biofilm control with antimicrobial agents · de hidrogénio, cloreto de benzalcónio (bac), cloreto...
TRANSCRIPT

BIOFILM CONTROL WITH ANTIMICROBIAL AGENTS: THE ROLE OF THE EXOPOLYMERIC MATRIX
A Dissertation presented to the UNIVERSITY OF PORTO
for the degree of Doctor in Chemical and Biological Engineering
by
Paula Alexandra da Silva Araújo
Supervisor: Professor Manuel Simões Co-supervisor: Professor Filipe Mergulhão
LEPABE – Laboratory for Process, Environment, Biotechnology and Energy Engineering
Department of Chemical Engineering Faculty of Engineering, University of Porto
June, 2014

II

III
“We live in an island surrounded by a sea of ignorance.
As our island of knowledge grows, so does the shore of our ignorance.”
John Archibald Wheeler (1992)

IV

V
“Ser poeta é ser mais alto, é ser maior
Do que os homens! Morder como quem beija!
É ser mendigo e dar como quem seja
Rei do Reino de Aquém e de Além Dor!
É ter de mil desejos o splendor
E não saber sequer que se deseja!
É ter cá dentro um astro que flameja,
É ter garras e asas de condor!
É ter fome, é ter sede de Infinito!
Por elmo, as manhãs de oiro e de cetim...
É condensar o mundo num só grito!
E é amar-te, assim perdidamente...
É seres alma, e sangue, e vida em mim
E dizê-lo cantando a toda a gente!”
Florbela Espanca (1923)

VI

VII
ACKNOWLEDGEMENTS
This is the place reserved to make a retrospective on the last 3 years of my career, (life),
and PhD dissertation. This has been a journey where I gained knowledge from different
people who made a difference that mattered.
I wish to express my gratitude to my supervisor Professor Manuel Simões for his
help, patience and support. My co-supervisor Professor Filipe Mergulhão, and Professor
Luís Melo are recognized as well. Without you it would be impossible to complete this
thesis. You were all very helpful throughout the whole process by providing assistance
and guidance.
The Faculty of Engineering of University of Porto and LEPABE are acknowledged.
These were the places where the work was performed. My deepest gratitude in
particular to Joana Moreira, Carla Ferreira, Joana Malheiro, and Margarida Pereira. And
for the others from the old E303 lab and the new E007/008 labs who helped me a lot
going around the place (whether suggesting better methods or even when searching
for a much needed tool). Thanks for all the discussions which were extremely helpful.
The insights of Idalina on the manuscripts are comprehensively appreciated. The help
of Paula Pinheiro and Silvia Faia is fully acknowledged as well. To all of you thanks for
making the labs a pleasant place.
I gratefully acknowledge the financial support provided by the Operational
Programme for Competitiveness Factors (COMPETE), the European Regional
Development Fund (FEDER), and by the Portuguese Foundation for Science and
Technology (FCT) through the Project Bioresist— PTDC/EBB-EBI/105085/2008. The
project Susclean – KBBE.2011.2.3-01/287514 is also acknowledged.
Thank you Hans just for being there, giving me those pep talks, and a vision for
future endeavors.
Jorge Trigo, I thank you for being my teacher, friend and for the important lessons
about the walk of life.
To my friends Lobo, Bony, Zé, João and Rita thank you for your friendship, I cherish
all the moments spent with you, with all the laugh and warm feelings. Thanks for being
here.

VIII
To my family, Mamã, Papá, Sis, João, Beatriz, Luiz, Hugo, Marieta, and Ana thank
you for all the love, tenderness, support and incentive much needed to keep me going
with bravery even when everything seemed to be falling apart. I love you all.
I would like to reinforce my gratitude for Luiz for all the time spent double checking
the manuscripts.
And last but not the least, to my Bubas just for being there doing what you do,
which is mostly a bit of all things acknowledged before and a tad more. I love you!
March 13th 2014

IX
ABSTRACT
Biofilms, accumulated microorganisms and extracellular compounds on a surface, are
able to thrive in all environments. Biofilm presence in the food industry can cause
negative effects, being associated to lower industrial operational efficiencies, as well as
microbial contamination of the final product. There are many strategies that attempt to
control biofilm proliferation, however, no control strategy is completely effective. Thus,
the development of new and more effective treatments and improving of the
conventional strategies is in demand. In an effort to overcome biofilm resistance new
compounds must be discovered and their antimicrobial properties assessed.
Additionally, the association between different chemical agents could potentiate their
singular antimicrobial efficacy.
The main objective of this study was to develop biofilm control strategies and to
understand the biofilm behavior to these conditions. Therefore a selection of factors
associated with biofilm resistance were studied. Bacillus cereus and Pseudomonas
fluorescens are common contaminants in the food industry and were selected as
microbial models. Several antimicrobial agents were screened using a colony biofilm
test. These consisted as biofilms developed in as colonies in the top of polycarbonate
membranes. The efficacy of selected agents with putative antimicrobial quenching
substances was studied using respirometry. The killing and removal efficacy of
treatments with antimicrobial agents was assessed using 96-well microtiter plates. To
mimic close-to-practice conditions, biofilms were developed in a flow cell system and
characterized. Control strategies potentiating current antimicrobial agents, and new
agents were performed using biofilms developed in the referred bioreactors.
The diffusion of ethanol, isopropanol, sodium hypochlorite, chlorine dioxide,
hydrogen peroxide, bezalkonium chloride (BAC), benzyldimethyldodecylammonium
chloride (BDMDAC), cetyltrimethylammonium bromide (CTAB), ciprofloxacin,
erythromycin, streptomycin and tetracycline was assessed on colony biofilms.
Ciprofloxacin, streptomycin, BAC and CTAB were selected to assess their biofilm control
efficacy. These products had distinct abilities to diffuse through the biofilms (high
diffusion – BAC and ciprofloxacin; low diffusion – CTAB and streptomycin). It was
concluded that the diffusion ability of antimicrobial agents is not directly correlated with
biofilm killing and removal efficacy. BAC and CTAB were selected for the following
studies due use in industrial cleaning and disinfection practices.
Known constituents of the extracellular polymeric matrix of biofilms (alginate and
humic acids), and selected disinfection-interfering agents from the European Standard
EN – 1276 (bovine serum albumin and yeast extract) were used to challenge the
antimicrobial efficacy of the selected quaternary ammonium compounds as soiling

X
agents. The minimum bactericidal concentration of the chemicals was assessed. The
interfering agents simulated “clean” soiling conditions. Within the range of
concentrations tested the interfering substances mildly reduced the action of the
antimicrobial agents. Humic acids were able not only of reducing the antimicrobials
efficacy, but also to increase P. fluorescens respiratory activity. It was shown that humic
acids should be considered as a potential interfering agent when developing cleaning
and disinfection solutions, due to its strong interaction with the quaternary ammonium
compounds tested.
The biofilms grown in the flow cell system at varying linear flow velocities (applied
in food industry) showed different characteristics. The biofilms developed at the lowest
linear flow velocity (u = 0.1 m.s-1) differed by being thicker and more hydrated than the
biofilms developed at the two higher linear flow velocities (u = 0.4 m.s-1 and
u = 0.8 m.s-1). These biofilms were more compact, with higher bacterial cell numbers
and more exopolymeric substances (proteins and polysaccharides). In spite of these
differences, the dry biofilm mass per area was similar, as well as the expression of the
major outer membrane proteins from the biofilm cells. The biofilms developed at higher
linear flow velocities were selected for further studies of control strategies, due to
higher resistance characteristics (cells and exopolymeric substances).
Halogen-based products are recognized for their relevant antimicrobial properties.
Thus, selected halogen-based products (CTAB, 3-bromopropionyl chloride -BrCl, 3-
bromopropionic acid -BrOH and sodium hypochlorite -SH) were used in order to
understand their antimicrobial activity against both planktonic and biofilm cells of
P. fluorescens. The mode of action of these products is cell membrane disruption,
causing leakage of essential cellular constituents. The results demonstrate comparable
effects of BrCl and BrOH to those of sodium hypochlorite that makes them a potential
alternative to sodium hypochlorite. However, CTAB was the most efficient agent.
The addition of enzymes as an aid to biofilm control treatments, applied alone or
in combination with BAC and CTAB, had the ability to kill and remove the biofilms
developed in microtiter plates and in the flow cell system. The combination enzyme-
biocides was synergistic on biofilm control. The treatments allowed both long term
effects (additional biofilm removal and colony forming units reduction were observed
in the hours following the treatments), as well as biofilm regrowth.
The presented studies in this thesis clearly underline the importance to study
biofilm control strategies under representative conditions for practice, being stress
conditions determinants of different biofilm responses. Biofilm control should be a
multifactorial approach due to the many features that biofilms have that provides them
an increased protection. It is, therefore, necessary to incessantly find new control
strategies because microorganisms will adapt and find new ways to overcome the
biofilm control treatments.

XI
SUMÁRIO
Os biofilmes, acumulação de microrganismos e de substâncias exopoliméricas numa
superfície, são capazes de prosperar em todos os ambientes. A sua presença na
indústria alimentar pode causar efeitos negativos, devido à redução de eficiência de
processos industriais, assim como a contaminação do produto final. Há muitas
estratégias para controlar a proliferação de biofilmes, no entanto, nenhuma é
totalmente eficaz. Deste modo, é necessário otimizar os tratamentos convencionais e
também descobrir tratamentos novos e eficazes. Novos compostos devem ser
procurados e estudados, para que superem a resistência dos biofilmes. Adicionalmente, a
associação entre diferentes agentes químicos pode também potenciar a sua eficácia
antimicrobiana.
O objetivo principal deste estudo foi desenvolver e otimizar estratégias para o
controlo de biofilmes. Para tal, fatores relacionados com a resistência dos biofilmes
foram estudados. Bacillus cereus e Pseudomonas fluorescens são duas bactérias
responsáveis por contaminações na indústria alimentar, e por isso foram selecionadas
como modelos microbianos representativos. Vários agentes antimicrobianos foram
rastreados com um teste com biofilmes em colónia. A eficácia dos agentes selecionados
com possíveis substâncias interferentes foi estudada usando respirometria. A morte e
a remoção dos biofilmes foi avaliada recorrendo a placas de microtitulação de 96 poços.
Para simular condições reais, foram desenvolvidos e caracterizados biofilmes em células
de fluxo. Este sistema serviu para testar estratégias de controlo de biofilmes como a
potenciação de agentes antimicrobianos, assim como o desenvolvimento de novos
agentes.
A difusão de etanol, isopropanol, hipoclorito de sódio, dióxido de cloro, peróxido
de hidrogénio, cloreto de benzalcónio (BAC), cloreto benzilldimetildodecilamónio
(BDMDAC), brometo de cetiltrimetilamónio (CTAB), ciprofloxacina, eritromicina,
tetraciclina e estreptomicina foi avaliada nos biofilmes em colónia. Destes agentes,
foram selecionados a ciprofloxacina, a estreptomicina, o BAC e o CTAB para avaliar a
sua eficácia na morte e remoção de biofilmes. Estes produtos difundiam de forma
distinta através dos biofilmes (alta difusão - BAC e ciprofloxacina, baixa de difusão -
CTAB e estreptomicina). Concluiu-se que a capacidade de difusão de agentes
antibacterianos não está diretamente correlacionada com a sua capacidade para matar
ou remover. No entanto, BAC e CTAB foram selecionados para estudos adicionais,
devido ao seu uso corrente em práticas de limpeza e desinfeção.
Componentes da matriz extracelular dos biofilmes (alginato e ácidos húmicos) e
agentes interferentes da desinfeção selecionados da Norma Europeia EN-1276 (1997)
albumina de soro bovino e extrato de levedura) foram usadas, em condições “limpas”, para
desafiar a eficácia antimicrobiana dos compostos quaternários de amónio selecionados. A

XII
concentração mínima bactericida dos compostos foi aferida neste estudo. Dentro da
gama de concentrações testada, as substâncias interferentes reduziram levemente a ação
dos agentes antimicrobianos. Os ácidos húmicos foram capazes não só de reduzir a eficácia
dos agentes antimicrobianos, mas também de aumentar a atividade respiratória de P.
fluorescens. Foi mostrado que estes devem ser considerados como um agente
potencialmente interferente aquando do desenvolvimento de soluções de limpeza e
desinfeção, devido à sua forte interação com os compostos testados.
Os biofilmes desenvolvidos nas células de fluxo apresentaram características
diferentes de acordo com a velocidade do fluxo (semelhantes às aplicadas na industria
alimentar) à qual foram desenvolvidos. Os biofilmes formados a uma velocidade de fluxo
mais baixa (u = 0.1 m.s-1) apresentaram-se mais espessos e hidratados do que os biofilmes
desenvolvidos às duas velocidades de fluxo mais elevadas (u = 0.4 m.s-1 e u = 0.8 m.s-1).
Estes eram mais compactos, com mais células e substâncias exopoliméricas (proteínas
e polissacarídeos). Apesar destas diferenças, a massa de biofilme seco por unidade de
área foi semelhante, bem como a expressão das proteínas principais da membrana
externa das células. Os biofilmes desenvolvidos às velocidades de fluxo mais elevadas
apresentaram maiores densidades celulares e substancias exopoliméricas, razão pela
qual foram selecionados para estudos de estratégias de controlo subsequentes.
Produtos à base de halogénio são reconhecidos pelas suas propriedades
antimicrobianas. Portanto, os produtos à base de halogéneo selecionados (CTAB,
cloreto de 3-bromopropionilo – BrOH, ácido 3-bromopropiónico – BrCl, e hipoclorito de
sódio) foram utilizados com o intuito de compreender a sua atividade tanto contra
células planctónicas como em biofilmes de P. fluorescens. O modo de ação destes
produtos caracterizou-se essencialmente pelo rompimento das membranas celulares e
a libertação de constituintes celulares essenciais. Os resultados demonstram que os
efeitos de BrCl e BrOH são comparáveis aos do hipoclorito de sódio, o que lhes confere
potencial como substitutos do último. Mas, CTAB foi o agente mais eficaz.
A adição de enzimas é, quando aplicada isoladamente e combinadas com BAC e
CTAB, capaz de matar e remover os biofilmes formados nas placas de poliestireno e nas
células de fluxo. Após os tratamentos foi possível verificar efeitos de longo prazo na
redução da massa do biofilme e das células formadoras de colónias, no entanto, foi
também observada uma eventual recuperação destes mesmos parâmetros.
Os estudos apresentados nesta tese enfatizam que o estudo de estratégias para o
controlo de biofilmes deve ser feitos em condições representativas da prática, sendo
que o comportamento dos biofilmes é determinado pelas condições de stress a que é
sujeito. O controlo dos biofilmes deve ser realizado através de uma abordagem
multifatorial, devido aos muitos recursos que estes dispõem para a sua proteção. É
necessário desenvolver incessantemente novas estratégias de controlo, pois os
microrganismos vão sempre encontrar novos métodos para superar os tratamentos.

XIII
THESIS OUTPUTS
The results presented in this thesis are published and/or submitted as following:
PEER REVIEWED JOURNALS
Araújo PA, Lemos M, Mergulhão F, Melo L, Simões M. 2013. The influence of interfering
substances on the antimicrobial activity of selected quaternary ammonium
compounds. International Journal of Food Science; 2013:9.
Araújo PA, Mergulhão F, Melo L, Simões M. 2014. The ability of an antimicrobial agent
to penetrate a biofilm is not correlated with its killing or removal efficiency.
Biofouling 1-9.
Araújo PA, Malheiro J, Mergulhão F, Melo L, Simões M. The influence of flow velocity
on the characteristics of Pseudomonas fluorescens biofilms. Submitted
Araújo PA, Machado I, Mergulhão F, Melo L, Simões M. Evaluation of the synergistic
potential of enzymes with quaternary ammonium compounds for biofilm control.
Submitted
Araújo PA, Machado I, Mergulhão F, Melo L, Simões M. Decreased efficacy of enzymes
on the control of flow generated biofilms. Submitted
Malheiro J, Araújo PA, Machado I, Mergulhão F, Melo L, Simões M. The effects of
selected halogen-containing chemicals on Pseudomonas fluorescens planktonic
cells and flow-generated biofilms. Submitted
BOOK CHAPTERS
Araújo PA, Lemos M, Mergulhão F, Melo L, Simões M. 2011. Antimicrobial resistance in
biofilms to disinfectants. In: Science against microbial pathogens: communicating
current research and technological advances. Badajoz, Spain: Formatex. p. 826-
834.
Araújo PA, Lemos M, Simões M. 2012. Controlo químico de biofilmes industriais. In:
Biofilmes – Na saúde, no ambiente, na indústria Porto, Portugal. Publindustria Lda.

XIV
Araújo PA, Mergulhão F, Melo L, Simões M. 2013. Diffusivity of antimicrobial agents
through biofilms of Bacillus cereus and Pseudomonas fluorescens. In: Worldwide
Research Efforts in the Fighting against Microbial Pathogens: From Basic Research
to Technological Developments. Brown Walker Press. p. 129-133.
Gomes LC, Araújo PA, Teodósio JS, Simões M, FJ Mergulhão. 2014. The effect of
plasmids and other biomolecules on the effectiveness of antibiofilm agents. In:
Antibiofilm agents: from diagnosis to treatment and prevention. Eds: Ahmad I,
Rumbaugh K. Springer.
CONFERENCE PROCEEDINGS
Araújo PA, Lemos M, Mergulhão FJ, Melo LF, Simões M. Influence of interfering
substances in disinfection. Poster presentation in Microbiotec’11 in the panel
Industrial and Food Microbiology and Biotechnology PS1:120. Braga, Portugal, 1-3
December 2011.
Araújo PA, Mergulhão FJ, Melo LF, Simões M. Diffusivity of antimicrobial agents through
biofilms of Bacillus cereus and Pseudomonas fluorescens. Oral presentation in ICAR
in the panel Antimicrobial surfaces - Biofilms - Quorum sensing. Lisbon, Portugal 22
November 2012.

XV
TABLE OF CONTENTS
Acknowledgements ..................................................................................................... VII
Abstract ......................................................................................................................... IX
Sumário .......................................................................................................................... XI
Thesis outputs .............................................................................................................. XIII
Table of contents ......................................................................................................... XV
List of figures ................................................................................................................ XXI
List of tables ............................................................................................................... XXIII
CHAPTER 1
THESIS OUTLOOK ............................................................................................................ 1
1.1 Relevance and motivation .................................................................................. 3
1.2 Main objectives ................................................................................................... 3
1.3 Thesis organization ............................................................................................. 4
References ...................................................................................................................... 4
CHAPTER 2
INTRODUCTION ............................................................................................................... 7
2.1 Biofilms ............................................................................................................... 9
Biofilm formation ................................................................................................. 10
2.2 The impact of biofilm formation ....................................................................... 11
2.3 The exopolymeric matrix .................................................................................. 13
2.4 Resistance ......................................................................................................... 16
Biofilm resistance ................................................................................................. 17
Cell heterogeneity ................................................................................................ 18
Mass tranfer limitations ....................................................................................... 19
Specific resistance genes ...................................................................................... 20

XVI
Quorum sensing .................................................................................................. 21
Multidrug efflux pumps ....................................................................................... 21
Persister cells ...................................................................................................... 22
2.5 Biofilm control ................................................................................................. 23
Prevention ........................................................................................................... 23
Cleaning and disinfection .................................................................................... 25
Biocides ............................................................................................................... 27
Mechanisms of antimicrobial action ................................................................... 29
Factors affecting biocide action .......................................................................... 30
2.6 Innovative strategies for biofilm control .......................................................... 32
References ................................................................................................................... 35
CHAPTER 3
DIFFUSION OF ANTIMICROBIAL AGENTS THROUGH BIOFILMS .................................... 47
Abstract ........................................................................................................................ 48
3.1 Introduction ..................................................................................................... 49
3.2 Materials and methods .................................................................................... 50
Microorganisms and culture conditions .............................................................. 50
Antimicrobials ..................................................................................................... 51
Colony biofilm formation and penetration tests ................................................. 51
Biofilm formation in microtiter plates ................................................................. 52
Biofilm characterization ...................................................................................... 52
Biofilm control in microtiter plates ..................................................................... 53
Biomass and viability quantification .................................................................... 53
Statistical analysis ................................................................................................ 54
3.3 Results and discussion ..................................................................................... 55
Diffusion of antimicrobial agents through biofilms ............................................. 56
Biofilm activity screening .................................................................................... 60
3.4 Conclusions ...................................................................................................... 62
References ................................................................................................................... 62

XVII
CHAPTER 4
INFLUENCE OF INTERFERING SUBSTANCES ON DISINFECTION ..................................... 67
Abstract ........................................................................................................................ 68
4.1 Introduction ...................................................................................................... 69
4.2 Materials and Methods ..................................................................................... 71
Microorganisms and Culture Conditions .............................................................. 71
QACs and Interfering Agents ................................................................................ 71
Disinfection Procedure ......................................................................................... 72
QACs Neutralization ............................................................................................. 72
Respiratory Activity Assessment .......................................................................... 72
Statistical Analysis ................................................................................................ 73
4.3 Results............................................................................................................... 74
4.4 Discussion ......................................................................................................... 79
4.5 Conclusions ....................................................................................................... 83
References .................................................................................................................... 83
CHAPTER 5
HYDRODYNAMIC CONDITIONS AND BIOFILM DEVELOPMENT ..................................... 87
Abstract ........................................................................................................................ 88
5.1 Introduction ...................................................................................................... 89
5.2 Materials and Methods ..................................................................................... 90
Microorganism and culture conditions ................................................................ 90
Biofilm formation in a flow cell system ................................................................ 90
Biofilm sampling and analysis............................................................................... 92
Outer membrane proteins extraction .................................................................. 93
SDS-PAGE ............................................................................................................. 93
Scanning electron microscopy ............................................................................. 93
Determination of nutrient and cell load ............................................................... 94
Determination of the shear stress........................................................................ 94

XVIII
Determination of the mass transfer coefficients................................................. 94
Statistical analysis ................................................................................................ 95
5.3 Results .............................................................................................................. 95
5.4 Discussion ........................................................................................................ 99
5.5 Conclusions .................................................................................................... 101
References ................................................................................................................. 101
CHAPTER 6
HALOGEN-BASED COMPOUNDS IN BIOFILM CONTROL ............................................. 105
Abstract ...................................................................................................................... 106
6.1. Introduction ................................................................................................... 107
6.2. Materials and methods .................................................................................. 109
Antimicrobial agents ......................................................................................... 109
Microorganisms and culture conditions ............................................................ 109
Antibacterial susceptibility tests ........................................................................ 110
Physicochemical characterization of bacterial surfaces .................................... 110
Bacterial surface charge .................................................................................... 111
Potassium (K+) leakage ...................................................................................... 111
Outer membrane protein extraction and analysis ............................................ 111
Assessment of quorum sensing inhibition ......................................................... 112
Colony biofilm formation and penetration tests ............................................... 112
Biofilm formation in a flow cell system ............................................................. 112
Biofilm analysis .................................................................................................. 113
Statistical analysis .............................................................................................. 113
6.3. Results ............................................................................................................ 114
6.4. Discussion ...................................................................................................... 119
References ................................................................................................................. 123

XIX
CHAPTER 7
ENZYMATIC TREATMENTS FOR BIOFILM CONTROL .................................................... 129
Abstract ...................................................................................................................... 130
7.1 Introduction .................................................................................................... 131
7.2 Materials and methods ................................................................................... 132
Bacteria and culture conditions ......................................................................... 132
Antimicrobial agents and enzymes .................................................................... 133
Biofilm formation in microtiter plates ................................................................ 133
Biofilm control using enzymes ........................................................................... 134
Biofilm mass and viability assessment ............................................................... 134
Biofilm control activity classification .................................................................. 134
Biofilm formation in a flow cell system .............................................................. 135
Flow generated biofilm characterization ........................................................... 135
Tests with planktonic cells.................................................................................. 136
Statistical analysis............................................................................................... 136
7.3 Results............................................................................................................. 137
The effect of an enzymatic treatment on biofilm control .................................. 137
Control of biofilms developed in the flow cell system ....................................... 140
Planktonic tests with enzymes ........................................................................... 142
7.4 Discussion ....................................................................................................... 143
7.5 Conclusions ..................................................................................................... 147
References .................................................................................................................. 147
CHAPTER 8
CONCLUDING REMARKS AND PERSPECTIVES FOR FURTHER RESEARCH .................... 151
8.1 Final conclusions ............................................................................................. 153
8.2 Future work .................................................................................................... 155

XX
Nomenclature ............................................................................................................ 157
Abbreviations .................................................................................................... 157
Indexes .............................................................................................................. 158
Greek ................................................................................................................. 158

XXI
LIST OF FIGURES
CHAPTER 2
Figure 2.1 Microbial contaminations in food industry (adapted from [44]). 10
Figure 2.2 P. fluorescens biofilms developed for 7 days at a Re of 4000. Air dehydrated in a desiccator for two days, the thin layer covering the cells is believed to be EPS.
12
Figure 2.3 Biofilm resistance diversity to antimicrobial agents: (1) genetic expression of certain resistance genes, (2) restricted growth rates; level of metabolic activity within the biofilm; the existence of persisters, (3) mass transfer limitations, (4) quorum sensing and (5) multidrug efflux pumps. (Adapted from [95]).
16
Figure 2.4 Antimicrobial mode of action of biocides (adapted from [164]). CRAs – Chlorine removal agents; QACs – Quaternary ammonium compounds.
28
CHAPTER 3
Figure 3.1 Array of polycarbonate membranes and biofilms for the study of the diffusion of antimicrobial agents through biofilms (adapted from Anderl et al. [41] and Singh et al.[42]).
69
Figure 3.2
Inhibition halos on S. aureus using three antimicrobial agents. Condition 1 corresponds to the control where no biofilm is present and condition 2 represents the tests with biofilms. Both condition are duplicated in the same plate. The conditions tested were (a) BAC test in the presence of a P. fluorescens biofilm, showing that this compound is not retarded; (b) ciprofloxacin in the presence of a B. cereus biofilm, showing that this compound is not retarded, also that the inhibition halos are large taking into consideration the small amount used (5 µg); and (c) CTAB in the presence of a B. cereus biofilm, showing that there is antimicrobial activity in 1, however, the compound was totally retarded by the presence of the biofilm (no halos were observed).
73
CHAPTER 4
Figure 4.1 Chemical structures of benzalkonium chloride (A) and cetyltrimethyl ammonium bromide (B).
74
Figure 4.2 Inactivation of B. cereus by BAC (a), CTAB (b), and QAC combination (c), where solid white box is the control (no interfering substances), light grey box corresponds to BSA, grey box, is ALG dark grey box YE, and black box HA. ∗ means no inactivation. Average values ± standard deviation for at least three replicates are illustrated.
75

XXII
Figure 4.3
Inactivation of P. fluorescens by BAC (a), CTAB (b), and QAC combination (c), where solid white box is the control (no interfering substances), light grey box corresponds to BSA, grey box is ALG, dark grey box is YE, and black box is HA. ∗ means no inactivation. Values below zero are indication that the metabolic activity increased in comparison with the control experiment. Average values ± standard deviation for at least three replicates are illustrated.
76
Figure 4.4
Inactivation of the bacterial consortium by BAC (a), CTAB (b), and QAC combination (c), where solid white box is the control (no interfering substances), light grey box corresponds to BSA, grey box is ALG, dark grey box is YE, and black box is HA. ∗ means no inactivation. Values below zero are indication that the metabolic activity increased in comparison with the control experiment. Average values ± standard deviation for at least three replicates are illustrated.
77
CHAPTER 5
Figure 5.1 Depiction of the flow cell system used to develop biofilms. 91
Figure 5.2 Photographs of the stainless steel coupons with 7 day old biofilms grown at (a) u = 0.1 m.s-1, (b) u = 0.4 m.s-1 and (c) u = 0.8 m.s-1.
95
Figure 5.3 SEM micrographs of P. fluorescens biofilms developed on stainless steel surfaces at different flow conditions: (a) u = 0.1 m.s-1, (b) u = 0.4 m.s-1 and (c) u = 0.8 m.s-1. x 15000 magnification; bar = 5 µm.
97
Figure 5.4 OMP profiles of P. fluorescens bacteria developed in different modes of growth. Biofilms formed at three flow regimes (a) u = 0.1 m.s-1, (b) u = 0.4 m.s-1 and (c) u = 0.8 m.s-1.
98
CHAPTER 6
Figure 6.1. Chemical structures of the chemicals used: (a) Cetyltrimethylammonium bromide (CTAB), (b) Sodium Hypochlorite (SH), (c) 3-Bromopropionyl chloride (BrCl) and (d) 3-Bromopropionic acid (BrOH).
109
Figure 6.2.
OMP profile of P. fluorescens cells when exposed to MBC of different chemicals. The molecular weight market (a) was used to extrapolate the molecular weight of some lanes of the OMPs profile obtained from incubation in the (b) absence or in the presence of (c) BrCl, (d) BrOH, (e) CTAB and (f) NaOCl.
116
Figure 6.3.
Disc diffusion assays for the detection of quorum sensing inhibition of C. violaceum by (a) BrOH, (b) BrCl, (c) SH and (d) CTAB.
117
Figure 6.4.
Pseudomonas fluorescens biofilm log CFU.cm-2 (a) and mass (b) before and after treatment with CTAB ( ), BrCl ( ) and SH ( ). Samples were collected before treatment ( ), immediately after 1 hour treatment and after 2, 12 and 24 hours after chemical removal. Values are average ± SD.
118

XXIII
CHAPTER 7
Figure 7.1 Killing and removal percentages of B. cereus and P. fluorescens biofilms using the selected enzymes with and without the selected QAC. Where corresponds to β-glucanase, protease, lipase, α-amylase and QAC. The enzymatic and QAC (biocide) solutions were applied for 1 h. *means no killing. Average values ± standard deviation for at least three replicates are illustrated.
138
Figure 7.2 Killing and removal percentages for B. cereus and P. fluorescens biofilms using the selected enzymes. Where corresponds to β-glucanase, protease, lipase, α-amylase and QAC. The enzymatic solutions were applied for 30 min then removed and the biocide was applied for 30 min (30 + 30). Average values ± standard deviation for at least three replicates are illustrated.
139
Figure 7.3 Mass, and log CFU of P. fluorescens biofilms in time after the treatments with an enzymatic solution (left hand) and an enzymatic solution combined with CTAB. Where corresponds to β-glucanase, protease,
lipase, α-amylase and CTAB. *-means no reduction. Average values ± standard deviation is depicted.
140
Figure 7.4
Effect of chemical treatment for 1 h of B. cereus (a) and P. fluorescens (b) planktonic cultures. Different enzymes were used ( β-glucanase, protease, lipase, and α-amylase) alone and in combination with the two biocides. The positions of BAC and CTAB alone solutions are represented by arrows. Total inactivation of respiratory activity is indicated with an asterisk (*). Average values ± standard deviation for at least three replicates are depicted.
142
LIST OF TABLES
CHAPTER 2
Table 2.1 Functions of EPS in bacterial biofilms adapted from [2, 5, 51, 60, 61]. 13
Table 2.2 Mechanisms of interaction of several biocides according to their cellular targets and antimicrobial actions (adapted from [163]).
26
CHAPTER 3
Table 3.1 Characterization of B. cereus and P. fluorescens grown as colonies and as microtiter plate biofilms.
53
Table 3.2 Antimicrobial agents plus respective S. aureus inhibition halos, and percentage retardation caused by the presence of B. cereus and P. fluorescens biofilms. (Average ± SD)
55

XXIV
Table 3.3 Percentage killing and removal of B. cereus and P. fluorescens biofilms. The average ± SD is presented.
59
CHAPTER 4
Table 4.1 Minimum bactericidal concentration for P. fluorescens, B. cereus and the consortium with and without interfering substances
78
CHAPTER 5
Table 5.1 Characterization of P. fluorescens biofilms grown at different linear flow velocities.
96
Table 5.2
Hydrodynamic and mass transfer coefficients of 7 day-old P. fluorescens biofilms grown at different flow regimes.
98
CHAPTER 6
Table 6.1 Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) values of each chemical tested.
114
Table 6.2 Surface tension parameters, hydrophobicity (∆𝐺𝑠𝑤𝑠 ), apolar (𝛾𝑠
𝐿𝑊) and polar (𝛾𝑠
𝐴𝐵), of untreated P. fluorescens (control) and after 1 hour treatment with a chemical (BrCl, BrOH, CTAB or SH). The average ± SD is presented.
114
Table 6.3 Zeta potential and conductivity of P. fluorescens before and after 1 hour treatment with different chemicals. The average ± SD is presented.
115
Table 6.4 Concentration of K+ in solution before and after 1 hour incubation of P. fluorescens with each chemical. The average ± SD is presented.
115
Table 6.5 Retardation caused by P. fluorescens biofilms, for each chemical used. Data is presented as average ± SD of the percentage of diameter measurements for halo readings compared with controls (no biofilm).
116

CHAPTER 1 THESIS OUTLOOK

2 Chapter 1 _________________________________________________________________________________

Thesis Outlook 3 _________________________________________________________________________________
1.1 RELEVANCE AND MOTIVATION
It is a natural tendency of microorganisms to attach to surfaces, multiply and produce
extracellular polymeric substances (EPS), originating biofilms. Biofilm existence can
represent a beneficial or a detrimental factor, affecting many areas, from the
biomedical to the industrial [1, 2].
In food industry, process conditions are ideal for biofilm proliferation. Biofilm
formation is very common in industrial settings, even when manufacturers apply
comprehensive contingency plans. The EPS produced by bacteria in biofilms protects
microorganisms from control strategies by hindering diffusion of antimicrobial agents
and promoting antimicrobial quenching effects due to chemical reactions with the
antimicrobial agents [3].
Flemming [4] described EPS as very complex and dynamic. Its exact functions of
EPS are not yet clear e.g. because of the extreme heterogeneity. The EPS matrix is an
intricate network that provides sufficient mechanical stability to maintain spatial
arrangement for embedded-bacteria. It consists of various organic substances such as
polysaccharides, proteins, nucleic acids and lipids [5]. This composition is affected by
the environmental conditions under which biofilms are formed, and its arrangement is
affected by the hydrodynamic stress [6, 7].
Disinfection procedures are commonly designed based on experiments carried out
with planktonic bacterial cell cultures [8]. But, such tests do not mimic the biofilm and
environmental conditions on surfaces in industrial processes. Actually, the European
Standard EN-1276 (1997) [9], used as reference for the development of disinfection
strategies for food, industrial, domestic and institutional areas, only provides a short list
of potential interfering chemical substances to be considered when optimizing/
developing a disinfection process. Nevertheless, the conventional explanations for
biofilm resistance and recalcitrance against current control strategies are based on the
effects of the presence of a heterogeneous EPS matrix on transport limitations and
chemical interactions with antimicrobial agents [3, 10, 11].
Knowledge of EPS is needed to develop effective biofilm control strategies. This
knowledge can help overcome biofilm resistance. The treatment of biofilms with
enzymes to weaken the EPS structure may enhance the effectiveness of other
antimicrobial agents. Enzymes can degrade the EPS barrier and therefore increase the
diffusivity of the chemical agents [12]. Moreover, due to the recognized antimicrobial

4 Chapter 1 _________________________________________________________________________________
resistance problem it is of importance to search and identify new and more effective
antimicrobial agents and develop new control strategies [13].
Despite the definite importance of biofilms in microbial life style and their effects
on human beings, the present knowledge about their structure, composition and
behavior is still limited. Therefore there is a need to better understand biofilm
resistance, by the identification of parameters linked with it, so that control strategies
can be developed and optimized. Bioresist is a project financed by national funds
through the Portuguese Foundation for Science and Technology (FCT) and MCTES
(PIDDAC) and co-financed by the European fund of Regional Development (FEDER)
through COMPETE – Operational Programme for Competitiveness Factors (POFC), with
the reference PTDC/EBB-EBI/105085/2008. This project studied the influence of biofilm
phenotype on its resilience and resistance. This PhD thesis was developed within the
scope of this project.
1.2 MAIN OBJECTIVES
There are still no biofilm control strategies providing sustainable results in terms of
inactivation, removal and prevention of biofilm regrowth events [14]. Development of
approaches to control unwanted biofilms requires detailed knowledge about the
biofilms [15]. It is necessary to develop strategies to control biofilms native of food
industry, and simultaneously identify the resistance mechanisms associated with
control strategies. Thus, the main objective of this study is to provide a contribution for
the development of biofilm control strategies. Moreover, the outcomes of this thesis
provide insight into how biofilms are affected by the food industry process
hydrodynamics and how the biofilm phenotype is linked with its resistance.
1.3 THESIS ORGANIZATION
This thesis is essentially divided in eight chapters:
Chapter 1 describes the relevance and motivation, the objectives, and the work
structure presented throughout this thesis are exposed.

Thesis Outlook 5 _________________________________________________________________________________
Chapter 2 provides a review of the aspects of microbial resistance, and control
strategies currently/recently applied, with particular emphasis on biofilms. It is the
state of the art of major aspects related with the topic of this thesis.
Chapter 3 presents a study of the diffusivity of twelve biocides and antibiotics (ethanol,
isopropanol, sodium hypochlorite, chlorine dioxide, hydrogen peroxide, BAC,
BDMDAC, CTAB, ciprofloxacin, erythromycin, streptomycin and tetracycline) through
Bacillus cereus and Pseudomonas fluorescens biofilms. BAC and CTAB were selected to
be used in further experiments, taking into account their ability to diffuse through the
biofilms and to inactivate the embedded cells.
In chapter 4, the influence of alginic acid, bovine serum albumin, yeast extract, and
humic acids as interfering substances on the antimicrobial action of selected
antimicrobial agents was assessed on planktonic P. fluorescens and B. cereus.
Chapter 5 presents the characteristics of P. fluorescens biofilms developed in a flow cell
system. Three distinct linear flow velocities were used (u = 0.1, 0.4 and 0.8 m.s-1). The
biofilms demonstrating the highest complexity (cell numbers and EPS content) were
selected for further studies.
In chapter 6 halogen-based chemicals, BrCl, BrOH, sodium hypochlorite and CTAB were
tested on their potential to control P. fluorescens planktonic cells and biofilms. The
effects caused by the exposure to the chemicals were studied in order to understand
different aspects of the antimicrobial mode of action of these chemicals.
Chapter 7 presents a biofilm control strategy using enzymes (β-glucanase, protease,
lipase, and α-amylase) and biocides (BAC and CTAB). Different types of treatment were
tested, as an environmentally friendly method. The action of these treatments against
planktonic cells was also assessed in order to understand the antimicrobial action of
the enzymes and the interaction with the selected biocides.
In chapter 8, the main achievements of this thesis are exposed. Regards about follow
up research are provided as well.

6 Chapter 1 _________________________________________________________________________________
This thesis is structured as a paper dissertation, consisting of a number of scientific
articles. The chapters on the experimental work are presented in the way they have
been submitted and/or published upon acceptance. Some repetitions are consequently
unavoidable amongst individual chapters.
REFERENCES
[1] Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: from the natural environmental to infectious diseases. Nature Reviews in Microbiology, 2004. 2:95-108.
[2] Verran J. Biofouling in food processing: biofilm or biotransfer potential? Food and Bioproducts Processing, 2002. 80(4):292-298.
[3] Mah T, O'Toole G. Mechanisms of biofilm resistance to antimicrobial agents. Trends in Microbiology, 2001. 9(1):34-39.
[4] Flemming H. The EPS matrix: The "house of biofilm cells". Journal of Bacteriology, 2007. 189(22):7945-7947.
[5] Bridier A, Briandet R, Thomas V, Dubois-Brissonnet F. Resistance of bacterial biofilms to disinfectants: a review. Biofouling, 2011. 27(9):1017-1032.
[6] Simões M, Pereira MO, Vieira MJ. Effect of mechanical stress on biofilms challenged by different chemicals. Water Research, 2005. 39(20):5142-5152.
[7] Simões M, Simões LC, Vieira MJ. Physiology and behavior of Pseudomonas fluorescens single and dual strain biofilms under diverse hydrodynamics stresses. International Journal of Food Microbiology, 2008. 128(2):309-316.
[8] Palomino JC, Martin A, Camacho M, Guerra H, Swings J, Portaels F. Resazurin microtiter assay plate: simple and inexpensive method for detection of drug resistance in Mycobacterium tuberculosis. Antimicrobial Agents and Chemotherapy, 2002. 46(8):2720-2722.
[9] European Standard EN-1276. Chemical disinfectants and antiseptics-Quantitative suspension test for the evaluation of bactericidal activity of chemical disinfectants and antiseptics used in food, industrial, domestic, and institutional areas-Test method and requirements (phase 2, step 1). 1997.
[10] Simões M, Simões LC, Vieira MJ. Species association increases biofilm resistance to chemical and mechanical treatments. Water Research, 2009. 43(1):229-237.
[11] Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Science 1999. 284(5418):1318-22.
[12] Andersson S, Dalhammar G, Land C, Kuttuva Rajarao G. Characterization of extracellular polymeric substances from denitrifying organism Comamonas denitrificans. Applied Microbiology and Biotechnology, 2009. 82(3):535-543.
[13] Saavedra MJ, Borges A, Dias C, Aires A, Bennett RN, Rosa ES, Simões M. Antimicrobial activity of phenolics and glucosinolate hydrolysis products and their synergy with streptomycin against pathogenic bacteria. Medicinal Chemistry, 2010. 6(3):174-183.
[14] Simões M, Simões L, Vieira M. A review of current and emergent biofilm control strategies. LWT - Food Science and Technology, 2010. 43(4):573-583.
[15] Stewart P, Multicellular nature of biofilm protection from antimicrobial agents. Biofilm communities: Order or Chaos, ed. A.J. McBain, et al. 2003. 181-190.

CHAPTER 2 INTRODUCTION
This chapter was based on:
Araújo PA, Lemos M, Mergulhão F, Melo L, Simões M. 2011. Antimicrobial resistance in biofilms to
disinfectants. In: Science against microbial pathogens: communicating current research and
technological advances. Badajoz, Spain: Formatex. p. 826-834.
Araújo PA, Lemos M, Simões M. 2012. Controlo químico de biofilmes industriais. In: Biofilmes – Na saúde,
no ambiente, na indústria Porto, Portugal. Publindustria Lda.

8 Chapter 2
_________________________________________________________________________________

Introduction 9 _________________________________________________________________________________
2.1 BIOFILMS
Biofilm formation was reported early in fossil records [1]. The first recorded life forms
on earth are biofilms, dated approximately 3.5 billion years ago. They have faced
fluctuating and harsh conditions of primitive earth, such as extreme temperatures and
ultraviolet light exposure [1]. Nowadays, biofilms can be found in the widest range of
environments, on extremes of cold and hot temperatures, high pressures, high alkalinity
or acidity, and even radioactivity [2]. There are also reports where biofilms were found
in improbable environments such as a disinfectant solution [2, 3].
In 1684, Leeuwenhoek described in a report to the Royal Society of London
“animalcules” that were found in plaque scraped from his teeth [4]. Later in 1940
Heukelekian and Heller stated in the Journal of Bacteriology that bacteria develop with
bacterial slime or as colonies attached to surfaces [4]. In 1943, Zobell observed and
described fundamental characteristics of attached microbial communities [4]. In 1975,
the word “aufwuchs” (in German), meaning growth was the first conceptual term used
to describe biofilms, however, it was later discarded by implying to be situated “around
plants”. Studies associated with biofilms started to have more attention in 1978, were
the description of sessile communities was first described and termed by the group of
Bill Costerton. The group described them as microorganisms with the ability to adhere
to wet surfaces in ecosystems of fresh water [5]. Characklis and Marshall described
biofilms as a community of microorganisms, either single or multi species, being
anchored to a surface and entrapped in organic polymers excreted by them [6]. It is
now commonly agreed that bacteria have a natural capability to attach irreversibly to
surfaces, to multiply, and to embed themselves in a slimy matrix, establishing biofilms.
The biofilm population is enclosed in a matrix adhered to each other, to a substratum
or to an interface [7]. Although the population could be constituted by other organisms
besides bacteria [8], single bacterial biofilms are often found in industry and in medicine
[9]. Later on, Stoodley et al. distinguished some characteristics denominating these
structures as we currently know them, they include the association with a surface, a
high population density, and the presence of exopolymeric substances (EPS), which is
the ”glue” that holds biofilms together [8]. Yet, it is not uncommon to find biofilms
lacking one of these characteristics due to the environmental characteristics to which
the bacteria are exposed [10-13].

10 Chapter 2
_________________________________________________________________________________
BIOFILM FORMATION
Biofilm formation is a complex process that involves several stages [8]. Planktonic cells
passing along a “conditioned” surface are deposited. The persistent microorganisms
that remain on the surface bind irreversibly, and start to grow, multiplying and
producing signaling molecules, as well as EPS [8, 14]. As the biofilm matures, an
equilibrium between accumulation and erosion takes place. The biofilm could erode by
dispersing (cells) or by sloughing (biofilm pieces) phenomena. After this stage the
planktonic cells return to the beginning of their cycle on different locations [15-18].
The properties of the adhesion surface, and the surface of the bacteria, as well as
their stage of growth, are determining factors for biofilm formation [19]. A suitable
nutrient concentration, an optimum pH, and an appropriate hydrodynamic force
exerted on the cells, provide favorable characteristics for attracting microorganisms to
be adsorbed to the surfaces [20]. The genetic information of cells in biofilms is fairly
different from their planktonic counterparts, this change is thought to be triggered
when cells adhere to surfaces [21]. In some cases, the development of appendages such
as flagella, fimbriae and pili help in biofilm formation [22].
The hydrodynamic conditions and adjacent environment contribute to biofilm
formation [19, 23]. They affect the matrix structure, quantity and composition [13, 24].
The way how biofilms develop, the transfer of mass, the biofilm density and the
conversion of substrate are all dependent on these parameters [25]. Biofilms developed
under laminar regimes are different from the ones generated in a turbulent regime, as
the access to deeper layers is made by an open structure to ease mass transfer [26, 27].
The shear stress exerted on the biofilms by the passing fluid determines their shape due
to the erosion it causes. As new layers are formed, the force of the passing fluid
dislodges the top layers. In contrast, when the biofilms attain a certain growth size,
some of them are able to secrete surfactants with the ability to alter their internal
properties [24].
The adhesion of microorganisms to surfaces, forming biofilms, represents an
ecological advantage and is a prevalent form of survival in hostile environments. In fact,
it is estimated that 99% of bacteria live in biofilms [4].

