c 2012, julie bumbaugh - tdl
TRANSCRIPT

A Dynamical System Study of the Hodgkin-Huxley Model of Nerve Membranes
by
Julie Bumbaugh, B.S.
A Thesis
In
Mathematics and Statistics
Submitted to the Graduate Facultyof Texas Tech University in
Partial Fulfillment ofthe Requirements for the Degree of
Master of Science
Approved
Lih-Ing Roegerco-Chair
Sophia Jangco-Chair
Peggy Gordon MillerDean of the Graduate School
August, 2012

c©2012, Julie Bumbaugh

Texas Tech University, Julie Bumbaugh, August, 2012
ACKNOWLEDGEMENTS
I wish to acknowledge the help and support of Dr. Lih-Ing Roeger and Dr.
Sophia Jang. Their encouragement and advice since August of 2011 has made all of
this work possible for me. I also wish to thank my principal, Angie Inklebarger, for
being kind enough to allow me extra time when I needed it to work on this thesis.
She also allowed me to be away from the high school campus every Thursday during
my conference in order to meet some of the research requirements. I owe Dr. Jerry
Dwyer many thanks for being kind enough several years ago to be my mentor when
I first began to study the work of Hodgkin and Huxley. Finally, I could not have
accomplished what I have without the love and support of my husband, Alan
Bumbaugh.
ii

Texas Tech University, Julie Bumbaugh, August, 2012
TABLE OF CONTENTS
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ii
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . v
List of Tables . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vi
List of Figures . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vii
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
2. Membrane current for Hodgkin-Huxley model . . . . . . . . . . . . . . . 3
2.1 The membrane current equation . . . . . . . . . . . . . . . . . 3
2.2 Modeling potassium conductance . . . . . . . . . . . . . . . . 4
2.3 Modeling sodium conductance . . . . . . . . . . . . . . . . . . 9
3. Solving the Hodgkin-Huxley space clamp model . . . . . . . . . . . . . 14
4. Modern techniques to analyzing dynamical systems . . . . . . . . . . . . 17
4.1 One-dimensional models . . . . . . . . . . . . . . . . . . . . . 17
4.1.1 Stability of equilibria . . . . . . . . . . . . . . . . . . . . . 18
4.1.2 Bifurcations . . . . . . . . . . . . . . . . . . . . . . . . . . 20
4.2 Two-dimensional models . . . . . . . . . . . . . . . . . . . . . 21
4.2.1 Nullclines . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
4.2.2 Stability in 2-dimensions . . . . . . . . . . . . . . . . . . . 23
4.2.3 Limit cycles . . . . . . . . . . . . . . . . . . . . . . . . . . 27
4.2.4 Bifurcations in 2-dimensions . . . . . . . . . . . . . . . . . 27
4.2.5 Stable and unstable manifolds . . . . . . . . . . . . . . . . 29
4.2.6 Hodgkin-Huxley reduced model . . . . . . . . . . . . . . . 29
5. Problems for study and analysis . . . . . . . . . . . . . . . . . . . . . . 34
5.1 Proposed problems for study . . . . . . . . . . . . . . . . . . . 34
5.1.1 Problem 1- Derive Boltzmann’s function . . . . . . . . . . 34
5.1.2 Problem 2- Prove instability and stability of equilibria . . . 35
5.1.3 Problem 3- Find equilibrium and determine its stability . . 36
5.1.4 Problem 4- Demonstrate excitation block . . . . . . . . . . 36
5.1.5 Problem 5- Choose appropriate power for an activation vari-
able . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
iii

Texas Tech University, Julie Bumbaugh, August, 2012
5.1.6 Problem 6- Derive the formula Hodgkin and Huxley used
for n . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
5.2 Solutions to the problems for study and analysis . . . . . . . . 37
5.2.1 Solution to problem 1 . . . . . . . . . . . . . . . . . . . . . 37
5.2.2 Solution to problem 2 . . . . . . . . . . . . . . . . . . . . . 38
5.2.3 Solution to problem 3 . . . . . . . . . . . . . . . . . . . . . 38
5.2.4 Solution to problem 4 . . . . . . . . . . . . . . . . . . . . . 39
5.2.5 Solution to problem 5 . . . . . . . . . . . . . . . . . . . . . 40
5.2.6 Solution to problem 6 . . . . . . . . . . . . . . . . . . . . . 41
Bibliography . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
Appendix A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
Appendix B . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
Appendix C . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
Appendix D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
iv

Texas Tech University, Julie Bumbaugh, August, 2012
ABSTRACT
Equations of the Hodgkin and Huxley model of action potentials in squid giant
axons have been studied and described in this thesis. Explanation of the
development of the space clamp model that won its authors the Nobel Prize for
Physiology and Medicine in 1963 is provided since it is still one of the key
mathematical models in the study of neural communications today. A numerical
approach was used to obtain solutions to their model. The computer code used will
be provided. More modern approaches to analyzing these models will be explored
and discussed as a study of dynamical systems. A reduced Hodgkin-Huxley model
will be explained and analyzed using modern methods. A set of problems will be
included for those who wish to be guided through parts of the development of the
models.
v

Texas Tech University, Julie Bumbaugh, August, 2012
LIST OF TABLES
2.1 Computations for αn’s and βn’s . . . . . . . . . . . . . . . . . . . . . 8
3.1 List of variables and parameters for the Hodgkin-Huxley model . . . 15
4.1 Parameters for persistent sodium model . . . . . . . . . . . . . . . . 18
4.2 Parameters for 1-dimensional potassium model . . . . . . . . . . . . 21
4.3 Parameters for low-threshold INa,p + IK model . . . . . . . . . . . . 22
5.1 Parameters for high-threshold INa,p + IK model . . . . . . . . . . . . 35
vi

Texas Tech University, Julie Bumbaugh, August, 2012
LIST OF FIGURES
2.1 Hodgkin-Huxley potassium vs. time for fixed membrane potential . . 7
3.1 Action potentials in squid giant axons . . . . . . . . . . . . . . . . . . 16
4.1 Persistent sodium model graphs . . . . . . . . . . . . . . . . . . . . . 19
4.2 Failure to generate “all or nothing” spikes . . . . . . . . . . . . . . . 23
4.3 Failure to have fixed threshold potential . . . . . . . . . . . . . . . . 24
4.4 Bifurcations in high threshold INa,p + IK model . . . . . . . . . . . . 25
4.5 Nullclines for high threshold INa,p + IK model . . . . . . . . . . . . . 25
4.6 A stable limit cycle . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
4.7 Low threshold INa,p + IK traces leading to excitation block . . . . . . 28
4.8 Bistability in high threshold INa,p + IK model. . . . . . . . . . . . . . 30
4.9 Hodgkin-Huxley model reduced to 2 variables with I = 0. . . . . . . . 32
4.10 Hodgkin-Huxley model reduced to 2 variables with I = 12. . . . . . . 32
5.1 Boltzmann’s function . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
5.2 The intersection of nullclines . . . . . . . . . . . . . . . . . . . . . . . 39
5.3 Excitation block in the low-threshold INa,p + IK model. . . . . . . . 40
vii

Texas Tech University, Julie Bumbaugh, August, 2012
CHAPTER 1
INTRODUCTION
The seminal and classical work of Alan Lloyd Hodgkin and Andrew Fielding
Huxley, from 1942 to 1952, of developing a mathematical model that would yield
membrane potential vs. time following a perturbation from the resting state of
membrane potential in squid giant axons continues to be the biophysically accurate
model that influences the development of such models today:
−dVdt
= gkn4(V − Vk) + gNam
3h(vV − VNa) + gl(V − Vl), (1.1)
dn
dt= αn(1− n)− βn · n, (1.2)
dm
dt= αm(1−m)− βm ·m, (1.3)
dh
dt= αh(1− h)− βh · h, (1.4)
αn =1
10
[110
(V + 10)
e110
(V+10) − 1
], (1.5)
βn =1
8eV80 , (1.6)
αm =0.1(V + 25)
e0.1(V+25) − 1, (1.7)
βm = 4eV18 , (1.8)
αh = 0.07eV20 , (1.9)
βh =1
eV+30
10 + 1. (1.10)
The first goal of this thesis is to fully understand and explain the work done by
Hodgkin and Huxley in connection with their “voltage clamp” model in order to
more fully understand and appreciate the modern techniques used today. The
second goal in this thesis is to demonstrate some of the modern forms of analysis
used in the study of dynamical systems and to apply some of them to the
Hodgkin-Huxley model. Hodgkin and Huxley published a series of 5 papers
describing their work. The first paper dealt with their experimental method and the
1

Texas Tech University, Julie Bumbaugh, August, 2012
membrane in a normal ionic environment. The second paper describes their
technique of changing the sodium concentrations in the medium surrounding the
membrane which allowed them to resolve the ionic current into sodium and
potassium currents. Their third paper dealt with the effect that sudden changes in
membrane potential had on the ionic conductances. The fourth paper explained the
inactivation process which reduces sodium permeability during the falling phase of
the action potential. Their final paper concluded the series and revealed their
mathematical model of membrane current and specifically of the membrane’s ability
to generate action potentials. The development of the model will be explained in
this thesis. Curves from various particular solutions to the model along with the
Matlab program used are provided. Some modern methods used in the study of
dynamical systems in neuroscience today are also described in this thesis. The
Hodgkin-Huxley model was an important breakthrough in the study of neural
communication. Its historical significance aside; the review of just how this model
was arrived at would be valuable to students wanting to experience how
mathematical models are developed; therefore, a set of problems including the
derivation of one part of the models used today and some analysis of models using
more modern techniques will be included. The solutions to the set of problems will
be included.
2

