caracterização de microdomínios lipídicos de...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
CENTRO DE CIÊNCIAS DA SAÚDE / INSTITUTO DE BIOQUÍMICA MÉDICA
FERNANDA SOUZA DE OLIVEIRA CAMPOS
Caracterização de microdomínios lipídicos de membrana
(lipid rafts) da glia embainhante olfatória
Rio de Janeiro
2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
Fernanda Souza de Oliveira Campos
Caracterização de microdomínios lipídicos de membrana (lipid
rafts) da glia embainhante olfatória
Dissertação de Mestrado submetida ao Programa de Pós-
Graduação em Química Biológica, Instituto de Bioquímica
Médica, Universidade Federal do Rio de Janeiro.
Orientadoras: Georgia Corrêa Atella
Leny Alves Cavalcante
Rio de Janeiro
2009

iii
CARACTERIZAÇÃO DE MICRODOMÍNIOS LIPÍDICOS DE
MEMBRANA (LIPID RAFTS) DA GLIA EMBAINHANTE OLFATÓRIA
Fernanda Souza de Oliveira Campos
Dissertação de Mestrado apresentada ao Programa de Pós-Graduação em Química Biológica, Instituto de Bioquímica Médica, Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Mestre em Ciências Biológicas (Química Biológica).
Aprovada por:
____________________________________Orientadora
Profa. Georgia Corrêa Atella (Instituto de Bioquímica Médica - UFRJ)
____________________________________Orientadora
Profa. Leny Alves Cavalcante (Instituto de Biofísica Carlos Chagas Filho - UFRJ)
____________________________________Revisor / Suplente Interno
Prof. Aurélio Vicente Graça de Souza (Instituto de Bioquímica Médica - UFRJ)
____________________________________Examinador
Prof. André Marco de Oliveira Gomes (Instituto de Bioquímica Médica - UFRJ)
____________________________________Examinadora
Profa. Silvana Allodi (Instituto de Ciências Biomédicas - UFRJ)
____________________________________Examinadora
Profa. Penha Cristina Barradas Daltro Santos (Departamento Farmacologia e Psicobiologia- IB- UERJ)
____________________________________Suplente Externa
Profa. Cláudia dos Santos Mermelstein (Instituto de Ciências Biomédicas - UFRJ)
Rio de Janeiro - Agosto / 2009

iv
FICHA CATALOGRÁFICA
Campos, Fernanda Souza de Oliveira
Caracterização de microdomínios lipídicos de membrana (lipid rafts) da glia embainhante olfatória / Fernanda Souza de Oliveira Campos. Rio de Janeiro, 2009.
83pp, xvi
Orientadoras: Georgia Corrêa Atella e Leny Alves Cavalcante.
Dissertação (Mestrado) - Universidade Federal do Rio de Janeiro, Instituto de Bioquímica Médica, 2009.
1. Glia embainhante olfatória - 2. Microdomínios Lipídicos - 3. CNPase (2’, 3’-nucleotídeo cíclico 3’-fosfodiesterase).

v
O presente trabalho foi realizado nos Laboratório de Bioquímica de Lipídeos e Lipoproteínas
no Instituto de Bioquímica Médica e de Neurobiologia do Desenvolvimento do Instituto de
Biofisica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro - UFRJ, sob a
orientação das Professoras Geórgia Correa Atella e Leny Alves Cavalcante, na vigência de
auxílios concedidos pela Coordenação de Aperfeiçoamento de Pessoal de Nível Superior -
CAPES e pelo Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq).

vi
À minha família

vii
AGRADECIMENTOS
Agradeço primeiramente a Deus por estar sempre ao meu lado e nunca ter me
desamparado.
Agradeço principalmente aos meus pais, Jaime e Neusa. Tudo o que sou hoje na vida é
fruto do amor desse casal maravilhoso. Vocês não existem!!! Parte da minha formação
também depende das minhas irmãs (Maria José, Renata e Juliana). Obrigada pela força que
vocês me dão. Amo vocês!
Agradeço as minhas orientadoras, Geórgia Corrêa Atella e Leny Alves Cavalcante, pela
orientação eficiente ao longo desses anos e por todo o carinho depositado em mim nessa longa
caminhada.
Agradeço ao meu amor e ao meu companheiro de trabalho, Felipe. Sem ele, em minha
vida muitas histórias eu não teria vivido. Obrigada sempre por estar ao meu lado, ser
compreensivo e me ajudar nas minhas dificuldades. Te amo!
Agradeço aos meus amigos do laboratório NBD: Eliane, Fred, Hugo, Lítia, Paulinho,
Serginho, Tainá, Vivi e Wagner. Obrigada por tornar meus dias no laboratório mais
“divertidos”, por todos os bons momentos, pelas ajudas nos protocolos... E também a minha
ex-coorientadora, Alessandra Santos Silva, pelo início da minha iniciação científica.
Agradeço aos meus amigos do laboratório de Bioquímica: Nuccia, Nicole, Lívia, Karla,
Michele, Paulinha, Alessandra e Liliane. Obrigada por me ensinarem a base da bioquímica
para os meus experimentos. Agradeço também a Miria, companheira de trabalho das rafts.
Agradeço as minhas amigas, Marcella, Vinha e Bárbara pela amizade que começou na
faculdade e espero que dure por muitos outros longos anos.
Agradeço a Maria, Nanana e Marcella por toda força, carinho e amizade que foram
fundamentais na minha formação pessoal.
As minhas amigas Anna, Virgínia, Flávia, Fernanda e respectivos. Pelo carinho, amizade,
cumplicidade que duram mais de 9 anos.
A todos aqueles que me ajudaram de alguma maneira para que essa dissertação virasse
realidade.

viii
RESUMO
A glia embainhante olfatória (GEO) tem sido apresentada como uma ferramenta
promissora da bioengenharia para reparo de lesões no sistema nervoso. No entanto, o efetivo
uso desta célula em terapias neuroregenerativas ainda depende de um melhor conhecimento
de suas propriedades fundamentais. Desta forma, a caracterização das lipid rafts, estruturas
envolvidas na transdução de sinal, adesão celular, migração e organização do citoesqueleto,
podem representar um ponto fundamental no entendimento dos processos regenerativos da
GEO. Neste trabalho caracterizamos as lipid rafts da GEO, verificando a presença da CNPase
(2’3’-nucleotídeo cíclico 3’-fosfodiesterase), 9-O-acetil GD3 e os gangliosídeos reconhecidos
pelo anticorpo A2B5, expressos em células neurais de alta motilidade. As lipid rafts da GEO,
aqui identificadas pelos marcadores gangliosídeo GM1, flotilina-1 e caveolina-1,
apresentaram fosfatidilcolina, esfingomielina, alta quantidade de colesterol, baixo conteúdo
protéico e uma concentração preferencial em relação ao restante da membrana de
componentes como os gangliosídeos mencionados acima e CNPase. Foi possível ainda
constatar que a destruição das lipid rafts pela metil-beta-ciclodextrina altera a distribuição dos
componentes identificados nestes microdomínios lipídicos. Estes resultados sugerem que as
membranas resistentes a detergente da GEO podem estar, pelo menos parcialmente,
envolvidas em eventos relevantes na migração promovida por esta célula.

ix
ABSTRACT
The olfactory ensheathing cell (OEC) has been referred as a promising tool for
bioengineering in the repair of nervous system injuries. However, the effective use of this cell
in neuroregenerative therapies still depends on better understanding of its fundamental
properties. Thus, the characterization of lipid rafts, which are structures involved in signal
transduction, cell adhesion, migration and cytoskeleton organization, may represent a major
subject in the understanding of the regenerative events promoted by OEC. Our aim is the
purification and characterization of OEC’s lipid rafts, verifying the presence of CNPase (non-
compact myelin protein 2' 3' cyclic nucleotide 3' phosphodiesterase), 9-O-acetyl GD3 and
other gangliosides recognized by the A2B5 antibody, which are commonly expressed by high
motility neural cells. Lipid rafts were identified by the labeling of markers such as the GM1
ganglioside, flotillin and caveolin. OEC’s lipid rafts showed phosphatidylcholine,
sphingomyelin, high cholesterol, low protein content and a concentration of the above
mentioned gangliosides and CNPase. Furthermore, the destruction of lipid rafts by methyl-
beta-cyclodextrin alters the distribution of components here identified in OEC’s lipid
microdomains. These results suggest that the detergent resistant membranes of OEC may be
at least partly involved in relevant events of the migration regeneration promoted by this cell.

x
ABREVIAÇÕES E SIGLAS
BCA - ácido bicinchonínico
BDNF - fator neurotrófico derivado do cérebro
BO - bulbo olfatório
BOA - bulbo olfatório acessório
BOP - bulbo olfatório principal
BSA - albumina sérica bovina
Cav-1 - caveolina-1
Cav-2 - caveolina-2
Cav-3 - caveolina-3
CCGr - camada de células granulares
CCM - camada de células mitrais
cDNA – DNA complementar
CEMs - membranas enriquecidas de colesterol
CFO - camada de fibras olfatórias
Cgl - camada glomerular
CNPase - 2’ 3’ nucleotídeo cíclico 3’ fosfodiesterase
CPE - camada plexiforme externa
CPI - camada plexiforme interna
CT-B - toxina do cólera subunidade B
DAPI - 4’.6-diamidino-2-fenilindole-dilactate
DIGs - membranas enriquecidas de glicoesfingolipídios insolúveis a detergente
DRMs - membranas resistentes a detergentes
EDTA - ácido etilenodiaminotetracético
FGF - fator de crescimento de fibroblastos
FITC – isotiocianato de fluresceína
GDNF - fator de crescimento derivado de células gliais
GEMs - membranas enriquecidas de glicoesfingolipídios
GEO - glia embainhante olfatória
GFAP - proteína ácida do filamento glial
GM1- monossialogangliosídeo
GPI - glicosilfosfatidilinositol
HRP – peroxidase de rabanete

xi
HPTLC – cromatografia de camada fina de alta performace
KDa - kilo Dalton
M.E.T. - microscopia eletrônica de transmissão
MβCD - metil-beta-ciclodextrina
MO - mucosa olfatória
N-CAM - molécula de adesão celular neural
ND - não determinado
NGF - fator de crescimento de neurônios
NGS - soro normal de cabra
NROs - neurônios receptores olfatórios
OVN - órgão vomeronasal
OEC - olfactory ensheathing cell
PC - fosfatidilcolina
PBS- tampão fosfato com salina
PSA-NCAM - molécula de adesão celular neural polisiálica
p75-NGFR - receptor para fator de crescimento neuronal de baixa afinidade
RNAm - ácido ribonucléico mensageiro
SDS - lauril sulfato de sódio
SM - esfingomielina
SNC - sistema nervoso central
SNP - sistema nervoso periférico
SPFH - estomatina-proibitina-flotilinas-HflK/C
TBS - solução tris base
TRIS - tris-hidroximetil-aminometano
TRITON X-100 - t-octilfenoxipolietoxietanol
VIP21 – proteína de membrana vesicular integral

xii
LISTA DE ILUSTRAÇÕES
Figura 1- Camadas do bulbo olfatório de rato adulto......................................................................... 3
Figura 2- Representação do sistema olfatório ..................................................................................... 5
Figura 3- Etapas de regeneração axonal pela GEO ........................................................................... 7
Figura 4- Moléculas de glicolipídios ...................................................................................................11
Figura 5- Modelo esquemático do mosaico fluido das membranas celulares e seus componentes .............................................................................................................................................. 13
Figura 6- Esquema do modelo dos microdomínios de membrana ............................................... 15
Figura 7- Esquema da proteína caveolina-1 inserida na membrana plasmática ........................ 19
Figura 8- Esquema da proteína flotilina-1 inserida parcialmente na membrana plasmática .. 21
Figura 9- Estrutura do gangliosídeo GM1 ......................................................................................... 22
Figura 10- Representação da metil-β-ciclodextrina ......................................................................... 23
Figura 11- Microscopia de fluorescência da cultura de células isoladas da GEO apresentando imunorreatividade para a proteína S100 e CNPase ........................................................................... 36
Figura 12- Ilustração da obtenção do gradiente de sacarose ......................................................... 37
Figura 13- Densidade das frações do gradiente de sacarose da GEO .................................... 37
Figura 14- Distribuição do gangliosídeo GM1 nas frações do gradiente de sacarose ............. 39
Figura 15- Microscopia de fluorescência da GEO incubada na presença de toxina do cólera subunidade B conjugada à FITC a 4 e 37°C ...................................................................................... 40
Figura 16- Distribuição da proteína flotilina-1 nas frações do gradiente de sacarose ............. 41
Figura 17- Microscopia de fluorescência da GEO apresentando imunorreatividade para a proteína flotilina-1 ................................................................................................................................... 42
Figura 18- Distribuição da proteína caveolina-1 nas frações do gradiente de sacarose .......... 43
Figura 19- Microscopia de fluorescência da GEO apresentando imunorreatividade para a proteína caveolina-1 ................................................................................................................................ 44
Figura 20- Micrografias eletrônicas das frações do gradiente de sacarose ................................ 45
Figura 21- Concentração total de colesterol e proteínas nas frações do gradiente de sacarose da GEO .................................................................................................................................... 47
Figura 22- Caracterização lipídica das frações do gradiente de sacarose da GEO ................... 49

xiii
Figura 23- Distribuição da proteína CNPase nas frações do gradiente de sacarose ................. 51
Figura 24- Microscopia de fluorescência da GEO apresentando imunorreatividade para CNPase e flotilina-1 ............................................................................................................................. 52
Figura 25- Distribuição do gangliosídeo 9-O-acetil GD3 e dos gangliosídeos reconhecidos pelo anticorpo A2B5 (G-A2B5) nas frações do gradiente de sacarose ....................................... 54
Figura 26- Microscopia de fluorescência da GEO apresentando imunorreatividade para a proteína S100 e CNPase .......................................................................................................................... 57
Figura 27- Representação da obtenção dos gradientes de sacarose com MβCD e em condição controle ....................................................................................................................................................... 58
Figura 28- Distribuição dos marcadores de rafts ............................................................................. 59
Figura 29- Concentração total de colesterol e proteínas nas frações do gradiente de sacarose da GEO .................................................................................................................................... 60
Figura 30- Distribuição das moléculas candidatas no envolvimento da migração da GEO nas frações do gradiente de sacarose ....................................................................................................... 61
Figura 31- Microscopia de fluorescência da GEO apresentando imunorreatividade para CNPase e flotilina-1 ................................................................................................................................ 62
LISTA DE TABELAS
Tabela1- Anticorpos Primários .............................................................................................. 30
Tabela 2- Anticorpos Secundários …………...…………………...……………..…………. 31
Tabela 3 - Ligantes e Sondas …………………………………………………………..….. 31

xiv
SUMÁRIO
INTRODUÇÃO
1.1 Organização geral do sistema olfatório ............................................................................... 1
1.2 Anatomia do bulbo olfatório …........................................................................................... 2
1.2.1 Organização citoarquitetônica do bulbo olfatório principal ................................. 2
1.3 Glia embainhante olfatória (GEO) ...................................................................................... 4
1.4 Moléculas candidatas no envolvimento da migração da GEO ........................................... 8
1.4.1 2’ 3’ nucleotídeo cíclico 3’ fosfodiesterase – CNPase ......................................... 8
1.4.2 Gangliosídeos ..................................................................................................... 10
1.5 Visão geral da membrana plasmática ................................................................................ 12
1.6 Microdomínios lipídicos de membrana (lipid rafts) ......................................................... 14
1.7 Diversidade de componentes dos microdomínios lipídicos .............................................. 16
1.7.1 Cavéolas e Caveolinas ........................................................................................ 17
1.7.2 Flotilinas ............................................................................................................. 20
1.7.3 Gangliosídeo GM1 ............................................................................................. 22
1.8 Rafts desorganizadas: depleção de colesterol ................................................................... 23
1.9 A relevância do estudo .................................................................................................... 24
OBJETIVO
2.1 Objetivos geral................................................................................................................... 25
2.2 Objetivos específicos ........................................................................................................ 25
MATERIAIS E MÉTODOS
3.1 Animais ............................................................................................................................. 26
3.2 Cultura de células isoladas .................................................................................................26
3.3. Tratamento das células com metil-beta-ciclodextrina (MβCD) ........................................27
3.4 Isolamento das membranas resistentes a detergente da GEO ........................................... 27
3.5 Densidade das frações do gradiente de sacarose ............................................................... 28
3.6 Dot blotting das frações do gradiente de sacarose ............................................................ 28
3.7 Fluorescência da toxina do cólera subunidade B (CT-B) ................................................. 29
3.8 Imunocitoquímica ............................................................................................................. 29
3.9 Anticorpos e outros ligantes .............................................................................................. 30

xv
3.10 Microscopia Eletrônica de Transmissão (M.E.T.): Contrastação negativa ..................... 32
3.11 Dosagem de Proteína ...................................................................................................... 32
3.12- Análise Lipídica ............................................................................................................. 33
3.12.1 Extração de lipídios .......................................................................................... 33
3.12.2 Identificação dos lipídios ................................................................................. 33
3.13.3 Análise de Colesterol ....................................................................................... 34
RESULTADOS
4.1 Perfil do gradiente de sacarose .......................................................................................... 35
4.2 Detecção do gangliosídeo GM1 nas frações do gradiente de sacarose e em células
vivas......................................................................................................................................... 38
4.3 Detecção da proteína flotilina-1 nas frações do gradiente de sacarose e em células
permeabilizadas ....................................................................................................................... 41
4.4 Detecção da proteína caveolina-1 nas frações do gradiente de sacarose e em células
permeabilizadas ....................................................................................................................... 43
4.5 Morfologia das frações resistentes a detergente ............................................................... 45
4.6 Distribuição de proteína e colesterol nas frações do gradiente de sacarose ...................... 46
4.7 Análise dos lipídios totais das frações do gradiente de sacarose da GEO ........................ 48
4.8 Localização da proteína CNPase nas frações do gradiente de sacarose e em células
permeabilizadas ....................................................................................................................... 50
4.9 Localização do gangliosídeo 9-O-acetil GD3 e dos gangliosídeos reconhecidos pelo
anticorpo A2B5 (G-A2B5) nas frações do gradiente de sacarose .......................................... 53
4.10 Desorganização das lipid rafts ........................................................................................ 55
DISCUSSÃO
5.1 Diversas metodologias são usadas para o isolamento das rafts ........................................ 63
5.2 Membranas resistentes a detergentes da GEO são positivas para marcadores das lipid
rafts.......................................................................................................................................... 64
5.3 Perfil lipídico e dosagem protéica da membrana da GEO confirmam a presença de lipid
rafts ......................................................................................................................................... 65
5.4 As lipid rafts isoladas formam espontaneamente vesículas .............................................. 66
5.5 Moléculas candidatas no envolvimento da migração se associam as rafts da GEO ......... 67
5.6 A depleção de colesterol altera a distribuição de componentes protéicos e lipídicos das
rafts da GEO ........................................................................................................................... 68

xvi
5.7 Considerações finais …………..............…………………………...........……………… 69
CONCLUSÕES ..................................................................................................................... 71
PERSPECTIVAS………………………………………........…...…………........………… 72
REFERÊNCIAS..................................................................................................................... 73

1
1 Introdução
1.1 Organização geral do sistema olfatório
O olfato é um sentido que permite as espécies animais de receberem e processarem as
informações do ambiente, transmitidas por substâncias químicas. O sistema olfatório é
composto pelo epitélio olfatório, nervo olfatório e estruturas centrais como o bulbo olfatório
(Schwob, 2002).
O epitélio olfatório abrange os neurônios receptores olfatórios (NROs), células
sustentaculares, células basais (globosas e horizontais) e as glândulas de Bowman (Schwob,
2002). Os neurônios receptores olfatórios representam o principal componente do epitélio
olfatório, uma vez que são responsáveis pela recepção e transdução da informação olfatória. O
NRO apresenta morfologia bipolar, com axônio amielínico e de pequeno diâmetro (0,1-0,4
mm). Sabe-se que esses neurônios periféricos exibem contínua renovação a partir de células
basais precursoras, ao longo da vida dos mamíferos (Graziadei & Monti-Graziadei, 1979;
Schwob, 2002).
Os NROs emitem dendritos apicais, voltado para a cavidade nasal, e um axônio basal
que segue amielínico através da lâmina própria. Estes axônios são então organizados em
feixes por um tipo especial de glia, a glia embainhante olfatória (GEO) e a coalizão de feixes,
por vez, forma o primeiro par de nervos cranianos ou nervo olfatório. Em seguida, os axônios
olfatórios atravessam a placa cribiforme, entram no sistema nervoso central (SNC),
dispersam-se na camada de fibras olfatórias do bulbo olfatório (BO) e rumam para a camada
glomerular do BO, onde estabelecem sinapses com outros tipos neuronais (Henion &
Schwarting, 2007).
O bulbo olfatório recebe uma grande variedade de informações provenientes de
moléculas odoríferas, captadas pelos NROs na cavidade nasal e as redirecionam para o córtex
piriforme, onde os odores são processados (Johnson et al., 2000).

