baroreceptor control of heart rate in the awake toad: peripheral autonomic effectors and arterial...
TRANSCRIPT

Journal of the Autonomic Nervous System 80 (2000) 31–39www.elsevier.com/ locate / jans
Baroreceptor control of heart rate in the awake toad: peripheral autonomiceffectors and arterial baroreceptor areas
*Leila Maria Bianchi-da-Silva, Leda Menescal-de-Oliveira, Anette Hoffmann˜ ˜ ˜ ˜Department of Physiology, Faculty of Medicine of Ribeirao Preto, University of Sao Paulo, 14 049-900 Ribeirao Preto, Sao Paulo, Brazil
Received 17 May 1999; received in revised form 4 October 1999; accepted 25 October 1999
Abstract
Systemic injection of sodium nitroprusside (30 mg/kg, i.v.) in the awake Bufo paracnemis toad induced a fall in arterial blood pressureand tachycardia. This tachycardia, but not the hypotension, was significantly reduced in toads with bilateral electrolytic lesion of thecaudal and commissural regions of the solitary tract nucleus and in animals with transection of the spinal cord, 2 mm below the obex. Thisindicates that the tachycardia is reflex, depends on the integrity of the solitary tract nucleus and is due to descending spinal autonomicactivation. Pretreatment with propranolol (4 mg/kg, i.v.) significantly reduced the tachycardia but did not block it completely, showingthe importance of beta-adrenoceptors in its genesis. The reflex increase in heart rate induced by nitroprusside was not statisticallysignificant in animals with bilateral section of the laryngeal nerve, whose baroreceptor fibers originate from the pulmocutaneous artery orin animals in which the bilateral section of the laryngeal nerve was performed together with section of the glossopharyngeal nerves, whichincorporate fibers originating from the carotid labyrinth. The reduction of the reflex tachycardia was significant in toads with aortic archdenervation alone or combined with section of the laryngeal nerves or in animals with complete denervation of the three baroreceptorsareas. These results suggest that the region of the aortic arch, when submitted to unloading, is the most important baroreceptor zone forcardiac compensation in toads. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Anuran amphibians; Toads; Sodium nitroprusside; Baroreceptor; Reflex tachycardia; Solitary tract nucleus
1. Introduction (Ishii et al., 1966; Kusakabe, 1990; Kusakabe, 1992;Poullet, 1977; Rogers, 1964). However, the pressure
Anurans, like other vertebrates so far tested (Bagshaw, required to bring about baroreceptor fiber discharge was1985), have the ability to compensate for transient changes far higher than the normal blood pressure (Ishii et al.,in systemic arterial pressure, a fact that makes them more 1966; Segura, 1979; Smith et al., 1981). Some studies haveapt to respond to environmental stress. This compensation shown that the pulmocutaneous artery of anurans harboursimplies the existence of receptors sensitive to variations in baroreceptors (Hoffmann and Cordeiro de Sousa, 1982;arterial pressure, strategically located on the walls of the Ishii and Ishii, 1971; Smits et al., 1986; Van Vliet andgreat vessels. In anurans, some investigators have attempt- West, 1987b) whose primary function is to protect theed to locate and characterize these receptors based on lungs from excessive hydrostatic pressure (West and vananalogy with mammals. Thus, some studies were con- Vliet, 1983). This population of baroreceptors may beducted to detect the presence of baroreceptors in the wall considered to be homologous with the pulmonary arterialof the carotid labyrinth, which resembles externally the baroreceptor population in mammals (Van Vliet and West,carotid sinus of mammals. Internally, it is formed by a 1987a). Stimulation of the central stump of the laryngealdense network of vessels whose function seems to be to nerve containing fibers from the pulmocutaneous artery,dampen the pressure pulse to the internal carotid artery occlusion of the artery with perivascular balloons, or(Adams, 1958; Carman, 1955; Toews et al., 1982). In distension by direct perfusion induce reflex systemicaddition to this vascular regulatory function, the labyrinth cardiovascular changes in toads of the genus Bufo (Hoff-has been found to have chemo- and baroreceptor functions mann and Cordeiro de Sousa, 1982; Smith et al., 1981;
West and van Vliet, 1983). Ishii et al. (1985) showed aorticnerve firing in synchrony with fluctuations in arterial*Corresponding author. Tel.: 155-16-6023332; fax: 155-16-6330017.