Texas Tech University, Julie Bumbaugh, August, 2012
CHAPTER 2
MEMBRANE CURRENT FOR HODGKIN-HUXLEY MODEL
2.1 The membrane current equation
The measure of the change in electric charge across the membrane of a neuron,
nerve cell, is called the membrane potential or potential difference, and the unit of
measure is called voltage, V . Hodgkin and Huxley used “space-clamp” or
“voltage-clamp” experiments holding the voltage across the membrane fixed at a
constant value, Vc. These experiments revealed that ionic movements through
excitable tissues were associated with perturbations of membrane potential [2].
Their work involved experiments with squid giant axons threaded with two fine
silver electrodes down the axis of the axons to both short circuit currents that arise
within the membrane and to inject a current proportional to the difference, Vc − V ,
allowing dVdt
= V = 0 [7]. One electrode measured the membrane’s potential and the
ionic current during a space-clamp experiment. The second electrode was used to
inject current to maintain a constant membrane potential equal to a stepped
potential. The required current injected to maintain this potential provided a way
of measuring the ionic currents which were initiated by the perturbation from the
resting potential.
They found that the ionic current could be broken down primarily into a
movement of sodium ions and a potassium current each of which varied with
permeability of the membrane to each ion and with the membrane potential. To
determine which portion of the currents were due to sodium and which were due to
potassium, Hodgkin and Huxley pioneered some techniques involving controlling the
medium surrounding the tissue. If 90% of the sodium in the extracellular seawater
were replaced with choline (enough to significantly reduce the amount of sodium
but not enough to render the squid axon inexcitable), there could be little inflow of
sodium and they would know the currents they were measuring were mostly due to
potassium. Then from their current measurements using normal seawater in the
surrounding medium, they could separate the potassium from the sodium currents.
3

Texas Tech University, Julie Bumbaugh, August, 2012
For specific fixed values of membrane potential, they recorded ionic currents and
membrane potential measured against time. Their results showed that when they
depolarized the membrane and kept its potential constant in normal sea water,
sodium ions would flood into the cell first and then later would flow out. Potassium
was shown to flow outward during the later phase as well. Hodgkin and Huxley were
the first to assume that the ionic conductances might not all behave the same way
and developed a model that allowed each ion to have its own specific conductance.
The total membrane current equals membrane capacity current plus ionic currents:
I = CMdVdt
+ Ii. The ionic current is the sum of the potassium, sodium and other
leaking ion currents whose conductances are not dependent on membrane potential:
Ii = Ik + INa + Il. Ionic currents equal the product of conductance and potential
difference between the membrane’s potential and the resting potential for the ion:
Ii = Ik + INa + Il = gk(E − Ek) + gNa(E − ENa) + gl(E − El).
Note: V = (E − Er) and VNa = (ENa − ER) so (E − ENa) = (V − VNa).This allows the formula for the ionic currents to be:
Ii = gk(V − Vk) + gNa(V − VNa) + gl(V − Vl).
Hence the total membrane current equation is:
I = CMdV
dt+ gk(V − Vk) + gNa(V − VNa) + gl(V − Vl).
2.2 Modeling potassium conductance
In order to find a useful and relatively simple model of best fit, Hodgkin and
Huxley determined the potassium conductance to be proportional to the 4th power
of a variable that obeys a first order differential equation. Two factors are related to
the ability to excite the neurons to produce action potentials, the membrane
potential and the concentrations of the ions on either side of the membrane. The
variable n was chosen to represent the portion of particles inside the membrane
involved with the transfer of potassium to the outside and 1− n represents the
portion of particles outside affecting the transfer of potassium to the inside.
4

Texas Tech University, Julie Bumbaugh, August, 2012
Another set of reasonable definitions for n and 1− n is that they represent the
probability that gates for the transfer of potassium to the outside and inside,
respectively, are open. Perhaps they are easier to understand as the portion that
such gates are open. Hodgkin and Huxley had no way of knowing the actual
physical makeup of the membrane. Next, the rate of change of the portion of said
particles would equal the rate in minus the rate out. They chose to let αn and βn be
the voltage dependent rate constants (constant with respect to time) for the
potassium ions to enter and leave the membrane. Hence,
dn
dt= αn(1− n)− βnn.
A solution to such a differential equation has the form n = C1 − C2e−C3t. Hodgkin
and Huxley used 1− e−t as a simplified model to determine an appropriate power of
n. We can analyze this function and powers of it to understand how they selected
the appropriate power. The first power is obviously inappropriate due to the fact
that the rise of potassium has a sigmoid shape with a single inflection point and
1− e−t has no inflection point. Studying various powers of 1− e−t reveals that the
point of inflection of (1− e−t)p will be located at t = ln(p) for p > 1. Hodgkin and
Huxley had very limited computing capabilities and it took them a great deal of
time to test these models using the calculators that they had available. They
decided to use the 4th power of n and mentioned that the 5th or 6th power might be
an improvement, but they did not feel the improvement would be sizeable enough to
make it worth the extra computational effort. We can note that the larger the value
of p, the less difference there is between ln(p) and ln(p+ 1). If we do think of n as
being a probability that a gate is open or what fraction that gate is open, and if to
model the behavior of ion transfer through the membrane requires the fouth power
of n, then it would seem that there are 4 gates in the channel that must all be open
to some degree for the ions to pass through the membrane. In other words, n is the
fraction that each of 4 required gates are open. Hodgkin and Huxley therefore used
the maximum conductance for potassium times the probability for conductance
through 4 gates at a given membrane potential, gk = gk · n4, to represent the voltage
dependent conductance of potassium. To date, it is held that there are 4 gates for
the transfer of potassium through this membrane. Next is the matter of the rate
5

Texas Tech University, Julie Bumbaugh, August, 2012
constants αn and βn. While they are constant with respect to time, they are voltage
dependent. Since n was to represent the portion of particles inside the membrane
associated with transfer of potassium ions to the outside, Hodgkin and Huxley let
the ratio of rate inrate in + rate out
represent the limiting value of n for the corresponding
value of membrane potential associated with those rates, n∞ = αnαn+βn
. At time
t = 0, they assigned n = n0 = α0
α0+β0. They also assigned τn to be the ratio 1
αn+βn.
With these assignments, we can find the solution to
dn
dt= αn(1− n)− βnn (2.1)
is
n = n∞ − (n∞ − n0)e−tτn . (2.2)
They ultimately needed to find a formula for potassium conductance in terms of
membrane potential, but all of their measurements were against time at fixed
membrane potentials. So, they first derive their equation for potassium conductanc,
gK , for fixed membrane potential values as a function of time.
gK = gK · n4, (2.3)
gK = ( 4√gKn)4, (2.4)
(gK∞)14 = (gK)
14n∞, (2.5)
(gK0)14 = (gK)
14n0, (2.6)
gK = ( 4√gK · (n∞ − (n∞ − n0)e−
tτn ))4, (2.7)
gK = ( 4√gKn∞ − ( 4
√gKn∞ − 4
√gKn0)e−
tτn )4, (2.8)
gK = ((gK∞)14 − ((gK∞)
14 − (gK0)
14 )e−
tτn )4. (2.9)
See Figure 2.1 for the graph of this function. Next, Hodgkin and Huxley had to find
values for gk∞ , gko and τn (or for αn and βn) at different values of V . Potassium
conductance vs. time curves were plotted for different values of V . For each curve,
gk0 and gk∞ were recorded and corresponding values for τn were chosen for each
curve such that the conductance equations would fit the data best.
(Assuming n = 1 at the asymptote of the conductance curves, they chose to let gk
be a value which was 20% higher than the values of gk∞ when V = −100mV since
6

Texas Tech University, Julie Bumbaugh, August, 2012
Figure 2.1. Hodgkin and Huxley’s potassium conductance vs. time curve using pa-rameters of gK0 = .09, gK∞ = 7.06, and τn = .75 for 5s of rise and then gK0 = 7.06,gK∞ = .09, and τn = 1.1 for 5s of the falling phase of conductance. V was heldat −25mv for the first 5s and at 0mv for the next 5s. This graph is plotted usingequation (2.9) for gK with fixed value of V .
they were not concerned with depolarizations greater than 110mV .) So,
20.26 · (1.20) = gk · (n∞)4 = gk · (1) = gk = 24.31. Using their values for gk∞’s, gk0’s,
and τn’s at each V , they calculated corresponding values for αn and βn using the
relationships:
gk∞ = gk · (n∞)4
to solve for n∞’s and then
n∞ =αn
αn + βn= αn · τn
to solve for the αn’s and then
τn =1
αn + βn
to solve for the βn’s.
Next, they plotted curves of αn vs. V , βn vs. V , and τn vs. V in order to see their
shapes and try to find functions in terms of V to fit each of them. Hodgkin and
Huxley made reference to using a function involving “movements of charged
particles in a constant field” from a paper by Goldman to represent their graph of
7

Texas Tech University, Julie Bumbaugh, August, 2012
Table 2.1. Computations for αn’s and βn’s
Curve V (mV ) gk∞(m.mh0/cm2) τn n∞ αn βn—– (−∞) 24.31 —–
B −100 20.26 1.10 .955 .869 0.040
C −88 18.60 1.25 .935 .748 .052
D −76 17.00 1.50 .914 .610 .057
E −63 15.30 1.70 .891 .524 .064
F −51 13.27 2.05 .860 .419 .069
G −38 10.29 2.60 .807 .310 .074
H −32 8.62 3.20 .772 .241 .071
I −26 6.84 3.80 .728 .192 .071
J −19 5.00 4.50 .673 .150 .073
K −10 1.47 5.25 .496 .094 .096
L −6 0.98 5.25 .448 .085 .105
—– (0) (0.24) —– .315
αn vs. V . There is an equation 17 in the Goldman paper [1] on page 53 of that
paper which is referred to as a constant case. The equation is
J =∆V Λ+ − Λ−e
−β∆V
1− e−β∆V
where Λ+ and Λ− are limiting conductance values for large potentials in one
direction or the other. If we split this equation into
J =∆V Λ+
1− e−β∆V− ∆V Λ−e
−β∆V
1− e−β∆V=
∆V Λ+
1− e−β∆V− ∆V Λ−eβ∆V − 1
The second term of the Goldman formula: ∆V Λ−eβ∆V −1
resembles the function Hodgkin
and Huxley used to define αn. This term, from Goldman, describes movements in
one direction through the membrane and the factor Λ− is a conductance limiting
value, like gk, so the rest of that term involves the rate that the limiting value is
applied which is what αn does. If we take the Goldman term without Λ−, ∆Veβ∆V −1
,
and modify it by letting β = 1, vertically shrink it by a factor of 110
, horizontally
dilate it by a factor of 10, and finally translate it 10 units left, we arrive at Hodgkin
8

