biosynthesis of diterpenoids in tripterygium adventitious
TRANSCRIPT

Biochemistry, Biophysics and Molecular BiologyPublications Biochemistry, Biophysics and Molecular Biology
9-2017
Biosynthesis of Diterpenoids in TripterygiumAdventitious Root CulturesFainmarinat S. InabuyWashington State University
Justin T. FischedickWashington State University
Iris LangeWashington State University
Michael HartmannWashington State University
Narayanan SrividyaWashington State University
See next page for additional authors
Follow this and additional works at: http://lib.dr.iastate.edu/bbmb_ag_pubs
Part of the Molecular Biology Commons, and the Molecular Genetics Commons
The complete bibliographic information for this item can be found at http://lib.dr.iastate.edu/bbmb_ag_pubs/184. For information on how to cite this item, please visit http://lib.dr.iastate.edu/howtocite.html.
This Article is brought to you for free and open access by the Biochemistry, Biophysics and Molecular Biology at Iowa State University DigitalRepository. It has been accepted for inclusion in Biochemistry, Biophysics and Molecular Biology Publications by an authorized administrator of IowaState University Digital Repository. For more information, please contact [email protected].

Biosynthesis of Diterpenoids in Tripterygium Adventitious Root Cultures
AbstractAdventitious root cultures were developed from Tripterygium regelii Sprague & Takeda and growthconditions optimized for the abundant production of diterpenoids, which can be collected directly from themedium. An analysis of publicly available transcriptome data sets collected with T. regelii roots and rootcultures indicated the presence of a large gene family (with 20 members) for terpene synthases (TPSs). Ninecandidate diterpene synthase genes were selected for follow-up functional evaluation, of which two belongedto the TPS-c, three to the TPS-e/f and four to the TPS-b subfamily. These genes were characterized byheterologous expression in a modular metabolic engineering system in E. coli. Members of the TPS-csubfamily were characterized as copalyl diphosphate (diterpene) synthases and those belonging to the TPS-e/f family catalyzed the formation of precursors of kaurane diterpenoids. The TPS-b subfamily encompassedgenes coding for enzymes involved in abietane diterpenoid biosynthesis and others with activities asmonoterpene synthases. The structural characterization of diterpenoids accumulating in the medium of T.regelii adventitious root cultures, facilitated by searching the Spektraris online spectral database, enabled us toformulate a biosynthetic pathway for the biosynthesis of triptolide, a diterpenoid with pharmaceuticalpotential. Considering the significant enrichment of diterpenoids in the culture medium, fast-growingadventitious root cultures may hold promise as a sustainable resource for the large-scale production oftriptolide.
DisciplinesBiochemistry, Biophysics, and Structural Biology | Genetics and Genomics | Molecular Biology | MolecularGenetics
CommentsThis article is published as Inabuy, F.X., Fischedick, J.T., Lange, I., Hartmann, M., Srividya, N., Parrish, A.N.,Xu, M., Peters, R.J., Lange, B.M. (2017) “Biosynthesis of diterpenoids in Tripterygium adventitious rootcultures”, Plant Physiol., 175(1):92-103. doi: 10.1104/pp.17.00659. Posted with permission.
AuthorsFainmarinat S. Inabuy, Justin T. Fischedick, Iris Lange, Michael Hartmann, Narayanan Srividya, Amber N.Parrish, Meimei Xu, Reuben J. Peters, and B. Markus Lange
This article is available at Iowa State University Digital Repository: http://lib.dr.iastate.edu/bbmb_ag_pubs/184

Biosynthesis of Diterpenoids in TripterygiumAdventitious Root Cultures1[OPEN]
Fainmarinat S. Inabuy ,a Justin T. Fischedick,a,2 Iris Lange,a Michael Hartmann,a,3 Narayanan Srividya,a
Amber N. Parrish,a Meimei Xu,b Reuben J. Peters,b and B. Markus Langea,4
aInstitute of Biological Chemistry and M.J. Murdock Metabolomics Laboratory, Washington State University,Pullman, Washington 99164-6340bRoy J. Carver Department of Biochemistry, Biophysics, and Molecular Biology, Iowa State University, Ames,Iowa 50011-1079
ORCID IDs: 0000-0003-4828-958X (M.H.); 0000-0001-7934-7987 (N.S.); 0000-0002-4080-8608 (A.N.P.); 0000-0003-4691-8477 (R.J.P.);0000-0001-6565-9584 (B.M.L.).
Adventitious root cultures were developed from Tripterygium regelii, and growth conditions were optimized for the abundantproduction of diterpenoids, which can be collected directly from the medium. An analysis of publicly available transcriptomedata sets collected with T. regelii roots and root cultures indicated the presence of a large gene family (with 20 members) forterpene synthases (TPSs). Nine candidate diterpene synthase genes were selected for follow-up functional evaluation, of whichtwo belonged to the TPS-c, three to the TPS-e/f, and four to the TPS-b subfamilies. These genes were characterized byheterologous expression in a modular metabolic engineering system in Escherichia coli. Members of the TPS-c subfamily werecharacterized as copalyl diphosphate (diterpene) synthases, and those belonging to the TPS-e/f subfamily catalyzed theformation of precursors of kaurane diterpenoids. The TPS-b subfamily encompassed genes coding for enzymes involved inabietane diterpenoid biosynthesis and others with activities as monoterpene synthases. The structural characterization ofditerpenoids accumulating in the medium of T. regelii adventitious root cultures, facilitated by searching the Spektraris onlinespectral database, enabled us to formulate a biosynthetic pathway for the biosynthesis of triptolide, a diterpenoid withpharmaceutical potential. Considering the significant enrichment of diterpenoids in the culture medium, fast-growingadventitious root cultures may hold promise as a sustainable resource for the large-scale production of triptolide.
Tripterygium wilfordii, also known as léi gōng téng inMandarin Chinese (generally translated as “thundergod vine”), has a long history of use in traditional
Chinese medicine for the treatment of fever, chills,edema, and carbuncles (Helmstädter, 2013). The genusTripterygium (Celastraceae) is known to be a rich sourceof specializedmetabolites, of whichmore than 400 havebeen isolated, structurally characterized, and assessedin cell-based assays (Brinker et al., 2007). Root extractshave been evaluated as a medication for rheumatoidarthritis, cancer, hepatitis, nephritis, ankylosing spon-dylitis, polycystic kidney disease, and obesity; morethan a dozen clinical trials with such extracts (oftenreferred to as Tripterygium glycoside) have been com-pleted, but, in part due to shortcomings in study de-signs, the efficacy has remained a matter of debate(Chen et al., 2010; Liu et al., 2013; Zhu et al., 2013). Morepromising results have been obtained with purifiedconstituents, which are usually extracted from Trip-terygium roots. Semisynthetic chemical derivatives oftriptolide, a diterpenoid epoxide, have been evaluatedin phase I clinical trials (Zhou et al., 2012; Meng et al.,2014). Minnelide, a water-soluble prodrug analog oftriptolide, has shown very promising activity in mul-tiple animal models of pancreatic cancer (Chugh et al.,2012) and, in 2013, was advanced to phase I clinicaltrials in the United States (clinicaltrials.gov identifierNCT01927965).
One of the critical challenges for clinical evaluationsis a supply shortage for triptolide and other diterpe-noids. The most abundant specialized metabolites in
1 This work was supported by the National Institutes of Health(award nos. RC2GM092561 to B.M.L. and GM076324 to R.J.P.) andMcIntire-Stennis formula funds from the Agricultural Research Cen-ter at Washington State University (to B.M.L.).
2 Current address: Excelsior Analytical, Union City, CA 94587.3 Current address: Institute of Molecular Ecophysiology of Plants,
Heinrich Heine Universität Düsseldorf, 40225 Duesseldorf, Germany.4 Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:B. Markus Lange ([email protected]).
B.M.L. conceived of the project; F.S.I. cloned candidate genes,subcloned them into appropriate modular expression vectors, andanalyzed diterpenoids produced in E. coli cells harboring these con-structs; J.T.F. established the methods for the isolation, purification,identification, and quantitation of diterpenoids; I.L. isolated RNA fromadventitious root cultures, cloned candidate genes, and performed thescale-up and selection of adventitious root cultures for high diterpenoidproduction; M.H. established and A.N.P. maintained the adventitiousroot cultures; N.S. performed the functional characterization of mono-terpene synthase genes; M.X. and R.J.P. supplied modular vectors andcarried out diterpenoid characterizations; B.M.L. and F.S.I. wrote thearticle, with contributions from all authors.
