biochemical engeenering j_2016
TRANSCRIPT

Seediscussions,stats,andauthorprofilesforthispublicationat:http://www.researchgate.net/publication/282333838
Anoxic–aerobicSBRsystemfornitrate,phosphateandCODremovalfromhigh-strengthwastewateranddiversitystudyofmicrobialcommunities
ARTICLEinBIOCHEMICALENGINEERINGJOURNAL·JANUARY2016
ImpactFactor:2.47
READS
21
5AUTHORS,INCLUDING:
JyotsnaJena
InstituteofMineralsandMaterialsTechnol…
11PUBLICATIONS21CITATIONS
SEEPROFILE
RavindraKumar
InstituteofLifeSciences
3PUBLICATIONS1CITATION
SEEPROFILE
AnshumanDixit
43PUBLICATIONS455CITATIONS
SEEPROFILE
TruptiDas
InstituteofMineralsandMaterialsTechnol…
37PUBLICATIONS148CITATIONS
SEEPROFILE
Availablefrom:TruptiDas
Retrievedon:13October2015

Afc
Ja
b
a
ARR1AA
KSNPAAM
1
empag
atbbv
h1
Biochemical Engineering Journal 105 (2016) 80–89
Contents lists available at ScienceDirect
Biochemical Engineering Journal
jo ur nal home p age: www.elsev ier .com/ locate / bej
noxic–aerobic SBR system for nitrate, phosphate and COD removalrom high-strength wastewater and diversity study of microbialommunities
yotsnarani Jena a, Ravindra Kumar b, Md Saifuddin a, Anshuman Dixit b, Trupti Das a,∗
CSIR- Institute of Minerals and Materials Technology, Bhubaneswar, Odisha 751013, IndiaInstitute of Life Sciences, Bhubaneswar, Odisha 751023, India
r t i c l e i n f o
rticle history:eceived 2 May 2015eceived in revised form0 September 2015ccepted 14 September 2015vailable online 15 September 2015
eywords:BRitrate (NO3)hosphate and COD removalnoxicerobicicrobial diversity
a b s t r a c t
Anoxic–aerobic sequencing batch reactor (SBR) system was operated for 180 days (under ambient tem-perature, 20 days SRT, 24 h HRT, influent COD/nitrate: 4 and COD/phosphate: 137) to treat a high strengthwastewater (1000 mg/L nitrate and 4000 mg/L COD). The unique aspect was elimination of anaerobiccycle due to availability of nitrate (NO3) and oxygen as electron acceptors in anoxic and aerobic phasesrespectively. Simultaneous removal of nitrate (98%), phosphate (86%), and COD (72%) was achievedin anoxic phase. The subsequent aerobic phase experienced 26% of residual COD removal along withphosphate release (∼3.4 mg/L), reducing the overall P-removal to 76%. A long anoxic phase (18 h) couldsustain denitrifying dephosphatation with less MLSS generation. Pyrosequencing data were analyzedthrough Ribosomal database project (RDP) and DECIPHER while diversity of sampling was analyzed usingChao1 and Shannon index. Rarefaction curve reflected adequacy of sampling for total species diversitystudy. Overall analysis revealed Proteobacteria, Alphaproteobacteria, Rhodobacterales, Rhodobacteraceaeand Paracoccous as the prominent phylum, class, order, family, and genus respectively.
Surplus electron donor and acceptor in anoxic phase (feasting) were advantageous for enrichment of
DNPAOs over OHOs while nitrate exhaustion in the aerobic phase provided adequate fasting conditionto maintain DNPAOs dominance. Low specific denitrification rate values in comparison to other het-erotrophs, also supported enrichment of denitrifying phosphate accumulating organisms (DNPAOs) inthe anoxic–aerobic sequencing batch reactor. Diverse micro flora ensured robustness and performancestability in high strength wastewater.© 2015 Published by Elsevier B.V.
. Introduction
Enhanced Biological Phosphorus Removal (EBPR) processesnriched with DNPAOs have been under investigation as theseicroorganisms possess equal potential for simultaneous phos-
hate uptake and nitrate removal from wastewater [1–3]. DNPAOsre capable of utilizing nitrate as electron acceptor instead of oxy-en hence the former have been enriched by introducing an anoxic
Abbreviations: COD, chemical oxygen demand; DNPAO, denitrifying phosphateccumulating organisms; OHO, other denitrifying organisms; HRT, hydraulic reten-ion time; N2O, nitrous oxide; OTU, operational taxonomic unit; PHB, poly hydroxyutyrate; PolyP, polyphosphate; RDP, ribosomal database project; SBR, sequencingatch rector; SDNR, specific denitrification rate; TSS, total suspended solid; VSS,olatile suspended solid; yHD, anoxic yield; MLSS, mixed liquor suspended solid.∗ Corresponding author. Fax: +91 674 2567160.
E-mail addresses: [email protected], [email protected] (T. Das).
ttp://dx.doi.org/10.1016/j.bej.2015.09.007369-703X/© 2015 Published by Elsevier B.V.
phase between the anaerobic–aerobic cycles of the conventionalEBPR system [4]. In general, DNPAOs uptake short chain fatty acidin anaerobic condition to store it as polyhydroxyalknoates (PHA)during which Poly-P mutilates to supply the required ATP. Thereduction equivalents are provided by glycogen through glycolyticpathway. In the following anoxic condition microbes utilize theinternally stored PHA to uptake phosphate, generally termed as lux-ury phosphate uptake, with nitrate as an electron acceptor [1,2,5,6].
Various combinations of operational phases likeanaerobic–anoxic–aerobic (AAO), anaerobic–aerobic–anoxic(AOA) [7], aerobic–anoxic condition [8] have been evaluated forthe treatment of municipal wastewater [9]. However, applicationof anoxic–aerobic treatment process in SBR systems for simulta-neous removal of nitrate, phosphate and COD, is a less discussed
phenomenon. Presence of electron acceptor in both the phasesleads to elimination of the fasting/starvation phase (anaerobic)from the system which might alter the entire process mecha-
ineeri
ncothd[i[bipatKsaethutotthlsto
iilepeitEwsb
fiNudmcis[vTpSmd
aosp
i
J. Jena et al. / Biochemical Eng
ism. Therefore, the question is could treatment of wastewaterontaining nitrate, phosphate and COD be possible in the absencef an anaerobic phase? Can PAO population be enriched withouthe presence of an anaerobic cycle? There are few reports whichave discussed that presence of nitrate in aerobic phase mightisturb the phosphorus removal efficiency [7,10]. Casey et al.11] found that under anoxic–aerobic condition nitrate did notnhibit phosphate uptake and the kinetic model of Kuba et al.12] indirectly supported phosphate uptake in presence of nitratey reporting that exhaustion of nitrate under anoxic condition
nhibits cell growth, poly-p uptake and glycogen synthesis. Anoxichosphorus uptake by denitryfying PAOs is feasible when thenoxic zone was supplied with excess nitrate load so as to exceedhe denitrifying potential of other heterotrophic organisms [13].im et al. [14] reported that Ca. accumulibacter clades coulduccessfully uptake phosphorus in the presence of nitrate. Therere also other reports on successful EBPR using nitrate as terminallectron acceptor, in SBR systems using seed sludge from sewagereatment plants [15,16]. However, performance studies usingigh strength wastewater in biological SBR systems is still anntapped area of research, that needs attention, pertaining tohe exponential growth of industries generating huge amountf nutrient rich effluents [17]. Though municipal wastewaterreatment processes are well established, specific strategies haveo be adapted to develop a sustainable method for treatment ofigh strength wastewater from pulp and paper, pharmaceutical,
eather, steel, fertilizer and dairy industries. Vibrant growth ofuch industries in a rapidly developing country like India, demandshe implication of advanced treatment processes for safe disposalf industrial effluents.
Most of the conventional nutrient removal processes operat-ng under anaerobic–anoxic conditions have usually dealt with thenternal storage compounds such as glycogen and PHB, accumu-ated by the microorganisms during the anaerobic phase, to act aslectron donors. However, the real wastewater being a more com-lex formulation will have both carbon and nutrient load in theffluent. When such effluents are exposed to treatment processes its indeed difficult to provide a condition devoid of electron accep-or (anaerobic) prior to the anoxic phase as in the conventionalBPR systems. Therefore, the current anoxic–aerobic configurationill be very crucial to evaluate the SBR performance, designed for
imultaneous nitrate and phosphate removal in the presence ofoth electron acceptor and donor at the same time.
