c-type lectin receptors differentially induce th17 cells and
TRANSCRIPT

of March 28, 2018.This information is current as
the Endemic Mycosis of North AmericaInduce Th17 Cells and Vaccine Immunity to C-Type Lectin Receptors Differentially
Klein and Marcel WüthrichGalles, Shinobu Saijo, Xin Lin, Garry T. Cole, Bruce S. Huafeng Wang, Vanessa LeBert, Chiung Yu Hung, Kevin
http://www.jimmunol.org/content/192/3/1107doi: 10.4049/jimmunol.1302314January 2014;
2014; 192:1107-1119; Prepublished online 3J Immunol
MaterialSupplementary
4.DCSupplementalhttp://www.jimmunol.org/content/suppl/2014/01/03/jimmunol.130231
Referenceshttp://www.jimmunol.org/content/192/3/1107.full#ref-list-1
, 22 of which you can access for free at: cites 61 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2014 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
C-Type Lectin Receptors Differentially Induce Th17 Cells andVaccine Immunity to the Endemic Mycosis of North America
Huafeng Wang,*,1 Vanessa LeBert,*,1 Chiung Yu Hung,† Kevin Galles,* Shinobu Saijo,‡
Xin Lin,x Garry T. Cole,† Bruce S. Klein,*,{,‖ and Marcel Wuthrich*
Vaccine immunity to the endemic mycoses of North America requires Th17 cells, but the pattern recognition receptors and signaling
pathways that drive these protective responses have not been defined. We show that C-type lectin receptors exert divergent con-
tributions to the development of antifungal Th17 cells and vaccine resistance against Blastomyces dermatitidis, Histoplasma
capsulatum, and Coccidioides posadasii. Acquired immunity to B. dermatitidis requires Dectin-2, whereas vaccination against
H. capsulatum and C. posadasii infection depends on innate sensing by Dectin-1 and Dectin-2, but not Mincle. Tracking Ag-specific
T cells in vivo established that the Card9 signaling pathway acts indispensably and exclusively on differentiation of Th17 cells,
while leaving intact their activation, proliferation, survival, and migration. Whereas Card9 signaling is essential, C-type lectin
receptors offer distinct and divergent contributions to vaccine immunity against these endemic fungal pathogens. Our work
provides new insight into innate immune mechanisms that drive vaccine immunity and Th17 cells. The Journal of Immunology,
2014, 192: 1107–1119.
Despite several million new systemic fungal infectionsannually worldwide, there are no commercial vaccinesavailable. We and others have engineered vaccines that
protect against experimental infection with the dimorphic fungalpathogens Blastomyces dermatitidis (1), Histoplasma capsulatum(2), and Coccidioides posadasii (3). Our studies showed that vaccine-induced immunity is chiefly mediated by CD4+ T cells (4). Despitethe crucial roles of Th1 cells in protective immunity against fungalinfection (3, 5, 6) and the controversial roles of Th17 cells in someother infection models (7–13), in our vaccination model Th1 im-munity is dispensable, whereas fungus-specific Th17 cells arenecessary and sufficient for vaccine-induced protection againstthese three pathogenic fungi that cause the major endemic mycosesof North America (14). Thus, engaging Th17 cells could be a prom-ising strategy to develop effective fungal vaccines. However, themechanisms underlying the vaccine-induced Th17 immunity are
still largely unknown and need to be determined to develop ra-tionale strategies for antifungal vaccines.Fungi-specific T cell responses are initiated through the rec-
ognition of pathogen-associated molecular patterns by pattern
recognition receptors (PRRs) on innate immune cells. Among the
best-characterized PRRs that recognize fungi are the ITAM-
coupled receptors Dectin-1, Dectin-2, and Mincle. They are C-
type lectin receptors (CLRs), which are predominantly expressed
in myeloid cells (15, 16). There is accumulating evidence that
stimulation of the most-studied CLR, Dectin-1, by b-glucans
induces Th17 differentiation of naive CD4+ T cells (17). Recently,
Viriyakosol et al. (18) showed that Dectin-12/2 mice infected with
Coccidioides immitis have lower levels of Th17 cytokines in their
lungs. Patients who are homozygous for a single polymorphism of
Dectin-1 are susceptible to mucocutaneous Candida albicans infec-
tions (19, 20) and invasive aspergillosis (21, 22) owing to defective
IL-17 production. We have found, however, that Dectin-1 is un-
expectedly dispensable in the development of vaccine-induced
Th17 cell responses and resistance to B. dermatitidis (14). It is un-
known whether Dectin-1 is required for the development of vaccine-
induced Th17 cells and resistance to H. capsulatum and C. posadasii
infection.In contrast to Dectin-1, few reports describe the role of Dectin-2
in driving Th17 responses. In mice, Dectin-2 is required for the
differentiation of Th17 cells induced by C. albicans infection (23).
In human dendritic cells (DCs), Dectin-2 activation by Candida
results in the selective activation of the NF-kB subunit c-Rel and
the production of IL-1b and IL-23 p19, which skews CD4+ T cell
responses toward a Th17 profile (24). Although Mincle has been
reported to induce Th1/Th17 immunity in response to the myco-
bacterial cell wall glycolipid trehalose 6,69-dimycolate (TDM)
and its synthetic analog trehalose-6,6-dibehenate (TDB) (25), to
our knowledge its role in driving anti-fungal Th17 responses has
not been investigated.Although Dectin-1 recognizes fungi via b-1,3-glucan exposed on
the cell wall and recruits Syk directly through its hemITAM motif
(26), Dectin-2 and Mincle recognize mannose-like structures (23,
27–29) and need to pair with the ITAM-bearing adaptor FcRg to
*Department of Pediatrics, University of Wisconsin Medical School, University ofWisconsin Hospital and Clinics, Madison, WI 53792; †Department of Biology andSouth Texas Center for Emerging Infectious Diseases, University of Texas, SanAntonio, TX 78249; ‡Center for Experimental Medicine and Systems Biology, Uni-versity of Tokyo, Tokyo 108-8639 Japan; xDepartment of Molecular and CellularOncology, University of Texas, M.D. Anderson Cancer Center, Houston, TX77030; {Internal Medicine, University of Wisconsin Medical School, Universityof Wisconsin Hospital and Clinics, Madison, WI 53792; and ‖Medical Microbiologyand Immunology, University of Wisconsin Medical School, University of WisconsinHospital and Clinics, Madison, WI 53792
1H.W. and V.L. contributed equally to this work.
Received for publication September 3, 2013. Accepted for publication November 29,2013.
This work was supported by National Institutes of Health Grants R01 AI093553(to M.W.) and R01 AI071118 (to G.T.C.).
Address correspondence and reprint requests to Dr. Marcel Wuthrich, Universityof Wisconsin, Microbial Sciences Building, 1550 Linden Drive, Madison, WI53706. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: BMDC, bone marrow–derived dendritic cell; CLR,C-type lectin receptor; DC, dendritic cell; FKS, formalin-killed spherule; LN, lymphnode; PRR, pattern recognition receptor; sdLN, skin-draining lymph node; TDB,trehalose-6,6-dibehenate; TDM, trehalose 6,69-dimycolate; Tg, transgenic.
Copyright� 2014 by The American Association of Immunologists, Inc. 0022-1767/14/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1302314
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

activate the Syk-Card9 pathway (30–32). In mice, Card9 signalinginduces DC maturation, the production of proinflammatory cyto-kines, and the induction of Th17 responses (17). In humans, a Card9mutation results in susceptibility to chronic mucocutaneous can-didiasis (33). Notably, C. albicans–induced Th17 responses aredependent on Card9 (17), and greatly reduced numbers of Th17cells are associated with patients carrying homozygous Card9mutations (33), indicating that Card9 is essential for antifungalTh17 responses. However, it is unclear at what stage the Card9signaling pathway governs Th17 cell development in vivo. Todissect whether Card9 affects the priming, expansion, differentiation,contraction, and migration of vaccine-induced Th17 cells, weexploited an adoptive transfer system that we have previouslyestablished using TCR transgenic 1807 cells that recognize a sharedAg from B. dermatitidis, H. capsulatum, and C. posadasii (34). Onadoptive transfer into recipient mice, 1807 cells become activated,proliferate, and expand in the draining lymph node (LN); 1807 cellsdifferentiate into cytokine-producing effector T cells after traffick-ing to the site of vaccination and the lung upon challenge andconfer resistance against the three dimorphic fungi (14, 34, 35).Thus, the autologous adoptive transfer system offers a powerful toolto dissect normal or defective development of vaccine-induced Ag-specific T cells responsive to multiple dimorphic fungi.Although Card9 and CLRs have been implicated in mediating
innate resistance to primary fungal infection and priming of Th17cells, their role in vaccine-induced resistance to fungi and the effecton the sequential stages of T cell development has not been in-vestigated. In this study, we demonstrate that the adaptor Card9is indispensable for the acquisition of vaccine immunity and thedevelopment of Th17 cells against all three systemic dimorphicfungi of North America, but the upstream CLRs play distinctlydifferent roles for each pathogen. We also pinpoint at what stage ofthe immune response Card9 controls Th17 cell development andshow that this adaptor promotes the differentiation of antifungalTh17 cells, but does not influence downstream stages of T cellexpansion, activation, contraction, or migration to the lung uponchallenge. With B. dermatitidis, the upstream PRR Dectin-2, butneither Mincle nor Dectin-1, recognizes yeast and induces ITAMsignaling that initiates the development of Th17 cells and vaccineimmunity to fungal infection. Conversely, for the related dimor-phic fungi H. capsulatum and C. posadasii, while Card9 is re-quired for the induction of vaccine immunity, both Dectin-1 andDectin-2, but not Mincle, participate in the recognition and in-duction of protective Th17 cell responses. Our data offer insightinto the mechanisms that underpin acquisition of vaccine immu-nity to pathogenic fungi.
Materials and MethodsEthics statement
All animal procedures were performed in accordance with the recom-mendations in the Guide for the Care and Use of Laboratory Animals of theNational Institutes of Health. Care was taken to minimize animal suffering.The work was done with the approval of the Institutional Animal Care andUse Committee of the University of Wisconsin–Madison.
Mouse strains
Inbred wild type C57BL/6 and congenic B6.PL-Thy1a/Cy (stock #00406)mice carrying the Thy 1.1 allele were obtained from Jackson Laboratories(Bar Harbor, ME). B6.129P2-Fcer1gtm1Rav N12 mice (model no. 583) thatlack FcRg were purchased from Taconic. Blastomyces-specific TCR trans-genic (Tg) 1807 mice were generated in our laboratory and were back-crossed to congenic Thy1.1+ mice as described elsewhere (35). Card92/2
(36), Dectin-22/2 (23), Mincle2/2 (37), and Syk+/2 (38) were bred at ourfacility. Radiation chimeras were generated by transferring fetal liver cellsfrom day 15–17 Syk2/2 and Syk+/+ embryos (womb mates that wereidentified by PCR) into lethally irradiated C57BL6 recipients (38). Syk+/2
mice were a generous gift from Dr. Victor L. J. Tybulewicz. All mice were7–8 wk old at the time of experiments. Mice were housed and cared foraccording to guidelines of the University of Wisconsin Animal CareCommittee, who approved all aspects of this work.
Fungi and growth conditions
B. dermatitidis strains used were ATCC 26199, a wild type virulent strain,and the isogenic, attenuated mutant lacking BAD1, designated strain #55(39). Isolates of B. dermatitidis were maintained as yeast on Middlebrook7H10 agar with oleic acid-albumin complex (Sigma, St. Louis, MO) at39˚C. H. capsulatum strain G217B (provided by George S. Deepe, Uni-versity of Cincinnati, Cincinnati, OH) was maintained on brain-heart in-fusion slants supplemented with 5% sheep blood at 37˚C. C. posadasiiisolate C735 is a virulent, human clinical strain that was used to generatea live, attenuated vaccine strain (DT) by deleting the chitinases 2, 3, andD-arabinotol-2-dehydrogenase genes as described previously (3). Thesaprobic phase of both the parental and mutant strains were grown onglucose yeast extract medium (1% glucose, 0.5% yeast extract, 1.5% agar)at 30˚C for 3–4 wk to generate a confluent layer of arthroconidia (spores)on the agar surface. Formalin-killed endosporulated spherules of C. pos-adasii were generated as described (40).
