changes in properties of glutamate transport in trichoderma viride vegetative mycelia upon...
TRANSCRIPT

Changes in properties of glutamate transport in Trichoderma viridevegetative mycelia upon adaptation to glutamate as carbon source
Richard Pokorny¤ a, Daniela Hudecova¤ a, Wolfgang Burgstaller b, L’udov|¤t Varec›ka a;�
a Department of Biochemistry and Microbiology, Slovak University of Technology, Radlinske¤ho 9, 81102 Bratislava, Slovak Republicb Institute of Microbiology, Leopold-Franzens-University Innsbruck, Technikerstrasse 25, 6020 Innsbruck, Austria
Received 26 September 2003; received in revised form 3 November 2003; accepted 17 November 2003
First published online 5 December 2003
Abstract
The U-14C-labelled glutamate uptake was measured in both sucrose- and glutamate-grown mycelia of Trichoderma viride. The biomassyield was five-fold lower with glutamate as a sole carbon source. The rate of glutamate transport measured at a glutamate concentrationof 1 mM remained unchanged in glutamate-grown mycelia whereas the properties of the glutamate transport were substantially changedcompared to sucrose-grown mycelia. The glutamate uptake in both sucrose- and glutamate-grown mycelia was inhibited by an uncoupler(3,3P,4P,5-tetrachlorosalicylanilide) but the inhibitory efficiency was higher in the latter. The affinity of the permease to glutamateincreased approximately five-fold in the glutamate-grown mycelia (about 76 WM compared to about 16 WM). The pH optimum forglutamate uptake was 4 in sucrose-grown mycelia but the glutamate-grown mycelia had two pH optima, one at pH 4 and the secondbetween pH 6 and 7. The inhibition of glutamate uptake by other amino acids yielded different inhibitory patterns in the two myceliaunder study. The glutamate uptake in mycelia of different ages also showed differences in both transport rate and temporal pattern. Theresults show that the growth of mycelia on glutamate led to the appearance of an additional permease with different properties andsuggest that only this permease is operating in mycelia grown on glutamate.9 2003 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Trichoderma viride ; Glutamate uptake; Developmental change
1. Introduction
Glutamate (Glu) is the amino acid which was found tobe the predominant nitrogen-containing cell constituent ineukaryotic cells and to represent a major part of conidia ofNeurospora crassa [1]. Although its role in metabolism is,in general, known, there are several aspects of Glu metab-olism in fungi which were revealed previously. For exam-ple, in N. crassa, Glu seems to play an important role ingermination of conidia [1] using a Q-aminobutyric acidshunt. This may be true also for germination of Tricho-derma viride conidia [2]. In addition, decarboxylation ofGlu was found to take place in the exponential phase of T.viride submerged and aerial growth and conidiation [2].Further, Glu, when used as a sole carbon source in Tri-choderma atroviride, was found to alkalinize medium and
this was found to be a prerequisite for coal solubilization[3]. Thus, understanding of the character and regulation ofGlu transport may also have some application potential.
Transport of amino acids has been studied for decadesand has been reviewed [4^6]. There is genetic evidence forthe presence of three types of transporters conveying ami-no acids across the plasma membrane, i.e. general perme-ase and transporters for neutral-aromatic and for basicamino acids, respectively [7]. The general permease couldbe induced by short-term pre-incubation with basic aminoacids [8]. The transport of acidic amino acids was found tobe active [9] and mediated by the general permease [9,10].In Aspergillus nidulans, the transport of Glu [11^13] andbasic and neutral amino acids [14] and the genetic regu-lation have been described [15]. In Aspergillus ochraceus,the relationship of Glu transport and metabolism inochratoxin production was studied [16]. Sinoara reportedproperties of the transport of several amino acids intoconidia of Aspergillus oryzae [17]. Also, the amino acidtransport was described in Penicillium spp., such as P.chrysogenum [18^21] and P. cyclopium [22]. Unfortunately,
0378-1097 / 03 / $22.00 9 2003 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.doi :10.1016/S0378-1097(03)00884-X
* Corresponding author. Tel. : +421 (2) 5932 5514;Fax: +421 (2) 5249 3198.
