cartographie génétique des génomes eucaryotespdv.univ-lille1.fr/labo/figb4part1.pdf · structure...
TRANSCRIPT

Cartographie génétique des génomes eucaryotes
Marie-Christine QuilletSN2 3ème étage

Références bibliographiquesOuvrages
• An introduction to genetic analysis, 6th ed. A.F. Griffiths et al., 1998.Editeur: W.H. Freeman and Company, New York
• Genetics the continuity of life. D.J. Faibanks and W.R. Andersen, 1999.Editeur: Brooks/Cole Publishing Company, Pacific Grove, CA
• Gene V. B. Lewin, 1995Editeur: Oxford University Press Inc, New York
• Biologie moléculaire et médecine, 2ème ed. J.C. Kaplan et M. Delpech, 1998Editeur: Flamarion Médecine-sciences, Paris
• Génétique; gènes et génomes. J.L. Rossignol et al., 2000Editeur: Dunod, Paris
• Génétique. A. Oulmouden et al., 1999Editeur: Dunod, Paris
• Analyse de génomes, transcriptomes et protéomes, 3ème ed.. A. Bernot, 2001. Editeur: Dunod (collection Biotech. Info), Paris

Cartographie génétique des génomes eucaryotes
Construction de cartes génétique1. Marqueurs génétiques2. Types de descendances3. Méthodes d'établissement des cartes génétiques4. Caractéristiques des cartes génétiques

Exemple de pénétrance et d'expressivité incomplète :• pigmentation des graines chez le pois, sous le contrôle d'un gène
majeur avec A dominant/aEn cas de pénétrance et d'expressivité totale:
Génotype A• → phénotype , Génotype aa → phénotype
Phénotype des individus A•Fond génétique (milieu) 1Pénétrance incomplète (56%)
Fond génétique (milieu) 2Expressivité incomplète
Fond génétique (milieu) 3Pénétrance et expressivité incomplète
Pénétrance incomplète: impossible d'associer le phénotype à un génotype sans risque de se tromper

Structure d'un gène d'eucaryotes et de son promoteur
Site d'initiation de la transcription
Substitutionnucléotidique
Site d'initiation de la traduction
Site de terminaison de la transcription
promoteur exon 1 intron 1 intron 2exon 2 exon 3
Site donneur
Site donneur
Siteaccepteur
Siteaccepteur
Site depolyadénylation
Insertioncourte/E.T.
délétion
Substitutionnucléotidique
Site d'arrêt de la traduction (stop)
5'7-me-GTP AAAAAn 3'
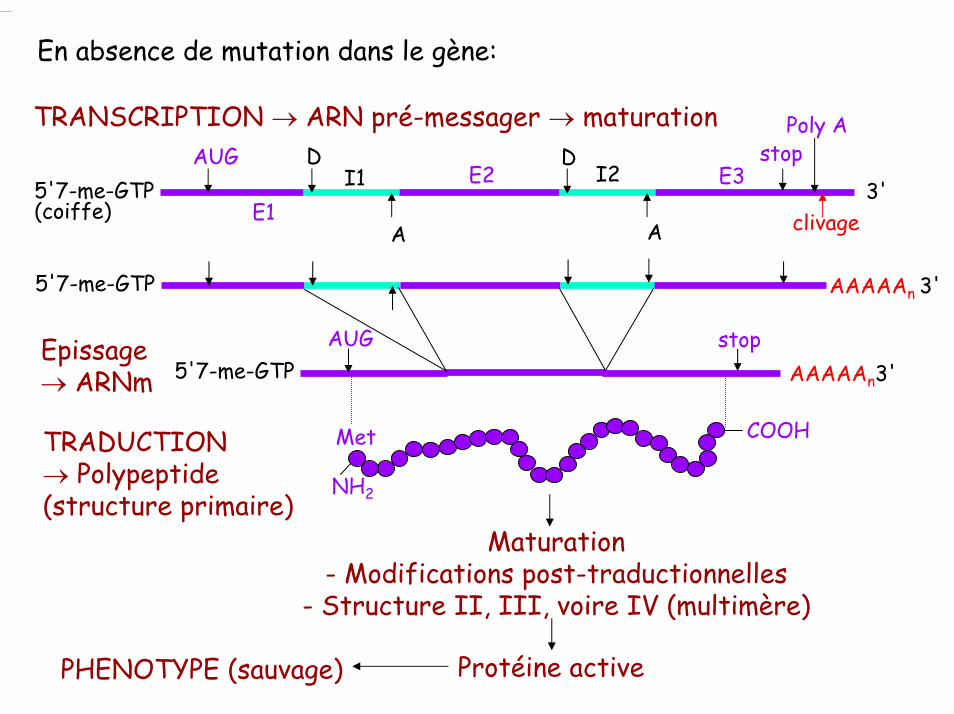
TRANSCRIPTION → ARN pré-messager → maturation
5'7-me-GTP(coiffe) clivageE1
E2 E3AUG D D
A A
stopPoly A
I1 I23'
AAAAAn3'5'7-me-GTPEpissage → ARNm
AUG stop
TRADUCTION→ Polypeptide (structure primaire)
NH2
COOHMet
Maturation- Modifications post-traductionnelles
- Structure II, III, voire IV (multimère)
Protéine activePHENOTYPE (sauvage)
En absence de mutation dans le gène:

Exemple d'activation de la transcription d'un gène après l'insertion d'un élément mobile à proximité du promoteur
→ Gène impliqué dans la pigmentation des grains du maïs
Cellule d'anthère ou de feuille
P gène
Pas d'expression du gène
Pas de pigmentation des anthères et des feuilles
PE.T. gène
Insertion élément mobile
Expression ectopique du gène
Pigmentation des anthères et des feuilles
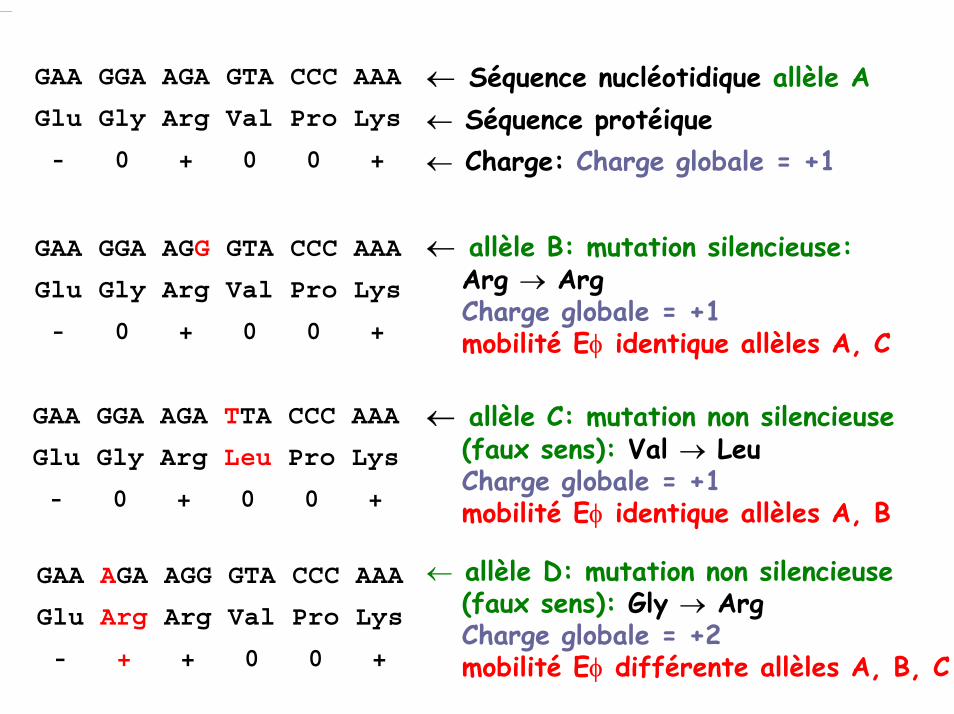
GAA GGA AGA GTA CCC AAA
Glu Gly Arg Val Pro Lys
- 0 + 0 0 +
← Séquence nucléotidique allèle A← Séquence protéique← Charge: Charge globale = +1
GAA GGA AGG GTA CCC AAA
Glu Gly Arg Val Pro Lys
- 0 + 0 0 +
← allèle B: mutation silencieuse:Arg → ArgCharge globale = +1mobilité Eφ identique allèles A, C
GAA GGA AGA TTA CCC AAA
Glu Gly Arg Leu Pro Lys
- 0 + 0 0 +
← allèle C: mutation non silencieuse(faux sens): Val → LeuCharge globale = +1mobilité Eφ identique allèles A, B
GAA AGA AGG GTA CCC AAA
Glu Arg Arg Val Pro Lys
- + + 0 0 +
← allèle D: mutation non silencieuse(faux sens): Gly → ArgCharge globale = +2mobilité Eφ différente allèles A, B, C