Vaccinations
Mice were vaccinated s.c. at two sites: dorsally and at the base of the tail asfollows. B. dermatitidis virulence attenuated BAD12/2 yeast, designatedstrain #55, were injected as live or heat-killed cells using a dose range of106–107 yeast per mouse. For vaccination with H. capsulatum G217B, 107
heat-killed or 106 live yeast per mouse was used. For vaccination withC. posadasii, mice were immunized with 5.0 3 104 viable spores of theDT strain (3). Formalin-killed spherules (FKS) of C. posadasii were usedat 106 per mouse for vaccination. Resistance experiments included one(B. dermatitidis and H. capsulatum) or two (C. posadasii) booster vacci-nation(s) of the same vaccine dose and site 2 weeks apart.
Experimental infection
Mice were infected intratracheally with 2 3 103 or 2 3 104 wild type26199 B. dermatitidis yeast as described previously (14). At day 4 afterinfection, coinciding with the peak of T cell influx (14), the mice weresacrificed and lung T cells were analyzed by FACS analysis. Extent of lunginfection was examined 2 wk after infection and determined by platinghomogenized lung and enumerating yeast CFU on brain-heart infusion(Difco) agar. For H. capsulatum infections, mice were challenged with a sub-lethal dose of 1–2 3 105 G217B intratracheally, and cellular analysis wasperformed 4 d later. Lung burden was determined 13 d after infection byplating lung homogenates on sheep blood containing Mycosel plates andenumerating CFUs. For C. posadasii infections, mice were challenged bythe intranasal route with a lethal dose of 50–80 spores of the virulent,parental strain 4 wk after completion of the vaccination protocol (34). Thefungal burden determined as CFUs in lungs and spleens of the vaccinatedand unvaccinated mice was conducted at day 14 after infection by platingorgan homogenates on glucose yeast extract plates containing 50 mg/mlchloramphenicol.
Adoptive transfer of transgenic 1807 T-cells and surfacestaining
Single-cell suspensions of 106 magnetic bead purified CD4+ cells from1807 Tg Thy1.1+ mice were injected i.v. into Thy1.2+ C57BL/6 recipients.When indicated, transgenic 1807 cells were labeled with CFSE (MolecularProbes) before adoptive transfer. After vaccination or infection, single-cellsuspensions from draining lymph nodes (brachial and inguinal) and lungcells of recipient mice were stained with mAbs directed against the fol-lowing surface markers: CD4, CD8, Thy1.1, CD44, CD62L, and B220 (asa dump marker to exclude events that are stained nonspecifically). mAbswere obtained from BD Pharmingen (San Diego, CA) and eBioscience(San Diego, CA), and cytometry data were gathered with an LSRII FlowCytometer (BD Biosciences, San Jose). Data were analyzed with FlowJosoftware (Tree Star, Ashland, OR). The number of 1807 CD4+ T cells ina lung was calculated by multiplying the percentage of Thy1.1+ CD4+ cellsby the number of viable cells as determined by trypan blue dye exclusion.
Intracellular cytokine staining
Effector T cells from the site of vaccination and lung were obtained asdescribed previously (14). An aliquot of isolated cells was stained forsurface CD4 and Thy 1.1 to determine the percentage of transferred 1807cells. The numbers of 1807 cells in the lung was derived by multiplying thepercentage of cells by the total number of cells per organ isolated. The rest
1108 DECTIN-2 INDUCES ANTIFUNGAL T CELL IMMUNITY
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

of the cells were stimulated with anti-CD3 and anti-CD28 mAb in thepresence of Golgi-Stop (BD Pharmingen). After 4–6 h of stimulation, cellswere stained for surface markers, fixed, and permeabilized using the inCytofix/Cytoperm kit (BD Pharmingen) and stained with anticytokine Absas described (14).
Generation of bone marrow dendritic cells
Bone marrow–derived dendritic cells (BMDCs) were obtained from thefemurs and tibias of individual mice. Each bone was flushed with 10 mlof 1% FBS in RPMI 1640 through a 22-gauge needle. RBCs were lysedfollowed by wash and resuspension of cells in 10% FBS in RPMI 1640medium. In a Petri dish, 2 3 106 bone marrow cells were plated in 10 mlof RPMI 1640 containing 10% FBS plus penicillin-streptomycin (HyClone),2-ME, and 20 ng/ml of rGM-CSF. The culture media were refreshed every3 d, and BMDCs were harvested after 10 d for in vitro coculture assays.
Ex vivo and in vitro coculture for cytokine protein measurement
Ex vivo cell culture supernatants were generated using the brachial andinguinal draining lymph nodes harvested from mice 10 d after vaccination,washed, and resuspended in complete RPMI 1640 containing 10 mg/mlyeast cell wall membrane Ag (CW/M) (1), and plated in 96-well plates ata concentration of 5 3 105 cells/well. Supernatants were collected fromex vivo cocultures after three days of incubation at 37˚C and 5% CO2 (14).In vitro cell culture supernatants were generated using BMDCs plated incomplete RPMI 1640 at a concentration of 105 cells/well with 3 3 105
heat-killed strain #55 yeast overnight. After 24 h, single-cell suspensionsof magnetic bead purified CD4+ naive 1807 cells from brachial, axillary,cervical, and inguinal lymph nodes were added at a concentration of2 3 105 cells/well. Supernatants were collected from in vitro coculturesafter three days of incubation at 37˚C and 5% CO2. IFN-g and IL-17(R&D Systems) were measured by ELISA according to the manu-facturer’s specifications (detection limits, 0.05 ng/ml and 0.02 ng/ml,respectively).
Isolation of effector cells at the site of vaccination
Mice were vaccinated once s.c. with 107 heat-killed strain #55 yeast ata single dorsal site. On day 10 after vaccination, inflamed s.c. tissue wasexcised from the site of vaccination, placed in ice-cold collagenase bufferand minced into fine pieces. To digest the tissue, 5 ml of dissociation buffer(0.025 mg/ml Liberase [Roche Diagnostics] and 50 mg/ml DNAse I [Sigma-Aldrich] in collagenase buffer) was added and samples were incubated at37˚C and 5% CO2 for 30 min. To release single cells further, the tissue wasmashed with the back of a 10-ml syringe plunger through a 70-mm cellstrainer with an additional 5 ml of dissociation buffer and incubated foranother 30 min. The dissolved tissue was washed with ice-cold PBScontaining 5 mM EDTA and 1% BSA and strained again. Filtered cellswere spun at 1500 rpm, the supernatant was carefully aspirated, and cellswere resuspended in complete media for stimulation.
RT-PCR analysis for the detection of Card9-associated genetranscripts
BMDCs were raised from 15 individual Card92/2 or wild type mice and3 3 105 nonadherent DCs from each individual mouse were coculturedwith 3 3 105 B. dermatitidis vaccine yeast or medium alone in a 24-wellplate for 24 h. Total RNA from the BMDC cultures described above waspurified using the Qiagen RNeasy kit (catalog #74106) according to themanufacturer’s protocol. Genomic DNA was removed using Turbo DNase(Ambion, catalog no. AM2238). Following DNase treatment, the RNAwascleaned up using Qiagen RNeasy columns according to the manufacturer’sprotocol and by performing additional washes with buffer RPE and 80%ethanol to remove contaminants. The n-fold change of gene expression foryeast stimulation versus no stimulation was calculated using the compar-ative CT method (41).
Generation of Fc fusion proteins
The Dectin-2-Fc expression vector pSecTag2(C)-mDectin-2-Fc was pro-vided by Dr. Philip R. Taylor (Cardiff University, Cardiff, U.K.) (27). Toconstruct the Mincle-Fc expression vector, the extracellular domain ofmouse Mincle (amino acids 46–214) was amplified by PCR from apEF1HISB-mMincle plasmid provided by Dr. Christine Wells (Universityof Queensland, Brisbane, QLD, Australia) (37) and introduced in-frame intothe KpnI and PstI sites of pSecTag2(C)-mDectin-2-Fc to replace the Dectin-2coding sequence. The pSecTag2(C)-mDectin-2-Fc and pSecTag2(C)-mMincle-Fc constructs were separately transfected into HEK293T cells using Lip-ofectamine 2000 (Invitrogen). Soluble Dectin-2-Fc and Mincle-Fcwere expressed and purified as described previously (27). Soluble
Dectin-1-Fc was a gift from Dr. Gordon D. Brown (University of Aber-deen, Aberdeen, U.K.) (42). Human IgG1 Fc recombinant protein waspurchased from Invitrogen.
FACS assay for the binding of Fc fusion proteins to fungi
Fc fragment alone or Fc fusion proteins were added at 10–20 mg/ml to 23106 B. dermatitidis, 107 H. capsulatum, or 2 3 105 C. posadasii in 100 mlbinding buffer (20 mM Tris-HCl, 150 mM NaCl, 10 mM CaCl2, 0.05%Tween-20, pH 7.4) and incubated for 2 h at room temperature or overnightat 4˚C. The fungal cells were washed twice with binding buffer and in-cubated for an additional 30 min at room temperature with 1:100 dilutedPE-conjugated goat anti-human Fc Ab (Jackson ImmunoResearch Labo-ratories) in binding buffer. The cells were then washed three times withbinding buffer before fixed with 1% formaldehyde in binding buffer.Bound Fc proteins were quantified by flow cytometry.
CLR reporter cell construction, stimulation, and binding byfungi
B3Z cells carrying an NFAT-lacZ reporter construct have been describedpreciously (43) and were provided by Dr. Nilabh Shastri (University ofCalifornia, Berkeley, CA). B3Z cells expressing Dectin-2 or a wild type ora signaling-defective FcRg-chain that carries two tyrosine-to-phenylalaninemutations within the ITAM motif (31), as well as BWZ cells expressingDectin-1–CD3z (the extracellular domain of mouse Dectin-1 fused to CD3zintracellular tail) (44) and the parent BWZ cells (45), were a generous giftfrom Dr. Caetano Reis e Sousa (London Research Institute, London, U.K.).To construct Mincle-expressing reporter cells, the full-length cDNA of mouseMincle was subcloned from the pEF1HISB-mMincle plasmid into a pFB-IRES-EGFP retroviral vector. B3Z cells were retrovirally transduced withpFB-mMincle-IRES-EGFP, selected based on EGFP expression, and ver-ified by cell surface staining and FACS analysis using an anti-mouseMincle mAb (MBL). To generate B3Z cells coexpressing Mincle and theadaptor FcRg, Mincle-expressing reporter cells were retrovirally trans-duced with pMX-IP-FcRg-chain (wild type) or pMX-IP-FcRgm (mutant)(31). FcRg expression was verified with Western blotting using an anti-FcRg Ab (Upstate).
For B3Z/BWZ cell stimulation, 105 B3Z/BWZ cells/well in a 96-wellplate were incubated for 18 h with heat-killed fungal cells. lacZ activitywas measured in total cell lysates using CPRG (Roche) as a substrate. OD560 was measured using OD 620 as a reference. For the B3Z/BWZ cellbinding assay, 2 3 105 B3Z/BWZ cells were prestained with CFSE(Molecular Probes) and incubated for 6 h with 106 B. dermatitidis, 23 106
H. capsulatum, or 2 3 105 C. posadasii prestained with LIVE/DEAD Violetstaining (Invitrogen). Bound fungal cells were quantified with flowcytometry.