2
1.2 Anatomia do bulbo olfatório
O BO é uma estrutura telencefálica laminada, situada na face anterior do lobo frontal,
imediatamente acima da placa crivosa do osso etmóide. O BO de animais macrosmáticos
pode ser dividido em duas regiões: bulbo olfatório principal (BOP) e bulbo olfatório acessório
(BOA), localizado em uma posição caudo-dorsal em relação ao BOP (Halpern & Martínez-
Marcos, 2003). Além disso, essas regiões são funcionalmente separadas e embora o BOA
apresente uma organização laminar semelhante à do BOP, este é formado por camadas mais
delgadas e menos distintas.
1.2.1 Organização citoarquitetônica do bulbo olfatório principal
A estrutura histológica do BOP consiste em seis camadas distintas (da camada mais
externa para interna): camada de fibras olfatórias, camada glomerular, camada plexiforme
externa, camada de células mitrais, camada plexiforme interna e camada de células granulares
(Meisami & Bhatnagar, 1998) (Figura 1).
A camada de fibras olfatórias, camada mais externa do BO, marca a transição entre o
sistema nervoso periférico (SNP) e o SNC, onde os fascículos dos axônios olfatórios
penetram no BO para inervar neurônios de segunda ordem. Esta camada também é composta
por astrócitos e pela glia embainhante olfatória (GEO).
A camada glomerular contém, além dos glomérulos, os corpos celulares das células
tufosas externas e pequenos interneurônios, denominados neurônios periglomerulares
(Doetsch & Alvarez-Buylla, 1996). Os glomérulos são formados por neurópilo, circundado
por neurônios, oligodendrócitos, astrócitos e, possivelmente, por glia embainhante olfatória
(Valverde & Lopez-Mascareque, 1991).

3
A camada plexiforme externa é a camada relativamente espessa de neurópilo, no qual
predominam dendritos de células mitrais, células tufosas e células granulares. Contém corpos
celulares esparsos de células tufosas (Lledo et al., 2004).
A camada de células mitrais forma uma lamina, delgada e compacta, contendo os
corpos celulares das células mitrais (Margrie et al., 2001).
A camada plexiforme interna é uma camada fina com baixa densidade celular, mas
com axônios colaterais de células mitrais e outras fontes não identificadas e dendritos das
células granulares.
A camada de células granulares, camada mais interna do BO, é composta por uma
grande quantidade de neurônios granulares desprovidos de axônios.
Figura 1- Camadas do bulbo olfatório de rato adulto. Coloração do núcleo por DAPI mostrando as seis camadas (da mais externa para mais interna) do bulbo olfatório de mamífero: camada de fibras olfatórias – CFO, camada glomerular – Cgl, camada plexiforme externa – CPE, camada de células mitrais – CCM, camada plexiforme interna – CPI e camada de células granulares – CCGr. Barra de calibração: 40µm.

4
1.3 Glia embainhante olfatória (GEO)
A GEO foi originalmente descrita como células estreladas ou fusiformes presentes
entre as fibras do BO próximas à pia máter (Ramón-Cueto & Nieto-Sampedro, 1994; revisto
por Levine & Marcillo, 2008). Embora mais tarde, estudos se refiram a essas células como
“glia de Blañes” ou “célula de Schwann olfatória”, a nomeclatura atualmente empregada é a
de glia embainhante olfatória, proposta por Doucette (1984).
A GEO origina-se de células progenitoras gliais que emigram do placódio olfatório
(Marin-Padilla & Amieva, 1989). Esse tipo celular é encontrado apenas no eixo olfatório,
mais especificamente na lâmina própria da mucosa olfatória, no nervo olfatório, na camada de
fibras olfatórias e possivelmente na camada glomerular (Valverde et al., 1992) do BO, sendo
portanto, a única glia capaz de atravessar o limite entre os sistemas nervosos periférico e
central (Doucette, 1984; Au & Roskams, 2003) (Figura 2). No SNP, a GEO acompanha
axônios de neurônios olfatórios, emitindo prolongamentos que embainham grupos destes
axônios organizando-os como fascículos, até sua entrada no bulbo olfatório.
No eixo olfatório é observada uma contínua reposição de neurônios receptores, bem
como a reorganização de sinapses ao longo da vida e embora todos os determinantes destes
eventos não sejam conhecidos, a GEO é apontada como um dos elementos chaves deste
processo. A GEO é capaz de produzir vários fatores de crescimento como fator neurotrófico
derivado do cérebro (BDNF), fator de crescimento de fibroblastos (FGF), fator de
crescimento de neurônios (NGF), fator de crescimento derivado de células gliais (GDNF),
neurotrofina 4/5, neuregulinas (Ramón-Cueto e Valverde, 1995; revisto por Ramón-Cueto e
Ávila, 1998; Kaftz e Greer, 1999; Chuah et al., 2000; Woodhall et al., 2001) e moléculas de
adesão celular como N-CAM (molécula de adesão celular neural), PSA-NCAM, laminina,
fibronectina e L1 (Miragall et al., 1988; Franceschini & Barnett, 1996). Estes fatores
possivelmente participam do crescimento fisiológico de novos axônios olfatórios primários.

5
Além de moléculas promotoras de crescimento, a GEO pode proporcionar uma barreira entre
fascículos de axônios olfatórios e respectivas moléculas quimiorepelentes (Schwarting et al.,
2000).
Figura 2- Representação do sistema olfatório. (A) Células basais (em marrom) dão origem a neurônios receptores olfatórios (em azul). A glia embainhante olfatória (em verde) embainha os axônios olfatórios ao longo de seu trajeto da membrana basal do epitélio olfatório até a camada glomerular do bulbo olfatório. (B) Seção sagital da cabeça de rato, mostrando o sistema olfatório. Bulbo olfatório acessório - BOA; bulbo olfatório - BO; mucosa olfatória –MO. Modificado e adaptado de Firestein, 2001 e Thuret et al., 2006.

6
Inicialmente, a idéia de que culturas de GEO apresentam diferentes tipos de células foi
sugerida por Ramón-Cueto e colaboradores (Pixley, 1992; Ramón-Cueto & Nieto-Sampedro,
1992; Ramón-Cueto et al., 1993), que relatou através de ensaios imunocitoquímicos a
presença de três morfologias celulares distintas nas culturas de GEO do BO de ratos adultos:
(1) “Schwann cell-like OECs” ou GEO similar à célula de Schwann fusiforme com
prolongamentos finos; (2) “astrocyte-like OECs” ou GEO semelhante a astrócitos com
morfologia aplanada com número variável de prolongamentos achatados que lembram
astrócitos tipo-1 e (3) “macrophage like cells” ou células similares a macrófagos, nem sempre
detectável in vitro. O primeiro grupo de GEO apresenta características morfológicas
semelhantes às das células de Schwann não-mielinizante do SNP e, tal como estas, estão
ligadas à regeneração axonal. Esse perfil morfológico expressa a proteína ligadora de cálcio
associada ao citoesqueleto S100, a molécula de adesão L1, o receptor para fator de
crescimento neuronal de baixa afinidade (p75-NGFR) e marcação fraca para proteína ácida do
filamento glial (GFAP) (Ramón-Cueto & Nieto-Sampedro, 1992). O segundo grupo
(semelhantes a astrócitos) apresenta forte expressão das proteínas GFAP, S100 e da molécula
de adesão celular neural polisiálica (PSA-NCAM) (Franceschini & Barnett, 1996).
Nos últimos anos, o interesse na GEO tem aumentado em estudos de transplante
celular para regeneração do SNC. Estes estudos revelaram um potencial regenerativo da GEO
ao proporcionar um ambiente favorável para o crescimento axonal nas áreas de lesão, o que
frequentemente resulta na regeneração axonal e em casos de recuperação funcional dos
animais (Franklin & Barnett, 1996; Rámon-Cueto at al., 2000; Li et al., 2003; Santos-Benito
& Ramón-Cueto, 2003; Keyvan-Fouladi et al., 2005; revisto por Franssen et al., 2007)
(Figura 3).
Embora a GEO represente uma potente ferramenta terapêutica, alguns autores relatam
falhas no processo de regeneração. O motivo desta falha não pode ser determinado, mas

7
coincidentemente foi observada uma concomitante interrupção na migração da GEO (Li et al.,
2003). Desta forma, a determinação das estruturas celulares e vias de sinalização envolvidas
na migração da GEO constitui um dos pontos centrais para o entendimento da capacidade e
limitações desta célula na promoção da regeneração axonal, bem como para seu uso
terapêutico.
Figura 3- Etapas de regeneração axonal pela GEO. Efeito regenerativo da GEO transplantada (em vermelho) acompanhada de fibroblastos (em verde) sobre axônios corticoespinhais lesionados (em cinza) de rato. (A) Durante a primeira semana, os axônios avançam sob aposição da GEO e fibroblastos. (B) Entre a terceira e a quarta semana, os axônios prosseguem através da lesão, envoltos pela GEO, formando uma ponte entre os domínios de mielina central produzidas por oligodendrócitos (em azul). Modificado de Li et al., 2003.
A
B

8
1.4 Moléculas candidatas no envolvimento da migração da GEO
1.4.1 2’ 3’ nucleotídeo cíclico 3’ fosfodiesterase - CNPase
Inicialmente, a 2’ 3’ nucleotídeo cíclico 3’ fosfodiesterase (CNPase) foi caracterizada
no baço e no pâncreas bovino (Whitfeld et al., 1955). Atualmente, sabe-se que a CNPase é
expressa em níveis mais baixos fora do SNC e foi verificada sua associação com proteínas do
citoesqueleto, mitocôndria e mielina (Bifulco et al., 2002; Lee et al., 2005, 2006).
Diversos estudos demonstraram que a CNPase (pertencente à super-família das 2H
fosfoesterases) tem capacidade de hidrolizar ligações fosfodiester em oligonucleotídeos,
nucleotídeos cíclicos e 2’, 3’ NADP cíclico. Entretanto, pela aparente ausência de
nucleotídeos 2’, 3’ (substrato) em tecidos animais, a atividade enzimática desta proteína tem
sido questionada (Tsukada & Kurihara, 1992).
Duas isoformas, CNPase I (46 kDa) e CNPase II (48 kDa) são codificadas pelo único
gene de CNPase, distinguindo em apenas 20 aminoácidos no domínio N-terminal (Tsukada &
Kurihara, 1992). Estas duas isoformas moleculares são resultantes de splicing alternativo.
Esses dados sugerem que o ácido ribonucléico mensageiro (RNAm) que codifica a CNPase I
pode ser expresso principalmente no SNC, enquanto que o RNAm que codifica a CNPase II
pode ser expresso em diferentes tecidos (Scherer et al., 1994).
A CNPase é reconhecida como marcadora precoce de oligodendrócitos, marcadora de
células de Schwann (revisto por Sprinkle, 1989) e GEO adultas (Santos-Silva & Cavalcante,
2001). A CNPase está aparentemente envolvida nas etapas iniciais de embainhamento
anteriores à compactação da mielina (Reynolds & Wilkin, 1988; Yoshino et al., 1989; Amur-
Urmarjee et al., 1990; Barradas et al., 1995).
Em oligodendrócitos, a CNPase aparece associada à membrana e mantém fortes
relações com microtúbulos e microfilamentos, distribuindo-se nas expansões de membrana
destas células, num padrão de nervuras e prolongamentos intercomunicantes mais finos, co-

9
localizada com tubulina e actina (Dyer & Benjamins, 1989; Reynolds et al., 1989). Desse
modo, os autores descrevem a CNPase como uma proteína membrano-esqueletal. Além disso,
diversos estudos sugerem um papel da CNPase na migração e/ou na expansão da membrana
celular durante a mielinização (De Angelis & Braun, 1996).
Além da associação com o citoesqueleto, a presença da CNPase em oligodendrócitos
também foi detectada em lipid rafts, microdomínios enriquecidos em colesterol e
esfingolipídios (Kim & Pfeiffer, 1999; Taylor et al., 2002; Hinman et al., 2008). Sabe-se que
as plataformas lipídicas estão envolvidas no controle do tráfego de proteínas pela célula e
modulam cascatas de sinalização intracelular (Simons & Toomre, 2000), como será detalhado
adiante. Desta forma, é possível que a CNPase esteja envolvida em vias de sinalização
membrano-esqueletais.
Apesar de evidências experimentais, a função biológica da CNPase ainda não é clara.
Lappe-Siefke e colaboradores (2003) demonstraram, por exemplo, que ratos transgênicos
incapazes de expressar CNPase apresentam redução na espessura do corpo caloso,
degeneração axonal, gliose reativa, diminuição do desempenho motor e aumento na
mortalidade. Por outro lado, a superexpressão de CNPase induz a formações aberrante de
membranas de mielina, evidenciando que a CNPase desempenhe um importante papel na
compactação de mielina (Yin et al., 1997).

10
1.4.2 Gangliosídeos
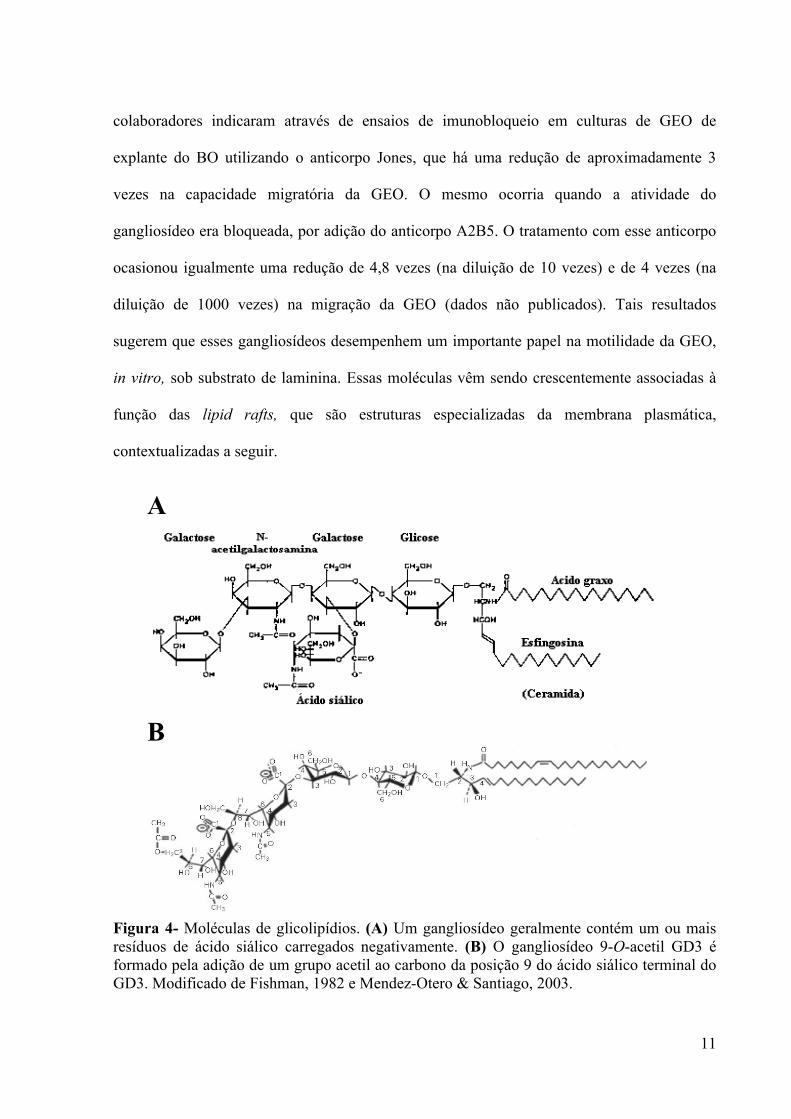
Os gangliosídeos compreendem um grande grupo de glicoesfingolipídios sialilados,
isto é, glicolipídios formados por uma cadeia de ácido graxo e uma cadeia de esfingosina
ligada ao ácido siálico (Wiegandt, 1985) (Figura 4A). Esses componentes são importantes na
estrutura das membranas biológicas, devido ao seu caráter anfipático e são amplamente
expressos em tecidos de mamíferos, particularmente no SNC (Loyd & Furukawa, 1998), onde
representam 5 a 10% da massa total de lipídios. Os gangliosídeos participam de várias
funções importantes, entre elas: reconhecimento, adesão e sinalização celular (Hakomori,
1990, Yu et al., 2004).
Dentre os gangliosídeos, pode-se citar o gangliosídeo reconhecido pelo anticorpo
monoclonal Jones, o 9-O-acetil GD3 (Constantine-Paton et al., 1986). Este gangliosídeo está
presente no sistema nervoso central e periférico, sendo particularmente abundante durante o
desenvolvimento (Mendez-Otero et al., 1988), e em algumas regiões específicas no adulto,
onde há migração celular e/ou crescimento neurítico. O 9-O-acetil GD3 é também expresso
em culturas de neurônios e precursores de oligodendrócitos (Farrer & Quarles, 1999; Santiago
et al., 2001). O 9-O-acetil GD3 é um derivado do gangliosídeo GD3 em que um grupo acetil é
anexado por uma ligação éster na posição 9 do resíduo de ácido siálico terminal (Ren et al.,
1992) (Figura 4B). A esfingosina do 9-O-acetil GD3 é uma lactosilceramida, que se insere na
membrana plasmática. Por sua vez, situadas na face externa da membrana plasmática,
encontram-se as duas moléculas de ácido siálico presas a lactosilceramida.
O anticorpo monoclonal A2B5 reconhece uma variedade de gangliosídeos expressos
por precursores oligodendrogliais de elevada motilidade, como o gangliosídeo GT3 e o 9-O-
acetil GT3 na superfície das células neuronais e gliais (Dubois et al., 1990), bem como o 9-O-
acetil GD3 (Farrer & Quarles, 1999).
Dados preliminares do grupo envolvido nesse estudo, realizados por Santos-Silva e
11
colaboradores indicaram através de ensaios de imunobloqueio em culturas de GEO de
explante do BO utilizando o anticorpo Jones, que há uma redução de aproximadamente 3
vezes na capacidade migratória da GEO. O mesmo ocorria quando a atividade do
gangliosídeo era bloqueada, por adição do anticorpo A2B5. O tratamento com esse anticorpo
ocasionou igualmente uma redução de 4,8 vezes (na diluição de 10 vezes) e de 4 vezes (na
diluição de 1000 vezes) na migração da GEO (dados não publicados). Tais resultados
sugerem que esses gangliosídeos desempenhem um importante papel na motilidade da GEO,
in vitro, sob substrato de laminina. Essas moléculas vêm sendo crescentemente associadas à
função das lipid rafts, que são estruturas especializadas da membrana plasmática,
contextualizadas a seguir.
Figura 4- Moléculas de glicolipídios. (A) Um gangliosídeo geralmente contém um ou mais resíduos de ácido siálico carregados negativamente. (B) O gangliosídeo 9-O-acetil GD3 é formado pela adição de um grupo acetil ao carbono da posição 9 do ácido siálico terminal do GD3. Modificado de Fishman, 1982 e Mendez-Otero & Santiago, 2003.
A
B

12
1.5 Visão geral da membrana plasmática
A membrana plasmática corresponde à interface por meio da qual a célula interage
com seu ambiente e células vizinhas. Esta estrutura é composta principalmente de proteínas,
lipídios e eventuais glícidos associados (Unwin et al., 1984).
As proteínas associadas à membrana, podem ser classificadas em: proteínas periféricas
ou extrínsecas (localizadas à superfície da membrana) e proteínas integrais ou intrínsecas
(localizadas entre os fosfolipídios, podendo atravessar completamente a bicamada). Além
disso, as proteínas podem também estar associadas a glicídios na monocamada externa, sendo
denominadas, dessa forma, glicoproteínas. Sabe-se que as proteínas e glicoproteínas
promovem importantes eventos celulares como o transporte de íons, armazenamento de
energia, migração e transdução de sinais (Zheng et al., 2009).
Os três principais grupos de lipídios da membrana são: os fosfolipídios, o colesterol e
os glicolipídios (Figura 5B), que podem variar em presença e quantidade, em função do tipo
celular ou especializações membranares de uma mesma célula. Os fosfolipídios são
compostos por um ácido fosfórico, uma molécula de glicerol esterificado e duas cadeias
hidrofóbicas de ácidos graxos. Este componente representa o principal componente lipídico
da membrana plasmática, conferindo propriedades de permeabilidade seletiva e organização
em bicamada da membrana celular e organelas. O colesterol é outro lipídio constituinte da
membrana plasmática, responsável pela fluidez e formação de microdomínios lipídicos
(Alberts et al., 2002). Por vez, os glicolipídios são moléculas de lipídios associados a glicídios,
localizados na monocamada externa da bicamada lipídica, cuja função está em grande parte
associada a sítios receptores específicos da membrana celular (Alberts et al., 2002).
O clássico modelo do mosaico fluido de Singer e Nicholson (1972) sugere que as
proteínas e os lipídios difundem livremente sob a superfície bidimensional da célula, sendo as
proteínas incorporadas randomicamente na bicamada de fosfolipídios (Figura 5A).

