caracterização dos danos celulares em gametas femininos e ... 1046.pdf · caracterização dos...
TRANSCRIPT

Acta Scientiae Veterinariae, 2012. 40(3): 1046.
REVIEW ARTICLE Pub. 1046
ISSN 1679-9216 (Online)
1
Received: December 2011 www.ufrgs.br/actavet Accepted: March 2012
Laboratório de Manipulação de Oócitos e Folículos Pré-Antrais (LAMOFOPA). Programa de Pós-graduação em Ciências Veterinárias (PPGCV). Univer-sidade Estadual do Ceará (UECE), Fortaleza, CE, Brazil. CORRESPONDENCE: A.A. Carvalho [[email protected] - Fax: +55 (85) 3101-9840]. LAMOFOPA, Faculdade de Veterinária - UECE. Av. Paranjana n. 1700, Campus do Itaperi. CEP 60.740-903 Fortaleza, CE, Brazil.
Caracterização dos danos celulares em gametas femininos e embriões após
criopreservação
Characterization of Cellular Damage in Female Gametes and Embryos after Cryopreservation
Adeline de Andrade Carvalho, Luciana Rocha Faustino, Simone Vieira Castro, José Ricardo de Figueiredo &
Ana Paula Ribeiro Rodrigues
ABSTRACT
Background: Cryopreservation is a biotech successfully employed in female gametes and embryos. This technique is of great importance for propagation of genetic material from animals with high-value livestock as well as to preserve the fertility of women undergoing cancer treatments. However, low temperatures can result in damage to different cellular compartments and organelles. This damage culminates in reduced viability, since they affect cell metabolism.Review: Cryopreservation consists of maintenance of biological material at low temperatures, in which chemical reactions are ceased, however, allowing the cells to preserve their viability. However, the decrease of temperature and subsequent warming may result in cellular damage. These damages occur in the cell membrane, cytoplasmic organelles and the cell nucleus. It is believed that the fi rst cell structure undergoing cryoinjury is the plasma membrane, responsible for main-taining homeostasis within the cell, and the loss of plasma membrane during the reduction temperature reported the main damage. The membrane damage due to cryopreservation appears to correlate with the reduction of thermal energy at low temperatures, thus limiting the movement of molecules through the phospholipids of lipid bilayer. Cryopreservation also alters the morphology, structure and cellular distribution of lipid droplets, reducing the survival of oocytes and embryos. The physical state changes, besides changing physicochemical properties of intracellular lipid may also result in damage to lipids associated with the cellular cytoskeleton. The interaction between cell lipid phase and components of cytoskeleton is complex and hardening of these lipids can cause deformation and disruption of cytoskeleton with consequent negative effect on cell survival and development. The disruption of cytoskeleton can also be intrinsic to change in dehydration and cellular form that follow the cryopreservation process. Among the cellular organelles, mitochondria are sensitive to cryopreservation procedures, since the formation of intracellular ice crystals considered one of the most relevant cryoin-jury, leading to damage to the mitochondrial cristae and matrix. Another important organelle undergoes damage as result of cryopreservation is the endoplasmic reticulum, which change in morphology has been described after vitrifi cation of embryos. It is also known that cryopreservation may result in fragmentation of DNA even in the absence of deformation of the cellular morphology, and that these changes can lead to delay in the development or cell death. In addition to the direct damage to double-stranded DNA, cryopreservation may trigger chromosomal abnormalities, these the aneuploidy is the most frequent and seems to compromise the developmental competence of oocytes in addition to being indicated as the main factor for the low achieve of live births.Conclusion: Cryopreservation is an important alternative for the preservation of gametes and embryos, however, the cells are subjected to unfavorable conditions that can compromise cell recovery after thawing/warming. The main cause of reduction in survival oocytes and embryos after cryopreservation appear to be related to damage in different cellular compartments and structures, which are essential for maintaining cell metabolism. Thus, it is observed the need to achieve cryopreservation protocols capable of maintaining the integrity of different cellular components.
Keywords: cryopreservation, cryoinjury cellular, oocytes, female gametes, embryos, ovarian tissue.Descritores: criopreservação, crioinjúria celular, oócitos, gametas femininos, embriões, tecido ovariano.

2
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
I. INTRODUÇÃO
II. BASES GERAIS DA CRIOPRESERVAÇÃO CELULAR
III. CARACTERÍSTICAS GERAIS DAS CÉLULAS EUCARI-
ÓTICAS
IV. PRINCIPAIS DANOS CELULARES CAUSADOS PELA
CRIOPRESERVAÇÃO
1. Danos de membrana2. Danos em gotículas lipídicas intracitoplasmáticas e citoesqueleto3. Danos em organelas celulares4. Danos nucleares
V. CONCLUSÃO
VI. REFERÊNCIAS
I. INTRODUÇÃO
A criopreservação é uma técnica que tem des-pertado grande interesse para a reprodução assistida nos últimos 15 anos, principalmente desde os primeiros relatos de nascimentos em ovinos [39] e humanos [25] após o transplante de tecido ovariano previamente criopreservado. Portanto, é compreensível a aplicação cada vez mais frequente dessa técnica com a fi nalidade de preservação de gametas femininos com o objetivo fi nal de manutenção da função reprodutiva da fêmea ou mesmo a produção de crias após impossibilidade de reprodução natural ou radicalmente, após a morte do animal.
Considerando a crescente incidência de do-enças cancerígenas, aliada ao sucesso atual dos trata-mentos oncológicos, a criopreservação tem ganhado ênfase na medicina reprodutiva humana por repre-sentar uma alternativa para que mulheres, sobretudo as jovens, possam gerar fi lhos após o tratamento do câncer [78]. Além disso, a criopreservação tem sido bastante difundida no âmbito da veterinária por per-mitir a disseminação de material genético de animais de alto valor genético e grande produtividade [117]. Adicionalmente, a criopreservação também tem sido considerada uma vantajosa ferramenta para preservar animais em vias de extinção [114].
Vários estudos têm sido realizados visando aperfeiçoar o processo de criopreservação de gametas femininos, seja na forma matura [72,78,102] ou ima-tura [11,49,74], como meio de permitir um melhor aproveitamento do potencial reprodutivo mesmo após
a morte ou impossibilidade da função reprodutiva da fêmea. Além da criopreservação de gametas femininos, a criopreservação de embriões também tem sido bas-tante estudada e amplamente difundida e estabelecida para aplicação tanto em humanos [86,87] quanto em animais [27,83]. Contudo, ainda observa-se a inexis-tência de um protocolo ideal [67], haja vista as baixas taxas de sobrevivência e desenvolvimento oocitário até embrião normal [60]. Além disso, também são baixas as taxas de sobrevivência e implantação embrionária após criopreservação, as quais ainda são consideradas muito inferiores àquelas obtidas quando se trata de embriões frescos [57].
A criopreservação de células germinativas pode ser realizada tanto por congelação lenta como por vitrifi cação. Ambos os métodos têm sido empre-gados em tecido ovariano [66,71], oócitos [12,29,30] e embriões [60,85]. Contudo, esse procedimento ain-da é apontado por alguns autores como uma técnica prejudicial às células, que pode resultar em danos em diferentes compartimentos celulares, como por exem-plo, na membrana celular [55], nas organelas [63,94] e no núcleo [45,67]. O conhecimento e a caracterização desses danos são de grande relevância para a crio-biologia, visto que a intensidade com que as lesões celulares ocorrem pode incapacitar as células de su-perarem grandes danos, consequentemente, perdendo sua viabilidade. Desta forma, a presente revisão tem o objetivo de descrever os principais danos originados em consequência dos procedimentos de criopreservação em tecido ovariano, oócito maturo e embriões oriundos de seres humanos e animais.
II. BASES GERAIS DA CRIOPRESERVAÇÃO CELULAR
A criopreservação consiste na manutenção de material biológico a temperaturas criogênicas, nas quais todas as reações químicas, processos biológicos e atividades físicas intra e extracelulares estão suspen-sas, contudo, as células conseguem manter-se íntegras, preservando sua viabilidade [91]. Essas temperaturas são denominadas criogênicas, sendo frequentemente utilizado o nitrogênio líquido (-196ºC) para alcançá-las [7]. Em temperaturas entre 0° e 25ºC a atividade celular torna-se lenta, mas ainda está presente. Con-tudo, abaixo de -40ºC as trocas físico-químicas são cessadas e, para garantir a preservação celular por tempo indeterminado, as células devem ser mantidas

3
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
a temperaturas inferiores a -130ºC na presença de agentes crioprotetores [7].
Em regiões frias, algumas plantas [9] e ani-mais [22,84] desenvolveram uma resistência natural às baixas temperaturas. Essa resistência consiste na capacidade de possibilitar que a água mantenha-se su-peresfriada em temperaturas próximas a -40ºC [9]. Esta adaptação é denominada de aclimatação e geralmente envolve a síntese de açúcares intra e extracelulares que aumentam a pressão osmótica das células e reduzem a injúria por plasmólise a baixas temperaturas [9,22]. Contudo, em 1949 foi realizada a descoberta de uma substância com a capacidade de atravessar a membrana celular e reduzir a formação de cristais de gelo [58,82]. Esta substância, isto é, o glicerol, bem como outras com propriedades similares, como o propanodiol, etileno-glicol e dimetilsulfóxido, passaram desde então a ser utilizadas com sucesso na criopreservação de células e tecidos [17,111].
