ceramide mediates inhibition of the akt/enos pathway by high levels of glucose in human vascular...
TRANSCRIPT

DOI 10.1515/jpem-2012-0144 J Pediatr Endocr Met 2013; 26(1-2): 31–38
Aimin Wang , Chun Li , Jie Liao , Min Dong , Zhiming Xiao and Minxiang Lei *
Ceramide mediates inhibition of the Akt/eNOS pathway by high levels of glucose in human vascular endothelial cells
Abstract Objective: To investigate how ceramide mediates the
effects of high-glucose-induced inhibition of the Akt/
endothelial nitric oxide synthase (eNOS) signalling
pathway in human vascular endothelial cells (HUVECs).
Materials and methods: NO levels were determined by
ELISA. Endogenous ceramide levels were determined
using a liquid chromatography-mass spectrometry assay.
Akt and eNOS protein expressions were determined by
Western blotting.
Results: High-glucose levels induce ceramide accumula-
tion in a dose- and time-dependent manner (p < 0.05). We
also show that exposure of HUVECs to high-glucose con-
ditions inhibits the insulin-mediated activation of Akt/
eNOS signalling and the subsequent NO generation in a
dose-dependent manner (p < 0.05). Preventing de novo
ceramide synthesis attenuated the antagonistic effects of
high-glucose levels on the Akt/eNOS signalling pathway
(p < 0.05); conversely, inducing ceramide build-up aug-
mented the inhibitory effects of high-glucose levels on the
Akt/eNOS signalling pathway (p < 0.05).
Conclusion: Ceramide is both necessary and sufficient
for mediating the inhibition of the Akt/eNOS signalling
pathway by high-glucose levels in endothelial cells.
Keywords: Akt/endothelial nitric oxide synthase (eNOS);
ceramide; glucose; nitric oxide (NO); type 2 diabetes mellitus.
*Corresponding author: Minxiang Lei, Department of Endocrinology,
Xiangya Hospital, Central South University, Xiangya Road, 87 # ,
Changsha, Hunan 410008, China, Phone: + 86-731-84327089,
Fax: + 86-731-84327089, E-mail: [email protected]
Aimin Wang: Department of Endocrinology , Xiangya Hospital,
Central South University, Changsha, Hunan, China
Chun Li: Department of Endocrinology , Xiangya Hospital, Central
South University, Changsha, Hunan, China
Jie Liao: Department of Endocrinology , Xiangya Hospital, Central
South University, Changsha, Hunan, China
Min Dong: School of Pharmaceutical Sciences , Guangxi Medical
University, Nanning, Guangxi, China
Zhiming Xiao: The Third Hospital of Xiangya , Central South
University, Changsha, China
Introduction Type 2 diabetes mellitus is characterised by chronic hyper-
glycaemia and inadequate β -cell responses to progressive
insulin resistance (1, 2) . Chronic hyperglycaemia is the
proximate cause of many diabetic complications, includ-
ing retinopathy, kidney failure, neuropathies, and vascu-
lar disease (3, 4) . Chronic hyperglycaemia also has toxic
effects on the vascular endothelium, another target of dia-
betic complications (5) . Oxidative stress induced by high-
glucose levels is a common pathophysiological pathway
found in diabetic complications, especially with respect
to vascular disease.
The vascular endothelium is an insulin-sensitive
tissue; in vascular endothelial cells, the phosphati-
dylinositol 3 kinase (PI3K) pathway activates the serine/
threonine protein kinase Akt, which phosphorylates
endothelial nitric oxide synthase (eNOS) leading to
increased nitric oxide (NO) production (6, 7) . Changes
in NO levels are the earliest known characteristic of
dysfunctional vascular endothelial cells (8, 9) . There-
fore, downregulation of insulin signalling can lead to
metabolic and cardiovascular dysfunction. Ceramide, a
metabolic byproduct of the signalling molecule sphin-
gomyelin, has been widely implicated in the regula-
tion of intracellular signalling pathways, and it plays
important roles in cell growth, proliferation, motility,
adhesion, differentiation, senescence, and apoptosis
(10 – 13) . As a second-messenger lipid, ceramide is sig-
nificantly associated with insulin resistance induced
by tumour necrosis factor- α (TNF- α ) and free fatty acid
(FFA) (14, 15) . Ceramide also blocks the insulin recep-
tor substrate-1 (IRS-1)/PI3K/eNOS signalling pathway at
several different stages. Our previous studies have dem-
onstrated that FFA can induce ceramide accumulation
in human umbilical vein endothelial cells (HUVECs),
leading to reductions in eNOS phosphorylation and
vascular endothelial dysfunction (16) . Dandan et al.
demonstrated that high-glucose levels could induce
apoptosis in vascular endothelial cells and increase
intracellular ceramide accumulation (17) . Chun et al.
Brought to you by | University of Queensland - UQ LibraryAuthenticated | 130.102.42.98
Download Date | 9/25/13 4:10 PM

32 Wang et al.: Ceramide and Akt/eNOS signalling pathway
found that inhibition of de novo ceramide synthesis
improved endothelium-dependent vasodilation (EDVD)
of the arteries of streptozotocin-induced diabetic rats
(18) . Therefore, ceramide accumulation is an important
mediator associated with vascular endothelial dysfunc-
tion. However, it remains unclear whether ceramide
accumulation causes high-glucose-induced endothelial
dysfunction through targeting of the insulin-induced
Akt/eNOS signalling pathway.
