biodiversity of intertidal soft-sediment habitats in the ......biodiversity of intertidal...
TRANSCRIPT

Biodiversity of intertidal soft-sediment habitats in the Auckland Region
NIWA Client Report: HAM2009-097 June 2009 NIWA Project: DOC09302

All rights reserved. This publication may not be reproduced or copied in any form without the permission of the client. Such permission is to be given only in accordance with the terms of the client's contract with NIWA. This copyright extends to all forms of copying and any storage of material in any kind of information retrieval system.
Biodiversity of intertidal soft-sediment habitats in the Auckland Region Judi Hewitt Scott Edhouse Julia Simpson
NWA contact/Corresponding author
Judi Hewitt
Prepared for
Department of Conservation NIWA Client Report: HAM2009-097 June 2009 NIWA Project: DOC09302
National Institute of Water & Atmospheric Research Ltd Gate 10, Silverdale Road, Hamilton P O Box 11115, Hamilton, New Zealand Phone +64-7-856 7026, Fax +64-7-856 0151 www.niwa.co.nz

Contents Executive Summary iv 1. Introduction 1 2. Methods 5
2.1 Sample areas 5 2.2 Sampling 5 2.3 Habitat types 6 2.4 Diversity definitions 7 2.5 Diversity calculations 8
3. Average habitat diversity at the replicate scale 10 3.1 Southern Kaipara 10 3.2 Kawau Bay 10 3.3 Mahurangi Harbour 13 3.4 Weiti Estuary 13 3.5 Okura Estuary 13 3.6 Central Waitemata Harbour 17 3.7 Upper Waitemata Harbour 17 3.8 Whitford Embayment 20 3.9 Tamaki Strait 20
4. Species accumulation rates and total species richness of habitats 23 4.1 Southern Kaipara 23 4.2 Kawau Bay 24 4.3 Mahurangi Harbour 25 4.4 Karepiro Bay (Weiti and Okura Estuaries) 26 4.5 Waitemata Harbour (Central and Upper) 27 4.6 Whitford Embayment 28 4.7 Tamaki Strait 29
5. Effect of scale on diversity 31 5.1 Within-habitat heterogeneity 31 5.2 Habitat, site and replicate comparisons 35
6. Summary of results 40 6.1 Patterns apparent at the replicate core scale 40 6.2 Diversity at the habitat scale 40 6.3 Bullet point summary 41 6.4 Conclusions 42
7. Acknowledgements 43 8. References 44
Reviewed by: Approved for release by:
Carolyn Lundquist Neale Hudson
Formatting checked

Biodiversity of intertidal soft-sediment habitats in the Auckland Region iv
Executive Summary
The allocation of Marine Protected areas is a major conservation and management priority in New
Zealand. This task requires some determination of both representativeness and uniqueness of areas in
terms of their biodiversity and ecology. At present, while the role of habitats at a smaller-scale is
considered, the main focus of the classifications is on habitats described by environmental variables
(depth and substrate type) rather than including biotic factors. In part, this emphasis is driven by a
lack of information both about the effect of biogenic habitats on diversity and on the distribution of
such habitats. This project seeks to remedy the lack of information regarding key-habitat forming
species on biodiversity in the soft-sediment intertidal zone, a particular information gap identified by
the Biodiversity and Biosecurity Outcome Based Investment (OBI). The habitats used in the study
included both environmentally driven (i.e., those defined by the Ministry of Fisheries (MFish) and
Department of Conservation (DoC)) and biogenic habitats. Data was available for six habitats:
Cockles, Macomona, Mud, Sand (medium to fine particle size), Seagrass and Tubeworms.
We found no strong patterns in alpha diversity (i.e., the average number of species per core) between
the habitats that applied across all locations. However, Cockle habitats were more likely to have a high
alpha diversity, closely followed by Seagrass and Tubeworm habitats. Importantly, the habitats
described by sediment characteristics (as mud and sand) were most likely to have a low alpha
diversity.
Gamma diversity (i.e., the total number of species predicted for a habitat if 100 samples were
collected) also showed no consistent patterns between the habitats across locations. The Tubeworm
and Cockle habitats were most likely to have high gamma diversity, while the Mud habitat most
frequently had the lowest gamma diversity.
The differences we observe between the replicate and habitat scale, in which habitats have more
species richness, are driven by the within-habitat heterogeneity. This is controlled by the number of
infrequently occurring species (beta diversity). Two consistent patterns were observed: the Seagrass
habitat never exhibited the highest or lowest beta diversity; and within sites, the species found in
individual cores varied little for the Cockle habitat at all locations.
While we did not find consistent patterns of diversity among different habitats, our results suggest that
some species do provide a habitat for other species and can, therefore, affect biodiversity. In
particular, the New Zealand cockle, seagrass and the mats created by maldanid and polydorid
tubeworms. Location-specific variables, rather than regional species pools, seemed to be most
important for controlling the effect of the relationship between a habitat and its biodiversity. This
highlights the need to preserve a variety of habitats rather than concentrating on those habitats that are
considered to be of high biodiversity. Indeed preserving habitat diversity at a variety of scales is likely
to be the key to conserving biodiversity and ecosystem function.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 1
1. Introduction
Recently, the Ministry of Fisheries (MFish) and the Department of Conservation
(DoC) defined important habitat types for biodiversity conservation and mapping
(Ministry of Fisheries and Department of Conservation 2008). These were based on a
few sedimentary types and depth categories, e.g., mud, sand, intertidal, <20 m depth
etc. However, such broad-scale habitat definitions blur over the small-scale
preferences exhibited by many benthic macrofaunal species. Particularly in soft-
sediments, biogenic habitats can be important for biodiversity, with many rare species
exhibiting habitat specificity (Ellingsen et al. 2007, Hewitt et al. 2005, Thrush et al.
2006a).
There is surprisingly little New Zealand data available from which to determine which
habitat features of intertidal coastal areas are of particular importance to biodiversity.
Although there are many popular accounts that assert the importance of specific
biogenic habitats, few of these accounts have quantified diversity. Early work in New
Zealand focussed on description of assemblages (see Rowden et al. 2007 for a review)
rather than biodiversity estimates per se. Assessments of effects of marine reserves
generally focus on species-specific comparisons of abundance and size. Assessments
of biodiversity have been made in some marine areas of New Zealand, e.g.,
MacDiarmid (2006) and Rowden et al. (2007); however, these assessments are
focussed on specific groups of organisms and/or determining the location of
biodiversity hotspots, rather than habitat dependency.
This project supports the research on biodiversity, habitat diversity and species
richness in estuaries and coasts within the Biodiversity & Biosecurity OBI. Work
within the OBI determined that while knowledge of biodiversity of many New
Zealand marine habitats was generally lacking, knowledge of biodiversity in the
habitats occurring within intertidal soft-sediment areas was particularly sparse. The
habitats to be utilised were chosen to include both environmentally determined (i.e.,
those defined by MFish and DoC) and biogenic habitats that had been proven to have
effects on macrofaunal species, or that were considered to be important species for
other reasons: mangroves, seagrass meadows, cockle beds (Austrovenus stutchburyi),
tubeworm mats (Cummings et al. 1993, Thrush et al. 1996), Macomona liliana beds
(Pridmore et al. 1990, Thrush et al. 2006b), pipi beds (Paphies australis) and shrimp
burrows (Berkenbusch et al. 2007). Available data reduced this list of habitats to six:
cockles, Macomona, mud, sand (medium to fine particle size), seagrass and
tubeworms (Fig. 1.1).

