biophysik der zelle - physik.uni-muenchen.de · biological energy energy consumption of humans:...
TRANSCRIPT

03.06.EnergieumwandlungMitochondrien, Atmungskette, Michell Hypothese, ATP Synthase
ATP Synthese
BIOPHYSIK DER ZELLE

Important Concepts and Terms
• ATP- Hydrolysis• Glykolysis• Oxidative Phosphorylierung
Elektronentransfer-Chain• Mitchell Hypothesis• ATP Synthase

Biological Energyenergy consumption of humans:
25-800 kcal/h or 2x60 Watt
Energie production is related to respiration / burning of sugar.
The energy units in cells is ATP ≈ 25kT.
The daily ATP production is 140 kg !!!

Merken:
1kBT300K = 25 meV = 4 pNnm = 0.6 kcal/Mol = 2.5 kJ/Mol
ThermalEnergy Scale(kBT300K) 1 10 100 1000
Myosinstroke
Molecularrecognition

6CO2 + 6H2O C6H12O6 + 6 O2
C6H12O6 + 6 O2 6CO2 + 6H2O + (30ATP energy)
C6H12O6 C2H5OH + 2CO2 + (2ATP energy)
Photosynthesis
aerobicrespiration
anaerobic respiration

Wasserspaltung, die erste ökologische Katastrophe der Erde
∆Sglobal > 0
∆Sglobal > 02. HS Thdyn.
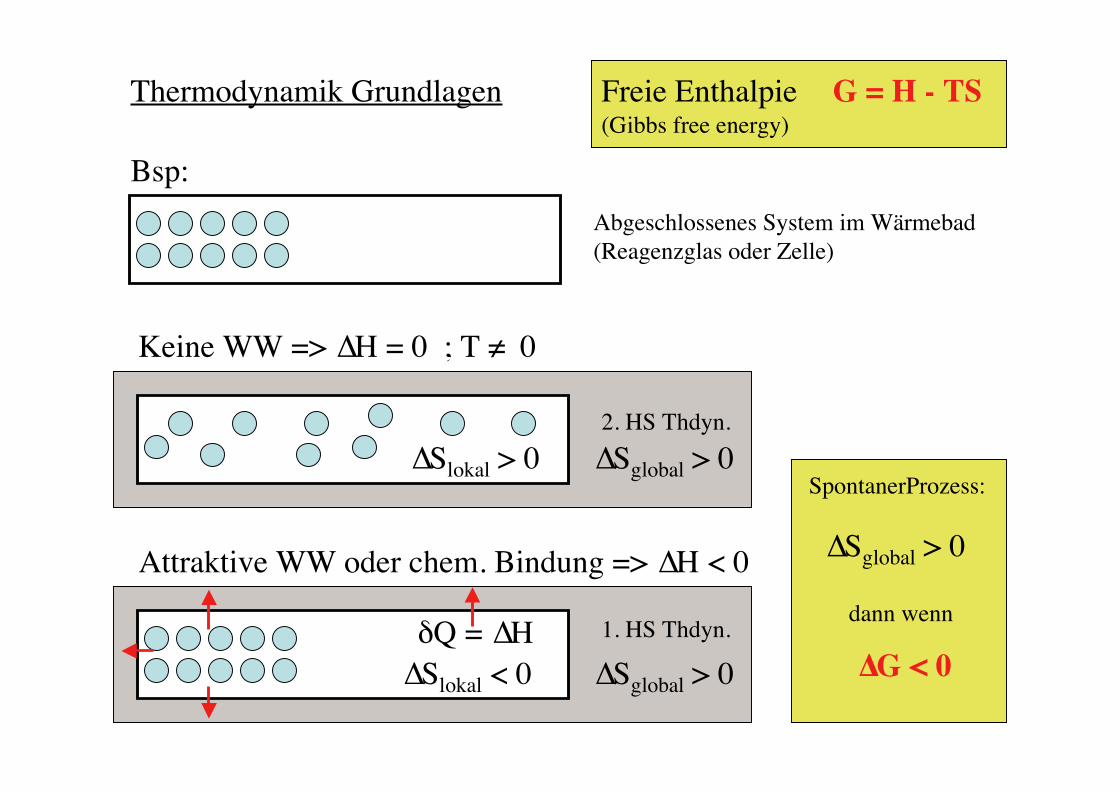
Thermodynamik Grundlagen Freie Enthalpie(Gibbs free energy)
G = H - TS
Keine WW => ∆H = 0 ; T ≠ 0
∆Slokal > 0
Attraktive WW oder chem. Bindung => ∆H < 0
∆Slokal < 0
Abgeschlossenes System im Wärmebad(Reagenzglas oder Zelle)
Bsp:
SpontanerProzess:
∆Sglobal > 0
dann wenn
∆G < 0δQ = ∆H 1. HS Thdyn.