13
Atualmente sabe-se que a estrutura física da bicamada pode ser profundamente alterada pela
concentração localizada de componentes como colesterol e esfingolipídios, um tipo de
glicolipídio. O colesterol naturalmente se insere na região hidrofóbica da bicamada lipídica,
reduzindo a permeabilidade membranar e inibindo possíveis transições de fase (Alberts et al.,
2002). Já os esfingolipídios possuem cadeias de ácidos graxos saturadas e mais longas do que
a maioria dos lipídios membranares, o que proporciona uma maior interação entre estes,
reduzindo, por vez, a difusão lateral de componentes membranares. A concentração destes
componentes pode eventualmente alterar o estado de fluidez da membrana, que passaria de
líquido desordenado, descrito no modelo tradicional do mosaico fluido, para um estado de
líquido ordenando (revisto por Munro et al., 2003).
Figura 5- Modelo esquemático do mosaico fluido das membranas celulares e seus componentes. (A) A membrana celular é uma estrutura complexa composta principalmente por proteínas e lipídios. (B) Os lipídios são separados em: fosfolipídios, glicolipídios e esterol. A quantidade relativa desses componentes varia de célula para célula. Modificado de Pietzsch, 2004 e Fantini et al., 2002.
A B

14
1.6 Microdomínios lipídicos de membrana (lipid rafts)
Em 1988, Simons e van Meer revelaram uma nova característica da membrana
plasmática quando propuseram a existência de microdomínios ordenados em células epiteliais
polarizadas. Estudos seguintes demonstraram que estes microdomínios lipídicos, designados
lipid rafts, correspondiam a uma separação de fase temporária na bicamada lipídica fluida
onde se concentravam grandes quantidades de esfingolipídios e colesterol (Simons & Ehehalt,
2002; Pike, 2006) (Figura 6). Dessa forma, a presença de microdomínios líquidos ordenados
nas células transformou o modelo do mosaico fluido em um sistema mais complexo. Em
outras palavras, as lipid rafts formam distintas fases líquidas ordenadas na bicamada lipídica,
inseridas em uma matriz líquida desordenada de lipídios insaturados (Schroeder et al., 1994;
revisto por Brown & London, 1998).
As rafts representam áreas especializadas de pequeno diâmetro (de 10 a 200 nm) das
membranas eucarióticas as quais são caracterizadas por uma composição química e
propriedades físicas únicas (Simons & Ikonen, 1997; Pike, 2006). Desta forma, as lipid rafts
criam um microambiente físico e bioquímico adequado para o funcionamento e recrutamento
por afinidade de diversas proteínas. Através da agregação dos diferentes lipídios e proteínas
que a constituem, as rafts medeiam muitos processos biológicos importantes, incluindo vias
de transdução de sinais, apoptose, adesão/migração celular, organização do citoesqueleto,
bem como eventos de endocitose e exocitose (revisto por Simons & Toomre, 2000; revisto
por Harris & Siu, 2002; Prag et al., 2007). Esses últimos eventos, em especial, são também
utilizados para a invasão e propagação de patógenos virais (Bavari et al., 2002) e bacterianos
(Fivaz et al., 2000).
A composição das rafts é melhor caracterizada em sua monocamada externa (revisto
por Parton & Richards, 2003). Estudos recentes sugerem que a monocamada interna tenha
uma organização equivalente à externa, sendo, portanto, rico em fosfolípidios com ácidos

15
graxos saturados e colesterol. A indeterminação persistente até o momento de propriedades
fundamentais das rafts é em grande parte resultado da dificuldade metodológica da
observação destes microdomínios em células vivas.
Figura 6- Esquema do modelo dos microdomínios de membrana. A lipid raft é uma região da membrana plasmática enriquecida de colesterol, glicoproteínas, glicolipídios, proteínas transmembranares e proteínas ancoradas por glicosilfosfatidilinositol (GPI). Modificado de Alberts et al., 2002.
LÚMEN
CITOSOL
LIPID RAFT

16
1.7 Diversidade de componentes dos microdomínios lipídicos
Na literatura encontramos inúmeros métodos para o isolamento das lipid rafts.
Entretanto, trabalhos realizados utilizando diferentes metodologias bioquímicas de extração
das rafts resultaram em dados heterogêneos quanto a concentrações de componentes e
densidades das rafts (Radeva & Sharom, 2004). Desse modo, a lipid raft é relatada como uma
estrutura dinâmica. Sob algumas circunstâncias, parece ser um domínio enriquecido de
colesterol e glicoesfingolipídios e sob outras, parece ser uma estrutura mais diversificada de
lipídios.
Dependendo da distribuição bioquímica, as rafts podem receber diversas nomeclaturas,
como: membranas enriquecidas de colesterol (cholesterol-enriched membranes, ou CEMs),
membranas enriquecidas de glicoesfingolipídios (glycosphingolipid-enriched membranes, ou
GEMs), membranas enriquecidas de glicoesfingolipídios insolúveis a detergente (detergent-
insoluble, glycosphingolipid-enriched membranes, ou DIGs) e membranas resistentes a
detergentes (detergent-resistant membranes, ou DRMs). Os dois últimos termos refletem a
observação de que esses domínios não são solubilizados em detergentes não-iônicos (revisto
por Pike, 2004).
A extração com 1% de Triton X-100 a 4°C consiste na ferramenta bioquímica
tradicional usada para estudar estas plataformas lipídicas. Análises desses microdomínios a
partir dessa preparação têm revelado uma composição similar entre diferentes tipos celulares.
Essa composição abrange basicamente lipídios como o colesterol (de 35-50% do total das
rafts); esfingomielina (de 10-15%); glicoesfingolipídios, como o gangliosídeo GM1 (de 10-
20%); glicerofosfolipídios, incluindo os maiores fosfolipídios da membrana, fosfatidicolina e
fosfatidiletanolamina (menos de 30%) (revisto por Pike, 2004).
Embora grande parte das proteínas membranares encontrem-se dispersas, algumas se
inserem preferencialmente em rafts. Essas proteínas atuam como importantes moduladores da

17
estrutura, estabilidade e funções desses microdomínios (revisto por Bauer & Pelkmans, 2006).
Exemplos típicos incluem proteínas ancoradas por glicosilfosfatidilinositol (GPI) (revisto por
Chatterjee & Mayor, 2001); cinases da família Src (Lck, Fyn e Lyn), que são ancoradas na
monocamada externa (revisto por Simons & Toomre, 2000); proteínas aciladas via S-
palmitoilação ou N-miristoilação como flotilinas (Rajendran et al., 2003) e proteínas
associadas a colesterol, como caveolinas (Kurzchalia & Parton, 1999).
Entre as proteínas e lipídios residentes das lipid rafts, daremos destaque para as
proteínas flotilina e caveolina, bem como para o gangliosídeo GM1, co-expressos em
diferentes tipos celulares de mamíferos e, portanto, utilizados como biomarcadores
convencionais destes microdomínios.
1.7.1 Cavéolas e Caveolinas
Uma das maiores subclasses das lipid rafts é a cavéola, uma estrutura identificável
morfologicamente (Kurzchalia & Parton, 1999), que contém um enriquecimento significativo
da proteína caveolina. As cavéolas quando observadas pela microscopia eletrônica mostram-
se como pequenas invaginações na membrana plasmática da célula de 50 a 100 nm de
diâmetro, não revestidas por moléculas de clatrina (Yamada, 1955; Anderson, 1998; Shin &
Abraham, 2001).
As cavéolas podem desempenhar funções de endocitose, transporte de colesterol e
sinalização. Dados da literatura sugerem a proteína integrante das cavéolas, caveolina
(ligadora de colesterol), seja parte integrante do maquinário de transporte do colesterol celular.
Uma vez complexada com o colesterol, a caveolina forma a estrutura de grampo (hairpin
loop) típica dos microdomínios caveolares, o que propicia os eventos de endocitose
característicos destas estruturas (Figura 7). A afinidade da caveolina pelo colesterol pode

18
variar de acordo com seu estado de polimerização, tendo sua forma monomérica ou em
oligômeros uma afinidade menor do que a forma polimerizada (Murata et al., 1995).
A família da caveolina é composta pelas proteínas caveolina-1, caveolina-2 e
caveolina-3 (Scherer et al., 1996, 1997; Couet et al., 1997). Todas as isoformas da caveolina
são integrais, apesar disso, não atravessam a membrana plasmática. Tanto a região carboxi,
quanto a região amino terminal ficam voltadas para o lado citosólico. Embora mostrem
semelhanças em estrutura e função, todas as caveolinas diferem em propriedades específicas e
na distribuição nos tecidos (Way & Parton, 1995; Scherer et al., 1996; Tang et al., 1996).
Além do colesterol, estas proteínas podem se ligar diretamente a glicoesfingolipídios e
moléculas sinalizadoras modificadas por lipídios (Li et al., 1995; Murata et al., 1995).
O primeiro membro da família caveolina a ser identificado foi a caveolina-1 (21 kDa),
também conhecida como VIP21 (vesicular integral membrane protein) ou Cav-1. Esta
proteína estrutural das cavéolas é a principal componente das vesículas derivadas da rede
trans-Golgi (Kurzchalia at al., 1992). A caveolina-1 (Figura 7) pode também participar de
diversas vias de sinalização ao ser fosforilada em resíduos de tirosina por membros da família
Src de cinases (Corley et al., 2001). Além disso, a caveolina-1 apresenta um domínio
conservado de 20 aminoácidos denominado de caveolin-scaffolding domain, que corresponde
a uma sub-região do domínio de oligomerização implicado na interação funcional da proteína
com moléculas de sinalização (Shin & Abraham, 2001; Williams & Lisanti, 2004).
A caveolina-2 (Cav-2) e a caveolina-3 (Cav-3) foram identificadas em 1996 usando
diferentes métodos experimentais. A Cav-2 foi descoberta pelo microsequenciamento de uma
proteína de 20 kDa que foi co-purificada de membranas caveolares derivadas de adipócitos
(Scherer et al., 1996). Outras caracterizações revelaram que a Cav-2 colocaliza com a Cav-1,
formando hetero-oligômeros em cavéola e é co-expressada em muitas células e tecidos, e
exige a presença da Cav-1 para a correta localização na membrana (Scherer et al., 1997;

19
Parolini et al., 1999). Cav-3, também conhecida como M-caveolina foi identificada através de
dados de pesquisas tradicionais com a biblioteca de cDNA numa tentativa de encontrar Cav-1
homólogas (Way & Parton, 1995; Tang et al., 1996). Esta proteína é encontrada de forma
específica no tecido muscular e apresenta peso molecular de 18 kDa.
Figura 7- Esquema da proteína caveolina-1 inserida na membrana plasmática. (A) A proteína caveolina, quando complexada com o colesterol forma a estrutura de um grampo de cabelo(hairpin loop), acredita-se para induzir a curvatura da bicamada lipídica para formar a cavéola. Observa-se os dois monômeros de caveolina-1, formando um dímero e que o domínio carbox e amino terminais estão direcionados para o citoplasma (B) Seqüência primária da Cav-1, evidenciando os seus domínios. O grupo palmitoil, auxilia na inserção na membrana plasmática. Modificado de Williams & Lisanti, 2004.

20
1.7.2 Flotilinas
Uma segunda família de proteínas, a família da flotilina, pode também ter um papel
estrutural na formação das lipid rafts, colaborando com a estabilização destes microdomínios.
Duas isoformas da flotilina tem sido caracterizadas, a flotilina-1 (45 kDa), também conhecida
como Reggie-2 e a flotilina-2 (42 kDa), também chamada de Reggie-1 (Bickel et al., 1997;
Volonte et al., 1999).
As flotilinas são proteínas pertencentes à superfamília SPFH (Stomatins, Prohibitins,
Flotillins, HflK/C), sendo altamente conservadas evolutivamente e com propensão à
oligomerização (Babuke & Tikkanen, 2007; revisto por Browman et al., 2007; Solis et al.,
2007). Esta proteína encontra-se associada à monocamada interna da membrana celular, na
qual se ancora por meio de acetilação (revisto por Langhorst et al., 2005). A interação entre a
flotilina e a monocamada interna ocorre em diferentes pontos hidrofóbicas de sua estrutura
alongada, o que colabora para a referida estabilização das rafts mediada por esta proteína. Os
2 tipos de flotilina podem valer-se de diferentes mecanismos de acetilações para seu
ancoramento na membrana, sendo a flotilina-1 ancorada por palmitoilização (Figura 8) e a
flotilina-2 por miristoilização, além de palmitoilização.
Embora se saiba que caveolinas e flotilinas possam interagir através de hetero-
oligomerização in vitro (Lisanti et al., 1993; Scherer et al., 1997; Volonte et al., 1999), o
mecanismo pelo qual estas proteínas facilitam a formação de vesículas ainda não foi
caracterizado. A flotilina-1 tem sido implicada na organização caveolar juntamente com
caveolina (Bickel et al., 1997; Volonte et al., 1999). Embora existam lipid rafts não
caveolares, toda cavéola se forma a partir de um arcabouço inicial destes microdomínios
lipídicos, o que é fundamental para a correta inserção da caveolina na membrana celular
(revisto por Ikonen, 2001).
Estudos recentes sugerem funções alternativas para flotilina, frequentemente utilizada

21
como proteína marcadora de rafts (Lang et al., 1998; Hazarika et al., 1999). Por fim, as
flotilinas também ligam ao colesterol, interagem com moléculas sinalizadoras através de
proteínas ancoradas por GPI (Anderson, 1998), regulam a reorganização do citoesqueleto de
actina, adesão celular dependente de actina e migração celular (Vassilieva et al., 2009).
Outros estudos, porém, têm implicado a flotilina-2 na formação de filopódios em células
neuronais (Lang et al., 1998; Hazarika et al., 1999).
Figura 8- Esquema da proteína flotilina-1 inserida parcialmente na membrana plasmática. A associação da flotilina-1 na membrana é mediada por duas regiões hidrofóbicas no domínio SPFH e um resíduo palmitato. Azul: domínio SPFH; marrom: domínio hidrofóbico; vermelho: sítio de palmitoilação; verde: domínio flotilina. Modificado de Browman et al., 2007.

22
1.7.3 Gangliosídeo GM1
O GM1 é um esfingolipídio presente na monocamda externa da membrana plasmática
e enriquecido em lipid rafts (Peterson, 2005) (Figura 9). Sabe-se que este gangliosídeo é
parte integrante do maquinário responsável pela endocitose da toxina do cólera. Esta toxina é
liberada pela bactéria Vibrio cholera e é formada por uma subunidade A (catalítica) e 5
subunidades B, responsáveis pela ligação ao gangliosídeo GM1 e consequente entrada na
célula-alvo (revisto por Fantini et al., 2002). A subunidade B reconhece o motivo
pentassacarídeo deste gangliosídeo (revisto por Fantini et al., 2002). Devido à sua afinidade
pelo GM1, a toxina do cólera, vêm sendo frequentemente utilizado como ferramenta de
identificação de lipid rafts que são, como mencionado, enriquecidas neste ganglisídeo.
Figura 9- Estrutura do gangliosídeo GM1. O esquema demonstra a inserção da esfingomielina, fosfolipídios saturados, colesterol e GM1 nas rafts. Por vez, na direita, há a representação bioquímica desse último componente lipídico. Modificado de Peterson, 2005.

23
1.8 Rafts desorganizadas: depleção de colesterol
Uma das formas de desfazer/desorganizar os microdomínios lipídicos de membrana é
através do uso da metil-β-ciclodextrina (MβCD). Essa droga depleta o colesterol da
membrana plasmática, desestruturando formações de lipid rafts do tipo caveolar ou não e,
consequentemente, bloqueando processos biológicos dependentes dessas regiões, o que
constitui uma importante ferramenta bioquímica para o estudo de funcionalidade desses
microdomínios lipídicos (Ilangumaran & Hoessli, 1998). A MβCD é um polímero constituído
de sete unidades de glicose que pode formar complexos solúveis em água com pequenas
moléculas e partes de grandes compostos (Pitha et al., 1988).
Diferentemente de outros agentes de ligação a colesterol que se incorporam em
membranas, as ciclodextrinas agem estritamente na superfície extraindo seletiva e
rapidamente o colesterol de membrana pela inclusão desta molécula em sua cavidade central
hidrofóbica (Zidovetzki & Levitan, 2007). O grau de depleção do colesterol na célula é
dependente da concentração da droga, tempo de incubação, temperatura e do tipo celular
utilizado no estudo.
Figura 10- Representação da metil-β-ciclodextrina. Essa estrutura é um polímero com uma cavidade não polar, onde o colesterol é armazenado na sua depleção na membrana plasmática. Fonte: www.chemblink.com

24
1.9 A relevância do estudo
O estudo sobre microdomínios lipídicos (lipid rafts) vem se intensificando nos últimos
anos. O envolvimento destas estruturas em importantes eventos celulares torna de particular
interesse sua caracterização em células como a glia embainhante olfatória (GEO), cujo efetivo
uso em terapias neuroregenrativas depende de um melhor entendimento de suas propriedades
fundamentais. O objeto de estudo deste trabalho é a caracterização das lipid rafts da GEO, por
critérios bioquímicos estabelecidos na literatura e por meio dos marcadores GM1, flotilina e
caveolina. Bem como, verificar as possíveis alterações dos microdomínios lipídicos da GEO
após a depleção de colesterol pelo fármaco metil-beta-ciclodextrina.
A conhecida falha na regeneração promovida pela GEO é atribuída a uma reduzida
motilidade dessa célula em determinadas circunstâncias. Dados preliminares do grupo
envolvido nesse estudo sugerem que o bloqueio funcional de gangliosídeos reconhecidos
pelos anticorpos Jones e A2B5, resulta na redução da motilidade da GEO. A identificação da
distribuição membranar destes componentes e da CNPase (proteína de mielina não-compacta)
em relação à microdomínios lipídicos, portanto, representa um importante passo no estudo de
seu envolvimento em vias de sinalização de influência na motilidade.

25
2 Objetivos
2.1 Geral
Isolar e caracterizar os microdomínios lipídicos (lipid rafts) presentes na membrana
plasmática da glia embainhante olfatória (GEO).
2.2 Específicos
Avaliar a presença de marcadores específicos nas frações resistentes a detergente;
Verificar a morfologia das lipid rafts;
Avaliar a composição lipídica das frações do gradiente de sacarose;
Investigar a presença da CNPase (proteína da mielina não-compacta característica da
GEO) nos microdomínios lipídicos da membrana;
Verificar se o gangliosídeo 9-O-acetil-GD3 e os gangliosídeos reconhecidos pelo
A2B5 co-localizam com lipid rafts, estabelecendo uma correlação no contexto da
migração da GEO;
Identificar as possíveis alterações dos microdomínios lipídicos da GEO após a
depleção de colesterol pela droga metil-beta-ciclodextrina.

