCC 6633 is encoded in an operon that contains a homolog of the hemolysin B transport protein. J. Bacteriol. 174: 1417-1422 Cole, K., Farnell, M.B., Donoghue, A.M., Stern, N.J., Svetoch, E.A., Eruslanov, B.N., Volodina, L.I., Kovalev, Y.N., Perelygin, V.V., Mitsevich, E.V., Mitsevich, I.P., Levchuk, V.P., Pokhilenko, V.D., Borzenkov, V.N., Svetoch, O.E., Kudryavtseva, T.Y., Reyes-Herrera, I., Blore, P.J., Solis de los Santos, F., Donoghue, D.J. 2006. Bacteriocins reduce Campylobacter colonization and alter gut morphology in turkey poults. Poult. Sci. 85: 1570-1575 Condon, C. 2003. RNA processing and degradation in Bacillus subtilis. Microbiol. Mol. Biol. Rev. 67: 157-174 Cotter, P. D., Hill, C., Ross, R. P. 2005. Bacteriocin: Developing innate immunity for food. Nat Rev Microbiol. 3: 777-788 Cotter, P.D., Hill, C., Ross, R.P. 2006. Author Reply: What's in a name? Class distinction for bacteriocins. Nat Rev Microbiol. 4 (doi:10.1038/nrmicro1273-c2) Davidson, A.L., Dassa, E., Orelle, C., Chen, J. 2008. Structure, function, and evolution of bacterial ATP-binding cassette systems. Microbiol. Mol. Biol. Rev. 72: 317-364 De Kwaadsteniet, M., Fraser, T., Van Reenen, C.A., Dicks, L.M.T. 2006. Bacteriocin T8, a Novel Class IIa sec-Dependent Bacteriocin Produced by Enterococcus faecium T8, Isolated from Vaginal Secretions of Children Infected with Human Immunodeficiency Virus. Appl. Environ. Microbiol. 72: 4761–4766 Debarbieux, L., Wandersman, C. 2001. Folded HasA inhibits its own secretion through its ABC exporter. EMBO J. 20: 4657-4663 Delves-Broughton, J. 2005. Nisin as a food preservative. Food Australia. 57: 525-527 de Hoon, M.J.L., Makita, Y., Nakai, K. Miyano, S. 2005. Prediction of Transcriptional Terminators in Bacillus subtilis and Related Species. PLoS Comput Biol. 1(e25): 212-221 de Vuyst, L. Vandamme, E.J. 1992. Influence of the carbon source on nisin production in Lactococcus lactis subsp. lactis batch fermentations. J. Gen. Microbiol. 138: 571-578 Diep, D.B., Axelsson, L., Grefsli, C., Nes, I.F. 2000. The synthesis of the bacteriocin sakacin A is a temperature-sensitive process regulated by a pheromone peptide through a three-component regulatory system. Microbiology. 146: 2155-2160 Diep, D.B., Skaugen, M., Salehian, Z., Holo, H., Nes, I.F. 2007. Common mechanisms of target cell recognition and immunity for class II bacteriocins. Proc. Natl. Acad. Sci. USA. 104:2384-2389 Drider, D., Fimland, G., Héchard, Y., McMullen L.M., Prévost, H. 2006. The continuing story of class IIa bacteriocins. Microbiol. Mol. Biol. Rev. 70 :564-582 Drosinos, E.H., Mataragas, M., Nasis, P., Galiotou, M., Metaxopoulos, J. 2005. Growth and bacteriocin production kinetics of Leuconostoc mesenteroides E131. J. Appl. Microbiol. 99: 1314-1323

82
Essential formulas Incorporated. 2011. [Online] http://www.essentialformulas.com/ [Accessed: May 12, 2012] Fariselli, P., Finocchiaro, G., Casadio, R. 2003. SPEPlip: the detection of signal peptide and lipoprotein cleavage sites. Bioinformatics. 19: 2498-2499 Farnham, P.J., Platt, T. 1981. Rho-independent termination: dyad symmetry in DNA causes RNA polymerase to pause during transcription in vitro. Nucleic. Acids. Res. 9: 563-577 Fink, A.L. 1999. Chaperone-Mediated Protein Folding. Physiol. Rev. 2: 425-449 Foulquié Moreno, M.R., Rea, M.C., Cogan, T.M., De Vuyst L. 2003. Applicability of a bacteriocin-producing Enterococcus faecium as a co-culture in Cheddar cheese manufacture. Int. J. Food Microbiol. 81: 73-84