E-mail address: [email protected] (L. Varec›ka).
FEMSLE 11334 29-12-03
FEMS Microbiology Letters 230 (2004) 123^128
www.fems-microbiology.org

only scarce data, if any, are available about the adaptiveresponses of the transport systems in ¢lamentous fungi,and no data about Glu transport in Trichoderma are avail-able. Therefore, the aim of this study was to characterizethe Glu uptake in T. viride and to test whether it ischanged when the fungus is cultivated with Glu as a solecarbon source.
2. Materials and methods
2.1. Strain
T. viride strain CCM F-534 (Czech Collection of Micro-organism, T.G. Masaryk University, Brno, Czech Repub-lic) was used in all experiments.
2.2. Culture conditions
The medium for submerged cultivation of T. viride(Czapek-Dox medium) consisted of (g l31) : 30 sucrose,2 NaNO3, 0.5 KCl, 0.5 MgSO4W7H2O, 0.01 FeSO4W7H2O,1 K2HPO4 and 5 yeast extract. When mycelia had to begrown on Glu as a sole carbon source, L-glutamic acid(13 g l31) was used instead of sucrose. All componentswere dissolved in distilled water and the pH was adjustedto 6.5 before sterilization. One hundred milliliters of eachsucrose- and Glu-containing medium were inoculated witha conidial glycerol stock suspension to a ¢nal density of1U106 conidia ml31 in 250-ml £asks in duplicate. Sub-merged cultivation was carried out for 30 h (or for thetime indicated in the developmental stage dependence ex-periment) at 26^28‡C in the dark on a rotary shaker (250rpm).
2.3. Uptake of [U-14C]L-glutamic acid
Growth was terminated by ¢ltration through a nylonmesh. Mycelia were washed three times with a mineralmedium consisting of (g l31): 2 NaNO3, 0.5 KCl, 0.5MgSO4W7H2O, 0.01 FeSO4W7H2O and 1 K2HPO4, pH 6.5(150 ml for each time) by re-suspension and subsequent¢ltration as above. After the ¢nal ¢ltration step, myceliawere re-suspended in the same mineral medium at pH 6.5(or in mineral media without K2HPO4 but containing 20mM of di¡erent Good bu¡ers (MES, MOPS, HEPES)adjusted with Tris to the pH indicated in the pH depen-dence experiment) to a ¢nal density of 5^7 mg dry weight(DW) ml31 (i.e. 30^40 mg wet weight ml31), or to a ¢naldensity required for the mass dependence experiment. Thesuspension had to show no apparent inhomogeneities. Sus-pensions were aliquoted at 1 ml into sets of Eppendorftubes (measurements of each value in the ¢gures werecarried out in triplicate) for subsequent uptake experi-ments and ¢ve 1-ml aliquots from each suspension wereused for DW estimation (3.5 h at 105‡C).
Uptake experiments were started after 10 min of pre-incubation at 26‡C (or at temperatures indicated in thetemperature dependence experiment) by the addition of0.2 ml of the mixture of [U-14C]L-Glu and non-radioactiveL-Glu in the mineral medium at pH 6.5 (¢nal concentra-tion of Glu was 1 mM or as indicated in the Glu concen-tration dependence experiment and ¢nal speci¢c radioac-tivity was about 40 cpm nmol31). In the case of inhibitionexperiments with L-amino acids and other Glu analogs (K-ketoglutaric acid), these were added from their stock so-lutions in the mineral medium at pH 6.5 to the mycelialsuspension just before adding radionuclide to a ¢nal con-centration of 1 mM. When uncoupler inhibition experi-ments were carried out, 3,3P,4P,5-tetrachlorosalicylanilide(TCS, at the indicated ¢nal concentrations in methanol(0.3% ¢nal)) was added to the mycelial suspension justbefore the radionuclide was added. The suspensions wereimmediately vortexed and incubated at 26‡C (if not indi-cated otherwise) for 60 min (or for the time indicated inkinetics experiments). The reaction was stopped by with-drawing 0.5-ml aliquots that were immediately ¢lteredthrough a membrane ¢lter with a pore size of 1.6 Wm(Whatman GF/A) and washed on ¢lter three times with1 ml of the mineral medium which had the same compo-sition and pH as the complete reaction mixture apart fromthe radioactively labelled L-Glu (and TCS for the uncou-pler inhibition experiment). At time zero (0 min), the ali-quots were withdrawn immediately after addition of radio-active mixture and vortexing (with no further incubation).The washed membrane ¢lters were transferred into a scin-tillation vial contained 0.5 ml of 10% trichloroacetic acidand incubated for 10 min at ambient temperature. Thescintillation liquid (2 ml) was then added and the radio-activity was measured by liquid scintillation counting. Thevalues were corrected for the vial background radioactivitymeasured in advance. The results are expressed as theaverages of triplicates T S.D. of a representative of threeexperiments.
