cells, tissues, and organs on chips challenges and opportunities for the cancer tumor...
TRANSCRIPT

LI
ISSN 1757-9694
www.rsc.org/ibiology Volume 5 | Number 9 | September 2013 | Pages 1089–1198
REVIEW ARTICLEEdmond W. K. YoungCells, tissues, and organs on chips: challenges and opportunities for the cancer tumor microenvironment
Interdisciplinary approaches for molecular and cellular life sciences

1096 Integr. Biol., 2013, 5, 1096--1109 This journal is c The Royal Society of Chemistry 2013
Cite this: Integr. Biol.,2013,5, 1096
Cells, tissues, and organs on chips: challenges andopportunities for the cancer tumor microenvironment
Edmond W. K. Young*
The transition to increasingly sophisticated microfluidic systems has led to the emergence of ‘‘organ-on-
chip’’ technology that can faithfully recapitulate organ-level function. Given the rapid progress at the
interface between microfluidics and cell biology, there is need to provide a focused evaluation of the
state-of-the-art in microfluidic systems for cancer research to advance development, accelerate discovery
of novel insights, and facilitate cooperation between engineers, biologists and oncologists in the
clinic. Here, we provide a focused review of microfluidics technology from cells- and tissues- to
organs-on-chips with application toward studying the tumor microenvironment. Key aspects of the tumor
microenvironment including angiogenesis, hypoxia, biochemical gradients, tumor–stromal interactions,
and the extracellular matrix are summarized for both solid tumors and non-solid hematologic malignancies.
An overview of microfluidic systems designed specifically to answer questions related to different
aspects of the tumor microenvironment is provided, followed by an examination of how these
systems offer new opportunities to study outstanding challenges related to the major cancer hallmarks.
Challenges also remain for microfluidics engineers, but it is hoped that cooperation between engineers
and biologists at the intersection of their respective fields will lead to significant impact on the utility of
organs-on-chips in cancer research.
Insight, innovation, integrationOrgan-on-chip technologies are attracting significant interest with existing reviews focusing mostly on technical aspects of the field. This review focuses insteadon cells, tissues, and organs-on-chips specifically for studying cancer biology. By discussing the advancement of microfluidic cell-based systems from thestandpoint of the disease and the key elements within the tumor microenvironment, this review offers a different perspective that may lead to new organ-on-chipinnovations as well as unexpected insights on how we approach cancer research in the future.
1. Introduction
The integration of microfluidics and cell biology research hasrecently reached another significant milestone with develop-ment of ‘‘organ-on-chip’’ technologies. What began at the turnof the millennium as simple demonstrations of biological cellsbeing transported and manipulated in microchannels for basicshort-term analysis1,2 has now advanced to the point where wecan engineer living cellular microsystems with controllablemicroenvironments that behave and function – with organ-levelcomplexity – like their counterparts in vivo.3–5 While advance-ment from cell- to tissue- to organ-level function in vitro hasbeen impressive from an engineering perspective, what is truly
compelling is the potential impact this technology will have onthe study of human diseases, and on clinical and therapeuticapplications that may ultimately improve overall human health.
Cancer research in particular has potential to reap immensebenefits from the application of organ-on-chip technologies.A major challenge in cancer research has been the need todevelop more accurate, more informative, and more predictiveexperimental models of human tumor development than thosecurrently available. Traditional in vitro systems and in vivoanimal models have both made major contributions to ourcurrent understanding of the disease, and to the discovery andpractical application of oncotherapies, but these models alsohave shortcomings that limit their overall effectiveness towardpredicting the success or failure of many candidate drugcompounds during the screening process, as well as that ofselected therapeutic strategies.6 Ineffectiveness in the formercase has led to a disappointingly high attrition rate of potential
Department of Mechanical & Industrial Engineering, University of Toronto,
5 King’s College Road, MC314B, Toronto, ON, M5S 3G8, Canada.
E-mail: [email protected]; Fax: +1 (416) 978-7753; Tel: +1 (416) 978-1521
Received 16th April 2013,Accepted 6th June 2013
DOI: 10.1039/c3ib40076j
www.rsc.org/ibiology
Integrative Biology
REVIEW ARTICLE
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
.
View Article OnlineView Journal | View Issue

This journal is c The Royal Society of Chemistry 2013 Integr. Biol., 2013, 5, 1096--1109 1097
candidates late in the drug discovery process that is both costlyand time consuming. Organ-on-chip technologies have potentialto offer significant improvements in in vitro models of thedisease, with better physiological mimicry of the tumor micro-environment than existing in vitro assays, and more speciesrelevance than in vivo animal models (if primary human cellsare employed). Importantly, the role of the tumor micro-environment in cancer initiation and progression, and therecognition that a tumor itself should be considered a complexorgan,7 suggests that physiological microenvironments withimproved relevance to humans and perhaps even organ-levelcomplexity are integral to our understanding of the disease,and must be given more consideration in future methodologiesof experimental research. The purpose of developing improvedexperimental models through organ-on-chip technologies is toaccelerate progress in fundamental research, to increase effi-ciency in drug discovery, and to advance the translation of newknowledge into clinical outcomes. To accelerate the potentialimpact of this technology on cancer research, it is importantand necessary to review the pertinent literature to date, parti-cularly work that has already shown the promise of microfluidicchip technologies in meeting the needs of cancer research.
This review discusses the current state-of-the-art in cells-,tissues-, and organs-on-chips technology specifically for cancermicroenvironments. While other recent reviews on this technol-ogy have provided more general overviews of technical advancesin device functionality and complexity,8,9 this review focuseson cancer, and categorizes various microsystems based on keyfactors relevant to the tumor microenvironment. The potentialfor these microsystems to contribute to cancer research isproposed in relation to major cancer hallmarks, and a discus-sion of major challenges facing those working at the interfacebetween microfluidics and cancer is provided. The combinationof these different aspects distinguishes this review from otherrecent articles summarizing the application of microfluidics forcancer research.10,11 For convenience and completeness, a briefoverview is provided on the essential elements of the tumormicroenvironment for both solid tumors and nonsolid malig-nancies. Regarding nomenclature, where appropriate, differentcellular microsystems are referred to in specific terms as cells-on-chips, tissues-on-chips, or organs-on-chips, depending onbiological complexity. However, for brevity, these microsystemsare also collectively referred to here as either microfluidicsystems or ‘‘biochips’’ to cover all three hierarchical levels.
2. Essentials of the tumor microenvironment
It has become increasingly evident that cancer development, inits remarkable complexity, is not only a matter of individualtumor cells evolving as a result of multiple genetic mutations,but also involves the complex interactions between the tumorcells and the many physical, biological, and molecular factorsof the surrounding tumor microenvironment. From a historicalperspective, the well-known seed-soil hypothesis by Paget in 1889,as a proposed explanation for site-specific metastases, was one ofthe first signs of recognition that the local microenvironment
(although distant from the primary site in this case) may con-tribute to tumorigenicity, enabling metastatic lesions to thrive inan otherwise foreign and hostile environment.12,13 Since then, anextensive body of literature has provided strong support for therole of the tumor microenvironment,14,15 leading to majorfields of research ranging from angiogenesis and hypoxia todimensionality and cancer mechanobiology. The tumor micro-environment as a whole has thus emerged as a central player incancer research, and as a consequence has become a majorpotential target for cancer therapy.
In experimental biology, our inability to accurately model thehuman tumor microenvironment in experimental systems is oneof the major and perhaps most important reason why many ofour results in the lab do not translate directly to improvedoutcomes in the clinic. The question of whether current in vitrosystems and in vivo animal models are sufficiently representativeof human physiological conditions to be predictive is a majorsource of debate, and stems from the recognition that thesemodels are limited in physiological context, either lacking impor-tant spatial and temporal cues in the case of simplistic planar(2D) in vitro models, or lacking the genotype specific to humansin the case of animal models. Thus, for decades, while bothexperimental paradigms provided significant contributions toour understanding of the disease, and resulted in novel dis-coveries in therapeutics, there remained an ongoing concernrelated to the limitations of our experimental repertoire.
The potential for organ-on-chip technology to provide arealistic, unconventional and potentially disruptive alternativeto existing in vitro and in vivo methods is therefore an excitingproposition. We now have the ability through microfabricationtechniques and microengineered control to construct withvarying levels of complexity living microsystems that recapitulatedifferent aspects of the microenvironment using human cells,perhaps even those derived directly from diseased individuals.This level of physiological relevance to humans is not achievablevia existing methodologies. Given such power to tailor themicroenvironment as desired for each experiment or applica-tion, the question is no longer whether we can do it. Instead, thequestion is how we do it, and what we do with the technology tobest serve the future of cancer research.
To begin to answer these questions, we briefly examine thevarious aspects of the developing tumor microenvironment(Fig. 1), first in the context of an epithelial-derived carcinoma.Carcinomas represent over 80% of all cancers, including thoseof the breast, colon, liver, lung, pancreas, and prostate, amongothers, and are collectively considered the largest class ofneoplasias. Subsequently, we will also examine aspects of themicroenvironment for non-solid hematologic malignancies, asecond major class of cancer that has unique features distinctfrom the microenvironment of solid tumors. For convenience,the discussion will proceed in sequence through the stepsleading from normal health to diseased state.
