behavioral adaptations to parasites: an ethological approach
TRANSCRIPT

Behavioral Adaptations to Parasites: An Ethological ApproachAuthor(s): Benjamin L. HartSource: The Journal of Parasitology, Vol. 78, No. 2 (Apr., 1992), pp. 256-265Published by: The American Society of ParasitologistsStable URL: http://www.jstor.org/stable/3283472 .
Accessed: 24/09/2013 20:21
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The American Society of Parasitologists is collaborating with JSTOR to digitize, preserve and extend access toThe Journal of Parasitology.
http://www.jstor.org
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions

J. Parasitol., 78(2), 1992, p. 256-265
? American Society of Parasitologists 1992
BEHAVIORAL ADAPTATIONS TO PARASITES: AN ETHOLOGICAL APPROACH*
Benjamin L. Hart Department of Physiological Sciences, School of Veterinary Medicine, University of California, Davis, California 95616
ABSTRACT: Wild vertebrate animals must live in an environment with the ever present threat of internal and external parasites. This threat by macroparasites is responsible for the natural selection of an array of behavioral adaptations that, together with the immune system and other physiological forms of resistance, enable the animals to survive and reproduce in this environment. Several lines of research, some quite recent, illustrate that specific behavioral patterns can be effective in helping animals or their offspring avoid or control macro- parasites that can affect adversely the animal's fitness. These behavioral patterns fall under the general strategies of avoidance behavior and mate selection.
Various aspects of social, feeding, and repro- ductive behavior of vertebrates are shaped by the forces of predation and resource limitation that animals must address to survive to repro- ductive age and successfully rear young. A rel- atively unappreciated force shaping behavior in much this same way is the existence of external and internal parasites. Animals that are studied by scientists usually are maintained in relatively clean laboratories, field stations, and domestic environments, where it is easy to forget that the animals evolved and thrived in environments with an array of parasites long before human protective measures were available. The behav- ioral patterns that animals may use to avoid, control or eliminate parasites or pathogens from their bodies can be categorized into 5 strategies (Hart, 1990). The first comprises those behaviors that enable animals to avoid or minimize their exposure to parasites. The second strategy is that of controlled exposure by which animals or their offspring may be exposed to a small dose of a particular pathogen or parasite resulting in de- velopment of immunity. A third strategy in- volves the behavioral patterns of anorexia and depression that are seen when animals are sick with a febrile disease. This behavior of the sick animal appears to potentiate the fever response in enabling the animal to overcome infections. The fourth strategy relates to the tendency of some animals to help group mates or kin that are sick or injured. The fifth strategy, and one that has received much recent attention, is that in which animals select mates on the basis of
evidence for resistance to parasite infestation thus providing offspring with the genetic basis for re- sisting parasites. These strategies focus on the ability of individual animals to live well enough to assure survival of their offspring to reproduc- tive age. The existence of behavioral strategies does not imply that individuals remain free of parasites or never succumb to parasitism. How- ever, an animal may carry a parasite load that does not affect its health, but when there are intense demands on body resources, such as in escaping from predators or undergoing a seasonal nutritional stress, a moderate or light parasite load may play a role in the animal's survival. As in avoiding predators, the fact that an animal may succumb to parasites does not mean that the parasite control strategies are ineffective any more than antipredator strategies may be con- sidered ineffective because an animal eventually succumbs to predation. Conversely, the display of behavioral strategies for the control of para- sites does not require that the parasites always be present any more than antipredator behavior requires experiencing predators.
In this paper I will discuss primarily the par- asite avoidance and mate selection strategies be- cause these are the most relevant for controlling parasites (macroparasites). The other 3 strategies are mostly involved in controlling viruses and bacteria, the so-called microparasites (Anderson and May, 1979). Another review (Hart, 1990) has dealt with all 5 strategies with regard to both microparasites and macroparasites.
To accept a particular behavioral pattern as having a parasite control function, 2 criteria must be met. One of these criteria is that the parasite in question must have a detrimental effect on the host's fitness. This may be difficult to demon- strate for macroparasites in that a few intestinal worms may be harmful only in times of socially
Received 5 July 1991; accepted 29 September 1991. * Paper from symposium on neurobiology of hel-
minths given at the 1991 annual meeting of the American Society of Parasitologists.
256
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions

HART-BEHAVIORAL ADAPTATIONS TO PARASITES 257
50 soo
40
30 so
2 4 6 8 10 12 14
Weeks After Initiation of Larval Feeding
FIGURE 1. Growth in 4-mo-old lambs given daily infections with 4,000 Ostertagia circumcinta larvae. This level of infection did not cause clinical symptoms. The intake restricted graph represents a group in which food intake in uninfected lambs was matched to that of infected lambs (data from Sykes and Coop [1977]). Circles, ad libitum control; squares, intake restricted; triangles, infected.
related or nutritional stress or in conjunction with occasional physical demands such as escaping from a predator or fighting with a conspecific animal. When animals are in competition, in es- caping from predators or fighting over territory for example, just a minor difference in parasite load may influence the outcome and hence in- fluence fitness. It should be remembered that in nature animals sustain infestations from several types of macroparasites, and it is the cumulative effect of parasites that constitutes the fitness cost.
The second criterion is that a behavior must be effective in helping the animal avoid or re- move the parasite in question. It is apparent that almost any parasite control behavior represents some cost in fitness to the animal from loss of feeding time, energy utilized, or distraction from predator vigilance. Presumably, the greater the cost of the parasite control behavior to the ani- mal the more beneficial should be the behavior.