E-mail address: [email protected] (A. Hoffmann) pressure in the anaesthetized or pithed Bufo vulgaris.
0165-1838/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0165-1838( 99 )00083-1

32 L.M. Bianchi-da-Silva et al. / Journal of the Autonomic Nervous System 80 (2000) 31 –39
However, based on other functional and anatomical find- laterally to the vertebral column in its lumbar portion forings in the same study, they consider the aortic arch to be a exposure of the iliac artery that was occlusively cannulatedpreponderantly chemoreceptor area. Thus, it is possible with a silastic cannula. The anterior abdominal vein wasthat this region harbours baroreceptors, although it is not also cannulated through an incision in the abdominal skinknown whether they are active in unanaesthetized prepara- at the midline level. The cannulae were filled withtions. heparinized saline (0.2 IU/mI) and exteriorised through
Contrary to structures located in the prosencephalon, the surgical incision. The two incisions were closed withmore caudal regions of the brain are phylogenetically sutures and the respective cannulae were fixed to the skinconserved in vertebrates. The fasciculus and the nucleus of to prevent twisting.the solitary tract are already well differentiated in anuransand receive mainly visceral afferences from the vagus andglossopharyngeal nerves and, to a lesser extent, from the 2.2. Spinal sectionfacial and the trigeminus nerves (Nieuwenhuys andOpdam, 1976). However, the topographic distribution of Some animals were submitted to transection of thethese afferents within the solitary tract fasciculus is little spinal cord immediately below the obex or to shamknown. The most rostral portion of the fasciculus is operation. The animals were anaesthetized and the 4thoccupied by gustatory projections in Rana catesbeiana ventricle was exposed after making a midline incision in(Hanamori and Ishiko, 1987; Herrick, 1944). In contrast to the cervical region, separating the skin and breaking thethe numerous studies that show the importance of the processes of the first cervical vertebrae. The spinal cordsolitary tract nucleus in baroreflex control in mammals, was transected with ophthalmic scissors 2 mm below thelittle is known about its importance in non-mammalian obex under visual control through a Zeiss stereoscopicvertebrates. Electrical or chemical stimulation of this microscope. Only the animals in which the transectionregion induces cardiovascular changes in anaesthetized B. proved to be complete and which did not bleed excessivelyparacnemis (Cordeiro de Souza and Hoffmann, 1982). were used. After the experiments the animals were deeply
The purpose of the present study was to validate a anaesthetized and again examined under the microscope tomethod of low invasiveness to induce the baroreflex in reconfirm the precision of the transections.awake toads. We opted for a method, currently used inmammals that evaluates the ability of the baroreceptorsystem to compensate for systemic changes in arterial 2.3. Bilateral lesion of the solitary tract nucleuspressure caused by drugs by means of changes in heartrate. Using this method, we set out to determine the The procedure described earlier for the exposure of theparticipation of caudal portions of the solitary tract nucleus 4th ventricle was also used in the animals submitted toand of the different barosensitive areas in baroreflex bilateral electrolytic lesion of the solitary tract nucleus orcompensation. We also investigated the contribution of to sham lesion. In both cases, the meninges covering thespinal and cranial autonomic outflow to baroreflex cardiac ventricle were removed with the aid of a forceps. Thecompensation and the nature of the postganglionic trans- toads to be submitted to lesion were then fixed to a Kopfmitters. stereotaxic apparatus and each of two parallel tungsten
electrodes (A-M Systems, Everett, WA, USA) spaced 0.4mm apart, fixed to a metal piece attached to the electrode
2. Materials and methods holder was positioned 0.2 mm lateral to the obex to a depthof 0.5 mm. Each electrode was alternately connected to the
The experiments were conducted on B. paracnemis anode of a source of continuous current and the cathodetoads weighing 180–250 g collected in the vicinity of was connected with a toothed clamp to the animal’s paw,