Texas Tech University, Julie Bumbaugh, August, 2012
and Huxley’s formula for αn,
∆V
eβ∆V − 1⇒ αn =
1
10
[110
(V + 10)
e110
(V+10) − 1
].
We know they were relying on Goldman’s work and that they were tweaking
parameters to get curves to fit their data. It seems reasonable that this is how they
arrived at their formula for αn. Hodgkin and Huxley found a simple exponential
curve to describe βn. They explained that the data values for βn were so much
smaller than those for αn that all they could assertain from the curves was that they
appeared to be simple exponentials. They said they did not worry too much about
whether that was the appropriate choice. Together, these two formulas,
αn =1
10
[110
(V + 10)
e110
(V+10) − 1
],
βn =1
8eV80 ,
along withdn
dt= αn(1− n)− βn · n,
and
gk = gk · n4,
complete the Hodgkin-Huxley definition for potassium conductance in terms of
membrane potential V and the activation variable n.
2.3 Modeling sodium conductance
The conductance of sodium is considerably different than that of potassium due
to its transient nature meaning that it seems to have both activation and
inactivation gates. Activation gates open to let sodium ions in and later inactivation
gates open to let it out. Potassium conductance rises to a limiting value and stays
there unless the membrane is repolarized. Sodium conductance, on the other hand,
rises and falls again during a maintained depolarization of the membrane. Hodgkin
and Huxley could only speculate as to what was causing this transient behavior, but
9

Texas Tech University, Julie Bumbaugh, August, 2012
they knew their model for sodium would need to include a factor that would cause
the conductance to return to resting values during long depolarizations. We need
first to see how they determined what power the activation variable and inactivation
variable should each be raised to. They used a similar form for sodium conductance
as they did for potassium: gNa = gNamphq. They named the activation variable
responsible for activating the carrying in of sodium, m, and the inactivation variable
responsible for shutting down the inflow of sodium and restoring the resting levels,
h. Similarly to potassium, they named the limiting value for the conductance
inward of sodium gNa. The rates of change of the activation and inactivation
variables have the same form as the potassium activation variable, n.
dm
dt= αm(1−m)− βm ·m
dh
dt= αh(1− h)− βh · h
αm and βh are rate constants (voltage dependent, but time constant) responsible for
the carrying of sodium ions into the membrane. βm and αh are the rate constants
resposible for the carrying of sodium ions to the outside of the membrane. Hodgkin
and Huxley did not know the physical makeup of the membrane, so their rationale
was that m could represent the portion of some particles on the inside associated
with the carrying of sodium ions to the outside and (1−m) would then be the
portion of the same type of particles on the outside associated with the carrying of
sodium ions to the inside during the rise of sodium conductance. Likewise, they
used h to represent the portion of some particle on the outside of the membrane
assosciated with the carrying of sodium ions to the inside and (1− h) as the
remaining portion of the same particle on the inside associated with the carrying of
sodium ions to the outside during the decline of sodium conductance. Solutions to
these differential equations have the same form as potassium’s activation variable.
m = m∞ − (m∞ −m0)e−tτm ,
h = h∞ − (h∞ − h0)e− tτh .
10

Texas Tech University, Julie Bumbaugh, August, 2012
Just as with n,
m∞ =αm
αm + βmand h∞ =
αhαh + βh
,
τm =1
αm + βmand τh =
1
αh + βh.
Before experimenting with various powers of m and h, they made some crucial
observations which simplified the process. They found that during rest, sodium
conductance was so small compared to its values during a large depolarization that
they chose to neglect m0 whenever depolarizations were over 30mV . Also,
inactivation seemed to be nearly complete whenever V < −30mV , so that h∞ would
be neglected as well. These simplifications adjust the formulas for m and h to be:
m = m∞ −m∞e−tτm ,
h = h0e− tτh .
At this point, they must have experimented with various combinations of powers of
m and h. They needed the right shape for the rise of sodium conductance. It
needed to have a rise with a single point of inflection and then a fall with no point
of inflection. Their choice was to use gk = gkm3h. This brings the sodium
conductance formula to be
gNa = gNa(m∞ −m∞e−tτm )3(h0e
− tτh ),
gNa = gNa(m∞)3h0(1− e−tτm )3e
− tτh .
Next, they assigned g′Na = gNam3∞h0, which is the maximum level that gNa would
attain if h remained at its resting level, h0. So, sodium conductance becomes:
gNa = g′Na(1− e−tτm )3e
− tτh .
In this definiton of gNa, we see a form of the familiar (1− e−t)p which only has a
point of inflection at t = ln p, so m had to be raised to a power greater than 1 since
a slight change of inflection is visible in the rise of sodium conductance on some of
the curves. If p1 < p2, then (1− e−t)p2 < (1− e−t)p1 for all t > 0; so, the maximum
value of sodium conductance will be lower for larger values of p at the moment
11

Texas Tech University, Julie Bumbaugh, August, 2012
sodium conductance starts to fall. It is assumed here that the 3rd power provided
the optimum shape needed to fit the data. Hodgkin and Huxley then plotted a
family of curves for gNa = g′Na(1− e−tτm )3e
− tτh each with a different ratio for τm to
τh in order to best fit each of their gNa vs. t sets of data plots. Each plot is
associated with a different depolarization, so the voltage dependent time constants
had to be adjusted for each curve. This gave them τm’s and τh’s for each V to be
used with the measured g′Na’s to calculate the corresponding αm’s and βm’s. From
the formula, m∞ = αmαm+βm
, we can get:
αm =m∞τm
and βm =1−m∞τm
.
From the maximum level of gNa = g′Nam3∞h0, each m∞ was obtained by using
3√g′Na and the assumption that gNa approaches 1 for large depolarizations. Now,
they had a set of data for αm’s and βm’s and needed a function to fit them. The
function of V that Hodgkin and Huxley used to define αm was again a
transformation of the curve y = VeV −1
, from Goldman’s work, which is asymptotic to
y = 0 and decreases with a shape very close to the shape of the αm vs. V data. This
is the same parent function used to define αn. This time they horizontally dilated
the parent curve by a factor of 10 and translated it 25 units left to arrive at
αm =0.1(V + 25)
e0.1(V+25) − 1.
Another simple exponential was used to define the βm’s:
βm = 4eV18 .
One can also test these formulas by graphing the measured values for m∞’s against
the curve m∞ = αmαm+βm
and see that it is a reasonable fit. To obtain inactivation
rate constants, αh and βh, Hodgkin and Huxley calculated values for them using:
αh =h∞τh
and βh =1− h∞τh
.
12

Texas Tech University, Julie Bumbaugh, August, 2012
The values were plotted against V and the following curves were used to fit them:
αh = 0.07eV20 and βh =
1
eV+30
10 + 1.
One can again test these formulas by plotting the measured h∞’s with the curve for
h∞ = αhαh+βh
and seeing that it too is a reasonable fit. Now we have the sodium
conductance well defined by the equations:
αm =0.1(V + 25)
e0.1(V+25) − 1, βm = 4e
V18 ,
αh = 0.07eV20 , βh =
1
eV+30
10 + 1,
dm
dt= αm(1−m)− βm ·m,
dh
dt= αh(1− h)− βh · h,
and gNa = gNam3h.
13

Texas Tech University, Julie Bumbaugh, August, 2012
CHAPTER 3
SOLVING THE HODGKIN-HUXLEY SPACE CLAMP MODEL
Remember that the total membrane current was defined as:
I = CMdV
dt+ gk(V − Vk) + gNa(V − VNa) + gl(V − Vl).
Now that we have the ionic concuctances well defined, we can give the final form of
this equation along with all of the other necessary defining equations all of which
are functions of V and t:
I = CMdV
dt+ gkn
4(V − Vk) + gNam3h(V − VNa) + gl(V − Vl), (3.1)
dn
dt= αn(1− n)− βn · n, (3.2)
dm
dt= αm(1−m)− βm ·m, (3.3)
dh
dt= αh(1− h)− βh · h, (3.4)
αn =1
10
[110
(V + 10)
e110
(V+10) − 1
], (3.5)
βn =1
8eV80 , (3.6)
αm =0.1(V + 25)
e0.1(V+25) − 1, (3.7)
βm = 4eV18 , (3.8)
αh = 0.07eV20 , (3.9)
βh =1
eV+30
10 + 1. (3.10)
During a “voltage clamp” experiment where membrane potential is held at a
constant value, dVdt
= 0 and the one-way conductance rates, α’s and β’s, which are
voltage dependent, should then also be constant. At time, t = 0, I = 0, V = 0, m,
n, and h will begin at their resting or, steady state, values of m0, n0, and h0. The
last term of the total current equation, current due to “leaking ions”, has a rate
constant that is not dependent on membrane potential. Hodgkin and Huxley
14

Texas Tech University, Julie Bumbaugh, August, 2012
created a term here that causes the resting potential of the membrane to be zero for
convenience. With the capacitance, C, of the membrane equal to 1 µF/cm2 and the
total current = 0, we can rewrite equation (3.1) as:
−dVdt
= gkn4(V − Vk) + gNam
3h(vV − VNa) + gl(V − Vl). (3.11)
To solve the system of equations (3.2) through (3.11) we will apply Euler’s method
to obtain successive values of the four variables, V , n, m, and h and then plot
curves of V vs. t for different depolarizations on the membrane. Table 3.1 holds the
constants needed for the calculations and the matlab program for solving the system
can be found in the Appendix A. Graphs of various membrane potential vs time
solutions to the Hodgkin-Huxley model for squid giant axons which were generated
by the Matlab program and are shown in Figure 3.1.
Table 3.1. List of variables and parameters for the Hodgkin-Huxley modelI Membrane Current Density (0A/cm2 during space clamp)
CM Membrane Capacitance (1 µF/cm2)
V Membrane Potential (mV )
t time (ms)
gk maximal conductance for potassium (36 m.mho/cm2)
n Proportion of particles in neuron involved with opening of potassium gate
Vk Resting Potential of potassium (12mV)
gna maximal conductance for sodium (120 m.mho/cm2)
m proportion of particles in neuron affecting opening of sodium gate
h proportion of particles outside of neuron affecting closing of “2nd” sodium gate
Vna Resting potential for Sodium (−115 mV)
gl maximal conductance for “leaking” ions (0.3 m.mho/cm2)
Vl Resting potential for “leaking” ions (−10.613 mV)
αn Voltage dependent rate for potassium ions to enter the cell per ms
βn Voltage dependent rate for potassium ions to exit the cell per ms
αm Voltage dependent rate for sodium to enter the cell per ms via the “1st” gate
βm Voltage dependent rate for sodium to exit the cell per ms via the “1st” gate
αh Voltage dependent rate for sodium to exit the cell per ms via the “2nd” gate
βh Voltage dependent rate for sodium to enter the cell per ms via the “2nd” gate
15

