can the hair follicle become a model for studying selected aspects
TRANSCRIPT

Can the Hair Follicle Become a Model for StudyingSelected Aspects of Human Ocular Immune Privilege?
Michael Kinori,1,2 Jennifer E. Kloepper,2 and Ralf Paus2,3
Immune privilege (IP) is important in maintaining ocularhealth. Understanding the mechanism underlying this dynamicstate would assist in treating inflammatory eye diseases. De-spite substantial progress in defining eye IP mechanisms, be-caue of the scarcity of human ocular tissue for research pur-poses, most of what we know about ocular IP is based onrodent models (of unclear relevance to human eye immunol-ogy) and on cultured human eye–derived cells that cannotfaithfully mirror the complex cell–tissue interactions that un-derlie normal human ocular IP in situ. Therefore, accessible,instructive, and clinically relevant human in vitro models areneeded for exploring the general principles of why and how IPcollapses under clinically relevant experimental conditions andhow it can be protected or even restored therapeutically.Among the few human IP sites, the easily accessible and abun-dantly available hair follicle (HF) may offer one such surrogatemodel. There are excellent human HF organ culture systemsfor the study of HF IP in situ that instructively complement invivo autoimmunity research in the human system. In this arti-cle, we delineate that the human eye and HF, despite theirobvious differences, share key molecular and cellular mecha-nisms for maintaining IP. We argue that, therefore, humanscalp HFs can provide an unconventional, but highly instruc-tive, accessible, easily manipulated, and clinically relevant pre-clinical model for selected aspects of ocular IP. This essay is anattempt to encourage professional eye researchers to turn theirattention, with appropriate caveats, to this candidate surrogatemodel for ocular IP in the human system. (Invest OphthalmolVis Sci. 2011;52:4447–4458) DOI:10.1167/iovs.10-7154
It is now obvious that the eye possesses immune privilege(IP) characteristics that are important for eye health. The
credit for coining this term belongs to Peter Medawar,1 whoshowed over a half century ago that skin allografts are notrejected by the host’s immune system when transplanted het-erotopically into defined anatomic sites, such as the rabbit eyeor brain. However, even 130 years ago, Van Dooremaal,2 aDutch ophthalmologist, had already discovered that mouseskin grafts show significantly prolonged survival if transplantedinto the anterior chamber (AC) of a dog’s eye. These pioneer-
ing studies have opened the door for robust research in thefield of IP. Consequently, ocular IP has become a subject ofmajor recent interest, and its fundamental importance in in-flammatory eye diseases is now widely accepted.3–15
However, as every investigative ophthalmologist painfullyexperiences sooner or later, as a tissue on which to perform invitro research, the human eye is an exceptionally rare com-modity that is very difficult to come by. In the rare cases inwhich eyes are enucleated, the damage due to trauma, infec-tion, or tumor growth raises questions as to how useful suchtissue is in helping us to understand the physiology of humanocular IP. With very few exceptions (see below), therefore,most currently available data and concepts on ocular IP arebased on the systematic analysis of rodent models.16–26 Giventhe very substantial immunologic differences between rodentand human systems,27,28 it is inherently problematic to extrap-olate from rodent to human eyes. Moreover, cultured humaneye–derived cells, yet another source of ocular IP re-search,29–33 are also problematic, since IP is an in situ statebased on complex cell–tissue interactions and not a conditiondisplayed by isolated cell populations in vitro.15,34,35
Therefore, our understanding of human ocular IP remainsrather limited, and all extrapolation from rodent and cell cul-ture work must be interpreted with caution.
Thus, good, clinically relevant human in vitro models areurgently needed to enable the study of the general principles ofwhy and how IP collapses and how it can be protected or evenrestored in situ. Although the search for such human surrogatemodels meets with evident obstacles, since no other organ isquite like the eye, at least some aspects of human ocular IP maybe studied in other, more accessible and more abundantlyavailable human tissues.
SEARCHING FOR A SURROGATE PRECLINICAL MODEL
OF OCULAR IP
The eye is not the only mammalian site of IP. Other sitesinclude parts of the testis and ovary, the adrenal cortex, partsof the brain, the fetomaternal placental unit, the hamster cheekpouch, and probably the proximal nail matrix.34,36–41 Notably,the human hair follicle (HF) also qualifies as an IP site34 (fordebate, see below). This miniorgan is unique among all otherIP sites, in that it is massively distributed over the human bodyand is highly accessible to experimental analysis and manipu-lation. Our integument has approximately 5 million HFs andthus a correspondingly vast number of potential (human) IPorgans.42 Moreover, there are excellent human HF organ cul-ture systems that can instructively complement in vivo re-search for the study of autoimmunity in the human sys-tem.43–45
Since common mechanisms of IP in different organs havebeen elucidated,46–48 an obvious question is whether one IPsite can hold lessons for other, less easily explored sites. Al-though the IP of human HFs has been much less well studiedthan that of the eye, the relative ease with which human HFs
From the 1Department of Ophthalmology, Chaim Sheba MedicalCenter, Tel-Hashomer, Israel; the 2Department of Dermatology, Uni-versity of Lubeck, Lubeck, Germany; and the 3School of TranslationalMedicine, University of Manchester, Manchester, United Kingdom.
Supported in part by a “Cluster of Excellence” grant fromDeutsche Forschungsgemeinschaft (DFG) (“Inflammation at Inter-faces”) and by a DFG Graduate College grant (“Autoimmunity”) (RP).
Submitted for publication December 31, 2010; revised April 4,2011; accepted April 7, 2011.
Disclosure: M. Kinori, None; J.E. Kloepper, None; R. Paus,None
Corresponding author: Michael Kinori, Department of Ophthal-mology, Chaim Sheba Medical Center, Tel-Hashomer, 52621, Israel;[email protected].
Perspectives
Investigative Ophthalmology & Visual Science, June 2011, Vol. 52, No. 7Copyright 2011 The Association for Research in Vision and Ophthalmology, Inc. 4447
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

can be obtained, microdissected, organ cultured, and immuno-logically manipulated in vitro (see below) invites one to exploithuman HF organ culture44 as an unusual, but highly instructiveand clinically relevant preclinical research model (e.g., forfollowing up in vitro studies of ocular IP in rodent models,before entering into clinical trials on the human eye).
Following in the footsteps of earlier suggestions along thisvein,34,49 the current essay attempts to shed some light onboth the similarities and important differences between ocularand HF IP and to attract eye researchers and clinical ophthal-mologists to systematically employ, with appropriate cautionand circumspection, the human HF as an accessible, relativelyeasily manipulated surrogate model for carefully selected as-pects of ocular IP model.
IMMUNOLOGIC HOMEOSTASIS, IGNORANCE,AND PRIVILEGE
Before critically exploring the usefulness of human HFs as acandidate surrogate model for ocular IP, it helps to rememberthat many tissues have their own mechanisms for preservingthe immunologic status quo for proper functioning (i.e., im-mune homeostasis).7 For example, the lung and the gut,which are chronically exposed to foreign antigens that mayincite inflammatory responses must allow gas exchange andfood processing, respectively, without provoking undesiredlevels of inflammation.7 Another important concept in thiscontext is immunologic ignorance, which emphasizes the keyrole of anatomic barriers and peculiarities in a given anatomicsite (e.g., the absence of patent lymphatics), which preventsthe entry of immune cells resulting in graft rejection.8,50
In contrast, immune privilege classically describes tissuesites within which foreign tissue grafts can survive for ex-tended periods, whereas similar grafts placed in conventionalsites are acutely rejected by the host.46,50,51 Today, the term IPis generally understood in a much broader sense and indicatesthe presence of multiple active mechanisms for preventing theinduction and expression of both innate and adaptive immuneresponses.7,8,11,34,36,50
Although there are phenomenologic indications that the HFevades some and actively suppresses other potentially autoag-gressive immune responses (see below), for evident method-ological reasons, rejection and survival of heterologous tissuetransplants within this tiny miniorgan have not yet been stud-ied. (In fact, due to insufficiently refined microsurgery and
microinjection techniques, previous attempts in our laboratoryto inject melanocytes from C57BL/6 mice into the vibrissaehair bulb of white, immunocompetent Balb/c mice have failedmiserably.) Thus, one can only lament that Billingham’s land-mark experiment and its visionary interpretation remain theonly currently available functional evidence that HFs can in-deed shelter heterotransplants from immune rejection. He ob-served that, while donor melanocytes within the epidermis arerapidly eliminated, heterologous epidermal melanocytes showlong-term survival if they manage to escape into the anagenhair bulbs of (white) host guinea pigs, which thus began toproduce black hair shafts.34,52,53
OCULAR IMMUNE PRIVILEGE: BASIC CONCEPTS AND
CURRENT PERSPECTIVES
IP has turned out to be a complex and dynamic tissue state andthe list of “players” involved in it is ever-growing (Table 1, Fig. 1).One key mechanism of ocular IP, AC associated immune devi-ation (ACAID), was identified by Kaplan and Streilein et al.81–84
ACAID means that injection of antigens into the AC of rodents(and even of monkeys28) produces a stereotypic systemic im-mune response that is selectively deficient in antigen-specificdelayed-type hypersensitivity (DTH), whereas other conven-tional effector modalities of immunity (such as cytotoxic Tcells and non–complement-fixing antibody isotypes) are pre-served.28,85
ACAID Has Ocular and Systemic Pathways
It is thought that antigens inoculated into the eye are processedin a distinctive fashion by stromal antigen-presenting cells(APCs) of the iris and ciliary body.14,86 This phenomenonappears to be largely under the control of transforming growthfactor (TGF)-�, a key immunosuppressive cytokine in the aque-ous humor.61,62,87 In fact, in mice, a deficit of total TGF-�2 inaqueous humor correlates with loss of ACAID.51 These APCsthen emigrate from the eye by traversing the trabecular mesh-work (TM) and directly enter the blood stream to reach thespleen.88 Access to the blood stream (without an access to thelymphatic system thorough the uveoscleral pathway) seems tobe crucial for the induction of ACAID. Indeed, monkeys treatedwith topical prostaglandins (which redirects a substantial frac-tion of aqueous humor into the uveoscleral path89) failed toinduce ACAID.28
TABLE 1. Important Characteristics of Ocular and Hair Follicle Immune Privilege
Eye Hair Follicle
Specific location of IP Anterior chamber, iris and ciliary body, subretinalspace7,22,54,55
Hair bulb, hair bulge34,56,57
Cyclic phenomena No YesResults of IP collapse Corneal transplant rejection, immune mediated
microbial keratitis and uveitis7,8,58,59AA, lichen planopilaris, scleroderma, skin
manifestations of SLE, and GVHD34
ACAID Yes NoStrong expression of immunoinhibitory
molecules�-MSH60; TGF�261,62,63*; TSP121,64*; VIP65; SOM23;
MIF66; CGRP67; CRP32; PEDF19; IDO30; IL-1Ra68;GITRL16; TRAIL25; B7–2(CD86)69; cortisol62
�-MSH, TGF�1, IL-10, IGF-134,43,44; MIF,TGF�2, CD200, IDO57; cortisol70;downregualtion of MICA71; TSP-1?72,CGRP? (currently being investigated)
Downregulation of APC molecules Low MHC class Ia, weak MHC class Ib, no MHCclass II cells24,59,73–76
Low MHC class Ia, weak MHC class Ib(HLA-E and G), no MHC classII34,44,56,57,77
Apoptosis induction of lymphocytes FasL(CD95L)78�sFasL(sCD95L)79;B7-H1(CD86)20,29; B7-H359
FasL?80
AA, alopecia areata; ACAID, anterior chamber-associated immune deviation; APC, antigen presenting cell; GVHD, graft versus host disease; IP,immune privilege; SLE, systemic lupus erythematosus. The abbreviations for proteins are as described in Figure 1.