Ceramide biosynthesis requires the coordinated
action of two enzymes: serine palmitoyltransferase and
ceramide synthase (19) . Serine palmitoyltransferase
catalyses the initial step, which involves the condensa-
tion of serine and palmitoyl-CoA to form 3-keto-sphin-
ganine. Studies have shown that high-glucose-induced
mechanical dysfunction can be significantly attenu-
ated by both the membrane-permeant ceramide analog
C2-ceramide or the ceramide glucosyltransferase inhibi-
tor D, L-threo-1-pheny-2-decanoylamino-3-morpholino-
1-propanol (PDMP). We hypothesised that ceramide
might be the principal effector molecule that mediates
high-glucose-induced Akt/eNOS inhibition. Therefore,
we first determined whether myriocin and desipramine
(DES) – inhibitors of de novo ceramide synthesis – could
prevent high-glucose-induced inhibition of the Akt/PKB
signalling pathway. Next, we determined whether PDMP
and N-oleoylethanolamine (NOE) – specific inhibitors
of ceramide glucosyltransferase – could also influence
high-glucose-induced inhibition of the Akt/PKB signal-
ling pathway.
Based on the experiments described above, we inves-
tigated how ceramide affects the high-glucose-induced
inhibition of the Akt/eNOS signalling pathway in cultured
HUVECs. Our study suggests that ceramide is an impor-
tant mediator molecule that plays a crucial role in high-
glucose-induced inhibition of the Akt/eNOS signalling
pathway.
Materials and methods
Cell culture HUVECs were cultured in Dulbecco ’ s modified eagle media
(DMEM) supplemented with 10 % foetal calf serum (FCS). The cells
were maintained at 37 ° C under a humidified atmosphere of 5 % CO 2
and 95 % air in a Forma Scientific incubator (Biosciences, Dublin,
Ireland). Differentiated HUVECs were seeded into six-well plates
at 5 × 10 5 cells per well and allowed to adhere overnight. When
cultures reached 90 % confluence, the supernatants were harvest-
ed and/or cell lysates were prepared for assaying as described
below.
High-glucose treatment and insulin stimulation First, HUVECs were exposed to 5 mmol/L D-glucose and 30 mmol/L
D-glucose (high glucose) medium for 4, 8, 12, 16, or 24 h to observe
ceramide accumulation levels. Next, HUVECs were incubated with
various concentrations of D-glucose (5, 10, 20, or 30 mmol/L) for 16 h
to observe ceramide accumulation levels and NO production. Finally,
HUVECs were stimulated with 50 nmol/L insulin for 30 min to observe
changes in NO levels. The groups were as follows: (i) HUVECs treated
with 25 mmol/L L-glucose and 5, 10, 20, or 30 mmol/L D- glucose;
(ii) to observe the changes of NO levels, HUVECs were stimulated with
50 nmol/L insulin for 30 min in media supplemented with 25 mmol/L
L-glucose and 5, 10, 20, or 30 mmol/L D-glucose; (iii) select samples
were treated with 10 μ mol/L myriocin or 2 μ mol/L DES prior to the ad-
dition of high levels (30 mmol/L) of glucose; and (iv) select samples
were treated with or without high levels of glucose in the presence of
the ceramidase inhibitor (NOE) (250 μ mol/L) or the glucosylceramide
synthase inhibitor (PDMP) (50 μ mol/L), which were added prior to
glucose addition.
Ceramide assays Confl uent HUVEC cultures were incubated in 175 cm 2 tissue culture
fl asks at 37 ° C for 30 min in 20 mL of serum-free DMEM, followed by
rinsing with the same medium. Next, the cells were detached with
0.05 % trypsin/0.53 mmol/L EDTA (Gibco-BRL) and pelleted by cen-
trifugation. The pellets were washed 3 times with 10 mL cold phos-
phate buff ered saline (PBS), followed by another centrifugation step.
Approximately 5 × 10 6 washed cells were resuspended in 220 μ L cold
0.25 mmol/L sucrose in PBS, transferred into microfuge tubes, and
disrupted by sonication. The cell lysates were then centrifuged at
800 × g for 10 min, and the supernatants containing the cytosolic
fraction – made up of the cytoplasm and organelles – were with-
drawn. The pellets containing the membrane fractions were then
washed 3 times with 1 mL cold PBS, centrifuged, and suspended in
220 μ L PBS. Ceramide levels in each of the above fractions were de-
termined using the methods described by Zhou et al. (20) . Deriva-
tised ceramide was separated from byproducts using 3 % 2-propanol
in n-hexane as the mobile phase. The fl ow rate was 2.0 mL/min, and
the eluted compounds were monitored using a fl uorescence spectro-
photometer with an excitation wavelength of 230 nm and emission
wavelength of 352 nm.
Determination of NO concentrations Although NO is unstable, its end products – nitrite and nitrate –
are stabile. Therefore, in order to index NO generation, we deter-
mined the sum of the arterial concentrations of nitrite and nitrate.
The culture media were harvested and stored at – 80 ° C until used
in the assays. Nitrite and nitrate levels were measured as described
previously (21) . Briefl y, nitrate was converted to nitrite with nitrate
reductase, and total nitrite was reacted to the Griess reagent; the
coloured product could then be quantifi ed by its absorbance at 540
nm using a spectrophotometer. Each experiment was repeated in
triplicate.
