calcium dependent protein kinase (cdpk) expression during fruit development in cultivated peanut...
TRANSCRIPT

S
Cig
Ma
b
c
d
a
ARRA
KACGSSS
I
ptfgntCt
(pEt
MT
0d
Journal of Plant Physiology 168 (2011) 2272– 2277
Contents lists available at ScienceDirect
Journal of Plant Physiology
j ourna l ho mepage: www.elsev ier .de / jp lph
hort communication
alcium dependent protein kinase (CDPK) expression during fruit developmentn cultivated peanut (Arachis hypogaea) under Ca2+-sufficient and -deficientrowth regimens
ukesh Jaina,1, Bhuvan P. Pathaka,1, Alice C. Harmonb,c, Barry L. Tillmana,d, Maria Galloa,c,∗
Agronomy Department, University of Florida, Gainesville, FL 32610-3610, USABiology Department, University of Florida, Gainesville, FL 32611-8256, USAPlant Molecular and Cellular Biology Program, University of Florida, Genetics Institute, Cancer/Genetics Research Complex, Gainesville, FL 32610-3610, USANorth Florida Research and Education Center, University of Florida, 3925 Highway 71, Marianna, FL 32446-7906, USA
r t i c l e i n f o
rticle history:eceived 19 April 2011eceived in revised form 30 July 2011ccepted 31 July 2011
eywords:rachis hypogaeaell wall invertaseeocarpy
a b s t r a c t
Adequate soil calcium (Ca2+) levels are crucial for sustained reproductive development of peanut (Arachishypogaea). A role for calcium dependent protein kinase was evaluated during peanut fruit developmentunder sufficient and deficient soil Ca2+ conditions. Quantitative RT-PCR and protein gel blot analysesconfirmed transcriptional upregulation of CDPK in seeds developing under inadequate soil Ca2+ regimen,as well as spatiotemporal regulation of CDPK expression during early mitotic growth and later duringthe storage phase of seed development. However, a consistent basal level of CDPK was present duringsimilar developmental stages of pod tissue, irrespective of the soil Ca2+ status. Immunolocalization datashowed CDPK decoration primarily in the outer most cell layers of the pericarp and around vascular
eed developmentoil calciumucrose partitioning
bundles linked by lateral connections in developing pods, as well as the single vascular trace supplyingnutrients to the developing seed. Finally, carbohydrate analyses and qRT-PCR data are provided for peanutgenes encoding enzymes involved in sucrose cleavage (orthologs of Vicia faba, VfCWI1 and VfCWI2) andutilization (AhSuSy and AhSpS), and oleosin gene transcripts (AhOleo17.8 and AhOleo18.5) validating arole for CDPK in the establishment and maintenance of sink strength, and subsequent onset of storage
se du
product biosynthetic phantroduction
Cultivated peanut (Arachis hypogaea) differs from most otherlant species due to its geocarpic reproductive growth, marked byhe underground development of fruit following pollination andertilization of air-borne flowers. Consequently, the heterotrophicrowth of the young developing fruit is reliant entirely upon mater-al assimilate supply. Peanut yield and quality is highly sensitive
o calcium (Ca2+) availability in the soil pegging zone either due toa2+ deficiency per se or reduced uptake from the soil solution dueo water deficit, low soil pH, etc. (Gascho and Parker, 2001; ReddyAbbreviations: CDPK, calcium dependent protein kinase; CWI, cell wall invertase�-fructofuranosidase, EC 3.2.1.26); EF1a, elongation factor 1-alpha; SpS, sucrose-hosphate synthase (UDP-glucose: d-fructose-6-phosphate-2-glucosyl transferase,C 2.4.1.14); SuSy, sucrose synthase (uridine diphospho glucose-fructose glucosyl-ransferase, EC 2.4.1.13).∗ Corresponding author at: 303 Cancer and Genetics Research Complex, 1376owry Road, P.O. Box 103610, Gainesville, FL 32610-3610, USA.
el.: +1 352 273 8124; fax: +1 352 392 1840.E-mail address: [email protected] (M. Gallo).