2.4. Chemicals
Chemicals used were from the following sources:[U-14C]L-glutamic acid from ICN Radiochemicals,Aurora, OH, USA; Good’s bu¡ers MES and HEPESfrom Serva, Heidelberg, Germany and MOPS from Fluka,Buchs, Switzerland; Tris-(hydroxymethyl)-aminomethane(Tris) and L-glutamic acid from AppliChem, Darmstadt,Germany; other L-amino acids from Nutritional Biochem-ical Corporation, Cleveland, OH, USA and TCS fromEastman-Kodak, Rochester, NY, USA. All other reagentswere purchased from Lachema, Brno, Czech Republic.
3. Results
The maximal speci¢c growth rate of T. viride on Glu
FEMSLE 11334 29-12-03
R. Pokorny¤ et al. / FEMS Microbiology Letters 230 (2004) 123^128124

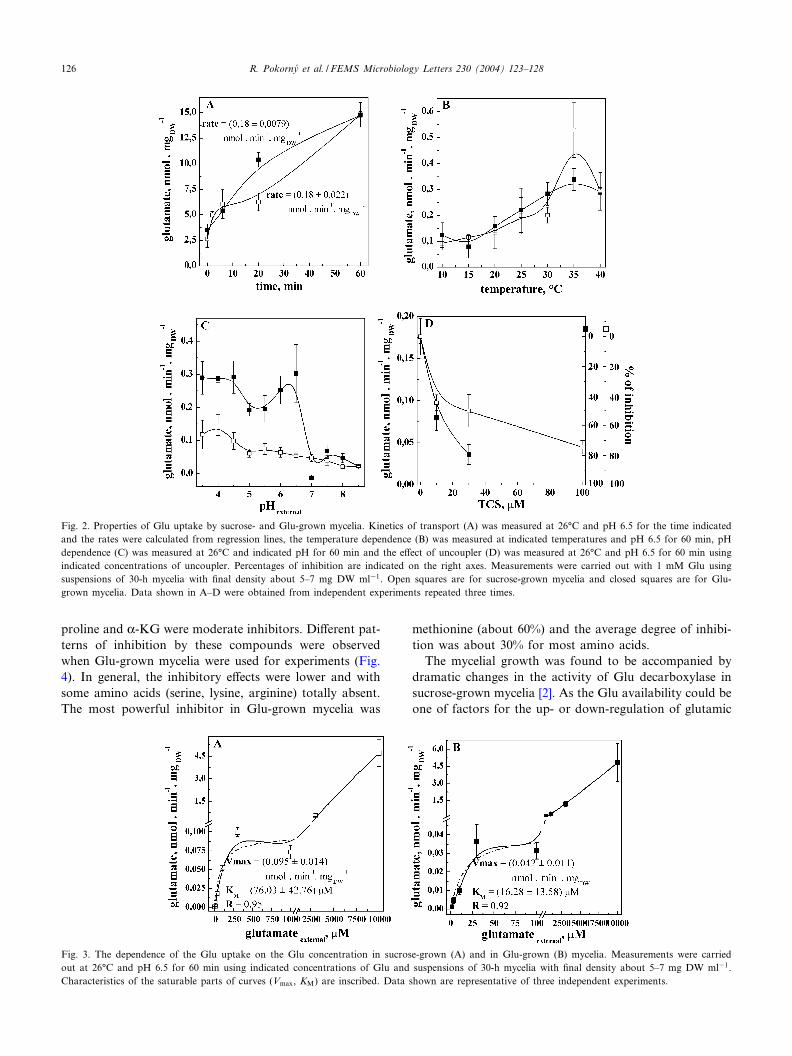
(Wmax = 0.15 h31) was higher than that on sucrose(Wmax = 0.073 h31) but the growth yield was about ¢vetimes lower (Fig. 1A,B). Hyphae, grown on sucrose asthe sole carbon source, were able to transport Glu propor-tionally to the concentration of mycelia in the concentra-tion range between 2.6 and 15 mg DW ml31 (correspondsto 1.1^6.3 mg DW in the withdrawn aliquots, Fig. 1C).Basic properties of Glu transport were measured in bothsucrose- and Glu-grown mycelia. The time course of up-take in sucrose-grown mycelia showed two phases: a rapidphase culminating at about 10 min following by a steadystate, and the second phase leading to a dramatic increasein the uptake (Fig. 2A). The Glu uptake into Glu-grownmycelia was monophasic and the steady state was notestablished until 60 min. The rates of uptake in both my-celia, measured in the presence of 1 mM Glu, were almostidentical, about 0.17 nmol min31 mg DW31 (Fig. 2A). Thetemperature dependence of the Glu uptake in both myceliawas similar with the temperature optimum at 35‡C andGlu uptake was arrested below 15‡C (Fig. 2B). The pHdependence of the Glu uptake in the sucrose-grown myce-lia had an optimum at pH 4 and decreased with pH (Fig.2C). On the other hand, Glu transport into Glu-grownmycelia was more complex with two pH optima, one at