˜ ˜Ribeirao Preto, Sao Paulo, Brazil, during the rainy summer previously wrapped in cotton moistened with physiologicalmonths and maintained in terraria at room temperature on saline. The lesions were made with 1 mA current applieda 12:12 h light–dark cycle. The animals had free access to for a period of 5 s. After lesion, the skin was closed withwater and were fed earthworms at 3-day intervals. Post- sutures. At the end of the experiments the animals wereoperative care, anaesthesia, surgery and euthanasia were deeply anaesthetized and their heads were removed and
´carried out in compliance with the lines set by Colegio stored in formaldehyde for 3 days. The brains were then˜Brasileiro de Experimentacao Animal (COBEA). removed, embedded in paraffin, cut into 10 mm sections
and stained with cresyl violet. The sections were examined2.1. Surgical procedure under the microscope to determine the extent of the
electrolytic lesions. An electrolytic rather than a neurotoxicThe toads were anaesthetized with ether vapour until lesion was chosen because the latter procedure requires a
cessation of locomotion and ventilation movements. A 6-day survival following injection of the drug in order tosmall incision was made on the skin of the dorsum dissipate the local infiltration of inflammatory cells that

L.M. Bianchi-da-Silva et al. / Journal of the Autonomic Nervous System 80 (2000) 31 –39 33
could act as an irritating stimulus. Due to impairment of was used to calibrate the pressure before and after eachautonomic functions, the physical state of the toad tends to experiment.become worse with time with the latter type of lesion. In one group of animals phenylephrine (30, 100 and 150
mg/kg, n56) was injected through the venous cannula inorder to determine possible reflex changes in heart rate
2.4. Isolation and section of the laryngeal and caused by the increase in arterial pressure induced by theglossopharyngeal nerves drug. In another group sodium nitroprusside (7.5, 15 and
60 mg/kg, n56; 30 mg/kg, n515) was injected in order toUnder anaesthesia, the nerves were identified and iso- induce a fall in arterial pressure and therefore provocative
lated through bilateral incisions in the axillary region. This baroreceptor unloading. The 30 mg/kg nitroprusside doseprocedure was carried out under visual control with the aid was selected as most appropriate for this study for itof a stereoscopic microscope. The laryngeal nerve crosses proved to induce consistently heart rate rises to repeatedthe pulmocutaneous artery obliquely and the glos- hypotensions induced by nitroprusside. The drug wassopharyngeal nerve crosses the external carotid immedi- injected into animals with an intact central nervous systemately above the carotid labyrinth. Depending on the (n56), into animals with high spinal section (n56) andexperiment planned, bilateral section of fibers of the with bilateral lesion of the solitary tract nucleus (n55), aslaryngeal nerve or of both the laryngeal and glos- well as into the respective sham-operated controls (n56sopharyngeal nerves, at their point of entry into the wall of and n55, respectively). In another group, nitroprussidethe pulmocutaneous or the carotid artery was performed. was injected 15 min after systemic injection of theAfter this procedure, the skin was closed with sutures. automatic blockers atropine sulphate (2 mg/kg, n57) or
propranolol hydrochloride (4 mg/kg, n58) or of the twodrugs combined (n510). Finally, in order to determine the
2.5. Aortic denervation participation of the different barosensitive areas in thegenesis of the reflex tachycardia, nitroprusside was in-
The axillary opening also permitted the visualization of jected into toads with bilateral section of the laryngealthe aortic arches to whose surface cotton wool soaked in nerves (n514), with combined bilateral section of the20% phenol solution was applied to destroy the remaining laryngeal and glossopharyngeal nerves (n510), with aorticnervous fibers after removal of the collagen fiber layer. arch deafferentation (n512), with bilateral section of theThis procedure was used because of the difficulty in laryngeal nerve plus aortic deafferentation (n510), andvisualising the baroreceptor aortic fibers. with bilateral section of the laryngeal nerve plus bilateral
section of the glossopharyngeal nerve and aortic archdeafferentation (n59), as well as into sham-denervated