[OPEN] Articles can be viewed without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.17.00659
92 Plant Physiology�, September 2017, Vol. 175, pp. 92–103, www.plantphysiol.org � 2017 American Society of Plant Biologists. All Rights Reserved. www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from
Copyright © 2017 American Society of Plant Biologists. All rights reserved.

Tripterygium roots are quinone methide triterpenoidsand sesquiterpene pyridine alkaloid macrolides,whereas triptolide and other diterpenoids occur only atvery low concentrations ranging from 0.0001% to0.002% of dry weight biomass (Zhou et al., 2012; Zenget al., 2013; Guo et al., 2014). Accordingly, the extractionyields of diterpenoids from Tripterygium roots are poorand alternative, sustainable production methods needto be developed. Tissue cultures represent a promisingalternative for the production of high-value plant me-tabolites. Early studies with T. wilfordii suspensioncultures were designed to unravel the structures ofsmall-molecule constituents (Kutney et al., 1981, 1992,1993; Kutney and Han, 1996; Nakano et al., 1997, 1998).Similarly, hairy root cultures were initially employedfor phytochemical investigations (Nakano et al., 1998).It was recognized only recently that triptolide concen-trations produced by tissue cultures (up to 0.15% of dryweight; Miao et al., 2013, 2014; Zhu et al., 2014) far ex-ceed those reported for roots.Surprisingly, despite considerable pharmaceutical
interest, the biosynthesis of triptolide has only recentlyattracted the deserved attention. Several diterpenesynthases, some of which are relevant to triptolideformation, have now been characterized in T. wilfordii(Zerbe et al., 2013; Andersen-Ranberg et al., 2016;Hansen et al., 2017). However, the remaining genesinvolved in the biosynthesis of the highly functional-ized triptolide structure have remained enigmatic. Inthis article, we describe the development of Triptery-gium adventitious root cultures in which diterpenoidsare produced as principal metabolites that can be har-vested sustainably from the medium. Based on thestructures of these metabolites, we can now postulatethe biochemical steps leading to the main diterpenoidmetabolites. In addition, analysis of transcriptome datasets, acquiredwith diterpenoid-producing adventitiousroot cultures, enabled the selection and subsequentfunctional characterization of genes involved inthe early steps of diterpenoid biosynthesis. Our resultsindicate that Tripterygium tissue cultures offer oppor-tunities to both unravel diterpenoid biosyntheticpathways and produce target diterpenoids, such astriptolide, at larger scale.
RESULTS
Development of Diterpenoid-Secreting AdventitiousRoot Cultures
Tripterygium regelii adventitious root cultures wereinitiated based on standard protocols. Over a 1-yearperiod, during which mediumwas extracted with ethylacetate and hydrophobic metabolites were analyzed byHPLC-quadrupole time of flight-mass spectrometry(QTOF-MS), the highest triptolide producers were se-lected for further propagation. In contrast to roots ofmature Tripterygium plants, where triterpenoids occurat low concentrations (less than 1%) and diterpenoidsare trace constituents (less than 0.01%), our
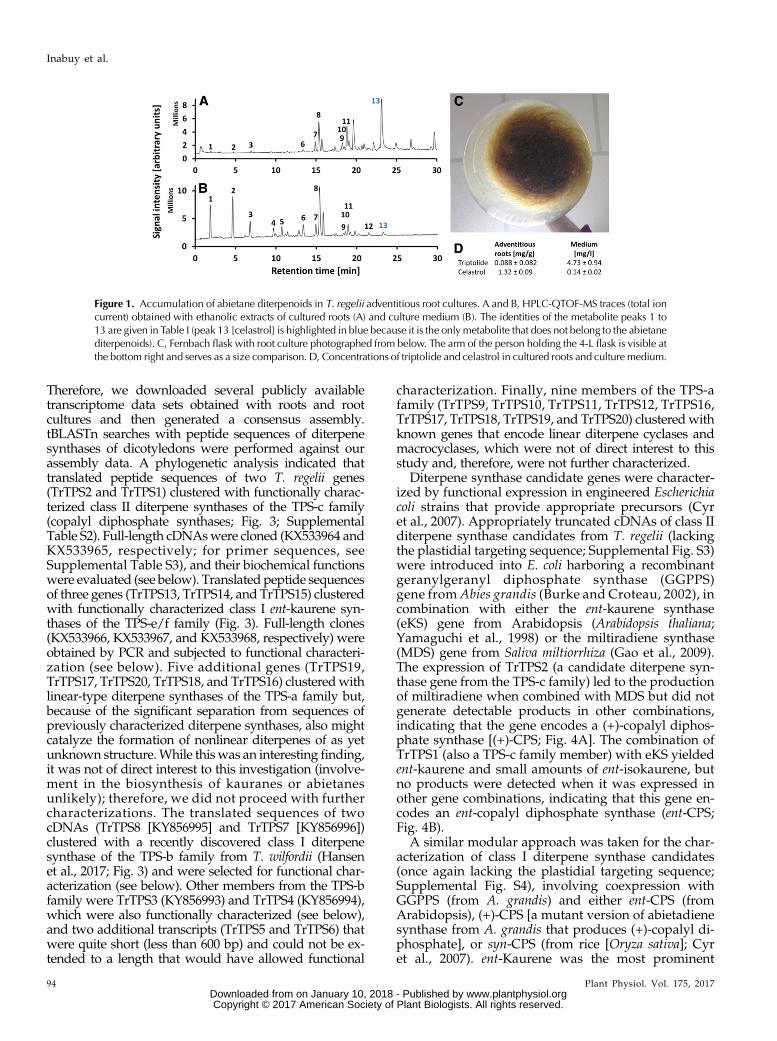
adventitious root cultures accumulated diterpenoidsabundantly in the medium and triterpenoids (in par-ticular celastrol) in roots (Fig. 1, A and B). By comparingadjusted peak areas from HPLC-QTOF-MS runs, weestimated that diterpenoids constitute approximately77% of all detected metabolites in an organic extract ofthe culture medium (Supplemental Fig. S1).
An accurate quantitation of all diterpenoids was notpossible because we did not have a complete set ofauthentic standards, but we performed absolutequantitations for signature metabolites (triptolide andcelastrol). Various common reagents and elicitors weretested (methyl jasmonate, methyl salicylate, chitosan,yeast extract, and cold exposure), but only the methylsalicylate treatment resulted in a significant increase intriptolide accumulation (2.2-fold; P value of 0.01) and aconcomitant decrease in celastrol concentration(Supplemental Table S1). The disadvantage of methylsalicylate treatments was that root color darkened andgrowth ceased, often for months. Therefore, we focusedour efforts on further selecting cultures that were highditerpenoid producers under noninducing conditionsand could be maintained for extended periods of time.These cultures were gradually scaled up to 2.8-L flasks(500mL of medium; Fig. 1C). Triptolide accumulated at4.7 mg L21 (Fig. 1D) and was readily extractable fromthe culture medium every 2 weeks (when adventitiousroots were transferred to fresh medium).
Hundreds of structurally diverse metabolites havebeen isolated and characterized from various Triptery-gium organs and tissues (Lange et al., 2017). Our ad-ventitious root cultures, in contrast, secreted primarilyditerpenoids into the culture medium and, therefore,are an excellent experimental model system in which tostudy the biosynthesis of these clinically relevant me-tabolites. As a first step to further assess the potential ofthese tissue cultures, it was important to identify themajor constituents of medium extracts. FollowingHPLC fractionation of medium extracts of T. regeliiadventitious root cultures, fractions containing metab-olites that corresponded to prominent peaks in HPLC-QTOF-MS chromatograms were characterized by massspectrometry (MS), tandem mass spectrometry (MS/MS), and 1H-NMR spectroscopy (Supplemental Fig. S2;Supplemental Methods and Data File S1), and metab-olites were identified by searches against the Spektrarisdatabase (Fischedick et al., 2015). We ascertained theidentities of five diterpenoids, were able to annotatethree additional metabolites with high confidence, andacquired tentative identifications for an additional twometabolites (Table I and Fig. 2). Threemetabolites showedthe mass fragmentation patterns typical of diterpenoids,but their identities remained unknown (Table I).
Functional Characterization of CandidateDiterpene Synthases
To obtain full-length candidate genes, we deemed itadvantageous to employ all available T. regelii sequences.
Plant Physiol. Vol. 175, 2017 93
Diterpenoid Biosynthesis in Tripterygium
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.