Détection du polymorphisme de site de restriction par RFLP
Paire de chromosomes homologues
Mutation ponctuelle disparition du site
Lignée BSonde marquée (radioactivité,
cheminuminescence)
Digestion de l'ADN des différents individus par l'enzyme
de restriction (quelques µg)
+
-Li
gnée
AL i
g né e
B
F1Ségrégation 1:2:1 parmi
les descendants F2
Miration sur gel d'agarose
Transfert sur une menbrane de nylon
(Southern blot)
Hybridation de la sonde Détection des signaux (autoradiographie)
Sites de restriction de l'enzyme
Lignée A

Détection du polymorphisme d'insertion/délétion par RFLP
InsertionSites de restriction de l'enzyme
Paire de chromosomes homologues
Lignée A Lignée BSonde marquée (radioactivité,
cheminumilescence)
Miration sur gel d'agarose
Transfert sur une menbrane de nylon
(Southern blot)
Hybridation de la sonde Détection des signaux (autoradiographie)
-
+
Lign
ée A
L ig n
é e B
F1Ségrégation 1:2:1 parmi
les descendants F2
Digestion de l'ADN des différents individus par l'enzyme
de restriction (quelques µg)

4.1.b. Marqueurs spécifiques de locus obtenus par PCR
• PCR: Polymerase Chain Reaction (Mullis et al. 1986)
5'3'
3'5'
5' 3' 3' 5'
ADN matrice (quelques ng)Amorces20-25 pb
dATP, dCTP, dGTP, dTTPTaq polymérase
Amplification exponentielle de la séquence cible (≤ 2 kpb)
Après n cycles de PCR:2n copies de la séquence cible
(× 10 ng)
1 cycle de PCR: 3 étapes- Dénaturation de l'ADN matrice- Hybridation des amorces- Elongation
Détection du produit de PCR après séparation par
électrophorèse

• Critère de sélection d'un "bon" marqueur spécifique de locus obtenu par PCR
Portion d'une paire de chromosomes homologues(individu hétérozygote)
polymorphisme
Séquences flanquantes
Site d'hybridation des amorces PCR

1. Le site d'hybridation des amorces PCR est une séquence unique, non polymorphe → ☺
-
+
AB AA BB1.
2. Le site d'hybridation des amorces PCR est une séquence unique, polymorphe →
-
+
A• A • aa2.
A• = AA ou Aaa: allèle nul

3. Le site d'hybridation des amorces PCR est une séquence répétée, non polymorphe (ou polymorphe) →
-
+
Ind 1 Ind 2 Ind 3

4.1.b.1. Marqueurs CAPS (Cleaved Amplified Polymorphic Sequence):
amorces PCR Site de restriction reconnu par l'enzymePortion d'une paire de chromosomes homologuesGénotype AB
Pas de reconnaissance du site
Allèle A Allèle B

• Amplification de l'ADN des individus dont on désire connaître le génotype (1 couple d'amorces spécifiques/locus)
• Dépôt des produits d'amplification sur un gel d'agarose
-
+
P1 P2 F1 BC1: F1 × P1
Homoplasie de taille
• Digestion des produits d'amplification par un enzyme de restriction et électrophorèse
P1 P2 F1 BC1: F1 × P1 (ségrégation 1:1)
AA BB AB Hors type
-
+
Résolution des gels d'agarose: ≥ 20 pb
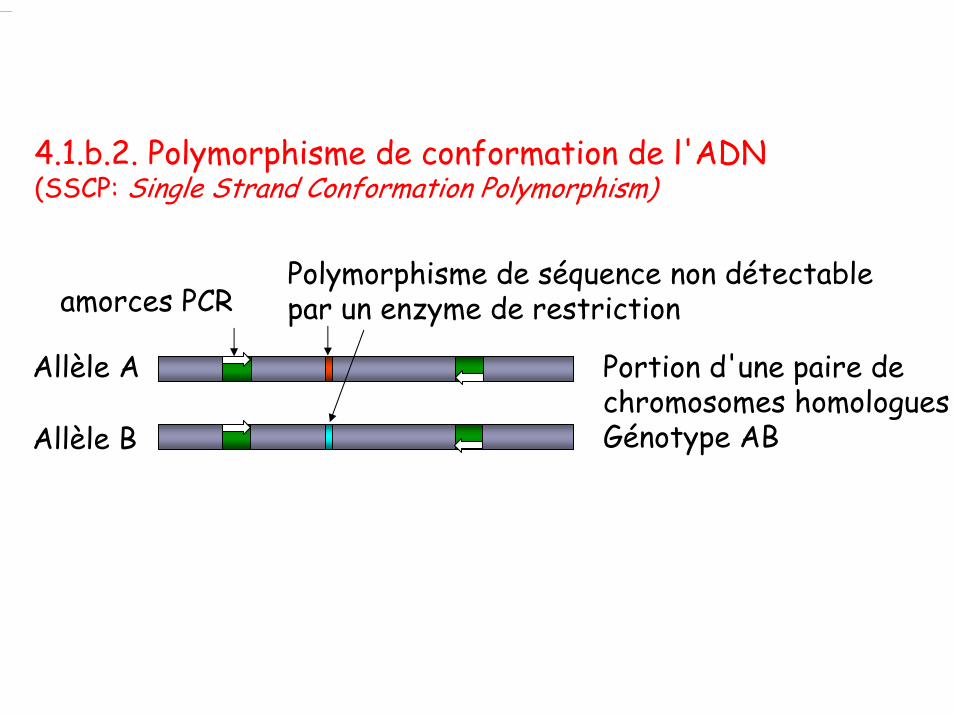
4.1.b.2. Polymorphisme de conformation de l'ADN (SSCP: Single Strand Conformation Polymorphism)
amorces PCR Polymorphisme de séquence non détectable par un enzyme de restriction
Allèle A
Allèle B
Portion d'une paire de chromosomes homologuesGénotype AB
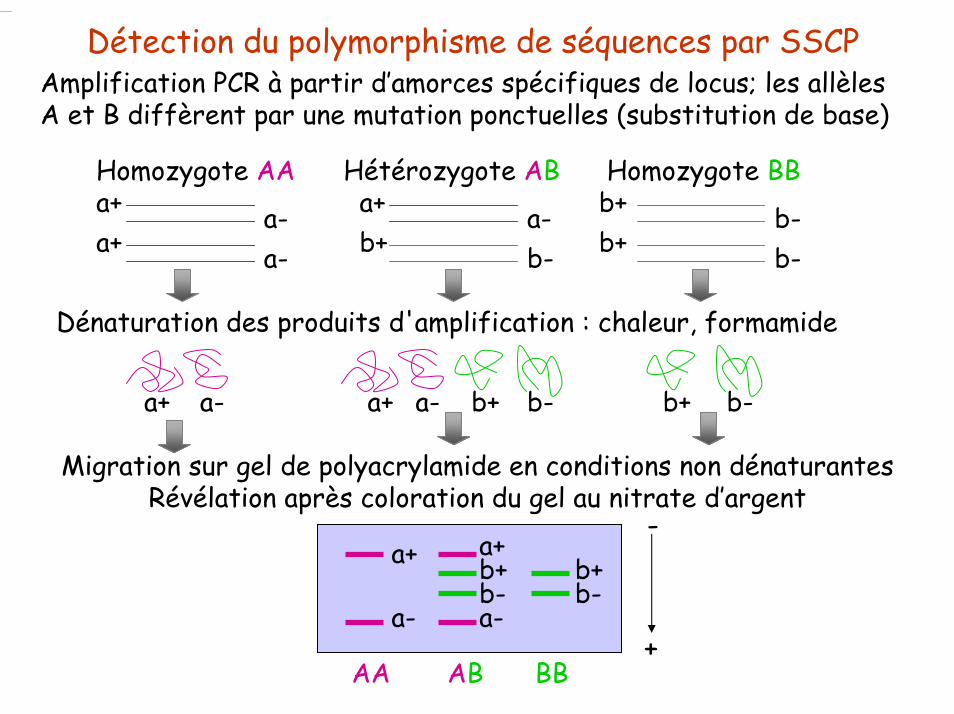
Détection du polymorphisme de séquences par SSCPAmplification PCR à partir d’amorces spécifiques de locus; les allèles A et B diffèrent par une mutation ponctuelles (substitution de base)
Homozygote AAa+ a-a+ a-
Hétérozygote ABa+ a-
b-b+
Homozygote BBb+b+
b-b-
Dénaturation des produits d'amplification : chaleur, formamide
a+ a+a- a- b+ b+b- b-
Migration sur gel de polyacrylamide en conditions non dénaturantesRévélation après coloration du gel au nitrate d’argent
a+ a+
a- a-
b+ b+b- b-
AA AB BB
-
+