Even though a host's behavior and immune system may be effective in protecting it from parasites, there are instances where the parasite's presence looks deceptively harmless because there is a standoff between the parasite's invasiveness and host resistance. Thus, any reduction in the host's defenses (including behavior) could be dis- astrous. This paper is devoted to illustrating the variety of behavioral patterns that have been shown to be effective in enabling vertebrate an- imals to reduce their exposure or vulnerability to helminth or arthropod parasites. However, in keeping with the first criterion above it is useful first to illustrate the types of costs to animals that
may be incurred with relatively light or moderate macroparasitic loads. It is maintained that be- havioral patterns operate not just to prevent out- right debilitating parasite loads, but to keep par- asite numbers to a minimum.
COSTS OF MACROPARASITES
Much of the work on the effects of intestinal and external parasites involves domestic animals where these issues are of economic concern. Growing animals may suffer impaired growth rate and reproductive efficiency with levels of intestinal parasites that do not necessarily result in clinical signs of disease. This is illustrated in a series of experiments on lambs infected with a gastric nematode (Ostertagia circumcinta) at a level reportedly low enough that the infections were considered clinically insignificant; there were no clinical signs that could be used to separate infected from uninfected sheep. After 14 wk the growth rate of infected lambs was about 50% that of controls (Fig. 1). Furthermore, protein and calcium deposition were much less than 50% of controls (Sykes and Coop, 1977).
Some helminth infections may impair an an- imal's fitness even though body weight may not be affected. In an experimental infection of mice with Trichinella spiralis, at levels of infection that did not produce any enduring change in body weight (Rau, 1983), open field activity and abil- ity to defend a territory against an intruder could still be impaired (Fig. 2).
With regard to external parasites much atten- tion is drawn to these parasites as vectors of disease. Although vector-borne diseases may be of importance to livestock introduced into cer- tain areas, wild animals in their native habitat usually are immune to these diseases so it would be difficult to assess the cost of such vector-borne diseases. For wild animals the direct cost of blood removed and depression of appetite could be the most important. Two examples serve to illustrate such costs. From a study of horses in upstate New York, investigators estimated that on a typ- ical summer day a horse could be bitten by as many as 4,000 tabanid flies that collectively would remove 0.5 liters of blood (Tashiro and Schwardt, 1953). A number of studies have analyzed the decrements in the growth of young animals from engorging ticks. Several studies, including one shown here (Fig. 3) have documented that 1 en- gorging tick (Boophilus microplus) produces a decrement in weight gain of about 0.6 kg on an annualized basis (Little, 1963; Seebeck et al.,
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions

258 THE JOURNAL OF PARASITOLOGY, VOL. 78, NO. 2, APRIL 1992
100 -
80-
60-
Larvae: 0 100 200 400 0 100 200 400 Resident 0 Resident 200 Resident 400 Intruder 0 Intruder 0 Intruder 100
Open Field Body Weight
Percent of Control Percent Wins Over Intruder
O .IN. I
(D 0 ;' iiii~~l~ll~i! 4 0
-.,i~i;iii;!llii~ - ~ ~ ~ ~ ~ ~ ~ X x;;•ii~~~~~~~~~~
• -•-• iiiiiiii............il
20- ae: 0 100 200 40 0 100 200 400 Resient 0*Resident*200 esident 400%*- Intrud r 0 Inruder
. .....er...
Larva: P0r10 e200400 010020o 400Pesident0 Weidnts200versidntruder
FIGURE 2. Effects of different doses of Trichinella spiralis larvae on body weight, open field activity, and winning fights over intruders in mice. Mice were 16-wk-old CD 1 (n = 8 per group) outbred males orally inoculated with different doses of larvae. Bars show measures or tests performed during the late chronic phase of T. spiralis infection (90-100 days postinoculation). Body weight did not differ at 90 days, but during the acute phase of infection mice treated with 400 larvae weighed significantly less. Open field tests (distance traveled) revealed differences between control (no larvae) and the 100- and 200-dose groups and between the 400-dose group and the 2 lighter dose groups (P < 0.01). Tests involving challenging a treated resident mouse with an untreated or lightly treated intruder revealed that residents failed to win over the intruders with lighter or no treatment more than would be expected (P < 0.005). Normally, residents would be expected to win 100% of the time under the test condition (data from Rau [1983]).
1971; Turner and Short, 1972). For animals such as slender gazelles or impalas on the East African plains that face the risk of predation daily, where lactating females must put out extra nutrients for nourishing their young, and where intermale competition for territory depends on size and endurance, the costs of just a half dozen engorg- ing ticks could result in failure to escape from the predator (especially as part of a group) or to win a territory.
THE AVOIDANCE STRATEGY
The behavioral patterns involved in the avoid- ance of internal and external macroparasites in- clude feeding, elimination, grooming, fly repel- ling, evasive behavior, grouping, and nest fumigation. In all of these areas there are at least some data that document the effectiveness of the avoidance strategy. Some selected examples will be discussed below.
Selective foraging and elimination
A relatively widely acknowledged behavioral pattern of grazing mammals that reduces their exposure to infectious stages of internal parasites is selective grazing where the animals avoid graz- ing next to areas that have been recently contam- inated with fecal material even though the forage may be quite lush. This behavior has been de- scribed for sheep, horses, and cattle, and it pre- sumably occurs in other wild and domestic ru- minants as well (Taylor, 1954; Odberg and Francis-Smith, 1977). Despite this widely rec- ognized behavior, there are surprisingly little data to support this presumption. One quantitative study was conducted by Michel (1955), who sam- pled pasture areas for lungworm larvae (Dictyo- caulus viviparus) and found that selective grazing by cattle reduces their intake of infectious larvae to 25% of what would have been consumed ran- domly (Fig. 4).