Therefore, we downloaded several publicly availabletranscriptome data sets obtained with roots and rootcultures and then generated a consensus assembly.tBLASTn searches with peptide sequences of diterpenesynthases of dicotyledons were performed against ourassembly data. A phylogenetic analysis indicated thattranslated peptide sequences of two T. regelii genes(TrTPS2 and TrTPS1) clustered with functionally charac-terized class II diterpene synthases of the TPS-c family(copalyl diphosphate synthases; Fig. 3; SupplementalTable S2). Full-length cDNAswere cloned (KX533964 andKX533965, respectively; for primer sequences, seeSupplemental Table S3), and their biochemical functionswere evaluated (see below). Translated peptide sequencesof three genes (TrTPS13, TrTPS14, and TrTPS15) clusteredwith functionally characterized class I ent-kaurene syn-thases of the TPS-e/f family (Fig. 3). Full-length clones(KX533966, KX533967, and KX533968, respectively) wereobtained by PCR and subjected to functional characteri-zation (see below). Five additional genes (TrTPS19,TrTPS17, TrTPS20, TrTPS18, and TrTPS16) clusteredwithlinear-type diterpene synthases of the TPS-a family but,because of the significant separation from sequences ofpreviously characterized diterpene synthases, also mightcatalyze the formation of nonlinear diterpenes of as yetunknown structure.While thiswas an interesting finding,it was not of direct interest to this investigation (involve-ment in the biosynthesis of kauranes or abietanesunlikely); therefore, we did not proceed with furthercharacterizations. The translated sequences of twocDNAs (TrTPS8 [KY856995] and TrTPS7 [KY856996])clustered with a recently discovered class I diterpenesynthase of the TPS-b family from T. wilfordii (Hansenet al., 2017; Fig. 3) and were selected for functional char-acterization (see below). Other members from the TPS-bfamily were TrTPS3 (KY856993) and TrTPS4 (KY856994),which were also functionally characterized (see below),and two additional transcripts (TrTPS5 and TrTPS6) thatwere quite short (less than 600 bp) and could not be ex-tended to a length that would have allowed functional
characterization. Finally, nine members of the TPS-afamily (TrTPS9, TrTPS10, TrTPS11, TrTPS12, TrTPS16,TrTPS17, TrTPS18, TrTPS19, and TrTPS20) clustered withknown genes that encode linear diterpene cyclases andmacrocyclases, which were not of direct interest to thisstudy and, therefore, were not further characterized.
Diterpene synthase candidate genes were character-ized by functional expression in engineered Escherichiacoli strains that provide appropriate precursors (Cyret al., 2007). Appropriately truncated cDNAs of class IIditerpene synthase candidates from T. regelii (lackingthe plastidial targeting sequence; Supplemental Fig. S3)were introduced into E. coli harboring a recombinantgeranylgeranyl diphosphate synthase (GGPPS)gene fromAbies grandis (Burke and Croteau, 2002), incombination with either the ent-kaurene synthase(eKS) gene from Arabidopsis (Arabidopsis thaliana;Yamaguchi et al., 1998) or the miltiradiene synthase(MDS) gene from Saliva miltiorrhiza (Gao et al., 2009).The expression of TrTPS2 (a candidate diterpene syn-thase gene from the TPS-c family) led to the productionof miltiradiene when combined with MDS but did notgenerate detectable products in other combinations,indicating that the gene encodes a (+)-copalyl diphos-phate synthase [(+)-CPS; Fig. 4A]. The combination ofTrTPS1 (also a TPS-c family member) with eKS yieldedent-kaurene and small amounts of ent-isokaurene, butno products were detected when it was expressed inother gene combinations, indicating that this gene en-codes an ent-copalyl diphosphate synthase (ent-CPS;Fig. 4B).
A similar modular approach was taken for the char-acterization of class I diterpene synthase candidates(once again lacking the plastidial targeting sequence;Supplemental Fig. S4), involving coexpression withGGPPS (from A. grandis) and either ent-CPS (fromArabidopsis), (+)-CPS [a mutant version of abietadienesynthase from A. grandis that produces (+)-copalyl di-phosphate], or syn-CPS (from rice [Oryza sativa]; Cyret al., 2007). ent-Kaurene was the most prominent
Figure 1. Accumulation of abietane diterpenoids in T. regelii adventitious root cultures. A and B, HPLC-QTOF-MS traces (total ioncurrent) obtained with ethanolic extracts of cultured roots (A) and culture medium (B). The identities of the metabolite peaks 1 to13 are given in Table I (peak 13 [celastrol] is highlighted in blue because it is the onlymetabolite that does not belong to the abietanediterpenoids). C, Fernbach flask with root culture photographed from below. The arm of the person holding the 4-L flask is visible atthe bottom right and serves as a size comparison. D, Concentrations of triptolide and celastrol in cultured roots and culturemedium.
94 Plant Physiol. Vol. 175, 2017
Inabuy et al.
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.

reaction product (with ent-isokaurene as a minor sideproduct) when TrTPS13 or TrTPS14 (TPS-e/f familymembers) were expressed in combination with ent-CPS(Fig. 5, A and B), indicating ent-kaurene synthasefunctions for these isoforms. When combined with(+)-CPS, TrTPS13 expression produced small amountsof sandaracopimaradiene and isopimaradiene, whilecoexpression of TrTPS13 with syn-CPS led to the for-mation of syn-pimara-7,15-diene and syn-stemod-13(17)-ene (Fig. 5, C and D). The combination of bothclass II and class I diterpene synthases from T. regelii(TrTPS2 and TrTPS13) in one modular construct resul-ted in the production of ent-kaurene (with smallerquantities of ent-isokaurene; Supplemental Fig. S5),thus confirming the function of these genes in ent-kaurane diterpenoid biosynthesis. TrTPS15 (also amember of the TPS-e/f family) produced small quan-tities of ent-manool, syn-manool, or (+)-manool whencoupledwith ent-CPS, syn-CPS, or (+)-CPS, respectively(Fig. 5, E–G). When coupled with (+)-CPS, TrTPS8(from the TPS-b family; Supplemental Fig. S6) pro-duced primarily miltiradiene (Fig. 6A) along with smallamounts of abietatriene, presumably arising from apreviously noted autoxidation (Zi and Peters, 2013). Bothproducts also were found to be present in the medium ofT. regelii adventitious root cultures (Supplemental Fig.S1). When combined with ent-CPS or syn-CPS, TrTPS8generated small quantities of ent-manool or syn-manool,respectively (Fig. 6, B and C).
TrTPS3 and TrTPS4, members of the TPS-b subfamily(Supplemental Fig. S6), did not form products in anycombination with CPSs. Instead, both TrTPS3 andTrTPS4 produced mixtures of monoterpenes upon re-action with geranyl diphosphate (substrate for mono-terpene synthases), with (2)-linalool and (+)-linalool asmajor products (Table II; Supplemental Fig. S7). Whenreacted with linalyl diphosphate (a reaction interme-diate of monoterpene synthases), TrTPS3 and TrTPS4both formed (2)-terpineol and (+)-terpineol. In assayswith linalyl diphosphate, TrTPS3, but not TrTPS4, alsogenerated appreciable amounts (2)-limonene (;20% oftotal products; Table II). TrTPS7 was expressed in therecombinant E. coli strains but was not active in anycombination with copalyl diphosphate synthases. Italso did not generate products from geranyl diphos-phate as a substrate in in vitro assays.
DISCUSSION
Proposing a Diterpenoid Biosynthetic Pathway Based onMetabolites That Accumulate in TripterygiumAdventitious Root Cultures
In contrast to roots of mature Tripterygium plants,where diterpenoids are only very minor constituents(less than 0.01%), our adventitious root cultures se-creted these metabolites at very high levels (approxi-mately 77% of the area under all peaks detected by
Table I. Metabolites detected by HPLC-QTOF-MS in the medium of T. regelii adventitious root cultures
MS/MS spectra were acquired with a fragmentation voltage of 30 eV.
Plant Physiol. Vol. 175, 2017 95
Diterpenoid Biosynthesis in Tripterygium
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.