Solid tumors
Carcinomas derived from epithelial tissue originate from singlecell clones, which proliferate and aggregate into a mass of
Review Article Integrative Biology
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

1098 Integr. Biol., 2013, 5, 1096--1109 This journal is c The Royal Society of Chemistry 2013
mutant cells that form the basis of a tumor. Because tumorcells continue to accumulate mutations during tumor progres-sion, the population of cells becomes increasingly hetero-geneous, with successive generations of mutant cells mixedwith the initial clonal population. Accumulating evidence sug-gests that this amalgam also consists of tumor initiating cells,more popularly known as cancer stem cells, which are capableof spawning new tumors when injected into previously healthymice.16 Tumor cells are surrounded by the stroma, whichconsists of the three-dimensional structural framework ofextracellular matrix (ECM) components and the various othercell types that support the associated connective tissue. TheECM allows anchorage of tumor and stromal cells via cell–surface integrins that transduce mechanical signals andmediate various mechanobiological responses. Non-tumorstromal cells make up an estimated 80% of the cells in the
tumor microenvironment, and are arranged in specific posi-tions and in specific proportions, creating a complex networkof heterotypic interactions that plays a central role in tumorprogression.17 Notable cell types in the stroma include: thevarious types of fibroblasts (e.g., normal stromal fibroblasts,myofibroblasts, cancer-associated fibroblasts), which are closelyrelated, yet display distinct phenotypic markers, and mayoriginate from different precursors;18 immune cells such asmacrophages, neutrophils, and T and B lymphocytes that sensepro-inflammatory cytokines and chemokines secreted by thetumor and infiltrate the microenvironment; adipocytes such asthose found in the mammary gland;19 and endothelial cells andsupporting pericytes, which are recruited into the micro-environment by the tumor-associated inflammatory cells thatsecrete soluble factors involved in inducing angiogenesis.20
Angiogenesis is the formation and extension of new blood
Fig. 1 The cancer tumor microenvironment consists of a heterogeneous mix of cells in complex spatial arrangement. Cell types include: tumor cells (e.g., carcinoma),cancer stem cells, invasive phenotypes, inflammatory immune cells, assorted fibroblasts, pericytes, and endothelial cells of angiogenic blood vessels. The cells aresupported by extracellular matrix. Regions far from blood supply are hypoxic and subject to oxygen gradients (dark shaded region). Invasive phenotypes may undergoepithelial–mesenchymal transition (EMT), migrate away from the tumor core toward the margins (dotted line), and intravasate into neighboring blood vessels wherethey circulate to ectopic locations and form metastases. (a) Prostate cancer coculture spheroids in microchannels.33 (b) 3D side-by-side coculture of human mammaryfibroblasts and ductal carcinoma in situ (DCIS) cells to study role of stromal interactions in transition to invasive ductal carcinoma.48 (c) Microfluidic chip for generatingperfusable microvessel networks.58 HUVECs stained for nuclei (blue), f-actin (green), and CD31 (red). (d) Microfluidic system for creating oxygen gradient by oxygen-generating and oxygen-scavenging chemical reactions.63 For (a) to (d), images reproduced in part from ref. 33, 48, 58, and 63, respectively, with permission of TheRoyal Society of Chemistry. (e) Microfluidic system for studying cancer cell (HT1080, red) invasion and intravasation through endothelium (human microvascularendothelial cells, MVECs, green). Adapted from ref. 71 Copyright 2012 National Academy of Sciences, USA.
Integrative Biology Review Article
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

This journal is c The Royal Society of Chemistry 2013 Integr. Biol., 2013, 5, 1096--1109 1099
vessels from existing vasculature that provides oxygen andnutrients to the tumor that are necessary for its growth. Asthe primary tumor grows and blood vessels infiltrate the tumor,internal oxygen and chemical gradients form in relation to theangiogenic network, creating regions of hypoxic and necroticcells. The complex set of microenvironmental cues contributeto the differentiation of some tumor cells into more migratoryand invasive phenotypes, possibly via epithelial–mesenchymaltransition (EMT), which gives rise to cells that escape the tumormass via intravasation into nearby blood vessels. These circulat-ing tumor cells (CTCs) then travel via the circulatory system todistant ectopic locations where they have the potential to reside,colonize, and form secondary tumors called metastases. Fig. 1illustrates these major elements of the tumor microenvironment.
Liquid tumors
A second major class of cancers is hematologic malignancies,or cancers related to the blood, bone marrow, and lymphnodes (i.e., ‘‘liquid tumors’’).21 While certain elements of themicroenvironment exist in both solid and liquid tumors (e.g.,angiogenesis), elements unique to hematologic cancers areimportant enough to warrant mention here. For example, interms of cell types, hematopoietic stem cells reside in the bonemarrow and have been implicated in the pathogenesis andprogression of hematologic malignancies,22 but have notbeen implicated in solid tumors. Other cell types such asmesenchymal stromal cells, bone marrow stromal cell (BMSCs),and monocyte-derived nurse-like cells belong to the hemato-poietic lineage, and thus are also only found in the micro-environments of hematologic malignancies. The presence oflymphoma-associated macrophages (LAMs) suggests a similarlyimportant role for inflammatory cells in liquid tumors as insolid tumors. Secondary lymphoid organs are characterized bypresence of T cells responsible for assisting in antigen recogni-tion. Naturally, the bone milieu also includes osteoblasts andosteoclasts that reside in the bone matrix. Whereas solidtumors can be described by the location of the primary site,hematologic cancers exist in the lymphatic and circulatorytransport systems, and are thus characterized by their presencein circulation. Of importance is the notion that the role of themicroenvironment in blood-related cancers is highly variablebetween cancer types, ranging from a very limited role inBurkitt lymphoma to a highly involved, coexisting role inmultiple myeloma (MM) and chronic lymphocytic leukemia(CLL), which has implications on the likely responsiveness ofa blood cancer type to therapeutic strategies that target themicroenvironment.23
3. Microfluidic technology for tumormicroenvironments
From a molecular and cell biology perspective, the briefoverview of tumor microenvironments presented above isadmittedly too simple to capture the inherent complexity of amulticellular system that can be influenced by so many
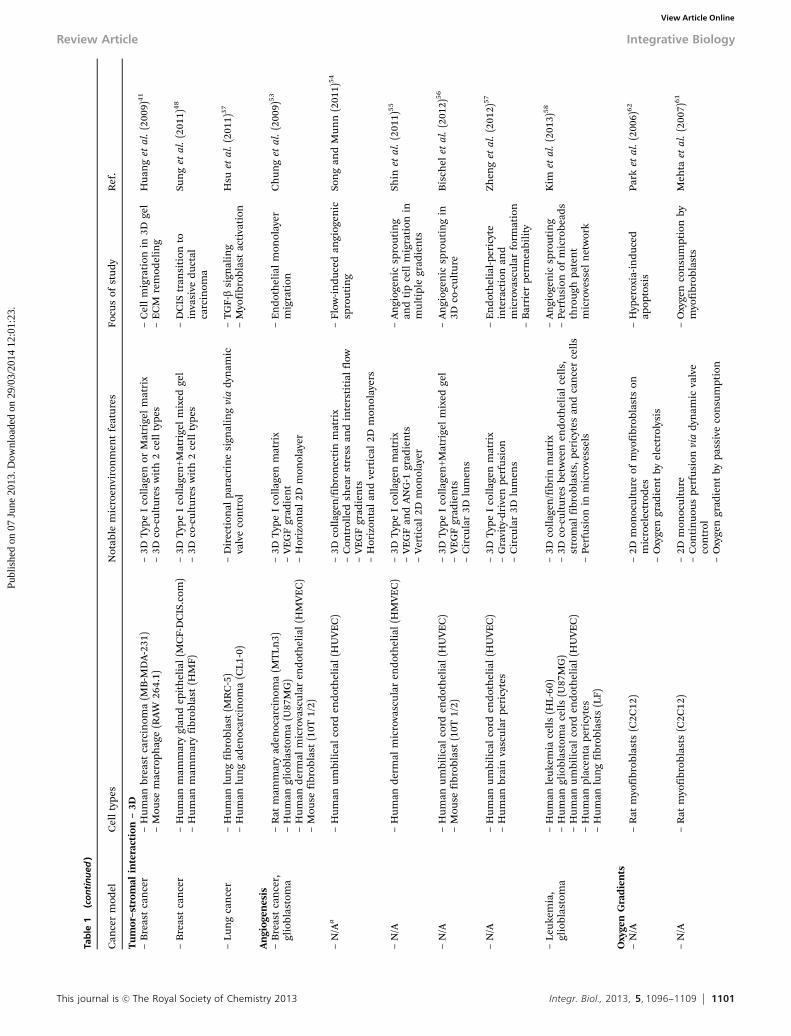
environmental factors. The important point to take from thesummary above is that development of the tumor micro-environment progresses through a series of major events (e.g.,tumor initiation; angiogenesis) that involve key features (tumornodule formation; new blood vessels) with specific functionalroles, and that the coordinated interplay of these features leadsto tumor progression. Advances in microfluidic cell-basedsystems or biochips have led to a new class of in vitro toolswith both demonstrated utility in recapitulating the variousaspects of the developing tumor microenvironment (Fig. 1), andthe potential to offer insight into unprecedented spatio-temporal dynamics of these events. Microfluidic systems havevarying levels of complexity, ranging from simple 2D cellculture microchips to more physiologically relevant 3D tissuemicrochips. Importantly, microfluidic systems offer variousadvantages over traditional in vitro systems, including morecomplex geometries, better spatiotemporal control of micro-environmental factors and stimuli, added functionality, andincreased throughput.24 In this section, recent research studiesemploying microfluidic systems to study aspects of the tumormicroenvironment are reviewed, with an attempt to organizethe studies in order of increasing physiologic complexity andmore advanced stage of tumor development (Table 1).