Thermodynamik Grundlagen Chemisches Potentialµ = ∂G/∂ n
Phase 1
Phasengleichgewicht
Bsp:
Phase 2
Gleichgewicht => µ1 = µ2
Für verdünnte Lösungen gilt:
Herleitung Biophysik 2
µLösung = µo + RT ln c
≈ ∂H / ∂ n
hand waiving:
Ω ≈ 1/cln c ≈ -ln Ω ≈ -S
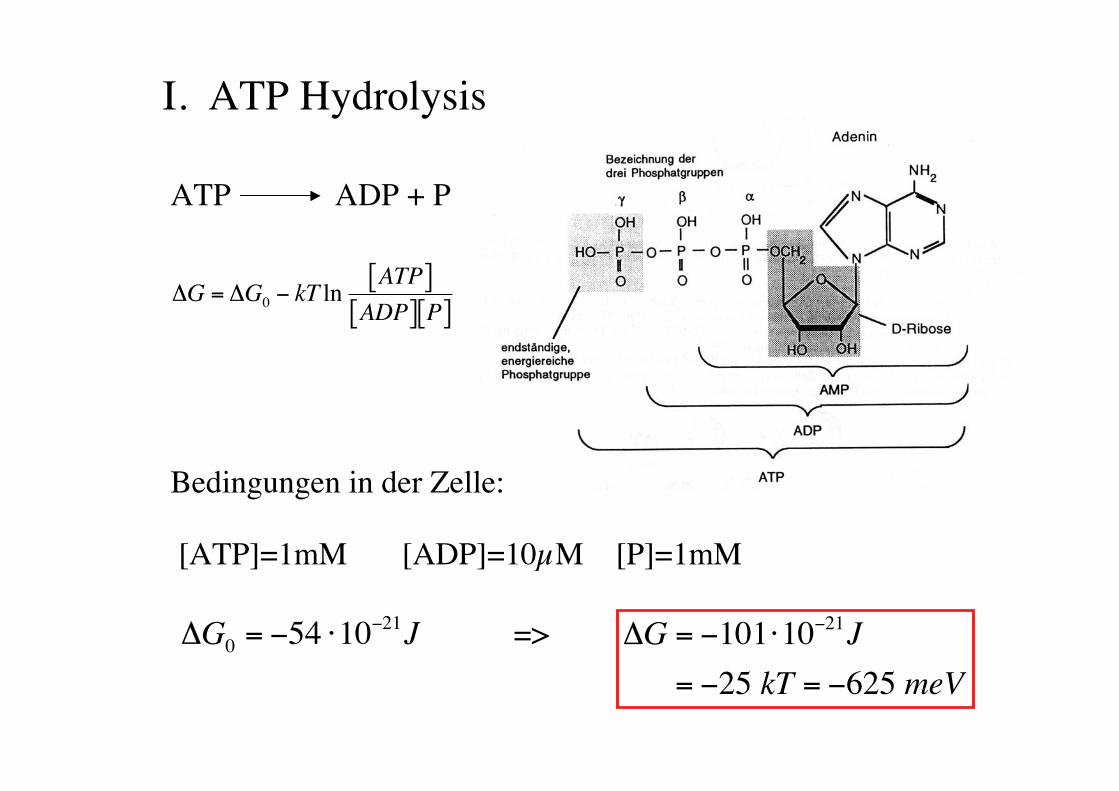
I. ATP Hydrolysis
ATP ADP + P
!
"G = "G0# kT ln
ATP[ ]ADP[ ] P[ ]
!
"G0
= #54 $10#21J
Bedingungen in der Zelle:
[ATP]=1mM [ADP]=10µM [P]=1mM
=>
!
"G = #101$10#21J
= #25 kT = #625 meV