Apesar dos avanços na criobiologia, tanto a redução de temperatura quanto o posterior aquecimento celular podem resultar em danos na estrutura da célula [9,28]. Esses danos podem ocorrer na membrana celu-lar [17], nas organelas citoplasmáticas [47,49,50] e até mesmo no núcleo da célula [23,67]. Em virtude disso, várias pesquisas têm sido realizadas com o intuito de conhecer as crioinjúrias ocasionadas e amenizar ou reverter os danos celulares decorrente dessas injúrias [29,30,107]. Devido à complexidade da estrutura ce-lular eucariótica e as várias organelas essenciais para a sobrevivência da mesma, cada organela será descrita em maiores detalhes a seguir.
III. CARACTERÍSTICAS GERAIS DAS CÉLULAS
EUCARIÓTICAS
As células de organismos multicelulares apresentam forma e estrutura variáveis e se diferen-ciam de acordo com sua função. No entanto, todas as células eucarióticas apresentam uma organização básica similar (Figura 1). Esta organização consiste em uma membrana celular externa, constituída por uma bicamada lipídica com proteínas dispostas nesta bicamada [37]. Esta membrana é responsável por de-limitar o citoplasma, composto por citosol, organelas celulares, proteínas e íons; e o núcleo, que alberga o material genético da célula, o qual é um compartimento altamente organizado e separado do citoplasma por uma membrana porosa denominada carioteca [41].
Figura 1. Imagem ilustrativa de uma célula eucariótica evidenciando as organelas internas no citoplasma e no núcleo. Imagem concedida por Thibodeau e Patton [103].
O citoplasma celular, além de albergar fl uidos e enzimas, também contém estruturas físicas altamente organizadas, denominadas organelas intracelulares [33]. Toda esta estrutura é mantida com auxílio de uma complexa rede fi lamentosa chamada de citoesqueleto. O citoesqueleto é uma estrutura constituída por micro-túbulos formados por tubulina α e β, microfi lamentos de actina e fi lamentos intermediários específi cos de certas células. Todos estes elementos assumem, além da função esquelética, outras funções de acordo com as proteínas as quais estão associados [49].
Nas células eucarióticas, o citoesqueleto tem como função principal coordenar a distribuição de organelas nas células e manter a forma celular. Sendo, portanto, o citoesqueleto responsável pela sustentação e resistência celular, bem como por alterações na forma e distribuição das organelas desencadeadas por interações entre a célula e seu meio e, entre células diferentes [41]. Os microtúbulos executam diversas funções, dentre elas, a manutenção da estrutura celular, o transporte de moléculas, bem como estão envolvidos na divisão celular, uma vez que participam da formação do fuso meiótico [30].
No citoplasma também estão presentes as or-ganelas responsáveis pela respiração e energia celular, conhecidas como mitocôndrias [20]. A distribuição destas organelas parece estar associada com o nível de metabolismo celular, através da conversão de piruvato à adenosina trifosfato (ATP), substância que atua como fonte de energia para a célula [41].
As células eucarióticas também possuem or-ganelas formadas por uma interconexão de túbulos e

4
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
vesículas. Estas estruturas, conhecidas como retículo endoplasmático, são constituídas por membrana, matriz endoplasmática e um meio aquoso que ocupa o espaço entre os túbulos e vesículas. O retículo en-doplasmático pode ou não ter sua superfície recoberta por ribossomos, sendo, então, denominado de retículo endoplasmático rugoso e liso, respectivamente [70]. O retículo endoplasmático rugoso ou granular possui a capacidade de sintetizar novas moléculas proteicas, enquanto o retículo endoplasmático liso, também chamado de agranular, tem como função a síntese de substâncias lipídicas, incluindo a produção de hormô-nios esteróides [70]. As substâncias produzidas pelo retículo endoplasmático são liberadas no meio extra-celular através de encapsulamento pelas vesículas do complexo de Golgi. Essa organela possui membranas semelhantes às membranas do retículo endoplasmático agranular e geralmente é composta por várias camadas de vesículas achatadas e empilhadas, sendo proemi-nente em células secretoras [96].
O núcleo celular é um compartimento orga-nizado no qual são encontrados os cromossomos, responsáveis por armazenar as informações genéti-cas [54]. Durante a divisão celular os cromossomos encontram-se aderidos ao fuso meiótico, formado por microtúbulos e microfi lamentos de actina [78]. O núcleo é considerado o centro de controle da célula por conter os genes que determinam as características proteicas da célula [54], incluindo proteínas estruturais e enzimas que controlam as atividades nucleares e citoplasmáticas [41].
Cada compartimento celular citado desempe-nha um importante papel para manutenção da home-ostase celular. Entretanto, essa homeostase pode ser comprometida em virtude de alterações em diferentes estruturas celulares decorrentes da criopreservação, conforme detalhado no próximo tópico.
IV. PRINCIPAIS DANOS CELULARES CAUSADOS PELA
CRIOPRESERVAÇÃO
1. Danos de membrana
De forma geral, todas as membranas celulares são sensíveis às crioinjúrias, porém, a sobrevivência da célula parece estar mais diretamente relacionada com a integridade da membrana plasmática do que com as membranas que delimitam as organelas. A membrana celular possui maior relevância por atuar no procedimento de criopreservação como uma barreira
semipermeável entre o citoplasma e o meio extracelu-lar, permitindo o movimento (efl uxo/infl uxo) de água e agentes crioprotetores, fundamental para a preservação da célula a temperaturas ultra-baixas [105]. Acredita-se que a membrana seja o primeiro compartimento a sofrer injúrias decorrentes do frio e que estas injúrias estejam correlacionadas com a redução de temperatura a qual a célula é exposta [99]. Um dos graves danos decorrentes da criopreservação é a perda de parte da membrana plasmática durante a redução de temperatu-ra [1]. Esta perda membranária é resultado da intensa desidratação celular que precisa ocorrer com o intuito de reduzir a formação intracelular de gelo de modo a garantir o sucesso da criopreservação, desde que esta não seja excessiva. Durante a desidratação excessiva e consequente estresse hídrico, as células se retraem e ocorre uma contração da membrana celular. Esta con-tração resulta na aproximação de porções de membrana permitindo uma interação da bicamada lipídica com ocasional fusão da membrana [94], ocorrendo conse-quentemente formação de vesículas e perda de parte do material da membrana. Essa alteração pode culminar com a perda de elasticidade da membrana e lise da célula ao retornar às condições isotônicas normais [1].
Os danos de membrana decorrentes da crio-preservação também parecem estar relacionados com a redução da energia térmica a baixas temperaturas e consequente limitação do movimento de molécu-las na bicamada lipídica da membrana [37]. Esses danos também são infl uenciados pela composição lipídica da membrana, que infl uencia diretamente suas propriedades, inclusive a resistência ao estresse térmico [5,119,121]. Esta composição parece variar de acordo com o desenvolvimento e tipo celular. Já foi demonstrado, na espécie ovina, que a membrana plasmática de zigotos e oócitos em diferentes estágios de desenvolvimento possuem diferentes temperaturas de solidifi cação, sugerindo ainda que essa variação está relacionada com a composição fosfolipídica e concentração de colesterol da membrana [120].
Ao avaliar ovários humanos criopreservados utilizando a congelação lenta na presença de dimetil-sulfóxido (DMSO), Martinez-Madri [62] observaram que 96,7% dos folículos ovarianos apresentavam membranas celular e nuclear intactas. A congelação lenta de tecido ovariano de cutias (Dasyprocta aguti) com propanodiol também foi efi caz em preservar as membranas do oócito e das células da granulosa [114].

5
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
Contudo, em caprinos foram relatadas alterações na membrana nuclear (Figura 2A) após congelação em meio contendo DMSO e sacarose [17].
Figura 2. Fotomicrografi as demonstrativas da ultraestrutura de folículos ovarianos e oócitos submetidos a procedimentos de criopreservação. (A) Folículo primordial de mulher incluso em ovário inteiro congelado/descon-gelado evidenciando um oócito (O) com ruptura de membrana nuclear e condensação da cromatina (seta preta) no núcleo do oócito (N). Entre as células foliculares (Fc) percebe-se célula folicular alterada com condensa-ção da cromatina (seta branca). Imagem concedida por Martinez-Madrid [62] e autorizada pela Elsevier. (B) Oócito bovino imaturo congelado/descongelado com gotícula lipídica intacta (1) e com lipólise parcial (2). Observam-se áreas de lipólise (seta branca) e mitocôndrias (m). Aumento de 8000x. Figura adaptada de Isachenko [48] e autorizada por Wiley Job Network. (C) Oócitos equinos maturos vitrifi cados/aquecidos com enturgecimento de mitocôndrias e redução de elétron-densidade da matriz mitocondrial (setas pretas). Figura concedida por Hochi [46] e autorizada pela Elsevier. (D) Folículo suíno incluso em tecido ovariano congelado/descongelado com mitocôndrias túrgidas (m). (l): gotículas lipídicas, (Nu): núcleo, (CG): células da granulosa e (*): espaços vazios. Figura concedida por Borges [13] e autorizada pela Elsevier. (E) Folículo ovariano humano incluso em tecido vitrifi cado/aquecido com presença de poros na membrana nuclear (setas pretas) e retículo endoplasmático (ER) e mitocôndrias (M) bem preservados, semelhante ao tecido fresco. Figura concedida por Sheikhi [95] e autorizada pela Oxford Journals.