Texas Tech University, Julie Bumbaugh, August, 2012
Figure 3.1. Membrane action potentials in squid giant axons: the upper left graphis of membrane potential vs. time with the initial condition V (0) = 20mV . In theupper right graph, V (0) = −20mV . Lower left graph uses V (0) = 2.9mV and thelower left graph uses V (0) = 1mV .
16

Texas Tech University, Julie Bumbaugh, August, 2012
CHAPTER 4
MODERN TECHNIQUES TO ANALYZING DYNAMICAL SYSTEMS
Much progress has been made in the study of spike generation since the work of
Hodgkin and Huxley. Working without the knowledge of the membrane structure,
scientists tried to build models of a neuron by adjusting parameters that were
measured and not necessarily from the same neuron. The study of dynamical
systems today asks and tries to answer the questions of why two seemingly similar
neurons can behave so differently under the same conditions. A dynamical system
consists of a set of variables that describe its state and a law that describes the
evolution of the state variables with time [7].
The Hodgkin Huxley model is a dynamical system consisting of the state
variables V , n, m, and h with a four-dimensional system of ordinary differential
equations governing the evolution of the state variables. We will see that the
Hodgkin Huxley model can be reduced to a two-dimensional model and still produce
the same action potentials. Then we will perform analysis on the reduced model to
explain some of the dynamics of the squid giant axons. We begin by seeing how
other models are analyzed.
4.1 One-dimensional models
An example of a one-dimensional system is the persistent sodium model,
CV = I − gL(V − EL)− gNam∞(V )(V − ENa)
with m∞(V ) =1
1 + e(V1/2−V )/k.
This formula for m∞(V ) is much easier to find parameters for than the way
Hodgkin and Huxley did it from formulas for αm’s and βm’s. We show the
derivation of this Boltzmann function and how to get the parameters in chapter 5.
The state of this model is described by the membrane potential V . Using the
following experimental parameter values, we can plot the I − V , steady state
current-voltage relations and the voltage vs. time relations to better understand
17

Texas Tech University, Julie Bumbaugh, August, 2012
this system. The parameters used are shown in Table 4.1.
Table 4.1. Parameters for persistent sodium model
C = 10µF I = 0pA gL = 19ms EL = −67mVgNa = 74ms V1/2 = 1.5mV k = 16mV ENa = 60mV
4.1.1 Stability of equilibria
Finding equilibria and determining their stability is an important step in
analyzing a dynamical system. The graph of V vs. V in Figure 4.1.1 shows that the
persistent sodium model has 3 equilibria due to the fact that V = 0 for 3 values of
V .
The smallest potential for which the membrane is in a state of equilibrium is a
stable equilibrium since small perturbations to the left of it would place the
potential in a region where V is positive and would thus send the potential back to
the right to the equilibrium point. Small perturbations to the right of it would place
the potential in a region where V is negative and would send the potential back to
the left towards the equilibrium.
The signs of V on either side of the middle equilibrium point show that it is
unstable. Small perturbations of the membrane potential from this equilibrium
point would send the potential into regions where V would send the potential in the
opposite direction towards one of the other two equilibria. The same argument
applies to the largest equilibrium point making it also a stable equilibrium. The
smallest of the stable points holds the potential at a resting value, while the larger
one holds it in an excited state.
We can conclude, in the one-dimensional system, that when the slope of the V
vs. V is negative at an equilibrium, it will be stable and when the slope is postive at
an equilibrium, it will be unstable. If we define V = F (V ), then for steady state,
F (V ) = 0. When F ′(V ) < 0, the equilibrium is stable and when F ′(V ) > 0, the
18

Texas Tech University, Julie Bumbaugh, August, 2012
Figure 4.1. Graphs of persistent sodium model - Upper Left: Bistability trajectorieswith initial current I = 0. Upper Right: Monostabilty trajectories with initial currentI = 60. Lower Left: V vs. V . Lower Right: Steady State I − V relation (Currentvs. Membrane Potential).
equilibrium is unstable. We define the pararmeter, λ, the simplest eigenvalue for an
equilibrium, as
λ = F ′(V ). (V is an equilibrium)
If λ < 0, the equilibrium is stable. If λ > 0, the equilibrium is unstable. We will see
the same relationship between eigenvalues and equilibria with two-dimensional
systems.
19

Texas Tech University, Julie Bumbaugh, August, 2012
Equivalently, if the slopes of the steady-state I − V curve are positive at an
equilibrium, the equilibrium would be stable due to the fact that the signs of the
steady-state I − V curve and the V vs. V curve are opposite of each other. When
V 6= 0, then the sign of V at a particular value of V is an indication of flow of
current in a particular direction. To maintain a steady-state at that same value of
membrane potential, V , there would need to be an injected current flowing in the
opposite direction. Therefore, the signs of the graphs of steady state I − V and V
vs. V would be opposite and their zeros would be at the same membrane potential
values.
4.1.2 Bifurcations
Determining the parameter values that can change a system’s phase portrait
qualitatively is another important step in analyzing a dynamical system. The
parameter which affects whether the phase portrait contains one or two stable
equilibria is I, the injected current. There are values of I for which the phase
portrait changes from 3 to 2 to 1 equilibrium. When the phase portrait changes
qualitatively like this, the system is said to undergo a bifurcation.
The graphs in Figure 4.1.1 depicting Bistability and Monostability of the
persistent sodium model are easily explained since the case of Bistability occurred
with an injected current, I = 0pA, which has been shown to have 2 stable equilibria.
If a current of I = 60pA were to be injected into the membrane, the steady state
curve would be translated up 60pA and would then have only 2 zeros corresponding
to one unstable equilibrium and the excited state stable equilibrium. It is not likely
that the membrane could stay at the unstable equilibrium potential since small
perturbations are very common and would send the potential to the excited state.
The 2 graphs depicting monostability and bistability of Figure 4.1.1 depict 2
qualitatively different phase portraits. The type of bifurcation depicted by the
persistent sodium model is known as a saddle-node bifurcation which is one in
which the system can change from having one stable resting equilibrium, one
unstable equilibrium and one stable excited state to a system with only one stable
excited state.
20

Texas Tech University, Julie Bumbaugh, August, 2012
The one-dimensional potassium model, IK-model,
CV = −gL(V − EL)− gKm4∞(V )(V − EK)
does not have a saddle-node bifurcation. It is relatively easy to establish that it
cannot have a saddle-node bifurcation for V > EK . The parameters for this model
are found in Table 4.2. In order to have a saddle-node bifurcation when V > EK ,
Table 4.2. Parameters for 1-dimensional potassium model
C = 1 gL = 1 EL = −80 gK = 1 EK = −90 k = 15
the V vs. V graph would have to have an N-shaped curve and, if V = F (V ), then
F ′(V ) must equal zero more than once when V > EK . For this model,
F ′(V ) =dF
dV= −gL − gK(m4
∞(V )− 4gkm3∞(V )(V − EK).
If V > EK , then all of the terms of this derivative have a negative sign. Therefore,
F ′(V ) < 0 for all V > EK . So, F (V ), or V , is monotonic when V > EK and cannot
equal zero more than once on that interval; therefore, the model does not have a
saddle-node bifurcation when V > EK .
4.2 Two-dimensional models
Let us consider the 2-dimensional persistent sodium + potassium model denoted
as: INa,p + IK .
CV = I − gL(V − EL)− gNam∞(V )(V − ENa)− gKn(V − EK), (4.1)
n = (n∞(V )− n)/τ(V ), (4.2)
n∞(V ) =1
1 + exp(V1/2 − V )/k, (4.3)
m∞(V ) =1
1 + exp(V1/2 − V )/k. (4.4)
The state of this system is a 2-dimensional vector (V, n) on the phase plane R2
21

Texas Tech University, Julie Bumbaugh, August, 2012
since the Na+ current has instantaneous activation kinetics, such that its
conductance may be considered to be maximal, m∞, over most all of the time
interval. The activation kinetics for n are much slower and so it must be defined by
its derivative. Parameters for this low-threshold model are given in table (4.3)
Table 4.3. Parameters for low-threshold INa,p + IK modelC = 1 I = 0 EL = −78mV gL = 8 gNa = 20gK = 10 m∞has V1/2 = −20 k = 15 n∞has V1/2 = −45 k = 5τV = 1 ENa = 60mV EK = −90mV
4.2.1 Nullclines
We first need to establish the equilibrium points by studying the null clines of the
state variables. A nullcline is all of the locations in the phase plane where a state
variable is at rest. In this system, that would be when V = 0 and when n = 0. The
equation for the V nullcline is:
0 = I − gL(V − EL)− gNam∞(V )(V − ENa)− gKn(V − EK),
n =I − gL(V − EL)− gNam∞(V )(V − ENa)
gK(V − EK),
where m∞(V ) =1
1 + e(V1/2−V )/K.
The n nullcline is:0 = n∞(V )− n,
n = n∞(V ) =1
1 + e(V1/2−V )/K.
The intersections of these nullclines will be the points where neither of the state
variables is changing, so the membrane is at equilibrium. In Figure 4.2, we see the
cubic-shaped nullcline for V in red and the sigmoid-shaped nullcline for n in green.
Their intersection is the equilibrium for this model.
This figure also is demonstrating that some models fail to have “all or nothing
spikes”. We can see trajectories of the phase plane which have varying initial values
for membrane potential but the same initial value for the potassium activation
22