Locus 1
Locus 2
L1 L2F1 F2
a+, b+a-b-
a+b+
a-b-
1 2 3 4 5 6 7 8 9 10 11
Exemple de profils SSCP; co-migration des produits d'amplification obtenus pour 2 locus (carte génétique de la chicorée, LPDV)
Locus 1
Locus 2
L1BB
AA
L2AA
BB
F1AB
AB
AA AB AB BB AA AB AB BB AA AB AA
AB AB AA AA AB BB AB AA AB AB AA
1 2 3 4 5 6 7 8 9 10 11

• Structure d'un locus microsatellite
Nombre variable de répétitions du motif CA
Séquences flanquantes
Portion d'une paire de chromosomes homologuesGénotype: (CA)14 /(CA)10
Amorce PCR
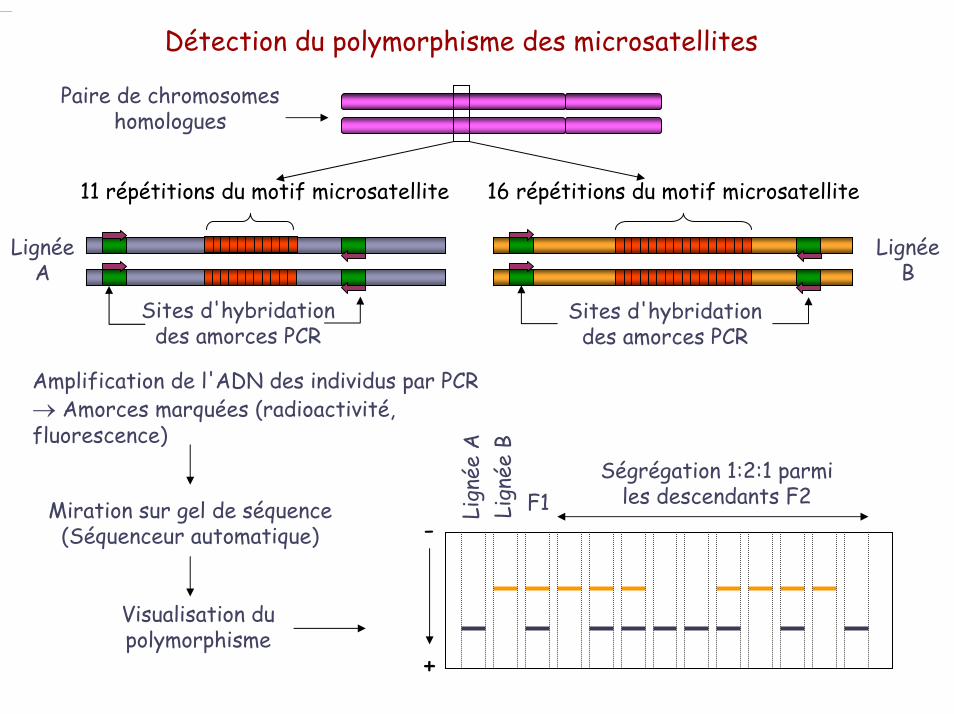
Détection du polymorphisme des microsatellites
Paire de chromosomes homologues
Amplification de l'ADN des individus par PCR→ Amorces marquées (radioactivité, fluorescence)
Miration sur gel de séquence(Séquenceur automatique) -
+
Lign
ée A
L ig n
é e B
F1Ségrégation 1:2:1 parmi
les descendants F2
Visualisation du polymorphisme
11 répétitions du motif microsatellite
Lignée A
Sites d'hybridation des amorces PCR
Lignée B
16 répétitions du motif microsatellite
Sites d'hybridation des amorces PCR
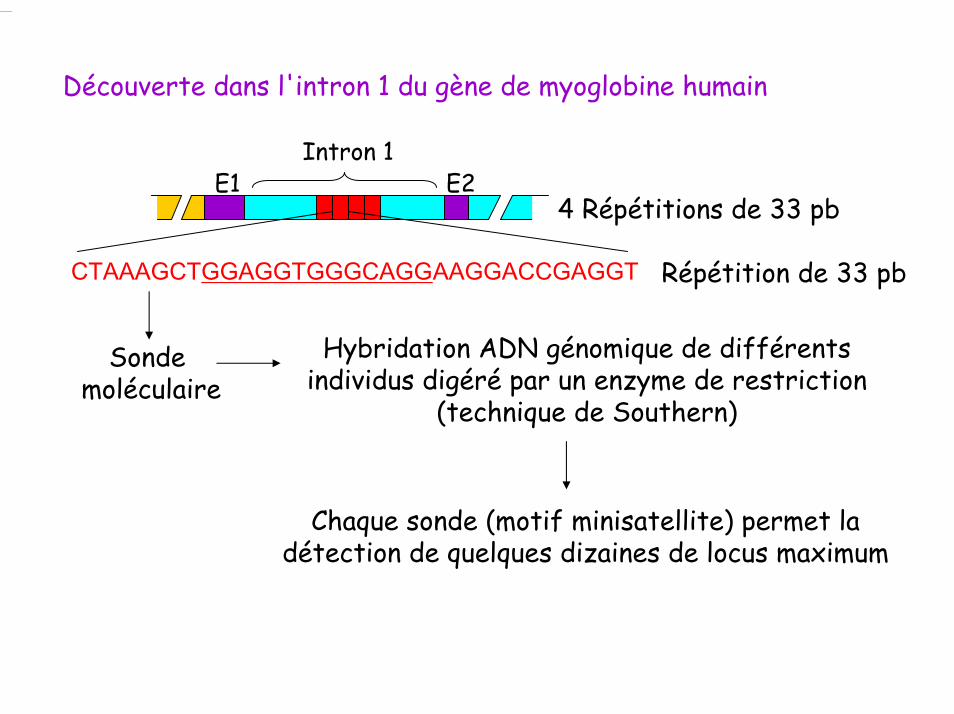
CTAAAGCTGGAGGTGGGCAGGAAGGACCGAGGT
E1Intron 1
E2
Répétition de 33 pb
4 Répétitions de 33 pb
Découverte dans l'intron 1 du gène de myoglobine humain
Sonde moléculaire
Hybridation ADN génomique de différents individus digéré par un enzyme de restriction
(technique de Southern)
Chaque sonde (motif minisatellite) permet la détection de quelques dizaines de locus maximum

VNTR 1 VNTR 2 VNTR 3 VNTR 4
3 chromosomes du génome de l'espèce X qui contiennent un motif VNTR identique (i.e. reconnu par la même sonde)
Digestion de l'ADN de différents individus par un enzyme de restriction
Ind 1 Ind 2 Ind 3
Migration des l'ADN dans un gel d'agaroseTransfert sur une membrane de nylon
Hybridation avec la sonde correspondant au motif VNTR
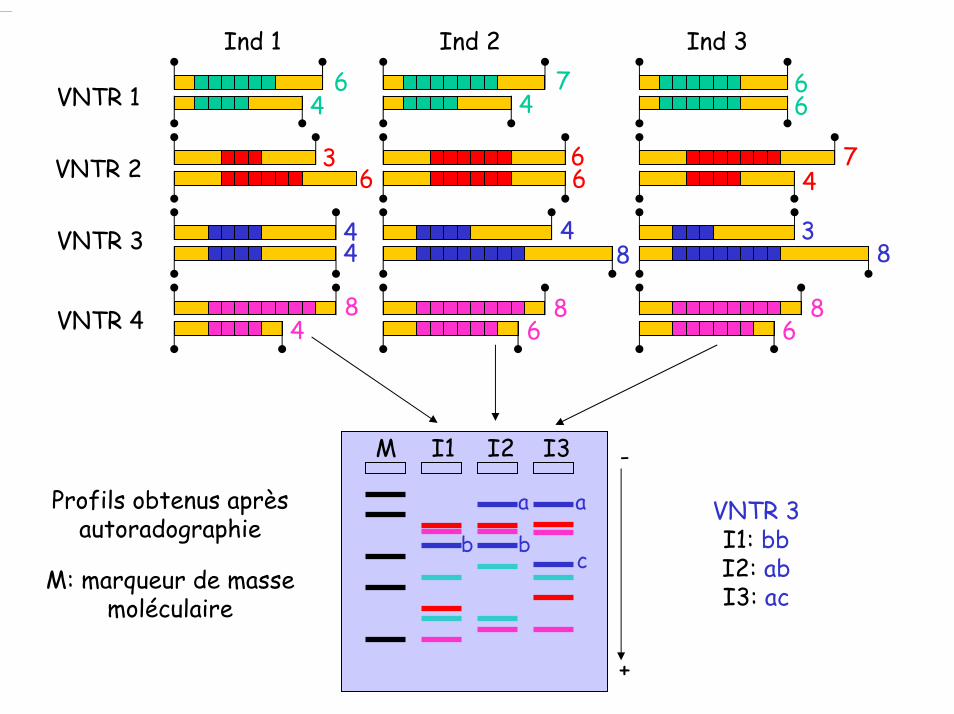
Profils obtenus après autoradographie
M: marqueur de masse moléculaire
M I1 I2 I3 -
+
bb
a a
c
VNTR 3I1: bbI2: abI3: ac
VNTR 1
VNTR 2
VNTR 3
VNTR 4
Ind 1 Ind 2 Ind 36
47
466
36
66
74
44
48
38
84
86
86