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 2
Figure 1.1: The 6 habitat types used in the analysis. a: Cockle, b: Macomona showing feeding tracks on the surface, c: Mud, d: Sand, e: Seagrass and f: Tubeworm.
It is possible that the relationship between habitats and biodiversity is affected by both
regional species pools and local environmental characteristics. For example, Turner et
al. (1999) noted differences in the effect of seagrass on diversity of intertidal
macrofauna between Manukau and Whangapoua harbours. van Houte-Howes et al.
(2004) observed that, while there were differences between the composition of
seagrass patches with surrounding sand areas, differences in composition between

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 3
harbours was greater still. Lundquist et al. (2003) noted that while diversity of muddy
estuarine intertidal areas was generally expected to be lower than that of sandy areas,
location-specific sedimentation rates were an important factor. Sites with higher
sedimentation rates exhibited lower diversity, independent of grain size. For this
reason, only locations where most of the habitats present had been sampled were
considered in this project (Fig. 1.2). Furthermore, habitat types had to be defined at the
scale of the site prior to sampling of sediment. A search was made for coastal
locations for which the majority of their intertidal areas had been sampled in the past
10 years. Data from these locations were investigated to determine whether most of
the habitats were present and whether samples had been collected in a consistent
fashion. Because the aim of the study was the calculation of biodiversity, it was
imperative that data of high taxonomic resolution were used. This resulted in the
exclusion of some locations. In a few cases, locations had been sampled but not yet
analysed, and in another case the results had already been published (Alfaro 2006).
Figure 1.2: Coastal locations for which information was available.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 4
This report investigates the relationship between different types of diversity (species
richness, evenness and diversity indices) relative to key habitat types and habitat
forming species (mud, sand, seagrass, cockle beds, wedge shell beds and tube-worm
beds). Results are presented for nine North Island locations around the Auckland area:
Southern Kaipara, Kawau Bay, Mahurangi Harbour, Weiti Estuary, Okura Estuary,
Upper Waitemata Harbour, Central Waitemata Harbour, Whitford Embayment and
Tamaki Strait (Fig. 1.2). Insight gained from this analysis will be useful in:
• defining appropriate measures of biodiversity;
• illustrating the efficacy of rapid assessment techniques for determining
habitats; and
• demonstrating the value of maintaining habitat diversity.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 5
2. Methods
2.1 Sample areas
Intertidal soft-sediment benthic macrofauna information was available for 9 North
Island locations (Southern Kaipara, Tamaki Strait, Whitford Embayment, Okura
Estuary, Kawau Bay, Weiti Estuary, Mahurangi Harbour, and the Central and Upper
Waitemata Harbour (see Fig. 1.2)). All of these coastal areas are within 100 km of
each other. The Southern Kaipara is in a different regional species pool, being on the
west coast, while all the other locations are on the east coast of the North Island.
Sampling of each location had been carried out under different studies, often in
different years (Table 2.1), but all samples were collected using a 13 cm diam., 15 cm
deep corer.
Table 2.1: Information on the data available. ARC contact Grant Barnes, NIWA contact Judi Hewitt Marine Ecology group, Hamilton.
Location Area to mean high water (km2) Estuary Type Year(s) sampled Data holder
Southern Kaipara Harbour
440 Drowned Valley 2004 ARC
Kawau Bay 121.5 Coastal Embayment
2006 ARC
Mahurangi Harbour 24.57 Drowned Valley 1994 - ongoing
2005-6
ARC
NIWA
Okura Estuary 1.36 Tidal Lagoon 1999 NIWA
Weiti Estuary 2.83 Tidal Lagoon 2008 ARC
Upper Waitemata Harbour
Drowned Valley 2003-ongoing ARC
Central Waitemata Harbour
Drowned Valley 2000-ongoing
2006
ARC
NIWA
Whitford Embayment
11.06 Drowned Valley 2000
2000-1
ARC
NIWA
Tamaki Strait 335.36 Coastal Embayment
2002, 2007-8
2001, 2006
ARC
NIWA
2.2 Sampling
The number of replicates collected at a site varied from 3 to 12. Where more than 3
replicates were collected, 3 replicates were randomly chosen. In all studies, the
benthic macrofauna had been identified in the same laboratory to the same taxonomic

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 6
level (generally species) and enumerated. More importantly, different studies used
differing mesh sizes (either 1 mm or 0.5 mm). Fortunately, in the four locations for
which the 1 mm mesh was not the primary method there had been some studies using
both mesh sizes. This information was used to determine the species found when a
0.5 mm mesh was used that were not found when a 1 mm mesh was used, and these
species were removed from the data before analysis.
2.3 Habitat types
For all the studies used, the core data was accompanied by general habitat
descriptions, mainly consisting of biogenic features.
Six habitat types were common enough for analysis: seagrass meadows and patches
(Zostera muelleri), tube worm beds (most frequently the spionid polychaete Boccardia
syrtis, but also the maldanid polychaetes Macroclymenella stewartensis and Asychis
spp.), adult cockle beds (Austrovenus stutchburyi >20 mm longest shell dimension),
adult wedge shell beds (Macomona liliana >20 mm longest shell dimension),
unvegetated mudflats (>20% mud content) and unvegetated fine-sand (>80% fine-
medium sand) flats that did not contain sufficient densities of cockles, Macomona or
tubeworms to be allotted to one of these habitats. This definition of the Sand habitat
may result in low species richness simply because of the exclusion of cockles,
Macomona and tubeworms (common intertidal species) from their species lists.
Habitats based on fauna were defined as follows: Tubeworm >5 in a 20 x 20 cm
quadrat; Austrovenus > 5 in a quadrat; Macomona >5 feeding tracks in 2 quadrats. In a
few cases a site belonged to more than one faunal habitat type (e.g., cockles and
Macomona or tube worms-Macomona) and the site was allocated to both habitat
types. Seagrass habitats frequently contained cockles and Macomona but were
defined as Seagrass only, in order to aid comparisons with other published studies.
All of the habitats occurred at 4 of the locations. No seagrass occurred at Okura
Estuary, Weiti Estuary, Upper Waitemata Harbour and Whitford Embayment and
while a patch occurred in the Central Waitemata, it had not been sampled. The
Tubeworm habitat did not occur in Okura and the Macomona habitat was not found in
the Upper Waitemata Harbour. Generally, 3 – 9 sites had been sampled from each
habitat in each location but only one site was found for the Macomona habitat in the
Central Waitemata Harbour, the Sand habitat in Mahurangi Harbour and the
Tubeworm habitat in Weiti Estuary.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 7
A number of other habitats were recorded but either occurred infrequently in an area
or did not occur in many of the areas. It is also possible that a number of biogenic
habitats were not recognised as such when sampling.
2.4 Diversity definitions
Many methods exist for measuring biodiversity. Of these, the most used is species
richness or, more simply, the number of species. Even this simple measure can be
expressed in a number of ways: average number of species observed; total number of
species observed, Margalef’s species richness, and number of species predicted, using
species accumulation curves, from a set number of samples (Colwell & Coddington
1994), areas (Ugland et al. 2003) or habitats (Thrush et al. 2006a).
In both terrestrial and marine systems, most individuals belong to a few abundant
species and most species are represented by a small number of individuals. Thus
another way to consider biodiversity is how abundances are distributed amongst
species (Pielou’s evenness). Low evenness generally indicates dominance by a single
species.
A number of indices have been developed to combine both the number of species and
evenness. Of these, the most frequently used is the Shannon-Weiner index.
Shannon-Weiner is a logarithmic index and care must be taken when making
comparisons between studies to ensure that a similar log base has been used (here we
use loge). The second most frequently used is Simpson’s index, calculated simply
from probabilities of occurrence. The Shannon-Weiner index is more sensitive to rare
species, while Simpson’s index is more sensitive to more abundant species.
Measures can also be made at a number of different scales: alpha or gamma. The
frequency distributions of species observed are typically strongly right-skewed, with a
large number of species present in only a few samples. Thus much of biodiversity as
measured by species number is represented by rare (infrequently occurring species).
How these species are spatially distributed and what proportions they represent
primarily affect the difference between the average and total number of species. The
average number of species is often termed alpha diversity, the total number of
species, gamma diversity and the difference between the two, beta diversity (Crist et
al. 2003, Klimek et al. 2008, Lande 1996). The latter is a representation of how
heterogeneous the sampled area is. Note that species turnover along a gradient can
also be termed beta diversity; in this case it is generally calculated as the ratio of the
average to the total number of species (Ricotta 2008, Whittaker 1960).