Presence of both carbon source and nitrate will facilitate denitri-cation. During complete denitrification of NO3/NO2, intracellularO accumulation is prevented. This in turn facilitates the substratetilization rate in the subsequent aerobic period [11]. Completeenitrification depends on various factors, like availability of opti-um substrate, population density of denitrifying bacteria in the
onsortium and length of anoxic phase. Critical anoxic phase lengths considered to be an important aspect as anoxic phase of veryhort or very long time period impedes the reactor performance18,19] and denitrification by normal heterotrophs may fail to pro-ide the required advantage of phosphate removal in the SBR.herefore, population dynamics of the microbial consortium andresence of phosphate in waste effluent exposed to treatment inBR system, will evaluate the adaptability and efficiency of theixed consortium to achieve simultaneous phosphate removal and
enitrification.The experimental plan for this study was designed to evalu-
te the performance of SBR under anoxic–aerobic process in termsf simultaneous nitrate, phosphate and COD removal from high
trength synthetic wastewater representing the effluents fromharmaceutical as well as dairy industries. The main objectives are:ng Journal 105 (2016) 80–89 81
i To determine whether replacement of anaerobic phase withanoxic–oxic cycling can help in enriching Denitrifying PhosphateAccumulating Organisms (DNPAOs) in a SBR system for simulta-neous removal of nitrate, phosphate and COD from high strengthwastewater, exposed to anoxic–aerobic treatment.
ii To evaluate the effect of high COD/phosphate ratio on DNPAOpopulation in the SBR.
ii To evaluate the SBR performance and reaction dynamics in thepresence of both electron donor and acceptor in the anoxic andaerobic cycles.
iv To study the microbial diversity in the SBR to identify the dom-inant microbial species.
2. Materials and methods
2.1. Wastewater and seed sludge
The synthetic feeding medium used as influent in the reac-tor contained 6 g/L CH3COONa, 1.63 g/L KNO3,0.043 g/L of KH2PO4,1.86 g/L MgSO4, 0.38 g/L peptone and 0.3 mL of trace element solu-tion (0.15 g/L FeCl3.6H2O, 0.15 g/L H3BO3, 0.03 g/L CuSO4.5H2O,0.18 g/L KI, 0.12 g/L MnCl2 4H2O, 0.06 g/L Na2MoO4.2H2O, 0.12 g/LZnSO4 7H2O, 0.15 g/L CoCl2.H2O, 10 g/L EDTA).
The seed MLSS was collected from the anaerobic phase of abench scale 2 L SBR performing nutrient removal under anaerobic(1 h)—aerobic(4 h)—settle/decant/refill (1 h) process for more than2 months prior to the set-up of the current anoxic–aerobic process.
2.2. SBR set up
Experiments were performed in a lab scale SBR with a workingvolume of 2 L with a cycle time of 24 h under ambient tempera-ture. Keeping in view the high initial nitrate (1000 mg/L) and COD(4000 mg/L) load in the influent, each cycle consisted 18 h anoxicfollowed by 5 h aerobic period and 1 h settle/decant/refill. Pertain-ing to the long anoxic phase followed by the short aerobic phase, thecurrent SBR is termed as the LASA (Long Anoxic Short Aerobic)-SBRThe main purpose of the long anoxic phase length was:
i To provide sufficient time for complete exhaustion of the highnitrate load from the system.
ii To study the effect of a long anoxic phase on the overall reactorperformance.
iii To enrich the denitrifying population in the sludge.
Anoxic period in the LASA–SBR was maintained by purging ofNitrogen gas (purity 99.99%) and air was provided during the aer-obic stage through aquarium air bubbler. After settling period, 1 Lof the supernatant was removed, resulting in a HRT of 24 h fol-lowed by addition of 1 L synthetic wastewater to the reactor bytwo different peristaltic pumps connected to a timer. The wastingrate was 50 mL/day to keep the solid retention time (SRT) at about20 days. Synthetic wastewater and MLSS were constantly mixedwith a magnetic stirrer except for settling/decanting period. InitialpH was maintained at 7.0 (adding 1 N H2SO4) and during operationpH was recorded but not controlled.
2.3. Physicochemical analysis
Nitrate, phosphate, nitrite, ammonia, COD, pH, TSS and VSS wasmeasured according to the standard method [20].
2.4. Glycogen and PHB analysis
Activated MLSS was drawn from the SBR and analyzed byphenol–sulphuric acid technique to determine the concentration

8 ineering Journal 105 (2016) 80–89
owt((
2
bwwavspstpfm
2
S
Sr
i
a
y
wra
(
F
w
[
U
2
FchppcpL
Fig. 1. (a) phosphate removal (b) nitrate removal (c) COD removal.
Table 1Quality filters resulting in RDP.
Initialreads
Exponentialqualityfilter
Avg.lengthaftertrimming
S.D. oflength
ChimericSequences
Reads afterpreprocessing
11377 1051 394 41.26 541 9785
2 J. Jena et al. / Biochemical Eng
f internally stored glycogen [21]. Extraction and estimation of PHBas performed according to chloroform extraction method [22]
he concentration was determined through a spectrophotometerPerkin Elmer). A standard curve was prepared using purified PHBSigma Aldrich).
.5. N2O analysis
N2O emission was quantified in representative small air tightatch reactors (250 ml volume). Activated MLSS from the LASA–SBRas used as inoculums for the batch tests and anoxic conditionsere maintained by purging N2 gas before sealing the reactors
nd this was followed by removal of air from head space by aacuum pump. The reactors were then incubated in a water bathhaker (Julabo) operated at 120 rpm under room temperature. Sam-ling was done at regular intervals by drawing air from the headpace and analyzed in a Simadzu AA30 gas chromatography sys-em equipped with Electron Capture Detector. Both NIST traceablerimary and laboratory prepared secondary standards were used
or quality assurance. N2O dissolved in the reaction medium waseasured by a process mentioned elsewhere [23].
.6. Determination of reaction parameters in SBR
SDNR for the system was determined as per Eq. (1)
DNR = SNO3−1 − SNO3−2
t × VSS(1)
NO3-1, SNO3-2 are the initial and final concentration of nitrateespectively under anoxic condition.
t = duration of anoxic condition, VSS = Volatile Suspended Solidn g/L
Similarly under anoxic growth conditions yHD for the longnoxic period was determined as per Eq. (2) [24,25].
HD = 1 − 2.86(NO3)SS
(2)
here, Ss is the substrate COD that is consumed for maximumeduction of �NO3 and 2.86 is a factor that represents the O2 equiv-lent of NO3–N consumed to utilize 1 mg of substrate COD.
F/M (Food/microorganism) ratio was calculated according to Eq.3) [26].
/M = So/�X (3)
here, S0 = initial substrate (mg/L), � = HRT (h), X = VSS (mg/L)The substrate utilization rate was calculated according to Eq. (4)
27].
= (F/M) × E/100 (4)
where, U = Utilization rate (mg COD/mgVSS h),F/M = Food microorganism ratio (COD/mgVSS.h),E = Process efficiency (%)
.7. Microbial community analysis
Total DNA was isolated from MLSS (during steady state) withast DNA Spin Kit, MP Biomedicals [28]. Lysing Matrix andhaotropic buffer available in the kit were used for the optimalomogenization of the sludge sample. The released DNA was thenurified by a silica based GeneClean® SPIN procedure, with sup-
lied filter buckets and catch tubes. 16S rRNA collected from theatch tube were amplified by PCR (27F forward and 1492R reverserimer) followed by pyrosequencing at the Research and Testingaboratory, Texas (USA) to obtain 250–400 nt sequence reads.2.7.1. Post-run analysisTwo software programs, Ribosomal Database Project (RDP) [29]
and DECIPHER [30] have been used to analyze the sequencingdata. While RDP is a collection of various tools to analyze datafor microbial population from large sequencing libraries, DECIPHERhas extensive capabilities for identification of chimeric sequences.