1 M.J. and B.P.P. contributed equally to this work.
176-1617/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.jplph.2011.07.005
ring seed maturation.© 2011 Elsevier GmbH. All rights reserved.
et al., 2003; Murata et al., 2008), and Ca2+ deficiency adverselyaffects seed viability and germination the following season (Wrightet al., 2009). Calcium cannot be repartitioned from older to youngertissues via the phloem route. Therefore, actively growing organsmust utilizing immediately available xylem-mobile Ca2+, thus pos-ing an additional burden on the hypogeal peanut fruit since it lackssignificant transpirational pull.
Calcium is involved in regulating a myriad of plant functionsincluding cell elongation and division, membrane fluidity andpermeability, ion fluxes, cellular pH, source-sink translocation ofcarbohydrates, N-metabolism, reproductive development, stressresponses and apoptosis (Hepler, 2005). Additionally, Ca2+ repre-sents an important node for the crosstalk involving an array ofdiverse second messengers in plants such as H+ (pH), lipids, inos-itol tri- and hexaphosphates (IP3 and IP6), cAMP, cADP-ribose,cGMP and nitric oxide (Kudla et al., 2010). The crux of the cal-cium conundrum, i.e. its ubiquitous presence and versatility asa nutrient and signaling specificity, is dependent upon stringent
regulation of free cytosolic Ca2+ concentration and appropriateperception of variability in amplitude, duration, location, kineticsand frequency of Ca2+ spikes during a signal transduction event(Hirschi, 2004). Amplification and downstream transduction of
Physi
daaiatitcmavptds(glstpsr
r(sdmflads2ssArad
Cr(ctreiifmalbre
M
P
g
M. Jain et al. / Journal of Plant
ynamic cytosolic Ca2+ transients in plants is predominantly medi-ted through calcium dependent protein kinases (CDPKs) (Harpernd Harmon, 2005). The CDPKs possess four architectural hallmarksncluding an N-terminal domain, an ATP binding kinase domain,n autoinhibitory and a C-terminal calmodulin-like domain con-aining four Ca2+-binding EF-hands. CDPKs perceive changes inntracellular Ca2+ concentrations as a result of direct binding toheir calmodulin-like domain. This allows for a conformationalhange resulting in activation of the kinase domain and displace-ent of the autoinhibitory domain, that otherwise engages the
ctive site as a pseudosubstrate. The N-terminal domain is highlyariable and often contains myristoylation or palmitoylation sitesresumably associated with subcellular targeting. CDPKs constitutehe largest Ca2+-sensing subfamily of the plant-specific, and highlyiverse and widespread Ser/Thr protein kinase superfamily, repre-enting for instance, 4% of the predicted 25,500 genes in ArabidopsisHrabak et al., 2003). Sequence annotations have revealed 34 CDPKenes in Arabidopsis and eight additional CDPK-related kinases, andikewise, multigene families encoding CDPKs are present acrosseveral other plant species (Ray et al., 2007; Li et al., 2008). Spa-iotemporal regulation of CDPK expression and isoform-specificerception of Ca2+ signatures may have a significant bearing onignal transduction outputs modulating plant development andesponses to the prevalent environmental cues.
A signaling function for Ca2+, above and beyond its nutritiveole, was documented during somatic embryogenesis in carrotOvervoorde and Grimes, 1994). Likewise, a CDPK was identified inandalwood embryogenic cultures that accumulated to high levelsuring somatic and zygotic embryogenesis, endosperm develop-ent and seed germination, and was undetectable in shoots and
owers (Anil et al., 2000). Two rice CDPK isoforms, OsCDPK2nd OsCDPK11, were differentially expressed during caryopsisevelopment (Frattini et al., 1999) and transgenic overexpres-ion of OsCDPK2 disrupted early seed development (Morello et al.,000). Transgenic suppression of rice CDPK, OsCDPK23, producedeeds with starch-deficient endosperm, suggestive of endosperm-pecific activation of sucrose synthase (SuSy) (Shimada et al., 2004).
broad functional analysis for the CDPK gene family members inice (Ray et al., 2007) and wheat (Li et al., 2008) showed discreets well as partially overlapping tissue-specific expression profilesuring reproductive development.