pH 4^4.5 and the second at pH 6.5. The uptake wasstrongly inhibited upon increasing pH over pH 6.5 (Fig.2C). The uptake was inhibited by the uncoupler TCS thatwas a more e⁄cient inhibitor in the Glu-grown mycelia(Fig. 2D). The Glu inward transport was dependent on theextracellular Glu concentration from 1 WM up to 10 mMwith biphasic responses (Fig. 3) in both sucrose-grown andGlu-grown mycelia. The KM(Glu) in sucrose-grown myce-lia was 76.0 WM and Vmax was 0.095 nmol min31 mgDW31, and correspondingly, 16.3 WM and 0.042 nmolmin31 mg DW31 in Glu-grown mycelia. Thus, multipleevidence was obtained for the di¡erence in properties ofthe Glu uptake in these mycelia. The linear componentwhich was observed at higher concentrations of Glu inboth mycelia studied (Fig. 3) was not analyzed.
The e¡ects of other amino acids and of K-ketoglutarate(K-KG) were tested in order to obtain information aboutthe speci¢city of Glu transport in both mycelia (Fig. 4). Insucrose-grown mycelia, all tested amino acids but not glu-tamine exerted the some inhibitory e¡ect on the Glu up-take. The most powerful inhibitory amino acid was notaspartic acid but non-polar amino acids, glycine, alanineand leucine, tryptophan and methionine. Basic aminoacids (lysine, histidine, arginine) as well as aspartic acid,
Fig. 1. Growth curves of T. viride grown on sucrose (A) or Glu (B) as the sole carbon source and the dependence of the Glu uptake on the mycelialmass of sucrose-grown mycelia (C). Growth curves (A,B) were measured with mycelia of the indicated age as described in Section 2. Parameters of the¢tted curves together with the maximal speci¢c growth rates (Wmax), which were estimated as the speci¢c growth rates in the in£ection points (opencircles) of the ¢t curves, are included. Glu transport (C) was measured as indicated in Section 2. Characteristics of three independent experiments areshown.
FEMSLE 11334 29-12-03
R. Pokorny¤ et al. / FEMS Microbiology Letters 230 (2004) 123^128 125
proline and K-KG were moderate inhibitors. Di¡erent pat-terns of inhibition by these compounds were observedwhen Glu-grown mycelia were used for experiments (Fig.4). In general, the inhibitory e¡ects were lower and withsome amino acids (serine, lysine, arginine) totally absent.The most powerful inhibitor in Glu-grown mycelia was
methionine (about 60%) and the average degree of inhibi-tion was about 30% for most amino acids.