Introduction 11 _________________________________________________________________________________
2.2 THE IMPACT OF BIOFILM FORMATION
Biofilms are able to thrive everywhere. The ability of biofilms to develop in nature or in
engineered porous media could be used as an advantage on man-made processes [4].
When biofilms are used this way, they are called beneficial biofilms. For example, when
planktonic cells are used in a reactor, their residence time is the same as the fluid flow
time, however, if cells are in a biofilm, their residence time is as long as the time the
biofilm is attached to a surface within the reactor [23]. Biofilms have been be used for
environmental applications that include the degradation of organic substances,
denitrification of waste or removal of phosphate and heavy metals [28, 29]. They have
been employed as bio-control agents in the rhizosphere of plants, particularly against
infections caused by fungi or bacteria. Biofilms formed by some microorganisms are
able to produce antifungal and antibacterial substances, which provide protection to
plants susceptible to phytopathogenic microorganisms. This has been a field of science
with plenty of interest, since it enables the exploitation of new physiologically active
products [30].
Conversely, biofilms could cause serious operation and management costs
depending on where they appear. When biofilms appear in food industry they are highly
unappreciated because they may contain pathogens and spoilage microorganisms,
which constitute a risk to humans when contaminated and spoiled products are
consumed. Typically, the emergence of biofilms is a result of an ineffective cleaning
plan, increasing production costs due to production downtime. In other areas, such as
the clinical area, biofilms are able to develop in medical devices, implants, venous or
urinary catheters, resulting in an increased risk of infection [31]. In clinical settings
biofilms have a higher importance due to their risk of causing infections, which could
turn chronic [32]. Notwithstanding, environmental biofilms could contain pathogens as
well. Foodborne diseases affect 48 million people in the United States of America each
year. In this group, 2612 people did not survive infections related with microbial
development, being estimated that 65% of all microbial diseases are a consequence of
biofilm development [33-35]. Food poisoning has associated costs, according to
Brooks and Flint these are difficult to estimate, however it was possible to make an
estimative of approximately $90 million for New Zeeland, which is a country with only
4.5 million people [36]. It was also estimated that 25% of the total food produced is lost

12 Chapter 2
_________________________________________________________________________________
due to microbial activity, in spite of the diverse methods employed for food
preservation, good manufacturing, quality control and hygienic measures [37].
Earlier in this chapter it was said that the characteristics of biofilms differ according
to the environmental conditions under which they were formed, i.e. temperature, pH,
type of nutrients available, and type of bacteria. It is also known that the type of
microorganisms that forms biofilms is different according to the location where the
biofilms were found. Dairy industries commonly have biofilms composed by
Pseudomonas fluorescens, Escherichia coli, Shigella spp., Staphylococcus aureus and
Bacillus cereus. Shrimp factories normally have P. fluorescens and P. putida as biofilm
colonizers. In fish factories it is common to find biofilms composed by
Enterobacteriaceae and Serratia liquefaciens. In caviar plants, biofilms of Neisseriaceae
spp., Pseudomonas spp., Vibrio spp. and Listeria spp. were reported [38, 39].
Pseudomonas spp., Klebsiella spp., Legionella spp., Helicobacter spp., Campylobacter
spp. and Escherichia coli were found in drinking water networks [2]. Pseudomonas spp.
are ubiquitous in food industry environments and have been reported to be found in
drains, and produce such as vegetables, meat and dairy products [40]. Bacillus spp. are
found throughout dairy processing plants, accumulating on joints and pipelines of the
equipment [40]. Both are able to form biofilms.
When these bacteria accumulate, they can cause other consequences besides
product spoilage or infections. Biofilms are able to cause detrimental effects on many
systems. Consequences are material corrosion and biodegradation, causing
contamination of the raw or processed products in food processing plants. In cooling
water towers and heat exchangers they cause energy loss due to increased fluid friction
and resistance to heat transfer. In drinking water distribution systems, an increase in
suspended solids and coliform contamination has been observed, in addition to pipe
corrosion and pressure drop [2]. In paper manufacturing, the quality of the product is
reduced. In ship hulls biofouling development increases drag and consequently energy
loss as in reverse osmosis membranes, where the reduced permeability and material
degradation are felt [41-43].
Industrial settings, particularly food processing plants, provide favorable
environmental conditions, i.e. hydrodynamics and nutrients abundance, for biofilm
proliferation (Figure 2.1). Biofilm contaminations are dangerous due to their mode of
life which includes partial sloughing or detachment. Once on the fluid stream these
could proliferate into other locations of the production line, restarting the process all

Introduction 13 _________________________________________________________________________________
over again [8]. This ease of proliferation results in both economic and public health
consequences, therefore, efforts have been directed for efficient industrial equipment
design and the development of effective disinfectants [45, 46].
Figure 2.1 Microbial contaminations in food industry (adapted from [44]).
2.3 THE EXOPOLYMERIC MATRIX
The most recognized characteristic of biofilms is the EPS matrix, which provides
favorable conditions to its inhabitants to thrive in the most diversified surroundings.
Figure 2.2 is a micrograph of P. fluorescens biofilm developed in a flow cell system, the
substance coating the cells is a dehydrated EPS matrix, as suggested in the work of
Flemming et al. [2].
Biofilm characteristics such as porosity, tortuosity, density, water content, charge,
sorption properties, hydrophobicity and mechanical stability are determined by
environmental conditions [2]. Biofilm structure and spatial heterogeneity are essential
to various biofilm processes such as convective and diffusional transport of oxygen and
nutrients into the biofilms [47]. Biofilm heterogeneity is defined as a non-uniform
structural, chemical and physical distribution within the biofilm [47].
The functions of the exopolymeric matrix are diverse, however, not all functions
are fully understood [48]. Some of these are described in Table 2.1. One of the functions
of EPS is to contribute to the mechanical stability of the biofilms, enabling them to
Chemical contaminants
Process water
Nutrients
Biological contaminants
Pathogenic or spoilage organisms
Contaminants in raw material Biofilm detachment
Environmental conditions
Processing time Retention time Temperature
pH
Product contamination
Product spoilage
Blockage of process lines
Product
Biofilm formation
Microbial growth

14 Chapter 2
_________________________________________________________________________________
withstand shear forces, dehydration or chemical attacks [49, 50]. EPS protects the
embedded cells from UV light, radiation, pH changes, osmotic shock, or drying [51].
Furthermore, the matrix reinforces biofilm attachment to the substratum and stabilizes
it, thereby reducing its susceptibility to sloughing by hydrodynamic shear stress [52, 53].
Figure 2.2 P. fluorescens biofilms developed for 7 days at a Re of 4000. Air dehydrated in a desiccator for two days, the thin layer covering the cells is believed to be EPS.
EPS are an intricate network formed essentially by polysaccharides and proteins
[54]. The matrix differs according the microbial producer. In addition, between genus
the matrix is likely to differ either in chemical composition or in terms of physical
characteristics [51]. The composition of the matrix may also contain glycoproteins,
lipoproteins, phospholipids, teichoic acids, nucleic acids and a variety of humic
substances [22, 24, 55]. Any particles passing by the biofilm may be incorporated into it
[56], therefore it is also possible to find mineral crystals, silt particles, milk residues as
calcium phosphate and, sometimes, blood components or dirt [57]. EPS is able to retain
water, the reason why biofilms are highly hydrated [2]. In fact, biofilms are composed
essentially by water, as up to 97% of biofilm volume and mass is water
[13, 58]. EPS composition is determined by the environmental conditions to which the

Introduction 15 _________________________________________________________________________________
biofilm microorganisms are exposed [19, 55]. EPS are excreted by the cells, but also
derive from natural cell lysis or hydrolytic activities [59]. Life in biofilms facilitates gene
transfer and the retention of extracellular enzymes, that are useful to degrade
biodegradable matter (lysed cells), that serve as nutrients for the living bacteria [60].
Table 2.1 Functions of EPS in bacterial biofilms. (Adapted from [2, 5, 51, 60, 61].)
Component function EPS components involved Relevance for biofilm organism
Aggregation of bacterial cells, formation of flocks and biofilms
Polysaccharides, proteins, DNA
Bridging between cells, immobilization of bacterial populations, basis for development of high cell densities; cell communication; biofouling and corrosion
Cell–cell recognition Polysaccharides, proteins, DNA
Symbiotic relation with animals and plants; possible pathogenic processes
Retention of water Hydrophilic polysaccharides/proteins
Maintenance of highly hydrated microenvironment organisms, desiccation tolerance in water-deficient environments
Protective barrier Polysaccharides, proteins Resistance to nonspecific and specific host defenses during infection, tolerance to various antimicrobial agents (e.g., disinfectants, antibiotics); protection against some grazers
Sorption of organic compounds
Charged or hydrophobic polysaccharides and proteins
Accumulation of nutrients from the environment; sorption of endogenous compounds
Sorption of inorganic ions
Charged polysaccharides and proteins, including inorganic substituents such as phosphate and sulphate
Promotion of polysaccharide gel formation; ion exchange; mineral formation; accumulation of toxic metal ions (detoxification)
Enzymatic activity Proteins Digestion of exogenous macromolecules for nutrient acquisition; degradation of structural EPS allowing release of cells
Accumulation, stabilization and retention of secreted enzymes on polysaccharides
Nutrient source Potentially all EPS components
Source of C, N and P compounds for utilization by biofilm community
Genetic information DNA Horizontal gene transfer between biofilm cells
The resistance mechanism provided by the EPS is further reviewed in the next
subsection.

16 Chapter 2
_________________________________________________________________________________
2.4 RESISTANCE
The survival of the fittest is a biological principle applicable to all living beings, and
although different organisms have developed their own survival mechanisms, all have
one common factor that relates survival with the ability to adapt to constant changes
in the environment. Microorganisms are particularly adaptable to environmental
changes because of their high reproduction rates, which allows them to transfer survival
characteristics to future generations in short periods of time [62]. When exposed to a
harmful and/or stressful environment, bacteria will do all within their power to survive
[63]. External stresses, such as environmental conditions, have different effects on
different organisms, leading to natural responses like inhibition and/or inactivation of
the cells. For instance, a deviation in the environmental conditions could result in
reduced growth rates [64]. When bacteria are exposed to sub-lethal levels of biocides,
and only minor cell damage is caused, a more resistant population could derive, with
consequences that may include changes in the global phenotype of the community [63].
Resistance mechanisms are the means that living organisms have to respond to
continuously changing environment in order to survive [65]. Resistance is the
description of the relative insusceptibility, viability or multiplication of a microorganism
to a certain chemical treatment under certain conditions. It may be temporary or
permanent and relates either to the first generation of organisms or to the next [66].
Thus, there are three documented types of resistance: (1) inherent resistance, also
termed natural or intrinsic to the microorganism, (2) adaptive resistance, due to the
occurrence of a mutation, by continuous exposure to certain environments, and finally
(3) acquired resistance which occurs through the acquisition of mobile genetic elements
(plasmids) [67-69]. An example of intrinsic resistance is the difference between Gram
positive and Gram negative cells. The main differences are in the outer cell layers. Gram
positive cells present a large peptidoglycan layer after the phospholipidic membrane,
where proteins and porins are located, while Gram negative have, from the inside to
the outside, a smaller peptidoglycan layer followed by periplasm, an outer
phospholipidic membrane and lipopolysaccharides, which gives the cells a hydrophobic
character. Gram positive outer membrane works as a permeability barrier [70]. This cell
wall is composed essentially by peptidoglycans and teichoic acids. Gram negative outer
hydrophobic membrane limits the entry of the most diverse chemicals by working as an
exclusion barrier [71]. Gram negative bacteria embedded in biofilms are known to have

Introduction 17 _________________________________________________________________________________
a higher ratio of unsaturated to saturated fatty acids, a typical profile of resistant
bacteria [72]. Their morphology limits the concentration of biocide to the
corresponding targets [67]. The adapted resistance could be due to the continuous use
of disinfectants, to which the embedded bacteria gain resistance as a consequence of
the repetitive use of these chemical agents [54]. Similarly, microorganisms may acquire
resistance to some antimicrobial agents through exposure to other agents of the same
type, which is called cross-resistance [73]. A documented case of acquired resistance,
provided by plasmids, is the horizontal transference of resistance to antibiotics from
Lactobacillus plantarum to Enterococcus faecalis [74].
Changes at the phenotypic level, i.e. by forming biofilms as a response to the
environmental conditions, is also a form of resistance [75]. As mentioned on the
previous section, the proliferation of biofilms in industrial settings, especially in food
industry, can result in serious operation and maintenance costs [36]. Their eradication
is proved to be difficult as biofilm cells are known to be highly resistant to antimicrobial
agents. Defense mechanisms against antimicrobial agents are frequently reported in
literature [76-89]. The study of the resistance mechanisms to antimicrobial agents
gradually unravels the mysteries of the biofilm tenacious nature and recalcitrance to
control [90]. A deeper understanding of biofilm resistance mechanisms is required in
order to develop new and more effective biofilm control strategies. Some resistance
mechanisms are described in the following sections.
BIOFILM RESISTANCE
There are several characteristics that underlie the increased resistance of bacterial
films, though some resistance mechanisms are shared with their planktonic
counterparts. Nonetheless, adhered cells have a phenotype that confers them an
increased resistance to antibiotics and biocides, when compared with suspended cells
[67, 72, 91, 92].
Figure 2.3 exhibits several biofilm defence mechanisms differentiating them from
their planktonic counterparts, such as specific resistance genes, restricted growth rates,
the existence of persister cells, quorum sensing communication, stress response
regulons, and the impervious EPS [93]. Cells in biofilms can be 10-1000 times more
resistant to antimicrobial agents than their planktonic counterparts [94]. For instance,

18 Chapter 2
_________________________________________________________________________________
control of biofilm cells of S. aureus requires 600 times more sodium hypochlorite than
their planktonic equivalents [17].
Figure 2.3 Biofilm resistance diversity to antimicrobial agents: (1) genetic expression of
certain resistance genes, (2) restricted growth rates; level of metabolic activity within the biofilm; the existence of persisters, (3) mass transfer limitations, (4) quorum sensing and (5) multidrug efflux pumps. (Adapted from [95].)
CELL HETEROGENEITY
Cell heterogeneity is frequent within biofilms. It is common to find cells at different
physiological states. In a given population, genetic and phenotypic diversity is triggered
by the surrounding conditions [96]. Parameters such as space, nutrients, and age could
contribute to the increased resistance of biofilms.
The unavailability of space can be a factor for resistance. The exopolymeric matrix
can be hindering cell division in mature biofilms. The cells prefer to produce EPS than
new cells, aging the population and increasing mass transfer limitations [97]. Bacteria
have different degrees of resistance according to their state. Resistance increased as
both Burkholderia cepacia planktonic and biofilm cultures approached the stationary
phase [98]. Additionally, P. fluorescens cells are known to produce an exopolysaccharide
lyase, which is triggered by the stress of feeling constrained. This enzyme degrades the

Introduction 19 _________________________________________________________________________________
matrix with the purpose of obtaining nutrients from it and, also to free cells from the
biofilm so that they can colonize other favorable locations [99].
In a high cell density population such inside biofilms [100], mass transfer might be
hindered by cell packing [81]. Higher cell numbers promote horizontal gene transfer of
resistance features [101]. The selection of resistant mutants by exposure to sub-lethal
concentrations of antimicrobial agents is higher in highly populated biofilms [102]. The
different microenvironments and depletion of nutrients and oxygen in the interior of
biofilms can alter the metabolic activity of the cells [94, 103]. The activity is high at the
surface and stratified going to to the interior, reaching its lowest levels in deeper layers.
As cells are buried in the biofilms and the supply of nutrients and oxygen starts to be
scarce, the cells go into a slow growing or dormant state [95, 104]. The antimicrobial
agents that target the disruption of microbial processes are more effective, against
rapidly growing cells [105]. When acting on slow growers or dormants, the activity of
these antimicrobials is antagonized. In a biofilm, only pockets of this type of
microorganism are found, however, some of these cells can be found with at least some
degree of cellular activity [1]. Nonetheless, this mechanism is usually observed by an
increase of resistance [106].
MASS TRANFER LIMITATIONS
The resistance mechanism is more evident in biofilms due to the presence of EPS. Both
structure and composition dictate its susceptibility to antimicrobial agents. The biofilm
constituents may act as an adsorbent or reactant, thus chemically interacting and
impairing diffusion and, its structure (porosity and tortuosity) may physically reduce
mass transport [30, 107-110].
The way how biofilms develop has influence on its degree of resistance: in a
biofilm, which environmental conditions make it to be very porous, with large channels,
the transport of substrates/nutrients would be easier than in a more compact one, with
tight pores [111]. The carbon and nitrogen availability, pH and temperature, all
influence EPS composition and, therefore, the resistance of biofilms can vary, by
developing zones with different densities and consequentially different mass transfer
parameters [28, 112]. The EPS matrix has sorption capacity for heavy metals, organic
substances, and particles, including nanoparticles, all of which can be caught and
accumulated within the biofilms [60].

20 Chapter 2
_________________________________________________________________________________
The polyanionic nature of the bacterial EPS may be responsible for binding the
antimicrobial agents before they have the opportunity to reach the cells [94, 113, 114].
Similarly, cells in biofilms are able to interact with the antimicrobial agents, wasting
them, so that less quantities are available to reach the under layers of the biofilm [115].
This way, it is not only the matrix, but also other biofilm components that are able to
hinder transport. Many examples are found to this reaction – diffusion phenomenon.
Davison et al. observed retardation of a QAC by the exopolymeric matrix [116]. The
work of Ciofu et al. demonstrated the increased resistance of mucoid biofilms.
P. aeruginosa that overproduces EPS showed to be up to 1000 times more resistant to
tobramycin than the normal EPS producers, in spite of similar planktonic MICs [117].
However, the selectivity of the biofilms might be related with the nature of the drugs.
A study by Anderl et al. showed that the chlorine ions and the antibiotic ciprofloxacin
were able to penetrate Klebsiella pneumonia biofilms, however ampicillin was retarded
by the biofilm components [114]. Furthermore, biofilm composition may be a factor, as
another group of investigators found that active chlorine ions were reacting with
organic matter at the surface of P. aeruginosa biofilms faster than they could diffuse
into deeper layers [118]. As a resistance mechanism, EPS works as barrier, hindering
penetration, however, the bacterial matrix could not fully inhibit penetration of the
antimicrobial agents, but instead it provides sufficient time for the induction of resistive
mechanisms to respond to the attack [119].
SPECIFIC RESISTANCE GENES
The adaptation of planktonic cells to different conditions may promote changes at their
genomic level [16]. These changes are the grounds for their specific adaptive response,
i.e. to change into biofilms [120]. Biofilm formation is regulated by many genes,
including some that are exclusive to biofilm growers [121, 122]. Mutations occur with
more frequency in biofilm embedded cells than in the planktonic state, as seen on
P. aeruginosa cells where mutations occur 105 times more when cell are in biofilms
[123]. In an effort to control P. fluorescens cells with ethylenediamine tetraacetic acid
(EDTA) and a QAC, Langsrud and Sundheim found that the cells developed resistance to
these chemicals [124].
The contact with sub-lethal concentrations of antibiotics could be the cause for
the adaptation of the outer structure of bacteria [115]. It was found that P. stutzeri had

Introduction 21 _________________________________________________________________________________
resistance to QACs, triclosan and antibiotics, after developing resistance to
chlorhexidine, probably through the alteration of the cell envelope [125]. The resistance
to several agents might be encoded into the cell genome, stress activates these genes,
deploying an active response [115].
In addition, plasmids were proven to be transferred inside biofilms. Kanamycin
resistance genes were transferred from a donor population to other cells within the
same biofilm, when this was exposed to sub-lethal concentrations of this antibiotic
[126].
QUORUM SENSING
Quorum sensing (QS) is the term used to describe cell-to-cell signaling or intracellular
signaling in bacteria. Small hormone-like signaling molecules termed auto-inducers are
produced, released and detected by the bacterial populations. High bacterial
populations inside the biofilms facilitate communication by signaling molecules
[2, 60, 97]. The detection of a threshold concentration of these molecules triggers the
expression or repression of genes, enabling bacteria in the biofilm to act as a
multicellular organism [24, 95].
Many cell regulatory systems are governed by QS, i.e. several physiological
processes such as formation, aggregation and dispersal of biofilms [127]. Deterioration
of food by enzymatic activity is also regulated by QS [128]. Cellular repair and defense
is also mediated by QS. The production of the enzymes catalase and superoxide
dismutase is regulated by QS [95]. These enzymes play a role in defense by neutralizing
toxic compounds [115]. Catalase breaks down hydrogen peroxide into water and
molecular oxygen, and superoxide dismutase promotes the degradation of superoxide
radicals [95]. P. aeruginosa biofilms presented higher catalase expression when
exposed to sub-lethal concentrations of hydrogen peroxide to biofilm cells, however
lethal to planktonic [129]. The exposure of biofilms to antimicrobials may trigger EPS
production as a defense mechanism to that stressful situation [130].
MULTIDRUG EFFLUX PUMPS
Porins are proteins forming channels that allow the transport of specific molecules
across the bacterial membrane [131]. These membranes could suffer structural

22 Chapter 2
_________________________________________________________________________________
alterations to act as a resistance mechanism to some antimicrobial agents [132]. This
system is ingenious enough to select what goes out the cell, leaving the agents outside
where the low permeability of the cells does not allow reentry, thus reducing the
accumulation inside the cell and increasing the concentration needed to inhibit the
bacteria [132]. The overproduction of efflux pumps has been reported as a biofilm
resistance mechanism [133]. QS may be involved in the protection against antimicrobial
agents, by increasing the production of these molecular pumps, with the intend to expel
drugs from the cells [95]. This is a well-known resistance mechanism [95]. Multidrug
efflux systems are usual on Gram-negative bacteria [134, 135].
PERSISTER CELLS
The indication of persister cells is the latest biofilm insusceptibility explanation [136].
Nonetheless, the first time persisters were mentioned was in 1944 by Bigger [137]. The
surviving cells from the incomplete inactivation of Staphylococcus spp. with penicillin
were able to regrow into a population with apparent penicillin susceptibility [137].
Recurring infection occurrences revived the interest into persister cells [138]. Small
subpopulations of bacteria within the biofilms may differentiate into persisters [4, 136].
These cells, present in all bacterial cultures [48], are essentially invulnerable to lethal
concentrations of antimicrobial agents [96, 139]. When exposed to antimicrobial
agents, persisters neither grow nor die [140]. Nonetheless, when the drug is removed
these bacteria give rise to a normal bacterial colony [95]. Typically, bacterial cells are
persisters against antibiotics, however, there is a study where the disinfection of
P. fluorescens biofilm embedded cells, treated with a multi-target biocide (ortho-
phthalaldehyde), resulted in ineffective killing due to the differentiation of some of
these cells into persisters [141]. The way how cells differentiate into persisters is not
well known [142]. Persisters are not believed to be mutants [143], but instead they
produce a toxin, RelE, that ceases bacterial activities, inducing an inactive state [95].
The formation of these cells could be considered as a method of adaptation to respond
to environmental alterations, and could follow two ways: in the first, the population
continues to grow, exhausting substrata risking extinction, and in the second, they
simply suppress their functions waiting for favorable conditions [30].

Introduction 23 _________________________________________________________________________________
The resistance mechanisms mentioned before, might explain biofilm persistence and
resistance to control [8]. The resistance of biofilms is defined as a multifactorial
mechanism and it is not universal, varying between different microorganisms [24].
2.5 BIOFILM CONTROL
Biofilms are a frequent source of infections and industrial process problems [105]. Many
studies have been performed in order to control biofilms in food industry. Some
strategies are already common practice in industry [144].
PREVENTION
Biofilms are difficult to eradicate, thus prevention of its occurrence is commonly
implemented in food industry. Biofilms are considered heterogeneous in time and
space, therefore research moved towards the study of biofilm mechanisms, covering
many fields of science in order to characterize the processes governing biofilms [96].
Contamination is normally caused by biofilm development due to ineffective or
complete lack of cleaning. Organic molecules are able to deposit in all types of surfaces,
including the water used for manufacturing, conditioning the surfaces and providing
favorable conditions for microbial growth [144]. The physical characteristics of the
substratum has influence on initial attachment. To prevent bacterial attachment to
surfaces, surface active substances are used. Surfactants provide uniform wetting of
surfaces, reducing the surface tension of water by adsorbing at the liquid-gas interface
and reducing the interfacial tension between the layers [24]. Additionally, the intricate
process lines of industrial plants have critical points where built up of fouling is
expected. These include gaskets, dead ends, joints, valves, corners, cracks, or crevices
[57, 145]. Rational equipment design helps reduce the risk of microbial development by
minimizing laminar product flow, reducing static product and facilitating cleaning and
disinfection processes, with the aim of reducing attachment [22]. However, a specific
design could be impractical or simply not implemented [40].
To prevent contamination, the materials that constitute the plant should be
carefully selected [57]. Good hygienic properties must be attained, either by the
material properties or by material modification to render them antimicrobial or to
reduce attachment [146, 147]. However, the application of coatings on industrial

24 Chapter 2
_________________________________________________________________________________
surfaces could be restricted by toxicity or infeasibility by increased costs [22]. The most
common materials used in food plants are stainless steel, grades 304 and 316, for their
chemical and mechanical stability at the diversified food processing temperatures, ease
of cleaning and resistance to corrosion [148]. The AISI 316 stainless steel shares the
same characteristics with the AISI 304 grade with the addition of a higher tolerance to
corrosion, given by the inclusion of molybdenum. Usually, corrosion could be caused by
food, detergents or disinfectants [22]. Polyvinylchloride is a material that is not
commonly used in food industry due to its increased risk of contamination, resultant
from its deterioration over time [44]. Stainless steel may be a better option because it
is more resistant to mechanical stresses like grinding, brushing, and electrolytic or
mechanical cleaning. For that reason, in a cleaning and disinfection plan, it is of major
importance to gather the maximum information about the system, together with flow
diagrams (information about volume, residence time, cycle time, half-life time, etc.) to
satisfy the sanitation regulations [57].
The risk of biofilm formation is increased in events such as intermittent operation,
unattended risk areas (i.e. filters), inconsistent raw water composition, lack of cleaning
after failures, and poor access to surfaces existing in the plant [149]. The risk can be
lessened by the exclusion of light, use of short piping systems, inert and smooth
materials, good air circulation, working at low temperatures, in fairly dry conditions,
and general quality control [149].
The early detection of biofilms is also used as a prompt response to outbreaks
[150]. There are currently many methods in the market. The conventional methods,
such as the count of total viable cells, microscopy and spectroscopy techniques,
impedance measurement and ATP analysis are broadly used [149, 151]. Pereira et al.
reviews the principles behind each method and the importance of monitoring in the
beverage and food industry [152].
Another strategy commonly used in food industry is the preconditioning of
surfaces to reduce/inhibit bacterial attachment [93]. For instance, biosurfactants are
known to have properties that prevent microbial attachment [153]. On the other hand,
functionalized materials, or compounds that could be blended into the material
surfaces have been reported in literature, the repulsion between the surfaces and the
bacteria also have been reported to be effective in biofilm prevention [93]. Araújo et al.
studied coated spacers used in reverse osmosis membranes with different antimicrobial
coatings, as copper, silver, gold and Polydopamine, plus a spacer infused with Triclosan.

Introduction 25 _________________________________________________________________________________
The objective was to prevent biofilm development, but this was instead only delayed in
time. Only the first layers of biofilms were killed, leaving a conditioning film that was
probably used for recolonization [154].
The frequency of cleaning is another strategy. The more often the food contact
surfaces are cleaned, the lesser the risk of microbial attachment [22]. The process of
irreversible attachment occurs so swiftly, that carefully determining the suitable
cleaning frequency becomes extremely important to avoid accumulation of microbial
and organic residues, influencing hygienic conditions as well as nutrient availability [22].
A thorough cleaning and disinfection process should occur several times a day on the
surfaces that contact directly with the product, in regular intervals varying from 4 to 24
hours [148, 155].
CLEANING AND DISINFECTION
In all industries, particularly in the food industry, the proliferation of microorganisms is
very common, even when manufacturers take all the “by-the-book” contingency plans.
Therefore, biofilms occurrence is common, leading to a need for their control using
cleaning and disinfection techniques. The aim of microbial control is the elimination or
reduction of microorganisms and their activity to acceptable levels, as well as the
prevention and control of the formation of biological deposits on process equipment
[44]. Therefore, programs are established to control microbial proliferation: two
examples are the Good Manufacturing Practice (GMP) and the Hazard Analysis and
Critical Control Points (HACCP) plans [39]. HACCP is presented in Codex Alimentarius
(CAC/RCP1- 1969- revision 2003), that was the basis for the Food Hygene directive by
the European Eunion [156]. An example of this is to include all surfaces and vectors,
such as the air or personnel that may disperse contaminations from areas such as the
floor or walls. They too should be cleaned and disinfected [93].
Biofilm control in food plants normally includes a process called Clean-in-Place
(CIP), which consists in cleaning of the plant without dismantling or opening the
equipment. During CIP, alternated cycles of detergent and disinfectant solutions run
throughout the plant with water rinses, with increased hydrodynamics (high turbulence
and flow velocities) [145]. This method typically uses caustic acids, surfactants, biocides
and, sometimes, includes enzymes [19, 145, 155, 157].

26 Chapter 2
_________________________________________________________________________________
The selection of a good cleaning and disinfection regime should be based in some
principles to be efficient. It is of utmost importance that the nature and age of
biofouling is known [158], as well as characteristics like location, type of microorganisms
(bacteria, spores, yeasts, molds or protozoa) or biological entities (prions, viruses) [159].
The characteristics of the surface to be cleaned are also relevant, because of side-
effects of cleaning on equipment materials (i.e. some cleaning agents can be corrosive)
[57]. The total number of target cells should be taken into account since high initial
numbers of bacteria may result in some persistent cells, which could result in biofilm
regrowth [63]. In cases where the concentration of the disinfectant is limited, the
bactericidal effect may be reduced in the presence of high numbers of bacteria. Also,
the growth phase of bacteria will influence their susceptibility to disinfectants. It is
known that bacteria in the exponential phase of growth are more sensitive to
disinfectants than in the stationary phase [160]. Biofilm cells are more resistant to
biocides as a result of their physiological heterogeneity and the presence of EPS, which
hinders the diffusion of biocides into the cells [161]. Cleaning is important because, in
general, disinfectants have poor diffusion and are not able to kill all the embedded cells
[93]. To clean biofilms, methods such as the use of alkali-based and acid cleaning,
scrubbing and brushing are used. However, when bases or acids are used, the
environmental conditions have to be thoroughly defined, otherwise its efficacy is
reduced [22]. Although biofilm removal can occur naturally by intrinsic processes,
mechanical removal by human action is a common strategy in food industry, though,
very expensive because of the need to open the process machinery [162]. Breaking up
and removing the deposits on surfaces is of major importance in food industry [148].
The incomplete removal could lead to reattachment and consequent biofilm regrowth
[155]. The right choice of disinfectants, single or in combination, point for biocide
injection, concentration, temperature, exposure time and hydrodynamics should be
carefully optimized for each system. The disinfectant should be kept at a concentration
equal or superior to the minimum inhibitory concentration for the period of time
defined as ideal for disinfection [44, 163]. All these parameters must be taken into
account when designing a disinfection plan.
Additionally, in order to achieve long lasting stable results, follow up actions are
required, such as monitoring the presence of microorganisms and the formation of
deposits on surfaces [42].

Introduction 27 _________________________________________________________________________________
BIOCIDES
The European Standard of 24 April 1998 (CE/8/98), defines biocidal products as active
substances, or preparations containing one or more active substances, presented to the
user in their final form, whose function is to destroy, stop the growth, render harmless,
avoid or control, by any means, the action of pathogenic organisms, by biological or
chemical processes. The use of biocides in biofilm control is well accepted and very
common [14]. Although biocides are used for the reduction of the number of
microorganisms, their simple use does not necessarily reduce the biofilm formation
rate. It is essential to use biocides correctly, because their incorrect application is
expensive and could lead to unwanted results [164].
The major groups of disinfectants used in the food industry are divided according
to their mode of action, (1) oxidizing agents e.g. chlorine-based disinfectants, ozone,
and hydrogen peroxide, (2) iodophores (iodine based disinfectants), (3) surface active
compounds like QACs and (4) weak acids [22, 40]. However, current methods of
disinfection include the application of other chemical compounds like alcohols,
aldehydes, anilides, biguanides, bis-phenols, diamidines, halophenols, and heavy metal
derivatives [165].
Most biocides kill bacteria by targeting the cytoplasmic membrane, resulting in
membrane damage such as disruption, dissipation of the proton motive force and
inhibition of membrane associated enzyme activity [70].
Each bacterial strain reacts differently to each chemical compound, either by its
phenotypic characteristics (e.g. properties of the cell wall) or due to resistance
mechanisms (coded by its genotype or induced). Thus, it is fundamental that upon
biocide selection, an evaluation of the efficacy against the dominant microorganisms
on the system is performed. Only after having information about the nature of the
microbial population to treat it is possible to determine the relation between the
minimum inhibitory concentration and the contact period of a biocide to a given
contaminant [167].
Table 2.2 provides information on the mechanisms of action, typical targets,
resulting effects and examples of biocides.

28 Chapter 2
_________________________________________________________________________________
Table 2.2 Mechanisms of interaction of several biocides according to their cellular targets and antimicrobial actions (adapted from [163]).
Cellular targets Antimicrobial action Interaction Mechanisms Examples
Chemical reactions
Thiol containing cytoplasmic and membrane bound enzymes e.g. dehydrogenases
Metabolic inhibition Oxidation of thiol groups (predominantly)
Isothiazolinone Organomercury Salts of heavy metals Hypochlorite
Biomolecules (e.g. proteins, RNA, DNA) with amino, imino, amide, carboxyl and thiol groups (nucleophilic)
Inhibition of cellular metabolism and replication. Possible cell wall damage
General alkylation reactions
Glutaraldehyde Formaldehyde Chloroacetamide
Amino groups in proteins Metabolic inhibition; lysis
Halogenation Hypochlorite Chlorine-releasing agents
Enzyme and protein thiol groups
Metabolic inhibition Free radical oxidation (e.g. hydroxyl radicals)
Hydrogen peroxide Peracetic acid
Divalent cation-mediated outer membrane integrity; principal target region Gram negative cell wall; metal ion-requiring enzyme processes
Release of cellular contents; high susceptibility to stress; metabolic inhibition.
Chelation of metal ions EDTA Oxine
Intercalation between DNA base pairs
Damage in replication Intercalation Aminoacridines
Ionic interactions
Cytoplasmic membrane integrity; membrane-bound enzyme environment and function
Leakage; respiratory inhibition; intracellular coagulation
Electrostatic interaction with phospholipids
QACs Clorhexidine Polyhexamethilene Biguanides
Physical interactions
Transmembrane pH gradient; membrane integrity
Leakage; disruption of transport, respiratory and energy coupling processes
Penetration/partition into phospholipid bilayer; possible displacement of phospholipid molecules; intra membrane molecular cycling
Phenols Weak acids Parabens Tetrachlorosalisylanilide Phenoxyethanol 2-phenylethanol
Membrane integrity Leakage Solution of phospholipids Aliphatic alcohols
Cytoplasmic membrane integrity; membrane-bound enzyme environment and function
Leakage, uncoupling of energy processes; lysis
Membrane-protein solubilization
Anionic surfactants

Introduction 29 _________________________________________________________________________________
MECHANISMS OF ANTIMICROBIAL ACTION
A classical approach which is used to determine the mechanism of action of a biocide
establishes a correlation between the minimum inhibitory concentration and the
resulting biochemical and physiological changes in the organism [163]. An antimicrobial
effect can be defined as an interaction between an active substance and specific targets
in the microbial cell. In target approach, the active ingredients contact with a variety of
cellular structures (cell wall, cytoplasmic membrane, membrane enzymes, cytoplasm,
and genetic material). Experiments conducted to compare different strains revealed
that Gram negative bacteria, which have the supplementary protection of the cell wall,
are more resistant to the bactericidal effects than Gram positive bacteria [168-170]. The
antimicrobial agents cross the cell wall through pores. This penetration, according to
Paulus, is dependent on the size, charge and lipophilic properties of molecules [71]. If a
substance is soluble in water and its molecular weight is around 600 Da, there is a
greater probability of passing through the channel formed by the porin. It is also
possible that the antimicrobial agent penetrates the cell wall after causing its
destabilization and disintegration. Finally, the biocide reaches the cytoplasmic
membrane as the primary site of action. Depending on the action spectrum, these
substances could be designated as biostatics (if they only inhibit the microorganism
growth or multiplication) or as biocides (if they are able to kill the microorganisms) [71].
The process of transporting the biocide to the cell surface, adsorption, diffusion,
penetration and interaction with the target cell component is not instantaneous and
the duration can be different according to the biocide. The differences depend on the
action mode, including the chemical composition and physicochemical properties of the
biocidal agent [163]. Biocidal compounds belong to different chemical classes.
Biocides could cause a series of self-destructive events in microorganisms,
resulting from sub-lethal/lethal damage to cell death. Typical damage caused by
biocidal compounds involves the disruption of the transmembrane proton motive force,
leading to an uncoupling of oxidative phosphorylation and inhibition of active transport
across the membrane inhibition of respiration or catabolic/anabolic reactions;
disruption of replication; loss of membrane integrity resulting in leakage of essential
intracellular constituents such as potassium cation, inorganic phosphate, pentoses,
nucleotides and nucleosides, and proteins; lysis and coagulation of intracellular material

30 Chapter 2
_________________________________________________________________________________
[163]. Figure 2.4 shows the antimicrobial mode of action of biocide on diverse types of
microorganisms.
Figure 2.4 Antimicrobial mode of action of biocides (adapted from [164]). CRAs – chlorine
removal agents; QACs – quaternary ammonium compounds.
FACTORS AFFECTING BIOCIDE ACTION
Cleaning is often inefficient in the removal of biofilms. Bactericidal activity is influenced
by the surrounding media, but a correct cleaning plan is also very important. The main
environmental factors that could influence the activity of a biocide are pH, water
hardness, presence of additives and temperature [164]. Biocide concentration,
exposure time, presence of organic compounds and type of microorganisms are key
factors of the antimicrobial action as well. Many biocides have an optimum pH range of
activity. For example, glutaraldehyde and cationic biocides such as chlorhexidine and

Introduction 31 _________________________________________________________________________________
QACs are most active at alkaline pH, whereas hypochlorites and phenolics are more
potent at acid pH. Additives, as corrosion inhibitors or conditioning agents may also
influence and even reduce or inactivate activity. The activity of biocides against Gram
negative organisms may be enhanced by permeabilizers that increase cell permeability.
Russell reported that EDTA chelates divalent cations from the outer membrane,
especially on P. aeruginosa [63]. Activity can also be increased by a combination of
biocides. In general, the efficacy of disinfectants increases with temperature [66]. When
disinfection occurs at low temperatures, the use of higher concentrations of biocides or
prolongation of the contact time may increase effectiveness [160]. The antibacterial
activity of biocides is determined by their chemical reactivity to certain organic groups.
Biocides do not react independently with fixed groups or groups of the cell surface.
Oxidizing biocides react with any oxidizing organic group, not only with living cells. In
food industries, deficient cleaning may not eliminate contaminating substances, such
as carbohydrates, fat, proteins, calcium phosphate, blood residues or dirt [57]. These
contaminants may have a high impact on the cleaning and disinfection steps. This
happens because the antimicrobial activity of chemical compounds may be reduced in
the presence of organic material, through reaction/neutralization [94, 171].
The effect of disinfectants is concentration dependent. Generally, a user-
concentration is given by the manufacturer based on simple laboratory tests that
typically measure the efficacy in suspension and without additives, which may not be
efficient to kill attached microorganisms [171]. In a practical disinfecting setting, the
disinfectant may be diluted due to residual water left after the cleaning process. In
order to avoid dilution, the equipment design should prevent, and thus facilitate,
running of water off the surfaces instead of its accumulation. Furthermore, surfaces
should be allowed to get reasonably dry before disinfection [160]. Biocides such as
phenolics or alcohols typically lose their potency with dilution, whereas QACs,
chlorhexidine, glutaraldehyde, ortho-pthalaldehyde retain much of their potency [63].
Nonetheless, the conventional protocols used for CIP have been unsatisfactory for
biofilm control [172].
Resistance is a survival mechanism that will continuously morph, ensuring
prevalence of the species. It will be necessary to find new antimicrobials and new ways
to employ them to overcome bacterial resistance [173].

32 Chapter 2
_________________________________________________________________________________
2.6 INNOVATIVE STRATEGIES FOR BIOFILM CONTROL
Many new developments have been made in order to overcome resistance, some of
these have already been put into practice in food industry.
In an effort to replace chemical disinfectants, alternative physical treatments have
been studied. Lately, the use of plasma radiation has been a theme in vogue. Ionizing
radiation consists in atmospheric plasma, that is generated using high voltage
discharges, to produce reactive oxygen species that kill microorganisms [22]. Another
method combining the action of a photosensitizer, a non-toxic dye, with visible light and
oxygen, is already used in food industry as a decontamination method. This procedure
causes DNA damage and the destruction of cellular membranes and organelles,
resulting in the leakage of cellular content [120]. In a paper by Buchovec et al. this
method was able to remove up to 3 logs L. monocytogenes biofilms [174].
Ultrasonication is another technique used for control. It has been used in various food
industry processes such as freezing, cutting, drying, softening, bleaching, sterilization,
and extraction [144]. This process is able to generate shock waves with the ability to
dislodge biofilms [175]. Besides the agitation, the ultrasounds are able to create small
vacuum bubbles that generate high temperatures when collapsed [176]. It is used as a
biofilm control strategy, already proven effective in cleaning the water in cooling towers
[177] and drinking water, without the generation of disinfection by-products [178].
Ultraviolet radiation (UV) has been employed in many systems, including the
treatment of municipal waters. Ultraviolet light is thought to be absorbed by the cells,
disrupting some processes such as replication [179]. Contrary to what would be
expected, the disinfection using ultraviolet to control biofilms present in water
distribution systems resulted in no significant biofilm reduction after the treatment, due
to the presence of other interfering substances in those systems [180, 181].
Nonetheless, UV radiation was told to be very effective eliminating planktonic and
sessile bacteria on another paper [182].
Electric fields cause a bioelectric effect that is reflected on the increase of cell
permeability [183] and, it is typically used to increase the intake of a drug into bacterial
cells [93]. Racyte et al. studied the effect of electric fields in combination with activated
carbon for disinfection of different types of bacteria in a fluidized bed electrode system
[184]. They found this system more effective against Gram positive than Gram negative
bacteria.

Introduction 33 _________________________________________________________________________________
Nanoparticles were employed as a suitable biofilm control strategy. Biofilm
formation may be inhibited in the presence of nanoparticles. In a study by
Kalishwaralal et al., biofilm formation of P aeruginosa and S. epidermidis was repressed
up to 95% using silver nanoparticles [185]. Moreover, nanoparticles have been used to
carry disinfection agents [186]. They are a promising antimicrobial strategy, because of
their high surface area to volume ratio they are thought to increase efficiency [187].
Ferreira et al. immobilized a QAC in nanoparticles. In a 1 hour disinfection process
utilizing these reusable nanoparticles, approximately 90% of a P. fluorescens biofilm was
killed.
The biofilm matrix is mainly composed of polysaccharides and proteins. The latest
studies show the potential of matrix degrading enzymes on biofilm control by the
disruption of the matrix components [120]. Enzymes hydrolyze the exopolimeric matrix
in which the bacteria are embedded. Formulations that contain enzymes are optimized
so that there is compatibility with low temperatures. They are efficient time-wise,
reducing the cleaning and disinfection time [24]. Moreover, they can work in mild pH,
temperature and high ionic strength without affecting, for instance, the membranes
used for water filtration, which are easily damaged by many chemical classes. Enzymes
were already used to control biofilms and were found to be enhancers of the action of
antimicrobial agents [10, 161, 188]. Nevertheless, enzymes are substrate specific [24],
and the efficacy of the method is dependent on the right use and right combination of
enzymes, being often suggested the characterization of EPS before the enzymatic
treatments [59].
The evolution of resistance [189], the possible failure of antimicrobial agents [79],
and the formation of harmful byproducts [190] translate into the need for new
antimicrobial agents [191]. These should be effective against the bacterial
contamination [192]. However, as legislation restricts the use of toxic biocides, eco-
friendly strategies represent a new approach for biofilm control [193, 194]. For
instance, chlorine could react with organic matter, resulting in cancer-forming
compounds that might enter the food chain [190]. Consequently, the exploitation of
“green” biocides, from plant sources has been on course [192]. Valeriano et al.
identified the antimicrobial properties of peppermint and lemongrass essential oils
against biofilm formation of S. enterica [195]. Rhodiola crenulata (arctic root),
Epimedium brevicornum (rowdy lamb herb), and Polygonum cuspidatum (Japanese
knotweed) extracts also showed anti-biofilm properties against Propionibacterium

34 Chapter 2
_________________________________________________________________________________
acnes, reducing the biofilms 64.8%, 98.5%, and 99.2%, respectively [196]. Also
Melia dubia (bead tree) bark extracts reduced E. coli formation and swarming by 84%
and 75% [197]. Ferulic, gallic and salicylic acids, considered to be phenolic compounds,
were tested against different bacterial biofilms with favorable control results [191, 198].
Chitosan, a polysaccharide, exhibited anti-biofilm properties against S. mutans. This
compound was able to reduce biofilms by approximately 95% [199]. It was also tested
againts L. monocytogenes, B. cereus, S. aureus, S. enterica, and P. fluorescens. The
biofilms developed by these bacteria were reduced from a maximum of 6 logs
(L. monocytogens) to a minimum of 1 log (S. aureus) [200].
QS interference is an alternative approach to biofilm control by targeting the
signaling molecules that control various cell processes, including biofilm formation
[127]. QS is a biochemical approach to a direct control of the rate and extent of biofilm
development, as opposed to cleaning and disinfection techniques [201]. QS inhibitors
impair the communication signals between cells in the biofilms [93]. One of the most
potent QS quenchers are the halogenated compounds secreted by the red algae Delisea
pulchra [202]. Other QS inhibitor substances are brominated furanones. Although the
mode of action of these drugs is not yet fully understood, it is thought that it inhibits QS
[203]. A biofilm treated with these chemicals is thought to have a higher susceptibility
to disinfectants [24]. The strategy is to take advantage of the quorum signals used for
biofilm regulatory mechanism [204]. In general, the study of which molecules regulate
QS in food industry, to find corresponding inhibitors, could increase food safety and
product shelf life [128]. The QS signaling molecules could be detected using biosensors
[128]. Then, providing the inhibiting signals, or manipulating their mechanisms
convincing bacteria not to form biofilms or triggering dispersal, biofilms could be
controlled. For instance, P. aeruginosa produces rhamnolipid biosurfactants to detach
from surfaces [205]. Inducing this bacteria to produce higher amounts of this chemical
could result in effective control [204].
Phages are very simple, and the most abundant organisms on earth. They are
viruses that infect bacteria, and like viruses they are only able to replicate inside their
host [206]. Phages have known to be applied, initially in the early 20th century, to treat
bacterial infections in Eastern Europe [144]. There are phages with extreme specificity
and others which specificity is broader [206]. Treatments with phages showed potential
to inhibit biofilm formation. As they are not chemical-based, their use eliminates the
risk of surface corrosion, and due to their high specificity and non-toxicity they are good

Introduction 35 _________________________________________________________________________________
candidates as therapy for biofilm infections in living hosts [144]. The co-existence of
phages and bacteria is known in biofilms, which is one of the reasons why the
combination of phages with disinfectants and polysaccharide depolymerases was
suggested to be a novel control strategy [207]. Moreover, phages can be engineered to
express biofilm degrading enzymes [208]. The use of phages as biofilm control resulted
in a removal of 99.997% of bacteria [208]. The only drawback of the usage of phages is
that their use for biofilm control might select resistant bacteria [36].
New strategies have been boosted by environmental restrictions. The combination
of two or more strategies could control biofilms synergistically, approaching the
problem in multiple fronts could be another alternative to overcome the persistence of
biofilms [144, 209]. This technique is referred to as hurdle technology and it is widely
used across industries. Any combination is valid as long as it is effective and abides to
the current law. The right combination of hurdles should prevent, reduce or completely
eliminate biofilms [144]. Therefore by combining different chemicals should broaden
their antimicrobial spectrum, if they are able to work synergistically [210]. For example,
treatments with ultrasounds in combination with enzymes and ozone were effective
against established biofilms [22]. To prevent clinical infections in the operating block,
surgical blades have been coated with a mixture of silver nanoparticles and lysozyme,
effectively reducing infections by many clinical pathogens. [211]. Oulahal-Lagsir et al.
reported the combination of an ultrasonic technique with a chelating agents (EDTA) to
be effective in the removal of E. coli biofilms [212].
REFERENCES
[1] Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: from the natural environmental to infectious diseases. Nature Reviews in Microbiology, 2004. 2:95-108.
[2] Flemming H-C, Biofilms, in Encyclopedia of Life Sciences (ELS). 2008, John Wiley & Sons, Ltd: Chichester.
[3] Simões M, Simões LC, Vieira MJ. Species association increases biofilm resistance to chemical and mechanical treatments. Water Research, 2009. 43(1):229-237.
[4] Paraje MG, Antimicrobial resistance in biofilms, in Science against microbial pathogens: communicating current research and technological advances, M.-V. A, Editor. 2011, Formatex: Badajoz, Spain. p. 826-834.
[5] Costerton J, Geesey G, Cheng K. How bacteria stick. Scientific American, 1978. 238(1):86-95.
[6] Characklis W, Marshall K. Biofilms: a basis for an interdisciplinary approach. Biofilms, 1990:3-15.
[7] Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappinscott HM. Microbial Biofilms. Annual Review of Microbiology, 1995. 49:711-745.