A partial cDNA (GenBank acc. Y18055) encoding a monomericDPK (Mr 53,000) and requiring Ca2+-dependent autophospho-ylation for activity was characterized from mature peanut seedChaudhuri et al., 1999). A homologous cDNA clone isolated fromultured peanut cells (GenBank acc. DQ074454) revealed a bipar-ite nuclear localization signal (NLS) within the autoinhibitoryegion in addition to atypical Ca2+-binding domains (Raichaudhurit al., 2006). Since hypogeal reproductive development in peanuts largely contingent upon the sensing and response of develop-ng fruit to soil Ca2+ levels, CDPK expression was examined duringruit development across Ca2+-sufficient and -deficient environ-
ents. Sugar analyses and qRT-PCR data for genes implicated inpoplastic sucrose cleavage (cell wall invertase, CWI) and uti-ization (SuSy; sucrose-phosphate synthase, SpS), and lipid bodyiogenesis (oleosin genes, AhOleo17.8 and AhOleo18.5) suggest aole for CDPK in maintaining source-to-sink assimilate unloadingnsuring reproductive success.
aterials and methods
lant material
Peanut (Arachis hypogaea L. cv. Georgia Green) plants wererown in a low soil Ca2+ location at the Plant Science Research
ology 168 (2011) 2272– 2277 2273
and Education Unit, University of Florida, Citra, FL. Soil samples(Candler fine sand; 97% sand-sized particles and 2% silt) wereanalyzed for mineral content 4 weeks before planting (Waters Agri-culture Laboratories, Camilla, GA). Gypsum (CaSO4, 25% Ca2+) (BenFranklin No. 1, United States Gypsum Company, Chicago, IL) wasapplied (1500 lb acre−1) twice at 30 and 60 days-after-planting(DAP). Developing fruits from both low Ca2+ and gypsum-treatedplots (221 and 2500 lb acre−1, respectively) were collected at 100DAP, scored for seed and pod developmental stages (Vim, veryimmature through Vma, very mature; according to Pattee et al.,1974) (supplementary Fig. 1) and flash frozen under liquid nitrogenuntil further analyses.
SDS-PAGE and protein gel blot analysis
Total proteins were isolated following the method of Anil et al.(2000) and quantified using the Dc Protein Assay kit (BioRad, Her-cules, CA). Pre-cast 4–12% NuPAGE gels (Invitrogen Corporation,Carlsbad, CA) were used for SDS-PAGE and proteins were blottedonto nitrocellulose membranes (GE Water and Process Technolo-gies, Trevose, PA) using the Mini Trans-Blot apparatus (BioRad).Protein gel blots were developed using the WesternBreeze® Chemi-luminescent immunodetection kit (Invitrogen Corp.) supplied withanti-rabbit alkaline phosphatase IgG-conjugate. Affinity purifiedpolyclonal CDPK antibodies against the calmodulin-like domain ofsoybean CDPK� were described in Bachmann et al. (1996).
Quantitative RT-PCR
Quantitative RT-PCR was performed using the Brilliant II SYBR®
Green qPCR mix on an Mx3000P platform (Stratagene, Agilent Tech-nologies, Santa Clara, CA) as previously described by Tirumalarajuet al. (2011). Elongation factor 1-alpha (EF1a) was used as the inter-nal reference gene for normalizing the transcript profiles followingthe 2−��Ct method for relative quantification of transcript abun-dance. Gene specific primers used for real-time PCR analyses arelisted in supplementary Table 1.
Carbohydrate analyses
Frozen tissue samples were homogenized in hot ethanol toseparate soluble sugars from starch. After centrifugation, the super-natant was used for glucose, sucrose and fructose analyses asdescribed by Datta et al. (2002). Starch content was determined inthe pellet fraction following amyloglucosidase digestion (EC 3.2.13,from Aspergillus niger, Roche Diagnostics, Indianapolis, IN), andquantification of released glucose moieties.
Immunolocalization of CDPK in peanut fruits
Immature fruits were harvested and fixed in cold formalin aceticalcohol (FAA; 3.7% formaldehyde, 5% acetic acid and 50% ethanol)for 24 h, followed by dehydration in a series of ethanol and tertiarybutyl alcohol (TBA), infiltration, and embedding in Paraplast Plusparaffin (Sherwood Medical Co., St. Louis, MO). Paraffin-embeddedtissues were sectioned with a rotary microtome (Microm 325, CarlZeiss, Germany), immunolabeled (using 1:3000 dilution of primaryantibody) and visualized using the SuperPictureTM Polymer Detec-tion Kit (Zymed, Invitrogen Immunodetection, Carlsbad, CA).