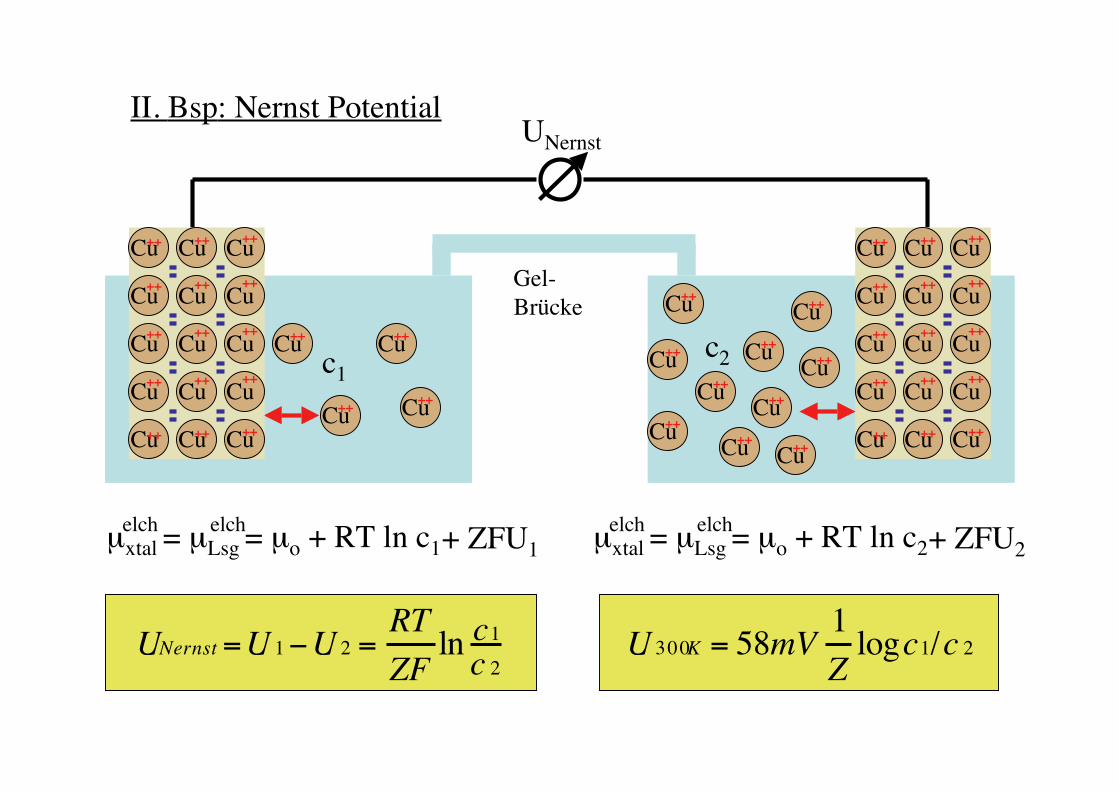
II. Bsp: Nernst Potential
Gel-Brücke
Cu Cu
CuCu
c1
µxtal = µLsg = µo + RT ln c1
Cu Cu Cu
Cu Cu Cu
Cu Cu Cu
Cu Cu Cu
Cu Cu Cu
elch elch + ZFU1
++++++
++++++
++++++
++++++
++++++
++ ++
++++
µxtal = µLsg = µo + RT ln c2elch elch + ZFU2
Cu Cu Cu
Cu Cu Cu
Cu Cu Cu
Cu Cu Cu
Cu Cu Cu
++++++
++++++
++++++
++++++
++++++
Cu++
Cu++
Cu++
Cu++
Cu++
Cu++
Cu++
Cu++Cu++
Cu++c2
!
UNernst =U 1"U 2 =RT
ZFlnc1
c 2
UNernst
!
U 300K = 58mV1
Zlogc1/c 2