2.7.2. PreprocessingRaw reads were treated by pyrosequencing pipeline of RDP
modules using default settings. Primers, adaptors and barcodeswere trimmed from each read. Sequence reads with poor qual-ity score were removed and DECIPHER was used to filter chimericsequences. Finally the remaining sequences reads were analyzed.Total 11,377 sequence reads were acquired from 454 pyrosequenc-ing and 9785 reads were finally selected for further investigation
(Table 1).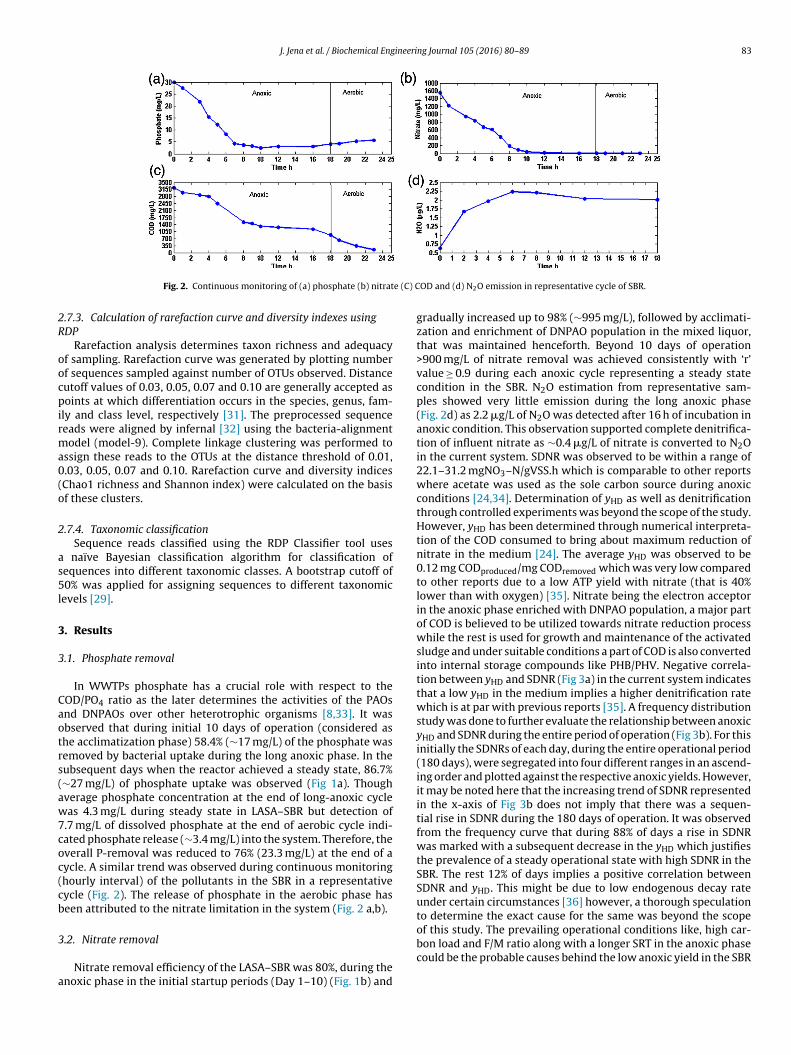
J. Jena et al. / Biochemical Engineering Journal 105 (2016) 80–89 83
e (C) C
2R
oocpirma0(o
2
as5l
3
3
Caotrs(aw7coc(cb
3
a
Fig. 2. Continuous monitoring of (a) phosphate (b) nitrat
.7.3. Calculation of rarefaction curve and diversity indexes usingDP
Rarefaction analysis determines taxon richness and adequacyf sampling. Rarefaction curve was generated by plotting numberf sequences sampled against number of OTUs observed. Distanceutoff values of 0.03, 0.05, 0.07 and 0.10 are generally accepted asoints at which differentiation occurs in the species, genus, fam-
ly and class level, respectively [31]. The preprocessed sequenceeads were aligned by infernal [32] using the bacteria-alignment
odel (model-9). Complete linkage clustering was performed tossign these reads to the OTUs at the distance threshold of 0.01,.03, 0.05, 0.07 and 0.10. Rarefaction curve and diversity indicesChao1 richness and Shannon index) were calculated on the basisf these clusters.
.7.4. Taxonomic classificationSequence reads classified using the RDP Classifier tool uses
naïve Bayesian classification algorithm for classification ofequences into different taxonomic classes. A bootstrap cutoff of0% was applied for assigning sequences to different taxonomic
evels [29].
. Results
.1. Phosphate removal
In WWTPs phosphate has a crucial role with respect to theOD/PO4 ratio as the later determines the activities of the PAOsnd DNPAOs over other heterotrophic organisms [8,33]. It wasbserved that during initial 10 days of operation (considered ashe acclimatization phase) 58.4% (∼17 mg/L) of the phosphate wasemoved by bacterial uptake during the long anoxic phase. In theubsequent days when the reactor achieved a steady state, 86.7%∼27 mg/L) of phosphate uptake was observed (Fig 1a). Thoughverage phosphate concentration at the end of long-anoxic cycleas 4.3 mg/L during steady state in LASA–SBR but detection of
.7 mg/L of dissolved phosphate at the end of aerobic cycle indi-ated phosphate release (∼3.4 mg/L) into the system. Therefore, theverall P-removal was reduced to 76% (23.3 mg/L) at the end of aycle. A similar trend was observed during continuous monitoringhourly interval) of the pollutants in the SBR in a representativeycle (Fig. 2). The release of phosphate in the aerobic phase haseen attributed to the nitrate limitation in the system (Fig. 2 a,b).
.2. Nitrate removal
Nitrate removal efficiency of the LASA–SBR was 80%, during thenoxic phase in the initial startup periods (Day 1–10) (Fig. 1b) and
OD and (d) N2O emission in representative cycle of SBR.
gradually increased up to 98% (∼995 mg/L), followed by acclimati-zation and enrichment of DNPAO population in the mixed liquor,that was maintained henceforth. Beyond 10 days of operation>900 mg/L of nitrate removal was achieved consistently with ‘r’value ≥ 0.9 during each anoxic cycle representing a steady statecondition in the SBR. N2O estimation from representative sam-ples showed very little emission during the long anoxic phase(Fig. 2d) as 2.2 �g/L of N2O was detected after 16 h of incubation inanoxic condition. This observation supported complete denitrifica-tion of influent nitrate as ∼0.4 �g/L of nitrate is converted to N2Oin the current system. SDNR was observed to be within a range of22.1–31.2 mgNO3–N/gVSS.h which is comparable to other reportswhere acetate was used as the sole carbon source during anoxicconditions [24,34]. Determination of yHD as well as denitrificationthrough controlled experiments was beyond the scope of the study.However, yHD has been determined through numerical interpreta-tion of the COD consumed to bring about maximum reduction ofnitrate in the medium [24]. The average yHD was observed to be0.12 mg CODproduced/mg CODremoved which was very low comparedto other reports due to a low ATP yield with nitrate (that is 40%lower than with oxygen) [35]. Nitrate being the electron acceptorin the anoxic phase enriched with DNPAO population, a major partof COD is believed to be utilized towards nitrate reduction processwhile the rest is used for growth and maintenance of the activatedsludge and under suitable conditions a part of COD is also convertedinto internal storage compounds like PHB/PHV. Negative correla-tion between yHD and SDNR (Fig 3a) in the current system indicatesthat a low yHD in the medium implies a higher denitrification ratewhich is at par with previous reports [35]. A frequency distributionstudy was done to further evaluate the relationship between anoxicyHD and SDNR during the entire period of operation (Fig 3b). For thisinitially the SDNRs of each day, during the entire operational period(180 days), were segregated into four different ranges in an ascend-ing order and plotted against the respective anoxic yields. However,it may be noted here that the increasing trend of SDNR representedin the x-axis of Fig 3b does not imply that there was a sequen-tial rise in SDNR during the 180 days of operation. It was observedfrom the frequency curve that during 88% of days a rise in SDNRwas marked with a subsequent decrease in the yHD which justifiesthe prevalence of a steady operational state with high SDNR in theSBR. The rest 12% of days implies a positive correlation betweenSDNR and yHD. This might be due to low endogenous decay rateunder certain circumstances [36] however, a thorough speculationto determine the exact cause for the same was beyond the scopeof this study. The prevailing operational conditions like, high car-
bon load and F/M ratio along with a longer SRT in the anoxic phasecould be the probable causes behind the low anoxic yield in the SBR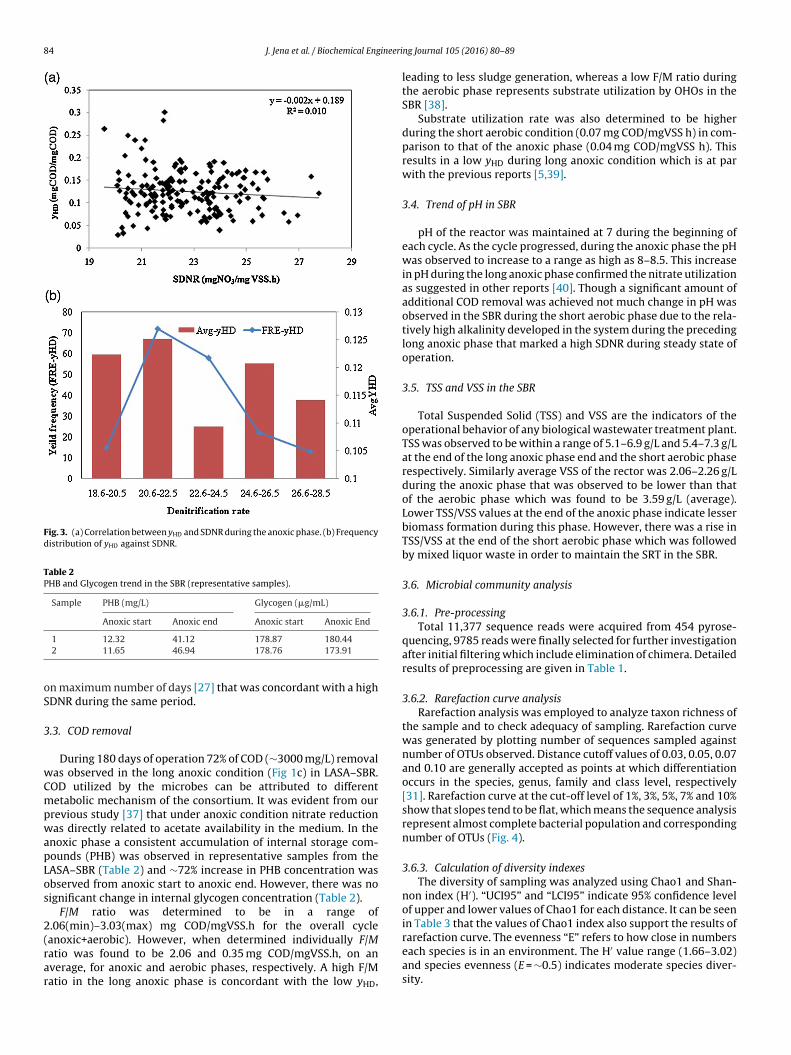
84 J. Jena et al. / Biochemical Engineeri
Fig. 3. (a) Correlation between yHD and SDNR during the anoxic phase. (b) Frequencydistribution of yHD against SDNR.
