chapter 7 diet and gape size
TRANSCRIPT

C H A P T E R 7
Diet and Gape Size
I n this chapter, we review what is known about the diets of black carpMylopharyngodon piceus. We address diets of wild and captive populations, ontoge-netic shifts in diet, and related subjects. Because black carp are gape-limited preda-
tors, in the second part of this chapter, we consider mouth size and the implications ithas on the diet of this species.
DIET
Black carp are bottom-feeders that predominantly prey on snails and bivalve mollusks.In spite of inadequacies, a substantial volume of literature exists to support the con-clusion that the species is indeed a feeding specialist (i.e., molluscivore ormalacophagous) (Table 7.1; Appendix 2). For instance, Hickling (1971) remarked thatthe species is “a most formidable eater of mollusks.” Because of its heavy reliance onmollusks as food, Sokolov (1983, 2002) classified black carp as a stenophagous spe-cies. Indeed, their heavy pharyngeal arches and large molariform teeth clearly haveevolved to crush the shells of their prey. Similar to most fishes, larval and small juve-nile black carp feed almost entirely on small invertebrates (e.g., zooplankton and aquaticinsects). In contrast, larger juveniles and adults feed mostly on mollusks, a changeclosely associated with development of the lower pharyngeal teeth (fixed teeth) andsuperior crushing ability (IHAS 1976; Liu et al. 1990). Much of what we know aboutthe black carp diet is based on studies of black carp as potential biological controlagents of mollusks and on cultured populations. These subjects will be covered inmore detail in other chapters (e.g., Chapter 13). Also see subsequent section on blackcarp gape size.
85
chap7.p65 5/4/2005, 4:11 PM85

DIET AND GAPE SIZE
86
Table 7.1.—Mollusks consumed by black carp under natural and captive situations. The table onlyincludes references that report generic or species names of food item. Abbreviations are as follows:N = natural food (items consumed while foraging in natural and other outdoor water bodies, includ-ing ponds); S = supplemental or supplied food consumed in outdoor settings (items, sometimesalready crushed, provided by humans); L = items provided to fish held in laboratory settings (e.g.,experiments conducted with fish in aquaria); ? = uncertain. Taxon in brackets represents name ofmollusk as reported by author. See text and Appendix 2 for additional information.
Class/family Taxa Locality and food source References
BivalviaCardiidae Cardium Kazakhstan: Aral Sea [N] 4Corbiculidae Corbicula China: Chang River-lake [N?], West River-Lake 3, 8, 12, 14
Fuxian [N], no locality data [N?] [S].Corbicula fluminea China: pond [S], farm ponds [S], rearing 1, 5, 12
ponds [S], no locality data [S]Dreissenidae Dreissena former USSR: cooling reservoir station [S]; 4, 11
Kazakhstan: Aral Sea [N]Dreissena polymorpha Moldova: hydropower station-cooling ponds [S] 10
Mytilidae Mytilus China: Chang River-lake [N?] 8Unionidae Anodonta China: Chang River-lake [N?] 8
Anodonta woodiana China: pond [S] 1Cristaria plicata Russia: Amur River [N] 7
[Cristcaria plicata]Hyriopsis China: Chang River-lake [N?] 8
GastropodaAmpullariidae Lanistes USA or Egypt [L] 13
Pomacea sp. USA: Texas-pond? [N?] 18, 19Lymnaeidae Lymnaea China: Chang River-lake [N?]; USA or Egypt: [L] 8, 13
Lymnaea auricularia Israel: reservoir [N] 9Physidae Physa USA or Egypt: [L] 13
Physella acuta Israel: [L] 16Planorbidae Biomphalaria [Biomphilaris] USA or Egypt [L] 13
Bulinus USA or Egypt [L] 13Bulinus truncatus Israel: reservoir [N] 9Helisoma (probably Planorbella) USA or Egypt [L] 13Planorbella spp. USA: Louisiana [L] 17Planorbella trivolvis USA: Louisiana ponds [N?] 15
Thiaridae Semisulcospira cancellata Russia: Amur River [N] 2[Melania amurensis]
Melanoides tuberculatus Israel: reservoir [N]; 9, 16[Melania tuberculata] research institute [L]
Melanoides USA or Egypt: [L] 13Viviparidae Bellamya USA or Egypt: [L] 13
Margarya China: West River-Lake Fuxian [N] 14Viviparus Russia: Amur River [N] 2, 6Viviparus quadratus China: rearing ponds [S] 12Viviparus (Idiopoma) quadratus China: farm ponds [S] 5Viviparus (Sinotaia) quadratus China: pond [S] 1
References: 1 = Liu (1955); 2 = Nikolsky (1956); 3 = Lee (1957); 4 = Markova (1964); 5 = Chang (1966); 6 = Nikolsky(1971); 7 = Strelkov and Shul’man (1971, in Evtushenko et al. 1994); 8 = IHAS (1976: Table 64); 9 = Leventer (1979, 1981); 10= Krepis et al. (1981); 11 = Korenev (1984); 12 = NACA (1985); 13 = Shelton et al. (1995); 14 = Yang and Chen (1995); 15 =Venable et al. (2000); 16 = Ben-Ami and Heller (2001); 17 = Ledford (2003); 18 = J. Davis (Texas A&M University, personalcommunication 1995); 19 = Collins (1996).