* Main inductors of ACAID.
4448 Kinori et al IOVS, June 2011, Vol. 52, No. 7
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

The eye-derived APCs reach the marginal zone in thespleen. There, a complex dialog involving multiple cells, in-cluding B cells, CD4� NKT cells, CD4� T cells, �� T cells, andCD8� T cells, ensues.58,90–92 When the latter recognize anti-gens presented by eye-derived APCs and/or marginal zone Bcells, they differentiate into ACAID-inducing regulatory T cells(ACAID-Tregs).58,59 CD4� ACAID-Tregs prevent the activationand differentiation of antigen-specific Th1 effector cells,85
whereas CD8� ACAID-Tregs inhibit the local function of effec-tor T cells (Th1 and Th2).15,93,94
Two other elements, are also important in the induction ofACAID: the thymus and the sympathetic nervous system.59
Although the thymus is an important source of the NKT cellsthat are needed for the splenic phase of ACAID,95 the sympa-thetic nervous system is believed to have a role in the gener-ation of the NKT-cell population that eventually enters thespleen and participates in the induction of CD8� ACAIDTregs.59,96
Clinical Aspects of ACAID
In mouse models, ACAID has been demonstrated to be in-volved in various clinical scenarios, such as acceptance ofcorneal transplants, autoimmune uveitis, acute retinal necrosis(ARN) in a fellow eye that experienced herpes virus infectionin the anterior segment, and progression of intraocular malig-nant melanoma.39 This mechanism may be the eye’s way ofprotecting its vital functions from immunopathogenic injury.97
ACAID has been widely studied in mice, rats, guinea pigs, andrabbits, implying that it is not a phenomenon restricted tolaboratory rodents.98,99 Significant progress was made whenEichhorn et al.28 demonstrated that ACAID can also occur inprimates. Even so, it should not be taken for granted that theseresults (although more convenient than those obtained in ro-dents) can be extrapolated to conclusions about the humaneye. Evidence compatible with the existence of an ACAID-likeresponse in humans indeed has been shown in patients withARN.100 Nevertheless, the evidence that these ACAID mecha-
nisms also apply to the human condition remains rather cir-cumstantial, since definitive experimental proof of the exis-tence of ACAID in humans would require (e.g., the ethicallyproblematic transfer of regulatory T cells from one individualto another).100 Thus, whether all the chief characteristics ofACAID established in animal models also occur in humansremains unknown.
IP and the Posterior Segment of the Eye
If one views the eye as an extension of the brain, it is notsurprising that there is also a close relationship between ocularIP and the nervous system. Indeed, ocular IP may mainly be theresult of neuroimmune interactions.8,101 The retina itself rep-resents a highly organized neuronal tissue that produces manyimmunosuppressive molecules generated by neurons and glialcells. As a cell layer, RPE can suppress the activation of by-stander T cells via soluble inhibitory factors such asTGF�21,102,103 and can induce apoptosis of activated T cells.104
Moreover, in mice, the (neonatal) RPE105 and the neuronalretina106 display inherent IP. It is now known that the vitreousand subretinal space are also IP sites107 and that antigensplaced in these sites can even lead to the induction of anACAID-like response.22,54,55 In this context, it is interesting tonote that (in mice) IP disruption occurs when RPE cells aredamaged by laser burns.18 This finding has raised the troublingquestion of whether laser-treated eyes are at increased risk ofocular inflammation, up to the collapse of eye IP.
Current Perspectives
The concept that “excessive“ ocular IP can facilitate life-threat-ening infections and the actual presence of autoimmune uveitisand corneal allograft rejection underscores the importance ofviewing IP as a relative, rather than absolute status, which cansometimes be bypassed.98 Indeed, it has been proposed thatthe immune system can “decide” that, sometimes, preservationof life supersedes preservation of vision (e.g., if there is a
FIGURE 1. Immune privilege of the eye and anagen VI hair follicle: key features. �-MSH, �-melanocytestimulating hormone; APC, antigen presenting cell; APM, arrector pili muscle; CGRP, calcitonin gene-related peptide; CRP, complement regulatory proteins; DP, dermal papilla; E, epidermis; FasL, Fas ligand;GITRL, glucocorticoid-induced TNF receptor family-related protein ligand; HLA, human leukocyte antigen;HS, hair shaft; IDO, indolamine 2,3-dioxygenase; IGF, insulin-like growth factor; IL-10, interleukin 10;IL-1Ra, interleukin 1 receptor antagonist; IRS, inner root sheath; MHC, major histocompatibility complex;MICA, MHC class I chain-related A gene; MIF, macrophage migrating inhibitory factor; ORS, outer rootsheath; PEDF, pigment epithelium derived factor; RPE, retinal pigment epithelium; sFasL, soluble Fasligand; SG, sebaceous gland; SOM, somatostatin; TGF, transforming growth factor; TRAIL, TNF-relatedapoptosis-inducing ligand; TSP-1, thrombospondin 1; and VIP, vasoactive intestinal peptide.
IOVS, June 2011, Vol. 52, No. 7 The Hair Follicle as a Model for Ocular Immune Privilege 4449
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

life-threatening ocular infection, such as trachoma, river blind-ness, or herpes simplex virus keratitis, resulting in ocular IPcollapse and inflammatory eye destruction7).
This example illustrates that IP mechanisms must generallybe carefully balanced, since we are continuously exposed topotentially deleterious environmental agents, including micro-organisms and antigenic insults; and yet, most humans do notgo blind. Moreover, corneal transplants are the least-rejectedamong all organ transplants,39,108 despite the use of HLA-unmatched transplants with minimal immunosuppression.109
This is probably due, not only to the anatomic characteristicsof the corneal tissue, but also to its low antigenicity.59,110
New frontiers in ocular IP research include the role ofocular IP in eye tumor development and progression. Giventhat IP can be acquired by tumor cells to evade immunesurveillance,111 it is interesting to note that modulating IP bythe injection of cytotoxic Fas ligand (FasL) in the vitreouscavity can prevent neovascularization in a mouse model ofchoroidal neovascularization.112
Thus, instructive experimental models in the human systemare urgently needed that allow deeper insights into IP biologyand pathology and that facilitate experimental manipulations.This is where the HF, one of the defining features of mamma-lian species, enters our vision.
EVIDENCE IN SUPPORT OF HF IP
It has been four decades since Billingham52 discovered that thehair bulb provides a special milieu that permits transplantedallogeneic cells (namely, donor melanocytes) to escape limita-tion by the host immune system while others are attacked.52
Black skin epidermis transplanted onto skin beds of geneticallyincompatible white guinea pigs quickly lost its pigmentation (asign that the foreign melanocytes had been rejected), whereasblack hair shafts soon thereafter began to pierce the (nowwhite) epidermis. This result indicates that at least some donormelanocytes had survived in the host hair bulbs and had re-sumed their transfer of melanosomes to HF keratino-cytes.34,52,53
Unfortunately, since then, no additional functional evidenceof the existence of HF IP has been published, perhaps becauseavailable pointers that HFs are a site of IP have long beenignored by the immunologic and transplantation research com-munities. Recently, however, HF IP has attracted more wide-spread interest,34,56,80,113–116 and there is an increasing com-munity of HF immunology authorities that has embraced theconcept of that the HF enjoys a relative IP and that a collapseof HF IP is of critical importance in alopecia areata (AA), one ofthe most frequent human autoimmune diseases,43,80,114–116
whereas a collapse of the putative IP of the epithelial stem cellarea of the HF (bulge)57 may be important in the pathogenesisof cicatricial alopecia.117
Compared with the eye, our understanding of the func-tional state of resident immune cells within the HF epithelium(Langerhans cells, T cells) and in the HF mesenchyme (macro-phages, mast cells) is still very limited, and current HF immu-nology concepts are largely based on immunophenomenologicanalyses. However, a few important facts have surfaced that,taken together, strongly support the concept that defined com-partments of the HF represent sites of relative IP. Some keyarguments can be summarized as follows (for full discussion,see Ref. 34).
MHC Class I and�2-Microglobulin Downregulation
A classic feature of IP sites is the downregulation of MHC classIa expression.34,44,116 The lower (proximal) epithelial com-
partments of the HF epithelium, the anagen hair bulb, shows astriking downregulation of both MHC class Ia and associated�2-microglobulin gene and protein expression.44,57,77,118
Since MHC class Ia-stabilization by �2-microgolubin is criticalfor the proper presentation of MHC class I–dependent anti-gens,27 any MHC class I molecules that may still be expressedin the anagen hair bulb probably cannot effectively presentautoantigens. In humans and mice, the HF’s stem cell zone, thebulge, even expresses nonclassic MHC class I molecules (MHCclass Ib molecules, such as Qa-2 and HLA-E), which inhibit, forexample, NK cell activities.57,77,119 To the best of our knowl-edge, no healthy mammalian tissue has been described thus farthat exhibits this phenomenon without enjoying relative IP.