26
3 Material e Métodos
3.1 Animais
Foram utilizados em nossos experimentos 53 ratos Wistar fêmeas e machos, com
idade de dois meses, a partir de cruzamentos feitos no biotério do Instituto de Bioquímica da
Universidade Federal do Rio de Janeiro (UFRJ).
Todos os procedimentos que utilizaram animais foram aprovados pelo Comitê de Ética
no Uso de Animais do Centro de Ciências da Saúde da UFRJ (protocolo IBCCF 020).
3.2 Cultura de células isoladas
Para aplicação dessa cultura, realizamos o método de isolamento de Nash e
colaboradores (2001) com algumas alterações.
Os animais foram sacrificados por deslocamento cervical, as cabeças foram cortadas
imediatamente e postas em PBS 10 mM, pH7.4 estéril (8 g de NaCl; 0,2 g de KCl; 1,44 g de
Na2HPO4. 2H2O; 0,2 g de KH2PO4 em 1 litro de água). A dissecção do BO foi realizada em
fluxo laminar, sendo o BO fatiado em um cortador de tecidos. As fatias (800 μm) foram
cortadas no plano transversal e tinham suas partes centrais retiradas, restando somente à
camada de fibras olfatórias (fatias enucleadas). As fatias enucleadas foram colocadas em
solução de Gey (0,17 g de CaCl2, 0,37 g KCl, 0,03 g de KH2PO4, 0,098 g de MgCl2, 0,139 g
de MgCO3 7.H2O, 8 g de NaCl, 0,227 g de NaHCO3 e 0,12 g de NaHPO4) gelada e estéril e
picotadas, onde o tecido foi dissociado através da adição de EDTA 0.01% por 15 minutos e
triturado através de várias passagens por uma seringa de 5 ml com agulha nº21. Obtida a
suspensão de células, foram feitas três centrifugações a 1.500 rpm por 5 minutos, onde os
sobrenadantes foram descartados e o pellet de células foi ressuspenso em meio DMEM/F12
(com 3,15 g/l de glicose, fungisona 25 g/ml, 10.000 g/ml unidades de penicilina, 10.000
g/ml de estreptomicina, 0,365 g/l de glutamina e 50 g/ml de gentamicina) com 10% soro

27
fetal bovino (Gibco, inativado a 56C por uma hora). Após a última centrifugação, o
sobrenadante foi desprezado e o pellet de células foi ressuspenso em 5 ml de DMEM/F12,
enriquecido com extrato de pituitária bovina (20 l/ml, Sigma-Aldrich) e forscolina (2 M,
Sigma-Aldrich). As células foram postas na garrafa de cultura e mantidas na estufa a 37C
submetidas a uma concentração de 5% de CO2. Passadas as primeiras 18 horas, o
sobrenadante da cultura foi passado para uma nova garrafa (ocorria adesão de fibroblastos na
primeira garrafa) e após 36 horas, o sobrenadante da cultura foi colocado em lamínulas
tratadas previamente com poli-L-lisina (20 g/ml, Sigma-Aldrich) cobertas com laminina (40
g/ml, Invitrogen), uma vez que ocorria adesão de astrócitos na segunda garrafa. O meio de
cultura era trocado a cada dois dias e após cinco dias em cultura após o plaqueamento
diferencial, as células foram fixadas em solução de paraformaldeído 4% em tampão fosfato
0,1 M pH 7, por 15 minutos, lavadas com PBS e guardadas na mesma solução a 4ºC.
3.3. Tratamento das células com metil-beta-ciclodextrina (MβCD)
Antes da realização do isolamento das membranas resistentes a detergente, 109 células
foram incubadas com metil-beta-ciclodextrina 20 mM por 30 minutos a 37ºC. Logo após,
essas células puderam seguir a metodologia de isolamento das membranas resistentes a
detergente, conferindo parte integrante do gradiente de sacarose em condição tratada.
3.4 Isolamento das membranas resistentes a detergente da GEO
Uma suspensão de 109 células (em condição controle ou tratada com MβCD 20 mM)
foi avolumada para 1 ml com tampão TNE [Tris-HCl 25 mM pH 7,3; NaCl 150 mM; EDTA
(ácido etilenodiaminotetracético) 5 mM], acrescida de inibidores de protease (PMSF 1mM.
Benzamidina 5 mM, pepstatina 10 μM, coquetel da Sigma e leupeptina 10 mM) a 4°C. Esse
material foi passado na seringa com agulha de 23G e sonicado 10% da amplitude total do

28
aparelho (Branson Sonifier 250) por 3 ciclos de 10 s e intervalo de 3 s para o rompimento
total das membranas. Ao término, foi realizada a extração com Triton X-100 1% em tampão
TNE a 4°C por 20 minutos. Finalizando o tratamento, a amostra foi misturada com um
tampão de sacarose 80% (2,3 M) em TNE na proporção de 1:1, de forma que a concentração
resultante da amostra fosse de 40%. No fundo do tubo da centrífuga foi colocada essa
solução, acima foram acrescentados 4 ml de sacarose 30% (0,88 M) e por último, 4 ml de
sacarose 5% (0,15 M). O gradiente foi centrifugado por 20-22 horas a 38.000 rpm com rotor
basculante do tipo SW40Ti em uma ultracentrífuga Beckman (Beckman Coulter Inc), a 4°C.
Depois da centrifugação, o material foi fracionado de 1 em 1 ml do topo do tubo ao final,
resultando em 13 frações.
3.5 Densidade das frações do gradiente de sacarose
Após o isolamento das membranas resistentes a detergente da GEO, 10 μl de cada
fração do gradiente controle foi analisada no refratômetro (American Optical Company). Os
valores obtidos nesse equipamento, índice de refração, foram convertidos com o uso da tabela
da Fiberlite (Piramoon Technologies Inc.) em densidade de sacarose (g/cm3).
3.6 Dot blotting das frações do gradiente de sacarose
Após o recolhimento das 13 frações dos gradientes de sacaroses controle e tratado com
MβCD, foram separados 500 µl de cada amostra. Na cuba do dot blotting, colocávamos duas
folhas de papel de filtro e uma membrana de nitrocelulose. Fechando a cuba, foi aplicado em
cada poço os 500 µl correspondentes das 13 frações. Com uma bomba de sucção acoplada na
cuba, as amostras foram passadas para a membrana de nitrocelulose, podendo assim iniciar o
bloqueio com TBS-Tween-Leite em pó desnatado (Tris-HCL 10 mM pH 7,2; NaCl 150 mM;
Tween 20 0,05% e leite em pó desnatado 15 g) per noite a 4°C. No dia seguinte, todo o

29
tampão foi retirado e incubado com o anticorpo primário (vide tabela 1) diluídos em TBS-
Tween-Leite em pó desnatado por 2 horas a temperatura ambiente. Agitando manualmente a
membrana foi lavada 5 vezes de 2 minutos com TBS-Tween-Leite em pó desnatado e logo em
seguida, incubada com o anticorpo secundário fluorescente por 1 hora à temperatura ambiente
(vide tabelas 2 e 3). Novamente, a membrana foi lavada 5 vezes de 2 minutos com TBS-
Tween-Leite em pó desnatado. Depois, lavada 3 vezes de 2 minutos com TBS-Tween (Tris-
HCL 10 mM pH 7,2; NaCl 150 mM; Tween 20 0,05%) e por último 3 lavagens de 2 minutos
de TBS (Tris-HCL 10 mM pH 7,2; NaCl 150 mM). A presença de fluorescência foi detectada
no aparelho Storm 860 (Molecular dynamics, Amersham Pharmacia Biotech).
3.7 Fluorescência da toxina do cólera subunidade B (CT-B)
As células purificadas de GEO foram previamente resfriadas em meio DMEM/F12 por
1 hora a 4°C. Posteriormente foram incubadas com a toxina do cólera (100 µg/ml) em meio
DMEM/F12 por 15 minutos a 4°C, donde uma lamínula foi retirada e fixada em
paraformaldeído 4% em PBS para observação no microscópio. O restante foi lavado em PBS
para remover o excesso do marcador não ligado e foi incubado por 30 minutos a 37°C. Por
fim, as células foram lavadas 2 vezes de 5 minutos em PBS, fixadas e incubadas com 4’,6-
Diamidino-2-phenylindole dihydrochloride (DAPI) por 5 minutos. Após lavagem com
solução de cloreto de sódio 0,9% por 5 minutos, o material foi montado em solução de p-
fenilenodiamino (5 mg de fenilenodiamino em 500 µl de PBS 0,15 M; 3,5 ml de glicerol e 1
ml de tampão carbonato-bicarbonato 0,5 M pH 9,2) e analisadas por microscopia de
fluorescência (Axioscope Zeiss).
3.8 Imunocitoquímica

30
As amostras (células isoladas fixadas da cultura de GEO) foram lavadas 5 vezes de 5
minutos com solução de PBS/Triton X-100 0,3%. Em seguida, foi feito o bloqueio das
reações inespecíficas com 10% de soro normal de cabra (NGS) e 2% de albumina sérica
bovina (BSA) em PBS/Triton X-100 durante 1 hora e meia. Após esse período, o material foi
novamente lavado e incubado com o(s) anticorpo(s) primário(s) de interesse (vide tabela 1)
diluído em PBS/Triton X-100 0,3%/BSA 1%, em pernoite. As amostras foram submetidas a
novas lavagens com PBS/Triton X-100 (5 vezes de 5 minutos cada). Ao término foi incubado
com anticorpo(s) secundário(s) adequado(s) (vide tabela 2), diluído em PBS/Triton X-100
0,3%/BSA 1% por 1 hora e em seguida, lavados 5 vezes de 5 minutos com PBS e incubado
novamente com DAPI (vide tabela 3) por 5 minutos. Após nova lavagem com solução de
NaCl 0,9% por 5 minutos, o material foi montado em solução de p-fenilenodiamino (5mg de
fenilenodiamino em 500 µl de PBS 0,15 M; 3,5 ml de glicerol e 1 ml de tampão carbonato-
bicarbonato 0,5 M pH9,2). As imagens foram obtidas por microscopia de fluorescência
(Axioscope Zeiss).
3.9 Anticorpos e outros ligantes
Tabela1- Anticorpos Primários
Anticorpo
PrimárioTipo Feito em Diluição Utilizada
Peso
MolecularOrigem
anti-CNPase monoclonal camundongo
1:100(imunocitoquímica)
1:1.000(immunoblotting)
46 kDa Sigma
anti-S100 policlonal coelho1:100
(imunocitoquímica) 20 kDa Sigma
anti-flotilina-1 policlonal ovelha
1:100(imunocitoquímica)
1:1.000(immunoblotting)
45 kDa
Santa Cruz
Biotechnology
anticorpo A2B5 monoclonal camundongo1:100
(immunoblotting)Abcam

31
anti-caveolina-1 policlonal coelho
1:100 (imunocitoquímica)
1:1.000(immunoblotting)
21 kDa Sigma
Jones monoclonal camundongo1:1000
(immunoblotting)Sigma
Tabela 2- Anticorpos Secundários
AnticorpoSecundário
Feito em Diluição Utilizada Origem
anti-IgG Alexa 488
coelho 1:200 Molecular
Probes
anti-IgG Alexa 488
camundongo1:200 Molecular
Probes
anti-IgG Alexa 546
camundongo1:200 Molecular
Probes
anti-IgG biotinilado
camundongo 1:1.000 Sigma
anti-IgG biotinilado
ovelha 1:1.000 Sigma
anti-IgG biotinilado
coelho 1:1.000 Sigma
anti-IgM μ-chain biotinilado
camundongo 1:1.000 Sigma
Tabela 3 - Ligantes e Sondas
Ligante/Sonda Diluição Utilizada Origem
DAPI 1:12.000 Molecular
Probes
toxina colérica conjugada a HRP (peroxidase de rabanete)
1:1.000 Sigma
toxina colérica conjugada a FITC (isotiocianato de fluresceína )
1:1.000 Sigma
extravidina conjugada a Cy31:500
Sigma
extravidina conjugada a FITC1:500
Sigma
extravidina conjugada a HRP1:1.000
Sigma

32
3.10 Microscopia Eletrônica de Transmissão (M.E.T.): Contrastação negativa
A quinta fração correspondente a raft e a fração pellet, correspondente à uma fração
não-raft foram depositadas sobre grades de cobre cobertas com filme de Formvar 0,4% em
clorofórmio e deixadas por 1 minuto. Depois de retirado o excesso de material com auxílio de
papel de filtro Whatman número 1 foi realizado contrastação negativa do material com 1% de
acetato de uranila aquosa por 1 minuto, seguida por secagem completa da solução. As
amostras foram observadas no microscópio eletrônico (Zeiss 900) na voltagem de 80 kV. Essa
técnica foi realizada com a colaboração do Professor José Garcia Ribeiro A. Júnior e sua
aluna de mestrado Alice Helena dos Reis, do ICB da UFRJ.
3.11 Dosagem de Proteína
As determinações protéicas foram estimadas através do kit baseado no ácido
bicinchonínico (BCA – Protein Assay Reagent, Pierce), tendo como proteína padrão a
albumina sérica bovina (BSA).
Para o procedimento da dosagem, foram misturados os reagentes de trabalho de
acordo com as instruções do fabricante: 25 partes do reagente MA; 24 partes do reagente MB
e 1 parte do reagente MC. Uma solução conservada em estoque de BSA foi preparada, para
após o experimento, realizar a curva padrão. A curva padrão foi constituída de 12 pontos de
0,5 µg/ml até 200 µg/ml de BSA. Nas amostras do padrão foram acrescentados, 150 µl de
reagente de reagente de trabalho e completados até 300 µl com SDS 0,2%. Para dosar as
frações dos gradientes controle e tratado com MβCD, foram usadas 150 µl do reagente de
trabalho, 40 µl de água, 10 µl das amostras e 100 µl de SDS 0,3%. Após 2 horas a 37°C, foi
realizada a leitura das amostras no comprimento de onda de 562 nm.

33
3.12- Análise Lipídica
3.12.1 Extração de lipídios
As frações recolhidas dos gradientes controles foram submetidas à extração lipídica de
acordo com o método de Bligh e Dyer (1959) em clorofórmio:metanol:água 1:2:0,8 (v/v),
cujo volume final foi de 1 ml, sob agitação de 5 em 5 minutos por 1 hora. Após o período da
extração, a solução foi centrifugada a 3000 rpm por 15 minutos, para separação do
sobrenadante. Esse sobrenadante (Sb1) foi separado e o pellet tinha sua continuidade na
extração. Novamente, foi usado clorofórmio:metanol:água 1:2:0,8, agitados por 1 hora e foi
centrifugado por 15 minutos a 3000 rpm. O pellet dessa centrifugação correspondia ás
proteínas desnaturadas. Por vez, esse sobrenadante (Sb2) foi acrescentado com o primeiro
(Sb1) para completar a re-extração, que consistiu em água:clorórmio 1:1 (v/v). Essa etapa
tinha uma nova centrifugação de 15 minutos a 3000 rpm. A fase orgânica (inferior)
apresentando os lipídios foi seca em vapores de nitrogênio gasoso.
3.12.2 Identificação dos lipídios
Os lipídios secos por nitrogênio gasoso da fase orgânica foram ressuspensos num
volume de 50-100 µl de clorofórmio e analisados numa cromatografia de camada fina de alta
performance (High Performance Thin-Layer Chromatography, HPTLC) unidimensional em
placas de Silica Gel 60 (Merck), sendo aplicados em faixas de 0,5 cm situadas a 1cm da
extremidade inferior da placa. Para a HPTLC de lipídios neutros, foi utilizado o sistema de
solventes hexano:éter etílico:ácido acético 60:40:1 (v/v/v; Kawooya e Law, 1988), por
aproximadamente 10 minutos de corrida. Após a corrida, as placas secas foram embebidas
com Reagente de Charring (CuSO4 10% + H3PO4 8%), e aquecidas a 200oC em estufa FAMO
Modelo ST200 por 20 minutos (Ruiz e Ochoa, 1997), permitindo a visualização quantitativa
dos lipídios presentes.

34
Para a análise de fosfolipídios, utilizamos o sistema de solventes
clorofórmio:acetona:metanol:ácido acético:água 40:15:13:12:8 (v/v; Ruiz e Ochoa, 1997) por
aproximadamente 30-40 minutos. O sistema foi revelado utilizando Reagente de Charring
(CuSO4 10% em H3PO4 8%) por 10 minutos a 100°C (Bitman e Wood, 1982).
As placas foram digitalizadas com o auxílio de um scanner e submetidas à análise
densitométrica, sendo utilizado o programa Image Master Total Lab v1.11 (Amersham
Pharmacia Biotech). Cada lipídio foi assinalado comparando-se com a migração de padrões
aplicados paralelamente.
3.13.3 Análise de Colesterol
Lipídios extraídos das frações dos gradientes de sacarose controle e tratado com
MβCD, como descrito no item 3.13.1, foram quantificados quanto ao seu conteúdo de
colesterol pelo método de Zlatkis e colaboradores, 1953. As amostras foram ressuspensas em
750 µl de ácido acético glacial e, em seguida, foram adicionados 500 µl da solução de FeCl3
0,2% em H3PO4 85% e H2SO4 concentrado. Após 5 a 10 minutos, as amostras apresentavam
uma cor amarronzada e desse modo à absorbância foram lidas com o comprimento de onda de
550 nm no espectrofotômetro (Analyser), utilizando colesterol (Sigma) como padrão para
análise.

