carbohydrate recognition mechanism of the mushroom
TRANSCRIPT

© 2018 FCCA (Forum: Carbohydrates Coming of Age)SE75
Trends in Glycoscience and GlycotechnologyVol. 30 No. 172 (January–May 2018) pp. SE75–SE88
Carbohydrate Recognition Mechanism of the Mushroom Galectin ACG
Jun Hirabayashi1; Dan Hu1, †; Hiroaki Tateno1; Naoyuki Kuwabara2; Ryuichi Kato2; and Fumio Yagi3
1 National Institute of Advanced Industrial Science and Technology, AIST Tsukuba Central 2, 1–1–1, Umezono, Tsukuba, Ibaraki 305–8568, Japan
2 High Energy Accelerator Research Organization (KEK), Tsukuba, Japan3 Faculty of Agriculture, Kagoshima University, Kagoshima, Japan
† Present address: Institute of Traditional Chinese Medicine and Natural Products, College of Pharmacy, Jinan University, Guangzhou 510632, China
FAX: +81–29–861–3125, TEL: +81–29–861–3124, E-mail: [email protected]
(Received on August 20, 2017, accepted on October 12, 2017)
Key Words: Agrocybe cylindrace, mushroom galectin, definition, cis-trans isomerization, molecular evolution, GalNAcα1-3Gal
AbstractThe fruiting body of the edible mushroom Agrocybe cylindracea contains a proto-type galectin named ACG, which shows af-
finity to a wide range of β-galactosides. Unlike other galectins, it also binds to disaccharides (GalNAcα1-3Gal/GalNAc) found in blood group A and Forssman epitopes. Structurally, ACG lacks an evolutionarily conserved Asn located on the S4 strand (Ala64) but is compensated for by Asn46, which is located on the extended loop region unique to this galectin. A recent site-directed mutagenesis study revealed that the N46A mutant had selective affinity to oligosaccharides containing GalNAcα1-3Gal/GalNAc epitopes (Hu et al., (2013) Biochem. J., 453, 261–270). Taking into consideration the previous observation that Pro45 takes the cis conformation, Hu et al. assumed that ACG has evolved to attain two conformations at the imide group of Pro45: a cis conformation, where it can recognize β-galactosides, and a Pro45-tras conformation while it binds to the unique GalNAcα1-3Gal/GalNAc. The proposed dual recognition mechanism was proved through further site-directed mutagenesis and X-ray crystallography analysis. Notably, N46A recognizes even non-reducing terminal disaccharides, GalNAcα1-3Gal/GalNAc. Thus, the one face of this “Janus-type” lectin fulfills the conventional definition of galectins, whereas the other face does not. A possible scenario of galectin deviation is discussed.
A. IntroductionSince the definition of a galectin was proposed in 1994 (1)*,
a basic question remains: “Why is the specificity of galectins confined to β-galactosides?” Binding to such β-galactosides oc-curs on the evolutionarily conserved region, which is composed of S4–S6 strands on the β-sheet surface of the β-sandwich struc-ture with jelly-roll topology (3). A few galectins are known to bind polysaccharides derived from citrus pectin, and thereby in-hibit galectin function, e.g., tumor promotion and metastasis by galectin-3 (4–8), in which binding involves galactosides and the canonical side on the galectin. However, recent reports suggest that this binding should occur at a noncanonical site, rather than the galectin-original carbohydrate-recognition domain (CRD; 9–11). Therefore, their conserved features in the canonical CRDs are primarily for the recognition of various β-galactoside disaccha-
rides, including type 1/2 LacNAc (Galβ1-3/4GlcNAc), LacdiNAc (GalNAcβ1-4GlcNAc), Galβ1-4Man, GalNAcβ1-4GlcA, Gal(NAc)β1-3Gal(NAc), and Galβ1-3Fuc (12, 13). Nevertheless, there are a few exceptional galectins with unique sugar-binding specificities. In this review, we will discuss a galectin from the mushroom Agrocybe cylindracea, which has broad specificity, and can bind to lactose and its 3′OH-modified derivatives, including α2-3sialyllactose, 3′-sulfo-lactose and A-tetrasaccharide, as well as non-β-galactoside disaccharides, such as GalNAcα1-3Gal(NAc).
The galectin known as ACG was originally isolated from the mushroom A. cylindracea by Yagi et al., who described it as a sol-uble lectin with strong interactions with glycoconjugates contain-ing Neu5Acα2,3Galβ1,3GlcNAc-/GalNAc sequences (14). Later, it was found to be a member of the galectin family (15). ACG is a proto-type galectin (16), forming a dimer of 16,500-Da subunit, which is somewhat larger than other proto-type galectins (i.e., 14,000–15,000 Da). This is due to the presence of an additional β-strand (designated here F0), as well as a redundant loop bridging the F2 and S3 strands (Fig. 1; 17).
The physiological function of ACG is not clear, although a galectin from another mushroom, Coprinopsis cinerea (CCG2), has anti-nematode activity through the recognition of Caenorhab-ditis elegans N-glycan core Galβ1-4Fuc structures (18). Antitumor
MINIREVIEWdoi: 10.4052/tigg.1736.1SE
* In 1994, galectins were defined based on the consensus between 21 galec-tin researchers at the time, as follows: membership in the galectin family requires fulfillment of two criteria: one is “affinity for β-galactosides” and the other is “significant sequence similarity in the carbohydrate-binding site, the relevant amino acid residues of which have been determined by X-ray crystallography (2).” They are represented by His44, Asn46, Arg48, Asn61, Trp68, Glu71, and Arg73 (residue numbers are those of human galectin-1) by consensus between studies related to galectins.
(Article for special issue on Galectins)

© 2018 FCCA (Forum: Carbohydrates Coming of Age) SE76
activity has also been reported in another mushroom galectin, AAG (designated as such for Agrocybe aegerita galectin to distinguish it from the well-known Aleuria aurantia lectin), isolated from the same genus as ACG (19, 20). AAG shows striking resemblance to ACG (90% amino-acid identity), and the above mentioned ad-ditional F0 β-strand and redundant loop between F2 and S3 strands are conserved between them (as described in section F). At pres-ent, however, the detailed sugar-binding specificity of AAG has not been investigated. A new mushroom galectin isolated from Laccaria amethystine (LAG) was recently described, which has 35–65% amino-acid identity with galectins from Agrocybe spp. and C. cinerea (21). Three galectin-related genes have been identi-fied in the genome of another Laccaria species (L. bicolar) (desig-nated LBG1-3r, 22). Although functional analysis of these genes has not yet been conducted, LBG1 and 2 have extensive similarity to other mushroom galectins, and conserve most amino acid resi-
dues that are essential for β-galactoside binding activity, including His44, Asn46, Arg48, Asn61, Trp78, Glu81, and Arg83 (residue numbers are for human galectin-1, the detailed binding features of which are presented in Fig. 2) (23).