2.6. Drugs control (n59).
All drugs were acquired from Sigma (St. Louis, MO, 2.8. Statistical analysisUSA) and diluted in physiological saline. The followingdoses were used: 30, 100 and 150 mg/kg of phenylephrine The cardiovascular parameters (blood pressure and heart(L-phenylephrine hydrochloride); 7.5, 15, 30 and 60 mg/kg rate) were analysed quantitatively on the basis of poly-of sodium nitroprusside (sodium nitroprusside dehydrate); graphic recordings of pulse arterial pressure. Mean arterial4 mg/kg of propranolol (propranolol hydrochloride), 2 pressure (MAP) was calculated as the sum of diastolicmg/kg of atropine (atropine sulphate). After the drug pressure plus 40% pulse pressure. Readings of bloodinjections, the venous cannula was flushed with 0.1 ml pressure and heart rate (HR) were taken over a period ofsaline. 15 s during the control period (before systemic injection of
nitroprusside) and during the phase of maximal hypoten-sion. HR is reported as beats /min. The dose–response
2.7. Experimental protocol and experimental groups curve was constructed on the basis of changes in MAP andHR induced by drugs. The results for each group are
The experiments were conducted in the summer at room reported as means of the differences between MAP andtemperature after a 24-h period of postoperative recovery. mean HR values observed during the phase of maximumThe animals were kept in boxes where they could move fall in arterial pressure induced by nitroprusside injectionfreely and the polygraphic recording was continued and those observed before the injection. The two-tailedthroughout the experiment. The arterial cannulae were t-test was used for comparisons between two groups. Whenconnected to a Nihon Kohden MPV-0.5A pressure trans- more than two groups were compared, we used one-wayducer placed at heart level outside the box and the Bonferroni analysis of variance (ANOVA). The limit oftransduced signal was amplified and recorded on a Nihon statistical significance was considered to be P,0.05. TheKohden model RM-6200 polygraph. A mercury column individual P values are reported for each test.