Results
Detailed biometric analyses describing effects of Ca2+-deficiencyon peanut reproductive development will be presented elsewhere.Although inadequate soil Ca2+ levels did not inhibit the devel-opmental progression of peanut pods (supplementary Table 2),

2274 M. Jain et al. / Journal of Plant Physiology 168 (2011) 2272– 2277
Fig. 1. Spatial and temporal specificity of CDPK expression during reproductive development in peanut. (a) Tissue specific expression of CDPK in vegetative and reproductiveorgans, and (b, c) effect of soil Ca2+ status on CDPK expression in seeds and pods during development. For protein gel blot analysis, the soybean CDPK� antiserum was usedat 1:6000 dilution. The qRT-PCR data are presented as average ± S.D., for three independently made RT preparations with each having three technical replicates. Transcripta om Car coredV
CiautpteadrhiCu
Fluv
bundance was calibrated against Vim (stage 1) seeds (a, b) and pods (c) obtained frow plot were pooled together, and the seed and pod developmental stages were sma, very mature; supplementary Fig. 1).
a2+-deficiency had a noticeably deleterious effect on seed fill-ng, and significantly increased the incidence of pod rot and seedbortion resulting in “pops” or empty fruits, or fruits with severelynderdeveloped seeds (supplementary Table 3). The discrimina-ory effect of soil Ca2+-status on developmental profiles of peanutod and seed tissues was further confirmed by qRT-PCR and pro-ein gel blot analyses. Quantitative RT-PCR data showed that CDPKxpression in developing fruits is fertilization-dependent, spatiallynd temporally regulated, and is also responsive to soil Ca2+ con-itions (Fig. 1). Expression of CDPK was observed to be mainlyestricted to underground organs of the plant such as roots and
ypocotyls, whereas leaves and unfertilized flowers showed min-mal CDPK expression and protein. Post-fertilization increase inDPK transcription in aerial pegs was followed by significantpregulation during penetration of pegs into the soil (Fig. 1a). In
ig. 2. Immunolocalization of CDPK in transverse sections of young (Vim, very immatureayers of exocarp and vascular bundles and (c) vascular supply to the developing seed, andsed at 1:3000 dilution (magnification bar: 1 mm (a, c and d), 0.25 mm (b); ex, exocarp; mt, vascular trace supplying the seed; lc, lateral vascular connections).
2+-sufficient soil. Developing fruits from 6 randomly collected plants from 1 m long according to Pattee et al. (1974) (Vim, very immature; Im, immature; Ma, mature;