35
4 Resultados
Com a finalidade de confirmar a identidade da GEO em culturas de células isoladas,
utilizamos o anticorpo policlonal anti-S100 e o anticorpo monoclonal anti-CNPase, um
marcador da GEO proposto pelo nosso grupo (Santos-Silva e Cavalcante, 2001). Os dados
demonstraram através da microscopia de imunofluorescência que, virtualmente todas as
células das culturas isoladas da GEO apresentaram imunorreatividade para a proteína S100 e
CNPase (Figura 11). Desse modo, os objetivos propostos e os resultados obtidos passam a ser
descritos.
4.1 Perfil do gradiente de sacarose
Com o propósito de caracterizar os microdomínios lipídicos, extratos de GEO foram
incubados com o detergente não iônico, Triton X-100, e submetidos a um gradiente de
sacarose. Após a centrifugação, pudemos verificar a presença de um material flutuante na
interface de 5-30% de sacarose (Figura 12B). Esse material em suspensão foi visualizado em
todas as purificações das lipid rafts. Desse modo, os gradientes de sacarose foram recolhidos
em 13 frações de 1 ml contendo diferentes densidades (Figura 13) e posteriormente
analisados.
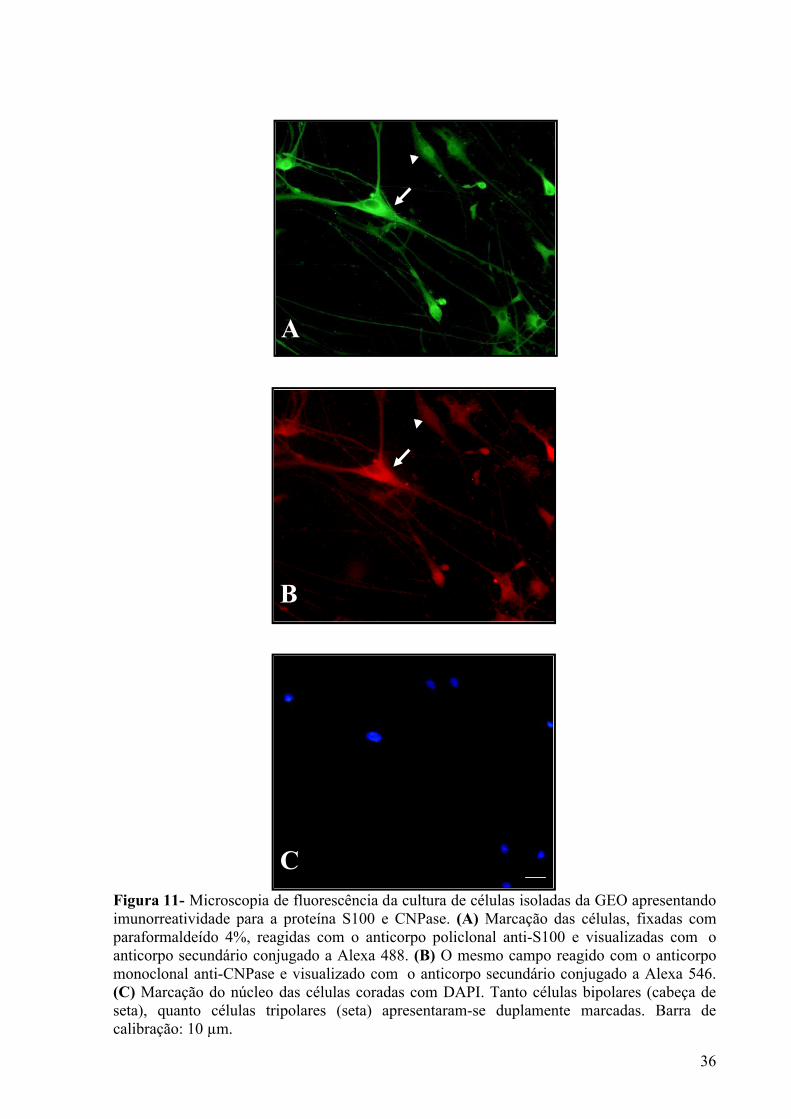
36
Figura 11- Microscopia de fluorescência da cultura de células isoladas da GEO apresentando imunorreatividade para a proteína S100 e CNPase. (A) Marcação das células, fixadas com paraformaldeído 4%, reagidas com o anticorpo policlonal anti-S100 e visualizadas com o anticorpo secundário conjugado a Alexa 488. (B) O mesmo campo reagido com o anticorpomonoclonal anti-CNPase e visualizado com o anticorpo secundário conjugado a Alexa 546. (C) Marcação do núcleo das células coradas com DAPI. Tanto células bipolares (cabeça de seta), quanto células tripolares (seta) apresentaram-se duplamente marcadas. Barra de calibração: 10 µm.
B
C
A

37
Figura 12- Ilustração da obtenção do gradiente de sacarose. (A) Esquema do tubo do rotor basculante do tipo SW40Ti da ultracentrífuga Beckman, com o perfil das soluções de sacarose. (B) Fotografia do resultado do gradiente de sacarose após 20-22 horas de centrifugação. Pode-se observar um material flutuante na interface de sacarose 5-30% (seta). Deste gradiente, treze frações de 1 ml foram recolhidas e analisadas nos experimentosseguintes.
Figura 13- Densidade das frações do gradiente de sacarose da GEO. As células (109
células/ml) foram lisadas, homogeneizadas e incubadas a 4°C com 1% Triton X-100 e submetidas à centrifugação em gradiente de sacarose. Após a retirada do gradiente de sacarose da ultracentrífuga, as treze frações foram analisadas no refratômetro e seus resultados convertidos em densidade (vide item 3.5 do material e métodos). Observa-se um aumento gradual da densidade ao longo do gradiente. A análise estatística foi realizada usando o programa GraphPad Prism. Cada fração representa a média e o desvio padrão de 5 experimentos diferentes.
Sacarose 5%
Sacarose 30%
Amostra em sacarose 40%
20-22hs a 38000rpm
A B
1 2 3 4 5 6 7 8 9
10
11
12
13
Densidade do gradiente de sacarose
1 2 3 4 5 6 7 8 9 10 11 12 130.95
1.00
1.05
1.10
1.15
1.20
Frações
g/cm
3
topo

38
4.2 Detecção do gangliosídeo GM1 nas frações do gradiente de sacarose e em células
vivas
Com o intuito de distinguir as frações lipid rafts isoladas foi realizado o método do dot
blotting usando a toxina colérica para o reconhecimento do gangliosídeo GM1 (esfingolipídio
marcador das lipid rafts). Na figura 14, pode se observar na forma do dot blotting (A) e
análise densitométrica (B), que a presença do gangliosídeo GM1 não é homogênea em todas
as frações do gradiente. As marcações foram mais intensas nas frações menos densas do
gradiente de sacarose.
Decidimos examinar a presença do gangliosídeo GM1 em células vivas, a partir do
resultado positivo do dot blotting para esse esfingolipídio. Com esse objetivo, nosso primeiro
ensaio consistiu na incubação das células com a toxina do cólera conjugado a FITC a 4°C, a
fim de verificar apenas os sítios de ligação da toxina com o gangliosídeo. A figura 15A
mostra marcação homogênea ao longo de toda a célula, não sendo possível observar
evidências de internalização da toxina na GEO. Logo após, foi realizada a mesma incubação
da toxina colérica, entretanto colocamos a GEO na temperatura ideal do metabolismo basal
(37°C). A marcação na região perinuclear observada nessa condição sugere possivelmente a
internalização da toxina nas células (Figura 15D).

39
A
B
Figura 14- Distribuição do gangliosídeo GM1 nas frações do gradiente de sacarose. As células (109 células/ml) foram lisadas, homogeneizadas e incubadas a 4°C com 1% Triton X-100 e submetidas à centrifugação em gradiente de sacarose. (A) Para averiguar a presença do gangliosídeo GM1, as frações foram submetidas ao dot blotting usando a subunidade B da toxina do cólera conjugado a FITC. Em destaque, as frações 4-6 que correspondem com o material flutuante do gradiente de sacarose. (B) A análise densitométrica foi realizada usando o programa Image Master TotalLab. Cada fração representa a média e o desvio padrão de 5 experimentos diferentes.
1 2 3 4 5 6 7 8 9 10 11 12 130
1 0
2 0
F r a ç õ e s
1 2 3 4 5 6 7 8 9 10 11 12 13
topo
Por
cen
tage
m

40
Figura 15- Microscopia de fluorescência da GEO incubada na presença de toxina do cólera subunidade B conjugada à FITC a 4 e 37°C. (A e D) Marcação com ligante do gangliosídeoGM1. (B e E) O mesmo campo mostra os núcleos das células corados pelo DAPI. (C e F) Sobreposição das imagens. Observa-se marcações mais evidentes principalmente na região perinuclear (setas). Barra de calibração: 5 μm.
B
4°C
A
E
D
37°C
C F
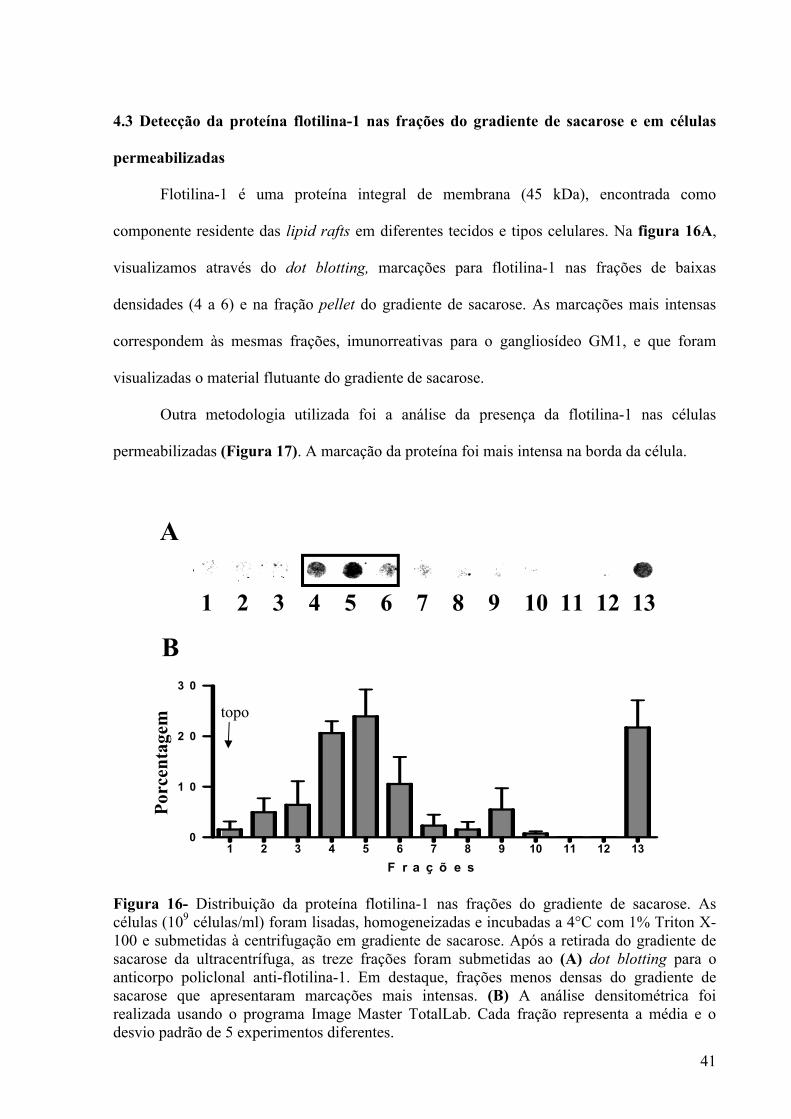
41
4.3 Detecção da proteína flotilina-1 nas frações do gradiente de sacarose e em células
permeabilizadas
Flotilina-1 é uma proteína integral de membrana (45 kDa), encontrada como
componente residente das lipid rafts em diferentes tecidos e tipos celulares. Na figura 16A,
visualizamos através do dot blotting, marcações para flotilina-1 nas frações de baixas
densidades (4 a 6) e na fração pellet do gradiente de sacarose. As marcações mais intensas
correspondem às mesmas frações, imunorreativas para o gangliosídeo GM1, e que foram
visualizadas o material flutuante do gradiente de sacarose.
Outra metodologia utilizada foi a análise da presença da flotilina-1 nas células
permeabilizadas (Figura 17). A marcação da proteína foi mais intensa na borda da célula.
A
B
Figura 16- Distribuição da proteína flotilina-1 nas frações do gradiente de sacarose. As células (109 células/ml) foram lisadas, homogeneizadas e incubadas a 4°C com 1% Triton X-100 e submetidas à centrifugação em gradiente de sacarose. Após a retirada do gradiente de sacarose da ultracentrífuga, as treze frações foram submetidas ao (A) dot blotting para o anticorpo policlonal anti-flotilina-1. Em destaque, frações menos densas do gradiente de sacarose que apresentaram marcações mais intensas. (B) A análise densitométrica foi realizada usando o programa Image Master TotalLab. Cada fração representa a média e o desvio padrão de 5 experimentos diferentes.
1 2 3 4 5 6 7 8 9 10 11 12 13
1 2 3 4 5 6 7 8 9 10 11 12 130
1 0
2 0
3 0
F r a ç õ e s
topo
Por
cen
tage
m

42
Figura 17- Microscopia de fluorescência da GEO apresentando imunorreatividade para a proteína flotilina-1. (A) Marcação da GEO, fixada com paraformaldeído 4%, para o anticorpo policlonal anti-flotilina-1, posterior incubação com anticorpo secundário conjugado à biotina e extravidina conjugada a FITC. (B) Marcação do núcleo da célula corado com DAPI. (C) Sobreposição das imagens. Notar a maior intensidade na borda da célula (seta). Barra de calibração: 5 μm.
A
B
C

43
4.4 Detecção da proteína caveolina-1 nas frações do gradiente de sacarose e em células
permeabilizadas
Outra proteína marcadora dos microdomínios lipídicos analisada nesse trabalho foi a
caveolina. Essa proteína é integrante das cavéolas, uma subclasse das lipid rafts. Com a
finalidade de verificar se a raft da GEO, podia ser do tipo caveolar foi realizada a metodologia
de dot blotting para a proteína caveolina-1. Na figura 18, visualizamos marcações para
caveolina-1 nas frações 4 a 9 do gradiente de sacarose.
Outra abordagem de estudo realizada com células da GEO permeabilizadas foi o
experimento de imunocitoquímica para caveolina-1 (Figura 19). Observa-se marcação da
proteína em toda a célula, com algumas regiões puntiformes na borda da célula.
A
B
Figura 18- Distribuição da proteína caveolina-1 nas frações do gradiente de sacarose. As células (109 células/ml) foram sonicadas, homogeneizadas e incubadas a 4°C com 1% Triton X-100 e submetidas à centrifugação em gradiente de sacarose. Em seguida, as treze foram submetidas ao (A) dot blotting para o anticorpo policlonal anti-caveolina-1. Em destaque, maior intensidade de marcação no meio do gradiente de sacarose. (B) A análise densitométrica foi realizada usando o programa Image Master TotalLab. Cada fração representa a média e o desvio padrão de 5 experimentos diferentes.
1 2 3 4 5 6 7 8 9 10 11 12 13
topo

44
Figura 19- Microscopia de fluorescência da GEO apresentando imunorreatividade para a proteína caveolina-1. (A) Marcação da GEO, fixada com paraformaldeído 4%, reagida com o anticorpo policlonal anti-caveolina-1 e visualizada com o anticorpo secundário conjugado a Alexa 488. (B) Marcação do núcleo da célula corado com DAPI. (C) Sobreposição das imagens. Observa-se marcações puntiformes na borda da célula (seta). Barra de calibração: 5 μm.
A
B
C

45
4.5 Morfologia das frações resistentes a detergente
Por meio da microscopia eletrônica de transmissão, visualizamos a morfologia das
lipid rafts e de componentes da fração de alta densidade (pellet) do gradiente de sacarose da
GEO. Estas frações foram previamente dialisadas e coradas negativamente com acetato de
uranila.
A micrografia eletrônica presente na figura 20A demonstra a natureza vesicular com
aparência multilamelar das membranas resistentes a detergente. Por vez, a fração pellet
(Figura 20B) foi analisada como fração controle negativo. Nesta fração não aparece formação
de vesículas, apenas estruturas irregulares e aglomerados de material amorfo. Provavelmente,
estas estruturas representam agregados macromoleculares de proteínas e/ou lipídios
solubilizados pelo detergente.
FRAÇÃO FRAÇÃO LIPID RAFT PELLET
Figura 20- Micrografias eletrônicas das frações do gradiente de sacarose. As células (109
células/ml) foram sonicadas, homogeneizadas e incubadas a 4°C com 1% Triton X-100 e submetidas à centrifugação por gradiente de sacarose. Após a retirada do gradiente de sacarose da ultracentrífuga foram recolhidas treze frações. A quinta fração (fração lipid raft) e uma fração de alta densidade (pellet) foram dialisadas e coradas negativamente com acetato de uranila, pela técnica de contrastação negativa. (A) Fração lipid raft mostrando perfis de membranas altamente organizados e (B) fração pellet apresentando estruturas pequenas de aparência granular. Barra de calibração: 0,2 μm.
A B

46
4.6 Distribuição de proteína e colesterol nas frações do gradiente de sacarose
Com o intuito de dar continuidade a caracterização das rafts foi realizada a dosagem
de colesterol pelo método colorimétrico. Na figura 21A, observamos a quantidade de
colesterol ao longo das frações do gradiente de sacarose. A concentração maior de colesterol
foi observada na fração 5, uma fração raft.
A dosagem de proteína foi realizada pela metodologia baseada no ácido
bicinchonínico (BCA). Essa identificação das frações ao longo do gradiente se torna
essencial, visto que as rafts apresentam poucas quantidades de proteínas. Na figura 21B,
notamos pequenas concentrações de proteínas nas frações de baixa densidade de sacarose,
comparado com as frações de alta densidade. A quantidade de proteínas tornou-se elevada,
principalmente a partir da fração 11.
Outra forma de avaliar a presença de colesterol e proteína nos microdomínios lipídicos
foi através da relação destes dois elementos (colesterol/proteína), como pode ser visto na
figura 21C. A fração com maior razão de colesterol por proteína foi a fração 5, uma fração
raft.

47
Figura 21- Concentração total de colesterol e proteínas nas frações do gradiente de sacarose da GEO. (A) A dosagem de colesterol foi realizada após a extração lipídica de acordo com Bligh e Dyer (1959). Nas frações 4-6 observa-se um pico de colesterol, características dasfrações rafts. (B) A dosagem de proteína foi quantificada de acordo com o método BCA. Pode-se observar que a concentração protéica aumenta em frações de maior densidade do gradiente. (C) Razão colesterol/proteína de todas as frações do gradiente de sacarose. Cada fração representa a média e o desvio padrão de 4 experimentos diferentes.
topo
topo
Dosagem de colesterol
1 2 3 4 5 6 7 8 9 10 11 12 130
5
10
15
20
25
30
35
Frações
g/m
l
A Dosagem de proteína
1 2 3 4 5 6 7 8 9 10 11 12 130
25
50
75
Frações
g/m
l
B
Colesterol/Proteína
1 2 3 4 5 6 7 8 9 10 11 12 130
1
2
Frações
N°
arb
itrá
rio
C

48
4.7 Análise dos lipídios totais das frações do gradiente de sacarose da GEO
Os microdomínios lipídicos foram localizados através dos marcadores específicos das
rafts e como próximo passo foi realizada a análise de lipídio total pela cromatografia de
camada fina de alta performance (HPTLC) das frações do gradiente de sacarose. A separação
ocorreu de acordo com o grau de afinidade dos lipídios com o substrato e sua comparação
com padrões comerciais.
A análise dos lipídios neutros revelou a presença de colesterol, a partir da quinta
fração (Figura 22A). Por vez, os dados de densitometria da HPTLC foram plotados em um
gráfico, no qual se demonstrou maior quantidade deste lipídio na quinta fração (Figura 22B).
Pela análise dos fosfolipídios (Figura 22C), observamos que na maioria das frações
do gradiente, a fosfatidilcolina (PC) e um fosfolipídio de natureza não determinada (ND)
correspondiam aos principais fosfolipídios presentes no gradiente de sacarose. Curiosamente,
a quinta fração foi a única que apresentou além desses componentes, a esfingomielina (SM).
A esfingomielina é um esfingolipídio encontrado no sistema nervoso, que age concentrando
moléculas sinalizadoras e também, como precursora de vários lipídios bioativos.

49
Lipidio Neutro - Colesterol
1 2 3 4 5 6 7 8 9 10 11 12 130
5
10
15
20
25
30
35
Frações
Por
cent
agem
B
Figura 22- Caracterização lipídica das frações do gradiente de sacarose da GEO. Após o gradiente de sacarose, as frações marcadas foram submetidas à extração lipídica e analisadas em cromatografia de camada fina de alta performance (HPTLC) para (A) lipídio neutro e (C)fosfolipídios. (B e D) Análise densitométrica da HPTLC, usando o programa Image Master TotalLab. Cada fração representa a média e o desvio padrão de 3 experimentos diferentes.CHO - Colesterol; PC - Fosfatidilcolina; SM - Esfingomielina; ND - Não determinado.
CHO
A
SM PC
C D
1 2 3 4 5 6 7 8 9 10 11 12 13
ND
1 2 3 4 5 6 7 8 9 10 11 12 13
Fosfolipídios
1 2 3 4 5 6 7 8 9 10 11 12 130
10
20PCNDSM
Frações
Po
rcen
tag
em

50
Após essa caracterização e sobreposição dos marcadores das lipid rafts, podemos
destacar, nesse estudo, que as frações 4-6 correspondem as frações rafts. Esses resultados são
corroborados pelos critérios bioquímicos: baixa quantidade de proteína e alta quantidade de
colesterol que essas frações apresentam.
4.8 Localização da proteína CNPase nas frações do gradiente de sacarose e em células
permeabilizadas
Dados na literatura sugerem que a CNPase pode participar da intermediação de
interações entre elementos do citoesqueleto (actina e microtúbulo) e da membrana plasmática
(Dyer e Benjamins, 1989). Desta forma, a CNPase poderia estar envolvida em eventos de
sinalização membrano-esqueletais. Por esta razão, decidimos verificar se a CNPase colocaliza
com as lipid rafts da GEO.
Para verificar a localização da CNPase nas rafts, foi realizada o método de dot blotting
(Figura 23). O resultado da marcação para CNPase apresentou similaridade ao encontrado
para a proteína flotilina-1 (Figura 16). A CNPase no dot blotting apresentou marcação mais
expressiva na quinta fração, correspondente a uma fração raft.
Outra abordagem apresentada neste estudo, foi a análise da distribuição das proteínas
flotilina-1 e CNPase na GEO. Para tal, foram realizadas imunocitoquímicas em culturas de
células isoladas (Figura 24). A distribuição da CNPase foi observada ao longo de toda a
célula, apresentando regiões de maior intensidade e a da flotilina foi mais limitada a
determinadas regiões. A sobreposição das marcações se deu em apenas alguns pontos da
célula.