34 L.M. Bianchi-da-Silva et al. / Journal of the Autonomic Nervous System 80 (2000) 31 –39
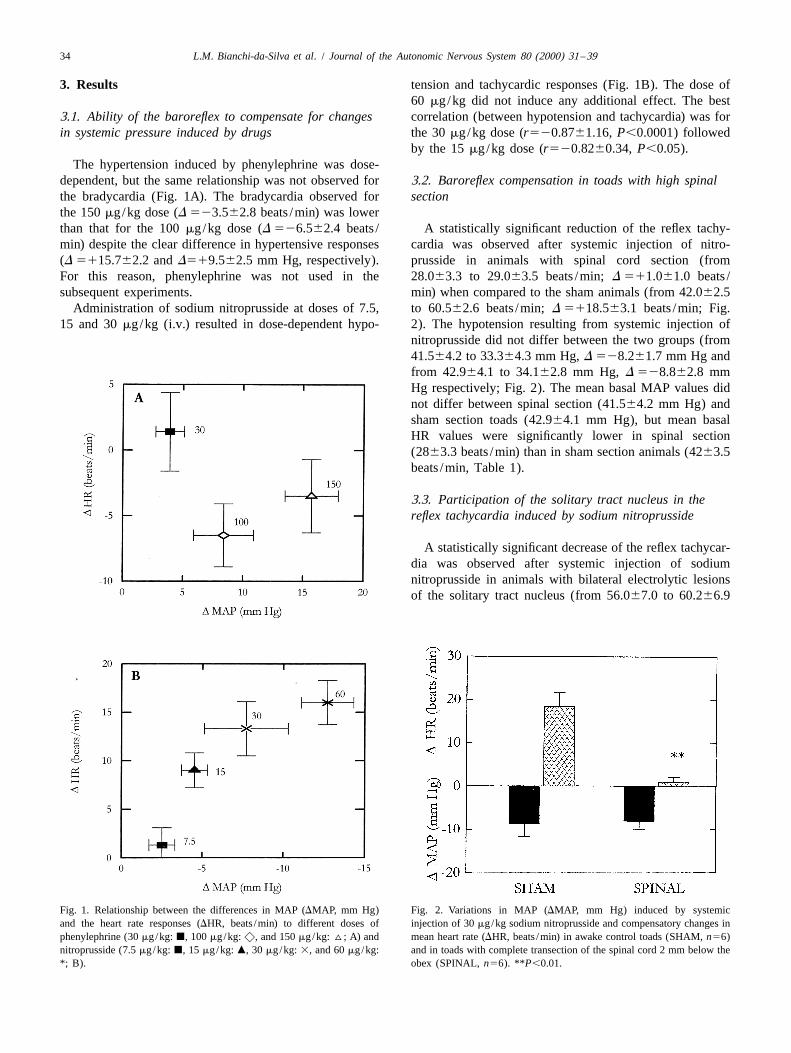
3. Results tension and tachycardic responses (Fig. 1B). The dose of60 mg/kg did not induce any additional effect. The best
3.1. Ability of the baroreflex to compensate for changes correlation (between hypotension and tachycardia) was forin systemic pressure induced by drugs the 30 mg/kg dose (r520.8761.16, P,0.0001) followed
by the 15 mg/kg dose (r520.8260.34, P,0.05).The hypertension induced by phenylephrine was dose-
dependent, but the same relationship was not observed for 3.2. Baroreflex compensation in toads with high spinalthe bradycardia (Fig. 1A). The bradycardia observed for sectionthe 150 mg/kg dose (D 523.562.8 beats /min) was lowerthan that for the 100 mg/kg dose (D 526.562.4 beats / A statistically significant reduction of the reflex tachy-min) despite the clear difference in hypertensive responses cardia was observed after systemic injection of nitro-(D 5115.762.2 and D519.562.5 mm Hg, respectively). prusside in animals with spinal cord section (fromFor this reason, phenylephrine was not used in the 28.063.3 to 29.063.5 beats /min; D 511.061.0 beats /subsequent experiments. min) when compared to the sham animals (from 42.062.5
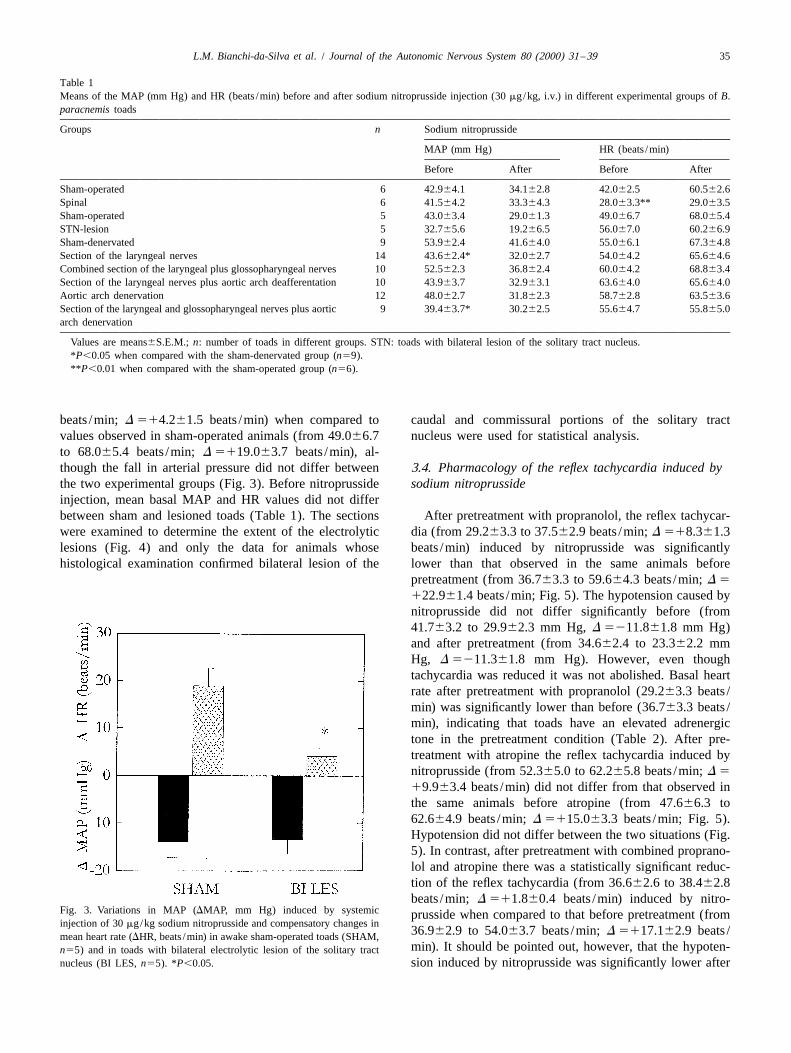
Administration of sodium nitroprusside at doses of 7.5, to 60.562.6 beats /min; D 5118.563.1 beats /min; Fig.15 and 30 mg/kg (i.v.) resulted in dose-dependent hypo- 2). The hypotension resulting from systemic injection of
nitroprusside did not differ between the two groups (from41.564.2 to 33.364.3 mm Hg, D 528.261.7 mm Hg andfrom 42.964.1 to 34.162.8 mm Hg, D 528.862.8 mmHg respectively; Fig. 2). The mean basal MAP values didnot differ between spinal section (41.564.2 mm Hg) andsham section toads (42.964.1 mm Hg), but mean basalHR values were significantly lower in spinal section(2863.3 beats /min) than in sham section animals (4263.5beats /min, Table 1).