HPLC-QTOF-MS in organic extracts of the culturemedium). The structure and abundance of the principalditerpenoids found in our tissue cultures can thus beemployed to develop hypotheses regarding the bio-synthesis of abietane-type diterpenoids of Tripterygium(Fig. 2). It has been proposed previously that the two-step cyclization of the universal diterpenoid precursorgeranylgeranyl diphosphate, via (+)-copalyl diphos-phate, yields miltiradiene as an abietane hydrocarbonintermediate (Hansen et al., 2017), and our data agreewith this suggestion. Oxidation at C18, like that de-scribed for CYP720 in conifers (Ro et al., 2005;Hamberger et al., 2011; Geisler et al., 2016), wouldproduce dehydroabietic acid (which we detected inT. regelii root cultures; Fig. 2). Demethylation at C4,carboxylation at C3, and hydroxylation at C14 wouldresult in the formation of triptinin B. This metabolitewas not detected in our root cultures, but its methylether, triptinin A, was identified tentatively as a sig-nificant constituent. The tentatively identified neo-triptophenolide and its glycoside, both likely derivedfrom triptinin B and/or triptinin A, accumulated assignificant by-products in our root cultures. The lacto-nization of triptinin B would generate triptophenolide,which accumulated as the most abundant abietanediterpenoid in our root cultures (Fig. 2). The function-alization of the aromatic ring would then producetriptolide and, following an additional hydroxylation,
tripdiolide, as the primary end products of the bio-synthetic conversions detected in our adventitious rootcultures.
Schemes for abietane diterpenoid biosynthesis in thegenus Tripterygium have been proposed before (Kutneyet al., 1981; Kutney and Han, 1996), but these werebased on the available phytochemical evidence at thetime and did not incorporate information about relativemetabolite abundance (Fig. 1; Supplemental Fig. S1). Byadding this new dimension, we are increasing theconfidence in predictions regarding the design princi-ples of the pathways leading to structurally unusualditerpene triepoxides. Because of the high abundance ofditerpenoids in the medium of T. regelii adventitiousroot cultures, we have been able to employ simple pu-rification protocols for intermediates (triptophenolide)and end products (triptolide and tripdiolide). Thisprovides commercially unavailable substrates andproducts for in vitro enzyme assays and, therefore,enables the functional characterization of candidategenes.
Identifying Genes with Roles in Generating DiterpenoidStructural Diversity
One T. regelii clone (TrTPS1) was characterized asencoding an ent-CPS that, in combination with eKSs(TrTPS13 and TrTPS14), generates ent-kaurene (and
Figure 2. Putative biosynthetic pathway toward abietane diterpenoids in Tripterygium. Metabolites found in this study are boxed.The numbering of the carbons forming the abietane skeleton is indicated using dehydroabietic acid as an example. Reactionsrequiring multiple steps are indicated by dotted arrows. The numbering of metabolites is the same as in Figure 1.
96 Plant Physiol. Vol. 175, 2017
Inabuy et al.
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.

very small amounts of ent-isokaurene; Fig. 5). Thesereactions are common to all vascular plants, whichrequire endogenous production of the derived GAhormones (Zi et al., 2014). However, ent-kauranediterpenoids also occur as specialized metabolites insome plant families, including the Celastraceae (whichcontains the genus Tripterygium). The previouslyreported metabolites can be grouped into two majorclasses: four-ring ent-kauranoic acids/alcohols and
five-ring oxygen bridge-containing ent-kauranes (Duanet al., 1999, 2001; Tanaka et al., 2004). In the closely re-lated species T. wilfordii, recent studies also identifiedan ent-CPS (TwTPS3) and several isoforms of ent-kaurene/ent-isokaurene synthase (TwTPS2, TwTPS16,TwTPS17, and TwTPS18; Zerbe et al., 2013; Hansenet al., 2017). Various biological activities of Tripterygiument-kauranes have been documented in in vitro assays(for review, see Brinker et al., 2007), but the in vivo
Figure 3. Molecular phylogeny of dicot (di)terpene synthases, with an emphasis on those from the genus Tripterygium. Functionallycharacterized terpene synthases from T. regelii are highlighted by gray boxes. The analysis was carried out using the maximumlikelihoodmethod based on amatrix-basedmodel (Jones et al., 1992). The bootstrap consensus tree inferred from1,000 replications(Felsenstein, 1985) is taken to represent the evolutionary history of the taxa analyzed. Branches corresponding to partitionsreproduced in less than 50%of bootstrap replicates are collapsed. Initial tree(s) for the heuristic search were obtained automaticallyby applying neighbor-joining and BioNJ algorithms to a matrix of pairwise distances and then selecting the topology with superiorlog likelihood value. The analysis involved 77 amino acid sequences. All positions containing gaps and missing data were elimi-nated. There were a total of 94 positions in the final data set. Evolutionary analyses were conducted inMEGA7 (Kumar et al., 2016).The filled triangle indicates the position of the sequence chosen as an outgroup for rooting the phylogenetic tree.
Plant Physiol. Vol. 175, 2017 97
Diterpenoid Biosynthesis in Tripterygium
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.

functions remain to be elucidated. When combinedwith a (+)-CPS, TrTPS13 formed sandaracopimaradieneand isopimaradiene; in combination with a syn-CPS,TrTPS13 showed activity for the production of syn-pimara-7,15-diene and syn-stemod-13(17)-ene (Fig. 5).We did not find a syn-CPS in T. regelii, nor does thereappear to be such an activity among the class IIditerpene synthases of T. wilfordii (Hansen et al., 2017);therefore, the biological relevance of this finding isunknown at this time.
Genes from T. wilfordiiwere characterized previouslyas coding for ent-copal-8-ol diphosphate synthase(termed TwTPS21) or kolavenyl diphosphate synthase(TwTPS10, TwTPS14, and TwTPS28; Andersen-Ranberg et al., 2016; Hansen et al., 2017). However,the stereochemistry at C8 and C9 (8S, 9S) is the oppositeof that of all labdane-type diterpenoids characterizedfrom Tripterygium thus far (8R, 9R; Duan et al., 1999,2001). In our functional assays, TrTPS15 producedent-manool, (+)-manool, and syn-manool when com-bined with ent-CPS, (+)-CPS, or syn-CPS, respectively.Interestingly, (+)-manool has the correct stereochemis-try (8R, 9R) to serve as a precursor for the known lab-dane diterpenoids of Tripterygium (Duan et al., 1999,2001). The othermanools andmanoyl oxides, generatedby diterpene synthases of T. regelii and T. wilfordii, re-spectively, may only be produced in vivo under specificenvironmental conditions or may simply reflect enzy-matic substrate promiscuity. However, the generationof structural diversity by combinations of class II andclass I diterpene synthases, whether with recognizablein vivo relevance, does provide biosynthetic access tothese and derived diterpenoids.
The gene coding for T. regelii TrTPS8 [converts(+)-copalyl diphosphate to miltiradiene, a likely inter-mediate in triptolide and tripdiolide biosynthesis] is anortholog of the recently characterized TwTPS27 genefrom T. wilfordii (Hansen et al., 2017). Based on phylo-genetic analyses, Hansen et al. (2017) concluded thatTwTPS27 diversified from members of the TPS-b sub-family, more specifically from terpene synthases thatcatalyze the formation of acyclic monoterpenes. Thelocation of TwTPS27 on the phylogenetic tree of an-giosperm terpene synthases was a surprise becauseall angiosperm diterpene synthases involved in abie-tane/labdane biosynthesis characterized until thenbelonged to the TPS-e/f subfamily (Chen et al., 2011).This evolutionary history also applies to TrTPS8, whichis very closely related to TwTPS27.
Hansen et al. (2017) expressed TwTPS23, TwTPS24,and TwTPS26 transiently in Nicotiana benthamiana andthen subjected volatiles to a head-space analysis.The authors did not have authentic standards formonoterpenes and, therefore, could only use spectralcomparisons for tentative identification. We are in thefortunate position to host a sizable repository of mon-oterpenes and, therefore, were able to perform in vitroassays with unequivocal results. TrTPS3 and TrTPS4catalyzed the formation of a mixture of (2)-linalool and(+)-linalool from geranyl diphosphate as a substrate.These results from early termination (by water capture)immediately following the formation of a linalyl cation(Supplemental Fig. S6). To test if these enzymes would becapable of catalyzing cyclization if initial water capturewas avoided, we also performed assays with linalyl di-phosphate as a substrate. Indeed, in these assays, TrTPS3
Figure 4. Modular in vivo assay results obtained with T. regelii class II diterpene synthase candidates of the TPS-c subfamily.From left to right, constructs (selectable markers in gray boxes; Chlor, chloramphenicol; Carb, carbenicillin; MDS [from Salviamiltiorrhiza]; eKS [from Arabidopsis]), gas chromatography (GC)-MS chromatograms, and mass spectra of assay products andauthentic standards. TrTPS2 was identified as a (+)-CPS (A), while TrTPS1 showed activity as an ent-CPS (B).
98 Plant Physiol. Vol. 175, 2017
Inabuy et al.