Table 2PHB and Glycogen trend in the SBR (representative samples).
Sample PHB (mg/L) Glycogen (�g/mL)
Anoxic start Anoxic end Anoxic start Anoxic End
oS
3
wCmpwapLos
2(rar
1 12.32 41.12 178.87 180.442 11.65 46.94 178.76 173.91
n maximum number of days [27] that was concordant with a highDNR during the same period.
.3. COD removal
During 180 days of operation 72% of COD (∼3000 mg/L) removalas observed in the long anoxic condition (Fig 1c) in LASA–SBR.
OD utilized by the microbes can be attributed to differentetabolic mechanism of the consortium. It was evident from our
revious study [37] that under anoxic condition nitrate reductionas directly related to acetate availability in the medium. In the
noxic phase a consistent accumulation of internal storage com-ounds (PHB) was observed in representative samples from theASA–SBR (Table 2) and ∼72% increase in PHB concentration wasbserved from anoxic start to anoxic end. However, there was noignificant change in internal glycogen concentration (Table 2).
F/M ratio was determined to be in a range of.06(min)–3.03(max) mg COD/mgVSS.h for the overall cycle
anoxic+aerobic). However, when determined individually F/Matio was found to be 2.06 and 0.35 mg COD/mgVSS.h, on anverage, for anoxic and aerobic phases, respectively. A high F/Matio in the long anoxic phase is concordant with the low yHD,ng Journal 105 (2016) 80–89
leading to less sludge generation, whereas a low F/M ratio duringthe aerobic phase represents substrate utilization by OHOs in theSBR [38].
Substrate utilization rate was also determined to be higherduring the short aerobic condition (0.07 mg COD/mgVSS h) in com-parison to that of the anoxic phase (0.04 mg COD/mgVSS h). Thisresults in a low yHD during long anoxic condition which is at parwith the previous reports [5,39].
3.4. Trend of pH in SBR
pH of the reactor was maintained at 7 during the beginning ofeach cycle. As the cycle progressed, during the anoxic phase the pHwas observed to increase to a range as high as 8–8.5. This increasein pH during the long anoxic phase confirmed the nitrate utilizationas suggested in other reports [40]. Though a significant amount ofadditional COD removal was achieved not much change in pH wasobserved in the SBR during the short aerobic phase due to the rela-tively high alkalinity developed in the system during the precedinglong anoxic phase that marked a high SDNR during steady state ofoperation.
3.5. TSS and VSS in the SBR
Total Suspended Solid (TSS) and VSS are the indicators of theoperational behavior of any biological wastewater treatment plant.TSS was observed to be within a range of 5.1–6.9 g/L and 5.4–7.3 g/Lat the end of the long anoxic phase end and the short aerobic phaserespectively. Similarly average VSS of the rector was 2.06–2.26 g/Lduring the anoxic phase that was observed to be lower than thatof the aerobic phase which was found to be 3.59 g/L (average).Lower TSS/VSS values at the end of the anoxic phase indicate lesserbiomass formation during this phase. However, there was a rise inTSS/VSS at the end of the short aerobic phase which was followedby mixed liquor waste in order to maintain the SRT in the SBR.
3.6. Microbial community analysis
3.6.1. Pre-processingTotal 11,377 sequence reads were acquired from 454 pyrose-
quencing, 9785 reads were finally selected for further investigationafter initial filtering which include elimination of chimera. Detailedresults of preprocessing are given in Table 1.
3.6.2. Rarefaction curve analysisRarefaction analysis was employed to analyze taxon richness of
the sample and to check adequacy of sampling. Rarefaction curvewas generated by plotting number of sequences sampled againstnumber of OTUs observed. Distance cutoff values of 0.03, 0.05, 0.07and 0.10 are generally accepted as points at which differentiationoccurs in the species, genus, family and class level, respectively[31]. Rarefaction curve at the cut-off level of 1%, 3%, 5%, 7% and 10%show that slopes tend to be flat, which means the sequence analysisrepresent almost complete bacterial population and correspondingnumber of OTUs (Fig. 4).
3.6.3. Calculation of diversity indexesThe diversity of sampling was analyzed using Chao1 and Shan-
non index (H′). “UCI95” and “LCI95” indicate 95% confidence levelof upper and lower values of Chao1 for each distance. It can be seenin Table 3 that the values of Chao1 index also support the results of
rarefaction curve. The evenness “E” refers to how close in numberseach species is in an environment. The H′ value range (1.66–3.02)and species evenness (E = ∼0.5) indicates moderate species diver-sity.
J. Jena et al. / Biochemical Engineering Journal 105 (2016) 80–89 85
Fig. 4. Rarefaction curve of MLSS sample at cutoff level of 1%, 3%, 5%, 7% and 10%.
Table 3The number of OTUs, Chao1 and Shanon index (H′).
Level clusters Chao1 LCI95 UCI95 H′ varH E
1% 284 316.58 301.63 344.22 3.02 0.04 × 10−2 0.533% 105 115.55 108.57 136.21 2.2 0.26 × 10−3 0.47
−3
3
ppbbiAbt(laPFt
4
nc
ldfiPdpbew
5% 66 70.5 67.03
7% 55 57.8 55.54
10% 40 41.87 40.29
.6.4. Bacterial community structureThe reads were classified into different taxa levels (from
hylum to genus) using RDP classifier at 50% threshold aser the recommendation of Cole et al. [29]. The total num-er of phyla, classes, order, family and genus were found toe 6,11,14,21 and 26 respectively. Total reads were classified
nto 4 major phyla named Proteobacteria (71%), TM7 (17.9%),ctinobacteria (5.5%) and Firmicuties (3.5%). The major class ofacteria was found to be Alphaproteobacteria (66.22%), actinobac-eria (5.55%), Betaproteobacteria (3.08%) and Gammaproteobacteria2.48%). Rhodobacteriales(62.21), Actinomycetales(5.55%) and Bacil-ales (3.29) were the dominant order in the sample. The mostbundant family and genus were Rhodobacteraceae (66.21%) andaracoccus (66.15%), respectively, The results are highlighted inig. 5 (A–E). The results of the analysis are provided in supplemen-ary material (Table S1–S5).
. Discussion
The SBR attained a steady state after 15 days of set up and 98%itrate, 72% COD and 86.7% of phosphate removal was achievedonsistently in the anoxic phase.