3.3. Participation of the solitary tract nucleus in thereflex tachycardia induced by sodium nitroprusside
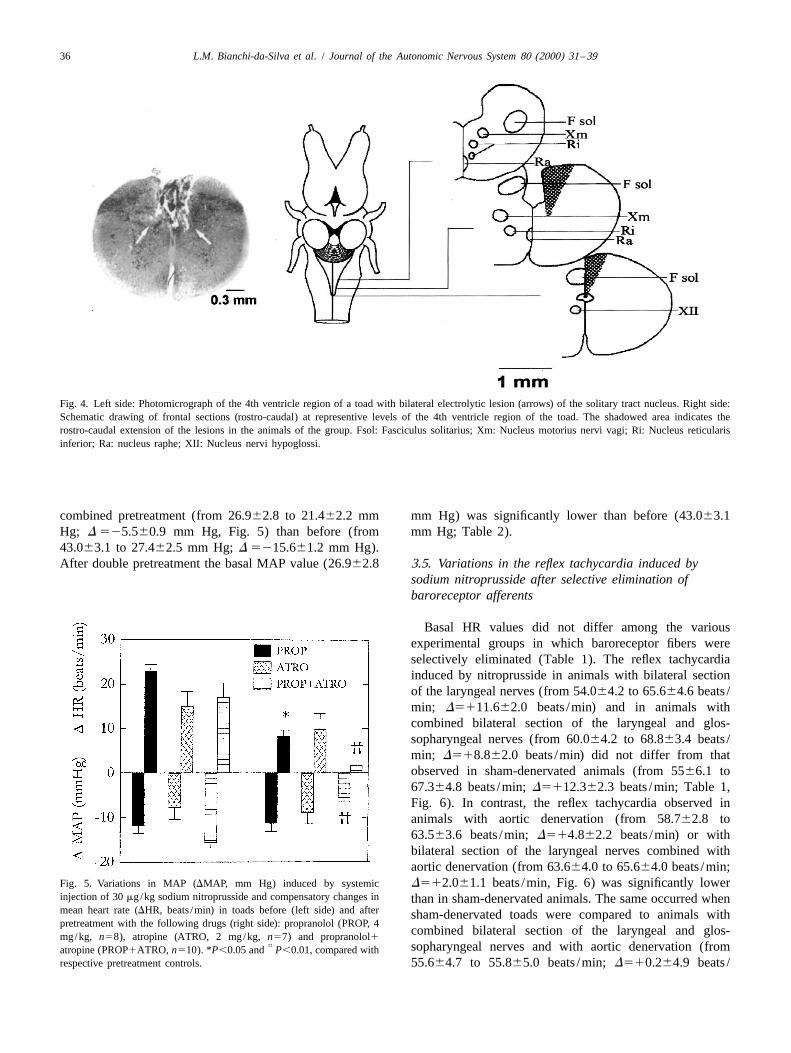
A statistically significant decrease of the reflex tachycar-dia was observed after systemic injection of sodiumnitroprusside in animals with bilateral electrolytic lesionsof the solitary tract nucleus (from 56.067.0 to 60.266.9
Fig. 1. Relationship between the differences in MAP (DMAP, mm Hg) Fig. 2. Variations in MAP (DMAP, mm Hg) induced by systemicand the heart rate responses (DHR, beats /min) to different doses of injection of 30 mg/kg sodium nitroprusside and compensatory changes inphenylephrine (30 mg/kg: j, 100 mg/kg: x, and 150 mg/kg: ^; A) and mean heart rate (DHR, beats /min) in awake control toads (SHAM, n56)nitroprusside (7.5 mg/kg: j, 15 mg/kg: m, 30 mg/kg: 3, and 60 mg/kg: and in toads with complete transection of the spinal cord 2 mm below the*; B). obex (SPINAL, n56). **P,0.01.