Franke, C.M., Leenhouts, K.J., Haandrikman, A.J., Kok, J., Venema, G., Venema, K. 1996. Topology of LcnD, a protein implicated the transport of bacteriocins from Lactococcus lactis. J. Bacteriol. 6: 1766-1769 Gajic, O., Buist, G., Kojic, M., Topisirovic, L., Kuipers, O.P., Kok, J. 2003. Novel Mechanism of Bacteriocin Secretion and Immunity Carried Out by Lactococcal Multidrug Resistance Proteins. J. Biol. Chem. 278: 34291-3498 Galvin, M., Hill, C., Ross, R.P. 1999. Lacticin 3147 displays activity in buffer against gram-positive bacterial pathogens which appear insensitive in standard plate assays. Lett. Appl. Microbiol. 28: 355–358 Ganz, T. 2003. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 3: 710-720 Gasteiger, E., Hoogland, C., Gattiker, A., Duvaud, S., Wilkins, M.R., Appel, R.D., Bairoch, A. 2005. Protein Identification and Analysis Tools on the ExPASy Server. The Proteomics Protocols Handbook. pp. 571-607 Gautheret, D., Lambert, A. 2001. Direct RNA Motif Definition and Identification from Multiple Sequence Alignments using Secondary Structure Profiles. J. Mol. Biol. 313: 1003–1011 Gentschev, I., Mollenkopf, H., Sokolovic, Z., Hess, J., Kaufmann, S.H., Goebel, W. 1994. Development of antigen-delivery systems, based on the Escherichia coli hemolysin secretion pathway. Gene. 179:133-140
Gomi, M., Sonoyama, M., Mitaku, S. 2004. High performance system for signal peptide prediction: SOSUIsignal. Chem-Bio Info. J. 4: 142-147 Gratia, J. P. 2000. André Gratia: a forerunner in microbial and viral genetics. Genetics. 156: 471–476 Halkier, T., Clausen, I.B., Nielsen, L.K. 14 October 1993. A detergent containing a protease and a protease inhibitor and novel inhibitors for use therein. World Intellectual Property Organization. WO 93/20175.
Hanson, P.I., Whiteheart, S.W. 2005. AAA+ proteins: have engine, will work. Nat. Rev. Mol. Cell Biol. 6: 519-529 Harley, T. K., Djordjevic, G. M., Tseng, T-T., Saier, M. H. 2002. MicroCorrespondence: Membrane-fusion protein homologues in Gram-positive bacteria. Mol. Microbiol. 36: 516-517 Havarstein, L.S., Diep, D.B., Nes, I.F. 1995. A family of bacteriocin ABC transporters carry out proteolytic processing of their substrates concomitant with export. Mol. Microbiol. 16: 229-240 Héchard, Y., Sahl, H-G. 2002. Mode of action of modified and unmodified bacteriocins from Gram-positive bacteria. Biochimie 84: 545–557 Heng, N.C.K., Tagg, J.R. 2006a. Correspondence: What's in a name? Class distinction for bacteriocins. Nat Rev Microbiol. 4 (doi:10.1038/nrmicro1273-c1)

83
Heng, N.C.K., Wescombe, P.A., Burton, J.P., Jack, R.W., Tagg, J.R. 2006b. Bacteriocins: Ecology and Evolution. Berlin. Springer. Chapter 4, pp 45-92 Herranz, C., Driessen, A.J.M. 2005. Sec-Mediated Secretion of Bacteriocin Enterocin P by Lactococcus lactis. Appl. Environ. Microbiol. 71: 1959-1963. Higgins, C. F. 1992. ABC transporters: From Microorganisms to Man. Annu. Rev. Cell Biol. 8: 67-113 Hille, M., Kies, S., Gotz, F., Peschel, A. 2001. Dual role of GdmH in producer immunity and secretion of the staphylococcal lantibiotics gallidermin and epidermin. Appl. Environ. Microbiol. 67: 1380-1383 Hiller, K., Grote, A., Scheer, M., Münch, R., Jahn, D. 2004. PrediSi: prediction of signal peptides and their cleavage positions. Nucleic acids res. 32(Web Server issue):W375-379 Hoffmann, A., Pag, U., Wiedemann, I., Sahl, H-G. 2002. Combination of antibiotic mechanism in lantibiotics. Il Farmaco. 57: 685-691. Horton, R. M., Cai, Z. L., Ho, S. N., Pease, L. R. 1990. Gene splicing by overlap extension: tailor-made genes using the polymerase chain reaction. Biotechniques. 8: 528-535 Ikai, A.J. 1980. Thermostability and aliphatic index of globular proteins. J. Biochem. 88: 1895-1898 Ishii, T., Yoshida, K., Terai, G., Fujita, Y., Nakai, K. 2001. DBTBS: a database of Bacillus subtilis promoters and transcription factors. Nucleic Acids Res. 29:278-280 Jack, R.W., Tagg, J.R., Ray, B. 1995. Bacteriocins of Gram-Positive Bacteria. Microbiol. rev. 59: 171-200 Johnsen, L., Fimland, G., Mantzilas, D., Nissen-Meyer, J. 2004. Structure-function analysis of immunity proteins of pediocin-like bacteriocins: C-terminal parts of immunity proteins are involved in specific recognition of cognate bacteriocins. Appl. Environ. Microbiol. 