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions

HART-BEHAVIORAL ADAPTATIONS TO PARASITES 259
S460 Infestation UReversed
420
o 380 ---- S380- Clean
- 340 --- Infested 4)
34
0
cc 301 E * 28
26 0 4 8 12 16 20 24 28 32 36 40
Weeks After Infestation
FIGURE 3. Effects of tick infestation on growth rate and hematocrit of growing cattle. Infested animals car- ried an average of 50 engorging ticks; clean cattle were treated to eliminate ticks. The overall suppression of body weight per engorging tick was 0.6 kg on an an- nualized basis. When the infestation was reversed, pre- viously infected cattle were treated and clean cattle artificially infested with ticks (data from Little [1963]).
Related to the topic of selective foraging is that of selective elimination, which avoids contam- ination with fecal-borne parasites. Horses, for example, use dunging areas in places that sel- dom are grazed (Odberg and Francis-Smith, 1977). Everyone who has raised a dog or cat is familiar with the behavior of animals that have nests or dens in being fastidious in not soiling these areas. Protection of the young from para- sitic larval exposure around the nest or den would be particularly important because the young are the most severely affected by internal parasites. The predisposition to keep the living area clean by eliminating away from the nest is evident in piglets when they are 4-5 days old (Buchenuer et al., 1982; Whatson, 1985) and in young pup- pies as soon as they are capable of walking (Ross, 1950; Rheingold, 1963).
Grooming
One of the most frequently and regularly per- formed behavioral patterns of rodents, felids, ru- minants, and primates is grooming. Laboratory rats have been found to spend up to one-third of their waking time grooming (Bolles, 1960). Depending upon species, antelope on the East African plains deliver 80-200 oral bouts of grooming, consisting of 600-2,000 grooms per 12-hr day (Hart and Hart, 1988). Clearly an im- portant function of grooming is removal of ec- toparasites. As a study by Bennett (1969) illus- trated, prevention of oral grooming in cattle by
100 90
p 80 S70
S60
Random Immediately Recently Foraged Foraged
Parasitic Larvae in Forage Sampled
S50
S40 o30 0)20
10r 0
Random Immediately Recently Foraged Foraged
Parasitic Larvae in Forage Sampled
FIGURE 4. Avoidance by cattle of forage contami- nated with feces and lungworm larvae (Dictyocaulus viviparus) as shown in samples from different regions of pastures grazed by cattle. Larval content, as ex- pressed as a percentage of that from random samples, is shown for regions analyzed immediately after being grazed by cattle and regions grazed recently by cattle (data from Michel [1955]).
means of a harness allows a tick load (B. micro- plus) to increase by more than 4 times over that of control animals without harnesses (Fig. 5). Prevention of grooming in cattle allows lice (Li- nognathus vituli; Damalinia bovis) to increase by about 6-fold over control levels; the louse load was reduced to baseline within 3 days after grooming was allowed once again (Lewis et al., 1967). Whereas cattle groom with their tongues, other ruminants, such as antelope, use the lower lateral incisors and canine teeth as a dental comb in grooming with an upward scraping action. The dental modification, which serves no function in grazing or browsing, is apparently effective in removing parasites (McKenzie, 1990). In impala that live in environments with many ticks, there may be excessive attrition on the dental comb, such that animals carry much higher parasite loads and are in poorer condition than normal impala (McKenzie, 1990).
Mice sustaining limb paralysis or limb am- putation, which prevents scratching, became in- fested with lice (Polyplax serrata) to about 30 times the control level when exposed to a seed population (Bell et al., 1966). Prevention of oral grooming by placing a spacer between the inci- sors used in combing the pelage results in an increase in lice to about 60 times that of controls (Murray, 1961, 1987). Infestation with mites (Myobia musculi) also increases when mice are prevented from grooming (Wiesbroth et al., 1974).
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions
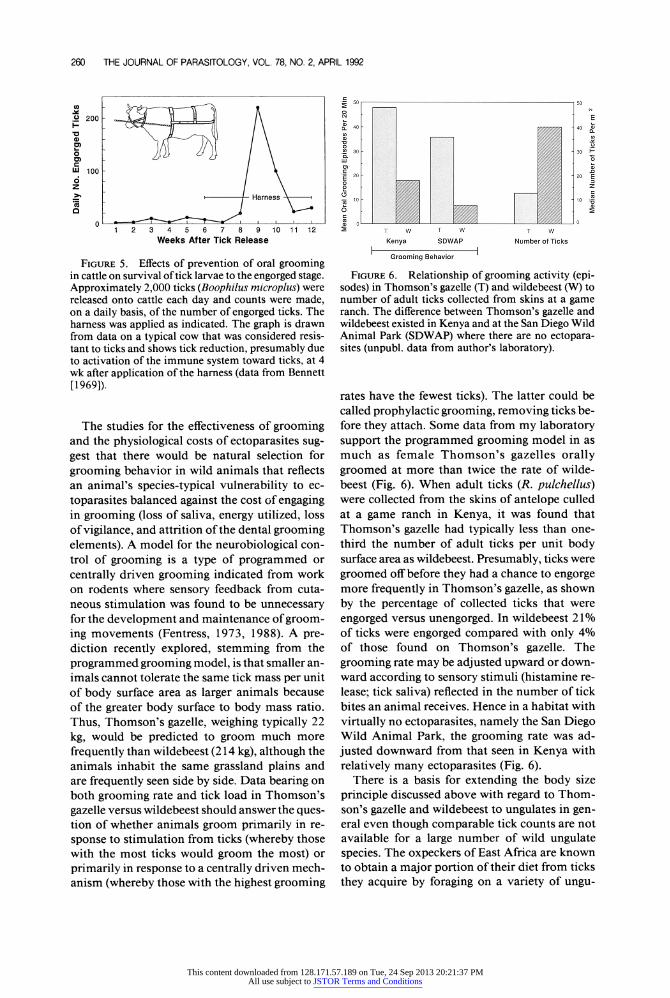
260 THE JOURNAL OF PARASITOLOGY, VOL. 78, NO. 2, APRIL 1992
.9 200 I--
a) 0)
W 100
z
1000 Harness
0 1 2 3 4 5 6 7 8 9 10 11 12
Weeks After Tick Release
FIGURE 5. Effects of prevention of oral grooming in cattle on survival of tick larvae to the engorged stage. Approximately 2,000 ticks (Boophilus microplus) were released onto cattle each day and counts were made, on a daily basis, of the number of engorged ticks. The harness was applied as indicated. The graph is drawn from data on a typical cow that was considered resis- tant to ticks and shows tick reduction, presumably due to activation of the immune system toward ticks, at 4 wk after application of the harness (data from Bennett [1969]).
