blerta [versidad autonoma metropolitana-iztapalapa148.206.53.84/tesiuami/uam3264.pdf ·...
TRANSCRIPT

blerta al tempo
[VERSIDAD AUTONOMA METROPOLITANA-IZTAPALAPA ARTAMENTO DE CIENCIAS DE LA SALUD
IJNIVERSIDAD AUTONOMA METROPOLITANA.
UNIDAD IZTAPALAPA.
CIENCIAS BIOLOGICAS Y DE LA SALUD.
LIC. EN BIOLOGIA EXPERIMENTAL
LEON KEMPIS MA. DEL ROCIO.
92330977
. I 1,. ' . I I 1. , , -;, . , . / . , , 1 . I
MONOCLONALES DIRIGIDOS CONTRA MOLECULAS DE
SUPERFICIE DE Snlmonella tyhpi "
ASESOR INT HECTOR F. SERRANO P
DEpF) . CaNC'IAS DE LA SALUD.
/77A
Bioquímica y Biología Molecular de la Fertilización 24 4733 1 [email protected] 124 4727

LEON KEMPIS MA. DEL ROCIO. MATRICULA: 92330977 LICENCIATURA EN BlOLOGlA EXPERIMENTAL.
"OBTENCI(5N DE HlBRlDOMAS PRODUCTORES DE ANTICUERPOS MONOCLONALES DIRIGIDOS CONTRA MOLÉCULAS DE SUPERFICIE DE Salmonella typhi"
Clave BEOQl.97 28 de Enero de 1998 ..
En el trabajo de investigación desarrollado se logró la fusión celular de c4lulas esplénicas inmunes con células murinas de mieloma, con el objeto de lograr un híbrido productor de anticuerpos dirigidos contra uno de los determinantes antigénicos expresados por Salmonella typhi, el antígeno Vi. Este antigen0 es de particular importancia ya que se ha visto que está estrechamente relacionado con la virulencia y patogenicidad de la bacteria, de modo que en los pacientes de fiebre tifoidea se han detectado anticuerpos contra este antígeno y además puede encontrársele en el medio en forma soluble. Después de ensayos de fusión se logró obtener 18 hibridomas productores de anticuerpos anti-Vi (detectados por ELISA); estos anticuerpos son de diveroso isotipos, predominando los isotipos IgM, IgG2a, lgG2b e lgG3. Del hibridoma productor de anticuerpos isotipo lgG3 se obtuvo sobrenadante de cultivo por agotamiento (concentrando la cantidad de -anticuerpos presentes en el medio) y se purificó por cromatografia de afinidad lograncjo obtener hasta 1 .I26 mg/ml de anticuerpo. De este eluído, se corrieron por SDS-PAGE 4, 6 y 8 pg de proteína tanto en condiciones reductoras como en condiciones no reductoras y se demostró fa presencia de moléculas de inmunoglobulina completas y la desnaturalización de éstas con S-mercaptoetanol en cadenas pesadas y en cadenas ligeras.
El desarrollo de anticuerpos monoclonales contra moléculas antigénicas permitirá diseñar una metodología apropiada para la detección del antígeno en diferentes fluidos biológicos, pudiéndose emplear para identificar portadores de S. fyphi mediante el uso de pruebas tamiz.

ABREVIATURAS.
Al F
F RPMI-HMT i .\ -, ” ‘C ELEA , 5 I SFB
HGPRT ;-r
Medio HMT
PBS
PEG
PH
T
DMSO
OPD
SDS-PAGE
Adyuvante incompleto de Freund
Medio RPMl suplementado con hipoxantina, metrotexato y timidina
Ensayo inmunoenzimático en fase sólida
Suero fetal bovino
Enzima Hipoxantina-guanina-fosforibosil transferasa
Medio de cultivo selectivo suplementado con hipoxantina, metrotexato y timidina
Solución salina amortiguada con fosfatos
Polietilenglicol
Potencial de Hidrógeno
Acido sulfúrico
Temperatura
Línea celular de mieloma murino
Dimetilsulfóxido
Ortofeniléndiamina
Electroforesis en gel de poliacrilamida

INDICE .
INTRODUCCION ............................................................................. 1
FIEBRE TIFOIDEA ................................................................ 1 ANTIGEN0 Vi ....................................................................... 2 ANTIGEN0 Vi Y DIAGNOSTICO DE FIEBRE TIFOIDEA .... 3 ANTICUERPOS MONOCLONALES ..................................... 3 OBTENCION DE ANTICUERPOS MONOCLONALES ......... 4
OBJETIVOS ..................................................................................... 6
METODOLOGIA ............................................................................... 7
METODOS BACTERIOLOGICOS ............................... 7 ESQUEMA DE INMUNIZACION .................................. 7 FUSION SOMATICA .................................................... 7 SELECCION POR €LISA ............................................. 9
CRIOPRESERVACION ................................................. 9
ISOTIPIFICACION ......................................................... 9 PRODUCCION MASIVA Y PURIFICACION .................. 9
1 .- PRODUCCION
2.- ALMACENAMIENTO .
3.- CARACTERIZACION Y PURIFICACION .
CARACTERIZACION POR SDS-PAGE ........................ 10
RESULTADOS ................................................................................... 1 1
CONCLUSIONES ............................................................................... 15
BIBLIOGRAFIA ................................................................................... 16