Texas Tech University, Julie Bumbaugh, August, 2012
variable, “n”. It is apparent that some trajectories follow a subthreshold path to the
equilibrium and others take a longer excursion with greater values of membrane
potential. So, there are varying amplitudes of action potentials, not “all or nothing”
spikes. In Figure 4.3, we observe the INa,p + IK low threshold system has
trajectories all starting with the same initial membrane potential of −48mV , but
varying values of initial K+ activation variable “n”. Some trajectories depict full
action potentials and some make small excursions and return to equilibrium with
subthreshold spikes. We can see that this model has no fixed threshold of membrane
potential. All of the action potentials exhibited in Figures 4.2 and 4.3 are transient;
they all return to equilibrium values.
Figure 4.2. Trajectories of a 2-dimensional system: I = 0. (low-threshold) INa,p + IKmodel: Failure to generate all-or-nothing action potentials is depicted.
4.2.2 Stability in 2-dimensions
Now we need to address how we can determine the stability of the equilibrium. If
at least one trajectory diverges from the equilibrium, then it is said to be unstable.
Unlike the one-dimensional systems, stability can not be determined from the slope
of the V vs. V curve. For example, the slope of the steady-state I − V curve for the
23
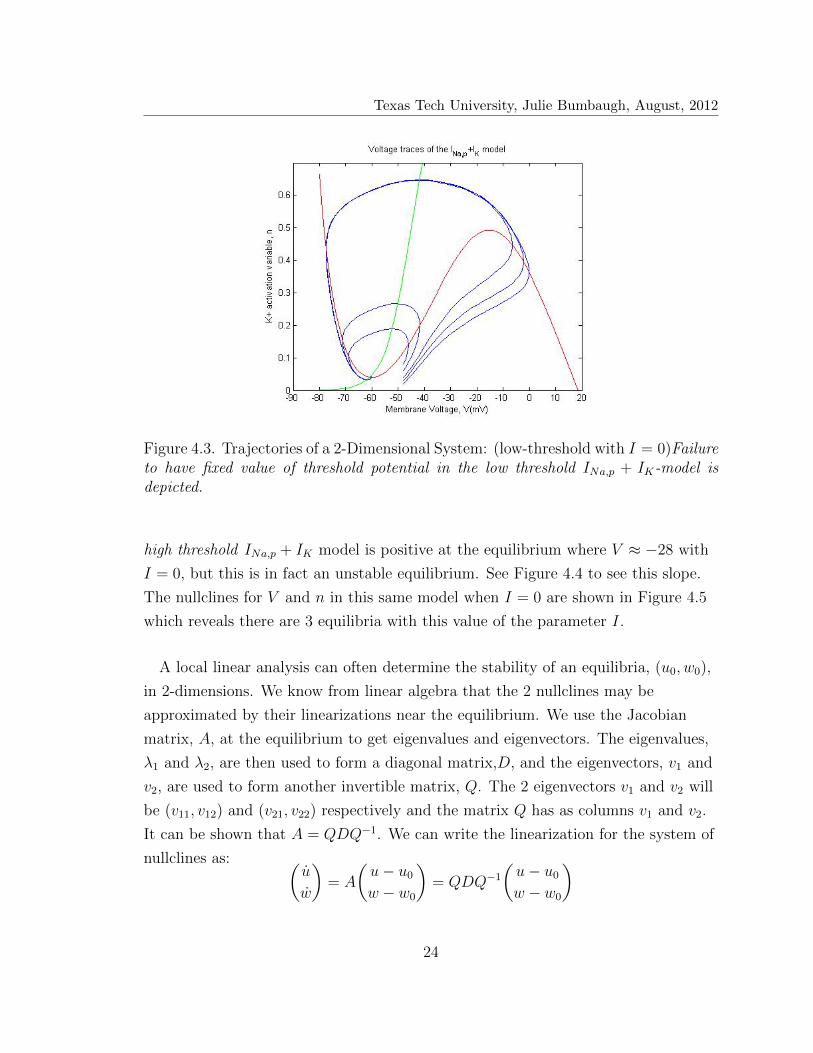
Texas Tech University, Julie Bumbaugh, August, 2012
Figure 4.3. Trajectories of a 2-Dimensional System: (low-threshold with I = 0)Failureto have fixed value of threshold potential in the low threshold INa,p + IK-model isdepicted.
high threshold INa,p + IK model is positive at the equilibrium where V ≈ −28 with
I = 0, but this is in fact an unstable equilibrium. See Figure 4.4 to see this slope.
The nullclines for V and n in this same model when I = 0 are shown in Figure 4.5
which reveals there are 3 equilibria with this value of the parameter I.
A local linear analysis can often determine the stability of an equilibria, (u0, w0),
in 2-dimensions. We know from linear algebra that the 2 nullclines may be
approximated by their linearizations near the equilibrium. We use the Jacobian
matrix, A, at the equilibrium to get eigenvalues and eigenvectors. The eigenvalues,
λ1 and λ2, are then used to form a diagonal matrix,D, and the eigenvectors, v1 and
v2, are used to form another invertible matrix, Q. The 2 eigenvectors v1 and v2 will
be (v11, v12) and (v21, v22) respectively and the matrix Q has as columns v1 and v2.
It can be shown that A = QDQ−1. We can write the linearization for the system of
nullclines as: (u
w
)= A
(u− u0
w − w0
)= QDQ−1
(u− u0
w − w0
)
24

Texas Tech University, Julie Bumbaugh, August, 2012
Figure 4.4. Bifurcations for high threshold INa,p + IK model. It appears that whenI = 4.5 and I = −85.5 the state of this neuron changes from 3 equilibria betweenthese two values of injected current to 2 and then 1 equilibria.
Figure 4.5. Nullclines for high threshold INa,p + IK model with I = 0: Red curve isnullcline for V and Green curve is nullcline for activation variable n. Curves appearto have equilibria at approximately: V = −66, V = −56, and at V = −28.
25

Texas Tech University, Julie Bumbaugh, August, 2012
Multiplying through by Q−1 on the left sides yields
Q−1
(u
w
)= Q−1A
(u− u0
w − w0
)= DQ−1
(u− u0
w − w0
).
Now, let y = Q−1(u−u0
w−w0
). This makes dy
dt= Q−1
(uw
). So, dy
dt= Dy. Solving for y will
lead us to finding a function of time to represent the magnitude of the deviations
from the equilibrium: (dy1
dtdy2
dt
)=
(λ1y1
λ2y2
),(
y1
y2
)=
(C1e
λ1t
C2eλ2t
).
Now that we have y, since y was defined to be Q−1(u−u0
w−w0
), we have(
u− u0
w − w0
)= Q · y,
=
(v11 v21
v12 v22
)·
(c1e
λ1t
c2eλ2t,
)
=
(v11c1e
λ1t + v21c2eλ2t
v12c1eλ1t + v22c2e
λ2t
)= (v11c1e
λ1t, v12c1eλ1t) + (v21c2e
λ2t, v22c2eλ2t)
= C1v1eλ1t + C2v2e
λ2t.
So, for any 2-dimensional system, if the eigenvalues are both negative or have
negative real parts, then as t→∞, the vector(u−u0
w−w0
)→(
00
)meaning that
deviations from the equilibrium will go to 0 making (u0, w0) stable. It is enough in
some cases, therefore, to find the λ’s to determine the stability of equilibria in
2-dimensional systems. Recall that an eigenvalue in one dimensional systems was
simply the slope of the V vs. V curve at the equilibrium and, if it was negative, it
meant the equilibrium was stable. (Equivalently, as explained earlier, if the slope of
the I − V curve is positive at equilibrium, it is stable because the signs of the V vs.
V curve and the I − V curve are opposite and their slopes would be opposite as
well.) Now, we know that negative eigenvalues also imply stability in 2-dimensions.
26

Texas Tech University, Julie Bumbaugh, August, 2012
4.2.3 Limit cycles
In 2 dimensions, it is possible to have trajectories that follow a repeating cycle. A
trajectory that forms a closed loop is called a periodic trajectory. An isolated
periodic trajectory is called a limit cycle. When a neuronal model exhibits a limit
cycle, it means the neuron would be firing repetitive action potentials in this state.
Figure 4.6 illustrates the low-threshold INa,p + IK model with I = 40 this time
rather than 0. With this change of the parameter, I, the trajectories follow such a
limit cycle. When I was zero, in this model, the eigenvalues at the intersection of
the nullclines were negative, indicating stability. With I = 40, the eigenvalues are
positive. The Figure 4.7 shows us snapshots of the limit cycle’s appearance and
disappearance between the values I = 0 and I = 400. As the injected current
continues to be raised, we see the amplitude of the limit cycle shrinks and
eventually the limit cycle vanishes and is replaced by a stable excited equilibrium.
Figure 4.6. The stable limit cycle of the persistent INa,p + IK (low-threshold) modelwith I=40. The equilibrium at the intersection of the nullclines is not stable.
4.2.4 Bifurcations in 2-dimensions
A zero eigenvalue arises when the equilibrium undergoes a bifurcation [7]. That
makes sense, since negative eigenvalues ensure stability and positive ensure
unstability. Naturally, if an eigenvalue’s real part is zero, the system must be
undergoing some sort of qualitative change. To study the bifurcation(s) a system
may have, we can analyze models where we change a parameter that leads to a
27

Texas Tech University, Julie Bumbaugh, August, 2012
Figure 4.7. Low threshold INa,p + IK traces leading to excitation block
bifurcation. We will look at the steady-state I − V curve for the system to see
where such bifurcations exist. Where the curve has a change in direction, is a
bifurcation since the number of equilibrium values of V which correspond to this
value of the current changes as we raise or lower the value of I from this location.
In Figure 4.4, we saw that the bifurcations for that model occur at approximately
I = 5 and I = −85. By determining potential values where the difference between
the two nullclines is zero near these bifurcation currents, we have found that when
28

Texas Tech University, Julie Bumbaugh, August, 2012
I = 4.496, the difference in nullclines is very close to zero, (we are near an
equlibrium and close a bifurcation), when V = −60.63398. Using I = −85.607, the
difference between the nullclines is very close to zero for V = −36.17021. We can
obtain corresponding values for n for these potentials and injected current values
and then compute each equilibrium’s eigenvalues to determine the stability of the
equilibria. At these equilibria, the eigenvalues are negative, indicating stability.
Trying again, with a slightly higher potential than the first one, V = −58.5, the
eigenvalues are positive indicating instability. What this means is that the equilibria
left of the left knee in the steady-state I − V curve are stable and right of it are
unstable. As the injected current approaches 5 from below, the equilibria are
getting closer together and when they become one, one of the eigenvalues will be
zero. The stable node and the unstable saddle coelesce to form the saddle-node
which is stable on one side and unstable on the other. If we now examine the
dynamics near the right knee of the steady state curve, the eigenvalues at the
bifurcation value of I are positive. This equilibrium is unstable.
4.2.5 Stable and unstable manifolds
It is of interest to examine at least two trajectories. The first being of an initial
potential, V , between the stable and middle unstable equilibria when I = 3 with
τ = 0.152. The second consists of an initial potential, V to the right of the middle
equilibria. See Figure 4.8. This figure demonstrates that there is a region, known as
a manifold which is the attraction domain for a stable limit cycle. It is also
apparent that the region outside of this manifold, (the unstable manifold) is the
attraction domain for the resting state at the leftmost equilibrium which is stable.
4.2.6 Hodgkin-Huxley reduced model
Finally, in this chapter on 2-dimensional systems, we address the fact that the
Hodgkin Huxley Model may be reduced to a two-dimensional model by replacing
one of its state variables having fast kinetics with its maximal value since it achieves
this value almost instantaneously. The other variable can be substituted by a
function of one of the remaining state variables. In order to successfully minimalize
a neuronal model, it must be able to still produce a spiking limit cycle. In order to
29