36 Chapter 2
_________________________________________________________________________________
[8] Stoodley P, Boyle JD, DeBeer D, Lappin-Scotta HM. Evolving perspectives of biofilm structure. Biofouling, 1999. 14(1):75-90.
[9] Oulahal‐Lagsir N, Martial‐Gros A, Bonneau M, Blum LJ. “Escherichia coli‐milk” biofilm removal from stainless steel surfaces: Synergism between ultrasonic waves and enzymes. Biofouling, 2003. 19(3):159-168.
[10] Andersson S, Dalhammar G, Land C, Kuttuva Rajarao G. Characterization of extracellular polymeric substances from denitrifying organism Comamonas denitrificans. Applied Microbiology and Biotechnology, 2009. 82(3):535-543.
[11] Gao BY, Zhu XB, Xu CH, Yue QY, Li WW, Wei JC. Influence of extracellular polymeric substances on microbial activity and cell hydrophobicity in biofilms. Journal of Chemical Technology and Biotechnology, 2008. 83(3):227-232.
[12] Liu Y, Tay J-H. The essential role of hydrodynamic shear force in the formation of biofilm and granular sludge. Water Research, 2002. 36(7):1653-1665.
[13] Sutherland I. The biofilm matrix - an immobilized but dynamic microbial environment. Trends in Microbiology, 2001. 9(5):222-227.
[14] Donlan R, Costerton J. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clinical Microbiology Reviews, 2002. 15(2):167-193.
[15] Sauer K, Camper AK, Ehrlich GD, Costerton JW, Davies DG. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. Journal of Bacteriology, 2002. 184(4):1140-1154.
[16] Stoodley P, Sauer K, Davies D, Costerton JW. Biofilms as complex differentiated communities. Annual Reviews in Microbiology, 2002. 56(1):187-209.
[17] Davies D. Understanding biofilm resistance to antibacterial agents. Nature Reviews in Drug Discovery 2003. 2:114-122.
[18] Teodósio JS, Silva FC, Moreira JMR, Simões M, Melo LF, Alves MA, Mergulhão FJ. Flow cells as quasi-ideal systems for biofouling simulation of industrial piping systems. Biofouling, 2013. 29(8):953-966.
[19] Shi X, Zhu X. Biofilm formation and food safety in food industries. Trends in Food Science & Technology, 2009. 20(9):407-413.
[20] Simões M, Simões LC, Vieira MJ. Physiology and behavior of Pseudomonas fluorescens single and dual strain biofilms under diverse hydrodynamics stresses. International Journal of Food Microbiology, 2008. 128(2):309-316.
[21] Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappin-Scott HM. Microbial biofilms. Annual Reviews in Microbiology, 1995. 49(1):711-745.
[22] van Houdt R, Michiels CW. Biofilm formation and the food industry, a focus on the bacterial outer surface. Journal of Applied Microbiology, 2010. 109(4):1117-1131.
[23] Bottero S, Storck T, Heimovaara TJ, van Loosdrecht MCM, Enzien MV, Picioreanu C. Biofilm development and the dynamics of preferential flow paths in porous media. Biofouling, 2013. 29(9):1069-1086.
[24] Anand S, Singh D, Avadhanula M, Marka S. Development and control of bacterial biofilms on dairy processing membranes. Comprehensive Reviews in Food Science and Food Safety, 2014. 13(1):18-33.
[25] Wäsche S, Horn H, Hempel DC. Influence of growth conditions on biofilm development and mass transfer at the bulk/biofilm interface. Water Research, 2002. 36(19):4775-4784.
[26] Beyenal H, Lewandowski Z. Internal and external mass transfer in biofilms grown at various flow velocities. Biotechnology Progress, 2002. 18(1):55-61.

Introduction 37 _________________________________________________________________________________
[27] Vieira MJ, Melo LF. Intrinsic kinetics of biofilms formed under turbulent flow and low substrate concentrations. Bioprocess and Biosystems Engineering, 1999. 20(4):369-375.
[28] Czaczyk K, Myszka K. Biosynthesis of extracellular polymeric substances (EPS) and its role in microbial biofilm formation. Polish Journal of Environmental Studies, 2007. 16(6):799-806.
[29] Ivanovic I, Leiknes T. The biofilm membrane bioreactor (BF-MBR)—a review. Desalination and Water Treatment, 2012. 37(1-3):288-295.
[30] Nikolaev Y, Plakunov V. Biofilm - “City of microbes” or an analogue of multicellular organisms? Microbiology, 2007. 76(2):125-138.
[31] Meyer B. Approaches to prevention, removal and killing of biofilms. International Biodeterioration & Biodegradation, 2003. 51(4):249-253.
[32] Dooley JSG, Roberts TA. Control of vegetative micro-organisms in foods. British Medical Bulletin, 2000. 56(1):142-157.
[33] Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV, Widdowson M-A, Roy SL, Jones JL, Griffin PM. Foodborne illness acquired in the United States-major pathogens. Emerging infectious diseases, 2011. 17(1).
[34] Potera C. Microbiology - Forging a link between biofilms and disease. Science, 1999. 283(5409):1837.
[35] Wolcott R, Costerton JW, Raoult D, Cutler SJ. The polymicrobial nature of biofilm infection. Clinical Microbiology and Infection, 2013. 19(2):107-112.
[36] Brooks JD, Flint SH. Biofilms in the food industry: problems and potential solutions. International Journal of Food Science & Technology, 2008. 43(12):2163-2176.
[37] Endersen L, O'Mahony J, Hill C, Ross RP, McAuliffe O, Coffey A. Phage therapy in the food industry. Annual Review of Food Science and Technology, 2014. 5:327-349.
[38] Simões M, Pereira M, Vieira M. Validation of respirometry as a short-term method to assess the efficacy of biocides. Biofouling, 2005. 21(1):9-17.
[39] Sharma M, Anand SK. Biofilms evaluation as an essential component of HACCP for food/dairy processing industry - a case. Food Control, 2001. 13(6-7):469-477.
[40] Chmielewski RAN, Frank JF. Biofilm formation and control in food processing facilities. Comprehensive Reviews in Food Science and Food Safety, 2003. 2(1):22-32.
[41] Srinivasan R, Stewart PS, Griebe T, Chen, . Biofilm parameters influencing biocide efficacy. Biotechnology and Bioengineering, 1995. 46:553–560.
[42] Kumar CG, Anand SK. Significance of microbial biofilms in food industry: a review. International Journal of Food Microbiology, 1998. 42(1-2):9-27.
[43] Bryers JD. Understanding and controlling detrimental bioreactor biofilms. Trends in Biotechnology, 1991. 9(12):422-426.
[44] Maukonen J, Mättö J, Wirtanen G, Raaska L, Mattila-Sandholm T, Saarela M. Methodologies for the characterization of microbes in industrial environments: a review. Journal of Industrial Microbiology & Biotechnology, 2003. 30(6):327-356.
[45] Jahid IK, Ha SD. A review of microbial biofilms of produce: Future challenge to food safety. Food Science and Biotechnology, 2012. 21(2):299-316.
[46] Gomes LC, Moreira JMR, Miranda JM, Simões M, Melo LF, Mergulhão FJ. Macroscale versus microscale methods for physiological analysis of biofilms formed in 96-well microtiter plates. Journal of Microbiological Methods, 2013. 95(3):342-349.
[47] Beyenal H, Lewandowski Z, Harkin G. Quantifying biofilm structure: facts and fiction. Biofouling, 2004. 20(1):1-23.

38 Chapter 2
_________________________________________________________________________________
[48] Karatan E, Watnick P. Signals, regulatory networks, and materials that build and break bacterial biofilms. Microbiology and Molecular Biology Reviews, 2009. 73(2):310-347.
[49] Molobela IP. Proteolytic and amyolytic enzymes for bacterial biofilm control. 2010, University of Pretoria: South Africa.
[50] Flemming H. The EPS matrix: The "house of biofilm cells". Journal of Bacteriology, 2007. 189(22):7945-7947.
[51] Smirnova T, Didenko L, Azizbekyan R, Romanova Y. Structural and functional characteristics of bacterial biofilms. Microbiology, 2010. 79(4):413-423.
[52] Bryers J. Biofilms and the technological implications of microbial cell-adhesion. Colloids and Surfaces B-Biointerfaces, 1994. 2(1-3):9-23.
[53] Bryers J, Mixed Population Biofilms, in Biofilms - Science and Technology, T.R.B. L. F. Melo, M. Fletcher, B. Capdeville, Editor. 1992. p. 277-289.
[54] Bridier A, Briandet R, Thomas V, Dubois-Brissonnet F. Resistance of bacterial biofilms to disinfectants: a review. Biofouling, 2011. 27(9):1017-1032.
[55] Donlan RM. Biofilms: microbial life on surfaces. Emerging Infectious Diseases 2002. 8(9): 881–890.
[56] Simões M, Simões LC, Vieira MJ. A review of current and emergent biofilm control strategies. LWT - Food Science and Technology, 2010. 43(4):573-583.
[57] Decreto-Lei no 306/2007 de 27 de Agosto, relativo ao controlo da qualidade da água destinada ao consumo humano.
[58] García López LA, Veiga MC, Nogueira R, Aparicio A, Melo LF. A technique using a membrane flow cell to determine average mass transfer coefficients and tortuosity factors in biofilms. Water Science and Technology, 2003. 47(5):61-67.
[59] Ras M, Lefebvre D, Derlon N, Paul E, Girbal-Neuhauser E. Extracellular polymeric substances diversity of biofilms grown under contrasted environmental conditions. Water Research, 2011. 45(4):1529-1538.
[60] Flemming HC. The perfect slime. Colloids and Surfaces B-Biointerfaces, 2011. 86(2):251-259.
[61] Wingender J, Neu TR, Flemming H-C, What are bacterial extracellular polymeric substances?, in Microbial extracellular polymeric substances. 1999, Springer. p. 1-19.
[62] Babu MM, Aravind L. Adaptive evolution by optimizing expression levels in different environments. Trends in Microbiology, 2006. 14(1):11-14.
[63] Russell AD. Biocide use and antibiotic resistance: the relevance of laboratory findings to clinical and environmental situations. The Lancet Infectious Diseases, 2003. 3(12):794-803.
[64] Gilbert P, Collier PJ, Brown M. Influence of growth rate on susceptibility to antimicrobial agents: biofilms, cell cycle, dormancy, and stringent response. Antimicrobial Agents and Chemotherapy, 1990. 34(10):1865.
[65] Gilbert P, Allison DG, McBain AJ. Biofilms in vitro and in vivo: do singular mechanisms imply cross-resistance? Journal of Applied Microbiology, 2002. 92:98s-110s.
[66] Russell A. Similarities and differences in the responses of microorganisms to biocides. Journal of Antimicrobial Chemotherapy, 2003. 52:750-763.
[67] Russell A. Bacterial resistance to disinfectants: present knowledge and future problems. Journal of Hospital Infection, 1999. 43(SUPPL. 1).
[68] Machado I, Graca J, Sousa AM, Lopes SP, Pereira MO. Effect of antimicrobial residues on early adhesion and biofilm formation by wild-type and benzalkonium chloride-adapted Pseudomonas aeruginosa. Biofouling, 2011. 27(10):1151-9.

Introduction 39 _________________________________________________________________________________
[69] Pogiatzis T, Vassiliadis V, Mergulhão F, Wilson D. When to clean, how to clean: Biofilms. in International Conference of heat exchanger fouling and cleaning 2013. Budapest, Hungary.
[70] Maillard J. Bacterial target sites for biocide action. Journal of Applied Microbiology, 2002. 92:16s-27s.
[71] Paulus W, Relationship between chemical structure and activity or mode of action of micro biocides, in Directory of microbicides for the protection of materials - A Handbook, W. Paulus, Editor. 1993, Kluwer Academic Publishers: Dordrecht, The Netherlands. p. 787.
[72] Cloete T. Resistance mechanisms of bacteria to antimicrobial compounds. International Biodeterioration & Biodegradation, 2003. 51(4):277-282.
[73] Chapman J. Biocide resistance mechanisms. International Biodeterioration & Biodegradation, 2003. 51(2):133-138.
[74] Jacobsen L, Wilcks A, Hammer K, Huys G, Gevers D, Andersen SR. Horizontal transfer of tet(M) and erm(B) resistance plasmids from food strains of Lactobacillus plantarum to Enterococcus faecalis JH2-2 in the gastrointestinal tract of gnotobiotic rats. FEMS Microbiology Ecology, 2007. 59(1):158-166.
[75] Stewart PS. Mechanisms of antibiotic resistance in bacterial biofilms. International Journal of Medical Microbiology, 2002. 292(2):107-113.
[76] Zhang L, Mah TF. Involvement of a novel efflux system in biofilm-specific resistance to antibiotics. Journal of Bacteriology, 2008. 190(13):4447-4452.
[77] Sundheim G, Langsrud S, Heir E, Holck AL. Bacterial resistance to disinfectants containing quaternary ammonium compounds. International Biodeterioration & Biodegradation, 1998. 41(3-4):235-239.
[78] Spratt BG. Resistance to antibiotics mediated by target alterations. Science, 1994. 264(5157):388-396.
[79] Simões M, Pereira M, Machado I, Simões L, Vieira M. Comparative antibacterial potential of selected aldehyde-based biocides and surfactants against planktonic Pseudomonas fluorescens. Journal of Industrial Microbiology & Biotechnology, 2006. 33(9):741-749.
[80] Shahverdi AR, Fakhimi A, Shahverdi HR, Minaian S. Synthesis and effect of silver nanoparticles on the antibacterial activity of different antibiotics against Staphylococcus aureus and Escherichia coli. Nanomedicine: Nanotechnology, Biology and Medicine, 2007. 3(2):168-171.
[81] Shafahi M, Vafai K. Synthesis of biofilm resistance characteristics against antibiotics. International Journal of Heat and Mass Transfer, 2010. 53(15-16):2943-2950.
[82] Russell AD. Bacterial adaptation and resistance to antiseptics, disinfectants and preservatives is not a new phenomenon. Journal of Hospital Infection, 2004. 57(2):97-104.
[83] Nakata K, Tsuchido T, Matsumura Y. Antimicrobial cationic surfactant, cetyltrimethylammonium bromide, induces superoxide stress in Escherichia coli cells. Journal of Applied Microbiology, 2010. 110:568-579.
[84] Gilbert P, McBain AJ. Potential impact of increased use of biocides in consumer products on prevalence of antibiotic resistance. Clinical Microbiology Reviews, 2003. 16(2):189-208.
[85] Beumer R, Bloomfield S, Exner M, Fara G, Nath K, Scott E. Microbial resistance and biocides - a review and consensus statement. 2000, International Scientific Forum on Home Hygiene.
[86] Sakagami Y, Yokoyama H, Nishimura H, Ose Y, Tashima T. Mechanism of resistance to benzalkonium chloride by Pseudomonas aeruginosa in applied and environmental microbiology. Jounal of Applied Microbiology, 1989. 55(8):2036–2040.
[87] Simões M, Simões LC, Pereira MO, Vieira MJ. Sodium dodecyl sulfate allows the persistence and recovery of biofilms of Pseudomonas fluorescens formed under different hydrodynamic conditions. Biofouling, 2008. 24(1):35-44.

40 Chapter 2
_________________________________________________________________________________
[88] Simões M, Pereira MO, Vieira MJ. Effect of mechanical stress on biofilms challenged by different chemicals. Water Research, 2005. 39(20):5142-5152.
[89] Diehl MA, Chapman JS. Association of the biocide 5-chloro-2-methyl-isothiazol-3-one with Pseudomonas aeruginosa and Pseudomonas fluorescens. International Biodeterioration and Biodegradation, 1999. 44(4):191-199.
[90] Simões M, Pereira M, Sillankorva S, Azeredo J, Vieira M. The effect of hydrodynamic conditions on the phenotype of Pseudomonas fluorescens biofilms. Biofouling, 2007. 23(3-4):249-58.
[91] Davies DG, Geesey GG. Regulation of the alginate biosynthesis gene algC in Pseudomonas aeruginosa during biofilm development in continuous culture. Applied and Environmental Microbiology, 1995. 61(3):860-867.
[92] Costerton J. Introduction to biofilm. International Journal of Antimicrobial Agents, 1999. 11(3-4):217-221.
[93] Cappitelli F, Polo A, Villa F. Biofilm formation in food processing environments is still poorly understood and controlled. Food Engineering Reviews, 2014. 6(1):29-42.
[94] Mah T, O'Toole G. Mechanisms of biofilm resistance to antimicrobial agents. Trends in Microbiology, 2001. 9(1):34-39.
[95] Harrison J, Turner R, Marques L, Ceri H. A new understanding of these microbial communities is driving a revolution that may transform the science of microbiology. Am Sci, 2005. 93(6):508-515.
[96] Stewart PS, Franklin MJ. Physiological heterogeneity in biofilms. Nature Reviews in Microbiology, 2008. 6(3):199-210.
[97] Watnick P, Kolter R. Biofilm, city of microbes. Jounal of Bacteriology, 2000. 182(10):2675-2679.
[98] Desai M, Buhler T, Weller PH, Brown MRW. Increasing resistance of planktonic and biofilm cultures of Burkholderia cepacia to ciprofloxacin and ceftazidime during exponential growth. Journal of Antimicrobial Chemotherapy, 1998. 42(2):153-160.
[99] Allison DG, Ruiz B, SanJose C, Jaspe A, Gilbert P. Extracellular products as mediators of the formation and detachment of Pseudomonas fluorescens biofilms. FEMS Microbiology Letters, 1998. 167(2):179-184.
[100] Stewart P. Diffusion in biofilms. Journal of Bacteriology, 2003. 185(5):1485-1491.
[101] Davey ME, O'Toole GA. Microbial biofilms: from ecology to molecular genetics. Microbiology and Molecular Biology Reviews, 2000. 64(4):847-867.
[102] Schulte S, Wingender J, Flemming H-C, Efficacy of biocides against biofilms, in Directory of Microbicides for the Protection of Materials, W. Paulus, Editor. 2005, Springer Netherlands. p. 93-120.
[103] Bester E, Kroukamp O, Wolfaardt GM, Boonzaaier L, Liss SN. Metabolic differentiation in biofilms as indicated by carbon dioxide production rates. Applied and Environmental Microbiology, 2010. 76(4):1189-1197.
[104] Brown MR, Allison DG, Gilbert P. Resistance of bacterial biofilms to antibiotics a growth-rate related effect? Journal of Antimicrobial Chemotherapy, 1988. 22(6):777-780.
[105] Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Science 1999. 284(5418):1318-22.
[106] Evans DJ, Allison DG, Brown MRW, Gilbert P. Susceptibility of Pseudomonas aeruginosa and Escherichia coli biofilms towards ciprofloxacin - effect of specific growth-rate. Journal of Antimicrobial Chemotherapy, 1991. 27(2):177-184.
[107] Stewart P, Grab L, Diemer J. Analysis of biocide transport limitation in an artificial biofilm system. Journal of Applied Microbiology, 1998. 85(3):495-500.

Introduction 41 _________________________________________________________________________________
[108] Russell A, Furr J, Maillard J. Microbial susceptibility and resistance to biocides. American Society for Microbiology News, 1997. 63(9):481-487.
[109] Flemming H, Wingender J. The biofilm matrix. Nature Reviews in Microbiology, 2010. 8(9):623-633.
[110] Costerton JW, Lewandowski Z, deBeer D, Caldwell D, Korber D, James G. Biofilms, the customized microniche. Journal of Bacteriology, 1994. 176(8):2137-2142.
[111] Melo L. Biofilm physical structure, internal diffusivity and tortuosity. Water Science and Technology, 2005. 52(7):77-84.
[112] Sandt C, Barbeau J, Gagnon MA, Lafleur M. Role of the ammonium group in the diffusion of quaternary ammonium compounds in Streptococcus mutans biofilms. Journal of Antimicrobial Chemotherapy, 2007. 60(6):1281-1287.
[113] Singh R, Ray P, Das A, Sharma M. Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. Journal of Antimicrobial Chemotherapy, 2010. 65:1955-1958.
[114] Anderl J, Franklin M, Stewart P. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Journal of Antimicrobial Chemotherapy, 2000. 44(7):1818-1824.
[115] McDonnell G, Russell A. Antiseptics and disinfectants: activity, action, and resistance. Clinical Microbiology Reviews, 1999. 12(1):147-179.
[116] Davison WM, Pitts B, Stewart PS. Spatial and temporal patterns of biocide action against Staphylococcus epidermidis biofilms. Antimicrobial Agents and Chemotherapy, 2010. 54(7):2920-2927.
[117] Ciofu O, Mandsberg LF, Wang H, Høiby N. Phenotypes selected during chronic lung infection in cystic fibrosis patients: implications for the treatment of Pseudomonas aeruginosa biofilm infections. FEMS Immunology & Medical Microbiology, 2012. 65(2):215-225.
[118] Chen X, Stewart PS. Chlorine penetration into artificial biofilm is limited by a reaction−diffusion interaction. Environmental Science and Technology, 1996. 30(6):2078-2083.
[119] Jefferson KK, Goldmann DA, Pier GB. Use of confocal microscopy to analyze the rate of vancomycin penetration through Staphylococcus aureus biofilms. Antimicrobial Agents and Chemotherapy, 2005. 49(6):2467-2473.
[120] Taraszkiewicz A, Fila G, Grinholc M, Nakonieczna J. Innovative strategies to overcome biofilm resistance. BioMed Research International, 2013. 2013(1): 1-13.
[121] Szczuka E, Urbańska K, Pietryka M, Kaznowski A. Biofilm density and detection of biofilm-producing genes in methicillin-resistant Staphylococcus aureus strains. Folia Microbiologica, 2013. 58(1):47-52.
[122] Yadav MK, Kwon SK, Cho CG, Park SW, Chae SW, Song JJ. Gene expression profile of early in vitro biofilms of Streptococcus pneumoniae. Microbiology and immunology, 2012. 56(9):621-629.
[123] Driffield K, Miller K, Bostock J, O'neill A, Chopra I. Increased mutability of Pseudomonas aeruginosa in biofilms. Journal of Antimicrobial Chemotherapy, 2008. 61(5):1053-1056.
[124] Langsrud S, Sundheim G. Factors contributing to the survival of poultry associated Pseudomonas spp. exposed to a quaternary ammonium compound. Journal of Applied Microbiology, 1997. 82(6):705-712.
[125] Russell A, Tattawasart U, Maillard J-Y, Furr J. Possible link between bacterial resistance and use of antibiotics and biocides. Antimicrobial agents and chemotherapy, 1998. 42(8):2151-2151.
[126] Ma H, Bryers JD. Non-invasive determination of conjugative transfer of plasmids bearing antibiotic-resistance genes in biofilm-bound bacteria: effects of substrate loading and antibiotic selection. Applied Microbiology and Biotechnology, 2013. 97(1):317-328.

42 Chapter 2
_________________________________________________________________________________
[127] Smith JL, Fratamico PM, Novak JS. Quorum sensing: a primer for food microbiologists. Journal of Food Protection®, 2004. 67(5):1053-1070.
[128] Bai AJ, Rai VR. Bacterial quorum sensing and food industry. Comprehensive Reviews in Food Science and Food Safety, 2011. 10(3):183-193.
[129] Szomolay B, Klapper I, Dockery J, Stewart PS. Adaptive responses to antimicrobial agents in biofilms. Environmental Microbiology, 2005. 7(8):1186-1191.
[130] Bagge N, Hentzer M, Andersen JB, Ciofu O, Givskov M, Høiby N. Dynamics and spatial distribution of β-lactamase expression in Pseudomonas aeruginosa biofilms. Antimicrobial Agents and Chemotherapy, 2004. 48(4):1168-1174.
[131] Wong K, Ma J, Rothnie A, Biggin PC, Kerr ID. Towards understanding promiscuity in multidrug efflux pumps. Trends in Biochemical Sciences, 2014. 39(1):8-16.
[132] Vila J, Martí S, Sánchez-Céspedes J. Porins, efflux pumps and multidrug resistance in Acinetobacter baumannii. Journal of Antimicrobial Chemotherapy, 2007. 59(6):1210-1215.
[133] Fernández L, Hancock RE. Adaptive and mutational resistance: role of porins and efflux pumps in drug resistance. Clinical Microbiology Reviews, 2012. 25(4):661-681.
[134] Poole K. Multidrug efflux pumps and antimicrobial resistance in Pseudomonas aeruginosa and related organisms. Journal of Molecular Microbiology and Biotechnology, 2001. 3(2):255-263.
[135] Poole K. Efflux-mediated multiresistance in Gram-negative bacteria. Clinical Microbiology and Infection, 2004. 10(1):12-26.
[136] Lewis K. Persister cells, dormancy and infectious disease. Nature Reviews in Microbiology, 2007. 5(1):48-56.
[137] Bigger JW. Treatment of Staphylococcal infections with penicillin by intermittent sterilisation. The Lancet, 1944. 244(6320):497-500.
[138] Balaban NQ, Merrin J, Chait R, Kowalik L, Leibler S. Bacterial persistence as a phenotypic switch. Science, 2004. 305(5690):1622-1625.
[139] José Miguel Sahuquillo Arce, Fernanda Yarad Auad, Cabezas AH, Biofilms: a biological antimicrobial resistance system, in Microbial pathogens and strategies for combating them: science, technology and education A. Méndez-Vilas, Editor. 2013, Formatex Research Center: Badajoz, Spain. p. 61-67.
[140] Keren I, Shah D, Spoering A, Kaldalu N, Lewis K. Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli. Journal of Bacteriology, 2004. 186(24):8172-8180.
[141] Simões LC, Lemos M, Pereira AM, Abreu AC, Saavedra MJ, Simões M. Persister cells in a biofilm treated with a biocide. Biofouling, 2011. 27(4):403-411.
[142] Gefen O, Balaban NQ. The importance of being persistent: heterogeneity of bacterial populations under antibiotic stress. FEMS microbiology reviews, 2009. 33(4):704-717.
[143] Lewis K. Riddle of biofilm resistance. Antimicrobial Agents and Chemotherapy, 2001. 45(4):999-1007.
[144] Srey S, Jahid IK, Ha SD. Biofilm formation in food industries: A food safety concern. Food Control, 2013. 31(2):572-585.
[145] Bremer PJ, Fillery S, McQuillan AJ. Laboratory scale Clean-In-Place (CIP) studies on the effectiveness of different caustic and acid wash steps on the removal of dairy biofilms. International Journal of Food Microbiology, 2006. 106(3):254-262.
[146] Araújo PA, Kruithof JC, van Loosdrecht MCM, Vrouwenvelder JS. The potential of standard and modified feed spacers for biofouling control. Journal of Membrane Science, 2012. 403–404(0):58-70.

Introduction 43 _________________________________________________________________________________
[147] Miller DJ, Araújo PA, Correia PB, Ramsey MM, Kruithof JC, van Loosdrecht MCM, Freeman BD, Paul DR, Whiteley M, Vrouwenvelder JS. Short-term adhesion and long-term biofouling testing of polydopamine and poly(ethylene glycol) surface modifications of membranes and feed spacers for biofouling control. Water Research, 2012. 46(12):3737-3753.
[148] Zottola EA, Sasahara KC. Microbial biofilms in the food processing industry—Should they be a concern? International Journal of Food Microbiology, 1994. 23(2):125-148.
[149] Verran J. Biofouling in food processing: biofilm or biotransfer potential? Food and Bioproducts Processing, 2002. 80(4):292-298.
[150] Vrouwenvelder JS, van Loosdrecht MCM, Kruithof JC. Early warning of biofouling in spiral wound nanofiltration and reverse osmosis membranes. Desalination, 2011. 265(1–3):206-212.
[151] Strathmann M, Mittenzwey K-H, Sinn G, Papadakis W, Flemming H-C. Simultaneous monitoring of biofilm growth, microbial activity, and inorganic deposits on surfaces with an in situ, online, real-time, non-destructive, optical sensor. Biofouling, 2013. 29(5):573-583.
[152] Pereira A, Melo, L. F., Fratamico, P. M., Annous, B. A., & Gunther, N. W. , Monitoring of biofilms in the food and beverage industries, in Biofilms in the food and beverage industries, P.M. Fratamico, Annous, B. A., & Guenther, N. W. , Editor. 2009, Elsevier. p. 131-151.
[153] Nitschke M, Costa S. Biosurfactants in food industry. Trends in Food Science & Technology, 2007. 18(5):252-259.
[154] P. A. Araújo, D. J. Miller, P. B. Correia, M. C. M. van Loosdrecht, J. C. Kruithof, B. D. Freeman, D. R. Paul, Vrouwenvelder JS. Impact of feed spacer and membrane modification by hydrophilic, bactericidal and biocidal coating on biofouling control. Desalination, 2012. 295(0):1-10.
[155] Gibson H, Taylor J, Hall K, Holah J. Effectiveness of cleaning techniques used in the food industry in terms of the removal of bacterial biofilms. Journal of Applied Microbiology, 1999. 87(1):41-48.
[156] World Health Organization, and Codex Alimentarius Commission. Codex Alimentarius, 2013. FAO/WHO.
[157] Otzen D. Protein-surfactant interactions: a tale of many states. Biochimica et Biophysica Acta - Proteins & Proteomics, 2011. 1814(5):562-591.
[158] Anwar H, Strap JL. Changing characteristics of aging biofilms. International Biodeterioration & Biodegradation, 1992. 30(2-3):177-186.
[159] Russell AD, McDonnell G. Concentration: a major factor in studying biocidal action. Journal of Hospital Infection, 2000. 44(1):1-3.
[160] Møretrø T, Heir E, Nesse LL, Vestby LK, Langsrud S. Control of Salmonella in food related environments by chemical disinfection. Food Research International, 2012. 45(2):532-544.
[161] Ganeshnarayan K, Shah SM, Libera MR, Santostefano A, Kaplan JB. Poly-N-Acetylglucosamine matrix polysaccharide impedes fluid convection and transport of the cationic surfactant cetylpyridinium chloride through bacterial biofilms. Applied and Environmental Microbiology, 2009. 75(5):1308-1314.
[162] Araújo PA, Lemos M, Mergulhão F, Melo L, Simões M, Antimicrobial resistance in biofilms to disinfectants, in Science against microbial pathogens: communicating current research and technological advances, A. Méndez-Vilas, Editor. 2011, Formatex: Badajoz, Spain. p. 826-834.
[163] Denyer S, Stewart G. Mechanisms of action of disinfectants. International Biodeterioration & Biodegradation, 1998. 41(3-4):261-268.
[164] Cloete TE, Jacobs L, Brözel VS. The chemical control of biofouling in industrial water systems. Biodegradation, 1998. 9:23-27.
[165] MacDonald R, Santa M, Brözel V. The response of a bacterial biofilm community in a simulated industrial cooling water system to treatment with an anionic dispersant. Journal of Applied Microbiology 2000. 89(2):225-235.

44 Chapter 2
_________________________________________________________________________________
[166] Ferreira C, Pereira A, Pereira M, Melo L, Simões M. Physiological changes induced by the quaternary ammonium compound benzyldimethyldodecylammonium chloride on Pseudomonas fluorescens. Journal of Antimicrobial Chemotherapy, 2011. 66(5):1036-1043.
[167] Brözel VS, Cloete TE. Resistance of bacteria from cooling waters to bactericides. Journal of Industrial Microbiology Biotechnology, 1991. 8(4):273-276.
[168] Sykes RB, Bonner DP, Bush K, Georgopapadakou NH. Azthreonam (SQ 26,776), a synthetic monobactam specifically active against aerobic gram-negative bacteria. Antimicrobial Agents and Chemotherapy, 1982. 21(1):85-92.
[169] Neu HC, Aswapokee N, Aswapokee P, Fu KP. HR 756, a new cephalosporin active against gram-positive and gram-negative aerobic and anaerobic bacteria. Antimicrobial Agents and Chemotherapy 1979. 15(2):273-81.
[170] Dahl TA, Midden WR, Hartman PE. Comparison of killing of gram-negative and gram-positive bacteria by pure singlet oxygen. Journal of Bacteriology, 1989. 171(4):2188-94.
[171] Møretrø T, Vestby LK, Nesse LL, Storheim SE, Kotlarz K, Langsrud S. Evaluation of efficacy of disinfectants against Salmonella from the feed industry. Journal Applied Microbiology, 2009. 106(3):1005-12.
[172] Lequette Y, Boels G, Clarisse M, Faille C. Using enzymes to remove biofilms of bacterial isolates sampled in the food-industry. Biofouling, 2010. 26(4):421-431.
[173] Cortés M, Bonilla J, Sinisterra R, Biofilm formation, control and novel strategies for erradication, in Science against microbial pathogens: communicating current research and technological advances, M.-V. A, Editor. 2011, Formatex: Badajoz, Spain. p. 826-834.
[174] Buchovec I, Paskeviciute E, Luksiene Z. Photosensitization-based inactivation of food pathogen Listeria monocytogenes in vitro and on the surface of packaging material. Journal of Photochemistry and Photobiology B: Biology, 2010. 99(1):9-14.
[175] Agarwal A, Jern Ng W, Liu Y. Removal of biofilms by intermittent low-intensity ultrasonication triggered bursting of microbubbles. Biofouling, 2014;(ahead-of-print):1-7.
[176] Qian Z, Sagers RD, Pitt WG. The effect of ultrasonic frequency upon enhanced killing of P. aeruginosa biofilms. Annals of Biomedical Engineering, 1997. 25(1):69-76.
[177] Broekman S, Pohlmann O, Beardwood E, de Meulenaer EC. Ultrasonic treatment for microbiological control of water systems. Ultrasonics Sonochemistry, 2010. 17(6):1041-1048.
[178] Simões LC, Simões M. Biofilms in drinking water: problems and solutions. RSC Advances, 2013. 3(8):2520-2533.
[179] Bolton JR, Linden KG. Standardization of methods for fluence (UV dose) determination in bench-scale UV experiments. Journal of Environmental Engineering, 2003. 129(3):209-215.
[180] Pozos N, Scow K, Wuertz S, Darby J. UV disinfection in a model distribution system:: biofilm growth and microbial community. Water Research, 2004. 38(13):3083-3091.
[181] Långmark J, Storey MV, Ashbolt NJ, Stenström T-A. The effects of UV disinfection on distribution pipe biofilm growth and pathogen incidence within the greater Stockholm area, Sweden. Water Research, 2007. 41(15):3327-3336.
[182] Bukhari Z, Hargy TM, Bolton JR, Dussert B, Clancy JL. Medium-pressure UV for oocyst inactivation. Journal-American Water Works Association, 2000. 91(3):86-94.
[183] Racyte J, Sharabati J-A-D, Paulitsch-Fuchs AH, Yntema DR, Mayer MJ, Bruning H, Rijnaarts HH. Combining fluidized activated carbon with weak alternating electric fields for disinfection. Carbon, 2011. 49(15):5321-5328.

Introduction 45 _________________________________________________________________________________
[184] Racyte J, Bernard S, Paulitsch-Fuchs AH, Yntema DR, Bruning H, Rijnaarts HH. Alternating electric fields combined with activated carbon for disinfection of Gram negative and Gram positive bacteria in fluidized bed electrode system. Water Research, 2013. 47(16):6395-6405.
[185] Kalishwaralal K, BarathManiKanth S, Pandian SRK, Deepak V, Gurunathan S. Silver nanoparticles impede the biofilm formation by Pseudomonas aeruginosa and Staphylococcus epidermidis. Colloids and Surfaces B: Biointerfaces, 2010. 79(2):340-344.
[186] Rai M, Yadav A, Gade A. Silver nanoparticles as a new generation of antimicrobials. Biotechnology Advances, 2009. 27(1):76-83.
[187] Kim JS, Kuk E, Yu KN, Kim J-H, Park SJ, Lee HJ, Kim SH, Park YK, Park YH, Hwang C-Y. Antimicrobial effects of silver nanoparticles. Nanomedicine: Nanotechnology, Biology and Medicine, 2007. 3(1):95-101.
[188] Molobela IP, Cloete TE, Beukes M. Protease and amylase enzymes for biofilm removal and degradation of extracellular polymeric substances (EPS) produced by Pseudomonas fluorescens bacteria. African Journal of Microbiology Research, 2010. 4(14):1515-1524.
[189] Livermore DM. Fourteen years in resistance. International Journal of Antimicrobial Agents, 2012. 39(4):283-294.
[190] Grant D, Bott T. Biocide Dosing Strategies for Biofilm Control. Heat Transfer Engineering, 2005. 26(1):44-50.
[191] Lemos M, Borges A, Teodósio J, Araújo PA, Mergulhão F, Melo L, Simões M. The effects of ferulic and salicylic acids on Bacillus cereus and Pseudomonas fluorescens single- and dual-species biofilms. International Biodeterioration & Biodegradation, 2014. 86(Part A):42–51.
[192] Simões M, Bennett RN, Rosa EAS. Understanding antimicrobial activities of phytochemicals against multidrug resistant bacteria and biofilms. Natural Product Reports, 2009. 26(6):746-757.
[193] Johnston MD, Lambert RJW, Hanlon GW, Denyer SP. A rapid method for assessing the suitability of quenching agents for individual biocides as well as combinations. Journal of Applied Microbiology, 2002. 92(4):784-789.
[194] Verhoef R, Schols HA, Blanco A, Siika-aho M, Ratto M, Buchert J, Lenon G, Voragen AGJ. Sugar composition and FT-IR analysis of exopolysaccharides produced by microbial isolates from paper mill slime deposits. Biotechnology and Bioengineering, 2005. 91(1):91-105.
[195] Valeriano C, De Oliveira TLC, De Carvalho SM, Cardoso MdG, Alves E, Piccoli RH. The sanitizing action of essential oil-based solutions against Salmonella enterica serotype Enteritidis S64 biofilm formation on AISI 304 stainless steel. Food Control, 2012. 25(2):673-677.
[196] Coenye T, Brackman G, Rigole P, De Witte E, Honraet K, Rossel B, Nelis HJ. Eradication of Propionibacterium acnes biofilms by plant extracts and putative identification of icariin, resveratrol and salidroside as active compounds. Phytomedicine, 2012. 19(5):409-412.
[197] Ravichandiran V, Shanmugam K, Anupama K, Thomas S, Princy A. Structure-based virtual screening for plant-derived SdiA-selective ligands as potential antivirulent agents against uropathogenic Escherichia coli. European Journal of Medicinal Chemistry, 2012. 48:200-205.
[198] Borges A, Saavedra M, Simões M. The activity of ferulic and gallic acids in biofilm prevention and control of pathogenic bacteria. Biofouling, 2012. 28(7):755-67.
[199] de Paz LEC, Resin A, Howard KA, Sutherland DS, Wejse PL. Antimicrobial effect of chitosan nanoparticles on Streptococcus mutans biofilms. Applied and Environmental Microbiology, 2011. 77(11):3892-3895.
[200] Orgaz B, Lobete MM, Puga CH, San Jose C. Effectiveness of chitosan against mature biofilms formed by food related bacteria. International Journal of Molecular Sciences, 2011. 12(1):817-828.

46 Chapter 2
_________________________________________________________________________________
[201] Kim S, Lee S, Hong S, Oh Y, Kweon J, Kim T. Biofouling of reverse osmosis membranes: microbial quorum sensing and fouling propensity. Desalination, 2009. 247(1):303-315.
[202] Dong Y-H, Wang L-H, Zhang L-H. Quorum-quenching microbial infections: mechanisms and implications. Philosophical Transactions of the Royal Society B: biological Sciences, 2007. 362(1483):1201-1211.
[203] Vestby LK, Johannesen KCS, Witsø IL, Habimana O, Scheie AA, Urdahl AM, Benneche T, Langsrud S, Nesse LL. Synthetic brominated furanone F202 prevents biofilm formation by potentially human pathogenic Escherichia coli O103:H2 and Salmonella ser. Agona on abiotic surfaces. Journal of Applied Microbiology, 2014. 116(2):258-268.
[204] Boyle KE, Heilmann S, van Ditmarsch D, Xavier JB. Exploiting social evolution in biofilms. Current Opinion in Microbiology, 2013. 16(2):207-212.
[205] Xavier JB, Picioreanu C, Rani SA, van Loosdrecht MCM, Stewart PS. Biofilm-control strategies based on enzymatic disruption of the extracellular polymeric substance matrix - a modelling study. Microbiology-SGM, 2005. 151:3817-3832.
[206] Azeredo J, Sutherland IW. The use of phages for the removal of infectious biofilms. Current Pharmaceutical Biotechnology, 2008. 9(4):261-266.
[207] Tait K, Skillman L, Sutherland I. The efficacy of bacteriophage as a method of biofilm eradication. Biofouling, 2002. 18(4):305-311.
[208] Lu TK, Collins JJ. Dispersing biofilms with engineered enzymatic bacteriophage Proceedings of the National Academy of Sciences, 2007. 104(27):11197-11202.
[209] Flemming H, Ridgway H, Biofilm control: conventional and alternative approaches, in Springer Series on Biofilms. 2009, Springer Berlin Heidelberg. p. 103-117.
[210] Lambert RJW, Johnston MD, Hanlon GW, Denyer SP. Theory of antimicrobial combinations: biocide mixtures - synergy or addition? Journal of Applied Microbiology, 2003. 94(4):747-759.
[211] Bang SH, Jang A, Yoon J, Kim P, Kim JS, Kim Y-H, Min J. Evaluation of whole lysosomal enzymes directly immobilized on titanium (IV) oxide used in the development of antimicrobial agents. Enzyme and Microbial Technology, 2011. 49(3):260-265.
[212] Oulahal N, Martial-Gros A, Bonneau M, Blum L. Combined effect of chelating agents and ultrasound on biofilm removal from stainless steel surfaces. Application to'Escherichia coli milk' and'Staphylococcus aureus milk' biofilms. Biofilms, 2004. 1(1):65-73.

CHAPTER 3 DIFFUSION OF ANTIMICROBIAL AGENTS THROUGH BIOFILMS
This chapter is published as:
Araújo PA, Mergulhão F, Melo L, Simões M. 2014. The ability of an antimicrobial agent to penetrate a biofilm
is not correlated with its killing or removal efficiency. Biofouling 30 (6): 677-683. DOI:
10.1080/08927014.2014.904294

48 Chapter 3 _________________________________________________________________________________
ABSTRACT
The penetration ability of twelve antimicrobial agents was determined against biofilms
of B. cereus and P. fluorescens using a colony biofilm assay. These antimicrobial agents
included antibiotics and biocides. The surfactants benzalkonium chloride (BAC) and
cetyltrimethyl ammonium bromide (CTAB), and the antibiotics ciprofloxacin and
streptomycin raised interest due to their distinct activities. Erythromycin and CTAB were
retarded by the presence of biofilms, conversely to ciprofloxacin and BAC (no
retardation due to presence of biofilms). The removal and killing efficacies of these four
selected agents was additionally evaluated against biofilms formed in microtiter plates.
The most efficient biocide was CTAB, which enabled a higher killing of both bacterial
biofilms. Ciprofloxacin was the best antibiotic although none of the selected
antimicrobial agents promoted total biofilm removal and/or killing. Comparative analysis
of the results obtained with colony biofilms and microtiter plate biofilms show that
although extracellular polymeric substances and the biofilm structure are considered a
determining factor in biofilm resistance, the ability of an antimicrobial agent to
penetrate a biofilm is not correlated with its killing or removal efficiency. Also, the results
reinforce the role of an appropriate antimicrobial selection as a key step in the design of
disinfection processes for biofilm control.

Diffusion of antimicrobial agents through biofilms 49 _________________________________________________________________________________
3.1 INTRODUCTION
A biofilm is commonly defined as a microbial community with cells irreversibly attached
to a substratum or attached to each other, and embedded in a matrix of extracellular
polymeric substances (EPS) [1]. EPS protects bacteria from environmental adversities
[2]. In all industries, especially in the food industry, the proliferation of microorganisms
is very common even when manufacturers diligently follow all contingency plans [3]. The
main objective of microbial control is to eliminate or reduce the numbers of
microorganisms to acceptable levels, as well as to prevent and control the formation of
biological deposits attached to the process equipment surfaces [4]. Currently, there is
no control strategy capable of entirely eradicating biofilms [5]. At the same time, there
is a need to continuously find new strategies to manage antimicrobial resistance [6, 7].
Resistance is the ability that microorganisms have to withstand antimicrobial
treatments. Russell [8] and Chapman [9] documented three types of resistance: intrinsic
resistance, e.g. Gram negative lipopolysaccharide layer [10]; acquired resistance, e.g.
manipulated resistance mediated via plasmids; and adaptive resistance, e.g. exposure
to sub-lethal concentrations of an antimicrobial agent that selects for mutation,
conferring resistance to that agent or others of the same type (cross-resistance). The
way how microorganisms develop resistance is not well understood. Biofilm formation,
a case of adaptive resistance is considered a microbial survival strategy, enabling them
to be 10-1000 fold more resistant to antimicrobial agents than their free-floating
equivalents [11-13]. Antimicrobial resistance is multi-factorial and usually does not
depend only on one specific mechanism [14, 15]. When biofilms are exposed to
antimicrobial agents, they present specific survival strategies. In comparison with their
planktonic counterparts, biofilm cells are physiologically distinct by having specific
resistance genes that express protective factors such as multi-drug efflux pumps, stress
response regulons and different cell physiognomies [16]. Moreover, they often present
decreased respiration and growth/replication rates, despite having higher cell densities.
Embedded cells are capable to communicate through quorum sensing, and the
existence of persister cells enables them to survive [17]. Biofilm cells are protected by
the EPS they produce. The functions of EPS are enabling the biofilm to withstand shear
forces, dehydration and chemical attacks [18]. EPS enhances robustness and survival of
the biofilm microorganisms on a substratum by serving as a chemically reactive
diffusional transport barrier slowing down the penetration of antimicrobial agents.