La fiebre tifoidea es una enfermedad febril aguda, causada por el bacilo Salmonella typhi, que afecta exclusivamente a los humanos y se caracteriza por fiebre, dolor de cabeza, apatia,tos, esplenomegalia, rash y leucopenia. El curso de la enfermedad es generalmente severo y prolongado, puede durar varias semanas si no es tratada (1 1 , 12).
Salmonella typhi es un bacilo citofílico, gram negativo, aerobio, no formador de espora;, móvil y mide de 2-3 por 0.6 micfómetros. Fermenta glucosa, pero no lactosa ni sacarosa. Su clasificación serológica se basa en los principales determinantes antigénicos presentes, el antígeno somático O y flagelar H; el antígeno O está presente en el lipopolisacárido (LPS) que consiste de un lípido A, un núcleo interno de oligosacárido y largas cadenas laterales de polisacárido. El lipido A o endotoxina constituye la principal porción en la lámina exterior de la membrana externa de organismos gram negativos, consiste de un disacárido de glucosamina unido por un puente de pirofosfato en el que los grupos amino e hidroxilo son Sustituidos por una cadena larga de ácidos grasos que es el principal determinante de toxicidad. El núcleo interno de oligosacárido es similar en todos los serotipos, mientras que las cadenas laterales O consisten de unidades repetitivas de cuatro o cinco residuos, algunos son determinados por conversión lisogénica, los cuales varían en los diferentes grupos y determinan la especificidad. El antígeno H muestra también diversidad en sus determinantes, las variaciones en la composición antigénica del flagelo de organismos con antígenos O comunes determinan los diferentes serotipos (7).
Epidemioloaia: la fuente de infección con Salmonella typhi es, generalmente, un paciente con fiebre tifoidea o un acarreador de bacilos, pues éstos excretan grandes cantidades de bacilos en heces (a menudo se excretan I O 6 o más bacilos viables por gramo de heces) o en orina. El bacilo viable puede estar presente también en vómito, secreciones respiratorias y pus (12).
Patoaenia: después de la ingestión de un inóculo adecuado, el bacilo pasa rápidamente a través de la mucosa intestinal hacia las regiones linfáticas, en donde es fagocitado. Algunos bacilos pueden escapar al englobamiento en los ganglios linfáticos regionales, pasando al torrente sanguíneo a través de la linfa y provocando una bacteremia temporal que es rápidamente abatida por remoción de dlulas retículoendoteliales en hígado, bazo, médula ósea y nódulos linfáticos. Salmonella typhi accesa a sitios intracelulares, en donde se multiplica. Cuando el número de bacterias intracelulares sobrepasa un punto o umbral crítico, se produce una bacteremia secundaria, que invade la vesícula biliar y las placas de Peyer, las bacterias vuelven a entrar en la luz intestinal y se recuperan en los coprocultivos a partir de la segunda semana de enfermedad clínica (1 1 , 12).
1

Diaanóstico: la reacción de Widal consiste de la aglutinación del antígeno somático y flagelar, el pico de aglutininas se dá en la tercera semana de la enfermedad. La interpretación de las pruebas de aglutinación es difícil debido a la reactividad cruzada con otras bacterias entéricas y por fa persistencia de altos títulos de aglutininas (por meses y hasta años) en sujetos inmunizados (5).
El diagnóstico definitivo se dá por aislamiento del agente causal de sangre, heces , orina u ocasionalmente de esputo o de exudados purulentos. Un incremento 4 veces mayor que un control normal del título de aglutininas, especialmente contra el antigeno O, sin inmunización reciente, confirma el diagnóstico (5,12).
ANTiGENO Vi.
La envoltura capsular bacteriana juega un papel importante en la patogenia de la bacteria, porque la protege de la fagocitosis y además algunos de sus componentes pueden funcionar como adhesinas. En algunas bacterias gram negativas, la cápsula consiste de polisacáridos relativamente simples con unidades repetitivas de 2 o 3 azúcares (8). El polisacárido capsular Vi se encuentra en varios miembros de la familia Enterobacteriaceae, incluyendo Salmonella fyphi, Salmonella parafyphi C, Salmonella dublin y Cifrobacter freundii (6, 7, 20 ,27). La presencia de este antígeno interfiere con la actividad bactericida del suero e inhibe la fagocitosis, por lo cual la cepas de S. fyphi Vi' son más virulentas que las Vi- (1 3,20). Daniels y cols. determinaron que Citrobacfer freundii es la bacteria que sintetiza el antígeno Vi en mayor cuantía y con mejor estabilidad, después de observar que varias cepas de Salmonella paratyphi C y Salmonella typhi dejaron de producir antígeno Vi después de 2 o 3 pases en cultivo (6).
El antígeno capsular Vi es un homopolímero lineal de ácido alfa-l,4-2-N- acetilgalacturónico variablemente O-acetilado en el carbono 3. La difracción por rayos X indica que esta molécula asume la forma característica de hélices con dos o tres pliegues simétricos, siendo 0.43 nm la distancia lineal entre los grupos carboxilo. Una característica importante de esta molécula es el papel inmunogénico que juegan los grupos O-acetilos y carboxilos (con una consecuente variación en el peso molecular). Una completa O-desacetilación resulta en una baja antigenicidad de Vi, mientras que una parcial O-desacetilación aumenta ligeramente su inmunogenicidad. La reducción de los carboxilos resulta en una reducción de la inmunogenicidad de Vi (20,22).
Los determinantes genéticos involucrados en la expresión del antigeno Vi ocupan loci cromosomales separados y se denominan v i d y viaB. El locus viaA se encuentra a 43 minutos del cromosoma de Salmonella fyphi, pero también se presenta en E. coli, en C. freundi y en cepas de Salmonella que expresan antígeno Vi. El locus via6 se localiza a 92 minutos del cromosoma de Salmonella fyphi sólo en bacterias que expresan el antígeno Vi (29). En un ensayo de detección de la
2