Especifi camente em oócitos, a camada glico-protéica denominada zona pelúcida, exerce a impor-tante função de impedir a polispermia e, também, se mostra sensível aos procedimentos de criopreservação [55,98,106]. A presença de vacúolos e debris na zona pelúcida, por exemplo, já foi relatada após criopre-servação de ovários de camundongas, seja através da congelação lenta ou da vitrifi cação [55]. Em geral,
sabe-se que a criopreservação de oócitos ocasiona uma redução na taxa de fecundação do oócito [98] provavelmente por mudanças estruturais da zona pe-lúcida. Essas alterações parecem ser ocasionadas pela liberação precoce dos grânulos corticais [16,36,52,64] devido a um aumento de cálcio após procedimentos de criopreservação [56], resultando no enrijecimento da zona pelúcida [52]. Contudo, o exato mecanismo que conduz a esse aumento de cálcio no oócito ainda não é completamente conhecido.
Em embriões criopreservados foi demonstra-da uma sensibilidade relativamente alta da membrana plasmática, uma vez que o processo de congelação/descongelação de embriões reduziu em cerca de cinco vezes a fl uidez da membrana de embriões de camundongos [2]. Contudo, ao avaliar a transição da fase cristalina líquida para a fase cristalina gel (Tm) de embriões e oócitos maturos (MII) e imaturos (VG) de mulheres, Ghetler et al. [37] observaram que a membrana celular de embriões possui uma Tm mais baixa que a de oócitos, tornando-os menos sensíveis às crioinjúrias. Esta variável está correlacionada com uma maior resistência ao frio, uma vez que a passagem dos lipídios da fase líquida para a fase gel ocorre a uma temperatura mais baixa, na qual os processos biológicos e a cinética de lesão são lentos. Desta forma, acredita-se que os danos na membrana possam ser reduzidos através da perfusão dos agen-tes crioprotetores a uma temperatura acima da Tm da membrana celular, possibilitando o transporte de crioprotetor e água através da membrana quando a bicamada lipídica ainda encontra-se em fase líquida, facilitando o transporte transmembrana [19,51,118].
2. Danos nas gotículas lipídicas intracitoplasmáticas e citoesqueleto
O sucesso dos protocolos de criopreservação de tecido ovariano, oócitos e embriões parece ser di-fi cultado, em grande parte, pela presença de lipídios (Figura 2B), tanto na forma de gotículas lipídicas intracitoplasmáticas [35] quanto na composição da membrana celular [37], principalmente nas espécies suína [13] e canina [50]. Além disso, o procedimento de vitrifi cação altera a morfologia, estrutura e distri-buição celular das gotículas de lipídios [34], reduzindo a sobrevivência de oócitos e embriões [37].
Na vitrifi cação de oócitos e embriões suínos, algumas pesquisas têm sido realizadas com o intuito de reduzir a quantidade de lipídio intracelular [67] e

6
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
aumentar a sobrevivência oocitária [35] e embrionária [67]. Contudo, a centrifugação e a remoção química utilizadas nos tratamentos de delipação podem causar danos aos oócitos e embriões, tornando as células mais frágeis ao processo, especialmente a membrana celular, que se torna mais sensível [35].
As gotículas lipídicas parecem estar relacionadas aos fi lamentos intermediários do citoesqueleto [15,43]. Estes fi lamentos formam uma rede que se estende do núcleo à membrana plasmática envolvendo grande parte das organelas celulares [15] e já foram descritos em as-sociação com o retículo endoplasmático [80], complexo de Golgi [65] e gotículas lipídicas intracitoplasmáticas [43]. A interação de fi lamentos intermediários com gotas lipídicas foi reforçada pela associação dessas estruturas com a vimentina (grupo de proteínas constituintes dos fi lamentos intermediários), observada por Traub [104], embora a fi nalidade dessa interação ainda seja desco-nhecida. No entanto, acredita-se que a interação lipídio-citoesqueleto possa estar relacionada à manutenção da morfologia celular [10], embora o mecanismo pelo qual este processo ocorra ainda também não esteja elucidado.
As mudanças de estado físico, além de alterarem as propriedades físico-químicas dos lipídios intracelula-res [49] podem resultar em danos também nos lipídios associados ao citoesqueleto celular [48] (Figura 3). A interação entre a fase lipídica das células e os elementos do citoesqueleto é complexa e o enrijecimento desses lipídios pode causar deformação e ruptura do citoesque-leto [49], com consequente infl uência na sobrevivência e desenvolvimento celular [47,88,112].
Figura 3. Imagens representativas do padrão de citoesqueleto de oócitos felinos imaturos submetidos à criopreservação evidenciando microfi la-mentos em vermelho e microtúbulos em verde. Microfi lamentos difusos no ooplasma com uma concentração ligeiramente maior na zona pelúcida (A), microtúbulos uniformemente distribuídos por todo ooplasma (A’). Microfi lamentos e microtúbulos distribuídos de forma irregular no cito-plasma (B e B’). [Barras = 50 µm]. Figura concedida por Luciano [59] e autorizada pela Elsevier.
O rompimento do citoesqueleto pode ser uma alteração intrínseca às mudanças na forma e desidratação celular que acompanham o processo de criopreservação [113]. Em oócitos, as crioinjúrias parecem afetar principalmente os microtúbulos [3] e a organização do citoesqueleto [75]. Esses criodanos podem também ser decorrentes da exposição aos agentes crioprotetores, que podem ocasionar a des-polimerização de microtúbulos e microfi lamentos e causar a destruição dos componentes do citoesqueleto [24]. A exposição ao crioprotetor, etapa fundamental da criopreservação, resultou em discreta redução no percentual de distribuição normal dos microfi lamentos em oócitos bovinos congelados [94] e oócitos suínos vitrifi cados [90]. Além disso, a congelação de oócitos bovinos demonstrou um efeito negativo sobre os micro-fi lamentos, observado pela drástica redução de oócitos com distribuição normal dos microfi lamentos [94].
Contudo, alguns estudos têm mostrado a efi ci-ência de protocolos de criopreservação com menores danos ao citoesqueleto celular. Em camundongas, a avaliação de oócitos imaturos criopreservados (conge-lação lenta), posteriormente submetidos à maturação in vitro, não revelou qualquer alteração signifi cativa na organização de microtúbulos e microfi lamentos [29]. Em oócitos maturos de equinos vitrifi cados por OPS (Open Pulled Straws), Tharasanit et al. [102] observa-ram um rompimento no citoesqueleto celular, porém, a frequência dessa alteração foi baixa.
Alterações irreversíveis na estrutura dos mi-crotúbulos podem ser ocasionadas em decorrência do estresse osmótico, do efeito solução e da formação de cristais de gelo intracelulares, que são efeitos intrínse-cos ao processo de criopreservação [94]. A manutenção da morfologia dos microfi lamentos é tão importante quanto a manutenção da morfologia dos microtúbulos. A preservação do fuso meiótico de oócitos em MII é a principal difi culdade da criopreservação destas estruturas. Nesta fase, os cromossomos encontram-se dispostos na placa equatorial e os microtúbulos ligados aos cromossomos estão estendidos [26], tornando-se mais vulneráveis às crioinjúrias. Vários estudos têm sugerido que a criopreservação causa alterações na polimerização da tubulina de microtúbulos do fuso de oócitos em MII, ocasionando uma ruptura no fuso meiótico e consequente desorganização da cromatina [81,100]. Danos no fuso meiótico já foram relatados após a vitrifi cação de oócitos de camundongas [38],

7
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
ovelhas [100] e mulheres [61]. Em oócitos de coelha ovulados e congelados não foi perceptível a presença de actina polimerizada (componente dos microfi lamen-tos), sendo observada a presença de microfi lamentos apenas no corpúsculo polar [111]. Contudo, algumas equipes sugerem que o citoesqueleto de oócitos em estágio de vesícula germinativa seja mais rígido que microtúbulos e microfi lamentos de oócitos em estágio MII [4]. Uma hipótese é que em função da complexa interação entre a fase lipídica das células e os elementos do citoesqueleto, o enrijecimento dos lipídios pode ocasionar uma deformação e até mesmo ruptura do citoesqueleto. No citoesqueleto rígido de oócitos em vesícula germinativa o enrijecimento em decorrência dos procedimentos de criopreservação torna-se, apa-rentemente, permanente, enquanto oócitos em MII pa-recem reverter estes danos em virtude da característica fl exível de seu citoesqueleto [48].
3. Danos em organelas celulares
Durante o procedimento de criopreservação, as células podem sofrer severos danos em sua estrutura. Esses danos estão relacionados com a integridade das organelas celulares e podem resultar em retardo no crescimento folicular [92] e embrionário [67] ou até mesmo culminar com a morte da célula [62].
As mitocôndrias, organelas essenciais para o metabolismo celular, são sensíveis aos procedimentos de criopreservação [12,73], apresentando criodanos ocasionados tanto pela congelação lenta [62] quanto pela vitrifi cação [66]. Aparentemente, a causa mais relevante desses danos é a formação de cristais de gelo intracelular, que induz a danos nas cristas e na matriz das mitocôndrias [69], acarretando em distanciamento das membranas mitocondriais e danos, principalmente, à membrana mitocondrial interna [79].