developing fruits, CDPK expression showed a bimodal transcriptprofile through seed maturation, irrespective of soil Ca2+ status(Fig. 1b). Vim and Ma seed stages showed high CDPK expression,with significantly lower levels in intermediary Im and the lowestlevels in the desiccated Vma stages. Under Ca2+-deficient condi-tions, a 2 to 3-fold increase in CDPK transcript abundance wasdiscernible across all stages of seed development. Unlike devel-oping seed, a basal level of CDPK transcriptional activity wasmaintained during pod development, and was unresponsive to soilCa2+ levels (Fig. 1c). Vma stages, characterized by the onset of fruitdesiccation, consistently showed negligible CDPK expression. Pro-
tein gel blot analysis corroborated the qRT-PCR data thus excludingthe likelihood of posttranscriptional regulation of CDPK expressionduring development of either the peanut seed or the pod. Cell-specific localization of immunoreactive CDPK in transverse sections; stage 1) fruit of peanut. a–c: immunolocalization of CDPK in (a, b) subepidermal d: represents the pre-immune control. The primary soybean CDPK� antiserum was
, mesocarp; en, endocarp; s, developing seed; sc, seed coat; vb, vascular bundles;
M. Jain et al. / Journal of Plant Physiology 168 (2011) 2272– 2277 2275
048
121620
048
121620
020406080
100
468
10121416see ds
pods
glucose fructose sucrosea
b
cVim Im Ma Vma
µg su
gar m
g-1FW
µg st
arch
mg-1
FWµg
suga
r mg-1
FW
Vim Im Ma VmaDevelopmental stages
see ds
pods
d
e
f
0.0
0.5
1.0
1.5
2.0
Vim Im Ma Vma Vi m Im Ma VmaTran
scrip
t abu
ndan
ce
see dspods
CWI1 CWI2
Tran
scrip
t abu
ndan
ceTr
ansc
ript a
bund
ance
Vim Im Ma VmaDevelopmental stages
Oleo17.8Oleo18.5
012345
0
1
2
3
4see dspods
SuSy SpSsee dspods
Vim Im Ma VmaDevelopmental stages
Vim Im Ma Vma
Developmental stages
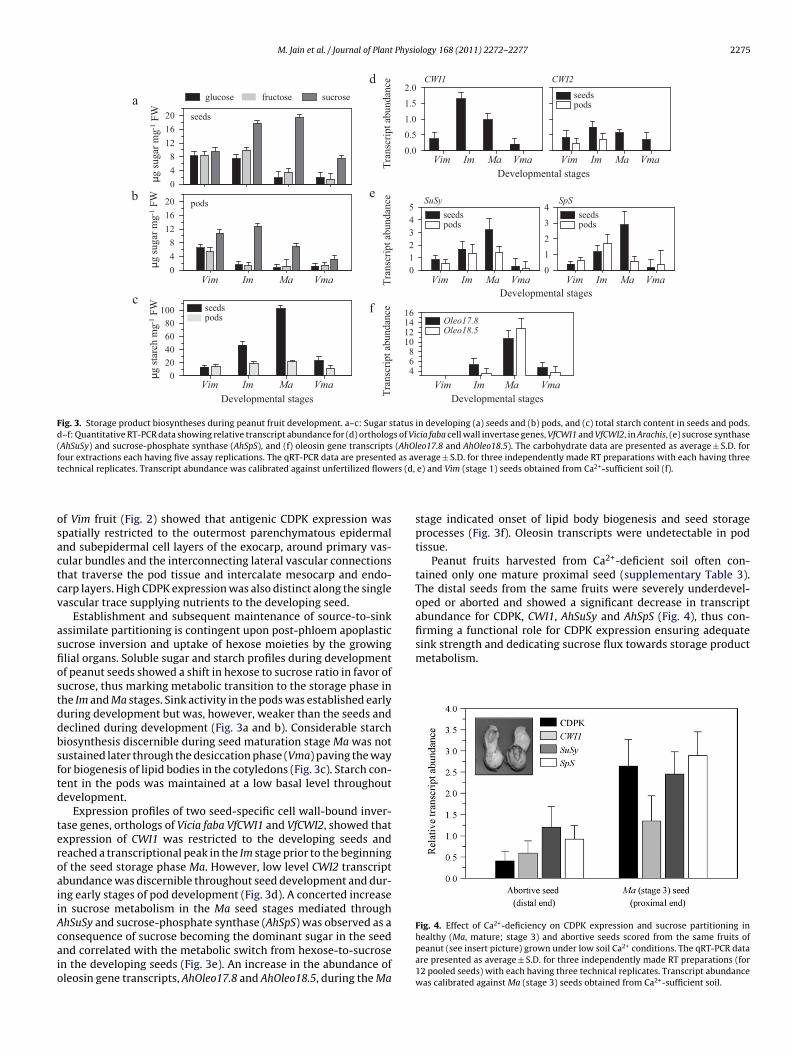
Fig. 3. Storage product biosyntheses during peanut fruit development. a–c: Sugar status in developing (a) seeds and (b) pods, and (c) total starch content in seeds and pods.d–f: Quantitative RT-PCR data showing relative transcript abundance for (d) orthologs of Vicia faba cell wall invertase genes, VfCWI1 and VfCWI2, in Arachis, (e) sucrose synthase(AhSuSy) and sucrose-phosphate synthase (AhSpS), and (f) oleosin gene transcripts (AhOleo17.8 and AhOleo18.5). The carbohydrate data are presented as average ± S.D. forfour extractions each having five assay replications. The qRT-PCR data are presented as average ± S.D. for three independently made RT preparations with each having threet rs (d,
osactcv
asfiostddbsftd
teroaiiAcaio
firming a functional role for CDPK expression ensuring adequatesink strength and dedicating sucrose flux towards storage productmetabolism.