Antígeno Vi y diagnóstico de fiebre tifoidea.
Ya que el antigen0 Vi se Da encontrado en la mayoría de cepas de S. fyphi aisladas de la sangre de pacienta con fiebre tifoidea y que además se encontraron títulos elevados de anticuerpos anti-Vi en el suero de estos pacientes, se propuso que la presencia de anticuerpo$ anti-vi estaría relacionada con la infección. En 1953 se realizó un ensayo de hmaglutinación pasiva con el antígeno purificado y se encontró un elevado título de anticuerpos en pacientes con fiebre tifoidea reciente o activa, no hallándolo en personas sanas (20). Se ha intentado la detección del bacilo en distinta fluidos biológicos haciendo modificaciones a diferentes ensayos serológicos tanto en zonas endémicas de fiebre tifoidea como en zonas donde su incidencia es baja (21 , 27, 28). En Chile, se realizó un ensayo de hemaglutinación pasiva que tuvo un 75% de sensibilidad pero careció de especificidad (I 4). Otro experimento, consistente en unir anticuerpos dirigidos contra los antígenos Vi, D y d 8 proteína A de Sfaphylococo y a enfrentarlas con muestras de orina provenientes de sujetos sanos y de pacientes, se obtiene, nuevamente, un alto porcentaje de sensibilidad, pero muy baja especificidad; estos resultados logran mejorar en ensayos de ELISA, que son capaces de detectar hasta 1 ng de antígeno Vi/ml, sin embargo, la desventaja de usar este antisuero (suero de conejo anti-Salmonella) sobrevino después de mostrar reactividad cruzada, pues se detectaron Salmonellas de otras especies (se obtuvo un 17% de falsos positivos) (2). Qadri y cols. desarrollaron anticuerpos monoclonales contra los antígenos O, H y Vi de S. typhi. Los anticuerpos anti-Vi reaccionaron a dos determinantes discretos, uno constituido por el grupo O-acetilo y el otro por los grupos N-acetilo y carboxilo. Estos anticuerpos se usaron en una prueba de ELEA de doble anticuerpo, para la detección de la bacteria en medio de cultivo inoculado y en orina, pudiendo detectar 0.001 mícrogramos de Vil ml y 6 x 10' bacterias/ml (1 9).
La detección de acarreadores crónicos de S. typhi no es fácil, el método tradicional que ha sido la coleccibn de una serie de cultivos diarios de heces, es costoso y sólo intermitentemente positivo debido a que S. fyphi es inhibida por la colonización de la flora normal (6). Por lo mencionado anteriormente, el empleo de anticuerpos monoclonales en el diagnóstico de fiebre tifoidea puede resultar en un buen sistema de detección oportuna de la presencia del antígeno Vi en fluidos fisiológicos de pacientes y acarreadores asintomáticos, quienes constituyen uno de los mecanismos más importantes de diseminación del bacilo, ya que un 50% de las personas infectadas continúan excretando Salmonella un mes después que los síntomas han desaparecido y 1 de cada 20 pacientes lo sigue haciendo hasta 5
I
3

meses despues (7). Por otra parte, el uso de anticuerpos monoclonales en la detecci6n de antígenos propios de algunas enfermedades ya está bien implementado en algunas patologías parasitarias como la malaria y la toxoplasmosis (1 8).
Anticuerpos monoclonales.
Cuqndo se está en contacto con un inmunógeno, el organismo es capaz de montar una respuesta cuyo efecto se dirige a la eliminación específica del inmunógeno. Esta respuesta inmune en un animal es de tipo policlonal, caracterizada por la producción de muchos anticuerpos con diferente afinidad y especificidad para el inmunógeno. La respuesta policlonal resulta de la activación de varias clonas de linfocitos B presentes en el organismo (1,4). Debido a que cada clona sintetiza un tipo único de anticuerpo específico para el inmunógeno, lo deseable es obtener híbridos de linfocitos 8 con c4lulas de mieloma que sintetizen anticuerpos homogéneos con alta especificidad y afinidad por el inmunógeno y que además, posean una vida mayor a las de las células B comunes. Este problema fué resuelto en 1975 por Ktihler y Milstein, quienes desarrollaron una técnica para producir una población homogénea de células secretoras de anticuerpos específicos (1,3,4,9, 13).
Obtención de anticuerpos monoclonales.
El fundamento de esta técnica es fusionar una célula plasmática (proveniente de un animal inmunizado con el antigen0 de interés y por ende, productora de un anticuerpo) con una célula neoplásica, de manera que se obtenga un híbrido cuyas características sean: sintetizar un anticuerpo específico y capacidad de dividirse indefinidamente. Es importante señalar que estas dos células progenitoras deben ser de líneas similares y aproximadamente en el mismo estadio de diferenciación para que el híbrido resultante retenga las características funcionales de las células parentales (3,9).
Ahora bien, las células de mieloma empleadas en este proceso de fusión tienen una deficiencia enzimática en hipoxantinaguanina fosforibosil transferasa (HGPRT), lo cual les impide utilizar timidina o hipoxantina del medio, ambas útiles en la síntesis de purinas. Esta deficiencia enzimática les impide crecer en un medio de selección como lo puede ser el medio HMT (hipoxantina-metrotexato-timidina). El metrotexato inhibe la actividad de la tetrahidrofolato reductasa, afectando directamente la síntesis de purinas, moléculas indispensables en la sintesis de DNA. Es un bloqueador de la síntesis de DNA por la vía de Novo. Las células tratadas con metrotexato pueden usar la vía de salvamento en la que la purina es
4

sintetizada a partir de hipoxantina suministrada exdgenamente, pero haciendo USO
de la enzima hipoxantinaguanina fosforibosil transferasa y el timilidato por timidina, empleando la enzima timidina cinasa (3,9). Una de las líneas celulares usadas es la P3-X63-Ag8, que no secreta cadenas pesadas ni ligeras de inmunoglobulinas y que desciende del mieloma de r a t h MOPC-21
Una vez fusionadas las células de mieloma con las células de bazo, se tendran tres poblaciones celulares:
a) bazo-bqzo, son células productoras de anticuerpo pero no inmortales. b) mieloma-bazo, híbridos productores de anticuerpos e inmortales. c) mieloma-mieloma, células inmortales pero no productoras de anticuerpos.
La selecci6n del hibridoma se realiza cultivando estas células en medio HMT, donde sólo sobrevivirán las poblaciones bazo-bazo y los mieloma-bazo. La población mieloma-mieloma muere por la deficiencia enzimática y las células bazo- bazo mueren en 5 días aproximadamente. Los hibridos mieloma-bazo sobreviven a las condiciones selectivas porque la deficiencia enzirnática de la célula neoplásica es compensada por el esplenocito (3,9).
Ya cultivados estos hibridomas, se colecta el sobrenadante y se analiza por ELISA, con el objeto de determinar la producción de anticuerpos; ya identificados los hibridomas productores del anticuerpo deseado, se les somete a clonaciones consecutivas para asegurar que el anticuerpo final sea monoclonal. Este hibridoma será probado nuevamente por ELISA; las clonas que arrojen resultados positivos se aislan y se cultivan en grandes volúmenes de medio de cultivo o se inyectan en cavidad intraperitoneal de ratones para producir líquido ascítico y obtener anticuerpos en grandes cantidades (3,9).
5