Imamura et al. previously attempted saturation mutagenesis of ACG focusing on Glu86 (residue number is of ACG), aiming at elimination of conventional β-galactoside-binding activity while preserving high affinity to acidic oligosaccharides (25). More re-cently, Hu et al. carried out comprehensive mutagenesis targeting the hydrophilic residues (i.e., Asn46, His62, Arg66, Asn75, and Glu86) involved in the sugar binding of ACG, and found that sub-stitution of Asn46 with alanine (N46A) resulted in successful sepa-ration of ACG’s unique specificity to GalNAcα1-3Gal/GalNAc from its conventional specificity to β-galactosides (26). Further studies made use of extensive mutagenesis and X-ray crystallog-raphy for both the wild-type ACG and its mutant N46A. ACG is
Fig. 1. a, Amino acid sequence alignment of ACG and other galectins. Abbreviations; ACG: Agrocybe cylindracea galectin; CCG-2: Coprinop-sis cinerea galectin isolectin-2; hGal-1: human galectin-1; hGal-7: human galectin-7; Cg-16: chicken 16K galectin. Five plus 1 β-strands named F0–F5 and six β-strands named S1–6 are shown in blue and red horizontal bars above the sequences. The redundant loop region located between F2 and S3 strands is unique to the mushroom galectin ACG, where the pentapeptide GSPNN (shown in red) is inserted. The truncated loop region between S4 and S5 strands here is shorter than in some vertebrate galectins (hGal-1 and Cg-16). b, Scheme of the β-strand features of ACG. F0–F5 and S1–S6 strands are shown with blue and red arrows, respectively. In mushroom galectins ACG and AAG, the Asn46 residue located in the redundant loop between F2 and S3 participates in carbohydrate recognition if the preceding Pro45 takes the cis conformation (residue numbers are of ACG). Mush-room galectins have a deletion of a tetrapeptide that exists in vertebrate galectins between the S4 and S5 strands, thus forming a truncated loop.

© 2018 FCCA (Forum: Carbohydrates Coming of Age)SE77
a good protein scaffold, and is equipped with potential diversity in carbohydrate specificity and productivity in Escherichia coli (>10 mg/mL culture as soluble protein; 27).
B. Taxonomy of Fungi and Fungal LectinsBefore discussing the molecular details of ACG, the bio-
logical position of fungi, to which A. cylindracea belongs, will be briefly described. The kingdom of fungi forms a large group of eukaryotic microorganisms, represented by yeast, molds, and mushrooms. Taxonomically, fungi are further classified into the following taxa: subkingdom Dikarya (phyla Ascomycota and Basidiomycota); the phyla Glomeromycota, Zygomycota, Blas-tocladiomycota, Neocallimastigomycota, and Microsporidia; and the Chytridiomycota. Although fungi are physically small, and are therefore often referred to as the ‘Forgotten Kingdom,’ they fulfill vital roles in the ecosystems where they live by contributing to car-bon cycling and forming symbiotic relationships with other nearby organisms (28).
Mushrooms have been used in traditional Chinese medicines or functional foods, and are attractive research subjects, especially
in Asian countries including China and Japan. Several bioactive components have been isolated from mushrooms, including poly-saccharides (e.g., β-glucan), glycoconjugates (e.g., polysaccharo-peptides, polysaccharide-protein complexes), hydrolytic enzymes (e.g., proteases, ribonucleases), ribosome-inactivating proteins, antifungal proteins, immunomodulatory proteins, and lectins. Mushroom lectins are often endowed with mitogenic, antiprolifera-tive, antitumor, antiviral, and immune stimulating properties (29). However, there remain barriers to their practical use in ‘evidence-based medicine.’ Lectins are a group of mushroom proteins for which their carbohydrate-binding activity is likely concerned with extensive biological phenomena as well as medicinal opportunities.
Varrot et al. (30) described various roles for fungal lectins, in-cluding storage, development, parasitism, infections, flocculation, and mating (18, 31). Fungal lectins have attracted increasing inter-est in biotechnological applications. They provide a rich source of new lectins with unique carbohydrate specificity and practical properties (e.g., stability and productivity). Well-studied and com-mercially available lectins include Agaricus bisporus lectin (ABL), Agrocybe cylindracea galectin (ACG), Aleuria aurantia lectin
Fig. 2. Binding features of human galectin-1 and N-acetyllactosamine. a, Evolutionarily conserved hydrophilic residues are involved in the sugar recognition via hydrogen bonding. The 4-OH group of Gal is hydrogen-bonded to His44, Asn46, and Arg48 on the S4 strand. The 6-OH of Gal is hydrogen-bonded to Asn61 on the S5 strand and to Glu71 on the S6 strand. The remaining hydroxyl group, the 3-OH of GlcNAc, is hydrogen-bonded to Arg48 on the S4 strand, and Glu71 and Arg73 on the S6 strand. Trp68 interacts with the hydrophobic side (B face) of the pyranose ring of Gal. Original data are from PDB: 1SLT (23). b, Schematic drawing of galectin-1 and N-acetyllactosamine binding. Original data are from Lobsanov and Rini (24).
© 2018 FCCA (Forum: Carbohydrates Coming of Age) SE78
(AAL), Aspergillus oryzae lectin (AOL), Clitocybe nebularis lectin (CNL), Marasmius oreades lectin (MOL), Psathyrella velutina lectin (PVL), Rhizopus stolonifer lectin (RSL), Pholiota squarrosa lectin (PhoSL), and Polyporus squamosus lectin (PSL). For com-prehensive descriptions of their properties and structures, see re-cent reviews by Varrot et al. (30), and Kobayashi and Kawagishi (32).
Fungi are endowed with an arsenal of lectins. According to Shubert et al., because of their lack of motility and thus inability to escape from predators and parasites, both multicellular fungi and plants developed very similar defense strategies. Without adaptive immune mechanisms and circulating immune cells, they must rely solely on “innate defense” (33). Previous reports have noted that among various potential fungal defense proteins so far identified, the number as well as the diversity of lectins is remarkably high (34–37). Interestingly, this may be related to their renowned abun-
dance and stability, which support reproducibility and long-term survival for fruiting bodies and sclerotia, respectively. Thus, fungal lectins are promising targets for both their medicinal and biotechni-cal applications.