Technische Anwendungdes Nernst Potentials:
pH Elektrode

Redoxreaktionen
H
e-H
O H
HO
H
H
O
Elektronentransferreaktionen spielen eine zentrale Rolle inlebenden Systemen z.B. Photosynthese, Metabolismus.
Aber: Lebensdauer des freien Elektrons in wässrigerUmgebung in Solvathülle nur ≈ 10-3 sec => Elektron Zwischenspeicher nötig
Allgemein:
!
Dred
+ Aox"D
ox+ A
red= D
++ A
#
!
D " e"# D
+
!
A + e"# A
"
k
Teilreaktionen:Oxidation(Elektronen-Abgabe)
Reduktion(Elektronen-Aufnahme)
Bsp Lactat Metabolismus:
O
C—C—C H||
O
O
-
3 +
H H
N
R
CONH2
++
H
H
OH
C—C—CH|
|O
O
-
3 +
+
R
CONH2
Lactat NAD Pyruvat NADH
H
N+


RedoxpotentialMass für die Tendenz eines Moleküls, Elektronen abzugeben, bzw. aufzunehmen
!
Kredox
=D
ox[ ] e"[ ]D
red[ ]
!
U ="G
0
F#RT
Fln
Dox
[ ]Dred[ ]
Gleichgewichts-Potential(vergl. Nernst)
!
"Gredox
= "G0+ RT ln
Dox[ ]
Dred[ ]
=> Bezogen auf 1 Mol
!
Gel
= "FUZur Umsetzung 1 Mols gegen das elektrischesPotential U benötigte Energie
!
Dox[ ] = D
red[ ] = 1mol
l
Def:
Redoxpotential ist dieSpannung, bei der gilt:

Redoxpotential Beispielereduzierte Form oxidierte Form Zahl der e- Uo (Volt)
H2 2 H+ + 2 e- 2 -0.42
NADH NAD+ + H+ + 2 e- 2 -0.32
NADPH NADP+ + H+ + 2 e- 2 -0.32
Ubichinon Ubihydrochinon 2 +0.1
2 Cytochrom bred 2 Cytochrom box,+ + 2 e- 2 +0.03
2 Cytochrom cred 2 Cytochrom cox,+ + 2 e- 2 +0.234
2 Cytochrom aa3red 2 Cytochrom aa3ox,+ + 2 e- +0.385
H2O 1/2 O2 + 2 H+ + 2 e- 2 +0.816
Standardwerte der Redoxpotentiale einiger wichtiger Partner der Elektronentransferkette, bezogenauf pH 7, T=25o C. Bei diesem pH ist das Potential der H2-Reduktion um 0.42 eV zu negativerenWerten verschoben.
Abkürzungen:NADH Nicotinamid-Adenin-DinucleotidNADPH: Nicotinamid-Adenin Dinucleotid-Phosphat.

III. The protomotiv force (pmf)
≈ 140 mV
≈ 60 mV
≈ 200 mV ≈ 8kBT pro H+
!
"µ(H +) = e#$ kT % "pH
!
pmf = "µ(H +) e = #$ kT e % "pH

The universal trick in energy conversion is couplingof reactions
Mitchell Hypothesis:„chemiosmotic coupling“