Objetivos: .
General: obtener hibridomas productores de anticuerpos monoclonales dirigidos contra ' . moléculas de la superficie de Salmonella typhi. . .
Particular: seleccionar aquellos hibridomas que sintetizen anticuerpos dirigidos contra : . '
- .
el antigen0 capsular Vi de Salmonella typhi.
6

Metodología utilizada:
1 .- Métodos bacteriológicos.
Citrobacter freundii , donada por el Instituto Nacional de Diagnóstico y Referencia Epidemiológica de la S.S.A. se sembró por estría sobre una placa de agar BHI , a las 24 horas de incubación a 37OC, se aisló una de las colonias crecidas y se sembró masivamente sobre otra placa de agar BHI. Después de 24 horas de incubación a 37 O C , se cosechó la bacteria y se resuspendió en SS1 estéril, esta suspensión se ajustó a una densidad bptica de 0.6 a 540 nm de longitud de onda. Se inactivaron por calor (incubando una hora a 56 O C ) las bacterias cosechadas.
Se realizó una mezcla 1 : I de la suspensión anterior con AIF y se homogenizó hasta lograr una emulsión.
2.- Esquema de inmunización.
Se inocularon vía intraperitoneal a los dias O, 15 y 30 cinco ratones BALB/c hembras con 100 pl de la mezcla anterior, lo cual equivale a 25 x IO6 bacterias. Se analizó el suero de los ratones mediante un ensayo inmunoenzimático (ELISA), comparando estos resultados con suero preinmune, con el objeto de determinar el título de anticuerpos generados contra el antígeno capsular Vi. Se eligió el ratón de mayor título y se inoculó con 10 pg de antígeno Vi (donado por el Dr. J. /B. Robbins) tres días antes de la fusión.
3.- Fusión somática.
- Obtención de macrófagos murinos de cavidad peritoneal.
A ratones BALB/c estimulados 8 días antes con 0.5 ml de AIF, se les sacrificó por dislocación cervical y se fijaron boca arriba a una tablilla. Se abrió la piel para exponer la pared abdominal y se les inyectaron vía ¡p. 5 ml de RPMl frío suplementado con 20 % de suero fetal bovino, glutamina 2 mM, 2- mercaptoetanol 50 pM , estreptomicina 100 pg/ml y penicilina 100 U.l./ml, se les dió un ligero masaje abdominal durante 5 minutos, tiempo después del cual se les extrajo el líquido introducido, se determina el número de células totales (conteo celular en cámara de Neubauer) y se centrifugó a 1200 rpm/ 5 min. La pastilla fue resuspendida en medio RPMI-20% y se dispensaron de 25000 a 30000 células por pozo en placas de cultivo celular de 96 pozos (Corning- Costar) y se incubaron a 37 O C , con atmósfera húmeda y 5% de Con .
I
7

- Línea celular de mieloma. Se empleó la línea celular murina P3X63Ag8, ATCC TIB 9 (deficiente en
HGPRT). Las células se mantuvieron en crecimiento hasta alcanzar la fase de crecimiento Jogaritrnico y se cosecharon por centrifugación a 1200 rpm/ 5 min para su uso en la fusión. Se lavaron 2 veces con medio RPMI sin suero fetal bovino.
- Obtención de células esplénicas inmunes. Tres días después de la última inmunización, se extrajo en condiciones
de esterilidad, el bazo del ratón y se depositó en una caja petri que contenía tela de organza y 5 rnl de RPMI-20%. Con ayuda de 2 jeringas con agujas se procedió a disgregar finamente el bazo, la suspensibn obtenida se depositó en un tubo de centrífuga cónico (Nunc Co.) y se centrifugó a 1200 rpml5 min. El botón celular obtenido se resuspendió en 5 ml de RPMl más cuatro ml de buffer de lisis para eritrocitos y se incubó a 37 O C durante 5 min, tiempo después del cual se volvió a centrifugar en las mismas condiciones y el sedimento se resuspendió en 20 ml de RPMI. Se determinó el número de células y su viabilidad en una cámara de Neubauer, mezclando 180 pl de azul tripán al 0.2 % con 20 pl de la suspensión celular.
- Purificación de células B Se transfirieron los 20 ml de la suspensión esplenocítica a una placa petri
que fue previamente recubierta con 20 ml de un anticuerpo monoclonal anti CD3 a una concentración de 2 pg/mI, (incubada 1 hora a T ambiente en la campana de flujo laminar). Se incubó la suspensión esplenocítica 1 hora a 37 O C y posteriormente se cosechó en un tubo cónico de 50 ml, se centrifugó a 1200 rpm/5 min y se determinó viabilidad y número total de células. Se lavó 2 veces con RPMl sin SFB.
- Fusión . En un tubo de centrífuga cónico de 50 ml se mezclaron 29.5 x I O 6
células de bazo con la misma cantidad de células de mieloma (proporción 1 :I ), después se centrifugaron a 1200 rpm/5 min y la pastilla celular se resuspendió en el medio que escurría por las paredes del tubo. Se agregó muy lentamente 1 m1 de polietilenglicol 1500 (Boehringer Manheim). Después de dejar reposar la mezcla durante 1 minuto a 37 O C se añadieron 8 ml de RPMl sin SFB, gota a gota y se dejó reposar 5 min a 37 "C. Posteriormente, se adicionaron 40 ml de medio, y después de 5 minutos se centrifugó a 800 rpm / 5 min. El sedimento se resuspendió en RPMI-20% y se depositaron 200,000 células por pozo en placas de cultivo que 48 horas antes habían sido recubiertas con macrófagos murinos de cavidad peritoneal, A las 24 horas se añadió un volumen igual de RPMI-HMT (suplementado con hipoxantina, metrotexato y timidina). Los cultivos fueron alimentados diariamente, retirándoles 100 pI de medio y atiadiéndoles el mismo volumen de medio fresco. De igual modo , a los 14 días de cultivo el medio HMT fue sustituido por Medio HT (RPMI suplementado eon hipoxantina y
8