A manutenção da morfologia e atividade mitocondrial são características indispensáveis para diversos eventos que estão envolvidos com a maturação citoplasmática e retomada da meiose [108], fertilização [115] e desenvolvimento embrionário [8]. Desta forma, a preservação dessas organelas caracteriza-se como um dos grandes desafi os da criopreservação, uma vez que as mitocôndrias estão relacionadas com a produ-ção de energia celular [8,41] e, consequentemente, a integridade mitocondrial está correlacionada com a viabilidade celular [88].
A vitrifi cação de tecido ovariano de camun-dongas está relacionada com a redução na quantidade
de cristas mitocondriais, alterações na morfologia mitocondrial [66] e aumento no número de mitocôn-drias [55,66] tanto nas células da granulosa quanto nos oócitos. Após criopreservação rápida (redução de 1°C/min) de ovário humano inteiro também foi observada redução nas cristas mitocondriais [62]. No entanto, em tecido ovariano humano submetido à congelação lenta observou-se um elevado número de mitocôn-drias em oócitos de folículos primordiais e primários morfologicamente normais, além de matriz com baixa densidade e poucas cristas periféricas, características consideradas normais em oócitos em estado quiescente ou em desenvolvimento inicial [71].
Em oócitos humanos maturos vitrifi cados foi verifi cada a presença de mitocôndrias com matriz elétron-densa e cristas bem preservadas. Contudo, foi observada uma fragmentação da matriz mitocondrial e desintegração da membrana de algumas mitocôndrias [12]. Já após a congelação lenta de oócitos humanos maturos (MII) as mitocôndrias encontravam-se unifor-memente distribuídas no citoplasma. No entanto, estas mitocôndrias possuíam aparentes danos nas cristas mitocondriais, caracterizadas por cristas que não atra-vessavam a matriz elétron-densa. Além disso, em um alto percentual de oócitos criopreservados (72%) foi observada uma redução na densidade da matriz mito-condrial [40]. Quando a congelação lenta foi utilizada em ovários de camundongas, foi observada presença de mitocôndrias túrgidas e com poucas cristas [66].
As mitocôndrias também têm sido bastante investigadas em embriões criopreservados em dife-rentes espécies, como bovinos [107] e suínos [31], uma vez que o ATP produzido pelas mitocôndrias é essencial para diversos processos celulares em embri-ões em desenvolvimento, incluindo a divisão celular e replicação do DNA genômico [8]. Em embriões de camundongos, após a congelação lenta, foi observado que a maioria das mitocôndrias apresentava-se agru-pada, principalmente, ao longo da membrana celular. Dentre as mitocôndrias presentes no citoplasma, grande parte exibiu um baixo potencial de membrana, evidenciando que a criopreservação é capaz de induzir a despolarização da membrana mitocondrial [2]. Em bovinos, a congelação lenta de embriões de 7 dias não demonstrou alterações nas mitocôndrias. Contu-do, ao se criopreservar embriões bovinos de 13 dias, observou-se danos na membrana das mitocôndrias e perda da homogeneidade de sua matriz [69].

8
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
Apesar da presença de danos nas mitocôndrias decorrentes do procedimento de criopreservação, um estudo com blastocistos bovinos vitrifi cados sugere que esse tipo de dano pode ser revertido após algumas horas de incubação, cultivando-se os embriões pós-aquecimento [107]. Nesse estudo, embriões analisados logo depois de aquecidos demonstraram alterações mitocondriais e redução de elétron-densidade da matriz ou mitocôndrias túrgidas e com aumento da elétron-densidade da matriz. Curiosamente, após 4 h de incubação as mitocôndrias retomaram seu tamanho normal e apenas uma minoria apresentava áreas com redução de elétron-densidade. Após 24 h de incuba-ção, as mitocôndrias apresentavam-se com formato normal e matriz elétron-densa. De forma contrária, em blastocistos suínos vitrifi cados observou-se o desaco-plamento da membrana mitocondrial interna e matriz mitocondrial com redução de elétron-densidade [53]. Ainda após vitrifi cação de blastocistos suínos, Fabian et al. [31] observaram uma degeneração extensiva das mitocôndrias com desligamento parcial das membranas interna e externa quando os embriões foram cultivados por 24 h após aquecimento.
Outra importante organela que sofre danos em decorrência da criopreservação é o retículo endoplas-mático, cuja modifi cação na morfologia foi observada após a vitrifi cação de embriões suínos, resultando em dilatação do retículo endoplasmático liso e presença de pequenas vesículas [31]. Alterações nessa organela também foram perceptíveis em oócitos suínos [13] e caprinos [17], com enturgecimento da mesma após a congelação do tecido ovariano (Figuras 2C e 2D). Acre-dita-se que essa deformação ocorre como resultado de mudanças no balanço iônico causado por alterações na permeabilidade da membrana plasmática [21]. O entumecimento de retículo endoplasmático e cristas mitocondriais [23,45] também foi observado após a vitrifi cação de oócitos bovinos maturos e imaturos, com estreita proximidade entre as gotículas lipídicas, mitocôndrias e retículo endoplasmático [23]. Contudo, a congelação lenta de oócitos imaturos inclusos em te-cido ovariano bovino mostrou-se efi ciente na preserva-ção dos retículos endoplasmáticos (liso e rugoso), não sendo perceptíveis alterações nessas organelas [18].
Em tecido ovariano ovino, após a congelação lenta foi observado um número variável de vesículas, complexo de Golgi bem desenvolvido e retículos en-doplasmáticos (liso e rugoso) agregados entre si ou
formando um complexo com mitocôndrias ou vesículas [93]. Essas características evidenciam a preservação efi caz destas organelas, uma vez que também foram observadas em tecido ovariano fresco de ovinos [32], caprinos [97] e humanos [44]. A preservação de mitocôndrias e retículos endoplasmáticos (liso e rugoso) também foi observada nos oócitos e células da granulosa de ovários de camundongas submetidos ao procedimento de vitrifi cação [42].
As crioinjúrias resultantes da congelação lenta de tecido ovariano humano mostram-se sutis, uma vez que após aplicação da técnica observou-se numerosos complexos de Golgi e retículo endoplasmático liso e rugoso preservados no citoplasma das células folicu-lares [71]. A vitrifi cação de tecido ovariano humano também não resultou em grandes alterações nas organe-las celulares, confi rmado pela presença de complexos de Golgi e retículos endoplasmáticos bem defi nidos e abundantes (Figura 2E) [95]. Contudo, quando a vitrifi cação foi empregada em oócitos maturos huma-nos, complexos de Golgi foram encontrados apenas ocasionalmente e o citoplasma oocitário mostrou-se ocupado de forma homogênea por vesículas dilatadas de retículo endoplasmático liso, em virtude da ruptu-ra destas organelas. Em adição, complexos retículo endoplasmático - mitocôndrias foram encontrados esporadicamente [12].
4. Danos nucleares
A sobrevivência imediata de uma célula de-pende da sua capacidade de evitar alterações no seu DNA. Apesar do grande esforço da célula para manter a integridade do seu material genético, diversos proces-sos desafi am essa capacidade intrínseca. Sabe-se que a criopreservação pode resultar em fragmentação do DNA, mesmo na ausência de deformação da morfolo-gia celular. Porém, os mecanismos subjacentes a esses danos ainda não são conhecidos [68]. Além disso, já foi demonstrado que os cromossomos são muito sus-ceptíveis a alterações decorrentes dos procedimentos de criopreservação [14]. Estas alterações podem ser divididas em estruturais e numéricas. As alterações numéricas podem ainda ser subdividas em alterações na qual todo o genoma é multiplicado (poliploidias) e alterações no qual apenas poucos cromossomos são duplicados ou perdidos (aneuploidias) [54].
Dentre as alterações cromossômicas, a aneu-ploidia é a mais frequente e parece comprometer o desenvolvimento competente do oócito [116], além
9
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
de ser indicada como o principal fator para a baixa obtenção de nascidos vivos [14,67]. Essas alterações cromossômicas também podem ser decorrentes da criopreservação de oócitos. Van der Elst et al. [109] observaram que ao criopreservarem oócitos de camun-dongas, a formação parcial do fuso meiótico ocasiona o deslocamento cromossômico em alguns oócitos, podendo resultar em aneuploidia ou retenção do se-gundo corpúsculo polar no momento da fecundação originando, consequentemente, embriões poliplóides.