Fig. 4. Effect of Ca2+-deficiency on CDPK expression and sucrose partitioning in
echnical replicates. Transcript abundance was calibrated against unfertilized flowe
f Vim fruit (Fig. 2) showed that antigenic CDPK expression waspatially restricted to the outermost parenchymatous epidermalnd subepidermal cell layers of the exocarp, around primary vas-ular bundles and the interconnecting lateral vascular connectionshat traverse the pod tissue and intercalate mesocarp and endo-arp layers. High CDPK expression was also distinct along the singleascular trace supplying nutrients to the developing seed.
Establishment and subsequent maintenance of source-to-sinkssimilate partitioning is contingent upon post-phloem apoplasticucrose inversion and uptake of hexose moieties by the growinglial organs. Soluble sugar and starch profiles during developmentf peanut seeds showed a shift in hexose to sucrose ratio in favor ofucrose, thus marking metabolic transition to the storage phase inhe Im and Ma stages. Sink activity in the pods was established earlyuring development but was, however, weaker than the seeds andeclined during development (Fig. 3a and b). Considerable starchiosynthesis discernible during seed maturation stage Ma was notustained later through the desiccation phase (Vma) paving the wayor biogenesis of lipid bodies in the cotyledons (Fig. 3c). Starch con-ent in the pods was maintained at a low basal level throughoutevelopment.
Expression profiles of two seed-specific cell wall-bound inver-ase genes, orthologs of Vicia faba VfCWI1 and VfCWI2, showed thatxpression of CWI1 was restricted to the developing seeds andeached a transcriptional peak in the Im stage prior to the beginningf the seed storage phase Ma. However, low level CWI2 transcriptbundance was discernible throughout seed development and dur-ng early stages of pod development (Fig. 3d). A concerted increasen sucrose metabolism in the Ma seed stages mediated throughhSuSy and sucrose-phosphate synthase (AhSpS) was observed as a
onsequence of sucrose becoming the dominant sugar in the seednd correlated with the metabolic switch from hexose-to-sucrosen the developing seeds (Fig. 3e). An increase in the abundance ofleosin gene transcripts, AhOleo17.8 and AhOleo18.5, during the Mae) and Vim (stage 1) seeds obtained from Ca2+-sufficient soil (f).
stage indicated onset of lipid body biogenesis and seed storageprocesses (Fig. 3f). Oleosin transcripts were undetectable in podtissue.
Peanut fruits harvested from Ca2+-deficient soil often con-tained only one mature proximal seed (supplementary Table 3).The distal seeds from the same fruits were severely underdevel-oped or aborted and showed a significant decrease in transcriptabundance for CDPK, CWI1, AhSuSy and AhSpS (Fig. 4), thus con-
healthy (Ma, mature; stage 3) and abortive seeds scored from the same fruits ofpeanut (see insert picture) grown under low soil Ca2+ conditions. The qRT-PCR dataare presented as average ± S.D. for three independently made RT preparations (for12 pooled seeds) with each having three technical replicates. Transcript abundancewas calibrated against Ma (stage 3) seeds obtained from Ca2+-sufficient soil.

2 Physi
D
bigfittscaeepewaip
ug1tyotCCtCaara(pmr
eloptmg2omsgp(icmmStuFmia
276 M. Jain et al. / Journal of Plant
iscussion
Significant CDPK expression in young developmental stages, inoth seeds and pods, and declining considerably in mature des-
ccated fruit, is reflective of its high metabolic status and steeprowth kinetics. Notably, young seeds showed a bimodal pro-le for CDPK transcript and protein abundance in sharp contrasto pods (Fig. 1b and c). While seed mitotic activity is restrictedo early developmental stages followed by cell expansion andtorage product accumulation later on, pods have to undergoontinued cellular proliferation and rapid expansion in order toccommodate cotyledonary growth during the storage phase, thusxplaining consistently high CDPK expression in pods throughoutarly development. Histological evidence for CDPK expression inod confirmed that the pods can absorb Ca2+ directly through thepidermal layers in addition to that supplied via the xylem route,hereas the growing seeds are solely dependent upon xylem-
vailable Ca2+ (Fig. 2). Transcriptional upregulation of CDPK onlyn seeds developing under inadequate Ca2+ conditions in the soilegging zone supports this argument (Fig. 1b and c).