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 8
2.5 Diversity calculations
A number of diversity measures were calculated at the replicate level: species
richness, evenness, Shannon-Weiner and Simpson index (Primer E, Clarke & Gorley
2006), from which the average at each site was reported. The statistical significance of
differences between habitats in each location was determined by one-way ANOVA
with habitat as a fixed factor. Averages and standard errors were calculated for each
habitat in each location.
Species richness and evenness were calculated at three spatial scales:
• The replicate (within-site) scale. Number of species and evenness of the
distribution of individuals across species were calculated for each replicate.
• The site scale. Number of species and evenness of the distribution of
individuals across species were calculated for the sum of the three replicates at
each site. Averages of these were then calculated for each habitat in each
location.
• The habitat scale.
o Evenness of the distribution of individuals across species was
calculated for the sum of the sites in each habitat in each location.
o Species richness was determined by prediction as it is highly
dependent on numbers of samples and each habitat had a different
number of samples, due to the different number of sites available for
each habitat. We derived species accumulation curves for each
habitat in each location using the commonly used Mao Tau estimation
(EstimateS, Colwell 2006). As few of the species accumulation
curves reached an asymptote (i.e., in most cases collecting more
samples continued to increase the number of species observed), we set
total species richness (gamma diversity) as the number of species
predicted to be found if 100 samples had been collected. N = 100 was
chosen, as the degree of separation between the curves had largely
stabilised by this point. Furthermore, for these comparisons it was
necessary to have as many samples per habitat as possible. For this
reason, Okura and Weiti estuaries which both flow into Karepiro Bay
were combined, and the Upper and Central Waitemata were also
combined.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 9
Beta diversity (heterogeneity of species richness occurring within each
habitat) was calculated as [total – average] species richness. Beta diversity
was calculated at two scales: within site; and within habitat.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 10
3. Average habitat diversity at the replicate scale
3.1 Southern Kaipara
In the southern Kaipara, the average number of species found was significantly lower
in the Mud and Sand habitats than the others (Fig. 3.1). Seagrass and Tubeworms had
the highest average number of species, though these differences were not statistically
significant due to high variability in these habitats. No differences were detected in
the average number of individuals observed between the habitats, due to the high
variability between sites. All habitats exhibited high evenness (>0.74), with the Mud
habitat the least even and the Tubeworm habitat the most even. The Tubeworm
habitat also had the highest Shannon-Weiner and Simpson index, within the habitats in
the Southern Kaipara, followed by the Seagrass habitat, while the Sand habitat had the
lowest Shannon-Weiner index and the Mud habitat had the lowest Simpson index.
3.2 Kawau Bay
In Kawau Bay, the average number of species found in the Sand habitat was
significantly lower than the number found in the Seagrass and Tubeworm habitats
(Fig. 3.2). These two habitats had the highest average number of species (11.5 and 9.4
respectively). Some differences were detected in the average number of individuals
observed between the habitats, with the Sand and Macomona habitats having the
lowest numbers and the Cockle habitat the highest. All habitats, except the Cockle
habitat, exhibited high evenness (>0.79), with the Tubeworm habitat again being the
most even. The Seagrass habitat had the highest Shannon-Weiner index and the
Tubeworm habitat had the highest Simpson index, while the Sand habitat again had
the lowest Shannon-Weiner index and the Cockle habitat had the lowest Simpson
index.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 11
Figure 3.1: Differences in diversity in Southern Kaipara Harbour based on six habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 12
Figure 3.2: Differences in diversity in Kawau Bay based on six habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 13
3.3 Mahurangi Harbour
In Mahurangi Harbour, the average number of species found was significantly lower
in the Seagrass and Tubeworm habitats than the Cockle habitat (Fig. 3.3). The Cockle
and the Sand habitats had the highest average number of species. No significant
differences were detected in the average number of individuals observed between the
habitats, due to the high variability between sites. All habitats exhibited high
evenness (>0.74), with the Mud habitat again the least even and the Seagrass habitat
the most even. The Cockle habitat had the highest Shannon-Weiner index and the
Seagrass had the highest Simpson index, while the Tubeworm habitat had the lowest
Shannon-Weiner and Simpson index.
3.4 Weiti Estuary
In Weiti Estuary, no Seagrass habitat was found. The average number of species found
was significantly higher in the Cockle habitat than in the all the other habitats (Fig.
3.4). No significant differences were detected in the average number of individuals
observed between the habitats, due to the high variability between sites. The Cockle
and the Macomona habitat exhibited a high evenness (> 0.74), while the remaining
habitats were less even. The Cockle habitat had the highest Shannon-Weiner
(statistically significant) and Simpson index, while the sand had the lowest Shannon-
Weiner and Simpson index.
3.5 Okura Estuary
In Okura Estuary, no Seagrass habitat was found and no Tubeworm habitats were
sampled. There were no significant differences between habitats in the average
number of species found due to high variability, however the Mud habitat exhibited
the lowest average number of species and the Cockle habitat exhibited the highest
average number of species (8.3 and 11.1 respectively), as shown in Fig. 3.5. Some
differences were also detected in the average number of individuals observed between
the habitats, with the Mud habitats having the lowest numbers and the Cockle habitat
the highest. All habitats exhibited high evenness > 0.74, with the Cockle habitat the
least even and the Mud habitat the most even. The Macomona habitat had the highest
Shannon-Weiner and Simpson index (although not significantly so).

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 14
Figure 3.3: Differences in diversity in Mahurangi Harbour based on six habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 15
Figure 3.4: Differences in diversity in Weiti Estuary based on five habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 16
Figure 3.5: Differences in diversity in Okura Estuary based on four habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 17
3.6 Central Waitemata Harbour
While a small area of seagrass does occur in the Central Waitemata Harbour area, this
had not been sampled. In the Central Waitemata Harbour, the average number of
species found in the Mud habitat was significantly lower than the others (Fig. 3.6).
The Cockle, Macomona and Tubeworm habitats had the highest average number of
species. Again no significant differences were detected in the average number of
individuals observed between the habitats, although the Mud and Sand habitats had
the lowest numbers and the Cockle habitat the highest. All habitats, except the Cockle
and Tubeworm habitat, exhibited high evenness, with the Sand habitat most even. The
Sand habitat also had the highest Shannon-Weiner and Simpson indices, followed by
the Macomona habitat, while the Mud habitat had the lowest Shannon-Weiner and
Simpson indices.
3.7 Upper Waitemata Harbour
In the Upper Waitemata Harbour, Seagrass and Macomona habitats are absent. The
Cockle habitat had the highest number of species and individuals (Fig. 3.7). However,
the Cockle habitat exhibited low evenness (<0.7); all others had evenness values
between 0.70 and 0.76. The Cockle habitat also exhibited a significantly higher
Shannon-Weiner diversity index than the Mud habitat, but there were no significant
differences in Simpson index between the four habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 18
Figure 3.6: Differences in diversity in Central Waitemata Harbour based on five habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 19
Figure 3.7: Differences in diversity in Upper Waitemata Harbour based on four habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 20
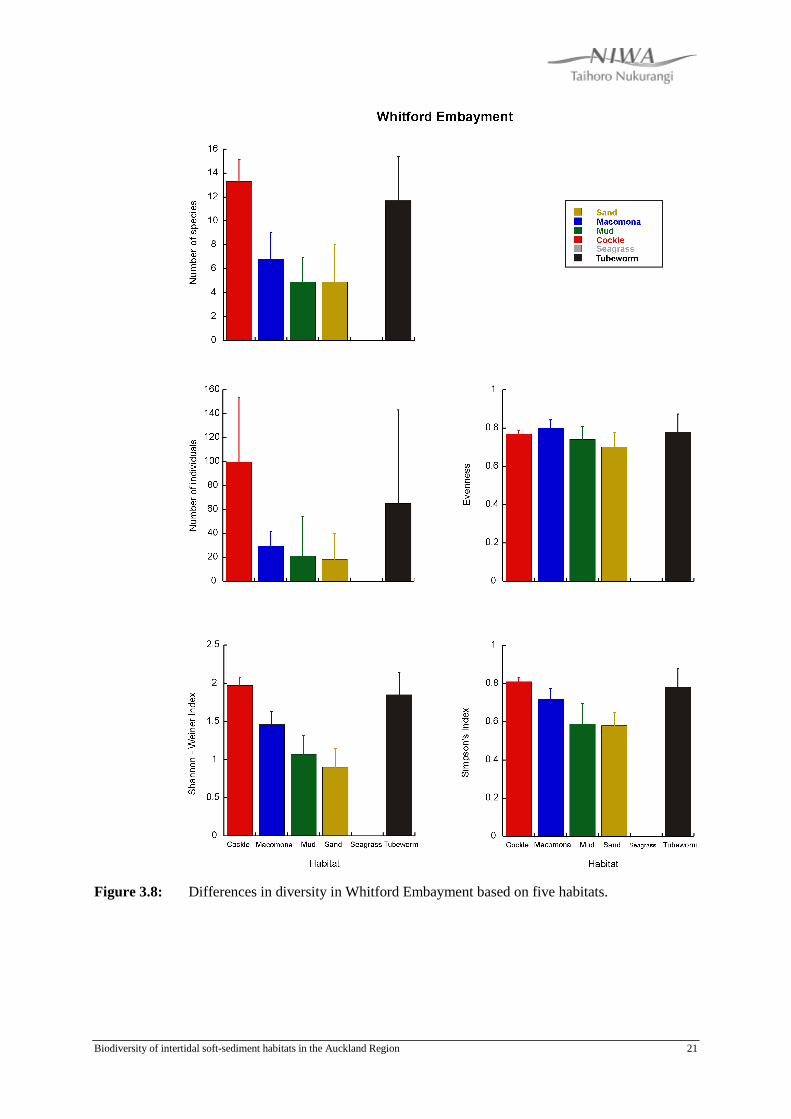
3.8 Whitford Embayment
Seagrass habitat is absent from the Whitford Embayment. The average number of
species found was significantly higher in the Cockle habitat than in the Macomona,
Mud and Sand habitats (Fig. 3.8). The average number of species found in the
Tubeworm habitat was not significantly higher than the other habitats due to high
variability. No differences were detected in the average number of individuals
observed between the habitats, due to the high variability observed in the Cockle and
Tubeworm habitats, although number of individuals was on average lower in the
Macomona, Sand and Mud habitats. Evenness ranged between 0.70 - 0.82, with the
Macomona habitat the most even. The Cockle habitat had the highest Shannon-Weiner
and Simpson index, followed by the Tubeworm habitat. The Sand and Mud habitats
had significantly lower Shannon-Weiner indices than the Cockle and Tubeworm
habitats, and also had the lowest Simpson index.
3.9 Tamaki Strait
In the Tamaki Strait, no significant differences were observed between habitats in the
average number of species found, with the average ranging between 6 and 10 (Fig.
3.9). No significant differences were detected in the average number of individuals
observed between the habitats, although the averages ranged between 18 and 50, due
to the high variability between sites. All habitats, except the Cockle habitat, exhibited
high evenness (> 0.74), with the Sand habitat being the most even. The Macomona
and the Mud habitats exhibited significantly higher Shannon-Weiner indices than the
Sand habitat and a significantly higher Simpson index than all the other habitats.
Biodiversity of intertidal soft-sediment habitats in the Auckland Region 21
Figure 3.8: Differences in diversity in Whitford Embayment based on five habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 22
Figure 3.9: Differences in diversity in Tamaki Strait based on six habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 23
4. Species accumulation rates and total species richness of habitats
Species accumulation curves for each habitat in each location were derived as
described in the Methods using Mao Tau estimation. In Section 4, we describe the
behaviour of these species accumulation curves for the number of samples collected at
each site. These curves are used to estimate total species richness (gamma diversity)
for each habitat by extrapolating these species accumulation curves out to 100 samples
for each location.
4.1 Southern Kaipara
The species accumulation curves for the habitats in Southern Kaipara show that the
Seagrass habitat has the greatest number of species present (Fig. 4.1). This is followed
by the Tubeworm habitat, then Cockle and Macomona habitats. The Mud and
Macomona habitats initially increase at a greater rate than the Sand habitat but appear
to be reaching an asymptote earlier than the Sand habitat, which continues to increase
at a rate that will result in higher species richness than the Macomona and Mud
habitats.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 24
Figure 4.1: Species accumulation curves for the different habitats in Southern Kaipara Harbour.
4.2 Kawau Bay
In Kawau Bay the Tubeworm habitat showed the greatest accumulation rate of species
(Fig. 4.2). The Cockle and Seagrass habitats have the next highest accumulation rates,
although the Seagrass habitat is reaching an asymptote while the Cockle habitat is
continuing to accumulate more species. The Mud habitat increased faster than the
Sand habitat at first but was flattening out, while the Sand habitat continued to
accumulate new species. The small number of samples taken in the Macomona
habitat makes it difficult to reach any conclusion, although it seems likely that the
species accumulation asymptote would lie between that of the Cockle and Sand
habitats (similar to the Southern Kaipara).

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 25
Figure 4.2: Species accumulation curves for the different habitats in Kawau Bay.
4.3 Mahurangi Harbour
In this location, the Cockle habitat exhibited the highest species accumulation rate
(Fig. 4.3). The number of species found in the Macomona habitat increased quickly as
well, but approached an asymptote earlier. Interestingly, the Mud habitat exhibited a
high species accumulation rate in this location, with a predicted maximum at 100
samples higher than that of the Macomona habitat. The predicted maximum of the
Tubeworm habitat at 100 samples is greater than that of both the Macomona or Mud
habitats. The few samples taken in the Seagrass and Sand habitats make it difficult to
assess their species accumulation rates accurately, although initially the Sand habitat
species accumulation curve is similar to that of the Macomona habitat while the
Seagrass habitat lies a little below that of the Mud habitat.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 26
Figure 4.3: Species accumulation curves for the different habitats in Mahurangi Harbour.
4.4 Karepiro Bay (Weiti and Okura Estuaries)
The Cockle habitat showed the greatest rate of species accumulation in Karepiro Bay
(Fig 4.4). The Mud habitat initially had the second highest rate, but this decreased
rapidly after 5 samples had been collected, while both the Sand and Macomona
habitats continued to accumulate species at high rates. The Tubeworm habitat
exhibited the lowest species accumulation rate for this location and there is no
Seagrass habitat here.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 27
Figure 4.4: Species accumulation curves for the different habitats in Karepiro Bay (Okura and Weiti Estuary).
4.5 Waitemata Harbour (Central and Upper)
In Waitemata Harbour, the Tubeworm habitat showed the greatest rate of species
accumulation, and does not appear to be reaching asymptote (Fig 4.5). The rate from
the Cockle habitat is initially higher but began to decrease after 15 samples. The Mud
habitat initially had a higher species accumulation rate than the Sand habitat but this
rate began to decrease faster than the Sand habitat after 30 samples. While the
Macomona habitat started accumulating at a rate similar to the Cockle and Tubeworm
habitats, the limited number of samples from this habitat makes it difficult to predict
accumulation rates. There is very little seagrass habitat in the Waitemata Harbour and
the few patches observed were not included in this analysis.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 28
Figure 4.5: Species accumulation curves for the different habitats in Waitemata Harbour.
4.6 Whitford Embayment
In the Whitford Embayment no Seagrass habitat was sampled. The Tubeworm habitat
displayed the greatest rate of species accumulation (Fig. 4.6). The Cockle habitat
initially exhibited the highest species accumulation rate, but this decreased markedly
after 5 samples had been collected. As a consequence the maximum number of species
predicted for 100 samples is higher for the Sand habitat. The Macomona habitat
followed a similar curve to the Cockle habitat at a lower level and the curve for the
Mud habitat was approximately parallel to that of the Macomona Habitat, but at a
lower level.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 29
Figure 4.6: Species accumulation curves for the different habitats in Whitford Embayment.
4.7 Tamaki Strait
In Tamaki Strait the Tubeworm habitat displayed the greatest rate of species
accumulation (Fig 4.7). The Mud, Cockle and Macomona habitats were next highest,
with the Sand and Seagrass habitats having the lowest rates.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 30
Figure 4.7: Species accumulation curves for the different habitats in Tamaki Strait.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 31
5. Effect of scale on diversity
5.1 Within-habitat heterogeneity
In the estimation of biodiversity, spatial scale is considered very important, with a
general increase in biodiversity estimates as we move from estimation at a replicate to
a regional scale. This increase in biodiversity with increase in spatial scale is
represented by the difference between the average species richness and the total
species richness of all the replicates combined. For species richness, this difference is
entitled beta diversity - the degree of heterogeneity. Beta diversity is crucially
important as it is largely a reflection of the number of rare (infrequently occurring)
species. Rare species, while having low numbers of individuals per species, generally
make up the largest proportion of the number of species. Thus, when making
comparisons, beta diversity is frequently converted to a percentage of the total
diversity.
In Section 5.1 we compare the proportion of the total species richness (gamma
diversity) represented by the beta diversity with that represented by the average
species richness (alpha diversity) (Fig. 5.1 & 2). In all cases, beta diversity is greater
than alpha diversity. In Section 5.2, we differentiate the beta diversity into that
occurring at the site scale (betasite) and that at the habitat scale (betahab), making
comparisons between them and the average species richness in a replicate core. This
indicates the scale at which we gain the most species.
In the Southern Kaipara Harbour, the proportion of beta diversity is always much
greater than the alpha diversity (87 - 92% Fig. 5.1). Within this narrow range, the
Mud and Sand habitats have the highest beta diversity, possibly due to the wider range
of communities that may inhabit these broad definitions.
In Kawau Bay, the proportion of beta diversity was again much higher than the alpha
diversity (81-90%, Fig. 5.1). Again the Sand habitat had the highest proportion of
beta diversity, but in this location the Tubeworm habitat also had a high proportion of
beta diversity.
In the Mahurangi Harbour, the range of proportions of beta diversity was small (80-
86% Fig. 5.1), with the exception of the Sand habitat for which very few samples had
been collected. The Tubeworm habitat had the highest proportion of beta diversity.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 32
In the Weiti Estuary (Fig. 5.1), the proportion of beta diversity was variable (80 –
91%), although still much greater than the proportion represented by alpha diversity,
with the highest proportion occurring in the Sand habitat.
However, in the Okura Estuary (Fig. 5.1), the proportion of beta diversity varied little
between the habitats (81 – 84%), with the exception of the Macomona habitat for
which few samples had been collected. The proportion of beta diversity was again
much higher than the alpha diversity and this result held for all locations.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 33
Figure 5.1: Percent alpha (i.e., average number of species in a replicate core) and beta diversity (within-habitat heterogeneity) for the different habitats in the Southern Kaipara, Kawau Bay, Mahurangi Harbour and Weiti and Okura Estuaries.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 34
In the Central Waitemata, the proportion of beta diversity varied between 80 – 88%
(Fig. 5.2). Similar to the Southern Kaipara the highest proportion occurred in the Mud
habitat.
In the Upper Waitemata, the proportion of beta diversity varied from 82 – 92% (Fig.
5.2), with the lowest proportion in the Cockle habitat and the highest proportions in
the Tubeworm and Mud habitats.
Figure 5.2: Percent alpha (i.e., average number of species in a replicate core) and beta diversity (within-habitat heterogeneity) for the different habitats in Central and Upper Waitemata Harbours, Whitford Embayment and the Tamaki Strait areas.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 35
In Whitford (Fig. 5.2), the Cockle habitat exhibited the lowest proportion of beta diversity (78%) while the Sand habitat had the highest (90%).
In Tamaki Strait (Fig. 5.2), the proportion of beta diversity was always high and
varied little (84 -89%); however, the Sand habitat had the highest proportion of beta
diversity.
5.2 Habitat, site and replicate comparisons
Here, we use the species accumulation curves to estimate total species richness at site
and habitat scales. We then calculate beta diversity as the increase in species richness
at the site (betasite) and habitat scales (betahab) when comparing between average
species richness in individual replicates and total species richness predicted to be
found in 100 samples for each site and habitat. Betasite refers to increases in diversity
when adding more replicates within a site. Betahab refers to increases in diversity
when adding more habitats.
For all habitats in all locations, the greatest gain in number of species occurred when
more habitats were sampled (betahab Table 5.1). The next greatest gain occurred at
the individual replicate level (alpha Table 5.1). The number of extra species being
added as more replicates were added at a site was generally low (betasite Table 5.1).
However, this did not hold for three of the habitats in the Kaipara (Mud, Sand and
Seagrass), or for Tubeworms and Mud habitats in the Upper Waitemata, Tubeworms
and Macomona habitats in Tamaki Strait, or Tubeworm and Sand habitats in Whitford.
These habitats all exhibited high variation between replicates at a site in the species
found, increasing the proportion of within-site beta diversity (betasite Table 5.1).

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 36
Table 5.1: Comparison of the increase in species richness as a proportion of the species richness predicted to be found in 100 samples for each habitat and location. Alpha = average % found in individual replicates. Betasite = % increase to average found in a site. Betahab = % increase to total found in a habitat.
Cockle Macomona Mud Sand Seagrass Tubeworm
Sth Kaipara alpha 12 13 9 8 10 12
betasite 9 9 8 8 8 9
betahab 79 78 83 84 82 79
Kawau alpha 13 16 14 10 19 12
betasite 10 13 10 9 15 8
betahab 77 71 76 82 67 80
Mahurangi alpha 18 17 19 32 17 14
betasite 12 8 8 16 14 6
betahab 70 74 73 52 69 80
Weiti alpha 18 13 17 9 20
betasite 11 5 10 7 15
betahab 71 82 74 84 64
Okura alpha 16 23 19 17
betasite 8 15 13 13
betahab 76 62 69 70
C.Waitemata alpha 20 20 12 18 18
betasite 9 14 8 14 12
betahab 71 66 79 68 70
U.Waitemata alpha 18 9 11 8
betasite 11 7 7 6
betahab 71 84 82 86
Tamaki alpha 16 15 14 11 17 15
betasite 13 13 11 8 24 13
betahab 71 73 75 81 59 72

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 37
Cockle Macomona Mud Sand Seagrass Tubeworm
Whitford alpha 22 15 14 10 13
betasite 10 11 10 10 11
betahab 68 73 75 80 76
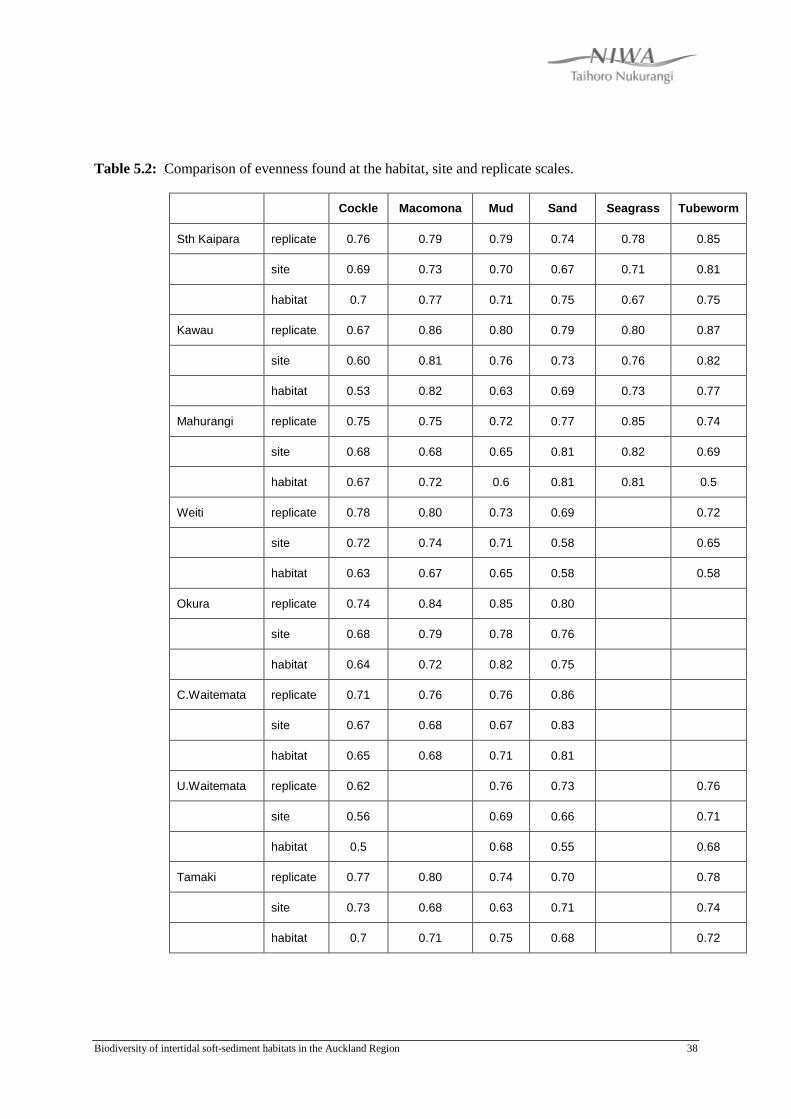
Generally the evenness with which individuals were distributed across species was
greatest at the replicate scale with the least even distribution occurring at the habitat
scale (Table 5.2). Some exceptions to this occurred for all habitat types and for all
locations, with the exception of the Upper Waitemata. In Southern Kaipara, the
habitat scale exhibited greatest evenness for the Sand habitat and moderate evenness
for the Cockle and Macomona habitats. Greatest evenness occurred at the site and
habitat scale for the Sand habitat in Mahurangi, at the habitat scale for the Mud habitat
in Whitford, and at the site scale for the Seagrass habitat in Tamaki. Greater evenness
was also found for the habitat scale rather than the site scale for the Mud habitat in
Okura and Central Waitemata, for the Macomona habitat in Kawau, Mahurangi and
Whitford, and for the Tubeworm habitat in Tamaki.
Biodiversity of intertidal soft-sediment habitats in the Auckland Region 38
Table 5.2: Comparison of evenness found at the habitat, site and replicate scales.
Cockle Macomona Mud Sand Seagrass Tubeworm
Sth Kaipara replicate 0.76 0.79 0.79 0.74 0.78 0.85
site 0.69 0.73 0.70 0.67 0.71 0.81
habitat 0.7 0.77 0.71 0.75 0.67 0.75
Kawau replicate 0.67 0.86 0.80 0.79 0.80 0.87
site 0.60 0.81 0.76 0.73 0.76 0.82
habitat 0.53 0.82 0.63 0.69 0.73 0.77
Mahurangi replicate 0.75 0.75 0.72 0.77 0.85 0.74
site 0.68 0.68 0.65 0.81 0.82 0.69
habitat 0.67 0.72 0.6 0.81 0.81 0.5
Weiti replicate 0.78 0.80 0.73 0.69 0.72
site 0.72 0.74 0.71 0.58 0.65
habitat 0.63 0.67 0.65 0.58 0.58
Okura replicate 0.74 0.84 0.85 0.80
site 0.68 0.79 0.78 0.76
habitat 0.64 0.72 0.82 0.75
C.Waitemata replicate 0.71 0.76 0.76 0.86
site 0.67 0.68 0.67 0.83
habitat 0.65 0.68 0.71 0.81
U.Waitemata replicate 0.62 0.76 0.73 0.76
site 0.56 0.69 0.66 0.71
habitat 0.5 0.68 0.55 0.68
Tamaki replicate 0.77 0.80 0.74 0.70 0.78
site 0.73 0.68 0.63 0.71 0.74
habitat 0.7 0.71 0.75 0.68 0.72

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 39
Cockle Macomona Mud Sand Seagrass Tubeworm
Whitford replicate 0.71 0.86 0.84 0.89 0.79 0.77
site 0.64 0.80 0.77 0.84 0.84 0.72
habitat 0.62 0.7 0.71 0.7 0.77 0.73

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 40
6. Summary of results
6.1 Patterns apparent at the replicate core scale
Alpha diversity (i.e., the average number of species per core) showed no strong
patterns between the habitats that applied across all locations. However, Cockle
habitats were more likely to have a high alpha diversity, closely followed by Seagrass
and Tubeworm habitats. Importantly, the habitats described by sediment
characteristics (as mud and sand) were most likely to have a low alpha diversity.
Shannon Weiner and Simpson indices were even less likely to show consistent
patterns across locations, in terms of ranking the habitats. The Cockle habitat was
ranked highest for Shannon-Weiner diversity three times (Weiti Estuary, Upper
Waitemata and Whitford). The Macomona and Sand habitats were both ranked
highest for Shannon-Weiner diversity twice, but the Macomona habitat was also
ranked lowest once and the Sand habitat was ranked lowest in all locations but
Mahurangi and Central Waitemata. The only habitat that never exhibited a high
ranking for Shannon-Weiner diversity was the Mud habitat. For the Simpson index,
the Tubeworm, Cockle and Sand habitats ranked highest in 2 locations each, they each
also ranked lowest in 2 locations. Even the Mud habitat ranked second highest in one
location (Tamaki Strait).
Very few differences were observed in the average number of individuals found in a
replicate, due mainly to high variability in all habitats and locations. Generally the
Tubeworm habitat exhibited the greatest evenness. Interestingly, the Cockle habitat
was the only habitat that never exhibited the highest evenness.
6.2 Diversity at the habitat scale
Gamma diversity (i.e., the total number of species predicted to be found in a habitat if
100 samples were collected) also showed no consistent patterns between the habitats
across locations. The Tubeworm and Cockle habitats were most likely to have high
gamma diversity, while the Mud habitat most frequently had the lowest gamma
diversity.
The differences we observe between the replicate and habitat scale in which habitats
have more species richness are driven by the within habitat heterogeneity. This is
controlled by the number of infrequently occurring species (beta diversity). The only
consistent pattern between habitats across sites was that the Seagrass habitat never