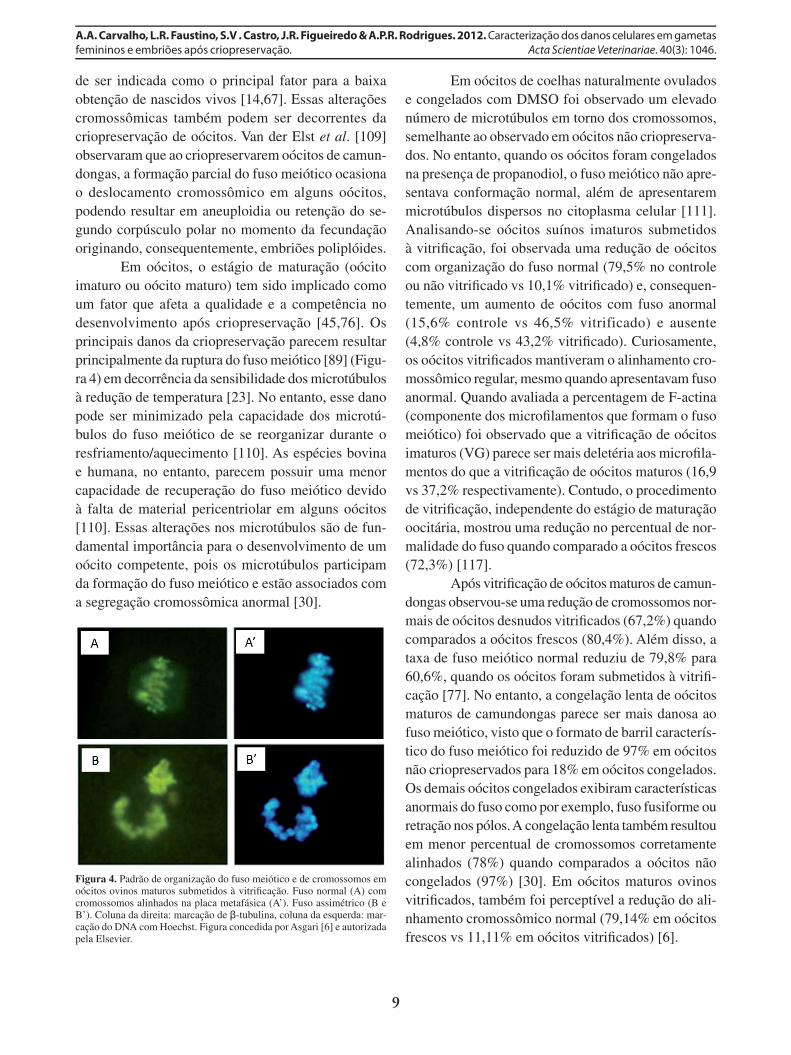
Em oócitos, o estágio de maturação (oócito imaturo ou oócito maturo) tem sido implicado como um fator que afeta a qualidade e a competência no desenvolvimento após criopreservação [45,76]. Os principais danos da criopreservação parecem resultar principalmente da ruptura do fuso meiótico [89] (Figu-ra 4) em decorrência da sensibilidade dos microtúbulos à redução de temperatura [23]. No entanto, esse dano pode ser minimizado pela capacidade dos microtú-bulos do fuso meiótico de se reorganizar durante o resfriamento/aquecimento [110]. As espécies bovina e humana, no entanto, parecem possuir uma menor capacidade de recuperação do fuso meiótico devido à falta de material pericentriolar em alguns oócitos [110]. Essas alterações nos microtúbulos são de fun-damental importância para o desenvolvimento de um oócito competente, pois os microtúbulos participam da formação do fuso meiótico e estão associados com a segregação cromossômica anormal [30].
Figura 4. Padrão de organização do fuso meiótico e de cromossomos em oócitos ovinos maturos submetidos à vitrifi cação. Fuso normal (A) com cromossomos alinhados na placa metafásica (A’). Fuso assimétrico (B e B’). Coluna da direita: marcação de β-tubulina, coluna da esquerda: mar-cação do DNA com Hoechst. Figura concedida por Asgari [6] e autorizada pela Elsevier.
Em oócitos de coelhas naturalmente ovulados e congelados com DMSO foi observado um elevado número de microtúbulos em torno dos cromossomos, semelhante ao observado em oócitos não criopreserva-dos. No entanto, quando os oócitos foram congelados na presença de propanodiol, o fuso meiótico não apre-sentava conformação normal, além de apresentarem microtúbulos dispersos no citoplasma celular [111]. Analisando-se oócitos suínos imaturos submetidos à vitrifi cação, foi observada uma redução de oócitos com organização do fuso normal (79,5% no controle ou não vitrifi cado vs 10,1% vitrifi cado) e, consequen-temente, um aumento de oócitos com fuso anormal (15,6% controle vs 46,5% vitrificado) e ausente (4,8% controle vs 43,2% vitrifi cado). Curiosamente, os oócitos vitrifi cados mantiveram o alinhamento cro-mossômico regular, mesmo quando apresentavam fuso anormal. Quando avaliada a percentagem de F-actina (componente dos microfi lamentos que formam o fuso meiótico) foi observado que a vitrifi cação de oócitos imaturos (VG) parece ser mais deletéria aos microfi la-mentos do que a vitrifi cação de oócitos maturos (16,9 vs 37,2% respectivamente). Contudo, o procedimento de vitrifi cação, independente do estágio de maturação oocitária, mostrou uma redução no percentual de nor-malidade do fuso quando comparado a oócitos frescos (72,3%) [117].
Após vitrifi cação de oócitos maturos de camun-dongas observou-se uma redução de cromossomos nor-mais de oócitos desnudos vitrifi cados (67,2%) quando comparados a oócitos frescos (80,4%). Além disso, a taxa de fuso meiótico normal reduziu de 79,8% para 60,6%, quando os oócitos foram submetidos à vitrifi -cação [77]. No entanto, a congelação lenta de oócitos maturos de camundongas parece ser mais danosa ao fuso meiótico, visto que o formato de barril caracterís-tico do fuso meiótico foi reduzido de 97% em oócitos não criopreservados para 18% em oócitos congelados. Os demais oócitos congelados exibiram características anormais do fuso como por exemplo, fuso fusiforme ou retração nos pólos. A congelação lenta também resultou em menor percentual de cromossomos corretamente alinhados (78%) quando comparados a oócitos não congelados (97%) [30]. Em oócitos maturos ovinos vitrifi cados, também foi perceptível a redução do ali-nhamento cromossômico normal (79,14% em oócitos frescos vs 11,11% em oócitos vitrifi cados) [6].

10
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
Em pesquisa realizada com embriões humanos foi observado que a sobrevivência embrionária pós-congelação está relacionada com o número de células, cujo núcleo foi preservado, e a redução da sobrevivên-cia embrionária foi relacionada com o maior número de células do embrião no momento da criopreserva-ção [101], sugerindo que embriões em estágio mais avançado podem ser mais susceptíveis às crioinjúrias. A vitrifi cação de embriões suínos resultou em um acréscimo na perda de gestações [67] correlacionada principalmente com aneuplodia [14]. No entanto, ainda não é conhecido o mecanismo exato pelo qual essas alterações cromossômicas são ocasionadas nem o estágio ao qual o embrião está mais vulnerável aos danos nucleares.
V. CONCLUSÃO
A criopreservação é uma importante alternativa para a preservação de gametas e embriões, sendo, por isso, bastante aplicada para armazenar o material gené-
tico de animais com alto valor zootécnico, bem como restaurar a fertilidade de mulheres após tratamentos oncológicos. Contudo, este procedimento submete as células a condições desfavoráveis que podem com-prometer a recuperação celular após descongelação/aquecimento. Conforme apontado no decorrer desta revisão, diversos autores têm demonstrado que a cau-sa na redução da sobrevivência de oócitos, folículos ovarianos (presentes ou isolados do tecido ovariano) e embriões após criopreservação está relacionada com danos em diferentes compartimentos e estruturas celulares. Essas estruturas são essenciais para o meta-bolismo e viabilidade da célula e, portanto, a aplicação de um protocolo de criopreservação capaz de manter essas estruturas intactas deve levar em conta o grau de desenvolvimento, tipo e número de células.
Declaration of interest. The authors report no confl icts of interest. The authors alone are responsible for the content and writing of the paper.
REFERÊNCIAS
1 Acker J.P. & McGann L.E. 2003. Protective effect of intracellular ice during freezing? Cryobiology. 46(2): 197-202.2 Ahn H.J., Sohn I.P., Kwon H.C., Jo D.H., Park Y.D. & Min C.K. 2002. Charateristics of the cell membrane fl uidity,
actin fi bers, and mitochondrial disfunctions of frozen-thawed two-cell mouse embryos. Molecular Reproduction and Development. 61(4): 466-476.
3 Albertini D.F. & Eppig J.J. 1995. Unusual cytoskeletal and chromatin confi gurations in mouse oocytes that are atypi-cal in meiotic progression. Developmental Genetics. 16(1): 13-19.
4 Allworth A.E. & Albertini D.F. 1993. Meiotic maturation in cultured bovine oocytes is accompanied by remodeling of the cumulus cell cytoskeleton. Developmental Biology. 158(1): 101-112.
5 Arav A., Pearl M. & Zeron Y. 2000. Does membrane lipid profi le explain chilling sensitivity and membrane lipid phase transition of spermatozoa and oocytes? Cryo Letters. 21(3): 179-186.
6 Asgari V., Hosseini S.M., Ostadhosseini S., Hajian M. & Nars-Esfahani M.H. 2011. Time dependent effect of post warming interval on microtubule organization, meiotic status, and parthenogenetic activation of vitrifi ed in vitro matured sheep oocytes. Theriogenology. 75(5): 904-910.
7 Bakhach J. 2009. The cryopreservation of composite tissues - Principles and recent advancement on cryopreservation of different type of tissues. Organogenesis. 5(3): 119-126.
8 Bavister B.D. 1995. Culture of pre-implantation embryo: facts and artifacts. Human Reproduction Update. 1(2): 91-148.9 Benson E.E. 2008. Cryopreservation Theory. In: Reed B.M. (Ed). Plant Cryopreservation: A Practical Guide. Corval-
lis: Springer, pp.15-32.10 Bodin S., Soulet C., Tronchère H., Sié P., Gachet C., Plantavid M. & Payrastre B. 2005. Integrin-dependent inter-
action of lipid rafts with the actin cytoskeleton in activated human platelets. Journal of Cell Science. 118(4): 759-769.11 Bogliolo L., Ariu F., Fois S., Rosati I., Zedda M.T., Leoni G., Succu S., Pau S. & Ledda S. 2007. Morphological and
biochemical analysis of immature ovine oocytes vitrifi ed with or without cumulus cells. Theriogenology. 68(8): 1138-1149.12 Bonetti A., Cervi M., Tomei F., Marchini M., Ortolani F. & Manno M. 2011. Ultrastructural evaluation of human
metaphase II oocytes after vitrifi cation: closed versus open devices. Fertility and Sterility. 95(3): 928-935.13 Borges E.N., Silva R.C., Futino D.O., Rocha-Junior C.M.C., Amorim C.A., Báo S.N. & Lucci C.M. 2009. Cryop-
reservation of swine ovarian tissue: Effect of different cryoprotectants on the structural preservation of preantral follicle oocytes. Cryobiology. 59(2): 195-200.