tirnidina) en el que se mantuvieron las células hasta el día 20 despuks de la fusión. Las colonias de hibridomas visibles microscópicamente fueron alimentadas y expandidas de modo que no murieran por sobrecrecimiento ni acidificación del medio de cultivo. 2Cypi26 4.- Selección de hibridomas secretores de anticuerpos monoclonales anti-Vi y expansión in vitro
I r
Una vez que los hibridomas cubrieron la tercera parte del pozo de cultivo se les col6ctó el sobrenadante (después de 72 horas de haber cambiado sólo 50 pl de medio de cultivo) para cuantificar la presencia de anticuerpos específicos por el método de ELISA. Los hibridomas productores fueron expandidos a botellas de cultivo celular de 25 cm2 (Nunc Co.)
5.- Almacenamiento de células en viales de criopreservación.
Con el propósito de conservar a los hibridomas obtenidos, se congelaron 4.5 x 1 O6 células en 1.5 ml de solución crioprotectora ( 9 partes de SFB + 1 parte de DMSO). Las células se colocaron en un criotubo de plástico y se guardaron a - 70 OC durante 24 horas, luego se transfirieron al tanque de nitrógeno líquido a -196OC.
6.- Isotipificación de los anticuerpos.
Para determinar el isotipo de los anticuerpos producidos por los híbridos de fusión, se analizó el sobrenadante de cultivo por medio de ELISA de doble anticuerpo que consiste en recubrir una placa con anticuerpos de cabra anti IgG1, 1gGnal lgGzbl lgG3, e IgM diluidos en PBS, se incubó 1 hora a 37OC, se lavó 2 veces y se bloqueó con PBS-gelatina 3% durante 2 horas a 37OC, se lavó 2 veces, se adicionó el sobrenadante de cultivo del hibridoma a analizar, se incubó 1 hora a T ambiente y después se lavó 2 veces; se añadió el anticuerpo anti-lg de ratón polivalente conjugado a peroxidasa diluido en PBS (1:1000), se incubó por 30 minutos a T ambiente, se lavó 4 veces y se reveló con buffer de citratos, H202 y OPD . Se detuvo la reacción a los 10 minutos por adición de H2S04 2.5 N y se leyeron las densidades ópticas a una longitud de onda igual a 492 nm.
7.- Producción masiva y purificación de anticuerpos .
Se cultivaron por agotamiento 1 x I O 8 células del hibridoma con mayores títulos de producción de anticuerpos por ELSA en cilindros de cultivo celular con medio RPMI-1640 suplementado al 5% con suero fetal bovino, la viabilidad y esterilidad del cultivo se determinó diariamente por inspección microscópica, así como el requerimiento de medio de cultivo fresco.
9