Brought to you by | University of Queensland - UQ LibraryAuthenticated | 130.102.42.98
Download Date | 9/25/13 4:10 PM
Wang et al.: Ceramide and Akt/eNOS signalling pathway 33
Western blot analyses To obtain total protein, cell extracts were prepared in lysis buff er con-
taining 50 mmol/L Tris HCl (pH 8.0), 150 mmol/L NaCl, 0.1 % SDS, 0.1
mmol/L phenylmethylsulphonyl fl uoride (PMSF), 10 lg/mL aprotinin,
1 % Nonidet (N)P-40, and 0.5 % sodium deoxycholate. The lysates
were cleared by centrifugation at 12,000 × g for 10 min at 4 ° C. Total
protein concentrations were measured using the Bradford method
(22) . The proteins were separated on 10 % SDS-PAGE gels, transferred
to polyvinylidene difl uoride membranes, and then probed with pri-
mary antibodies against Akt, eNOS, or GADPH; all primary antibod-
ies were polyclonal rabbit antibodies from Santa Cruz Biotechnology.
Evaluations were performed with the Fluor Chem. 8900 soft ware
system.
Statistical analyses All statistical analyses were carried out using SPSS soft ware (version
13.0 for Windows; SPSS, Chicago, IL, USA). Statistical signifi cance of
the mean values was evaluated using a one-way analysis of variance
(ANOVA) test for measurements and an LSD-t-test for comparisons
between datasets. All data are presented as means ± standard deviation
(SD). A p-value < 0.05 was considered to be statistically signifi cant.
Results
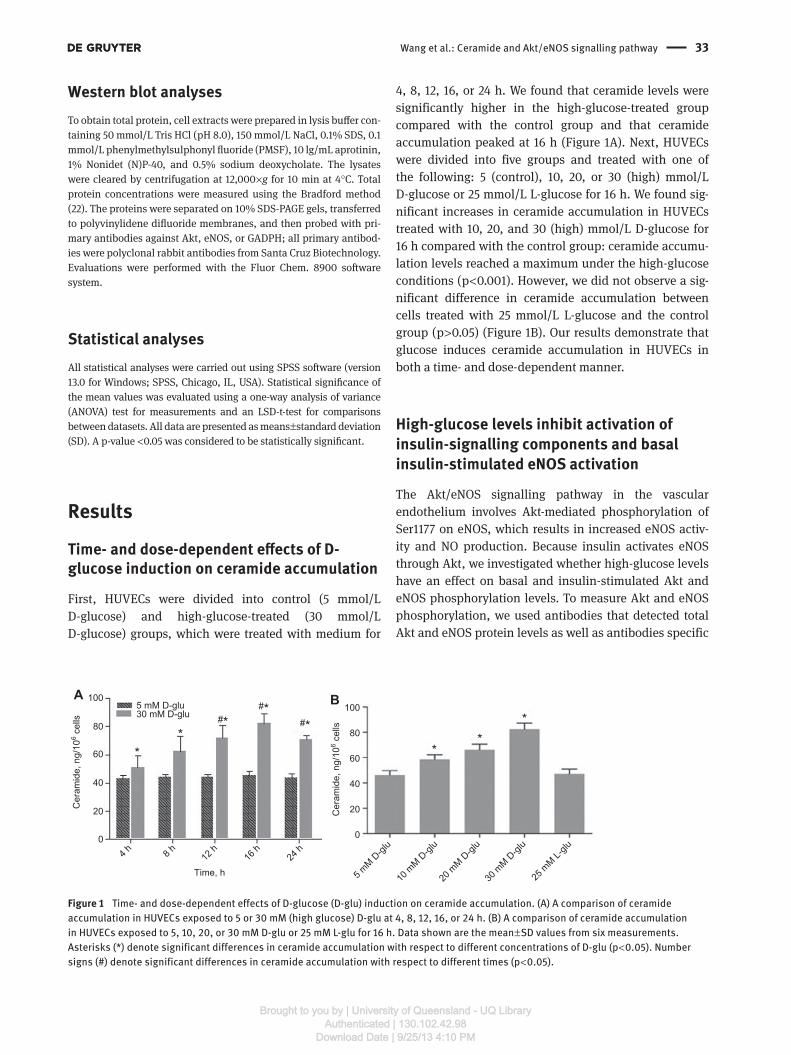
Time- and dose-dependent eff ects of D- glucose induction on ceramide accumulation
First, HUVECs were divided into control (5 mmol/L
D- glucose) and high-glucose-treated (30 mmol/L
D- glucose) groups, which were treated with medium for
4, 8, 12, 16, or 24 h. We found that ceramide levels were
significantly higher in the high-glucose-treated group
compared with the control group and that ceramide
accumulation peaked at 16 h (Figure 1 A). Next, HUVECs
were divided into five groups and treated with one of
the following: 5 (control), 10, 20, or 30 (high) mmol/L
D- glucose or 25 mmol/L L-glucose for 16 h. We found sig-
nificant increases in ceramide accumulation in HUVECs
treated with 10, 20, and 30 (high) mmol/L D-glucose for
16 h compared with the control group: ceramide accumu-
lation levels reached a maximum under the high-glucose
conditions (p < 0.001). However, we did not observe a sig-
nificant difference in ceramide accumulation between
cells treated with 25 mmol/L L-glucose and the control
group (p > 0.05) (Figure 1B). Our results demonstrate that
glucose induces ceramide accumulation in HUVECs in
both a time- and dose-dependent manner.