C. Sugar-binding Features of the A. cylindracea Galectin, ACG: Broad Specificity to β-galactosides and Non-β-galac-tosides
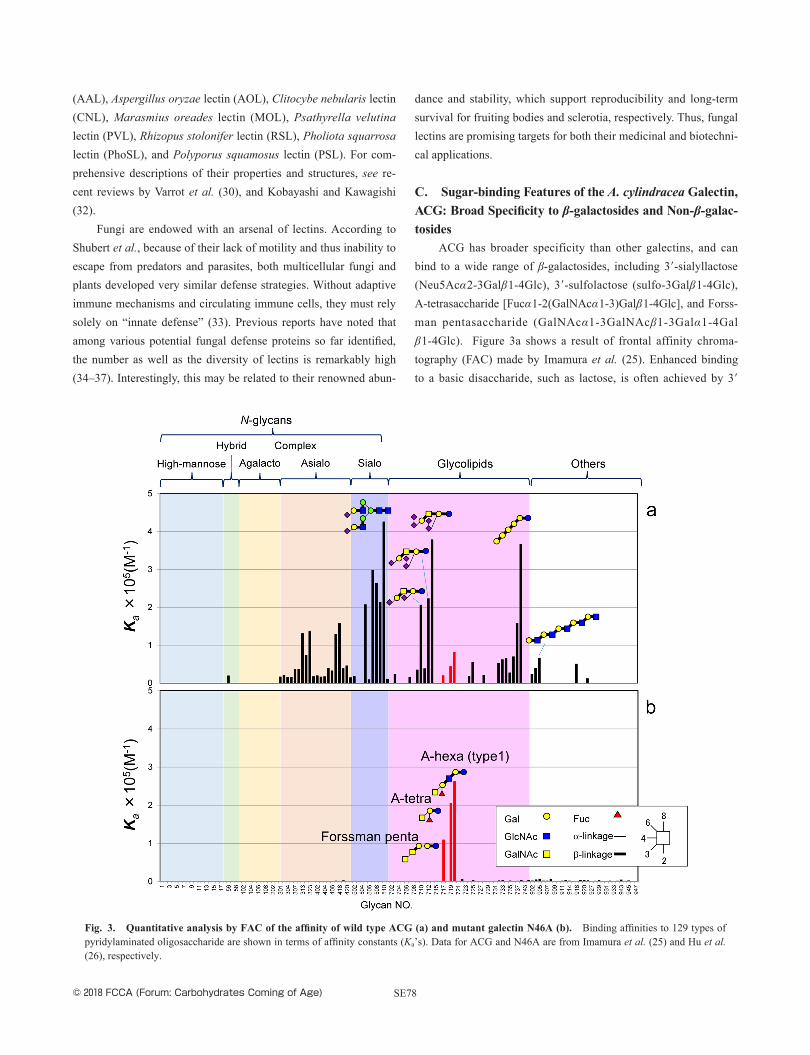
ACG has broader specificity than other galectins, and can bind to a wide range of β-galactosides, including 3′-sialyllactose (Neu5Acα2-3Galβ1-4Glc), 3′-sulfolactose (sulfo-3Galβ1-4Glc), A-tetrasaccharide [Fucα1-2(GalNAcα1-3) Galβ1-4Glc], and Forss-man pentasaccharide (GalNAcα1-3GalNAcβ1-3Galα1-4Galβ1-4Glc). Figure 3a shows a result of frontal affinity chroma-tography (FAC) made by Imamura et al. (25). Enhanced binding to a basic disaccharide, such as lactose, is often achieved by 3′
Fig. 3. Quantitative analysis by FAC of the affinity of wild type ACG (a) and mutant galectin N46A (b). Binding affinities to 129 types of pyridylaminated oligosaccharide are shown in terms of affinity constants (Ka’s). Data for ACG and N46A are from Imamura et al. (25) and Hu et al. (26), respectively.

© 2018 FCCA (Forum: Carbohydrates Coming of Age)SE79
substitution (e.g., sulfonyl group, αNeu5Ac, αGalNAc) with the non-reducing terminal Gal/GalNAc, as is understood in the con-cept of subsite specificity (38). In fact, various galectin inhibitors have been developed, with the aim of developing galectin-targeted pharmaceutical drugs (39–41). As in the cases of galectin-3 and galectin-9 CRDs, affinity enhancement is evident in ACG for the repeating LacNAc unit (i.e., LacNAcβ1-3), while affinity increases for oligolactosamines, such as LacNAc2, LacNAc3, and LacnAc5, are rather modest in ACG. As described in section D, however, the broad sugar specificity of ACG is not only attributed to the C3′ substitution of lactose, because we know that ACG has substantial binding in a glycoconjugate microarray to the Forssman disaccha-ride GalNAcα1-3GalNAc, which is no longer categorized as a β-galactoside, like lactose (26).
It is widely accepted that the evolutionarily conserved hydro-philic residues of galectins are important to exert their sugar bind-ing function through hydrogen bonding (Fig. 2). In fact, even when conservative substitutions are made for human galectin-1 at His44, Asn46, Arg48, Asn61, Glu71, and Arg73, the resultant mutants, H44Q, N46D, R48H, N61D, E71Q, and R73H completely lost their binding activity (42, 43). This is essentially true for galectins of evolutionarily distant species, e.g., Caenorhabditis elegans, be-cause these hydrophilic residues are strongly conserved in relation to β-galactoside-binding properties, together with Trp68 (residue number is of human galectin-1; 44), which contributes to stabiliza-tion by stacking to the hydrophobic face of β-galactosides (38).
So far, there are two reports of the amino acid sequences of Agrocybe galectins: one from A. cylindracea (ACG) and the other from A. aegerita (AAG). Interestingly, one of the evolutionarily conserved hydrophilic residues, corresponding to Asn46 in human galectin 1, is substituted with Ala in both ACG and AAG (residue number 64 in ACG with initial Met1, Fig. 1a). According to a previous X-ray crystallography analysis by Ban et al. (17), the ab-sence of this Asn is compensated for by another Asn (residue num-ber 46 in ACG). The crystallography analysis located this Asn46 in an extended loop region linked between F2 and S3 β-strands in ACG, i.e., Gly-Ser-Pro-Asn-Asn (Asn46 underlined), while other galectins lack this region. Intriguingly, Ban et al. found that Pro45, preceding Asn46, has an unusual cis conformation, which allows for binding of lactose (17).
D. Systematic Mutagenesis Unveiled an Alternative Binding Mechanism in ACG to Recognize GalNAcα1-3Gal/GalNAc
Hu et al. attempted comprehensive mutagenesis targeting hy-drophilic amino acid residues involved in the sugar-binding func-tion of ACG, with the aim of narrowing the broad sugar-binding
specificity of ACG. They included the evolutionarily conserved His62, Arg66, Asn75, and Glu86, and the non-conserved Asn46 (26). The results were quite unexpected: when Ala was substituted for either conserved His62, Arg66, Asn75, or Glu86 in ACG (cor-responding to His44, Arg48, Asn61, or Glu71, respectively, in hu-man galectin-1), all of the derived mutants (H62A, R66A, N75A) lost binding activity, except E86A, which retained slight affinity for a few oligosaccharides (45). On the other hand, substitution of non-conserved Asn46 with alanine (N46A) resulted in much narrower specificity with respect to GalNAcα1-3Gal/GalNAc-containing oligosaccharides, while the mutant could not bind β- galactosides at all, including lactose/LacNAc. Figure 3b shows the result of quantitative specificity analysis by FAC of the N46A mu-tant (26).