chap7.p65 5/4/2005, 4:11 PM86

DIET AND GAPE SIZE
87
Literature Review
Although we have a basic understanding of black carp diets, much of what is writtenon the subject is descriptive, brief, lacking necessary detail, and therefore difficult toevaluate and interpret. For example, scant information is available about importantfactors that might increase our understanding of black carp trophic ecology. Mostauthors fail to mention the numbers or sizes of fish examined. Furthermore, sourcesoften provide little or no information about location (river system, sometimes coun-try) or habitat of populations studied. Consequently, it is not always possible to deter-mine if diet data are based on examination of specimens taken from a natural system(i.e., rivers and associated water bodies), artificial open-water habitats (e.g., canals andreservoirs), or farm ponds and holding tanks.
The common practice of providing supplemental food to captive or pond-rearedfish, particularly the use of natural items, has added to our difficulty in trying to accu-rately determine or describe the natural diets and foraging patterns of black carp. Inparticular, in China (and a few other countries), black carp held in captivity frequentlyare fed snails and bivalves; these mollusks are collected by humans in local waters(ditches and natural lakes) and transported to farms where they are used as supple-mental feed for the cultured fish (Lee 1957:60; Wu et al. 1964b; Hickling 1968:58;Bardach et al. 1972; IHAS 1976; Chang 1987). Moreover, foreign authors reporting ondiets of captive fish frequently do not state whether their information is based onobservation of black carp foraging on prey animals already present in a particularpond or, rather, on observations of black carp simply approaching and eating mol-lusks or other food items added to the pond as a supplement. The distinction is im-portant. For instance, a reader briefly reviewing the literature may get the impressionthat black carp commonly forage on zebra mussels Dreissena polymorpha in nature.However, our research suggests that such a conclusion may not necessarily be accu-rate due to the fact that whole or crushed zebra mussels frequently are supplied tocaptive or pond-reared fish and the information is published without adequate orcomplete explanation of the mussel preparation technique.
The size and health of black carp populations in the wild are likely influenced bythe composition, diversity, and abundance of molluscan fauna present, but publishedresearch is lacking. There appears to be no useful quantitative data available relatingblack carp numbers with the kinds or numbers of snails and bivalves present. In hispaper on lakes of the middle and lower Chang River, Liu (1984) did note that “suitablemolluscan food” for black carp was not very abundant in these habitats; consequently,he suggested that only a moderate number of black carp should be stocked.
In spite of the recognition given to black carp as a mollusk-feeding specialist,there are surprisingly few data on the types of mollusks that are preyed on by thefish. Prey items found in the stomachs of juvenile and adult black carp are rarelyidentified to lower taxonomic levels (i.e., genus, species, or even family). Conse-quently, the diversity of mollusks documented as being consumed by black carp is
chap7.p65 5/4/2005, 4:11 PM87

DIET AND GAPE SIZE
88
relatively small, especially when compared with the high diversity of aquatic mollusksthat often co-occur with black carp in the wild (this includes many systems whereblack carp are native as well as areas where they have been introduced). Given what wedo know about black carp and mollusks, it is highly probable that black carp in naturalsystems feed on a much broader diversity of snails and bivalves than the relativelyshort list of taxa presented in Table 7.1.
In addition to the above, most authors made no attempt to quantitatively describediets, rarely reporting frequency of occurrence or numbers of the different prey con-sumed. Related to this, there have been no experimental studies that adequately defineor evaluate black carp feeding preferences or selectivity. Some information does existregarding black carp as a biological control agent, based on studies on fish stocked inponds or held in aquaria (see Chapter 12). A few of these investigations have shedlight on possible black carp preferences and even prey defenses (e.g., snails that bur-row apparently are less likely to be consumed). Nevertheless, almost nothing is knownabout black carp feeding preferences in relation to different types of prey (e.g., snailsversus bivalves; hard-shelled versus thin-shelled species) or prey size in natural sys-tems. Without providing supporting evidence, several authors have implied some de-gree of feeding preference or selectivity by black carp. For instance, based on researchin the Amur River in Russia, Nikolsky (1954, 1956) reported that black carp feed ongastropod mollusks, chiefly Viviparus, crushing their shells with powerful pharyngealteeth. Chang (1966) and Gangstad (1980) reported that black carp prey mainly onsnails, noting that the fish also take other small mollusks. Roberts et al. (1973) statedthat black carp feed primarily on gastropods with coiled shells, but the authors pro-vided no details. In their book on the fishes of Lake Fuxian, Yang and Chen (1995)stated that adult black carp feed mainly on mollusks, such as the snail Margarya and theclam Corbicula.