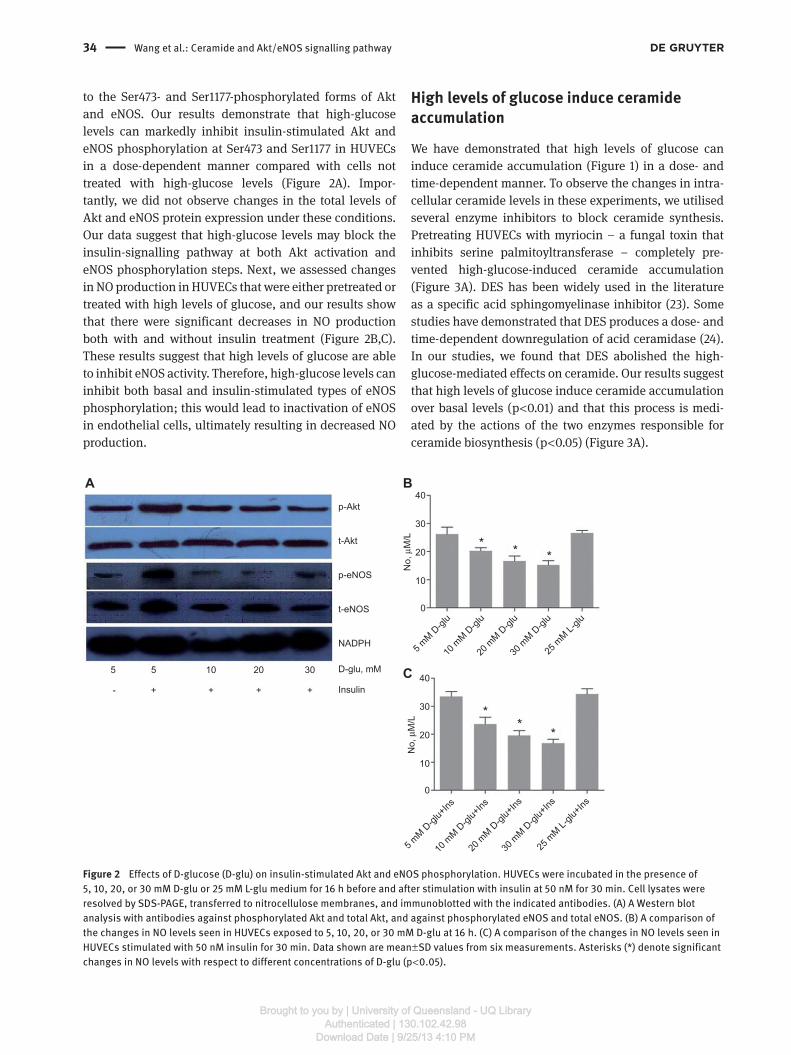
High-glucose levels inhibit activation of insulin-signalling components and basal insulin-stimulated eNOS activation
The Akt/eNOS signalling pathway in the vascular
endothelium involves Akt-mediated phosphorylation of
Ser1177 on eNOS, which results in increased eNOS activ-
ity and NO production. Because insulin activates eNOS
through Akt, we investigated whether high-glucose levels
have an effect on basal and insulin-stimulated Akt and
eNOS phosphorylation levels. To measure Akt and eNOS
phosphorylation, we used antibodies that detected total
Akt and eNOS protein levels as well as antibodies specific
100A B5 mM D-glu30 mM D-glu
#*#*#*
**
**
*
80
60
Cer
amid
e, n
g/10
6 cel
ls
40
20
Time, h
0
100
80
60
Cer
amid
e, n
g/10
6 cel
ls
40
20
0
4 h 8 h 12 h
16 h
24 h
5 mM D
-glu
10 m
M D-gl
u
20 m
M D-gl
u
30 m
M D-gl
u
25 m
M L-glu
Figure 1 Time- and dose-dependent effects of D-glucose (D-glu) induction on ceramide accumulation. (A) A comparison of ceramide
accumulation in HUVECs exposed to 5 or 30 mM (high glucose) D-glu at 4, 8, 12, 16, or 24 h. (B) A comparison of ceramide accumulation
in HUVECs exposed to 5, 10, 20, or 30 mM D-glu or 25 mM L-glu for 16 h. Data shown are the mean ± SD values from six measurements.
Asterisks (*) denote significant differences in ceramide accumulation with respect to different concentrations of D-glu (p < 0.05). Number
signs ( # ) denote significant differences in ceramide accumulation with respect to different times (p < 0.05).
Brought to you by | University of Queensland - UQ LibraryAuthenticated | 130.102.42.98
Download Date | 9/25/13 4:10 PM
34 Wang et al.: Ceramide and Akt/eNOS signalling pathway
to the Ser473- and Ser1177-phosphorylated forms of Akt
and eNOS. Our results demonstrate that high-glucose
levels can markedly inhibit insulin-stimulated Akt and
eNOS phosphorylation at Ser473 and Ser1177 in HUVECs
in a dose-dependent manner compared with cells not
treated with high-glucose levels (Figure 2 A). Impor-
tantly, we did not observe changes in the total levels of
Akt and eNOS protein expression under these conditions.
Our data suggest that high-glucose levels may block the
insulin-signalling pathway at both Akt activation and
eNOS phosphorylation steps. Next, we assessed changes
in NO production in HUVECs that were either pretreated or
treated with high levels of glucose, and our results show
that there were significant decreases in NO production
both with and without insulin treatment (Figure 2B,C).
These results suggest that high levels of glucose are able
to inhibit eNOS activity. Therefore, high-glucose levels can
inhibit both basal and insulin-stimulated types of eNOS
phosphorylation; this would lead to inactivation of eNOS
in endothelial cells, ultimately resulting in decreased NO
production.
