carrier-mediated transport · pdf file• lodish et al (2004) ... ed. pp 439-442 and...
TRANSCRIPT

Dale Sanders
6 March 2009
Module 0220502
Membrane Biogenesis and Transport
Lecture 14
Carrier-Mediated Transport

Aims:By the end of the lecture you should
understand…
• The mechanism of action of valinomycinand its significance for proteinaceouscarriers;
• The physiological significance of someimportant carriers (ion-coupled or not);
• The energetics of ion-coupled solutetransport;
• The basic molecular attributes of the lacpermease.

Reading
• Lodish et al (2004) Molecular Cell Biology, 5th
ed. pp.268-271
• Lodish et al.(2008) Molecular Cell Biology 6th
ed. pp 439-442 and 465-470
• Abramson et al (2003) Structure andmechanism of the lactose permease ofEscherichia coli. Science 301: 610-615
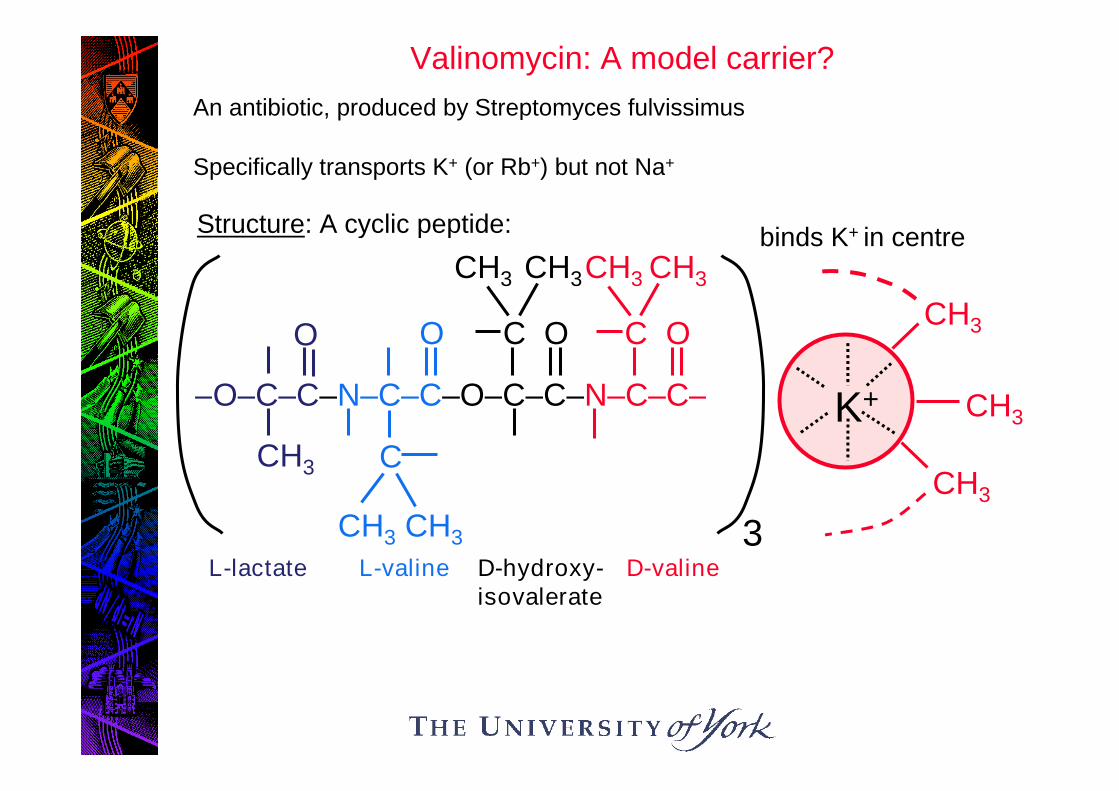
Structure: A cyclic peptide:
CH3
K+ CH3
CH3
–O–C–C–N–C–C–O–C–C–N–C–C–
CH3
O O OC
CH3 CH3
OC
CH3 CH3
C
CH3 CH3
L-lactate L-valine D-hydroxy-isovalerate
D-valine
binds K+ in centre
3
Valinomycin: A model carrier?
An antibiotic, produced by Streptomyces fulvissimus
Specifically transports K+ (or Rb+) but not Na+

Transport mechanism• Co-ordination by = O groups of K+ replaces its hydration shell.
• Selectivity for K+ (over Na+) because lower charge density of K+
gives weaker free energy of hydration.
• K+-val complex diffuses across bilayer and dissociates on otherside.
• Free val diffuses back. H2O H2OK+
H2O H2O
K+
K+
K+Valinomycin has been a usefulmodel for KINETIC descriptionof carrier action, but not forMOLECULAR MECHANISM

Kinetic Properties of Carriers……indicate that transport binding sites are exposed alternately toeach side of membrane – just as for valinomycin – i.e….
So
Co CSo
Ci
Si
CSi
S = transported soluteC = carrier binding site
out
in