Notably, the binding affinity to the following oligosaccharides apparently increased in the mutant, in the order of affinity in terms of dissociation constant (Kd): blood group A antigen type 1 hexasac-charide [Fuca1-2(GalNAcα1-3) Galβ1-3GlcNAcβ1-3Galβ1-4Glc, Kd determined by FAC=3.8 µM], A-tetrasaccharide [Fuca1-2(GalNAcα1-3) Galβ1-4Glc, 5 µM], and Forssman pentasaccharide (GalNAcα1-3GalNAcβ1-3Galα1-4Galβ1-4Glc, 10 µM). The com-mon structural feature of these oligosaccharides is the non-reducing terminal GalNAcα1-3Gal/GalNAc. Hu et al. found that the Forss-man disaccharide is also a good ligand for N46A using a glyco-conjugate microarray assay system (Fig. 4). It is therefore puzzling how the N46A mutant could bind the non-β-galactoside disaccha-ride GalNAcα1-3Gal/GalNAc. The wild-type ACG had substantial binding to A-tetrasaccharide, whereas it did not bind to Forssman disaccharides (GalNAcα1-3Gal). This observation implies that the mutant ACG (N46) attains a different structure from that of the wild type, which defines their sugar-binding specificities.
Based on the assumption that ACG takes two different struc-tural conformations related to cis-trans isomerization of Pro45 (Fig. 5), Hu et al. constructed further ACG mutants, N46X, in which Asn46 is replaced with every other amino acid. Hu et al. found that all of the 18 N46X mutants had almost the same binding profile as N46A according to the glycoconjugate microarray (Fig. 6), with each mutant showing selective binding to GalNAcα1-3Gal/Gal-NAc. It is therefore likely that Asn46 is involved in β-galactoside-binding to compensate for the lack of a conserved Asn64 (if con-served), which requires the cis conformation of Pro45 (17). On the other hand, all of the N46X mutants should exclusively take the trans conformation, because they can no longer bind β-galactosides, thus tethering the structurally more favorable trans conformation responsibility for a different specificity, like GalNAcα1-3Gal/GalNAc. If this is true, and Pro45 plays a key role in the specific-ity “switch,” P45X mutants, which are also expected to exclusively

© 2018 FCCA (Forum: Carbohydrates Coming of Age) SE80
Fig. 4. Glycoconjugate microarray analysis of wild type ACG (a) and mutant galectin N46A (b). Data are originally from Hu et al. (26)
Fig. 5. Proposed mechanism for the sugar-binding modes of ACG and its mutant N46A. Wild-type ACG binds to lactose with Pro45 in a cis conformation, so that the subsequent Asn46 can recognize the non-reducing terminal galactose by hydrogen bonding. The overall structures of ACG in the ligand-free form (yellow) and complex with lactose (green) are superimposed. The two β-sheets, F-sheet (F0–F5), and S-sheet (S1–S6) are indicated by letters. The area indicated by dotted lines in (a) is enlarged in (b). The relevant 3 amino acid residues, which are predicted to cause a sig-nificant conformational change upon Pro45-cis-trans transformation, i.e., Ser44, Pro45 and Asn46, are indicated by lines and labeled with letters. The peptide bond between Ser44 and Pro45 is emphasized by thick red lines. Data are from PDB: 1WW6 (ACG/lactose complex) and 1WW7 (ACG/free).

© 2018 FCCA (Forum: Carbohydrates Coming of Age)SE81
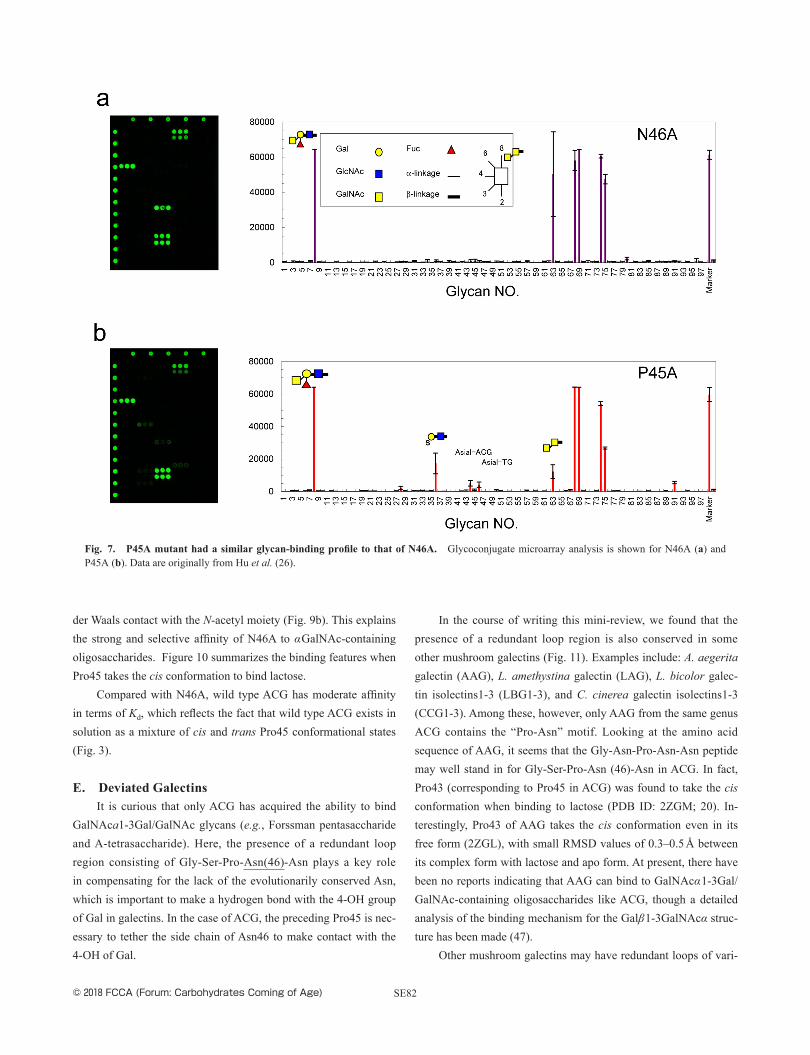
take the trans conformation, should show the same binding profile as N46X mutants. Continuing this idea, P45A mutant had a very similar binding profile to N46A, with selective binding to A-tetra-saccharide (Fucα1-2(GalNAcα1-3) Galβ1-4GlcNAc) and Forssman disaccharide (GalNAcα1-3GalNAc), but no binding to β-galacto-sides (Fig. 7).
These observations strongly suggest that Pro45 takes an un-usual cis conformation when binding to β-galactosides via Asn46, while it takes a trans conformation to attain a novel binding fea-ture and bind to GalNAcα1-3Gal/GalNAc, increasing the diver-sity of ACG’s specificity. In order to prove the above hypothesis, Kuwabara et al. performed detailed crystallography analysis for both wild type and mutant ACG (N46A) in their free forms, and in complexes with either lactose or A-tetrasaccharide, and com-pared the solved crystal structures with previously published ones by Ban et al. (17). Figure 8 shows the summary of overlap of all structures. Notably, the largest structural deviation is visible at residues 44–46, corresponding to Ser44-Pro45-Asn46. This clearly indicates that the redundant loop region, including residues 44–46, causes drastic conformational change upon cis-trans isomeriza-tion at Pro45. Kuwabara et al. concluded that: i) Pro45 takes the cis conformation when wild-type ACG is complexed with lactose, while it takes the trans conformation in a free state. ii) Wild-type
ACG also takes the trans conformation when it’s complexed with A-tetrasaccharide. iii) The trans conformation is consistently main-tained in the N46A mutant, both in its free state and in a complex with A-tetrasaccharide.