50 Chapter 3 _________________________________________________________________________________
Furthermore, this matrix reinforces the biofilm attachment to the substratum and
promotes its mechanical stability [19, 20]. Moreover, it is where the convective and
diffusional transport to the biofilm of oxygen, nutrients, and other substances takes
place [21]. EPS composition and architecture has influence on how oxygen, nutrients
and cell excreted products are transported [22]. The biofilm constituents may act as an
adsorbent or reactant, thus chemically impairing diffusion, and its structure (porosity
and tortuosity) may physically reduce transport [23-28].
In order to plan a disinfection procedure it is important to select a suitable
antimicrobial agent with an appropriate effectiveness against the contaminants [5, 29].
The objective of this study was to understand the role of biofilms on the effectiveness
of antimicrobial agents, with a specific focus on the selection of suitable chemical
compounds capable of passing the EPS barrier, killing and removing the biofilm
embedded cells of B. cereus and P. fluorescens. These bacteria are ubiquitous in
industrial systems causing numerous process and end product quality problems [30, 31].
The production of extracellular enzymes by these bacteria results in food spoilage
[30, 32, 33]. Moreover, they can represent a significant proportion of the contaminant
biofilm microflora of dairy plants [34-37].
3.2 MATERIALS AND METHODS
MICROORGANISMS AND CULTURE CONDITIONS
The bacteria used in this work were P. fluorescens ATCC 13525T and a B. cereus strain
isolated from a disinfectant solution and identified by 16S rRNA gene sequencing [31].
Bacterial growth conditions were 30 ± 3 ºC and pH 7, with glucose as the main
carbon source. Culture medium consisted of 5 g L-1 glucose, 2.5 g L-1 peptone and 1.25 g
L-1 yeast extract, in phosphate buffer (pH 7, 25 mM) [38]. All the culture medium
products were purchased to Merck (VWR, Carnaxide, Portugal). Bacterial suspensions
were prepared by gently removing a small portion of bacteria from solid medium, and
diluting it in a 1 L flask (Duran, VWR, Carnaxide, Portugal) containing 250 mL of sterile
culture medium. This bacterial suspension was incubated overnight (16 h) at the given
temperature, with agitation (120 rpm). After the growth period, the suspension was
washed with phosphate buffer in two consecutive steps of centrifugation (3999 g,
10 min) in an Eppendorf centrifuge 5810R (Göttingen, Germany), and resuspended in

Diffusion of antimicrobial agents through biofilms 51 _________________________________________________________________________________
phosphate buffer (20 mM), in order to obtain a final bacterial concentration of
1 × 109 cells mL-1.
ANTIMICROBIALS
The twelve antimicrobials used throughout the experiments (Table 3.2) were
cetyltrimethyl ammonium bromide (CTAB), benzalkonium chloride (BAC), sodium
hypochlorite, ethanol, hydrogen peroxide, streptomycin, and tetracycline that were
obtained from Sigma-Aldrich (Sintra, Portugal). Benzyl dimethyl dodecyl ammonium
chloride (BDMDAC) was obtained from Merck (VWR, Carnaxide, Portugal). Ciprofloxacin
was acquired from Fluka (Sintra, Portugal). Chlorine dioxide was obtained from
TwinOxide® (Salmon & Cia. Lda, Lisbon, Portugal) and, isopropanol and erythromycin
were purchased from AppliChem (VWR, Carnaxide, Portugal). When possible, the
compounds were used as they are commonly sold (♥). Some amounts of antimicrobial
were previously optimized to obtain a detectable inhibition halo (♣), and others were
used at the reported minimum inhibitory concentrations (MIC) (♦) [39, 40].
COLONY BIOFILM FORMATION AND PENETRATION TESTS
Colony biofilms were developed according to the method of Anderl et al. [41] and Singh
et al. [42]. Biofilms were grown in sterile Mueller-Hinton agar plates (24 h, 30 ± 3 oC). A
volume of 40 µL of cell suspension of B. cereus or P. fluorescens was placed on a 13 mm
polycarbonate membrane, pore size 0.2 µm (Merk, Millipore, Carnaxide, Portugal)
originating colony biofilms. Afterwards, the membranes with biofilms were transferred
to a fresh plate containing the same growth medium, seeded with Staphylococcus
aureus CECT 976 at a McFarland standard of 0.5 [41, 42]. Another polycarbonate
membrane was placed on the top of the biofilm so that the sterile discs (Biochemica,
VWR, Carnaxide, Portugal) were not in direct contact with the biofilms (Figure 3.1). The
antimicrobial discs were impregnated with a 15 µL drop containing the different
antimicrobials used, providing the amount per disc described in Table 3.2. The negative
controls contained a 15 µL drop of sterile distilled water and the positive controls were
obtained in the absence of biofilm.

52 Chapter 3 _________________________________________________________________________________
Figure 3.1 Array of polycarbonate membranes and biofilms for the study of the diffusion of antimicrobial agents through biofilms (adapted from Anderl et al. [41] and Singh et al.[42]).
The plates were incubated for 24 h at 30 ± 3 oC before the assessment of the
inhibition halos. The positive controls were taken as 100% penetration and used to
calculate the penetration rates when biofilms were present.
BIOFILM FORMATION IN MICROTITER PLATES
Biofilms were developed according to the modified microtiter plate test proposed by
Stepanović et al. [43]. For each bacterium, at least 16 wells of a sterile 96-wells flat-
bottomed polystyrene tissue culture plate with a lid (Orange Scientific, Braine-l'Alleud,
Belgium) were filled with 200 µL of bacterial suspension at a density of 1 × 109 cells mL-
1. The negative controls were wells containing culture medium without bacterial cells.
The plates were incubated for 24 h at 30 ± 3 ºC without agitation.
BIOFILM CHARACTERIZATION
Biofilms of B. cereus and P. fluorescens grown were removed from the polycarbonate
membranes or from the microtiter plates using a stainless steel scraper and, afterwards
resuspended in 10 mL of buffer solution (2 mM Na3PO4, 2 mM NaH2PO4, 9 mM NaCl and
1 mM KCl, pH 7) and homogenized by vortexing (Heidolph, model Reax top, Schwabach,
Germany) for 30 s with 100% power input, according to the method described by [31].
The homogenized biofilm suspensions were then characterized in terms of cell density,
total and extracellular proteins and polysaccharides. Thickness was measured for the
colony biofilms using a digital micrometer (VS-30H, Mitsubishi Kasei Corporation,
Nagoya, Japan). Cell densities were assessed in terms of colony forming units (CFU) on
Antimicrobial agent
Biofilm
Mueller-Hinton media seeded with S. aureus
13 mm polycarbonate membranes
Sterile disc

Diffusion of antimicrobial agents through biofilms 53 _________________________________________________________________________________
Plate Count Agar (PCA) (Merck, VWR, Carnaxide, Portugal), according to Simões et al.
[44]. The biofilm suspensions were diluted to the adequate cellular concentration in
buffer solution. A volume of 30 µL of the diluted suspension was transferred onto PCA
plates. Colony enumeration was carried out after 48 h at 27 ºC.
To assess the total and extracellular proteins and polysaccharides, the method
described by Simões et al. [45] was used. Biofilm extracellular proteins and
polysaccharides were extracted using Dowex resin [46]. Dowex® resin Marathon® C
sodium form, 20-50 mesh (Sigma, Sintra, Portugal) was added to the biofilm
suspensions. The extraction took place at 400 rpm and 4 oC for 4 h. The extracellular
components (present in the supernatant) were separated from the cells via
centrifugation (3777g, 5 min). The total (before extraction) and extracellular biofilm
proteins were determined using the Lowry et al. modified kit (Sigma, Sintra, Portugal),
with bovine serum albumin as standard. The procedure is essentially the Lowry method
[47] as modified by Peterson [48]. The total and extracellular polysaccharides were
quantified through the phenol-sulphuric acid method of Dubois et al. [49], using glucose
as standard.
BIOFILM CONTROL IN MICROTITER PLATES
To ascertain the adequacy of antimicrobial penetration results to develop biofilm control
strategies, 24 h aged biofilms formed in 96-well microtiter plates were exposed to
selected antimicrobial agents. Biofilms were exposed for 1 h at 30 ± 3 ºC, without
agitation, similarly to the colony biofilms. After antimicrobial exposure, the biofilms were
analysed in terms of biomass and viability and the results are presented as percentage
of biofilm reduction and killing.
BIOMASS AND VIABILITY QUANTIFICATION
The biomass was quantified using crystal violet (Merck VWR, Carnaxide, Portugal)
staining, according to Simões et al. [50]. The bacterial biofilms in the 96-wells plates were
fixed with 250 µL of 98% methanol (Vaz Pereira, Porto, Portugal) per well for 15 min.
Afterwards, the plates were emptied and left to dry. Then, the fixed bacteria were
stained for 5 min with 200 µL of crystal violet per well. Excess stain was rinsed off by
placing the plate under running tap water. After the plates were air dried, the dye bound

54 Chapter 3 _________________________________________________________________________________
to the adherent cells was resolubilized with 200 µL of 33% (v/v) glacial acetic acid (Merck,
VWR, Carnaxide, Portugal) per well. The absorbance was measured at 570 nm using a
microplate reader (Spectramax M2e, Molecular Devices, Inc., Sunnyvale, USA). All tests
were performed in three independent experiments with triplicates.
Biofilm removal was given by equation 1:
100%
C
WC
OD
ODODBR (eq. 3.1)
where %BR is the percentage of biofilm removal and OD is the optical density, ODC is the
OD570nm value for biofilms not exposed to antimicrobial agents and ODW is the OD570nm
value for biofilm exposed to the selected chemicals.
The modified alamar blue (Sigma-Aldrich, Sintra, Portugal) microtiter plate assay
was applied to determine the bacterial viability of the cells as reported by Borges et al.
[51]. For the staining procedure, fresh culture medium (190 µL) was added to the plates.
To each well 10 µL of alamar blue (400 mM) indicator solution were added. Plates were
incubated for 20 min in darkness at room temperature. Fluorescence was measured at
the wave lengths λexcitation = 570 nm and λemission = 590 nm with the same microplate
reader. The percentage of biofilm killing was given by equation 2:
100%
C
WC
FI
FIFIBI (eq. 3.2)
where %BI is the percentage of biofilm killing, FIC is the fluorescence intensity of biofilms
not exposed to antimicrobial agents and FIW is the fluorescence intensity value for
biofilms exposed to the selected chemicals.
STATISTICAL ANALYSIS
For each parameter tested, the average and the standard deviation were calculated. The
statistical significance of the results was evaluated using the t-test (confidence level of
95%) with the statistical program IBM SPSS Statistics software (Armonk, NY, USA),
version 20.0, to determine whether the differences between the controls and the
antimicrobial tests could be considered significant.

Diffusion of antimicrobial agents through biofilms 55 _________________________________________________________________________________
3.3 RESULTS AND DISCUSSION
In order to ascertain possible factors involved in biofilm resistance/susceptibility to the
selected antimicrobials, B. cereus and P. fluorescens biofilms were characterized in terms
of their biovolume, CFU, total and matrix proteins and polysaccharides (Table 3.1).
Table 3.1 Characterization of B. cereus and P. fluorescens grown as colonies and as microtiter plate biofilms.
B. cereus P. fluorescens
Colony Microtiter
plate Colony Microtiter
plate Biovolume /(cm3)
0.019 ± 0.002
0.018 ± 0.001
Log CFU/cm2 7.41 ± 0.52 7.20 ± 0.69 8.11 ± 0.13 8.89 ± 0.34
Matrix proteins/ (µg/cm2)
13.8 ± 1.5 16.7 ± 0.15 20.7 ± 0.78 15.2 ± 0.37
Total proteins/ (µg/cm2)
49.6 ± 2.3 26.3 ± 0.04 32.2 ± 2.7 27.1 ± 0.02
Matrix polysaccharides/ (µg/cm2)
20.4 ± 2.6 20.4 ± 0.09 7.17 ± 0.56 17.4 ± 3.0
Total polysaccharides/ (µg/cm2)
29.8 ± 3.1 26.2 ± 0.06 11.3 ± 0.19 23.6 ± 2.0
Colony biofilms of B. cereus covered approximately 5.5 ± 0.69 mm of the
membrane whilst those of P. fluorescens covered 6.3 ± 0.44 mm. B. cereus biofilms were
thicker than those of P. fluorescens (P < 0.05). The cell density of P. fluorescens biofilms
(8.11 ± 0.13 CFU cm-2) was significantly higher than for B. cereus (7.20 ± 0.69 CFU cm-2)
(P < 0.05). B. cereus biofilms had higher amounts of extracellular polysaccharides and
lower extracellular proteins content in comparison to those found in the P. fluorescens
biofilm matrix (P < 0.05). The resulting biofilms presented larger diameters, similar
thickness values, and lower cell numbers than those used in the studies of Singh et al.
[42] with S. epidermis. However, different growth conditions were used, particularly the
growth period. Singh et al. [42] used 48 h old biofilms while the biofilms used in this
study were 24 h old.

56 Chapter 3 _________________________________________________________________________________
DIFFUSION OF ANTIMICROBIAL AGENTS THROUGH BIOFILMS
When antimicrobial agents were applied to the biofilms, inhibition halos were produced
in the S. aureus culture underneath. The size of the halos was indicative of the ability of
antimicrobial agents to penetrate the biofilms. The same characteristic is related to the
antimicrobial potency of each antimicrobial agent against the S. aureus culture, i.e. a
larger inhibition halo was indicative of a more powerful antimicrobial agent, in terms of
penetration (Table 3.3).
In the diffusion test apparatus, ciprofloxacin and tetracycline were the
antimicrobial agents that produced the largest halos (about 22 mm) after passing
through the biofilms of B. cereus and P. fluorescens. This behavior was closely followed
by BAC and BDMDAC (19 mm halos) for both types of biofilms. Erythromycin and ethanol
were able to penetrate both biofilms (halos of about 13 mm were obtained).
Isopropanol, sodium hypochlorite, chlorine dioxide and streptomycin produced
inhibition halos of 5 mm. Hydrogen peroxide and CTAB caused insignificant inhibition
halos (P > 0.05). In terms of antimicrobial retardation, values comprised between 5%
and 20% were observed for ethanol, BDMDAC and tetracycline for both biofilms, and
erythromycin for B. cereus biofilms. Erythromycin was retarded approximately 30% by
P. fluorescens biofilms. The same percentage was only obtained with B. cereus biofilms
treated with chlorine dioxide. P. fluorescens biofilms retarded streptomycin diffusion by
40% and isopropanol and chlorine dioxide by 50%. Isopropanol was retarded more than
70% by B. cereus biofilms. Total antimicrobial retardation (100%) was achieved with
hydrogen peroxide and CTAB by both biofilms (for CTAB see Figure 3.2), and
streptomycin by B. cereus biofilms. The statistical analyses showed that the retardation
of hydrogen peroxide, BDMDAC, CTAB, streptomycin and tetracycline was significant for
both biofilms (P < 0.05). B. cereus biofilms with isopropanol and erythromycin, and
P. fluorescens biofilms with ethanol and chlorine dioxide also had significant effects on
chemical retardation (P < 0.05). These results show that the presence of a biofilm
markedly affected the diffusion of some antimicrobial agents. Biofilms have intrinsic
resistance to antimicrobial agents. Amongst those resistance mechanisms, mass transfer
limitations through biofilms is of utmost importance [52]. For the effective inactivation
of bacteria in the deeper layers of the biofilms it is essential that the antimicrobial agent
diffuses through the biofilm. In some cases, when biofilms are thick, cells can be in a
dormant/low metabolic active state in the deeper layer. Those cells can show a

Diffusion of antimicrobial agents through biofilms 57 _________________________________________________________________________________
remarkable resistance to antimicrobials [16, 21]. Moreover, EPS protects the cells
against an antimicrobial attack by hindering diffusion through the biofilms. The biofilm
matrix is known to have the ability to bind to antimicrobial agents [53]. Anderl et al. [41]
suggested that the diffusion of antimicrobial agents might be delayed because the
biofilm has the ability to chemically react with them, resulting in their inactivation. Thus,
less antimicrobial molecules are left to interact with the deeper layers of the biofilms.
Table 3.2 Antimicrobial agents and respective mass used for the biofilm colony tested. Inhibition halos (mm) of S. aureus due to antimicrobials in the presence of B. cereus and P. fluorescens biofilms. Percentage retardation caused by the presence of B. cereus and P. fluorescens biofilms. The average ± SD is presented
Antimicrobials
B. cereus P. fluorescens
Mass/ (µg)
Inhibition halos/ (mm)
Retardation/ (%)
Inhibition halos/
(mm) Retardation/
(%)
Alcohols
Ethanol♥ 8242 12 ± 1.3 12 ± 7.0 13 ± 0.96 9.3 ± 2.5
Isopropanol♥ 11700 2.3 ± 0.47 70 ± 8.8 4.3 ± 2.1 52 ± 22
Oxidising
Sodium
hypochlorite♣ 543 (Cl) 4.6 ± 0.50 5.0 ± 0.45 4.7 ± 0.10 1.9 ± 3.1
Chlorine
dioxide♥ 74 (Cl) 5.3 ± 0.84 32 ± 12 3.0 ± 0.71 47 ± 13
Hydrogen
peroxide♥ 500 0.0 ± 0.0 100 ± 0.0 0.0 ± 0.0 100 ± 0.0
Surfactants
BAC♣ 350 18 ± 1.3 0.11 ± 0.07 18 ± 0.25 0.0 ± 0.0
BDMDAC♣ 350 19 ± 0.25 15 ± 1.1 19 ± 0.10 13 ± 0.54
CTAB♣ 350 0.0 ± 0.0 100 ± 0.0 0.0 ± 0.0 99 ± 1.6
Antibiotics
Ciprofloxacin♦ 5 20 ± 1.0 0.0 ± 0.0 24 ± 0.50 0.0 ± 0.0
Erythromycin♦ 15 14 ± 0.05 14 ± 0.32 12 ± 0.50 28 ± 3.1
Streptomycin♦ 10 0.0 ± 0.0 100 ± 0.0 3.4 ± 0.05 40 ± 0.89
Tetracycline♦ 30 22 ± 0.94 6.9 ± 3.9 24 ± 0.47 12 ± 1.8
♥ Commonly available/standard concentration; ♣ optimized concentration; ♦ MIC; Cl means
chlorine

58 Chapter 3 _________________________________________________________________________________
Figure 3.2 Inhibition halos on S. aureus using three antimicrobial agents. Condition 1 corresponds to the control where no biofilm is present and condition 2 represents the tests with biofilms. Both condition are duplicated in the same plate. The conditions tested were: (a) BAC test in the presence of a P. fluorescens biofilm, showing that this compound is not retarded; (b) ciprofloxacin in the presence of a B. cereus biofilm, showing that this compound is not retarded, also that the inhibition halos are large taking into consideration the small amount used (5 µg); and (c) CTAB in the presence of a B. cereus biofilm, showing that there is antimicrobial activity in 1, however, the compound was totally retarded by the presence of the biofilm (no halos were observed).
Christensen et al. [54] reported that the presence of alginate, a common EPS,
caused mass transport limitations. Singh et al. [42] refers that the biofilm phenotype
provides antimicrobial resistance. These authors indicated the existence of spatial
heterogeneity in the biofilm structure as a possible explanation for the poor diffusion of
antimicrobial agents into biofilms. Diffusion in biofilms may be affected by charge
interactions between the matrix and the antimicrobial agents, by increasing the distance
between the antimicrobial and the bacteria, by size exclusion, and by the viscosity of the
matrix [55]. It has also been suggested that it is not the quantity of matrix that exclusively
causes resistance, but its polyanionic nature that hinders the antimicrobial agents [55].
For instance, the polysaccharides can hinder antimicrobial action due to their charge
and hydrophobic properties [21, 56]. In fact, the penetration of positively charged
hydrophilic drugs is known to be delayed by the EPS matrix [56].
In this study, retardation percentages often differed between the types of biofilm
(Table 3.2). Isopropanol, sodium hypochlorite and streptomycin diffused differently
through the biofilms of both species. The highest retardation rates, over 70%, occurred
for B. cereus biofilms. In fact, the distinct retardation rates are probably due to the
distinct biofilm characteristics, particularly the type of EPS produced by each bacterium
[57]. In addition, the amount of polysaccharides and proteins produced by both types of
1
1
2
2
1
1
2
2
1
1
2
2
a b c

Diffusion of antimicrobial agents through biofilms 59 _________________________________________________________________________________
bacteria is different. The high retardation rates observed for B. cereus biofilms might be
related to the high proteins content present (Table 3.2). As many antimicrobial agents
target protein-like structures [10], these might be adsorbed before penetrating the
biofilm.
The function of antimicrobial agents is to extinguish or to discontinue the growth
of an organism by biological or chemical processes [3]. The mode of action of
antimicrobial agents may be another important factor contributing to mass transfer
limitations through biofilms. Ethanol and isopropanol are membrane disruptors. These
chemicals act by penetrating into the cells through the hydrocarbon part of the
phospholipid bilayer, causing rapid release of intracellular components [15]. Even
though a higher mass of isopropanol than ethanol was used, isopropanol retardation
was higher, because it is slightly more reactive than ethanol against bacteria [10].
Chlorine based agents are the most broadly used disinfectants [10]. These chemicals are
highly active oxidizing agents destroying the cellular activity of proteins. Sodium
hypochlorite was slightly hindered (less than 5%) by the presence of a biofilm. In fact,
oxidizing agents react strongly with cell constituents such as amino, carboxyl, sulfhydryl
and hydroxyl groups in bacterial proteins as well as nucleic acids [10]. Hydrogen peroxide
damages ribosomes which are responsible for the translation of RNA into a peptide
chain, being also able to react with other cell constituents [15]. This compound has
oxidative potential, producing hydroxyl free radicals that target lipids, proteins and DNA
[10]. Peroxides are more active against Gram-positive bacteria than Gram-negative
bacteria [10]. However, the ability of both bacteria to produce catalase or other
peroxidases may increase tolerance to this compound [58, 59]. Quaternary ammonium
compounds (QACs) are classified according to the ionic physiognomies of their
hydrophilic group as anionic, cationic, non-ionic and zwitterionic [60]. The mechanism
of action of cationic surfactants (BAC, BDMDAC and CTAB) is the same as the general
mechanism of QACs. The hydrophilic headgroup of QACs is adsorbed to the cell wall and
reacts with the cytoplasmic membrane, allowing the release of intracellular constituents
[61-63]. The strong affinity of CTAB for proteins and lipid components of the membrane
suggests that this QAC is spent before it reaches the under-layers of the biofilm [64].
Cationic surfactants are also known to bind to DNA and DNA-protein mixtures [65].
Fluoroquinolones such as ciprofloxacin are generally not hindered by the EPS of the
matrix [14, 55]. This was also found in the present study. The penetration of
aminoglycosides (streptomycin) is known to be delayed by P. aeruginosa biofilms [15].

60 Chapter 3 _________________________________________________________________________________
Streptomycin was, in this case, 40% retarded by P. fluorescens biofilms and 100% when
applied to B. cereus biofilms. This antibiotic acts by binding to prokaryotic ribosomes and
has shown affinity for other nucleic acid targets [66]. Erythromycin, a macrolide, has the
same mechanism of action as streptomycin. The antimicrobial mode of action of
tetracycline is by binding to ribosomes [67]. In this study, lower retardation rates were
expected, since a higher mass of this compound was used (Table 3.2). However, it seems
that this antibiotic is strongly affected by the biofilms.
BIOFILM ACTIVITY SCREENING
The presence of inhibition halos on the S. aureus culture is indicative of the penetration
efficacy of the antimicrobial agents through the biofilms. In fact, this assay does not
allow the distinction between biofilm penetration and antimicrobial potency. Taking into
account the results obtained with the antimicrobial retardation tests, selected
antimicrobials agents were used in order to ascertain the reliability of the results
obtained with the colony biofilm system. Therefore, biofilms of B. cereus and
P. fluorescens were formed in microtiter plates. The effects of BAC, CTAB, ciprofloxacin
and streptomycin were assessed on biofilm removal and killing. These antibiotics and
biocides were those with the highest and lowest retardation values. The tests were
performed using 96-well microtiter plates. This bioreactor permits the assessment of the
biofilm killing and removal rates by the selected antimicrobial agents using a large
number of replicates [68, 69]. The biofilms developed in the 96-well microtiter plates
were characterized in terms of their cell density, total and matrix polysaccharides and
protein content (Table 3.2). The cell densities of B. cereus and P. fluorescens biofilms
formed in the microtiter plates were similar to those of colony biofilms (P > 0.05). Colony
biofilms had a higher amount of total proteins in comparison to those formed in
microtiter plates, for both biofilms (P < 0.05). B. cereus biofilms had similar
polysaccharide content in either biofilms formed as colony and microtiter plates
(P > 0.05), while the polysaccharides in P. fluorescens biofilms were lower in the colony
system (P > 0.05)
Table 3.3 depicts the killing and removal percentages with the selected
antimicrobial agents for the biofilms formed in the microtiter plates. For B. cereus
biofilms, removal was similar with ciprofloxacin and streptomycin (12-14%). Their killing
efficiency was statistically similar (P > 0.05), even if streptomycin was the most efficient

Diffusion of antimicrobial agents through biofilms 61 _________________________________________________________________________________
antibiotic (40 vs 36%). The killing percentages of B. cereus biofilms caused by both QACs
were approximately 50% and its removal was also similar (around 15%). P. fluorescens
biofilms were equally removed (17-23%) and killed (about 15%) by both antibiotics. The
same biofilm was easier to be killed by CTAB (26%) rather than by BAC (15%). The
removal of P. fluorescens was similar to what was verified for B. cereus biofilms (about
15%). The removal and killing was significantly different between antibiotics and
biocides (P < 0.05), which suggests that QACs are more efficient in biofilm killing than
antibiotics. In general, biocides are known to perform better in the killing of biofilms,
apparently due to their multi-target mode of action [70]. Between B. cereus and
P. fluorescens biofilms, the removal was statistically similar in all cases (P > 0.05).
B. cereus killing was higher for both QACs and antibiotics when compared to
P. fluorescens (P < 0.05). This bacterium, as a Gram negative, is known to have higher
tolerance to biocides [71], which is commonly explained by hindrances in penetration
due to the presence of the outer membrane [15]. Between BAC and CTAB the killing
percentages were not significant (P > 0.05).
Table 3.3 Percentage killing and removal of B. cereus and P. fluorescens biofilms. The average ± SD is presented.
B cereus P. fluorescens
Killing / (%) Removal / (%) Killing / (%) Removal / (%)
BAC 46.6 ± 13 15.3 ± 2.7 15.5 ± 7.5 13.8 ± 5.4
CTAB 51.8 ± 10 15.8 ± 1.3 26.5 ± 6.9 16.0 ± 2.4
Ciprofloxacin 36.2 ± 8.3 11.8 ± 3.7 13.5 ± 2.7 22.7 ± 8.5 Streptomycin 40.0 ± 5.9 14.3 ± 3.5 15.3 ± 7.0 16.6 ± 1.3
The resistance of a biofilm is a very complex phenomenon. EPS plays an important
role on antimicrobial interaction and mass transfer limitations; albeit, other phenomena
can contribute to biofilm resistance [17, 72-74]. An antimicrobial agent that efficiently
penetrates a biofilm does not necessarily kill the embedded cells. This means that the
high penetration ability of some antimicrobial agents is not directly related with their
efficiency, as proposed by the comparison between the results obtained with colony
biofilms and those formed in the microtiter plates.

62 Chapter 3 _________________________________________________________________________________
3.4 CONCLUSIONS
This study uses two simple biofilm formation systems (biofilm colonies and microtiter
plates) to provide insights into the role played by a biofilm in the interaction with
antimicrobial agents. The systems used formed biofilms with similar characteristics in
terms of CFU, proteins and polysaccharides. The overall results demonstrate that the
selection of a suitable antimicrobial agent, able to penetrate a biofilm and kill the
bacteria, is of utmost importance when developing disinfection plans. At the same time,
a diffusion test by itself does not provide enough information on the biofilm control
efficiency of an antimicrobial agent. This reinforces the fact that antimicrobial resistance
in biofilms is a multifactorial problem and transport limitations, although part of the
problem, should not be implicated alone. Moreover, the assessment of biofilm killing
and removal is important for the selection of an appropriate control strategy. Biofilm
killing and removal are distinct phenomena.
REFERENCES
[1] Donlan R, Costerton J. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clinical Microbiology Reviews, 2002. 15(2):167-193.
[2] Jefferson K. What drives bacteria to produce a biofilm? Fems Microbiology Letters, 2004. 236(2):163-173.
[3] Araújo PA, Lemos M, Mergulhão F, Melo L, Simões M, Antimicrobial resistance in biofilms to disinfectants, in Science against microbial pathogens: communicating current research and technological advances, A. Méndez-Vilas, Editor. 2011, Formatex: Badajoz, Spain. p. 826-834.
[4] Maukonen J, Mättö J, Wirtanen G, Raaska L, Mattila-Sandholm T, Saarela M. Methodologies for the characterization of microbes in industrial environments: a review. Journal of Industrial Microbiology & Biotechnology, 2003. 30(6):327-356.
[5] Simões M, Simões L, Vieira M. A review of current and emergent biofilm control strategies. LWT - Food Science and Technology, 2010. 43(4):573-583.
[6] Traba C, Chen L, Liang D, Azzam R, Liang JF. Insights into discharge argon-mediated biofilm inactivation. Biofouling, 2013. 29(10):1205-1213.
[7] Lee J-H, Cho HS, Joo SW, Regmi SC, Kim J-A, Ryu C-M, Ryu SY, Cho MH, Lee J. Diverse plant extracts and trans-resveratrol inhibit biofilm formation and swarming of Escherichia coli O157:H7. Biofouling, 2013. 29(10):1189-1203.
[8] Russell A. Bacterial resistance to disinfectants: present knowledge and future problems. Journal of Hospital Infection, 1999. 43(SUPPL. 1).
[9] Chapman J. Biocide resistance mechanisms. International Biodeterioration & Biodegradation, 2003. 51(2):133-138.
[10] McDonnell G, Russell A. Antiseptics and disinfectants: activity, action, and resistance. Clinical Microbiology Reviews, 1999. 12(1):147-179.

Diffusion of antimicrobial agents through biofilms 63 _________________________________________________________________________________
[11] Mah T, O'Toole G. Mechanisms of biofilm resistance to antimicrobial agents. Trends in Microbiology, 2001. 9(1):34-39.
[12] Nikolaev Y. Extracellular factors of bacterial adaptation to unfavorable environmental conditions. Applied Biochemistry and Microbiology, 2004. 40(4):327-336.
[13] Araújo PA, Kruithof JC, van Loosdrecht MCM, Vrouwenvelder JS. The potential of standard and modified feed spacers for biofouling control. Journal of Membrane Science, 2012. 403–404(0):58-70.
[14] Drenkard E. Antimicrobial resistance of Pseudomonas aeruginosa biofilms. Microbes and Infection, 2003. 5(13):1213-1219.
[15] Maillard J. Bacterial target sites for biocide action. Journal of Applied Microbiology, 2002. 92:16s-27s.
[16] Davies D. Understanding biofilm resistance to antibacterial agents. Nature Reviews Drug Discovery 2003. 2:114-122.
[17] Simões LC, Lemos M, Pereira AM, Abreu AC, Saavedra MJ, Simões M. Persister cells in a biofilm treated with a biocide. Biofouling, 2011. 27(4):403-411.
[18] Flemming H. The EPS matrix: The "house of biofilm cells". Journal of Bacteriology, 2007. 189(22):7945-7947.
[19] Bryers J. Biofilms and the technological implications of microbial cell-adhesion. Colloids and Surfaces B-Biointerfaces, 1994. 2(1-3):9-23.
[20] Bryers J, Mixed Population Biofilms, in Biofilms — Science and Technology, L.F. Melo, et al., Editors. 1992, Springer Netherlands. p. 277-289.
[21] Stewart P. Diffusion in biofilms. Journal of Bacteriology, 2003. 185(5):1485-1491.
[22] Andersson S, Dalhammar G, Land C, Kuttuva Rajarao G. Characterization of extracellular polymeric substances from denitrifying organism Comamonas denitrificans. Applied Microbiology and Biotechnology, 2009. 82(3):535-543.
[23] Russell A, Furr J, Maillard J. Microbial susceptibility and resistance to biocides. American society for microbiology news, 1997. 63(9):481-487.
[24] Costerton J. Introduction to biofilm. International Journal of Antimicrobial Agents, 1999. 11(3-4):217-221.
[25] Melo L. Biofilm physical structure, internal diffusivity and tortuosity. Water science and technology, 2005. 52(7):77-84.
[26] Flemming H, Wingender J. The biofilm matrix. Nature Reviews in Microbiology, 2010. 8(9):623-633.
[27] Stewart P, Grab L, Diemer J. Analysis of biocide transport limitation in an artificial biofilm system. Journal of Applied Microbiology, 1998. 85(3):495-500.
[28] Stewart PS. Mini-review: Convection around biofilms. Biofouling: The Journal of Bioadhesion and Biofilm Research, 2012. 28(2):187-198.
[29] Araújo PA, Lemos M, Mergulhão F, Melo L, Simões M. The influence of interfering substances on the antimicrobial activity of selected quaternary ammonium compounds. International Journal of Food Science, 2013:9.
[30] Lindsay D, Brozel VS, Mostert JF, von Holy A. Differential efficacy of a chlorine dioxide-containing sanitizer against single species and binary biofilms of a dairy-associated Bacillus cereus and a Pseudomonas fluorescens isolate. Journal of Applied Microbiology, 2002. 92(2):352-361.
[31] Simões M, Simões L, Pereira M, Vieira M. Antagonism between Bacillus cereus and Pseudomonas fluorescens in planktonic systems and in biofilms. Biofouling, 2008. 24(5):339-349.

64 Chapter 3 _________________________________________________________________________________
[32] Teh KH, Flint S, Palmer J, Andrewes P, Bremer P, Lindsay D. Proteolysis produced within biofilms of bacterial isolates from raw milk tankers. International Journal of Food Microbiology, 2012. 157(1):28-34.
[33] Teh KH, Flint S, Palmer J, Lindsay D, Andrewes P, Bremer P. Thermo-resistant enzyme-producing bacteria isolated from the internal surfaces of raw milk tankers. International Dairy Journal, 2011. 21(10):742-747.
[34] Kreske AC, Ryu JH, Pettigrew CA, Beuchat LR. Lethality of chlorine, chlorine dioxide, and a commercial produce sanitizer to Bacillus cereus and Pseudomonas in a liquid detergent, on stainless steel, and in biofilm. J Food Prot, 2006. 69(11):2621-34.
[35] Dogan B, Boor KJ. Genetic diversity and spoilage potentials among Pseudomonas spp. isolated from fluid milk products and dairy processing plants. Applied and environmental microbiology, 2003. 69(1):130-138.
[36] Sharma M, Anand SK. Characterization of constitutive microflora of biofilms in dairy processing lines. Food Microbiology, 2002. 19(6):627-636.
[37] Wijman JG dLP, Moezelaar R, Zwietering MH, Abee T. Air–liquid interface biofilms of Bacillus cereus: formation, sporulation, and dispersion. Appl Environ Microbiol, 2007. 73:1481–1488.
[38] Simões M, Cleto S, Pereira M, Vieira M. Influence of biofilm composition on the resistance to detachment. Water science and technology, 2007. 55:473-480.
[39] Spangler S, Visalli M, Jacobs M, Appelbaum P. Susceptibilities of non-Pseudomonas aeruginosa gram-negative nonfermentative rods to ciprofloxacin, ofloxacin, levofloxacin, D-ofloxacin, sparfloxacin, ceftazidime, piperacillin, piperacillin-tazobactam, trimethoprim-sulfamethoxazole, and imipenem. Antimicrobial Agents and Chemotherapy, 1996. 40(3):772-775.
[40] Turnbull P, Sirianni N, LeBron C, Samaan M, Sutton F, Reyes A, Peruski L. MICs of selected antibiotics for Bacillus anthracis, Bacillus cereus, Bacillus thuringiensis, and Bacillus mycoides from a range of clinical and environmental sources as determined by the Etest. Journal of Clinical Microbiology, 2004. 42(8):3626-3634.
[41] Anderl J, Franklin M, Stewart P. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Journal of Antimicrobial Chemotherapy, 2000. 44(7):1818-1824.
[42] Singh R, Ray P, Das A, Sharma M. Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. Journal of Antimicrobial Chemotherapy, 2010. 65:1955-1958.
[43] Stepanović S VD, Dakić I, Savić B, Švabić-Vlahović M A modified microtiter-plate test for quantification of Staphylococcal biofilm formation. Journal of Microbiological Methods, 2000. 40(2):175-179.
[44] Simões M, Pereira M, Vieira M. Validation of respirometry as a short-term method to assess the efficacy of biocides. Biofouling, 2005. 21(1):9-17.
[45] Simões M, Pereira M, Sillankorva S, Azeredo J, Vieira M. The effect of hydrodynamic conditions on the phenotype of Pseudomonas fluorescens biofilms. Biofouling, 2007. 23(3-4):249-58.
[46] Frølund B, Palmgren R, Keiding K, Nielsen P. Extraction of extracellular polymers from activated sludge using a cation exchange resin. Water Research, 1996. 30(8):1749-1758.
[47] Lowry O, Rosebrough N, Farr A, Randall R. Protein measurement with the Folin phenol reagent. Journal of biological chemistry, 1951. 193(1):265-275.
[48] Peterson G. Review of the Folin phenol protein quantitation method of Lowry, Rosebrough, Farr and Randall. Analytical Biochemistry 1979. 100(2):201-220.
[49] Dubois M, Gilles K, Hamilton J, Rebers P, Smith F. Colorimetric method for determination of sugars and related substances. Anal. Chem., 1956. 28(3):350-356.

Diffusion of antimicrobial agents through biofilms 65 _________________________________________________________________________________
[50] Simões LC, Simões M, Vieira MJ. Biofilm interactions between distinct bacterial genera isolated from drinking water. Applied Environmental Microbiology, 2007. 73(19):6192-6200.
[51] Borges A, Saavedra M, Simões M. The activity of ferulic and gallic acids in biofilm prevention and control of pathogenic bacteria. Biofouling, 2012. 28(7):755-67.
[52] Flemming H, Ridgway H, Biofilm control: conventional and alternative approaches, in Springer Series on Biofilms. 2009, Springer Berlin Heidelberg. p. 103-117.
[53] Nikolaev Y, Plakunov V. Biofilm - “City of microbes” or an analogue of multicellular organisms? Microbiology, 2007. 76(2):125-138.
[54] Christensen B, Ertesvag H, Beyenal H, Lewandowski Z. Resistance of biofilms containing alginate-producing bacteria to disintegration by an alginate degrading enzyme (AlgL). Biofouling, 2001. 17(3):203-210.
[55] Cloete T. Resistance mechanisms of bacteria to antimicrobial compounds. International Biodeterioration & Biodegradation, 2003. 51(4):277-282.
[56] Smirnova T, Didenko L, Azizbekyan R, Romanova Y. Structural and functional characteristics of bacterial biofilms. Microbiology, 2010. 79(4):413-423.
[57] Sutherland I. The biofilm matrix - an immobilized but dynamic microbial environment. Trends in Microbiology, 2001. 9(5):222-227.
[58] Anderson D. Infections, Infertility and Assisted Reproduction Journal of Andrology, 2006. 27(1):2-2.
[59] Hall JF. Methods in biotechnology in Biochemical Education, H.-P. Schmauder., Editor. 1998, Taylor and Francis: London. p. 257.
[60] Atay N, Yenigün O, Asutay M. Sorption of anionic surfactants SDS, AOT and cationic surfactant hyamine 1622 on natural soils. Water air and soil pollution, 2002. 136(1):55-68.
[61] Ferreira C, Pereira A, Pereira M, Melo L, Simões M. Physiological changes induced by the quaternary ammonium compound benzyldimethyldodecylammonium chloride on Pseudomonas fluorescens. Journal of Antimicrobial Chemotherapy, 2011. 66(5):1036-1043.
[62] Salton M. Lytic agents, cell permeability, and monolayer penetrability. Journal of General Physiology, 1968. 52(1):227-252.
[63] Trueba A, Otero FM, González JA, Vega LM, García S. Study of the activity of quaternary ammonium compounds in the mitigation of biofouling in heat exchangers–condensers cooled by seawater. Biofouling, 2013. 29(9):1139-1151.
[64] Simões M, Pereira M, Machado I, Simões L, Vieira M. Comparative antibacterial potential of selected aldehyde-based biocides and surfactants against planktonic Pseudomonas fluorescens. Journal of Industrial Microbiology & Biotechnology, 2006. 33(9):741-749.
[65] Gani S, Chattoraj D, Mukherjee D. Binding of cationic surfactants to DNA, protein and DNA-protein mixtures. Indian Journal of Biochemistry & Biophysics, 1999. 36(3):165-176.
[66] Willis B, Arya D, An expanding view of aminoglycoside–nucleic acid recognition, in Advances in Carbohydrate Chemistry and Biochemistry, H. Derek, Editor. 2006, Academic Press. p. 251-302.
[67] Smythies J, Benington F, Morin R. On the molecular mechanism of action of the tetracyclines. Experientia, 1972. 28(10):1253-1254.
[68] Stepanović S VD, Hola V, Di Bonaventura G, Djukić S, Ćirković I, Ruzicka F. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007. 115(8):891-899.
[69] Kumar S, Wittmann C, Heinzle E. Minibioreactors. Biotechnology Letters, 2004. 26(1):1-10.
[70] Denyer S, Stewart G. Mechanisms of action of disinfectants. International Biodeterioration & Biodegradation, 1998. 41(3-4):261-268.

66 Chapter 3 _________________________________________________________________________________
[71] Paulus W, Relationship between chemical structure and activity or mode of action of micro biocides, in Directory of microbicides for the protection of materials - A Handbook, W. Paulus, Editor. 1993, Kluwer Academic Publishers: Dordrecht, The Netherlands. p. 787.
[72] Simões M. Antimicrobial strategies effective against infectious bacterial biofilms. Current Medicinal Chemistry, 2011. 18(14):2129-2145.
[73] Lemos M, Borges A, Teodósio J, Araújo PA, Mergulhão F, Melo L, Simões M. The effects of ferulic and salicylic acids on Bacillus cereus and Pseudomonas fluorescens single- and dual-species biofilms. International Biodeterioration & Biodegradation, 2014. 86(Part A):42–51.
[74] Borges A, Serra S, Cristina Abreu A, Saavedra MJ, Salgado A, Simões M. Evaluation of the effects of selected phytochemicals on quorum sensing inhibition and in vitro cytotoxicity. Biofouling, 2014. 30:183-195.

CHAPTER 4 INFLUENCE OF INTERFERING SUBSTANCES ON DISINFECTION
This chapter is published as:
Araújo PA, Lemos M, Mergulhão F, Melo L, Simões M. 2013. The influence of interfering
substances on the antimicrobial activity of selected quaternary ammonium compounds.
International Journal of Food Science; 2013:9. DOI: 10.1155/2013/237581

68 Chapter 4 _________________________________________________________________________________
ABSTRACT
Standard cleaning processes may not remove all the soiling typically found in food
industry, such as carbohydrates, fats, or proteins. Contaminants have a high impact in
disinfection as their presence may reduce the activity of disinfectants. The influence of
alginic acid, bovine serum albumin, yeast extract, and humic acids was assessed on the
antimicrobial activities of benzalkonium chloride and cetyltrimethyl ammonium
bromide against Bacillus cereus vegetative cells and Pseudomonas fluorescens. The
bacteria (single and consortium) were exposed to surfactants (single and combined) in
the absence and presence of potential disinfection interfering substances. The
antimicrobial effects of the surfactants were assessed based on the bacterial respiratory
activity measured by oxygen uptake rate due to glucose oxidation. The tested
surfactants were efficient against both bacteria (single and consortium) with minimum
bactericidal concentrations ranging from 3 to 35 mg.L−1. The strongest effect was
caused by humic acids that severely quenched antimicrobial action, increasing the
minimum bactericidal concentration of the surfactants on P. fluorescens and the
consortium. The inclusion of the other interfering substances resulted in mild
interferences in the antibacterial activity. This study clearly demonstrates that humic
acids should be considered as an antimicrobial interfering substance in the
development of disinfection strategies.

Influence of interfering substances in disinfection 69 _________________________________________________________________________________
4.1 INTRODUCTION
In order to prevent and control microbial proliferation in industrial settings, cleaning
and disinfection plans are applied on a regular basis [1, 2]. In food processing plants,
the control of microbial contamination generally involves clean-in-place (CIP)
procedures which consist of running alternated cycles of detergent and disinfectant
solutions with water rinses in high turbulence regimes through the plant and pipeline
circuits without dismantling or opening the equipment [2-5].
Biocides are currently used in industrial processes as the most significant
countermeasure to control microbial growth and proliferation [6]. Industry moved
progressively towards the use of surfactants that are less toxic and more biodegradable
[7]. Surfactants are classified according to the ionic physiognomies of their hydrophilic
group as anionic, cationic, nonionic, and zwitterionic [6, 8]. Quaternary ammonium
compounds (QACs) are cationic surfactants that are commonly used because of their
hard-surface cleaning, odor removal and antimicrobial properties [9]. Besides killing
bacteria, the chemical nature of QACs can cause modifications on the properties of
abiotic surfaces, decreasing their tension and therefore preventing attachment of
microorganisms [7]. The antimicrobial mode of action of cationic surfactants is
proposed by some authors as a sequence of events: attraction by the negatively
charged cell surface; adsorption to the cell wall through the hydrophobic headgroup;
reaction with the lipids and proteins that compose the cytoplasmic membrane; and cell
penetration and interaction with intracellular constituents [10, 11]. Thus, QACs damage
the outer layers of bacteria [9], thereby promoting the release of intracellular
constituents [12].
Antimicrobial efficacy tests require planning of an adequate strategy and should
include all the parameters found in real settings [13]. Aspects such as the proper contact
time under known water hardness and conditions of high or low soil content should be
considered [14]. For an effective cleaning and disinfection plan, the choice of the
disinfectant must follow specific criteria such as compatibility with the surfaces to be
disinfected, economic constraints, safety in the workplace, toxicological safety, and
biological degradability [15]. It should, most of all, target the type of bacteria and the
type of soiling [16]. In fact, disinfectants can be seriously affected by the presence of
organic matter [17].