This kinetic scheme predicts Michaelis-Mentenkinetics, which are observed for almost all carriers.
Binding site becoming saturated:transmembrane reactions rate-limiting
Binding site unsaturated:linear increase in Flux ([S])
Influx ofradiolabelled
S(mol . s-1)
[S]o (M)
Therefore, there is a Km and Vmax for transport

NOPERSONis
allowed
toaccompany
in
this
LIFT
LIQUID
NITROGEN
(or
any
other
asphyxiant
gases)
Mechanistic implications
for large protein carriers, transport must involve
CONFORMATIONAL CHANGE in protein e.g.
SCo CSo CiCSi
S
S
S

Physiologically Important Carriers
A. Systems not coupled to “Driver ions”
1. Glucose carrier, mammalian cells
Function Glucose entry to cells, from blood.
Regulation Glucose uptake is insulin-stimulated in muscle,adipose tissue, liver:
insulin recruits more carrier to plasmamembrane
glu glu

2. Cl–/HCO3- exchanger
Location: red blood cells (and others)
Function CO2 removal from tissues.
Catalyses uptake of HCO3- at high PCO2
(tissues)
release of HCO3- at low PCO2
(lungs)
Cl–
HCO3
Cl–HCO3
i.e. Functionally reversible
Sometimes known as “Band 3”: Comprises 25% membrane protein

In some cells, Cl-/HCO3- exchanger contributes to pHi
regulation:
Cl–
H2CO3
CO2
H+ + HCO3
i.e. generatesH+ to cope withalkaline stress

B. Systems coupled to “Driver ions”
1. Na+ - coupled sugar and amino acid transporters(many)
• Location: brush border membranes of epithelia
• Function: absorption of nutrients from food and urine
nutrients then accumulated in cells lining lumen
Na+
Sugar

2. Na+/ H+ antiporter
Na+
H+
• Location: Ubiquitous in animal cell plasma membranes
• Function: regulation of pHi : removes H+ from cell
ie copes with acid stress; also urinaryacidification
Antiporter acutely controlled by pHi….
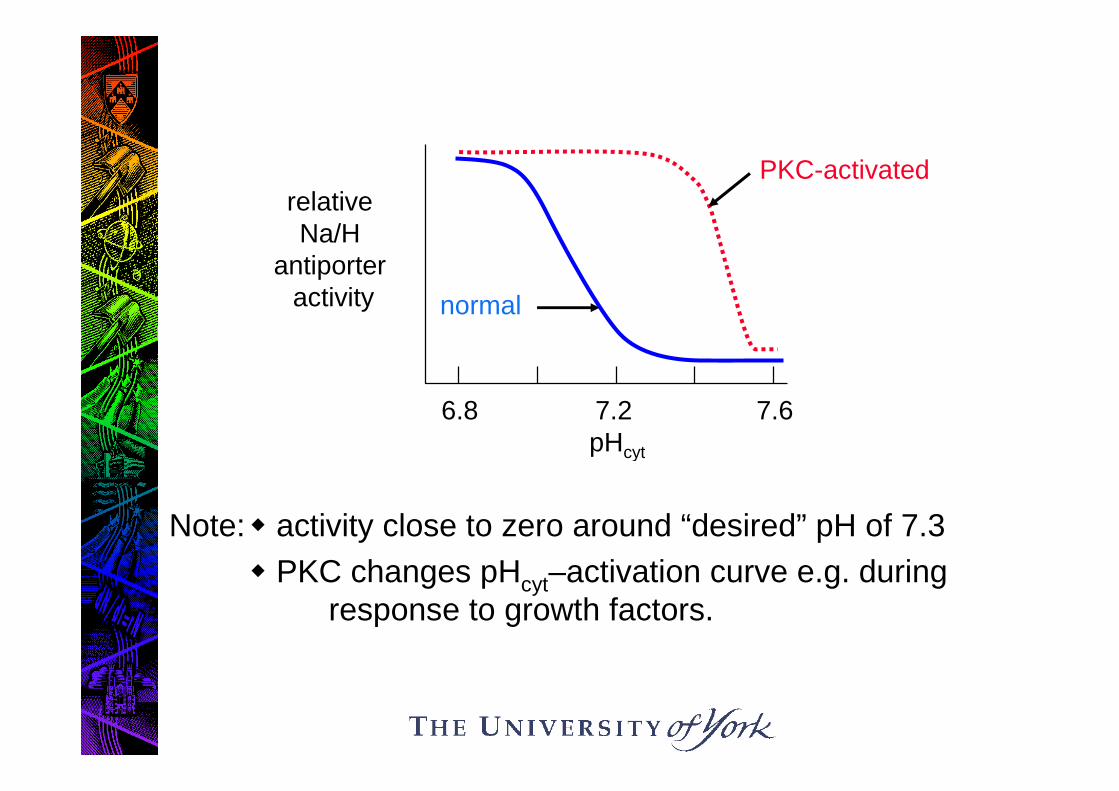
Note: activity close to zero around “desired” pH of 7.3
PKC changes pHcyt–activation curve e.g. duringresponse to growth factors.
normal
relativeNa/H
antiporteractivity
6.8 7.2 7.6pHcyt
PKC-activated