Local Generation of Potent Immunosuppressants
A key feature of all recognized IP sites is that they expresspotent, locally generated immunoinhibitory molecules.7,120,121
Therefore, it is important to note that the anagen hair bulb andeven the bulge prominently express potent immunosuppres-sants such as TGF�, �-melanocyte-stimulating hormone (�-MSH), IL-10, and others.34,44,122,123
Functionally Impaired Langerhans Cells
Given the importance of APCs in ocular IP (see above), it isintriguing to note that, in striking contrast to the APCs of the distalHF epithelium (i.e., the upper outer root sheath), the very fewintraepithelial Langerhans cells that are detectable ultrastructur-ally or by CD1a immunohistochemistry in the proximal anagenhair bulb of human scalp HFs do not express detectable MHCclass II or I molecules.57,123 Even the distal outer root sheath ofhuman HFs harbors immature Langerhans cell populations.124
That at least all professional APCs in the proximal human HFepithelium lack evidence of full antigen-presentation capacityperfectly fits the characteristic impaired antigen presentationby APCs in IP sites.
Autoreactive CD8� T Cells Are Key Protagonistsin HF Autoimmunity
Since AA is a T-cell-mediated, organ-specific autoimmune dis-ease,125 it offers an excellent model for probing the functionalrelationship between autoreactive T cells and the putative HF IP.AA also promises pointers to the functional relevance of HF IP.
T lymphocytes isolated from human scalp lesions and ex-panded in vitro with homogenates of HFs reproduce AA lesionswhen transferred into scalp explants in SCID mice.126 ThatCD8� T cells, but not CD4� T cells alone, can produce AAlesions43,127 indicates that a prior collapse of HF IP must haveoccurred, which exposes previously sequestered, MHC classI–presented autoantigens to CD8� T cells. The same is alsoseen in the best-characterized mouse model of AA128 (see alsoRef. 116). This, in turn, suggests that the striking downregula-tion of MHC class Ia and �2-microglobulin in healthy anagenhair bulbs is functionally important.
As in most autoimmune diseases, identification of theepitopes that trigger the autoimmune response remains a ma-jor goal. In AA, much current interest centers on melanocyte-associated antigens: Melanocyte peptide epitopes (such asGp100-derived G9–209 and G9–280 and MART-1 (27–35))injected into autologous lesional human scalp grafts on SCIDmice induce AA lesions.129 Moreover, skin-derived CD8� Tcells obtained from AA patients co-cultured with MAGE3 showa significant increase in intracellular interferon (IFN)-� expres-sion compared with the control.130 IFN�, in turn, is the mostpotent stimulator of ectopic MHC class Ia expression identifiedso far.34,44
Since melanocyte-associated, MHC class I–presented au-toantigens recognized by CD8� T cells27 are key immune
4450 Kinori et al IOVS, June 2011, Vol. 52, No. 7
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

targets in vitiligo and melanoma, these findings in AA patientsand animal models provide functional evidence that the ecto-pic expression and presentation of MHC class I–presentedautoantigens within the HF entails the danger of autoaggressiveimmune responses against the HF, if cognate, autoreactiveCD8� T cells are present. This finding strongly supports theconcept that the prominent downregulation of MHC class Iaand �2-microglobulin in healthy anagen HFs is critical to ward-ing off deleterious immune attacks on the HF.34,131
NK Cell Activities Appear to Be Suppressed inHealthy HFsSince NK cells are primed to recognize and eliminate cells withabsent or low MHC class I expression,27,71,132–138 immunolog-ically privileged HF compartments constitute a basic problemin self-/non–self-discrimination and self-tolerance.139 Onewould expect that MHC class I-negative or MHC class I ‘‘low”anagen HFs are under constant attack by NK cells. However,this is clearly not the case, since very few perifollicular NKcells can ever be found around healthy human anagen HFs.123
In contrast, in AA, CD56� NK cells prominently aggregatearound HFs and show an increased expression of NKG2D (NKcell-activating receptor) and decreased expression of KIR-2D2/2D3 (NK cell-inhibitory receptor).71 Moreover, macrophagemigrating inhibitory factor (MIF) may suppress NK cells in andaround healthy HFs71 (just like in the eye66). In contrast,excessive NK cell stimulation by NKG2D-activating ligandssuch as MHC class I chain–related A gene (MICA) by the HFepithelium71 and/or other MICA-related NKG2D ligands114 inand around AA HFs may contribute to HF IP collapse. There-fore, the failure to adequately suppress undesired NK cellsactivities directed against MHC class I–negative HF cells duringanagen may be an important additional element in AA patho-genesis.56 A recent genome-wide association study114 identi-fied several genetic susceptibility loci for AA. Among these,significant associations include the ULBP genes, which encodeactivating ligands for NKG2D. Normally, ULBP3 is not presentin HFs, but ULBP3 proteins were abundant in and aroundhuman HFs affected by AA. NKG2D/ULBP3 engagement, andthus inappropriate NK cell stimulation, may contribute to thedevelopment of AA.
Taken together, this combination of immunophenomeno-logic in situ observations and functional data from the relevantmodel disease (AA) constitutes sufficient evidence to suggestthat HFs do indeed enjoy relative IP.
A unique, key feature of IP in the anagen hair bulb is that itis temporary: The HF epithelium rhythmically generates, main-tains, and deconstructs an area of relative IP in the region ofthe HF, which is present only during a defined segment of thehair cycle—that is, the anagen phase (growth stage)—butabsent during HF regression (catagen) and the “resting” phase(telogen).34,44,122,123 In contrast, the bulge, the HF’s seat ofepithelial and melanocyte stem cells, seems to continuouslyenjoy a relative IP.57,140 In this anatomic HF landmark, MHCclass Ia, �2-microglobulin, and MHC class II molecules aredownregulated, whereas MHCIb (HLA-E) is upregulated. Inaddition, immunoinhibitory molecules, like the “no danger”signal CD200,117,141 �-MSH, MIF, and indoleamine-2,3-dioxyge-nase (IDO) are markedly overexpressed in the bulge region.57,142
All these are features of ocular IP as well.30,60,66,73,143
Thus, within the microcosmos of HF immunology, the rel-ative IP of the bulge may actually be more closely related toocular IP than that of the anagen hair bulb. Bulge IP collapseand the subsequent immunologically mediated destruction ofepithelial HF stem cells may play a key role in the pathogenesisof irreversible, scarring alopecia,117 just as ocular IP collapsecan irreversibly destroy the eye (instead, bulb IP collapse in AAtypically only induces reversible HF damage34,43).
WHY DOES THE HF NEED IMMUNE PRIVILEGE?
As vision is one of the most important qualities and survivalrequirements of most living creatures, it is easy to understandconceptually why the eye must be protected against autoag-gressive immune attacks, especially in its delicate, apparentlynonregenerating compartments (i.e., corneal endothelium andretinal cells). But what about the HF? Which selection advan-tage might mammals have had during evolution for establishingan area of IP in HF?
Although this may have been different for humanoids andprehistoric early humans, in our current climates and cultures,hair is clearly dispensable for human survival and propagationof the species. However, during �99% of the total duration ofmammalian evolution, an environmentally perfectly adaptedhair coat seems to have been vital for numerous of our mam-malian ancestors and their reproduction (suffice it here toenvision the poor survival and reproduction chances of ahairless polar bear, Arctic fox, or seal). Given that the HF is oneof the most frequent targets of immune-mediated tissue in-jury,34,144 the HF may therefore have established its IP as asafeguarding system against immune injury of this importantminiorgan. Viewed from this angle, ocular and HF IP may bothbe necessary (even though this is much less evident for thelatter than for the former).
SIMILARITIES AND DIFFERENCES BETWEEN OCULAR
AND HF IP
Thus, ocular and HF IP show some striking similarities—namely, in the bulge region of the HF:
● Classic MHC class I and �2-microglobulin are downregu-lated, which renders cells relatively invisible to CD8� cyto-toxic T cells.34,44,57,66,74,145
● APCs are both sparse and functionally impaired (e.g., theylack MHC class II expression).34,44,57,59,74
● IFN-� exposure causes ectopic upregulation of MHC classIa and II expression in cells lining the AC and in theHF.44,59,146,147
● Nonclassic MHC class I molecules (e.g., HLA-E and -G) aredownrebulated. Together with MIF, they are known to inhibita potential attack of NK cells on MHC class Ia (i.e., HLA-A, -B,and -C)–negative cells.57,73,75,119
● There is a strong expression of similar immunoinhibitorymolecules (see Table 1 and Fig. 1 for details).
In these specific areas, the human HF may well serve as anattractive surrogate model for ocular IP (see below).
Of course, there are also many important differences be-tween follicular and ocular IP that one needs to keep in mindwhen studying the HF as a surrogate model. Mainly, an ACAID-like response has not been demonstrated (yet) to be associatedwith antigens introduced into the HF. Also, Fas–FasL interac-tions, which seem to be an important element of ocular IP,78
are unlikely to play a major role in HF IP.34,49,123 Interestingly,however, FasL is indeed significantly decreased in lesional skinof AA patients, compared with nonlesional skin.80
Other apparent dissimilarities between ocular and HF IPmay also be less pronounced than one may be inclined tothink. Since neuroimmune interactions play a key role in ocularIP (see above), it is reasonable to ask whether these have anyrole in HF IP. Although this question has so far only beenaddressed very incompletely, the HF does represent a proto-typic neuroectodermal–mesodermal tissue interaction systemand is one of the most densely and intricately innervated of allperipheral tissues.148 In fact, HF biology has multiple neurobi-ological dimensions along what has been termed the “brain–HF
IOVS, June 2011, Vol. 52, No. 7 The Hair Follicle as a Model for Ocular Immune Privilege 4451
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

axis” 149 (for a review, see Ref. 150). Namely, HF development,growth and pigmentation are regulated in part by neurotro-phins and neuropeptides, whereas HF-derived neurotrophinscontrol HF innervation. In addition, human scalp HFs even displaya fully functional, peripheral equivalent of the central hypotha-lamic–pituitary–adrenal (HPA) stress response axis.44,151 It wouldbe unreasonable to expect that this brain–HF axis plays no role inHF IP.