Una vez que al microscopio se determinó una viabilidad celular menor del 5% por campo visual, se procedió a colectar el sobrenadante de cultivo, centrifugando a 3000 rpm para retirar restos celulares y se pasó por columna de proteína A acoplada a sepharosa (MonoAb rec-Protein A Column Purification System, ZYMED 10-1 160) 6 veces con un flujo de 350 pl/min. Se eluyó con 6 ml de buffer de elución y se colectaron diversas fracciones de 1 ml a las que se les midió absorbancia a una longitud de onda de 280 nm y se dialisaron contra PBS pH=7.4 durante una noche.
8.- Caract6rización de los anticuerpos por SDS-PAGE.
Se determinó la presencia de moléculas de inmunoglobulinas en el eluído mediante electroforesis en geles de poliacrilamida al IO%, corriéndolos en condiciones reductoras y no reductoras, con el objetivo de observar la molécula completa de inmunoglobulina y las cadenas pesadas y ligeras.
10
RESULTADOS.
- AnAlisis del suero de ratones inmunes.
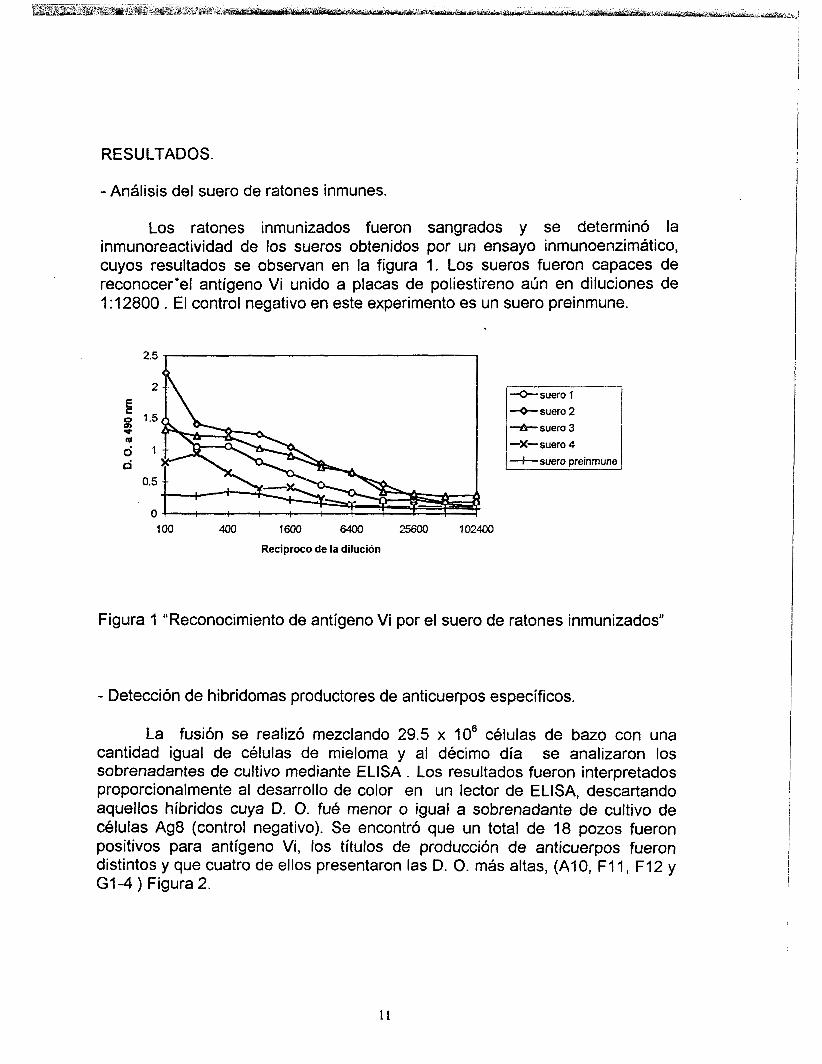
Los ratones inmunizados fueron sangrados y se determinó la inmunoreactividad de los sueros obtenidos por un ensayo inmunoenzimático, cuyos resultados se observan en la figura 1. Los sueros fueron capaces de reconocer‘el antígeno Vi unido a placas de poliestireno aún en diluciones de 1 : 12800 . El control negativo en este experimento es un suero preinmune.
2.5 1 I 2 A
E o 1.5 8 O ’ o
e
0.5
O
-0- suero 2 -Lt- suero 3 -X- suero 4
loo 400 1600 6400 25600 102400
Recíproco de la dilución
Figura 1 “Reconocimiento de antígeno Vi por el suero de ratones inmunizados”
- Detección de hibridomas productores de anticuerpos específicos.
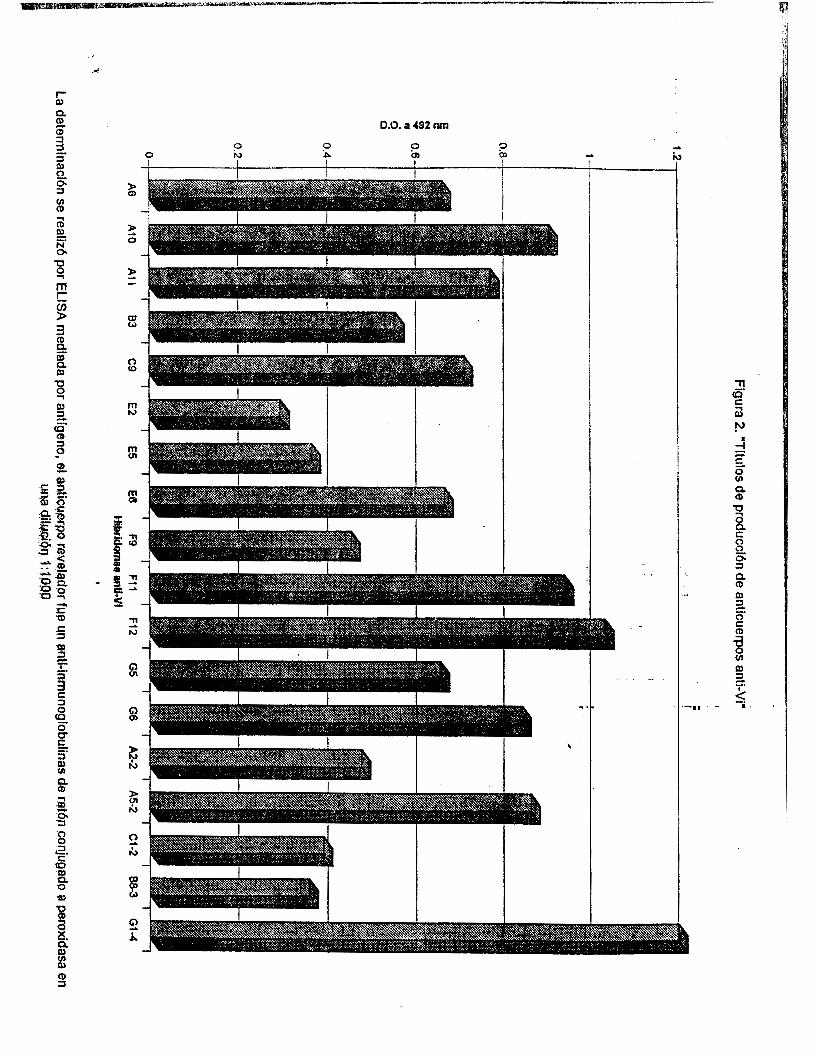
La fusión se realizó mezclando 29.5 x I O 6 células de bazo con una cantidad igual de células de mieloma y al décimo día se analizaron los sobrenadantes de cultivo mediante ELISA . Los resultados fueron interpretados proporcionalmente al desarrollo de color en un lector de ELISA, descartando aquellos híbridos cuya D. O. fué menor o igual a sobrenadante de cultivo de células Ag8 (control negativo). Se encontró que un total de 18 pozos fueron positivos para antígeno Vi, los títulos de producción de anticuerpos fueron distintos y que cuatro de ellos presentaron las D. O. más altas, (AIO, F11, F12 y G I 4 ) Figura 2.
11
. -
A-
0.0. a 192 m
O
2
2 O
. .
. -
.”
-11