Tumor spheroids
To model a tumor, a popular method is to employ multicellulartumor spheroids (MCTSs), which are derived from a collectionof cells that aggregate under non-adherent culture conditionsto form three-dimensional cellular masses. The morphology,growth kinetics, and cell–cell and cell–matrix interactionswithin these spheroids resemble tumor nodules, making themexcellent models of tumor initiation and growth, and a usefultool for testing drug delivery and efficacy.25 Various methodshave been developed over decades to achieve efficient spheroidformation, including rotating cultures in roller tubes andspinner flasks, as well as stationary cultures in hanging drops,96-well plates, and related organotypic cultures.26 Techniquesinvolving 96-well plates have been adapted to enable high-throughput spheroid-based drug screens, with protocolsbecoming increasingly standardized.27 Although microtiterplates have many advantages, such as enabling testing andmonitoring of spheroids in independent wells, and allowingconsiderable reduction in reagent consumption compared torotating cultures, the static, diffusion-dominant conditions inwells lead to accelerated nutrient depletion and waste accumu-lation, which can affect spheroid formation and function, andpotentially lead to false identification of drug candidates.27
Microfluidic systems have recently been shown to be effective atfacilitating the formation of tumor spheroids on chip. High-throughput arrays of hanging drops can be generated in well-plate format with the use of microfluidics to create uniform andindependently addressable spheroids.28–30 Microscale geo-metries can be designed to hydrodynamically trap cells to createspheroids with highly uniform size distributions, and cansimultaneously allow continuous perfusion to maintain long-term spheroid culture.31,32 In addition, porous membranes
Review Article Integrative Biology
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

1100 Integr. Biol., 2013, 5, 1096--1109 This journal is c The Royal Society of Chemistry 2013
Tab
le1
Sum
mar
yo
fm
icro
fluid
icte
chn
olo
gy
for
stu
dyi
ng
tum
or
mic
roen
viro
nm
ents
Can
cer
mod
elC
ell
type
sN
otab
lem
icro
envi
ron
men
tfe
atu
res
Focu
sof
stu
dy
Ref
.
Tu
mor
sph
eroi
d–
Bre
ast
can
cer
–H
um
anbr
east
can
cer
(MC
F-7)
–C
onti
nu
ous
perf
usi
on–
Mu
ltic
ellu
lar
aggr
egat
efo
rmat
ion
in3D
gel
Toh
etal
.(2
007)
34
–Li
ver
can
cer
–H
um
anli
ver
carc
inom
a(H
epG
-2)
–3D
cell
–mat
rix
and
cell
–cel
lin
tera
ctio
ns
–C
ell
viab
ilit
y
–B
reas
tca
nce
r–
Hu
man
brea
stca
nce
r(M
CF-
7)–
Con
tin
uou
spe
rfu
sion
–Sp
her
oid
form
atio
nW
uet
al.
(200
8)3
1
–Pr
osta
teca
nce
r–
Hu
man
pros
tate
can
cer
(PC
-3)
–Sp
her
oid
co-c
ult
ure
wit
h3
cell
type
s–
Sph
eroi
dfo
rmat
ion
Hsi
aoet
al.
(200
9)3
3
–H
um
anu
mbi
lica
lco
rden
dot
hel
ial
(HU
VE
C)
–C
onti
nu
ous
perf
usi
onin
adja
cen
tco
mpa
rtm
ent
sepa
rate
dby
poro
us
mem
bran
e–
2Dvs
.3D
–M
ouse
pre-
oste
obla
st(M
C3T
3-E
1)–
Mon
o-vs
.co
-cu
ltu
re
–C
olon
can
cer
–H
um
anco
lon
can
cer
(Col
o205
)–
Flow
-in
du
ced
shea
rst
ress
–Sp
her
oid
form
atio
nA
gast
inet
al.
(201
1)3
2
–St
atic
cult
ure
and
con
tin
uou
spe
rfu
sion
–E
pid
erm
oid
can
cer
–H
um
anep
ider
moi
dca
rcin
oma
(A43
1.H
9)–
Lon
g-te
rmsp
her
oid
cult
ure
(7d
)–
Sph
eroi
dfo
rmat
ion
via
han
gin
gd
rops
Tu
ng
etal
.(2
011)
29
–M
icro
flu
idic
wel
lpl
ate
des
ign
(3D
Bio
mat
rix)
–C
olon
can
cer
–H
um
anco
lore
ctal
aden
ocar
cin
oma
(HT
-29,
HC
T-1
16)
–A
uto
mat
edw
orkf
low
wit
hm
icro
flu
idic
wel
lpl
ate
des
ign
(In
Sph
ero)
–Sp
her
oid
/sph
eric
alm
icro
tiss
ue
form
atio
nD
rew
itz
etal
.(2
011)
28
–Li
ver
can
cer
–H
um
anli
ver
carc
inom
a(H
epG
-2)
–Pr
osta
teca
nce
r–
Hu
man
pros
tate
can
cer
(DU
-145
)–
Lon
g-te
rmsp
her
oid
cult
ure
(14
d)
–M
icro
tiss
ue
stai
nin
gan
dgr
owth
prof
iles
–K
idn
eyca
nce
r–
Hu
man
kid
ney
carc
inom
a(A
-498
)–
Gli
obla
stom
a–
Hu
man
glio
blas
tom
a(S
NB
-19)
–D
rug
sen
siti
vity
–Pr
osta
teca
nce
r–
Hu
man
pros
tate
can
cer
(PC
-3)
–E
xtra
lon
g-te
rmsp
her
oid
cult
ure
(22
d)
faci
lita
ted
bym
icro
-rin
gs–
Sph
eroi
dfo
rmat
ion
via
han
gin
gd
rops
Hsi
aoet
al.
(201
2)3
0
–H
um
anu
mbi
lica
lco
rden
dot
hel
ial
(HU
VE
C)
–M
ouse
pre-
oste
obla
st(M
C3T
3-E
1)–
Mod
ifie
dm
icro
flu
idic
wel
lpl
ate
des
ign
Tu
mor
–str
omal
inte
ract
ion
–2D
–C
ervi
cal
can
cer
–H
um
ance
rvic
alca
nce
r(H
eLa)
–2D
co-c
ult
ure
sw
ith
det
ach
able
subs
trat
es–
Cel
lm
igra
tion
inco
-cu
ltu
reK
aji
etal
.(2
008)
39
–H
um
anu
mbi
lica
lco
rden
dot
hel
ial
(HU
VE
C)
–M
ouse
fibr
obla
st(N
IH-3
T3)
–Pr
osta
teca
nce
r–
Hu
man
pros
tate
can
cer
(PC
-3)
–D
iffu
sion
dom
inan
t2D
co-c
ult
ure
sw
ith
mic
rofl
uid
icco
mpa
rtm
ents
–O
steo
clas
toge
nes
isby
mon
ocyt
ed
iffe
ren
tiat
ion
Dom
enec
het
al.
(200
9)3
6
–M
ouse
mon
ocyt
e(R
AW
264.
7)
–Pr
osta
teca
nce
r–
Hu
man
pros
tate
aden
ocar
cin
oma
(LN
CaP
)–
Dif
fusi
ond
omin
ant
2Dco
-cu
ltu
res
wit
hm
icro
flu
idic
com
part
men
ts–
Hed
geh
ogsi
gnal
ing
inm
yofi
brob
last
san
def
fect
oftu
mor
grow
th
Dom
enec
het
al.
(201
2)3
8
–M
ouse
pros
tate
myo
fibr
obla
sts
(UG
SM-2
,U
Gli
3�/�
)–
Nor
mal
pros
tate
fibr
obla
sts
(NPF
,N
2-1,
N5-
2)
–M
ult
iple
mye
lom
a–
Hu
man
mu
ltip
lem
yelo
ma
(RPM
I822
6)–
Dif
fusi
ond
omin
ant
2Dco
-cu
ltu
res
wit
hm
icro
flu
idic
com
part
men
ts–
NF-kB
and
STA
T3
acti
vati
onin
MM
cell
sY
oun
get
al.
(201
2)4
2
–C
hro
nic
lym
phoc
ytic
leu
kem
ia(C
LL)
–H
um
anbo
ne
mar
row
stro
mal
(HS-
5)–
Hu
man
prim
ary
CLL
–Su
spen
sion
cell
cult
ure
enab
led
bylo
wsh
ear
stre
ssge
omet
ries
–Li
ver
can
cer
–H
um
anli
ver
carc
inom
a(H
epG
-2)
–2D
co-c
ult
ure
sw
ith
up
to3
cell
type
s–
Com
peti
tive
cell
mig
rati
onin
tri-
cult
ure
Ma
etal
.(2
012)
43
–Sa
liva
rygl
and
aden
oid
cyst
icca
rcin
oma
–H
um
anm
uco
saep
ith
elia
(GE
S-1)
–H
um
anad
enoi
dcy
stic
carc
inom
a(A
CC
-2/M
)–
Hu
man
embr
yon
iclu
ng
fibr
obla
st(H
FL-1
)
Integrative Biology Review Article
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online
This journal is c The Royal Society of Chemistry 2013 Integr. Biol., 2013, 5, 1096--1109 1101
Tab
le1
(con
tinu
ed)
Can
cer
mod
elC
ell
type
sN
otab
lem
icro
envi
ron
men
tfe
atu
res
Focu
sof
stu
dy
Ref
.
Tu
mor
–str
omal
inte
ract
ion
–3D
–B
reas
tca
nce
r–
Hu
man
brea
stca
rcin
oma
(MB
-MD
A-2
31)
–3D
Typ
eI
coll
agen
orM
atri
gel
mat
rix
–C
ell
mig
rati
onin
3Dge
lH
uan
get
al.
(200
9)4
1
–M
ouse
mac
roph
age
(RA
W26
4.1)
–3D
co-c
ult
ure
sw
ith
2ce
llty
pes
–E
CM
rem
odel
ing
–B
reas
tca
nce
r–
Hu
man
mam
mar
ygl
and
epit
hel
ial
(MC
F-D
CIS
.com
)–
3DT
ype
Ico
llag
en+M
atri
gel
mix
edge
l–
DC
IStr
ansi
tion
toin
vasi
ved
uct
alca
rcin
oma
Sun
get
al.
(201
1)4
8
–H
um
anm
amm
ary
fibr
obla
st(H
MF)
–3D
co-c
ult
ure
sw
ith
2ce
llty
pes
–Lu
ng
can
cer
–H
um
anlu
ng
fibr
obla
st(M
RC
-5)
–D
irec
tion
alpa
racr
ine
sign
alin
gvi
ad
ynam
icva
lve
con
trol
–T
GF-b
sign
alin
gH
suet
al.