Indeed, psychoemotional stress causes prominent perifol-licular neurogenic inflammation and hair growth inhibition inmice, which depends on mast cells, substance P, and nervegrowth factor (NGF) and goes along with phenomenologicindications of HF IP collapse.152–156 Mice subjected to noisestress show dense inflammatory cell infiltrates around theirHFs, as early as 24 hours after exposure, with an increase in thenumber and activation of perifollicular mast cells as well asMHC class II–positive inflammatory cell clusters. Noise stressalso increases the number of intradermal dendritic cells andinduces their maturation.152,155,157,158
Of special interest here is substance P, which is released inresponse to stress by sensory skin nerves, plays a key role in thecutaneous neuroimmune network and influences immune cellfunctions through the neurokinin-1 receptor (NK-1R).153,159 Inmurine AA, NK-1R is prominently expressed on CD8� lympho-cytes and macrophages that accumulate around lesional HFs.Thus, currently available murine data suggest that substance P andNK-1R are important elements in the pathogenesis of in autoim-mune hair loss and the associated collapse of HF IP.159
Moreover, in organ-cultured human scalp HFs, substance Pdirectly induces HF IP collapse, as evidenced by ectopic MHCclass I and class II expression.45 Although the role of substanceP in ocular IP has not yet been systematically explored, theimmunoinhibitory neuropeptide calcitonin gene-related pep-tide (CGRP), which is co-expressed with substance P in sen-sory skin nerves, is involved in the maintenance of ocular IP.67
Preliminary evidence from our laboratory suggests that CGRPmay exert a similar protective function in the context of hu-man HF IP (Kinori et al., manuscript in preparation). Thus, it isquite likely that neuroimmune interactions are an importantcomponent, not only of ocular, but also of HF IP.
HOW MAY INSIGHTS FROM HUMAN HF IPGENERATE THERAPEUTIC BENEFITS FOR CLINICAL
OPHTHALMOLOGY?
Although it would be unreasonable to claim that most aspectsof ocular IP can be investigated in human HFs in vitro, wepropose here that selected IP-related insights from human HForgan culture could be put to excellent clinical use in ophthal-mology.
Figure 2 illustrates how relatively easy it is to microdissectand organ culture human scalp HFs obtained, for example,from excess scalp skin during routine face-lift surgery. In these,IP collapse can easily be induced by the proinflammatorycytokine IFN�. This effect induces rapid, massive, ectopicMHC class Ia and II expression in the epithelium of normalanagen (stage VI) HFs147 (see Fig. 2), thus seriously endanger-ing maintenance of the HF IP (see above). The same phenom-enon can also be induced by adding substance P to the HFculture medium,45 or in vivo by injecting IFN� into the backskin of mice with HFs in the anagen stage of the hair cycle.147
Thus, human HF organ culture permits one to screen forcandidate agents that effectively downregulate IFN�-inducedectopic MHC class I expression in human anagen HFs. A “pro-tection” or (perhaps more important) “restoration” assay de-sign can be chosen: In the former, the candidate IP protectantis added to the medium before IFN� is introduced, whereas
candidate “IP restoration” agents can be tested by adding themafter IFN� administration. In fact, three immunomodulatorsknown to be locally produced in the anagen hair bulb—�-MSH,TGF�1, and insulin-like growth factor 1 (IGF-1)122,163,164—areall capable of downregulating ectopic MHC class Ia expression,on both the protein and the mRNA level, in the IP restorationassay design.44 This human organ culture assay, therefore, iswell-suited as a clinically relevant preclinical screening systemto identify novel candidate IP-restoring or IP-protecting agents.Once identified, these can then further be explored as candi-date therapeutics for ocular IP protection and restoration.
WHICH SPECIFIC ASPECTS RELEVANT TO OCULAR IPCAN BE STUDIED IN HUMAN HF ORGAN CULTURE?
The human HF is hardly suitable as a surrogate model forstudying ACAID or for evaluating the effects of test agents onthe delicate retinal neuronal tissue. However, given that the HFmay be more dispensable than any other human organ (privi-leged or not), it offers investigators interested in IP (in the eyeand elsewhere) an unparalleled opportunity to directly studyand manipulate a complex but easily accessible and widelyavailable human IP site. In this site, the following specificquestions that are directly relevant to ocular IP may be studiedin situ:
1. How is MHC class Ia, Ib, and II and �2-microglobulinexpression regulated in situ in a normal human neuroec-todermal–mesodermal interaction unit?
2. How can (experimentally induced) ectopic upregulationof these molecules be effectively downregulated again?
3. Vice versa, how can the local expression of IP-protec-tive, immunoinhibitory molecules that are also of rele-vance in ocular IP (e.g., IDO; immunoinhibitory neuro-peptides, such as �-MSH, vasoactive intestinal peptide[VIP], and CGRP, and TGF�1, IGF-1, and CD200) beeffectively upregulated in a normal human neuroecto-dermal–mesodermal interaction unit?
4. Psychoemotional stress may be implicated in the relapseof anterior autoimmune uveitis.165 Shouldn’t it then bepossible to exploit human HF organ culture to furtherexplore in this human miniorgan the direct impact ofwell-defined stress mediators (including neuropeptideslike VIP that have not yet been studied in a hair researchcontext, but are important in ocular IP) on key IP char-acteristics, such as MHC Ia/�2-microglobulin expressionand the local generation of immunoinhibitory com-pounds?
5. What is the relative contribution of human NK cells,NKT cells, intraepithelial T cells, Langerhans cells, andmast cells to IP (e.g., via local tissue interactions withthe epithelium)? For example, a recent new concept inmurine AA pathobiology suggests that some NK cellsubpopulations, as opposed to IFN-�-secreting CD49b�
T-cell subsets, may actually award relative protectionfrom AA development.125
6. How do drugs or operative techniques that are alreadyused in the management of inflammatory eye diseasesaffect human HF IP? This may help to predict desired andundesired effects on ocular IP, whose study in humaneyes would require enucleation and is thus essentiallyimpossible. For example, in mice, retinal laser burn ab-rogates IP in both the burned and nonburned eye.18
Applying laser burns to scalp HFs in vitro may indicatewhether laser treatment is likely to exert similar effectson human IP. Also, at least some underlying mechanismsof action could be studied in HF organ culture, buthardly in human eyes.
4452 Kinori et al IOVS, June 2011, Vol. 52, No. 7
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018
7. Along the same vein, can HFs from a given patient withautoimmune ocular disease be used to predict the likelyresponse of that patient to the intended therapy, as apotential means to predict whether that therapeutic in-tervention is likely to restore or protect ocular IP?
8. IP in the proximal HF epithelium is a cyclic phenome-non that appears to be restricted to a defined segment ofthe hair cycle. Although the human eye is not thought toundergo cyclic transformations in adult life, one won-ders whether ocular IP also possesses some cyclic char-acteristics—for example, being maximal or minimal dur-ing certain periods on a circadian or perennial timescale. Could it be that autoimmune uveitis preferentiallyrelapses during such hypothetical periods of constitu-tively “minimal ocular IP”?
ADULT EPITHELIAL STEM CELLS AND IMMUNE
PRIVILEGE: YET ANOTHER EYE–HAIR CONNECTION?