70: 2647-2652 Jones, D. T. 1999. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 292: 195-202. Joseph, P., Guiseppi, A., Sorokin, A., Denizot, F. 2004. Characterization of the Bacillus subtilis YxdJ response regulator as the inducer of expression for the cognate ABC transporter YxdLM. Microbiology. 150: 2609-2617 Kenny, B., Haigh, R., Holland, I.B. 1991. Analysis of the haemolysin transport process through the secretion from Escherichia coli of PCM, CAT or beta-galactosidase fused to the Hly C-terminal signal domain. Mol. Microbiol. 5: 2557-2568
Kingsford, C., Ayanbule, K., Salzberg, S.L. 2007. Rapid, accurate, computational discovery of Rho-independent transcription terminators illuminates their relationship to DNA uptake. Genome Biol. 8: R22 Kjos, M., Borrero, J., Opsata, M., Birri, D.J., Holo, H., Cintas, L.M., Snipen, L., Hernández, P.E., Nes, I.F., Diep, D.B. 2011. Target recognition, resistance, immunity and genome mining of class II bacteriocins from Gram-positive bacteria. Microbiology. 157: 3256-3267 Kjos, M., Snipen, L., Salehian, Z., Nes, I.F., Diep, D.B. 2010. The Abi Proteins and Their Involvement in Bacteriocin Self-Immunity. J. Bacteriol. 192: 2068–2076 Klaenhammer, T.R. 1988. Bacteriocins of lactic acid bacteria. Biochimie. 70: 337-349 Klaenhammer, T.R. 1993. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 12: 39–85 Kleanthous, C. 2010. Swimming against the tide: progress and challenges in our understanding of colicin translocation. Nat. Rev. Microbiol. 8: 843-848

84
Kleerebezem, M. 2004. Quorum sensing control of lantibiotic production; nisin and subtilin autoregulate their own biosynthesis. Peptides. 25: 1405-1414 Kramer, N.E., Hasper, H.E., van den Bogaard, P.T., Morath, S., de Kruijff, B., Hartung, T., Smid, E.J., Breukink, E., Kok, J., Kuipers, O.P. 2008. Increased D-alanylation of lipoteichoic acid and a thickened septum are main determinants in the nisin resistance mechanism of Lactococcus lactis. Microbiology. 154: 1755-1762 Krogh, A., Larsson, B., von Heijne, G., Sonnhammer, E.L. 2001. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305: 567-580 Krogh, S., O’Reilly, M., Nolan, N., Devine, K.M. 1996. The phage-like element PBSX and part of the skin element, which are resident at different positions on the Bacillus subtilis chromosome, are highly homologous. Microbiology. 142: 2031–2040 Kuchler, K., Thorner, J. 1992. Secretion of peptides and proteins lacking hydrophobic signal sequences: the role of adenosine triphosphate-driven membrane translocators. Endocrine Reviews. 13: 499-514 Kumar, J.K. 2008. Lysostaphin: an antistaphylococcal agent. Appl. Microbiol. Biotechnol. 80: 555–561
Kunkel, B., Losick, R., Stragier, P. 1990. The Bacillus subtilis gene for the developmental transcription factor σK is generated by excision of a dispensable DNA element containing a sporulation recombinase gene. Genes Dev. 4: 525–535 Kuipers, O.P., Beerthuyzen, M.M., Deruyter, P.G.G.A., Luesink, E.J., de Vos, W.M. 1995. Autoregulation of nisin biosynthesis in Lactococcus lactis by signal-transduction. J. Biol. Chem. 270: 27299–27304 Koponen, O., Takala, T.M., Saarela, U., Qiao, M., Saris, P.E. 2004. Distribution of the NisI immunity protein and enhancement of nisin activity by the lipid-free NisI. FEMS Microbiol Lett. 231: 85-90 Koronakis, V., E. Koronakis, and C. Hughes. 1989. Isolation and analysis of the C-terminal signal directing export of Escherichia coli hemolysin protein across both bacterial membranes. EMBO Journal. 8:595-605 Kyte, J., Doolittle, R.F. 1982. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 157: 105-132