51
A
B
Figura 23- Distribuição da proteína CNPase nas frações do gradiente de sacarose. As células (109 células/ml) foram lisadas, homogeneizadas e incubadas a 4°C com 1% Triton X-100 e submetidas à centrifugação em gradiente de sacarose. Em seguida, as treze frações foram submetidas ao (A) dot blotting para o anticorpo monoclonal anti-CNPase. Observa-se uma marcação mais intensa na fração 5, que corresponde a uma fração raft. (B) A análise densitométrica foi realizada usando o programa Image Master TotalLab. Cada fração representa a média e o desvio padrão de 4 experimentos diferentes.
1 2 3 4 5 6 7 8 9 10 11 12 130
1 0
2 0
3 0
4 0
5 0
6 0
F r a ç õ e s
1 2 3 4 5 6 7 8 9 10 11 12 13
topo
Por
cen
tage
m

52
Figura 24- Microscopia de fluorescência da GEO apresentando imunorreatividade para CNPase e flotilina-1. (A) Marcação da célula, fixada com paraformaldeído 4%, para o anticorpo monoclonal anti-CNPase reagido com o anticorpo secundário conjugado a Alexa 488. (B) Marcação da célula com o anticorpo policlonal anti-flotilina-1, posterior incubação com anticorpo secundário biotinilado e reação deste com extravidina conjugada a Cy3. (C) Marcação do núcleo da célula corada com DAPI. (D) Sobreposição das imagens. Notar, sobreposição das proteínas em uma região da célula (seta). Barra de calibração: 5µm.
B
A
C
D

53
4.9 Localização do gangliosídeo 9-O-acetil GD3 e dos gangliosídeos reconhecidos pelo
anticorpo A2B5 (G-A2B5) nas frações do gradiente de sacarose
Trabalhos preliminares do grupo envolvido neste estudo, demonstraram que células
migratórias da GEO cultivadas sobre o substrato de laminina expressam 9-O-acetil GD3 e os
gangliosídeos reconhecidos pelo anticorpo A2B5. Um ponto importante para entender a
influência desses gangliosídeos no processo de migração da GEO é verificar a relação destes
esfingolipídios com as lipid rafs. Dessa forma, para verificar se as rafts da GEO possuem
esses gangliosídeos foi realizado o método dot blotting.
Os resultados obtidos para 9-O-acetil GD3 (através do anticorpo Jones) e G-A2B5
apresentaram marcação mais evidente na quinta fração, uma fração raft, e no pellet, onde se
encontram os outros componentes celulares pesados, tais como: núcleo e organelas (Figura
25).

54
A
B
C
D
Figura 25- Distribuição do gangliosídeo 9-O-acetil GD3 e dos gangliosídeos reconhecidos pelo anticorpo A2B5 (G-A2B5) nas frações do gradiente de sacarose. As células (109
células/ml) foram sonicadas, homogeneizadas e incubadas a 4°C com 1% Triton X-100 e submetidas à centrifugação em gradiente de sacarose. Em seguida, as treze foram submetidas ao dot blotting para (A) o gangliosídeo 9-O-acetil GD3 e (C) G-A2B5. (A e C) Observa-seuma marcação mais intensa na fração 5, uma fração raft. (B e D) A análise densitométrica foi realizada usando o programa Image Master TotalLab. Cada fração representa a média e o desvio padrão de 4 experimentos diferentes.
1 2 3 4 5 6 7 8 9 10 11 12 130
5
1 0
1 5
2 0
2 5
3 0
3 5
F r a ç õ e s
1 2 3 4 5 6 7 8 9 10 11 12 13
9-O-acetil GD3
G-A2B5
1 2 3 4 5 6 7 8 9 10 11 12 13
1 2 3 4 5 6 7 8 9 10 11 12 130
1 0
2 0
3 0
4 0
5 0
F r a ç õ e s
topo
topo
Por
cen
tage
mP
orce
nta
gem

55
4.10 Desorganização das lipid rafts
Uma das formas de desfazer as lipid rafts é através do uso da MβCD que depleta o
colesterol da membrana plasmática. Para investigar alterações na morfologia celular
associadas à retirada de colesterol, GEOs foram tratadas com MβCD 20 mM durante 30
minutos a 37°C e em seguida, foram realizadas ensaios de imunocitoquímica com as proteínas
marcadoras: S100 e CNPase (Figura 26). As células tratadas com MβCD mostraram-se mais
aplanadas no substrato, se comparadas com as controles. Outra diferença resultante do uso da
MβCD nas células foi observada no gradiente de sacarose. Os gradientes das células tratadas,
diferentemente dos gradientes controles, não apresentaram o material flutuante na interface de
5-30% de sacarose (Figura 27).
Para analisar o efeito da depleção de colesterol na organização dos componentes
encontrados nos microdomínios lipídicos, realizamos a mesma metodologia de dot blotting
com os gradientes de células tratadas com MβCD. Observa-se na figura 28A, uma
distribuição mais homogênea dos componentes analisados das rafts (GM1, flotilina-1 e
caveolina-1) ao longo do gradiente, se comparado com o gradiente controle (Figura 28B).
Com o intuito de dar continuidade ao estudo do efeito da depleção de colesterol, foram
realizadas dosagens de colesterol e de proteínas (Figura 29). Ambos componentes tiveram
sua distribuição alterada com o uso da droga. Houve uma redução de colesterol de quase 22%
no gradiente com células tratadas com MβCD (Figura 29A). Por vez, a dosagem de proteína
apresentou uma distribuição homogênea ao longo das frações do gradiente (Figura 29B).
Outra forma de avaliar a presença de colesterol e proteína foi através da razão destes dois
elementos (Figura 29C).
A distribuição das moléculas candidatas ao envolvimento da migração da GEO
encontradas nos microdomínios lipídicos, também foram analisadas nos gradientes de células
tratadas com MβCD. Na figura 30A, pudemos observar uma distribuição homogênea dos

56
componentes analisados (CNPase, o gangliosídeo 9-O-acetil GD3 e G-A2B5) ao longo das
frações do gradiente, se comparado com o gradiente controle (Figura 30B).
Outra abordagem apresentada neste estudo, foi a análise da distribuição das proteínas
flotilina-1 e CNPase em culturas de células isoladas tratadas com MβCD (Figura 31). Para
tal, foram realizados ensaios de imunocitoquímicas, que revelaram para ambas as proteínas
um padrão de marcação distribuído mais intensamente no corpo celular (colocalização em
amarelo). Em células sem o tratamento com a droga, a sobreposição acontece em alguns
pontos da célula.

57
MβCD Controle
Figura 26- Microscopia de fluorescência da GEO apresentando imunorreatividade para a proteína S100 e CNPase. (A-C) Células tratadas com MβCD 20 mM e (D-F) células controles. (A e D) Marcação das células, fixadas com paraformaldeído 4%, para o anticorpo policlonal anti-S100, reagido com o anticorpo secundário conjugado a Alexa 488. (B e E) Marcação das células com o anticorpo monoclonal anti-CNPase, reagido com o anticorpo secundário conjugado a Alexa 546. (C e F) Marcação do núcleo das células coradas com DAPI. Tanto células bipolares (seta), quanto células tripolares (cabeça de seta) apresentaram-se duplamente marcadas. Barra de calibração: 5 µm.
D
E
A
B
C F

58
Figura 27- Representação da obtenção dos gradientes de sacarose com MβCD e em condição controle. (A) Esquema do tubo do rotor basculante do tipo SW40Ti da ultracentrífuga Beckman, com o perfil das soluções de sacarose. (B) Fotografia do resultado do gradiente de sacarose da GEO (109 células/ml) tratada com MβCD 20 mM por 30 minutos e (C) gradiente controle, após 20-22 horas de centrifugação. Pode-se observar um material flutuante na interface de sacarose 5-30% no gradiente controle (seta), dado não observado no tratado com a droga.
Sacarose 5%
Sacarose 30%
Amostra em sacarose 40%
20-22hs a 38000rpm
1 2 3 4 5 6 7 8 9
10
11
12
13
A CB1
2
34
5
6
7
8
9
10
11
12
13

59
A
MβCD
Gangliosídeo GM1
Flotilina-1
Caveolina-1
B
Gangliosídeo GM1
Flotilina-1
Caveolina-1
Figura 28- Distribuição dos marcadores de rafts. (A) GEOs (109 células/ml) tratadas com MβCD 20 mM por 30 minutos ou (B) em condição controle foram lisadas, homogeneizadas e incubadas a 4°C com 1% Triton X-100 e submetidas à centrifugação em gradiente de sacarose. Para averiguar a presença do gangliosídeo GM1, flotilina-1 e caveolina-1 as treze frações foram submetidas ao dot blotting. (A) Observa-se distribuição dos componentes das rafts por todas as frações do gradiente, quando as células são tratadas com a droga depletora de colesterol. (B) Em destaque, as frações 4-6 que correspondem com as lipid rafts da GEO.
1 2 3 4 5 6 7 8 9 10 11 12 13
1 2 3 4 5 6 7 8 9 10 11 12 13

60
MβCD Controle
Figura 29- Concentração total de colesterol e proteínas nas frações do gradiente de sacarose da GEO. (A-C) Gradiente de sacarose da GEO tratada com MβCD e (D-F) gradiente em condição controle. (A e D) A dosagem de colesterol foi realizada após a extração lipídica de acordo com Bligh e Dyer (1959). (A) Observa-se distribuição heterogênea de colesterol entre as frações. (B e E) A dosagem de proteína foi quantificada de acordo com o método BCA. (B)Observa-se uma distribuição de proteína homogênea pelo gradiente de sacarose, com um pico na 13ª fração. (C e F) Razão colesterol/proteína de todas as frações do gradiente de sacarose.
Colesterol/Proteína
1 2 3 4 5 6 7 8 9 10 11 12 130
1
2
Frações
Nº
arb
itrá
rio
Dosagem de proteína
1 2 3 4 5 6 7 8 9 10 11 12 130
25
50
75
Frações
g
/ml
Dosagem de colesterol
1 2 3 4 5 6 7 8 9 10 11 12 130
5
10
15
20
25
Frações
g/m
l
Dosagem de colesterol
1 2 3 4 5 6 7 8 9 10 11 12 130
5
10
15
20
25
30
35
Frações
g/m
l
Dosagem de proteína
1 2 3 4 5 6 7 8 9 10 11 12 130
25
50
75
Frações
g/m
l
Colesterol/Proteína
1 2 3 4 5 6 7 8 9 10 11 12 130
1
2
Frações
N°
arbi
trár
io
A
B
C
E
F
Dtopo topo
topo topo
topo topo

61
A
MβCD
B
Figura 30- Distribuição das moléculas candidatas no envolvimento da migração da GEO nas frações do gradiente de sacarose. (A) As células (109 células/ml) tratadas com MβCD 20 mM por 30 minutos ou (B) em condição controle foram lisadas, homogeneizadas e incubadas a 4°C com 1% Triton X-100 e submetidas à centrifugação em gradiente de sacarose. As treze frações foram submetidas ao dot blotting para CNPase, o gangliosídeo 9-O-acetil GD3 e G-A2B5. (A) Observa-se distribuição das moléculas candidatas no envolvimento da migração da GEO por todas as frações do gradiente, quando as rafts são desfeitas por depleção de colesterol. (B) Em destaque, as frações 4-6 que correspondem com as lipid rafts da GEO.
1 2 3 4 5 6 7 8 9 10 11 12 13
CNPase
9-O-acetil GD3
G-A2B5
1 2 3 4 5 6 7 8 9 10 11 12 13
CNPase
9-O-acetil GD3
G-A2B5

62
MβCD Controle
Figura 31- Microscopia de fluorescência da GEO apresentando imunorreatividade para CNPase e flotilina-1. (A-D) Células tratadas com MβCD 20 mM por 30 minutos e (E-H) células controles. (A e E) Marcação da célula, fixada com paraformaldeído 4%, para o anticorpo monoclonal anti-CNPase, reagido com o anticorpo secundário conjugado a Alexa 488. (B e F) Marcação da célula para o anticorpo policlonal anti-flotilina-1, posterior incubação com anticorpo secundário biotinilado e reação deste com extravidina conjugada a Cy3. (C e G) Marcação do núcleo das células coradas com DAPI. (D e H) Sobreposição das imagens. Notar em D, a distribuição das duas proteínas ao longo do corpo celular (seta). Por vez, em H, a sobreposição das proteínas acontece em alguns pontos da célula (seta). Barra de calibração: 5 µm.
A
B
C
F
E
G
D H

63
5 Discussão
Este trabalho apresenta um estudo de caracterização dos microdomínios lipídicos de
membrana enriquecidos de colesterol e esfingolipídios (lipid rafts) da glia embainhante
olfatória (GEO). Constatamos que os marcadores das lipid rafts se localizam em frações
menos densas do gradiente de sacarose (fração 4-6), quando submetidos à metodologia de dot
blotting e marcação fluorescente para gangliosídeo GM1, proteínas flotilina-1 e caveolina-1.
Esses resultados enquadram-se nos critérios bioquímicos estabelecidos na literatura (Persaud-
Sawin et al., 2009): baixa quantidade de proteína e alta quantidade de colesterol. Do ponto de
vista morfológico, pelo método de contrastação negativa, constatamos que as lipid rafts
formam espontaneamente vesículas, ao passo que frações não-rafts formam aglomerados
macromoleculares de proteínas e/ou lipídios solubilizados pelo detergente Triton X-100.
Foram realizados imunoblottings para moléculas candidatas no envolvimento da migração da
GEO (gangliosídeos 9-O-acetil GD3 e os reconhecidos pelo anticorpo A2B5 e a CNPase),
resultando em marcações positivas nas frações características de rafts. Por fim, a distribuição
e/ou estrutura dos microdomínios lipídicos de membrana da GEO foram alterados, em
decorrência da depleção de colesterol pela droga metil-beta-ciclodextrina (MβCD).
5.1 Diversas metodologias são usadas para o isolamento das rafts
Diferentes metodologias são utilizadas para o isolamento desses microdomínios
lipídicos, sendo que o procedimento tradicional consiste na utilização do detergente não
iônico Triton-X100 a 4ºC. Sabe-se que nesta preparação as lipid rafts flutuam em gradiente de
5 a 30% de sacarose (Brown & Rose, 1992). Outros detergentes como o Tween 20, Lubrol
WX, Brij 58, Brij 96, Nonindet P40 e o CHAPS são também utilizados em diferentes
concentrações para o isolamento das rafts. Schuck e colaboradores (2003) levantaram
evidências de que as rafts preparadas em Triton X-100 ou CHAPS são fortemente

64
enriquecidas em esfingolipídios e colesterol, quando comparadas ao total da membrana. Em
contraste, a extração das membranas com Tween 20, Brij 98 ou Lubrol WX revelaram frações
de menor densidade das membranas com menor enriquecimento dos componentes
tradicionalmente observados em lipid rafts. Rafts preparadas pela solubilização de lisados
celulares com Brij 96 ou Brij 98 mostraram enriquecimento moderado de colesterol e
esfingomielina, a qual está particularmente mais enriquecida em preparados com Triton X-
100 ou CHAPS. Schuck e colaboradores (2003) descrevem ainda que estas diferenças
observadas refletem o grau de seletividade dos detergentes, onde o Triton X-100 e o CHAPS
se mostraram mais seletivos a lipídios não-rafts ao passo que o Tween 20 e o Brij 98 menos
seletivos.
5.2 Membranas resistentes a detergentes da GEO são positivas para marcadores das
lipid rafts
Frações de microdomínios lipídicos isolados por extração de detergentes submetidas a
gradiente de sacarose e a ultracentrifugação são tipicamente enriquecidas em esfingolipídios
saturados, colesterol e gangliosídeos. Por vez, dentre as proteínas consideradas marcadores
das rafts estão proteínas citoplasmáticas, como a flotilina e a caveolina, frequentemente
ancoradas à bicamada lipídica por isoprenilação (Brown & London, 1998, 2000). Essas duas
proteínas, em especial, conferem os microdomínios lipídicos um potencial endocítico
independente de clatrina (Anderson, 1998; Frick et al., 2007). Devido a este potencial,
patógenos como Vibrio cholera utilizam-se dessas estruturas, mais especificamente do
gangliosídeo GM1, como porta de entrada para as células. Dessa forma, a toxina do cólera
vem também sendo utilizada para a identificação das rafts, uma vez que concentram grandes
quantidades de GM1 (Radeva & Sharom, 2004).
As frações do gradiente de sacarose de lisados da GEO, realizado nesse trabalho,

65
revelaram que as frações 4, 5 e 6 apresentaram maior marcação de flotilina-1 e GM1, sendo
portanto, frações rafts (Figuras 14 e 16). A caveolina também mostrou-se concentrada nessas
regiões, embora frações mais pesadas (7, 8 e 9) tenham apresentado uma marcação mais
intensa, com um pico menos pronunciado de flotilina (Figura 18), sugerindo a existência de
microdomínios heterogêneos. Dados da literatura relatam à existência de lipid rafts não
caveolares, as quais podem eventualmente servir como um arcabouço inicial para a formação
de rafts caveolares (revisto por Ikonen, 2001). Dessa forma, a menor concentração de
caveolina nas frações 4, 5 e 6 em relação as frações 7, 8 e 9 pode representar estágios da
transição dinâmica entre microdomínios não caveolares e caveolares, embora estas últimas
frações sejam mais pesadas do que as comumente associadas às rafts. A presença de caveolina
também foi identificada em outras células neurais como astrócitos, oligodendrócitos e
microglia, embora acredite-se que neurônios maduros não as possuam (Cameron et al., 1997).
5.3 Perfil lipídico e dosagem protéica da membrana da GEO confirmam a presença de
lipid rafts
Pudemos observar que lisados da GEO submetidos ao gradiente de sacarose revelaram
que as frações resistentes a detergentes (4, 5 e 6) possuem uma elevada concentração de
colesterol e baixa densidade protéica, com uma razão máxima de 2:1 (respectivamente) na
fração 5 (Figura 21). Essa composição é característica das lipid rafts, como verificado por
Persaud-Sawin et al., 2009. A concentração de proteínas nos revela que apenas 43,58 µg/ml
estão localizadas nos microdomínios lipídicos da GEO. Por vez, dados na literatura são
bastante variáveis quanto à composição protéica, demonstrando uma diferença significativa
dos microdomínios lipídicos de diferentes células. Radeva & Sharom (2004) demonstraram
que em células leucêmicas basofílicas (RBL-2H3), a quantidade protéica não ultrapassava de
20 µg/ml nas frações resistentes a detergente Triton X-100. No entanto, Arvanitis e

66
colaboradores (2005) demonstraram que os microdomínios lipídicos de mielina apresentavam
concentração de proteínas de aproximadamente 300 µg/ml.
O enriquecimento em colesterol das rafts, observado na dosagem de colesterol e pela
análise de lipídio neutro por HPTLC (Figuras 21A e 22A), contribui para a menor fluidez
desses microdomínios lipídicos, o que é importante para a sua funcionalidade como
plataforma de sinalização. Essa funcionalidade é em grande parte atribuída ao recrutamento
seletivo de proteínas as rafts.
Outro ponto importante na análise lipídica é a presença fosfolipídios como:
fosfatidilcolina (PC) e esfingomielina (SM) apresentados nas rafts da GEO (Figura 22C). A
esfingomielina é um tipo de esfingolipídio presente nos microdomínios lipídicos, que age
concentrando moléculas sinalizadoras e também, como precursora de vários lipídios
bioativos. A presença desses fosfolipídios nas frações menos densas do gradiente de sacarose
também é observado em neurônios (Wu et al., 1997).
5.4 As lipid rafts isoladas formam espontaneamente vesículas
Neste trabalho constatamos que as rafts isoladas da GEO formam vesículas de
estrutura unilamelar com diâmetro que varia de 0,09 a 0,17 µm (Figura 20A). Esses
resultados estão de acordo com dados da literatura relatando que rafts isoladas de lisados
celulares de mamíferos formam espontaneamente vesículas com diâmetro variável (0,11-0,24
µm) (Radeva & Sharom, 2004). Essa formação de vesículas, no entanto, pode ser resultado da
ação do detergente, que estaria promovendo a fusão de rafts menores em estruturas de maior
diâmetro, possibilitando a formação das vesículas (Heerklotz, 2002). In situ, as rafts possuem
morfologia planar e incontínua, com interrupções de sua estrutura rígida por regiões de
membrana plasmática normal (Mayor & Maxfield, 1995).