Although mollusks are by far the main natural prey of black carp, an overview ofthe literature suggests a certain degree of flexibility in their foraging behavior. Lee(1957) reported that the favorite food of black carp includes snails, young mussels,and clams. Cheng and Zhou (1997) and Chen (1998) commented that black carpfeed primarily on snails, clams, and other mollusks (e.g., young freshwater mussels)and occasionally take shrimp and (aquatic) insects. They added that captive fish willeat wheat and beans. Ding (1994) stated that the natural food of black carp con-sisted of mollusks and also shrimp, crabs, and insect larvae, adding that juvenilesmainly consumed plankton. According to Ling (1977), the black carp “is a bottomfeeder, subsisting primarily on snails and clams, and sometimes on worms.” Atkinson(1977) indicated that it feeds on shell animals (snails, mollusks, crustaceans, etc.).Okada (1960) stated that black carp feed mostly on snails that live in small streams,pools, and lakes. He added that the fish sometimes take crustaceans. According toChu (1930), the black carp is a carnivorous fish, eating aquatic insects, snails, andshrimp. He reported that no plant material had been found in its stomach, but Chudid not mention the number of specimens examined. In likely reference to black
chap7.p65 5/4/2005, 4:11 PM88

DIET AND GAPE SIZE
89
carp larger than about 15 cm long, IHAS (1976) reported that juveniles feed prima-rily on mollusks such as clams, Corbicula, and spiral shells, although they found thatthe fish also take shrimp, crabs, and insect larvae. Gut contents of black carp takenfrom Honghu Lake in Hubei Province (Chang River basin) in 1959 were found toconsist almost entirely of mollusks. Identified items consisted of the algae Rivularia(42.8% frequency occurrence) and various mollusks, including Mytilus (28.6%), Cor-bicula (28.6%), Lymnaea (14.3%), Anodonta (14.3%), and Hyriopsis (14.3%) (IHAS 1976).
Nikolsky (1956) provided results from one of the few quantitative studies of blackcarp diet. Included in his report on fishes of the Amur River basin and black carp,Nikolsky analyzed the gut contents of seven black carp from the Yelabuga area. Hefound that the most important diet component were two types of snails, Viviparus(100% frequency of occurrence, average 23.3 snails per gut) and Melania amurensis (ajunior synonym of Semisulcospira cancellata) (85.7% frequency [i.e., in six of seven fish],9.1 snails per gut). Other items encountered in the gut of the fish were bivalve mol-lusks (28.7% [in two of the seven fish], 0.3 per gut [i.e., one in each]), one dragonflylarvae (14.3%), one chironomid larvae (14.3%), hundreds of bryozoan statoblasts(28.6% [in two fish]), and mollusk glochidia (28.6% [in two fish]). The glochidia werelikely ingested incidentally by black carp as the fish fed on adult mussels. Nikolsky alsoexamined the gut of two black carp caught in the Bolon area of the Amur River. Bothcontained only Viviparus, numbering 52 snails in one specimen and 76 in the other.Amur River fishermen reported that the intestines of black carp caught in the Bolonarea contained almost exclusively “pa�y a” (shells?). Similarly, Konstantinov (1950)inspected the digestive tract of a single black carp from the Amur River and found itto be filled completely with pieces of mollusk shells.
Based on a literature review, Evtushenko et al. (1994) reported that adult black carpprey most heavily on mollusks. The authors noted that bivalve mollusks recorded inadult diets included Cardium, Dreissena, and Cristaria plicata. They remarked that, inpond culture (presumably somewhere in the former Soviet Union), black carp havebeen found to take zebra mussel; for instance, 4-year-old black carp were reported asfeeding on an average of 1.4–1.8 kg of Dreissena per day (Evtushenko et al. 1994, andreferences therein). In his review of fishes of the Volga River, Poddubnyi (1979a)stated, without providing details, that black carp feed on Dreissena. Cristaria plicata (Leach1815) is a unionid species that superficially is very similar to many of our U.S. nativespecies (e.g., heelsplitters) and is characterized by having a very large, somewhat thinshell, shell length to 230 mm, height (with wing) 176 mm; it belongs to the subfamilyAnodontinae (Zhadin 1952).