The studies for the effectiveness of grooming and the physiological costs of ectoparasites sug- gest that there would be natural selection for
grooming behavior in wild animals that reflects an animal's species-typical vulnerability to ec- toparasites balanced against the cost of engaging in grooming (loss of saliva, energy utilized, loss of vigilance, and attrition of the dental grooming elements). A model for the neurobiological con- trol of grooming is a type of programmed or centrally driven grooming indicated from work on rodents where sensory feedback from cuta- neous stimulation was found to be unnecessary for the development and maintenance of groom- ing movements (Fentress, 1973, 1988). A pre- diction recently explored, stemming from the
programmed grooming model, is that smaller an- imals cannot tolerate the same tick mass per unit of body surface area as larger animals because of the greater body surface to body mass ratio. Thus, Thomson's gazelle, weighing typically 22
kg, would be predicted to groom much more frequently than wildebeest (214 kg), although the animals inhabit the same grassland plains and are frequently seen side by side. Data bearing on both grooming rate and tick load in Thomson's gazelle versus wildebeest should answer the ques- tion of whether animals groom primarily in re- sponse to stimulation from ticks (whereby those with the most ticks would groom the most) or primarily in response to a centrally driven mech- anism (whereby those with the highest grooming
50 50
N E 40 40 0
a n
0 o .T 30 -30 W"
20 20 E 0o z ? 10 10 :5
o 0
T w T W T W
Kenya SDWAP Number of Ticks
Grooming Behavior
FIGURE 6. Relationship of grooming activity (epi- sodes) in Thomson's gazelle (T) and wildebeest (W) to number of adult ticks collected from skins at a game ranch. The difference between Thomson's gazelle and wildebeest existed in Kenya and at the San Diego Wild Animal Park (SDWAP) where there are no ectopara- sites (unpubl. data from author's laboratory).
rates have the fewest ticks). The latter could be called prophylactic grooming, removing ticks be- fore they attach. Some data from my laboratory support the programmed grooming model in as much as female Thomson's gazelles orally groomed at more than twice the rate of wilde- beest (Fig. 6). When adult ticks (R. pulchellus) were collected from the skins of antelope culled at a game ranch in Kenya, it was found that Thomson's gazelle had typically less than one- third the number of adult ticks per unit body surface area as wildebeest. Presumably, ticks were groomed off before they had a chance to engorge more frequently in Thomson's gazelle, as shown by the percentage of collected ticks that were engorged versus unengorged. In wildebeest 21% of ticks were engorged compared with only 4% of those found on Thomson's gazelle. The grooming rate may be adjusted upward or down- ward according to sensory stimuli (histamine re- lease; tick saliva) reflected in the number of tick bites an animal receives. Hence in a habitat with virtually no ectoparasites, namely the San Diego Wild Animal Park, the grooming rate was ad- justed downward from that seen in Kenya with relatively many ectoparasites (Fig. 6).
There is a basis for extending the body size principle discussed above with regard to Thom- son's gazelle and wildebeest to ungulates in gen- eral even though comparable tick counts are not available for a large number of wild ungulate species. The oxpeckers of East Africa are known to obtain a major portion of their diet from ticks they acquire by foraging on a variety of ungu-
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions

HART-BEHAVIORAL ADAPTATIONS TO PARASITES 261
100 100 Calf 2 Avoiding Actions
0 0 a 80 -,pCalf 1 Flies 80
z
o0 -0
S60 60 cm0 0
? 40 40
0 M >0
6 20 C~alf 1 Avoiding Actions 2 6
20 odi20 0
z0 C 'Calf 2 Flies
-E" 00 1 2 3 4 5 6 7 8 9 10
Time in min FIGURE 7. Effect of avoidance behavior by Friesian calves (tail flicks, foot stamps, head-swings) on number
of flies observed on the calves at the end of each minute. Data are shown for a sample of 2 calves paired in the same environment when 100 Stomoxys calcitrans flies were released into the experimental chamber (70 m2 x 2.5 m high). Calf 2, with the most avoiding actions, had the fewest flies (data from Warnes and Finlayson [1987]).
lates. It stands to reason that the greater the tick mass per unit ofbody surface area for an ungulate species, the more profitable it is for tick birds to forage on that species. Thus, if a species of larger animals harbors a greater tick mass per unit of body surface area, more oxpeckers would be ex- pected to attend animals of that species. In sup- port of this concept, a regression analysis of counts of red-billed oxpeckers foraging for ticks on dif- ferent species of African ungulates revealed a positive relationship between oxpeckers per unit of body surface area and species-typical body mass (Hart el al., 1990).