Statistical analysis
Differences in the number and percentage of activated, proliferating orcytokine-producing T cells, and in the number of CFUs were analyzed usingthe Wilcoxon rank test for nonparametric data or the t test when datawere normally distributed (46). Factorial ANOVA analysis using the SAFsoftware was used to compare the vaccine-induced reduction in lung CFUsbetween different strains of mice; p , 0.05 is considered statisticallysignificant.
ResultsVaccine-induced immunity against systemic dimorphic fungirequires the adaptor Card9
We previously reported that vaccine-induced Th17 cells werenecessary and sufficient to protect mice against the three majorsystemic mycoses in North America (14). Because Card9 has beenreported to be required for the development of Th17 cells inresponses to primary C. albicans infection (17), we investigatedwhether Card92/2 mice can acquire vaccine-induced immunityagainst infection with the systemic dimorphic fungi. Card92/2
mice were unable to control the live, virulence-attenuated #55vaccine strain of B. dermatitidis and succumbed to widespreaddissemination and infiltration of the lungs by the yeast (Fig. 1A).To circumvent susceptibility of Card92/2 mice to the vaccinestrain, we immunized them with heat-killed yeast and tested theirability to resist a lethal pulmonary infection with wild type yeast.Vaccinated Card92/2 mice had a burden of lung infection similar
The Journal of Immunology 1109
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
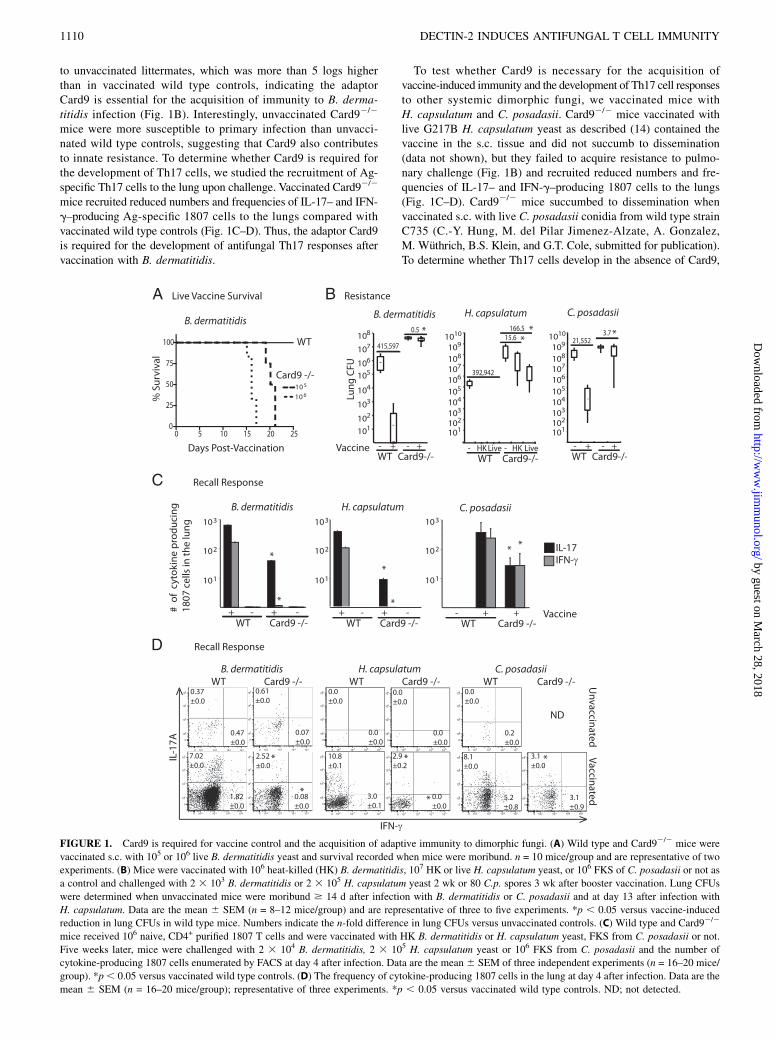
to unvaccinated littermates, which was more than 5 logs higherthan in vaccinated wild type controls, indicating the adaptorCard9 is essential for the acquisition of immunity to B. derma-titidis infection (Fig. 1B). Interestingly, unvaccinated Card92/2
mice were more susceptible to primary infection than unvacci-nated wild type controls, suggesting that Card9 also contributesto innate resistance. To determine whether Card9 is required forthe development of Th17 cells, we studied the recruitment of Ag-specific Th17 cells to the lung upon challenge. Vaccinated Card92/2
mice recruited reduced numbers and frequencies of IL-17– and IFN-g–producing Ag-specific 1807 cells to the lungs compared withvaccinated wild type controls (Fig. 1C–D). Thus, the adaptor Card9is required for the development of antifungal Th17 responses aftervaccination with B. dermatitidis.
To test whether Card9 is necessary for the acquisition ofvaccine-induced immunity and the development of Th17 cell responsesto other systemic dimorphic fungi, we vaccinated mice withH. capsulatum and C. posadasii. Card92/2 mice vaccinated withlive G217B H. capsulatum yeast as described (14) contained thevaccine in the s.c. tissue and did not succumb to dissemination(data not shown), but they failed to acquire resistance to pulmo-nary challenge (Fig. 1B) and recruited reduced numbers and fre-quencies of IL-17– and IFN-g–producing 1807 cells to the lungs(Fig. 1C–D). Card92/2 mice succumbed to dissemination whenvaccinated s.c. with live C. posadasii conidia from wild type strainC735 (C.-Y. Hung, M. del Pilar Jimenez-Alzate, A. Gonzalez,M. Wuthrich, B.S. Klein, and G.T. Cole, submitted for publication).To determine whether Th17 cells develop in the absence of Card9,
FIGURE 1. Card9 is required for vaccine control and the acquisition of adaptive immunity to dimorphic fungi. (A) Wild type and Card92/2 mice were
vaccinated s.c. with 105 or 106 live B. dermatitidis yeast and survival recorded when mice were moribund. n = 10 mice/group and are representative of two
experiments. (B) Mice were vaccinated with 106 heat-killed (HK) B. dermatitidis, 107 HK or live H. capsulatum yeast, or 106 FKS of C. posadasii or not as
a control and challenged with 2 3 103 B. dermatitidis or 2 3 105 H. capsulatum yeast 2 wk or 80 C.p. spores 3 wk after booster vaccination. Lung CFUs
were determined when unvaccinated mice were moribund $ 14 d after infection with B. dermatitidis or C. posadasii and at day 13 after infection with
H. capsulatum. Data are the mean 6 SEM (n = 8–12 mice/group) and are representative of three to five experiments. *p , 0.05 versus vaccine-induced
reduction in lung CFUs in wild type mice. Numbers indicate the n-fold difference in lung CFUs versus unvaccinated controls. (C) Wild type and Card92/2
mice received 106 naive, CD4+ purified 1807 T cells and were vaccinated with HK B. dermatitidis or H. capsulatum yeast, FKS from C. posadasii or not.
Five weeks later, mice were challenged with 2 3 104 B. dermatitidis, 2 3 105 H. capsulatum yeast or 106 FKS from C. posadasii and the number of
cytokine-producing 1807 cells enumerated by FACS at day 4 after infection. Data are the mean 6 SEM of three independent experiments (n = 16–20 mice/
group). *p, 0.05 versus vaccinated wild type controls. (D) The frequency of cytokine-producing 1807 cells in the lung at day 4 after infection. Data are the
mean 6 SEM (n = 16–20 mice/group); representative of three experiments. *p , 0.05 versus vaccinated wild type controls. ND; not detected.
1110 DECTIN-2 INDUCES ANTIFUNGAL T CELL IMMUNITY
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

knockout mice were instead vaccinated with either FKS or liveconidia of an attenuated DT vaccine strain. This strain lacks thechitinases 2, 3, and D-arabinotol-2-dehydrogenase genes (3) andfails to endosporulate and disseminate in s.c. vaccinated Card92/2
mice. Card92/2 mice vaccinated with FKS failed to acquire re-sistance to challenge (Fig. 1B) and recruited lower numbers andfrequencies of IL-17– and IFN-g–producing 1807 cells to thelungs upon challenge compared with wild type controls (Fig. 1C–D). Vaccination with the attenuated DT vaccine strain yieldedsimilar results (data not shown). In contrast to primary infectionwith B. dermatitidis and H. capsulatum, unvaccinated Card92/2
mice were not more susceptible than wild type controls to infectionwith C. posadasii. Thus, the Card9 signaling pathway is required forthe development of vaccine-induced resistance and antifungal Th17responses to the three major systemic dimorphic fungi.
Expansion and activation of Ag-specific cells does not requireCard9
Receptor signaling pathways can affect T cell development atmultiple stages. In the setting of LCMV infection, Tc1 cells requireintrinsic TLR/Myd88 signals not for activation, expansion, ordifferentiation, but to survive apoptosis after these events (47).Because Card9 is required for the development of vaccine-inducedanti-fungal Th17 cells, we investigated the stage of the immuneresponse at which Card9 governs T cell priming. To do so, we adop-tively transferred magnetic bead-purified CD4+, CFSE-labelednaive T cells from 1807 mice into Card92/2 and wild type recipi-ents prior to vaccination with B. dermatitidis and analyzed T celldevelopment at serial time points thereafter. At day 7 after vacci-nation, mice lacking Card9 showed no deficit in the proliferationand activation, but had a small reduction in expansion of Ag-specific1807 cells (Fig. 2A–C), indicating that Card9 minimally affects theearly stages of vaccine-induced T cell development. At subsequent
time points (days 14 and 35 after vaccination), activated 1807 cellsunderwent similar contraction. Upon recall at day 4 after infection,the frequency and number of activated CD44+ 1807 cells werecomparable in the skin-draining lymph node (sdLN) and in the lungof Card92/2 and wild type recipient mice (Fig. 2B–C). These dataindicate that the adaptor Card9 is not required for the activation,proliferation, and contraction of Ag-specific 1807 cells in the sdLN.Although the expansion of 1807 cells was minimally affected by theabsence of Card9, this did not affect the number of 1807 cells in thesdLN and the lung at day 4 after challenge.
Th17 cell differentiation requires Card9
Because Card92/2 mice failed to recruit IL-17– and IFN-g–producing 1807 cells to the lung (Fig. 1C–D), we investigatedwhether Card9 governs the differentiation of effector T cellsduring the vaccine-priming phase. We analyzed T cell differenti-ation by three independent approaches. First, we harvested acti-vated T cells at the peak of expansion from the sdLN andstimulated them ex vivo with cell wall membrane (CW/M) Ag.T cells from Card92/2 mice produced .8-fold less IL-17 proteinin response to fungal Ag than did wild type T cells (Fig. 3A).Similarly, IFN-g protein levels were reduced by at least 30-foldcompared with wild type controls. Although Card9 is primarilyresponsible for receptor signaling in myeloid but not lymphoidcells, we sought to exclude formally a T cell–intrinsic role ofCard9 for the differentiation of Th17 cells. Thus, in a secondapproach we cocultured BMDCs from Card92/2 and wild type micetogether with vaccine yeast and naive wild type 1807 cells in vitro.1807 cells cocultured with Card92/2 DCs produced ∼7-fold less IL-17 than did cocultures with wild type DCs (Fig. 3B). Third, toinvestigate Th17 cell differentiation in vivo, we determined thefrequency of cytokine-producing wild type 1807 cells at the siteof vaccination. The frequency of IL-17– and IFN-g–producing
FIGURE 2. Card9 is dispensable for T cell ex-
pansion and activation. (A) Wild type and Card92/2
mice received an adoptive transfer of 106 CD4+-purified,
CFSE-labeled, naive 1807 Tg cells and were vaccinated
with 106 HK B. darmatttidis yeast or not. Transferred
1807 cells were harvested from the sdLN and pro-
liferation was assessed by enumerating the frequency
of CFSElow cells (mean6 SEM; n = 4–6 mice/group;
representative of three experiments) at day 7 after
vaccination. (B) At serial times after vaccination, the
frequency of activated (CD44hi) 1807 Tg cells was
determined in the sdLN and in the lung at day four
after infection. Data are the mean 6 SEM (n = 4–6
mice/group); representative of five experiments. (C)
The number of activated (CD44hi) 1807 Tg cells for
(B). Data are the mean 6 SEM (n = 4–6 mice/group)
and are representative of five experiments. *p , 0.05
versus vaccinated wild type controls.