Käll, L., Krogh, A., Sonnhammer, E.L.L. 2004. A Combined transmembrane Topology and Signal Peptide Prediction Method. J. Mol. Biol. 338: 1027-1036 Käll, L., Krogh, A., Sonnhammer, E. L. L. 2007.Advantages of combined transmembrane topology and signal peptide prediction--the Phobius web server. Nucleic Acids Res. 35: W429-432 Langille, M.G., Brinkman, F.S. 2009. IslandViewer: an integrated interface for computational identification and visualization of genomic islands. Bioinformatics. 25: 664-665 Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A., McWilliam, H., Valentin, F., Wallace, I.M., Wilm, A., Lopez, R., Thompson, J.D., Gibson, T.J., Higgins, D.G. 2007. Clustal W and Clustal X version 2.0. Bioinformatics. 23: 2947-2948 Lemay, M-J., Choquette, J., Delaquis, P.J., Gariépy, C., Rodrigue, N., Saucier, L. 2002. Antimicrobial effect of natural preservatives in a cooked and acidified chicken meat model. Int J Food Microbiol. 78: 217-226 Limonet, M., Revol-Junelles, A.M., Millière, J.B. 2002. Variations in the membrane fatty acid composition of resistant or susceptible Leuconostoc or Weissella strains in the presence or absence of Mesenterocin 52A and Mesenterocin 52B produced by Leuconostoc mesenteroides subsp. mesenteroides FR52. Appl. Environ. Microbiol. 68: 2910-2916

85
Liu, L., Nakano, M.M., Lee, O.H., Zuber, P. 1996. Plasmid-amplified comS enhances genetic competence and suppresses sinR in Bacillus subtilis. J. Bacteriol. 178: 5144-5152 Looijesteijn, P.J., Boels, I.C., Kleerebezem, M., Hugoholtz, J. 1999. Regulation of Exopolysaccharide Production by Lactococcus lactis subsp. cremoris by the Sugar Source. Appl. Environ. Microbiol. 65: 5003–5008
Macke, T., Ecker, D., Gutell, R., Gautheret, D., Case, D.A., Sampath, R. 2001. RNAMotif – A new RNA secondary structure definition and discovery algorithm. Nucleic Acids Res. 29: 4724–4735 Maisnier-Patin, S., Richard, J. 1996. Cell wall changes in nisin-resistant variants of Listeria innocua grown in the presence of high nisin concentrations. FEMS Microbiol. Lett. 140: 29–35 Martínez-Cuesta, M.C., Kok, J., Herranz, E., Peláez, C., Requena, T., Buist, G. 2000. Requirement of Autolytic Activity for Bacteriocin-Induced Lysis. Appl. Environ. Microbiol. 66: 3174–3179. Martínez, B., Böttiger, T., Schneider, T., Rodríguez, A., Sahl, H-G., Wiedemann, I. 2008. Specific Interaction of the Unmodified Bacteriocin Lactococcin 972 with the Cell Wall Precursor Lipid II. Appl. Environ. Microbiol. 74: 4666-4670 Martínez, B., Fernández, M., Suárez, J. E., Rodríguez, A. 1999. Synthesis of lactococcin 972, a bacteriocin produced by Lactococcus lactis IPLA 972,depends on the expression of a plasmid encoded bicistronic operon. Microbiology. 145: 3155-3161 Martínez, B., Rodríguez, A., Suárez, J. E. 2000. Lactococcin 972, a bacteriocin that inhibits septum formation in lactococci. Microbiology. 146: 949-955 Martínez, B., Suárez, J. E., Rodríguez, A. 1996. Lactococcin 972, a homodimeric lactococcal bacteriocin whose primary target is not the plasma membrane. Microbiology. 14: 2393-2398. McAuliffe, O., O'Keeffe, T., Hill, C., Ross, R.P. 2001a. Regulation of immunity to the two-component lantibiotic, lacticin 3147, by the transcriptional repressor LtnR. Mol. Microbiol. 39: 982-993 McAuliffe, O., Ross, R.P., Hill, C. 2001b. Lantibiotics: structure, biosynthesis and mode of action. FEMS Microbiol. Rev. 25: 285-308 McAuliffe, O., Ryan, M.P., Ross, P., Hill, C., Breeuwer, P., Abee, T. 1998. Lacticin 3147, a Broad-Spectrum Bacteriocin Which Selectively Dissipates the Membrane Potential. Appl. Environ. Microbiol. 64: 439-445 Meile, J-C., Wu, L. J., Ehrlich, S. D., Errington, J., Noirot, P. 2006. Systematic localisation of proteins fused to the green fluorescent protein in Bacillus subtilis: Identification of new proteins at the DNA replication factory. Proteomics. 6: 2135-2146 Mikkelsen, K. M., Svendsen, A., Diderichsen, B., Clausen, G. 2 April 1992. Detergent containing protease and inhibitor and novel inhibitors for use therein. World Intellectual Property Organization. WO 92/05239.