- Criopreservación.
Una vez expandidos a botellas de cultivo de 25 cm2 , los hibridomas obtenidos se congelaron en solución crioprotectora, elaborada con 9 partes de suero fetal bovino y 1 parte de DMSO, filtrada por membrana de 0.22 pm. Se procedib a congelar a -70 O C y luego se transfirieron los viales al tanque de nitrógeno liquido a -196 OC. Se almacenaron al menos tres viales por hibridoma productor.
- Determinación del isotipo de los anticuerpos. 2 2 2 5 2 6
En la generalidad de los casos, se detectó una mezcla de isotipos, con una predominancia de los anticuerpos cuyos isotipos son IgM, I s G ~ ~ , lgG2b e lgG3 , lo que demuestra la necesidad de clonaciones por dilución limitante para obtener anticuerpos de un soloisotipo. Los 18 híbridos de fusión obtenidos demostraron positividad para IgM, 6 para IgGna, 5 para lgG2b y 6 para lgG3 . El control negativo en los ensayos de ELISA de doble anticuerpo fue, nuevamente sobrenadante de cultivo de células Ag8.
- Purificación de los anticuerpos.
Se obtuvieron 5 alícuotas del eluído, cuyas densidades ópticas a 280 nm fueron las siguientes:
MUESTRA DENSIDAD OPTICA. 1 2.1 81 5 2 1 .8958 3 0.7558 4 0.2605 5 O. 1350
Como se puede observar, en las primeras tres fracciones eluyeron los anticuerpos, de modo que se promediaron las tres absorbancias y con el resultado se determinó por proporcionalidad la concentración de anticuerpos contenidos en estas fracciones (el coeficiente de extinción molar de las inmunoglobulinas de isotipo IgG es de 1.43 y esta absorbancia equivale a 1 mg/ml, ver ref. 16). Esta proporcionalidad resulta en una concentración aproximada de 1 .I26 mglml de inmunoglobulinas de isotipo IgG.
13

- Caracterización de los anticuerpos presentes en las fracciones eluídas.
En el gel de poliacrilamida se evidencia la presencia de inmunoglobulinas y de otras proteínas (como albúmina). Se dispensaron muestras de 4, 6 y 8 pg de proteínas eluídas en los carriles 2, 3 y 4 , en condiciones no reductoras así como en los carriles 6, 7 y 8 en condiciones reductoras. En los carriles 1 y 5, como estsndar, se dispensaron 20 pg de inmunoglobulinas de cabra en condiciones no reductoras y en condiciones reductoras, respectivamente (ver Figura 3). .
Figura 3. SDS-PAGE 10%. Caracterización de anticuerpos dirigidos contra el antígeno Vi.
14

Actividades realizadas: se llevaron a cabo tres ensayos de fusión somática, de los cuales, el tercero fue en el que se obtuvieron los hibridomas.
Recomendaciones: Como se sabe, los polisa&ridos son antígenos timo- independientes, por lo que no promueven el cambio de isotipo en los linfocitos 6; en el presente trabajo, con el fin de obtener anticuerpos de isotipo IgG, las primeras dos inmunizaciones se realizaron con bacterias completas y la tercera y última inmunización se realizó con antígeno soluble y en una baja concentración ( I O pg), aprovechando la mayor afinidad característica de los anticuerpos de isotipo IgG con respecto a los anticuerpos de isotipo IgM. En los experimentos de fusión somática es importante recibir a las células recién fusionadas en c6lulas alimentadoras, que hayan condicionado el medio de cultivo con factores de crecimiento y que además fagociten el detritus de las células que van muriendo a causa del medio selectivo. Por otra parte, el haber purificado células 6 del bazo economiza tiempo y trabajo en el periodo post- fusión, debido a que el número celular se ve considerablemente reducido y se amplifica la posibilidad de fusionar una célula 8 secretora de anticuerpos con una célula de mieloma.
Conclusiones: se obtuvieron 18 híbridos de fusión sercretores de anticuerpos dirigidos contra el antígeno Vi de los que al menos se tienen 6 hibridomas secretores de anticuerpos de isotipo IgG. La obtención de anticuerpos monoclonales de este isotipo tiene relevancia acadkmica y práctica, debido a la mayor afinidad y estabilidad de los anticuerpos de isotipo IgG respecto a los anticuerpos de isotipo IgM; además de la mayor facilidad de purificación de IgG.
Los isotipos detectados son, principalmente IgM, lgG2, e IgG,; se hace necesaria la posterior clonación por dilución limitante para obtener clonas productoras de anticuerpos de un solo isotipo.
Debido a la presencia de albúmina en las fracciones eluídas (Figura 3), se necesita recurrir a la conjugación de métodos de purificación que permitan obtener los anticuerpos libres de otras proteínas.
Los anticuerpos monoclonales constituyen una herramienta de gran utilidad para diversos tipos de estudios, debido a su alta especificidad de reconocimeinto a un determinante antigénico en particular. Su desarrollo permitirá diseñar una metodología apropiada para la detección del antígeno en diferentes fluidos biológicos, pudiéndose emplear para identificar portadores de Salmonella typhi mediante el uso de pruebas tamiz.
15

BIBLIOGRAFIA.
1 .- Abbas, A. K. , Lichtman, A. H. , Pober, J. S. Cellular and molecular immunology (1991) W B Saunders Co. Philadelphia 39-40
2.- Baret, T. , Snyder, J., Blake, P. and Filley, J. (1982) Enzyme-linked immunosorbent assay for detection of Salmonella typhi Vi antigen in urine from typhoid fev'er. J. Clin. Microbiol. 15(2): 235-37
3.- Brown, G., Ling, N. Murine monoclonal antibodies. En: Antibodies, a practical approach (1989) Catty, A. IRLPRESS, Oxford. 81 -106
4.- Berzofsky, J. A., Berkower, I. J., Epstein, S. L. Antigen-antibody interactions and monoclonal antibodies. En: Fundamental immunology (1993) Paul, W. E. Third ed. Raven Press, New York 455-65
5.- Dacso, C., Nolan, P., Liu, P. Enfermedades infecciosas. En: Manual de pruebas diagnósticas (I 988) Liu, P. Interamericana-McGraw Hill. Madrid. 335-48
6.- Daniels, E. M., Schneerson, R., Egan, W., Szu, S. C., Robbins, J. B. (1989) Characterization of the Salmonella paratyphi C Vi polysaccharide. Infect. Immun. Vol 57: 31 59-63
7.- Falkow, S., Mekalanos, J. The enteric bacilli and vibrios. En: Microbiology (1990) Davis, B., Dulbecco, R., Eisen, H., Ginsberg, H. S. (Eds) Fourth ed. J.B. Lippincott Co. Philadelphia. 39-40
8.- Freshney, R. I . Culture of animal cells (1 994) Third ed. Wiley-Liss New York 394-97 I
9.- Harlow, E. Lane, D. Antibodies, a laboratory manual. (1 988) Cold Spring Harbor lab. USA, 71 1 pp
16