The Journal of Immunology 1111
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
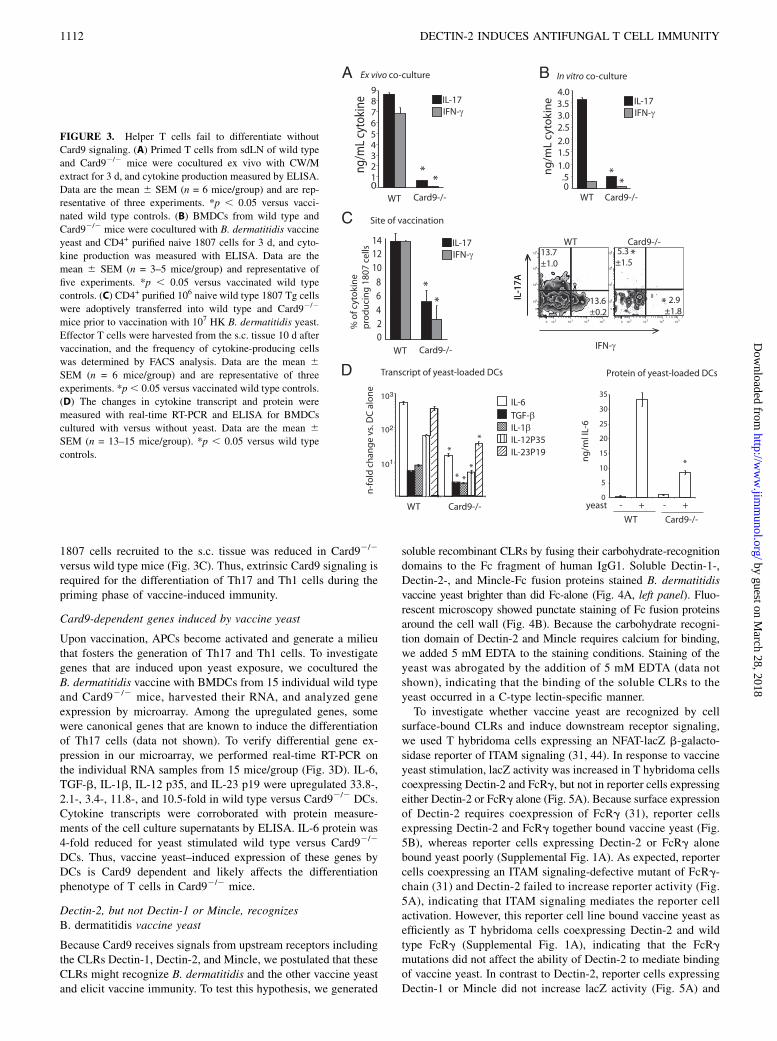
1807 cells recruited to the s.c. tissue was reduced in Card92/2
versus wild type mice (Fig. 3C). Thus, extrinsic Card9 signaling isrequired for the differentiation of Th17 and Th1 cells during thepriming phase of vaccine-induced immunity.
Card9-dependent genes induced by vaccine yeast
Upon vaccination, APCs become activated and generate a milieuthat fosters the generation of Th17 and Th1 cells. To investigategenes that are induced upon yeast exposure, we cocultured theB. dermatitidis vaccine with BMDCs from 15 individual wild typeand Card92/2 mice, harvested their RNA, and analyzed geneexpression by microarray. Among the upregulated genes, somewere canonical genes that are known to induce the differentiationof Th17 cells (data not shown). To verify differential gene ex-pression in our microarray, we performed real-time RT-PCR onthe individual RNA samples from 15 mice/group (Fig. 3D). IL-6,TGF-b, IL-1b, IL-12 p35, and IL-23 p19 were upregulated 33.8-,2.1-, 3.4-, 11.8-, and 10.5-fold in wild type versus Card92/2 DCs.Cytokine transcripts were corroborated with protein measure-ments of the cell culture supernatants by ELISA. IL-6 protein was4-fold reduced for yeast stimulated wild type versus Card92/2
DCs. Thus, vaccine yeast–induced expression of these genes byDCs is Card9 dependent and likely affects the differentiationphenotype of T cells in Card92/2 mice.
Dectin-2, but not Dectin-1 or Mincle, recognizesB. dermatitidis vaccine yeast
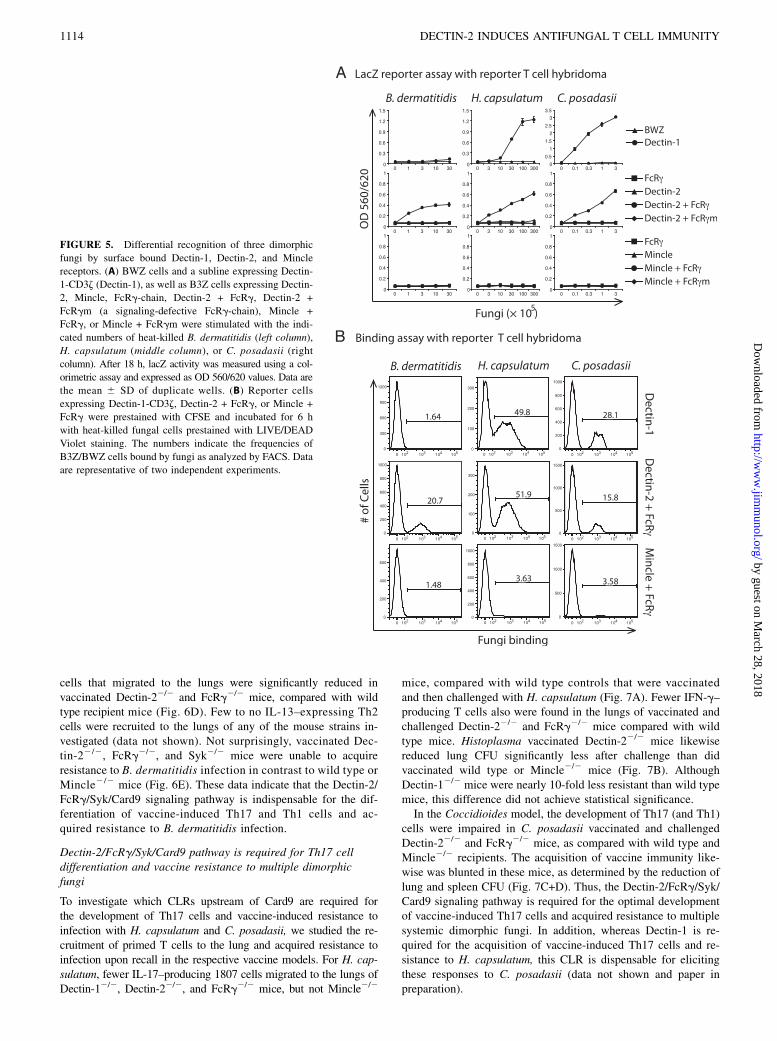
Because Card9 receives signals from upstream receptors includingthe CLRs Dectin-1, Dectin-2, and Mincle, we postulated that theseCLRs might recognize B. dermatitidis and the other vaccine yeastand elicit vaccine immunity. To test this hypothesis, we generated
soluble recombinant CLRs by fusing their carbohydrate-recognitiondomains to the Fc fragment of human IgG1. Soluble Dectin-1-,Dectin-2-, and Mincle-Fc fusion proteins stained B. dermatitidisvaccine yeast brighter than did Fc-alone (Fig. 4A, left panel). Fluo-rescent microscopy showed punctate staining of Fc fusion proteinsaround the cell wall (Fig. 4B). Because the carbohydrate recogni-tion domain of Dectin-2 and Mincle requires calcium for binding,we added 5 mM EDTA to the staining conditions. Staining of theyeast was abrogated by the addition of 5 mM EDTA (data notshown), indicating that the binding of the soluble CLRs to theyeast occurred in a C-type lectin-specific manner.To investigate whether vaccine yeast are recognized by cell
surface-bound CLRs and induce downstream receptor signaling,we used T hybridoma cells expressing an NFAT-lacZ b-galacto-sidase reporter of ITAM signaling (31, 44). In response to vaccineyeast stimulation, lacZ activity was increased in T hybridoma cellscoexpressing Dectin-2 and FcRg, but not in reporter cells expressingeither Dectin-2 or FcRg alone (Fig. 5A). Because surface expressionof Dectin-2 requires coexpression of FcRg (31), reporter cellsexpressing Dectin-2 and FcRg together bound vaccine yeast (Fig.5B), whereas reporter cells expressing Dectin-2 or FcRg alonebound yeast poorly (Supplemental Fig. 1A). As expected, reportercells coexpressing an ITAM signaling-defective mutant of FcRg-chain (31) and Dectin-2 failed to increase reporter activity (Fig.5A), indicating that ITAM signaling mediates the reporter cellactivation. However, this reporter cell line bound vaccine yeast asefficiently as T hybridoma cells coexpressing Dectin-2 and wildtype FcRg (Supplemental Fig. 1A), indicating that the FcRgmutations did not affect the ability of Dectin-2 to mediate bindingof vaccine yeast. In contrast to Dectin-2, reporter cells expressingDectin-1 or Mincle did not increase lacZ activity (Fig. 5A) and
FIGURE 3. Helper T cells fail to differentiate without
Card9 signaling. (A) Primed T cells from sdLN of wild type
and Card92/2 mice were cocultured ex vivo with CW/M
extract for 3 d, and cytokine production measured by ELISA.
Data are the mean 6 SEM (n = 6 mice/group) and are rep-
resentative of three experiments. *p , 0.05 versus vacci-
nated wild type controls. (B) BMDCs from wild type and
Card92/2 mice were cocultured with B. dermatitidis vaccine
yeast and CD4+ purified naive 1807 cells for 3 d, and cyto-
kine production was measured with ELISA. Data are the
mean 6 SEM (n = 3–5 mice/group) and representative of
five experiments. *p , 0.05 versus vaccinated wild type
controls. (C) CD4+ purified 106 naive wild type 1807 Tg cells
were adoptively transferred into wild type and Card92/2
mice prior to vaccination with 107 HK B. dermatitidis yeast.
Effector T cells were harvested from the s.c. tissue 10 d after
vaccination, and the frequency of cytokine-producing cells
was determined by FACS analysis. Data are the mean 6SEM (n = 6 mice/group) and are representative of three
experiments. *p, 0.05 versus vaccinated wild type controls.
(D) The changes in cytokine transcript and protein were
measured with real-time RT-PCR and ELISA for BMDCs
cultured with versus without yeast. Data are the mean 6SEM (n = 13–15 mice/group). *p , 0.05 versus wild type
controls.
1112 DECTIN-2 INDUCES ANTIFUNGAL T CELL IMMUNITY
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

bound vaccine yeast poorly (Fig. 5B; Supplemental Fig. 1B–C),although the respective soluble CLRs bound the yeast. Thus,Dectin-2, but not Dectin-1 or Mincle, recognized B. dermatitidisvaccine yeast and triggered downstream ITAM signaling throughFcRg.