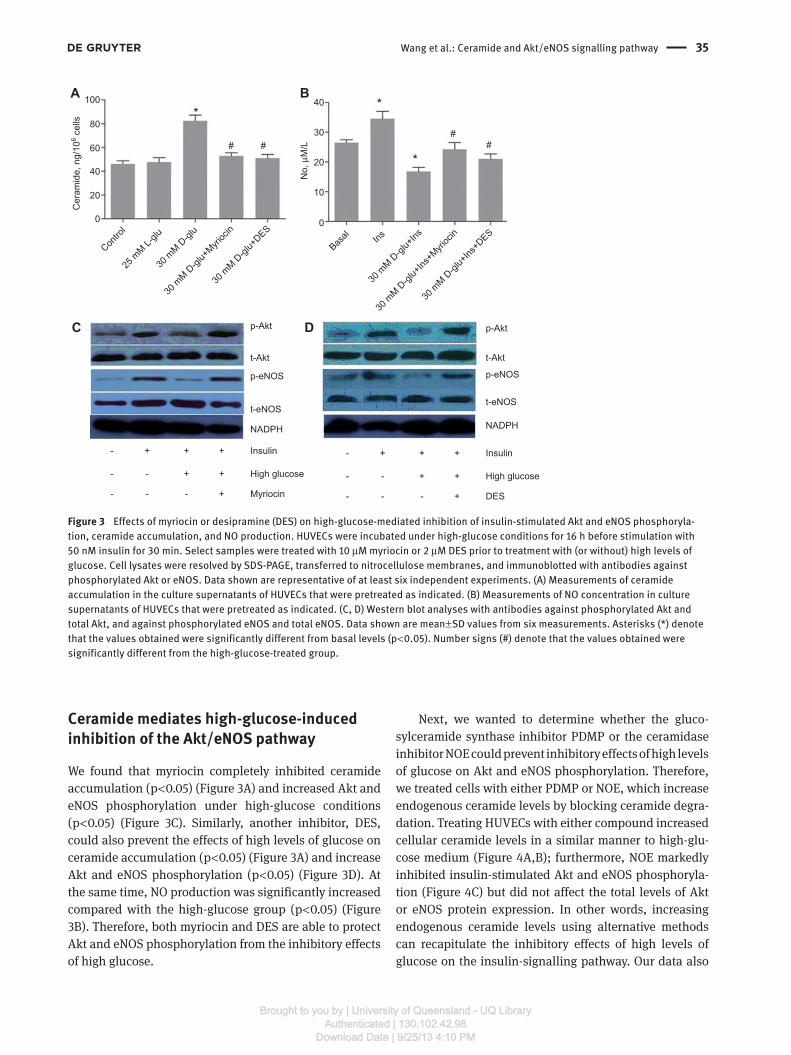
High levels of glucose induce ceramide accumulation
We have demonstrated that high levels of glucose can
induce ceramide accumulation (Figure 1) in a dose- and
time-dependent manner. To observe the changes in intra-
cellular ceramide levels in these experiments, we utilised
several enzyme inhibitors to block ceramide synthesis.
Pretreating HUVECs with myriocin – a fungal toxin that
inhibits serine palmitoyltransferase – completely pre-
vented high-glucose-induced ceramide accumulation
(Figure 3 A). DES has been widely used in the literature
as a specific acid sphingomyelinase inhibitor (23) . Some
studies have demonstrated that DES produces a dose- and
time-dependent downregulation of acid ceramidase (24) .
In our studies, we found that DES abolished the high-
glucose-mediated effects on ceramide. Our results suggest
that high levels of glucose induce ceramide accumulation
over basal levels (p < 0.01) and that this process is medi-
ated by the actions of the two enzymes responsible for
ceramide biosynthesis (p < 0.05) (Figure 3A).
40
30
20
No,
μM
/L
10
5 5 10 20 30
p-Akt
A B
C
t-Akt
p-eNOS
t-eNOS
NADPH
D-glu, mM
Insulin- + + + +
0
40
30
20
*
* * *
**
No,
μM
/L
10
0
5 mM D
-glu
5 mM D
-glu+
Ins
10 m
M D-gl
u+Ins
20 m
M D-gl
u+Ins
30 m
M D-gl
u+Ins
25 m
M L-glu
+Ins
10 m
M D-gl
u
20 m
M D-gl
u
30 m
M D-gl
u
25 m
M L-glu
Figure 2 Effects of D-glucose (D-glu) on insulin-stimulated Akt and eNOS phosphorylation. HUVECs were incubated in the presence of
5, 10, 20, or 30 mM D-glu or 25 mM L-glu medium for 16 h before and after stimulation with insulin at 50 nM for 30 min. Cell lysates were
resolved by SDS-PAGE, transferred to nitrocellulose membranes, and immunoblotted with the indicated antibodies. (A) A Western blot
analysis with antibodies against phosphorylated Akt and total Akt, and against phosphorylated eNOS and total eNOS. (B) A comparison of
the changes in NO levels seen in HUVECs exposed to 5, 10, 20, or 30 mM D-glu at 16 h. (C) A comparison of the changes in NO levels seen in
HUVECs stimulated with 50 nM insulin for 30 min. Data shown are mean ± SD values from six measurements. Asterisks (*) denote significant
changes in NO levels with respect to different concentrations of D-glu (p < 0.05).
Brought to you by | University of Queensland - UQ LibraryAuthenticated | 130.102.42.98
Download Date | 9/25/13 4:10 PM
Wang et al.: Ceramide and Akt/eNOS signalling pathway 35
Ceramide mediates high-glucose-induced inhibition of the Akt/eNOS pathway
We found that myriocin completely inhibited ceramide
accumulation (p < 0.05) (Figure 3A) and increased Akt and
eNOS phosphorylation under high-glucose conditions
(p < 0.05) (Figure 3C). Similarly, another inhibitor, DES,
could also prevent the effects of high levels of glucose on
ceramide accumulation (p < 0.05) (Figure 3A) and increase
Akt and eNOS phosphorylation (p < 0.05) (Figure 3D). At
the same time, NO production was significantly increased
compared with the high-glucose group (p < 0.05) (Figure
3B). Therefore, both myriocin and DES are able to protect
Akt and eNOS phosphorylation from the inhibitory effects
of high glucose.