The mycelial growth was found to be accompanied bydramatic changes in the activity of Glu decarboxylase insucrose-grown mycelia [2]. As the Glu availability could beone of factors for the up- or down-regulation of glutamic
Fig. 2. Properties of Glu uptake by sucrose- and Glu-grown mycelia. Kinetics of transport (A) was measured at 26‡C and pH 6.5 for the time indicatedand the rates were calculated from regression lines, the temperature dependence (B) was measured at indicated temperatures and pH 6.5 for 60 min, pHdependence (C) was measured at 26‡C and indicated pH for 60 min and the e¡ect of uncoupler (D) was measured at 26‡C and pH 6.5 for 60 min usingindicated concentrations of uncoupler. Percentages of inhibition are indicated on the right axes. Measurements were carried out with 1 mM Glu usingsuspensions of 30-h mycelia with ¢nal density about 5^7 mg DW ml31. Open squares are for sucrose-grown mycelia and closed squares are for Glu-grown mycelia. Data shown in A^D were obtained from independent experiments repeated three times.
Fig. 3. The dependence of the Glu uptake on the Glu concentration in sucrose-grown (A) and in Glu-grown (B) mycelia. Measurements were carriedout at 26‡C and pH 6.5 for 60 min using indicated concentrations of Glu and suspensions of 30-h mycelia with ¢nal density about 5^7 mg DW ml31.Characteristics of the saturable parts of curves (Vmax, KM) are inscribed. Data shown are representative of three independent experiments.
FEMSLE 11334 29-12-03
R. Pokorny¤ et al. / FEMS Microbiology Letters 230 (2004) 123^128126

acid decarboxylase, the changes in the speci¢c Glu uptakewere measured in sucrose-grown and Glu-grown myceliaat various ages (between 6 and 120 h of submerged culti-vation). The speci¢c uptake in sucrose-grown mycelia de-creased monophasically with the age of culture but thedecrease occurred between 6 and 12 h of cultivation andthe uptake remained fairly constant until the end of theexperiment (Fig. 5). The initial decrease in the Glu uptakewas observed also in Glu-grown mycelia. The uptake de-creased until the culture reached about 25 h and rose al-most to the original value between 30 and 40 h. The sec-ond depression in the Glu uptake occurred after about 70h of cultivation and the uptake rose again reaching thesecond peak after about 100 h (Fig. 5).
4. Discussion
It would be fair to stress that the cultivation of anyfungus with a single amino acid as the sole carbon sourceis non-physiological. Nevertheless this situation was usedas a ¢rst step to analyze the role of Glu uptake in thedevelopment and di¡erentiation of T. viride suggested bydevelopmental changes in glutamic acid decarboxylase [2].
The growth yield of the mycelia decreased ¢ve timeswhen Glu was used as the carbon source (Fig. 1A,B).This could be ascribed to the fact that the Glu concentra-tion in the cultivation medium represents a lower carbonconcentration (0.44 M, but the real carbon concentrationavailable for growth is even lower because some Glu hasto be kept for gluconeogenesis) than that of sucrose (1.05M) and that the Glu has a lower energy content thansucrose. The rate of transport did not change dramatically(Fig. 2A), but its other properties, such as a⁄nity (Fig. 3),pH dependence (Fig. 2C) and inhibition by other amino
acids (Fig. 4) as well as developmental changes in thetransport rate (Fig. 5) were di¡erent in fungi grown withthese substrates. This supports the view that the cultiva-tion with Glu caused the induction of a di¡erent transportsystem for Glu uptake. It could be expected that a high-a⁄nity transport system should be operating in non-Glu-grown mycelia, whereas the low-a⁄nity transport withhigher capacity should be present in Glu-grown mycelia.