70 Chapter 4 _________________________________________________________________________________
Interfering substances have been studied in the last years and included in cleaning
and disinfection plans regulated by the authorities such as the European Standard EN-
1276 (1997) [18]. There are already some reports on the effects of interfering
substances in disinfection. However, most of these studies only address the effects of
bovine serum albumin (BSA) and water hardness [9, 14, 15, 19-21]. Aal et al. [15]
evaluated the bactericidal activity of disinfectants referred in the German Veterinary
Society guidelines as references for testing disinfectants used in dairy and food
industries. In order to simulate the conditions found in practice, they used low fat milk
as an organic load and reported the significance in choosing an appropriate disinfectant
since the inclusion of a challenging substance (organic material) is important to assess
the proper bactericidal activity. Bessems [14] demonstrated that a QAC tested on three
microorganisms (Pseudomonas aeruginosa, Staphylococcus aureus, and Candida
albicans) had a similar killing rate in the absence of interfering substances and after the
inclusion of 17 dH water hardness, a strong reduction of the killing activity was found
for the Gram-negative bacteria. However, the same behavior was not verified for the
other two microorganisms. Jonõ et al. [19] assessed the effect of dried yeast and human
serum on the activity of benzalkonium chloride and concluded that the bactericidal
activity of the QAC was inhibited by solutions of both interfering substances. The
inhibition by yeast extract was more pronounced than the inhibition by human serum.
This work provides information on the influence of potential interfering
substances (bovine serum albumin - BSA, alginate - ALG, yeast extract - YE, and humic
acids - HA) on the antimicrobial activity of two QACs (benzalkonium chloride and
cetyltrimethyl ammonium bromide) against Bacillus cereus and Pseudomonas
fluorescens, as they are two major contaminants in the food industry, particularly the
dairy industry, and are a known cause of produce spoilage and foodborne illnesses
[2, 22-26]. Some of the interfering substances used throughout the experiments are
proposed in the European Standard EN-1276 (1997) [18] as potential interfering agents
in disinfection while the others are extracellular polymeric substances (EPS) from the
biofilm matrix that have an important role in antimicrobial resistance [27].

Influence of interfering substances in disinfection 71 _________________________________________________________________________________
4.2 MATERIALS AND METHODS
MICROORGANISMS AND CULTURE CONDITIONS
The bacteria used in this work were Pseudomonas fluorescens ATCC 13525T and a
Bacillus cereus strain, isolated from a disinfectant solution and identified by 16S rRNA
gene sequencing [28].
Bacterial strains were grown at a temperature of 30 ± 3 oC and pH 7, with glucose
as the main carbon source. Culture medium consisted of 5 g.L−1 glucose, 2.5 g.L−1
peptone, and 1.25 g.L−1 yeast extract in phosphate buffer (PB) (pH 7, 0.025 M) [29]. A
bacterial suspension was prepared by inoculation of a single colony grown on solid
medium into a 1 L flask containing 250 mL of sterile nutrient medium. This bacterial
suspension was incubated overnight at the given temperature with agitation (120 rpm).
QACS AND INTERFERING AGENTS
The QACs used throughout the experiments were benzalkonium chloride (BAC) and
cetyltrimethyl ammonium bromide (CTAB) (Sigma, Portugal) (Figure 4.1). Preliminary
studies with a concentration range between 0 and 5000 mg.L−1 were initially made. In
order to ascertain the behaviour of bacteria to the QAC, the selected concentrations for
further studies were 3, 5, 10, 20, and 35 mg.L−1. The QACs were used individually and in
combination (both chemicals were combined in equal volumes and concentrations).
The interfering substances used throughout the experiments were alginic acid
sodium salt -ALG (Sigma, Portugal), bovine serum albumin - BSA (Sigma, Portugal),
humic acids -HA (Acros organics, Fisher Chemical, Portugal), and yeast extract - YE
(Merck, Portugal).
Figure 4.1 Chemical structures of benzalkonium chloride (A) and cetyltrimethyl ammonium
bromide (B).
A B

72 Chapter 4 _________________________________________________________________________________
DISINFECTION PROCEDURE
After the growth period, the suspensions were centrifuged (3999 g, 5 minutes), washed
two times, and resuspended in PB to a final cell density of approximately 1 × 109 cells.
mL−1. In the case of the consortium, both bacterial suspensions were washed two times
resuspended in PB to a final cell density of approximately 1 × 109 cells.mL−1, and
combined in equal volumes to obtain the same cell concentrations of the single species
tests. Afterwards, all bacterial suspensions were exposed to several concentrations of
QAC for a period of 30 minutes [30]. The effects of the chemicals were evaluated by the
assessment of the oxygen uptake rate due to glucose oxidation, according to Simões et
al. [30].
To investigate the influence of interfering substances on the antimicrobial efficacy,
the same procedure was followed with the addition of 300 mg.L−1 of BSA, ALG, YE, or
HA to the bacterial suspension, simulating low concentrations of interfering substances
according to the European Standard EN-1276 (1997) [18]. Three independent
experiments, each with duplicate samples, were performed for each condition tested.
QACS NEUTRALIZATION
A neutralization process was performed after the disinfection procedure. The
methodology was performed according to Johnston et al. [31] for a period of 10
minutes. BAC and CTAB were chemically neutralized by a sterile solution of (w/v)
0.1% peptone, 0.5% Tween 80, 0.1% sodium thiosulphate, and 0.07% lecithin dissolved
in PB. All the chemicals were obtained from Sigma (Portugal). Control experiments were
performed to ascertain the effects of the 10-minute exposure to the neutralization
solution, and no effects were detected on the respiratory activity of B. cereus and
P. fluorescens. After the neutralization step, the bacterial suspensions were centrifuged
(3999 g, 5 min) and resuspended in the same volume of PB.
RESPIRATORY ACTIVITY ASSESSMENT
The respiratory activity was ascertained by measuring oxygen uptake rates in a
biological oxygen monitor (Yellow Springs Instruments 5300A). Simões et al. [30]
demonstrated that this procedure is more adequate and rapid than the assessment of

Influence of interfering substances in disinfection 73 _________________________________________________________________________________
colony forming units to characterize the antimicrobial activity of biocides against
heterotrophic aerobic bacteria [21]. Samples were placed in the temperature-
controlled vessel of the biological oxygen monitor (T= 25 ± 1 oC) each containing a
dissolved oxygen probe connected to a dissolved oxygen meter. Before measuring, the
samples were aerated for 10 minutes to ensure oxygen saturation ([O2] = 8.6 mg.L−1).
The vessel was closed, and the decrease of oxygen concentration was monitored over
time. The initial linear decrease corresponds to the endogenous respiration rate. To
determine the oxygen uptake due to substrate oxidation, 12.5 µL of a 5 g.L−1 glucose
solution was added to each vessel. The slope of the initial linear decrease in dissolved
oxygen, after glucose injection, corresponds to the total respiration rate. The difference
between these two rates is the oxygen uptake rate due to glucose oxidation [9].
The inactivation was calculated using metabolic activities according to the following equation:
100m
)m(monInactivati
c
tc
% (eq. 4.1)
where mc is the metabolic activity of the control experiments (without antimicrobial
exposure) and mt is the metabolic activity of the bacterial solutions exposed to the
antimicrobial. If % inactivation > 0 there was inactivation of the microorganisms
whereas if % inactivation < 0 there was metabolic potentiation. The MBC for each
situation was determined as the lowest concentration of QAC or QAC combination
where no respiratory activity was detected [31].
STATISTICAL ANALYSIS
For each parameter tested the average and the standard deviation were calculated. The
statistical significance of the results was evaluated using the Wilcoxon test (confidence
level ≥ 95%), and for the MBC the independent t-test was used to investigate whether
the differences between the resulting experimental values could be considered
significant.

74 Chapter 4 _________________________________________________________________________________
4.3 RESULTS
The antibacterial activity of BAC, CTAB, and their combination was investigated in the
absence and in the presence of four selected interfering substances.
In the absence of interfering substances BAC caused the inactivation of B. cereus
at 10 mg.L−1, P. fluorescens at 35 mg.L−1, and the consortium at 20 mg.L−1. CTAB at
20 mg.L−1 completely inactivated B. cereus and at 35 mg.L−1 inactivated the total
population of P. fluorescens and the consortium. The combination of both QACs was
synergistic in the inactivation of B. cereus (total inactivation with 3 mg.L−1) and
indifferent for P. fluorescens (35 mg.L−1) and the bacterial consortium (35 mg.L−1). The
inclusion of the selected interfering substances influenced the antimicrobial activity of
the QACs to some extent (Figures 4.2-4.4). The inactivation of B. cereus (Figure 4.1) was
not affected by the presence of any interfering substances (P > 0.05), except with HA.
This interfering substance decreased the antimicrobial efficacy of BAC and the
combination of QACs. The antimicrobial action of the QACs against P. fluorescens
(Figure 4.3) was not significantly influenced by the presence of most potential
interfering substances (P > 0.05), except for HA where interference was observed
(P < 0.05). The antimicrobial activity of the QACs against the bacterial consortium
(Figure 4.3) was affected by the presence of interfering substances. ALG and HA
reduced significantly the activity of BAC (P < 0.05). HA reduced significantly the activity
of CTAB at higher concentrations (P < 0.05). BSA and YE resulted in a significant
reduction of the activity of the combination of QACs (P < 0.05).
Linear correlations were determined to assess the relationship between QAC
concentrations and the inactivation data. The effect of increasing QAC concentration
on bacterial inactivation shows that there are strong linear correlations (R > 0.850) for
the control assays, with the exception of B. cereus (this bacterium was inactivated with
low QAC concentrations). When interfering substances were added, the correlations
decreased. The most extreme cases are the treatments with CTAB to P. fluorescens with
ALG as an interfering substance (R = 0.771) and the bacterial consortium in the presence
of YE (R = 0.738). Likewise, this decrease of linear correlation factors was found for
P. fluorescens and for the consortium exposed to HA where the lowest correlation factor
was 0.153, which was obtained for P. fluorescens treated with CTAB.
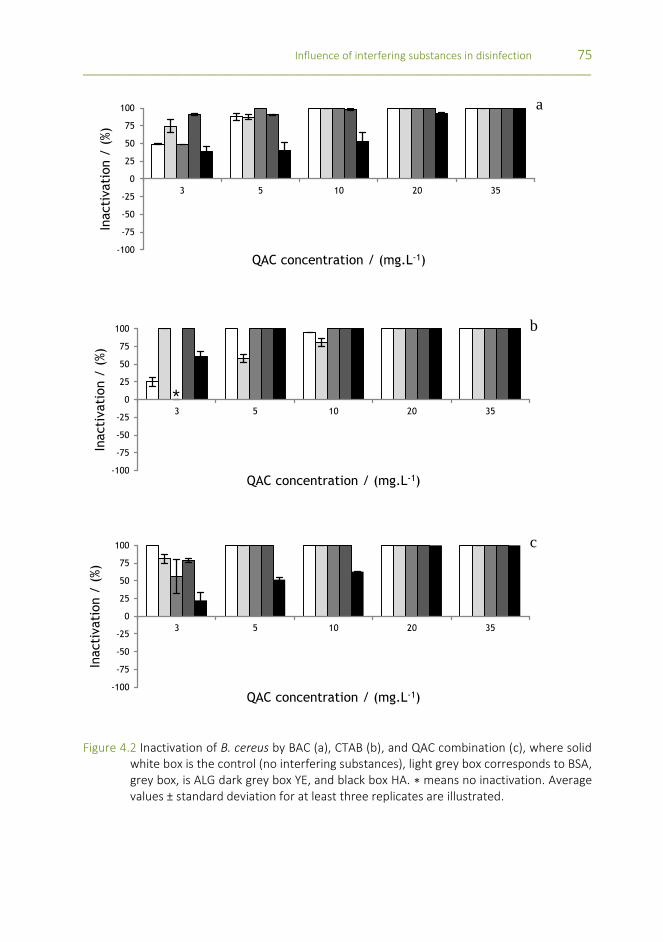
Influence of interfering substances in disinfection 75 _________________________________________________________________________________
Figure 4.2 Inactivation of B. cereus by BAC (a), CTAB (b), and QAC combination (c), where solid white box is the control (no interfering substances), light grey box corresponds to BSA, grey box, is ALG dark grey box YE, and black box HA. ∗ means no inactivation. Average values ± standard deviation for at least three replicates are illustrated.
-100
-75
-50
-25
0
25
50
75
100
3 5 10 20 35
Inacti
vati
on /
(%)
QAC concentration / (mg.L-1)
-100
-75
-50
-25
0
25
50
75
100
3 5 10 20 35
Inacti
vati
on /
(%)
QAC concentration / (mg.L-1)
*
-100
-75
-50
-25
0
25
50
75
100
3 5 10 20 35
Inacti
vati
on /
(%)
QAC concentration / (mg.L-1)
a
b
c

76 Chapter 4 _________________________________________________________________________________
Figure 4.3 Inactivation of P. fluorescens by BAC (a), CTAB (b), and QAC combination (c), where solid white box is the control (no interfering substances), light grey box corresponds to BSA, grey box is ALG, dark grey box is YE, and black box is HA. ∗ means no inactivation. Values below zero are indication that the metabolic activity increased in comparison with the control experiment. Average values ± standard deviation for at least three replicates are illustrated.
-100
-75
-50
-25
0
25
50
75
100
3 5 10 20 35
Inacti
vati
on /
(%)
QAC concentration / (mg.L-1)
* ****
-100
-75
-50
-25
0
25
50
75
100
3 5 10 20 35
Inacti
vati
on /
(%)
QAC concentration / (mg.L-1)
** *
-100
-75
-50
-25
0
25
50
75
100
3 5 10 20 35
Inacti
vati
on /
(%)
QAC concentration / (mg.L-1)
** * *
a
b
c

Influence of interfering substances in disinfection 77 _________________________________________________________________________________
Figure 4.4 Inactivation of the bacterial consortium by BAC (a), CTAB (b), and QAC combination (c), where solid white box is the control (no interfering substances), light grey box corresponds to BSA, grey box is ALG, dark grey box is YE, and black box is HA. ∗ means no inactivation. Values below zero are indication that the metabolic activity increased in comparison with the control experiment. Average values ± standard deviation for at least three replicates are illustrated.
-100
-75
-50
-25
0
25
50
75
100
3 5 10 20 35
Inacti
vati
on /
(%)
QAC concentration / (mg.L-1)
* *
-100
-75
-50
-25
0
25
50
75
100
3 5 10 20 35
Inacti
vati
on /
(%)
QAC concentration / (mg.L-1)
* *
-100
-75
-50
-25
0
25
50
75
100
3 5 10 20 35
Inacti
vati
on /
(%)
QAC concentration / (mg.L-1)
a
b
c

78 Chapter 4 _________________________________________________________________________________
The results also demonstrate the occurrence of metabolic potentiation
(inactivation below 0 %). This phenomenon only happened when the QACs were used
on P. fluorescens and the bacterial consortium in the presence of YE and HA. The most
significant cases of oxygen uptake rate increase were verified for P. fluorescens exposed
to BAC (5 to 35 mg.L−1) and CTAB (3 to 35 mg.L−1) in the presence of HA and combination
of QACs (3 to 10 mg.L−1) in the presence of YE. A similar metabolic behavior was found
for the bacterial consortium exposed to BAC (3 to 35 mg.L−1) and CTAB (5 and 10 mg.L−1)
for HA and QAC combination (3 to 20 mg.L−1) with YE.
The MBC values for the different conditions tested (single and combined QACs, in
the absence and presence of potential disinfection interfering substances) are shown in
Table 4.1.
Table 4.1 Minimum bactericidal concentration for P. fluorescens, B. cereus and the consortium with and without interfering substances.
MBC (mg.L-1)
BAC CTAB QAC combination
Control
B. cereus 10 20 3
P. fluorescens 35 35 35
Consortium 20 35 35
BSA
B. cereus 10 20 5
P. fluorescens 35 20 35
Consortium 20 35 >35
ALG
B. cereus 5 5 5
P. fluorescens 35 35 35
Consortium >35 35 20
YE
B. cereus 20 3 5
P. fluorescens 35 35 >35
Consortium 35 >35 >35
HA
B. cereus 35 5 20
P. fluorescens >35 >35 >35
Consortium >35 >35 >35

Influence of interfering substances in disinfection 79 _________________________________________________________________________________
The presence of BSA increased the MBC of the combination of QACs for B. cereus
(3 to 5 mg.L−1) and the consortium. ALG increased the MBC of BAC for the consortium
(20 to over 35 mg.L−1) and QACs combination (3 to 5 mg.L−1) for B. cereus. YE increased
the MBC of BAC for B. cereus (10 to 20 mg.L−1) and QAC combination (3 to 5 mg.L−1).
P. fluorescens MBC increased with the inclusion of YE with the combination of QACs.
The MBC values for the consortium of cells increased in the presence of YE (BAC - 20 to
35 mg.L−1, CTAB - 35 to over 35 mg.L−1, and QAC combination - 35 to over 35 mg.L−1).
HA increased the MBC for all the scenarios, except of CTAB when applied to B. cereus
(in this situation the MBC was reduced). The MBC was reduced in other situations such
as, for B. cereus, in the presence of ALG when using BAC and CTAB (10 to 5 mg.L−1 and
20 to 5 mg.L−1, respectively) and in the presence of YE when using CTAB (20 to 3 mg.L−1).
P. fluorescens inactivation by CTAB was reduced by BSA (35 to 20 mg.L−1). ALG also
reduced the antimicrobial activity of the combination of QACs against the bacterial
consortium (35 to 20 mg.L−1).
4.4 DISCUSSION
In disinfection practices, the environmental characteristics can influence the
antimicrobial activity of biocides [32]. It is assumed that the organic material can
potentially interfere with the antimicrobial agents by chemical and/or ionic interactions
[15, 33]. Therefore, it is necessary to know the role of each potential interfering
substance in the antimicrobial activity in order to develop effective disinfection
strategies. The interfering substances tested are commonly found as residuals in the
food industry (from food products and from microbial contaminants, biofilms) [18, 27].
In this study, higher inactivation rates were verified for B. cereus in comparison to
P. fluorescens at the same QAC concentration. The inactivation profiles of the cell
consortium are similar to P. fluorescens. In fact, when B. cereus and P. fluorescens are
combined in a 1:1 bacterial suspension, it is expected that the first is more affected than
the second. B. cereus is more susceptible due to the fact that it is a Gram positive
bacterium that lacks an outer membrane, which typically provides increased protection
to Gram negative bacteria. This fact is corroborated by previous reports which stated
that Gram positive bacteria are more susceptible to cationic surfactants than Gram
negative bacteria [34, 35].

80 Chapter 4 _________________________________________________________________________________
BSA was already studied as an interfering substance in disinfection practices
[9, 14, 19-21, 36]. The negative effect of BSA on the action of biocides against
P. fluorescens was demonstrated by Simões et al. [9, 21]. P. fluorescens treatment with
CTAB with the addition of 3 g.L−1 of BSA resulted in a 10-fold increase on the MBC of
this QAC [9, 21]. In the present study, low BSA concentrations decreased the
antimicrobial activity of the QACs. The efficacy of the combination of QACs against
B. cereus and the cell consortium was also reduced. This effect of BSA as an
antimicrobial quencher is apparently due to the strong ability of QACs to react with
proteins [21]. Proteins can precipitate in the form of their anions. In this way, the
negative-charged protein ions will cling to the positively charged molecules of the
cationic compounds [37]. CTAB is a biocide that targets the membrane and has a strong
affinity for proteins [21]. BAC is composed of a positively charged hydrophobic
headgroup which clings to opposite charged surfaces [8, 37]. Jonõ et al. [19] studied the
effect of the alkyl chain of BAC binding to BSA and dried yeast. Their conclusions were
that BAC is often inactivated by organic matter, either by adsorption to the bacterial
surface or by adsorption to the organic matter in general. These authors also suggested
that the reduction in the activity of BAC was probably related to more than one physical
property of the compounds like the chain length (longer chains result in more
adsorption to the bacterial surface).
ALG is a common constituent of the extracellular polymeric substances of the
biofilm matrix [38-40]. A function frequently attributed to EPS is their general protective
effect on biofilm microorganisms against adverse conditions. The EPS matrix delays or
prevents antimicrobials from reaching target microorganisms within the biofilm by
diffusion limitation and/or chemical interaction with the extracellular proteins and
polysaccharides [32, 41]. In this study, ALG either potentiated or hindered the
antimicrobial activity of the selected QACs. The presence of this interfering substance
was not obvious on the inactivation of P. fluorescens. On the other hand, the
inactivation of B. cereus by BAC and CTAB and the consortium by the combination of
QACs was easier in the presence of this interfering substance. The bacterial consortium
treatments with BAC and B. cereus with the combination of QACs were hampered by
the presence of ALG. Davies et al. [42, 43] found that the production of ALG was
triggered by membrane perturbation induced by ethanol stress, nitrogen limitation,
attachment to surfaces, or even high oxygen tension. This substance is suggested as one
of the main biofilm resistance vectors either by reacting with the antimicrobials or by

Influence of interfering substances in disinfection 81 _________________________________________________________________________________
hindering antimicrobials diffusion to the cells [44].The antimicrobial interference
caused by ALG is apparently due to electrostatic interactions between the anionic ALG
and the cationic-selected QACs [45].
The presence of YE as interfering substance resulted in three different outcomes
on the antimicrobial activity of the QACs: (1) no effect/indifference, (2) the respiratory
activity reduced, and (3) the respiratory activity potentiated. This interfering substance
worked mainly as a hinderer of the antimicrobial activity by increasing the MBC of
B. cereus in all cases except for CTAB, of P. fluorescens with the combination of QACs,
and of the consortium of cells with CTAB and the combination of QACs. These results
are in accordance with the available studies. YE is listed in the European Standard EN-
1276 (1997) as an interfering substance native to the brewery industry [18]. The
constituents of YE are very similar to the components of the bacterial cells, thus, it is
expected that the antimicrobial agents that target the bacterial cells are also drawn to
YE. In a similar study by Jonõ et al. [19] it was shown that the presence of dried yeast
decreased the biocidal effectiveness of BAC.
Humic substances are found ubiquitously in the environment and can be found in
the biofilm matrix [2, 46]. HA reduced the antimicrobial activity of the QACs in most of
the cases, although in some cases it promoted the respiratory activity (potentiation).
The presence of these compounds had the strongest effect compared to the remaining
interfering substances. Like ALG, HA are known to be a part of the EPS composition [47].
Atay et al. [8] studied the sorption mechanisms of anionic and cationic surfactants to
natural soils concluding that the dominant sorption mechanism of surfactants to clay is
cation exchange. Ishiguro et al. [48] reported that cationic surfactants bind strongly to
humic substances. Koopal et al. [49] also verified the formation of complexes HA-
cationic surfactant. These observations are consistent with the present results.
Respiratory activity potentiation was verified with the addition of HA to
P. fluorescens, and YE to the bacterial consortium. It is known that HA participates in
cellular metabolism processes such as growth, respiration, photosynthesis, and
nitrogen fixation [50]. On the other hand, HA were proposed to replace synthetic
surfactants such as SDS, Tween 80, and Triton X-100 in industrial applications such as
textile dying or washing [51]. It is therefore possible that the inclusion of humic
substances in a solution of QACs may interfere with the chemical characteristics of the
solution. The resultant mixture, with an apparent reduced antimicrobial efficacy, seems
to potentiate the respiratory activity of the bacteria, particularly of P. fluorescens. As

82 Chapter 4 _________________________________________________________________________________
QACs are membrane active agents, their use at sub-lethal concentrations could improve
membrane permeability and consequently the nutrient influx, without compromising
the bacterial viability. Also, there is the hypothesis that the potentially interfering
agents could be used as nutrients. In fact, it was found that the growth rates of
anaerobic and aerobic microorganisms increased when humic substances were added,
which stimulated enzyme activity [52, 53]. In a similar way, YE is a nitrogen source
widely used as a component of growth media [54]. HA are likely to be used for growth
in the same way as YE, these might be broken down to smaller molecules that can be
used by cells as a carbon [55] or nitrogen sources [51].
The antimicrobial activity of the tested QACs was enhanced in some cases, where
the interfering substances were present. This is an unexpected result due to the
recognized and observed potential of ALG, BSA, HA, and YE to interfere with
disinfection. This effect is probably due to the low concentration of interfering
substances tested that caused both respiratory activity reduction and potentiation.
Cases of antimicrobial enhancement are widely known. Ethylenediamine tetraacetate
(EDTA) was reported as early as 1965 to increase the biocidal effects of BAC and
chlorhexidine diacetate on Pseudomonas aeruginosa [56]. Sagoo et al. [57] reported
that chitosan (a polysaccharide) potentiated the antimicrobial action of sodium
benzoate on spoilage yeasts. In dairy plants, disinfection is potentiated by prewashes
with alkali or enzyme-based cleaning agents [58]. The antimicrobial potentiation of the
QACs occurred in some cases. Most of these cases were observed for B. cereus (four
occurrences), one was observed for P. fluorescens, and another one was observed for
the consortium of cells. The MBC was improved by more than 50% in the cases of
B. cereus and less than 30% for P. fluorescens and the consortium of cells. To our
knowledge there are no reported cases of antimicrobial agents potentiation by BSA, YE,
or ALG. Concerning the effects of HA, these molecules are reported to have detergent
properties [51]. Although the exact chemical structure of HA has not yet been
determined, HA could be chemically similar to the tested QACs, presenting a positive
hydrophilic head and a hydrophobic tail. With this structure HA could act as detergents
in conditions such as those observed in the treatment of B. cereus with CTAB [51].
The present work shows that increasing QACs concentrations lead to an increase
in antimicrobial effectiveness. This is valid mainly when the QACs were applied in the
absence of interfering substances. This means that disinfection was concentration
dependent, as found for most of the antimicrobial chemicals [59]. However, the linear

Influence of interfering substances in disinfection 83 _________________________________________________________________________________
dependency of inactivation versus concentration is not verified for most of the tests
where interfering substances were added. This result evidences that the mathematical
modelling of disinfection strategies requires a case-to-case analysis when interfering
substances are present.
4.5 CONCLUSIONS
The overall results demonstrate that a disinfection process in the presence of the
selected interfering substances can reduce the effectiveness of BAC, CTAB, and their
combination. The bacteria were inactivated equally by all QACs, although in the absence
of interfering substances CTAB was the most efficient solution. P. fluorescens was the
bacterium with the highest resistance to inactivation, followed by the bacterial
consortium. The tested interfering substances, referred in the European Standard
EN1276 (BSA and YE), and known EPS constituents related with biofilm resistance (ALG)
resulted in mild interferences on the activity of the QACs. HA were the interfering
substance that resulted in the most severe effect by reducing the activity of QACs,
causing, in some circumstances, significant respiratory activity potentiation. This
interfering substance should, therefore, be considered when developing disinfection
protocols.
REFERENCES
[1] Dooley JSG,Roberts TA. Control of vegetative micro-organisms in foods. British Medical Bulletin, 2000. 56(1):142-157.
[2] Simões M, Simões L, Vieira M. A review of current and emergent biofilm control strategies. LWT - Food Science and Technology, 2010. 43(4):573-583.
[3] Bremer PJ, Fillery S, McQuillan AJ. Laboratory scale Clean-In-Place (CIP) studies on the effectiveness of different caustic and acid wash steps on the removal of dairy biofilms. International Journal of Food Microbiology, 2006. 106(3):254-262.
[4] Srey S, Jahid IK, Ha SD. Biofilm formation in food industries: A food safety concern. Food Control, 2013. 31(2):572-585.
[5] Forsythe SJ, Hayes PR, Hayes PR. Food hygiene, microbiology, and HACCP. 3rd ed. A Chapman & Hall food science book. Gaithersburg, Md.: Aspen Publishers. 1998 xviii, 449 p.
[6] Simões M, Simões L, Machado I, Pereira M, Vieira M. Control of flow-generated biofilms with surfactants: Evidence of resistance and recovery. Food and Bioproducts Processing, 2006. 84(4):338-345.
[7] Simões M, Pereira MO, Vieira MJ. Effect of mechanical stress on biofilms challenged by different chemicals. Water Research, 2005. 39(20):5142-5152.

84 Chapter 4 _________________________________________________________________________________
[8] Atay N, Yenigün O, Asutay M. Sorption of anionic surfactants SDS, AOT and cationic surfactant hyamine 1622 on natural soils. Water air and soil pollution, 2002. 136(1):55-68.
[9] Simões M, Pereira MO, Vieira MJ. Action of a cationic surfactant on the activity and removal of bacterial biofilms formed under different flow regimes. Water Research, 2004. 39(2-3):478-486.
[10] Salton M. Lytic agents, cell permeability, and monolayer penetrability. Journal of General Physiology, 1968. 52(1):227-252.
[11] Paulus W, Relationship between chemical structure and activity or mode of action of micro biocides, in Directory of microbicides for the protection of materials - A Handbook, W. Paulus, Editor. 1993, Kluwer Academic Publishers: Dordrecht, The Netherlands. p. 787.
[12] Ferreira C, Pereira A, Pereira M, Melo L, Simões M. Physiological changes induced by the quaternary ammonium compound benzyldimethyldodecylammonium chloride on Pseudomonas fluorescens. Journal of Antimicrobial Chemotherapy, 2011. 66(5):1036-1043.
[13] Cloete TE, Jacobs L, Brözel VS. The chemical control of biofouling in industrial water systems. Biodegradation, 1998. 9:23-27.
[14] Bessems E. The effect of practical conditions on the efficacy of disinfectants. International Biodeterioration & Biodegradation, 1998. 41(3-4):177-183.
[15] Aal SFAAE, Hunsinger B, Böhm R. Determination of the bactericidal activity of chemical disinfectants bacteria in dairies according to the DVG-guidelines. Hygiene & Medizin 2008. 33(11).
[16] Gram L, Bagge-Ravn D, Ng YY, Gymoese P, Vogel BF. Influence of food soiling matrix on cleaning and disinfection efficiency on surface attached Listeria monocytogenes. Food Control, 2007. 18(10):1165-1171.
[17] Lambert RJW,Johnston MD. The effect of interfering substances on the disinfection process: a mathematical model. Journal of Applied Microbiology, 2001. 91(3):548-555.
[18] European standard EN-1276. Chemical disinfectants and antiseptics-Quantitative suspension test for the evaluation of bactericidal activity of chemical disinfectants and antiseptics used in food, industrial, domestic, and institutional areas-Test method and requirements (phase 2, step 1). 1997.
[19] Jonõ K, Takayama T, Kuno M, Higashide E. Effect of alkyl chain-length of benzalkonium chloride on the bactericidal activity and binding to organic materials. Chemical & Pharmaceutical Bulletin, 1986. 34(10):4215-4224.
[20] Gani S, Chattoraj D, Mukherjee D. Binding of cationic surfactants to DNA, protein and DNA-protein mixtures. Indian Journal of Biochemistry & Biophysics, 1999. 36(3):165-176.
[21] Simões M, Pereira M, Machado I, Simões L, Vieira M. Comparative antibacterial potential of selected aldehyde-based biocides and surfactants against planktonic Pseudomonas fluorescens. Journal of Industrial Microbiology & Biotechnology, 2006. 33(9):741-749.
[22] Simões M, Simões LC, Vieira MJ. Species association increases biofilm resistance to chemical and mechanical treatments. Water Research, 2009. 43(1):229-237.
[23] Kumar CG,Anand SK. Significance of microbial biofilms in food industry: a review. International Journal of Food Microbiology, 1998. 42(1-2):9-27.
[24] Simões LC, Lemos M, Araujo P, Pereira AM, Simões M. The effects of glutaraldehyde on the control of single and dual biofilms of Bacillus cereus and Pseudomonas fluorescens. Biofouling, 2011. 27(3):337-346.
[25] Johnson KM. Bacillus cereus foodborne illness-an update. Journal of Food Protection, 1984. 47(2):145-153.

Influence of interfering substances in disinfection 85 _________________________________________________________________________________
[26] Notermans S, Dufrenne J, Teunis P, Beumer R, te Giffel M, Peeters Weem P. A risk assessment study of Bacillus cereus present in pasteurized milk. Food Microbiology, 1997. 14(2):143-151.
[27] Cloete T. Resistance mechanisms of bacteria to antimicrobial compounds. International Biodeterioration & Biodegradation, 2003. 51(4):277-282.
[28] Simões M, Simões L, Pereira M, Vieira M. Antagonism between Bacillus cereus and Pseudomonas fluorescens in planktonic systems and in biofilms. Biofouling, 2008. 24(5):339-349.
[29] Simões M, Cleto S, Pereira M, Vieira M. Influence of biofilm composition on the resistance to detachment. Water science and technology, 2007. 55:473-480.
[30] Simões M, Pereira M, Vieira M. Validation of respirometry as a short-term method to assess the efficacy of biocides. Biofouling, 2005. 21(1):9-17.
[31] Johnston MD, Lambert RJW, Hanlon GW, Denyer SP. A rapid method for assessing the suitability of quenching agents for individual biocides as well as combinations. Journal of Applied Microbiology, 2002. 92(4):784-789.
[32] Heinzel M. Phenomena of biocide resistance in microorganisms. International Biodeterioration & Biodegradation, 1998. 41(3-4):225-234.
[33] Otzen D. Protein-surfactant interactions: a tale of many states. Biochimica et Biophysica Acta - Proteins & Proteomics, 2011. 1814(5):562-591.
[34] McDonnell G,Russell A. Antiseptics and disinfectants: activity, action, and resistance. Clinical Microbiology Reviews, 1999. 12(1):147-179.
[35] Russell AD. Biocide use and antibiotic resistance: the relevance of laboratory findings to clinical and environmental situations. The Lancet Infectious Diseases, 2003. 3(12):794-803.
[36] Møretrø T, Vestby LK, Nesse LL, Storheim SE, Kotlarz K, Langsrud S. Evaluation of efficacy of disinfectants against Salmonella from the feed industry. Journal Applied Microbiology, 2009. 106(3):1005-12.
[37] Lawrence CA. Mechanism of Action and Neutralizing Agents for Surface-Active Materials Upon Microorganisms. Annals of the New York Academy of Sciences, 1950. 53(1):66-75.
[38] Christensen B, Ertesvag H, Beyenal H, Lewandowski Z. Resistance of biofilms containing alginate-producing bacteria to disintegration by an alginate degrading enzyme (AlgL). Biofouling, 2001. 17(3):203-210.
[39] Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Science 1999. 284(5418):1318-22.
[40] Watnick P,Kolter R. Biofilm, city of microbes. Jounal of Bacteriology, 2000. 182(10):2675-2679.
[41] Mah T,O'Toole G. Mechanisms of biofilm resistance to antimicrobial agents. Trends in Microbiology, 2001. 9(1):34-39.
[42] Davies DG, Chakrabarty AM, Geesey GG. Exopolysaccharide production in biofilms - substratum activation of alginate gene-expression by Pseudomonas aeruginosa. Applied and Environmental Microbiology, 1993. 59(4):1181-1186.
[43] Davies DG,Geesey GG. Regulation of the alginate biosynthesis gene Algc in Pseudomonas aeruginosa during biofilm development in continuous culture. Applied and Environmental Microbiology, 1995. 61(3):860-867.
[44] Stewart P, ed. Multicellular nature of biofilm protection from antimicrobial agents. Biofilm communities: Order or Chaos, ed. A.J. McBain, et al. 2003. 181-190.
[45] Shinde UA,Nagarsenker MS. Characterization of gelatin-sodium alginate complex coacervation system. Indian Journal of Pharmaceutical Sciences, 2009. 71(3):313-317.

86 Chapter 4 _________________________________________________________________________________
[46] Ghabbour EA, Davies G, International Humic Substances Society. Humic substances : nature's most versatile materials. New York: Taylor and Francis. 2004 xxii, 372 p.
[47] Tong M, Zhu P, Jiang X, Kim H. Influence of natural organic matter on the deposition kinetics of extracellular polymeric substances (EPS) on silica. Colloids and Surfaces B: Biointerfaces, 2011. 87(1):151-158.
[48] Ishiguro M, Tan W, Koopal LK. Binding of cationic surfactants to humic substances. Colloids and Surfaces A: Physicochemical and Engineering Aspects, 2007. 306(1-3):29-39.
[49] Koopal LK, Goloub TP, Davis TA. Binding of ionic surfactants to purified humic acid. Journal of Colloid and Interface Science, 2004. 275(2):360-367.
[50] Visser SA. Physiological action of humic substances on microbial cells. Soil Biology & Biochemistry, 1985. 17(4):457-462.
[51] Salati S, Papa G, Adani F. Perspective on the use of humic acids from biomass as natural surfactants for industrial applications. Biotechnology Advances, 2011. 29(6):913-922.
[52] Hartung HA. Stimulation of anaerobic-digestion with peat humic substance. Science of the Total Environment, 1992. 113(1-2):17-33.
[53] Pouneva ID. Effect of humic substances on the growth of microalgal cultures. Russian Journal of Plant Physiology, 2005. 52(3):410-413.
[54] Hakobyan L, Gabrielyan L, Trchounian A. Yeast extract as an effective nitrogen source stimulating cell growth and enhancing hydrogen photoproduction by Rhodobacter sphaeroides strains from mineral springs. International Journal of Hydrogen Energy, 2012. 37(8):6519-6526.
[55] Camper AK. Involvement of humic substances in regrowth. International Journal of Food Microbiology, 2004. 92(3):355-364.
[56] Brown MR,Richards RM. Effect of ethylenediamine tetraacetate on the resistance of Pseudomonas aeruginosa to antibacterial agents. Nature, 1965. 207(5004):1391-3.
[57] Sagoo SK, Board R, Roller S. Chitosan potentiates the antimicrobial action of sodium benzoate on spoilage yeasts. Letters in Applied Microbiology 2002. 34(3):168-172.
[58] Langsrud S, Baardsen B, Sundheim G. Potentiation of the lethal effect of peroxygen on Bacillus cereus spores by alkali and enzyme wash. International Journal of Food Microbiology, 2000. 56(1):81-86.
[59] Chapman J. Biocide resistance mechanisms. International Biodeterioration & Biodegradation, 2003. 51(2):133-138.

CHAPTER 5 HYDRODYNAMIC CONDITIONS AND BIOFILM DEVELOPMENT
This chapter was submitted as:
Araújo PA, Malheiro J, Mergulhão F, Melo L, Simões M. The influence of linear flow velocity on the
characteristics of Pseudomonas fluorescens biofilms.

88 Chapter 5 _________________________________________________________________________________
ABSTRACT
The characteristics of Pseudomonas fluorescens biofilms formed under three different
linear flow velocities (u = 0.1, 0.4 and 0.8 m.s-1) were studied. A flow cell reactor system
was used to form biofilms. These biofilms were characterized in terms of thickness,
morphological structure, mass, cell density, outer membrane proteins expression, and
matrix and total proteins and polysaccharide content. The external mass transfer
coefficients were also calculated.
Biofilms developed at the higher velocities (u = 0.4 and 0.8 m.s-1) had similar
characteristics, but different from those developed at the lower flow rate. High flow
velocities formed thinner biofilms with higher cell densities, and higher contents of
matrix proteins and polysaccharides. The external mass transfer coefficients suggest
mass transfer limitations from the bulk fluid for the lowest velocity. Scanning electron
microscopy images show cell-surface and cell-cell attachment structures appearing
more frequently in biofilms formed at the two higher velocities. No major differences
were found in the outer membrane proteins expression of biofilm cells, regardless of
the linear flow velocity under which they were formed. The overall results show the
effect of the hydrodynamic conditions under which biofilms were formed on selected
macromolecular characteristics, demonstrating that higher flow velocities originate
more complex and dense biofilms. However, cellular aspects as the outer membrane
proteins expression are not affected by the flow velocity.
Understanding biofilm formation and corresponding characteristics allows the
manipulation of hydrodynamic conditions as a control parameter in the improvement
of biofilm control strategies in many engineered systems.

Biofilm development under different flow velocities 89 _________________________________________________________________________________
5.1 INTRODUCTION
Bacterial attachment to surfaces and the consequent biofilm formation is a well-
recognized phenomenon in diverse areas such as the food and biomedical fields [1-3].
Especially in food industry, bacterial spoilage is a major concern with both economic
and public health consequences. Therefore, efforts must be directed for efficient
industrial equipment design and the development of effective cleaning and disinfection
strategies [1, 4, 5].
Biofilms can be described as dense microbial communities associated to surfaces,
which are highly hydrated clusters of bacterial cells surrounded by a matrix of
extracellular polymeric substances (EPS) [6-8]. EPS, a result of bacterial secretion, cell
lysis and hydrolysis, are constituted by biopolymers such as polysaccharides, proteins,
extracellular DNA and lipids. These substances are responsible for the protection of
bacteria from environmental stress, dehydration and chemical exposure and mediate
bacterial adhesion to surfaces [6, 9-11]. In addition, EPS are essential for biofilm stability
and architecture since their composition, structure and properties influences oxygen
penetration and substrate absorption and transport [9]. Moreover, the biofilm
structure depends on the microbial constituents and environmental factors like
composition, pH and temperature of the contact fluid, surface properties and
hydrodynamic conditions [12-14].
The hydrodynamic effects can induce a detachment force as a consequence of gas
or liquid flow and particle-particle collision [10, 15]. Therefore, shear force has been
considered a pivotal factor in biofilm formation, since it leads to equilibrium between
biofilm thickness and density resulting in a steady state structure. Several authors found
that higher shear force caused thinner and denser biofilms [10, 14, 16, 17]. Considering
that the biofilm structure is influenced by the existing hydrodynamic conditions, the
latter also influences the efficacy of substrate diffusion and the ecological selection
within the biofilm [10, 14, 16, 18]. In fact, the three-dimensional biofilm structure has a
physical impact on internal mass diffusivity, since it is dependent on the biofilm density
[19] and tortuosity [20]. On one hand, substrate diffusion through biofilms could be
enhanced by high turbulence which tends to produce thinner biofilms, but on the other
hand it could be reduced by a shear-compacted biofilm structure [10]. A consequence
of the hydrodynamic conditions is also the overproduction of EPS under high shear
stress [10]. Thus, the diffusivity of a substance into the biofilm would be a result of

90 Chapter 5 _________________________________________________________________________________
internal and external mass transfer effects [10]. In addition, the overproduction of EPS,
especially polysaccharides, is useful in initial cell adhesion [10, 21]. Understanding the
relationship between biofilm structure and function and also the factors that physically
shape biofilms is important to the use and control of biofilms in industrial and
biomedical fields [14, 22]. This study provides insights on macromolecular aspects of
biofilms formed under three different linear flow velocities.
5.2 MATERIALS AND METHODS
MICROORGANISM AND CULTURE CONDITIONS
The bacterium used in this work was Pseudomonas fluorescens ATCC 13525T. This
bacterium is ubiquitous in industrial settings and has a high ability to form biofilms [23].
This strain was grown at 30 ± 3 oC, pH 7, with glucose as the main carbon source. Culture
media consisted in 5 g.L-1 glucose, 2.5 g.L-1 peptone and 1.25 g.L-1 yeast extract, in
phosphate buffer (PB), pH 7, 25 mM [24]. Bacterial suspensions were prepared by gently
removing a small portion of bacteria from solid medium (agar at 10%), and diluting it in
a 1 L flask containing 250 mL of sterile nutrient medium. This bacterial suspension was
incubated overnight (16 h) with agitation (120 rpm). All medium components were
purchased from Merck (VWR, Portugal).
BIOFILM FORMATION IN A FLOW CELL SYSTEM
The flow cell system used consisted of a 3.5 L recirculating bioreactor, two vertical
perspex flow cells operating in parallel, one 0.5 L bioreactor, one peristaltic and two
centrifuge pumps (Figure 5.1). The cross-section of the flow cells is semi-circular with
diameter (d) of 2 cm. P. fluorescens was used to inoculate the smaller bioreactor
(Bioreactor I), containing the culture medium defined previously, that operated
continuously, dripping into the larger bioreactor (Bioreactor II) at a flow rate of
10 mL.h-1. This larger bioreactor was fed with a medium that consisted of 0.05 g.L-1
glucose, 0.025 g.L-1 peptone, and 0.0125 g.L-1 yeast extract in PB (pH 7, 25 mM), at a
flow rate of 0.833 L.h-1. The dilution rate applied ensured that biofilm formation
predominated over planktonic growth [25]. The flow cells were designed so that
stainless steel coupons (1 × 2 cm) could be glued into structures to be inserted in
specific slots of the flow cells. The coupons were fitted flush with the rest of the surface.