Mitotic growth of young developing seeds may be dependentpon CDPK activity as suggested by Ca2+-dependent dynamic reor-anization of cytoskeleton elements in maize (Smertenko et al.,998) and soybean (Lee et al., 2003a). Lee et al. (2003b) iden-ified NtRpn3 and NtCDPK1 as interacting partners following aeast two-hybrid screen in tobacco. Rpn3, a regulatory subunitf 26S proteasome involved in proteolysis of cell cycle regula-ory proteins, physically interacts with NtCDPK1 in vitro in aa2+-independent manner while undergoing phosphorylation in aa2+-dependent manner, suggesting a role for CDPK in the con-rol of cell division and differentiation. Developmental polarity ofWI-mediated apoplastic sucrose inversion generates appropri-te sugar signals, a pre-requisite for maintaining mitotic status inctively growing sink tissues (Cheng and Chourey, 1999). A tempo-al overlap in the transcriptional profiles of CDPK and CWI suggests
congruent role in control of mitotic growth of immature seedsFigs. 1b and 3d). Appraisal of brassinosteroid signaling pathwayrovides additional evidence linking CDPK expression and CWI-ediated assimilate partitioning in determining the source/sink
egulation (Friedrichsen and Chory, 2001).In addition to a role in early mitotic growth, elevated CDPK
xpression (Fig. 1b) may have a significant bearing on starch andipid biosyntheses reactions (Fig. 3) during peanut seed devel-pment. SuSy-mediated reversible conversion of sucrose in theresence of UDP to UDP-glucose and fructose is the commit-ed step in storage starch biosynthesis, and marks cessation of
itotic activity, transcriptional activation of storage-associatedenes and the beginning of starch accumulation (Weber et al.,005). Phosphorylation of SuSy by CDPK is implicated in activationf SuSy-dependent assimilate unloading during transition fromitotic to the growth/expansion and storage phase. Antisense-
uppression of CDPK in transgenic rice showed normal vegetativerowth and reproductive phase transition, but displayed com-romised sink strength and failed to accumulate storage starchShimada et al., 2004). Transient upregulation of a sucrose-nducible CDPK, and transcript localization in sucrose accumulatingell layers were observed in potato stolon tips during tuber for-ation (Raíces et al., 2001). Anguenot et al. (2006) reportedembrane-associated and CDPK-mediated phosphorylation of
uSy in young tomato fruits. Regulation of primary metabolismhrough phosphorylation of SpS by CDPK is somewhat poorlynderstood (Pagnussat et al., 2002) and merits further attention.
inally, higher CDPK expression in the Ma seed stage (Fig. 3f)ay also be related to the biogenesis of lipid bodies accumulat-ng during storage phase. Oleosins (AhOleo17.8 and AhOleo18.5)re small molecular mass proteins located on the oil body surface
ology 168 (2011) 2272– 2277
preventing the phospholipid layers from contacting and coalescingwith each other (Li et al., 2009), and co-precipitate with CDPK in theoil body-associated protein fraction of mature peanut seeds (Anilet al., 2003).
Comparative analysis in aborted and developmentally normalseeds scored from the same pods grown under Ca2+-deficient con-ditions (Fig. 4) further reiterates the functional association betweenCDPK expression, assimilate unloading and utilization, thus inte-grating soil Ca2+ cues and developmental progression. Notably,after soil penetration of the pegs, initial mitotic activity and cel-lular expansion is restricted to the proximal end of the fertilizedovary that helps in positioning the developing pod horizontally. Itis not surprising that in an event of compromised Ca2+ status, devel-opmental progression of only one fertilized ovule at the same endis favored to allow pragmatic utilization of available assimilates.
In conclusion, evidence is provided that (a) CDPK expression isresponsive to soil Ca2+ status only in the developing peanut seed,as opposed to the pod tissue which is capable of absorbing Ca2+
directly from the soil solution; and (b) quantitative changes in spa-tiotemporal expression of CDPK during maturation of geocarpicpeanut fruits reflects its functional correlation with assimilatepartitioning and utilization for storage product biosyntheses. How-ever, the possibility of multiple CDPK isoforms addressing diverseenvironmental and nutritional signals, as well as tissue-specificcues during peanut fruit development cannot be ruled out in ensur-ing reproductive success.