(201
1)3
7
–H
um
anlu
ng
aden
ocar
cin
oma
(CL1
-0)
–M
yofi
brob
last
acti
vati
on
An
giog
enes
is–
Bre
ast
can
cer,
glio
blas
tom
a–
Rat
mam
mar
yad
enoc
arci
nom
a(M
TLn
3)–
3DT
ype
Ico
llag
enm
atri
x–
En
dot
hel
ial
mon
olay
erm
igra
tion
Ch
un
get
al.
(200
9)5
3
–H
um
angl
iobl
asto
ma
(U87
MG
)–
VE
GF
grad
ien
t–
Hu
man
der
mal
mic
rova
scu
lar
end
oth
elia
l(H
MV
EC
)–
Hor
izon
tal
2Dm
onol
ayer
–M
ouse
fibr
obla
st(1
0T1/
2)
–N
/Aa
–H
um
anu
mbi
lica
lco
rden
dot
hel
ial
(HU
VE
C)
–3D
coll
agen
/fib
ron
ecti
nm
atri
x–
Flow
-in
du
ced
angi
ogen
icsp
rou
tin
gSo
ng
and
Mu
nn
(201
1)5
4
–C
ontr
olle
dsh
ear
stre
ssan
din
ters
titi
alfl
ow–
VE
GF
grad
ien
ts–
Hor
izon
tal
and
vert
ical
2Dm
onol
ayer
s
–N
/A–
Hu
man
der
mal
mic
rova
scu
lar
end
oth
elia
l(H
MV
EC
)–
3DT
ype
Ico
llag
enm
atri
x–
An
giog
enic
spro
uti
ng
and
tip
cell
mig
rati
onin
mu
ltip
legr
adie
nts
Shin
etal
.(2
011)
55
–V
EG
Fan
dA
NG
-1gr
adie
nts
–V
erti
cal
2Dm
onol
ayer
–N
/A–
Hu
man
um
bili
cal
cord
end
oth
elia
l(H
UV
EC
)–
3DT
ype
Ico
llag
en+M
atri
gel
mix
edge
l–
An
giog
enic
spro
uti
ng
in3D
co-c
ult
ure
Bis
chel
etal
.(2
012)
56
–M
ouse
fibr
obla
st(1
0T1/
2)–
VE
GF
grad
ien
ts–
Cir
cula
r3D
lum
ens
–N
/A–
Hu
man
um
bili
cal
cord
end
oth
elia
l(H
UV
EC
)–
3DT
ype
Ico
llag
enm
atri
x–
En
dot
hel
ial-p
eric
yte
inte
ract
ion
and
mic
rova
scu
lar
form
atio
n
Zhen
get
al.
(201
2)5
7
–H
um
anbr
ain
vasc
ula
rpe
ricy
tes
–G
ravi
ty-d
rive
npe
rfu
sion
–C
ircu
lar
3Dlu
men
s–
Bar
rier
perm
eabi
lity
–Le
uke
mia
,gl
iobl
asto
ma
–H
um
anle
uke
mia
cell
s(H
L-60
)–
3Dco
llag
en/f
ibri
nm
atri
x–
An
giog
enic
spro
uti
ng
Kim
etal
.(2
013)
58
–H
um
angl
iobl
asto
ma
cell
s(U
87M
G)
–3D
co-c
ult
ure
sbe
twee
nen
dot
hel
ial
cell
s,st
rom
alfi
brob
last
s,pe
ricy
tes
and
can
cer
cell
s–
Perf
usi
onof
mic
robe
ads
thro
ugh
pate
nt
mic
rove
ssel
net
wor
k–
Hu
man
um
bili
cal
cord
end
oth
elia
l(H
UV
EC
)–
Hu
man
plac
enta
peri
cyte
s–
Perf
usi
onin
mic
rove
ssel
s–
Hu
man
lun
gfi
brob
last
s(L
F)
Oxy
gen
Gra
die
nts
–N
/A–
Rat
myo
fibr
obla
sts
(C2C
12)
–2D
mon
ocu
ltu
reof
myo
fibr
obla
sts
onm
icro
elec
trod
es–
Hyp
erox
ia-i
nd
uce
dap
opto
sis
Park
etal
.(2
006)
62
–O
xyge
ngr
adie
nt
byel
ectr
olys
is
–N
/A–
Rat
myo
fibr
obla
sts
(C2C
12)
–2D
mon
ocu
ltu
re–
Oxy
gen
con
sum
ptio
nby
myo
fibr
obla
sts
Meh
taet
al.
(200
7)6
1
–C
onti
nu
ous
perf
usi
onvi
ad
ynam
icva
lve
con
trol
–O
xyge
ngr
adie
nt
bypa
ssiv
eco
nsu
mpt
ion
Tab
le1
(co
nti
nu
ed)
Review Article Integrative Biology
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

1102 Integr. Biol., 2013, 5, 1096--1109 This journal is c The Royal Society of Chemistry 2013
Tab
le1
(con
tinu
ed)
Can
cer
mod
elC
ell
type
sN
otab
lem
icro
envi
ron
men
tfe
atu
res
Focu
sof
stu
dy
Ref
.
–Lu
ng
can
cer
–C
arci
nom
ich
um
anal
veol
arba
sal
epit
hel
ial
cell
s(A
549)
–2D
mon
ocu
ltu
re–
Oxy
gen
grad
ien
tby
oxyg
en-g
ener
atin
gan
dox
ygen
-sca
ven
gin
gre
acti
ons
–H
yper
oxia
-in
du
ced
apop
tosi
s–
Hyp
oxia
-in
du
ced
cyto
toxi
city
byti
rapa
zam
ine
Ch
enet
al.
(201
1)6
3
Inva
sion
and
Met
asta
sis
–B
reas
tca
nce
r–
Hu
man
brea
stca
nce
r(M
CF-
7;n
on-m
etas
tati
c)–
2Dm
onoc
ult
ure
–C
ell
mig
rati
on,
inva
sion
thro
ugh
mic
roga
psC
haw
etal
.(2
007a
)70
–H
um
anbr
east
carc
inom
a(M
B-M
DA
-231
)–
3Dm
atri
gel
mat
rix
coat
ing
inm
icro
gaps
–B
reas
tca
nce
r–
Hu
man
brea
stca
rcin
oma
(MD
A-M
B-4
35S)
–2D
mon
ocu
ltu
re–
Cel
lm
igra
tion
and
inva
sion
thro
ugh
mic
roga
ps
Ch
awet
al.
(200
7b)6
9
–Li
ver
can
cer
–H
um
anli
ver
carc
inom
a(H
epG
-2)
–3D
mat
rige
lm
atri
xco
atin
gin
mic
roga
ps–
Cer
vica
lca
nce
r–
Hu
man
cerv
ical
can
cer
(HeL
a)–
Subc
onfl
uen
ten
dot
hel
ial
lin
ing
acro
ssm
icro
gaps
–H
um
anm
icro
vasc
ula
ren
dot
hel
ial
(HM
EC
)
–B
reas
tca
nce
r–
Hu
man
brea
stca
rcin
oma
(MD
A-2
31)
–3D
hyd
roge
lm
atri
x–
Tu
mor
–en
dot
hel
ial
inte
ract
ion
san
d3D
intr
avas
atio
n
Zerv
anto
nak
iset
al.
(201
2)7
1
–Fi
brob
last
icsa
rcom
a–
Hu
man
fibr
osar
com
a(H
T10
80)
–3D
cocu
ltu
res
betw
een
end
oth
elia
lce
lls,
mon
ocyt
esan
dca
nce
rce
lls
–H
um
anu
mbi
lica
lco
rden
dot
hel
ial
(HU
VE
C)
–H
um
anm
icro
vasc
ula
ren
dot
hel
ial
(MV
EC
)–
Hor
izon
tal
and
vert
ical
2Dm
onol
ayer
s–
Mou
sem
onoc
yte
(RA
W26
4.7)
Com
bin
atio
n–
Col
onca
nce
r–
Hu
man
colo
nca
rcin
oma
(LS1
74T
)–
Lon
g-te
rmsp
her
oid
cult
ure
(4d
)–
Com
bin
edsp
her
oid
sw
ith
grad
ien
tsW
alsh
etal
.(2
008)
60
–D
iffu
sed
grad
ien
tsof
dox
oru
bici
n
–Sa
liva
rygl
and
aden
oid
cyst
icca
rcin
oma
–H
um
anad
enoi
dcy
stic
carc
inom
a(A
CC
-M)
–3D
mat
rix
subs
titu
te(b
asem
ent
mem
bran
eex
trac
t)–
CA
F-m
edia
ted
tum
orce
llin
vasi
onLi
uet
al.
(201
0)3
5
–H
um
anem
bryo
nic
lun
gfi
brob
last
(HFL
-1)
–Pr
imar
yis
olat
edca
rcin
oma-
asso
ciat
edfi
brob
last
s(C
AFs
)–
3Dsp
her
oid
sin
co-c
ult
ure
wit
hn
eigh
bori
ng
CA
Fs–
Com
bin
edsp
her
oid
sw
ith
stro
mal
inte
ract
ion
–B
reas
tca
nce
r–
Mou
sem
amm
ary
tum
or(4
T1)
–3D
Mat
rige
lm
atri
x–
Hyp
oxia
-dep
end
ent
mig
rati
onof
tum
oran
den
dot
hel
ial
cell
s
Gao
etal
.(2
011)
40
–H
um
and
erm
alm
icro
vasc
ula
ren
dot
hel
ial
(HD
ME
C)
–C
onst
ant
med
iape
rfu
sion
–In
du
ced
hyp
oxia
byC
oCl 2
–A
ttem
pted
mod
elof
com
bin
edh
ypox
iaan
dan
giog
enes
is
aN
/A–
not
appl
icab
leor
not
spec
ifie
d.