Fly repelling behavior
Animals may repel or dislodge biting flies by ear twitching, head-tossing, leg stamping, muscle flicking, muscle twitching, and tail switching (Okumura, 1977; Harvey and Launchbaugh, 1982). When fly biting intensity is high, fly re- pelling or dislodging behavior increases (Harris et al., 1987). Data actually documenting the ef- fectiveness of fly repelling activities are few. In a study by Warnes and Finlayson (1987) pairs of calves were exposed to the same number of biting flies; the calf that engaged in the most fly repelling actions had the fewest flies (Fig. 7).
Evasive behavior
Animals may lie down as a means of avoiding flies as revealed by observations on reindeer showing that when harassed by head flies, rein- deer more than doubled their time lying down (Espmark and Langvatn, 1979). The investiga- tors maintained that by lying down the animals do not dislodge flies from foliage, produce less carbon dioxide or other chemoattractants, and present a reduced area of exposed body surface. Seeking a microhabitat where there are fewer bit- ing flies also has been observed in feral horses that move to windy ridges during the time of day when flies are particularly bad even though for- age is less desirable (Keiper and Berger, 1982). During periods of intense fly activity, caribou are known to move to a higher altitude where there are fewer flies (Downes et al., 1986).
Gadding behavior is one of the more drastic forms of evasive behavior. This behavior occurs in cattle and some wild cervids upon the ap- proach of warble or hypodermal flies that deposit eggs on the hairs of the legs and lower abdomen. The would-be hosts act wild and run frantically in apparent attempts to evade the flies (Soulsby, 1982; Schmidt and Roberts, 1985). Interestingly, this gadding behavior in response to warble flies,
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions
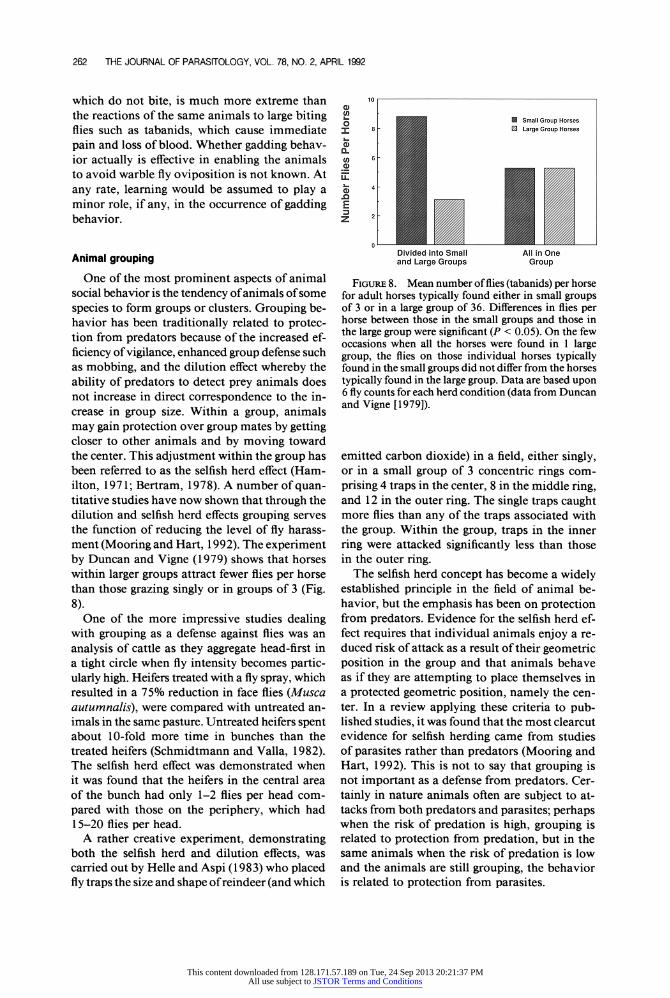
262 THE JOURNAL OF PARASITOLOGY, VOL. 78, NO. 2, APRIL 1992
which do not bite, is much more extreme than the reactions of the same animals to large biting flies such as tabanids, which cause immediate pain and loss of blood. Whether gadding behav- ior actually is effective in enabling the animals to avoid warble fly oviposition is not known. At any rate, learning would be assumed to play a minor role, if any, in the occurrence of gadding behavior.
Animal grouping
One of the most prominent aspects of animal social behavior is the tendency of animals of some species to form groups or clusters. Grouping be- havior has been traditionally related to protec- tion from predators because of the increased ef- ficiency of vigilance, enhanced group defense such as mobbing, and the dilution effect whereby the ability of predators to detect prey animals does not increase in direct correspondence to the in- crease in group size. Within a group, animals may gain protection over group mates by getting closer to other animals and by moving toward the center. This adjustment within the group has been referred to as the selfish herd effect (Ham- ilton, 1971; Bertram, 1978). A number of quan- titative studies have now shown that through the dilution and selfish herd effects grouping serves the function of reducing the level of fly harass- ment (Mooring and Hart, 1992). The experiment by Duncan and Vigne (1979) shows that horses within larger groups attract fewer flies per horse than those grazing singly or in groups of 3 (Fig. 8).