67
5.5 Moléculas candidatas no envolvimento da migração se associam as rafts da GEO
A CNPase é uma proteína periférica da monocamada interna. O significado fisiológico
das relações da CNPase estabelece com o citoesqueleto e com as organelas celulares, assim
como sua função exata não são conhecidas. Alguns trabalhos sugerem que a CNPase associe-
se com a tubulina através das porções hidrofílicas e por meio de seus domínios hidrofóbicos,
desse modo poderia mediar a comunicação dos microtúbulos com a membrana plasmática
(Bifulco et al., 2002). É possível, portanto, que a CNPase esteja envolvida com sinalizações
membrano-esqueletais, incluindo movimentos direcionais. A constatação nesse trabalho de
que a CNPase se localiza preferencialmente em rafts (Figura 23) vai de acordo com dados na
literatura que associam esses microdomínios a eventos de adesão, importantes para migração
celular (Harris & Siu, 2002; Prag et al., 2007). Essa colocalização da CNPase e marcadores de
rafts foi da mesma forma observada em experimentos de caracterização desses
microdomínios lipídicos em oligodendrócitos que, tal como a GEO, são células de
propriedades embainhantes (Kim & Pfeiffer, 1999).
Farrer & Quarles (1999) observaram que os gangliosídeos 9-O-acetil GD3 e os
reconhecidos pelo anticorpo A2B5 são importantes para a migração de precursores de
oligodendrócitos. Sabe-se que o ambiente iônico proporcionado por gangliosídeos como o 9-
O-acetil GD3 é importante para plena ativação de componentes membranares envolvidos na
adesão celular (Mendez-Otero & Santiago, 2003). Além da expressão desses gangliosídeos, in
vitro, foi possível também constatar que a migração da GEO depende ao menos parcialmente
da atividade destes componentes uma vez que, o imunobloqueio dos mesmos levou a uma
redução na migração celular (dados de Santos-Silva e colaboradores não publicados). Desta
forma a migração da GEO parece compartilhar um mecanismo de controle comum com
neurônios e precursores de oligodendrócitos com capacidade migratória (Farrer & Quarles,
1999; Santiago et al., 2001; revisto por Mendez-Otero & Cavalcante, 2003). Desse modo, a

68
concentração desses gangliosídeos nas rafts da GEO, constatada neste trabalho, pode,
portanto, corroborar ainda mais o envolvimento desses microdomínios lipídicos em eventos
de migração e adesão desta célula (Figura 25).
5.6 A depleção de colesterol altera a distribuição de componentes protéicos e lipídicos
das rafts da GEO
A depleção de colesterol da membrana plasmática da GEO por meio da droga metil-
beta-ciclodextrina (MβCD) a 20 mM resulta na redistribuição de componentes enriquecidos
em microdomínios lipídicos (colesterol, gangliosídeo GM1, flotilina-1 e caveolina-1) de uma
forma mais homogênea ao longo do gradiente de sacarose (Figura 28). Algumas das funções
do colesterol na membrana plasmática são proporcionar a menor permeabilidade, bem como
manter o estado líquido ordenado dos microdomínios enriquecidos de esfingolipídios (Alberts
et al., 2002). Desse modo, a depleção de colesterol destruiria as rafts, ao facilitar a dispersão
de seus componentes na membrana plasmática (Zidovetzki & Levitan, 2007) de forma a
redistribuí-los de maneira mais randômica ou internalizando-os. Neste trabalho, a depleção de
colesterol foi na ordem de 22% (Figura 29A), o que pode ser ainda corroborado pela ausência
do material flutuante resistente ao detergente nas frações 4, 5 e 6 do gradiente de sacarose
(Figura 27). Em condições similares, diversos trabalhos obtiveram resultados semelhantes,
confirmando a destruição dos microdomínios lipídicos pela MβCD (Ilangumaran & Hoessli,
1998; DeBruin et al., 2005).
Além dos marcadores convencionais das lipid rafts, diversos componentes celulares
podem ter a sua distribuição afetada pela a destruição das rafts. Essa observação transformou
a MβCD em uma valiosa droga para o estudo da associação espacial/funcional de
componentes celulares a microdomínios de membrana (Cooper et al., 2003). Neste trabalho,
como já discutido, pudemos constatar que a proteína de mielina não-compacta CNPase, os

69
gangliosídeos 9-O-acetil GD3 e os reconhecidos pelo anticorpo A2B5 (G-A2B5) localizam-se
preferencialmente em rafts da GEO. Complementando esses resultados, constatamos que o
uso de MβCD resulta na redistribuição desses componentes de forma semelhante ao que
ocorreu com os marcadores das rafts mencionado anteriormente (Figura 30). Esses dados
ressaltam que os microdomínios lipídicos da GEO requerem de colesterol para sua formação e
função. Da mesma forma que observamos com a CNPase, trabalhos como o de DeBruin e
colaboradores (2005), também demonstraram que proteínas de mielina, além da CNPase, têm
sua distribuição afetada pelo uso de MβCD em oligodendrócitos. Juntos esses dados
evidenciam que as rafts apresentam um importante papel em células embainhantes. Com
relação aos gangliosídeos, sabe-se que o GM1 pode se dispersar na membrana após a
depleção de colesterol (Fujita et al., 2007). Contudo, ainda não existem relatos na literatura a
respeito da redistribuição dos gangliosídeos 9-O-acetil GD3 e os G-A2B5, observada neste
trabalho.
Por meio de ensaios de imunocitoquímicas, pudemos também avaliar o efeito da
depleção de colesterol sobre células individuais (Figura 31). Em uma situação controle,
observamos que a CNPase e a flotilina-1 colocalizam-se em regiões distintas da célula.
Células tratadas, por vez, apresentam um padrão de colocalização distribuída
homogeneamente pela célula. Estes resultados estão de acordo com nossos experimentos de
dot blotting, bem como com os dados de DeBruin e colaboradores (2005), ao demonstrar que
a distribuição espacial da CNPase pode ser afetada pela destruição das rafts.
5.7 Considerações finais
Este trabalho abordou a caracterização das lipid rafts da GEO. Essa célula vem sendo
objeto de muito interesse devido ao seu potencial uso em terapias celulares
neuroregenerativas. Pudemos constatar que as rafts da GEO se localizam nas frações 4, 5 e 6

70
no gradiente de sacarose e que a proteína de mielina não compacta CNPase, os gangliosídeos
9-O-acetil GD3 e G-A2B5 estão localizados nessas regiões (Figura 32). Tal fato indica que
tais componentes devam se associar as rafts, provavelmente em vias de sinalização
membrano-esqueletais. Constatamos ainda que a destruição das rafts resulta não só na
dispersão dos seus marcadores, como também na dispersão da CNPase e dos gangliosídeos
abordados, candidatos no envolvimento da migração da GEO. Dessa forma, esse trabalho
deve contribuir para o melhor entendimento do maquinário celular envolvido em vias de
sinalização membrano-esqueletais da GEO.
Figura 32- Resumo esquemático dos resultados apresentados nesta dissertação. Os microdomínios lipídicos da membrana da glia embainhante olfatória (GEO) pode apresentar colesterol, fosfolipídios (fosfatidilcolina e esfingomielina), esfingolipídios (gangliosídeos GM1, 9-O-acetil GD3 e os G-A2B5) e as proteínas flotilina, caveolina e CNPase.
• Fosfatidilcolina
• Esfingomielina
•Gangliosídeos: GM1,
•9-O-acetil GD3 e os G-A2B5

71
6 Conclusões
As marcações das proteínas flotilina-1 e caveolina-1, e do gangliosídeo GM1
(esfingolipídio) confirmam a presença das lipid rafts na glia embainhante olfatória (GEO);
A análise por M.E.T. revelou que as membranas resistentes a detergente do gradiente de
sacarose da GEO formam vesículas que variam de 0,09 a 0,17 µm de diâmetro;
As frações positivas para marcadores das lipid rafts apresentaram baixa quantidade de
proteína e alta concentração de colesterol, fosfatidilcolina e esfingomielina;
A presença da CNPase (proteína da mielina não-compacta), dos gangliosídeos 9-O-acetil-
GD3 e os reconhecidos pelo anticorpo A2B5 sugerem que as lipid rafts da GEO parecem
estar, pelo menos parcialmente, envolvidas na migração.
A depleção de colesterol por meio da droga metil-beta-ciclodextrina altera a distribuição dos
componentes dos microdomínios lipídicos da GEO.

72
7 Perspectivas
Com o intuito de dar continuidade aos estudos aqui mencionados, serão realizados os
seguintes experimentos:
Análise dos gangliosídeos das lipid rafts por HPTLC ;
Avaliar nas frações do gradiente de sacarose da GEO a expressão das proteínas lipid rafts
(flotilina-1 e caveolina-1) e da CNPase por western blotting;
Experimento com a sonda Laurdan para a verificação de formação de domínios nas células
controle e tratada com metil-beta-ciclodextrina (MβCD);
Experimentos funcionais de bloqueio das células com o uso de MβCD e quantificação da
taxa de migração;
Ensaios com citocalasina D e colchicina para o rompimento da estrutura e organização do
citoesqueleto visando o entendimento da via de sinalização ligada aos gangliosídeos
abordados nesta dissertação e as lipid rafts.

73
8 Referências Bibliográficas
Alberts, B., Lewis, J., Johnson, A., Raff, M., Roberts, K. & Walter, P. (2002) Molecular Biology of the Cell (Garland Science, New York).
Amur-Umarjee, S.G., Dasu, R.G. & Campagnoni, A.T. (1990) Temporal expression of myelin specific components in neonatal mouse brain cultures: evidence that 2’3’-cyclic nucleotide 3’-phosphodiesterase appears prior galactocerebroside. Dev. Neuroscience. 12: 251-262.
Anderson, R.G.W. (1998) The caveolae membrane system. Ann. Rev. Biochem. 67: 199-225.
Arvanitis, D.N., Min, W., Gong, Y., Heng, Y.M. & Boggs, J.M. (2005) Two types of detergent-insoluble, glycosphingolipid/cholesterol-rich membrane domains from isolated myelin. J. Neurochem. 94(6): 1696-710.
Au, E. & Roskams, A.J. (2003) Olfactory ensheating cells of the lamina propria in vivo and in vitro. Glia, 41: 224-236.
Babuke, T. & Tikkanen, R. (2007) Dissecting the molecular function of reggie/flotillin proteins. Eur. J. Cell Biol. 86(9): 525-532.
Barradas, P.C., Gomes, S.S. & Cavalcante, L.A. (1995) CNPase expression in the developing opossum brain stem and cerebellum. Neuroreport. 6: 289-292.
Bauer, M., Pelkmans, L. (2006) A new paradigm for membrane-organizing and -shaping scaffolds. FEBS Lett. 580(23): 5559-5564.
Bavari, S., Bosio, C.M., Wiegand, E., Ruthel, G., Will, A.B., Geisbert, T.W., Hevey, M., Schmaljohn, C., Schmaljohn, A. & Aman, M.J. (2002) Lipid raft microdomains: a gateway for compartmentalized trafficking of Ebola and Marburg viruses. J. Exp. Med. 195: 593-602.
Bickel, P.E., Scherer, P.E., Schnitzer, J.E., Oh, P., Lisanti, M.P. & Lodish, H.F. (1997) Flotillin and epidermal surface antigen define a new family of caveolaeassociated integral membrane proteins. J Biol Chem. 272: 13793–13802.
Bifulco, M., Laezza, C., Stingo, S. & Wolff, J. (2002) 2’,3’-Cyclic nucleotide 3’-phosphodiesterase: A membrane-bound, microtubule-associated proteins and membrane anchor for tubulin. Proc. Natl. Acad. Sci. 99: 1807–1812.
Bitman, J. & Wood, D.L. (1982) An improved copper reagent for quantitative sensitometric thin-layer chromatography of lipis. J. Liquid Chromatogr. 5: 1155-1162.
Bligh, E.G. & Dyer, W.J. (1959) A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37: 911-913.
Brown, D.A. & Rose, J.K. (1992) Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 68(3): 533-44.

74
Brown, D.A. & London, E. (1998) Functions of lipid rafts in biological membranes. Annu. Rev. Cell Dev. Biol. 14: 111-136.
Brown, D.A. & London, E. (2000) Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J. Biol. Chem. 275(23): 17221–17224.
Browman, D.T., Hoegg, M.B. & Robbins, S.M. (2007) The SPFH domain-containing proteins: more than lipid raft markers. Trends Cell Biol. 17: 394–402.
Cameron, P.L., Ruffin, J.W., Bollag, R., Rasmussen, H. & Cameron, R.S. (1997) Identification of caveolin and caveolin-related proteins in the brain. J. Neurosci. 17(24): 9520-35.
Chatterjee, S. & Mayor, S. (2001) The GPI-anchor and protein sorting. Cell. Mol. Life Sci. 58: 1969-1987.
Chuah, M.I., Cossins, J.M., Woodhall, E., Tennet, R., Nash, G. & West, A.K. (2000) Glial growth factor 2 induces proliferation and structural changes in ensheathing cells. Brain Res., 857: 264-274.
Constantine-Paton, M., Blum, A.S., Mendez-Otero, R. & Barnstable, C.J. (1986) A cell surface molecule distributed in a dorsoventral gradient in the perinatal rat retina. Nature. 324: 459-462.
Corley Mastick, C., Sanguinetti, A.R., Knesek, J.H., Mastick, G.S. & Newcomb, L,F. (2001) Caveolin-1 and a 29-kDa caveolin-associated protein are phosphorylated on tyrosine in cells expressing a temperature-sensitive v-Abl kinase. Exp. Cell Res. 266(1):142-154.
Couet, J., Li, S., Okamoto, T., Ikezu, T. & Lisanti, M.P. (1997) Identification of peptide and protein ligands for the caveolin-scaffolding domain. Implications for the interaction of caveolin with caveolae-associated proteins. J. Biol. Chem. 272(10): 6525–6533.
DeBruin, L.S., Haines, J.D., Wellhauser, L.A., Radeva, G., Schonmann, V., Bienzle, D. & Harauz, G. (2005) Developmental partitioning of myelin basic protein into membrane microdomains. J. Neurosci. Res. 80(2): 211-25.
De Angelis, D.A. & Braun, P.E. (1996) 2’,3’-Cyclic nucleotide 3’-phosphodiesterase binds to actin-based cytoskeletal elements in an isoprenylation-independent manner. J. Neurochem. 67: 943-951.
Doetsch, F. & Alvarez-Buylla, A. (1996) Network of tangential pathways for neuronal migration in adult mammalian brain. Proc. Natl. Acad. Sci. U.S.A. 93(25): 14895-14900.
Doucette, J.R. (1984) The glial cells in the nerve fiber layer of the rat olfactory bulb. Anat. Rec. 210: 385-391.
Dubois, C., Manuguerra, J. C., Hauttecoeur, B. & Maze, J. (1990) Monoclonal antibody A2B5, which detects cell surface antigens, binds to ganglioside GT3 (II3 (NeuAc)3LacCer) and to its 9-O-acetylated derivative. J. Biol.Chem. 265(5): 2797-2803.

75
Drummond, G.I., Iyer, N.T. & Keith, J. (1962) Hydrolysis of ribonucleoside 2’, 3’-cyclic phosphates by a diesterase from brain. J. Biol. Chem., 237: 3535-3539.
Dyer, C.A. & Benjamins, J.A. (1989) Organization of oligodendroglial membrane sheets. I: association of myelin Basic protein and 2’3’-cyclic nucleotide 3’-phosphohydrolase with cytoskeleton. J. Neurosci. Research 24: 201-211.
Fantini, J., Garmy, N., Mahfoud, R. & Yahi, N. (2002) Lipid rafts: structure, function and role in HIV, Alzheimer’s and prion diseases. Expert. Rev. Mol. Med.4(27): 1-22.
Farrer, G. & Quarles, R. H. (1999) GT3 and its o-acetylated derivative are the principal A2B5-reactive gangliosides in culture O2A lineage cells and are down-regulated along with O-acetyl GD3 during differentiation to oligodendrocytes. J. Neurosci. Research. 57: 371-380.
Firestein, S. (2001) How the olfactory system makes sense of scents. Nature. 413(6852): 211-218.
Fishman, P.H. (1982) Role of Membrane Gangliosides in the Binding and Action of Bacterial Toxins. J. of Membrane Biology 69: 85-97.
Fivaz, M., Abrami, L. & van der Gool, F.G. (2000) Pathogens, toxins, and lipid rafts. Protoplasma 212: 8-14.
Franceschini, I.A. & Barnett, S.C. (1996) Low-affinity NGF-receptor and E-N-CAM expression define two types of olfactory nerve ensheathing cells that share a common lineage. Dev Biol. 173(1): 327–343.
Franklin, R.J.M. & Barnett, S.C. (1996) Do olfactory glia have advantages over Schwann cell-like myelination following transplantation of an olfactory bulb-ensheathing cell line into areas of demyelination in the adult CNS. Glia. 17: 217-224.
Franssen, E.H., de Bree, F.M. & Verhaagen, J. (2007) Olfactory ensheathing glia: their contribution to primary olfactory nervous system regeneration and their regenerative potential following transplantation into the injured spinal cord. Brain Res. Rev. 56(1): 236-58.
Frick, M., Bright, N.A., Riento, K., Bray, A., Merrified, C. & Nichols, B.J. (2007) Coassembly of flotillins induces formation of membrane microdomains, membrane curvature, and vesicle budding. Curr. Biol. 17(13): 1151-6.
Fujita, A., Cheng, J., Hirakawa, M., Furukawa, K., Kusunoki, S. & Fujimoto, T. (2007) Gangliosides GM1 and GM3 in the living cell membrane form clusters susceptible to cholesterol depletion and chilling. Mol. Biol, Cell. 18(6): 2112-22.
Gómez-Moutón, C., J. Abad, E. Mira, R. Lacalle, E. Gallardo, S. Jiménez-Baranda, I. Illa, A. Bernad, S. Mañes, & C. Martínez-A. (2001) Segregation of leading-edge and uropod components into specific lipid rafts during T cell polarization. Proc. Natl. Acad. Sci. USA. 98: 9642–9647.

76
Graziadei, P.P.C. & Monti-Graziadei, G.A. (1979) Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J. Neurocytol. 8: l-l8.
Hakomori, S. (1990) Bifunctional role of glycosphingolipids. Modulators for transmembrane signaling and mediators for cellular interactions. J. Biol. Chem. 265: 18713-18716.
Halpern, M. & Martinez-Marcos, A. (2003) Structure and function of the vomeronasal system: an update. Progress in Neurobiology 70: 245-318.
Harris, T.J. & Siu, C.H. (2002) Reciprocal raft-receptor interactions and the assembly of adhesion complexes. Bioessays 24: 996-1003.
Hazarika, P., Dham, N., Patel, P., Cho, M., Weidner, D., Goldsmith, L. & Duvic, M. (1999) Flotillin 2 is distinct from epidermal surface antigen (ESA) and is associated with filopodia formation. J. Cell Biochem. 75: 147-159.
Heerklotz, H. (2002) Triton promotes domain formation in lipid raft mixtures. Biophys. J. 83(5): 2693-701.
Henion, T.R. & Schwarting, G.A. (2007) Patterning the developing and regenerating olfactory system. J Cell Physiol. 210(2): 290-7.
Hinman, J.D., Chen, C.D., Oh, S.Y., Hollander, W. & Abraham, C.R. (2008) Age-dependent accumulation of ubiquitinated 2',3'-cyclic nucleotide 3'-phosphodiesterase in myelin lipid rafts. Glia. 56(1): 118-133.
Ikonen, E. (2001) Roles of lipid rafts in membrane transport. Curr. Opin. Cell Biol. 13(4): 470-477.
Ilangumaran, S. & Hoessli, D.C. (1998) Effects of cholesterol depletion by cyclodextrin on the sphingolipid microdomains of the plasma membrane. Biochem. J. 335(Pt 2): 433-440.
Johnson, D.M., Illig, K.R., Behan, M. & Haberly, L.B. (2000) New features of connectivity in piriform cortex visualized by intracellular injection of pyramidal cells suggest that "primary" olfactory cortex functions like "association" cortex in other sensory systems. J Neurosci. 20(18): 6974-82.
Kaftz, K.W. & Greer, C.A. (1999) Olfactory ensheathing cells promote neurite extension from embryonic olfactory receptor cells in vitro. Glia. 25(2): 99-110.
Kawooya, J.K. & Law, J.H. (1988) Role of lipophorin in lipid transport to the insect egg. J. Biol. Chem. 263: 8748-8753.
Keyvan-Fouladi, N., Raisman, G. & Li, Y. (2005) Delayed repair of corticospinal tract lesions as an assay for the effectiveness of transplantation of Schwann cells. Glia. 51: 306-311.