10.- Washimoto, V., Itho, Y. , Fujinaga, Y., Khan, A., Sultana, F., Miyake, M., Hirose, K., Yamamoto, f-l. and Ezaki, T. (1995) Development of nested PCR based on the via6 sequence to detect Salmonella tyhpi. J. Clin. Microbiol. 33(3): 775-77
11 .- Hornick, R. B. Typhoid fever. En: Hoeprich, P. D. , Jordan, M. C. , Ronald, A. R. Infectious diseases. (1994) Fifth ed. J. B. Lippincott Co. Philadelphia 747- 53 *
12.- Keusch, G. Salmonetosis. En: Isselbacher, Braunwald, Wilson, Martin, Fauci, Kasper. Harrison, principios de medicina interna (1 994) 1 3a ed. Interamericana-McGraw Hill. España 782-88
13.- Kohler, G. and Milstein, C. (1975) Continuos cultures of fused cells secreting antibodies of predefined specificity. Nature 256:495
14.- Lanata, C., Ristori, C., Jiménez, L. Garcia, J., Levine, M. , Black, R., Salcedo, M. and Sotomayor, V. (1983) Vi serology in detection of chronic Salmonella typhi carriers in an endemic area. The Lancet, august 20:441-43
15.- Liao, J., Nickerson, K., Bistricky, S., Robbins, J., Schneerson, R., Szu, S. and Kabat, E. (1995) Characterization of a human monoclonal immunoglobulin M (IgM) antibody (IgM BEN) specific for Vi capsular polysaccharide of Salmonella typhi Infect. Immun. 63( 1 1 ):4429-32
16.- Nilsson, B. Current opinion in immunology. Enzyme-linked immunosorbent assays (1 990) 2:898-904
17.- Peters, J. H. and Baumgarten, H. Monoclonal antibodies (1992) Springer Verlag. Germany. 171.
18.- Phillips, M. S., Zodda, D. M. Monoclonal antibodies and immunoparasitology. En: Monoclonal antibodies and functional cell lines. Progress and applications (1984) Kennet, R. H., Bechtol, K. B. , McKearn, T. J. (Eds) Plenum Press, USA 239-68
17

19.- Qadri, A., Ghosh, S., Prakash, K, Ramesh, K., Moudgil, K. And Talwar, P. (1990) Sandwich enzyme immunoassays for detection of Salmonella fyphi . J. Immunoassay 1 1 (2):251-70
2;<2526
n
21.- Rockhill, R., Rumans, L., Lesmana, M. and Dennis , D. (1980) Detection of .;'.--
Salmonella fyphi Dl Vi and d antigens, by slide coagglutination, in urine from patients with typhoid fever. J. Cli. Microbiol. 11(3):213-I6
x- ?Et>,
, ,!" . . I d
22.- Szu, S. C., Li, X-R., Stone, A. and Robbins, J. (1991) Relation between structure and immunologic properties of the capsular Vi polysaccharide. Infect. Immun. 59( 12): 4555-61
23.- Szu, S. C., Stone, A. L., Robbins, J. O., Schneerson, R., Robbins, J. B. (1 987) Vi capsular polysaccharide-protein conjugates for prevention of typhoid fever. J. Exp. Med. 166: 1510-24
24.- Szu, S. C., Taylor, D., Trota, A., Clements, J., Shiloach, J., Sadoff, J., Bryla, D. and Robbins, J. (1994) Laboratory and preliminary clinical characterization of Vi capsular polysaccharide-protein conjugates vaccines. Infect. Immun. 62( 10):4440-44
25.- Tacket, C., Ferreccio, C., Robbins, J., Tsai, C-M., Schulz, D., Cadoz, M., Goudeau, A. and Levine, M. (I 986) Safety and immunogenicity of two Salmonella typhi Vi capsular polysaccharide vaccines. J. Infect. Dis. 154(2):342- 45
26.- Taylor, Ch. And Bright, R. (1989) T-cell modulation of the antibody response to bacterial polysaccharide antigens Infect. Immun. 57(1):180-85
27.- Tsang, R. S. W. And Chau, P. Y. (1987) Production of Vi monoclonal antibodies and their application as diagnostic reagents. J. Clin. Microbiol. 25 (3):531-35
18

28.- Taylor, D., Harris, J., Barret, T., Hargrett, N., Prentzel, l . , Valdivieso, C., Palomino, C., Levine, M. and Blake, P.(1983) Detection of urinary Vi antigen as a diagnostic test for typhoid fever. J. Clin. Microbiol. 18(4):872-76
29.- Virlogeux, l., Waxin, H., Ecobichon, Ch., Lee, J. And Popoff, M. (1996) Characterization of the rcsA and res6 genes from Salmonella typhi : rcs6 through tviA is involved in regulation of Vi antigen synthesis. J. Bacteriol. 178 (6): 1691 -98
30.- Wong, K. H. and Feeley, J. C. (1974) Adhesion of Vi antigen and toxicity in typhoid vaccines inactivated by acetona or by heat .and phenol. J. Infect. Dis. 129(5):501-O6
31.- Wong, K. H., Feeley, J., Northrup, R. and Forlines, M. E. (1974) Vi antigen from Salmonella typhosa and immunity against typhoid fever. Infect. Immun. 9(2):348-53
32.- ATCC QC. and reference strains. First ed. 1997 M. J. Edwards M. L. S. editor. USA, 120 pp.
19