The detailed recognition mechanisms of this system are as follows: when wild-type ACG binds to lactose (a classic mode), Pro45 takes the cis conformation, and the subsequent residue Asn46 can compensate for the lack of the evolutionarily conserved Asn64 by forming hydrogen bonds with the 3-OH and 4-OH groups of the non-reducing terminal Gal. Other binding features are similar to conventional ones: the 4-OH of Gal is hydrogen-bonded to His62 (corresponding to His44 of human galetin-1), the 6-OH is hydrogen-bonded to Asn75 (Asn61), and the 3-OH of the reducing terminal glucose is hydrogen-bonded to Arg 66 (Arg48) and Arg 88 (Arg73). In addition, Trp83 (Trp69) and Glu86 (Glu71) are also involved in substrate recognition (Fig. 9a). Conversely, when wild-type ACG and the N46A mutant bind to A-tetrasac-charide (a novel mode), Pro45 takes the trans conformation, and the side chain of Asn46 is tethered in the opposite direction to the binding site. In this situation, a nitrogen atom of the indole group of Trp83 makes a new contact with the 6-OH group of GalNAc by hydrogen bonding. At the same time, the side chains of Asn47, Ala49, Asn140, and The142 come close to GalNAc and make Van
Fig. 6. All Asn46 mutants (N46X) had binding profiles similar to that of N46A. Glycoconjugate microarray analysis was performed for wild-type ACG and all the N46X mutants. The latter 19 mutants had essentially the same glycan-binding profiles. Data are originally from Hu et al. (26).
© 2018 FCCA (Forum: Carbohydrates Coming of Age) SE82
der Waals contact with the N-acetyl moiety (Fig. 9b). This explains the strong and selective affinity of N46A to αGalNAc-containing oligosaccharides. Figure 10 summarizes the binding features when Pro45 takes the cis conformation to bind lactose.
Compared with N46A, wild type ACG has moderate affinity in terms of Kd, which reflects the fact that wild type ACG exists in solution as a mixture of cis and trans Pro45 conformational states (Fig. 3).
E. Deviated GalectinsIt is curious that only ACG has acquired the ability to bind
GalNAca1-3Gal/GalNAc glycans (e.g., Forssman pentasaccharide and A-tetrasaccharide). Here, the presence of a redundant loop region consisting of Gly-Ser-Pro-Asn(46)-Asn plays a key role in compensating for the lack of the evolutionarily conserved Asn, which is important to make a hydrogen bond with the 4-OH group of Gal in galectins. In the case of ACG, the preceding Pro45 is nec-essary to tether the side chain of Asn46 to make contact with the 4-OH of Gal.
In the course of writing this mini-review, we found that the presence of a redundant loop region is also conserved in some other mushroom galectins (Fig. 11). Examples include: A. aegerita galectin (AAG), L. amethystina galectin (LAG), L. bicolor galec-tin isolectins1-3 (LBG1-3), and C. cinerea galectin isolectins1-3 (CCG1-3). Among these, however, only AAG from the same genus ACG contains the “Pro-Asn” motif. Looking at the amino acid sequence of AAG, it seems that the Gly-Asn-Pro-Asn-Asn peptide may well stand in for Gly-Ser-Pro-Asn (46)-Asn in ACG. In fact, Pro43 (corresponding to Pro45 in ACG) was found to take the cis conformation when binding to lactose (PDB ID: 2ZGM; 20). In-terestingly, Pro43 of AAG takes the cis conformation even in its free form (2ZGL), with small RMSD values of 0.3–0.5 Å between its complex form with lactose and apo form. At present, there have been no reports indicating that AAG can bind to GalNAcα1-3Gal/GalNAc-containing oligosaccharides like ACG, though a detailed analysis of the binding mechanism for the Galβ1-3GalNAcα struc-ture has been made (47).
Other mushroom galectins may have redundant loops of vari-
Fig. 7. P45A mutant had a similar glycan-binding profile to that of N46A. Glycoconjugate microarray analysis is shown for N46A (a) and P45A (b). Data are originally from Hu et al. (26).

© 2018 FCCA (Forum: Carbohydrates Coming of Age)SE83
Fig. 8. Overlap of the crystal structures for wild type ACG and the N46 mutant, both in free and complexed with ligand saccharides (lac-tose and A-tetrasaccharide). Data are originally from Kuwabara et al. (44). PDB IDs: 3WG1 (wild type and lactose), 3WG2 (N46A mutant, free), 3WG3 (wild type and A-tetrasaccharide), 3WG4 (N46A mutant and A-tetrasaccharide). Data for the wild type bound to lactose, PDB ID: 1WW6, from 15, is supplied as a reference.
Fig. 9. Two modes of binding mechanisms of ACG observed for lactose (wild type, a) and A-tetrasaccharide (wild type and N46A mutant, b). Original data are from Kuwabara et al. (46). PDB IDs: 3WG1 (wild type and lactose) and 3WG3 (wild type and A-tetrasaccharide).

© 2018 FCCA (Forum: Carbohydrates Coming of Age) SE84
ous lengths, but they lack the “Pro-Asn” motif in the exact cor-responding position. Nevertheless, it has been noted that the “Pro-Asn” motif may be found in somewhat distant positions (Fig. 11), such as Pro-Asn (36) in L. amethystina galectin (LAG) and Pro-Asn (37) in C. cinerea galectin-related isolectin 3 (CCG3r). Al-though crystallography analysis has not been performed for these galectins, it is possible to speculate that these Asn residues are involved in sugar-binding, with the cis conformation of the preced-ing Pro located in a flexible loop region encompassed by F2 and S3 strands. Detailed carbohydrate specificity has not been investigated for these mushroom galectins, except for ACG and AAG. Feng et al. previously reported that AAL binds to a Galβ1-3GalNAcα structure known as the Thomsen-Friedenreich antigen via a unique hydrogen bond network consisting of an Arg85-water-Glu66-water motif, where Arg85 corresponds to the evolutionarily conserved Arg73 (residue number is of human galectin-1; 47). On the other hand, Glu66 is extensively conserved in mushroom galectins, while it is not conserved in vertebrate galectins (Fig. 11). While Feng et al. noted that Pro42, Asn43, His59, Arg63, Asn72, Trp80, and Glu83 are involved in the recognition of non-reducing terminal galactose, they did not mention the special function of Pro45 in its unusual cis conformation.
F. Possible ScenarioBased on the above information, a possible evolutionary
pathway for ACG and related galectins can be speculated upon. As described, the current ACG possesses both β-galactoside and αGalNAc-binding activities, which are mediated by cis and trans-conformed Pro45, respectively. Thus, it fulfills the current defini-tion of galectins (1), and its deviation can be understood as varia-tion among galectin descendants. However, it should be noted that this Janus-type dual specificity is dependent on the integrity of both Pro45 (cis conformer) and Asn46 (to compensate for the lack of conserved Asn66), located in the redundant loop between F2-S3 strands unique to Agrocybe galectins. Ancestral mushroom galectins are assumed to be functional and equipped with Asn66. In future, ACG may lose its conventional β-galactoside-binding activity by a substitution event, possibly occurring on either Pro45 or Asn46, i.e., a non-synonymous substitution, such as the current CCA (Pro)-AAC (Asn) mutating to CGA (Arg)-AAC (Asn) or CCA (Pro)-ATC (Ile), while maintaining GalNAcα1-3Gal(NAc)-binding activity (substituted nucleotide and resultant amino acid are underlined). In that case, the resultant molecules (P45X or N46X) no longer fulfill the classical definition of a galectin (Fig. 12).