One of the more impressive studies dealing with grouping as a defense against flies was an analysis of cattle as they aggregate head-first in a tight circle when fly intensity becomes partic- ularly high. Heifers treated with a fly spray, which resulted in a 75% reduction in face flies (Musca autumnalis), were compared with untreated an- imals in the same pasture. Untreated heifers spent about 10-fold more time in bunches than the treated heifers (Schmidtmann and Valla, 1982). The selfish herd effect was demonstrated when it was found that the heifers in the central area of the bunch had only 1-2 flies per head com- pared with those on the periphery, which had 15-20 flies per head.
A rather creative experiment, demonstrating both the selfish herd and dilution effects, was carried out by Helle and Aspi (1983) who placed fly traps the size and shape of reindeer (and which
10 a
o
o ]
Small Group Horses S 8
[ Large Group Horses
6
M 2 0)
Divided into Small All in One and Large Groups Group
FIGuRE 8. Mean number of flies (tabanids) per horse for adult horses typically found either in small groups of 3 or in a large group of 36. Differences in flies per horse between those in the small groups and those in the large group were significant (P < 0.05). On the few occasions when all the horses were found in 1 large group, the flies on those individual horses typically found in the small groups did not differ from the horses typically found in the large group. Data are based upon 6 fly counts for each herd condition (data from Duncan and Vigne [1979]).
emitted carbon dioxide) in a field, either singly, or in a small group of 3 concentric rings com- prising 4 traps in the center, 8 in the middle ring, and 12 in the outer ring. The single traps caught more flies than any of the traps associated with the group. Within the group, traps in the inner ring were attacked significantly less than those in the outer ring.
The selfish herd concept has become a widely established principle in the field of animal be- havior, but the emphasis has been on protection from predators. Evidence for the selfish herd ef- fect requires that individual animals enjoy a re- duced risk of attack as a result of their geometric position in the group and that animals behave as if they are attempting to place themselves in a protected geometric position, namely the cen- ter. In a review applying these criteria to pub- lished studies, it was found that the most clearcut evidence for selfish herding came from studies of parasites rather than predators (Mooring and Hart, 1992). This is not to say that grouping is not important as a defense from predators. Cer- tainly in nature animals often are subject to at- tacks from both predators and parasites; perhaps when the risk of predation is high, grouping is related to protection from predation, but in the same animals when the risk of predation is low and the animals are still grouping, the behavior is related to protection from parasites.
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions

HART-BEHAVIORAL ADAPTATIONS TO PARASITES 263
Nest fumigation for parasites
Animal movement to avoid or reduce the level of infestation with parasites is not the only way animals may reduce their exposure to ectopar- asites. A virtually unexplored area of parasite control is represented by work on the phenom- enon of nest fumigation by European starlings that weave fresh plant material into their nests. Plants contain a variety of biocidal chemicals that protect them from parasites. The starling is one of those kinds of animals that reuse nest sites and, as such, are susceptible to parasites that may build up in the nests. Clark and Mason (1985) noted that in collecting fresh green material to add to the nest matrix at the beginning of the breeding season, a few plants were selected much more frequently than would have been expected by chance. In laboratory studies it was found that the hatching of louse eggs (Menacanthus sp.) was retarded by extracts from the selected plants much more than by extracts from nonpreferred plants. A further laboratory study involved placing pre- ferred plants in a plastic bag with nest matrix material taken from nest boxes. The emergence of larval mites (Ornithonyssus sylvarum) from the nest matrix was delayed by the preferred plants but not by nonpreferred plants (Clark and Mason, 1988). Field experiments in which fresh plant material was removed from the matrix of starling nests resulted in an increase in mites (Clark and Mason, 1988). The degree to which nest fumigation may play a role in the nest build- ing and maintenance by other avian species and burrowing mammals that reuse old nests remains to be examined.
MATE SELECTION FOR PARASITE RESISTANCE
I have touched upon a few examples illustrat- ing the strategy by which animals may avoid or reduce their exposure to macroparasites. Strat- egies involving controlled exposure that poten- tiates the immune system, the behavior of ani- mals when they are sick, and the care of sick or injured animals relate mostly to control of mi- croparasites and are reviewed elsewhere (Hart, 1990). The strategy of mate selection (usually a function of female behavior) is relevant to the control of macroparasites. Fitness of females may be enhanced directly by avoiding contacting or
sharing nests with mates that are carrying heavy ectoparasite loads and indirectly by choosing mates with genes for resistance to parasites. It is difficult to separate the proximate benefit of an
animal avoiding parasitized mates from the more ultimate benefit of providing offspring with genes for resistance to parasites, and quite likely both functions go together. Some examples serve to illustrate that this type of selection occurs at least in avian species. Sage grouse with noticeable in- festations of lice were found to secure fewer cop- ulations than those free of these parasites (Boyce, 1990). Rock doves infested with lice (Columbi- cola columbae) were chosen less often by females than were uninfested males (Clayton, 1990). That animals may be judging resistance to parasites by signs other than the visual presence of para- sites is indicated in an experiment on jungle fowl that were infected experimentally with an intes- tinal nematode (Ascaridia galli). Infected males were chosen by females only half as much as uninfected males (Zuk et al., 1990). Of course, in these studies a confounding issue is the degree to which parasitism influences how a male fares in competing for territory or females. Both satin bowerbirds (Borgia, 1986) and red grouse (Jen- kins et al., 1963) that are free of ectoparasites are more likely to have territories than those that carry parasites. The effect of a parasite in influ- encing a male's success in acquiring access to females undoubtedly acts simultaneously in the same direction as the parasite's effect on the at- tractiveness of the male to females. A theoretical approach that has gathered considerable atten- tion was presented by Hamilton and Zuk (1982). They suggested that ornamental secondary sex- ual characteristics or showy displays, by which females sometimes select mates, may be indi- cators of a male's freedom from parasites. From recent studies there is some confirming evidence for this perspective. In sage grouse, louse infes- tation causes hematomas on air sacs, which are used as ornaments in courtship displays. Females choose males that have artificially applied he- matomas on their air sacs less often than control males (Boyce, 1990). Red jungle fowl that have had a history of infection with intestinal nema- todes are selected by females less frequently than uninfected males. This selection is based upon ornament-related characteristics of shorter tail feathers, paler hackle feathers, and duller and shorter combs (Zuk et al., 1990). The long tail feathers of barn swallows are a sexually dimor- phic trait that is used by females when choosing mates, and recent evidence reveals that the in- crease of tail length from one year to another is negatively related to experimentally manipulat- ed mite numbers in nests during the preceding
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions

264 THE JOURNAL OF PARASITOLOGY, VOL. 78, NO. 2, APRIL 1992
season (Moller, 1990). Resistance of swallows to mites appears to be heritable in that cross-fos- tering experiments show that the mite loads are more closely related to the mite infestations of parents than to the number of mites in the nests.