Tab
le1
(co
nti
nu
ed)
Integrative Biology Review Article
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

This journal is c The Royal Society of Chemistry 2013 Integr. Biol., 2013, 5, 1096--1109 1103
can be incorporated to create additional channel compartmentsthat permit indirect perfusion, which has been applied to yieldformation of coculture spheroids with as many as three celltypes to model tumor heterogeneity within metastatic prostatecancer (Fig. 1a).33 Besides providing perfusion and physiologicalflow,32 microfluidic systems offer the potential for more advancedin vitro models of the microenvironment, where spheroids can beembedded within a 3D matrix,34 and can even support the cultureof surrounding carcinoma-associated fibroblasts (CAFs) to allowstudies on tumor–stromal interactions.35 These examples offerevidence that we are beginning to witness more integration oftumor spheroid cultures into microfluidic systems.
Tumor–stromal interactions
A large body of work within the area of microfluidic systems hasfocused on leveraging the high spatial resolution afforded bymicrofabrication techniques to create precise structures andarrangements for more complex in vitro coculture models tostudy tumor–stromal interactions. Coculture models havetraditionally been achieved by (i) placing one cell type in directphysical contact to the second cell type, with one forming amonolayer of underlying ‘‘feeder’’ cells, or (ii) using Transwellmembrane inserts to compartmentalize two cell types whileallowing soluble factor signaling through the membrane. WhileTranswell inserts are useful for decoupling paracrine andmechanical signals that are coupled in cultures having directcell–cell contacts, Transwell-based assays are static, are limitedto two compartments, and the typical culture volumes anddiffusion distances of these assays often lead to significantdilution of factors, and loss of effectiveness of factors withshort half-lives. Microfluidic systems, even with the most basicdesigns, can achieve improved spatial organization, increasedlevel of compartmentalization, and more controlled diffusion offactors than Transwell inserts, leading to improved sensitivity,36
and importantly, more control over the dynamics of the micro-environment, including fluid flow. Examples of microfluidiccoculture models for tumor–stromal interactions are quitevaried, and include: lung cancer37 and prostate cancer cells38
cocultured with myofibroblasts; cervical cancer cells withfibroblasts;39 breast cancer cells with either endothelial cells40
or macrophages;41 and for hematologic cancers in the bonemarrow microenvironment, multiple myeloma cells with bonemarrow stromal cells.42 While these examples demonstrateimproved compartmentalization, including a case with threeseparated cell types,43 as well as improved dynamic control viamicrovalves,37 the majority (except for ref. 41) have focused onparacrine signaling at the cellular level in 2D, neglecting theinfluence of dimensionality and 3D cell–matrix interactions atthe tissue and organ levels. Nevertheless, microfluidic coculturesystems offer an alternative to existing coculture assays withadvantages that motivate continued interest in this area.
ECM and three-dimensionality
The addition of ECM components to create a 3D structuralframework for cells in culture adds mechanical context anddimensionality to the in vitro microenvironment. While 2D
compartmentalized cocultures are advantageous because theyare simple to use, have straightforward readouts, and areamenable to high-throughput applications, there is little doubtas to the importance of the third dimension in culture,26
and the role of mechanobiology in tumorigenesis.44,45 Tumorgrowth is associated with biomechanical alterations in themicroenvironment, including increased solid stresses in theneighboring tissue, increased matrix stiffness, and aberrantinterstitial fluid flow.46 In addition, matrix remodeling andother mechanobiological cues affect the invasiveness and meta-static potential of tumor cells.47 Tumor spheroids, discussedabove, is one approach to achieve three-dimensionality, but themorphology of spheroids does not necessarily apply to alltumor microenvironments, nor does it represent the optimalform for testing and experimentation. Various microfluidicsystems have incorporated 3D matrix components and hydrogelsinto compartments, and thus advanced 2D cells-on-chip technol-ogy toward 3D tissues-on-chip. By incorporating collagen-basedgels, cancer cell invasion can be monitored in 3D, and matrixremodeling can be visualized by second harmonic generationimaging of the changing collagen structure.41 This can now beachieved quite easily with pipette-based passive pumping into asimple Y-shaped microchannel (Fig. 1b).48 The gel has the capa-city to support stromal cells such as mammary fibroblasts, andadditional microchannel geometries can also be incorporated tomodify diffusion distances, allowing greater spatial controlbetween cell types, even in 3D. Given the mounting evidencetoward the importance of 3D culture, and the continued advance-ment of technology to facilitate its adoption, future microfluidicsystems will likely shift even more toward 3D cultures.
Angiogenesis
One feature of the tumor microenvironment that requiresspecialized 3D tissue structure beyond the simple addition ofa third dimension is the network of angiogenic blood vesselsthat supply oxygen and nutrients to the tumor. While stromalfibroblasts and immune cells can be simply embedded into 3Dgels by re-suspending in the matrix solution prior to loading,endothelial cells must form confluent monolayers, withappropriate formation of cell–cell junctions and basementmembrane layers, in tubular lumen structures in order toresemble the actual structure of blood vessels. 2D endothelialmonolayers have been popular for decades for studying flow-induced shear stress on vascular endothelial cells. Thisapproach has been applied to enable shear stress studies onendothelial cells in microfluidic systems that promised toincrease throughput, lower reagent consumption, and reducesample quantities.49,50 Recently, microfluidic cell culturesystems have advanced toward endothelial-lined blood vesselmimics that are potentially useful for cancer-related angiogen-esis studies.51 Many of these microsystems employed simple 2Dendothelial monolayers as before, but in a vertical orientationto allow visualization of horizontal angiogenic vessel growthinto hydrogels.52–55 One design in particular superposed VEGFgradients and shear flow over the vertical monolayers to studyflow-induced angiogenic sprouting.54 To create more advanced
Review Article Integrative Biology
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

1104 Integr. Biol., 2013, 5, 1096--1109 This journal is c The Royal Society of Chemistry 2013
3D lumens with cylindrical cross-section, a simple viscousfingering method via micropipetting was employed in onedesign,56 while an elaborate microfluidic assembly was con-structed in another design to house a slab of collagen gel withmolded microfeatures for the seeding and remodeling ofendothelial-lined microvessels.57 Interestingly, by embeddingthe collagen gel with pericytes in this latter design, endothelialcells were able to recruit pericytes to the microvessel walls, andan appropriate basal lamina between pericytes and endothelialcells was allowed to form. This was also accompanied byangiogenic sprouting into the collagen gel slab. Altogether, thisexample demonstrates how complex interplay between distinctmicroenvironmental cues can lead to natural remodeling of amicrovessel network that displays many in vivo-like characteri-stics, some of which can be applied to the engineering of atumor microenvironment. Others have since been able to createfull angiogenic microvessel networks in a 3D chip (Fig. 1c) thatcan be cocultured with different cell types and be perfused withmicrobeads,58 showing further advancement in complexity formodels involving angiogenesis.
Gradients
In addition to aspects of the microenvironment related tocellular organization, dimensionality, ECM architecture, andmorphological properties, microfluidic systems are also naturallywell suited for generating and maintaining spatiotemporal gradi-ents of biochemical and physicochemical components. Becauseof well-recognized laminar flow characteristics in low-Reynoldsnumber microchannel flows, microfluidic gradient generatorswere one of the first and most popular types of systems developedin the field.59 Several microfluidic angiogenesis systems haveemployed gradients of VEGF to induce sprouting,53,54,56 includingone in particular that used a combination of VEGF and ANG-1gradients to examine cooperative effects between the two solublefactors.55 Furthermore, it is possible to superpose gradients ontumor spheroids to further add complexity to the spheroidmodel.60 Aside from biochemical gradients, the control of oxygengradients have also been demonstrated as a way to model hypoxicconditions within the tumor. Oxygen gradients can either beproduced passively by allowing oxygen consumption by cellscultured in the microchannels,61 or actively by controlling waterelectrolysis via patterned electrodes,62 or by oxygen-generatingand oxygen-scavenging chemical reactions (Fig. 1d), the latter ofwhich was used to investigate oxygen tension on alveolar basalcarcinoma cells.63 The power of microfluidics will become evenmore evident as we continue to create more complex micro-environments through superposition of multiple gradients, aswell as other microenvironment factors.
Metastases
While the focus of this section is to highlight advances specificto the tumor microenvironment, it should be noted thatmicrofluidic systems have also been valuable for studies relatedto invasion, intravasation, extravasation, and metastasis, stagesof tumor progression where the events have begun to differ-entiate, or are differentiated, from the primary tumor region.
Steps of the invasion-metastasis cascade have thus far beenchallenging to examine in detail with in vitro systems, withTranswell inserts again being a popular choice as an invasionassay.64,65 Metastasis studies have almost exclusively beenconducted with in vivo mouse models because of the lack ofin vitro methods.66–68 Recently, a few microfluidic systems weredeveloped to study specific dynamic events within the metastasiscascade, including migration through gels and transmigrationthrough subconfluent endothelial linings,69,70 and real-timeimaging of invasion and extravasation steps with insight intotumor cell–endothelial cell interactions (Fig. 1e).71 Given theenormous challenges that remain in elucidating mechanismsof metastasis, these examples are likely just the beginning formicrofluidic metastasis systems, with much research needed tofurther improve models.
In summary, numerous studies of microfluidic systems,from cells-on-chips to tissues-on-chips, have been performedto date to tackle one specific aspect of the tumor micro-environment. Only a few systems have attempted to tackle morethan one aspect in the same device (Table 1, ‘‘Combination’’).35,40,60
Much research is necessary to further advance cells-, tissues-,and organs-on-chips toward even more complex micro-environments. Doing so will likely allow us to pursue our goalof acquiring more knowledge and answering more outstandingquestions in cancer research.