11
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
14 Burgoyne P.S., Holland K. & Stephens R. 1991. Incidence of numerical chromosome anomalies in human pregnancy estimation from induced and spontaneous abortion data. Human Reproduction. 6(4): 555-565.
15 Carmo-Fonseca M. & David-Ferreira J.F. 1990. Interactions of intermediate fi laments with cell structures. Electron Microscopy Reviews. 3(1): 115-141.
16 Carroll J., Depypere H. & Matthews C.D. 1990. Freeze-thaw-induced changes of the zona pellucida explains de-creased rates of fertilization in frozen-thawed mouse oocytes. Journal of Reproduction and Fertility. 90(2): 547-553.
17 Castro S.V., Carvalho A.A., Silva C.M., Faustino L.R., Campello C.C., Lucci C.M., Báo S.N., Figueiredo J.R. & Rodrigues A.P.R. 2011. Freezing solution containing dimethylsulfoxide and fetal calf serum maintains survival and ultrastructure of goat preantral follicles after cryopreservation and in vitro culture of ovarian tissue. Cell and Tissue Research. 346(2): 283-92.
18 Celestino J.J.H., Santos R.R., Lopes C.A.P., Martins F.S., Matos M.H.T., Melo M.A.P., Báo S.N., Rodrigues A.P.R., Silva J.R.V. & Figueiredo J.R. 2008. Preservation of bovine preantral follicle viability and ultra-structure after cooling and freezing of ovarian tissue. Animal Reproduction Science. 108(3-4): 309-318.
19 Cha K.Y., Chung H.M., Lim J.M., Ko J.J., Han S.Y., Choi D.H. & Yoon T.K. 2000. Freezing immature oocytes. Molecular and Cellular Endocrinology. 169(1-2): 43-47.
20 Chan D.C. 2006. Mitochondria: Dynamic organelles in disease, aging, and development. Cell. 125(7): 1241-1252.21 Coss R.A., Dewey W.C. & Bamburg J.R. 1979. Effects of hyperthermia (41,5°) on Chinese hamster ovary cells
analyzed in mitosis. Cancer Research. 39: 1911-1918.22 Crowe J.H., Hoekstra F.A. & Crowe L.M. 1992. Anhydrobiosis. Annual review of Physiology. 54: 579-599.23 Diez C., Duque P., Gómez E. Hidalgo C.O., Tamargo C., Rodríguez A., Fernández L., De La Varga S., Frenández
A., Facal N. & Carbajo M. 2005. Bovine oocyte vitrifi cation before or after meiotic arrest: effects on ultrastructure and developmental ability. Theriogenology. 64 (2): 317-333.
24 Dobrinsky J.R. 2002. Advancements in cryopreservation of domestic animal embryos. Theriogenology. 57(1)-285-302.25 Donnez J., Dolmans M.M., Demylle D., Jadoul P., Pirard C., Squiffl et J., Martinez-Madrid B. & Van Langendonckt
A. 2004. Livebirth after orthotopic transplantation of cryopreserved ovarian tissue. The Lancet. 364(9443): 1405-1410.26 Ducibella T., Schultz R.M. &Ozil J.P. 2006. Role of calcium signals in early development. Seminars in Cell & De-
velopmental Biology. 17(2): 324-332.27 Dulioust E., Toyama K., Busnel M.C., Moutier R., Carlier M., Marchaland C., Ducot B., Roubertoux P. &
Auroux M. 1995. Long-term effects of embryo freezing in mice. Proceedings of the National Academy of Sciences of the United States of America. 92(2): 589-593.
28 Elliott J.A.W. 2010. Introduction to the special issue: Thermodynamic aspects of cryobiology. Cryobiology. 60(1): 1-3.
29 Eroglu A., Toner M., Leykin L. & Toth T.L. 1998. Cytoskeleton and polyploidy after maturation and fertilization of cryopreserved germinal vesicle-stage mouse oocytes. Journal of Assisted Reproduction and Genetics. 15(7):447-454.
30 Eroglu A., Toth T.L. & Toner M. 1998. Alterations of the cytoskeleton and polyploidy induced by cryopreservation of metaphase II mouse oocytes. Fertility and Sterility. 69(5): 944-957.
31 Fabian D., Gjørret J.O., Berthelot F., Martinat-Botté F. & Maddox-Hyttel P. 2005. Ultrastructure and cell death of in vivo derived and vitrifi ed porcine blastocysts. Molecular Reproduction and Development. 70(2): 155-165.
32 Fair T., Hulshof S.C.J., Hyttel P., Greve T. & Boland M. 1997. Oocyte ultrastructure in bovine primordial to early tertiary follicles. Anatomy and Embryology. 195(4): 327-336.
33 Foster L.J., Hoog C.L., Zhang Y., Zhang Y., Xie X., Mootha V.K. & Mann M. 2006. A mammalian organelle map by protein correlation profi ling. Cell. 125(1): 187-199.
34 Fu X.W., Shi W.Q., Zhang Q.J., Zhao X.M., Yan C.L., Hou Y.P., Zhou G.B., Fan Z.Q., Suo L., Wusiman A., Wang Y.P & Zhu S.E. 2009. Positive effects of Taxol pretreatment on morphology, distribution and ultrastructure of mitochondria and lipid droplets in vitrifi cation of in vitro matured porcine oocytes. Animal Reproduction Science. 115(1-4): 158-168.
35 Fu X.W., Wu G.Q., Li J.J., Hou Y.P., Zhou G.B., Suo L., Wang Y.P. & Zhu S.E. 2011. Positive effects of Forskolin (stimulator of lipolysis) treatment on cryosurvival of matured porcine oocytes. Theriogenology. 75(2): 268-275.
36 Fuku E., Xia L. & Downey B.R. 1995. Ultrastructural changes in bovine oocytes cryopreserved by vitrifi cation. Cryobiology. 32(2): 139-156.

12
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
37 Ghetler Y., Yavin S., Shalgi R. & Arav A. 2005. The effect of chilling on membrane lipid phase transition in human oocytes and zygotes. Human Reproduction. 20(12): 3385-3389.
38 Gomes C.M., Silva C.A.S., Acevedo N., Baracat E., Serafi ni P. & Smith G.D. 2008. Infl uence of vitrifi cation on mouse metaphase II oocyte spindle dynamics and chromatin alignment. Fertility and Sterility. 90(4): 1396-1404.
39 Gosden R.G., Baird D.T., Wade J.C. & Webb R. 1994. Restoration of fertility to oophorectomized sheep by ovarian autografts stored at -196°C. Human Reproduction. 9(4): 597-603.
40 Gualtieri R., Iaccarino M., Mollo V., Prisco M., Iaccarino S. & Talevi R. 2009. Slow cooling of human oocytes: ultrastructural injuries and apoptotic status. Fertility and Sterility. 91(4): 1023-1034.
41 Guyton A.C. & Hall J.E. 2006. The Cell and Its Functions. In: Textbook of Medical Physiology. Philadelphia: Elsevier Saunders, pp.11-26.
42 Haidari K. Salehnia M. & Valojerdi M.R. 2008. The effect of leukemia inhibitory factor and coculture on the in vitro maturation and ultrastructure of vitrifi ed and nonvitrifi ed isolated mouse preantral follicles. Fertility and Sterility. 90(6): 2389-2397.
43 Hall P.F. 1997. The roles of calmodulin, actin, and vimentin in steroid synthesis by adrenal cells. Steroids. 62(1): 185-189. 44 Hertig A.T. & Adams E.C. 1967. Studies on the human oocyte and its follicle. I. Ultrastructural and histochemical
observation on the primordial follicle stage. The Journal of Cell Biology. 34(2): 647-675.45 Hochi S., Ito K., Hirabayashi M., Ueda M., Kimura K. & Hanada A. 1998. Effect of nuclear stages during IVM
on the survival of vitrifi ed-warmed bovine oocytes. Theriogenology. 49(4): 787-796.46 Hochi S., Kozawa M., Fujimoto T., Hondo E., Yamada J. & Oguri N. 1996. In vitro maturation and transmission
electron microscopic observation of horse oocytes after vitrifi cation. Cryobiology. 33(3): 300-310.47 Hyttel P., Vajta G. & Callesen H. 2000. Vitrifi cation of bovine oocytes with the open pulled straw method: ultra-
structural consequence. Molecular Reproduction and Development. 56(1): 80-88.48 Isachenko V., Isachenko E., Michelmann H.W., Alabart J.L., Vazquez I., Bezugly N. & Nawroth F. 2001. Lipolysis
and ultrastructural changes of intracellular lipid vesicles after cooling of bovine and porcine GV-oocytes. Anatomia, Histologia, Embryologia. 30(6): 333-338.