In his discussion on fishes of the Saratov Reservoir (Volga River), Yermolin (1979)also noted that black carp consume mollusks, adding that, in the reservoir, the speciesfeeds in the same area as several other benthic fishes, including roach Rutilus rutilus,bream Abramis brama, and white bream Blicca bjoerkna. In a report by NACA (1985),there is a comment suggesting that black carp, because they feed on mollusks, preferclear water. Unfortunately, the English translation of that report is not definite.
chap7.p65 5/4/2005, 4:11 PM89

DIET AND GAPE SIZE
90
Response to Declines in Mollusk Abundance
In places where mollusks are few or absent, black carp are known to forage on otheritems. Most information on the subject is based on study of pond-reared fish. Innature, black carp probably migrate to other areas, if possible, as a response to adecline in mollusks. Savina (1975), based on a study of black carp raised in ponds inMoldovia, noted that black carp began feeding on macrophytes and insects whenmollusks were limited. However, he noted that, in such instances, growth rates ofblack carp declined. Leventer (1979, 1981), describing the feeding behaviors of blackcarp and common carp Cyprinus carpio stocked as biological control agents in reser-voirs in Israel, noted that both species search for food on the reservoir bottom. Hefound that the two fishes switched to feeding on insect larvae and shrimps once snailpopulations decreased to some low point that the authors did not define. Fish farmersin the United States reported that black carp can be raised on commercial feeds with-out mollusks as a supplement. Nevertheless, Chinese researchers commonly providewhole and crushed snails and bivalves to captive black carp and often consider suchnatural items as indispensable to successful production (see, for example, NACA 1985).
Captive Fish
While other bottom-dwelling cyprinids, such as common carp, feed on detritus and amixture of small benthic organisms, Chinese fish culturists have long recognized thatblack carp prefer mollusks. Thus, the Chinese have traditionally provided pond-rearedblack carp with snails and bivalves, even though the species reportedly requires littlesupplemental feeding under the right conditions (Hora 1953; Lin 1955; Lee 1957;Bardach et al. 1972; NACA 1985). Hora (1953) commented that small snails are ex-pressly collected in Jiangsu and Zhejiang provinces for feeding black carp. Lin (1955)noted that pond snails as food for black carp are easily obtained from nearby canalsand rivers. Similarly, Hora and Pillay (1962) commented that black carp raised in pondswere often fed snails collected from streams and lakes. Lee (1957:52) listed commoncommercial feeds available in China for culture of fingerlings. Included in his list aresnails, clams, Corbicula spp., as well as other natural and artificial foods.
According to Lee (1957:60), because of the abundance of snails, clams, and Cor-bicula spp. in some regions, such as Wu-shi and Soo-chow of Jiangsu Province and LinLake of Zhejiang Province, local fish farmers favor culture of black carp. Echoing Lee(1957), Wu et al. (1964b) stated that there are many streams in Jiangsu Province andZhejiang Province that have a great production of snails. In fact, some Chinese fish-ermen reportedly specialize in gathering snails, particularly in regions where there isalso a great amount of rearing black carp, especially in the village of Beizhuangji inSuzhou City (Wu et al. 1964b). According to the IHAS (1976), growth of captive blackcarp fingerlings reared in the Chinese provinces of Jiangsu and Zhejiang is enhancedby providing crushed spiral shells and clams supplemented with bean cake and silk-worm chrysalis. Hickling (1968:58) reported that black carp are stocked to feed on
chap7.p65 5/4/2005, 4:11 PM90

DIET AND GAPE SIZE
91
snails, which are common in well-fertilized farm ponds. He added that wild snails andclams are collected by the basketfull in ditches for use as supplementary feed. Atkinson(1977) briefly reported on experimental studies in a Chinese fish farm near Shanghaiwhere freshwater clams were being reared for freshwater pearl production and tosupply food for black carp and other fishes. Network of Aquaculture Centres in Asia(NACA 1985) stated that black carp and grass carp Ctenopharyngodon idella, under cul-ture conditions, both tend to be more or less omnivorous, feeding on oil seed cakes,brans, dregs, and animal feeds such as silkworm pupae, earthworms, and animal en-trails (see Appendix 2 for listing of other food items given to captive black carp).Chen (1934) reported that, when kept in an aquarium, black carp refuse to eat. Never-theless, there have been several recent studies of black carp held in aquaria whereregular feeding occurred (e.g., Ledford 2003).
Early Life Stages and Ontogenetic Shifts in Diet
As previously noted, larval and small juvenile black carp feed mostly on zooplanktonand aquatic insects, later shifting to a diet of mollusks as they grow and their pharyn-geal teeth become more developed. As discussed in Chapter 3, several authors havestudied the developmental stages of black carp pharyngeal teeth, noting that the teethare formed during postlarval development and grow on pharyngeal bones on thelower part of the posterior pharyngeal cavity (Liu et al. 1990; Yue and Nakajima 1994,1995; Nakajima and Yue 1995). Detailed descriptions of the diet of early life stagesand subsequent ontogenetic changes in feeding have been provided by various Sovietand Chinese scientists, including Motenkova (1970, 1972), Savina (1975), IHAS (1976),Liu et al. (1990), and others. All or most of these investigations appear to be basedentirely on examination of captive fish reared in ponds or other artificial settings. Ingeneral, larval and small juvenile black carp have a diet that is basically identical to thatof the larvae and early juveniles of the other Chinese carp species (Chang 1966;Gangstad 1980).