4. Cancer hallmarks: longstandingchallenges meet new opportunities
Our understanding of the biology of cancer has been crystal-lizing gradually over the years with the recognition that amidstall its complexity, cancer can be described in terms of basicunderlying principles that revolve around six essential acquiredcapabilities of cancer cells. These so-called ‘‘hallmarks of cancer’’,namely (i) the self-sufficiency in growth signals, (ii) insensitivity toanti-growth signals, (iii) ability to evade apoptosis, (iv) sustainedangiogenesis, (v) limitless replicative potential, and (vi) tissueinvasion and metastasis, were first summarized in 2000,72
and then subsequently revisited a decade later in an updatedreview in 2011.15 The two landmark reviews have not onlyserved as a guide to cancer research, but have also providedtwo mileposts on the cancer research timeline that offers avaluable perspective of the progress made in the interveningyears. Since 2000, new insights on cancer stem cells have beendiscovered; the importance of tumor heterogeneity has beenrecognized; major progress has been made in understandingEMT, invasion, and metastasis; and two new ‘‘emerging hall-marks’’ and ‘‘enabling characteristics’’ have been identified.15
Of particular relevance to this review is the clearly emergentrole of the tumor microenvironment, a topic which seemed topervade the discussion across all the different hallmarks. Thefact that these recognized hallmarks are essential for tumordevelopment, yet remain inadequately understood, providesmajor challenges on one hand, but may also present newresearch opportunities on the other. To provide a glimpse of
Integrative Biology Review Article
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

This journal is c The Royal Society of Chemistry 2013 Integr. Biol., 2013, 5, 1096--1109 1105
the types of opportunities that may be available, a brief journeyis made through some outstanding problems associated withthese well-known hallmarks.
The ability of cancer cells to simultaneously (i) sustain theirown growth and proliferation, (ii) evade anti-growth signals,and (iii) resist cell death, three distinct yet connected acquiredhallmarks of cancer, demonstrates their commitment to breakthe delicate balance of cell population demographics that iscrucial to homeostasis. Notably, such population demo-graphics, which encompass different cell types, their numbers,and their locations, are important to the spatial and temporalpatterns of regulatory signals governing proliferation, apoptosisand necrosis, and vice versa. The fact that our knowledge andunderstanding of the precise signals, their interactions, and themechanisms regulating those interactions remains incompletemay be attributed to difficulties in reconstructing and testingin vitro cellular microenvironments that capture the properdemographics found in vivo. Furthermore, it is suggested thatthe phenomenon of contact inhibition commonly observed in2D culture may be related to mechanisms in vivo that controlcell numbers. These problems may be difficult to address withconventional cultures, but the ability to use 3D cultures withimproved spatial patterning of cell populations in microfluidicsystems provides a method that can potentially resolve thesecomplex interactions, and shed light on the importance ofpopulation demographics.
The acquired capability of cancer cells to avoid senescenceand effectively become immortalized with ‘‘limitless replicativepotential’’ may involve molecular factors of the tumor milieuthat have yet to be fully identified. Experimentally, it has alsobeen suggested that culture conditions may be tuned in amanner that can influence the onset of cell senescencein vitro.15 Microscale systems can be applied either (i) to createtumor microenvironments suitable for testing different bio-chemical factors on normal cells in an accurate physiologiccontext and measuring their replicative potential, possibly bymonitoring cell divisions within the engineered micro-environments, or (ii) to test various culture conditions withdifferent microchannel dimensions, media components, serumand glucose levels, in a high-throughput, physiologically rele-vant manner to elucidate how culture conditions affect cellsenescence. In both cases, the flexibility of microfluidic systemscan be leveraged to help elucidate the role of the micro-environment, and the culture conditions within it.
One cancer hallmark that has already received considerableattention in the microfluidics community is angiogenesis.Paradoxically, in vitro models of angiogenesis are becomingincreasingly sophisticated in some cases,57 yet also becomingmore routine and accessible in others.56 Both of these 3Dcoculture models have potential to elucidate important rolesfor stromal and perivascular cells in angiogenic sprouting.Given that the proper microvascular structure composed ofprecise apposition between endothelial cells and pericytes canbe faithfully recapitulated in vitro as demonstrated, new ques-tions can be posed regarding the effect of pericytes on endo-thelial barrier function, leakiness, and the susceptibility of the
vessel wall to intravasation and extravasation. Real-time moni-toring of engineered angiogenesis systems may allow us to tracethe origin, progression, and/or transdifferentiation of normalendothelial cells to tumor-associated endothelial cells, or trackthe interactions between endothelial tip and stalk cells withinangiogenic sprouts. As more complex in vitro models become areality, it may be possible to combine tumor spheroids andangiogenic microvessels on the same microchip, allowing thesimultaneous monitoring of tumor growth and neovascularrecruitment, and investigations into vascular normalization atthe cellular level.73
While much progress has been made over the past decade inunderstanding invasion and metastasis, it is notable that manychallenging questions remain unanswered.15 Appropriately,these questions present interesting opportunities for micro-fluidic systems to serve as novel experimental platforms thatcan offer fresh perspectives on the multistep metastatic pro-cess. For example, paracrine signals from stromal cells of thetumor microenvironment may be involved in activating a panelof transcription factors that coordinate EMT. Moreover, aspatial pattern exists where cancer cells that have undergoneEMT appear more frequently near invasive margins of thetumor rather than near the tumor core, suggesting that spatialposition and the local microenvironment are important factorsof the EMT process. In both cases, spatial control offered bymicrofluidic systems may be exploited. Since dissemination oftumor cells from the primary site often occurs via the lymphaticsystem, research is also needed to create in vitro models oflymphatic vessels for potential application in studies of lymphaticmetastasis. Additionally, a major phase within metastasis thatis particularly fraught with experimental challenges is coloniza-tion, the development of micrometastases into secondarytumors. Interesting and unanswered questions related to dormancyand mechanisms of the seed-soil hypothesis must inherentlyinvolve both the seed (i.e., circulating tumor cell (CTC) originat-ing from the primary tumor) and the soil (i.e., the tissue bed ofthe distant organ on which the CTC settles). While isolationand handling of CTCs may be the subject of other microfluidicmanipulations,74,75 development of engineered tissue beds thatserve as the soil in colonization experiments may be attainablewith advances in organ-on-chip technologies. This work willlikely be essential to the successful modeling of metastaticcolonization in vitro.
An important topic that has been identified as both anemerging hallmark and an enabling characteristic of canceris inflammation.15 The incorporation of inflammatory cells intoin vitro microfluidic coculture systems was reviewed for severalexamples involving macrophages (Table 1), providing a glimpseof the potential utility of biochips to understanding tumor–stromal interactions involving other inflammatory cell typessuch as neutrophils and lymphocytes. In fact, other micro-fluidic systems developed to study neutrophil migration forother diseases76 can potentially be modified for cancer studies.Using these systems, the signals between tumor cells andimmune cells can be monitored in real-time, and used toelucidate the temporal dynamics of the inflammatory response.
Review Article Integrative Biology
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

1106 Integr. Biol., 2013, 5, 1096--1109 This journal is c The Royal Society of Chemistry 2013
The other emerging hallmark is the ability for cancer cells toreprogram metabolism, and this is an area that can benefitsignificantly from the application of microfluidic systems.Much research is currently underway on the development ofanalytical methods for metabolomics in microfluidic systems,with the goal of being able to sample, detect, and measurequantities of metabolites in high throughput.77–79 Within thesesystems, the front-end cultures are so far basic cell cultureswithout elaborate microenvironment features, but potentialdefinitely exists to incorporate more sophisticated physiologicmodels that can then be sampled downstream for metabolitesin the sample media. Since the most commonly used materialfor microfluidic devices is poly(dimethylsiloxane) (PDMS), andit is well-known to absorb hydrophobic molecules from themedia,80 other device materials such as polystyrene (PS) andother thermoplastics may need to be considered for any studyinvolving the measure of metabolites. In addition, recentresearch has shown that cells in microculture are underincreased cell stress due to metabolic constraints,81 indicatingthat improved understanding of cellular metabolism in micro-fluidic cultures is fundamentally important to the future micro-environment studies on the effects of metabolism in cancer.
5. Remaining technical challenges
While many outstanding questions await cancer researchers, asoutlined above, it should be emphasized that outstandingchallenges also remain for microfluidic technologists. Someof these challenges are more technical, and apply to micro-fluidic technologies in general, while others are more prag-matic, and apply specifically to chips designed for cancer tumormicroenvironments. Regardless, these challenges must first beovercome at the technology end, if biochips are to realize theirfull potential in providing a new and useful platform forresearch in cancer and the tumor microenvironment. Thefollowing discussion aims to synthesize the various challengesfacing engineers, biologists, and those working at the interface,and importantly start the dialogue needed to further advancethe application of biochip technologies.
Starting from a technical standpoint, what remains at thecore of engineered microsystems is microfabrication. Con-tinued research in microfabrication methods is necessary forfuture advancements in biochips, and it simply cannot beneglected. The fact that soft lithography and PDMS offer manymore pros than cons has been demonstrated time and again,but this fact should not limit us from pursuing innovativefabrication methods that may be faster, cheaper, more reliable,and/or more amenable to high volume manufacturing.Although polystyrene (PS) offers familiarity to biologists, andis gaining recognition in the microfluidics world,82,83 PS has itsshortcomings as well, as its high modulus of elasticity pre-cludes its use in applications that require deformations in thedevice structure, such as for a breathing lung-on-a-chip.4 Never-theless, more research on how to fabricate microdevices in PSand other thermoplastics will still be valuable, both to serve asa practical alternative to PDMS in academic research, and to
offer insights on how to potentially enable mass manufacturingcapabilities for future commercialization. Besides selection ofdevice material, other technical issues such as throughput,world-to-chip interfacing, and the cost-benefit ratio of inte-grating additional functionality are engineering challenges thatmust be considered for each specific application, and will nodoubt be an important research focus if organs-on-chip plat-forms, particularly for cancer drug screening and personalizedmedicine, are to become a reality.
While engineers will need to commit some effort to advan-cing device fabrication, they must also commit to designing for(i) user operability and (ii) endpoint measurements, aspectsthat are likely of greater concern to the biologists and clinicianswho will actually be using the tools to acquire useful data. Themajority of the microfluidic systems reviewed here were proof-of-concept designs, many with primitive prototyping setupsthat do not translate immediately and conveniently into biologylaboratories. Operability may not be a high priority duringprototyping and proof-of-concept stages, but because itbecomes critical during translation and adoption stages, it isprudent to design operability directly into the system as early aspossible during development.