Texas Tech University, Julie Bumbaugh, August, 2012
Figure 4.8. The trajectories of the persistent sodium + potassium (high threshold)model with I = 3 and τ = 0.152. Bistability is exhibited with a resting equilitbriumwhich 3 of the trajectories shown in light blue end at and a spiking state shown bythe staable limit cyde which is the trajectory in yellow. The dark blue curve is thenullcline for V and the red is the nullcline for n.
do this, the model must have one amplifying and one resonant variable. One to
amplify a depolarization and one to restore or reset values so that another action
potential can occur.
In the case of the Hodgkin-Huxley model with its 4 state variables, m, n, h, and
V , an amazing near linear relationship was found between the variables h and n by
Krinskii and Kokoz in 1973 [7]. If these variables are plotted in the (n, h) plane, the
curve is seen to be very close to the line h = 0.89− 1.1n. This discovery along with
the determination that the action kinetics for sodium are practically instantaneous,
such that m may be replaced with m∞(V ), reduces the Hodgkin-Huxley model to 2
dimensions. See equations (4.5) and (4.6).
Some modern conventions have been applied here. Hodgkin and Huxley’s model
used (V − VK) rather than (V − EK) because it was what they could measure.
30

Texas Tech University, Julie Bumbaugh, August, 2012
Their model also used the opposite signs for membrane potentials. The ions’ Nernst
potentials are used today, EK , ENa, and El which have the opposite values to the
values of VK , VNa, and Vl used in the older model. This reflects the V vs. t curves
over the t axis. Modern convention to correct the signs of the potential also involve
changing (V − 10) to (10− v), (V − 25) to (25− V ),(V − 30) to (30− V ) and V to
−V in equations (3.5) through (3.10).
CV = I − gKn4(V − EK)− gNa(m∞(V ))3(0.89− 1.1n)(V − ENa)−
gL(V − EL), (4.5)
n =(n∞(V )− n)
τn(V ). (4.6)
This definition for n is equivalent to equation (3.2) using the definitions for τn and
n∞ stated earlier. (Note: the Hodgkin-Huxley formula for n∞ is not as easy to work
with as using the Boltzmann function.) To analyze this minimalized model of the
Hodgkin-Huxley model, we first find the intersections of its nullclines to find the
equilibria. The V nullcline must satisfy:
n = 4
√I − gNam3
∞(V )(V − ENa)− gL(V − EL)
gK(V − EK). (4.7)
The n nullcline must satisfy:
n = n∞(V ). (4.8)
We will see that there is one equilibrium and that it is stable when I = 0 but
becomes unstable as seen when I = 12. See figures 4.9 and 4.10.
Calculations of the eigenvalues for each of the equilibria of the Hodgkin-Huxley
models, with I = 0 and I = 12 are negative and positive respectively, confirming
what we see geometrically, that the equilibrium is stable when I = 0 and unstable
when I = 12. See chapter 5 for these calculations. There is an Andronov-Hopf
bifurcation somewhere between these 2 values of I which means that the unique
equilibrium is stable for small values of I but becomes unstable when I increases
31

Texas Tech University, Julie Bumbaugh, August, 2012
Figure 4.9. Hodgkin-Huxley space-clamp model reduced to 2 variables with I = 0.The intersection of the nullclines and 2 traces show that the equilibrium appears tobe stable.
Figure 4.10. Hodgkin-Huxley space-clamp model reduced to 2 variables with I = 12.The intersection of the nullclines and 2 traces show that the equilibrium appears tobe unstable. A stable limit cycle appears somewhere between I = 0 and I = 12.
32

Texas Tech University, Julie Bumbaugh, August, 2012
past 12 giving birth to a limit cycle attractor [7].
It can be shown that, in models like the Hodgkin-Huxley model, as I continues to
increase, the amplitude of the stable limit cycle decreases until the excitation state,
ability to produce action potentials, is blocked. See the next chapter for simulations
and analysis to justify these claims.
33

Texas Tech University, Julie Bumbaugh, August, 2012
CHAPTER 5
PROBLEMS FOR STUDY AND ANALYSIS
Modeling the real world with mathematical equations is one of the greatest
reasons we study mathematics. Students of Mathematics may follow the directions
below to practice some of the modern techniques to studying dynamical systems
and some of the mathematical steps that Hodgkin and Huxley used to derive their
model of action potentials in squid giant axons.
5.1 Proposed problems for study
5.1.1 Problem 1- Derive Boltzmann’s function
Today, the Boltzmann function is used to describe the steady-state activation
variables m∞, n∞, and inactivation variable h∞. To derive this formula, we note
that the shape of the curves from measured values of m,n and h resemble logistics
curves with their sigmoid shape. Boltzmann’s function also has the form of a
logistics curve with the asymptote at y = 1:
m∞ =1
1 + e−1k
(V−V1/2).
The limiting value for all of these curves is 1 because these variables represent the
ratio of these activation variables in membrane at each value of V . The maximum
ratio would be 1. Plotted curves of measured values for m,n, and h vs. V are used
to find the slopes of the curves at the inflection points to determine the parameter
C1. Solve the logistics Differential equation (5.1) for m∞(V ) to derive Boltmann’s
function:dm∞dV
= C1(m∞)(1− m∞C2
). (5.1)
Use C2 = 1 as the limiting value and call the tangent slope at the inflection point1/22k
since we know the vertical location of an inflection point on a logistics curve
occurs at half the limiting value. They represented the run as 2k for a nicer looking
result. See Figure 5.1 to understand how 2k is obtained from the graphs. V1/2
represents the value of the membrane potential at the inflection point which is
where the value of the activation/inactivation variable is 1/2.
34
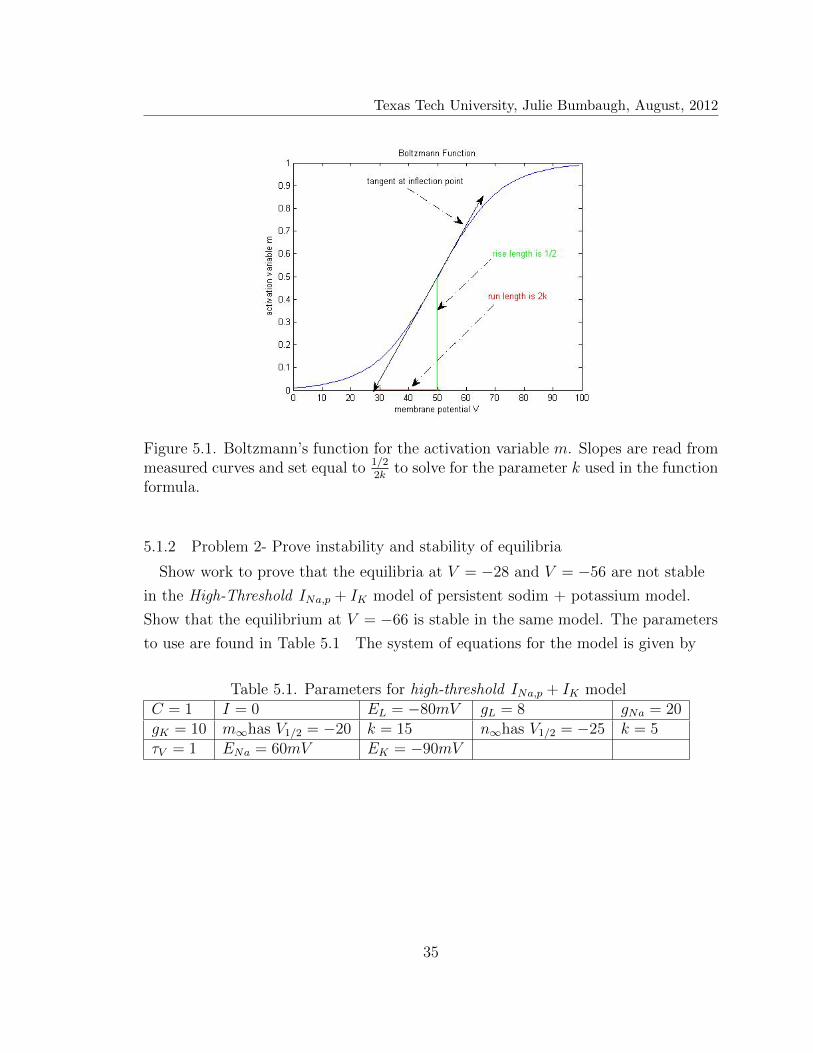
Texas Tech University, Julie Bumbaugh, August, 2012
Figure 5.1. Boltzmann’s function for the activation variable m. Slopes are read frommeasured curves and set equal to 1/2
2kto solve for the parameter k used in the function
formula.
5.1.2 Problem 2- Prove instability and stability of equilibria
Show work to prove that the equilibria at V = −28 and V = −56 are not stable
in the High-Threshold INa,p + IK model of persistent sodim + potassium model.
Show that the equilibrium at V = −66 is stable in the same model. The parameters
to use are found in Table 5.1 The system of equations for the model is given by
Table 5.1. Parameters for high-threshold INa,p + IK modelC = 1 I = 0 EL = −80mV gL = 8 gNa = 20gK = 10 m∞has V1/2 = −20 k = 15 n∞has V1/2 = −25 k = 5τV = 1 ENa = 60mV EK = −90mV
35

Texas Tech University, Julie Bumbaugh, August, 2012
equations (5.2) to (5.5).
CV = I − gL(V − EL)− gNam∞(V )(V − ENa)− gKn(V − EK), (5.2)
n = (n∞(V )− n)/τV , (5.3)
n∞(V ) =1
1 + e(V1/2−V )/k, (5.4)
m∞(V ) =1
1 + e(V1/2−V )/k. (5.5)
5.1.3 Problem 3- Find equilibrium and determine its stability
Use the Hodgkin-Huxley 2-dimensional reduced model parameters in Table 3.1
with I = 12 to write a program that will locate the value of V at the equilibrium of
the model with this injected current. The equations for the nullclines of this model
are (4.7) and (4.8). Then find the corresponding value for n. Finally, determine the
stability of this equilibrium. Finding the partial derivatives at the equilibrium is
quite complex for this model. Writing a program to perform the numerous
calculations would be very helpful.
5.1.4 Problem 4- Demonstrate excitation block
Show that as the injected current increases in the low threshold INa,p + IK model,
that an excitation block occurs which decreases the amplitude of the action
potentials until the limit cycle vanishes. See equations (4.1) through (4.4) and Table
4.3 for the equations defining this model and its parameters.
5.1.5 Problem 5- Choose appropriate power for an activation variable
Hodgkin and Huxley chose the 4th power of the activation variable n in their
model for potassium conductance. They were using the fact that 1− e−t is a
particular solution to the differential equation describing the rate of change of
potassium. Show/explain why the first power of this solution does not work as a
good model and theorize why it is reasonable that they would have stopped and
been satisfied with the 4th power of this solution.
36