49 Isachenko V., Soler C., Isachenko E., Perez-Sanchez F. & Grishchenko V. 1998. Vitrifi cation of immature porcine oocytes: effects of lipid droplets, temperature, cytoskeleton, and addition and removal of cryoprotectant. Cryobiology. 36(3): 250-253.
50 Ishijima T., Kobayashi Y., Lee D.S., Ueta Y.Y., Matsui M., Lee J.Y., Suwa Y., Miyahara K. & Suzuki H. 2006. Cryopreservation of canine ovaries by vitrifi cation. Journal of Reproduction and Development. 52(2): 293-299.
51 Jelinkova L., Selman H.A., Arav A., Strehler E., Reeka N. & Sterzik K. 2002. Twin pregnancy after vitrifi cation of 2-pronuclei human embryos. Fertility and Sterility. 77(2): 412-414.
52 Johnson M.H., Pickering S.J. & George M.A. 1988. The infl uence of cooling on the properties of the zona pellucida of the mouse oocyte. Human Reproduction. 3(3): 383-387.
53 Kaidi S., Bernard S., Lambert P., Massip A., Dessy F. & Donnay I. 2001. Effect of conventional controlled-rate freezing and vitrifi cation on morphology and metabolism of bovine blastocysts produced in vitro. Biology of Reproduc-tion. 65(4): 1127-1134.
54 Karp G. 2002. O núcleo da célula e o controle da expressão gênica. In: Biologia Celular e Molecular. 3.ed. Barueri: Manole, pp.494-550.
55 Kim G.A., Kim H.Y., Kim J.W., Lee G., Lee E. & Lim J.M. 2010. Ultrastructural deformity of ovarian follicles induced by different cryopreservation protocols. Fertility and Sterility. 94(4): 1548-1550.
56 Larman M.G., Sheehan C.B. & Gardner D.K. 2006. Calcium-free vitrifi cation reduces cryoprotectant-induced zona pellucida hardening and increases fertilization rates in mouse oocytes. Reproduction. 131(1): 53-61.
57 Lin T.A., Chen C.H., Sung L.Y., Carter M.G., Chen Y.E., Du F., Ju J.C. & Xu J. 2011. Open-pulled straw vitrifi cation differentiates cryotolerance of in vitro cultured rabbit embryos at the eight-cell stage. Theriogenology. 75(4): 760-768.
58 Lovelock J.E. 1953. The haemolysis of human red blood cells by freezing and thawing. Biochimica et Biophysica Acta. 10(3): 414-426.
59 Luciano A.M., Chigioni S., Lodde V., Franciosi F., Luvoni G.C. & Modina S.C. 2009. Effect of different cryop-reservation protocols on cytoskeleton and gap junction mediated communication integrity in feline germinal vesicle stage oocytes. Cryobiology. 59(1): 90-95.

13
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
60 Magli M.C., Lappi M., Ferraretti A., Capoti A., Ruberti A. & Gianaroli L. 2010. Impact of oocyte cryopreserva-tion on embryo development. Fertility and Sterility. 93(2): 510-516.
61 Martinéz-Burgos M., Herrero L., Magías D., Salvanes R., Mantoya M.C., Cobo A.C. & Garcia-Velasco J. 2011. Vitrifi cation versus slow freezing of oocytes: effects on morphologic appearance, meiotic spindle confi guration, and DNA damage. Fertility and Sterility. 95(1): 374-377.
62 Martinez-Madrid B., Camboni A., Dolmans M.M., Nottola S., Van Langendonckt A. & Donnez J. 2007. Apoptosis and ultrastructural assessment after cryopreservation of whole human ovaries with their vascular pedicle. Fertility and Sterility. 87(5): 1153-1165.
63 Massip A., Mermillod P. & Dinnyes A. 1995. Morphology and biochemistry of in vitro produced bovine embryos: implications for their cryopreservation. Human Reproduction. 10(11): 3004-3011.
64 Matson P.L., Graefl ing J., Junk S.M., Yovich J.L. & Edirisinghe W.R. 1997. Cryopreservation of oocytes and embryos: use of a mouse model to investigate effects upon zona hardness and formulate treatment strategies in an in vitro fertilization programme. Human Reproduction. 12(7): 1550-1553.
65 Mayerson P.L. & Brumbaugh J.A. 1981. Lavender, chick melanocyte mutant with defective melanosome transloca-tion: a possible role for 10 nm fi laments and microfi laments but not microtubules. Journal of Cell Science. 51: 25-51.
66 Mazoochi T., Salehnia M., Valojerdi M.R. & Mowla J. 2008. Morphologic, ultrastructural, and biochemical iden-tifi cation of apoptosis in vitrifi ed-warmed mouse ovarian tissue. Fertility and Sterility. 90(4): 1480-1486.
67 Men H., Agca Y., Riley L. & Crister J.K. 2006. Improved survival of vitrifi ed porcine embryos after partial delipa-tion through chemically stimulated lipolysis and inhibition of apoptosis. Theriogenology. 66(8): 2008-2016.
68 Men H., Monson R.L. & Rutledge. 2002. Effect of meiotic stages and maturation protocols on bovine oocyte’s resistance to cryopreservation. Theriogenology. 57(3): 1095-1103.
69 Mohr L.R. & Trounson A.O. 1981. Structural changes associated with freezing of bovine embryos. Biology of Re-production. 25(5): 1009-1025.
70 Mothes W., Heinrich S.U., Graf R., Nilsson I., Von Heijne G., Brunner J. & Rapoport T.A. 1997. Molecular mechanism of membrane protein integration into the endoplasmic reticulum. Cell. 89(4): 523-533.
71 Nottola S.A., Camboni A., Van Langendonkt A., Demylle D., Macchiarelli G., Dolmans M.M., Martinez-Madrid B., Correr S. & Donnez J. 2008. Cryopreservation and xenotransplantation of human ovarian tissue: an ultrastructural study. Fertility and Sterility. 90(1): 23-32.
72 Nottola S.A., Cotticchio G., Sciajno R., Gambardella A., Maione M., Scaravelli., Bianchi S., Macchiarelli G. & Borini A. 2009. Ultrastructural markers of quality in human mature oocytes vitrifi ed using cryoleaf and cryoloop. Reproductive BioMedicine Online. 19(Suppl 3): 17-27.
73 Nottola S.A., Macchiarelli G., Coticchio G., Bianchi S., Cecconi S, De Santis L. Scaravelli G., Flamigni C. & Borini A. 2007. Ultrastructure of human mature oocytes after slow cooling cryopreservation using different sucrose concentrations. Human Reproduction. 22(4):1123-1133.
74 Oktay K., Newton H., Aubard Y., Salha O. & Gosden R.G. 1998. Cryopreservation of immature human oocytes and ovarian tissue: an emerging technology? Fertility and Sterility. 69(1): 1-7.
75 Overstrom E.W., Paqui-Platls D., Toner M. & Cravacho E.G. 1990. Cryoprotectant and thermal effect on cytoskel-etal organization and IVF rate of mouse oocytes. Biology of Reproduction. 42(Suppl 1):175.
76 Palasz A.T. & Mapletoft R.J. 1996. Cryopreservation of mammalian embryos and oocytes: Recent advances. Bio-technology Advances. 14(2): 127-149.
77 Park S.E., Chung H.M., Cha K.Y., Hwang W.S., Lee E.S. & Lim J.M. 2001. Cryopreservation of ICR mouse oocytes: improved post-thawed preimplantation development after vitrifi cation using Taxol™, a cytoskeleton stabilizer. Fertility and Sterility. 75(6): 1177-1184.
78 Park S.E., Son W.Y., Lee SH., Lee K.A., Ko J.J. & Cha K.Y. 1997. Chromosome and spindle confi gurations of human oocytes matured in vitro after cryopreservation at the germinal vesicle stage. Fertility and Sterility. 68(5): 920-926.
79 Partridge R.J., Wrathall A.E. & Leese H.J. 1995. Glucose uptake and lactate production by single frozen-thawed bovine embryos produced in vivo or by in vitro fertilization. Journal of Reproduction and Fertility. 13:41.
80 Pelliniemi L.J., Dym M. & Paranko J. 1982. Cytoplasmic fi laments in fetal Leydig cells. Annals of the New York Academy of Sciences. 383(1): 486-487.

14
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
81 Pickering S.J. & Johnson M.H. 1987. The infl uence of cooling on the organization of themeiotic spindle of the mouse oocyte. Human Reproduction. 2(3): 207-216.
82 Polge C., Smith A.U., & Parkes A.S. 1949. Revival of spermatozoa after vitrifi cation and dehydration at low tem-peratures. Nature. 164(4172): 666.
83 Popelková M., Chrenek P., Pivko J., Makarevich A.V., Kubovičová E. & Kačmárik J. 2005. Survival and ul-trastructure of gene-microinjected rabbit embryos after vitrifi cation. Zygote. 13(4): 283-293.
84 Potts M. 1994. Desiccation tolerance of prokaryotes. Microbiological Reviews. 58(4): 755-805.85 Ptak G., Dattena M., Loi P., Tischner M. & Cappai P. 1999. Ovum pick-up in sheep: effi ciency of in vitro embryo
production, vitrifi cation and birth of offspring. Theriogenology. 52(6): 1105-1114.86 Quintans C.J., Donaldson M.J., Bertolino M.V. & Pasqualini R.S. 2002. Birth of two babies using oocytes that
were cryopreserved in a choline-based freezing médium. Human Reproduction. 17(12): 3149-3152.87 Reed M.L., Lane M., Gardner D.K., Jensen N.L. & Thompson J. 2002. Vitrifi cation of human blastocysts using the
cryoloop method: successful clinical application and birth of offspring. Journal of Assisted Reproduction and Genetics. 19(6): 304-306.