There are increasing numbers of galectin-related proteins that have apparently lost sugar-binding function. They include Charcot-Leyden crystal protein (48) and GRIFIN (galectin-related interfiber protein; 49). The former was found to be a member of the galectin family and named galectin 10, but it showed almost no β-galacto-side-binding activity with multiple substitutions at evolutionarily conserved hydrophilic residues, i.e., H44R, N46R, R48H, and R73T (residue numbers are of those of human galectin-1). Interest-ingly, this protein is strongly associated with placental immunity, and is named PP13. Recent studies support the idea that PP13 is a placental immunoregulatory protein that protects pregnancy (50).
GRIFIN is specifically expressed in the lens, and is located at the interface between lens fiber cells. However, it lacks affinity for β-galactosides, which is consistent with the absence of evolution-arily conserved residues, i.e., N46K and R73V. Unlike Agrocybe galectins, neither PP13 nor GRIFIN possess the redundant loop region located between F2 and S3 strands, and thus lack the com-pensatory Pro-Asn sequence. Intriguingly, the zebrafish homologue to mammalian GRIFIN is specifically localized to the lens, but maintains β-galactoside-binding activity and has all seven critical amino acids; i.e., His44, Asn46, Arg48, Asn61, Trp68, Glu71, and Arg73 (51). More recently, the chicken counterpart, C-GRIFIN, has been identified by the Gabius group, and has both lens-specific localization and β-galactoside-binding ability, albeit with a small sequence change at R73V (52). However, most other galectin-
Fig. 10. Summary of the binding features of ACG and N-acetyl-lactosamine. With the lack of evolutionarily conserved Asn64 (Ala64 in ACG), it becomes insufficient to recognize the 4-OH group of the non-reducing terminal galactose. Instead, the Asn46 side chain can contact the 4-OH group of Gal, which is induced by the preceding Pro45 in its cis conformation.

© 2018 FCCA (Forum: Carbohydrates Coming of Age)SE85
Fig. 11. Amino-acid sequence alignment of ACG and related galectins in mushroom. Residue numbers are those of ACG. (∗) denotes evo-lutionarily conserved sugar-binding residues. (P) denotes Pro45 of ACG, which takes cis-conformation when binding to β-galactoside (e.g., lactose). (+) indates Asn46 unique to Agrocybe galectins compensatory for the lack of evolutionarily conserved Asn (resudue number 64 in ACG). Glu69 and Arg88 involved in the unique water-mediated hydrogen bond to Galβ1-3GalNAc recognition are shown in red in AAL. Redundant and truncated loop regions are boxed in blue. For reference, human galectin-1 (hGal-1) is aligned in the bottom. Abbreviations; ACG: Agrocybe cylindracea galectin; AAG: Agrocybe aegerita galectin; LAG: Laccaria amethystina galectin; LBG1-3: Laccaria bicolor galectin isolectins1-3; CCG1-3: Coprinopsis ci-nerea galectin isolectins1-3. CCG3r and LBG1r-3r are putative galectins predicted from the genome data base, for which carbohydrate-binding ability has not been demonstrated.
Fig. 12. A possible evolutionary scenario to generate ACG, and its speculative further deviation to a non-galectin protein. The current ACG possesses both β-galactoside-binding and αGalNAc-binding activities, mediated by cis and trans-conformed Pro45, respectively, which located in the redundant loop region between F2–S3 strands that are unique to Agrocybe galectins. Ancestral mushroom galectins containing Agrocybe ga-lectins are assumed to be functional and equipped with Asn64. In the future, it is expected that ACG may lose conventional β-galactoside-binding activity by a substitution event possibly occurring on either Pro45 or Asn46, while maintaining GalNAcα-binding activity. In that case, the resultant molecules (P45X or N46X) no longer fulfill the definition of a galectin (1).

© 2018 FCCA (Forum: Carbohydrates Coming of Age) SE86
related proteins appear to be non-functional as lectins, but retain their structures and biological distribution (53, 54). Apparently, the observed features imply that evolutionary pathways and carbohy-drate-binding properties are not necessarily coincident in divergent biological species (for more comprehensive discussion, see the chapter by Gabius et al. in this special issue).
G. ConclusionsThe definition of a galectin, which was described in 1994 (1),
is perhaps due for reconsideration before facing its quarter-century anniversary. According to the classical definition, two criteria for galectins are affinity for β-galactosides and sequence similarity. With more extensive genomic coverage, many exceptions to these criteria have been found, and many more have been suggested, and so the first criterion should be extended or modified. Galectin β-galactoside specificity has been confirmed in many organisms, including vertebrates, invertebrates, plants, and fungi (55, 56). Recent insights have led to the suggestion that galectins originally functioned as recognition and effector factors in innate immunity,
through binding to surface glycans of pathogenic microbes, while parasites attempted subvert the recognition roles of host galectins (57). It will be interesting to investigate whether proteins with a galectin scaffold consisting of β-sandwich/jerry roll structures re-ally have greater binding potential for β-galactosides than others. For this, we must first generate non-β-galactoside-binding affinity using the galectin scaffold. In the current article, we have empha-sized the ability of the fungal galectin ACG to switch its specificity from β-galactoside to α-GalNAc by using cis-trans conversion of Pro45, which is located in a redundant loop region. This may be a common strategy in molecular evolution, not only for carbohy-drate-binding proteins but also other functional proteins.
AcknowledgmentsWe acknowledge Drs. Yuka Kobayashi (J-oil Mils, Co., Ltd.)
and Jun Iwaki (Tokyo Chemical Industry, Co., Ltd.) for helpful discussion about fungi classification and galectin specificity, re-spectively. We also thank Editage (www.editage.jp) for English language editing.
References 1. Barondes, S. H., Castronovo, V., Cooper, D. N. W., Cummings, R. D., Drickamer, K., Felzi, T., Gltt, M. A., Hirabayashi, J., Hughes, C., Kasai, K.,
Leffler, H., Liu, F.-T., Lotan, R., Mercuri, A. M., Monsigny, M., Pillail, S., Poirer, F., Raz, A., Rigby, P. W. J., Rini, J. M., and Wang, J. L. (1994) Cell 76, 597–598.