CONCLUSIONS
Animals in the natural environment must live with an ever present threat of parasitism. This threat is viewed as being responsible for the nat- ural selection of behavioral adaptations, some examples of which are reviewed here. These ex- amples undoubtedly just scratch the surface of the total inventory of behavioral patterns that exist. Probably, animals do not knowingly or cognitively employ the control strategies, al- though they may be enhanced or shaped by learn- ing. The control strategies may be particularly important in enhancing fitness in marginal sit- uations, such as in producing behaviorally com- petitive offspring in the shortest time possible or in competing with conspecific animals in running from a predator. The behavior of mate selection provides a basis for choosing partners that max- imizes the genetic endowment for parasite resis- tance in offspring.
What have not been covered in this paper are the counterstrategies coevolving in parasites. These counterstrategies of parasites provide physiological and behavioral mechanisms that allow the parasite to evade the behavioral and physiological resistance of the hosts.
LITERATURE CITED
ANDERSON, R. M., AND R. M. MAY. 1979. Population biology of infectious diseases: Part I. Nature 280: 361-367.
BELL, J. F., C. M. CLIFFORD, G. J. MOORE, AND G. RAYMOND. 1966. Effects of limb disability on lousiness in mice. III. Gross aspects of acquired resistance. Experimental Parasitology 18: 49-60.
BENNETT, G. F. 1969. Boophilus microplus (Acarina: Ixodidae): Experimental infestations on cattle re- strained from grooming. Experimental Parasitol- ogy 26: 323-328.
BERTRAM, B. C. R. 1978. Living in groups: Predators and prey. In Behavioral ecology, an evolutionary approach, J. R. Krebs and N. B. Davies (eds.). Sinauer, Sunderland, Massachusetts, p. 64-97.
BOLLES, R. C. 1960. Grooming behavior in the rat. Journal of Comparative Physiological Psychology 53:306-310.
BORGIA, G. 1986. Satin bowerbird parasites: A test of the bright male hypothesis. Behavioral Ecology and Sociobiology 19: 355-358.
BOYCE, M. S. 1990. The red queen visits sage grouse leks. American Zoologist 30: 263-270.
BUCHENUER, D., C. LuFr, AND A. GRAUVOGL. 1982.
Investigations on the eliminative behavior of pig- lets. Applied Animal Ethology 9:153-164.
CLARK, L., AND J. R. MASON. 1985. Use of nest ma- terial as insecticidal and anti-pathogenic agents by the European starling. Oecologia 67: 169-176.
, AND . 1988. Effect of biologically ac- tive plants used as nest material and the derived benefit to starling nestlings. Oecologia 77: 174- 180.
CLAYTON, D. 1990. Mate choice in experimentally parasitized rock doves. American Zoologist 30: 251-262.
DOWNES, C. M., J. B. THEBERGE, AND S. M. SMITH. 1986. The influence of insects on the distribution, microhabitat choice, and behavior of the Burwash caribou herd. Canadian Journal of Zoology 64: 622-629.
DUNCAN, P., AND N. VIGNE. 1979. The effect of group size in horses on the rate of attacks by blood- sucking flies. Animal Behavior 27: 623-625.
ESPMARK, Y., AND R. LANGVATN. 1979. Lying down as a means of reducing fly harassment in red deer (Cervus elaphus). Behavioral Ecology and Socio- biology 5: 51-54.
FENTRESS, J. C. 1973. Development of grooming in mice with amputated forelimbs. Science 179: 704- 705.
.1988. Expressive contexts, fine structure and control mediation of rodent grooming. Annals of the New York Academy of Science 525: 18-26.
HAMILTON, W. D. 1971. Geometry for the selfish herd. Journal of Theoretical Biology 31: 295-311.
, AND M. ZUK. 1982. Heritable true fitness and
bright birds: A role for parasites? Science 218: 384- 387.
HARRIS, J. A., J. E. HILLERTON, AND S. V. MORANT. 1987. Effect on milk production of controlling muscid flies, and reducing fly-avoidance behav- iour by use of Fenvalerate ear tags during the dry period. Journal of Dairy Research 54: 165-171.
HART, B. L. 1990. Behavioral adaptations to patho- gens and parasties. Neuroscience and Biobehav- ioral Reviews 14: 273-294.