Dectin-1 and Dectin-2 recognize multiple dimorphic fungi
The three CLRs differentially recognize B. dermatitidis; therefore,we investigated how they recognize H. capsulatum and C. pos-adasii. We incubated the latter two fungi with Fc fusion proteinsand analyzed them by FACS. All three soluble CLR fusion pro-teins stained H. capsulatum yeast to various degrees (Fig. 4A).Soluble Dectin-1–Fc stained the yeast strongly, Dectin-2–Fc stainedintermediately and Mincle-Fc stained the least. All three solubleCLR fusion proteins stained C. posadasii efficiently (Fig. 4A).Fluorescent microscopy showed punctate staining of Fc fusionproteins on the cell wall of H. capsulatum (Fig. 4B), whereasa more evenly distributed staining pattern was observed on thesurface of C. posadasii (Fig. 4B). Dectin-1– and Dectin-2–expressingT hybridoma cells showed increased lacZ reporter activity whenstimulated with H. capsulatum or C. posadasii (Fig. 5A), whereasno increase in reporter activity was detected with Mincle-expressingreporter cells. Dectin-1– and Dectin-2–expressing T hybridomacells consistently bound H. capsulatum and C. posadasii, whereasMincle-expressing and control reporter cells failed to do so (Fig.5B; Supplemental Fig. 1). Because all three dimorphic fungi failedto increase reporter activity of Mincle-expressing T hybridomacells, we used TDB, a known ligand for Mincle and syntheticanalog of the mycobacterial cord factor, as a positive controlto stimulate the lacZ reporter cells. In an additional approach,Dr. Sho Yamasaki’s laboratory tested the recognition of B. derma-titidis yeast by surface-bound Mincle using a NFAT-GFP reporterT cells as described (28). TDB increased the reporter activity ofthe Mincle expressing lacZ reporter cells (Supplemental Fig. 1D)
and the GFP reporter cells confirmed our results with the lacZreporting cells (data not shown). Thus, Dectin-1 and Dectin-2, butnot Mincle, recognized and responded to both H. capsulatum andC. posadasii.
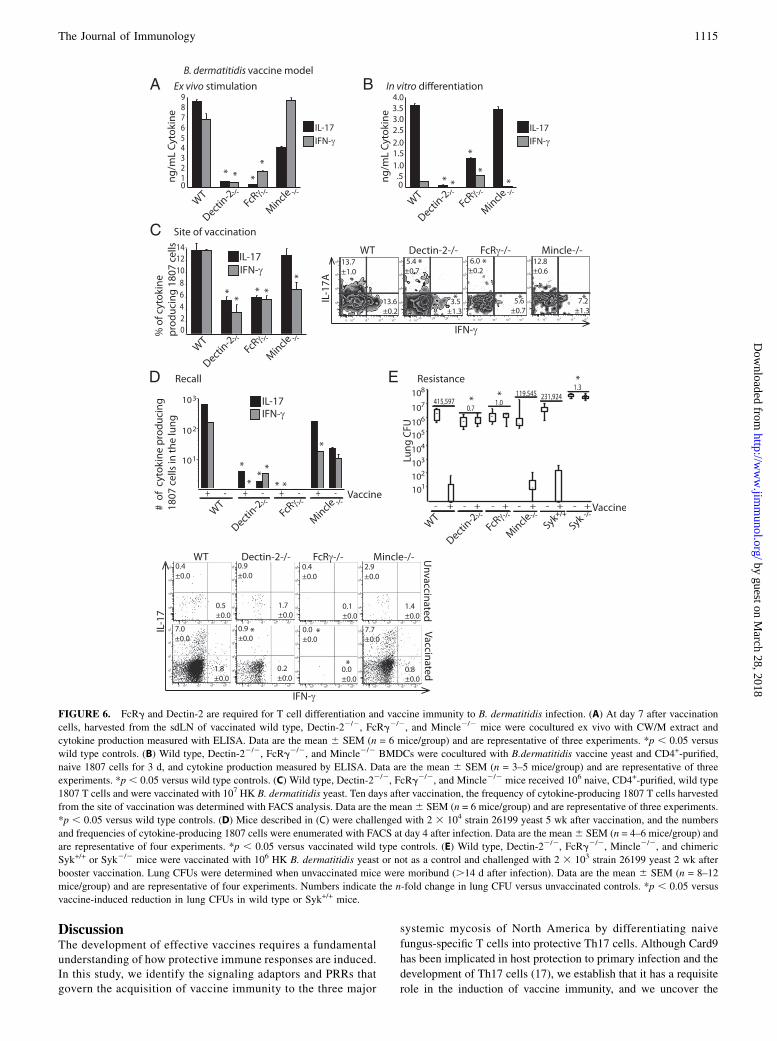
Th17 cell differentiation and vaccine immunity toB. dermatitidis requires Dectin2 and FcRg
Because Dectin-2 recognizes and binds B. dermatitidis vaccineyeast and triggers functional ITAM signaling, we tested to deter-mine whether Dectin-2 and its downstream signaling componentsare necessary to prime Th17 cells and acquire vaccine-inducedresistance. We studied Th17 cell differentiation using severalapproaches. First, we adoptively transferred naive magneticbead purified CD4+ 1807 cells into Dectin-22/2, FcR-g2/2, andMincle2/2 mice before vaccination, harvested the primed T cellsfrom the sdLN at the peak of expansion, and stimulated them withCW/M Ag in vitro as described above. T cells primed in Dectin-2and FcRg-deficient mice produced significantly less IL-17 andIFN-g than did T cells from wild type or Mincle2/2 mice (Fig.6A). In this assay, both endogenous and adoptively transferredCD4+ T cells contributed to cytokine production and a T cell in-trinsic contribution cannot be evaluated. Thus, second, to excludea T cell intrinsic role of either Dectin-2 or FcRg, we studied T celldifferentiation of naive wild type 1807 cells in vitro. When cocul-tured with BMDCs from Dectin-2 and FcRg deficient mice, 1807cells produced significantly less IL-17 compared with cocultureswith wild type or Mincle-deficient DCs (Fig. 6B). Third, westudied differentiation of wild type 1807 cells in vivo at the site ofvaccination. The frequency of 1807 cells producing IL-17 andIFN-g was significantly reduced in Dectin-22/2 and FcRg2/2
compared with wild type recipient mice (Fig. 6C). Fourth, wedetermined the number and frequencies of IL-17– and IFN-g–producing 1807 cells that migrated to the lung upon recall. Boththe numbers and frequencies of Ag-specific Th17 and Th1 1807
FIGURE 4. Soluble Dectin-1-Fc, Dectin-2-Fc, and Min-
cle-Fc proteins bind three dimorphic fungi. Fungal cells
were incubated with Fc fragment alone or Fc fusion pro-
teins as indicated by different colors, followed by staining
with PE-conjugated anti-human Fc Ab, and analyzed with
FACS (A) and microscopy (B). As a negative control, Ab-
stained fungal cells without any Fc proteins are indicated by
gray filled histograms in (A). Data are representative of two
independent experiments. Scale bar, 10 mm.
The Journal of Immunology 1113
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
cells that migrated to the lungs were significantly reduced invaccinated Dectin-22/2 and FcRg2/2 mice, compared with wildtype recipient mice (Fig. 6D). Few to no IL-13–expressing Th2cells were recruited to the lungs of any of the mouse strains in-vestigated (data not shown). Not surprisingly, vaccinated Dec-tin-22/2, FcRg2/2, and Syk2/2 mice were unable to acquireresistance to B. dermatitidis infection in contrast to wild type orMincle2/2 mice (Fig. 6E). These data indicate that the Dectin-2/FcRg/Syk/Card9 signaling pathway is indispensable for the dif-ferentiation of vaccine-induced Th17 and Th1 cells and ac-quired resistance to B. dermatitidis infection.
Dectin-2/FcRg/Syk/Card9 pathway is required for Th17 celldifferentiation and vaccine resistance to multiple dimorphicfungi
To investigate which CLRs upstream of Card9 are required forthe development of Th17 cells and vaccine-induced resistance toinfection with H. capsulatum and C. posadasii, we studied the re-cruitment of primed T cells to the lung and acquired resistance toinfection upon recall in the respective vaccine models. For H. cap-sulatum, fewer IL-17–producing 1807 cells migrated to the lungs ofDectin-12/2, Dectin-22/2, and FcRg2/2 mice, but not Mincle2/2
mice, compared with wild type controls that were vaccinatedand then challenged with H. capsulatum (Fig. 7A). Fewer IFN-g–producing T cells also were found in the lungs of vaccinated andchallenged Dectin-22/2 and FcRg2/2 mice compared with wildtype mice. Histoplasma vaccinated Dectin-22/2 mice likewisereduced lung CFU significantly less after challenge than didvaccinated wild type or Mincle2/2 mice (Fig. 7B). AlthoughDectin-12/2 mice were nearly 10-fold less resistant than wild typemice, this difference did not achieve statistical significance.In the Coccidioides model, the development of Th17 (and Th1)
cells were impaired in C. posadasii vaccinated and challengedDectin-22/2 and FcRg2/2 mice, as compared with wild type andMincle2/2 recipients. The acquisition of vaccine immunity like-wise was blunted in these mice, as determined by the reduction oflung and spleen CFU (Fig. 7C+D). Thus, the Dectin-2/FcRg/Syk/Card9 signaling pathway is required for the optimal developmentof vaccine-induced Th17 cells and acquired resistance to multiplesystemic dimorphic fungi. In addition, whereas Dectin-1 is re-quired for the acquisition of vaccine-induced Th17 cells and re-sistance to H. capsulatum, this CLR is dispensable for elicitingthese responses to C. posadasii (data not shown and paper inpreparation).
FIGURE 5. Differential recognition of three dimorphic
fungi by surface bound Dectin-1, Dectin-2, and Mincle
receptors. (A) BWZ cells and a subline expressing Dectin-
1-CD3z (Dectin-1), as well as B3Z cells expressing Dectin-
2, Mincle, FcRg-chain, Dectin-2 + FcRg, Dectin-2 +
FcRgm (a signaling-defective FcRg-chain), Mincle +
FcRg, or Mincle + FcRgm were stimulated with the indi-
cated numbers of heat-killed B. dermatitidis (left column),
H. capsulatum (middle column), or C. posadasii (right
column). After 18 h, lacZ activity was measured using a col-
orimetric assay and expressed as OD 560/620 values. Data are
the mean 6 SD of duplicate wells. (B) Reporter cells
expressing Dectin-1-CD3z, Dectin-2 + FcRg, or Mincle +
FcRg were prestained with CFSE and incubated for 6 h
with heat-killed fungal cells prestained with LIVE/DEAD
Violet staining. The numbers indicate the frequencies of
B3Z/BWZ cells bound by fungi as analyzed by FACS. Data
are representative of two independent experiments.
1114 DECTIN-2 INDUCES ANTIFUNGAL T CELL IMMUNITY
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
DiscussionThe development of effective vaccines requires a fundamentalunderstanding of how protective immune responses are induced.In this study, we identify the signaling adaptors and PRRs thatgovern the acquisition of vaccine immunity to the three major
systemic mycosis of North America by differentiating naive
fungus-specific T cells into protective Th17 cells. Although Card9
has been implicated in host protection to primary infection and the
development of Th17 cells (17), we establish that it has a requisite
role in the induction of vaccine immunity, and we uncover the
FIGURE 6. FcRg and Dectin-2 are required for T cell differentiation and vaccine immunity to B. dermatitidis infection. (A) At day 7 after vaccination
cells, harvested from the sdLN of vaccinated wild type, Dectin-22/2, FcRg2/2, and Mincle2/2 mice were cocultured ex vivo with CW/M extract and
cytokine production measured with ELISA. Data are the mean 6 SEM (n = 6 mice/group) and are representative of three experiments. *p , 0.05 versus
wild type controls. (B) Wild type, Dectin-22/2, FcRg2/2, and Mincle2/2 BMDCs were cocultured with B.dermatitidis vaccine yeast and CD4+-purified,
naive 1807 cells for 3 d, and cytokine production measured by ELISA. Data are the mean 6 SEM (n = 3–5 mice/group) and are representative of three
experiments. *p , 0.05 versus wild type controls. (C) Wild type, Dectin-22/2, FcRg2/2, and Mincle2/2 mice received 106 naive, CD4+-purified, wild type
1807 T cells and were vaccinated with 107 HK B. dermatitidis yeast. Ten days after vaccination, the frequency of cytokine-producing 1807 T cells harvested
from the site of vaccination was determined with FACS analysis. Data are the mean6 SEM (n = 6 mice/group) and are representative of three experiments.