M I1 I2 I3
Malheureusement les profils obtenus sont en
noir et blanc !
-
+
⇒ Impossibilité de déterminer les relations
d'allélisme entre les différents niveaux de
bandes
VNTR 1
VNTR 2
VNTR 3
VNTR 4
Ind 1 Ind 2 Ind 36
47
466
36
66
74
44
48
38
84
86
86

Interprétation du résultat
système binaire permettant une notation
de chaque niveau de bande→ marqueurs dominants1 = présence du signal0 = absence du signal
9 1 1 0
7 0 0 1
5 0 1 1
3 1 1 1
1 0 1 12 1 1 1
4 1 1 0
6 1 0 1
8 1 0 0
10 0 1 111 1 0 0
I1 I2 I3M I1 I2 I3
VNTR 1
VNTR 2
VNTR 3
VNTR 4
Ind 1 Ind 2 Ind 36
47
466
36
66
74
44
48
38
84
86
86

Utilisation des marqueurs minisatellites• Peu utilisés pour la construction de cartes génétiques• Souvent utilisés pour établir des empreintes génétiques (DNA fingerprints) chez de nombreux organismes
ADN extrait de sang humain prélevé sur les lieux d'un crime
ADN extrait du sang de plusieurs suspects
Identification variétale chez le maïs

4.2.b. Marqueurs d'empreintes génétiques obtenus par PCR
4.2.b.1. RAPD (Random Amplified Polymorphic DNA)
• PCR dans l'objectif d'amplifier une région unique du génome →marqueur spécifique de locus
5'3'
3'5'
5' 3' 3' 5'
ADN matrice = cible
Amorces 20-25 pb

Amplification exponentielle
PCR :ADN matrice (quelques ng)
5' 3'+ 1 amorce 10 pb
+ dATP, dCTP, dGTP, dTTP+ Taq polymérase
Distance < 2 kpb3'5'
5'3'
Cas 1: ☺
Visualisation des produits de PCR après migration sur un gel d'agarose
-
+

PCR :ADN matrice (quelques ng)
5' 3'+ 1 amorce 10 pb
+ dATP, dCTP, dGTP, dTTP+ Taq polymérase
Distance > 2 kpb3'5'
5'3'
Cas 2:
Cas 3: 3'5'
5'3'
Cas 4: 3'5'
5'3'
Pas d'amplification exponentielle
Aucun produit de PCR détectable après
migration sur un gel d'agarose

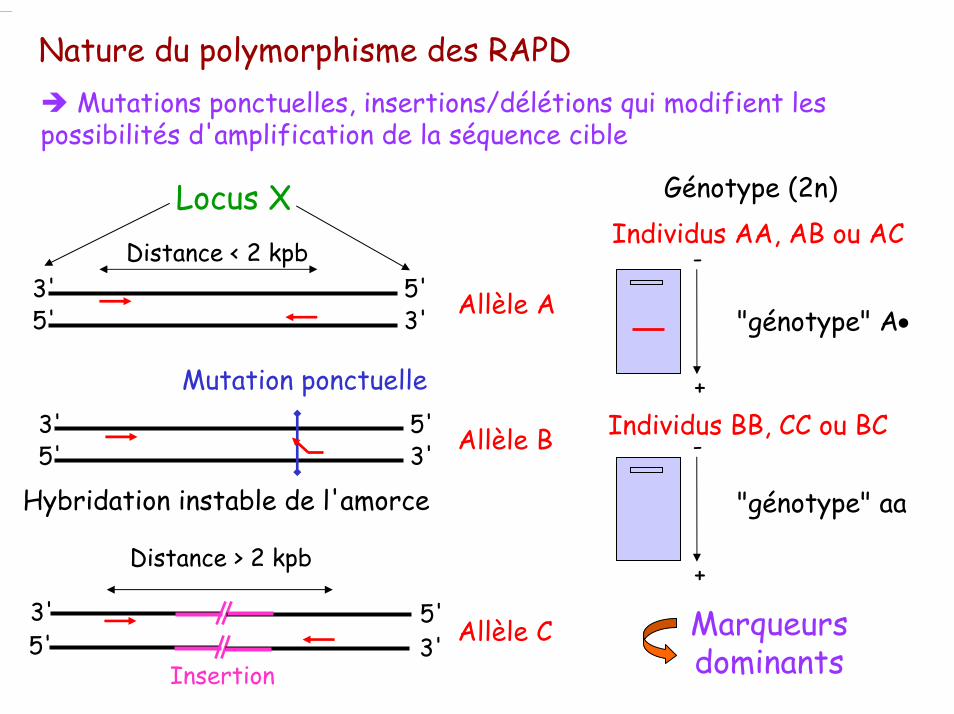
Nature du polymorphisme des RAPDMutations ponctuelles, insertions/délétions qui modifient les
possibilités d'amplification de la séquence cible
Distance < 2 kpb3'5'
5'3'
Locus X
Allèle A
Hybridation instable de l'amorce
3'5'
5'3'
Mutation ponctuelle
Allèle B
5'3'
3'5'
Distance > 2 kpb
Insertion
Allèle C
Amplification exponentielle
-
+
Pas d'amplification
-
+
Nature du polymorphisme des RAPDMutations ponctuelles, insertions/délétions qui modifient les
possibilités d'amplification de la séquence cible
Distance < 2 kpb3'5'
5'3'
Locus X
Hybridation instable de l'amorce
3'5'
5'3'
Mutation ponctuelle
5'3'
3'5'
Distance > 2 kpb
Insertion
Allèle A
Allèle B
Allèle C
Génotype (2n)
-
+
Individus AA, AB ou AC
"génotype" A•
-
+
Individus BB, CC ou BC
"génotype" aa
Marqueurs dominants
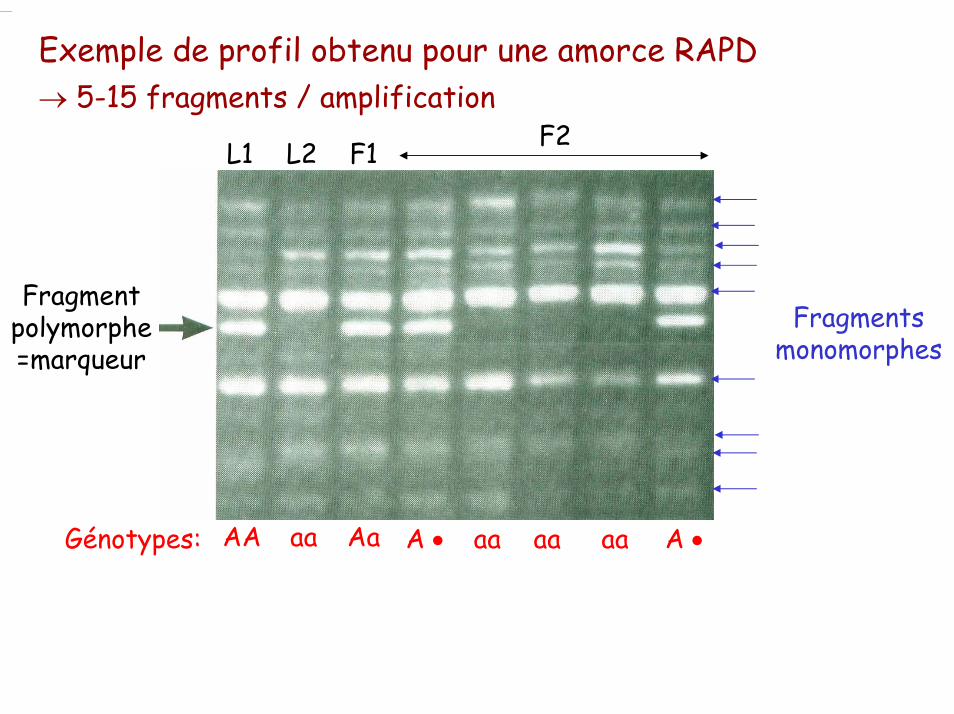
Exemple de profil obtenu pour une amorce RAPD→ 5-15 fragments / amplification
Fragment polymorphe=marqueur
L1 L2 F1 F2
AA aa Aa A • A •aa aa aaGénotypes:
Fragments monomorphes