Next, we wanted to determine whether the gluco-
sylceramide synthase inhibitor PDMP or the ceramidase
inhibitor NOE could prevent inhibitory effects of high levels
of glucose on Akt and eNOS phosphorylation. Therefore,
we treated cells with either PDMP or NOE, which increase
endogenous ceramide levels by blocking ceramide degra-
dation. Treating HUVECs with either compound increased
cellular ceramide levels in a similar manner to high-glu-
cose medium (Figure 4 A,B); furthermore, NOE markedly
inhibited insulin-stimulated Akt and eNOS phosphoryla-
tion (Figure 4C) but did not affect the total levels of Akt
or eNOS protein expression. In other words, increasing
endogenous ceramide levels using alternative methods
can recapitulate the inhibitory effects of high levels of
glucose on the insulin-signalling pathway. Our data also
100A
C D
B
# ##
#
**
*
80
40
30
20
No,
μM
/L
10
p-Akt
t-Akt
p-eNOS
t-eNOS
NADPH
Insulin- + + +
High glucose- - + +
Myriocin- - - +
p-Akt
t-Akt
p-eNOS
t-eNOS
NADPH
Insulin- + + +
High glucose- - + +
DES- - - +
Basal Ins
30 m
M D-gl
u+Ins
30 m
M D-gl
u+Ins
+Myri
ocin
30 m
M D-gl
u+Ins
+DES
0
60
40
Cer
amid
e, n
g/10
6 cel
ls
20
0
Contro
l
25 m
M L-glu
30 m
M D-gl
u
30 m
M D-gl
u+Myri
ocin
30 m
M D-gl
u+DES
Figure 3 Effects of myriocin or desipramine (DES) on high-glucose-mediated inhibition of insulin-stimulated Akt and eNOS phosphoryla-
tion, ceramide accumulation, and NO production. HUVECs were incubated under high-glucose conditions for 16 h before stimulation with
50 nM insulin for 30 min. Select samples were treated with 10 μ M myriocin or 2 μ M DES prior to treatment with (or without) high levels of
glucose. Cell lysates were resolved by SDS-PAGE, transferred to nitrocellulose membranes, and immunoblotted with antibodies against
phosphorylated Akt or eNOS. Data shown are representative of at least six independent experiments. (A) Measurements of ceramide
accumulation in the culture supernatants of HUVECs that were pretreated as indicated. (B) Measurements of NO concentration in culture
supernatants of HUVECs that were pretreated as indicated. (C, D) Western blot analyses with antibodies against phosphorylated Akt and
total Akt, and against phosphorylated eNOS and total eNOS. Data shown are mean ± SD values from six measurements. Asterisks (*) denote
that the values obtained were significantly different from basal levels (p < 0.05). Number signs ( # ) denote that the values obtained were
significantly different from the high-glucose-treated group.
Brought to you by | University of Queensland - UQ LibraryAuthenticated | 130.102.42.98
Download Date | 9/25/13 4:10 PM

36 Wang et al.: Ceramide and Akt/eNOS signalling pathway
demonstrate that PDMP and NOE can augment the effects
of high-glucose-induced NO generation by blocking cera-
mide glucosylation or deacylation.
Discussion The study reported here demonstrates that ceramide plays
an important role in high-glucose-induced inhibition of
the Akt/eNOS signalling pathway in vascular endothelial
cells. Our findings include the following: (i) high levels of
glucose induce ceramide accumulation; (ii) high levels of
glucose inhibit Akt and eNOS activity through the Akt/
eNOS signalling pathway in cultured HUVECs, leading to
significant decreases in NO generation; (iii) the inhibitory
effects of high levels of glucose on Akt/eNOS signalling
pathway activation can be reversed by inhibiting de novo
ceramide synthesis; and (iv) blocking ceramide metabo-
lism both increases ceramide accumulation and inhibits
the Akt/eNOS pathway. Our findings suggest that cera-
mide is a key effector molecule that links high levels of
glucose to endothelial cell dysfunction through the Akt/
eNOS signalling pathway.
There exist many potential mechanisms whereby high
levels of glucose might induce endothelial cell damage.
High levels of glucose can cause chronic oxidative stress,
which in turn leads to the apoptosis of vascular endothelial
cells. The Akt/eNOS pathway involves a series of phospho-
rylation reactions dependent on insulin signalling (25, 26) .