Texas Tech University, Julie Bumbaugh, August, 2012
5.1.6 Problem 6- Derive the formula Hodgkin and Huxley used for n
Show how Hodgkin and Huxley arrived at the formula for n, in equation (2.2)
from the differential equation (2.1).
5.2 Solutions to the problems for study and analysis
5.2.1 Solution to problem 1
We start with the fact that the limiting value, C2, will be 1 and solve the logistics
differential equation (5.1).
First, we separate the variables:
dm∞m∞(1−m∞)
= (C1)dV.
Integrate and use inflection point coordinates (V1/2,12) and slope 1/2
2kto finish finding
the particular solution.∫(
1
m∞+
1
1−m∞)dm∞ =
∫C1dV,
lnm∞ − ln(1−m∞) = C1V + C3,
lnm∞
1−m∞= C1V + C3,
m∞1−m∞
= C4eC1V ,
m∞ =C4e
C1V
C4eC1V + 1,
m∞ =1
1 + C5e−C1V.
Using dm∞dV
= 1/22k
in the differential equation yields 1k
= C1.
Using (V,m∞) = (V1/2,12) in the general solution yields C5 = e
1kV1/2 . So, the
particular solution is :
m∞ =1
1 + e1kV1/2e−
1kV
or
m∞ =1
1 + e−1k
(V−V1/2).
37

Texas Tech University, Julie Bumbaugh, August, 2012
5.2.2 Solution to problem 2
The partial derivatives needed for the Jacobian matrix elements are as follows:
∂V
∂V=
1
C(−gL − gNa ·m∞(V )− gNa(V − ENa)(−1)(1 + e(V1/2−V )/k)−2·
(e(V1/2−V )/k)(−1
k)− gK · n),
∂V
∂n=− gK(V − EK),
∂n
∂V=
1
τV(1 + e(V1/2−V )/k)−2(e(V1/2−V )/k)(
1
k),
∂n
∂n=− 1
τV.
The corresponding values for n can be calculated using the values of V at each
equilibria in the equation (5.3). The coordinates of the equilibria are (−28, .3543),
(−56, .002025), and (−66, .00027458). Here are the Jacobian matrices for V = −28,
V = −56, and V = −66 respectively: A =
(8.40466 −620
.0457568 −1
),
A =
(2.1104 −340
.00040424 −1
), and A =
(−1.74898 −240
.0000549 −1
). We calculate the trace
and determinant of each matrix. For V = −28, Tr(A) = 7.40466 and
|A| = 19.964556. When V = −56, Tr(A) = 1.1104 and |A| = −1.9729584. When
V = −66, Tr(A) = −2.74898 and |A| = 1.762156. Finally we compute the
eigenvalues for each equilibrium usingTr(A)±
√(Tr(A))2−4det(A)
2. The eigenvalues for
V = −28 are 3.70233±√−25.029234
2. When V = −56 the eigenvalues are 2.065566 and
−.955165. The eigenvalues for V = −66 are −1.018026 and −1.7309543. These
results tell us that the equilibrium at V = −28 is not stable, the equilibrium at
V = −56 is a saddle since one eigenvalue is positive and the other is negative. The
equilibrium at V = −66 is indeed stable since both eigenvalues are negative.
5.2.3 Solution to problem 3
See a computer code in appendix B that locates the value of V at the equilibrium.
Figure 5.2 shows that the equilibrium is at a value of V = 22.357 to the nearest
thousandth. Now we can now determine the corresponding value of n using
38

Texas Tech University, Julie Bumbaugh, August, 2012
Figure 5.2. Location of intersection of nullclines is at the zero of difference betweennullclines for HH reduced model. The plot shows this to be nearest to V = 22.357
equation (4.7). This yields n = 0.2998158738. Next, we find the Jacobian matrix.
Another program can be found in appendix C that aids in finding the partial
derivatives of V and n. Jacobian matrix, A =(1628.6135 −1162.5718
.0052275 −.2687
).
The Tr(A) = 1628.3448 and the determinant |A| = −431.5311034. The eigenvalues
areTr(A)+
√(Tr(A))2−4det(A)
2= 1628.609769 and
Tr(A)−√
(Tr(A))2−4det(A)
2= 1.000614876.
Since both eigenvalues are positive, this equilibrium is unstable.
5.2.4 Solution to problem 4
We simply need to write a program to plot V vs. I for this model. See Appendix
D for possible programs. The following figure demonstrates the results of one such
program.
39

Texas Tech University, Julie Bumbaugh, August, 2012
Figure 5.3. Excitation block in the low-threshold INa,p + IK model.
5.2.5 Solution to problem 5
The particular solution 1− e−t to the differential equation,dndt
= αn(1− n)− βn · n does not have an inflection point since
d
dt(1− e−t) = e−t
andd2(1− e−t)
dt2= −e−t < 0.
The second derivative of this solution is always negative; hence, no inflection point.
The data collected showed a sigmoid shaped curve which has an inflection point. If
we raise this solution to any integral power p > 1, there will be an inflection point
at t = ln(p). Let y = (1− e−t)p. Then
dy
dt= p(1− e−t)p−1(e−t),
40

Texas Tech University, Julie Bumbaugh, August, 2012
d2y
dt2= p(p− 1)(1− e−t)p−2 · e−2t − p(1− e−t)p−1 · e−t,
= p(1− e−t)p−2 · e−2t((p− 1)− (1− e−t · et)),
= p(1− e−t)p−2 · e−2t(p− et),
= 0 only when p− et = 0.
p− et = 0 when t = ln p.
Due to the slow rise of the natural log function, we know that the greater the value
of p, the smaller the difference is between ln p and ln (p+ 1). By raising the value of
p, we move the inflection point further and further to the right in our curve, but it
moves smaller and smaller amounts to the right such that eventually, there is no
practical benefit of using a higher power.
ln 2 ≈ .693 ln 3 ≈ 1.0986 ln 4 ≈ 1.386 ln 5 ≈ 1.609 ln 6 ≈ 1.792
Choosing 3rd power over 2nd pushes the inflection point ≈ .4 units further to the
right. Choosing 4th power over 3nd moves it an additional ≈ .3 units. 5th power
would have moved it another ≈ .2 units and 6th power would have moved it another
≈ .18 units. It must have been the case that the 4th power placed the inflection
point within ≈ .2unit of where it needed to be such that using higher powers
became unnecessary. They did make comment in their paper that they were
choosing a power to compensate for the “delayed rise of potassium”. We assume
here that they are referring to the point at which the rate of change in the rise of
the curve changes, the inflection point.
5.2.6 Solution to problem 6
First, we separate the variables:
dn
αn − n(αn + betan)= dt.
Next we integrate:
− 1
αn + βnln |αn − n(αn + βn)| = t+ C.
41

Texas Tech University, Julie Bumbaugh, August, 2012
Substitute for their definition of τn and continue solving for n:
−τn · ln |αn − n(αn + βn)| = t+ C,
ln |αn − n(αn + βn)| = − 1
τn· t+− 1
τn· C.
Apply the initial condition that when t = 0, n = n0 :
ln |αn − n0(αn + βn)| = − 1
τn· C,
−τn · ln |αn − n0(αn + βn)| = C.
Now we substitute this value of C back into the general solution:
ln |αn − n(αn + βn)| = −tτn− 1
τn· −τn · ln |αn − n0(αn + βn)|,
=−tτn
+ ln |αn − n0(αn + βn)|,
αn − n(αn + βn) = e−tτn · eln |αn−n0(αn+βn)|,
αn − n(αn + βn) = e−tτn · (αn − n0(αn + βn)).
Now, finish solving for n and make substitutions for n∞ = αnαn+βn
:
n · (αn + βn) = αn − (αn − n0(αn + βn)) · e−tτn ,
n =αn
(αn + βn)−(
αn(αn + βn)
− n0
)· e−
tτn ,
n = n∞ − (n∞ − n0) · e−tτn .
42

Texas Tech University, Julie Bumbaugh, August, 2012
BIBLIOGRAPHY
[1] D. E. Goldman,Potential, Impedence, and Rectification in Membranes, J. Gen.Physiology, 26(1943), pp. 37-60.
[2] A. L. Hodgkin and A. F. Huxley, A Quantitative Description of MembraneCurrent and its Application to Conduction and Excitation in Nerve, J.Physiology, 117(1952), pp. 500–544.
[3] A. L. Hodgkin, A. F. Huxley, and B. Katz, Measurement of Current-VoltageRelations in the Membrane of the Giant Axon of Loligo, J. Physiology,116(1952), pp. 424-448.
[4] A. L. Hodgkin and A. F. Huxley, Currents Carried by Sodium and PotassiumIons through the Membrane of the Giant Axon of Loligo, J. Physiology,116(1952), pp. 449-472.
[5] A. L. Hodgkin and A. F. Huxley,The Components of Membrane Conductancein the Giant Axon of Loligo, J. Physiology, 116(1952), pp. 473-496.
[6] A. L. Hodgkin and A. F. Huxley,The Dual Effect of Membrane Potential onSodium Conductance in the Giant Axon of Loligo, J. Physiology, 116(1952),pp. 497-506.
[7] E. M. Izhikevich, Dynamical Systems in Neuroscience: The Geometry ofExcitability and Bursting, Cambridge, Massachusetts, 2007.
[8] J. D. Murray, Mathematical Biology, Springer-Verlag, Berlin, Heidelberg, 1993.
43