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.
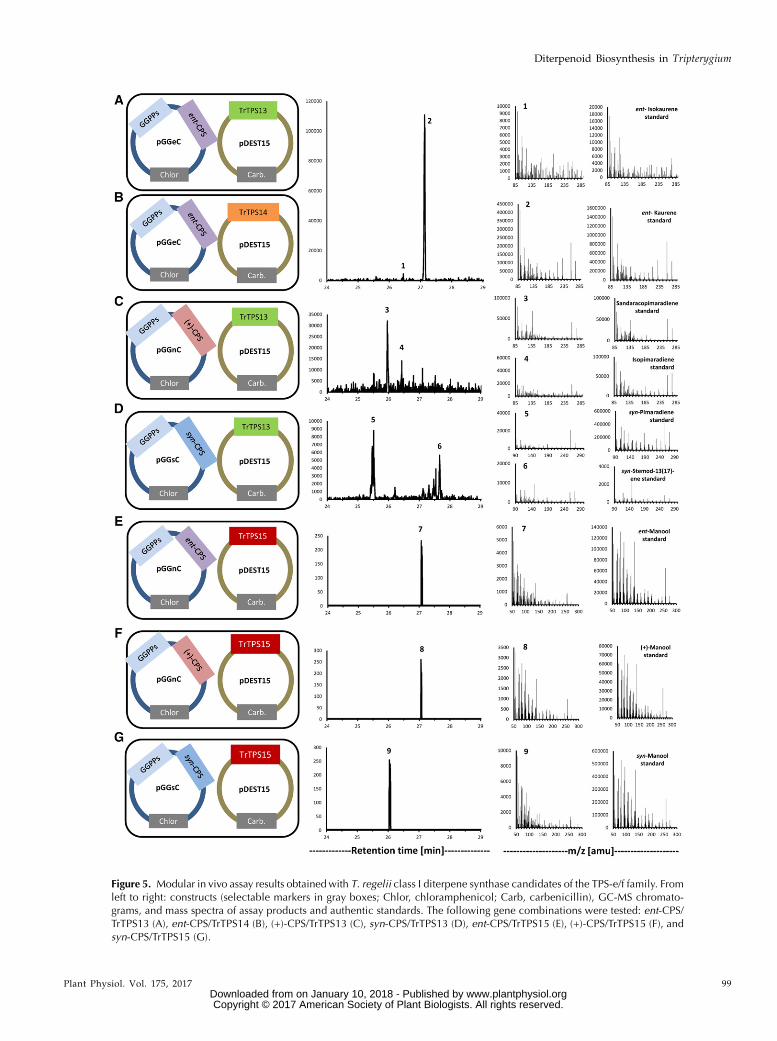
Figure 5. Modular in vivo assay results obtainedwith T. regelii class I diterpene synthase candidates of the TPS-e/f family. Fromleft to right: constructs (selectable markers in gray boxes; Chlor, chloramphenicol; Carb, carbenicillin), GC-MS chromato-grams, and mass spectra of assay products and authentic standards. The following gene combinations were tested: ent-CPS/TrTPS13 (A), ent-CPS/TrTPS14 (B), (+)-CPS/TrTPS13 (C), syn-CPS/TrTPS13 (D), ent-CPS/TrTPS15 (E), (+)-CPS/TrTPS15 (F), andsyn-CPS/TrTPS15 (G).
Plant Physiol. Vol. 175, 2017 99
Diterpenoid Biosynthesis in Tripterygium
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.
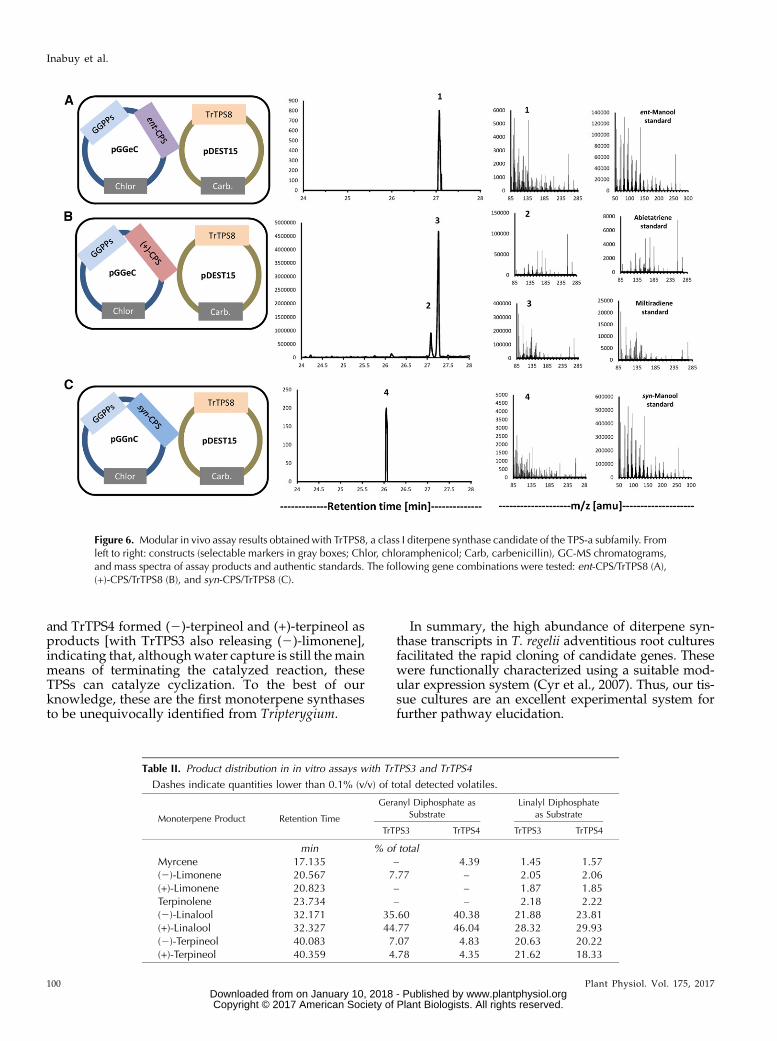
and TrTPS4 formed (2)-terpineol and (+)-terpineol asproducts [with TrTPS3 also releasing (2)-limonene],indicating that, althoughwater capture is still themainmeans of terminating the catalyzed reaction, theseTPSs can catalyze cyclization. To the best of ourknowledge, these are the first monoterpene synthasesto be unequivocally identified from Tripterygium.
In summary, the high abundance of diterpene syn-thase transcripts in T. regelii adventitious root culturesfacilitated the rapid cloning of candidate genes. Thesewere functionally characterized using a suitable mod-ular expression system (Cyr et al., 2007). Thus, our tis-sue cultures are an excellent experimental system forfurther pathway elucidation.
Table II. Product distribution in in vitro assays with TrTPS3 and TrTPS4
Dashes indicate quantities lower than 0.1% (v/v) of total detected volatiles.
Monoterpene Product Retention Time
Geranyl Diphosphate as
Substrate
Linalyl Diphosphate
as Substrate
TrTPS3 TrTPS4 TrTPS3 TrTPS4
min % of totalMyrcene 17.135 – 4.39 1.45 1.57(2)-Limonene 20.567 7.77 – 2.05 2.06(+)-Limonene 20.823 – – 1.87 1.85Terpinolene 23.734 – – 2.18 2.22(2)-Linalool 32.171 35.60 40.38 21.88 23.81(+)-Linalool 32.327 44.77 46.04 28.32 29.93(2)-Terpineol 40.083 7.07 4.83 20.63 20.22(+)-Terpineol 40.359 4.78 4.35 21.62 18.33
Figure 6. Modular in vivo assay results obtainedwith TrTPS8, a class I diterpene synthase candidate of the TPS-a subfamily. Fromleft to right: constructs (selectable markers in gray boxes; Chlor, chloramphenicol; Carb, carbenicillin), GC-MS chromatograms,and mass spectra of assay products and authentic standards. The following gene combinations were tested: ent-CPS/TrTPS8 (A),(+)-CPS/TrTPS8 (B), and syn-CPS/TrTPS8 (C).
100 Plant Physiol. Vol. 175, 2017
Inabuy et al.
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.

Can Adventitious Root Cultures Serve as SustainableResources for the Production of PharmaceuticallyRelevant Diterpenoids?