Acknowledgements
Authors wish to thank J. McKinney and C. J. Boggs at the PlantScience Research and Education Unit, Citra, FL for their help with themaintenance of field plots. This research was partially supported bythe Florida Peanut Producers Association.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.jplph.2011.07.005.
References
Anguenot R, Nguyen-Quoc B, Yelle S, Michaud D. Plant Physiol Biochem2006;44:294–300.
Anil VS, Harmon AC, Rao KS. Plant Physiol 2000;122:1035–43.Anil VS, Harmon AC, Rao KS. Plant Cell Physiol 2003;44:367–76.Bachmann M, Shiraishi N, Campbell WH, Yoo BC, Harmon AC, Huber SC. Plant Cell
1996;8:505–17.Chaudhuri S, Seal A, DasGupta M. Plant Physiol 1999;120:859–66.Cheng WH, Chourey PS. Theor Appl Genet 1999;98:485–95.Datta R, Chamusco KC, Chourey PS. Plant Physiol 2002;130:1645–56.Frattini M, Morello L, Breviario D. Plant Mol Biol 1999;41:753–64.Friedrichsen D, Chory J. BioEssays 2001;23:1028–36.Gascho GJ, Parker MB. Agron J 2001;93:1305–15.Harper JF, Harmon A. Nat Rev Mol Cell Biol 2005;6:555–66.Hepler PK. Plant Cell 2005;17:2142–55.Hirschi KD. Plant Physiol 2004;136:2438–42.Hrabak EM, Chan CWM, Gribskov M, Harper JF, Choi JH, Halford N, et al. Plant Physiol
2003;132:666–80.Kudla J, Oliver Batistic O, Hashimoto K. Plant Cell 2010;22:541–63.Lee S, Park J, Lee Y. Mol Cells 2003a;15:313–9.Lee SS, Cho HS, Yoon GM, Ahn J-W, Kim H-H, Pai H-S. Plant J 2003b;33:825–40.Li A-L, Zhu Y-F, Tan X-M, Wang X, Wei B, Guo H-Z, et al. Plant Mol Biol
2008;66:429–43.Li C, Wu K, Fu G, Li Y, Zhong Y, Lin X, et al. J Exp Bot 2009;60:4371–82.Morello L, Frattini M, Gianì S, Christou P, Breviario D. Transgenic Res 2000;9:453–62.Murata MR, Zharare GE, Hammes PS. J Plant Nutr 2008;31:69–79.Overvoorde PJ, Grimes HD. Plant Cell Physiol 1994;35:135–44.Pagnussat GC, Fiol DF, Salemo GL. Physiol Plant 2002;115:183–9.
Pattee HE, Johns EB, Singleton JA, Sanders TH. Peanut Sci 1974;1:57–62.Raíces M, Chico JM, Téllez-Inón MT, Ulloa RM. Plant Mol Biol 2001;46:591–601.Raichaudhuri A, Bhattacharya R, Chaudhuri S, Chakrabarti P, DasGupta M. J Biol Chem
2006;281:10399–409.

Physi
R
RS
S
Weber H, Borisjuk L, Wobus U. Annu Rev Plant Biol 2005;56:253–79.
M. Jain et al. / Journal of Plant
ay S, Agarwal P, Arora R, Kapoor S, Tyagi AK. Mol Genet Genomics 2007;278:493–505.
eddy TY, Reddy VR, Anbumozhi V. Plant Growth Regul 2003;41:75–88.himada H, Koishihara H, Saito Y, Arashima Y, Furukawa T, Hayashi H. Plant Cell
Physiol 2004;45:1105–9.mertenko AP, Jiang C-J, Simmons NJ, Weeds AG, Davies DR, Hussey PJ. Plant J
1998;14:187–93.
ology 168 (2011) 2272– 2277 2277
Tirumalaraju SV, Jain M, Gallo M. J Plant Physiol 2011;168:481–92.
Wright DL, Tillman B, Jowers E, Marois J, Ferrell JA, Katsvairo T, et al. Agron-omy Department, Florida Cooperative Extension Service, Institute of Food andAgricultural Sciences, University of Florida; 2009. SS-AGR-74 (http://if-srvv-edis.ifas.ufl.edu/pdffiles/AA/AA25800.pdf).