, L. A. HART, AND M. S. MOORING. 1990. Dif-
ferential foraging of oxpeckers on impala com- pared with sympatric antelope species. African Journal of Ecology 28: 240-249.
HART, L. A., AND B. L. HART. 1988. Autogrooming and social grooming in impala. Annals of the New York Academy of Science 525: 399-402.
HARVEY, T. L., AND J. L. LAUNCHBAUGH. 1982. Effect of horn flies on behavior of cattle. Journal of Eco- nomic Entomology 75: 25-27.
HELLE, T., AND J. AsPI. 1983. Does herd formation reduce insect harassment among reindeer? A field experiment with animal traps. Acta Zoologica Fennica 175: 129-131.
JENKINS, D. A., A. WATSON, AND G. R. MILLER. 1963. Population studies on red grouse, Lagopus lagopus scoticus (Lath.), in north-east Scotland. Journal of Animal Ecology 32: 317-376.
KEIPER, R. R., AND J. BERGER. 1982. Refuge-seeking and pest avoidance by feral horses in desert and island environments. Applied Animal Ethology 9: 111-120.
LEWIS, L. F., D. M. CHRISTENSON, AND G. W. EDDY.
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions

HART-BEHAVIORAL ADAPTATIONS TO PARASITES 265
1967. Rearing the long-nosed cattle louse and cat- tle biting louse on host animals in Oregon. Journal of Economic Entomology 60: 755-757.
LITTLE, D. A. 1963. The effect of cattle tick infesta- tion on the growth rate of cattle. Australian Vet- erinary Journal 39: 6-10.
MCKENZIE, A. A. 1990. The ruminant dental groom- ing apparatus. Zoological Journal of the Linnean Society 99: 117-128.
MICHEL, J. F. 1955. Parasitological significance of bovine grazing behaviour. Nature 175:1088-1089.
MOLLER, A. P. 1990. Effects of a haematophagous mite on the barn swallow (Hirundo rustica): A test of the Hamilton and Zuk hypothesis. Evolution 44:771-784.
MOORING, M. S., AND B. L. HART. 1992. The selfish herd: Predators and parasites. Behaviour (in press).
MURRAY, M. D. 1961. The ecology of the louse Poly- plax serrate (Burm.) on the mouse Mus musculus L. Australian Journal of Zoology 9:1-11.
. 1987. Effects of host grooming on louse pop- ulations. Parasitology Today 3: 276-278.
ODBERG, F. O., AND K. FRANCIS-SMITH. 1977. Studies on the formation of ungrazed eliminative areas in fields used by horses. Applied Animal Ethology 3: 27-34.
OKUMURA, T. 1977. The relationship of attacking fly abundance to behavioral responses of grazing cat- tle. Japanese Journal of Applied Entomology and Zoology 21: 119-122.
RAU, M. E. 1983. Loss of behavioral dominance in male mice infected with Trichinella spiralis. Par- asitology 88: 371-373.
RHEINGOLD, H. L. 1963. Maternal behavior in the dog. In Maternal behavior in mammals, H. L. Rheingold (ed.). John Wiley & Sons, Inc., New York, p. 169-202.
Ross, S. 1950. Some observations of the lair dwelling behavior of dogs. Behaviour 2: 144-162.
SCHMIDT, G. D., AND L. S. ROBERTS. 1985. Foun- dations of parasitology. Times Mirror/Mosby Col- lege Publishing, St. Louis, 775 p.
SCHMIDTMANN, E. T., AND M. E. VALLA. 1982. Face- fly pest intensity, fly-avoidance behavior (bunch- ing) and grazing time in Holstein heifers. Applied Animal Ethology 8: 429-438.
SEEBECK, R. M., P. H. SPINGELL, AND J. C. O'KELLY. 1971. Alterations in host metabolism by the spe- cific and anorectic effects of the cattle ticks (Booph- ilus microplus). I. Food intake and body weight growth. Australian Journal of Biological Sciences 24: 373-380.
SOULSBY, E. J. L. 1982. Helminths, arthropods and protozoa of domesticated animals, 7th ed. Lea and Febiger, Philadelphia, 809 p.
SYKES, A. R., AND R. L. COOP. 1977. Chronic para- sitism and animal efficiency. ARC Research Re- views 3: 41-46.
TASHIRO, H., AND H. H. SCHWARDT. 1953. Biological studies of horse flies in New York. Journal of Eco- nomic Entomology 46: 813-822.
TAYLOR, E. L. 1954. Grazing behavior and helmin- thic disease. British Journal of Animal Behavior 2: 61-62.
TURNER, H. C., AND A. J. SHORT. 1972. Effects of field infestations of gastrointestinal helminths and of the cattle ticks (Boophilus microplus) on growth of three breeds of cattle. Australian Journal of Ag- ricultural Research 23: 177-193.
WARNES, M. L., AND L. H. FINLAYSON. 1987. Effect of host behaviour on host preference in Stomoxys calcitrans. Medical and Veterinary Entomology 1: 53-57.
WHATSON, T. S. 1985. Development of eliminative behavior in piglets. Applied Animal Behavioral Science 14: 365-377.
WIESBROTH, S. H., S. FRIEDMAN, AND M. POWELL. 1974. The parasitic ecology of the rodent mite Myobia mulculi. I. Grooming factors. Laboratory Animal Science 24: 510-516.
ZUK, M., R. THORNHILL, AND J. D. LIGON. 1990. Par- asites and mate choice in red jungle fowl. Amer- ican Zoologist 30: 235-244.
This content downloaded from 128.171.57.189 on Tue, 24 Sep 2013 20:21:37 PMAll use subject to JSTOR Terms and Conditions