A high initial nitrate concentration (1000 mg/L) during theong anoxic phase did not impede anoxic phosphate uptakeue to enrichment of DNPAOs over other heterotrophic denitri-ers like GAOs (that generally compete for substrate with theAOs/DNPAOs) due to availability of nitrate and acetate in surplusuring the long anoxic phase. GAOs are also incapable to assimilate
hosphate; however, 86.7% of phosphate uptake in the LASA–SBReyond 10 days of operation also provided a clear indication of thenrichment of DNPAOs over GAOs in the system. Further the SDNRere observed to be significantly lower when compared to SDNR85.61 1.94 0.21 × 10 0.4669.49 1.9 0.19 × 10−3 0.4751.98 1.66 0.16 × 10−3 0.45
of OHOs reported elsewhere supporting dominance of DNPAOs[13]. The phosphate accumulation phenomenon was termed asluxury phosphate uptake in this case, due to the availability ofelectron donor (acetate) as well as accepter (nitrate) in surplus.However, the phosphate uptake trend breaks with onset of aero-bic phase and accounted for 3.4 mg/L of phosphate release to themedium. This is evident of a stressful condition for DNPAOs whichis also in agreement with previous reports [41] and might occurdue to two possible reasons. Firstly under nitrate limiting condi-tions (nitrate ≤ 5 mg/L in the current SBR), the dominant microfloraunable to utilize oxygen as electron acceptor slide into a stresscondition (as electron transport chain is disrupted) where internalPoly P pools break down to supply energy required for cell mainte-nance. The stress generated might be due to lack of proper electronacceptor because substrate (COD) is available in the medium inaerobic phase. It is also apparent from our previous study whereaddition of extra nitrates followed by its complete exhaustion, facil-itated phosphate uptake by the microorganisms [42]. Secondly inthe beginning of aerobic cycle pH of the medium is high (8.5) dueto active denitrification in the preceding anoxic phase. At higherpH energy demand for substrate utilization by the microorgan-ism is higher [43] which induced the breakdown of PolyP poolin the microbes. Further the dominant DNPAOs in the activatedsludge utilize nitrate as the electron acceptors instead of oxygen.However, the later (oxygen) might act as electron acceptors duringthe metabolism of other heterotrophs (since it is mixed consortia)which is supported by the COD utilization in the aerobic phase. Butthe dominant DNPAOs experience stress due to nitrate limitationand phosphate accumulated in the preceding anoxic (feast) phase
are utilized for energy leading to phosphate release in the aero-bic phase. Although the aerobic phase abets phosphate release, italso plays a crucial role in maintaining the selective dominance of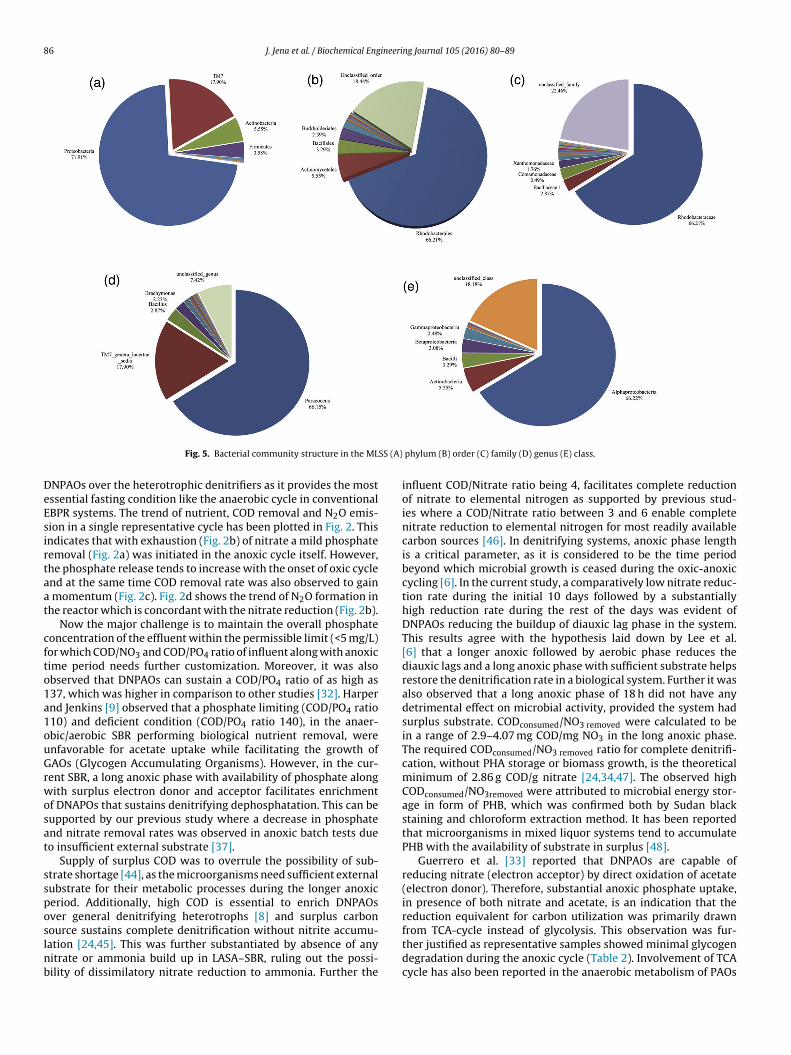
86 J. Jena et al. / Biochemical Engineering Journal 105 (2016) 80–89
SS (A)
DeEsirtaat
cfto1a1ouGrwosat
ssposlnb
Fig. 5. Bacterial community structure in the ML
NPAOs over the heterotrophic denitrifiers as it provides the mostssential fasting condition like the anaerobic cycle in conventionalBPR systems. The trend of nutrient, COD removal and N2O emis-ion in a single representative cycle has been plotted in Fig. 2. Thisndicates that with exhaustion (Fig. 2b) of nitrate a mild phosphateemoval (Fig. 2a) was initiated in the anoxic cycle itself. However,he phosphate release tends to increase with the onset of oxic cyclend at the same time COD removal rate was also observed to gain
momentum (Fig. 2c). Fig. 2d shows the trend of N2O formation inhe reactor which is concordant with the nitrate reduction (Fig. 2b).
Now the major challenge is to maintain the overall phosphateoncentration of the effluent within the permissible limit (<5 mg/L)or which COD/NO3 and COD/PO4 ratio of influent along with anoxicime period needs further customization. Moreover, it was alsobserved that DNPAOs can sustain a COD/PO4 ratio of as high as37, which was higher in comparison to other studies [32]. Harpernd Jenkins [9] observed that a phosphate limiting (COD/PO4 ratio10) and deficient condition (COD/PO4 ratio 140), in the anaer-bic/aerobic SBR performing biological nutrient removal, werenfavorable for acetate uptake while facilitating the growth ofAOs (Glycogen Accumulating Organisms). However, in the cur-
ent SBR, a long anoxic phase with availability of phosphate alongith surplus electron donor and acceptor facilitates enrichment
f DNAPOs that sustains denitrifying dephosphatation. This can beupported by our previous study where a decrease in phosphatend nitrate removal rates was observed in anoxic batch tests dueo insufficient external substrate [37].
Supply of surplus COD was to overrule the possibility of sub-trate shortage [44], as the microorganisms need sufficient externalubstrate for their metabolic processes during the longer anoxiceriod. Additionally, high COD is essential to enrich DNPAOsver general denitrifying heterotrophs [8] and surplus carbonource sustains complete denitrification without nitrite accumu-
ation [24,45]. This was further substantiated by absence of anyitrate or ammonia build up in LASA–SBR, ruling out the possi-ility of dissimilatory nitrate reduction to ammonia. Further thephylum (B) order (C) family (D) genus (E) class.
influent COD/Nitrate ratio being 4, facilitates complete reductionof nitrate to elemental nitrogen as supported by previous stud-ies where a COD/Nitrate ratio between 3 and 6 enable completenitrate reduction to elemental nitrogen for most readily availablecarbon sources [46]. In denitrifying systems, anoxic phase lengthis a critical parameter, as it is considered to be the time periodbeyond which microbial growth is ceased during the oxic-anoxiccycling [6]. In the current study, a comparatively low nitrate reduc-tion rate during the initial 10 days followed by a substantiallyhigh reduction rate during the rest of the days was evident ofDNPAOs reducing the buildup of diauxic lag phase in the system.This results agree with the hypothesis laid down by Lee et al.[6] that a longer anoxic followed by aerobic phase reduces thediauxic lags and a long anoxic phase with sufficient substrate helpsrestore the denitrification rate in a biological system. Further it wasalso observed that a long anoxic phase of 18 h did not have anydetrimental effect on microbial activity, provided the system hadsurplus substrate. CODconsumed/NO3 removed were calculated to bein a range of 2.9–4.07 mg COD/mg NO3 in the long anoxic phase.The required CODconsumed/NO3 removed ratio for complete denitrifi-cation, without PHA storage or biomass growth, is the theoreticalminimum of 2.86 g COD/g nitrate [24,34,47]. The observed highCODconsumed/NO3removed were attributed to microbial energy stor-age in form of PHB, which was confirmed both by Sudan blackstaining and chloroform extraction method. It has been reportedthat microorganisms in mixed liquor systems tend to accumulatePHB with the availability of substrate in surplus [48].