4.2.b.2. AFLP (Amplified Fragment Lenght Polymorphism) (1995)
Digestion de l'ADN génomique par 2 enzymes de
restriction
Amplifications PCR
Site de reconnaissance à
6 pb: EcoR1
Site de reconnaissance à
4 pb: Mse1
5' 5'
Adaptateurs; fragments d'ADN de synthèse bicaténaires (≈ 25 pb)
Extémitécompatible EcoR1
Extrémitécompatible Mse1
Ligation d'adaptateursAdaptateur 1
Adaptateur 2
5'
5'
3'3'5'5'
NN N
N
G5'
T 5'
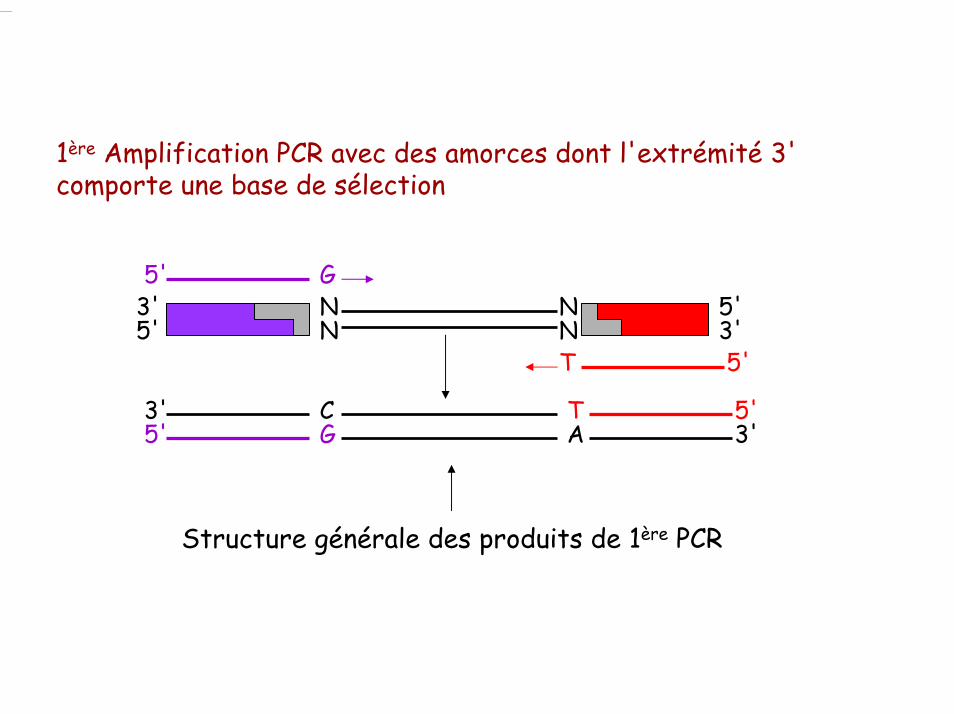
1ère Amplification PCR avec des amorces dont l'extrémité 3' comporte une base de sélection
T 5'CG5' A 3'
3'
Structure générale des produits de 1ère PCR

2ème Amplification PCR avec des amorces dont l'extrémité 3' comporte 2 bases de sélection supplémentaires
NNT 5'CNNGNN5' NNA 3'
3'GGA5'
GTT 5'
T 5'CG5' A 3'
3'Produits de 1ère PCR
GGA5' CAA 3'CCT GTT3' 5'
Structure générale des produits de 2ème PCR

Migration des produits de la seconde PCR dans un gel de séquence
Résolution de bonne qualité entre 50 et 500 pb
Vue d'ensemble des profils d'une série d'individus obtenus à l'issue de la seconde PCR
-
+
Plusieurs dizaines de fragments révélés par individu/couple
d'amorces utilisé pour réaliser la seconde PCR
Nombre de marqueurs potentiels ≈ illimité

Interprétation en tant que marqueurs dominants
M1
M2
M3M4
F1 F2
25 pb
Détail d'une portion d'un gel AFLP (carte génétique chicorée, LPDV)
aaAa A• A• A• aa aa aa A• A•

M1
L2
L1
M2
Cellule avant la méïose2n = 4, 2C
Génotype: L1L2 M1M2
M1
L2
L1
M2
M1
L2
M2
L1Phase S: réplication de l'ADN; 2n = 4, 4C2 chromatides sœurs/ chromosomes
Prophase I méiose:appariement des
chromosomes homologues2n = 4, 2C
Echanges de segments d'ADN entre chromatides non sœurs = crossing-over =
Brassages génétiques intra-chromosomiques(pas de conséquences sur la ségrégation des
allèles dans ce cas)
M1
M2
M1
M2L2
L1
L2
L1
Plaque équatoriale
Métaphase I méiose:ségrégation aléatoire des
chromosomesBrassages génétiques inter-chromosomiques
P = 1/2
ou
M1
M2
M1
M2
L2
L1
L2
L1
P = 1/2
2 marqueurs , L et M, présents sur des chromosomes différents
M1
M2L2
L1
M1
M2L2
L1 M1
M2
L2
L1 M1
M2
L2
L1
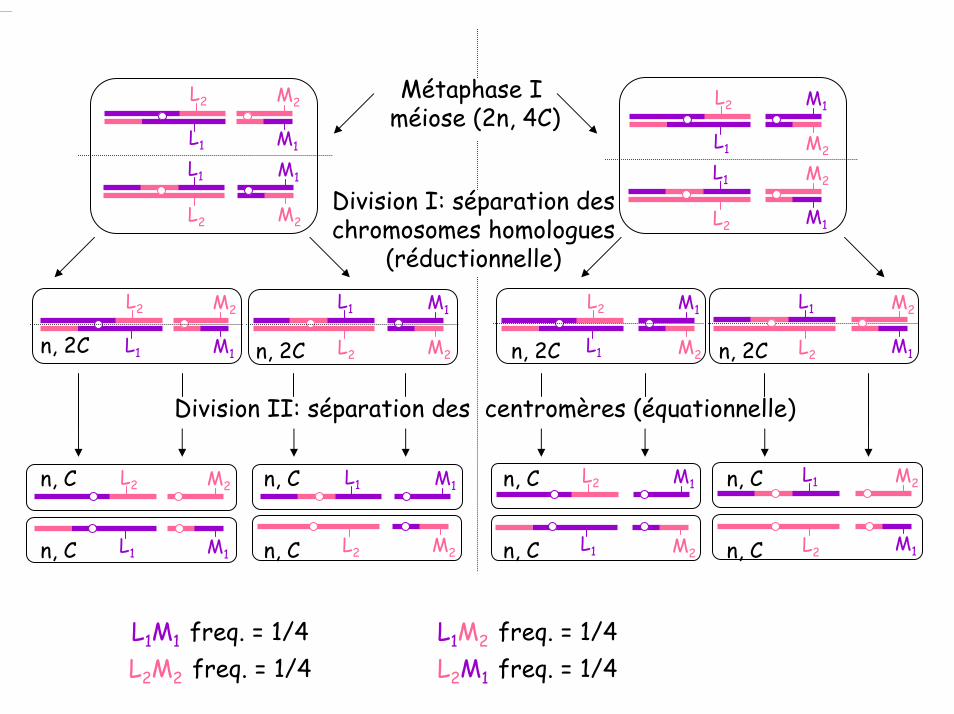
Division I: séparation des chromosomes homologues
(réductionnelle)
n, 2C n, 2C n, 2C n, 2C
n, C
n, C
n, C n, C
L1M1 freq. = 1/4L2M2 freq. = 1/4
L1M2 freq. = 1/4L2M1 freq. = 1/4
M2L2
M1L1
M1L2
M2L1
M1L1
M2L2 M1L2
M2L1
Division II: séparation des centromères (équationnelle)
n, C
n, C n, C n, C
M1
M2
M1
M2L2
L1
L2
L1
M1
M2
M1
M2
L2
L1
L2
L1
Métaphase Iméiose (2n, 4C)