*p , 0.05 versus wild type controls. (D) Mice described in (C) were challenged with 2 3 104 strain 26199 yeast 5 wk after vaccination, and the numbers
and frequencies of cytokine-producing 1807 cells were enumerated with FACS at day 4 after infection. Data are the mean6 SEM (n = 4–6 mice/group) and
are representative of four experiments. *p , 0.05 versus vaccinated wild type controls. (E) Wild type, Dectin-22/2, FcRg2/2, Mincle2/2, and chimeric
Syk+/+ or Syk2/2 mice were vaccinated with 106 HK B. dermatitidis yeast or not as a control and challenged with 2 3 103 strain 26199 yeast 2 wk after
booster vaccination. Lung CFUs were determined when unvaccinated mice were moribund (.14 d after infection). Data are the mean 6 SEM (n = 8–12
mice/group) and are representative of four experiments. Numbers indicate the n-fold change in lung CFU versus unvaccinated controls. *p , 0.05 versus
vaccine-induced reduction in lung CFUs in wild type or Syk+/+ mice.
The Journal of Immunology 1115
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
stage of the immune response at which it governs priming of Th17cells in vivo. Finally, we report distinctly different requirementsfor PRRs upstream of Card9 to induce the development of Th17cells and vaccine immunity to the three systemic dimorphic fungiof the United States.We found that Card9 is essential for the acquisition of vaccine-
induced resistance to infection with the dimorphic fungi B. derma-titidis, H. capsulatum, and C. posadasii. Card92/2 mice failed tocontrol s.c. delivery of live attenuated vaccine yeast of B. dermatitidisand wild type conidia of C. posadasii, but survived vaccination withlive H. capsulatum yeast. In response to vaccination with heat killedB. dermatitidis yeast, live H. capsulatum yeast or formalin-fixedspherules of C. posadasii, Card92/2 mice failed to acquire vaccineresistance. These results indicate that the Card9 signaling pathway isessential and can be harnessed for the induction of vaccine-inducedprotection against dimorphic fungal infection.Because the generation of Th17 cells directly correlates with
vaccine-induced resistance to the systemic dimorphic fungi (14),
we determined the number of IL-17–producing CD4+ T cells inthe lungs of vaccinated mice upon recall after challenge. We alsoenumerated the number of IFN-g–producing CD4+ T cells be-cause they can contribute to vaccine-induced resistance (4), al-though Th1 cells are dispensable, unlike Th17 cells (14). Weobserved that the numbers of Th17 and Th1 cells in all threevaccine models were significantly reduced in vaccinated Card92/2
versus wild type mice. Using our recently developed adoptivetransfer system with TCR transgenic 1807 cells, which recognizea shared and protective Ag among the systemic dimorphic fungi(34), we demonstrated that although Card9 is required for thedifferentiation of naive T cells into Th17, the adaptor is largelydispensable for the activation, expansion, proliferation, and sur-vival during the contraction phase, and for recruitment to the lungupon recall. Moreover, Card9 not only affected the differentiationof Th17 cells; it likewise affected Th1 cells. Although the role ofthe CLR-Card9 signaling pathway on the development of effectorTh17 and Th1 cells has not been studied in vivo, the TLR-Myd88
FIGURE 7. FcRg and Dectin-2 are required for
T cell differentiation and adaptive immunity to C.
posadasii and H. capsulatum. (A) 106 naıve CD41
purified, wild type 1807 Tg cells were transferred
into wild type, Dectin-22/2, FcRg2/2, and Mincle2/2
mice prior to vaccination with 107 HK H. capsulatum
G217B yeast and challenged with 105 H. capsulatum
yeast. At day four postinfection, the numbers and
frequencies of cytokine-producing 1807 cells in vac-
cinated mice were determined by FACS analysis.
Data are the mean 6 SEM (n 5 4–6 mice/group);
representative of two experiments. *p , 0.05 versus
vaccinated wild type controls. (B) Wild type,
Dectin-22/2, FcRg2/2, and Mincle2/2 mice were
vaccinated with 107 HK H. capsulatum G217B
yeast (or not as a control) and challenged with 105
H. capsulatum yeast 2 wk after booster vaccination.
Lung CFU were determined 13 d postinfection.
Data are the mean 6 SEM (n 5 8–12 mice/group);
representative of two experiments. Numbers indi-
cate the n-fold change in lung CFU versus unvac-
cinated controls. *p , 0.05 versus vaccine-induced
reduction in CFU in wild type mice. (C) Same as
(A), but mice were vaccinated with 106 FKS of
C. posadasii. Two wk after the boost, the mice were
challenged with 106 FKS. Data are the mean6 SEM
(n 5 4–6 mice/group); representative of two
experiments. *p , 0.05 versus vaccinated wild type
controls. (D) Wild type, Dectin-22/2, FcRg2/2, and
Mincle2/2 mice were vaccinated with 106 FKS or 5
3 104 viable spores of the ΔT strain (or not as
a control) and challenged with 50–80 C. posadasii
spores 4 wk later. Lung and spleen CFU were de-
termined 14 d postinfection. Data are the averages 6SEM of two independent experiments (n 5 8–12
mice/group/experiment); representative of two experi-
ments. Numbers indicate the n-fold change in lung
CFU versus unvaccinated controls. *p, 0.05 versus
vaccine-induced reduction in lung CFU in wild type
mice.
1116 DECTIN-2 INDUCES ANTIFUNGAL T CELL IMMUNITY
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

pathway has been shown to exert an incremental impact on thedifferentiation of A. fumigatus–specific CD4+ T cells during pul-monary infection (48). Myd88-mediated signals were dispensablefor the recruitment and expansion of Ag-specific T cells to themediastinal lymph nodes (MLN) and trafficking to the lung afterpriming. However, Myd88 signaling was required to enhance T-bet induction and differentiation of Th1 cells in the MLN. Furthermaturation of MLN-primed T cells into fully differentiated IFN-g–producing Th1 cells occurred upon arrival of the cells in theairways and was TLR and Myd88 independent.To discern how Card9 induces T helper differentiation, we per-
formed a microarray analysis and verified differential gene ex-pression by real-time RT-PCR. Among those differentially expressedwere genes known to induce Th17 and Th1 cells, such as IL-6, IL-23, TGF-b, IL-1b, and IL-12 p35. Thus, Card9 governs the ex-pression of cytokines that are instrumental in driving differentia-tion of Th17 and Th1 cells. Using the microarray data, we alsofound noncanonical genes that have not been linked previously toTh17 differentiation (V. LeBert, H. Wang and M. Wuthrich, un-published data). We intend to study the role of novel candidates inelucidating how Card9 governs the expression of the Th17 andTh1 priming cytokines.To investigate which PRRs upstream of Card9 are required to
induce the differentiation of Th17 cells and vaccine-induced im-munity, we first tested whether soluble fusion proteins consistingof the extracellular domain of Dectin-1, Dectin-2, and Mincle andhuman Fc can stain fungal yeast and spherules. Soluble Dectin-1and Dectin-2 have been used previously to stain A. fumigatus (42),Fonsecaea pedrosoi (49), C. albicans (27, 30), and C. immitis (18).Here, all three soluble Fc chimeras tested were able to bind thethree dimorphic fungi; however, the binding efficiency betweensoluble receptors and fungal species varied presumably based onrespective ligand expression and size of the organisms. Second, toexplore whether cell surface–bound CLRs can recognize and bindthe fungal vaccine strains we cocultured T cell hybridoma cellsthat were engineered to express a b-galactosidase NFAT-lacZ re-porter of ITAM signaling (43). By using this nonmyeloid reportersystem, we were able to distinguish between signaling eventstriggered by putative CLR ligands versus other ligands. To oursurprise, some of the hybridoma cell–expressed CLRs showed a rec-ognition pattern different from soluble CLR chimera. For example,soluble Dectin-1 stained B. dermatitidis vaccine yeast, but surfaceexpressed receptor did not recognize the vaccine yeast. Similarly,soluble Mincle stained all three dimorphic fungi, but Mincleexpressing reporter cells did not become activated and failed tobind the three fungi. On the other hand, soluble and surface-expressed Dectin-2 yielded consistent results for each of the fungitested. Although soluble CLR chimeras gave valid binding results,they were less reliable than surface expressed CLRs in forecastingthe in vivo role of CLRs in vaccine resistance.One potential explanation for the discrepant findings between
the two assays could be that only surface-exposed fungal ligandscan be recognized by cell surface–expressed receptors, but notligands that are embedded deeper in the cell wall; however, thelatter might still be accessible by soluble CLRs. Thus, cell sur-face–expressed receptors on the T hybridoma reporter cells helpedto predict the importance of the receptor for the priming of Thelper responses in vivo. Indeed, the failure of cell surface–expressed Dectin-1 to recognize B. dermatitidis is consistent withour previous data showing that Dectin-1 is dispensable for theinduction of Th17 and Th1 cells and vaccine immunity (14).Similarly, the phenotypes of Dectin-2 and Mincle in the vacci-nation model with B. dermatitidis showed good fidelity with theresults of the NFAT reporter cells. Dectin-22/2 mice showed
a severe defect in differentiation of fungus-specific Th17 cells andvaccine immunity, indicating that Dectin-2 is indispensable for theacquisition of adaptive immunity to B. dermatitidis infection. Theinability of T hybridoma cell–expressed Mincle to trigger NFAT-reporter activity or bind to the three dimorphic fungi correlatedwith the dispensability of this CLR in the vaccine models.Similar to Card9, the upstream receptor Dectin-2 affected the
development of both B. dermatitidis–specific Th17 and Th1 butnot Th2 cells during vaccination. Therefore, Dectin-2 and Card9deficiency did not shift the balance of Th cell differentiation as hasbeen reported for Th17 responses that depended on Dectin-1during a pulmonary infection of mice with A. fumigatus (50).Dectin-1 deficiency disproportionally increased Th1 responses anddecreased Th17 differentiation. The authors showed that Dectin-1mediated signals alter CD4 T cell responses to fungal infection byreducing the production of IFN-g and IL-12p40 in innate cells,thereby decreasing T-bet expression in responding CD4 T cellsand enhancing Th17 responses (50).Despite the fact that cell surface–expressed Dectin-1 and Dectin-