Guerrero et al. [33] reported that DNPAOs are capable ofreducing nitrate (electron acceptor) by direct oxidation of acetate(electron donor). Therefore, substantial anoxic phosphate uptake,in presence of both nitrate and acetate, is an indication that thereduction equivalent for carbon utilization was primarily drawnfrom TCA-cycle instead of glycolysis. This observation was fur-
ther justified as representative samples showed minimal glycogendegradation during the anoxic cycle (Table 2). Involvement of TCAcycle has also been reported in the anaerobic metabolism of PAOs
ineeri
erwwTPua
aaeapPlymaiPae
tiedicglictwmaoa
mlebs(Tgu
ttpttgcrmpaB
bt
J. Jena et al. / Biochemical Eng
lsewhere [49]. PHB accumulation, instead of utilization, was alsoecorded during the long anoxic phase (Table 2). It is concordantith our previous studies, where PHB accumulation was observedith increase in acetate concentration in the anoxic state [42].
herefore, it was assumed that a portion of COD was utilized forHB accumulation in the microbial consortia along with substratetilization towards dissimilatory nitrate reduction to nitrogen gasnd for the growth/maintenance of the MLSS as well.
A consistent rise in PHB concentration was observed during thenoxic phase which indicates that microbial consortia experience
feasting phase in the presence of surplus carbon along with anlectron acceptor that facilitates PHB storage, otherwise termeds luxury storage. Further it is also evident of the fact that phos-hate accumulation during the anoxic condition is independent ofHB metabolism. It was found that for Paracoccus (described in the
ater section), a dominant species in the LASA–SBR, PHB hydrol-sis might not be a prerequisite for phosphate uptake from theedium [41]. This unusual functioning might be attributed to the
daptive measures acquired by the enriched microbial populationn the presence of both electron donor and acceptor. Apart fromHB storage, presence of other heterotrophic organisms (OHOs),s evident from bioinformatics analysis, may contribute towardsxtra COD uptake than the reported stochiometric value.
It has been reported that DNPAOs adapt dynamically tohe reactor condition further facilitating phosphate removaln anaerobic–anoxic, anaerobic–anoxic–aerobic, oxic–anoxic-xtended idle processes [39,50,51]. However study of microbialiversity in SBRs operational under anoxic–aerobic conditions
s rare [11]. Reactor performance under different operationalondition is a function of microbial diversity in mixed liquor. Ineneral the upward slope on the rarefaction curve reflects that aarge number of species remain to be discovered while flatter linendicates that almost all of the species in the sample have been dis-overed. It also indicates about the adequacy of sampling requiredo discover the total species diversity in the particular ecosystemhich was shown in Fig. 4. Diversity index calculations indicateoderate species diversity in the system. This is comprehensible
s the current conditions are suitable to augment a specific arrayf microbial consortia to persistently achieve nutrient removallong with PHB accumulation.
Bacterial diversity analysis also indicates that at phylum levelost of the sequences were classified, but at the family and genus
evel they were not completely categorized. It indicates the pres-nce of some novel families (unclassified 22.46%) or genus ofacteria (unclassified genus 7.42%). The drastic decrease of unclas-ified species under genus level (7.42%) in comparison to family22.46%) is due to hierarchical assignment of TM7 population. TheM7 genera incertae sedi was only classified under phylum andenus level by RDP, which sharply decrease the percentage of thenclassified population to 7.42%.
As a contradiction to some of the previous reports, Alphapro-eobacteria instead of Betaproteobacteria [52,53] was found to behe dominant group in LASA–SBR. However, this result is also atar with some of the previous reports [54,55] where Betaproteobac-eria population in the mixed liquor were limited in comparisono Alphaproteobacteri. It was also reported [55] that Rhodocyclusroup belonging to the beta-subclass were dominant in biologi-al systems with a high phosphate load, whereas their populationeduced in low phosphate mixed liquor. This observation is very
uch in agreement with the current LASA–SBR where the phos-hate load was low, in comparison to COD, which perhaps wasdvantageous for enrichment of Alphaproteobacteria rather than
etaproteobacteria family.Paracocous, found to be the predominant genus in this reactor,elongs to Alphaproteobacteria (Fig. 4), and are capable of simul-aneous phosphate and nitrate removal in wastewater treatment
ng Journal 105 (2016) 80–89 87
system under anoxic conditions [41]. This species has also beenwidely reported to uptake short chain fatty acid and store it as(PHB) in both WWTP and single culture system [41,56]. Paracoc-cus also prefers the TCA cycle over glycogen degradation pathwayfor acetate uptake which in turn reduces phosphate release underanoxic condition. Therefore, anoxic phosphate uptake can beattributed to these unique metabolic activities of the dominantmicroorganism.
Betaproteobacteria have been frequently reported as PAOs. Sim-ilarly Rhodocyclus, Actinobactor, found to be present in the reactor,have been reported earlier as possible PAOs in full scale EBPRs[50,16]. Functioning of LASA–SBR under different condition thanthe conventional EBPR might have been the cause of the lower fre-quency of Beta-proteobacteria population. This finding again provesthat the diverse microorganism can adapt and metabolically func-tion differently under different operational condition which is atpar with previous reports [55].
Other species found in the reactor like, Azoarocous, Azonexusand Thaurea are members of family Rhodocyclaceae, and has beenreported as potential denitrifiers abundantly found in municipalWWTP performing nitrogen and phosphorus removal [57]. Thesedenitrifiers are also capable of internal PHA storage by using acetateas a substrate. Availability of both DNPAOs and PAOs indicates thatthe mixed liquor is a true consortium of different microorganismsand provides operational flexibility without compromising withperformance.
Gammaproteobacteria found in the mixed liquor are reportedas the prime competitor of PAOs/DNPAOs for substrate [58].Some microbes (unidentified) of Gammaproteobacteria class wereinterestingly confirmed by staining techniques, as capable of accu-mulating PHB however, it lacks the ability to recycle phosphate [59].Acenatobactor a genus of Gamma-proteobacteria found minutely(0.16%) in this system were reported for polyphosphate accu-mulation with nitrate as electron acceptor [60]. Population ofGamma-proteobacteria is very limited in this reactor. Therefore, theprobable disturbance caused by this group to the reactor perfor-mance is negligible. Pseudomonas detected in this study has beenreported previously which required external carbon supplementa-tion, unlike the putative DNPAOs, to uptake phosphate and nitrate[41,50].
A large portion of the sequences were identified as TM7,reported frequently in wastewater treatment plants [58,61], pos-sess the capability of using both oxygen and nitrate as electronacceptor and have a limited ability to store short chain fatty acidas PHB. TM7 referred as uncultured filamentous bacteria are oftenreported to be present in low numbers in WWTP [62]. They werealso reported to be incapable of Poly P as well as PHB uptake, byTEM analysis [63]. However, these observations may need furtherclarifications.
It was confirmed that Actinobactor could accumulate phosphatein full scale EBPR [50] and was also reported as the most abundantmicrobe instead of typical PAOs in WWTP [64]. Presence of Acti-nobactor as one of the major populations in the current system canalso be supported by previous reports [62]. Corynebacterium (order-actinomycetels) has also been reported earlier as a PAO [65,66].
5. Conclusion
Surplus electron donor and acceptor in anoxic phase (feastingphase) was advantageous for enrichment of DNPAOs while nitrateexhaustion in the aerobic phase provided an adequate fasting state
to maintain their dominance in MLSS. Hence nitrate, phosphate andCOD removal could be achieved simultaneously without an anaero-bic cycle. Diverse micro flora ensured robustness and performancestability in high strength wastewater.