88 Rho G.J., Kim S.N, Yoo J.G., Balasubramanian S., Lee H.J & Choe S.Y. 2002. Microtubulin confi guration and mitochondrial distribution after ultra-rapid cooling of bovine oocytes. Molecular Reproduction and Development. 63(4): 464-470.
89 Rienzi L., Martinez F., Ubaldi F., Minasi M.G., Iacobelli M., Tesarik J. & Greco E. 2004. Polscop analysis of meiotic spindle changes in living metaphase II human oocytes during the freezing and thawing procedures. Human Reproduction. 19: 655-659.
90 Rojas C., Palomo M.J., Albarracín J.L. & Mogas T. 2004. Vitrifi cation of immature and in vitro matured pig oocytes: study of distribution of chromosomes, microtubules, and actin microfi laments. Cryobiology. 49(3): 211-220.
91 Rubinsky B. 1989. The energy equation for freezing of biological tissue. Journal of Heat Transfer. 111(4): 988-997.92 Santos R.R., Knijn H.M., Vos P.L.A.M., Oei C.H.Y., Van Loon T., Colenbrander B., Gadella B.M., Van den
Hurk R. & Roelen B.A.J. 2009. Complete follicular development and recovery of ovarian function of frozen-thawed, autotransplanted caprine ovarian cortex. Fertility and Sterility. 91(Suppl 4): 1455-1458.
93 Santos R.R., Rodrigues A.P.R., Costa S.H.F., Silva J.R.V., Matos M.H.T., Lucci C.M., Báo S.N., Van Den Hurk R. & Figueiredo J.R. 2006. Histological and ultrastructural analysis of cryopreserved sheep preantral follicles. Animal Reproduction Science. 91(3-4): 249-263.
94 Saunders K.M. & Parks J.E. 1999. Effects of cryopreservation procedures on the cytology and fertilization rate of in vitro-matured bovine oocytes. Biology of Reproduction. 61(1): 178-187.
95 Sheikhi M., Hultenby., Niklasson B., Lundqvist. & Hovatta O. 2011. Clinical grade vitrifi cation of human ovarian tissue: an ultrastructural analysis of follicles and stroma in vitrifi ed tissue. Human Reproduction. 26(3): 594-603.
96 Shorter J. & Warren G. 2002. Golgi architecture and inheritance. Annual Review of Cell and Developmental Biology. 18:379-420.
97 Silva J.R.V., Báo S.N., Lucci C.M., Carvalho F.C.A., Andrade E.R., Ferreira M.A.L. & Figueiredo J.R. 2001. Morphological and ultrastructural changes occurring during degeneration of goat preantral follicles preserved in vitro. Animal Reproduction Science. 66(3-4): 209-223.
98 Somfai T., Ozawa M., Noguchi J., Kaneko H., Karja N.W.K., Farhudin M., Dinnyés A., Nagai T. & Kikuchi K. 2007. Developmental competence of in vitro-fertilized porcine oocytes after in vitro maturation and solid surface vitrifi cation: Effect of cryopreservation on oocyte antioxidative system and cell cycle stage. Cryobiology. 55(2): 115-126.
99 Steponkus P.L. 1984. Role of the plasma membrane in freezing injury and cold acclimation. Annual Review of Plant Physiology. 35(1): 543-584.
100 Succu S., Leoni G.G., Bebbere D., Berlinguer F., Mossa F., Bogliolo L., Madeddu M., Ledda S. & Naitana S. 2007. Vitrifi cation devices affect structural and molecular status of in vitro matured ovine oocytes. Molecular Repro-duction and Development. 74(10): 1337-1344.
101 Testart J., Lassalle B., Belaisch-Allart J., Hazout A., Forman R., Rainhorn J.D. & Frydman R. 1986. High pregnancy rate after early human embryo freezing. Fertility and Sterility. 46(2): 268-272.
102 Tharasanit T., Colenbrander B. & Stout T.A.E. 2006. Effect of maturation stage at cryopreservation on post-thaw cytoskeleton quality and fertilizability of equine oocytes. Molecular Reproduction and Development. 73(5): 627-637.

15
A.A. Carvalho, L.R. Faustino, S.V . Castro, J.R. Figueiredo & A.P.R. Rodrigues. 2012. Caracterização dos danos celulares em gametas
femininos e embriões após criopreservação. Acta Scientiae Veterinariae. 40(3): 1046.
www.ufrgs.br/actavetPub. 1046
103 Thibodeau G.A. & Patton K.T. 2006. Células e Tecidos. In: Estrutura e Funções do Corpo Humano. 11.ed. Barueri: Manole, pp.20-27.
104 Traub P., Perides G., Kuhn S. & Scherberth A. 1987. Effi cient interaction of nonpolar lipids with intermediate fi laments of the vimentin type. European Journal of Cell Biology. 43(1): 55-64.
105 Uemura M. & Steponkus P.L.1994. A contrast of the plasma membrane lipid composition of oat and rye leaves in relation to freezing tolerance. Plant Physiology. 104(2): 479-496.
106 Vajta G., Holm P., Kuwayama M., Booth P.J., Jacobsen H., Greve T. & Callesen H. 1998. Open pulled straw (OPS) vitrifi cation: a new way to reduce cryoinjuries of bovine ova and embryos. Molecular Reproduction and De-velopment. 51(1): 53-58.
107 Vajta G., Hyttel P. & Callesen H. 1997. Morphological changes of in-vitro-produced bovine blastocysts after vitri-fi cation, in-straw direct rehydration, and culture. Molecular Reproduction and Development. 48(1): 9-17.
108 Van Blerkom J. 1991. Microtubule mediation of cytoplasmic and nuclear maturation during the early stages of resumed meiosis in cultured mouse oocyte. Proceedings of the National Academy of Sciences of the United States of America. 88(11): 5031-5035.
109 Van der Elst J., Nerinckx S. & Van Steirteghem A. 1993. Association of ultrarapid freezing of mouse oocytes with increased polyploid at the pronucleate stage, reduced cell numbers in blastocysts, and impaired fetal development. Journal of Reproduction and Fertility. 99(1): 25-32.
110 Vincent C. & Johnson M.H. 1992. Cooling, cryoprotectants, and the cytoskeleton of the mammalian oocytes. Oxford Reviews of Reproductive Biology. 14:73-100.
111 Vincent C., Garnier V., Heyman H. & Renard J.P. 1989. Solvent effects on cytoskeletal organization and in-vivo survival after freezing of rabbit oocytes. Journal of Reproduction and Fertility. 87(2): 809-820.
112 Vincent C., Pickering S.J. & Johnson M.H. 1990. The hardening effect of dimethylsulfoxide on the mouse zona pellucida requires the presence of an oocyte and is associated with a reduction in the number of cortical granules present. Journal of Reproduction and Fertility. 89(1): 253-259.
113 Vincent C., Pickering S.J., Johnson M.H. & Quick S.J. 1990. Dimethylsulphoxide affects the organization of microfi laments in the mouse oocyte. Molecular Reproduction and Development. 26(3): 227-235.
114 Wanderley L.S., Luz H.K.M., Faustino L.R., Lima I.M.T., Lopes C.A.P., Silva A.R., Báo S.N., Campello C.C., Rodrigues A.P.R. & Figueiredo J.R. 2012. Ultrastructural features of agouti (Dasyprocta aguti) preantral follicles cryopreserved using dimethyl sulfoxide, ethylene glycol and propanediol. Theriogenology. 77(2): 260-267.
115 Whitaker M. 2006. Calcium at fertilization and in early development. Physiological Reviews. 86(1): 25-88.116 Wu B., Tong J. & Leibo S.P. 1999. Effects of cooling germinal vesicle-stage bovine oocytes on meiotic spindle
formation following in vitro fertilization. Molecular Reproduction and Development. 54(4): 388-395.117 Wu C., Rui R., Dai J., Zhang C., Ju S., Xie B., Lu X. & Zheng X. 2006. Effects of cryopreservation on the de-
velopmental competence, ultrastructure and cytoskeletal structure of porcine oocytes. Molecular Reproduction and Development. 73(11): 1454-1462.
118 Yoon T.K., Chung H.M., Lim J.M., Han S.Y., Ko J.J. & Cha K.Y. 2000. Pregnancy and delivery of healthy infants developed from vitrifi ed oocytes in a stimulated in vitro fertilization-embryo transfer program. Fertility and Sterility. 74(1): 180-181.
119 Zeron Y., Ocheretny A., Kedar O., Borochov A., Sklan D. & Arav A. 2001. Seasonal changes in bovine fertility: relation to developmental competence of oocytes, membrane properties and fatty acid composition of follicles. Repro-duction. 121(3): 447-454.
120 Zeron Y., Sklan D. & Arav A. 2002. Effect of polyunsaturated fatty acid supplementation on biophysical parameters and chilling sensitivity of ewe oocytes. Molecular Reproduction and Development. 61(2): 271-278.
121 Zeron Y., Tomczak M., Crowe J. & Arav A. 2002. The effect of liposomes on thermotropic membrane phase transi-tions of bovine spermatozoa and oocytes: implications for reducing chilling sensitivity. Cryobiology. 45(2): 143-152.