2. Lobsanov, Y. D., Gitt, M. A., Leffler, H., Barondes, S. H., and Rini, J. M. (1993) J. Biol. Chem. 268, 27034–27038. 3. Barondes, S. H., Cooper, D. N., Gitt, M. A., and Leffler, H. (1994) J. Biol. Chem. 269, 20807–20810. 4. Inohara, H., and Raz, A. (1994) Glycoconj. J. 11, 527–532. 5. Glinsky, V. V., and Raz, A. (2009) Carbohydr. Res. 344, 1788–1791. 6. Sathisha, U. V., Jayaram, S., Harish Nayaka, M. A., and Dharmesh, S. M. (2007) Glycoconj. J. 24, 497–507. 7. Li, Y., Liu, L., Niu, Y., Feng, J., Sun, Y., Kong, X., Chen, Y., Chen, X., Gan, H., Cao, S., and Mei, Q. (2012) Eur. J. Nutr. 51, 107–117. 8. Venkateshaiah, S. U., Eswaraiah, M. S., Annaiah, H. N., and Dharmesh, S. M. (2017) Clin. Exp. Metastasis 34, 141–154. 9. Miller, M. C., Ippel, H., Suylen, D., Klyosov, A. A., Traber, P. G., Hackeng, T., and Mayo, K. H. (2016) Glycobiology 26, 88–99.10. Zhang, T., Zheng, Y., Zhao, D., Yan, J., Sun, C., Zhou, Y., and Tai, G. (2016) Int. J. Biol. Macromol. 91, 994–1001.11. Stegmayr, J., Lepur, A., Kahl-Knutson, B., Aguilar-Moncayo, M., Klyosov, A. A., Field, R. A., Oredsson, S., Nilsson, U. J., and Leffler, H. (2016)
J. Biol. Chem. 291, 13318–13334.12. Hirabayashi, J., Hashidate, T., Arata, Y., Nishi, N., Nakamura, T., Hirashima, M., Urashima, T., Oka, T., Futai, M., Muller, W. E., Yagi, F., and
Kasai, K. (2002) Biochim. Biophys. Acta 1572, 232–254.13. Iwaki, J., Tateno, H., Nishi, N., Minamisawa, T., Nakamura-Tsuruta, S., Itakura, Y., Kominami, J., Urashima, T., Nakamura, T., and Hirabayashi,
J. (2011) Biochim. Biophys. Acta 1810, 643–651.14. Yagi, F., Miyamoto, M., Abe, T., Minami, Y., Tadera, K., and Goldstein, I. J. (1997) Glycoconj. J. 14, 281–288.15. Yagi, F., Hiroyama, H., and Kodama, S. (2001) Glycoconj. J. 18, 745–749.16. Hirabayashi, J., and Kasai, K. (1993) Glycobiology 3, 297–304.17. Ban, M., Yoon, H. J., Demirkan, E., Utsumi, S., Mikami, B., and Yagi, F. (2005) J. Mol. Biol. 351, 695–706.18. Butschi, A., Titz, A., Wälti, M. A., Olieric, V., Paschinger, K., Nöbauer, K., Guo, X., Seeberger, P. H., Wilson, I. B., Aebi, M., Hengartner, M. O.,
and Künzler, M. (2010) PLoS Pathog. 6, e1000717.19. Zhao, C., Sun, H., Tong, X., and Qi, Y. (2006) Biochem. J. 374, 321–327.20. Yang, N., Li, D. F., Feng, L., Xiang, Y., Liu, W., Sun, H., and Wang, D. C. (2009) J. Mol. Biol. 387, 694–705.21. Lyimo, B., Yagi, F., and Minami, Y. (2011) Biosci. Biotechnol. Biochem. 75, 62–69.22. Martin, F., Aerts, A., Ahrén, D., Brun, A., Danchin, E. G., Duchaussoy, F., Gibon, J., Kohler, A., Lindquist, E., Pereda, V., Salamov, A., Shapiro,
H. J., Wuyts, J., Blaudez, D., Buée, M., Brokstein, P., Canbäck, B., Cohen, D., Courty, P. E., Coutinho, P. M., Delaruelle, C., Detter, J. C., Deveau, A., DiFazio, S., Duplessis, S., Fraissinet-Tachet, L., Lucic, E., Frey-Klett, P., Fourrey, C., Feussner, I., Gay, G., Grimwood, J., Hoegger, P. J., Jain, P., Kilaru, S., Labbé, J., Lin, Y. C., Legué, V., Le Tacon, F., Marmeisse, R., Melayah, D., Montanini, B., Muratet, M., Nehls, U., Niculita-Hirzel, H., Oudot-Le Secq, M. P., Peter, M., Quesneville, H., Rajashekar, B., Reich, M., Rouhier, N., Schmutz, J., Yin, T., Chalot, M., Henrissat, B., Kües, U., Lucas, S., Van de Peer, Y., Podila, G. K., Polle, A., Pukkila, P. J., Richardson, P. M., Rouzé, P., Sanders, I. R., Stajich, J. E., Tunlid, A., Tuskan, G., and Grigoriev, I. V. (2008) Nature 452, 88–92.

© 2018 FCCA (Forum: Carbohydrates Coming of Age)SE87
23. Liao, D. I., Kapadia, G., Ahmed, H., Vasta, G. R., and Herzberg, O. (1994) Proc. Natl. Acad. Sci. U.S.A. 91, 1428–1432.24. Lobsanov, Y. D., and Rini, J. M. (1997) Trends Glycosci. Glycotechnol. 9, 145–154.25. Imamura, K., Takeuchi, H., Yabe, R., Tateno, H., and Hirabayashi, J. (2011) J. Biochem. 150, 545–552.26. Hu, D., Tateno, H., Sato, T., Narimatsu, H., and Hirabayashi, J. (2013) Biochem. J. 453, 261–270.27. Hu, D., Tateno, H., and Hirabayashi, J. (2015) Molecules 20, 7637–7656.28. de Mattos-Shipley, K. M. J., Ford, K. L., Alberti, F., Banks, A. M., Bailey, A. M., and Foster, G. D. (2016) Stud. Mycol. 85, 125–157.29. Singh, S. S., Wang, H., Chan, Y. S., Pan, W., Dan, X., Yin, C. M., Akkouh, O., and Ng, T. B. (2014) Molecules 20, 446–469.30. Varrot, A., Basheer, S. M., and Imberty, A. (2013) Curr. Opin. Struct. Biol. 23, 678–685.31. Liang, Y., Lin, J. C., Wang, K., Chen, Y. J., Liu, H. H., Luan, R., Jiang, S., Che, T., Zhao, Y., Li, F., Wang, C., Guo, L., and Sun, H. (2010) Biochim.
Biophys. Acta 1800, 474–480.32. Kobayashi, Y., and Kawagishi, H. (2014) Methods Mol. Biol. 1200, 15–38.33. Schubert, M., Bleuler-Martinez, S., Butschi, A., Wälti, M. A., Egloff, P., Stutz, K., Yan, S., Collot, M., Mallet, J. M., Wilson, I. B., Hengartner, M.