Mitra, A., Kesarwani, A.K., Pal, D., Nagaraja, V. 2011. WebGeSTer DB—a transcription terminator database. Nucleic Acids Res. 39(Database Issue): 1-7 Münch, R., Hiller, K., Grote, A., Scheer, M., Klein, J., Schobert, M. & Jahn D. 2005. Virtual Footprint and PRODORIC: an integrative framework for regulon prediction in prokaryotes. Bioinformatics. 21: 4187-4189. Natale, P., Brüser, T., Driessen, A.J.M. 2008. Sec- and Tat-mediated protein secretion across the bacterial cytoplasmic membrane—Distinct translocases and mechanisms. Biochim. Biophys. Acta. 1778: 1735-1756
Nes, I. F., Diep, D. B., Håvarstein, L. S., Brurberg, M. B., Eijsink, V., Holo, H. 1996. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie Van Leeuwenhoek. 70: 113-128

86
Nes, I.F., Yoon, S-S., Diep, D.B. 2007. Ribosomally Synthesiszed Antimicrobial Peptides (Bacteriocins) in Lactic Acid Bacteria: A Review. Food Sci. Biotechnol. 16: 675-690 Nicaud, J-M., Mackman, N., Gray, L., Holland, I.B. 1986. The C-terminal, 23 kDa peptide of E. coli haemolysin 2001 contains all the information necessary for its secretion by the haemolysin (Hly) export machinery. FEBS Letters. 204: 331-335
Nielsen, H., Krogh, A. 1998. Prediction of signal peptides and signal anchors by a hidden Markov model. Proc. Int. Conf. Intell. Syst. Mol. Biol. (ISMB 6). pp. 122-130 Nikaido, H., Zgurskaya, H. I. 2001. AcrAB and Related Multidrug Efflux Pumps of Escherichia coli. J. Mol. Microbiol. Biotechnol. 3: 215-218 Nilsen, T., Nes, I.F., Holo, H. 2003. Enterolysin A, a Cell Wall-Degrading Bacteriocin from Enterococcus faecalis LMG 2333. Appl. Environ. Microbiol. 69: 2975–2984 Nilsson, I., Sääf, A., Whitley, P., Gafvelin, G., Waller, C., von Heijne, G. 1998. Proline-induced disruption of a transmembrane alpha-helix in its natural environment. J. Mol. Biol. 284: 1165-1175 Nissen-Meyer, J., Rogne, P., Oppegård, C., Haugen, H.S., Kristiansen, P.E. 2009. Structure-function relationships of the non-lanthionine-containing peptide (class II) bacteriocins produced by Gram-positive bacteria. Curr. Pharm. Biotechnol. 10: 10–37 Novacta News. November 2, 2011. Clinical Studies NVB302 [Online] http://www.novactabio.com./news.php?year=2011 [Accessed: May 12, 2012] Nugent, T., Jones, D.T. 2009. Transmembrane protein topology prediction using support vector machines. BMC Bioinformatics. 10: 159 Omori, K., Idei, A., Akatsuka, H. 2001. Serratia ATP-binding cassette protein exporter, Lip, recognizes a protein region upstream of the C terminus for specific secretion. J. Biol. Chem. 276: 27111-27119
Ortiz de Orué Lucana, D., Groves, M.R. 2009. The three-component signalling system HbpS-SenS-SenR as an example of a redox sensing pathway in bacteria. Amino Acids. 37: 479-86 Oswald, C., Holland, I.B., Schmitt, L. 2006. The motor domain of ABC-transporters: What can structure tell us? Naunyn-Schmiedeberg’s Arch. Pharmacol. 6: 385-399 Otto, M., Peschel, A. & Gotz, F. 1998. Producer self-protection against the lantibiotic epidermin by the ABC transporter EpiFEG of Staphylococcus epidermidis Tu3298. FEMS Microbiol. Lett. 166: 203–211 Paik, S.H., Chakicherla, A., Hansen, J.N. 1998. Identification and Characterization of the Structural and Transporter Genes for, and the Chemical and Biological Properties of, Sublancin 168, a Novel Lantibiotic Produced by Bacillus subtilis 168. J. Biol. Chem. 273: 23134-23142 Palva, I., Pettersson, R.F., Kalkkinen, N., Lehtovaara, P., Sarvas, M., Soderlund, H., Takkinen, K., Kaariainen, L. 1981. Nucleotide sequence of the promoter and NH2-terminal signal peptide region of the α-amylase gene from Bacillus amyloliquefaciens. Gene. 15: 43-51.
Pašić, L., Rodriguez-Mueller, B., Martin-Cuadrado, A-B., Mira, A., Rohwer, F., Rodriguez-Valera, F. 2009. Metagenomic islands of hyperhalophiles: the case of Salinibacter ruber. BMC Genomics. 10: 570 Petersen, T.N., Brunak, S., von Heijne, G., Nielsen, H. 2011. SignalP 4.0: discriminating signal peptides from Transmembrane regions. Nature Methods. 8: 785-786

87
Pietiäinen, M., Gardemeister, M., Mecklin, M., Leskelä, S., Sarvas, M., Kontinen, V.P. 2005. Cationic antimicrobial peptides elicit a complex stress response in Bacillus subtilis that involves ECF-type sigma factors and two-component signal transduction systems. Microbiology. 151: 1577-1592 Pohl, S., Harwood, C.R. 2010. Heterologous Protein Secretion by Bacillus Species: From the Cradle to the Grave. Adv. Appl. Microbiol.. 73: 1-25
Pongtharangkul, T., Demirci, A. 2004. Evaluation of agar diffusion bioassay for nisin quantification. Appl. Microbiol. Biotechnol. 65: 268-272 Price, M.N., Huang, K.H., Alm, E.J., Arkin, A.P. 2005. A novel method for accurate operon predictions in all sequenced prokaryotes. Nucleic Acids Res. 33: 880-892 Punta, M., Coggill, P.C., Eberhardt, R.Y., Mistry, J., Tate, J., Boursnell, C., Pang, N., Forslund, K., Ceric, G., Clements, J., Heger, A., Holm, L., Sonnhammer, E.L., Eddy, S.R., Bateman, A., Finn, R.D. 2012. The Pfam protein families database. Nucleic Acids Res. 40(D1): D290-D301
Quevillon, E., Silventoinen, V., Pillai, S., Harte, N., Mulder, N., Apweiler, R., Lopez, R. 2005. InterProScan: protein domains identifier. Nucleic Acids Res. 33: W116-W120 Rasmussen, M. D., Bjoernvad, M. E., Diers, I. 14 December 2000. Pectate lyase fusion for expression and secretion of polypeptides. World Intellectual Property Organization. WO 00/75344 Reddy, K.V., Yedery, R.D., Aranha, C. 2004. Antimicrobial peptides: premises and promises. Int. J. Antimicrob. Agents. 24: 536-547.