Cultures of Tripterygium hairy roots, adventitiousroots, or cell suspensions all have the advantage of ac-cumulating abietane diterpenoids at high concentra-tions (Miao et al., 2013; Zhu et al., 2014) and, incomparison with the unsustainable harvest of roots(where the concentrations of these specialized metab-olites are very low), should be considered as commer-cial sources. We demonstrate here that diterpenoids aresecreted into the culture medium of T. regelii adventi-tious root cultures, which significantly reduces thecomplexity of the matrix for extraction. Triptolide isdetected as the peak with the second highest intensity(16% of the total peak area) in our HPLC-QTOF-MSchromatograms of culture medium extracts. Further-more, it is well separated from other medium constit-uents, which enabled a one-step HPLC processing togreater than 95% purity (as judged byHPLC-QTOF-MSand 1H-NMR). In our hands, the production of tripto-lide in adventitious root cultures also has been reliable(grown continuously for more than 2 years) and readilyscalable (from 50- to 500-mL volumes within weeks).Others have scaled up Tripterygium cultures to 10- to20-L bioreactors (Kutney et al., 1992; Miao et al., 2014);therefore, it appears that typical shortcomings of tissueculture (reliability and scalability) have already beenaddressed.It is probable that triptolide levels could be enhanced
further if flux through the abietane diterpenoid path-way was increased in transgenic tissue cultures, wheregenes involved in diterpenoid biosynthesis are alreadyexpressed at fairly high levels (Supplemental Fig. S8).One approach would be the overexpression of genesinvolved in providing diterpenoid precursors. A dif-ferent and potentially complementary approach wouldbe the down-regulation of genes involved in competingpathways (e.g. reducing triterpenoid formation).We are currently investigating methods for the trans-formation of Tripterygium to enable such endeavors.Alternatively, the genes required for triptolide biosyn-thesis, once discovered, could be transferred to anengineered microbial strain, akin to the successful ef-forts to produce another diterpenoid, forskohlin(Pateraki et al., 2017). At this point in time, yields ofhighly functionalized plant diterpenoids in synthetichosts have been relatively low; therefore, plant tissuecultures would seem to be a competitive option fortriptolide production.
MATERIALS AND METHODS
Chemicals
Triptolide and celastrol were purchased from Cayman Chemical. Dehy-droabietic acid was synthesized according to a literature protocol (Gonzálezet al., 2010). Acetone was of HPLC grade (Fisher Scientific), ethyl acetate wasACS quality (Avantor Performance Materials), ethanol was Omnisolv (EMDSerono), and acetonitrile and water were liquid chromatography-mass
spectrometry grade (Sigma-Aldrich). CDCl3 was obtained from CambridgeIsotope Laboratories. All other reagents were purchased from Sigma-Aldrich.
Adventitious Root Cultures
Young Tripterygium regelii plants were purchased from Woodlanders andmaintained under greenhouse conditions (illumination, 16-h day/8-h night[250-500 mE]; temperature, 24°C day/20°C night; relative humidity, 45%–55%).To initiate tissue cultures, leaf material was harvested, rinsed with sterile water,and surface sterilized by soaking in 20% (v/v) commercial bleach. Sterilizedleaves were cut into 1-cm2 pieces and placed onto Murashige and Skoog me-dium with macronutrients and micronutrients plus Gamborg’s vitamins(Caisson), Suc (20 g L21; Sigma-Aldrich), 1-naphthaleneacetic acid (0.2 mg L21;Caisson), 6-benzylaminopurine (1 mg L21; Sigma-Aldrich), and phytagel(2.4 g L21; Sigma-Aldrich), with pH adjusted to 5.8. After 4 to 6 weeks, callusthat developed along the leaf square edges was moved to plates containing rootinduction medium (Murashige and Skoog medium with macronutrients andmicronutrients plus Gamborg’s vitamins, Suc [20 g L21], 1-naphthaleneaceticacid [1 mg L21], 6-benzylaminopurine [0.2 mg L21], and phytagel [2.4 g L21],with pH adjusted to 5.8). Developing adventitious roots were transferred tofresh plates every 4 to 6 weeks until they appeared strong enough for transferto liquid medium (Murashige and Skoog medium with macronutrients andmicronutrients plus Gamborg’s vitamins, Suc [20 g L21], and 1-naphthaleneaceticacid [1mg L21], pH 5.8). Adventitious roots were partially submerged inmedium(50 mL in a 250-mL Erlenmeyer flask) and maintained at 25°C, by shaking at80 rpm, in the dark, with transfer to freshmedium every 2weeks. As adventitiousroots grew, the size of the flask and the volume of liquid medium were adjustedup to 500 mL in a 2-L Fernbach flask.
Metabolite Extraction from Adventitious Root Cultures
Solid materials from adventitious root cultures were washed with water(volume equivalent to thewetweight ofmaterial) and freeze dried for 3 d (Lyph-Lock 12L; LabConco), and the resulting dry matter was stored at 280°C untilfurther use. Aliquots of 506 0.5 mgwere transferred to receptacles of anMM01Dry Mill (Retsch) and further homogenized by ball shaking for 45 s at a rate of20 shakes per second. The homogenate was then transferred to glass test tubeswith Teflon-lined caps. Each sample was extracted four times at 23°Cwith 5mLof acetone for 30 min (Ultrasound Bath FS30H; Fisher Scientific). After eachextraction step, samples were centrifuged at 3,000g for 5 min, and the combinedsupernatants were collected in a separate glass test tube. The solvent was re-moved under reduced pressure (EZ Bio; GeneVac), and the remaining residuewas redissolved in 1mL of 90% acetonitrile containing 10mgmL21 9-anthracenecarboxylic acid as an internal standard. Prior to further analysis, samples werepassed through syringe filters (polytetrafluoroethylene; 0.22-mm pore size) andstored at 4°C for a maximum of 2 d.
A medium aliquot of adventitious root cultures (generally 5 mL) wasextracted twicewith ethyl acetate (2mL each time) by thoroughmixing for 1minat 23°C. The combined organic extracts (4 mL) were extracted against water(2 mL) and transferred to a new glass vial, and the solvent was removed underreduced pressure (EZ Bio; GeneVac). The residue was redissolved in 200 mL of90% aqueous acetonitrile containing 10 mg mL21 9-anthracene carboxylic acidas an internal standard. Prior to further analysis, samples were passed throughsyringe filters (polypropylene; 0.22-mm pore size) and stored at 4°C for amaximum of 2 d.
HPLC-QTOF-MS and MS/MS Data Acquisition andMethod Validation
The separation of metabolites was almost identical to that described previ-ously (Fischedick et al., 2015). The conditions for metabolite separation weremodified slightly: the initial conditions were 70% solvent A (water with 0.1%[v/v] formic acid) and 30% solvent B (acetonitrile with 0.1% [v/v] formic acid).A linear gradient (flow rate, 0.6 mL min21) was used to increase solvent B to80% over 35 min, followed by a more rapid gradient to 95% solvent B at 40 min.The diode array detector was set to record at 219, 254, and 424 nm, with UV/visible spectra being recorded from 200 to 500 nm. Mass spectral data wereobtained based on the protocols outlined previously (Fischedick et al., 2015)with the following differences: the electrospray ionization source was operatedin positive polarity and MS/MS data were acquired with a collision energy of30 eV. Data analysis was performed using the MassHunter software version
Plant Physiol. Vol. 175, 2017 101
Diterpenoid Biosynthesis in Tripterygium
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.

B.03.01 (Agilent Technologies). The approach for validating the quantitation oftriptolide and celastrol was reported previously for various analytes in plantmatrices (Cuthbertson et al., 2013), and only the relevant values are given here:recovery from adventitious roots (n = 3): 85.6% 6 5.6% for triptolide, 96.8% 62.8% for celastrol; recovery from culture medium (n = 3): 106.6% 6 3.9% fortriptolide, 86.1%6 2.9% for celastrol; reproducibility of extraction/detection (asrelative SD; n = 3): 6.5% (interday) and 0.9% (intraday) for triptolide, 0.5%(interday) and 0.6% (intraday) for celastrol; limit of detection at signal-to-noiseratio of 1: 5 (n = 3): 0.01 ng for triptolide (MS detection) and 1 ng for celastrol(diode array detection at 424 nm); limit of quantitation at signal-to-noise ratio of1: 10 (n= 3): 0.05 for triptolide (MS detection) and 10 ng for celastrol (diode arraydetection at 424 nm); regression equation for calibration curve (n = 3): y =25,938x + 16,475x (R2 = 0.989) for triptolide (corrected for matrix effects) and y =0.9077x (R2 = 0.999) for celastrol (matrix effects irrelevant for diode array de-tection); and linear range, 0.05 to 50 ng for triptolide and 1 to 2,500 ng forcelastrol.