8 ineeri
tautce
C
A
istDat
A
t
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
8 J. Jena et al. / Biochemical Eng
DNPAOs could be enriched in the MLSS under suitable condi-ions and simultaneous nitrate, phosphate and COD removal waschieved from high strength wastewater in a single SBR systemnder a long anoxic short aerobic phase. Further the results ofhis study will facilitate future research on setting up of biologi-al processes for treatment of nutrient rich high strength industrialffluents.
onflict of interest
The authors have no conflict of interest to declare.
cknowledgement
Ms. Jyotsnarani Jena is thankful to CSIR and Mr. Ravindra Kumars thankful to Department of Biotechnology (DBT) for their fellow-hips. Authors are grateful to SETCA–CSIR for funding. Authors alsohank the Head, Environment & Sustainability Department and theirector, CSIR–IMMT, Bhubaneswar for their encouragement. Theuthors are extremely grateful to the two anonymous reviewers forheir constructive comments and suggestions.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at http://dx.doi.org/10.1016/j.bej.2015.09.007.
eferences
[1] J. Ahn, T. Daidou, S. Tsuneda, A. Hirata, Characterization of denitrifyingphosphate accumulating organisms cultivated under different electronacceptor conditions using polymerase chain reaction-denaturing gradient gelelectrophoresis assay, Water Res. 36 (2002) 403–412.
[2] F. Freitas, M. Temudo, M.A.M. Reis, Microbial population response to changesof the operating condition in a dynamic nutrient removal sequencing batchreactor, Bioprocess. Biosyst. Eng. 28 (2005) 199–209.
[3] J.A. Baeza, A. Guisasola, J. Guerrero, water online Guest Column Sept (2014)Environ. Microbiol. 10.1111/1462-2920.12614.
[4] H.N. Chang, R.K. Moon, B.G. Park, S.J. Lim, D.W. Choi, W.G. Lee, S.L. Song, Y.H.Ahn, Simultaneous sequential batch reactor(SBR) operation for simultaneousremoval of nitrogen and phosphorous, BioPro Eng. 23 (2000) 513–521.
[5] R.J. Seviour, T. Mino, M. Onuki, The microbiology of biological phosphorousremoval in activated sludge systems, FEMS Microbiol. Rev. 27 (2003) 99–127.
[6] S.H. Lee, S. Svoronos, B. Koopman, Influence of alternating oxic/anoxicconditions on growth of denitrifying bacteria, Water Res. 44 (2010)1819–1824.
[7] S. Tsuneda, T. Ohno, K. Soejima, A. Hirata, Simultaneous nitrogen andphosphorous removal using denitrifying phosphate accumulating organismsin sequencing batch reactor, Biochem. Eng. J. 27 (2005) 191–196.
[8] J. Shi, X. Lu, R. Yu, Q. Gu, Y. Zho, Influence of wastewater composition onnutrient removal behaviors in the new anaerobic–anoxic/nitrifying/inducedcrystallizationprocess, Saudi J. Biol. Sci. 21 (2014) 71–80.
[9] W.F. Harper, D. Jenkins, The effect of an initial anaerobic zone on the nutrientrequirements of activated sludge, Water Environ. Res. 75 (2003) 216–224.
10] J.P. Kerrn-Jespersen, M. Henze, Biological phosphorus uptake under anoxicand aerobic conditions, Water Res. 27 (1993) 617–624.
11] T.G. Casey, M.C. Wentzel, G.A. Ekama, R.E. Loewenthal, G.R. Marais, Ahypothesis for the cause and control of anoxic–aerobic (AA) filament bulkingin nutrient removal activated sludge systems, Water Sci. Technol. 29 (1994)203–212.
12] T. Kuba, E. Murnleitner, M.C.M. Van Loosdrecht, J.J. Heijnen, A metabolicmodel for the biological phosphorus removal by denitrifying organisms,Biotech. Bioeng. 52 (1996) 685–695.
13] Z. Hu, M.C. Wentzel, G.A. Ekama, Anoxic growth of phosphate accumulatingorganisms (PAOs) in biological nutrient removal activated MLSS systems,Water Res. 36 (19) (2002) 4927–4937.
14] J.M. Kim, H.J. Lee, D.S. Lee, C.O. Jeon, Characterization of thedenitrification-associated phosphorus uptake properties of CandidatusAccumulibacter phosphatis clades in sludge subjected to enhanced biologicalphosphorus removal, Appl. Environ. Microbiol. 79 (6) (2013) 1969–1979.
15] R.J. Zeng, A.M. Saunders, Z. Yuan, L.L. Blackall, J. Keller, Identification and
comparison of aerobic and denitrifying polyphosphate accumulatingorganisms, Biotechnol. Bioeng. 83 (2003) 140–148.16] A. Oehmen, P.C. Lemos, G. Carvalho, Z.G. Yuan, J. Keller, L.L. Blackall, M.A.M.Reis, Advances in enhanced biological phosphorus removal: from micro tomacro scale, Water. Res. 41 (2007) 2271–2300.
[
ng Journal 105 (2016) 80–89
17] R. Kaur, S.P. Wani, A.K. Singh, K. Lal, Wastewater Production, Treatment andUse in India, Country report India, 2012.
18] O.A. Fadali, M.M. Nassar, I.E. Ibrahim, I.A. Ashour, N. Shehata, Effect oflag-phase in the transition from oxic to anoxic conditions on the performanceof the sequencing batch reactor, Int. J. Innov. Appl. Stud. 7 (2014) 393–400.
19] Y. Linjiang, H. Wei, W. Lei, Y. Yongzhe, W. Zhiying, Simultaneous denitrifyingphosphorus accumulation in a sequencing batch reactor, Front. Environ. Sci.Eng. I (1) (2007) 23–27, http://dx.doi.org/10.1007/s11783-007-0004-6.
20] American Public Health Association (APHA) Standard methods for theexamination of water and wastewater, 18th ed., Washington, DC, USA.
21] S. Lo, J.C. Russell, A.W. Taylor, Determination of glycogen in small tissueample, J. Appl. Phys. 28 (2) (1970) 234–236.
22] S.M. Kumar, S.N. Mudliar, K.M.K. Chakrabarti, Production of biodegradableplastics from activated sludge generated from a food processing industrialwastewater treatment plant, Bioresour. Technol. 95 (2004) 327–330.
23] S. Ray, A. Mohanty, S.S. Mohanty, S. Mishra, G.R. Choudhury, Optimization ofbiological elimination of ammonia and chemichal oxygen demand fromwastewater using response surface methodology, Clean Soil Air Water 41(9999) (2014) 1–7.
24] D. Guven, Effects of different carbon sources on denitrification efficiencyassociated with culture adaptation and C/N ratio, Clean 7 (2009) 565–573.
25] H. Melcer, P. Dold, R.M. Jones, C.M. Bye, I. Takacs, et al., Methods for WaterCharacterization in Activated Sludge Modeling, Water Environment ResearchFoundation, Alexandria, VA, USA, 2003.
26] D. Dionisi, M. Majone, R. Ramadori, M. Becarri, The storage of acetate underanoxic condition, Water Res. 35 (2001) 2661–2668.
27] G. Tchobanoglous, F.L. Burton, H.D. Stensel, third ed, in: Metcalf & Eddy,Wastewater engineering; Treatment disposal reuse, McGraw-Hill bookcompany, New York, USA, 2003.
28] Y. Wang, J. Geng, Z. Ren, G. Guo, C. Wang, H. Wang, Effect of COD/N and COD/Pon the PHA transformation and dynamics of microbial community structurein a denitrifying phosphorous removal process, J. Chem. Technol. Biotecnol.88 (2012) 1228–1236.
29] J.R. Cole, Q. Wang, E. Cardenas, J. Fish, B. Chai, R.J. Farris, A.S.Kulam-Syed-Mohideen, D.M. McGarrell, T. Marsh, G.M. Garrity, J.M. Tiedje,The Ribosomal Database Project: improved alignments and new tools forrRNA analysis, Nucleic. Acids. Res. 37 (2009).
30] E.S. Wright, L.S. Yilmaz, D.R. Noguera, DECIPHER-A search-based approach tochimera identification for 16S rRNA sequences, Appl. Environ. Microb. 78(2011) 717–725.
31] S. Samantaray, J.K. Nayak, N. Mallick, ater utilization forPoly-ß-Hydroxylbutyrate production by Cyanobacterium AulosiraFertilissimain a recirculatory aquaculture system, Appl. Environ. Microb. 77 (2011)8735–8743.