Rees, D. C. Johanson, E., Lewinson, O. 2009. ABC transporters: the power of change. Nat. Rev. Mol. Cell Biol. 10: 218-227 Riley, M.A., Wertz, J. E. 2002. Bacteriocins: Evolution, Ecology and Application. Ann. Rev. Microbiol. 56:117-37 Rogers, L.A., Whittier, E. D. 1928. Limiting factors in lactic fermentation. J. Bacteriol. 16: 211–229 Sablon, E., Contreras, B., Vandamme, E. 2000. New Products and New Areas of Bioprocess Engineering. Advances in Biochemical Engineering Biotechnology. Springer. Volume 68. Pages: 44-45 Sahl, H-G., Bierbaum, G. 1998. Lantibiotics: Biosynthesis and biological activities of uniquely modified peptides from Gram-positive bacteria. Annu. Rev. Microbiol. 52: 41-79 Sanger, F., Nicklen, S., Coulson, A.R. 1977.DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA.. 74: 5463-5467
Shine, J., Dalgarno, L. 1975. Determinant of cistron specificity in bacterial ribosomes. Nature. 254: 34-38 Simonen, M., Palva, I. 1993. Protein secretion in Bacillus species. Microbiol. Rev. 57: 109-137 Stein, T., Heinzmann, S., Düsterhus, S., Borchert, S., Entian, K-D. 2005. Expression and Functional Analysis of the Subtilin Immunity Genes spaIFEG in the Subtilin-Sensitive Host Bacillus subtilis MO1099. J. Bacteriol. 187: 822-828 Stoddard, G.W., Petzel, J.P., Belkum, J.K., McKay, J.L. 1992. Molecular Analyses of the Lactococcin A Gene Cluster from Lactococcus lactis subsp. lactis Biovar Diacetylactis WM4. Appl. Environ. Microbiol. 58: 1952-1961

88
Sugai, M., Fujiwara, T., Ohta, K., Komatsuzawa, H., Ohara, M., Suginaka, H. 1997. epr, which encodes glycylglycine endopeptidase resistance, is homologous to femAB and affects serine content of peptidoglycan cross bridges in Staphylococcus capitis and Staphylococcus aureus. J Bacteriol. 179: 4311–4318
Teixeira, V., Feio, M.J., Bastos, M. 2012. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid Res. 51: 149-177 The Clinical and Laboratory Standards Institute (formerly the National Committee for Clinical Laboratory Standards). 2011. Guideline M07–A8: Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically. + Guideline M26-A: Methods for Determining Bactericidal Activity of Antimicrobial Agents (http://www.nccls.org) The UniProt Consortium. 2012. Reorganizing the protein space at the Universal Protein Resource (UniProt). Nucleic Acids Res. 40: D71-D75.