Also, more research is needed toward more endpointmeasurements. The majority of microsystems discussed so farhave relied heavily on microscopy to acquire endpoint readouts.This is reasonable and desirable given that direct visualization ofcells and their behavior, perhaps with different dyes and labels,offers the most reliable and direct proof that cells are viable andfunctioning as expected in these systems. Furthermore, func-tional readouts such as the number and length of angiogenicsprouts, or the migration rates of cells or monolayer fronts, canonly be detected and measured via microscopy. However, muchinformation at the molecular level that cannot be acquired bymicroscopy has yet to be uncovered in many microfluidic cellculture systems. Some microscale platforms allow cell lysis sothat contents can be collected for qRT-PCR and western blottingoff-chip;38,84,85 and a combination of immunocytochemicalstaining and protein localization analysis can be employed toquantify nuclear translocation as a substitute for electrophoreticmobility shift assays (EMSAs) used to study transcription factoractivation;42 Moreover, other platforms have achieved single-cellresolution qPCR performed completely on-chip.86 However,it remains a challenge to harvest cells cultured from micro-fluidic systems with more complex 3D microenvironments. Forexample, it would be useful and powerful to have the ability toisolate and independently harvest the endothelial cells fromangiogenic vessels without contaminating the sample withneighboring cocultured stromal cells, but this has yet to bedemonstrated. In addition, an ongoing challenge has been todetect and quantify secreted factors from microcultures. While thishas been shown as proof-of-concept with simple microcultures,87,88
more research is necessary to demonstrate this for complextumor microenvironments.
The interdisciplinary nature of our current discussionmeans inherently that the challenge not only exists from atechnical perspective, but also from a pragmatic perspective.
Integrative Biology Review Article
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

This journal is c The Royal Society of Chemistry 2013 Integr. Biol., 2013, 5, 1096--1109 1107
Even if the technical hurdles are overcome, many questionsawait regarding the application of the technology in cancerresearch. For example, assuming the technology has reached alevel of maturity and robustness sufficient for serious applica-tions in biology beyond the proof-of-concept, what tumors andtumor microenvironments are the most interesting and imme-diately important to develop in vitro? Lung, breast, prostate,colon, and liver cancers appear to be excellent candidates tostart given that they have the highest mortality rates, and alsobecause lung and gut-on-a-chip devices have already beenreported.4,5 The bone marrow microenvironment is also ofinterest because, as mentioned above, it is responsible forproviding the niche within which the class of hematologiccancers are developed. Furthermore, the bone microenvironmentalso serves as a major site for metastases of the lung, breast,and prostate,89 and thus will be an important model for on-chipmetastasis studies.
Another question is how much complexity is required alongthe cell-, tissue-, and organ-level spectrum to yield useful,functional readouts that can offer new insights in humandiseases like cancer. There is clearly a compromise betweenhaving more complex physiologic microenvironments withmore convoluted interactions and less straightforward readoutsversus less physiologic context, and thus less accuracy, in themicroenvironment model, but more control over interactionsand more basic but similarly useful readouts. This question canonly be addressed by rigorous validation and comparisonbetween in vitro systems offering varying levels of complexity,in addition to comparisons with in vivo animal models thatserve as positive controls.
Regardless of physiologic complexity, the basic units of anybiochip are the basic units of life itself: the cells. While themajority of microdevices have demonstrated feasibility usingimmortalized cell lines or commercially available primary cellslike HUVECs, the promise that organs-on-chips can signifi-cantly improve drug screens and provide a future involvingpersonalized medicine depends on our ability to integrate intothe chip technologies primary human cells obtained directlyfrom individuals. This issue, and the others described before it,may not be specific to cancer research, but they should beemphasized for their importance to the successful technologicaladvancement of these systems for cancer research, and otherresearch areas.
Summary
The ability for cells-, tissues-, and organs-on-chips to recapitulatecomplex and controlled microenvironments enables them tooffer new and innovative approaches to cancer research and thestudy of the role of the tumor microenvironment. As evidencedby the extensive literature already amassed in the last severalyears on microfluidic systems for tumor microenvironmentstudies, the field is growing at a remarkable rate, and keepingpace is a challenge itself. Both cancer research and micro-fluidics technology development continue to face numerouschallenges, but the intersection of these fields provides a major
opportunity for significant impact in both areas, with potentialfor scientific breakthroughs. The impact of organs-on-chips oncancer research may not be apparent in the next few years, butif the current rate of progress is any indication, interestingdevelopments are certainly on the horizon.
Acknowledgements
The author acknowledges financial support from the NaturalSciences and Engineering Research Council of Canada (NSERC)through a Discovery Grant. The author also apologizes for anyunintended omissions of pertinent literature in the currentdiscussion.
References
1 P. Li and D. Harrison, Anal. Chem., 1997, 69, 1564–1568.2 D. Beebe, G. Mensing and G. Walker, Annu. Rev. Biomed.
Eng., 2002, 4, 261–286.3 P. J. Lee, P. J. Hung and L. P. Lee, Biotechnol. Bioeng., 2007,
97, 1340–1346.4 D. Huh, B. D. Matthews, A. Mammoto, M. Montoya-Zavala,
H. Y. Hsin and D. E. Ingber, Science, 2010, 328, 1662–1668.5 H. J. Kim, D. Huh, G. Hamilton and D. E. Ingber, Lab Chip,
2012, 12, 2165–2174.6 A. Rapisarda and G. Melillo, Nat. Rev. Clin. Oncol., 2012, 9,
378–390.7 M. Egeblad, E. S. Nakasone and Z. Werb, Dev. Cell, 2010, 18,
884–901.8 D. Huh, G. A. Hamilton and D. E. Ingber, Trends Cell Biol.,
2011, 21, 745–754.9 C. Moraes, G. Mehta, S. C. Lesher-Perez and S. Takayama,
Ann. Biomed. Eng., 2012, 40, 1211–1227.10 D. Wlodkowic and J. M. Cooper, Curr. Opin. Chem. Biol.,
2010, 14, 556–567.11 H. Ma, H. Xu and J. Qin, Biomicrofluidics, 2013, 7, 011501.12 S. Paget, Lancet, 1889, 133, 571–573.13 I. Fidler, Nat. Rev. Cancer, 2003, 3, 453–458.14 T. D. Tlsty and L. M. Coussens, Annu. Rev. Pathol.: Mech.
Dis., 2006, 1, 119–150.15 D. Hanahan and R. A. Weinberg, Cell, 2011, 144, 646–674.16 N. A. Lobo, Y. Shimono, D. Qian and M. F. Clarke, Annu. Rev.
Cell Dev. Biol., 2007, 23, 675–699.17 R. A. Weinberg, The Biology of Cancer, Garland Sciences,
New York, NY, 2007.18 A. Ostman and M. Augsten, Curr. Opin. Genet. Dev., 2009, 19,
67–73.19 P. Iyengar, V. Espina, T. W. Williams, Y. Lin, D. Berry,
L. A. Jelicks, H. Lee, K. Temple, R. Graves, J. Pollard,N. Chopra, R. G. Russell, R. Sasisekharan, B. J. Trock,M. Lippman, V. S. Calvert, E. F. Petricoin 3rd, L. Liotta,E. Dadachova, R. G. Pestell, M. P. Lisanti, P. Bonaldo andP. E. Scherer, J. Clin. Invest., 2005, 115, 1163–1176.
20 K. S. Siveen and G. Kuttan, Immunol. Lett., 2009, 123, 97–102.21 B. Vogelstein and K. W. Kinzler, Nat. Med., 2004, 10,
789–799.
Review Article Integrative Biology
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

1108 Integr. Biol., 2013, 5, 1096--1109 This journal is c The Royal Society of Chemistry 2013
22 M. Dong and G. C. Blobe, Blood, 2006, 107, 4589–4596.23 J. A. Burger, P. Ghia, A. Rosenwald and F. Caligaris-Cappio,
Blood, 2009, 114, 3367–3375.24 E. W. K. Young and D. J. Beebe, Chem. Soc. Rev., 2010, 39,
1036–1048.25 G. Mehta, A. Y. Hsiao, M. Ingram, G. D. Luker and
S. Takayama, J. Controlled Release, 2012, 164, 192–204.26 K. Schmeichel and M. Bissell, J. Cell Sci., 2003, 116,
2377–2388.27 J. Friedrich, C. Seidel, R. Ebner and L. A. Kunz-Schughart,
Nat. Protocols, 2009, 4, 309–324.28 M. Drewitz, M. Helbling, N. Fried, M. Bieri, W. Moritz,
J. Lichtenberg and J. M. Kelm, Biotechnol. J., 2011, 6,1488–1496.
29 Y.-C. Tung, A. Y. Hsiao, S. G. Allen, Y.-S. Torisawa, M. Hoand S. Takayama, Analyst, 2011, 136, 473–478.
30 A. Y. Hsiao, Y.-C. Tung, C.-H. Kuo, B. Mosadegh, R. Bedenis,K. J. Pienta and S. Takayama, Biomed. Microdevices, 2012, 14,313–323.
31 L. Y. Wu, D. Di Carlo and L. P. Lee, Biomed. Microdevices,2008, 10, 197–202.
32 S. Agastin, U.-B. T. Giang, Y. Geng, L. A. Delouise andM. R. King, Biomicrofluidics, 2011, 5, 24110.
33 A. Y. Hsiao, Y.-S. Torisawa, Y.-C. Tung, S. Sud,R. S. Taichman, K. J. Pienta and S. Takayama, Biomaterials,2009, 30, 3020–3027.
34 Y. Toh, C. Zhang, J. Zhang, Y. Khong, S. Chang, V. Samper,D. van Noort, D. Hutmacher and H. Yu, Lab Chip, 2007, 7,302–309.