2 marqueurs, L et M, présents sur le même chromosome
Pas de C-Oentre chromatides
non sœurs
1 C-O impliquant 2 chromatides
1
3
Autres possibilités
Génotype des gamètes
Association des allèles
Prophase I méiose
L1M1L1M1L2M2L2M2
PPPP
Aucune
1-42-32-4
L1M1L1M2L2M2L2M1
1-31-42-4
L1M1L1M2L2M1L2M2
PRPR
PRRP
2 C-O impliquant 2 chromatides
23
Génotype: L1L2 M1M22n =2, 2CL1 M1
L2 M2
L2
L1
L1
M1
M1
L2
M2
M2
12
34
Phase S
2n = 2, 4C2 chromatides
sœurs/ chromosomes

2 marqueurs, L et M, présents sur le même chromosome
2 C-O impliquant les 4
chromatides
Génotype: L1L2 M1M22n =2, 2CL1 M1
L2 M2
L2
L1
L1
M1
M1
L2
M2
M2
12
34
Phase S
2n = 2, 4C2 chromatides
sœurs/ chromosomes
Autres possibilités
Génotype des gamètes
Association des allèles
Prophase I méiose
2 C-O impliquant 3 chromatides
123
1-2-42-3-41-3-4
Aucune
L1M1L1M2L2M1L2M2
PRRP
L1M2L1M2L2M1L2M1
RRRR
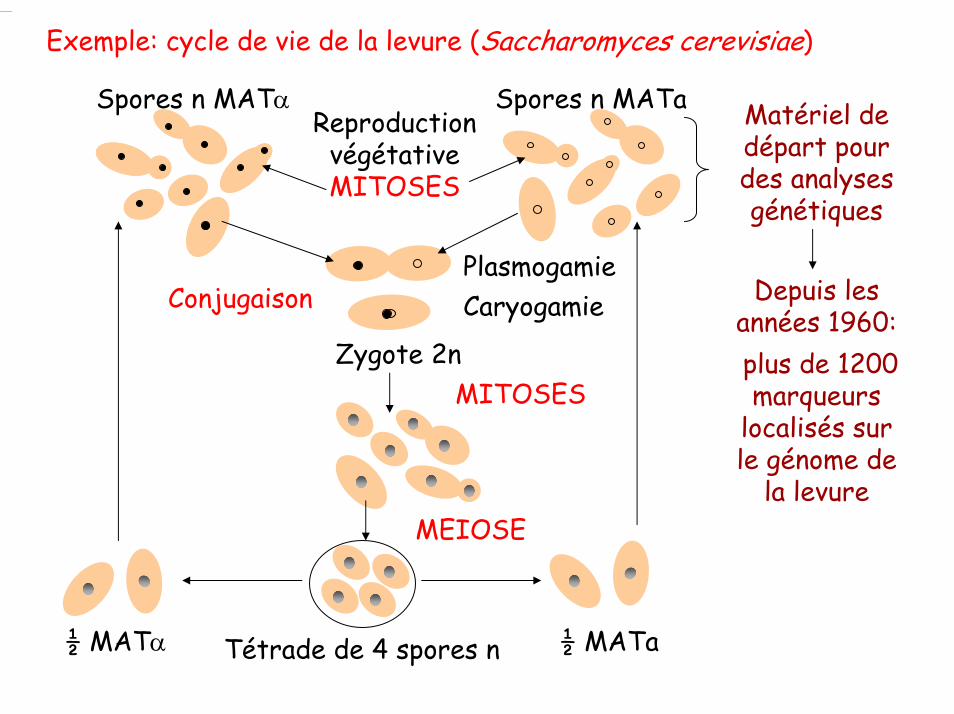
Exemple: cycle de vie de la levure (Saccharomyces cerevisiae)
Spores n MATα Spores n MATaReproduction
végétativeMITOSES
Plasmogamie CaryogamieConjugaison
Zygote 2nMITOSES
Tétrade de 4 spores n
MEIOSE
½ MATα ½ MATa
Matériel de départ pour des analyses génétiques
Depuis les années 1960:plus de 1200 marqueurs
localisés sur le génome de
la levure

Analyse directe des produits de méiose chez la levure
MAT: locus de compatibilité sexuelL: marqueur RAPD; L: présence du fragment, •: absence du fragment
Génotype dessouches haploïdes L MATa × • MATα
souche 1 × souche 2
Génotype du zygote L MATa• MATα
Génotype des spores L MATa • MATα L MATα • MATa
Effectifs observés 145 161 35 43
• Les locus L et MAT sont-ils liés ou indépendants ? (démonstration)• Si ces 2 locus sont liés, estimer la fréquence de recombinaison • Si le locus L est un marqueur de type microsatellite, polymorphe entre les souches 1 et 2, cela change t'il le génotype des spores et les effectifs observés ?

Plantes: mégagamétophyte femelle des gymnospermes
Cellule mère des mégasopres (2n) =mégasporocyte
ovule
Mégasoprefonctionnelle (n)
MEIOSE
Cellule œuf (n)
Mégagamétophyte (n)
MITOSES
… A maturité de la graine
Aile membraneuse
Embryon (2n)Mégagamétophyte (n) Extraction de l'ADN
ou des protéines

Individu hétérozygoteà de nombreux locus
Échantillon des produits de méiose
MEIOSE
Androgenèse / Gynogenèse
Individushaploïdes
Doublement chromosomique
Individus diploïdes
homozygotes
Collection de lignées HD
Pérennisation de la descendance par
autofécondation des lignées

• 2 locus, marqueurs codominants
L1 M1
L2 M2
L2 M2
L2 M2
L1 M2
L2 M2
L2 M1
L2 M2
L1 M1
L1 M1
L2 M2
L2 M2× P2:P1: L1 M1
L2 M2F1: L2 M2
L2 M2× P2: BC1
(1-r) N2
(1-r) N2
r N2
r N2
a b c d
A2 B2
Effectifs attendus
L1 M1 (γP) L2 M2 (γP) L1 M2 (γR) L2 M1 (γR)γ F1:
γ P2:
Effectifs observés
r = Nb. individus issus de γRNb. individus total
Rem. : résultat identique si on utilise P1 comme parent récurrent
= c + dN
1 génotype γF1
1 génotype BC1

• 2 locus, marqueurs dominants, phase de couplage (1)
(1-r) N2
(1-r) N2
r N2
r N2
a b c d
Effectifs attendus
l m
L M (γP) l m (γP) L m (γR) l M (γR)γ F1:
γ P2:
Effectifs observés
r = Nb. individus issus de γRNb. individus total
= c + dN
1 génotype γF1
1 génotype BC1
× P2:P1: L ML M
l ml m
L Ml m
F1: × P2: l ml m
Homozygote récessif ☺
L ml m
L Ml m
l ml m
l Ml m

• 2 locus, marqueurs dominants, phase de couplage (2)
× P2:P1: L ML M
l ml m
L Ml m
F1:
Phénotypes
L M
L M (γP) l m (γP) L m (γR) l M (γR)γ F1:
γ P2: L mL M
L ML M
l mL M
l ML M
P1: Homozygote dominant ×
L ML M
100% [L M]
⇒ Impossible d'estimer r

• 2 locus, marqueurs dominants, phase de répulsion
Phénotypes
L m
L m (γP) l M (γP) L M (γR) l m (γR)γ F1:
γ P2:
× P2:P1: L mL m
l Ml M
L ml M
F1:
L ML m
L mL m
l ML m
l mL m
× P1: L mL m
⇒ Impossible d'estimer r
[L m] [L M] [L M] [L m]
Rem. : résultat identique si on utilise P2 comme parent récurrent

3.2. Descendances dérivées de F2 chez les plantes: lignées recombinantes (LR)
P1 × P2(P1, P2 = lignées) Individus F2 Générations
F2
F3
F4
F5
F8-F10Lignées
recombinantes
× F1F1 Autofécondation
→ Chaque lignée = descendance monograine ou SSD (Single seed Descent)→ Pérennisation de la collection de lignées par autofécondation

Probabilité qu'un individu soit hétérozygote à un locus donné après n générations d'autofécondation: p = 1/2n
AA × aa F1: 100% Aa
F2: 25% AA 50% Aa 25% aa
F3: 37,5% AA 25% Aa 37,5% aa
F4: 43,75% AA 12,5% Aa 43,75% aa
F5: 46,88% AA 6,25% Aa 46,88% aa
F6: 48,44% AA 3,12% Aa 48,44% aa
F7: 49,22% AA 1,56% Aa 49,22% aa
F8: 49,61% AA 0,78% Aa 49,61% aap = 1/27

Individu F1 hétérozygoteà de nombreux
locus
Structure chromosomique des lignées recombinantes
MEIOSE
γ γ
n n méioses n méioses
F2
F8-F10
→ Probabilité de détecter une liaison entre 2 locus plus élevée que dans le cas des autres types de descendances