Stem cells rank among the most exciting current researchfrontiers in experimental and clinical ophthalmology. Althoughbeyond the scope of this essay, it therefore should at least bementioned briefly that the study of adult human epithelial stemcells (eSCs) in the bulge region of the HF166–170 may alsobenefit ophthalmology research. After all, at least in rodents,HF-derived eSCs can differentiate into cells with a cornealepithelial phenotype when given appropriate stimuli.171–173
On this background, it is interesting to note that physiolog-ical concentrations of thyroid hormones enhance expressionof CD200 on human HF eSCs.174 This important immunoin-hibitory and tolerogenic surface molecule175 is a crucial ele-ment of bulge IP maintenance, since its targeted knockout inmice causes massive inflammation and irreversible HF destruc-tion.57,117,141
Therefore, one might learn from HF-associated eSCs howcell-based therapies for the treatment of ocular disease couldbe engineered so as to reduce the risk of immune rejection orundesired immune deviation by progenitor cells introducedinto the human eye, for example, by promoting their expres-sion of CD200. This could, for example, become important inthe treatment of limbal stem cell deficiency (LSCD)173 andage-related macular degeneration (AMD),176 especially if one
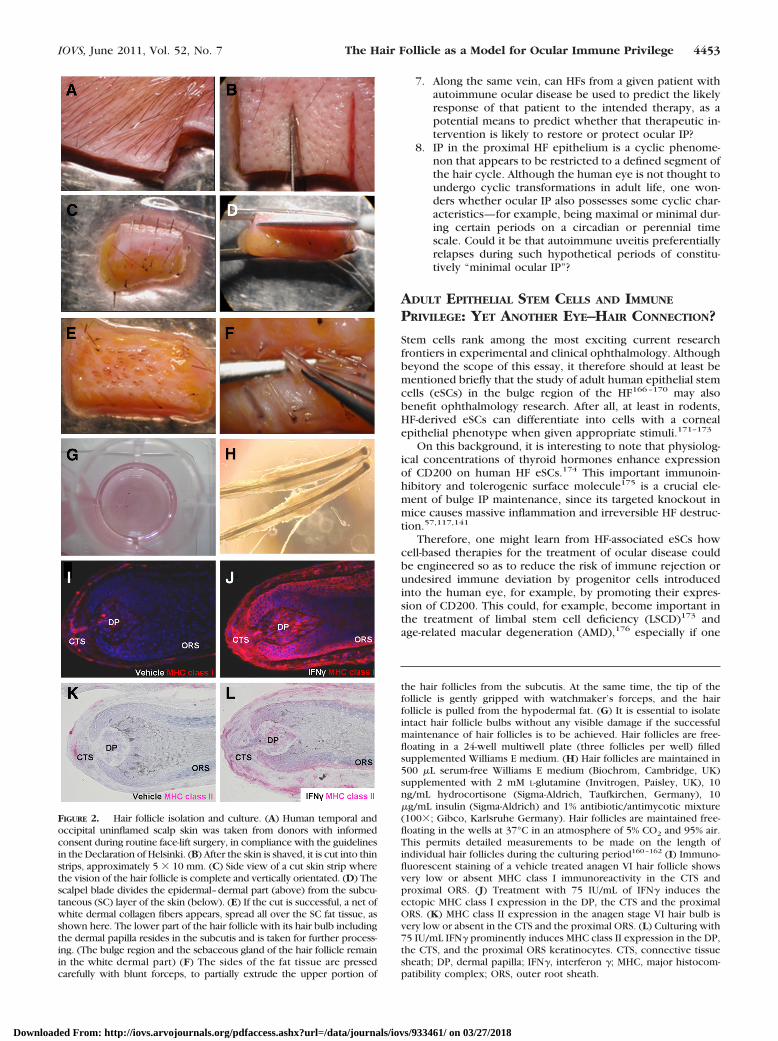
FIGURE 2. Hair follicle isolation and culture. (A) Human temporal andoccipital uninflamed scalp skin was taken from donors with informedconsent during routine face-lift surgery, in compliance with the guidelinesin the Declaration of Helsinki. (B) After the skin is shaved, it is cut into thinstrips, approximately 5 � 10 mm. (C) Side view of a cut skin strip wherethe vision of the hair follicle is complete and vertically orientated. (D) Thescalpel blade divides the epidermal–dermal part (above) from the subcu-taneous (SC) layer of the skin (below). (E) If the cut is successful, a net ofwhite dermal collagen fibers appears, spread all over the SC fat tissue, asshown here. The lower part of the hair follicle with its hair bulb includingthe dermal papilla resides in the subcutis and is taken for further process-ing. (The bulge region and the sebaceous gland of the hair follicle remainin the white dermal part) (F) The sides of the fat tissue are pressedcarefully with blunt forceps, to partially extrude the upper portion of
the hair follicles from the subcutis. At the same time, the tip of thefollicle is gently gripped with watchmaker’s forceps, and the hairfollicle is pulled from the hypodermal fat. (G) It is essential to isolateintact hair follicle bulbs without any visible damage if the successfulmaintenance of hair follicles is to be achieved. Hair follicles are free-floating in a 24-well multiwell plate (three follicles per well) filledsupplemented Williams E medium. (H) Hair follicles are maintained in500 �L serum-free Williams E medium (Biochrom, Cambridge, UK)supplemented with 2 mM L-glutamine (Invitrogen, Paisley, UK), 10ng/mL hydrocortisone (Sigma-Aldrich, Taufkirchen, Germany), 10�g/mL insulin (Sigma-Aldrich) and 1% antibiotic/antimycotic mixture(100�; Gibco, Karlsruhe Germany). Hair follicles are maintained free-floating in the wells at 37°C in an atmosphere of 5% CO2 and 95% air.This permits detailed measurements to be made on the length ofindividual hair follicles during the culturing period160–162 (I) Immuno-fluorescent staining of a vehicle treated anagen VI hair follicle showsvery low or absent MHC class I immunoreactivity in the CTS andproximal ORS. (J) Treatment with 75 IU/mL of IFN� induces theectopic MHC class I expression in the DP, the CTS and the proximalORS. (K) MHC class II expression in the anagen stage VI hair bulb isvery low or absent in the CTS and the proximal ORS. (L) Culturing with75 IU/mL IFN� prominently induces MHC class II expression in the DP,the CTS, and the proximal ORS keratinocytes. CTS, connective tissuesheath; DP, dermal papilla; IFN�, interferon �; MHC, major histocom-patibility complex; ORS, outer root sheath.
IOVS, June 2011, Vol. 52, No. 7 The Hair Follicle as a Model for Ocular Immune Privilege 4453
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

explores the use of autologous, easily accessible human HFeSCs as a potential source for such cell therapy. Do HF-derivedhuman eSCs (or any other epithelial progenitor cell type ex-ploited for cell-based regeneration strategies in experimentalophthalmology) retain the relatively immune-privileged statusthat they had enjoyed in situ,142 once they are isolated, prop-agated, and treated in cell culture?
CONCLUSIONS AND PERSPECTIVES
It is now widely accepted that understanding ocular IP willcontribute to the development of new therapeutic approachesto tissue transplantation and autoimmune diseases, not only ofthe eye, but also of other organs.4,7,12,59 The same may beclaimed for HF IP, and this in a clinically relevant and preclini-cally much more accessible and available model system. De-spite the many evident differences between the eye and theHF, the limited, but persuasive similarities between ocular andfollicular IP delineated above raise the possibility that theirrespective responses to test agents, and the selected aspects of IPlisted above also are quite similar—probably at least on the samelevel of similarity as that of rodent versus human ocular IP.
Collaboration between ocular and skin scientists in the jointexploration of IP therefore promises to be very fruitful to bothcommunities. Let us remember: Van Dooremaal andMedawar,1,2 the pioneers of IP research, made their seminaldiscoveries on ocular IP by placing skin allografts into the AC,thus paving the way for IP research that combines a cutaneousand an ocular perspective. It is in this tradition that we advo-cate the study and manipulation of a cutaneous miniorgan, theHF, as an unconventional, but highly instructive, accessible,and clinically relevant preclinical surrogate model for definedaspects of ocular IP. No doubt, the surrogate model we areproposing here has major limitations and cannot fully satisfy adevoted ocular IP researcher; but it is the best preclinicalsurrogate model that we have so far (and may ever have) in thehuman system.
Acknowledgments
MK thanks Joseph Moisseiev for his continued encouragement, profes-sional advice, and support.
References
1. Medawar PB. Immunity to homologous grafted skin; the fate ofskin homografts transplanted to the brain, to subcutaneous tissue,and to the anterior chamber of the eye. Br J Exp Pathol. 1948;29:58–69.
2. van Dooremaal J. Die Entwicklung der in fremden Grund versetz-ten lebenden Geweba. Graefes Arch Ophthalmol. 1873;19:358–373.
3. Hazlett LD, Hendricks RL. Reviews for immune privilege in theyear 2010: immune privilege and infection. Ocul Immunol In-flamm. 2010;18:237–243.
4. McKenna KC, Chen PW. Influence of immune privilege on oculartumor development. Ocul Immunol Inflamm. 2010;18:80–90.
5. Taylor AW. Ocular immune privilege. Eye (Lond). 2009;23:1885–1889.
6. Niederkorn JY. Immune escape mechanisms of intraocular tu-mors. Prog Retin Eye Res. 2009;28:329–347.
7. Niederkorn JY, Stein-Streilein J. History and physiology of im-mune privilege. Ocul Immunol Inflamm. 2010;18:19–23.
8. Hori J, Vega JL, Masli S. Review of ocular immune privilege in theyear 2010: modifying the immune privilege of the eye. OculImmunol Inflamm. 2010;18:325–333.
9. Forrester JV. Privilege revisited: an evaluation of the eye’s de-fence mechanisms. Eye (Lond). 2009;23:756–766.
10. Niederkorn JY. High-risk corneal allografts and why they losetheir immune privilege. Curr Opin Allergy Clin Immunol. 2010;10:493–497.
11. Forrester JV, Xu H, Lambe T, Cornall R. Immune privilege orprivileged immunity? Mucosal Immunol. 2008;1:372–381.
12. Cobbold SP. Future therapeutics for the induction of peripheralimmune tolerance in autoimmune disease and organ transplanta-tion. Immunotherapy. 2009;1:447–460.
13. Cunnusamy K, Chen PW, Niederkorn JY. Paradigm shifts in therole of CD4� T cells in keratoplasty. Discov Med. 2010;10:452–461.
14. Caspi RR. Ocular autoimmunity: the price of privilege? ImmunolRev. 2006;213:23–35.
15. Masli S, Vega JL. Ocular immune privilege sites. Methods MolBiol. 2011;677:449–458.
16. Hori J, Taniguchi H, Wang M, Oshima M, Azuma M. GITR ligand-mediated local expansion of regulatory T cells contributes toimmune privilege of corneal allografts. Invest Ophthalmol VisSci. 2010;51:6556–6565.
17. Cunnusamy K, Chen PW, Niederkorn JY. IL-17 promotes immuneprivilege of corneal allografts. J Immunol. 2010;185:4651–4658.
18. Qiao H, Lucas K, Stein-Streilein J. Retinal laser burn disruptsimmune privilege in the eye. Am J Pathol. 2009;174:414–422.
19. Zamiri P, Masli S, Streilein JW, Taylor AW. Pigment epithelialgrowth factor suppresses inflammation by modulating macro-phage activation. Invest Ophthalmol Vis Sci. 2006;47:3912–3918.
20. Hori J, Wang M, Miyashita M, et al. B7–H1-induced apoptosis as amechanism of immune privilege of corneal allografts. J Immunol.2006;177:5928–5935.
21. Zamiri P, Masli S, Kitaichi N, Taylor AW, Streilein JW. Thrombos-pondin plays a vital role in the immune privilege of the eye.Invest Ophthalmol Vis Sci. 2005;46:908–919.
22. Sonoda KH, Sakamoto T, Qiao H, et al. The analysis of systemictolerance elicited by antigen inoculation into the vitreous cavity:vitreous cavity-associated immune deviation. Immunology. 2005;116:390–399.
23. Taylor AW, Yee DG. Somatostatin is an immunosuppressive fac-tor in aqueous humor. Invest Ophthalmol Vis Sci. 2003;44:2644–2649.
24. Streilein JW, Arancibia-Caracamo C, Osawa H. The role of minorhistocompatibility alloantigens in penetrating keratoplasty. DevOphthalmol. 2003;36:74–88.
25. Lee HO, Herndon JM, Barreiro R, Griffith TS, Ferguson TA. TRAIL:a mechanism of tumor surveillance in an immune privileged site.J Immunol. 2002;169:4739–4744.
26. Kawanaka N, Taylor AW. Localized retinal neuropeptide regula-tion of macrophage and microglial cell functionality. J Neuroim-munol. 2011;232:17–25.
27. Murphy K, Travers P, Walport M. Janeway’s Immunobiology.7th ed. New York: Garland Science Press; 2008;711–729.
28. Eichhorn M, Horneber M, Streilein JW, Lutjen-Drecoll E. Anteriorchamber-associated immune deviation elicited via primate eyes.Invest Ophthalmol Vis Sci. 1993;34:2926–2930.