3. Na+, K+, 2CI- symporter
• Location: Plasma membrane, animal cells.
• Function: activated in hypertonic conditions (cellshrinkage)
Salt absorption by cells water uptake, hencevolume recovery
Na+
K+
2Cl–

4. H+ - sucrose symporter
Drawing 17
• Location: Plasma membrane of plant cells
• Function: Loads sucrose into phloem of leavesfor transport to roots, growing leaves,fruits.
H+
sucrose

How Big are the Solute Accumulation Ratios that can beGenerated by Coupled Transport? (1)
nH+
S
e.g. Proton symportin a bacterium
Can write as nH+o + So nH+
i + Si
G = n H + S, so at equilibrium, -n H = S
Since PMF = H/F,
-nF(PMF) = S
= RT.ln ([S]i/[S]o)
Thus -(nF/RT) (PMF) = 2.3 log10 ([S]i/[S]o)
and [S]i/[S]o = 10
Substituting constants:
[S]i/[S]o = 10 Eq (1)
((-nF/2.3RT)(PMF))
(-17n (PMF))

For bacterial plasma membranes, typically, (pH0 – pHi) might be-1 unit and might be –200 mV so
PMF = 59 (pH0 – pHi) + = -259 mV or -0.259 V
How Big are the Solute Accumulation Ratios that can beGenerated by Coupled Transport? (2)
2.00x10151.58x10101.26x10513
1.58x10106.31x1062.51x10312
1.26x1052.51x1035011
PMF =
-0.3 V
PMF =
-0.2 V
PMF =
-0.1 V
PMF =
0 Vn
Applying Eq 1, we can calculate maximum solute accumulationratios ([S]I/[S]o) for various values of n and PMF

Evolutionary Relationships of Carriers
Homology between many carriers suggests that they belong tothe same superfamily:
The Major Facilitator Superfamily (MFS)
• 25% of bacterial membrane transporter genes
Typified by:
• Single transport-active polypeptide
• Very hydrophobic
• Often12 transmembrane spans, sometimes with evidence of aninternal repeat and hydrophilic insert between helices 6 & 7

Key: Tet A: Tetracycline resist. efflux E. coli
AraE: Arabinose – H+ symport E. coli
Hup-1: Hexose - H+ symport Chlorella
Gtr-2: Glucose uniport mouse liver
Cit A: Citrate - H+ symport E. coli
UhpT: Hexose – phosphate antiport E. coli
LacY: Lactose - H+ symport E. coli
Simplified tree:
5 subfamilies (of a totalof 58) [Law et al (2008) Annu.
Rev. Microb. 62: 289]
12
3
45
Lac Y
Uhp T
Cit A
Gtr2
Hup-1Ara E
Tet A

Note: The MFS does not include Na+ coupledtransporters of animal cells.
Sub-group 2 comprises both H+ coupled and“passive” (uniport) systems.
Conclude: H+ - coupled and Na+ -coupled transport evolvedindependently
H+ - coupled transport and passive transportsystems closely related.
Note also: LacY system rather displaced from other MFSmembers. Nevertheless, a paradigm for molecularmechanistic studies.

Molecular Mechanisms: The LacPermease
Lactose: β 1, 4 dimer of glu & gal
LacY gene product: 417 residues
Mr = 46,500, sequenced 1980
12 transmembrane spans (I – XII)
Functional monomer
Landmark study in 2003: 3D crystalstructure solved…

Structure of the Lac Permease
Abramson et al (2003) Science 301: 610-615

Structural changes between the inwardand outward facing conformations of the
lac permease: The “Rocker Switch”Mechanism
Abramson et al (2003) Science 301: 610-615

Mechanism of H+ Coupling
This structure, together with site directed mutagenesis/covalentprobes possible model
4 Critical residues, interacting
R302 (IX) – E325 (X) Salt bridge
H322 (X) - E269 (VIII) H – bond
Substrate binding: VIII – V
H binds to E269
S binds, structural change between VIII and V disrupts E269 –H322 interaction
E269 donates H+ to E325, inaccessible to water, anddisplacing E325 from R302
E269 salt bridges with R144, leading to binding sitereorientation
H+ lost from E325, through crevice between IX and X
S released
R302 – E325 interaction re-established, binding sitereorientation

Possible lactose/proton symportmechanism
Abramson et al (2003) Science 301: 610-615

Summary
1. The carrier valinomycin exhibits carrier type kinetics, but isnot a good molecular model for most carriers.
2. Some physiologically important carriers are uniporters, whilesome are symporters and antiporters; present in all kingdoms
3. By coupling solute transport to that of a “driver” ion, verylarge solute accumulation ratios are possible
4. Most carriers belong to the Major Facilitator Superfamily
5. H+- coupled lactose transport by the lac permease involves a“rocker switch” mechanism and intramembrane chargedresidues