Tagg, J.R., Dajani, A.S., Wannamaker, L.W. 1976. Bacteriocins of Gram-Positive bacteria. Bacteriol. rev. 40: 722-756 Tam, R., Saier, M.H. 1993. Structural, Functional, and Evolutionary Relationships among Extracellular Solute-Binding Receptors of Bacteria. Microbiol. Rev. 57: 320-346 Tjalsma, H., Antelmann, H., Jongbloed, J. D. H., Braun, P. G. Dormon, E., Dorenbos, R., Dubois, J-Y. F. Westers, H., Zanen, G., Quax, W.J., Kuipers, O. P. Baron, S., Hecker, M., van Dijl, J. M. 2004. The proteomics of protein secretion by Bacillus subtilis: separating the “secrets” of the secretome. Microbiol. Mol. Biol. Rev. 68: 207–233. Tjalsma, H., Bolhuis, A., Jongbloed, J.D., Bron, S., van Dijl, J.M. 2000. Signal peptide-dependent protein transport in Bacillus subtilis: A genome-based survey of the secretome. Microbiol. Mol. Biol. Rev. 64: 515–547 Torreblanca, M., Meseguer, I., Ventosa, A. 1994. Production of halocin is a practically universal feature of archael halophilic rods. Lett. Appl. Microbiol. 19: 201–205
Tran, L-S. Szabó, L. Ponyi, T., Orosz, L., Sík, T., Holczinger, A. 1999. Phage abortive infection of Bacillus licheniformis ATCC 9800;identication of the abiBL11 gene and localisation and sequencing of its promoter region. Appl. Microbiol. Biotechnol. 52: 845-852 Tramer, J., and Fowler, G.G. 1964. Estimation of nisin in foods. J. Sci. Food. Agric. 8: 522-528 Tusnády, G.E., Simon, I. 2001. The HMMTOP transmembrane topology prediction server. Bioinformatics. 17: 849-850 Vadyvaloo, V., Arous, S., Gravesen, A., Héchard, Y., Chauhan-Haubrock, R., Hastings, J.W., Rautenbach, M. 2004. Cell-surface alterations in class IIa bacteriocin-resistant Listeria monocytogenes strains. Microbiology. 150: 3025-3033 Vadyvaloo, V., Hastings, J.W., van der Merwe, M.J., Rautenbach, M. 2002. Membranes of class IIa bacteriocin-resistant Listeria monocytogenes cells contain increased levels of desaturated and short-acyl-chain phosphatidylglycerols. Appl. Environ. Microbiol. 68:5223-5230 Varcamonti, M., Nicastro, G., Venema, G., Kok, J. 2001. Proteins of the lactococcin A secretion system: lcnD encodes two in-frame proteins. FEMS Microbiol. Lett. 204: 259-263 Veith, B., Herzberg, C., Steckel, S., Feesche, J., Maurer, K.H., Ehrenreich, P., Bäumer, S., Henne, A., Liesegang, H., Merkl, R., Ehrenreich, A., Gottschalk, G.L. 2004. The complete genome sequence of Bacillus licheniformis DSM13, an organism with great industrial potential. J. Mol. Microbiol. Biotechnol. 7: 204–211
Viklund, H., Bernsel, A., Skwark, M., Elofsson, A. 2008a. SPOCTOPUS: A combined predictor of signal peptides and membrane protein topology. Bioinformatics. 24: 2928-2929

89
Viklund, H., Elofsson, A. 2008b. Improving topology prediction by two-track ANN-based preference scores and an extended topological grammar. Bioinformatics. 24: 1662-1668
Walker, J.E., Saraste, M., Runswick, M.J., Gay, N.J. 1982. Distantly related sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. EMBO J. 1: 945-951 Wang, B.Y., Kuramitsu, H.K. 2005. Interactions between oral bacteria: inhibition of Streptococcus mutans bacteriocin production by Streptococcus gordonii. Appl. Environ. Microbiol. 71: 354–362 Wen, P-C., Tajkhorshid, E. 2011. Conformational coupling of the nucleotide-binding and the transmembrane domains in ABC transporters. Biophys. J. 101: 680-690 Wescombe, P.A., Upton, M., Dierksen, K.P., Ragland, N.L., Sivabalan, S., Wirawan, R.E., Inglis, M.A., Moore, C.J., Walker, G.V., Chilcott, C.N., Jenkinson, H.F., Tagg JR. 2006. Production of the lantibiotic salivaricin A and its variants by oral streptococci and use of a specific induction assay to detect their presence in human saliva. Appl. Environ. Microbiol. 72: 1459-1466 Whitehead, H.R. 1933. A substance inhibiting bacterial growth, produced by certain strains of lactic streptococci. Biochem. J. 27: 1793–1800 Widner, B., Thomas, M., Sternberg, D., Lammon, D., Behr, R., Sloma, A. 2000. Development of marker-free strains of Bacillus subtilis capable of secreting high levels of industrial enzymes. J. Ind. Microbiol. Biotechnol. 25:204-212. Worobo, R.W., Van Belkum, M.J., Sailer, M., Roy, K.L., Vederas, J.C., Stiles, M.E. 1995. A signal peptide secretion-dependent bacteriocin from Carnobacterium divergens. J. Bacteriol. 177:3143–3149
Yohannan, S., Faham, S., Yang, D., Whitelegge, J.P., Bowie, J.U. 2004. The evolution of Transmembrane helix kinks and the structural diversity of G protein-coupled receptors. Proc. Nat. Acad. Sci. USA. 101: 959-963 Young, J., Holland, B.I. 1999. ABC transporters: bacterial exporters-revisited five years on. Biochim. Biophys. Acta. 1461: 177-200

90
APPENDIX I-IV APPENDIX I
Figure 1. The G+C content of the region 4135k-4180k bases in the chromosome of Bacillus licheniformis determined as the G+C content of every 1000bases. The grey line indicates the average G+C content of B. liceniformis (46.2%).