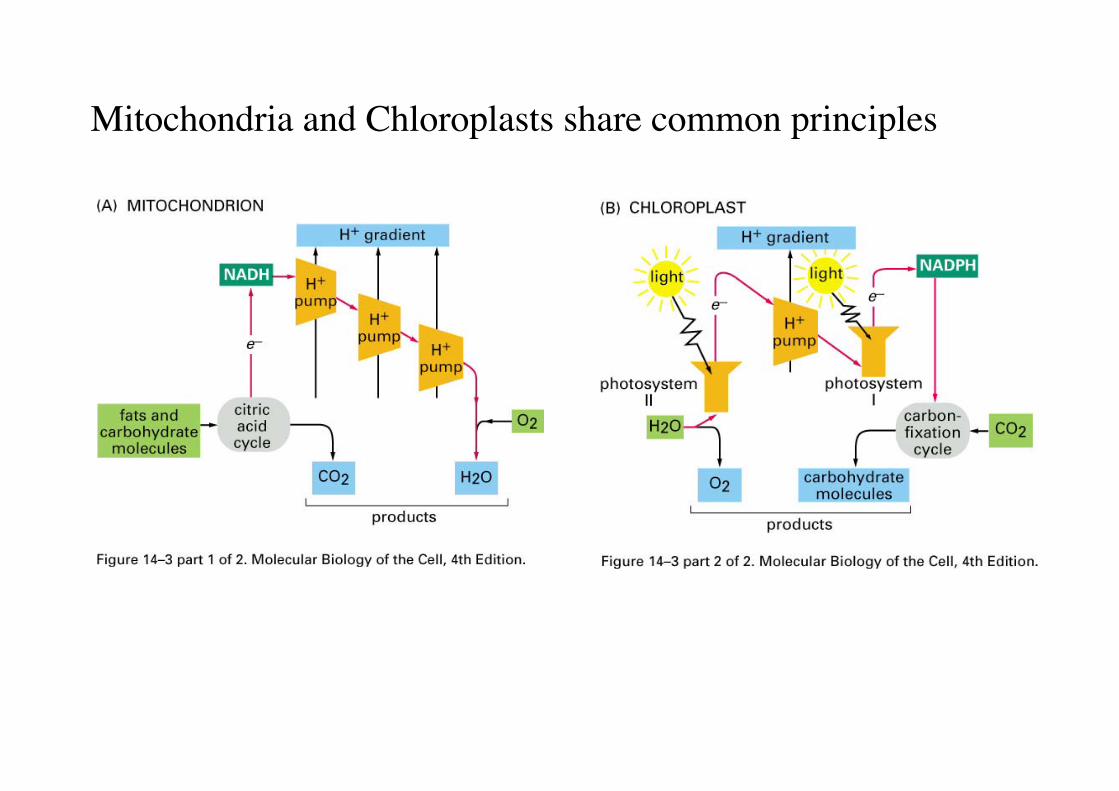
Mitochondria and Chloroplasts share common principles

Electron transport and oxidative phosphorylation
Mitchell Hypothese: Mitchell proposed, that ATP production andthe respiratory Elektronentransferchain is indirectly coupled by aproton gradient (chemiosmotic mechanism)

Electron transport in mitochondria is coupled to protontranslocation

Experiments withmembrane vesicles supportthe chemiosmoticmechanism of ATPformation


Bacteria, mitochondria and chloroplasts all usechemiosmosis to generate ATP

The energy conversion machinery uses common energy intermediates for generation and distribution

Membrane orientationand the direction ofproton movement inbacteria, mitochondria,and chloroplasts

Mitochondria possess two structurally and functionallydistinct membranes

Cytosolic enzymes convert glucose to pyruvate (steps 4-9)


Themitochondrialinner membraneand matrix arethe sites of mostreactionsinvolving theoxidation ofpyruvate and thecoupledsynthesis ofATP

Glycolysis

Oxidation of glucose and fatty acids to CO2
• Glycolysis is the initial stage of glucose metabolism -- it takes place in thecytosol and does not involve O2
• Glycolysis produces a small amount of ATP and the 3-carbon compoundpyruvate
• In aerobic cells, the pyruvate is transported into mitochondria, where it isoxidized by O2 to CO2 to generate even more ATP

Cytosolic enzymes convert glucose to pyruvate (steps 1-3)

Cytosolic enzymes convert glucose to pyruvate (steps 4-9)
Substrate-level phosphorylation generates ATP during glycolysis

During glycolysis, 2 protons and 4 electrons are transferredto 2 molecules of NAD+ to produce 2 molecules of NADH
NicotinamideAdeninDinucleotide

NADH works as Elektrondonor

Anaerobic metabolism of each glucose molecule yieldsonly two ATP molecules

Mitochondrial oxidation of pyruvate begins with theformation of acetyl CoA

Oxidative Phosphorylation

The multienzyme complex pyruvate dehydrogenase convertspyruvate and coenzyme A into CO2 and acetyl CoA

Oxidation of theacetyl group of acetylCoA in the citric acidcycle yields CO2 andreduced coenzymes