100A
C
# #
*80
p-Akt
t-Akt
p-eNOS
t-eNOS
NADPH
- + + + Insulin
- - - 30 D-glu
NOE- - + -
+
30
+
60
40
Cer
amid
e, n
g/10
6 cel
ls
20
0
Contro
l
25 m
M L-glu
30 m
M D-gl
u
30 m
M D-gl
u+NOE
30 m
M D-gl
u+PDMP
B
* * *
40
30
20
No,
μM
/L
10
Basal Ins
30 m
M D-gl
u+Ins
30 m
M D-gl
u+Ins
+NOE
30 m
M D-gl
u+Ins
+PDMP
0
Figure 4 Effects of inhibitors on high-glucose-mediated inhibition of insulin-signalling component phosphorylation, ceramide accumula-
tion, or NO production. HUVECs were incubated in 30 mM D-glu (high glucose) conditions for 16 h before stimulation with 100 nM insulin
for 10 min. Select samples were treated either with or without high glucose in the presence of NOE (250 μ M) or PDMP (50 mM). Cell lysates
were resolved by SDS-PAGE, transferred to nitrocellulose membranes, and immunoblotted with antibodies against phosphorylated Akt or
eNOS. Data shown are representative of at least four independent experiments. (A) Western blot analyses with antibodies against phos-
phorylated Akt and total Akt. (B) Western blot analyses with antibodies against phosphorylated eNOS and total eNOS. (C) Measurements of
ceramide accumulation in culture supernatants of HUVECs that were pretreated as indicated. Measurements of NO concentrations in culture
supernatants of HUVECs that were pretreated as indicated. Data shown are mean ± SD values from six measurements. Asterisks (*) denote
that the values obtained were significantly different from the high-glucose-treated group (p < 0.05). Number signs ( # ) denote that the values
obtained were significantly different from the high-glucose-treated group.
Brought to you by | University of Queensland - UQ LibraryAuthenticated | 130.102.42.98
Download Date | 9/25/13 4:10 PM

Wang et al.: Ceramide and Akt/eNOS signalling pathway 37
After insulin binds to the insulin receptor (IR) on the cell
surface, the IR becomes activated and phosphorylates
IRS-1 by PTK inside the cell, which then phosphorylates
IRS-1 and PI3K, which in turn phosphorylate and activate
Akt (27, 28) . Because phosphorylated Akt is the kinase
that directly phosphorylates eNOS, Akt phosphorylation
levels may be an indicator of eNOS activity; therefore, we
investigated the effect of high levels of glucose on basal
and insulin-stimulated Akt activation by monitoring Akt
phosphorylation at Ser473. We observed that high levels
of glucose inhibited Akt phosphorylation in a dose-
dependent manner and also inhibited insulin-stimulated
eNOS phosphorylation/activation. We also showed that
high levels of glucose could inhibit eNOS activity through
the Akt/eNOS pathway; this resulted in a significant
decrease in NO generation, which could promote the
onset of endothelial dysfunction. High levels of glucose
inhibited eNOS phosphorylation at Ser1177 and activation
in a dose-dependent manner and also inhibited insulin-
stimulated eNOS phosphorylation. Because NO is a key
regulator of endothelial function, eNOS inhibition by high
levels of glucose could provide a mechanism to explain
endothelial dysfunction in diabetic vascular compliance.
Our results suggest that high levels of glucose are likely
to be a major risk factor for diabetic vascular compliance,
consistent with previous reports.
Ceramide is capable of inhibiting both Akt and Akt-
mediated eNOS phosphorylation. Our findings demon-
strate that inhibiting de novo ceramide synthesis can
prevent high-glucose-induced ceramide accumulation as
well as its antagonistic effects on the Akt/eNOS pathway,
resulting in eNOS activation and NO generation. We also
found that blocking ceramide metabolism while simul-
taneously adding high levels of glucose augmented both
ceramide accumulation and the phosphorylation of Akt
and eNOS.
Several convincing studies have demonstrated
that high levels of glucose cause increases in ceramide
concentration and endothelial cell apoptosis (17, 18) . In
this study, we employed several inhibitors to block spe-
cific enzymes involved in ceramide metabolism. First, the
fungal toxins myriocin and DES inhibit separate enzymes
required for de novo ceramide synthesis; both toxins
were able to protect HUVECs from ceramide accumula-
tion induced by high glucose. Furthermore, both of the
ceramide glucosyltransferase inhibitors PDMP and NOE,
which inhibit glucosylceramide synthase and ceramidase,
respectively, were able to induce ceramide accumulation.
Our data indicate that high levels of glucose can induce
ceramide accumulation and that ceramide is an important
mediator molecule associated with inhibition of the high-
glucose-induced Akt/eNOS pathway.
In summary, we demonstrate that high levels of
glucose induce ceramide accumulation, and we report
the novel finding that ceramide plays an important role
with respect to the inhibition of the Akt/eNOS pathway
by high levels of glucose. Further efforts will be neces-
sary to explore the precise mechanisms behind diabetic
vascular compliance and high-glucose-induced vascu-
lar endothelial cell dysfunction as mediated by the Akt/
eNOS signalling pathway on both the cellular and organ-
ismal levels.
Acknowledgements: We would like to thank all of the par-
ticipants in this study. This work was supported by the
National Natural Science Foundation of China (30871190)
and the Hunan Provincial Finance Department and Edu-
cation Steering Committee under grant no. (2009).
Conflict of interest statement The authors declare no conflicts of interest.
Received May 4, 2012; accepted September 21, 2012; previously
published online November 9, 2012
References 1. Stumvoll M, Goldstein BJ, van Haeften TW. Type 2 diabetes:
principles of pathogenesis and therapy. Lancet 2005;365:
1333 – 46.
2. Kannel WB, McGee DL. Diabetes and cardiovascular disease: the
Framingham Study. J Am Med Assoc 1978;241:2035 – 8.
3. Brownlee M. Biochemistry and molecular cell biology of diabetic
complications. Nature 2001;414:813 – 20.
4. King GL, Loeken MR. Hyperglycaemia-induced oxidative stress in
diabetic complications. Cell Biol 2004;122:333 – 8.
5. Robertson RP. Chronic oxidative stress as a central mechanism
for glucose toxicity of pancreatic islet beta cells in diabetes. J Biol
Chem 2004;279:42351 – 4.