32] E.P. Nawrocki, S.R. Eddy, Query-dependent banding (QDB) for faster RNAsimilarity searches, PLoS Comput. Biol. 3 (2007), http://dx.doi.org/10.1371/journal.pcbi.0030056.
33] J. Guerrero, A. Guisasola, J.A. Baeza, The nature of the carbon source rules thecompetition between PAO and denitrifiers in systems for simultaneousbiological nitrogen and phosphorous removal, Water Res. 45 (2011)4793–4802.
34] Y. Fernandez-Nava, E. Maranon, J. Soons, L. Castrillon, Denitrification of highnitrate concentration wastewater using alternative carbon source, J. Hazard.mater. 173 (2010) 682–688.
35] A. Muller, M.C. Wentzel, R.E. Lowenthal, G.A. Ekama, Heterotrophic anoxicyield in anoxic aerobic activated sludge system treating municipalwastewater, Water Res. 37 (2003) 2435–2441.
36] A. Li, G. Wu, Effects of sludge retention times on nutrient removal and nitrousoxide emission in biological nutrient removal processes Int, Int. J. Environ.Res. Public Health 11 (2014) 3553–3569.
37] J. Jena, S. Ray, S. Pandey, T. Das, Effect of COD/N ratio on simultaneousremoval of nutrients and COD from synthetic high strength wastewater underanoxic conditions, J. Sci. Ind. Res. 72 (2013) 127–131.
38] V. Tandoi, D. Jenkins, J. Wanner, Activated Sludge Separation Problems:Theory, Control Measures, in: Practical Experiences, IWA Publishing, 2006,2015.
39] T. Kuba, M.C.M. Van Loosdrecht, E. Murnleitner, J. Heijnen, Occurrence ofdenitrifying phosphorus removing bacteria in modified UCT-type wastewatertreatment plants, Water Res. 31 (1997) 777–786.
40] T. Watanabe, H. Motoyama, M. Kuroda, Denitrification and neutralizationtreatment by direct feeding of an acidic wastewater containing copper ionand high-strength nitrate to a bio-electrochemical reactor process, Water Res.17 (2001) 4102–4110.
41] Y. Barak, J. Rijn, A typical polyphosphate accumulation by the denitrifyingbacterium Paracocous denitrificance, Appl. Env. Microbiol. (2000) 1209–1212.
42] J. Jena, R. Kumar, A. Dixit, S. Pandey, T. Das, Evaluation of simultaneousnutrient and COD removal with Poly hydroxybutyrate accumulation usingmixed microbial consortia under anoxic condition and their bioinformaticsanalysis, Plos One 10 (2) (2015) 1–20, http://dx.doi.org/10.1371/journal.pone.0116230.
43] C.O. Jeon, D.S. Lee, M.W. Lee, J.M. Park, Enhanced biological phosphorous
removal in an anaerobic–aerobic sequencing batch reactor: effect of pH,Water Environ. Res. 73 (2001) 301–306.44] E.R. Coats, A. Mockos, F.J. Loge, Post-anoxic denitrification driven by PHA andglycogen within enhanced biological phosphorus removal, Bioresour. Tech.102 (2011) 1019–1027.

ineeri
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[microbial community in a full-scale enhanced biological phosphorus removalplant, Microbiology. 153 (2007) 4061–4073.
[66] M. Hu, X. Wang, X. Wen, Y. Xia, Microbial community structure in differentwastewater treatment plants as revealed by 454-pyrosequencing analysis,
J. Jena et al. / Biochemical Eng
45] S. Sozen, D. Orhon, The effect of nitrite correction on the evaluation of the rateof nitrate utilization under anoxic condition, J. Chem. Technol. Biotechnol. 74(1999) 790–800.
46] J.V. Rijn, Y. Tal, H.J. Schreier, Denitrification in recirculating systems: theoryand applications, Aquacult. Eng. 34 (2006) 364–376.
47] M.C.M. Van Loosdrecht, M.A. Pot, J.J. Heijnen, Importance of bacterial storagepolymers in bioprocess, Water Sci. Technol. 35 (1997) 41–47.
48] M.V. Reddy, S.V. Mohan, Influence of aerobic and anoxic microenvironmentson polyhydroxyalkanoates (PHA) production from food waste and acidogeniceffluents using aerobic consortia, Bioresour. Technol. 103 (2012) 313–321.
49] Y. Zhou, M. Pijuan, R.J. Zeng, Z. Yuan, Involvement of the TCA cycle inanaerobic metabolism of phosphate accumulating organisms (PAOs), WaterRes. 43 (5) (2009) 1330–1340.
50] S.Y. Gebremariam, M.W. Beutel, D. Christian, T.F. Hess, Research advances andchallenges in the microbiology of enhanced biological phosphorousremoval-a critical review, Water Environ. Res. 83 (2011) 195–219.
51] X.M. Li, H. Chena, Q. Yanga, D. Wanga, K. Luoa, G.-M. Zenga, Biologicalnutrient removal in a sequencing batch reactor operated asoxic/anoxic/extended-idle regime, Chemosphere 105 (2014) 75–81.
52] P. Bond, R. Erhart, M. Wagner, J. Keller, L.L. Blackall, Identification of some ofthe major groups of bacteria in efficient and non-efficient biologicalphosphorus removal activated sludge systems, Appl. Environ. Microbiol. 65(1999) 4077–4084.
53] Y.H. Kong, S.L. Ong, W.J. Ng, W.T. Liu, Diversity and distribution of a deeplybranched novel proteobacterial group found in anaerobic-aerobic activatedsludge process, Environ. Microbiol. 4 (2002) 743–757.
54] R.P.X. Hesselmann, C. Werlen, D. Hahn, J.R. van der Meer, A.J.B. Zehnder,Phylogenetic Analysis and Detection of a Bacterium that Performs EnhancedBiological Phosphate Removal in Activated Sludge, Syst. Appl. Microbiol. 22(1999) 454–465.
55] T. Mino, Microbial selection of polyphosphate-accumulating bacteria in
activated sludge wastewater treatment process for enhanced biologicalphosphate removal, Biochemistry 65 (2000) 341–348.56] S.T. Ferguson, D.G. Gadian, Evidence from 31P magnetic resonance thatpolyphosphate synthesis is a slip reaction in Paracoccusdenitrificance,Biochem. Soc. Trans. 7 (1979) 176–179.
ng Journal 105 (2016) 80–89 89
57] T. Thomsen, Y. Kong, P.H. Nielsen, Ecophysiology of abundant denitrifyingbacteria inactivated sludge, FEMS Microbiol. Ecol. 60 (2007) 370–382.
58] H.P. Nielsen, A.T. Mielczarek, J.L. Neilsen, A.M. Saunders, Y. Kong, A.A. Hansen,J. Vollertsen, A conceptual ecosystem model of microbial communities inenhanced biological phosphorus removal plants, Water Res. 44 (2010)5070–5088.
59] W. Liu, K.D. Linning, K. Nakamura, T. Mino, T. Matsuo, L.J. Forney, Microbialcommunity changes in biological phosphate-removal systems on alteringsludge phosphorus content, Microbiology 146 (2000) 1099–1107.
60] G.C. Knight, R.J. Seviour, J.A. Soddell, S. McDonnell, R.C. Bayly, Metabolicvariation among strains of acinetobacter isolated from activated sludge,Water Res. 29 (1995) 2081–2084.
61] Y. Xia, Y. Kog, T.R. Thomson, P.H. Nielsen, Identification and ecophysiologicalcharacterization of epiphytic protein-hydrolyzing SaprospiraceaeCandidataus Epiflobacter, Appl. Environ. microbial. 74 (2008) 2229–2238.
62] T. Zhang, M.F. Shao, L. Ye, 454 Pyrosequencing reveals bacterial diversity ofactivated sludge from 14 sewage treatment plants, ISME J. 6 (2012)1137–1147.
63] P. Hugenholtz, G.W. Tyson, R.I. Webb, A.M. Wagner, L.L. Blackall, Investigationof candidate division TM7. a recently recognized major lineage of the domainbacteria with no known pure-culture representatives, Appl. Environ.Microbiol. 67 (2001) 411–419.
64] M. Beer, H.M. Staratton, P.C. Griffiths, R.J. Seviour, Which are thepolyphosphate accumulating organism in full scale activated sludgeenhanced biological phosphate removal system in Astralia? J. Appl. Microbiol.100 (2006) 233–243.
65] Y. Kong, X. Xia, J.L. Nielsen, P.H. Nielsen, Structure and function of the
Bioresour. Technol. 11 (7) (2012) 72–79.