Cloning and Functional Characterization ofTerpene Synthases
Raw data from several transcriptome sequencing projects performed pre-viously with T. regelii tissues (National Center for Biotechnology InformationShort Read Archive accession no. SRP075639 [adventitious root cultures]) anddata sets available at http://www.medplantrnaseq.org/ (roots and root cul-tures) were downloaded, and a consensus assembly was generated using theTrinity (Haas et al., 2013) and TransABySS (Robertson et al., 2010) assemblers.tBLASTn searches with peptide sequences of characterized class I and class IIditerpene synthases (Supplemental Table S2) were performed against theT. regelii consensus assembly. RNAwas isolated from T. regelii adventitious rootcultures (harvested at 10 d after transfer to fresh medium) using the Plant RNAPurification reagent (Invitrogen) and the RNEasy Mini Kit (Qiagen) accordingto each manufacturer’s instructions, converted to cDNA usingMaxima ReverseTranscriptase (Thermo Fisher Scientific), and PCR with gene-specific primerswas then employed to clone candidate diterpene synthase sequences (details inSupplemental Table S3). RACE-PCR was performed according to the manu-facturer’s instructions (SMARTer-RACE 59/39 kit; Clontech/TaKaRa). TheGateway cloning system (Invitrogen) was used to insert TrTPS1 and TrTPS2into a previously described pGG-DEST vector, and the other putative diterpenesynthases into pDEST15, for functional expression in the Escherichia coli C41(DE3) (Cyr et al., 2007). Diterpenoids were extracted directly from 50-mL cul-tureswith an equal volume of n-hexanes (Fisher Scientific). The extract was thenrun through silica gel 60 and magnesium sulfate columns as described else-where (Cyr et al., 2007). The eluents were dried under a flow of nitrogen, andthe residue was dissolved in 200 mL of n-hexanes. Aliquots (1 mL) were injectedonto an HP-5MS column (30-m length 3 0.25-mm diameter, 0.25-mm filmthickness; J&W Scientific, distributed through Agilent) of a 6890N gas chro-matograph (operated in splitless mode) coupled to a 5973 inert mass selectivedetector (Agilent). Settings were as follows: helium as carrier gas at a flow rateof 1 mL min21; inlet temperature set to 250°C; oven program with 50°C for1 min, first linear gradient to 300°C at 7°C min21, second linear gradient to330°C at 20°C min21, and final hold of 5 min; transfer line set to 180°C; electronimpact spectra recorded at 70 eV with MS data collection from mass-to-chargeratio of 50 to 450. Peakswere identified based on comparisons of retention timesand MS fragmentation patterns with those of authentic standards. The func-tional evaluation of monoterpene synthases was performed according toSrividya et al. (2015).
Accession Numbers
All genes characterized as part of this study have been deposited inGenBankwith the following accession numbers: KX533964 (TrTPS2), KX533965 (TrTPS1),KX533966 (TrTPS13), KX533967 (TrTPS14), KX533968 (TrTPS15), KY856993(TrTPS3), KY856994 (TrTPS4), KY856995 (TrTPS8), and KY856996 (TrTPS7).
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. Accumulation of abietane diterpenoids in themedium of T. regelii adventitious root cultures.
Supplemental Figure S2. 1H-NMR spectra of abietane diterpenoids.
Supplemental Figure S3. Sequences of T. regelii diterpene synthases of theTPS-c family.
Supplemental Figure S4. Sequences of T. regelii diterpene synthases ofthe TPS-e/f family.
Supplemental Figure S5. Sequences of T. regelii diterpene synthases of theTPS-b family.
Supplemental Figure S6. Modular in vivo combination of T. regelii class II(TrTPS2) and class I (TrTPS13) diterpene synthases.
Supplemental Figure S7. Proposed mechanism for T. regelii monoterpenesynthases.
Supplemental Figure S8. Comparison of expression levels of candidatediterpene synthase genes in T. regelii adventitious root cultures and rootsbased on RNA sequence by expectation-maximization analysis.
Supplemental Figure S9. Alignment of diterpene synthase sequences in-cluded in the phylogenetic analysis.
Supplemental Table S1. Changes in triptolide and celastrol concentrationsin T. regelii adventitious root cultures following various elicitor treat-ments.
Supplemental Table S2. List of dicot diterpene synthase protein sequencesused in the phylogenetic analysis.
Supplemental Table S3. List of primers used in T. regelii terpene synthasegene cloning.
Supplemental Methods and Data File S1. Purification and characteriza-tion of diterpenoids from T. regelii adventitious root cultures.
ACKNOWLEDGMENTS
We thank Sean R. Johnson for generating sequence assemblies with publiclyavailable transcriptome data sets, Washington State University’s NMR CoreFacility for access to instruments, and Dr. Greg Helms for expert support.
Received May 17, 2017; accepted July 18, 2017; published July 27, 2017.
LITERATURE CITED
Andersen-Ranberg J, Kongstad KT, Nielsen MT, Jensen NB, Pateraki I,Bach SS, Hamberger B, Zerbe P, Staerk D, Bohlmann J, et al (2016)Expanding the landscape of diterpene structural diversity throughstereochemically controlled combinatorial biosynthesis. Angew ChemInt Ed Engl 55: 2142–2146
Brinker AM, Ma J, Lipsky PE, Raskin I (2007) Medicinal chemistry andpharmacology of genus Tripterygium (Celastraceae). Phytochemistry68: 732–766
Burke C, Croteau R (2002) Interaction with the small subunit of geranyldiphosphate synthase modifies the chain length specificity of ger-anylgeranyl diphosphate synthase to produce geranyl diphosphate. JBiol Chem 277: 3141–3149
Chen F, Tholl D, Bohlmann J, Pichersky E (2011) The family of terpenesynthases in plants: a mid-size family of genes for specialized metabo-lism that is highly diversified throughout the kingdom. Plant J 66: 212–229
Chen YZ, Gao Q, Zhao XZ, Chen XM, Zhang F, Chen J, Xu CG, Sun LL,Mei CL (2010) Meta-analysis of Tripterygium wilfordii Hook F in theimmunosuppressive treatment of IgA nephropathy. Intern Med 49:2049–2055
Chugh R, Sangwan V, Patil SP, Dudeja V, Dawra RK, Banerjee S,Schumacher RJ, Blazar BR, Georg GI, Vickers SM, et al (2012) Apreclinical evaluation of Minnelide as a therapeutic agent against pan-creatic cancer. Sci Transl Med 4: 156ra139
Cuthbertson DJ, Johnson SR, Piljac-�Zegarac J, Kappel J, Schäfer S, WüstM, Ketchum RE, Croteau RB, Marques JV, Davin LB, et al (2013) Ac-curate mass-time tag library for LC/MS-based metabolite profiling ofmedicinal plants. Phytochemistry 91: 187–197
Cyr A, Wilderman PR, Determan M, Peters RJ (2007) A modular approachfor facile biosynthesis of labdane-related diterpenes. J Am Chem Soc129: 6684–6685
102 Plant Physiol. Vol. 175, 2017
Inabuy et al.
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.