Di¡erences in the speci¢city of both observed Glu-trans-porting systems could be inferred from the inhibition withother amino acids (Fig. 4). This indicates that the Glu-grown uptake is more speci¢c for Glu than that in su-crose-grown mycelia. It must be mentioned here that thecompeting amino acids could inhibit transport of Glu by adirect competition with the molecule of the transporter, orvia the consumption of proton-motive force (pmf) as wasdemonstrated with uracil transport and this microorgan-ism earlier [23]. This possibility is also feasible as the Gluuptake in sucrose-grown mycelia was inhibited by a broadspectrum of amino acids and was dependent on the pmf(Fig. 2D). However, the Glu uptake in Glu-grown myce-lia, which is even more dependent on the pmf (Fig. 2D), ismuch less susceptible to inhibition by competing aminoacids indicating higher speci¢city of the inducible Glu-transporting system. It has to be established quantitativelywhether the pmf increases during Glu adaptation andwhether it is dissipated by the addition of di¡erent aminoacids.
The decrease of the inward Glu transport rate was ob-served in both sucrose- and Glu-grown conidia duringtheir germination (Fig. 5) suggesting that conidia are com-petent for Glu uptake. However, the accumulation of Gluduring mycelial growth di¡ered dramatically between thecompared mycelia. The minute changes in the transportcapacity of Glu in sucrose-grown mycelia during develop-ment indicate that this transport system/pathway is notoperational during mycelial growth on sucrose as a carbon
Fig. 4. E¡ect of L-amino acids and K-KG on Glu uptake by sucrose-and Glu-grown mycelia. Glu transport was measured at 26‡C and pH6.5 for 60 min with 1 mM Glu and suspensions of 30-h mycelia with ¢-nal density about 5^7 mg DW ml31 using 1 mM of the indicated L-ami-no acids or K-KG. Open columns are for sucrose-grown mycelia andclosed columns are for Glu-grown mycelia. Percentages of inhibition (orstimulation) are indicated on the right axes. Data shown were obtainedfrom three independent experiments.
Fig. 5. Developmental changes in the Glu uptake in sucrose- and Glu-grown mycelia. Measurements were carried out at 26‡C and pH 6.5 for60 min with 1 mM Glu using suspensions of mycelia of indicated agewith ¢nal density about 5^7 mg DW ml31. Open squares are for su-crose-grown mycelia and closed squares are for Glu-grown mycelia.Data were obtained from three independent experiment.
FEMSLE 11334 29-12-03
R. Pokorny¤ et al. / FEMS Microbiology Letters 230 (2004) 123^128 127

source. On the other hand, the growth on Glu was accom-panied by major changes in Glu transport and their timecourse was very similar to that of glutamic acid decarbox-ylase described earlier [2]. This indicates that the Glu me-tabolism is strongly coupled to its transport in T. viride ifused as a sole carbon source. The mechanism of couplinghas not been analyzed yet but pmf generation seems to bean acceptable candidate (Fig. 5). Thus, the up- and down-regulation of pmf generation across the cell membranecould be expected.
Acknowledgements
This work was supported in part by Grant VEGA 01/0109/03, Project VTR 2/9012/21 and by the bilateral grantprogram Austria^Slovakia (dissertation network No. 42/DN).
References
[1] Schmit, J.C. and Brody, S. (1975) Neurospora crassa conidial germi-nation: role of endogenous amino acid pools. J. Bacteriol. 124, 232^242.
[2] Striga¤c›ova¤, J., Chovanec, P., Liptaj, T., Hudecova¤, D., Tursky¤, T.,SY imkovic›, M. and Varec›ka, L’. (2001) Glutamate decarboxylase ac-tivity in Trichoderma viride conidia and developing mycelia. Arch.Microbiol. 175, 32^40.
[3] Ho«lker, U., Ludwig, S., Scheel, T. and Ho«fer, M. (1999) Mechanismsof coal solubilization by the deuteromycetes Trichoderma atrovirideand Fusarium oxysporum. Appl. Microbiol. Biotechnol. 52, 57^59.
[4] Hora¤k, J. (1986) Amino acid transport in eucaryotic microorganisms.Biochim. Biophys. Acta 864, 223^256.