Biofilm development under different flow velocities 91 _________________________________________________________________________________
In this way, biofilm sampling was facilitated. The bacterial suspension from the larger
bioreactor was allowed to recirculate in the flow cells, in order to form biofilms on the
stainless steel (AISI 316) coupons at linear flow velocities (u) of 0.1, 0.4 and 0.8 m.s-1.
Figure 5.1 Depiction of the flow cell system used to develop biofilms at different linear flow
velocities.
The linear velocity was calculated as a function of the duct design, using the
hydraulic equivalent diameter (Dh) and the flow rate (Q):
𝐷ℎ = 4 ×𝑓𝑙𝑜𝑤 𝑎𝑟𝑒𝑎
𝑤𝑒𝑡 𝑝𝑒𝑟𝑖𝑚𝑒𝑡𝑒𝑟 (eq. 5.1)
For the semicircular duct [26]:
𝐷ℎ = 4 ×𝜋×𝑑2
8𝜋𝑑
2+𝑑
(eq. 5.2)
where d is the semicircular duct diameter. The linear flow velocity was calculated as:
24
hD
Qu
(eq. 5.3)
where u is the linear flow velocity (m.s-1), Q is the flow rate (m3.s-1) and Dh is the
hydraulic equivalent diameter (m). Biofilms were allowed to grow for 7 days to ensure

92 Chapter 5 _________________________________________________________________________________
steady-state cell density and mass [27]. Two parallel similar flow cells were used
simultaneously. Reynolds numbers were 1000 (u = 0.1 m.s-1), 4000 (u = 0.4 m.s-1) and
8000 (u = 0.8 m.s-1).
BIOFILM SAMPLING AND ANALYSIS
The biofilms of P. fluorescens were characterized in terms of mass, thickness, cell
density, total and extracellular proteins and polysaccharides.
The coupons were removed from the flow cell reactor and their thickness was
immediately assessed, using a needle connected to a digital micrometer (VS-30H,
Mitsubishi Kasei Corporation), as decribed by Teodósio et al. [28].
Afterwards, the biofilms that covered the coupons were completely scraped off,
using a sterile scalpel and resuspended in extraction buffer (EB) (2 mM Na3PO4.12H2O,
2 mM Na2HPO4.H2O, 9 mM NaCl and 1 mM KCl) to assess biofilm mass, cell density, total
and extracellular proteins and polysaccharides.
The dry biofilm mass accumulated on the slides was assessed by the determination
of the total volatile solids (TVS) of the homogenised biofilm suspensions according to
the Standard Methods (American Public Health Association [APHA], American Water
Works Association [AWWA], Water Pollution Control Federation [WPCF]), method
number 2540 A–D [29]. According to this method the TVS assessed at 550 ± 5 oC in a
furnace (Lenton thermal designs) for 2 h are equivalent to the amount of biological mass
(cells and EPS). The dry biofilm mass accumulated was expressed in terms of biofilm
mass per slide surface area (mg.cm-2).
The biofilm number of cultivable cells was assessed in terms of colony forming
units (CFU) in Plate Count Agar (Merck, Portugal), according to Ferreira et al. [30].
Biofilm extracellular proteins and polysaccharides were extracted from the cells
suspension in EB using a Dowex Marathon® resin, C sodium form, 20-50 mesh (Sigma,
Portugal), using the method described by Frølund et al. [31]. The extraction of proteins
and polysaccharides took place at 4°C for 4 h, at 400 rpm. The extracellular components
(present in the supernatant) were separated from the cells via centrifugation (3999 g,
5 min). The total (biofilm suspension before EPS extraction) and extracellular biofilm
proteins were determined using the Lowry modified method (Sigma), with bovine
serum albumin as standard. The procedure is essentially the Lowry et al. [32] method
as modified by Peterson [33]. The total (biofilm suspension before EPS extraction) and

Biofilm development under different flow velocities 93 _________________________________________________________________________________
extracellular polysaccharides were quantified through the phenol-sulphuric acid
method of Dubois et al. [34], using glucose as standard.
OUTER MEMBRANE PROTEINS EXTRACTION
The outer membrane proteins (OMP) were isolated according to the method described
by Winder et al. [35]. Sessile cells were harvested by centrifugation (3999 g, 5 min,
4 oC). The pellet was suspended in 25 mM Tris and 1 mM MgCl2 buffer (pH 7.4). The
bacterial suspension was sonicated for 2 min, 50% power (Bandelin generator with a
Microtip MS 72 probe) on ice to promote cell lysis. After sonication, the solution was
centrifuged (7000 g, 10 min, 4oC) in order to remove non-lysed cells. The supernatant
was collected and sarcosine (Sigma) was added to a final concentration of 2% (w/v), in
order to solubilize the OMP. This solution was left on ice for 20 min. The solution was
then centrifuged (13000 g, 1 h, 4 oC) to recover the OMP. The pellet containing the OMP
was resuspended in 25 mM Tris buffer (pH 7.4) and stored at -20 oC until needed.
SDS-PAGE
The biofilm OMP were separated by sodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE), as reported by Laemmli [36], using a 12% (w/v) acrylamide
gel. The proteins content of each sample was standardized to 240 ± 10 µg.ml-1 for each
sample. Electrophoresis was performed at a constant current of 170 mV. After
electrophoresis, the proteins were stained with Coomassie blue for protein profile
detection [37].
SCANNING ELECTRON MICROSCOPY
Twelve stainless steel slides covered with biofilms (four for each linear flow velocity)
were observed by scanning electron microscopy (SEM). Prior to SEM observations,
biofilm samples were fixed with 3% (w/v) glutaraldehyde in cacodylate buffer pH 7.2
[38] for 10 min and exposed to an ethanol dehydration series of 50, 60, 70, 80, 90 and
twice 100% (v/v) ethanol, followed by a chemical dehydration series of 100% ethanol +
hexamethyldisilazane (HMDS, Ted Pella, USA) at 50, 60, 70, 80, 90 and 2 × 100% (v/v)
HMDS [39], 5 min for each concentration. The coupons were then air-dried for 1 day in
a desiccator. Each coupon was sputter-coated with a palladium-gold thin film [23] using

94 Chapter 5 _________________________________________________________________________________
the SPI Module Sputter Coater equipment for, 90 s at 15 mA. The biofilms were analysed
using a SEM/EDS (FEI Quanta 400FEG ESEM/EDAX Genesis X4M) under high-vacuum
mode, at 10 kV. SEM observations were documented through the acquisition of, at
least, 20 representative microphotographs.
DETERMINATION OF NUTRIENT AND CELL LOAD
The cell and nutrient loads were calculated as the number of cells, or the glucose mass
per cross section area in a time unit. The cell number was calculated for Bioreactor II,
which contained planktonic cells. The nutrient load was considered as the glucose
content of the medium fed to bioreactor II.
DETERMINATION OF THE SHEAR STRESS
The shear stress was calculated using the dimensionless Darcy friction factor (f)
obtained from the work of Teodósio et al. [40] applied to the following equation:
𝑓 =4∙𝜏𝑤
𝜌∙𝑢2
2⁄ (eq. 5.6)
where τw is the wall shear stress (Pa), ρ is the density of water at 25 oC (Kg.m-3) and u is
the fluid velocity (m.s-1).
DETERMINATION OF THE MASS TRANSFER COEFFICIENTS
The external mass transfer coefficient, km (m.s-1) was obtained using the mathematical
model described by Moreira et al. [41]. The correlation uses the Sherwood (Sh) number
which is a function of the Reynolds (Re) and Schmidt (Sc) numbers (for 2100 < Re < 3500
and 0.6 < Sc < 3000). For tubes with a fully developed concentration profile in laminar
flow, Sh = 3.66:
𝑘𝑚 = 𝑆ℎ∙𝐷
𝐷ℎ (eq. 5.7)
where D is the molecular diffusivity of glucose (m2.s-1) , the growth-limiting nutrient in
the medium and Dh is the hydraulic diameter of the flow channel (m) [42]. For turbulent
flow in tubes:
𝑆ℎ = 0.023 ∙ 𝑅𝑒0.83 ∙ 𝑆𝑐1
3⁄ (eq. 5.8)
𝑆𝑐 = 𝜇
𝜌∙𝐷 (eq. 5.9)

Biofilm development under different flow velocities 95 _________________________________________________________________________________
𝑅𝑒 =𝐷ℎ𝑢𝜌
µ (eq. 5.10)
where µ is the viscosity of the water at 25 oC (kg.m-1.s-1).
STATISTICAL ANALYSIS
The data were analyzed using the statistical program SPSS version 20.0 (Statistical
Package for the Social Sciences). The mean and standard deviation (SD) within samples
were calculated in all cases. The experiments were replicated at least 3 times. The
statistical significance of the results was evaluated using the t-test. Statistical
calculations were based on confidence level equal to or higher than 95% (P ≤ 0.05 was
considered statistically significant).
5.3 RESULTS
The flow cell reactor was operated at three different flow velocities u = 0.1, 0.4 and
0.8 m.s-1. The biofilms developed under each different condition were characterized in
terms of thickness, dry and wet mass, cell density, matrix and total proteins and
polysaccharides (Table 5.1). The influence of flow conditions, on biofilm superficial
structure and morphology was assessed by SEM (Figure 5.3). Furthermore, the OMP
expression was analyzed by SDS-PAGE electrophoresis (Figure 5.4). The hydrodynamic
and external mass transfer coefficients were also calculated (Table 5.2).
Figure 5.2 Photographs of the stainless steel coupons with 7 days old biofilms grown at
(a) u = 0.1 m.s-1, (b) u = 0.4 m.s-1 and (c) u = 0.8 m.s-1.
Figure 5.2 demonstrates the morphological differences of biofilms developed on
the coupons induced by different flow conditions. Biofilms developed at the lowest
linear velocity (u = 0.1 m.s-1) showed an apparent partial coverage of the stainless steel
a c b

96 Chapter 5 _________________________________________________________________________________
surface (Figure 5.2a). Biofilms formed at u = 0.4 and 0.8 m.s-1 (Figures 5.2b and 5.2c)
also showed a patchy appearance, but they appeared more homogeneous than those
formed under low hydrodynamic stress.
Table 5.1 Characterization of P. fluorescens biofilms grown at different linear flow velocities.
Linear flow velocity (u) (m.s-1)
0.1 0.4 0.8
Thickness (mm)
0.213 ± 0.05 0.207 ± 0.06 0.178 ± 0.02
Biofilm mass (mg.cm-2)
Dry 0.297 ± 0.00 0.262 ± 0.04 0.269 ± 0.07
Wet 33.6 ± 6.00 30.0 ± 4.56 30.6 ± 3.94
Log cell density (CFU.gbiofilm
-1) 8.11 ± 0.68
12.2 ± 0.63
11.5 ± 0.12
Matrix (mg.gbiofilm
-1)
Proteins 104 ± 15.2 125 ± 4.23 211 ± 11.5
Polysaccharides 88.0 ± 20.1 135 ± 6.14 265 ± 9.39
Total (mg.gbiofilm
-1)
Proteins 294 ± 39.6 532 ± 42.8 471 ± 39.9
Polysaccharides 198 ± 18.6 512 ± 30.7 621 ± 15.8
The thickness of the biofilms decreased with an increase of the linear flow velocity
(Table 5.1). The thickness of the biofilms generated at the lowest velocity was different
when compared with the thickness of those formed at u = 0.4 and 0.8 m.s-1 (P < 0.05).
The biofilms wet and dry mass also differed with the flow velocity under which they
were formed. The biofilm mass values were not statistically distinct (P > 0.05).
Concerning the cell density of the biofilms, the biofilms formed at u = 0.4 m.s-1 had the
highest cell density, being followed by those formed at u = 0.8 m.s-1. Biofilms generated
at the highest flow velocities had similar cell density values (P > 0.05), and different from
those formed at 0.1 m.s-1 (P < 0.05). At
u = 0.1 m.s-1 the amounts of extracellular proteins and polysaccharides in the biofilms
were the lowest. In fact, the productivity of extracellular products increased with the
flow velocity. Also, for most of the cases the total protein and polysaccharide content
increased with increasing flow velocity, even if there was no statistical difference
between the total proteins and polysaccharides content for the biofilms formed under
the highest flow velocities (P > 0. 05).
SEM micrographs highlight the morphological aspects of the biofilms developed
under different flow regimes (Figure 5.3). These images show the presence of
extracellular structures that apparently connect the cells to each other and to the

Biofilm development under different flow velocities 97 _________________________________________________________________________________
stainless steel surface. These structures are more frequent when the flow velocity
increases. The selected SEM micrographs are intended to provide the representative
inspection of the evidences of the existence of extracellular appendages connecting
cells to each other and to the surface and are not representative of the numbers of cells
in each biofilm.
Figure 5.3 SEM micrographs of P. fluorescens biofilms developed on stainless steel surfaces at
different flow conditions: (a) u = 0.1 m.s-1, (b) u = 0.4 m.s-1 and (c) u = 0.8 m.s-1. × 15000 magnification; bar = 5 µm.
Biofilm formation under different linear flow velocities had no apparent effects on
the type of the OMP expressed (Figure 5.4). The major OMPs expressed by the biofilm
bacteria had apparent molecular weights of 32, 36, 80 and 250 (±2) kDa. However, the
results suggest that the low flow velocities induced the formation of lower quantities of
a
c b

98 Chapter 5 _________________________________________________________________________________
the major OMP. In fact, the protein with the apparent weight of 80 kDa appears to not
be present in the gel, for the lowest velocity.
Figure 5.4 OMP profiles of P. fluorescens bacteria developed in different modes of growth. Biofilms formed at (a) u = 0.1 m.s-1, (b) u = 0.4 m.s-1 and (c) u = 0.8 m.s-1.
Some hydrodynamic and mass transfer coefficients, such as the feed flow, nutrient
and cell loads, friction factor, shear stress, of the biofilms developed in the flow cell
system are presented in Table 5.2.
Table 5.2 Hydrodynamic and external mass transfer coefficients of 7 days-old P. fluorescens
biofilms grown at different flow velocities.
u / m.s-1 0.1 0.4 0.8
Re 1000 4000 8000
Feed flow L.h-1 41 174 331
Nutrient load g glucose.m -2.s-1 3.65 15.4 29.2
Cell load Log cells.m-2.s-1 13.3 13.9 14.2
Friction factor 0.063 0.037 0.033
Shear stress Pa 0.042 0.44 1.43
Sc n/a 1295 1295
Sh 3.66 255 435
km m.s-1 2.07×10-7 1.44×10-5 2.46×10-5
250 kDa
25
20
15
37
75
50
150
100
a b c

Biofilm development under different flow velocities 99 _________________________________________________________________________________
The shear stress variation inside the flow cells next to the biofilm surface is shown
in Table 5.2. The shear stress increase is more significant between the u = 0.1 m.s-1
(10.5 times lower) and 0.4 m.s-1, than between 0.4 and 0.8 m.s-1 (3.25 times higher).
The friction factor is higher in the low flow velocity biofilms (0.063), and similar
(P < 0.05) for those formed under u = 0.4 m.s-1 (0.037) and u = 0.8 m.s-1 (0.033). The
external mass transfer coefficient of biofilms developed at the lower flow velocity were
100 times lower than those calculated for the biofilms developed at the two highest
flow velocities (P < 0.05). For these the km values were statistically distinct (P < 0.05).
5.4 DISCUSSION
The objective of this work was to determine how the hydrodynamic conditions under
which biofilms were formed could influence their resistance characteristics. The
biofilms were developed in a flow cell system at three different flow velocities, 0.1, 0.4
and 0.8 m.s-1. In general, a linear velocity increase promoted a reduction of the biofilm
thickness, however, the biofilm mass was kept constant despite the flow velocity.
Therefore, a direct relationship between the increase of fluid flow velocity and the
formation of more compact and denser biofilms was observed. This is in accordance
with a previous study with Escherichia coli biofilms where the thickness of biofilms
developed in a similar flow cell system was higher at lower flow velocities [41]. Biofilms
grown at lower velocities are subjected to lower shear forces, growing faster and
forming more open structures [43]. However, low flow-stressed biofilms are also known
to have low mechanical strength, being more prone to sloughing events than those
formed under higher flow rates [40]. Other authors also stated that the flow regime has
a high impact on biofilm morphology; at lower flow rates the biofilms formed tend to
be fluffy and thicker and, in opposition, higher flow rates yield compact, dense and
smooth biofilms [44]. Verran [45] proposed that these structures with low mechanical
resistance are critical on cleaning and disinfection practices. When biofilm erosion or
sloughing occurs, bacteria are released to the bulk phase. These cells can attach to
surfaces downstream and reseed a biofilm.
The cell load obtained from the bulk fluid containing planktonic cells, and the
biofilm cell density increased with the flow velocity under which the biofilms were
formed. The results showed that more cells were available to colonize the stainless steel
for the higher flow velocities. The highest flow velocities resulted in biofilms with higher

100 Chapter 5 _________________________________________________________________________________
cell densities. This fact is exacerbated by the shear stress, imposed by higher flow
regimes, in the microorganisms, resulting in higher adhesion, and ultimately in biofilms
with a higher cell density. Studies on electron transport systems provided evidence that
the catabolic activity of biofilms can be stimulated by high shear forces, which could
lead to higher cell numbers within the biofilms [46]. Simões et al. [23] studied the effects
of hydrodynamic conditions on P. fluorescens biofilms. These authors showed that
biofilm development under turbulent conditions gave origin to biofilms with more cells
per unit area than those generated at laminar flow. That study also demonstrated an
decreased bacterial metabolic activity of the biofilms developed under higher
hydrodynamic stress conditions. They also proposed that higher flow velocities increase
the availability of nutrients in the bulk fluid, stimulating bacterial metabolism. In this
way, higher cell replication or EPS production was affordable.
The amount of matrix proteins and polysaccharides apparently increased with a
feed flow increase. Chmielewski and Frank [47] stated that the biofilm structure and
content are influenced by the flow regime, associating high turbulence with increased
EPS production. Shear stress is the predominant force acting on biofilms [15] and an
increase in linear velocity, reflected by an increase of shear stress, may influence biofilm
accumulation [44]. Vrouwenvelder et al. [44] also reported that biofilms developed
under high shear stresses are very stable against mechanical disturbances. In the
present study, three distinct flow velocities were tested, corresponding to three
different shear stress values. These hydrodynamic conditions allowed the formation of
biofilms with different thicknesses. This result is in agreement with a previous study
where higher shear stresses originated compact biofilms, characterized by low
thicknesses values and high cell densities [44].
SEM micrographs showed structures that the biofilm-embedded cells use,
apparently, to attach to each other and to the surface. Winn et al. [48] used a similar
dehydration process as that used in this study and also observed microtubular-like
structures used for microbial attachment and relevant for biofilm mechanical stability.
The hydrodynamic conditions used to form the biofilms had no significant effects
on the OMP expression of biofilm cells. The OMP of 32 and 36 kDa is similar for the
three biofilms. The protein with the apparent weight of 36 kDa could be correspondent
to the one described by Kragelund et al. [49] as being the OprF, an outer membrane
porin [50] known to be implicated in biofilm formation [51].

Biofilm development under different flow velocities 101 _________________________________________________________________________________
The external mass transfer coefficients increased with the flow velocity. A direct
consequence of increasing the flow velocity is that the transport rate of nutrients to the
biofilm surface was higher at the highest velocities [52]. The higher external mass
transfer effects observed in the biofilms developed at the two higher flow velocities
were a higher amount of cells and EPS. These results are in accordance with the findings
of Simões et al. [23]. The higher flow velocities are correlated with higher shear stress
imposed to the biofilm that is related by thinner biofilms. In spite of having more cells
and EPS, they also have less limitations to mass transfer by being thinner, than the
thicker biofilm (u = 0.1 m.s-1) that have adapted its structure to be fluffier in order to
facilitate the access to nutrients [13, 23]. Due to the higher shear stress they rather
produce EPS, as seen on the results (u = 0.8 m.s-1), than new cells to increase its
cohesion and withstand the forces of the passing fluid [41].
5.5 CONCLUSIONS
In this study several characteristics of P. fluorescens biofilms were studied when formed
at three distinct linear flow velocities. The biofilms developed at the two highest linear
velocities were thinner, and both had approximately the same mass as that formed at
the lowest velocity. The cell density and EPS content was also superior for the biofilms
generated at the highest velocities. These features make these biofilms denser. The
external mass transfer coefficient increased with the flow velocity. In general, biofilms
formed under higher flow velocities were more complex, including the presence of
attributes that can contribute to their antimicrobial resistance (higher cell density and
EPS content) in a higher extent than those formed under lower flow velocities.
REFERENCES
[1] Gibson H, Taylor J, Hall K, Holah J. Effectiveness of cleaning techniques used in the food industry in terms of the removal of bacterial biofilms. Journal of Applied Microbiology, 1999. 87(1):41-48.
[2] Simões M, Simões L, Vieira M. A review of current and emergent biofilm control strategies. LWT - Food Science and Technology, 2010. 43(4):573-583.
[3] Van Houdt R, Michiels CW. Biofilm formation and the food industry, a focus on the bacterial outer surface. Journal of Applied Microbiology, 2010. 109(4):1117-1131.
[4] Jahid IK, Ha SD. A review of microbial biofilms of produce: Future challenge to food safety. Food Science and Biotechnology, 2012. 21(2):299-316.

102 Chapter 5 _________________________________________________________________________________
[5] Gomes LC, Moreira JMR, Miranda JM, Simões M, Melo LF, Mergulhão FJ. Macroscale versus microscale methods for physiological analysis of biofilms formed in 96-well microtiter plates. Journal of Microbiological Methods, 2013. 95(3):342-349.
[6] Andersson S, Dalhammar G, Land C, Kuttuva Rajarao G. Characterization of extracellular polymeric substances from denitrifying organism Comamonas denitrificans. Applied Microbiology and Biotechnology, 2009. 82(3):535-543.
[7] Flemming H. The EPS matrix: The "house of biofilm cells". Journal of Bacteriology, 2007. 189(22):7945-7947.
[8] Tolker-Nielsen T, Molin S. Spatial Organization of Microbial Biofilm Communities. Microbial Ecology, 2000. 40(2):75-84.
[9] Gao BY, Zhu XB, Xu CH, Yue QY, Li WW, Wei JC. Influence of extracellular polymeric substances on microbial activity and cell hydrophobicity in biofilms. Journal of Chemical Technology and Biotechnology, 2008. 83(3):227-232.
[10] Liu Y, Tay J-H. The essential role of hydrodynamic shear force in the formation of biofilm and granular sludge. Water Research, 2002. 36(7):1653-1665.
[11] Sutherland I. The biofilm matrix - an immobilized but dynamic microbial environment. Trends in Microbiology, 2001. 9(5):222-227.
[12] Araújo PA, Lemos M, Mergulhão F, Melo L, Simões M, Antimicrobial resistance in biofilms to disinfectants, in Science against microbial pathogens: communicating current research and technological advances, A. Méndez-Vilas, Editor. 2011, Formatex: Badajoz, Spain. p. 826-834.
[13] Melo LF, Vieira MJ. Physical stability and biological activity of biofilms under turbulent flow and low substrate concentration. Bioprocess Engineering, 1999. 20(4):363-368.
[14] Stoodley P, Boyle JD, DeBeer D, Lappin-Scotta HM. Evolving perspectives of biofilm structure. Biofouling, 1999. 14(1):75-90.
[15] Stewart PS. Mini-review: Convection around biofilms. Biofouling, 2012. 28(2):187-198.
[16] Wäsche S, Horn H, Hempel DC. Influence of growth conditions on biofilm development and mass transfer at the bulk/biofilm interface. Water Research, 2002. 36(19):4775-4784.
[17] Vieira MJ, Melo LF, Pinheiro MM. Biofilm formation: Hydrodynamic effects on internal diffusion and structure. Biofouling, 1993. 7(1):67-80.
[18] Stewart P. Diffusion in biofilms. Journal of Bacteriology, 2003. 185(5):1485-1491.
[19] Sandt C, Barbeau J, Gagnon MA, Lafleur M. Role of the ammonium group in the diffusion of quaternary ammonium compounds in Streptococcus mutans biofilms. Journal of Antimicrobial Chemotherapy, 2007. 60(6):1281-1287.
[20] Melo L. Biofilm physical structure, internal diffusivity and tortuosity. Water Science and Technology, 2005. 52(7):77-84.
[21] Lopes FA, Vieira MJ, Melo L. Chemical composition and activity of a biofilm during the start-up of an airlift reactor. Water Science and Technology, 2000. 41(4-5):105-111.
[22] Cunningham AB, Characklis WG, Abedeen F, Crawford D. Influence of biofilm accumulation on porous-media hydrodynamics. Environmental Science & Technology, 1991. 25(7):1305-1311.
[23] Simões M, Pereira M, Sillankorva S, Azeredo J, Vieira M. The effect of hydrodynamic conditions on the phenotype of Pseudomonas fluorescens biofilms. Biofouling, 2007. 23(3-4):249-58.
[24] Simões M, Cleto S, Pereira M, Vieira M. Influence of biofilm composition on the resistance to detachment. Water science and technology, 2007. 55:473-480.
[25] Simões M, Carvalho H, Pereira MO, Vieira MJ. Studies on the behaviour of Pseudomonas fluorescens biofilms after ortho-phthalaldehyde treatment. Biofouling, 2003. 19(3):151-157.

Biofilm development under different flow velocities 103 _________________________________________________________________________________
[26] Tosun I, Uner D, Ozgen C. Critical Reynolds number for Newtonian flow in rectangular ducts. Industrial & Engineering Chemistry Research, 1988. 27(10):1955-1957.
[27] Pereira M, Kuehn M, Wuertz S, Neu T, Melo L. Effect of low regime on the Architecture of a Pseudomonas fluorescens biofilm. Biotechnology and Bioengineering 2002. 78:164-171.
[28] Teodósio J, Simões M, Melo L, Mergulhao F. Flow cell hydrodynamics and their effects on E-coli biofilm formation under different nutrient conditions and turbulent flow. Biofouling, 2011. 27(1):1-11.
[29] APHA/AWWA/WPCF. Standard methods for the examination of water and wastewater, 17th edition, Clesceri, LS, Greenberg, AE and Trussel, RR, (eds). American Public Health Association, Washington DC, USA. 1989.
[30] Ferreira C, Pereira AM, Pereira MC, Simões M, Melo LF. Biofilm control with new microparticles with immobilized biocide. Heat Transfer Engineering, 2013. 34(8-9):712-718.
[31] Frølund B, Palmgren R, Keiding K, Nielsen P. Extraction of extracellular polymers from activated sludge using a cation exchange resin. Water Research, 1996. 30(8):1749-1758.
[32] Lowry O, Rosebrough N, Farr A, Randall R. Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry, 1951. 193(1):265-275.
[33] Peterson G. Review of the Folin phenol protein quantitation method of Lowry, Rosebrough, Farr and Randall. Analytical Biochemistry 1979. 100(2):201-220.
[34] Dubois M, Gilles K, Hamilton J, Rebers P, Smith F. Colorimetric method for determination of sugars and related substances. Analitical Chemistry, 1956. 28(3):350-356.
[35] Winder CL, Al-Adham ISI, Abdel Malek SMA, Buultjens TEJ, Horrocks AJ, Collier PJ. Outer membrane protein shifts in biocide-resistant Pseudomonas aeruginosa PAO1. Journal of Applied Microbiology, 2000. 89(2):289-295.
[36] Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 1970. 227(5259):680-5.
[37] Simões M, Pereira M, Machado I, Simões L, Vieira M. Comparative antibacterial potential of selected aldehyde-based biocides and surfactants against planktonic Pseudomonas fluorescens. Journal of Industrial Microbiology & Biotechnology, 2006. 33(9):741-749.
[38] Manuel C. Biofilm Dynamics and drinking water stability: Effects of hydrodynamics and surface materials, in Department of Chemical Engineering. 2007, University of Porto: Faculty of Engineering.
[39] Thorn RMS, Greenman J. A novel in vitro flat-bed perfusion biofilm model for determining the potential antimicrobial efficacy of topical wound treatments. Journal of Applied Microbiology, 2009. 107(6):2070-2079.
[40] Teodósio JS, Silva FC, Moreira JMR, Simões M, Melo LF, Alves MA, Mergulhão FJ. Flow cells as quasi-ideal systems for biofouling simulation of industrial piping systems. Biofouling, 2013. 29(8):953-966.
[41] Moreira JMR, Teodósio JS, Silva FC, Simões M, Melo LF, Mergulhão FJ. Influence of flow rate variation on the development of Escherichia coli biofilms. Bioprocess and Biosystems Engineering, 2013. 36(11):1787-1796.
[42] Perry RH, Green DW, Maloney JO. Perry's chemical engineers' handbook. 7th ed. New York: McGraw-Hill. 1999.
[43] Vieira MJ, Melo LF. Intrinsic kinetics of biofilms formed under turbulent flow and low substrate concentrations. Bioprocess and Biosystems Engineering, 1999. 20(4):369-375.
[44] Vrouwenvelder JS, Buiter J, Riviere M, van der Meer WGJ, van Loosdrecht MCM, Kruithof JC. Impact of flow regime on pressure drop increase and biomass accumulation and morphology in membrane systems. Water Research, 2010. 44(3):689-702.

104 Chapter 5 _________________________________________________________________________________
[45] Verran J. Biofouling in food processing: biofilm or biotransfer potential? Food and Bioproducts Processing, 2002. 80(4):292-298.
[46] Liu Y, Tay J-H. Detachment forces and their influence on the structure and metabolic behaviour of biofilms. World Journal of Microbiology and Biotechnology, 2001. 17(2):111-117.
[47] Chmielewski RAN, Frank JF. Biofilm formation and control in food processing facilities. Comprehensive Reviews in Food Science and Food Safety, 2003. 2(1):22-32.
[48] Winn M, Casey E, Habimana O, Murphy CD. Characteristics of Streptomyces griseus biofilms in continuous flow tubular reactors. FEMS Microbiology Letters, 2014. in press.
[49] Kragelund L, Leopold K, Nybroe O. Outer membrane protein heterogeneity within Pseudomonas fluorescens and P. Putida and use of an Oprf antibody as a probe for rRNA homology group I Pseudomonads. Applied and Environmental Microbiology, 1996. 62(2):480-485.
[50] Winsor GL LD, Fleming L, Lo R, Whiteside MD, Yu NY, Hancock RE, Brinkman FS (2011). Pseudomonas Genome Database: improved comparative analysis and population genomics capability for Pseudomonas genomes. Nucleic Acids Res. 2011 Jan; 39 (Database issue): D596-600. Pubmed.
[51] Seyer D, Cosette P, Siroy A, Dé E, Lenz C, Vaudry H, Coquet L, Jouenne T. Proteomic comparison of outer membrane protein patterns of sessile and planktonic Pseudomonas aeruginosa cells. Biofilms, 2005. 2(01):27-36.
[52] Beyenal H, Lewandowski Z. Internal and external mass transfer in biofilms grown at various flow velocities. Biotechnology Progress, 2002. 18(1):55-61.

CHAPTER 6 HALOGEN-BASED COMPOUNDS IN BIOFILM CONTROL
This chapter was submitted as:
Malheiro J, Araújo PA, Machado I, Mergulhão F, Melo L, Simões M. The effects of selected halogen-
containing chemicals on Pseudomonas fluorescens planktonic cells and flow-generated biofilms.
ww
w.p
erio
dic
table
.co
m

106 Chapter 6
_________________________________________________________________________________
ABSTRACT
Microbial biofilms are ubiquitous in nature and inherently resistant to an increasing
range of antimicrobial agents. One of the most widely used disinfectants is sodium
hypochlorite (SH), however, despite its proven efficacy, SH is surface corrosive, presents
several health concerns, and resistance phenomena is already emerging. Therefore, the
development of innovative biofilm control strategies are needed. Little is known about
the usefulness of brominated products as food industry disinfectants. In this study, the
control of Pseudomonas fluorescens biofilms was assessed with the halogen-based
chemicals: cetyltrimethylammonium bromide (CTAB), sodium hypochlorite (SH),
3-bromopropionyl chloride (BrCl) and 3-bromopropionic acid (BrOH). The influence of
these chemicals was assessed on several physiological aspects of planktonic cells,
particularly their antimicrobial action, influence on cell surface properties and
potassium release. While CTAB had the highest antimicrobial activity, BrOH had the
lowest. All the chemicals promoted cellular disruption, with apparent pore formation in
the cell membranes and consequent leakage of essential intracellular constituents. Only
CTAB, BrCl and BrOH led to irreversible changes in membrane properties (charge and
physicochemical properties) through hydrophobicity changes and decrease of negative
surface charge. When these chemicals were applied to biofilms, no significant killing or
removal was achieved (maximum killing of 1 log and 15% removal). Moreover, the
chemicals allowed the biofilms to regrow after exposure. In fact, the overall results
demonstrated similar effects with all selected chemicals. The overall data demonstrate
that both BrCl and CTAB are advantageous alternatives to the currently used
disinfectant, SH, since they present comparable efficiency with potentially less health
and surface equipment damage concerns.

Halogen- containing chemicals 107 _________________________________________________________________________________
6.1. INTRODUCTION
The World Health Organization (WHO) refers to food safety as one of the top priorities
and challenges of the century [1]. Nowadays, foodborne diseases are a prime public
health concern in developing and developed countries. WHO reported 1.8 million
mortality cases of diarrheal diseases worldwide. In the United States of America it is
estimated that every year, about 48 million people suffer from foodborne diseases
[1-3]. The Centers for Disease Control and Prevention (CDC) and the US National Health
Institute (NIH), documented that biofilms are involved in over 65% of all microbial
diseases. In addition, foodborne pathogens can form biofilms in produce, which makes
them resistant to commonly used disinfectants [1, 4]. Consequently, the formation of
biofilms has severe implications in several areas, from industrial processes to health-
related fields, with huge economic losses [5, 6].
The sanitizers and biocides used in industry do not control microorganisms in
biofilms as they are typically 10-1000 times more resistant than their planktonic
counterparts [7, 8]. The efficacy of a disinfectant depends on several factors, such as
the type of target microorganism and its susceptibility, the adhesion surface,
temperature, exposure time, concentration and pH [9]. Also, antimicrobial resistance
occurs as a multifactorial aspect that includes slow or incomplete penetration of the
biocide into the biofilm, physiological alterations of the biofilm cells, expression of stress
response with adaptive molecules, or even, differentiation as persister cells [10-12].
Biofilm prevention and control is, therefore, a priority in food industry, prompting a
need to search for new biocides and/or sanitizers and to understand their potential to
prevent and control biofilms.
Food contact surfaces are normally disinfected and cleaned with agents containing
peroxides, chloramines or hypochlorite [5]. The free chlorine obtained by the use of
hypochlorite can be very aggressive to stainless steel, interfering with its surface, and
might facilitate further bacterial adhesion and biofilm formation [5, 13]. Furthermore,
chlorine is the most widely used disinfectant in industry, however, there is a possibility
that during disinfection it reacts with natural organic matter or contaminants in surface
waters, and it can also produce a complex mixture of disinfection by-products which
already demonstrated carcinogenic, mutagenic and teratogenic (abnormalities of
physiological development ) activity in animal studies [1, 14].

108 Chapter 6
_________________________________________________________________________________
Quaternary ammonium compounds (QACs), cationic compounds with a basic
structure (NH4+) and a strong antimicrobial potential, are frequently used for
disinfection and sanitation in a wide range of fields, such as hospitals and food
manufacturing [15, 16]. Cetyltrimethylammonium bromide (CTAB) is a relatively safe
and inexpensive product [17]. Nevertheless, it has been shown that its increasing use in
a wide range of applications contributed to the emergence of resistant bacteria and,
occasionally, multidrug resistance [17, 18]. Comparatively to chlorine, QACs are more
expensive, however, they are an attractive alternative as they are less affected by the
presence of organic matter, are not corrosive at low concentrations, are more stable,
and could be stored for longer periods of time without compromising their
antimicrobial activity [19].
The formation of biofilms is a microbial community behavior coordinated through
cell-to-cell communication mediated by small, diffusible signals, a phenomenon called
quorum sensing. Several phenotypes regulated by cell-to-cell communication are
implicated in bacterial colonization and virulence [20]. Therefore, eukaryotes have
developed a defense mechanism based on chemicals, including secondary metabolites
that inhibit these phenotypes [20-22]. For example, furanones produced by the marine
algae Delisea pulchra [23], oxidize halogen compounds produced by Laminaria digitata
and, haloperoxidases produced by seaweeds are responsible for the production of the
microbicidal compounds hypobromous acid (BrOH) and hypochlorous acid (ClOH) [22].
The natural furanones are halogenated at several positions by bromine, iodide or
chloride and, as observed in field experiments, the concentration of furanones is
inversely correlated with the degree of bacterial colonization [20]. Stabilized halogen
antimicrobials are extensively used to control biofouling in industry and they have been
shown to be more effective in penetrating and disinfecting biofilms than free halogen
[22]. Considering this assumption, and the fact that 3-bromopropionic acid (BrOH) is
used to synthetize several compounds with antimicrobial properties [24, 25], and that
3-bromopropionyl chloride (BrCl) has a comparable structure, these two halogenated
compounds were selected for this study to be assessed on their antimicrobial properties
against planktonic cells and biofilms of Pseudomonas fluorescens (Figure 6.1). Little is
known about the utility of bromine as a disinfectant for food industry. Previous studies
demonstrated that dibromodimethyl hydrantoin was as effective as chlorine against
Streptococcus faecalis [26], however, less effective against Bacillus cereus spores [27].
Similarly to free chlorine, there are safety concerns about the production of brominated

Halogen- containing chemicals 109 _________________________________________________________________________________
organic compounds and their impact on human and environmental safety [28]. Because
chlorine is widely used in industry as a disinfectant and sanitizer, it was used for
comparison purposes.
The goal of this study was to assess the antimicrobial action of selected halogen-
based chemicals against planktonic cells and biofilms of Pseudomonas fluorescens. This
bacterium is a major contaminant in food industry, causing produce spoilage and
foodborne illnesses [29, 30].
6.2. MATERIALS AND METHODS
ANTIMICROBIAL AGENTS
Sodium hypochlorite solution (SH) and cetyltrimethylammonium bromide (CTAB) were
purchased from Sigma (Portugal), 3-bromopropionic acid (BrOH) was purchased from
Merck (VWR, Portugal), 3-bromopropionyl chloride (BrCl) was purchased from Alfa
Aesar (VWR, Portugal). All dilutions were performed using sterile distilled water.
Figure 6.1 Chemical structures of the chemicals used: (a) cetyltrimethylammonium bromide (CTAB), (b) sodium hypochlorite (SH), (c) 3-bromopropionyl chloride (BrCl) and (d) 3-bromopropionic acid (BrOH).
MICROORGANISMS AND CULTURE CONDITIONS
The bacterium used in this study was Pseudomonas fluorescens ATCC 13525. Bacterial
growth was obtained from overnight cultures (16 h) in culture medium (5 g.L-1 glucose,
2.5 g.L-1 peptone and 1.25 g.L-1 yeast extract in 0.025 M phosphate buffer, pH 7) and
incubated at 30 ± 3°C, and 150 rpm of agitation [31].

110 Chapter 6
_________________________________________________________________________________
ANTIBACTERIAL SUSCEPTIBILITY TESTS
The minimum inhibitory concentration (MIC) of each agent was determined by the
microdilution method according to the Clinical and Laboratory Standards Institute (CLSI)
guidelines [32] using 96-well microtiter plates. Bacteria at a density of 109 colony
forming units (CFU) per ml were inoculated into fresh culture medium. A volume of
200 µl was inserted in each well, along with the different concentrations of the
chemicals (10% v/v). The bacterial growth was determined at 600 nm using a microplate
reader (Spectramax M2e, Molecular Devices, Inc.). The MIC was determined as the
lowest concentration at which microbial growth was inhibited [33]. The cell suspension
was plated in Plate Count Agar (PCA, Merck, Germany) and incubated overnight at
30 ± 3°C, after a neutralization step to quench the chemicals antimicrobial activity, by
dilution, to sub-inhibitory concentrations [34]. The minimum bactericidal concentration
(MBC) was considered the lowest concentration of the antimicrobial agent where no
growth was detected on the solid medium [33].
PHYSICOCHEMICAL CHARACTERIZATION OF BACTERIAL SURFACES
The physicochemical properties of P. fluorescens cell surface were assessed by the
sessile drop contact angle measurement on bacteria lawns, performed as described by
Busscher et al. [35]. Contact angles were determined using an OCA 15 Plus
(DATAPHYSICS) video-based optical measuring instrument, allowing image acquisition
and data analysis. The measurements (≥ 15 per liquid and chemical) were performed
according to Simões et al. [36], after bacterium incubation (1 h) with the chemical at
the MBC. The liquid surface tension components reference values were obtained from
the literature [37]. Hydrophobicity was assessed after contact angle measurement,
following the van Oss method [38-40], where the degree of hydrophobicity of a given
surface (s) is expressed as the free energy of interaction between two entities of that
surface, when immersed in water (w)−(∆Gsws mJ. m−2). The surface is considered
hydrophobic if the interaction between two entities is stronger than the interaction of
each with water ∆𝐺𝑠𝑤𝑠 < 0. Otherwise, if ∆Gsws > 0, the material is considered
hydrophilic. ∆Gsws can be calculated using the surface tension components of the
interacting entities of equation 6.1:

Halogen- containing chemicals 111 _________________________________________________________________________________
∆Gsws = − 2 (√γsLW − √γw
LW)2
+ 4(√γs+γw
− + √γs−γw
+ − √γs+γs
− − √γw+ γw
− ) (eq. 6.1)
where γLW, represents the Lifshitz-van der Waals component of the surface free energy
and γ+ and γ− are the electron acceptor and donor parameters, respectively, of the
Lewis acid-based component (γAB), where γAB = 2√γ+γ−. The surface tension
components, of a solid material, can be obtained by measuring the contact angles of
three liquids with different polarities and known surface tension components
(1): α-bromonaphtalene (apolar), formamide (polar), and water (polar). Upon obtaining
the data, three equations of the type below can be solved:
(1 + cos θ)γLTot = 2 (√γS
LWγLLW + √γS
+γL− + √γS
−γL+) (eq. 6.2)
where θ is the contact angle. The total surface energy is calculated as γTot = γLW + γAB.
BACTERIAL SURFACE CHARGE
The zeta potential of bacterial suspensions was determined in sterile water using a Nano
Zetasizer (Malvern Instruments). This determination was performed before and after
1 h bacterial exposure to the chemicals at the corresponding MBC.
POTASSIUM (K+) LEAKAGE
The quantification of K+ in bacterial solutions, before and after 1 h exposure to the MBC
of each biocide was determined by flame emission and atomic absorption spectroscopy.
Samples were filtrated (Whatman, pore size 0.2 µm) and analyzed in a GBC AAS 932
plus device using GBC Avante 1.33 software.
OUTER MEMBRANE PROTEIN EXTRACTION AND ANALYSIS
Outer membrane proteins (OMP) were isolated based on the method described by
Winder et al. [41]. Briefly, an overnight inoculum of P. fluorescens was washed with
8.5% NaCl solution, diluted to approximately 109 CFU.ml-1 and incubated with each
chemical at the MBC, for 1 h at 30 ± 3°C and 150 rpm of agitation. All suspensions were
then harvested by centrifugation (3202 g, 25 min) and ressuspended twice with Tris-
HCl 25 mM, pH 7.4 with 1 mM MgCl2. Then, the suspension was sonicated for 2 min,
50% power (Bandelin generator with a Microtip MS 72 probe) on ice, to promote cell

112 Chapter 6
_________________________________________________________________________________
lysis. Next, the solution was centrifuged (7000 g, 10 min, 4°C) to discard cell debris.
Sarcosine (Sigma, final concentration of 2%) was added to the supernatant and
incubated for 20 min at 4 °C, to solubilize the OMPs. The solution was centrifuged
(13000 g, 4 °C, 1h), to recover the OMP that were ressuspended in Tris-HCl pH 7.4 [42].
The concentration of proteins was determined by Bicinchoninic Acid Protein Assay Kit
(BCA) (BCA - PIERCE Cat. No. 23225) and standardized to 240 ± 10 µg.ml-1 in each sample
and applied to a sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE)
with 12% bisacrylamide [43, 44]. The proteins were stained with Coomassie blue [45].
Electrophoresis was accomplished at constant 170 V. All electrophoresis components
were purchased from BioRad (Portugal).
ASSESSMENT OF QUORUM SENSING INHIBITION
Quorum sensing inhibition was determined by the disc diffusion assay. The inoculum of
Chromobacterium violaceum (ATCC 12472) was grown overnight (approximately 16 h)
in Luria-Bertani broth (LB), Liofilchem, Italy (30 ± 3°C, 150 rpm). The MIC and MBC for
all chemicals were determined with the antibacterial susceptibility tests as described
before, with minor modifications. LB broth was used and C. violaceum growth was
determined at 620 nm. Standard disc diffusion assay was performed for all chemicals at
the MBC. Briefly, the bacterial suspension (approximately 108 CFU.ml-1), was seeded on
LB agar plates, using a sterilized swab. Next, sterile paper discs (6 mm diameter) were
placed over the LB agar plates and 15 µl of each biocide was added. Antimicrobial and
quorum sensing inhibition (halo of colorless but viable cells) halos were measured after
24 h of incubation at 30 ± 3°C [46, 47].
COLONY BIOFILM FORMATION AND PENETRATION TESTS
These tests were performed as explained in chapter 3, in the corresponding sub-section
of material and methods.
BIOFILM FORMATION IN A FLOW CELL SYSTEM
Biofilm formation in the flow cell system was executed as explained in chapter 5, in the
corresponding sub-section of material and methods.

Halogen- containing chemicals 113 _________________________________________________________________________________
BIOFILM CONTROL USING DIFFERENT CHEMICALS
The flow-generated biofilms of P. fluorescens were submitted to a disinfection process
with the selected chemicals (CTAB, BrCl, SH) at their MBC. The flow cell was carefully
emptied and the disinfection of the biofilms was made by recirculation of the chemical
at the MBC, with a flow rate of 3.4 L.h-1, for 1 h. After that period, the initial conditions
were restored in the flow cell system. Control experiments with phosphate buffer were
also performed. Four coupons, two from each flow cell were removed at different time
periods: before chemical exposure, immediately after the antimicrobial exposure and
2, 12 and 24 h post-antimicrobial treatment. After biofilm chemical exposure, a
neutralization step by diluting to sub-inhibitory concentrations was performed,
according to Johnston et al. [34].
BIOFILM ANALYSIS
The P. fluorescens biofilms were characterized in terms of organic mass and cell density.
The stainless steel coupons were removed from the flow cell and the biofilms that
covered the coupons surface were completely scraped using a sterile scalpel and
resuspended in 10 mL of phosphate buffer. The suspensions were vortexed (IKA TTS2)
for 30 s at 100% input. The biofilm mass was determined according to the standard
methods (American Public Health Association [APHA], American Water Works
Association [AWWA], Water Pollution Control Federation [WPCF]) [52]. The biofilm cell
densities were assessed in terms of CFUs in Plate Count Agar (Merck, Portugal) [53].
STATISTICAL ANALYSIS
Data were analyzed applying the parametric paired t-test using the statistical program
SPSS version 22.0 (Statistical Package for the Social Sciences). The average and standard
deviation (SD) within samples were calculated for all cases. At least three independent
experiments were performed for each condition tested. Statistical calculations were
based on a confidence level ≥ 95% (P < 0.05) which was considered statistically
significant.