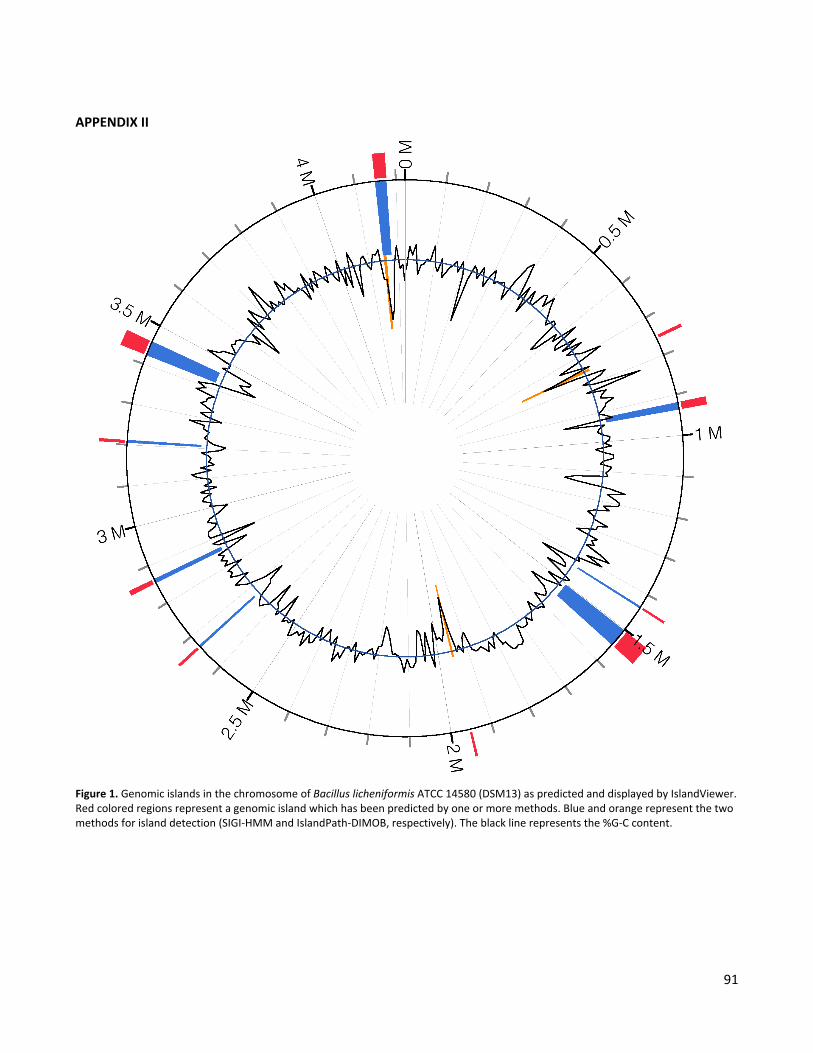
91
APPENDIX II
Figure 1. Genomic islands in the chromosome of Bacillus licheniformis ATCC 14580 (DSM13) as predicted and displayed by IslandViewer. Red colored regions represent a genomic island which has been predicted by one or more methods. Blue and orange represent the two methods for island detection (SIGI-HMM and IslandPath-DIMOB, respectively). The black line represents the %G-C content.

92
APPENDIX III
Figure 1. Sequence based topology prediction of the transmembrane protein, ForG. Four different methods of topology prediction were used as illustrated in the picture: OCTOPUS (blue), SCAMPI-msa (green), HMMTOP (red), and TMHMM (yellow). The transmembrane regions predicted for each program can be seen as a line connected with either weak lines of “O” and “I” which indicates if the loop between the transmembrane helixes is predicted to be outside (O) or inside (I) the cell. The sequence that all four programs agree on is a transmembrane region is indicated in the sequence as underlined and bold.

93
Figure 2. Multiple sequence based topology prediction of the transm
embrane protein, ForG
. The five proteins aligned in this figure is (from the top) ForG
from B. licheniform
is, an unknow
n protein (38% identity) from
Jonesia denitrificans, an unknown protein (36%
identity) from G
eobacillus termoglucosidasius, an unknow
n protein (29% identity) from
Bifidobacterium
longum, and a putative lantibiotic protection ABC transporter (18%
identity) from Parascardovia denticolens. The alignm
ent was perform
ed using ClustalW,
where *(asterisk) represents a conserved am
ino-acid, a : (colon) indicates conservation between am
ino-acids of strongly similar properties, and a . (period) represents
conservation between am
ino-acids of weakly sim
ilar properties. Prediction of transmem
brane regions of the five different proteins (seen with different colors) w
as made by the
topology prediction program M
EMSAT and the signal peptide prediction w
as made using Phobius. The predicted signal peptide is underlined in the sequence of each protein.

94
APPENDIX IV
Figure 1. Sequence alignment of formosin (BL00275) from Bacillus licheniformis seen in green and the formosin homologue Jden_0041 from Jonesia denitrificans seen in purple. The alignment was performed using ClustalW and similarities in the sequences was shown as *(asterisk) representing a conserved amino-acid, a : (colon) indicating conservation between amino-acids of strongly similar properties, and a . (period) representing conservation between amino-acids of weakly similar properties.