Texas Tech University, Julie Bumbaugh, August, 2012
APPENDIX A
COMPUTER CODE FOR HODGKIN-HUXLEY MODEL
This matlab program uses Euler’s method to plot Action Potentials in “Voltage
Clamp” experiments:
1. % This program plots Voltage vs. Time of an action potential
%of a neuron.
2. % Input necessary constant values.
3. % This program should be run multiple times using different
% initial values for membrane potential.
4. Vna = -115;
5. Vk = 12;
6. Vl = -10.613;
7. gna = 120;
8. gk = 36;
9. gl = 0.3;
10. V=20;
11. %Calculate needed initial values from the constants.
12. an = 0.01*(V+10)/(exp(1+0.1*V)-1);
13. bn = 0.125*exp(V/80.0);
14. am = 0.1*(V+25)/(exp(0.1*V+2.5)-1);
15. bm = 4.0*exp(V/18.0);
16. ah = 0.07*exp(V/20.0);
17. bh = 1.0/(exp(3+0.1*V)+1);
18. % Initialize values for n,m,h, & V.
19. n = an/(an + bn);
20. m=am/(am + bm);
21. h = ah/(ah + bh);
22. V(1) = V;
23. t(1) = 0;
24. nn(1) = n;
25. mm(1) = m;
44

Texas Tech University, Julie Bumbaugh, August, 2012
26. hh(1) = h;
27. ann(1) = an;
28. bnn(1) = bn;
29. ninf(1) = an/(an + bn);
30. %Use Euler’s method to get the next 1000 values of the
%variables using a step size of 0.01.
31. for i = 1:1000
32. an = 0.01*(V(i)+10)/(exp(1+0.1*V(i))-1);
33. bn = 0.125*exp(V(i)/90.0);
34. am = 0.1*(v(i)+25)/(exp(0.1*V(i)+2.5)-1);
35. bm = 4.0*exp(V(i)/18.0);
36. ah = 0.07*exp(V(i)/20.0);
37. bh = 1.0/(exp(3+0.1*V(i))+1);
38. dmdt = am*(1-m)-bm*m;
39. dndt = an*(1-n)-bn*n;
40. dhdt = ah*(1-h)-bh*h;
41. m = m + dmdt&0.01;
42. n = n+dndt*0.01;
43. h = h+dhdt*0.01;
44. dvdt = -gk*n^4*(V(i)-Vk)-gna*m^3*h*(V(i)-Vna)-gl*(V(i)-Vl);
45. V(i+1) = (V(i) + dvdt*0.01);
46. t(i+1) = (i)*0.01;
47. nn(i+1) = n;
48. mm(i+1) = m;
49. hh(i+1) = h;
50. ann(i+1) = an;
51. bnn(i+1) = bn;
52. ninf(i+1) = an/(an+bn);
53. end
54. % One could now plot alphas and betas versus V or
%could plot Membrane potential, Versus Time, t.
55. plot (t,-V)
45

Texas Tech University, Julie Bumbaugh, August, 2012
APPENDIX B
COMPUTER CODE FOR FINDING INTERSECTION OF NULLCLINES
This matlab program finds intersections of nullclines by locating the zero of their
difference.
%This program will plot differences in the nullclines for n and V
%of the reduced HH model to help locate the equilibrium for
%given values of injected current I.
1. I=12;
2. gna=120;
3. Ena=120;
4. gl=.3;
5. El=10.6;
6. gk=36;
7. Ek=-12;
8. h=.00001;
9. N=1000;
10. V=22.35;
11. for i=1:N
12. v(i)= V;
13. a(i)=.1*(25-v(i))/(exp((25-v(i))/10)-1);
14. b(i)= 4*exp(-1*v(i)/18);
15. minf(i)=a(i)/(a(i)+b(i));
16. Vnull(i)=((I-gna*(minf(i))^3*(v-Ena)-gl*(v-El))/
(gk*(v-Ek)))^(.25);
17. nnull(i)=.01*(10-v)/(exp((10-v)/10)-1)/(.01*(10-v)/
((exp((10-v)/10)-1)+1/8*exp(-v/80)));
18. D(i)=Vnull(i)-nnull(i);
19. V = V+h;
20. end
21. plot(v,D,’r’)
22. hold on
23. plot(v,0)
24. xlabel(’V (mv)’)
46

Texas Tech University, Julie Bumbaugh, August, 2012
25. ylabel(’Difference in nullclines’)
26. title(’Locating zero for difference in nullclines of reduced HH’)
27. axis([22.35 22.36 -.001 .001])
47

Texas Tech University, Julie Bumbaugh, August, 2012
APPENDIX C
COMPUTER CODE FOR CALCULATING PARTIAL DERIVATIVES AT
EQUILIBRIUM
This is a matlab program for calculating partial derivatives of dv/dt and dn/dt atthe equilibrium in Hodgkin-Huxley reduced model.
%Calculates partial of dv/dt of hhreduced model with respect to v.
1. I=12;
2. C=1;
3. gna=120;
4. Ena=120;
5. gl=.3;
6. El=10.6;
7. gk=36;
8. Ek=-12;
9. v=22.357;
10. n=.2998158738;
11. m1=(.1*(25-v)); %numerator of alpha_m
12. m2=(exp((25-v)/10)-1); %denominator of alpha_m
13. mm=m1/m2; %alpha_m
14. dmm=(m2*-.1-m1*-.1*exp((25-v)/10))/m2^2; %deriv of alpha_m
%with respect to v
15. d=mm+4*exp(-v/18); %alpha_m + beta_m
16. dd=dmm+4*exp(-v/18)*-1/18; %derivative of
%alpha_m +beta_m with respect to v
17. minf=mm/d; %m_infty(V)
18. n1=.01*(10-v); %numerator of alpha_n
19. n2=exp(.1*(10-v))-1; %denominator of alpha_n
20. nn=n1/n2; %alpha_n
21. e=nn+.125*exp(-v/80); %alpha_n + beta_n
22. ninf=nn/e; %n_infty(V)
23. dnn=(n2*-.01-n1*exp(.1*(10-v))*-.1)/n2^2; %derivative of
%alpha_n with respect to v
24. de=dnn+.125*exp(-v/80)*-1/80; %derivative of
%alpha_n +beta_n with respect to v
25. dninf=(e*dnn-nn*de)/e^2; %derivative of n_infty with
%respect to v
48

Texas Tech University, Julie Bumbaugh, August, 2012
26. dminf=(d*dmm-mm*dd)*d^2; %derivative of m_infty(V)
% with respect to v
%dminf is derivative of m_infty(V) with respect to V
27. w=1/C*(-gk*n^4-gna*minf^3*(.89-1.1*n)-gna*(v-Ena)
*(.89-1.1*n)*3*minf^2*dminf-gl);
%w is partial of dv/dt with respect to V
28. x=1/C*(-gk*(v-Ek)*4*n^3-gna*minf^3*(-1.1)*(v-Ena))
%x is partial of dv/dt with respect to n.
29. y=(1-n)*dnn-n*.125*exp(-v/80)*-1/80;
%y is partial of dn/dt with respect to v
30. z=-nn-.125*exp(-v/80); %partial of dn/dt with respect to n
31. h=.1
32. a=1
33. for i=1:10
34. q(i)=a
35. L(i)=z
36. a=a+h
37. end
38. plot(q,L)
39. axis([1 2 -.269 -.2685])
%Calculates the partial of dn/dt with respect to n.
49

Texas Tech University, Julie Bumbaugh, August, 2012
APPENDIX D
COMPUTER CODE TO PLOT EXAMPLES OF EXCITATION BLOCK
The following program(s) will allow one to plot geometric proof of excitation block
when currents are ramped up in some models.
%This program is to plot low-threshold persistent sodium
%+potassium model’s potential vs. time graph with increasing
% injected current.
1. [t,y]=ode45(’my_lowthresholdtrajectoryfn’,[0 10000],[-75 .2 40]);
2. plot(t,y(:,1),’b’)
3. xlabel(’Time, t(ms)’)
4. ylabel(’Membrane potential, V(mv)’)
5. title(’V vs. t in I_{Na,p}+I_K low threshold model with
increasing values of I’)
6. axis([0 100 -75 0])
%% This program is to be used with the previous one.
% function dydt=my_highthresholdtrajectoryfn(t,y)
1. dydt=zeros(size(y));
2. C=1; I=0; El=-78; gl=8; gNa=20; gk=10; mV=-20; mk=15;
nV=-45; nk=5; Tau=1;
3. Ena=60; Ek=-90;
4. A=y(1)
5. B=y(2)
6. D=y(3)
50

Texas Tech University, Julie Bumbaugh, August, 2012
7. dydt(1)=(1/C)*(D-gl*(A-El)-gNa*(1/(1+exp((mV-A)/mk)))*(A-Ena)-
gk*B*(A-Ek));
8. dydt(2)=(1/(1+exp((nV-A)/nk))-B)/Tau;
9. dydt(3)=3.6;
%%The next two programs go together to plot traces of n vs. V of
% the same model with fixed values of I.
%This program is to plot Traces of low-threshold
%persistent sodium +potassium model.
%Plots n vs. V graphs with different values for I.
1. [t,y]=ode45(’my_lowthresholdtrajectoryfn2’,[0 500],[-60 .1]);
2. h = 1
3. g = -89
4. for i=1:109
5. k(i)= g
6. j(i)= (300-8*(g+78)-20*
(1/(1+exp((-20-g)/15)))*(g-60))/(10*(g+90))
7. l(i) = 1/(1+exp((-45-g)/5))
8. g = g+h
9. end
10. plot(y(:,1),y(:,2),’b’)
11. hold on
12. plot(k,j,’r’)
13. hold on
14. plot(k,l,’g’)
15. hold off
16. xlabel(’Membrane potential, V(mv)’)
17. ylabel(’K+ activation variable, n’)
18. title(’n vs. V in I_{Na,p}+I_K low threshold model with I=0’)
19. axis([-89 20 0 1.2])
%% to be used with the program above.
1. function dydt=my_highthresholdtrajectoryfn(t,y)
2. dydt=zeros(size(y));
51

Texas Tech University, Julie Bumbaugh, August, 2012
3. C=1; I=300; El=-78; gl=8; gNa=20; gk=10; mV=-20; mk=15; nV=-45;
4. nk=5; Tau=1;
5.Ena=60; Ek=-90;
6. A=y(1)
7. B=y(2)
8. dydt(1)=(1/C)*(I-gl*(A-El)-gNa*(1/(1+exp((mV-A)/mk)))*(A-Ena)-
gk*B*(A-Ek));
9. dydt(2)=(1/(1+exp((nV-A)/nk))-B)/Tau;
52