[5] Sophianopoulou, V. and Diallinas, G. (1995) Amino acid transport-ers of lower eukaryotes: regulation, structure and topogenesis. FEMSMicrobiol. Rev. 16, 53^75.
[6] Wol¢nbarger, L. (1980) Transport and utilization of amino acids byfungi. In: Microorganisms and Nitrogen Sources (Payne, J.W., Ed.),pp. 63^87. John Wiley and Sons, New York.
[7] Wol¢nbarger Jr., L. and De Busk, A.G. (1971) Molecular transport.In vivo studies of transport mutants of Neurospora crassa with al-tered amino acid competition patterns. Arch. Biochem. Biophys. 144,503^511.
[8] Wol¢nbarger Jr., L. and De Busk, A.G. (1971) An inducible aminoacid transport system in Neurospora crassa. Biochim. Biophys. Acta241, 677^681.
[9] Wol¢nbarger Jr., L., Jervis, H.H. and De Busk, A.G. (1971) Activetransport of L-aspartic acid in Neurospora crassa. Biochim. Biophys.Acta 249, 63^68.
[10] Wol¢nbarger, L.Jr. and De Busk, A.G. (1972) The kinetics of L-aspartate transport in Neurospora crassa conidia. Biochim. Biophys.Acta 290, 355^367.
[11] Robinson, J.H., Anthony, C. and Drabble, W.T. (1973) The acidicamino-acid permease of Aspergillus nidulans. J. Gen. Microbiol. 79,53^63.
[12] Robinson, J.H., Anthony, C. and Drabble, W.T. (1973) Regulationof the acidic amino-acid permease of Aspergillus nidulans. J. Gen.Microbiol. 79, 65^80.
[13] Pateman, J.A., Kinghorn, J.R. and Dunn, E. (1974) Regulatory as-pects of L-glutamate transport in Aspergillus nidulans. J. Bacteriol.119, 534^542.
[14] Piotrowska, M., Stepien, P.P., Bartnik, E. and Zakrzewska, E. (1976)Basic and neutral amino acid transport in Aspergillus nidulans. J. Gen.Microbiol. 92, 89^96.
[15] Kinghorn, J.R. and Pateman, J.A. (1975) Mutations which a¡ectamino acid transport in Aspergillus nidulans. J. Gen. Microbiol. 86,174^184.
[16] Bacon, C.W., Robbins, J.D. and Burdick, D. (1975) Metabolism ofglutamic acid in Aspergillus ochraceus during the biosynthesis ofochratoxin A. Appl. Microbiol. 29, 317^322.
[17] Sinoara, H. (1974) Amino acid transport in spores of Aspergillusoryzae. J. Biochem. (Tokyo) 76, 513^522.
[18] Hunter, D.R. and Segel, I.H. (1971) Acidic and basic amino acidtransport systems of Penicillium chrysogenum. Arch. Biochem. Bio-phys. 144, 168^183.
[19] Hunter, D.R. and Segel, I.H. (1973) Control of the general aminoacid permease of Penicillium chrysogenum by transinhibition andturnover. Arch. Biochem. Biophys. 154, 387^399.
[20] Hunter, D.R. and Segel, I.H. (1973) E¡ect of weak acids on aminoacid transport by Penicillium chrysogenum : evidence for a proton orcharge gradient as the driving force. J. Bacteriol. 113, 1184^1192.
[21] Benko, P.V., Wood, T.C. and Segel, I.H. (1969) Multiplicity andregulation of amino acid transport in Penicillium chrysogenum.Arch. Biochem. Biophys. 129, 498^508.
[22] Roos, W. (1989) Kinetic properties, nutrient-dependent regulationand energy coupling of amino-acid transport systems in Penicilliumcyclopium. Biochim. Biophys. Acta 978, 119^133.
[23] Lakatos›, B., SYimkovic›, M., Betina, V. and Varec›ka, L’. (1999) Prop-erties of uracil transport by vegetative mycelium of Trichoderma vi-ride. FEMS Microbiol. Lett. 171, 161^165.
FEMSLE 11334 29-12-03
R. Pokorny¤ et al. / FEMS Microbiology Letters 230 (2004) 123^128128