77
Kim, T. & Pfeiffer, S. E. (1999) Myelin glycosphingolipid/cholesterol-enrichedmicrodomains selectively sequester the non-compact myelin proteins CNP and MOG. J. Neurocytol 28: 281-293.
Kurzchalia, T.V., Dupree, P., Parton, R.G., Kellner, R., Virta, H., Lehnert, M. & Simons, K. (1992) VIP 21, A 21-kDa membrane protein is an integral component of trans-Golgi-network-derived transport vesicles. J. Cell Biol. 118: 1003-1014.
Kurzchalia, T.V. & Parton, R.G. (1999) Membrane microdomains and caveolae. Curr. Opin. Cell Biol. 11, 424-431.
Lang, D.M., Lommel, S., Jung, M., Ankerhold, R., Petrausch, B., Laessing, U., Wiechers, M.F., Plattner, H. & Stuermer, C.A. (1998) Identification of reggie-1 and reggie-2 as plasmamembrane-associated proteins which cocluster with activated GPI-anchored cell adhesion molecules in non-caveolar micropatches in neurons. J. Neurobiol. 37(4): 502-523.
Langhorst, M.F., Reuter, A. & Stuermer, C.A.O. (2005) Scaffolding microdomains and beyond: the function of reggie/flotillin proteins. Cell. Mol. Life Sci. 62: 2228–2240.
Lappe-Siefke, C., Goebbels, S., Gravel, M., Nicksch, E., Lee, J., Braun, P.E.. Griffiths, I. R.. & Nave, K.A. (2003) Disruption of Cnp1 uncouples oligodendroglial functions in axonal support and myelination. Nature Genetics. 33: 366-374.
Lee, J., Gravel, M., Zang, R., Thibault, P. & Braun, P.E. (2005) Process outgrowth in oligodendrocytes is mediated by CNP, a novel microtubule assembly myelin protein. J. Cell Biol. 170: 661–673.
Lee, J., O’Neill, R.C., Park, M.W., Gravel, M. & Braun, P.E. (2006) Mitochondrial localization of CNP2 is regulated by phosphorylation of the N-terminal targeting signal by PKC: Implications of a mitochondrial function for CNP2 in glial and non-glial cells. Mol. Cell. Neurosci. 31: 446–462.
Levine, C. & Marcillo, A. (2008) Regarding several points of doubt of the structure of the olfactory bulb: as described by T. Blanes. Anat. Rec. (Hoboken). 291(7):751-62.
Li, S., Okamoto, T., Chun, M., Sargiacomo, M., Casanova, J.E., Hansen, S.H., Nishimoto, I. & Lisanti, M.P. (1995) Evidence for a regulated interaction between heterotrimeric G proteins and caveolin. J. Biol. Chem. 270(26): 15693–15701.
Li, Y., Sauve, Y., Li, D., Lund, R.D. & Raisman, G. (2003) Transplanted olfactory ensheathing cells promote regeneration of cut adult rat optic nerve axons. J. Neurosci. 23: 7783-7788.
Lisanti, M.P., Tang, Z. & Sargiacomo, M. (1993)Caveolin forms a hetero-oligomeric protein complex that interacts with an apical GPI-linked protein: implications for the biogenesis of caveolae. J. Cell Biol. 123: 595-604.
Lledo, P.M., Saghatelyan, A. & Lemasson, M. (2004) Inhibitory interneurons in the olfactory bulb: from development to function. Neuroscientist. 10(4): 292-303.

78
Loyd, K.O. &Furukawa, K. (1998) Biosynthesis and functions of gangliosides: recent advances. Glycoconj. 7: 627-636.
Margrie, T.W., Sakmann, B. & Urban, N.N. (2001) Action potential propagation in mitral cell lateral dendrites is decremental and controls recurrent and lateral inhibition in the mammalian olfactory bulb. Proc. Natl. Acad. Sci. U.S.A. 98(1): 319-24.
Marin-Padilla, M.. & Amieva, M.R. (1989) Early neurogenesis of the mouse olfactory nerve: Golgi and electron microscopic studies. J. Comp. Neurol. 288: 339-352.
Mayor, S. & Maxfield, F.R. (1995) Insolubility and redistribution of GPI-anchored proteins at the cell surface after detergent treatment. Mol. Biol. Cell. 6(7): 929–944.
Meisami, E. & Bhatnagar, K.P. (1998) Structure and diversity in mammalian accessory olfactory bulb. Microsc. Res. Tech. 43(6): 476-99.
Mendez-Otero R, Schlosshauer B, Barnstable CJ & Constantine-Paton M (1988). A developmentally regulated antigen associated with neural cell and process migration. Journal of Neuroscience, 8: 564-579.
Mendez-Otero, R. & Cavalcante, L.A. (2003) Functional role of gangliosides in neuronal motility. Prog. Mol, Subcell. Biol. 32: 97-124.
Mendez-Otero, R. & Santiago, M.F. (2003) Functional role of a specific ganglioside in neuronal migration and neurite outgrowth. Braz. J. Med. Biol. Res. 36(8): 1003-1013.
Miragall, F., Kadmon, G., Husmann, M. & Schachner, M. (1988) Expression of cell adhesion molecules in the olfactory system of the adult mouse: presence of the embryonic form of NCAM. Dev Biol. 129(2):516-531.
Munro S. (2003) Lipid rafts: elusive or illusive? Cell. 115(4): 377-388.
Murata, M., Peränen, J., Schreiner, R., Wieland, F., Kurzchalia, T.V. & Simons, K. (1995) VIP21/caveolin is a cholesterol-binding protein. Proc. Natl. Acad. Sci. U.S.A. 92(22): 10339-10343.
Nash, H.H., Borke, R.C. & Anders, J.J. (2001) New method of purification for establishing primary cultures of ensheathing cells from the adult olfactory bulb. Glia. 34: 81-87.
Parolini, I., Sargiacomo, M., Galbiati, F., Rizzo, G., Grignani, F., Engelman, J.A., Okamoto, T., Ikezu, T., Scherer, P.E., Mora, R., Rodriguez-Boulan, E., Peschle, C. & Lisanti, M.P. (1999) Expression of caveolin-1 is required for the transport of caveolin-2 to the plasma membrane. Retention of caveolin-2 at the level of the Golgi complex. J Biol Chem, 274: 25718-25725.
Parton, R.G. & Richards, A.A. (2003) Lipid rafts and caveolae as portals for endocytosis: new insights and common mechanisms. Traffic 4, 724-738.
Persaud-Sawin, D.A., Lightcap, S. & Harry, G.J. (2009) Isolation of rafts from mouse brain tissue by a detergent-free method. J. Lipid Res. 50(4):759-67.

79
Peterson, B.R. (2005) Synthetic mimics of mammalian cell surface receptors: prosthetic molecules that augment living cells. Org. Biomol. Chem. 3(20): 3607-12.
Pietzsch, J. (2004) Mind the membrane. Horizon Symposia. pp, 1-4. London, UK: Nature publishing group.
Pike, L.J. (2004) Lipid rafts: heterogeneity on the high seas. J. Biochem. 378: 281-292.
Pike, L.J. (2006) Rafts defined: a report on the keystone symposium on lipid rafts and cell function. J. Lipid Res. 47, 1597–1598.
Pitha, J., Irie, T., Sklar, P.B. & Nye, J.S. (1988) Drug solubilizers to aid pharmacologists: amorphous cyclodextrin derivatives. Life Sci. 43(6): 493-502.
Pixley, S.K. (1992) The olfactory nerve contains two populations of glia, identified both in vivo and in vitro. Glia. 5(4): 269-284.
Prag, S., Parsons, M., Keppler, M.D., Ameer-Beg, S.M., Barber, P., Hunt, J., Beavil, A.J., Calvert, R., Arpin, M., Vojnovic, B. & Ng, T. (2007) Activated ezrin promotes cell migration through recruitment of the GEF dbl to lipid rafts and preferential downstream activation of Cdc42. Mol. Biol. Cell. 18(8): 2935-2948.
Santiago, M.F., Berredo-Pinho, M., Costa, M.R., Gandra, M., Cavalcante, L.A. & Mendez-Otero, R. (2001). Expression and function of ganglioside 9-O-acetyl GD3 in postmitotic granule cell development. Molecular and Cellular Neurosciences, 17: 488-499.
Santos-Benito, F.F. & Ramón-Cueto, A. (2003) Olfactory ensheathing glia transplantation: a therapy to promote repair in the mammalian central nervous system. Anat. Rec. B. New Anat. 271(1): 77-85.
Santos-Silva, A. & Cavalcante, L.A. (2001) Expression of the non-compact myelin protein 2’3’-cyclic nucleotide 3’-phosphodiesterase (CNPase) in the olfactory bulb ensheathing glia from explant cultures. Neurosci. Research. 40: 189-193.
Scherer, P.E., Okamoto, T., Chun, M., Nishimoto, I., Lodish, H.F. & Lisanti, M.P. (1996) Identification, sequence and expression of caveolin-2 defines a caveolin gene family. Proc. Natl. Acad. Sci. U.S.A. 93: 131-135.
Scherer, P.E., Lewis, R.Y., Volonte, D., Engelman, J.A., Galbiati, F., Couet, J., Kohtz, D.S., van Donselaar, E., Peters, P. & Lisanti, M.P. (1997) Cell-type and tissue-specific expression of caveolin-2. Caveolins 1 and 2 co-localize and form a stable hetero-oligomeric complex in vivo. J Biol Chem. 272(46): 29337-29346.
Scherer, S.S., Braun, P.E., Grinspan, J., Collarini, E., Wang, D.Y. & Kamholz, J. (1994) Differential regulation of the 2’,3’-cyclic nucleotide 3’-phosphodiesterase gene during oligodendrocyte development. Neuron 12: 1363-1375.
Schuck, S., Honsho, M., Ekroos, K., Shevchenko, A. & Simons K. (2003) Resistance of cell membranes to different detergents. Proc. Natl. Acad. Sci. U.S.A. 100(10): 5795–5800.

80
Shin, J.S. & Abraham, S.N. (2001) Caveolae as portals of entry for microbes. Microbes Infect. 3(9):755-61.
Schroeder, R., London, E. & Brown, D. (1994) Interactions between saturated acyl chains confer detergent resistance on lipids and glycosylphosphatidylinositol (GPI)-anchored proteins: GPI-anchored proteins in liposomes and cells show similar behavior. Proc. Natl Acad. Sci. USA 91: 12130–12134.
Schwarting, G.A., Kostek, C., Ahmad, N., Dibble, C., Pays, L. & Püschel, A.W. (2000) Semaphorin 3A is required for guidance of olfactory axons in mice. J. Neurosci. 20(20): 7691-7697.
Schwob, J.E. (2002) Neural regeneration and the peripheral olfactory system. Anat. Rec. 269(1): 33-49.
Simons, K. & Ehehalt, R. (2002) Cholesterol, lipid rafts, and disease. J. Clin. Invest. 110: 597-603.
Simons, K. & Ikonen, E. (1997) Functional rafts in cell membranes. Nature 387, 569–572.
Simons, K. & Toomre, D. (2000) Lipid rafts and signal transduction. Nat. Rev. Mol. Cell Biol. 1: 31-41.
Simona, K. & van Meer, G. (1988) Lipid sorting in epithelial cells. Biochemistry. 27(17): 6197-6202.
Simons, K. & Vaz, W. L. (2004) Model systems, lipid rafts, and cell membranes. Annu. Rev.Biophys. Biomol. Struct. 33: 269-295.
Singer, S.J. & Nicolson, G.L. (1972) The fluid mosaic model of the structure of cell membranes. Science, 175: 720-731.
Solis, G.P., Hoegg, M., Munderloh, C., Schrock, Y., Malaga-Trillo, E., Rivera-Milla, E. & Stuermer, C.A. (2007). Biochem. J. 403: 313–322.
Sprinkle, T.J. (1989) 2’,3’-Cyclic nucleotide 3’-phosphodiesterase, an oligodendrocyte-Schwann cell and myelin-associated enzyme of the nervous system. CRC Crit. Rev. Neurobiol. 4: 235-301.
Radeva, G. & Sharom, F. J. (2004) Isolation and characterization of lipid rafts with different properties from RBL-2H3 (rat basophilic leukaemia) cells. J. Biochem. 380: 219–230.
Rajendran, L., Masilamani, M., Solomon, S., Tikkanen, R., Stuermer, C.A., Plattner, H. & Illges, H. (2003) Asymmetric localization of flotillins/reggies in preassembled platforms confers inherent polarity to hematopoietic cells. Proc. Natl. Acad. Sci. USA 100: 8241-8246.
Ramón-Cueto, A. & Nieto-Sampedro, M. (1992) Glial cells from adult rat olfactory bulb: immunocytochemical properties of pure cultures of ensheathing cells. Neurosci. 47(1): 213-220.

81
Ramón-Cueto, A., Pérez, J. & Nieto-Sampedro, M. (1993) In vitro enfolding of olfactory neurites by p75 NGF receptor positive ensheathing cells from adult rat olfactory bulb. Eur. J. Neurosci. 5(9): 1172-1180.
Ramón-Cueto, A. & Nieto-Sampedro, M. (1994) Regeneration into the spinal cord of transected dorsal root axons is promoted by ensheathing glia transplants. Exp. Neurol. 127(2): 232-244.
Ramón-Cueto, A. & Valverde, F. (1995) Olfactory bulb ensheathing glia: a unique cell type with axonal growth-promoting properties. Glia. 14(3): 163-173.
Ramón-Cueto, A. & Avila, J. (1998) Olfactory ensheathing glia: properties and function. Brain Res Bull. 46(3): 175-187.
Ramón-Cueto, A., Cordero, M.L., Santos-Benito, F.F. & Avila, J. (2000) Functional recovery of paraplegic rats and motor axon regeneration in their spinal cords by olfactory ensheathing glia. Neuron. 25: 425-435.
Ren, S., Scarsdale, J.N., Ariga, T., Zhang, Y., Klein, R.A., Hartmann, R., Kushi, Y., Egge, H.& Yu, R.K. (1992) O-acetylated gangliosides in bovine buttermilk. Characterization of 7-O-acetyl, 9-O-acetyl, and 7,9-di-O-acetyl GD3. J. Biol. Chem. 267(18): 12632-8.
Reynolds, R. & Wilkin, G.P. (1988) Development of macroglial cells in rat cerebellum. II. An in situ immunohistochemical study of oligodendroglial precursors to mature myelination cell. Development. 102: 409-425.
Reynolds, R., Carey, E.M. & Herschkowitz, N. (1989) Immunohistochemical localization of myelin basic protein and 2’3’-cyclic nucleotide 3’-phosphohydrolase in flattened membrane expansions produced by cultured oligodendrocyte. Neuroscience 28 (1): 181-188.
Ruiz, J.I. & Ochoa, B. (1997) Quantification in the subnanomolar range of phospholipids and neutral lipids by monodimensional thin-layer chromatography and image analysis. J. Lipid Res. 38: 1482-1489.
Tang, Z., Scherer, P.E., Okamoto, T., Song, K., Chu, C., Kohtz, D.S., Nishimoto, I., Lodish, H.F. & Lisanti, M.P. (1996) Molecular cloning of caveolin-3, a novel member of the caveolin gene family expressed predominantly in muscle. J. Biol. Chem. 271: 2255-2261.
Taylor, C.M., Coetzee, T., Pfeiffer, S.E. (2002) Detergent-insoluble glycosphingolipid/cholesterol microdomains of the myelin membrane. J. Neurochem. 81: 993-1004.
Thuret, S., Moon, L.D. & Gage, F.H. (2006) Therapeutic interventions after spinal cord injury. Nat Rev Neurosci. 7(8): 628-643.
Tsukada, Y. & Kurihara, T. (1992) 2’,3’-Cyclic nucleotide 3’-phosphodiesterase : Molecular characterization and possible functional significance. In: Myelin – Biology and Chemistry. Martenson, R.E. (ed.) CRC Press, Boca Raton, 449-480.

82
Unwin, N., Henderson, R. (1984) The structure of proteins in biological membranes. Sci. Am. 250: 78-94.
Valverde, F. & Lopez-Mascaraque, L. (1991) Neuroglial arrangements in the olfactory glomeruli of the hedhog. J. Comp. Neurol. 307(4):658-671.
Valverde, F., Santacana, M. & Heredia, M. (1992) Formation of na olfactory glomerulus: morphological aspects of development and organization. Neurosci. 49(2): 255-275.
Vassilieva, E.V., Ivanov, A.I., Nusrat, A. (2009) Flotillin-1 stabilizes caveolin-1 in intestinal epithelial cells. Biochem. Biophys. Res. Commun. 379(2): 460-465.
Volonte, D., Galbiati, F., Li, S., Nishiyama, K., Okamoto, T., Lisanti, M.P. (1999) Flotillins/ cavatellins are differentially expressed in cells and tissues and form a hetero-oligomeric complex with caveolins in vivo. J Biol Chem. 274: 12702–12709.
Way, M. & Parton, R. (1995) M-caveolin: a muscle-specific caveolinrelated protein. FEBS Lett, 376: 108-112.
Whitfeld, P.R., Heppel, L.A. & Markham, R. (1955) The enzymic hydrolysis of ribonucleoside-2’:3’ phosphates. Biochem. J., 60: 15-19.
Wiegandt, H., Gangliosides, in: Wiegandt H. (Ed.), Glycolipids, Elsevier Science Publishers, Amsterdam, 1985, pp.199-260.
Williams, T.M. & Lisanti, M.P. (2004) The Caveolin genes: from cell biology to medicine. Ann Med. 36(8): 584-595.
Woodhall, E., West, A.K. & Chuah, M.I. (2001) Cultured olfactory ensheathing cells express nerve growth factor, brain-derived neurotrophic factor, glia cell line-derived neurotrophic factor and their receptors. Mol. Brain Research. 88: 203-213.
Wu, C., Butz, S., Ying, Y-s & Anderson, R.G.W. (1997) Tyrosine kinase receptors concentrated in caveolae-like domains from neuronal plasma membrane. J. Biol. Chem. 272: 3554–59.
Yamada, E. (1955) The fine structure of the gall bladder epithelium of the mouse. J. Biophys. Biochem. Cytol., 1: 445-458.
Yin, X., Peterson, J., Gravel, M., Braun, P.E. & Trapp, B.D. (1997) CNP overexpression induces aberrant oligodendrocyte membranes and inhibits MBP accumulation and myelin compaction. J Neurosci Res. 50(2): 238-247.
Yoshino, J.E., Dinneen, M.P., Sprinkle, T.J. & DeVries, G.H. (1989) Localization of 2’3’-cyclic nucleotide 3’-phosphohydrolase on cultured Schwann cells. Brain Research. 325: 199-203.
Yu, R.K., Bieberich, E., Xia, T. & Zeng, G. (2004) Regulation of ganglioside biosynthesis in the nervous system. J. Lipid Res. 45(5): 783-793.

83
Zheng, Y.Z., Berg, K.B. & Foster, L.J. (2009) Mitochondria do not contain lipid rafts, and lipid rafts do not contain mitochondrial proteins. J. Lipid. Res. 50(5): 988-998.
Zidovetzki, R. & Levitan, I. (2007) Use of cyclodextrins to manipulate plasma membrane cholesterol content: evidence, misconceptions and control strategies. Biochim. Biophys. Acta. 1768(6): 1311-1324.
Zlatkis, A., Zak, B. & Boyle, A.J. (1953) A new method for the direct determination of serum
cholesterol. J. Lab. Clin. Med. 41(3): 486-492.
Site consultado:
www.chemblink.com

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo