biochemical study of lipids in chyle, blood, and liver of corn oil- and butter-fed rats with phase...
TRANSCRIPT

EXPERIMENTAL AND MOLECUL4R PATHOLOGY 2, 14-31 ( 1963)
Biochemical Study of Lipids in Chyle, Blood, and Liver of Corn Oil-
and Butter-Fed Rats with Phase and Electron Microscopy
Correlation’
ROSE JONES, K. F. SCOTT, E. S. MORRISON, M. PROMS, AND UT. A. THOMAS
Department of Pathology, iilbany Medical College, Albany, New York
Received March 15, 1962
INTRODUCTION
Myocardial and renal infarcts associated with arterial thrombi can be produced in rats fed a complex diet containing, among other ingredients, propylthiouracil, bile salts, cholesterol and 40% butter (Thomas and Hartroft, 1959). When corn oil is substituted for the butter component, the thrombogenic and infarct producing effects of this diet are abolished (Thomas and Hartroft, 1959; O’Neal et al., 1959). Probably related to the difference in thrombogenic properties is the significantly longer “clot-lysis” time, measured in the thrombelastograph, that occurs in rats receiving the butter as compared to the corn oil diet (Scott et al., 1961). In addi- tion, the presence of butter in place of corn oil in these diets results in much higher serum lipid levels and a greatly modified adipose tissue fatty acid pattern (Scott et al., 1962a). We believe that the anatomic, hematologic, and chemical effects of corn oil and butter in the rat are worth further investigation because of their pos- sible relationship to thrombosis and infarction in man.
Using phase and electron microscopy, the physical appearance of corn oil and butter (or derivatives) in the jejunal mucosa and chyle of rats was investigated by us in a previously reported experiment (Jones et al., 1962). Following gastric tube feedings of either 2 ml of corn oil or butter, the osmiophilic material in both the jejunal mucosa and chyle of rats had appearances that differed with the type of fat given.
In the present study we have extended the previous phase and electron microscopy observations by examining blood, blood vessels, and liver of rats after single tube feedings with butter, corn oil, or water. In addition, the lipid composition of the corn oil and butter, and of the chyle, blood, and liver of rats given these fats has been analyzed using silica gel thin layer and vapor phase chromatography. These analyses were done for two reasons: (1) to learn how the composition, particularly the fatty acid composition, of corn oil and butter is modified as these substances travel from the gastrointestinal tract to chyle, through blood, and to the liver, and (2) to determine whether or not the physical appearance of these lipids, as seen in fresh preparations by phase microscopy and in osmium tetroxide fixed material b> electron microscopy, could be related to their chemical composition.
1 This work was supported by PHS Grant H-7155, National Heart Institute, Bethesda, Maryland.
14

LIPIDS IN CORN OIL- AND BUTTER-FED RATS 15
MATERIALS AXD METHODS
Male Wistar rats weighing between 250 and 400 gms were maintained in wire bottomed cages on Purina rat pellets and fed water ad Zibitur~. After 18 hours of fasting, but with water being offered ad libitum, individual rats in sets were fed by transoral gastric intubation either 2 ml of corn oil, melted butter, or tap water. All sets of rats contained at least one water-, one corn oil-, and one butter-fed animal. In the experiment 17 rats received water, 23 rats received corn oil and 23 rats, butter. After 4-6 hours the rats were anesthetized with pentobarbital sodium, 5 mg 100gms of body weight.
From rats prepared in this manner: material for electron microscopy was obtained from jejunum, a visceral lymphatic or the thoracic duct, thoracic aorta, anterior left ventricle of the heart, including the coronary artery in this area, and anterior left lobe of liver. The method of obtaining jejunum for electron microscopy was the same as described previously (Jones et al., 1962). Chyle was obtained by cannulat- ing either a visceral lymphatic in the area of the jejunum or the thoracic duct. A fine polyethylene catheter, previously rinsed out with heparin to prevent clotting of the chyle, was used, and the first drop of chyle was discarded. The chyle was then allowed to drip into a test tube containin, v osmium tetroxide. All tissues were fixed for one and one-half hours in Dalton’s osmium tetroxide (with two changes of osmium) and the tissue was dehydrated with graded concentrations of a tnixture of methyl and butyl alcohol. The material was then embedded in Epon, sectioned with a Porter-Blum microtome, stained with uranyl acetate (Watson, 1958a) or lead hydroxide (Watson, 1958b). and examined with an RCA Model EMU-3F electron microscope.
In order to examine chylomicra not subject to the artifactual changes possibly induced by fixation in osmium tetroxide and embedding in Epon, chyle, whole blood, and plasma from water, corn oil- and butter-fed rats were examined in as natural a state as possible by phase microscopy. Chyle, obtained by cannulation as described above, and whole blood from the aorta were examined after being spread gently on a siliconized glass slide. Plasma was obtained by mixing whole blood and 3.8% sodium citrate in a 9: 1 ratio, and then centrifuging the mixture at 4°C for 15 min at 3000 rpm in an International centrifuge usin, u a 413A head. The plasma from different levels (top layer and bottom layer) was then gently spread on siliconized glass slides and examined for chylomicra.
IV-hole chyle was analyzed for its lipid composition in 6 water-, 6 corn oil-, and 6 butter-fed animals. Because only a small amount of chyle could be obtained from each rat, the chyle from each group of 6 animals was pooled quantitatively into 2 groups of 3 each. Serum from 9 water-, 9 corn oil-, and 9 butter-fed rats was pooled in 3 groups of 3 each and analyzed. From each of these rats an average of 7 ml of blood was collected in a centrifuge tube after aortic puncture. The blood was allowed to clot at room temperature for 4 hours and then centrifuged at 4°C for 15 min at 2000rpm in an International centrifuge using a 413A head, after which the serum was pipetted off. The pools of chyle and serum were stored in liquid nitrogen at -195 J until analyses were done.
The superior left lobe of the liver from the bled rats was obtained immediately after death. The livers from 6 water-fed rats were quantitatively pooled in 2 pools

16 JONES, SCOTT, MORRISON, KROMS, AND THOMAS
of 3 each, as were the liver lobes from 6 corn oil- and 6 butter-fed rats. The pools of liver tissue were immediately homogenized using a Waring Blendor, extracted in 2: 1 chloroform-methanol and the residue filtered off. The chloroform-methanol extract was then treated as described below.
For analysis, the chyle and serum samples were taken from liquid nitrogen, thawed at room temperature and extracted according to Folch, with 20 volumes of 2 : 1 chloroform-methanol. After an hour they were layered with l”/* saline and equilibrated overnight under nitrogen to remove nonlipid contaminants. Aliquots of the chloroform extract were taken for spectrophotometric determination of sterol (Zlatkis et al., 1954), lipid phosphorus (Bartlett, 1959), triglycerides (Van Handel and Zilversmit, 1957), and for separation of lipid classes on thin layer silica gel plates.
The fatty acid compositions of the three esterified fractions (triglyceride, choles- terol ester, and phospholipid) of chyle, serum and superior left lobe of the liver were determined in the following manner. Chloroform extract from the Folch procedure was evaporated at 60°C under reduced pressure on a rotary evaporator and then taken up in about 1 ml of chloroform. This was applied to 20 X 20 cm silica gel plates in about 30 discrete spots of 10 microliters each, per plate. Two plates were used for each sample. Silica gel G” was applied in a layer of approximately 275 lt thickness. The developing solvent used was 80: 20: 2 n-hexane : diethyl ether: glacial acetic acid. The system gave good separation of phospholipids, cholesterol, nonesteri- fied fatty acids, triglycerides, and cholesterol esters in standard test systems and also in serum extracts. Running time was about 50 min.
Solvent was evaporated from the plates at room temperature, and a narrow strip at the ends and in the middle of each plate was sprayed with 0.2% solution of 2,7-dichlorofluorescein in 9576 ethanol in order to mark the lipid bands. Unstained portions of each of the five bands, in addition to a blank area, were scraped from the plates and transferred to 40 ml centrifuge tubes. Ten ml of 2% sulfuric acid in absolute methanol were added to each tube; the tubes were shaken intermittently for an hour and permitted to stand overnight at room temperature in a nitrogen atmosphere. After heating to 65°C with agitation, the silica gel was removed by centrifugation, the methyl esters of the fatty acids extracted with n-hexane, and analyzed as previously described (Scott et al., 1962a) on a Barber-Coleman Model 10 gas chromatograph. The machines were checked each morning with standard mixtures of methyl esters supplied by the National Institutes of Health. Machines showing greater deviation than 270 for any component of the standard were not used until they were adjusted. Hydrogenation of some samples was done to identify unsaturated fatty acids.
RESULTS
The results of the lipid analysis of the corn oil and butter fed to the rats in this experiment are given in Tables I and II. Gross examination of the stomach and small intestine of the corn oil- and butter-fed rats showed them to be moderately full of lipid 4-6 hours after being fed. The stomach and small bowel of the water-fed animals appeared grossly to contain no food particles.
3 Obtained from Brinkmann Instruments Inc., Great Neck, Long Island.

LIPIDS IN CORN OIL- AND BUTTER-FED RATS 17
Electron microscopy sections of jejunum from water-fed animals seldom showed osmiophilic masses suggestive of lipid. In the corn oil-fed animals, sections showed lipid to be present within and between mucosal cells (Fig. 1) and in what appeared to be lymphatics. The morphologic appearance of the lipid was similar to that seen in previous experiments. The droplets had smooth, regular borders, stained regu- larly, and did not show fragmentation or drop-out. Sections of jejunum from the butter-fed rats (Figs. 2 and 3) showed the distribution of the presumed lipid to be the same as in the corn-oil fed animals, but its appearance to be quite different, thereby confirming previous experiments (Scott et al., 1962b).
TABLE I
QUANTITATIVE TRIGLYCERIDE, STEROL, AKD LIPID PHOSPHORUS IN CORN OIL AND BUTTER FED TO RATS
Lipid Triglyceride Sterol phosphorus
(IL mols/gm) b mols/gm) ( !I mols/gm)
Corn oil 803 52 0.6
Butter 774 15 3
TABLE II
RELATIVE PERCENTAGE OP FATTY ACIDS OF TRIGLYCERIDE, PHOSPHOLIPIII, AND STEROL IN CORN OIL
AND BUTTER FED TO RATS
Mono- Poly-
Satu- unsatu- unsatu-
C14:O C16:O C16:l C18:O C18:l C18:2 C20:4 rates rates rates --
Triglyceride
Corn oil Trace” 12 Trace 1 26 61 None 13 26 61
Butter 14 44 3 13 26 Trace None 71 29 0
Phospholipid
Corn oil Trace 20 4 4 29 43 Trace 24 33 43
Butter 13 50 7 15 15 Trace None 78 22 0 -
Sterol
Corn oil 3 15 4 4 22 52 Trace 22 26 52
Butter 15 41 5 16 23 Trace None 72 28 0
u Amount less than 0.5%.
Only a small amount of osmiophilic material was found by either phase or electron microscopy in the chyle of water-fed rats. In the corn oil-fed rats, smooth bordered chylomicra measuring from 1 p down to the limits of resolution were seen by both phase and electron microscopy (Fig. 4). Phase microscopy of the chyle of butter- fed rats showed a similar picture, but in addition, some non-nucleated masses meas- uring up to 30 p were seen; these masses were presumably lipid. Electron micro- graphs of the chyle from butter-fed rats showed predominantly two forms of fat: irregular fragments much smaller than the chylomicra seen in corn oil-fed rats by phase microscopy, and large masses measuring up to 30 p in diameter (Fig. 5).
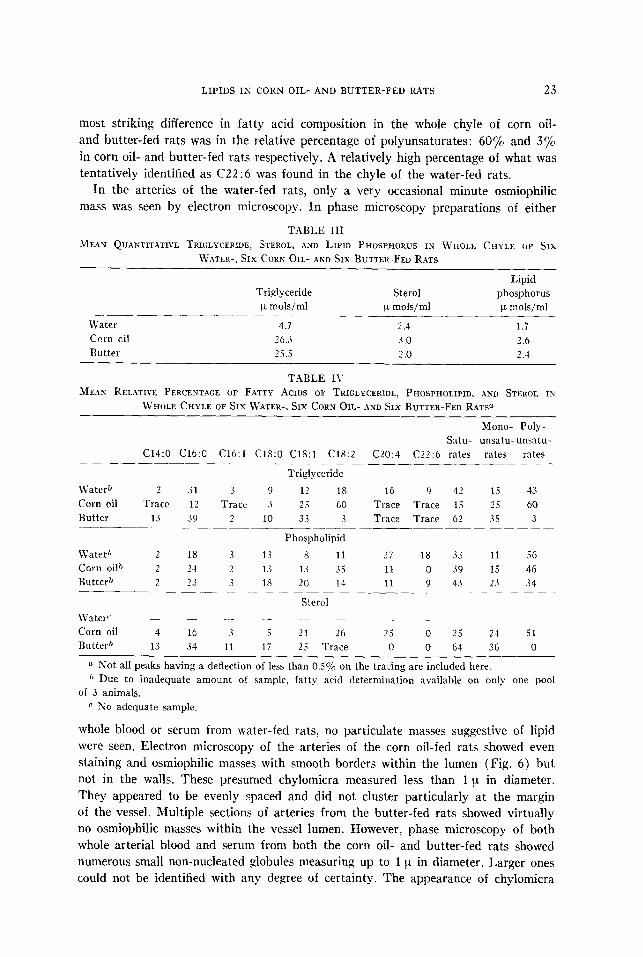
The results of the lipid analyses of whole chyle in rats fed water, butter, and corn oil are given in Tables III and IV. There appears to be fairly close correlation between the fatty acid composition of the dietary lipid, either corn oil or butter, and the fatty acid composition of the whole chyle of rats receiving these lipids. The

1s JONES, SCOTT, MORRISON, KROMS, AND THOMAS
Fm. 1. Jejunum of a rat fed 2 ml of corn oil 4 hours before death. .4 moderate amount of
densely osmiophilic material can be seen within the lumen as well as in and between the cells.
The terminal webb appears to contain a small amount of lipid. The chemical composition of the corn oil and butter fed to the rats in this experiment is given in Tables I and II. Uranyl acetate.
X 23,C@O.

LIPIDS IN CORN OIL- AND BUTTER-FED RATS 19
FIG. 2. Jejunum of butter-fed rat. The presumed lipid shows considerable artifactual “fall out,” a feature not seen in the jejunal lipid of corn oil-fed rats. Some small lipid droplets are seen
either in an intercellular position or within a lymphatic. Uranyl acetate. x 21,500.

20 JONES, SCOTT, MORRISON, KROMS, AND THOMAS
FIG. 3. From deep within the jejunum of a butter-fed rat. A large mass of densely osmiophilic
material showing considerable drop-out can be seen within a cell. Between this and an adjacent
cell are smaller droplets of either butter or lipid derived from butter. Uranyl acetate. x 21,000.

LIPIDS IN CORN OIL- AND BUTTER-FED RATS 21
FIG. 4. Chyle cannulated from the visceral lymphatic of a corn oil-fed rat. The chylomicra
vary in size from 1 u down to the limits of resolution. No giant chylomicra were found either by
electron or phase microscopy in the chyle of corn oil-fed rats. The lipid composition of the chyle
of corn oil- and butter-fed rats is given in Tables III and IV. Two lymphocytes and a few strands of what is possibly fibrin can be seen. Uranyl acetate. x 1200.

22 JONES, SCOTT, MORRISON, KROMS, AND THOMAS
FIG. 5. Chyle cannulated from the visceral lymphatic of a butter-fed rat. The picture was
deliberately greatly magnified to show the homogeneous appearance of the “giant chylomicron” seen at the top, The chylomicron measured approximately 30 u in diameter. The minute irregular
fragments of fat seen by electron microscopy in the chyle of butter-fed rats are also shown.
Uranyl acetate. X 52,000.
LIPIDS IN CORN OIL- AND BUTTER-FED RATS 23
most striking difference in fatty acid composition in the whole chyle of corn oil- and butter-fed rats was in the relative percentage of polyunsaturates: 60% and 3% in corn oil- and butter-fed rats respectively. A relatively high percentage of what was tentatively identified as C22 :6 was found in the chyle of the water-fed rats.
In the arteries of the water-fed rats, only a very occasional minute osmiophilic mass was seen by electron microscopy. In phase microscopy preparations of either
T.4BLE III
MEAN QUANTITXTIVE TRIGLYCERIDE, STEROL, AND LIPID PHOSPHORUS IN WHOLE CHYLE OF SIX
WATER-, SIX CORE OIL- AND SIX BUTTER-FED RATS
Lipid Triglyceride Sterol phosphorus
FL mols/ml F mols/ml p mols/ml _______-
Water 4.7 2.4 1.7
Ccrn oil 2601 3.0 2.6
Butter 25.5 2 .o 2.4
TABLE I\
MEAN RELATIVE PERCENTAGE OF FATTY ACIDS OF TRIGLYCERIDE, PHOSPHOLIPID, AND STEROL IN
WHOLE CHYLE OF SIX WATER-, SIX CORN OIL- AND SIX BUTTER-FED RATP
Mono- Poly-
Satu- unsatu- unsatu-
C14:O C16:O C16:l C18:O C18:l C18:2 C20:4 C22:6 rates rates rates --
Triglyceride
\Vaterb 2 31 3 9 12 18 16 9 42 15 43
Corn oil Trace 12 Trace 3 2 5 60 Trace Trace 15 25 60
Butter 13 39 2 10 33 3 Trace Trace 62 35 3 _____
Phospholipid
LVater” 2 1s 3 13 s 11 27 18 33 11 56
Corn oil{) 2 24 2 1.s 1.i 35 11 0 39 15 46
Butter” 2 23 3 18 20 11 11 9 43 2.1 34 -__ -__~- -~~- ~~ - -
Sterol
L\yatert’ _ _ - - - ._ - -
Corn oil 4 I6 3 .i 21 26 2 5 0 25 24 51
Butter” 13 3-i 11 li 25 Trace 0 0 64 36 0
o Not all peaks having a deflection of less than 0.5% on the tracing are included here. ?’ Due to inadequate amount of sample, fatty acid determination available on only one pool
of 3 anima!s.
r SO adequate sample.
whole blood or serum from water-fed rats, no particulate masses suggestive of lipid were seen. Electron microscopy of the arteries of the corn oil-fed rats showed even staining and osmiophilic masses with smooth borders within the lumen (Fig. 6) but not in the walls. These presumed chylomicra measured less than 1 p in diameter. They appeared to be evenly spaced and did not cluster particularly at the margin of the vessel. Multiple sections of arteries from the butter-fed rats showed virtually no osmiophilic masses within the vessel lumen. However, phase microscopy of both whole arterial blood and serum from both the corn oil- and butter-fed rats showed numerous small non-nucleated globules measuring up to 1 p in diameter. Larger ones could not be identified with any degree of certainty. The appearance of chylomicra

24 JONES, SCOTT, MORRISON, KROMS, AND THOMAS
FIG. 6. Myocardium of a corn oil-fed rat. Within the lumen of a vessel are seen masses of
lipid measuring less than 1 cl. These masses of lipid were not seen penetrating within the wall of
any vessel. Distal to the vessel wall are strands of collagen , some of them cut end-on. Uranyl
acetate. X 19,500.

LIPIDS IN CORN OIL- AND BUTTER-FED RATS 2.5
in whole blood, serum, or plasma, as seen by phase microscopy, was the same whether the rat had been fed butter or corn oil. These globules were thought to be lipid and not platelets because of their appearance and because they were seen only in the corn oil- or butter-fed rats and not in the water-fed animals,
The results of the serum lipid analyses of the water-, corn oil-, and butter-fed rats are shown in Tables V and VI. The total triglyceride appeared to be slightly
TABLE 1’
MEAN QUANTITATIVE TRIGLYCERIDE, PHOSPHOLIPID, AND STEROL IN SERUM OF SIX WATER-, SIX CORN OIL- AND SIX BUTTER-FED RATS
Water Corn oil
Butter
Triglyceride
p mols/ml
0.6
1.7
0.8
Phospholipid
p mols/ml
1.4
I.5 1.6
Sterol
w mols/ml
2.0
2.6
1.9
TABLE VI
MEAN RELATIVE PERCENTAGE OF FATTY ACIDS ESTERIFIED WITH TRIGLYCERIDE, PHOSPHOLIPID, AND
STEROL IN SERUM OF SIX WATER-. SIX CORN OIL-, AND SIX BUTTER-FED RATS
Mono- Poly- Satu- unsatu- unsatu-
C14:O C16:O C16:l C18:O C18:l C18:2 C20:4 rates rates rates
Triglyceride
\Vater 1 30 3 3 22 33 8 34 25 41
Corn oil 1 15 1 1 23 S7 2 17 24 S9
Butter 7 29 4 s 28 17 7 44 32 24
Phospholipid
Water 1 27 1 28 6 14 23 56 7 37
Corn oil 1 23 1 22 s 25 20 46 9 45
Butter 1 26 1 24 10 16 22 51 11 38 - ~~__.
Sterol
\Vater 1 9 2 2 10 15 61 12 12 76
Corn oil 1 10 3 2 11 22 51 13 14 73
Butter 2 12 3 2 ‘? 1 5 58 16 11 73
higher in the corn oil-fed animals. The serum fatty acid patterns, to a lesser extent than those of the chyle, tended to reflect the fatty acid pattern of the kind of lipid fed.
Electron microscopy section of jejunum from water-fed animals seldom showed fed rats were unremarkable. In those rats fed corn oil or butter, no material sug- gestive of lipid could be found within the vessel wall.
The electron microscopic appearance of the liver of the water-fed rats showed virtually no osmiophilic bodies suggestive of lipid. The appearance of the liver of the corn oil-fed rats was similar to that of the butter-fed animals (Fig. 7). Dense osmiophilic masses of varying size, with smooth borders and no fragmentation, were seen. These masses often seemed to be associated with mitochondria. The results of the lipid analyses of the superior left lobe of the liver in the water-, corn coil-, and butter-fed rats are given in Tables VII and VIII.

26 JONES, SCOTT, MORRISON, KROMS, AND THOMAS
FIG. 7. Liver of a corn oil-fed rat. The physical appearance and the lipid composition of the
livers of the water-, corn oil- and butter-fed rats were similar. Lead hydroxide. x 44,000.

LIPIDS IN CORN OIL- AND BUTTER-FED RATS 27
Electron microscopy sections of the myocardium of the water- (Fig. 8), corn oil-, and butter-fed rats showed no consistent differences. Most of the lipid seen was associated with mitochondria.
TABLE VII
MEAN QUANTITATIVE TRIGLYCERIDE, PHOSPHOLIPID, AND STEROL IN LIVER OF SIX WATER-, SIX CORK OIL- AND SIX BUTTER-FED RATS
Water
Corn oil
Butter
Triglyceride Phospholipid
11 mols/gm y mols/pm
16.5 40.8
18.4 39.9
18.3 54.6
Sterol
v mols/gm ___.~
7.8
6.8
7.3
TABLE VIII
MEAX RELATIVE PERCENTAGE OF FATTY .\CIDS OF TRIGLYCERIDE, PHOSPIIOLIPID, AND STEROL IN LIVER
OF SIX WATER-, SIX CORN OIL-, AND SIX BUTTER-FED RATS
Mono- Poly-
Satu- unsatu- unsatu-
C14:O Cl6:O C16:l C1Y:O C18:l Cl8:Z C20:4 rates rates rates
CVater
Corn oil Butte+
Water
Corn oil
Butter
27
25
34
20
19
20
Triglyceride
3 24 35 8 31 26 43
2 21 45 a 28 22 50
3 24 30 7 38 25 37 .__
Phospholipid
29 5 14 30 50 6 44
30 5 14 30 50 6 44
30 5 13 30 51 6 43
- Sterol
Water’& Trace 36 i 8 26 23 0 44 33 23
Corn oilG Trace 36 4 9 2s 26 0 45 29 26
Butter” - - - - -
o One pool of three animals only.
b Samples too small to measure.
DISCUSSION
Experiments using purified lipids in in vitro systems have suggested that osmium tetroxide combines with double bonds of fatty acids and possibly also with polar end groups such as those of phospholipids (Riemersma and Booij, 1962). In this study, the electron microscopy appearance of certain osmium tetroxide-fixed lipids has been correlated with chemical composition of these lipids as they exist in dif-
ferent locations in an in viva system, specifically in rats fed either corn oil or butter. The fatty acid pattern of chyle in the corn oil- and butter-fed rats resembled that
of ingested lipids. Triglyceride was the largest lipid class in both groups, and the polyunsaturated fatty acid content was high in corn oil-fed and low in butter-fed rats. The short chain fatty acids present in the butter were absent in the chyle of the butter-fed rats; the short chain fatty acids were presumably absorbed directly into the venous system from the jejunum (Frazer, 1961). The chylomicra of corn oil-fed rats had a similar physical appearance in both fresh material seen by phase microscopy and in fixed material seen by electron microscopy. The chylomicra of

28 JONES, SCOTT, MORRISON, KROMS, AA-D THOMAS
FIG. 8. Myocardium of a water-fed rat. As in the corn oil- and butter-fed rats, a moderate amount of lipid was found associated with the mitochondria, hut no morphologic differences
between water-, corn oil- and butter-fed rats were found in the myocardium. Uranyl acetate.
x 18,000.

LIPIDS IN CORN OIL- AND BUTTER-FED RATS 29
the butter-fed rats, however, were not similar in appearance in fresh phase and fixed electron microscopy preparations. The virtual disappearance of chylomicra measuring 0.5 to 1 IL in the osmium fixed preparations of chyle from butter-fed rats may have been due to the relatively small total number of double bonds in these chylomicra. Despite the presence of some monounsaturated fatty acids (mostly oleic acid), the osmium tetroxide appeared to be unable to combine and form visible complexes with these globules of fat under the conditions of this experiment. How- ever, the effect of the alcohols used in clearing and the Epon used in embedding may also be important in the eventual physical appearance of the lipid.
In serum, the fatty acid patterns of the corn oil- and butter-fed rats still tended to resemble the lipid fed. In corn oil-fed rats, the appearance of chylomicra as seen in unfixed whole blood by phase microscopy and as seen after fixation in blood trapped in blood vessels by electron microscopy was similar. In butter-fed rats the chylomicra as seen by phase microscopy in uncentrifuged whole blood and plasma were similar in size and number to those seen in the corn oil-fed rats, although serum triglyceride was lower in the butter-fed group. By electron microscopy, despite many sections, no osmiophilic masses were found within the vessel lumen in the butter-fed rats. Because these butter chylomicra were readily seen in unfixed phase microscopy preparations and not in osmium fixed preparations as the corn oil chylomicra were, it appears that qualitative properties of the butter chylomicra prevented them from being fixed with osmium tetroxide.
The livers of water-, corn oil-, and butter-fed rats all showed a relatively large total number of polyunsaturated fatty acids, although differences did exist in fatty acids between the three groups of animals. In keeping with the high polyunsaturated fatty acid content of these livers, no consistent differences as yet have been seen in electron micrographs from water-, corn oil-, or butter-fed animals. The elevated phospholipid content of the livers of the butter-fed rats is not reflected so far in the electron microscopy preparations from these animals.
In this study, lipid complexes in an in zGoo system containing substantial numbers of molecules with multiple double bonds were demonstrated readily by electron microscopy at all stages of absorption and transport. Those complexes with fewer molecules containing double bonds were either artifactually distorted, as judged by comparison with their appearance in fresh phase microscopy preparations, or not visible at all. Since it has been shown that osmium tetroxide will combine with oleic acid introduced per trachea into the lung of a rat (Imai, 1962), it is likely that the relatively oleic acid-rich lipid in the chyle and blood of butter-fed rats is capable of being demonstrated. Possibly a longer fixation time than was used in this study, enabling the osmium tetroxide to penetrate and combine with the single double bond of more oleic acid molecules, would be effective.
The giant chylomicra noted previously in unstained sections of chyle from butter- fed rats were also observed in stained sections in this study. We still cannot be certain whether these occur in viva or whether they simply represent fusion in vitro of smaller chylomicra. No osmiophilic masses suggestive of lipid were found in the wall of the aorta or coronary arteries in either butter- or corn oil-fed rats. In butter-fed rats, the absence of chylomicra in vessel walls may be artifactual, but this does not seem likely in corn oil-fed animals because chylomicra were readily seen within the

30 JONES, SCOTT, MORRISON, KROMS, AND THOMAS
vessel lumen in electron microscopy preparations of corn oil-fed rats. The lipid probably passes into the arterial walls in much smaller particles (p-lipoprotein size or smaller). The manner in which lipid is transported into a vessel wall is of great importance with regard to arteriosclerosis and should be investigated further using combined electron microscopy and biochemical techniques. The resolving potenti- alities of the electron microscope are such that with appropriate techniques, particles at least near the molecular size of most lipids can be seen. Such techniques need to be developed for use in current investigations concerning atherosclerosis.
SUMMARY
In both whole chyle and serum of corn oil- and butter-fed rats, the fatty acid patterns resembled
that of the lipid fed. In vitro studies have suggested that osmium tetroxide combines with polar
end groups of lipids or double bonds of fatty acids. In the corn oil-fed rats, the lipid masses in
chyle and within blood vessels had a similar appearance in fresh phase microscopy preparations
and in osmium tetroxide-fixed material seen by electron microscopy. In the butter-fed rats, how-
ever, lipid in chyle and serum was either artifactually distorted or was not stained at all by
osmium tetroxide. In the livers of the water-, corn oil-, and butter-fed rats, a relatively large
number of double bonds were demonstrated by lipid analysis in all three groups. In keeping
with this finding, osmium tetroxide stainable lipid of similar appearance was found in electron
microscopy preparations of the livers from all three groups of rats. The lack of double bonds in
the lipid of chyle and serum of the butter-fed rats compared to the corn oil-fed rats possibly
explains the behavior of these lipids when fixed with osmium tetroxide.
.4s in a previous study, giant chylomicra were seen both by phase and electron microscopy in
the chyle of butter-fed rats. It was not possible, however, to determine whether these chylomicra
were not, at least in part, artifactual. No lipid masses were found passing through the vessel
walls of either the corn oil- or butter-fed rats, but in this study the maximum resolving po’wer
of the electron microscope was not used, and it is possible that further studies may demonstrate
lipid masses of specific chemical composition within arterial walls.
REFERENCES
BARTLETT, G. R. (1959). Phosphorus assay in column chromatography. /. Biol. Chew. 334,
466-471.
FR;ZZER, A. C. (1961). Role of lipids in normal metabolism. Fed. Proc. 5th International Congress
on Nutrition. Vol. 20, No. 1, Part II, 146-151.
IJIAI, H. (1963). Study of electron microscopy of oleic and linoleic acids injected intrabron- chially into the lungs of rats. Exptl. and Mol. Pathol. In press.
JONES, R., THOMAS, W. .4., and SCOTT, R. F. (1962). Electron microscopy study of chyle from
rats fed butter or corn oil. EnptE. and Mol. Pathol. 1, 65-83.
O’NEAL, R. M. THOMAS, W. A., and HARTROFT, W. S. (1959). Dietary production of myocardial
infarction in rats: Anatomic features of the disease. Am. J. Cardiol. 3, 94-100.
RIE~YERSMA, J. C., and Boors, H. L. (1962). The reaction of osmium tetroxide with lecithin:
Application of staining procedures. /. Histochem. and Cytochem. 10, 89-95.
SCOTT, R. F., ALOUSI, K., and THOMAS, W. A. (1961). Chemico-pathologic studies of dietary
lipids, thrombosis, and clot-lysis. I. In vitro methods, utilizing liquid nitrogen refrigeration and
the thrombelastograph, for demonstrating with rats the antilytic effect of butter-containing
atherothrombogenic diets. Arch. Pathol. 71, 594-599. SCOTT, R. F., HALE, C., HALE, T., GOODALE, F., and TEJADA, C. (1962a). Chemico-anatomic
studies in the geographic pathology of arteriosclerosis: A comparison of fatty acids of adipose tissue and of plasma lipids in ncndiabetics and diabetics from Guatemala and the United States.
Exptl. and Mol. Pathol. 1, 44-56.
SCOTT, R. F., IMAI, H., GOODALE, F., LEE, K. T., and MORRISON, E. (1962b). Dietary-induced myocardial infarction in rats: the possible role of essential fatty acid deficiency. Exptl. and Mol.
Path. 1. l-14.

LIPIDS IS CORN OIL- AND BUTTER-FED RATS 31
THOMAS, W. A., and HARTROFT, W. S. (1959). Myocardial infarction in rats fed diets con-
taining high fat, cholesterol, thiouracil, and sodium cholate. Circ. 19, 65-72.
VAN HANDEL, E., and ZILVERSMIT, D. B. (195i). Micromethod for the direct determination of
serum triglycerides. .I. Lab. Clin. Med. 56, 152-157.
WATSON, M. L. (1958a). Staining of tissue sections for electron microscopy with heavy metals,
J. Biophys. Biochenz. Cytol. 4, 465-478.
WATSON, M. L. (195Sb). Staining of tissue sections for electron microscopy with heavy metals. II. .4pplicaticns of solutions containing lead and barium. J. Bioph.y,. Riorhtw. Cyfol. 4, 727-730.
ZLATXIS, J., Z.41i, B., and BOYLE, A. J. (1934). A new method for the determination of
cholesterol. J. Lab. Clin. Med. 41. 486-492.