29. Usui Y, Okunuki Y, Hattori T, et al. Functional expression ofB7H1 on retinal pigment epithelial cells. Exp Eye Res. 2008;86:52–59.
30. Ryu YH, Kim JC. Expression of indoleamine 2,3-dioxygenase inhuman corneal cells as a local immunosuppressive factor. InvestOphthalmol Vis Sci. 2007;48:4148–4152.
31. Sugita S, Keino H, Futagami Y, et al. B7� iris pigment epithelialcells convert T cells into CTLA-4�, B7-expressing CD8� regula-tory T cells. Invest Ophthalmol Vis Sci. 2006;47:5376–5384.
32. Sohn JH, Kaplan HJ, Suk HJ, Bora PS, Bora NS. Complementregulatory activity of normal human intraocular fluid is mediatedby MCP, DAF, and CD59. Invest Ophthalmol Vis Sci. 2000;41:4195–4202.
33. Yang W, Li H, Chen PW, et al. PD-L1 expression on human ocularcells and its possible role in regulating immune-mediated ocularinflammation. Invest Ophthalmol Vis Sci. 2009;50:273–280.
34. Paus R, Nickoloff BJ, Ito T. A ‘hairy’ privilege. Trends Immunol.2005;26:32–40.
4454 Kinori et al IOVS, June 2011, Vol. 52, No. 7
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

35. Huang L, Baban B, Johnson BA, 3rd, Mellor AL. Dendritic cells,indoleamine 2,3 dioxygenase and acquired immune privilege. IntRev Immunol. 2010;29:133–155.
36. Barker CF, Billingham RE. Immunologically privileged sites. AdvImmunol. 1977;25:1–54.
37. Erlebacher A. Why isn’t the fetus rejected? Curr Opin Immunol.2001;13:590–593.
38. Mellor AL, Munn DH. Immunology at the maternal-fetal interface:lessons for T cell tolerance and suppression. Annu Rev Immu-nol. 2000;18:367–391.
39. Streilein JW. Ocular immune privilege: therapeutic opportunitiesfrom an experiment of nature. Nat Rev Immunol. 2003;3:879–889.
40. Meinhardt A, Hedger MP. Immunological, paracrine and endo-crine aspects of testicular immune privilege. Mol Cell Endocrinol.2011;335:60–68.
41. Ito T, Ito N, Saathoff M, et al. Immunology of the human nailapparatus: the nail matrix is a site of relative immune privilege.J Invest Dermatol. 2005;125:1139–1148.
42. Schneider MR, Schmidt-Ullrich R, Paus R. The hair follicle as adynamic miniorgan. Curr Biol. 2009;19:R132–R142.
43. Gilhar A, Paus R, Kalish RS. Lymphocytes, neuropeptides, andgenes involved in alopecia areata. J Clin Invest. 2007;117:2019–2027.
44. Ito T, Ito N, Bettermann A, Tokura Y, Takigawa M, Paus R.Collapse and restoration of MHC class-I-dependent immuneprivilege: exploiting the human hair follicle as a model. Am JPathol. 2004;164:623–634.
45. Peters EM, Liotiri S, Bodo E, et al. Probing the effects of stressmediators on the human hair follicle: substance P holds centralposition. Am J Pathol. 2007;171:1872–1886.
46. Kaplan HJ, Niederkorn JY. Regional immunity and immune priv-ilege. Chem Immunol Allergy. 2007;92:11–26.
47. Niederkorn JY, Wang S. Immune privilege of the eye and fetus:parallel universes? Transplantation. 2005;80:1139–1144.
48. Arck PC, Gilhar A, Bienenstock J, Paus R. The alchemy of immuneprivilege explored from a neuroimmunological perspective. CurrOpin Pharmacol. 2008;8:480–489.
49. Niederkorn JY. Mechanisms of immune privilege in the eye andhair follicle. J Investig Dermatol Symp Proc. 2003;8:168–172.
50. Simpson E. A historical perspective on immunological privilege.Immunol Rev. 2006;213:12–22.
51. Welge-Lussen U, Wilsch C, Neuhardt T, Wayne Streilein J, Lutjen-Drecoll E. Loss of anterior chamber-associated immune deviation(ACAID) in aged retinal degeneration (rd) mice. Invest Ophthal-mol Vis Sci. 1999;40:3209–3214.
52. Billingham RE. Transplantation immunity evoked by skin ho-mografts and expressed in intact skin. Adv Biol Skin. 1971;11:183–198.
53. Billingham RE, Silvers WK. A biologist’s reflections on dermatol-ogy. J Invest Dermatol. 1971;57:227–240.
54. Anand V, Duffy B, Yang Z, Dejneka NS, Maguire AM, Bennett J. Adeviant immune response to viral proteins and transgene productis generated on subretinal administration of adenovirus and ad-eno-associated virus. Mol Ther. 2002;5:125–132.
55. Wenkel H, Chen PW, Ksander BR, Streilein JW. Immune privilegeis extended, then withdrawn, from allogeneic tumor cell graftsplaced in the subretinal space. Invest Ophthalmol Vis Sci. 1999;40:3202–3208.
56. Paus R, Ito N, Takigawa M, Ito T. The hair follicle and immuneprivilege. J Investig Dermatol Symp Proc. 2003;8:188–194.
57. Meyer KC, Klatte JE, Dinh HV, et al. Evidence that the bulgeregion is a site of relative immune privilege in human hair folli-cles. Br J Dermatol. 2008;159:1077–1085.
58. Niederkorn JY. Role of NKT cells in anterior chamber-associatedimmune deviation. Expert Rev Clin Immunol. 2009;5:137–144.
59. Hori J. Mechanisms of immune privilege in the anterior segmentof the eye: what we learn from corneal transplantation. J OculBiol Dis Infor. 2008;1:94–100.
60. Taylor AW, Streilein JW, Cousins SW. Identification of alpha-melanocyte stimulating hormone as a potential immunosuppres-sive factor in aqueous humor. Curr Eye Res. 1992;11:1199–1206.
61. Cousins SW, McCabe MM, Danielpour D, Streilein JW. Identifica-tion of transforming growth factor-beta as an immunosuppressivefactor in aqueous humor. Invest Ophthalmol Vis Sci. 1991;32:2201–2211.
62. Denniston AK, Kottoor SH, Khan I, et al. Endogenous cortisol andTGF-� in human aqueous humor contribute to ocular immuneprivilege by regulating dendritic cell function. J Immunol. 2011;186:305–311.
63. Granstein RD, Staszewski R, Knisely TL, et al. Aqueous humorcontains transforming growth factor-beta and a small (less than3500 daltons) inhibitor of thymocyte proliferation. J Immunol.1990;144:3021–3027.
64. Sheibani N, Sorenson CM, Cornelius LA, Frazier WA. Thrombos-pondin-1, a natural inhibitor of angiogenesis, is present in vitre-ous and aqueous humor and is modulated by hyperglycemia.Biochem Biophys Res Commun. 2000;267:257–261.
65. Taylor AW, Streilein JW, Cousins SW. Immunoreactive vasoactiveintestinal peptide contributes to the immunosuppressive activityof normal aqueous humor. J Immunol. 1994;153:1080–1086.
66. Apte RS, Sinha D, Mayhew E, Wistow GJ, Niederkorn JY. Cuttingedge: role of macrophage migration inhibitory factor in inhibitingNK cell activity and preserving immune privilege. J Immunol.1998;160:5693–5696.
67. Taylor AW, Yee DG, Streilein JW. Suppression of nitric oxidegenerated by inflammatory macrophages by calcitonin gene-re-lated peptide in aqueous humor. Invest Ophthalmol Vis Sci.1998;39:1372–1378.
68. Kennedy MC, Rosenbaum JT, Brown J, et al. Novel production ofinterleukin-1 receptor antagonist peptides in normal human cor-nea. J Clin Invest. 1995;95:82–88.
69. Sugita S, Streilein JW. Iris pigment epithelium expressing CD86(B7–2) directly suppresses T cell activation in vitro via binding tocytotoxic T lymphocyte-associated antigen 4. J Exp Med. 2003;198:161–171.
70. Ito N, Ito T, Kromminga A, et al. Human hair follicles display afunctional equivalent of the hypothalamic-pituitary-adrenal axisand synthesize cortisol. Faseb J. 2005;19:1332–1334.
71. Ito T, Ito N, Saatoff M, et al. Maintenance of hair follicle immuneprivilege is linked to prevention of NK cell attack. J InvestDermatol. 2008;128:1196–1206.
72. Velasco P, Huegel R, Brasch J, et al. The angiogenesis inhibitorthrombospondin-1 inhibits acute cutaneous hypersensitivity re-actions. J Invest Dermatol. 2009;129:2022–2030.
73. Kim CY, Masli S, Streilein JW. Qa-1, a nonclassical MHC moleculewith immunomodulatory functions, is ubiquitously expressed inthe immune-privileged anterior chamber of the eye. Ocul Immu-nol Inflamm. 2005;13:271–277.
74. Streilein JW, Toews GB, Bergstresser PR. Corneal allografts fail toexpress Ia antigens. Nature. 1979;282:326–327.
75. Niederkorn JY, Chiang EY, Ungchusri T, Stroynowski I. Expres-sion of a nonclassical MHC class Ib molecule in the eye. Trans-plantation. 1999;68:1790–1799.
76. Wang HM, Kaplan HJ, Chan WC, Johnson M. The distribution andontogeny of MHC antigens in murine ocular tissue. Invest Oph-thalmol Vis Sci. 1987;28:1383–1389.
77. Paus R, Eichmuller S, Hofmann U, Czarnetzki BM, Robinson P.Expression of classical and non-classical MHC class I antigens inmurine hair follicles. Br J Dermatol. 1994;131:177–183.
78. Griffith TS, Brunner T, Fletcher SM, Green DR, Ferguson TA. Fasligand-induced apoptosis as a mechanism of immune privilege.Science. 1995;270:1189–1192.
79. Sugita S, Taguchi C, Takase H, et al. Soluble Fas ligand and solubleFas in ocular fluid of patients with uveitis. Br J Ophthalmol.2000;84:1130–1134.