6. Tousoulis D, Kampoli AM, Tentolouris C, Papageorgiou N,
Stefanadis C. The role of nitric oxide on endothelial function. Curr
Vasc Pharmacol 2012;10:4 – 18.
7. Fulton D, Gratton JP, McCabe TJ, Fontana J, Fujio Y, et al.
Regulation of endothelium-derived nitric oxide production by the
protein kinase Akt. Nature 1999;399:597 – 601.
Brought to you by | University of Queensland - UQ LibraryAuthenticated | 130.102.42.98
Download Date | 9/25/13 4:10 PM

38 Wang et al.: Ceramide and Akt/eNOS signalling pathway
8. Calles-Escandon J, Cipolla M, Cipolla M. Diabetes and
endothelial dysfunction, a clinical perspective. Endocr Rev
2001;22:36 – 52.
9. Dimmeler S, Fleming I, Fisslthaler B, Hermann C, Busse R,
et al. Activation of nitric oxide synthase in endothelial cells by
Akt-dependent phosphorylation. Nature 1999;399:601 – 5.
10. Jayadev S, Liu B, Bielawska AE, Lee JY, Nazaire F, et al. Role for
ceramide in cell cycle arrest. J Biol Chem 1995;270:2047 – 52.
11. Hannun YA. The sphingomyelin cycle and the second messenger
function of ceramide. J Biol Chem 1994;269:3125 – 8.
12. Mathias S, Pena LA, Kolesnick RN. Signal transduction of stress
via ceramide. Biochem J 1998;335:465 – 80.
13. Hannun YA, Obeid LM. Ceramide: an intracellular signal for
apoptosis. Trends Biochem Sci 1995;20:73 – 7.
14. Soeda S, Tsunoda T, Kurokawa Y, Shimeno H. Tumor necrosis
factor- α -induced release of plasminogen activator inhibitor-1
from human umbilical vein endothelial cells: involvement of
intracellular ceramide signalling event. Biochim Biophys Acta
1998;1448:37 – 45.
15. Xiao-Yun X, Zhuo-Xiong C, Min-Xiang L, Xingxuan H, Schuchman
EH, et al. Ceramide mediates inhibition of the AKT/eNOS
signalling pathway by palmitate in human vascular endothelial
cells. Med Sci Monit 2009;15:BR254 – 61.
16. Stratford S, Hoehn KL, Liu F, Summers SA. Regulation of
insulin action by ceramide: dual mechanisms linking ceramide
accumulation to the inhibition of Akt/protein kinase B. J Biol
Chem 2004;279:36608 – 15.
17. Dandan H, Minxiang L, Xingyuan H, et al. Effects of pioglitazone
on high glucose-induced vascular endothelial cell apoptosis
and the involved mechanisms. Chin J Diabetes 2009;17:709 – 11.
18. Chun L, Junlin Z, Aimin W, Niansheng L, Benmei C, et al.
Inhibition of ceramide synthesis reverses endothelial
dysfunction and atherosclerosis in streptozotocin-induced
diabetic rats. Diabetes Res Clin Pract 2011;93:77 – 85.
19. Merrill AH Jr. De novo sphingolipid biosynthesis: a necessary,
but dangerous pathway. J Biol Chem 2002;277:25843 – 6.
20. Zhou Q, Zhang L, Fu X-Q, Chen GQ. Quantitation of yeast
ceramides using high-performance liquid chromatography-
evaporative light-scattering detection. J Chromatogr B Analyt
Technol Biomed Life Sci 2002;780:161 – 9.
21. Nims RW, Cook JC, Krishna MC, Christodoulou D, Poore CM,
et al. Colorimetric assays for nitric oxide and nitrogen oxide
species formed from nitric oxide stock solutions and donor
compounds. Methods Enzymol 1996;268:93 – 105.
22. Silv é rio SC, Moreira S, Milagres AM, Macedo EA, Teixeira JA,
et al. Interference of some aqueous two-phase system phase-
forming components in protein determination by the Bradford
method. Anal Biochem 2012;421:719 – 24.
23. Albouz S, Hauw JJ, Berwald-Netter Y, Boutry JM, Bourdon R, et al.
Tricyclic antidepressants induce sphingomyelinase deficiency
in fibroblast and neuroblastoma cell cultures. Biomedicine
1981;35:218 – 20.
24. Elojeimy S, Holman DH, Liu X, El-Zawahry A, Villani M, et al.
New insights on the use of desipramine as an inhibitor for acid
ceramidase. FEBS Lett 2006;580:4751 – 6.
25. McVeigh GE, Cohn JN. Endothelial dysfunction and the
metabolic syndrome. Curr Diab Rep 2003;3:87 – 92.
26. Kobayashi T, Taguchi K, Yasuhiro T, Matsumoto T, Kamata
K. Impairment of PI3-K/Akt pathway underlies attenuated
endothelial function in aorta of type 2 diabetic mouse model.
Hypertension 2004;44:956 – 62.
27. Montagnani M, Chen H, Barr VA, Quon MJ. Insulin-stimulated
activation of eNOS is independent of Ca 2 + but requires
phosphorylation by Akt at Ser (1179). J Biol Chem 2001;276:
30392 – 8.
28. Zeng G, Quon MJ. Insulin-stimulated production of nitric oxide
is inhibited by wortmannin: direct measurement in vascular
endothelial cells. J Clin Invest 1996;98:894 – 8.
Brought to you by | University of Queensland - UQ LibraryAuthenticated | 130.102.42.98
Download Date | 9/25/13 4:10 PM