Of particular note is the research of Liu et al. (1990), who analyzed changes inpharyngeal apparatus development associated with changes in diet. They indicatedthat black carp shift to a diet of mollusks at 3–33 cm long, whereas IHAS (1976)remarked that the change occurred when the fish were about 15 cm long. We suspectthat the actual change varies, depending on developmental rate of fish and the typesand abundance of prey available.
Soviet Studies.—Motenkova (1970, 1972) studied the diets of larval black carp raised inponds in southern Russia, analyzing the data by grouping larval sizes based on the differ-ent developmental stages described in Sukhanova (1967). Evtushenko et al. (1994) re-viewed and summarized some of the literature, including that of Motenkova. Evtushenkoet al. (1994) indicated that at the later stages of development (probably late larvalstage), food items included cladocerans, copepods, other crustaceans, larval chirono-mids and other insect larvae, aquatic hemipterans, statoblasts of bryozoans, crayfish
chap7.p65 5/4/2005, 4:11 PM91

DIET AND GAPE SIZE
92
shells, rotifers, mollusks, and food pellets. The major food items of young of the year inponds are chironomid larvae. The diets of 2-year-old black carp included mollusks,oligochaetes, chironomids, aquatic macrophytes, larval insects, and zooplankton.
During the period 1972–1973, Savina (1975) examined the diets of young black carpraised in ponds at an aquaculture facility in what is now the Republic of Moldova. Henoted that first-year fish fed mainly on zooplankton, later switching to bottom zoo-plankton and benthos. By their second summer, the fish preyed mainly on mollusks.
Evtushenko et al. (1994), in their review of black carp biology, summarized thenatural and artificial diets of black carp at different ages and sizes. During the larvalstage of development, black carp feed mainly on microcrustaceans and rotifers. Earlystage larvae take mainly copepod nauplii and cladocerans, especially Moina and Bosmina.Middle stage larvae (7–12 d old) prey on copepods, cladocerans (Cladocera), and largerrotifers such as Asplanchna and Branchionus. Chironomid larvae are also taken. Latestage black carp larvae feed heavily on ostracods and aquatic insects, mainly chirono-mid larvae, odonate nymphs, and ephemeroptera larvae. Black carp larvae also occa-sionally take phytoplankton.
Chinese Studies.—According to Chang (1966) and Gangstad (1980), smaller blackcarp feed on various benthic organisms such as dragonfly nymphs and other insectlarvae, gonospores of bryozoa, and even mollusk larvae. The authors also provided abrief summary of changes in the diets of young black carp and grass carp, underabout 17 mm long.
Also based on research in China, Bardach et al. (1972:98) summarized the naturalfoods of black carp fry as follows: at size range of 7–9 mm, fry feed on protozoa,rotifers, and nauplii; at 10–12 mm, the diet includes protozoa, rotifers, nauplii, plussmall daphnids and Cyclops; at 13–17 mm, the diet consists of large daphnids andminute benthic animals; at 18–23 mm, they feed on large daphnids, minute benthicanimals, plus organic detritus; and at 24–30 mm, the fry take minute benthic animals,insect larvae, and organic detritus. The IHAS (1976) also stated that the major food ofblack carp larvae and fingerlings is zooplankton, adding that juveniles of about 15 cmlong begin to feed on small spiral shells and clams (a change that they associated withdevelopment of the lower pharyngeal teeth and the enhanced crushing ability).
Liu et al. (1990) recognized three stages in the development of black carp pharyn-geal teeth and associated each with a particular type diet. Black carp 6.7–7.8 mm totallength (TL) have “original teeth” and are incapable of masticating food. During thisstage, they feed mainly on zooplankton. Black carp measuring 8.9–25 mm TL have“transitional teeth” and the pharyngeal apparatus has only a weak ability to masticatefood and is not capable of grinding. During this stage, the diet of juvenile black carpshifts from zooplankton to benthic invertebrates (e.g., chironomid larvae). Black carp31–330 mm TL have “fixed teeth” and a pharyngeal apparatus capable of masticatingand grinding benthic animals with shells, such as snails. For additional details on thestudy by Liu et al. (1990), and others, refer to the section on pharyngeal apparatusappearing in Chapter 4.
chap7.p65 5/4/2005, 4:11 PM92

DIET AND GAPE SIZE
93
Wu et al. (1992) examined the diel feeding pattern of fingerling black carp. Theresearch is published in Chinese and awaits translation.
Artificial Foods and Nutritional Information
Although black carp are largely carnivorous, black carp fry reared in captivity readilyeat bean cake, grain residue from alcohol fermentation, and other similar plant food(Chang 1966; Gangstad 1980). Bardach et al. (1972:96) also reported that captive blackcarp fry in China are given barley, bean cake, and small snails as food for purposes ofrearing and growth. To keep at least a few fish of all sizes available for sale and to meetdemands for small fish, some Asian fry farmers intentionally attempt to slow growthof black carp fry and other Chinese carp. Maintenance in a stunted condition, withoutcausing malnutrition, is accomplished by using limited amounts (0.5–2.0% of weightof fish/d) of artificial feed, including rice bran, soybean milk, and peanut cake oil(Bardach et al. 1972:95–96). Hora and Pillay (1962) provided information on feedingschedules for black carp. Foods listed included mussels, barley, and bean cake. Theynoted that small quantities of artificial food can be given to black carp by the middleof April, but intensive feeding is conducted from May to October when water tem-perature is about 20°C. Hora and Pillay commented that black carp can be given onlymussels and snails, foods of low sustenance ratio, in May and June when it is relativelycold. They added that the fish need to be given a diet consisting of half vegetable andhalf animal foods toward the end of the year.
There is little information on the digestive physiology of black carp. In a compara-tive study of activities of hepatic alcohol dehydrogenase (ADH), Hseu and Ting (1997)found that black carp had the highest hepatic ADH activity among 10 fishes tested.Results were inconclusive but suggested that hepatic ADH activity of fishes, unlikemammals, might not parallel diets. Nevertheless, high ADH activity in black carp maybe related to its carnivorous diet. Wang and Song (1984) reported that the digestiverate of protein in black carp fingerlings is affected by the quantity of carbohydratecontents in the feed.
For additional information on nutrition of black carp, see the section entitled Fac-tors Influencing Growth Rate in Chapter 6.
GAPE SIZE
Although the soft bodies of mollusks are a nutritious source of food, each is pro-tected by a shell that forms a relatively hard and indigestible barrier. Fishes that preyon mollusks employ a variety of methods to exploit these invertebrates as food (Fryerand Iles 1972). Cyprinids do not have jaw teeth and, therefore, must rely on theirpharyngeal or throat teeth and chewing pad to crush hard items (see section on pha-ryngeal apparatus in Chapter 4). Although their mouth is somewhat protruscible, blackcarp are apparently incapable of sucking out soft tissues of their prey. Black carp usetheir pharyngeal mill to crush the hard shells and expose the soft parts for digestion.
chap7.p65 5/4/2005, 4:11 PM93

DIET AND GAPE SIZE
94
The pharyngeal apparatus of black carpis particularly massive, accounting for theirability to specialize on snails and bivalvemollusks as food (Figures 1.4 and 4.2–4.4).The black carp is a gape-limited predatorbecause its mouth diameter, or gape, es-sentially determines the maximum size ofmollusks it can consume (Figures 4.3 and7.1; see Zaret 1980). Because they aregape-limited predators, any hard items thatblack carp feed upon would, by necessity,have to be smaller than the fish’s mouthwidth. Nevertheless, gape size is relatedto body size and increases as the fishgrows. Therefore, larger fish have largermouths capable of exploiting a broader range of mollusk sizes. Information on gapewidth is critical to better understand potential impacts black carp might have on nativemolluscan faunas.
The actual morphological feature of the predator that limits maximum prey sizemay vary across taxa. For instance, within sunfishes, the distance between the cleithralbones has been used as a determining characteristic (Lawrence 1958; Wainwright 1996).Based on his investigation of black carp, W. Shelton (University of Oklahoma, per-sonal communication 2002) has noted that the pharyngeal arches of black carp areproportionally much larger than that of a similarly sized grass carp. Nevertheless, themouth of the black carp is relatively small (Shelton et al. 1995). If a black carp is ableto engulf a large mollusk, the prey may still be too large to be consumed as the fish’soverall throat dimensions (i.e., cleithral distance and space between the pharyngealarches) are slightly smaller than that of mouth gape. Black carp may be able to over-come this limitation by manipulating larger prey. Shelton et al. (1995) observed blackcarp initially engulfing and holding large snails within the buccal-pharyngeal cavitywhile using pharyngeal teeth to chip away at the edges of the prey’s shell. Given theabove information, actual gape size might be expected to be a reliable estimator of thelargest size mollusk that a particular black carp can ingest, crush, and swallow (Sheltonet al. 1995).
Shelton et al. (1995) conducted their studies in laboratories in the United States andEgypt, testing various aspects of black carp feeding biology as a basis for predictingtheir potential as biological control agents. They approached the study first from thestandpoint of physical limitations to engulfing mollusks by examining prey sizes inrelation to mouth capacity of various-sized fish. Because black carp crush their foodbefore swallowing, Shelton et al. (1995) also recognized that the limiting factor couldbe prey size relative to pharyngeal gape as well as the force necessary for shell crush-ing. They conducted feeding experiments with various species and sizes of snails (Physa
Figure 7.1.—Black carp with mouth agape.(Photograph by Kaoru Moteki.)
chap7.p65 5/4/2005, 4:11 PM94
DIET AND GAPE SIZE
95
spp., Heliosoma spp., Bellamya, Melanoides spp., Lanistes, Lymnaea spp., Bulinus, andBiomphalaria spp.) and demonstrated that mouth gape was a reliable estimator of thelargest size snail that a particular fish could consume. To calculate gape size, the re-searchers measured mouth circumference and mouth width of 80 black carp (120 toabout 500 mm TL). Results, including a graph of TL versus mouth gape (GW), indi-cated that the relationship for the size range of fish tested was linear (GW = 0.0471TL + 1.60) and highly correlated (r = 0.89; r 2 = 0.79). Fish between 100 and 500 mmhad gapes from 7 to 25 mm, and consumed snails measuring 7–17 mm.