2 recognized and bound H. capsulatum yeast and C. posadasiispherules, the effect on Th17 cell priming and the acquisition ofvaccine immunity was modest, albeit statistically significant. Our dataare compatible with the interpretation that these two CLRs are re-dundant or cooperate in the innate recognition of these two dimorphicfungi. Receptor collaboration has been reported for Dectin-1 andmultiple TLRs (51), and for the recognition ofMalassezia by Dectin-2 and Mincle through distinct ligands (29). The fact, that H. capsu-latum and C. posadasii vaccinated FcRg2/2 mice showed strongerphenotypes in acquiring vaccine-induced immunity and Th17 cellscompared with Dectin-22/2 mice is compatible with the notion ofreceptor cooperation. Alternatively, other FcRg coupled CLR aloneor together with Dectin-2 could play a prominent role in innate fungalrecognition as well. A recently identified CLR, MCL, is a potentialcandidate, and mediates TDM-induced acquired immune responses(52); however, its role in vaccine immunity to fungi has not beenstudied.A prior study showed that b-glucan on H. capsulatum yeast was
shielded by a-glucan and not recognized by Dectin-1 (53), whereaswe observed that Dectin-1 expressing reporter cells bound to H.capsulatum, which triggered lacZ reporter activity in vitro. Conse-quently, in vivo, the induction of Th17 cells, the acquisition ofvaccine-induced resistance to H. capsulatum infection, and resis-tance to primary infection were all blunted in Dectin-12/2 versuswild type control mice. The differing results between the twostudies may be due to strain specific differences in the cell wallcomposition. We used the North American, RFLP class 2 andchemotype I strain G217B (ATCC 26032) that mostly lacks a-1,3-glucan on the yeast surface. In contrast, Rappleye et al. (53) usedthe Central and South American, RFLP class 3 and chemotype IIyeast strain G186AR (ATCC 26029), which expresses surfacea-1,3-glucan that shields b-glucan from detection by Dectin-1. Ina recent report (54), liquid grown G217B yeast bound to Dectin-1–expressing 3T3 fibroblasts when yeast were harvested duringthe exponential phase, but not during the stationary phase (54). Inour study, we grew G217B yeast for 3 to 7 d on slants and heat-killed them to perform overnight incubation in the lacZ reporterassay. Surface staining of yeast with Dectin-1 fusion protein wascomparable in live versus heat-killed cells, indicating that heat in-activation did not increase exposure of b-glucan. However, we haveseen large differences in the gross appearance of the yeast cell wallby electron microscopy (H. Wang and M.Wuthrich, unpublished data)when comparing liquid versus agar grown yeast, which couldaccount for the different Dectin-1 binding phenotypes. Nonethe-less, our data suggest that b-glucan is not shielded in H. capsu-
The Journal of Immunology 1117
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

latum strain G217B yeast in vivo, and it is recognized by Dectin-1during primary and adaptive immunity to recruit and prime Th17cells, respectively.Dectin-1 induces two independent signaling pathways—one
through Syk and one through Raf-1—to control adaptive immu-nity (55, 56), but we have not determined whether the FcRg/Card9and Dectin-1 phenotypes in the vaccination model with H. cap-sulatum are functionally linked or unrelated/Raf-1 dependent.The fungal ligands of most CLRs remain largely unclear. It is
likely that differences in the cell wall composition and structurebetween the endemic fungi account for the differential recognitionby Dectin-1, Dectin-2, andMincle. In addition, individual CLR canrecognize several distinct ligands within one fungal species andamong different fungi and microorganisms. For example, Minclerecognizes TDM frommycobacteria, corynebacteria, andNocardia(57), its synthetic analog TDB (25) and two structurally differentglycolipids 44-1 and 44-2 fromMalassezia (29). Similarly, Dectin-2recognizes the terminal mannose of N-linked glycan (27) and O-linked a-1,2 mannosyl residues from Malassezia (29). Thus, it isconceivable that Dectin-2 recognizes ligands in the systemic en-demic fungi that are structurally distinct from those that have beendescribed above.CLRs have been of burgeoning interest as targets to enhance
vaccine efficacy (56, 58). A promising strategy is to use carbo-hydrate ligands of CLRs to drive vaccine responses (59, 60). Forexample, the particulate b-glucan curdlan, a well-defined ligandfor Dectin-1, has been reported to be a potent adjuvant that primesanti-Candida Th17 responses (61). Moreover, porous b-glucanparticles that consist primarily of b-1,3-D-glucans and allow forhigh Ag loading have been exploited as a receptor-targeted vac-cine delivery system (62). Thus, the glucan particle-based vaccineplatform combines adjuvanticity and Ag delivery to induce strongTh17- and Th1-biased CD4+ T cell responses. Because mannansare recognized by Dectin-2 (23, 27, 29), we postulate that they canbe harnessed as adjuvants for vaccination to combat infection withthe systemic dimorphic fungi of North America that collectivelyaccount for nearly a million new infections annually.
AcknowledgmentsWe thank the laboratory of Dr. Sho Yamasaki for validating the recogni-
tion of B. dermatitidis by Mincle expressing NFAT-GFP reporter cells;
Dr. Victor L. J. Tybulewicz for Syk2/2 breeder mice; Dr. George S. Deepe
for H. capsulatum G217B yeast; Dr. Philip R. Taylor for the Dectin-2-Fc
expression vector pSecTag2(C)-mDectin-2-Fc; Dr. Christine A. Wells and
Robert B. Ashman for the pEF1HISB-mMincle plasmid; Dr. Gordon
D. Brown for soluble Dectin-1-Fc fusion protein; Dr. Nilabh Shastri for
B3Z parent cells; and Dr. Caetano Reis e Sousa for the pFB-IRES-EGFP ret-
roviral vector, the B3Z reporter cells expressing Dectin-2 and negative
control constructs, and BWZ cells expressing Dectin-1.
DisclosuresThe authors have no financial conflicts of interest.
References1. Wuthrich, M., H. I. Filutowicz, and B. S. Klein. 2000. Mutation of the WI-1 gene
yields an attenuated blastomyces dermatitidis strain that induces host resistance.J. Clin. Invest. 106: 1381–1389.
2. Wuthrich, M., H. I. Filutowicz, T. Warner, G. S. Deepe, Jr., and B. S. Klein.2003. Vaccine immunity to pathogenic fungi overcomes the requirement forCD4 help in exogenous antigen presentation to CD8+ T cells: implications forvaccine development in immune-deficient hosts. J. Exp. Med. 197: 1405–1416.
3. Xue, J., X. Chen, D. Selby, C. Y. Hung, J. J. Yu, and G. T. Cole. 2009. A ge-netically engineered live attenuated vaccine of Coccidioides posadasii protectsBALB/c mice against coccidioidomycosis. Infect. Immun. 77: 3196–3208.
4. Wuthrich, M., H. I. Filutowicz, T. Warner, and B. S. Klein. 2002. Requisiteelements in vaccine immunity to Blastomyces dermatitidis: plasticity uncoversvaccine potential in immune-deficient hosts. J. Immunol. 169: 6969–6976.
5. Chai, L. Y., F. van de Veerdonk, R. J. Marijnissen, S. C. Cheng, A. L. Khoo,M. Hectors, K. Lagrou, A. G. Vonk, J. Maertens, L. A. Joosten, et al. 2010. Anti-Aspergillus human host defence relies on type 1 T helper (Th1), rather than type17 T helper (Th17), cellular immunity. Immunology 130: 46–54.
6. Allendorfer, R., G. D. Brunner, and G. S. Deepe, Jr. 1999. Complex requirementsfor nascent and memory immunity in pulmonary histoplasmosis. J. Immunol.162: 7389–7396.
7. Lin, L., A. S. Ibrahim, X. Xu, J. M. Farber, V. Avanesian, B. Baquir, Y. Fu,S. W. French, J. E. Edwards, Jr., and B. Spellberg. 2009. Th1-Th17 cells mediateprotective adaptive immunity against Staphylococcus aureus and Candidaalbicans infection in mice. PLoS Pathog. 5: e1000703.
8. Zelante, T., A. De Luca, C. D’Angelo, S. Moretti, and L. Romani. 2009. IL-17/Th17 in anti-fungal immunity: what’s new? Eur. J. Immunol. 39: 645–648.
9. Zelante, T., A. De Luca, P. Bonifazi, C. Montagnoli, S. Bozza, S. Moretti,M. L. Belladonna, C. Vacca, C. Conte, P. Mosci, et al. 2007. IL-23 and the Th17pathway promote inflammation and impair antifungal immune resistance. Eur.J. Immunol. 37: 2695–2706.
10. Hernandez-Santos, N., A. R. Huppler, A. C. Peterson, S. A. Khader, K. C. McKenna,and S. L. Gaffen. 2013. Th17 cells confer long-term adaptive immunity to oralmucosal Candida albicans infections. Mucosal Immunol. 6: 900–910.
11. Pandiyan, P., H. R. Conti, L. Zheng, A. C. Peterson, D. R. Mathern,N. Hernandez-Santos, M. Edgerton, S. L. Gaffen, and M. J. Lenardo. 2011. CD4(+)CD25(+)Foxp3(+) regulatory T cells promote Th17 cells in vitro and enhancehost resistance in mouse Candida albicans Th17 cell infection model. Immunity34: 422–434.
12. Rudner, X. L., K. I. Happel, E. A. Young, and J. E. Shellito. 2007. Interleukin-23(IL-23)-IL-17 cytokine axis in murine Pneumocystis carinii infection. Infect.Immun. 75: 3055–3061.
13. Deepe, G. S., Jr., and R. S. Gibbons. 2009. Interleukins 17 and 23 influence thehost response to Histoplasma capsulatum. J. Infect. Dis. 200: 142–151.
14. Wuthrich, M., B. Gern, C. Y. Hung, K. Ersland, N. Rocco, J. Pick-Jacobs,K. Galles, H. Filutowicz, T. Warner, M. Evans, et al. 2011. Vaccine-inducedprotection against 3 systemic mycoses endemic to North America requiresTh17 cells in mice. J. Clin. Invest. 121: 554–568.
15. Vautier, S., D. M. MacCallum, and G. D. Brown. 2012. C-type lectin receptorsand cytokines in fungal immunity. Cytokine 58: 89–99.
16. Osorio, F., and C. Reis e Sousa. 2011. Myeloid C-type lectin receptors inpathogen recognition and host defense. Immunity 34: 651–664.
17. LeibundGut-Landmann, S., O. Gross, M. J. Robinson, F. Osorio, E. C. Slack,S. V. Tsoni, E. Schweighoffer, V. Tybulewicz, G. D. Brown, J. Ruland, andC. Reis e Sousa. 2007. Syk- and CARD9-dependent coupling of innate immunityto the induction of T helper cells that produce interleukin 17. Nat. Immunol. 8:630–638.
18. Viriyakosol, S., Mdel. P. Jimenez, M. A. Gurney, M. E. Ashbaugh, and J. Fierer.2013. Dectin-1 is required for resistance to coccidioidomycosis in mice. MBio.4: e00597–e12.
19. Ferwerda, B., G. Ferwerda, T. S. Plantinga, J. A. Willment, A. B. van Spriel,H. Venselaar, C. C. Elbers, M. D. Johnson, A. Cambi, C. Huysamen, et al. 2009.Human dectin-1 deficiency and mucocutaneous fungal infections. N. Engl.J. Med. 361: 1760–1767.
20. Plantinga, T. S., W. J. van der Velden, B. Ferwerda, A. B. van Spriel, G. Adema,T. Feuth, J. P. Donnelly, G. D. Brown, B. J. Kullberg, N. M. Blijlevens, andM. G. Netea. 2009. Early stop polymorphism in human DECTIN-1 is associatedwith increased candida colonization in hematopoietic stem cell transplantrecipients. Clin. Infect. Dis. 49: 724–732.
21. Chai, L. Y., M. G. de Boer, W. J. van der Velden, T. S. Plantinga, A. B. vanSpriel, C. Jacobs, C. J. Halkes, A. G. Vonk, N. M. Blijlevens, J. T. van Dissel,et al. 2011. The Y238X stop codon polymorphism in the human b-glucan re-ceptor dectin-1 and susceptibility to invasive aspergillosis. J. Infect. Dis. 203:736–743.
22. Cunha, C., M. Di Ianni, S. Bozza, G. Giovannini, S. Zagarella, T. Zelante,C. D’Angelo, A. Pierini, L. Pitzurra, F. Falzetti, et al. 2010. Dectin-1 Y238Xpolymorphism associates with susceptibility to invasive aspergillosis in hema-topoietic transplantation through impairment of both recipient- and donor-dependent mechanisms of antifungal immunity. Blood 116: 5394–5402.