A summary of the reactions of glycolysis and thecitric acid cycle
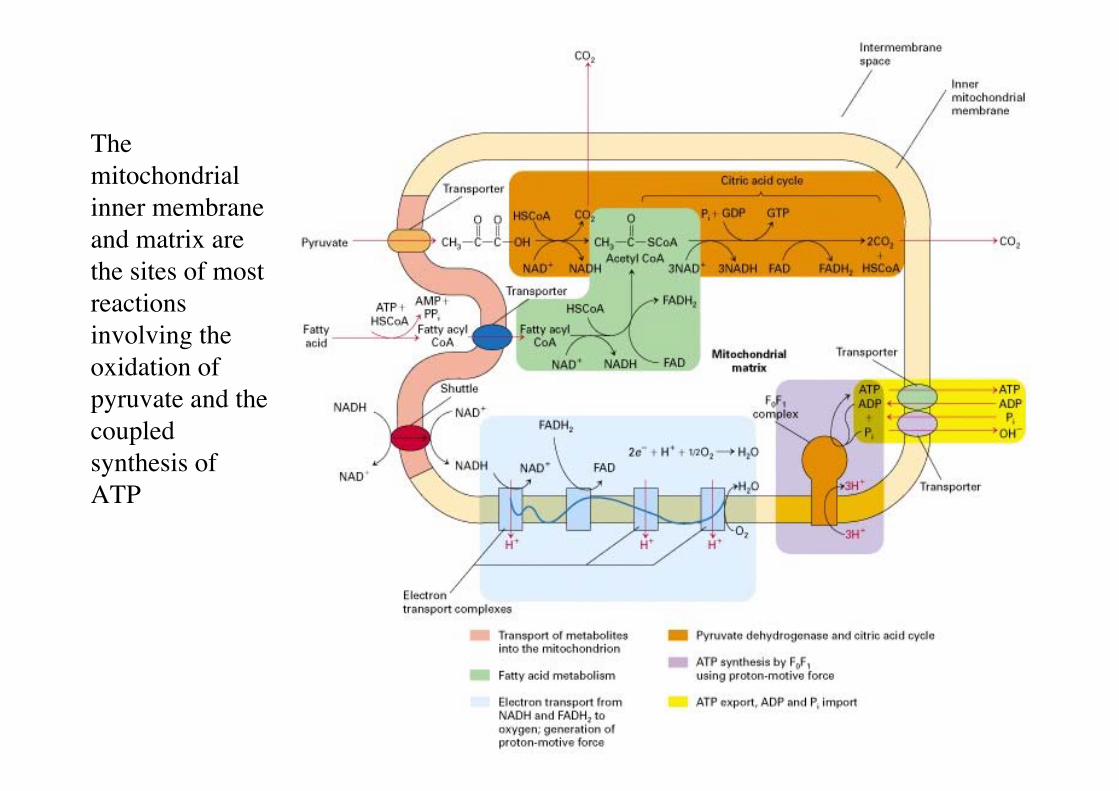
Themitochondrialinner membraneand matrix arethe sites of mostreactionsinvolving theoxidation ofpyruvate and thecoupledsynthesis ofATP