80. Kang H, Wu WY, Lo BK, et al. Hair follicles from alopecia areatapatients exhibit alterations in immune privilege-associated geneexpression in advance of hair loss. J Invest Dermatol. 2010;130:2677–2680.
81. Streilein JW, Niederkorn JY. Induction of anterior chamber-asso-ciated immune deviation requires an intact, functional spleen. JExp Med. 1981;153:1058–1067.
82. Jones TB, Kaplan AM. Immunologic tolerance to HGG in mice. I.Suppression of the HGG response in normal mice with spleen
IOVS, June 2011, Vol. 52, No. 7 The Hair Follicle as a Model for Ocular Immune Privilege 4455
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

cells or a spleen cell lysate from tolerant mice. J Immunol.1977;118:1880–1885.
83. Kaplan HJ, Streilein JW, Stevens TR. Transplantation immunologyof the anterior chamber of the eye. II. Immune response toallogeneic cells. J Immunol. 1975;115:805–810.
84. Streilein JW, Dana MR, Ksander BR. Immunity causing blindness:five different paths to herpes stromal keratitis. Immunol Today.1997;18:443–449.
85. Wilbanks GA, Streilein JW. Characterization of suppressor cells inanterior chamber-associated immune deviation (ACAID) inducedby soluble antigen; evidence of two functionally and phenotypi-cally distinct T-suppressor cell populations. Immunology. 1990;71:383–389.
86. Wilbanks GA, Mammolenti M, Streilein JW. Studies on the induc-tion of anterior chamber-associated immune deviation (ACAID).III. Induction of ACAID depends upon intraocular transforminggrowth factor-beta. Eur J Immunol. 1992;22:165–173.
87. Wahl SM, Wen J, Moutsopoulos N. TGF-beta: a mobile purveyorof immune privilege. Immunol Rev. 2006;213:213–227.
88. Wilbanks GA, Streilein JW. Studies on the induction of anteriorchamber-associated immune deviation (ACAID). 1. Evidence thatan antigen-specific, ACAID-inducing, cell-associated signal existsin the peripheral blood. J Immunol. 1991;146:2610–2617.
89. Crawford K, Kaufman PL, Gabelt BT. Effects of topical PGF2alpha on aqueous humor dynamics in cynomolgus monkeys. CurrEye Res. 1987;6:1035–1044.
90. Faunce DE, Sonoda KH, Stein-Streilein J. MIP-2 recruits NKT cellsto the spleen during tolerance induction. J Immunol. 2001;166:313–321.
91. Faunce DE, Stein-Streilein J. NKT cell-derived RANTES recruitsAPCs and CD8� T cells to the spleen during the generation ofregulatory T cells in tolerance. J Immunol. 2002;169:31–38.
92. Sonoda KH, Faunce DE, Taniguchi M, Exley M, Balk S, Stein-Streilein J. NK T cell-derived IL-10 is essential for the differentia-tion of antigen-specific T regulatory cells in systemic tolerance.J Immunol. 2001;166:42–50.
93. Nakamura T, Sonoda KH, Faunce DE, et al. CD4� NKT cells, butnot conventional CD4� T cells, are required to generate efferentCD8� T regulatory cells following antigen inoculation in animmune-privileged site. J Immunol. 2003;171:1266–1271.
94. Skelsey ME, Mayhew E, Niederkorn JY. CD25�, interleukin-10-producing CD4� T cells are required for suppressor cell produc-tion and immune privilege in the anterior chamber of the eye.Immunology. 2003;110:18–29.
95. Wang Y, Goldschneider I, O’Rourke J, Cone RE. Blood mononu-clear cells induce regulatory NK T thymocytes in anterior cham-ber-associated immune deviation. J Leukoc Biol. 2001;69:741–746.
96. Li X, Taylor S, Zegarelli B, Shen S, O’Rourke J, Cone RE. Theinduction of splenic suppressor T cells through an immune-privileged site requires an intact sympathetic nervous system.J Neuroimmunol. 2004; 153:40–49.
97. Lutjen-Drecoll E, Bergstrom A, Ehinger B. Ultrastructure of retinalcells transplanted to the rabbit choroid. Ophthalmologica. 2000;214:70–77.
98. Streilein JW. Immune regulation and the eye: a dangerous com-promise. FASEB J. 1987;1:199–208.
99. Streilein JW. Ocular immune privilege and the Faustian dilemma.The Proctor lecture. Invest Ophthalmol Vis Sci. 1996;37:1940–1950.
100. Kezuka T, Sakai J, Usui N, Streilein JW, Usui M. Evidence forantigen-specific immune deviation in patients with acute retinalnecrosis. Arch Ophthalmol. 2001;119:1044–1049.
101. Streilein JW, Okamoto S, Sano Y, Taylor AW. Neural control ofocular immune privilege. Ann N Y Acad Sci. 2000;917:297–306.
102. Sugita S, Futagami Y, Smith SB, Naggar H, Mochizuki M. Retinaland ciliary body pigment epithelium suppress activation of Tlymphocytes via transforming growth factor beta. Exp Eye Res.2006;83:1459–1471.
103. Streilein JW, Ma N, Wenkel H, Ng TF, Zamiri P. Immunobiologyand privilege of neuronal retina and pigment epithelium trans-plants. Vision Res. 2002;42:487–495.
104. Jorgensen A, Wiencke AK, la Cour M, et al. Human retinal pig-ment epithelial cell-induced apoptosis in activated T cells. InvestOphthalmol Vis Sci. 1998;39:1590–1599.
105. Wenkel H, Streilein JW. Evidence that retinal pigment epitheliumfunctions as an immune-privileged tissue. Invest Ophthalmol VisSci. 2000;41:3467–3473.
106. Ng TF, Osawa H, Hori J, Young MJ, Streilein JW. Allogeneicneonatal neuronal retina grafts display partial immune privilegein the subcapsular space of the kidney. J Immunol. 2002;169:5601–5606.
107. Jiang LQ, Jorquera M, Streilein JW. Subretinal space and vitreouscavity as immunologically privileged sites for retinal allografts.Invest Ophthalmol Vis Sci. 1993;34:3347–3354.
108. Niederkorn JY. See no evil, hear no evil, do no evil: the lessons ofimmune privilege. Nat Immunol. 2006;7:354–359.
109. The Collaborative Corneal Transplantation Studies ResearchGroup. The collaborative corneal transplantation studies: effec-tiveness of histocompatibility matching in high-risk corneal trans-plantation. Arch Ophthalmol. 1992;110:1392–1403.
110. Sonoda Y, Streilein JW. Orthotopic corneal transplantation inmice–evidence that the immunogenetic rules of rejection do notapply. Transplantation. 1992;54:694–704.
111. Mellor AL, Munn DH. Creating immune privilege: active localsuppression that benefits friends, but protects foes. Nat RevImmunol. 2008;8:74–80.
112. Roychoudhury J, Herndon JM, Yin J, Apte RS, Ferguson TA.Targeting immune privilege to prevent pathogenic neovascular-ization. Invest Ophthalmol Vis Sci. 2010;51:3560–3566.
113. Paus R, Christoph T, Muller-Rover S. Immunology of the hairfollicle: a short journey into terra incognita. J Investig DermatolSymp Proc. 1999;4:226–234.
114. Petukhova L, Duvic M, Hordinsky M, et al. Genome-wide associ-ation study in alopecia areata implicates both innate and adaptiveimmunity. Nature. 2010;466:113–117.
115. Harries MJ, Sun J, Paus R, King LE Jr. Management of alopeciaareata. BMJ. 2010;341:c3671.
116. Gilhar A. Collapse of immune privilege in alopecia areata: coin-cidental or substantial? J Invest Dermatol. 2010;130:2535–2537.
117. Harries MJ, Paus R. The pathogenesis of primary cicatricial alo-pecias. Am J Pathol. 2010;177:2152–2162.
118. Harrist TJ, Ruiter DJ, Mihm MC Jr, Bhan AK. Distribution of majorhistocompatibility antigens in normal skin. Br J Dermatol. 1983;109:623–633.
119. Lee N, Llano M, Carretero M, et al. HLA-E is a major ligand for thenatural killer inhibitory receptor CD94/NKG2A. Proc Natl AcadSci U S A. 1998;95:5199–5204.
120. Carson MJ, Doose JM, Melchior B, Schmid CD, Ploix CC. CNSimmune privilege: hiding in plain sight. Immunol Rev. 2006;213:48–65.
121. Foulds LM, Boysen RI, Crane M, et al. Molecular identification oflyso-glycerophosphocholines as endogenous immunosuppres-sives in bovine and rat gonadal fluids. Biol Reprod. 2008;79:525–536.
122. Paus R, Cotsarelis G. The biology of hair follicles. N Engl J Med.1999;341:491–497.
123. Christoph T, Muller-Rover S, Audring H, et al. The human hairfollicle immune system: cellular composition and immune privi-lege. Br J Dermatol. 2000;142:862–873.
124. Gilliam AC, Kremer IB, Yoshida Y, et al. The human hair follicle:a reservoir of CD40� B7-deficient Langerhans cells that repopu-late epidermis after UVB exposure. J Invest Dermatol. 1998;110:422–427.
125. Kaufman G, d’Ovidio R, Kaldawy A, et al. An unexpected twist inalopecia areata pathogenesis: are NK cells protective andCD49b� T cells pathogenic? Exp Dermatol. 2010;19:e347–349.
126. Gilhar A, Shalaginov R, Assy B, Serafimovich S, Kalish RS. Alope-cia areata is a T-lymphocyte mediated autoimmune disease: le-sional human T-lymphocytes transfer alopecia areata to humanskin grafts on SCID mice. J Investig Dermatol Symp Proc. 1999;4:207–210.
127. Gilhar A, Ullmann Y, Berkutzki T, Assy B, Kalish RS. Autoimmunehair loss (alopecia areata) transferred by T lymphocytes to humanscalp explants on SCID mice. J Clin Invest. 1998;101:62–67.
4456 Kinori et al IOVS, June 2011, Vol. 52, No. 7
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

128. McElwee KJ, Yu M, Park SW, Ross EK, Finner A, Shapiro J. Whatcan we learn from animal models of Alopecia areata? Dermatol-ogy. 2005;211:47–53.