Duan H, Takaishi Y, Momota H, Ohmoto Y, Taki T, Jia Y, Li D (1999)Immunosuppressive diterpenoids from Tripterygium wilfordii. J NatProd 62: 1522–1525
Duan HQ, Takaishi Y, Momota H, Ohmoto Y, Taki T, Tori M, Takaoka S,Jia YF, Li D (2001) Immunosuppressive terpenoids from extracts ofTripterygium wilfordii. Tetrahedron 57: 8413–8424
Felsenstein J (1985) Confidence limits on phylogenies: an approach usingthe bootstrap. Evolution 39: 783–791
Fischedick JT, Johnson SR, Ketchum REB, Croteau RB, Lange BM (2015)NMR spectroscopic search module for Spektraris, an online resource forplant natural product identification: taxane diterpenoids from Taxus 3media cell suspension cultures as a case study. Phytochemistry 113:87–95
Gao W, Hillwig ML, Huang L, Cui G, Wang X, Kong J, Yang B, Peters RJ(2009) A functional genomics approach to tanshinone biosynthesisprovides stereochemical insights. Org Lett 11: 5170–5173
Geisler K, Jensen NB, Yuen MM, Madilao L, Bohlmann J (2016) Modu-larity of conifer diterpene resin acid biosynthesis: P450 enzymes ofdifferent CYP720B clades use alternative substrates and converge on thesame products. Plant Physiol 171: 152–164
González MA, Pérez-Guaita D, Correa-Royero J, Zapata B, Agudelo L, Mesa-Arango A, Betancur-Galvis L (2010) Synthesis and biological evaluation ofdehydroabietic acid derivatives. Eur J Med Chem 45: 811–816
Guo L, Duan L, Liu K, Liu EH, Li P (2014) Chemical comparison ofTripterygium wilfordii and Tripterygium hypoglaucum based onquantitative analysis and chemometrics methods. J Pharm BiomedAnal 95: 220–228
Haas BJ, Papanicolaou A, Yassour M, Grabherr M, Blood PD, Bowden J,Couger MB, Eccles D, Li B, Lieber M, et al (2013) De novo transcriptsequence reconstruction from RNA-seq using the Trinity platform forreference generation and analysis. Nat Protoc 8: 1494–1512
Hamberger B, Ohnishi T, Hamberger B, Séguin A, Bohlmann J (2011)Evolution of diterpene metabolism: Sitka spruce CYP720B4 catalyzesmultiple oxidations in resin acid biosynthesis of conifer defense againstinsects. Plant Physiol 157: 1677–1695
Hansen NL, Heskes AM, Hamberger B, Olsen CE, Hallström BM, Andersen-Ranberg J, Hamberger B (2017) The terpene synthase gene family in Trip-terygium wilfordii harbors a labdane-type diterpene synthase among themonoterpene synthase TPS-b subfamily. Plant J 89: 429–441
Helmstädter A (2013) Tripterygium wilfordii Hook. f.: how a traditionalTaiwanese medicinal plant found its way to the West. Pharmazie 68:643–646
Jones DT, Taylor WR, Thornton JM (1992) The rapid generation of mu-tation data matrices from protein sequences. Comput Appl Biosci 8: 275–282
Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular EvolutionaryGenetics Analysis version 7.0 for bigger datasets. Mol Biol Evol 33:1870–1874
Kutney JP, Han K (1996) Studies with plant-cell cultures of the Chineseherbal plant, Tripterygium wilfordii: isolation and characterization ofditerpenes. Recl Trav Chim Pays Bas 115: 77–93
Kutney JP, Hewitt GM, Kurihara T, Salisbury PJ, Sindelar RD, Stuart KL,Townsley PM, Chalmers WT, Jacoli GG (1981) Cyto-toxic diterpenestriptolide, tripdiolide, and cyto-toxic triterpenes from tissue-cultures ofTripterygium wilfordii. Can J Chem 59: 2677–2683
Kutney JP, Hewitt GM, Lee G, Piotrowska K, Roberts M, Rettig SJ (1992)Studies with tissue-cultures of the Chinese herbal plant, Tripterygium-wilfordii: isolation of metabolites of interest in rheumatoid-arthritis,immunosuppression, and male contraceptive activity. Can J Chem 70:1455–1480
Kutney JP, Samija MD, Hewitt GM, Bugante EC, Gu H (1993) Anti-inflammatory oleanane triterpenes from Tripterygium wilfordii cellsuspension cultures by fungal elicitation. Plant Cell Rep 12: 356–359
Lange BM, Fischedick JT, Lange MF, Srividya N, Šamec D, Poirier BC (2017)Integrative approaches for the identification and localization of specializedmetabolites in Tripterygium roots. Plant Physiol 173: 456–469
Liu YF, Tu SH, Gao WN, Wang Y, Liu PL, Hu YH, Dong H (2013) Extractsof Tripterygium wilfordii Hook F in the treatment of rheumatoid ar-thritis: a systemic review and meta-analysis of randomised controlledtrials. Evid Based Complement Alter Med 2013: 410793
Meng C, Zhu H, Song H, Wang Z, Huang G, Li D, Ma Z, Ma J, Qin Q, SunX, et al (2014) Targets and molecular mechanisms of triptolide in cancertherapy. Chin J Cancer Res 26: 622–626
Miao GP, Zhu CS, Feng JT, Han J, Song XW, Zhang X (2013) Aggregate cellsuspension cultures of Tripterygium wilfordii Hook f for triptolide, wilfor-gine, and wilforine production. Plant Cell Tissue Organ Cult 112: 109–116
Miao GP, Zhu CS, Yang YQ, Feng MX, Ma ZQ, Feng JT, Zhang X (2014)Elicitation and in situ adsorption enhanced secondary metabolites pro-duction of Tripterygium wilfordii Hook. f. adventitious root fragmentliquid cultures in shake flask and a modified bubble column bioreactor.Bioprocess Biosyst Eng 37: 641–650
Nakano K, Oose Y, Takaishi Y (1997) A novel epoxy-triterpene and nor-triterpene from callus cultures of Tripterygium wilfordii. Phytochemis-try 46: 1179–1182
Nakano K, Yoshida C, Furukawa W, Takaishi Y, Shishido K (1998) Ter-penoids in transformed root culture of Tripterygium wilfordii. Phyto-chemistry 49: 1821–1824
Pateraki I, Andersen-Ranberg J, Jensen NB, Wubshet SG, Heskes AM,Forman V, Hallström B, Hamberger B, Motawia MS, Olsen CE, Staerk D,Hansen J, et al (2017) Total biosynthesis of the cyclic AMP booster forskolinfrom Coleus forskohlii. eLife 6: e23001
Ro DK, Arimura G, Lau SYW, Piers E, Bohlmann J (2005) Loblolly pineabietadienol/abietadienal oxidase PtAO (CYP720B1) is a multifunc-tional, multisubstrate cytochrome P450 monooxygenase. Proc Natl AcadSci USA 102: 8060–8065
Robertson G, Schein J, Chiu R, Corbett R, Field M, Jackman SD, MungallK, Lee S, Okada HM, Qian JQ, et al (2010) De novo assembly andanalysis of RNA-seq data. Nat Methods 7: 909–912
Srividya N, Davis EM, Croteau RB, Lange BM (2015) Functional analysisof (4S)-limonene synthase mutants reveals determinants of catalyticoutcome in a model monoterpene synthase. Proc Natl Acad Sci USA 112:3332–3337
Tanaka N, Ooba N, Duan H, Takaishi Y, Nakanishi Y, Bastow K, Lee KH(2004) Kaurane and abietane diterpenoids from Tripterygium doianum(Celastraceae). Phytochemistry 65: 2071–2076
Yamaguchi S, Sun Tp, Kawaide H, Kamiya Y (1998) The GA2 locus ofArabidopsis thaliana encodes ent-kaurene synthase of gibberellin bio-synthesis. Plant Physiol 116: 1271–1278
Zeng F, Wang W, Guan S, Cheng C, Yang M, Avula B, Khan IA, Guo DA(2013) Simultaneous quantification of 18 bioactive constituents in Trip-terygium wilfordii using liquid chromatography-electrospray ionization-mass spectrometry. Planta Med 79: 797–805
Zerbe P, Hamberger B, Yuen MM, Chiang A, Sandhu HK, Madilao LL,Nguyen A, Hamberger B, Bach SS, Bohlmann J (2013) Gene discoveryof modular diterpene metabolism in nonmodel systems. Plant Physiol162: 1073–1091
Zhou ZL, Yang YX, Ding J, Li YC, Miao ZH (2012) Triptolide: structuralmodifications, structure-activity relationships, bioactivities, clinical de-velopment and mechanisms. Nat Prod Rep 29: 457–475
Zhu B, Wang Y, Jardine M, Jun M, Lv JC, Cass A, Liyanage T, Chen HY,Wang YJ, Perkovic V (2013) Tripterygium preparations for the treat-ment of CKD: a systematic review and meta-analysis. Am J Kidney Dis62: 515–530
Zhu C, Miao G, Guo J, Huo Y, Zhang X, Xie J, Feng J (2014) Establishmentof Tripterygium wilfordii Hook. f. hairy root culture and optimization ofits culture conditions for the production of triptolide and wilforine. JMicrobiol Biotechnol 24: 823–834
Zi J, Mafu S, Peters RJ (2014) To gibberellins and beyond! Surveying theevolution of (di)terpenoid metabolism. Annu Rev Plant Biol 65: 259–286
Zi J, Peters RJ (2013) Characterization of CYP76AH4 clarifies phenolic di-terpenoid biosynthesis in the Lamiaceae. Org Biomol Chem 11: 7650–7652
Plant Physiol. Vol. 175, 2017 103
Diterpenoid Biosynthesis in Tripterygium
www.plantphysiol.orgon January 10, 2018 - Published by Downloaded from Copyright © 2017 American Society of Plant Biologists. All rights reserved.