114 Chapter 6
_________________________________________________________________________________
6.3. RESULTS
The MIC and MBC values obtained for the tested chemicals against P. fluorescens are
presented in Table 6.1. Overall, the MBC was higher than the MIC, with the exception
of SH which a MIC and MBC were 500 µg.ml-1. CTAB was the most efficient antimicrobial
with a MBC 10 to 20 times lower than the other chemicals.
Table 6.1 Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) values of each chemical tested.
MIC / µg.mL-1 MBC / µg.mL-1
BrCl 650 700
BrOH 850 900
CTAB 20 50
SH 500 500
The parameters of the bacterial surface tension before and after the treatment
with each chemical were determined to ascertain the effects of the selected chemical
on the bacterial surface properties (Table 6.2).
Table 6.2 Surface tension parameters, hydrophobicity (∆𝐺𝑠𝑤𝑠
), apolar (𝛾𝑠𝐿𝑊) and polar (𝛾𝑠
𝐴𝐵), of untreated P. fluorescens (control) and after 1 h treatment with the chemicals (BrCl, BrOH, CTAB or SH). The average ± SD is presented.
Surface tension parameters / mJ.m-2 ∆𝑮𝒔𝒘𝒔
/ mJ.m-2 𝜸𝒔
𝑳𝑾 𝜸𝒔𝑨𝑩 𝜸𝒔
+ 𝜸𝒔−
Control 22.8 ± 4.43 30.3 ± 4.79 4.14 ± 1.25 57.0 ± 4.71 30.7 ± 6.30
BrCl 19.4 ± 0.43 31.4 ± 2.92 4.40 ± 1.07 57.0 ± 3.96 29.8 ± 5.50
BrOH 20.3 ± 0.80 34.1 ± 3.61 6.10 ± 1.25 53.0 ± 3.51 23.4 ± 4.90
CTAB 12.0 ± 1.35 47.0 ± 7.10 10.4 ± 2.98 54.0 ± 0.81 14.0 ± 5.00
SH 29.4 ± 4.68 13.3 ± 1.83 0.89 ± 0.33 51.0 ± 6.00 33.5 ± 8.70
P. fluorescens is naturally hydrophilic (∆Gsws > 0 mJ.m-2), however, this property
was less pronounced when the cells were in contact with BrOH and CTAB (P < 0.05).
Regarding the apolar parameter (γsLW), only CTAB promoted a small decrease of the
apolar component compared to the untreated cells. The polar parameter (γsAB) of the
bacterium increased with the application of CTAB and decreased in the presence of SH

Halogen- containing chemicals 115 _________________________________________________________________________________
(P < 0.05). Moreover, when the capacity to accept (γs+) or donate (γs
−) electrons was
analyzed, it was possible to observe that the treatment with SH significantly decreased
the surface capacity of the cell to accept or donate electrons (P < 0.05), while BrOH and
CTAB increased the electron acceptor component of P. fluorescens surface (P < 0.05).
P. fluorescens untreated cells had a negative surface charge of -13.53 mV with a
conductivity of 0.05 mS.cm-1 (Table 6.3). The exposure to CTAB, BrCl or BrOH modified
P. fluorescens surface charge to less negative and increased its conductivity (P < 0.05),
with the exception of CTAB that had no effects on the cell surface conductivity
(P > 0.05). Conversely, SH enhanced conductivity (P < 0.05) without interfering with the
cell surface charge (P > 0.05).
Table 6.3 Zeta potential and conductivity of P. fluorescens before and after 1 h treatment with
different chemicals. The average ± SD is presented.
Zeta Potential / mV Conductivity / mS.cm-1
Control -13.5 ± 2.32 0.05 ± 0.02
BrCl -2.88 ± 0.66 2.25 ± 0.06
BrOH -4.96 ± 0.95 0.46 ± 0.09
CTAB -8.14 ± 0.42 0.05 ± 0.01
SH -13.0 ± 1.41 31.1 ± 0.14
To ascertain the effects of the chemicals in the cell integrity the intracellular K+
release was assessed. Table 6.4 shows the K+ concentration with and without exposure
to the chemicals. All chemicals tested promoted an alteration in the cytoplasmic
membrane permeability, causing K+ release, regardless the chemical used (P < 0.05).
Table 6.4 Concentration of K+ in solution of the untreated and after 1 h incubation of P. fluorescens with each chemical. The average ± SD is presented.
Concentration of K+ in solution / µg.ml-1
Control 1.21 ± 0.08
BrCl 1.99 ± 0.21
BrOH 1.96 ± 0.25
CTAB 2.09 ± 0.28
SH 2.07 ± 0.26
The OMP expression, using 1-D SDS-PAGE, was assessed before and after biocide
exposure for 1 h (Figure 6.2). No significant differences were found in the expression of

116 Chapter 6
_________________________________________________________________________________
the major OMP of P. fluorescens with and without the exposure to the selected
chemicals, with the exception that CTAB and SH reduced significantly the amount of
OMP expressed (Figure 6.2).
Figure 6.2 OMP profile of P. fluorescens cells when exposed to the MBC of different chemicals.
The molecular weight marker (a) was used to extrapolate the molecular weight of some lanes of the OMP profile obtained from incubation in the (b) absence or in the presence of (c) BrCl, (d) BrOH, (e) CTAB and (f) SH.
The percentage of retardation gives an estimate on the efficacy of chemical
products to cross the biofilm (Table 6.5). In this study, the penetration of BrCl was the
most efficient followed closely by SH, with 0 and 1.90% retardation respectively. The
biofilm penetration was retarded by 15% for BrOH and 100 % for CTAB (P < 0.05).
Table 6.5 Retardation caused by P. fluorescens biofilms, for each chemical used. Data is presented as average ± SD of the percentage of diameter measurements for halo readings compared with controls (no biofilm).
Retardation / %
BrCl 0.00 ± 0.00
BrOH 15.7 ± 4.40
CTAB 100 ± 0.00
SH 1.90 ± 3.20

Halogen- containing chemicals 117 _________________________________________________________________________________
The disc diffusion assay for the detection of quorum sensing inhibition is depicted
in Figure 6.3. Quorum sensing was not affected by the chemicals tested.
Figure 6.3 Disc diffusion assays for the detection of quorum sensing inhibition of C. violaceum by
(a) BrOH, (b) BrCl, (c) SH and (d) CTAB.
BrCl was selected over BrOH for the biofilm assay, due to the lower MIC and MBC,
higher capacity to change the surface properties of P. fluorescens and ability to
penetrate the biofilm without being retarded. Therefore, BrCl, CTAB and SH were tested
against 7-day old flow-generated biofilms, formed under conditions mimicking those
found in industry, chapter 5. The effectiveness of the chemicals was assessed in terms
of number of biofilm CFU (Figure 6.4a) and mass (Figure 6.4b).The results obtained for
the number of biofilm CFU revealed a reduction after 1 h exposure to the MBC of CTAB
and SH (Figure 6.4a). However, this effect was more pronounced for SH with 1-log
reduction (P > 0.05). For CTAB a CFU log reduction of 0.4 was achieved and for BrCl the
difference was almost negligible. In order to ascertain the role of the chemicals tested
on biofilm regrowth, the CFU were determined during the 2, 12 and 24 h after chemical
exposure. Two hours after the treatment the number of CFU increased for all chemicals
tested, attaining similar values to those before the treatment (P > 0.05, Figure 6.4a).

118 Chapter 6
_________________________________________________________________________________
The number of biofilm CFU, remained constant overtime for all the conditions tested,
except for the 24 h BrCl-treated biofilms. In this case the number of CFU increased
significantly (P < 0.05, Figure 6.4a) when compared to the control values (untreated
biofilms). In terms of biofilm mass, the three chemicals promoted similar biomass
removal (16%, Figure 6.4b). These values remained unchanged 2 h after the treatment
(P > 0.05). When analyzing the biofilm, 12 h after the treatment, no significant biomass
changes were found for the biofilms treated with CTAB and BrCl, in comparison to the
biofilms immediately after exposure. The SH treated biofilms recovered significantly in
terms of biomass (P < 0.05). However, 24 h after the treatment, the values obtained for
the biomass, were as low as the values achieved with the SH treatment after the same
time. During the recovery period, the biomass of CTAB-treated biofilms was similar to
the value immediately after the treatment, while significant biomass regrowth of the
BrCl-treated biofilms was found (P > 0.05).
Figure 6.4 P. fluorescens biofilm log CFU.cm-2 (a) and mass (b) before and after treatment with
CTAB ( ), BrCl ( ) and SH ( ). Samples were collected before treatment ( ), immediately after 1 h treatment and after 2, 12 and 24 h after chemical removal. Values are average ± SD.
4,0
4,5
5,0
5,5
6,0
6,5
7,0
BeforeTreatment
Imediatly after 2 hours 12 hours 24 hours
Log
CFU
.cm
-2
0,0
0,5
1,0
1,5
2,0
2,5
BeforeTreatment
Imediatly after 2 hours 12 hours 24 hours
Bio
mas
s /
mg.
cm-2
a
b

Halogen- containing chemicals 119 _________________________________________________________________________________
6.4. DISCUSSION
Antimicrobial resistance to conventional antimicrobial agents such as SH has been
documented [54]. Also, the use of SH can result in the production of harmful
disinfection by-products through their reaction with organic matter [55]. The best
known and characterized products are the trihalomethanes, which include chloroform,
bromoform, bromodichloromethane and chlorodibromomethane [56]. Drinking water
quality regulation was specified to limit trihalomethane levels to 100 µg.L-1 [57].
In food industry, outbreaks of foodborne pathogens have been increasing in the
last decades due to the high food demand to match population needs [58]. Therefore,
the susceptibility to microbial contamination and biofilm formation requires major
investments for produce decontamination and sanitation of the facilities [59]. However,
resistance to disinfectants has been increasing, urging the necessity for the
development of new formulations [10-12, 17, 18]. It is necessary to find alternative
compounds capable of removing and/or killing undesired resistant microorganisms
[60]. Brominated compounds might be a suitable replacement to the conventional
chlorinated antimicrobials. In this work, the antimicrobial activity and capacity for
biofilm control of chlorine (as sodium hypochlorite) and three bromine based chemicals
(CTAB, BrCl and BrOH), against planktonic and biofilm embedded P. fluorescens, was
studied. Additionally, several aspects of their interaction with the bacteria were
assessed. The three brominated chemicals were selected based on their structure,
particularly the presence of bromine [61, 62], chlorine [62, 63] and carboxyl group [64]
known for their antimicrobial properties. P. fluorescens was chosen as a well-studied
Gram negative bacterium, and ubiquitous in the natural, medical and industrial
environments, that can cause serious problems in either its planktonic or biofilm states
[33, 65]. In addition, this bacterium is known to form biofilms resistant to disinfectants
[66]. Several bacterium physiological characteristics were assessed such as the MIC,
MBC, hydrophobicity, potassium (K+) release, and surface charge.
The MIC and MBC values of CTAB against P. fluorescens were 20 and 50 µg.ml-1,
respectively (Table 6.1). Previous use of CTAB, revealed MIC values of 4 µg.mL-1 against
the Gram negative Salmonella typhimurium and P. aeruginosa and 18 µg.mL-1 for the
yeast Candida albicans [67]. The MIC and MBC of SH were 500 µg.mL-1 (Table 6.1). A
range of SH concentrations from 50 to 5000 µg.mL-1 has been determined by several
authors, for a variety of conditions, and bacteria [68-71]. Differences between the

120 Chapter 6
_________________________________________________________________________________
values obtained within the previous studies can be explained by the use of different
methods and bacteria to determine these parameters. Moreover, it is widely assumed
that no strain can characterize the behavior of a species [72]. To our knowledge, this is
the first study reporting the antimicrobial properties of BrOH and BrCl. It was found that
both chemicals have antimicrobial activity, with a MIC and MBC against P. fluorescens
of 650 and 700 µg.mL-1 for BrCl and 850 and 900 µg.mL-1 for BrOH. The lower MIC and
MBC of BrCl is possibly due to the presence of chlorine, known for its antimicrobial
properties [63, 73].
In order to understand the action of the selected chemicals on P. fluorescens,
several aspects of the interaction between the chemicals and the bacterial cells were
assessed, particularly the surface physicochemical properties, charge and K+ release.
CTAB is a compound that binds to the negative cell surface of bacteria due to
electrostatic attraction by chemisorption [74-76]. Azeredo et al. [75] revealed that
when a concentration of CTAB higher than the MBC is used, hydrophobicity and surface
charge properties can be enhanced and bacteria becomes hydrophilic and positively
charged. Upon interaction with the surface, CTAB promotes cell membrane
disorganization [77] or even disruption [78]. In this work, the effect of cell disruption
was verified measuring by the amount of K+ released.
Conversely to the effect caused by CTAB, P. fluorescens exposure to SH decreased
the bacterial polar (𝛾𝑠𝐴𝐵) character and, consequently the capacity to accept (𝛾𝑠
+) or
donate (𝛾𝑠−) electrons. According to Gottardi et al. [63], the action of active chlorine
(hypochlorous acid - HOCl) in bacteria can be divided in two effects, non-lethal and
lethal. The first implies reversible chlorination of the bacterial surface and the second is
based on penetration into the bacteria combined with irreversible alterations. SH can
also promote aggregation of essential proteins [73]. The present study corroborates the
findings of Winter et al. [73] on the membrane destabilization effects. In fact, K+ release
is a consequence of membrane leakage. The interaction of active chlorine does not
interfere with the cell surface charge, suggesting covalent links between the biocide
and the bacterial membrane [63]. Moreover, SH dissociation in ions can originate salt
formation that can help explain the increase in conductivity. BrOH decreased bacterial
hydrophilic characteristics, and improved electron acceptance (𝛾𝑠+). These results,
together with reduction in negative surface charge suggest that, electrostatic
interactions of bromine based chemicals with the membrane occur after Br- dissociation

Halogen- containing chemicals 121 _________________________________________________________________________________
from the structure, as it was described for BrOH [79]. The membrane interaction of the
chemical may also promote destabilization and consequently potassium release into
the solution. The effects of BrCl were less noticeable on the hydrophobicity values, but
this was the chemical that most affected the bacterial cells charge. The difference in
antimicrobial activity and mode of action of BrOH and BrCl can be due to the
presence/absence of the chemical entities OH- or Cl-. It seems that the presence of Cl-
improves the antimicrobial activity of the molecules, causing a significant decrease in
the cell surface charge and the leakage of intracellular K+. Even if significant effects on
the cell surface properties and charge were promoted by the biocides, no changes were
induced on OMP expression. This may indicate that these compounds may not
potentiate antimicrobial resistance. This hypothesis is based on the knowledge of the
OMP importance in bacterial resistance to biocides and antibiotics [41, 80-83].
The penetration of CTAB through P. fluorescens biofilms was completely retarded
due to the presence of biofilms. In fact, bacteria in biofilms exhibit less susceptibility to
antimicrobials due to their spatial heterogeneity, which consequently originates
nutrient depletion within the biofilm, reduced access of the chemicals to the bacteria
inside the biofilm, or biocide interaction with extracellular polymeric substances, and
the existence of degradative enzymes and neutralizing chemicals [65, 84, 85]. Simões
et al. [86] supports the accessibility hypothesis as it was verified that less dense biofilms
were more susceptible than denser biofilms. The retardation of SH was negligible. A
previous study showed that chlorine effectively diffuses through biofilms with a
diffusion coefficient in water estimated to be 0.84 cm2.s-1 [87]. BrOH was moderately
retarded by the biofilm, while BrCl was able to penetrate the biofilm without
retardation. Again, this result supports the observations reported previously in this
study, that the presence of ions Br-, Cl- or OH- may define the activity of the molecule.
Biofilms are organized cell aggregates in a self-produced extracellular matrix and,
can form on living or inert surfaces, which can create serious problems in several fields
if disinfection protocols fail [65]. As biofilms are a major problem in industry, it is
important to understand the chemical mechanism of disinfection and its efficacy on
pre-established biofilms. Flow generated P. fluorescens biofilms were exposed, for 1 h,
to the chemicals at their MBC values. Only modest reductions in the log CFU.cm-2 were
obtained. This fact reinforces the higher resistance of biofilm cells compared to their
planktonic counterparts [7, 88]. In terms of mass removal, the use of BrCl, CTAB or SH
promoted low removal of the total biofilm mass (15%). It can be hypothesized that CTAB

122 Chapter 6
_________________________________________________________________________________
acts eroding the biofilm, which may consist on eradication of the superficial bacteria,
and disruption of the matrix. This assumption is based on the fact that CTAB is the most
efficient chemical despite lacking the capacity to cross the biofilm layers. The possibility
of BrCl and SH to pass through the biofilm and the lack of efficacy may be a consequence
of an intrinsic or acquired resistance of biofilms [73, 89]. Furthermore, these chemicals
did not interfere with quorum sensing. A screening protocol developed by McLean et
al. [46] was used for the detection of quorum signal inhibition (targeting acylated
homoserine lactones dependent signaling), no effects were found other than
antimicrobial action.
To ascertain the ability of the biofilms to regrow after 1 h exposure to the
antimicrobial agents, the initial conditions were reestablished, so that a disinfection
practice in industry was mimicked. Biofilm regrowth was found on SH treated biofilms
after 12h h, and on BrCl treated biofilms after 24 h. It was verified that CFU recovered
to its initial values 2 h after exposure to all chemicals. This result can be explained by
the possible presence of starved or injured cells or potentially viable but not culturable
cells [10, 90, 91]. Also, Pereira et al. [92] found that 7-days old P. fluorescens biofilms
formed in a flow cell system are in the stationary or stabilization phase. This means that
the loss of biomass due to physical stresses is balanced by the growth of new cells at
the edge of the biofilm [93]. The results obtained show low to moderate effects of the
selected biocides on biofilm removal and killing, and rapid regrowth to the stabilization
phase. These results suggest that the selected biocides, at the concentrations tested,
had no significant effects on the dynamic behavior of the biofilms. It is apparent that
the promising results obtained with the tests on planktonic cells did not provide
relevant insights on their application in biofilm control, even if the same strain and
antimicrobial concentrations were used in both tests. Moreover, in this study, it seems
that the chemical nature of the biocide was not relevant on biofilm control, and the
bromine-based products had no clear antimicrobial advantage on biofilm control over
SH. It is possible that the combination of bromine-based products and chlorine might
potentiate their antimicrobial action. Pioneer studies [94, 95], demonstrated synergistic
antimicrobial relationship when bromine was added to chlorine solutions. However, as
with free chlorine, there are safety concerns about the production of brominated
organic compounds and their impact on human and environmental safety, even if
bromide ion has a low degree of toxicity [28].

Halogen- containing chemicals 123 _________________________________________________________________________________
The effect of the halogenated compounds tested was similar between each other.
Despite that BrOH and BrCl require higher MIC and the MBC than CTAB, they are both
similar to SH. Also, the results propose that the chemicals tested share a similar mode
of antimicrobial action against P. fluorescens. The overall results propose that the
selected bromine-based products can be a potential alternative to chlorine-based
products.
REFERENCES
[1] Jahid I, Ha S-D. A review of microbial biofilms of produce: Future challenge to food safety. Food Science and Biotechnology, 2012. 21(2):299-316.
[2] Stein C, Kuchenmüller T, Hendrickx S, Prüss-Űstün A, Wolfson L, Engels D, Schlundt J. The Global Burden of Disease Assessments—WHO Is Responsible? PLoS Neglected Tropical Diseases, 2007. 1(3):e161.
[3] Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV, Widdowson MA, Roy SL, Jones JL, Griffin PM. Foodborne illness acquired in the United States - major pathogens. Emerging Infectious Diseases, 2011. 17(1):7-15.
[4] Potera C. Forging a link between biofilms and disease. Science, 1999. 283(5409):1837-1839.
[5] Van Houdt R, Michiels CW. Biofilm formation and the food industry, a focus on the bacterial outer surface. Journal of Applied Microbiology, 2010. 109(4):1117-1131.
[6] Kuehn C, Graf K, Heuer W, Hilfiker A, Chaberny IF, Stiesch M, Haverich A. Economic implications of infections of implantable cardiac devices in a single institution. European Journal of Cardio-thoracic Surgery, 2010. 37(4):875-9.
[7] Davies D. Understanding biofilm resistance to antibacterial agents. Nature Reviews in Drug Discovery, 2003. 2(2):114-22.
[8] Mah T, O'Toole G. Mechanisms of biofilm resistance to antimicrobial agents. Trends in Microbiology, 2001. 9(1):34-39.
[9] Araújo P, Lemos M, Mergulhão F, Melo Ld, Simões M, Antimicrobial resistance to disinfectants in biofilms, in Science against microbial pathogens: communicating current research and technological advances, A. Méndez-Vilas, Editor. 2011, Formatex. p. 826-834.
[10] Simões LC, Lemos M, Pereira AM, Abreu AC, Saavedra MJ, Simões M. Persister cells in a biofilm treated with a biocide. Biofouling, 2011. 27(4):403-411.
[11] Bremer PJ, Monk I, Butler R. Inactivation of Listeria monocytogenes/Flavobacterium spp. biofilms using chlorine: impact of substrate, pH, time and concentration. Letters in Applied Microbiology, 2002. 35(4):321-5.
[12] Fraise AP. Biocide abuse and antimicrobial resistance--a cause for concern? Journal of Antimicrobial Chemotherapy, 2002. 49(1):11-2.
[13] Zottola EA, Sasahara KC. Microbial biofilms in the food processing industry - Should they be a concern? International Journal of Food Microbiology, 1994. 23(2):125-148.
[14] Ferraris M, Chiesara E, Radice S, Giovara A, Frigerio S, Fumagalli R, Marabini L. Study of potential toxic effects on rainbow trout hepatocytes of surface water treated with chlorine or alternative disinfectants. Chemosphere, 2005. 60(1):65-73.

124 Chapter 6
_________________________________________________________________________________
[15] Moore SL, Payne DN, Types of Antimicrobial Agents, in Russell, Hugo & Ayliffe's Principles and Practice of Disinfection, Preservation & Sterilization. 2008, Blackwell Publishing Ltd. p. 8-97.
[16] Russell AD, Factors Influencing the Efficacy of Antimicrobial Agents, in Russell, Hugo & Ayliffe's Principles and Practice of Disinfection, Preservation & Sterilization. 2008, Blackwell Publishing Ltd. p. 98-127.
[17] Nakata K, Tsuchido T, Matsumura Y. Antimicrobial cationic surfactant, cetyltrimethylammonium bromide, induces superoxide stress in Escherichia coli cells. Journal of Applied Microbiology, 2011. 110(2):568-79.
[18] Bore E, Hebraud M, Chafsey I, Chambon C, Skjaeret C, Moen B, Moretro T, Langsrud O, Rudi K, Langsrud S. Adapted tolerance to benzalkonium chloride in Escherichia coli K-12 studied by transcriptome and proteome analyses. Microbiology, 2007. 153(Pt 4):935-46.
[19] Chaidez C, Lopez J, Castro-del Campo N. Quaternary ammonium compounds: an alternative disinfection method for fresh produce wash water. Journal of Water and Health, 2007. 5(2):329-33.
[20] Hentzer M, Givskov M. Pharmacological inhibition of quorum sensing for the treatment of chronic bacterial infections. The Journal of Clinical Investigation, 2003. 112(9):1300-7.
[21] Rasmussen TB, Givskov M. Quorum sensing inhibitors: a bargain of effects. Microbiology, 2006. 152(Pt 4):895-904.
[22] Borchardt SA, Allain EJ, Michels JJ, Stearns GW, Kelly RF, McCoy WF. Reaction of acylated homoserine lactone bacterial signaling molecules with oxidized halogen antimicrobials. Applied and Environmental Microbiology, 2001. 67(7):3174-9.
[23] Givskov M, de Nys R, Manefield M, Gram L, Maximilien R, Eberl L, Molin S, Steinberg PD, Kjelleberg S. Eukaryotic interference with homoserine lactone-mediated prokaryotic signalling. Journal of Bacteriology, 1996. 178(22):6618-22.
[24] Salama MA, Almotabacani LA. Synthesis and chemistry of some new 2-mercaptoimidazole derivatives of possible antimicrobial activity. Phosphorus, Sulfur, and Silicon and the Related Elements, 2004. 179(2):305-319.
[25] Latif NAA, Saeed MM, Ahmed NS, Batran RZ, El-Mouhty NRA. Synthesis of some pyridine, pyrimidine and cyclohexenone derivatives as antibacterial agents. International Journal of Innovative Research in Science, Engineering and Technology, 2014. 3(1):8517 - 8529.
[26] Ortenzio L, Stuart L. A standard test for efficacy of germicides and acceptability of residual disinfecting activity in swimming pool water. Journal of the Association of Official Analytical Chemists, 1964. 47:540-547.
[27] Cousins CM, Allan CD. Sporicidal Properties of Some Halogens. Journal of Applied Bacteriology, 1967. 30(1):168-174.
[28] WHO. Bromide in Drinking-water, in Background document for development of WHO Guidelines for Drinking-water Quality. 2009, WHO Press: Geneva, Switzerland.
[29] Simões M, Simões LC, Vieira MJ. Species association increases biofilm resistance to chemical and mechanical treatments. Water Research, 2009. 43(1):229-237.
[30] Mundo MA, Padilla-Zakour OI, Worobo RW. Growth inhibition of foodborne pathogens and food spoilage organisms by select raw honeys. International Journal of Food Microbiology, 2004. 97(1):1-8.
[31] Simões M, Cleto S, Pereira MO, Vieira MJ. Influence of biofilm composition on the resistance to detachment. Water Science and Technology, 2007. 55(8-9):473-480.
[32] CLSI, Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically: Approved Standard - Ninth Edition M07- A09. 2012, NCCLS.

Halogen- containing chemicals 125 _________________________________________________________________________________
[33] Ferreira C, Pereira AM, Pereira MC, Melo LF, Simões M. Physiological changes induced by the quaternary ammonium compound benzyldimethyldodecylammonium chloride on Pseudomonas fluorescens. Journal of Antimicrobial Chemotherapy, 2011. 66(5):1036-43.
[34] Johnston MD, Lambert RJW, Hanlon GW, Denyer SP. A rapid method for assessing the suitability of quenching agents for individual biocides as well as combinations. Journal of Applied Microbiology, 2002. 92(4):784-789.
[35] Busscher HJ, Weerkamp AH, van der Mei HC, van Pelt AW, de Jong HP, Arends J. Measurement of the surface free energy of bacterial cell surfaces and its relevance for adhesion. Applied and Environmental Microbiology, 1984. 48(5):980-3.
[36] Simões LC, Simões M, Oliveira R, Vieira MJ. Potential of the adhesion of bacteria isolated from drinking water to materials. Journal of Basic Microbiology, 2007. 47(2):174-183.
[37] Janczuk B, Chibowski E, Bruque JM, Kerkeb ML, Caballero FG. On the Consistency of Surface Free Energy Components as Calculated from Contact Angles of Different Liquids: An Application to the Cholesterol Surface. Journal of Colloid and Interface Science, 1993. 159(2):421-428.
[38] van Oss CJ, Chaudhury MK, Good RJ. Monopolar surfaces. Advances in Colloid and Interface Science, 1987. 28(1):35-64.
[39] Van Oss CJ, Good RJ, Chaudhury MK. Additive and nonadditive surface tension components and the interpretation of contact angles. Langmuir, 1988. 4(4):884-891.
[40] Van Oss CJ, Ju L, Chaudhury MK, Good RJ. Estimation of the polar parameters of the surface tension of liquids by contact angle measurements on gels. Journal of Colloid and Interface Science, 1989. 128(2):313-319.
[41] Winder CL, Al-Adham ISI, Abdel Malek SMA, Buultjens TEJ, Horrocks AJ, Collier PJ. Outer membrane protein shifts in biocide-resistant Pseudomonas aeruginosa PAO1. Journal of Applied Microbiology, 2000. 89(2):289-295.
[42] Simões M, Pereira MO, Sillankorva S, Azeredo J, Vieira MJ. The effect of hydrodynamic conditions on the phenotype of Pseudomonas fluorescens biofilms. Biofouling, 2007. 23(3-4):249-58.
[43] Laemmli UK. Characterization of DNA condensates induced by poly(ethylene oxide) and polylysine. Proceedings of the National Academy of Sciences, 1975. 72(11):4288-4292.
[44] Simões M, Pereira MO, Machado I, Simões LC, Vieira MJ. Comparative antibacterial potential of selected aldehyde-based biocides and surfactants against planktonic Pseudomonas fluorescens. J Ind Microbiol Biotechnol, 2006. 33(9):741-9.
[45] Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Short Protocols in Molecular Biology - Fifth Edition. Vol. 1: Jonh Wiley and Sons Press. 2002.
[46] McLean RJC, Pierson III LS, Fuqua C. A simple screening protocol for the identification of quorum signal antagonists. Journal of Microbiological Methods, 2004. 58(3):351-360.
[47] Adonizio AL, Downum K, Bennett BC, Mathee K. Anti-quorum sensing activity of medicinal plants in southern Florida. Journal of Ethnopharmacology, 2006. 105(3):427-435.
[48] Anderl JN, Franklin MJ, Stewart PS. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrobial Agents and Chemotherapy, 2000. 44(7):1818-24.
[49] Singh R, Ray P, Das A, Sharma M. Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. Journal of Antimicrobial Chemotherapy, 2010. 65(9):1955-1958.
[50] Teodósio JS, Simões M, Melo LF, Mergulhão FJ. Flow cell hydrodynamics and their effects on E. coli biofilm formation under different nutrient conditions and turbulent flow. Biofouling, 2011. 27(1):1-11.

126 Chapter 6
_________________________________________________________________________________
[51] Simões M, Carvalho H, Pereira MO, Vieira MJ. Studies on the behaviour of Pseudomonas fluorescens biofilms after ortho-phthalaldehyde treatment. Biofouling, 2003. 19(3):151-157.
[52] APHA, AWWA, WPCF, Standard methods for the examination of water and wastewater, L.S. Clesceri, A.E. Greenberg, and R.R. Trussel, Editors. 1989, American Public Health Association: Washington DC, USA.
[53] Simões M, Pereira MO, Vieira MJ. Validation of respirometry as a short-term method to assess the efficacy of biocides. Biofouling, 2005. 21(1):9-17.
[54] Ridgway H, Olson B. Chlorine resistance patterns of bacteria from two drinking water distribution systems. Applied and Environmental Microbiology, 1982. 44(4):972-987.
[55] Grant D, Bott T. Biocide dosing strategies for biofilm control. Heat Transfer Engineering, 2005. 26(1):44-50.
[56] Rodrigues PM, Esteves da Silva JC, Antunes MCG. Factorial analysis of the trihalomethanes formation in water disinfection using chlorine. Analytica Chimica Acta, 2007. 595(1):266-274.
[57] Decreto-Lei no 306/2007 de 27 de Agosto, relativo ao controlo da qualidade da água destinada ao consumo humano.
[58] Bhat RV. Human health problems associated with current agricultural food production. Asia Pacific Journal of Clinical Nutrition, 2008. 17 Suppl 1:91-4.
[59] Todd EC, Michaels BS, Holah J, Smith D, Greig JD, Bartleson CA. Outbreaks where food workers have been implicated in the spread of foodborne disease. Part 10. Alcohol-based antiseptics for hand disinfection and a comparison of their effectiveness with soaps. Journal of Food Protection, 2010. 73(11):2128-2140.
[60] Simões LC, Simões M. Biofilms in drinking water: problems and solutions. RSC Advances, 2013. 3(8):2520-2533.
[61] Petruso S, Bonanno S, Caronna S, Ciofalo M, Maggio B, Schillaci D. Oxidative halogenation of substituted pyrroles with Cu(II). Part IV. Bromination of 2-(2′-hydroxybenzoyl)pyrrole. A new synthesis of bioactive analogs of monodeoxypyoluteorin. Journal of Heterocyclic Chemistry, 1994. 31(4):941-945.
[62] Ward FE, Garling DL, Buckler RT, Lawler DM, Cummings DP. Antimicrobial 3-methylene flavanones. Journal of Medicinal Chemistry, 1981. 24(9):1073-1077.
[63] Gottardi W, Nagl M. Chlorine covers on living bacteria: the initial step in antimicrobial action of active chlorine compounds. Journal of Antimicrobial Chemotherapy, 2005. 55(4):475-82.
[64] Kabara JJ, Swieczkowski DM, Conley AJ, Truant JP. Fatty acids and derivatives as antimicrobial agents. Antimicrobial Agents and Chemotherapy, 1972. 2(1):23-8.
[65] Simões M, Simões LC, Vieira MJ. A review of current and emergent biofilm control strategies. LWT - Food Science and Technology, 2010. 43(4):573-583.
[66] Simões M, Simões LC, Pereira MO, Vieira MJ. Sodium dodecyl sulfate allows the persistence and recovery of biofilms of Pseudomonas fluorescens formed under different hydrodynamic conditions. Biofouling, 2008. 24(1):35-44.
[67] Lindstedt M, Allenmark S, Thompson RA, Edebo L. Antimicrobial activity of betaine esters, quaternary ammonium amphiphiles which spontaneously hydrolyze into nontoxic components. Antimicrobial Agents and Chemotherapy, 1990. 34(10):1949-54.
[68] Maillard J-Y. Innate resistance to sporicides and potential failure to decontaminate. Journal of Hospital Infection, 2011. 77(3):204-209.
[69] Perez J, Springthorpe VS, Sattar SA. Activity of selected oxidizing microbicides against the spores of Clostridium difficile: Relevance to environmental control. American Journal of Infection Control, 2005. 33(6):320-325.

Halogen- containing chemicals 127 _________________________________________________________________________________
[70] Rutala WA, Weber DJ. Uses of inorganic hypochlorite (bleach) in health-care facilities. Clinical Microbiology Reviews, 1997. 10(4):597-610.
[71] Rasmussen LH, Kjeldgaard J, Christensen JP, Ingmer H. Multilocus sequence typing and biocide tolerance of Arcobacter butzleri from Danish broiler carcasses. BMC Research Notes, 2013. 6:322.
[72] Fux CA, Shirtliff M, Stoodley P, Costerton JW. Can laboratory reference strains mirror "real-world" pathogenesis? Trends in Microbiology, 2005. 13(2):58-63.
[73] Winter J, Ilbert M, Graf PC, Ozcelik D, Jakob U. Bleach activates a redox-regulated chaperone by oxidative protein unfolding. Cell, 2008. 135(4):691-701.
[74] Rodrigues A, Nogueira R, Melo LF, Brito AG. Effect of low concentrations of synthetic surfactants on polycyclic aromatic hydrocarbons (PAH) biodegradation. International Biodeterioration & Biodegradation, 2013. 83(0):48-55.
[75] Azeredo L, Pacheco AP, Lopes I, Oliveira R, Vieira MJ. Monitoring cell detachment by surfactants in a parallel plate flow chamber. Water Science and Technology, 2003. 47(5):77-82.
[76] Neu TR. Significance of bacterial surface-active compounds in interaction of bacteria with interfaces. Microbiological Reviews, 1996. 60(1):151-66.
[77] McDonnell G, Russell AD. Antiseptics and disinfectants: activity, action, and resistance. Clinical Microbiology Reviews, 1999. 12(1):147-79.
[78] Simões M, Pereira MO, Vieira MJ. Action of a cationic surfactant on the activity and removal of bacterial biofilms formed under different flow regimes. Water Research, 2005. 39(2–3):478-486.
[79] Cooks RG, Ronayne J, Williams DH. Studies in mass spectrometry. Part XXI. Migrations to carbonium ion centres generated upon electron impact. Journal of the Chemical Society C: Organic, 1967;(0):2601-2607.
[80] Walsh SE, Maillard J-Y, Russell AD, Catrenich CE, Charbonneau DL, Bartolo RG. Development of bacterial resistance to several biocides and effects on antibiotic susceptibility. Journal of Hospital Infection, 2003. 55(2):98-107.
[81] Delcour AH. Outer membrane permeability and antibiotic resistance. Biochimica et Biophysica Acta, 2009. 1794(5):808-16.
[82] Cha JS, Cooksey DA. Copper resistance in Pseudomonas syringae mediated by periplasmic and outer membrane proteins. Proceedings of the National Academy of Sciences of the United States of America, 1991. 88(20):8915-9.
[83] Masuda N, Sakagawa E, Ohya S. Outer membrane proteins responsible for multiple drug resistance in Pseudomonas aeruginosa. Antimicrobial Agents and Chemotherapy, 1995. 39(3):645-9.
[84] Fazlara A, Ekhtelat M. The Disinfectant Effects of Benzalkonium Chloride on Some Important Foodborne Pathogens. American-Eurasian Journal of Agricultural & Environmental Sciences, 2012. 12(1):23-29.
[85] Brown MR, Gilbert P. Sensitivity of biofilms to antimicrobial agents. Journal of Applied Bacteriology, 1993. 74 Suppl:87S-97S.
[86] Simões M, Simões LC, Machado I, Pereira MO, Vieira MJ. Control of Flow-Generated biofilms with surfactants: Evidnece of Resistance and Recovery. Food and Bioproducts Processing, 2006. 84(C4):338-345.
[87] Stewart PS, Rayner J, Roe F, Rees WM. Biofilm penetration and disinfection efficacy of alkaline hypochlorite and chlorosulfamates. Journal of Applied Microbiology, 2001. 91(3):525-532.
[88] Simões LC, Simões M, Vieira MJ. Adhesion and biofilm formation on polystyrene by drinking water-isolated bacteria. Antonie van Leeuwenhoek, 2010. 98(3):317-329.

128 Chapter 6
_________________________________________________________________________________
[89] Gibson H, Taylor JH, Hall KE, Holah JT. Effectiveness of cleaning techniques used in the food industry in terms of the removal of bacterial biofilms. Journal of Applied Microbiology, 1999. 87(1):41-8.
[90] Banning N, Toze S, Mee BJ. Escherichia coli survival in groundwater and effluent measured using a combination of propidium iodide and the green fluorescent protein. Journal of Applied Microbiology, 2002. 93(1):69-76.
[91] Gião MS, Wilks S, Azevedo NF, Vieira MJ, Keevil CW. Incorporation of natural uncultivable Legionella pneumophila into potable water biofilms provides a protective niche against chlorination stress. Biofouling, 2009. 25(4):335-41.
[92] Pereira MO, Morin P, Vieira MJ, Melo LF. A versatile reactor for continuous monitoring of biofilm properties in laboratory and industrial conditions. Letters in Applied Microbiology, 2002. 34(1):22-26.
[93] Capdeville B, Nguyen KM, Rols JL, Biofilm Modelling : Structural, Reactional and Diffusional Aspects, in Biofilms — Science and Technology, L.F. Melo, et al., Editors. 1992, Springer Netherlands. p. 251-276.
[94] Kristoffersen T. Mode of action of hypochlorite sanitizers with and without sodium bromide. Journal of Dairy Science, 1958. 41(7):942-949.
[95] Shere L, Kelley MJ, Richardson JH. Effect of bromidehypochlorite bactericides on microorganisms. Applied Microbioogyl, 1962. 10:538-41.

CHAPTER 7 ENZYMATIC TREATMENTS FOR BIOFILM CONTROL
This chapter was submitted as:
Araújo PA, Machado I, Mergulhão F, Melo L, Simões M. Evaluation of the synergistic potential of enzymes with quaternary ammonium compounds for biofilm control.
Araújo PA, Machado I, Mergulhão F, Melo L, Simões M. Decreased efficacy of enzymes on the control of flow generated biofilms.

130 Chapter 7
_________________________________________________________________________________
ABSTRACT
Current biofilm control strategies are inefficient in removing biofilms from equipment
surfaces. The use of enzymes is considered a new and environmentally friendly
approach for biofilm control. This work investigates the effects of a β-glucanase, a
protease, a lipase, and α-amylase, alone and in combination with benzalkonium chloride
(BAC) and cetyltrimethyl ammonium bromide (CTAB) in the control of biofilms formed
by Bacillus cereus and Pseudomonas fluorescens.
Synergistic effects of the combination of the biocides with the enzymes were
found against the biofilms developed using the microtiter tests. This biofilm control
strategy was also applied against P. fluorescens biofilms formed in a flow cell system. In
this case, the enzymes, when applied alone, resulted in low to moderate biofilm
removal and CFU reduction. Following the enzymatic or CTAB-enzyme treatments, it
was found biofilm regrowth and long term control events. The effect of the enzymes
against planktonic cells was also evaluated by respirometry. Most enzymes showed to
have the ability to reduce the respiratory activity of planktonic cells, except α-amylase
that, instead, increased the activity of P. fluorescens. Moreover, protease, lipase and
α-amylase hindered the antimicrobial activity of the biocides, apparently due to
chemical neutralization.
The overall results show the synergistic potential of selected enzymes with BAC
and CTAB in the control of biofilms and demonstrate that a careful selection and
application of enzymes must be considered since these molecules can quench the
activity of antimicrobial agents.

The effect of enzymes in biofilm control 131 _________________________________________________________________________________
7.1 INTRODUCTION
Biofilms can be defined as microbial sessile communities, characterized by cells
embedded in self-produced extracellular polymeric substances (EPS) [1]. The type and
amount of EPS constituents can be strain dependent and vary with the environmental
conditions under which biofilms are formed. However, there is unanimity in considering
polysaccharides, proteins and DNA as its main constituents. EPS contribute to the
mechanical stability of biofilms, enabling them to withstand shear forces, dehydration
and chemical attacks [2]. EPS also enhance robustness and survival of the embedded-
microorganisms on a substratum, by acting as a chemically reactive diffusional transport
barrier, slowing the penetration of antimicrobial agents. Furthermore, the
exopolymeric matrix increases biofilm attachment to the substratum and stabilizes it,
thereby reducing its susceptibility to sloughing by hydrodynamic stress [3].
The use of water for diverse processes of food industry increases the chance for
microbial contamination and biofilm formation. Typical consequences of biofouling are
reduced operational efficacy in heat exchangers, increased operational pressure,
blockage of tubes, increased energy consumption, accelerated metal surfaces
corrosion, final product contamination and consequential potential health problems
[4, 5]. Biofouling deposits can contain pathogenic and spoilage bacteria, increasing the
risks for human consumption. Consequently, contingency plans to control microbial
contaminations need to be applied. The main objective of microbial control is to
eliminate and/or reduce the number of microorganisms and their activity, as well as to
prevent and control the formation of biological deposits on process equipment [6]. As
a result, programs such as Good Manufacturing Practice (GMP) and Hazard Analysis and
Critical Control Points (HACCP) [7] are currently employed to control microbial
proliferation. Biofilm control in food industry is usually performed without dismantling
or opening the equipment in a Clean-in-Place (CIP) process. CIP consists on running
alternated cycles of detergent and disinfectant solutions with water rinses with
increased turbulence [8]. This method typically uses caustic acids, surfactants, biocides
and, occasionally, enzymes [4, 8-10]. As the continued use of biocides can induce
microbial resistance, new control strategies must be developed. Also, as regulation
restricts the use of toxic biocides, eco-friendly strategies represent a new approach for
the control of biofilms of food industry [11, 12].

132 Chapter 7
_________________________________________________________________________________
Enzymes are already used in a wide range of applications, including the production
of food and beverages, detergents, clothing, paper products, pharmaceuticals, fuel and
monitoring devices [13]. For instance, glucanases are used to reduce the viscosity in
barley and oats used in animal feed, enhancing their digestibility. Proteases are the
most used enzymes in dairy industry, and in cleaning detergents. They are also used for
protein hydrolysis, milk clotting, low-allergenic infant-food formulation, flavor
improvement in milk and cheese, meat tenderization and prevention of chill haze
formation in brewing. Lipases are used for flavoring cheese, in-situ emulsification for
dough conditioning, support for lipid digestion in young animals, and synthesis of
aromatic molecules. Amylases are used for starch liquefaction and sacharification, and
also increase shelf life and, by retaining moist, they improve product quality. The
elasticity and softness of bread is provided by amylases. In addition, flour adjustment,
and low calorie beer are also processes that use amylases. Amylases are the second
most widely used group of enzymes, along with cellulases [13, 14], in the formulation
of enzyme detergents, mainly to remove food residues of starch-based foods. Enzymes
have already been tested in biofilm control, and are proven to potentiate the action of
some antimicrobial agents [15-18].
This work evaluates the effectiveness of an enzymatic treatment in the control of
Bacillus cereus and Pseudomonas fluorescens biofilms. The combined action of enzymes
and quaternary ammonium compounds (QACs) was assessed against biofilms formed
in microtiter plates and in a flow cell system. Additionally, tests with planktonic bacteria
were performed in order to ascertain the supposed action of enzymes as quenchers of
antimicrobial agents.
7.2 MATERIALS AND METHODS
BACTERIA AND CULTURE CONDITIONS
P. fluorescens ATCC 13525T and a B. cereus strain isolated from a disinfectant solution
and identified by 16S rRNA gene sequencing [19] were used in this study. Bacteria were
grown at 30 ± 3 oC, in the medium composed by 5 g.L-1 glucose, 2.5 g.L-1 peptone,
1.25 g.L-1 yeast extract, in phosphate buffer (PB), pH 7, 25 mM, that uses glucose (Merk)
as the main carbon source [20]. Bacterial suspensions were prepared by inoculation of
a single colony grown on solid medium (above medium supplemented with 10% agar)

The effect of enzymes in biofilm control 133 _________________________________________________________________________________
into a 1 L flask containing 250 mL of sterile nutrient medium. This bacterial suspension
was incubated overnight at 30 ± 3 oC in an orbital shaker (120 rpm).
ANTIMICROBIAL AGENTS AND ENZYMES
Two QACs were used: benzalkonium chloride (BAC) and cetyltrimethyl ammonium
bromide (CTAB) (Sigma, Portugal). In order to ascertain the effect of the enzymes on
the bacteria, QACs were used at their minimum bactericidal concentrations (MBC),
previously assessed by Araújo et al. [21], chapter 4. The MBC of BAC was 10 mg.L-1 for
B. cereus, and 35 mg.L-1 for P. fluorescens; CTAB was used at 20 mg.L-1 on the first and
at 35 mg.L-1 on the second bacterium. QACs were prepared as concentrated aqueous
solutions.
The enzymes tested were provided by Novozymes (Denmark). Their commercial
names are Ultraflo® (3.2.1.6, β-glucanase), Alcalase® (3.4.21.62, protease-subtisilin),
Lecitase® (3.1.1.3, lipase), and Fungamyl® (3.2.1.1, α-amylase), supplied as aqueous
solutions containing 5-30% active protein. The enzymes were used diluted (1:100) in PB
[17]. The solutions combining enzymes with biocides started as enzymatic solutions to
which a concentrated solution of biocide was added to achieve the desired
concentration.
BIOFILM FORMATION IN MICROTITER PLATES
After the growth period, bacterial suspensions were harvested by centrifugation, using
an Eppendorf 5810R centrifuge with the A-4-62 rotor (Göttinger, Germany) (3999 g,
10 minutes) and resuspended in fresh culture medium to a final density of
1 × 109 cells.mL-1. The methodology used to grow biofilms was based on the modified
microtiter plate test, as proposed by Stepanović et al. [22]. For each bacterium, 200 µL
of the bacterial suspension, 100-fold diluted, were transferred to the wells of sterile
96-wells flat-bottomed polystyrene tissue culture plates (Orange Scientific, Portugal).
Plates were incubated in an orbital shaker (120 rpm) for 24 h at 30 ± 3 oC. Negative
controls consisted of culture medium with no bacterial cells.

134 Chapter 7
_________________________________________________________________________________
BIOFILM CONTROL USING ENZYMES
The effect of each enzyme and QAC, as well as different combinations of enzymes with
antimicrobial agents on biofilms formed in microtiter plates was screened using the
methods presented by Simões et al. [23] and Lequette et al. [17]. Briefly, the medium
was removed and the wells were gently washed twice with PB to remove reversibly
adhered bacteria. The remaining attached bacteria were submitted to a process that
consisted in biofilm exposure to: (1) an enzymatic solution for 1 h, (2) a solution of
enzyme and biocide for 1 h, and (3) an enzymatic solution for 30 minutes, followed by
a gentle washing step with PB, and then, the biocide for 30 minutes (30 + 30 min). These
tests were performed in an orbital shaker at 30 ± 3 oC, 120 rpm. Controls consisted of
PB or biocide solutions, in the absence of enzymes. QACs were used at their MBC values
[21]. This concentration will affect the biofilms only to a certain extent, not killing all the
embedded bacteria, so that any effect caused by the selected enzymes, other than
killing, could be recognized.
BIOFILM MASS AND VIABILITY ASSESSMENT
The biofilm mass was quantified using crystal violet staining (Merck, Portugal), and the
modified Alamar blue (Sigma-Aldrich, Portugal) microtiter plate assay was applied to
determine the bacterial viability of the biofilm-cells. Both methods were described by
Araújo et al. [24], see chapter 3.
BIOFILM CONTROL ACTIVITY CLASSIFICATION
The effects of the enzymes, biocides, and combination of enzymes with biocides on
biofilms were classified based on a ranking proposed by Lemos et al. [25]. Values of
killing and removal percentage, or CFU reduction inferior to 10% represent insignificant
efficacy of the product tested, 10% to 30% low efficacy, 30% to 60% moderate efficacy,
60% to 85% high efficacy and values superior to 85% very high efficacy.
The effect of the combination of enzymes with biocides was classified as described
by Saavedra et al. [26], to elucidate the effects of the interaction. The combination of
enzyme with biocide is considered antagonistic if [effect of combination enzyme and
biocide – (effect biocide + effect enzyme)/2] < 0; indifferent if the 0 ≤ (effect of

The effect of enzymes in biofilm control 135 _________________________________________________________________________________
combination enzyme and biocide) – (effect biocide + effect enzyme)/2 < effect enzyme
or effect biocide; additive if (effect biocide) < [(effect of combination enzyme and
biocide) – (effect biocide + effect enzyme)/2] < 2× (enzyme effect or biocide effect); and
synergistic if (effect of combination enzyme and biocide) > 2× (effect enzyme or effect
biocide).
BIOFILM FORMATION IN A FLOW CELL SYSTEM
To test these strategies using biofilms similar to those found in industrial environments,
biofilms were developed in a flow cell system, see chapter 5 (Figure 5.1). The biofilms
were left to develop for 7 days at a Reynolds number (Re) of 4000, corresponding to a
flow velocity of 0.4 m.s-1. Following this period, two coupons were analyzed as control
(without any treatment) for each experiment; subsequently the biofilms were
submitted to a similar treatment as the one used for microtiter plate biofilms. The flow
cell was carefully emptied, and then the solutions of CTAB, enzymes, and solutions
combining CTAB with enzymes ran through the flow cells at a flow velocity of
0.006 m.s-1, for 1 h. After that period, the initial conditions of the system were restored.
Control experiments with PB were also performed. Four coupons, two from each flow
cell were removed at different time periods: before chemical exposure (control),
immediately after the exposure, and then 2, 12 and 24 h post treatment, in order to
observe putative long-term effects or regrowth events following the treatment.
FLOW GENERATED BIOFILM CHARACTERIZATION
Biofilm mass and cell density were assessed as indicators of control. The biofilms
covering the coupons were completely scraped using a sterile scalpel, resuspended in
10 mL of PB, and vortexed for 30 s. The organic mass was determined according to the
standard methods - American Public Health Association [APHA], American Water Works
Association [AWWA], Water Pollution Control Federation [WPCF] [27]. The cell density
was assessed in terms of colony forming units (CFUs) in Plate Count Agar (Merck,
Portugal), according to Simões et al. [28]. The effectiveness of the control strategy was
classified according to the rank based on the classification proposed by Lemos et al.
[25].