129. Ito T, Hashizume H, Takigawa M. Significant increase of MAGE 3specific-CTLs in alopecia areata patients. Presented at the 6thWorld Congress for Hair Research. Cairns, Australia; 2010.
130. Gilhar A, Landau M, Assy B, Shalaginov R, Serafimovich S, KalishRS. Melanocyte-associated T cell epitopes can function as autoan-tigens for transfer of alopecia areata to human scalp explants onPrkdc(scid) mice. J Invest Dermatol. 2001;117:1357–1362.
131. Paus R, Slominski A, Czarnetzki BM. Is alopecia areata an autoim-mune-response against melanogenesis-related proteins, exposedby abnormal MHC class I expression in the anagen hair bulb? YaleJ Biol Med. 1993;66:541–554.
132. Yokoyama WM, Kim S. Licensing of natural killer cells by self-major histocompatibility complex class I. Immunol Rev. 2006;214:143–154.
133. Vivier E. What is natural in natural killer cells? Immunol Lett.2006;107:1–7.
134. Khakoo SI, Carrington M. KIR and disease: a model system orsystem of models? Immunol Rev. 2006;214:186–201.
135. Johansson MH, Hoglund P. The dynamics of natural killer celltolerance. Semin Cancer Biol. 2006;16:393–403.
136. Gasser S, Raulet DH. Activation and self-tolerance of natural killercells. Immunol Rev. 2006;214:130–142.
137. Bryceson YT, March ME, Ljunggren HG, Long EO. Activation,coactivation, and costimulation of resting human natural killercells. Immunol Rev. 2006;214:73–91.
138. Borrego F, Masilamani M, Marusina AI, Tang X, Coligan JE. TheCD94/NKG2 family of receptors: from molecules and cells toclinical relevance. Immunol Res. 2006;35:263–278.
139. Boehm T. Quality control in self/nonself discrimination. Cell.2006;125:845–858.
140. Kloepper JE, Tiede S, Brinckmann J, et al. Immunophenotypingof the human bulge region: the quest to define useful in situmarkers for human epithelial hair follicle stem cells and theirniche. Exp Dermatol. 2008;17:592–609.
141. Rosenblum MD, Olasz EB, Yancey KB, et al. Expression of CD200on epithelial cells of the murine hair follicle: a role in tissue-specific immune tolerance? J Invest Dermatol. 2004;123:880–887.
142. Tiede S, Koop N, Kloepper JE, Fassler R, Paus R. Nonviral in situgreen fluorescent protein labeling and culture of primary, adulthuman hair follicle epithelial progenitor cells. Stem Cells. 2009;27:2793–2803.
143. Copland DA, Calder CJ, Raveney BJ, et al. Monoclonal antibody-mediated CD200 receptor signaling suppresses macrophage ac-tivation and tissue damage in experimental autoimmune uveoreti-nitis. Am J Pathol. 2007;171:580–588.
144. Norris DA. How close are we to solving the puzzle?—review ofthe Alopecia Areata Research Workshop. J Investig DermatolSymp Proc. 2003;8:222–225.
145. Whitsett CF, Stulting RD. The distribution of HLA antigens onhuman corneal tissue. Invest Ophthalmol Vis Sci. 1984;25:519–524.
146. Barez S, Boumpas DT, Percopo CM, Anastassiou ED, Hooks JJ,Detrick B. Modulation of major histocompatibility complex class1 genes in human retinoblastoma cells by interferons. InvestOphthalmol Vis Sci. 1993;34:2613–2621.
147. Ruckert R, Hofmann U, van der Veen C, Bulfone-Paus S, Paus R.MHC class I expression in murine skin: developmentally con-trolled and strikingly restricted intraepithelial expression duringhair follicle morphogenesis and cycling, and response to cytokinetreatment in vivo. J Invest Dermatol. 1998;111:25–30.
148. Paus R, Theoharides TC, Arck PC. Neuroimmunoendocrine cir-cuitry of the ‘brain-skin connection’. Trends Immunol. 2006;27:32–39.
149. Arck PC, Handjiski B, Hagen E, Joachim R, Klapp BF, Paus R.Indications for a ‘brain-hair follicle axis (BHA)’: inhibition ofkeratinocyte proliferation and up-regulation of keratinocyte apo-ptosis in telogen hair follicles by stress and substance P. FASEB J.2001;15:2536–2538.
150. Botchkarev VA, Botchkareva NV, Peters EM, Paus R. Epithelialgrowth control by neurotrophins: leads and lessons from the hairfollicle. Prog Brain Res. 2004;146:493–513.
151. Slominski A, Wortsman J, Tuckey RC, Paus R. Differential expres-sion of HPA axis homolog in the skin. Mol Cell Endocrinol.2007;265–266:143–149.
152. Arck PC, Handjiski B, Peters EM, et al. Stress inhibits hair growthin mice by induction of premature catagen development anddeleterious perifollicular inflammatory events via neuropeptidesubstance P-dependent pathways. Am J Pathol. 2003;162:803–814.
153. Arck PC, Handjiski B, Kuhlmei A, et al. Mast cell deficient andneurokinin-1 receptor knockout mice are protected from stress-induced hair growth inhibition. J Mol Med. 2005;83:386–396.
154. Paus R, Arck P, Tiede S. (Neuro-)endocrinology of epithelial hairfollicle stem cells. Mol Cell Endocrinol. 2008;288:38–51.
155. Peters EM, Arck PC, Paus R. Hair growth inhibition by psychoe-motional stress: a mouse model for neural mechanisms in hairgrowth control. Exp Dermatol. 2006;15:1–13.
156. Paus R. A neuroendocrinological perspective on human hairfollicle pigmentation. Pigment Cell Melanoma Res. 2011;24:89–106.
157. Joachim RA, Kuhlmei A, Dinh QT, et al. Neuronal plasticity of the“brain-skin connection”: stress-triggered up-regulation of neuro-peptides in dorsal root ganglia and skin via nerve growth factor-dependent pathways. J Mol Med. 2007;85:1369–1378.
158. Peters EM, Handjiski B, Kuhlmei A, et al. Neurogenic inflamma-tion in stress-induced termination of murine hair growth is pro-moted by nerve growth factor. Am J Pathol. 2004;165:259–271.
159. Siebenhaar F, Sharov AA, Peters EM, et al. Substance P as animmunomodulatory neuropeptide in a mouse model for autoim-mune hair loss (alopecia areata). J Invest Dermatol. 2007;127:1489–1497.
160. Kloepper JE, Hendrix S, Bodo E, et al. Functional role of beta 1integrin-mediated signalling in the human hair follicle. Exp CellRes. 2008;314:498–508.
161. Philpott M, Green MR, Kealey T. Studies on the biochemistry andmorphology of freshly isolated and maintained rat hair follicles.J Cell Sci. 1989;93:409–418.
162. Philpott MP, Green MR, Kealey T. Human hair growth in vitro.J Cell Sci. 1990;97:463–471.
163. Botchkarev VA, Botchkareva NV, Slominski A, Roloff B, Luger T,Paus R. Developmentally regulated expression of alpha-MSH andMC-1 receptor in C57BL/6 mouse skin suggests functions beyondpigmentation. Ann N Y Acad Sci. 1999;885:433–439.
164. Slominski A, Wortsman J, Luger T, Paus R, Solomon S. Cortico-tropin releasing hormone and proopiomelanocortin involvementin the cutaneous response to stress. Physiol Rev. 2000;80:979–1020.
165. Carrim ZI, Ahmed TY, Taguri AH. The relationship betweenstress and acute anterior uveitis. Acta Ophthalmol Scand. 2006;84:795–798.
166. Tiede S, Kloepper JE, Bodo E, Tiwari S, Kruse C, Paus R. Hairfollicle stem cells: walking the maze. Eur J Cell Biol. 2007;86:355–376.
167. Gutierrez-Rivera A, Pavon-Rodriguez A, Jimenez-Acosta F, et al.Functional characterization of highly adherent CD34� keratino-cytes isolated from human skin. Exp Dermatol. 2010;19:685–688.
168. Ohyama M, Zheng Y, Paus R, Stenn KS. The mesenchymal com-ponent of hair follicle neogenesis: background, methods andmolecular characterization. Exp Dermatol. 2010;19:89–99.
169. Biernaskie J. Human hair follicles: “bulging” with neural crest-likestem cells. J Invest Dermatol. 2010;130:1202–1204.
170. Tiede S, Kloepper JE, Ernst N, Poeggeler B, Kruse C, Paus R.Nestin in human skin: exclusive expression in intramesenchymalskin compartments and regulation by leptin. J Invest Dermatol.2009;129:2711–2720.
171. Blazejewska EA, Schlotzer-Schrehardt U, Zenkel M, et al. Corneallimbal microenvironment can induce transdifferentiation of hairfollicle stem cells into corneal epithelial-like cells. Stem Cells.2009;27:642–652.
IOVS, June 2011, Vol. 52, No. 7 The Hair Follicle as a Model for Ocular Immune Privilege 4457
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018

172. Yang K, Jiang Z, Wang D, Lian X, Yang T. Corneal epithelial-liketransdifferentiation of hair follicle stem cells is mediated by pax6and beta-catenin/Lef-1. Cell Biol Int. 2009;33:861–866.
173. Meyer-Blazejewska EA, Call MK, Yamanaka O, et al. From hair tocornea: towards the therapeutic use of hair follicle-derived stemcells in the treatment of limbal stem cell deficiency. Stem Cells.2011;29:57–66.
174. Tiede S, Bohm K, Meier N, Funk W, Paus R. Endocrine controls ofprimary adult human stem cell biology: thyroid hormones stim-
ulate keratin 15 expression, apoptosis, and differentiation inhuman hair follicle epithelial stem cells in situ and in vitro. EurJ Cell Biol. 2010;89:769–777.
175. Kawasaki BT, Mistree T, Hurt EM, Kalathur M, Farrar WL. Co-expression of the toleragenic glycoprotein, CD200, with markersfor cancer stem cells. Biochem Biophys Res Commun. 2007;364:778–782.
176. Lee E, Maclaren RE. Sources of retinal pigment epithelium (RPE)for replacement therapy. Br J Ophthalmol. 2011;445–449.
4458 Kinori et al IOVS, June 2011, Vol. 52, No. 7
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933461/ on 03/27/2018