O., Aebi, M., Allain, F. H., and Künzler, M. (2011) PLoS Pathog. 8, e1002706.34. Renko, M., Sabotic, J., Mihelic, M., Brzin, J., Kos, J., and Turk, D. (2010) J. Biol. Chem. 285, 308–316.35. Lacadena, J., Alvarez-García, E., Carreras-Sangrà, N., Herrero-Galán, E., Alegre-Cebollada, J., García-Ortega, L., Oñaderra, M., Gavilanes, J. G.,
and Martínez del Pozo, A. (2007) FEMS Microbiol. Rev. 31, 212–237.36. Berne, S., Lah, L., and Sepcić, K. (2009) Protein Sci. 18, 694–706.37. Bleuler-Martínez, S., Butschi, A., Garbani, M., Wälti, M. A., Wohlschlager, T., Potthoff, E., Sabotiĉ, J., Pohleven, J., Lüthy, P., Hengartner, M. O.,
Aebi, M., and Künzler, M. (2011) Mol. Ecol. 20, 3056–3070.38. Lobsanov, Y. D., and Rini, J. M. (1997) Trends Glycosci. Glycotechnol. 9, 145–154.39. van Hattum, H., Branderhorst, H. M., Moret, E. E., Nilsson, U. J., Leffler, H., and Pieters, R. J. (2013) J. Med. Chem. 56, 1350–1354.40. Rajput, V. K., MacKinnon, A., Mandal, S., Collins, P., Blanchard, H., Leffler, H., Sethi, T., Schambye, H., Mukhopadhyay, B., and Nilsson, U. J.
(2016) J. Med. Chem. 59, 8141–8147.41. Atmanene, C., Ronin, C., Téletchéa, S., Gautier, F. M., Djedaïni-Pilard, F., Ciesielski, F., Vivat, V., and Grandjean, C. (2017) Biochem. Biophys.
Res. Commun. 489, 281–286.42. Hirabayashi, J., and Kasai, K. (1991) J. Biol. Chem. 266, 23648–23653.43. Hirabayashi, J., and Kasai, K. (1994) Glycoconj. J. 11, 437–442.44. Nemoto-Sasaki, Y., Hayama, K., Ohya, H., Arata, Y., Kaneko, M. K., Saitou, N., Hirabayashi, J., and Kasai, K. (2008) Biochim. Biophys. Acta
1780, 1131–1142.45. Hu, D., Huang, H., Tateno, H., Nakakita, S., Sato, T., Narimatsu, H., Yao, X., and Hirabayashi, J. (2015) J. Biochem. 157, 197–200.46. Kuwabara, N., Hu, D., Tateno, H., Makyio, H., Hirabayashi, J., and Kato, R. (2013) FEBS Lett. 587, 3620–3625.47. Feng, L., Sun, H., Zhang, Y., Li, D. F., and Wang, D. C. (2010) FASEB J. 24, 3861–3868.48. Ackerman, S. J., Corrette, S. E., Rosenberg, H. F., Bennett, J. C., Mastrianni, D. M., Nicholson-Weller, A., Weller, P. F., Chin, D. T., and Tenen, D.
G. (1993) J. Immunol. 150, 456–468.49. Ogden, A. T., Nunes, I., Ko, K., Wu, S., Hines, C. S., Wang, A. F., Hegde, R. S., and Lang, R. A. (1998) J. Biol. Chem. 273, 28889–28896.50. Than, N. G., Balogh, A., Romero, R., Kárpáti, E., Erez, O., Szilágyi, A., Kovalszky, I., Sammar, M., Gizurarson, S., Matkó, J., Závodszky, P.,
Papp, Z., and Meiri, H. (2014) Front. Immunol. 5, 348.51. Ahmed, H., and Vasta, G. R. (2008) Biochem. Biophys. Res. Commun. 371, 350–355.52. García Caballero, G., Kaltner, H., Michalak, M., Shilova, N., Yegres, M., André, S., Ludwig, A. K., Manning, J. C., Schmidt, S., Schnölzer, M.,
Bovin, N. V., Reusch, D., Kopitz, J., and Gabius, H. J. (2016) Biochimie 128–129, 34–47.53. García Caballero, G., Flores-Ibarra, A., Michalak, M., Khasbiullina, N., Bovin, N. V., André, S., Manning, J. C., Vértesy, S., Ruiz, F. M., Kaltner,
H., Kopitz, J., Romero, A., and Gabius, H. J. (2016) Biochim. Biophys. Acta 1860, 2285–2297.54. Kaltner, H., García Caballero, G., Sinowatz, F., Schmidt, S., Manning, J. C., André, S., and Gabius, H. J. (2016) Biochim. Biophys. Acta 1860,
2298–2312.55. Cooper, D. N. (2002) Biochim. Biophys. Acta 1572, 209–231.56. Vasta, G. R., Ahmed, H., and Odom, E. W. (2004) Curr. Opin. Struct. Biol. 14, 617–630.57. Vasta, G. R., Feng, C., González-Montalbán, N., Mancini, J., Yang, L., Abernathy, K., Frost, G., and Palm, C. (2017) Pathog. Dis. 75. doi: 10.1093/
femspd/ftx046

© 2018 FCCA (Forum: Carbohydrates Coming of Age) SE88
Jun Hirabayashi Born in Feb 28, 1958 in Tochigi prefecture in Japan. Education: Bachelor of Science, Tohoku University (Mar., 1980), Master of Science (chemistry), Tohoku University Graduate School (Mar., 1982). Professional Carrier: Assistant professor, Faculty of Pharmaceutical Sciences, Teikyo Uni-versity, Sagamiko, Kanagawa, Japan (Apr., 1982), Lecturer (Apr., 1989), PhD in science (chemistry), Tohoku University (Dec., 1989); Team leader, National Institute of Advanced Industrial Science and Technology (AIST), Tsukuba, Ibaraki, Japan (Nov., 2002); Visiting professor, Kagawa University, Kaga-wa, Japan (Sep., 2003); Deputy director of Research Center for Medical Glycoscience, AIST (Dec., 2006), Prime senior researcher, AIST (Apr., 2012). Main research fields: Biochemistry, Glycoscience, Glycotechnology, Lectin Engineering, evolutionary chemistry.Councilor, The Japanese Society of Carbohydrate Research (JSCR)Director, Japan Consortium for Glycoscience and Glycotechnology (JCGG)
Dr. Dan Hu was born in Jan 13, 1979 at Hunan Province in China. He received his Master degree from the Shenyang Pharmaceutical University in 2006 under the direction of Prof. Tatsuya Yamagata. Then he moved to Japan and obtained his Ph. D from the University of Tokyo in 2010 under the guidance of Prof. Kazuo Yamamoto. Then, he worked as a JSPS postdoctoral fellow at National Institute of Ad-vanced Industrial Science and Technology (AIST) with Prime Senior Researcher, Jun Hirabayashi. In 2013, he moved back to China and was appointed as an Associate Professor at Jinan University. He has been working on the functional characterization of gangliosides in the cell membranes and endoplasmic reticulum-resident lectins. He was also interested in lectin engineering using an approach of directed evolution. Now he is focused on the biosynthesis of fungi-derived natural products and expanding chemical diversity by combinational biosynthesis.
Naoyuki Kuwabara Structural Biology Research Center, Institute of Material Structure Science, High Energy Accelerator Research Organization, KEK. Research interest: Structure-function relationships on carbohydrate-recognition proteins and glycosyltransferases.
Information of the Authors