Inner-membraneproteins allow theuptake ofelectrons fromcytosolic NADH
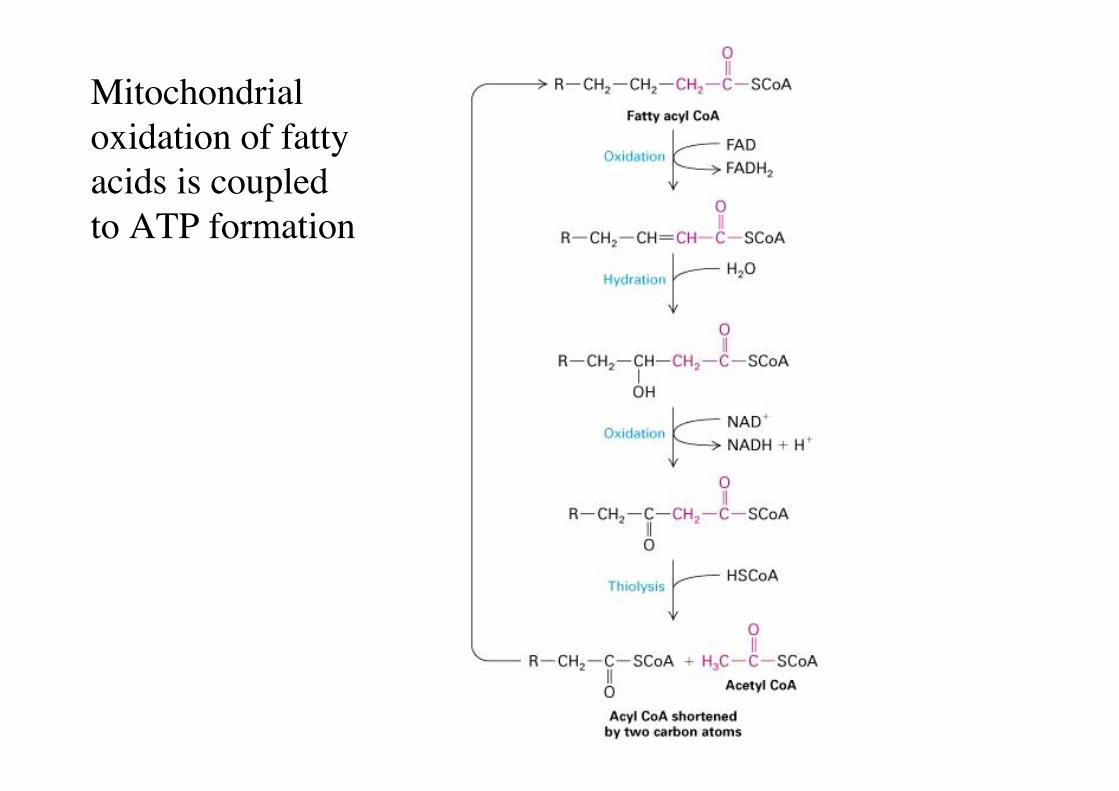
Mitochondrialoxidation of fattyacids is coupledto ATP formation
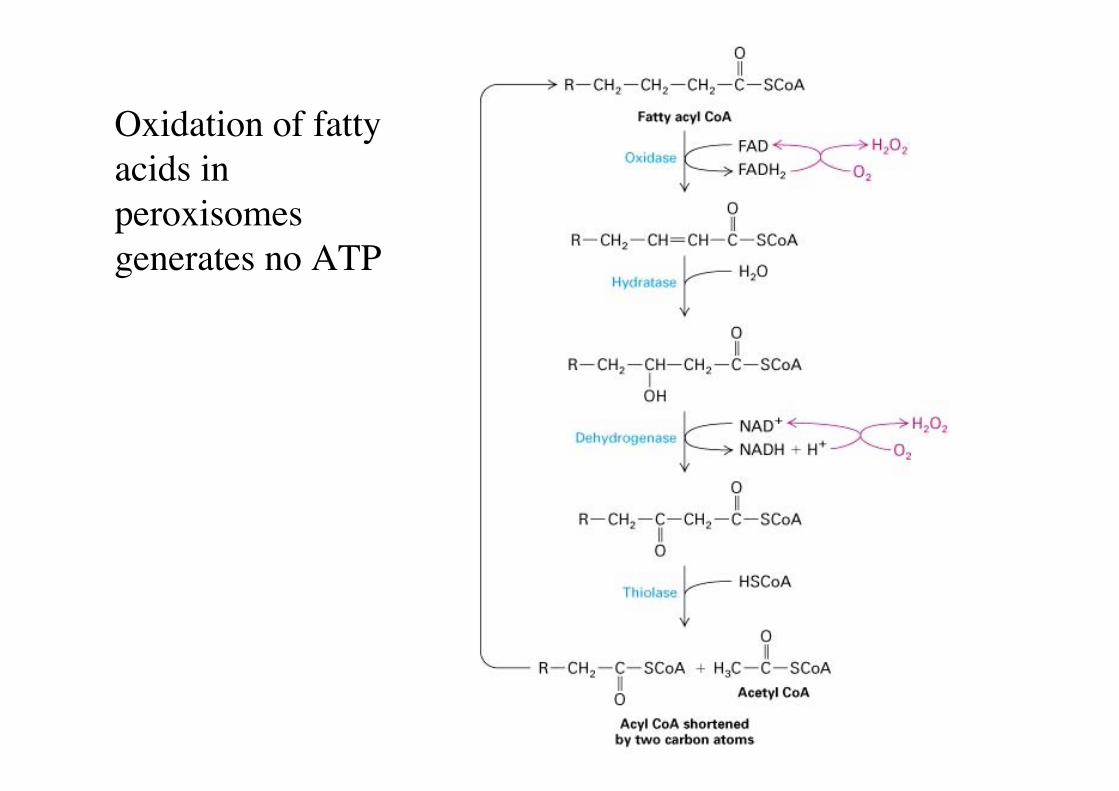
Oxidation of fattyacids inperoxisomesgenerates no ATP
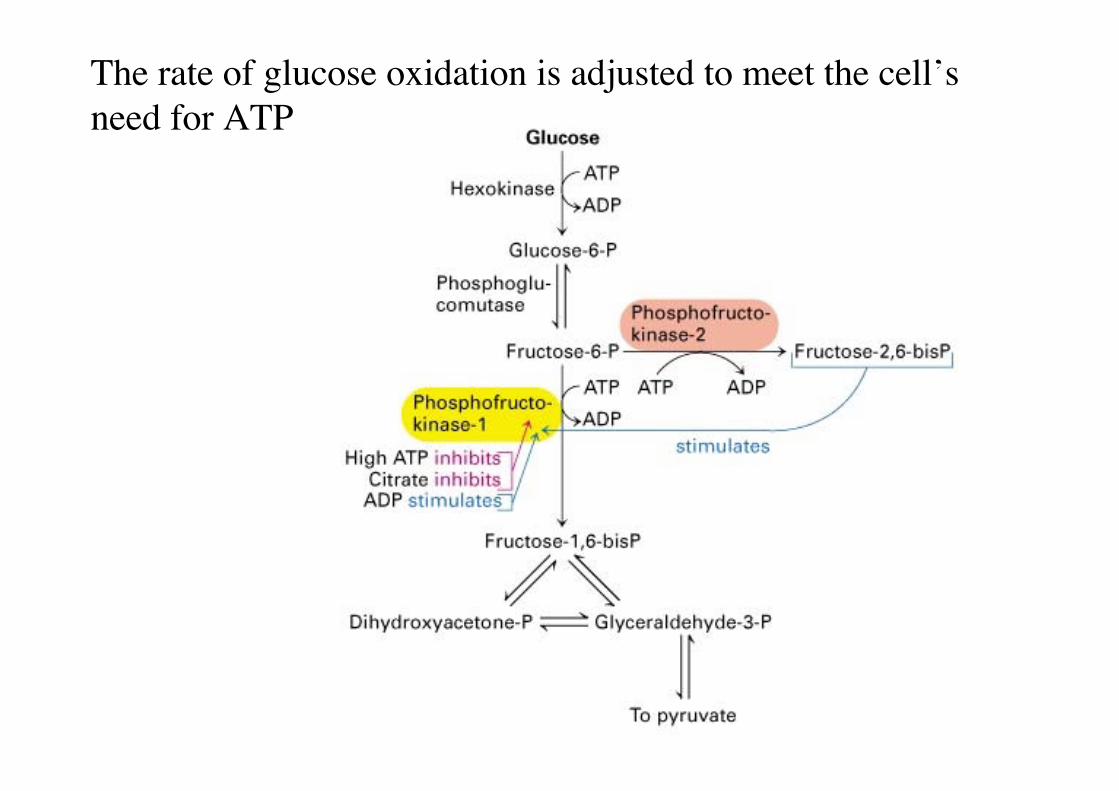
The rate of glucose oxidation is adjusted to meet the cell’sneed for ATP


Electron transport and oxidative phosphorylation
• Most of the free energy released during oxidation of glucose to CO2 is retained inNADH and FADH2
• During respiration, electrons are released from NADH and FADH2 andeventually are transferred to O2 (forming H2O)
• The step-by-step transfer of electrons via the electron transport chain allows thelarge amount of free energy produced by the transfer of electrons to O2 to bereleased in small increments
• Several electron transport chain components convert these small increments intoa proton and voltage gradient across the inner membrane
• The movement of protons down their electrochemical gradient drives thesynthesis of ATP from ADP and Pi