23. Saijo, S., S. Ikeda, K. Yamabe, S. Kakuta, H. Ishigame, A. Akitsu, N. Fujikado,T. Kusaka, S. Kubo, S. H. Chung, et al. 2010. Dectin-2 recognition of alpha-mannans and induction of Th17 cell differentiation is essential for host defenseagainst Candida albicans. Immunity 32: 681–691.
24. Gringhuis, S. I., B. A. Wevers, T. M. Kaptein, T. M. van Capel, B. Theelen,T. Boekhout, E. C. de Jong, and T. B. Geijtenbeek. 2011. Selective C-Rel ac-tivation via Malt1 controls anti-fungal T(H)-17 immunity by dectin-1 and dectin-2. PLoS Pathog. 7: e1001259.
25. Schoenen, H., B. Bodendorfer, K. Hitchens, S. Manzanero, K. Werninghaus,F. Nimmerjahn, E. M. Agger, S. Stenger, P. Andersen, J. Ruland, et al. 2010.Cutting edge: Mincle is essential for recognition and adjuvanticity of the my-cobacterial cord factor and its synthetic analog trehalose-dibehenate. J. Immunol.184: 2756–2760.
26. Reid, D. M., N. A. Gow, and G. D. Brown. 2009. Pattern recognition: recentinsights from Dectin-1. Curr. Opin. Immunol. 21: 30–37.
27. McGreal, E. P., M. Rosas, G. D. Brown, S. Zamze, S. Y. Wong, S. Gordon,L. Martinez-Pomares, and P. R. Taylor. 2006. The carbohydrate-recognitiondomain of Dectin-2 is a C-type lectin with specificity for high mannose.Glycobiology 16: 422–430.
28. Yamasaki, S., M. Matsumoto, O. Takeuchi, T. Matsuzawa, E. Ishikawa,M. Sakuma, H. Tateno, J. Uno, J. Hirabayashi, Y. Mikami, et al. 2009. C-type
1118 DECTIN-2 INDUCES ANTIFUNGAL T CELL IMMUNITY
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

lectin Mincle is an activating receptor for pathogenic fungus, Malassezia. Proc.Natl. Acad. Sci. USA 106: 1897–1902.
29. Ishikawa, T., F. Itoh, S. Yoshida, S. Saijo, T. Matsuzawa, T. Gonoi, T. Saito,Y. Okawa, N. Shibata, T. Miyamoto, and S. Yamasaki. 2013. Identification ofdistinct ligands for the C-type lectin receptors Mincle and Dectin-2 in thepathogenic fungus Malassezia. Cell Host Microbe 13: 477–488.
30. Sato, K., X. L. Yang, T. Yudate, J. S. Chung, J. Wu, K. Luby-Phelps,R. P. Kimberly, D. Underhill, P. D. Cruz, Jr., and K. Ariizumi. 2006. Dectin-2 isa pattern recognition receptor for fungi that couples with the Fc receptor gammachain to induce innate immune responses. J. Biol. Chem. 281: 38854–38866.
31. Robinson, M. J., F. Osorio, M. Rosas, R. P. Freitas, E. Schweighoffer, O. Gross,J. S. Verbeek, J. Ruland, V. Tybulewicz, G. D. Brown, et al. 2009. Dectin-2 isa Syk-coupled pattern recognition receptor crucial for Th17 responses to fungalinfection. J. Exp. Med. 206: 2037–2051.
32. Yamasaki, S., E. Ishikawa, M. Sakuma, H. Hara, K. Ogata, and T. Saito. 2008.Mincle is an ITAM-coupled activating receptor that senses damaged cells. Nat.Immunol. 9: 1179–1188.
33. Glocker, E. O., A. Hennigs, M. Nabavi, A. A. Schaffer, C. Woellner, U. Salzer,D. Pfeifer, H. Veelken, K. Warnatz, F. Tahami, et al. 2009. A homozygousCARD9 mutation in a family with susceptibility to fungal infections. N. Engl.J. Med. 361: 1727–1735.
34. Wuthrich, M., C. Y. Hung, B. H. Gern, J. C. Pick-Jacobs, K. J. Galles,H. I. Filutowicz, G. T. Cole, and B. S. Klein. 2011. A TCR transgenic mousereactive with multiple systemic dimorphic fungi. J. Immunol. 187: 1421–1431.
35. Wuthrich, M., K. Ersland, T. Sullivan, K. Galles, and B. S. Klein. 2012. Fungisubvert vaccine T cell priming at the respiratory mucosa by preventingchemokine-induced influx of inflammatory monocytes. Immunity 36: 680–692.
36. Hsu, Y. M., Y. Zhang, Y. You, D. Wang, H. Li, O. Duramad, X. F. Qin, C. Dong,and X. Lin. 2007. The adaptor protein CARD9 is required for innate immuneresponses to intracellular pathogens. Nat. Immunol. 8: 198–205.
37. Wells, C. A., J. A. Salvage-Jones, X. Li, K. Hitchens, S. Butcher, R. Z. Murray,A. G. Beckhouse, Y. L. Lo, S. Manzanero, C. Cobbold, et al. 2008. Themacrophage-inducible C-type lectin, mincle, is an essential component of theinnate immune response to Candida albicans. J. Immunol. 180: 7404–7413.
38. Turner, M., P. J. Mee, P. S. Costello, O. Williams, A. A. Price, L. P. Duddy,M. T. Furlong, R. L. Geahlen, and V. L. Tybulewicz. 1995. Perinatal lethality andblocked B-cell development in mice lacking the tyrosine kinase Syk. Nature 378:298–302.
39. Brandhorst, T. T., M. Wuthrich, T. Warner, and B. Klein. 1999. Targeted genedisruption reveals an adhesin indispensable for pathogenicity of Blastomycesdermatitidis. J. Exp. Med. 189: 1207–1216.
40. Levine, H. B., Y. C. Kong, and C. Smith. 1965. Immunization of Mice toCoccidioides Immitis: Dose, Regimen and Spherulation Stage of Killed SpheruleVaccines. J. Immunol. 94: 132–142.
41. Johnson, M. R., K. Wang, J. B. Smith, M. J. Heslin, and R. B. Diasio. 2000.Quantitation of dihydropyrimidine dehydrogenase expression by real-time re-verse transcription polymerase chain reaction. Anal. Biochem. 278: 175–184.
42. Steele, C., R. R. Rapaka, A. Metz, S. M. Pop, D. L. Williams, S. Gordon,J. K. Kolls, and G. D. Brown. 2005. The beta-glucan receptor dectin-1 recog-nizes specific morphologies of Aspergillus fumigatus. PLoS Pathog. 1: e42.
43. Karttunen, J., S. Sanderson, and N. Shastri. 1992. Detection of rare antigen-presenting cells by the lacZ T-cell activation assay suggests an expressioncloning strategy for T-cell antigens. Proc. Natl. Acad. Sci. USA 89: 6020–6024.
44. Sancho, D., O. P. Joffre, A. M. Keller, N. C. Rogers, D. Martınez, P. Hernanz-Falcon, I. Rosewell, and C. Reis e Sousa. 2009. Identification of a dendritic cellreceptor that couples sensing of necrosis to immunity. Nature 458: 899–903.
45. Sanderson, S., and N. Shastri. 1994. LacZ inducible, antigen/MHC-specificT cell hybrids. Int. Immunol. 6: 369–376.
46. Fisher, L. D., and G. van Belle. 1993. Biostatistics: A Methodology for theHealth Sciences. John Wiley & Sons, New York, p. 611–613.
47. Quigley, M., J. Martinez, X. Huang, and Y. Yang. 2009. A critical role for directTLR2-MyD88 signaling in CD8 T-cell clonal expansion and memory formationfollowing vaccinia viral infection. Blood 113: 2256–2264.
48. Rivera, A., G. Ro, H. L. Van Epps, T. Simpson, I. Leiner, D. B. Sant’Angelo, andE. G. Pamer. 2006. Innate immune activation and CD4+ T cell priming duringrespiratory fungal infection. Immunity 25: 665–675.
49. Sousa, Mda. G., D. M. Reid, E. Schweighoffer, V. Tybulewicz, J. Ruland,J. Langhorne, S. Yamasaki, P. R. Taylor, S. R. Almeida, and G. D. Brown. 2011.Restoration of pattern recognition receptor costimulation to treat chromo-blastomycosis, a chronic fungal infection of the skin. Cell Host Microbe 9: 436–443.
50. Rivera, A., T. M. Hohl, N. Collins, I. Leiner, A. Gallegos, S. Saijo, J. W. Coward,Y. Iwakura, and E. G. Pamer. 2011. Dectin-1 diversifies Aspergillus fumigatus-specific T cell responses by inhibiting T helper type 1 CD4 T cell differentiation.J. Exp. Med. 208: 369–381.
51. Gantner, B. N., R. M. Simmons, S. J. Canavera, S. Akira, and D. M. Underhill.2003. Collaborative induction of inflammatory responses by dectin-1 and Toll-like receptor 2. J. Exp. Med. 197: 1107–1117.
52. Miyake, Y., K. Toyonaga, D. Mori, S. Kakuta, Y. Hoshino, A. Oyamada,H. Yamada, K. I. Ono, M. Suyama, Y. Iwakura, Y. Yoshikai, and S. Yamasaki.2013. C-type lectin MCL is an FcRgamma-coupled receptor that mediates theadjuvanticity of mycobacterial cord factor. Immunity. 38: 1050–1062.
53. Rappleye, C. A., L. G. Eissenberg, and W. E. Goldman. 2007. Histoplasmacapsulatum alpha-(1,3)-glucan blocks innate immune recognition by the beta-glucan receptor. Proc. Natl. Acad. Sci. USA 104: 1366–1370.
54. Edwards, J. A., E. A. Alore, and C. A. Rappleye. 2011. The yeast-phase viru-lence requirement for a-glucan synthase differs among Histoplasma capsulatumchemotypes. Eukaryot. Cell 10: 87–97.
55. Geijtenbeek, T. B., and S. I. Gringhuis. 2009. Signalling through C-type lectinreceptors: shaping immune responses. Nat. Rev. Immunol. 9: 465–479.
56. Wuthrich, M., G. S. Deepe, Jr., and B. Klein. 2012. Adaptive immunity to fungi.Annu. Rev. Immunol. 30: 115–148.
57. Ishikawa, E., T. Ishikawa, Y. S. Morita, K. Toyonaga, H. Yamada, O. Takeuchi,T. Kinoshita, S. Akira, Y. Yoshikai, and S. Yamasaki. 2009. Direct recognition ofthe mycobacterial glycolipid, trehalose dimycolate, by C-type lectin Mincle.J. Exp. Med. 206: 2879–2888.
58. Hardison, S. E., and G. D. Brown. 2012. C-type lectin receptors orchestrateantifungal immunity. Nat. Immunol. 13: 817–822.
59. Cutler, J. E., G. S. Deepe, Jr., and B. S. Klein. 2007. Advances in combatingfungal diseases: vaccines on the threshold. Nat. Rev. Microbiol. 5: 13–28.
60. Lang, R., H. Schoenen, and C. Desel. 2011. Targeting Syk-Card9-activatingC-type lectin receptors by vaccine adjuvants: findings, implications and openquestions. Immunobiology 216: 1184–1191.
61. Bar, E., A. Gladiator, S. Bastidas, B. Roschitzki, H. Acha-Orbea, A. Oxenius, andS. LeibundGut-Landmann. 2012. A novel Th cell epitope of Candida albicansmediates protection from fungal infection. J. Immunol. 188: 5636–5643.
62. Huang, H., G. R. Ostroff, C. K. Lee, C. A. Specht, and S. M. Levitz. 2010.Robust stimulation of humoral and cellular immune responses following vac-cination with antigen-loaded beta-glucan particles. MBio. 1: e00164–e10.
The Journal of Immunology 1119
by guest on March 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from