Estimation de la fréquence de recombinaison entre 2 locus (LR)→ Estimation indépendante du type de marqueur
× P2:P1: L mL m
l Ml M
L ml M
F1: × F1: L ml M
F2 F8, F10 …
P P R R
L mL m
l Ml M
L ML M
l ml m
[L m] [l M] [L M] [l m]
(1-R) N2
(1-R) N2
R N2
R N2
type:
Génotype des lignéesEffectifs attendus
Eff. obs. a b c d
R = Nb. lignées recombinantesNb. lignées total
= c + dN
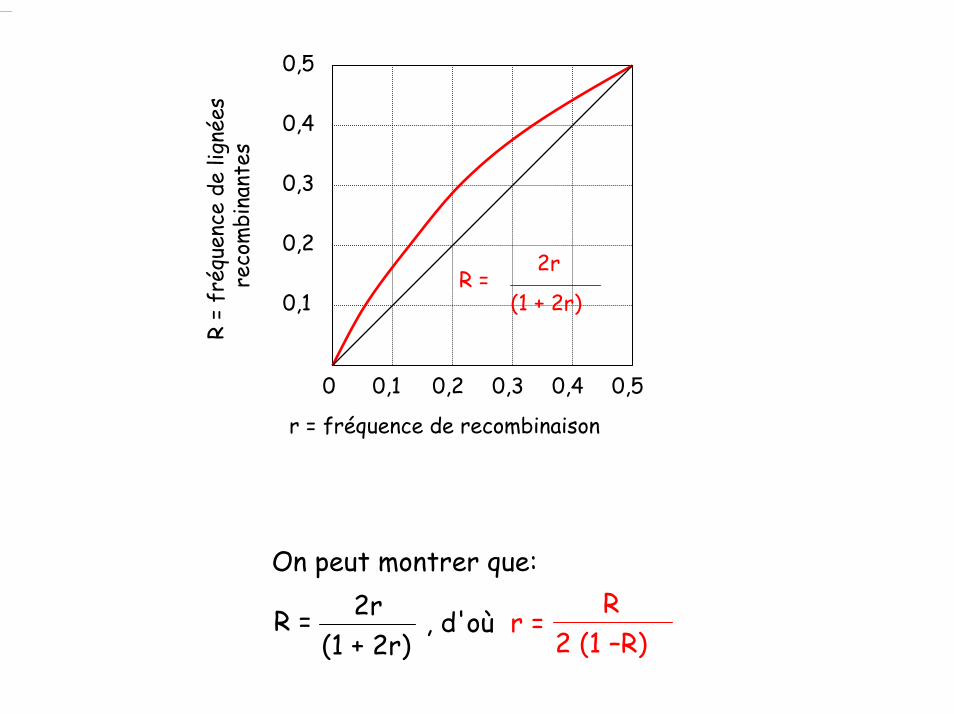
On peut montrer que:2r
(1 + 2r)R = , d'où r =
R2 (1 –R)
0,1 0,2 0,3 0,4 0,50
0,1
0,2
0,3
0,4
0,5
2r
(1 + 2r)R =
r = fréquence de recombinaison
R =
fréq
uenc
e de
lign
ées
reco
mbi
nant
es

• Tests 2 points: tests de liaison génétiques en considérant tous les couples de locus possibles
Ex: 100 marqueurs → 4900 combinaisons
Assigner chacun des marqueurs à un groupe de liaison
Groupe 1Groupe 2

Tests multipoint: ordres relatifs de 4, 5, 6, … locus
Les logiciels de cartographie utilisent le principe suivant afin de réduire le nombre de calculs à réaliser:
Test 3 points? ? ?
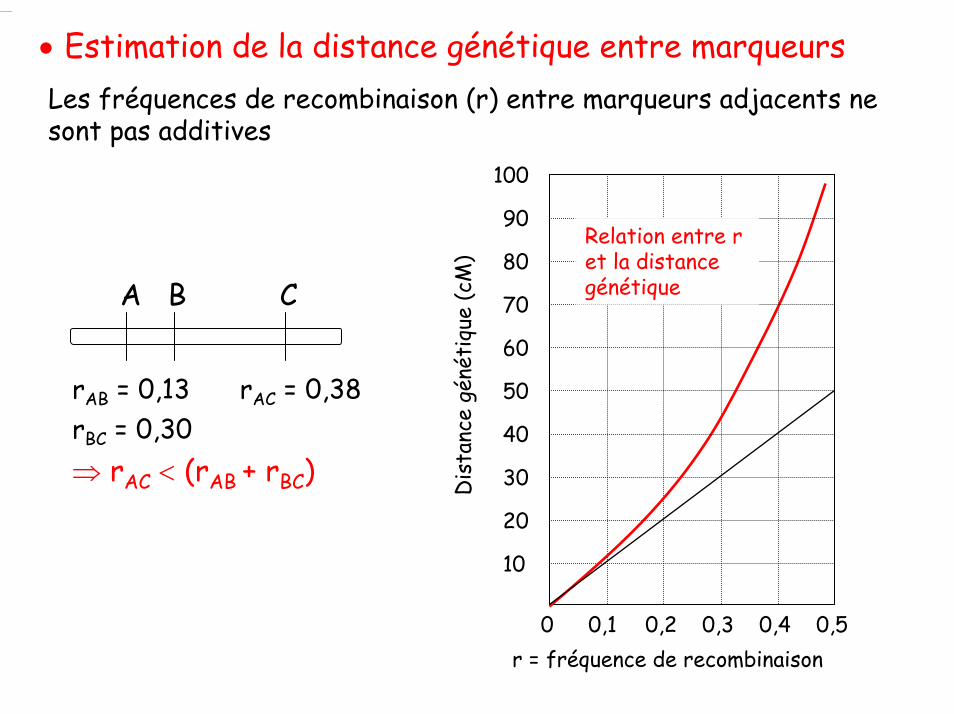
• Estimation de la distance génétique entre marqueursLes fréquences de recombinaison (r) entre marqueurs adjacents ne sont pas additives
A B C
rAB = 0,13rBC = 0,30
rAC = 0,38
⇒ rAC < (rAB + rBC)
0,1 0,2 0,3 0,4 0,50
10
20
30
40
50
r = fréquence de recombinaison
Dis
tanc
e gé
néti
que
(cM
)
60
70
80
90
100
Relation entre r et la distance génétique