136 Chapter 7
_________________________________________________________________________________
TESTS WITH PLANKTONIC CELLS
To assess the antimicrobial potential of the selected enzymes in planktonic cells, the
respiratory activity of B. cereus and P. fluorescens cell suspensions was ascertained by
measuring oxygen uptake rates in a biological oxygen monitor (Yellow Springs
Instruments 5300A) after the exposure to the QACs and the enzymes for 1 h. This
method was described previously by Araújo et al. [21], see chapter 4. The concentration
of O2 used by the bacteria for the oxidation of glucose corresponds to the exogenous
respiration rate, which is obtained by the difference between the total and endogenous
rates [28]. The respiratory activity, in mgO2. mgorganic mass -1. min-1, was calculated
according to the following equation:
𝑅𝑒𝑠𝑝𝑖𝑟𝑎𝑡𝑜𝑟𝑦 𝑎𝑐𝑡𝑖𝑣𝑖𝑡𝑦 =𝑒𝑥𝑜𝑔𝑒𝑛𝑜𝑢𝑠 𝑟𝑎𝑡𝑒× 𝑂
2𝑠𝑜𝑙𝑢𝑏𝑖𝑙𝑖𝑡𝑦
𝑏𝑎𝑐𝑡𝑒𝑟𝑖𝑎𝑙 𝑚𝑎𝑠𝑠 (eq. 7.1)
where the metabolic rate was measured as the percentage of O2 consumed by the cells
in time, the O2 solubility was considered 8.6 mg.L-1 [29], and the bacterial mass obtained
by the determination of total volatile solids (TSV) according to standard methods (APHA,
AWWA, and WPCF, [27], expressed as mgorganic mass.L-1. If respiratory activity < control,
there was killing of the microorganisms, whereas if respiratory activity > control, there
was metabolic potentiation.
STATISTICAL ANALYSIS
The experimental data was analyzed using the statistical program SPSS - Statistical
Package for the Social Sciences, Version 22.0 (Armonk, NY, USA). The average and
standard deviation were calculated for all cases, from at least three independent
experiments performed for each condition tested. Normality of data distribution was
assessed by the Kolmogorov-Smirnov method. The statistical significance of the average
values obtained for biofilm biomass, biofilm activity and cell number were evaluated
using the t-test. Statistical calculations were based on confidence level equal to or
higher than 95% (P ≤ 0.05 was considered statistically significant).

The effect of enzymes in biofilm control 137 _________________________________________________________________________________
7.3 RESULTS
Several tests were performed in order to control B. cereus and P. fluorescens biofilms
using selected enzymes: β-glucanase, protease, lipase and α-amylase, alone and
combined with two QACs. An initial screening was performed using biofilms grown in
96-wells polystyrene microtiter plates (Figures 7.1 and 7.2). Afterwards, the enzymes
were tested against biofilms developed in a flow-cell system (Figure 7.3). The effects of
the enzymatic treatments were also assessed against planktonic cells (Figure 7.4).
THE EFFECT OF AN ENZYMATIC TREATMENT ON BIOFILM CONTROL
The 24 h-old biofilms formed in the microtiter plates were subjected to three different
types of treatments (1) 1 h exposure to an enzymatic solution, (2) 1 h exposure to an
enzymatic solution containing a QAC, and (3) exposure to an enzymatic solution for 30
min followed by 30 min exposure to a QAC (30 + 30). Control tests were performed with
PB as negative controls, and the QAC solution as the positive. The control treatments
were made to assess if the enzymes were acting in synergy with the QACs.
In Figure 7.1, biofilm killing and removal after the treatments with the duration of
1 h are depicted. The treatment with lipase resulted in no killing of B. cereus biofilms,
and the treatments with β-glucanase, protease, and α-amylase promoted low killing
(P > 0.05). The removal of B. cereus with the enzymatic solutions was low, with the
exception of α-amylase that was insignificant (P > 0.05). P. fluorescens biofilms killing
was insignificant with β-glucanase, protease and lipase (P > 0.05). The killing percentage
was low with α-amylase (P > 0.05). The enzymatic solutions of β-glucanase, protease
and α-amylase produced moderate removal (P < 0.05), while lipase caused low biofilm
removal (P > 0.05) (Figure 7.1).
Both biofilms of B. cereus and P. fluorescens showed susceptibility to BAC and
CTAB, when the biocides were applied alone. B. cereus killing was moderate with both
biocides (P < 0.05), and removal was low with BAC (P > 0.05) and insignificant with CTAB
(P > 0.05). P. fluorescens was insignificantly killed by both biocides (P > 0.05), and its
removal was low with BAC and insignificant with CTAB (P > 0.05).
The combination of the selected biocides with the enzymes increased, in most
cases, B. cereus and P. fluorescens killing percentage. The combined treatment BAC-
enzymes improved B. cereus killing from low to moderate (P < 0.05), however, with

138 Chapter 7
_________________________________________________________________________________
β-glucanase killing remained low (P > 0.05). With the addition of CTAB, the killing of
B. cereus was insignificant with α-amylase, and moderate with the other enzymes.
P. fluorescens biofilms killing with BAC-α-amylase was insignificant (P > 0.05), with BAC-
protease and BAC-lipase remained insignificant (P > 0.05), and with BAC-β-glucanase
increased to moderate (P < 0.05). Killing improved in all cases with CTAB: with protease,
lipase and α-amylase increased to low, and with β-glucanase increased to moderate
(P < 0.05). The removal of B. cereus with the combinations of BAC with all enzymes
remained low despite an increase, in average of 5% (P > 0.05). B. cereus removal with
CTAB improved slightly when combined with all enzymes (P > 0.05). The removal of
P. fluorescens with BAC was low when combined with lipase and α-amylase, and
moderate with β-glucanase and protease (P < 0.05). The removal of P. fluorescens with
the combination CTAB-enzymes was similar to the case when only enzymes were used
(P > 0.05).
Figure 7.1 Killing and removal percentages of B. cereus and P. fluorescens biofilms using the
selected enzymes with and without the selected QACs. Where corresponds to β-glucanase, protease, lipase, α-amylase and QAC. The enzymatic and QAC (biocide) solutions were applied for 1 h. *means no killing. Average values ± standard deviation for at least three replicates are illustrated.
The effect of the combinations of enzymes with biocides is antagonistic, when
biofilm killing was reduced in comparison with the tests with the enzymes or the biocide

The effect of enzymes in biofilm control 139 _________________________________________________________________________________
alone. This happened for B. cereus with the combinations of α-amylase with CTAB, and
β-glucanase with BAC. The effect is indifferent as happened for B. cereus biofilms
control using CTAB with β-glucanase, protease, and lipase, and for P. fluorescens using
BAC with protease. The protease was an additive to killing when was used along with
BAC on B. cereus.
The combination of enzymes with biocides was synergistic against B. cereus
biofilms when control was improved. This effect happened with all enzymes when
combined with BAC, except β-glucanase. P. fluorescens killing improved with the
synergistic combinations of BAC with β-glucanase and lipase, and of CTAB with all
enzymes tested. B. cereus removal with BAC and CTAB was indifferent with all enzymes.
P. fluorescens removal was indifferent with BAC, but the enzymes worked as additives
to CTAB.
The procedure that consisted in 30 minutes exposure to an enzymatic solution,
followed by the same period of exposure (30 + 30 min) to the QACs resulted in higher
killing and removal percentages (Figure 7.2).
Figure 7.2 Killing and removal percentages for B. cereus and P. fluorescens biofilms using the
selected enzymes. Where corresponds to β-glucanase, protease, lipase and α-amylase. The enzymatic solutions were applied for 30 min then removed and the biocide was applied for 30 min (30 + 30). Average values ± standard deviation for at least three replicates are illustrated.

140 Chapter 7
_________________________________________________________________________________
High killing percentages of B. cereus were observed with the pre-treatment of
β-glucanase and protease followed by BAC (P < 0.05), and for CTAB with all enzymes
tested (P < 0.05). The pre-treatments of lipase and amylase followed by BAC were
classified as moderate (P > 0.05). For P. fluorescens, biofilms killing with BAC combined
with all enzymes was low, except with protease which was moderate (P < 0.05). With
CTAB the killing efficacy was still low for all cases (P > 0.05). The removal increased in
comparison with the approach, when the treatments were applied for 1 h, for B. cereus
in all situations, and for P. fluorescens with BAC (all increased to moderate). Contrary to
when the treatments were applied for 1 h, biofilm removal was, on average, more
significant with BAC than with CTAB (B. cereus 41% vs. 20%; P. fluorescens 31% vs. 21%).
CONTROL OF BIOFILMS DEVELOPED IN THE FLOW CELL SYSTEM
P. fluorescens formed biofilms with the highest resistance to killing by QACs and
enzymes. This bacterium was selected to develop biofilms in a flow cell system in order
to understand the killing and removal efficacies of QACs and enzymatic treatments
against biofilms with characteristics mimicking those found in industrial systems. The
results before (control), immediately after 1 h exposure to the enzymatic and biocidal
solutions, and up to 24 h post treatment were compared in terms of CFU and biofilm
mass. In Figure 7.3 the biofilm mass reduction percentage and log CFU reduction are
represented for the different treatments.
The treatments with the enzymes resulted, in most cases, in biofilm removal.
β-glucanase, protease and α-amylase caused moderate biofilm removal, while lipase
resulted in insignificant biofilm removal, immediately after exposure. CTAB caused low
removal, 15% of total biofilm mass. When CTAB was applied in combination with the
enzymes, the removal was insignificant for protease and lipase, and moderate with
β-glucanase (57%) and α-amylase (36%), P < 0.05.
The application of enzymes caused log CFU reductions from 1 to 1.7. CTAB caused
a log CFU reduction of 1.3, immediately after treatment. When CTAB was combined
with the selected enzymes the efficacy of the treatment increased, except for the
combination with lipase which reduced the efficacy of the treatment (P < 0.05). The
CTAB-β-glucanase and CTAB-protease combinations caused the highest efficacy
increase (P < 0.05). The log CFU reduction between these treatments is statistically
similar (P > 0.05), and different from the treatment with the biocide (P < 0.05). The log

The effect of enzymes in biofilm control 141 _________________________________________________________________________________
CFU reduction was slightly higher for the combination CTAB-lipase than the reduction
caused by CTAB alone.
Figure 7.3 Mass and log CFU reduction of P. fluorescens biofilms overtime after the treatments
with an enzymatic solution (left hand) and an enzymatic solution combined with CTAB (right hand). Where corresponds to β-glucanase, protease, lipase, α-amylase and CTAB. *means no reduction. Average values ± standard deviation are depicted.
The biofilm behavior, in terms of biofilm mass and CFU numbers, was further
analyzed 2, 12 and 24 h following the treatment, after the initial biofilm growth
conditions were reestablished (Figure 7.3). Significant (P < 0.05) biofilm regrowth
(biofilm mass reduction percentage was lower than immediately after the treatment)
following the enzymatic treatment was found for α-amylase, 12 h after treatment. A
long-term biofilm removal effect (biofilm mass reduction percentage was higher than
immediately after the treatment) was found with lipase, 2 and 12 h after treatment,
α-amylase, 2 h after treatment and with β-glucanase and protease, 24 h after treatment
(P < 0.05). For the treatments with CTAB and CTAB-enzyme combinations, significant
(P < 0.05) biofilm mass regrowth was found on the biofilms treated with β-glucanase,
2 and 24 h after treatment, and CTAB-lipase and CTAB-α-amylase, 24 h after treatment.
Significant long-term effects were also found for CTAB, 2 and 12 h after treatment, for
CTAB-lipase and CTAB-α-amylase, 12 h after treatment.
In terms of CFU regrowth, following the enzymatic treatment, no significant
(P > 0.05) CFU increase was found. A long-term effect (P < 0.05) on CFU reduction was
found 2, 12 and 24 h after β-glucanase, and 2 h after α-amylase treatments. For the

142 Chapter 7
_________________________________________________________________________________
treatment of CTAB, alone, and combined with enzymes, regrowth was found 2 h after
the application of CTAB alone and CTAB-β-glucanase and CTAB-protease. This regrowth
behavior persisted for the 12 h (CTAB alone and in combination with protease) and
24 h (CTAB-β-glucanase) following treatment. Long-term effects (P < 0.05) in CFU
reduction were found for CTAB-lipase, 2, 12 and 24 h after treatment.
PLANKTONIC TESTS WITH ENZYMES
Using respirometry, the respiratory activity of B. cereus and P. fluorescens was studied
after the esposure to solutions of (1) enzymes, (2) biocides and (3) the combination of
both (Figure 7.4).
Figure 7.4 Effect of chemical treatment for 1 h of B. cereus (a) and P. fluorescens (b) planktonic
cultures. Control (no treatment). The different enzymes β-glucanase, protease, lipase, and α-amylase, were used alone and in combination with the two QACs.
The results obtained with BAC and CTAB alone solutions are represented by arrows. Total inactivation of respiratory activity is indicated with an asterisk (*). Average values ± standard deviation for at least three replicates are depicted.
The use of enzymatic solutions, for 1 h, on B. cereus suspensions resulted in a
decrease of the respiratory activity with all enzymes relatively to the control (no
treatment). The respiratory activity was reduced by approximately 30% with

The effect of enzymes in biofilm control 143 _________________________________________________________________________________
β-glucanase, 40% with lipase and 50% with both protease and α-amylase solutions. The
presence of enzymes did not affect the antibacterial activity of BAC and CTAB (P > 0.05),
with the exception of lipase (P < 0.05). In the presence of lipase, BAC at its MBC reduced
the respiratory activity to 92% (P < 0.05).
For P. fluorescens there were reductions of the respiratory activity with the
enzymatic solutions of protease (75%, P < 0.05) and lipase (30%, P < 0.05). No changes
were observed in the respiration of the cells treated with β-glucanase (P > 0.05). Albeit,
the exposure to α-amylase promoted cell activation (24%, P < 0.05) as the cells exposed
to this enzyme were more active than the cells with no treatment. No total reduction
of respiratory activity was observed with the combined solutions of BAC with protease
and lipase (P < 0.05). Moreover, the effects of CTAB on the bacterial respiratory activity
decreased when protease and α-amylase were present (P < 0.05).
7.4 DISCUSSION
The removal of B. cereus increased with the same enzymes. In this case, lipase was the
best for biofilm removal. P. fluorescens biofilm removal was best with the enzymatic
solution of protease, followed by β-glucanase, lipase and α-amylase solutions
(Figure 7.1). Oulahal-Lagsir et al. [30] found that protease, α-amylase, and β-glucanase
were effective in cleaning a simulated industrial biofilm formed during paper pulp
manufacture. In another study [31], a lipase was unsuccessful when tested on the
control of biofilm formed by a Pseudoalteromonas strain. In a study by Marcato-Romain
et al. [18] two lipases were inefficient, or only slightly efficient for microbial multi-
species biofilm removal. These results show that the efficiency of enzymatic treatments
is strongly dependent on the biofilm type, particularly the species colonizers and the
EPS they produce. Enzyme specificity is a fundamental stepping stone in designing an
enzyme-based control strategy. The specific mode of action of enzymes makes the
search for the correct enzymes for control challenging, because of the complex diversity
of biofilm constituents [49] that differs between biofilms [50].
The preliminary studies with microtiter plates suggest that the enzymes worked
synergistically with the biocides, even if biofilm killing and removal was modest. The
biofilms of B. cereus were more effectively removed by BAC than the biofilms formed
by P. fluorescens that were more effectively removed by CTAB. Simões et al. [32]

144 Chapter 7
_________________________________________________________________________________
showed that a P. fluorescens biofilms exhibited reduced susceptibility after a treatment
with BAC.
A previous study by Walker et al. [33] proposed that treating biofilms with only
one type of enzyme would only loosen the cells. These authors also suggested that
enzymatic soaking should be used as a pre-treatment prior to the use of biocides. This
corroborates the results obtained with the 30+30 min test performed in the present
study (the tests providing the highest biofilm removal results).
Jacquelin et al. [34] reported the use of enzymes with surfactants to improve
disinfection efficacy. However, the inclusion of enzymes is not certain to increase
efficacy, because there are several factors contributing to the success of such
treatment. Kim et al. [35] demostrated that an enzymatic treatment with proteinase K
and acylase I was not suitable against P. aeruginosa biofilms, as these were actually
increasing the amount of proteins present. Marcato-Romain [18] tested the effect of
concentration and contact time to discover that proteases were the best enzymes to
remove biofilms, and that glycosidases and lipases only slightly removed biofilms from
paper industry. It is known that exopolysaccharides and glycoproteins contribute to the
adhesion of bacteria to a surface and bacterial accumulation in the biofilm [30]. In
P. fluorescens biofilms α- and β- polysaccharides contribute to its cohesiveness [36].
Christensen et al. [36] studied the effect of alginate degrading enzymes to control pure
culture biofilms. These authors concluded that bacterial alginate does not contribute to
the cohesiveness of the biofilms tested. However, Johansen et al. [37] managed to
successfully remove Pseudomonas spp. biofilms attached to stainless steel using
polysaccharide hydrolysases. Lequette et al. [17] studied the effect of polysaccharidases
and proteolitic enzymes on a CIP procedure, using bacteria commonly found in food
industry. These authors found that proteolitic enzymes and polysaccharidases removed
P. fluorescens biofilms attached to stainless steel. These enzymes were tested in several
scenarios that differed from the current work, in parameters such as the pH and
temperature.
In the current work, the selected enzymatic treatments were able to reduce low
to moderately the flow-generated biofilms; however, this reduction is apparently lower
than the removal observed for the biofilms developed in the microtiter plates
(Figures 7.1 vs. 7.3 immediately after the treatment). The biofilms generated in the flow
cell system are older, thicker and with resistance characteristics (high EPS content and
cell density), see chapter 5. Flow-generated biofilms are known to be firmly attached to

The effect of enzymes in biofilm control 145 _________________________________________________________________________________
the surfaces enabling them to withstand the shear stress caused by the passing fluid
[38]. This result reinforces that the biofilm characteristics, particularly its age (7 days
old biofilms formed in the flow cell system vs. 1 day old biofilms formed in the microtiter
plates) are relevant in the control process. This comparison proposes that an adequate
biofilm formation system, with the ability to simulate the conditions found in practice
should be used in the development of control strategies.
The analysis of the flow generated biofilms following enzymatic and CTAB-enzyme
treatments was followed up to 24 h after treatment. The results demonstrated some
long-term effects on biofilm removal and CFU reductions. It is possible that the enzymes
or CTAB-enzyme caused sustained effects in biofilm control. Even if the solutions are
removed from the system, residual concentrations remain acting on the biofilms. Also,
regrowth was found following some treatments. However, the long-term and regrowth
effects found in this study were not specific for any particular enzyme and/or CTAB-
enzyme combination. Also, none of the strategies was effective in completely killing
and/or removing the flow generated biofilms. Parkar et al. [39] proposed that in
cleaning industrial plants, a large decrease of cells (removal and killing) is not indicative
of a successful treatment because the treatments leave cell debris that act as an organic
conditioning film able to assist microorganism attachment and regrowth.
The function of the biocide is to kill bacteria, while the enzymes can cause EPS
disruption. A part of biofilm resistance relies on internal mass transfer limitations
caused by the intricate nature of biofilm architecture. The polyanionic nature of the
bacterial EPS may be responsible for binding the antimicrobial agents before they have
the opportunity to reach the cells, hindering their diffusion [40-42]. The antimicrobial
agent can also react and be neutralized by components of the biofilms [43]. Augustin et
al. [44] points out the possibility that inside biofilms there are altered chemical
microenvironments able to inactivate enzymes. These authors also proposed that EPS
may be impairing the diffusion of enzymes through biofilms, similarly to how oxygen
does not fully penetrate biofilms [44]. The amount of enzymes available could be spent
before these reach the under-layers of the biofilms [44, 45]. In order to overcome this
issue, Pechaud et al. [46] allowed contact times of 20 hours to ensure total enzyme
penetration inside the biofilms, which is a long downtime for cleaning and disinfection
practices. Despite the long contact time, they observed that the enzymatic treatments
were not efficiently removing biofilms.

146 Chapter 7
_________________________________________________________________________________
The use of enzymes depends on several factors such as the type of biofilm to treat,
the surface to which it is attached, the contact time, the pH and the temperature [33].
The action of some agents could be antagonized by local accumulation of acidic waste
products that might lead to pH dissimilarities. Differences greater than 1, between the
bulk fluid and the biofilm interior, were previously found [44]. In this case, the neutral
pH was selected because it is the pH commonly used to grow the selected bacteria [47].
In fact, the effects of the enzymes could only be assessed if the parameters tested were
only the presence/absence of enzymes. Lequette et al. [17] found that alkaline buffers,
promoted biofilm removal.
The results obtained with respirometry, demonstrated some inhibition of
respiratory activity in the treatments where only enzymes were applied, except for
P. fluorescens treated with α-amylase (which potentiated modestly the respiratory
activity). However, when protease, lipase and α-amylase were applied with the QACs,
the respiratory activity values of B. cereus and P. fluorescens indicated that the bacteria
were in a viable state. In fact, the respiratory activity was higher than with the use of
BAC and CTAB alone. Therefore, no advantage on microbial inactivation was found for
these cases compared to the use of BAC and CTAB alone. These QACs were used at their
MBC values and caused total bacterial inactivation. The antimicrobial activity of
enzymes was already described [48]. In the last years many studies have been
performed on the antimicrobial properties of bacterial cell wall hydrolases [49]. The
hydrolysis of a sufficient number of specific bonds in the peptidoglycan layer results in
weakening, or serious cell damage that ultimately results in bacteriolysis [50]. For
instance, lysozyme is a bacteriolytic enzyme that hydrolyses polysaccharides which
compose the cell wall [30]. The hydrolases used in this study could be acting in the same
way. The exposure to protease resulted in high respiratory reduction of P. fluorescens
cells. The other enzymes were unable to produce the same effect, with the exception
of lipase that reduced the bacterial respiration in a low extent. This result is probably
due to the presence of the outer membrane in P. fluorescens (Gram negative) compared
with B. cereus (Gram positive).
It is currently known that environmental characteristics can influence the activity
of antimicrobial agents [51]. These hindrances can be caused by organic material that
potentially interferes with antimicrobial agents by chemical and/or ionic interactions
[9, 52]. The results from the present study, the combination of lipase with BAC resulted
in lower inactivation effects on B. cereus and that protease reduced the effects of BAC

The effect of enzymes in biofilm control 147 _________________________________________________________________________________
on P. fluorescens. The antimicrobial effects of CTAB on P. fluorescens were reduced with
protease and α-amylase. The work of Araújo et al. [21] (chapter 4) elucidates how the
antimicrobial function of BAC and CTAB antimicrobial is affected by low concentrations
of organic material such as bovine serum albumin (BSA), alginate, yeast extract and
humic acids. The antimicrobial mechanism of QAC involves the disruption and
denaturation of structural proteins and enzymes [53] that could lead to the impairment
of the enzyme function and activity. A similar phenomenon could have happened in the
present study. In the case of bacterial metabolism activation the enzymes might have
been taken as nutrients, since these could be carbon and nitrogen sources. Bacterial
activation was observed by Kim et al. [35] when using enzymatic treatments for biofilm
control. In this case, the release of fatty acids from bacterial EPS was promoted by the
treatments which in its turn were utilized by bacteria.
7.5 CONCLUSIONS
The preliminary studies with microtiter plates suggest that enzymes work synergistically
with the QACs, even if biofilm killing and removal was modest. Similar effects were
found on the treatments of P. fluorescens biofilms developed in the flow cell system.
Long term effects were observed apparently due the interaction between enzymes and
the biofilm components. On the other hand, for some treatments, the biofilms
recovered some of their characteristics over the course of 24 h after the treatments.
The enzymes were proven to work as antimicrobial agents against planktonic cells;
however, when combined with biocides, some enzymes acted as interfering agents
decreasing the activity of the QACs. The increase of concentration and contact time
could be a solution for the low efficacy rates; however, higher concentrations of
enzymes would be overly expensive, or the optimized contact time could be infeasible
for industrial applications.
REFERENCES
[1] Donlan R, Costerton J. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clinical Microbiology Reviews, 2002. 15(2):167-193.
[2] Flemming H. The EPS matrix: The "house of biofilm cells". Journal of Bacteriology, 2007. 189(22):7945-7947.
[3] Bryers J, Mixed Population Biofilms, in Biofilms - Science and Technology, LF Melo, et al., Editors. 1992. p. 277-289.

148 Chapter 7
_________________________________________________________________________________
[4] Shi X, Zhu X. Biofilm formation and food safety in food industries. Trends in Food Science & Technology, 2009. 20(9):407-413.
[5] Kumar CG, Anand SK. Significance of microbial biofilms in food industry: a review. International Journal of Food Microbiology, 1998. 42(1-2):9-27.
[6] Maukonen J, Mättö J, Wirtanen G, Raaska L, Mattila-Sandholm T, Saarela M. Methodologies for the characterization of microbes in industrial environments: a review. Journal of Industrial Microbiology & Biotechnology, 2003. 30(6):327-356.
[7] Sharma M, Anand SK. Biofilms evaluation as an essential component of HACCP for food/dairy processing industry - a case. Food Control, 2001. 13(6-7):469-477.
[8] Bremer PJ, Fillery S, McQuillan AJ. Laboratory scale Clean-In-Place (CIP) studies on the effectiveness of different caustic and acid wash steps on the removal of dairy biofilms. International Journal of Food Microbiology, 2006. 106(3):254-262.
[9] Otzen D. Protein-surfactant interactions: a tale of many states. Biochimica et Biophysica Acta - Proteins & Proteomics, 2011. 1814(5):562-591.
[10] Gibson H, Taylor J, Hall K, Holah J. Effectiveness of cleaning techniques used in the food industry in terms of the removal of bacterial biofilms. Journal of Applied Microbiology, 1999. 87(1):41-48.
[11] Johnston MD, Lambert RJW, Hanlon GW, Denyer SP. A rapid method for assessing the suitability of quenching agents for individual biocides as well as combinations. Journal of Applied Microbiology, 2002. 92(4):784-789.
[12] Verhoef R, Schols HA, Blanco A, Siika-aho M, Ratto M, Buchert J, Lenon G, Voragen AGJ. Sugar composition and FT-IR analysis of exopolysaccharides produced by microbial isolates from paper mill slime deposits. Biotechnology and Bioengineering, 2005. 91(1):91-105.
[13] Gurung N, Ray S, Bose S, Rai V. A broader view: microbial enzymes and their relevance in industries, medicine, and beyond. BioMed Research International, 2013. 2013:18.
[14] Fernandes P. Enzymes in food processing: a condensed overview on strategies for better biocatalysts. Enzyme Research 2010. 2010:862537.
[15] Ganeshnarayan K, Shah SM, Libera MR, Santostefano A, Kaplan JB. Poly-N-Acetylglucosamine matrix polysaccharide impedes fluid convection and transport of the cationic surfactant cetylpyridinium chloride through bacterial biofilms. Applied and Environmental Microbiology, 2009. 75(5):1308-1314.
[16] Andersson S, Dalhammar G, Land C, Kuttuva Rajarao G. Characterization of extracellular polymeric substances from denitrifying organism Comamonas denitrificans. Applied Microbiology and Biotechnology, 2009. 82(3):535-543.
[17] Lequette Y, Boels G, Clarisse M, Faille C. Using enzymes to remove biofilms of bacterial isolates sampled in the food-industry. Biofouling, 2010. 26(4):421-431.
[18] Marcato-Romain CE, Pechaud Y, Paul E, Girbal-Neuhauser E, Dossat-Letisse V. Removal of microbial multi-species biofilms from the paper industry by enzymatic treatments. Biofouling, 2012. 28(3):305-314.
[19] Simões M, Simões L, Pereira M, Vieira M. Antagonism between Bacillus cereus and Pseudomonas fluorescens in planktonic systems and in biofilms. Biofouling, 2008. 24(5):339-349.
[20] Simões M, Cleto S, Pereira M, Vieira M. Influence of biofilm composition on the resistance to detachment. Water Science and Technology, 2007. 55:473-480.
[21] Araújo PA, Lemos M, Mergulhão F, Melo L, Simões M. The influence of interfering substances on the antimicrobial activity of selected quaternary ammonium compounds. International Journal of Food Science, 2013:9.

The effect of enzymes in biofilm control 149 _________________________________________________________________________________
[22] Stepanović S VD, Dakić I, Savić B, Švabić-Vlahović M A modified microtiter-plate test for quantification of Staphylococcal biofilm formation. Journal of Microbiological Methods, 2000. 40(2):175-179.
[23] Simões L, Simões M, Vieira M. Influence of the diversity of bacterial isolates from drinking water on resistance of biofilms to disinfection. Applied and Environmental Microbiology, 2010. 76(19):6673-6679.
[24] Araújo PA, Mergulhão F, Melo L, Simões M. The ability of an antimicrobial agent to penetrate a biofilm is not correlated with its killing or removal efficiency. Biofouling, 2014:1-9.
[25] Lemos M, Borges A, Teodósio J, Araújo PA, Mergulhão F, Melo L, Simões M. The effects of ferulic and salicylic acids on Bacillus cereus and Pseudomonas fluorescens single- and dual-species biofilms. International Biodeterioration & Biodegradation, 2014. 86(Part A):42–51.
[26] Saavedra MJ, Borges A, Dias C, Aires A, Bennett RN, Rosa ES, Simoes M. Antimicrobial activity of phenolics and glucosinolate hydrolysis products and their synergy with streptomycin against pathogenic bacteria. Medicinal Chemistry, 2010. 6(3):174-183.
[27] APHA/AWWA/WPCF. Standard methods for the examination of water and wastewater, 17th edition, Clesceri, LS, Greenberg, AE and Trussel, RR, (eds). American Public Health Association, Washington DC, USA. 1989.
[28] Simões M, Pereira M, Vieira M. Validation of respirometry as a short-term method to assess the efficacy of biocides. Biofouling, 2005. 21(1):9-17.
[29] Perry RH, Green DW, Maloney JO. Perry's chemical engineers' handbook. 7th ed. New York: McGraw-Hill. 1999.
[30] Oulahal‐Lagsir N, Martial‐Gros A, Bonneau M, Blum LJ. “Escherichia coli‐milk” biofilm removal from stainless steel surfaces: Synergism between ultrasonic waves and enzymes. Biofouling, 2003. 19(3):159-168.
[31] Leroy C, Delbarre C, Ghillebaert F, Compere C, Combes D. Effects of commercial enzymes on the adhesion of a marine biofilm-forming bacterium. Biofouling, 2007. 24(1):11-22.
[32] Simões M, Pereira MO, Vieira MJ. Effect of mechanical stress on biofilms challenged by different chemicals. Water Research, 2005. 39(20):5142-5152.
[33] Walker SL, Fourgialakis M, Cerezo B, Livens S. Removal of microbial biofilms from dispense equipment: The effect of enzymatic pre-digestion and detergent treatment. Journal of the Institute of Brewing, 2007. 113(1):61-66.
[34] Jacquelin LF, Le Magrex E, Brisset L, Carquin J, Berthet A, Choisy C. Synergy effect of enzymes or surfactants in association with a phenolic disinfectant on a bacterial biofilm. Patholologie Biologie, 1994. 42425-431.
[35] Kim LH, Kim S-J, Kim C-M, Shin MS, Kook S, Kim IS. Effects of enzymatic treatment on the reduction of extracellular polymeric substances (EPS) from biofouled membranes. Desalination and Water Treatment, 2013:1-7.
[36] Christensen B, Ertesvag H, Beyenal H, Lewandowski Z. Resistance of biofilms containing alginate-producing bacteria to disintegration by an alginate degrading enzyme (AlgL). Biofouling, 2001. 17(3):203-210.
[37] Johansen C, Falholt P, Gram L. Enzymatic removal and disinfection of bacterial biofilms. Applied and Environmental Microbiology, 1997. 63(9):3724-8.
[38] Bryers J. Biofilms and the technological implications of microbial cell-adhesion. Colloids and Surfaces B-Biointerfaces, 1994. 2(1-3):9-23.
[39] Parkar SG, Flint SH, Brooks JD. Evaluation of the effect of cleaning regimes on biofilms of thermophilic bacilli on stainless steel. Journal of Applied Microbiology, 2004. 96(1):110-116.

150 Chapter 7
_________________________________________________________________________________
[40] Singh R, Ray P, Das A, Sharma M. Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. Journal of Antimicrobial Chemotherapy, 2010. 65:1955-1958.
[41] Anderl J, Franklin M, Stewart P. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Journal of Antimicrobial Chemotherapy, 2000. 44(7):1818-1824.
[42] Mah T, O'Toole G. Mechanisms of biofilm resistance to antimicrobial agents. Trends in Microbiology, 2001. 9(1):34-39.
[43] Simões M, Simões L, Vieira M. A review of current and emergent biofilm control strategies. LWT - Food Science and Technology, 2010. 43(4):573-583.
[44] Augustin M, Ali-Vehmas T, Atroshi F. Assessment of enzymatic cleaning agents and disinfectants against bacterial biofilms. Journal of Pharmacy and Pharmaceutical Sciences, 2004. 7(1):55-64.
[45] Xavier JB, Picioreanu C, Rani SA, van Loosdrecht MCM, Stewart PS. Biofilm-control strategies based on enzymatic disruption of the extracellular polymeric substance matrix - a modelling study. Microbiology, 2005. 151:3817-3832.
[46] Pechaud Y, Marcato-Romain CE, Girbal-Neuhauser E, Queinnec I, Bessiere Y, Paul E. Combining hydrodynamic and enzymatic treatments to improve multi-species thick biofilm removal. Chemical Engineering Science, 2012. 80:109-118.
[47] Simões M, Pereira MO, Vieira MJ. Action of a cationic surfactant on the activity and removal of bacterial biofilms formed under different flow regimes. Water Research, 2004. 39(2-3):478-486.
[48] Thallinger B, Prasetyo EN, Nyanhongo GS, Guebitz GM. Antimicrobial enzymes: An emerging strategy to fight microbes and microbial biofilms. Biotechnology Journal, 2013. 8(1):97-109.
[49] Parisien A, Allain B, Zhang J, Mandeville R, Lan CQ. Novel alternatives to antibiotics: bacteriophages, bacterial cell wall hydrolases, and antimicrobial peptides. Journal of Applied Microbiology, 2008. 104(1):1-13.
[50] Shockman G, Daneo-Moore L, Kariyama R, Massidda O. Bacterial walls, peptidoglycan hydrolases, autolysins, and autolysis. Microbial Drug Resistance, 2009. 2(1):95-98.
[51] Heinzel M. Phenomena of biocide resistance in microorganisms. International Biodeterioration & Biodegradation, 1998. 41(3-4):225-234.
[52] Aal SFAAE, Hunsinger B, Böhm R. Determination of the bactericidal activity of chemical disinfectants bacteria in dairies according to the DVG-guidelines. Hygiene & Medizin 2008. 33(11).
[53] Buffet-Bataillon S, Tattevin P, Bonnaure-Mallet M, Jolivet-Gougeon A. Emergence of resistance to antibacterial agents: the role of quaternary ammonium compounds—a critical review. International Journal of Antimicrobial Agents, 2012. 39(5):381-389.

CHAPTER 8 CONCLUDING REMARKS AND PERSPECTIVES FOR FURTHER RESEARCH

152 Chapter 8 _________________________________________________________________________________

Conclusions and future remarks 153 _________________________________________________________________________________
8.1 FINAL CONCLUSIONS
The results obtained in this thesis contribute to a better understanding of the
phenomena associated with biofilm resistance to cleaning and disinfection. These
studies also add information to develop cleaning and disinfection strategies to be
applied in industry. Biofilm control was shown to be extremely complex, because
embedded bacteria have dynamic resistance, enabling adaptation to varying
environmental and engineered conditions.
When resilient contaminations occur, they are often related to biofilms.
Furthermore, the embedded bacterial cells may gain additional resistance to
conventional control treatments. From the work presented in this thesis some major
conclusions can be drawn:
Antimicrobial agent penetration hindrances may be caused by the interaction of
the agents with the biofilm components, or by diffusional limitations caused by the
three-dimensional structure of the biofilm.
The ability of an antimicrobial agent to penetrate through a biofilm is not
correlated with biofilm killing or removal efficacy.
Selected QAC (BAC and CTAB) showed reduced antimicrobial efficacy in the
presence of BSA, YE and ALG. In the disinfection process of P. fluorescens, CTAB was
severely hindered by low concentrations of HA, that inclusively increased the metabolic
activity of this bacterium. The inclusion of HA substantially reduced the antimicrobial
efficacy of the QAC. This substance is proposed to be included as an antimicrobial
interfering agent for the testing protocols to develop disinfecting strategies.
The flow monitor system used for the development of flow generated biofilms at
different linear flow velocities, resulted in biofilms with clearly different characteristics
from those formed in microtiter plates. Nonetheless, both bioreactor systems
demonstrated to be suitable to perform biofilm control studies.
The flow regime influences biofilm development. The biofilms developed at the
highest linear flow velocities (u = 0.4 and 0.8 m.s-1) have similar characteristics, and are
different from those developed at the lowest linear flow velocity (u = 0.1 m.s-1).
Specifically, characteristics such as a higher occurrence of micro-tubular structures that
cells are thought to use to adhere to the stainless steel surface and to each other, and
more bacterial cells and EPS that were condensed in a more compact biofilm structure.

154 Chapter 8 _________________________________________________________________________________
The most complex biofilms, developed in the flow monitor system at the highest linear
flow velocities, were selected as a worst case scenario for the ensuing experiments on
biofilm control strategies.
BrCl and BrOH, CTAB and SH demonstrated the ability to change the cellular
membrane properties, changing hydrophobicity, and decreasing surface charge. The
action mechanism of BrCl and BrOH is thought to be cellular disruption, with pore
formation, as leakage of the intracellular constituents was observed.
BrOH, BrCl, CTAB and SH showed antimicrobial efficacy against planktonic
P. fluorescens. The MIC and MBC values of SH were similar to the values for BrCl and
BrOH, but the MIC and MBC of CTAB was the lower. Using the flow cell system, biofilm
control with CTAB, SH and BrCl was modest at the tested concentrations. Also, the
biofilms were able to recover after the treatments. The overall efficacy of BrCl and BrOH
in biofilm control was comparable to that of SH, proposing that these brominated-based
chemicals can be alternatives to SH. CTAB was the best antimicrobial agent.
The enzymatic treatments were able to reduce low to moderately the biofilm
quantity of both B. cereus and P. fluorescens developed in microtiter plates and in the
flow cell system. The enzymes showed synergistic potential with both BAC and CTAB.
The effects of the enzymatic treatments were observed for the biofilms developed in
the flow cell system. In the subsequent hours, the treatments alone and combined with
CTAB showed both long term effects and biofilm regrowth. These effects occurred with
no particular specificity to an enzyme or enzyme-CTAB treatment. Nonetheless, the
potential of the application of enzymes on biofilm control was found for the selected
bacteria.
When the enzymatic solutions were used on bacterial cell suspensions, all enzymes
showed antimicrobial activity against the bacteria tested, except β-glucanase and
α-amylase on P. fluorescens. The metabolism of this bacterium was stimulated when
α-amylase was applied. Nevertheless, when enzymes were combined with the selected
QACs, their antimicrobial potency was reduced. This phenomenon occurred when BAC
was combined with lipase for both bacteria, BAC combined with protease, and CTAB
combined with protease and α-amylase for P. fluorescens. A careful application of
enzymes must be considered since these molecules can quench the activity of the
antimicrobial agents.

Conclusions and future remarks 155 _________________________________________________________________________________
8.2 FUTURE WORK
Biofilms can be a problem in industries where water is involved in the manufacturing
process. The chemical control of biofilms is an important issue because of the severe
operation, management and public health impact.
Biofilm control is proven to be even more challenging than the approaches taken
against planktonic bacteria, however, the tests for chemicals efficacy are, most of the
times, performed on planktonic bacteria. The selection of a suitable antimicrobial agent
is of utmost importance for the development of a disinfection strategy. The
antimicrobial efficacy should be tested against particular contaminations, because the
loss of efficacy depends on many factors. Thus, it should be tested in conditions as close
to practice as possible, as in many cases, the influence of interfering agents could
severely hinder the antimicrobial efficacy. Therefore, it is proposed to widen the list of
disinfecting interfering substances, through the investigation of the mechanisms of
action of the antimicrobial agents and their interactions with different cellular targets
and soiling agents.
Industrial processes have a need for biocides able to retain their activity in soiled
conditions, work in low volumes, have low costs, and reduced corrosion. Particularly in
food industry, consumers and governmental agencies demand chemical agents that are
less toxic, less susceptible to microbial resistance, and stable so that disinfection by-
products do not enter natural systems. Therefore, anti-biofilm specific compounds
should be sought as alternative drugs with the function to selectively blocking virulence,
quorum sensing, and biofilm formation. Consequently, natural products such as
phytochemicals, have already been introduced into the market, however, in general,
their effects are limited compared to conventional disinfectants. In this work, the
combination of enzymes showed biofilm control potential, the addition/ combination
of phenolic or other new chemicals to potentiate the action of the conventional
antimicrobials is suggested as follow up research. Both strategies (combinations with
enzymes and new products) need optimization for complete control.
When the biofilms were scaled up to the flow cell system, it was stressed that the
way how biofilms develop is strongly connected with its degree of resistance. The study
of the process of biofilm formation is required from the early stages to maturation, by
a combined perspective of their physical, chemical and biological phenomena. When

156 Chapter 8 _________________________________________________________________________________
the correlation of the processing characteristics is made with contamination
occurrences, some solutions can be found: (1) the study of the effects that the
environmental parameters have on biofilm should be deepened and related with the
food processing line characteristics. Plant performance should be optimized to find an
equilibrium when production is maximized and the microbial contaminations are
minimized, (2) design materials with ability to inhibit soil accumulation must be sought,
and (3) the development of improved cleaning regimes, incorporated with
conventional/new but efficient chemical agents is in demand. A cleaning and
disinfection plan should be developed complying with certain principals: the nature of
the equipment (material and design), nature of the soiling agent, selection of a suitable
antimicrobial agent, and optimum operational conditions at which the agent has
maximum efficacy (temperature, concentration, hydrodynamics and exposure time).
These suggestions should be performed not only for the model bacteria used for this
thesis, but others such as Escherichia coli, Salmonella spp. or Listeria monocytogenes,
commonly found in food industry, and their combinations. Understanding the different
constituents that could emerge in food industry, will lead to a faster and more efficient
response, by tailoring treatments to specific situations.
New strategies are currently being researched and many more will appear as a
response to new resistance mechanisms or technological advances.

Nomenclature 157 _________________________________________________________________________________
NOMENCLATURE
ABBREVIATIONS
ALG alginate - APHA American Public Health Association - ATCC American Type Culture Collection - AVG average - AWWA American Water Works Association - BAC bezalkonium chloride - BDMDAC benzyldimethyldodecylammonium chloride - BrCl 3-bromopropionyl chloride - BrOH 3-bromopropionic acid - BSA bovine serum albumin - CDC Centers for Disease Control and Prevention - CFU colony forming units CFU.mL-1 CIP clean-in-place - CLSI Clinical and Laboratory Standards Institute - CTAB cetyltrimethylammonium bromide - DNA deoxyribonucleic acid - EB extraction buffer - EDTA ethylenediamine tetraacetate - EPS exopolymeric substances - GMP Good Manufacturing Practice - HA humic acids - HACCP Hazard Analysis and Critical Control Points - HDMS hexamethyldisilazane - LB Luiria Bertrani - MBC minimum bactericidal concentration µg.mL-1 MIC minimum inhibitory concentration µg.mL-1 NIH National Health Institute - OD optical density nm OMP outer membrane proteins - PB phosphate buffer - PCA plate count agar - QAC quaternary ammonium compound - QS quorum sensing - rRNA ribosomal ribonucleic acid - SD standard deviation - SDS-PAGE sodium dodecyl sulfate polyacrylamide gel electrophoresis SEM scanning electron microscopy - SH sodium hypochlorite - SPSS Statistical Package for the Social Sciences - TVS total volatile solids mg biomass. L-1 USA United States of America -

158 Nomenclature _________________________________________________________________________________
UV ultraviolet - WHO World Health Organization - WPCF Water Pollution Control Federation - YE yeast extract -
INDEXES
BI biofilm inactivation % BR biofilm removal % D molecular diffusivity of glucose m2.s-1 Dh hydraulic equivalent diameter m FIC fluorescence intensity of biofilms not exposed to
antimicrobial agents -
FIW fluorescence intensity value for biofilms exposed to the antimicrobial agents
-
km external mass transfer coefficient m.s-1 mc metabolic activity of the control experiments mg O2. mgorganic mass -1. min-1
mt metabolic activity of bacteria exposed to the antimicrobial
mg O2. mgorganic mass -1. min-1
ODC OD570nm value for biofilms not exposed to agents - ODW OD570nm value for biofilm exposed to the selected
chemicals -
Q flow rate m3.s-1 Re Reynolds number - Sc Schmidt number - Sh Sherwood number - u linear flow velocity m.s-1
GREEK
∆Gsws free energy of interaction between two entities mJ.m-2 γ− electron donor parameter mJ.m-2 γ+ electron acceptor parameter mJ.m-2 γAB Lewis acid-based component mJ.m-2
γLW Lifshitz-van der Waals component mJ.m-2
γTot total surface energy mJ.m-2 µ water viscosity kg.m-1.s-1 ɛ porosity - f Darcy friction factor - θ contact angle - ρ density of water Kg.m-3 ρd true density of dry biomass Kg.m-3 ρdw mass per unit of wet volume Kg.m-3 τ tortuosity - τw wall shear stress Pa