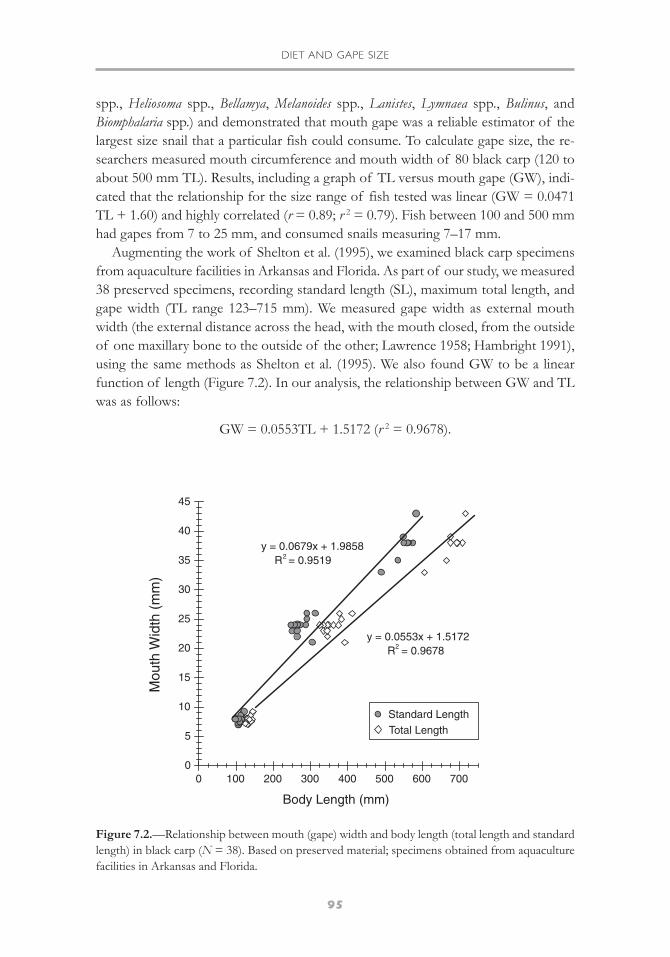
Augmenting the work of Shelton et al. (1995), we examined black carp specimensfrom aquaculture facilities in Arkansas and Florida. As part of our study, we measured38 preserved specimens, recording standard length (SL), maximum total length, andgape width (TL range 123–715 mm). We measured gape width as external mouthwidth (the external distance across the head, with the mouth closed, from the outsideof one maxillary bone to the outside of the other; Lawrence 1958; Hambright 1991),using the same methods as Shelton et al. (1995). We also found GW to be a linearfunction of length (Figure 7.2). In our analysis, the relationship between GW and TLwas as follows:
GW = 0.0553TL + 1.5172 (r 2 = 0.9678).
0 100 200 300 400 500 600 7000
5
10
15
20
25
30
35
40
45
Mou
th W
idth
(m
m)
Body Length (mm)
Total Length
Standard Length
y = 0.0553x + 1.5172
y = 0.0679x + 1.98582R = 0.9519
R = 0.96782
Figure 7.2.—Relationship between mouth (gape) width and body length (total length and standardlength) in black carp (N = 38). Based on preserved material; specimens obtained from aquaculturefacilities in Arkansas and Florida.
chap7.p65 5/4/2005, 4:11 PM95

DIET AND GAPE SIZE
96
The relationship between GW and SL was
GW = 0.0679SL + 1.9858 (r 2 = 0.9519).
Our linear trendline results differed slightly from those of Shelton et al. (1995).These differences might be the result of various factors, for instance, use of fresh orlive versus preserved material, differences in fish health and condition, or slight differ-ences in techniques used to measure the gape. It should be noted that our sample sizewas smaller (N = 38) than that of Shelton and his colleagues (N = 80), but it includeda series of eight specimens (605–715 mm TL) that were larger than the largest fish(about 50 cm TL) measured in the earlier study.
One possible use of the resulting data is to extrapolate the approximate gape sizeof much larger black carp. The species is reported to attain lengths of over 1.5 m andmore than 70 kg. Based on our equation, a black carp 1 m long (TL) would have a gapesize of about 57 mm and a fish 1.5 m long would have a gape of approximately 84mm. Using the equation of Shelton et al. (1995), the estimated gape sizes for thesesame-sized fish would be slightly smaller (49 and 72 mm wide, respectively). Never-theless, results indicate that larger black carp would likely be able to consume all butthe largest of native mussels and snails, assuming that their pharyngeal apparatuscould exert the crushing force necessary to break the heavier shells. Further investiga-tion is needed to determine if the linear model is appropriate for extrapolating tosignificantly larger fish. The Japanese angler K. Moteki (Black Carp Aouo Club, Tokyo,personal communication 2004) informed us that a man’s entire fist can fit into themouth of a black carp about 1.5 m long. He also agreed that a fish this size would havea gape width of the approximate size our model predicts.