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 41
exhibited the highest or lowest beta diversity. Beta diversity was generally high
(>80% of the total number of species), with very little difference occurring between
locations.
For all habitats in all locations, the greatest gain in number of species occurred when
more sites and/or habitat patches were sampled (Table 5.1). Within sites, the species
found in individual cores varied little for the Cockle habitat in all locations. Other
habitats in the Southern Kaipara and the Sand and Tubeworm habitats in other
locations were likely to have more variation between replicates at a site in the species
found, increasing the proportion of within-site beta diversity.
Generally the evenness with which individuals were distributed across species was
greatest within replicates and this decreased to a less even distribution within sites
with another decrease at the habitat scale.
6.3 Bullet point summary
• No habitat had highest diversity across all locations.
• Cockle habitats were most likely to have highest average number of species at
a site, closely followed by Seagrass and Tubeworm habitats.
• Mud habitats were most likely to have low Shannon Weiner and Simpson
indices.
• The Tubeworm habitat was most likely to have high evenness.
• The Tubeworm and Cockle habitats were most likely to have highest predicted
number of species, while the Mud habitat most frequently had the lowest.
• The Seagrass habitat never exhibited the highest or lowest beta diversity.
• Beta diversity was generally high (>80% of the total number of species), with
very little difference occurring between locations. The greatest gain in number
of species occurred when more habitat patches were sampled.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 42
6.4 Conclusions
At present, conservation and management strategies in New Zealand are focussing on
the allocation of Marine Protected Areas. This task requires some determination of
representativeness versus uniqueness of areas in terms of their biodiversity and
ecology. As little information is generally available with which to select specific
areas, New Zealand has developed a classification based on environmental variables
for offshore areas (Snelder et al. 2006) and a biogeographical classification for both
offshore and coastal areas (Ministry of Fisheries and Department of Conservation
2008). While the role of habitats at a smaller-scale is considered, the main focus of the
classifications is on habitats described by environmental variables (depth and substrate
type) rather than including biotic factors (Ministry of Fisheries and Department of
Conservation 2008).
Our results suggest, however, that some species do provide a habitat for other species
and can, therefore, affect biodiversity. The New Zealand cockle, seagrass and the
mats created by maldanid and polydorid tubeworms all created habitats more likely to
have higher average numbers of species (alpha diversity) than the other habitats.
Cockles and tubeworm mats also created habitats likely to have higher total numbers
of species (gamma diversity). Our results are consistent with other published data.
For example, Macomona has been demonstrated to have a negative effect on a
number of species (Thrush et al. 2006b, Thrush et al. 1992). Similarly we did not find
this habitat to have high species richness. Seagrass is internationally considered to be
a habitat of high biodiversity. However similar to our findings, published New
Zealand studies show variable results. Henriques (1980) showed that seagrass habitats
in the Manukau Harbour had higher species diversity than comparable non-vegetated
habitat. Turner et al. (1999) did not always find greater diversity and number of
species inside seagrass patches than on bare sandflats. Alfaro (2006) found higher
average species richness in seagrass than in coarse sand, channels, mud, mangroves, or
pneumatophore zones.
Effects of habitat-creating species in soft sediments, however, go beyond that of
simply affecting biodiversity of the sediment. Effects on oxygen and nutrient fluxes
have been demonstrated for both cockles and Macomona (Thrush et al. 2006b).
Cockles have also been demonstrated to affect benthic-pelagic coupling, with their
feeding changing sedimentation rates (Hewitt & Norkko 2007). Pawson (2004)
suggested that feeding by cockles controlled the availability of food in the water
column (as algal biomass) in Papanui Inlet on the Otago Peninsula. Sandwell (2006)
observed that Macomona feeding and movement destabilised the sediment. Seagrass
habitats have been demonstrated to trap sediment (Larkum et al. 2006, Matheson et al.
2008); polydorid tubeworm mats were observed to stabilise sediment (Thrush et al.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 43
1996). Furthermore, tubeworms, Macomona and cockles are a food source for fish
and birds.
Location-specific variables rather than regional species pools seemed to be most
important for controlling the effect of the relationship between a habitat and its
biodiversity. The Southern Kaipara did not give results that were markedly different
to the other locations, nor did locations spatially close to each other produce results
that were more similar than locations further apart. While local hydrodynamics and
anthropogenic activities are the factors most likely to affect the biodiversity of a
specific habitat, factors such as patch size and density are also likely to be important.
For the Tubeworm habitat, species of tubeworm may also be important; unfortunately
there was not enough information to be able to analysis the relative importance of this
factor. For the seagrass habitat, the presence of other species may also be important.
For example, many of the seagrass sites in the Southern Kaipara contained high
numbers of cockles or tubeworms.
In particular, this report highlights the need to preserve a variety of habitats rather than
concentrate on those that are considered to be of high biodiversity. Indeed preserving
habitat diversity at a variety of scales is likely to be the key to conserving biodiversity
and ecosystem function (Thrush & Dayton 2002, Thrush et al. 2006a).
7. Acknowledgements
Without the data produced by the ARC this report could not have been written.
Thanks also go to James Dare for dataset preparation.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 44
8. References
Alfaro, A.C. (2006). Benthic macro-invertebrate community composition within a mangrove / seagrass estuary in northern New Zealand. Estuarine, Coastal and Shelf Science 66: 97-110.
Berkenbusch, K.; Rowden, A.A. (2007). An examination of the spatial and temporal generality of the influence of ecosystem engineers on the composition of associated assemblages. Aquatic Ecology 41: 129-147.
Clarke, R.T.; Gorley, R.N. (2006). Primer v6. PrimerE, Plymouth.
Colwell, R.K. & Coddington, J.A. (1994). Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London 345: 101-118.
Colwell, R.K. (2006). EstimateS: Biodiversity estimation. 8.0. http://viceroy.eeb.uconn.edu/EstimateS,
Crist, T.O.; Veech, J.A.; Gering, J.C.; Summerville, K.S. (2003). Partitioning species diversity across landscapes and regions: a hierarchical analysis of α, β, and γ diversity. American Naturalist 162: 734-743.
Cummings, V.J.; Pridmore, R.D.; Thrush, S.F.; Hewitt, J.E. (1993). Emergence and floating behaviours of post-settlement juveniles of Macomona liliana (Bivalvia: Tellinacea). Marine Behaviour and Physiology 24: 25-32.
Ellingsen, K.E.; Hewitt, J.E.; Thrush, S.F. (2007). Rare species, habitat diversity and functional redundancy in marine benthos. Journal of Sea Research 58: 291-301.
Henriques, P.R. (1980). Faunal community structure of eight soft shore, intertidal habitats in the Manukau harbour. New Zealand Journal of Ecology 3: 97-103.
Hewitt, J.E.; Norkko, J. (2007). Incorporating temporal variability of stressors into studies: An example using suspension-feeding bivalves and elevated suspended sediment concentrations. Journal of Experimental Marine Biology and Ecology 341: 131-141.
Hewitt, J.E.; Thrush, S.F.; Halliday, J.; Duffy, C. (2005). The importance of small-scale biogenic habitat structure for maintaining beta diversity. Ecology 86: 1618-1626.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 45
Klimek, S.; Marini, L.; Hofman, M.; Isselstein, J. (2008). Additive partitioning of plant diversity with respect to grassland management regime, fertilisation and abiotic factors. Basic and Applied Ecology 9: 626-634.
Lande, R. (1996). Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos 76: 5-13.
Larkum, A.W.D.; Orth, R.J.; Duarte, C.M. (eds) (2006). Seagrasses: Biology, Ecology and Conservation. Dordrecht, The Netherlands. Springer.
Lundquist, C.J.; Vopel, K.; Thrush, S.F.; Swales, A. (2003). Evidence for the physical effects of catchment sediment runoff preserved in estuarine sediments: Phase III macrofaunal communities. NIWA Client Report HAM2003-051 prepared for Auckland Regional Council June 2003. (NIWA Project ARC03202).
MacDiarmid, A. (ed.) (2006). An assessment of biodiversity in the New Zealand marine ecoregion. NIWA Client Report for WWF WLG2006–44. http://www.treasuresofthesea.org.nz/
Matheson, F.; Dos Santos, V.; Pilditch, C. (2008). Environmental stressors of New Zealand seagrass: an introduction and progress report. NIWA Client Report #HAM2008-053 prepared for the Department of Conservation, project #DOC08202.
Ministry of Fisheries, Department of Conservation (2008). Marine protected areas: Classification, protection standard and implementation guidelines. 54 p.
Pawson, M.M. (2004). The cockle Austrovenus stutchburyi and chlorophyll depletion in a southern New Zealand inlet. University of Otago, Dunedin. p.
Pridmore, R.D.; Thrush, S.F.; Hewitt, J.E.; Roper, D.S. (1990). Macrobenthic community composition of six intertidal sandflats in Manukau Harbour, New Zealand. New Zealand Journal of Marine and Freshwater Research 24: 81-96.
Snelder, T.H.; Leathwick, J.R.; Dey, K.L.; Rowden, A.A.; Weatherhead, M.A.; Fenwick, G.D.; Francis, M.P.; Gorman, R.M.; Grieve, J.M.; Hadfield, M.G. (et al.) (2006). Development of anecologic marine classification in the New Zealand region. Environmental Management 39(1): 12-29.
Ricotta, C. (2008). Computing additive diversity from presence-absence scores: a critique and alternative parameters. Theoretical Population Biology 73: 244-249.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 46
Rowden, A.A.; Berkenbusch, K.; Brewin, P.E.; Dalen, J.; Neill, K.F.; Nelson, W.A.; Oliver, M.D.; Probert, P.K.; Schwarz, A-M.; Sui, P.H.; Sutherland, D. (2007). A review of the marine soft-sediment assemblages of New Zealand. NZ Aquatic Environment and Biodiversity Report. 184 pp.
Sandwell, D.R. (2006). Austrovenus stutchburyi, regulators of estuarine benthic-pelagic coupling. University of Waikato, Hamilton. p.
Thrush, S.F.; Gray, J.S.; Hewitt, J.E.; Ugland, K.I. (2006a). Predicting the effects of habitat homogenization on marine biodiversity. Ecological Applications 16: 1636-1642.
Thrush, S.F.; Whitlatch, R.B.; Pridmore, R.D.; Hewitt, J.E.; Cummings, V.J.; Maskery, M. (1996). Scale-dependent recolonization: the role of sediment stability in a dynamic sandflat habitat. Ecology 77: 2472-2487.
Thrush, S.F.; Hewitt, J.E.; Gibb, M.; Lundquist, C.; Norkko, A. (2006b). Functional role of large organisms in intertidal communities: Community effects and ecosystem function. Ecosystems 9: 1029-1040.
Thrush, S.F.; Pridmore, R.D.; Hewitt, J.E.; Cummings, V.J. (1992). Adult infauna as facilitators of colonization on intertidal sandflats. Journal of Experimental Marine Biology and Ecology 159: 253-265.
Thrush, S.F.; Dayton, P.K. (2002). Disturbance to marine benthic habitats by trawling and dredging - Implications for marine biodiversity. Annual Review of Ecology and Systematics 33: 449-473.
Turner, S.J.; Hewitt, J.E.; Wilkinson, M.R.; Morrisey, D.J.; Thrush, S.F.; Cummings, V.J.; Funnell, G. (1999). Seagrass patches and landscapes: the influence of wind-wave dynamics and hierarchical arrangements of spatial structure on macrofaunal seagrass communities. Estuaries 22: 1016-1032.
Ugland, K.I.; Gray, J.S. & Ellingsen, K.E. (2003). The species-accumulation curve and estimation of species richness. Journal of Animal Ecology 72:888-897.

Biodiversity of intertidal soft-sediment habitats in the Auckland Region 47
van Houte-Howes, K.S.; Turner, S.J.; Pilditch, C.A. (2004). Spatial differences in Macroinvertebrate communities in intertidal seagrass habitats and unvegetated sediment in three New Zealand estuaries. Estuaries 27: 945-957.
Whittaker, R.H. (1960). Vegetation of the Siskiyou Mountains, Oregon and California. Ecological Monographs 30: 279-338.