35 T. Liu, B. Lin and J. Qin, Lab Chip, 2010, 10, 1671–1677.36 M. Domenech, H. Yu, J. Warrick, N. Badders,
I. Meyvantsson, C. Alexander and D. Beebe, Integr. Biol.,2009, 1, 267–274.
37 T.-H. Hsu, J.-L. Xiao, Y.-W. Tsao, Y.-L. Kao, S.-H. Huang,W.-Y. Liao and C.-H. Lee, Lab Chip, 2011, 11, 1808–1814.
38 M. Domenech, R. Bjerregaard, W. Bushman and D. J. Beebe,Integr. Biol., 2012, 4, 142–152.
39 H. Kaji, T. Yokoi, T. Kawashima and M. Nishizawa, LabChip, 2009, 9, 427–432.
40 Y. Gao, D. Majumdar, B. Jovanovic, C. Shaifer, P. C. Lin,A. Zijlstra, D. J. Webb and D. Li, Biomed. Microdevices, 2011,13, 539–548.
41 C. Huang, J. Lu, H. Seon, A. Lee, L. Flanagan, H. Kim,A. Putnam and N. Jeon, Lab Chip, 2009, 9, 1740–1748.
42 E. W. K. Young, C. Pak, B. S. Kahl, D. T. Yang,N. S. Callander, S. Miyamoto and D. J. Beebe, Blood, 2012,119, E76–E85.
43 H. Ma, T. Liu, J. Qin and B. Lin, Electrophoresis, 2010, 31,1599–1605.
44 E. Cukierman and D. E. Bassi, Semin. Cancer Biol., 2010, 20,139–145.
45 J. I. Lopez, J. K. Mouw and V. M. Weaver, Oncogene, 2008, 27,6981–6993.
46 A. C. Shieh, Ann. Biomed. Eng., 2011, 39, 1379–1389.47 S. Kumar and V. M. Weaver, Cancer Metastasis Rev., 2009,
28, 113–127.
48 K. E. Sung, N. Yang, C. Pehlke, P. J. Keely, K. W. Eliceiri,A. Friedl and D. J. Beebe, Integr. Biol., 2011, 3, 439–450.
49 E. W. K. Young and C. A. Simmons, Lab Chip, 2010, 10, 143–160.50 K. H. K. Wong, J. M. Chan, R. D. Kamm and J. Tien, Annu.
Rev. Biomed. Eng., 2012, 14, 205–230.51 E. W. K. Young, JALA, 2013, DOI: 10.1177/2211068213495206.52 V. Vickerman, J. Blundo, S. Chung and R. Kamm, Lab Chip,
2008, 8, 1468–1477.53 S. Chung, R. Sudo, P. Mack, C. Wan, V. Vickerman and
R. Kamm, Lab Chip, 2009, 9, 269–275.54 J. W. Song and L. L. Munn, Proc. Natl. Acad. Sci. U. S. A.,
2011, 108, 15342–15347.55 Y. Shin, J. S. Jeon, S. Han, G.-S. Jung, S. Shin, S.-H. Lee,
R. Sudo, R. D. Kamm and S. Chung, Lab Chip, 2011, 11,2175–2181.
56 L. Bischel, E. W. K. Young, B. Mader and D. J. Beebe,Biomaterials, 2013, 34, 1471–1477.
57 Y. Zheng, J. Chen, M. Craven, N. W. Choi, S. Totorica,A. Diaz-Santana, P. Kermani, B. Hempstead, C. Fischbach-Teschl, J. A. Lopez and A. D. Stroock, Proc. Natl. Acad. Sci.U. S. A., 2012, 109, 9342–9347.
58 S. Kim, H. Lee, M. Chung and N. L. Jeon, Lab Chip, 2013, 13,1489–1500.
59 T. Keenan and A. Folch, Lab Chip, 2008, 8, 34–57.60 C. L. Walsh, B. M. Babin, R. W. Kasinskas, J. A. Foster,
M. J. McGarry and N. S. Forbes, Lab Chip, 2009, 9, 545–554.61 G. Mehta, K. Mehta, D. Sud, J. Song, T. Bersano-Begey,
N. Futai, Y. Heo, M. Mycek, J. Linderman andS. Takayama, Biomed. Microdevices, 2007, 9, 123–134.
62 J. Park, T. Bansal, M. Pinelis and M. M. Maharbiz, Lab Chip,2006, 6, 611–622.
63 Y.-A. Chen, A. D. King, H.-C. Shih, C.-C. Peng, C.-Y. Wu,W.-H. Liao and Y.-C. Tung, Lab Chip, 2011, 11, 3626–3633.
64 R. Sikes, B. Nicholson, K. Koeneman, N. Edlund,E. Bissonette, M. Bradley, G. Thalmann, M. Cecchini,K. Pienta and L. Chung, Int. J. Cancer, 2004, 110, 497–503.
65 D. Price, T. Miralem, S. Jiang, R. Steinberg and H. Avraham,Cell Growth Differ., 2001, 12, 129–135.
66 Y. Kang, P. Siegel, W. Shu, M. Drobnjak, S. Kakonen,C. Cordon-Cardo, T. Guise and J. Massague, Cancer Cell,2003, 3, 537–549.
67 A. Minn, G. Gupta, P. Siegel, P. Bos, W. Shu, D. Giri, A. Viale,A. Olshen, W. Gerald and J. Massague, Nature, 2005, 436,518–524.
68 P. D. Bos, X. H.-F. Zhang, C. Nadal, W. Shu, R. R. Gomis,D. X. Nguyen, A. J. Minn, M. J. van de Vijver, W. L.Gerald, J. A. Foekens and J. Massague, Nature, 2009, 459,1005–1009.
69 K. Chaw, M. Manimaran, E. Tay and S. Swaminathan, LabChip, 2007, 7, 1041–1047.
70 K. C. Chaw, M. Manimaran, F. E. H. Tay andS. Swaminathan, Biomed. Microdevices, 2007, 9, 597–602.
71 I. K. Zervantonakis, S. K. Hughes-Alford, J. L. Charest,J. S. Condeelis, F. B. Gertler and R. D. Kamm, Proc. Natl.Acad. Sci. U. S. A., 2012, 109, 13515–13520.
72 D. Hanahan and R. Weinberg, Cell, 2000, 100, 57–70.
Integrative Biology Review Article
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online

This journal is c The Royal Society of Chemistry 2013 Integr. Biol., 2013, 5, 1096--1109 1109
73 S. Goel, D. G. Duda, L. Xu, L. L. Munn, Y. Boucher,D. Fukumura and R. K. Jain, Physiol. Rev., 2011, 91, 1071–1121.
74 S. Nagrath, L. V. Sequist, S. Maheswaran, D. W. Bell, D. Irimia,L. Ulkus, M. R. Smith, E. L. Kwak, S. Digumarthy,A. Muzikansky, P. Ryan, U. J. Balis, R. G. Tompkins,D. A. Haber and M. Toner, Nature, 2007, 450, 1235–1239.
75 D. Issadore, J. Chung, H. Shao, M. Liong, A. A. Ghazani,C. M. Castro, R. Weissleder and H. Lee, Sci. Transl. Med.,2012, 4, 141ra92, DOI: 10.1126/scitranslmed.3003747.
76 E. Sackmann, E. Berthier, E. W. K. Young, M. A. Shelef,S. A. Wernimont, A. Huttenlocher and D. J. Beebe, Blood,2012, 120, e45–e53.
77 J. R. Kraly, R. E. Holcomb, Q. Guan and C. S. Henry, Anal.Chim. Acta, 2009, 653, 23–35.
78 A. Amantonico, P. L. Urban and R. Zenobi, Anal. Bioanal.Chem., 2010, 398, 2493–2504.
79 L. Shintu, R. Baudoin, V. Navratil, J.-M. Prot, C. Pontoizeau,M. Defernez, B. J. Blaise, C. Domange, A. R. Pery,P. Toulhoat, C. Legallais, C. Brochot, E. Leclerc andM.-E. Dumas, Anal. Chem., 2012, 84, 1840–1848.
80 K. J. Regehr, M. Domenech, J. T. Koepsel, K. C. Carver,S. J. Ellison-Zelski, W. L. Murphy, L. A. Schuler, E. T. Alaridand D. Beebe, Lab Chip, 2009, 9, 2132–2139.
81 X. Su, A. B. Theberge, C. T. January and D. J. Beebe,Anal. Chem., 2013, 85, 1562–1570.
82 E. W. K. Young, E. Berthier, D. J. Guckenberger,E. Sackmann, C. Lamers, I. Meyvantsson, A. Huttenocherand D. J. Beebe, Anal. Chem., 2011, 83, 1408–1417.
83 E. Berthier, E. W. K. Young and D. Beebe, Lab Chip, 2012, 12,1224–1237.
84 X. Su, E. W. K. Young, H. A. S. Underkofler, T. J. Kamp,C. T. January and D. J. Beebe, J. Biomol. Screening, 2011, 16,101–111.
85 W. S. Hong, E. W. K. Young, W. H. Tepp, E. A. Johnson andD. J. Beebe, Toxicol. Sci., 2013, DOI: 10.1093/toxsci/kft082.
86 A. K. White, M. VanInsberghe, O. I. Petriv, M. Hamidi,D. Sikorski, M. A. Marra, J. Piret, S. Aparicio andC. L. Hansen, Proc. Natl. Acad. Sci. U. S. A., 2011, 108,13999–14004.
87 H. Zhu, G. Stybayeva, M. Macal, E. Ramanculov,M. D. George, S. Dandekar and A. Revzin, Lab Chip, 2008,8, 2197–2205.
88 N.-T. Huang, W. Chen, B.-R. Oh, T. T. Cornell, T. P. Shanley,J. Fu and K. Kurabayashi, Lab Chip, 2012, 12, 4093–4101.
89 K. Hess, G. Varadhachary, S. Taylor, W. Wei, M. Raber,R. Lenzi and J. Abbruzzese, Cancer, 2006, 106, 1624–1633.
Review Article Integrative Biology
Publ
ishe
d on
07
June
201
3. D
ownl
oade
d on
29/
03/2
014
12:0
1:23
. View Article Online