Fonction de Haldane
0,1 0,2 0,3 0,4 0,50
10
20
30
40
50
r = fréquence de recombinaison
Dis
tanc
e gé
néti
que
(cM
)
60
70
80
90
100
Fonction de Kosambi
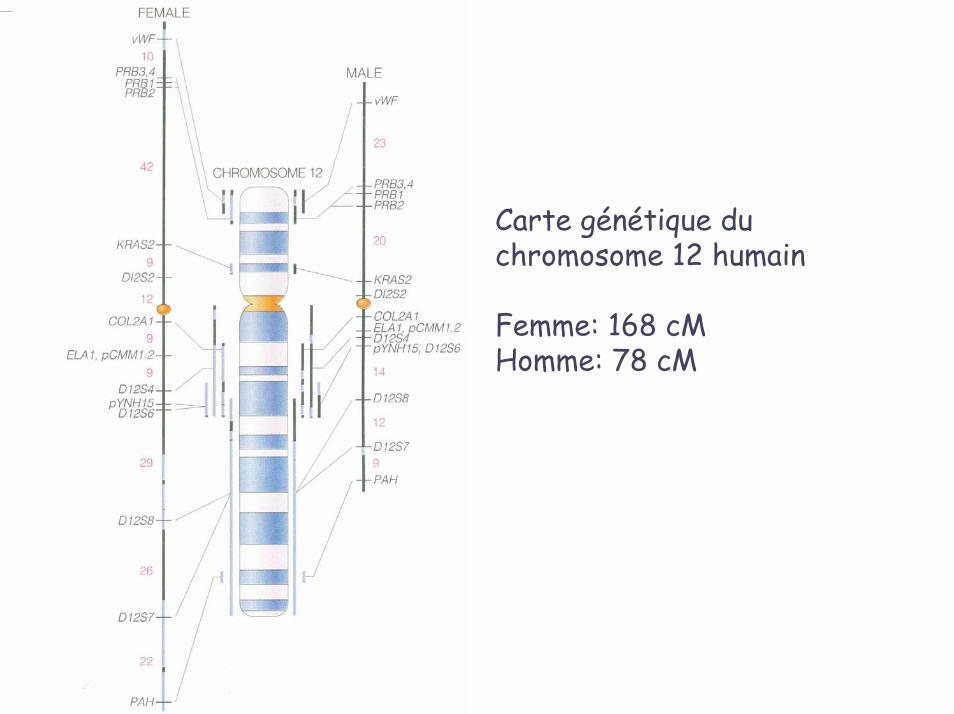
Carte génétique du chromosome 12 humain
Femme: 168 cMHomme: 78 cM
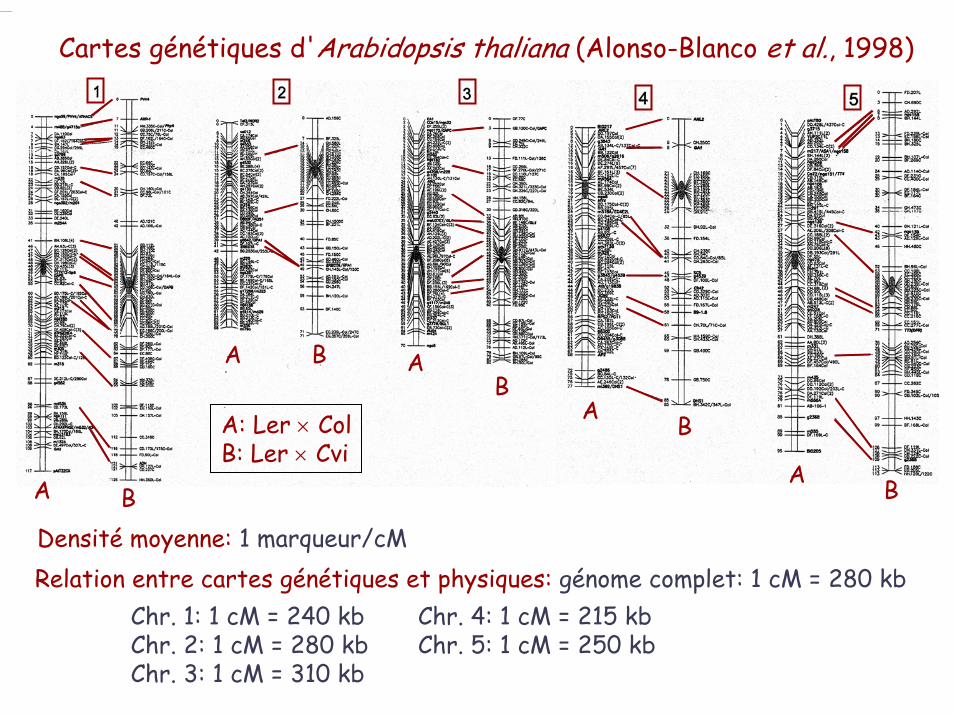
Cartes génétiques d'Arabidopsis thaliana (Alonso-Blanco et al., 1998)
A
A A
A
AB
BB
B
B
A: Ler × ColB: Ler × Cvi
Densité moyenne: 1 marqueur/cMRelation entre cartes génétiques et physiques: génome complet: 1 cM = 280 kb
Chr. 1: 1 cM = 240 kb Chr. 4: 1 cM = 215 kbChr. 2: 1 cM = 280 kb Chr. 5: 1 cM = 250 kbChr. 3: 1 cM = 310 kb
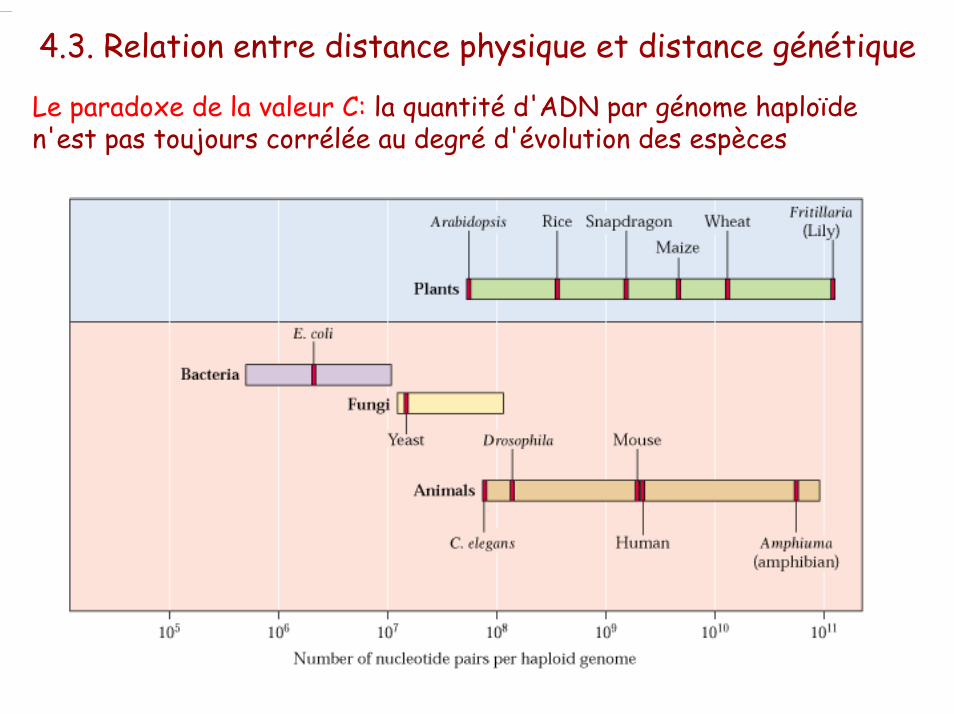
Le paradoxe de la valeur C: la quantité d'ADN par génome haploïde n'est pas toujours corrélée au degré d'évolution des espèces
4.3. Relation entre distance physique et distance génétique
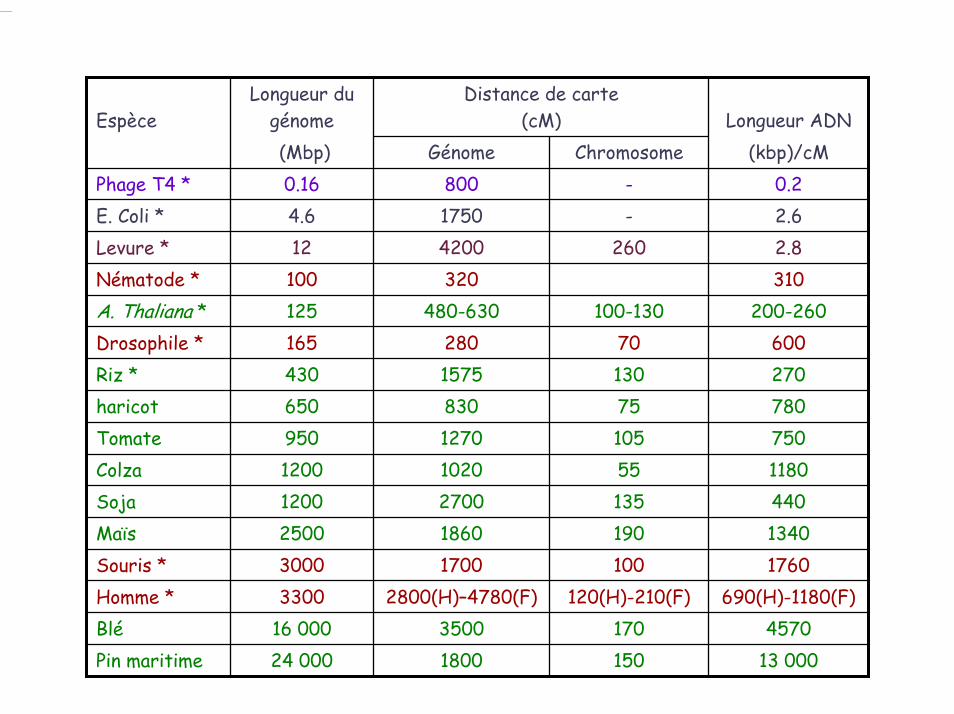
EspèceLongueur du
génomeDistance de carte
(cM) Longueur ADN(Mbp) Génome Chromosome (kbp)/cM
Phage T4 * 0.16 800 - 0.2
haricot 650 830 75 780
Soja 1200 2700 135 440
Homme * 3300 2800(H)–4780(F) 120(H)-210(F) 690(H)-1180(F)Blé 16 000 3500 170 4570
Maïs 2500 1860 190 1340Souris * 3000 1700 100 1760
Tomate 950 1270 105 750Colza 1200 1020 55 1180
E. Coli * 4.6 1750 - 2.6Levure * 12 4200 260 2.8 Nématode * 100 320 310A. Thaliana * 125 480-630 100-130 200-260Drosophile * 165 280 70 600Riz * 430 1575 130 270
1800Pin maritime 24 000 150 13 000