In addition to measuring mouth gape size, we removed the pharyngeal arches froma young black carp (345 mm TL), keeping the two arches intact and connected byligature (Figure 4.4). Outer dimensions of the entire apparatus measured 40 mm wideand 29 mm high; inner dimensions (open passage or gape within the arches, exclusiveof teeth) measured 20 mm wide and 18 mm high. The gape size of this individualmeasured 22 mm wide, only slightly larger than the pharyngeal arch opening. Becauseblack carp are capable of using their pharyngeal apparatus to crush items, the gapesize between the arches (as well as the distance between the cleithral bones) is lessimportant than its actual mouth gape.
To date, we are unaware of any studies evaluating crushing power exerted by thepharyngeal apparatus of large black carp during their feeding on various mollusks. Intheir study of the postlarval development of the masticating apparatus of black carp,Liu et al. (1990) presented data comparing tooth enamel hardness of juvenile blackcarp (75–195 mm long) with the hardness of one or more unnamed snails. Theyconcluded that after the fish lose their transitional teeth and are passing through sev-eral generations of fixed teeth, the pharyngeal apparatus becomes capable of masti-cating and grinding animals with shells. According to Liu et al. (1990), fixed teeth
chap7.p65 5/4/2005, 4:11 PM96

DIET AND GAPE SIZE
97
appear when black carp are about 26 d old (31–301 mm long). There are publishedpapers containing information on the crushing resistance or hardness of differentsnails in diets of other fishes, for instance, redear sunfish Lepomis microlophus (Frenchand Morgan 1995; Huckins 1997).
W. Shelton (University of Oklahoma, personal communication 1995), discussinghis past work with captive-bred black carp in Israel and Egypt, emphasized that thespecies was clearly gape-limited in terms of the size of mollusks that could be eaten.He indicated that black carp crush food items (with their pharyngeal teeth) beforeswallowing, and because of the black carp’s powerful pharyngeal apparatus, the fishcan usually handle any food item that it can get into its mouth. However, snails that aredifficult to crush, such as those with tight coils (e.g., Melanoides sp.), may require exten-sive manipulation. Based on examination of gut contents, black carp tend to swallowcrushed shells along with the soft parts (see earlier part of this chapter). Still, accord-ing to Shelton, food items that cannot be handled are spit out. Moreover, in someinstances, the fish is able to crack the edge of the shell, extract soft parts, and then spitout shell fragments. Based on Shelton’s comments, clearly black carp are able to ma-nipulate food items to some extent, but it is unclear if black carp are able to consumemussels whose shell dimensions are much longer than high or wide (i.e., shell lengthgreatly exceeds the mouth gape of the predator but where the maximum shell width iswithin gape limits). Additional discussion on this subject appears in Chapter 14.
Although black carp reportedly have small mouths for their size, they attain sizesmuch larger than most native mollusk-eating fish. For instance, French and Morgan(1995) examined eight adult redear sunfish (200–222 mm long) and reported a meangape of the pharyngeal cavity of 11.3 mm. They did not provide measurements ofmouth width.
As will be discussed in Chapter 13, there has been attention, but little research, ofthe effects of black carp predation on nonindigenous zebra mussels. Although blackcarp are capable of consuming individual zebra mussels, there have been no definitivestudies to determine if black carp can break apart large rafts that zebra mussels char-acteristically form (Figure 7.3). Therefore, black carp may not be an effective biologi-cal control agent for this invasive species.
The implications of the research presented in this chapter are important relative tothe native molluscan fauna of North America. The literature on their anatomy anddiet, combined with our data on gape size, suggests that black carp would be fullycapable of preying on adults of many native mussels and snails, even those specieswith thicker shells (see Chapter 14 concerning an analysis relating the size of bivalvesin the Midwest to black carp gape size). There are no known native fish with the samecombination of size, morphology, and diet. Consequently, black carp would fill a nichein North American rivers currently unoccupied, which may alter food webs substan-tially. Even though black carp are molluscivores, the literature suggests that the speciesis somewhat flexible in diet. In areas where mollusks are less common, introducedblack carp could potentially shift their diet and consume native crayfishes and shrimp.
chap7.p65 5/4/2005, 4:11 PM97

DIET AND GAPE SIZE
98
Figure 7.3.—Small aggregate (druse) of zebra mussels attached to native mussel (dimensions about10 × 8 × 7 cm). (Photograph by Leo G. Nico.)
chap7.p65 5/4/2005, 4:11 PM98