L.M. Bianchi-da-Silva et al. / Journal of the Autonomic Nervous System 80 (2000) 31 –39 35
Table 1Means of the MAP (mm Hg) and HR (beats /min) before and after sodium nitroprusside injection (30 mg/kg, i.v.) in different experimental groups of B.paracnemis toads
Groups n Sodium nitroprusside
MAP (mm Hg) HR (beats /min)
Before After Before After
Sham-operated 6 42.964.1 34.162.8 42.062.5 60.562.6Spinal 6 41.564.2 33.364.3 28.063.3** 29.063.5Sham-operated 5 43.063.4 29.061.3 49.066.7 68.065.4STN-lesion 5 32.765.6 19.266.5 56.067.0 60.266.9Sham-denervated 9 53.962.4 41.664.0 55.066.1 67.364.8Section of the laryngeal nerves 14 43.662.4* 32.062.7 54.064.2 65.664.6Combined section of the laryngeal plus glossopharyngeal nerves 10 52.562.3 36.862.4 60.064.2 68.863.4Section of the laryngeal nerves plus aortic arch deafferentation 10 43.963.7 32.963.1 63.664.0 65.664.0Aortic arch denervation 12 48.062.7 31.862.3 58.762.8 63.563.6Section of the laryngeal and glossopharyngeal nerves plus aortic 9 39.463.7* 30.262.5 55.664.7 55.865.0arch denervation
Values are means6S.E.M.; n: number of toads in different groups. STN: toads with bilateral lesion of the solitary tract nucleus.*P,0.05 when compared with the sham-denervated group (n59).**P,0.01 when compared with the sham-operated group (n56).
beats /min; D 514.261.5 beats /min) when compared to caudal and commissural portions of the solitary tractvalues observed in sham-operated animals (from 49.066.7 nucleus were used for statistical analysis.to 68.065.4 beats /min; D 5119.063.7 beats /min), al-though the fall in arterial pressure did not differ between 3.4. Pharmacology of the reflex tachycardia induced bythe two experimental groups (Fig. 3). Before nitroprusside sodium nitroprussideinjection, mean basal MAP and HR values did not differbetween sham and lesioned toads (Table 1). The sections After pretreatment with propranolol, the reflex tachycar-were examined to determine the extent of the electrolytic dia (from 29.263.3 to 37.562.9 beats /min; D 518.361.3lesions (Fig. 4) and only the data for animals whose beats /min) induced by nitroprusside was significantlyhistological examination confirmed bilateral lesion of the lower than that observed in the same animals before
pretreatment (from 36.763.3 to 59.664.3 beats /min; D 5
122.961.4 beats /min; Fig. 5). The hypotension caused bynitroprusside did not differ significantly before (from41.763.2 to 29.962.3 mm Hg, D 5211.861.8 mm Hg)and after pretreatment (from 34.662.4 to 23.362.2 mmHg, D 5211.361.8 mm Hg). However, even thoughtachycardia was reduced it was not abolished. Basal heartrate after pretreatment with propranolol (29.263.3 beats /min) was significantly lower than before (36.763.3 beats /min), indicating that toads have an elevated adrenergictone in the pretreatment condition (Table 2). After pre-treatment with atropine the reflex tachycardia induced bynitroprusside (from 52.365.0 to 62.265.8 beats /min; D 5
19.963.4 beats /min) did not differ from that observed inthe same animals before atropine (from 47.666.3 to62.664.9 beats /min; D 5115.063.3 beats /min; Fig. 5).Hypotension did not differ between the two situations (Fig.5). In contrast, after pretreatment with combined proprano-lol and atropine there was a statistically significant reduc-tion of the reflex tachycardia (from 36.662.6 to 38.462.8beats /min; D 511.860.4 beats /min) induced by nitro-
Fig. 3. Variations in MAP (DMAP, mm Hg) induced by systemic prusside when compared to that before pretreatment (frominjection of 30 mg/kg sodium nitroprusside and compensatory changes in
36.962.9 to 54.063.7 beats /min; D 5117.162.9 beats /mean heart rate (DHR, beats /min) in awake sham-operated toads (SHAM,min). It should be pointed out, however, that the hypoten-n55) and in toads with bilateral electrolytic lesion of the solitary tract
nucleus (BI LES, n55). *P,0.05. sion induced by nitroprusside was significantly lower after
36 L.M. Bianchi-da-Silva et al. / Journal of the Autonomic Nervous System 80 (2000) 31 –39
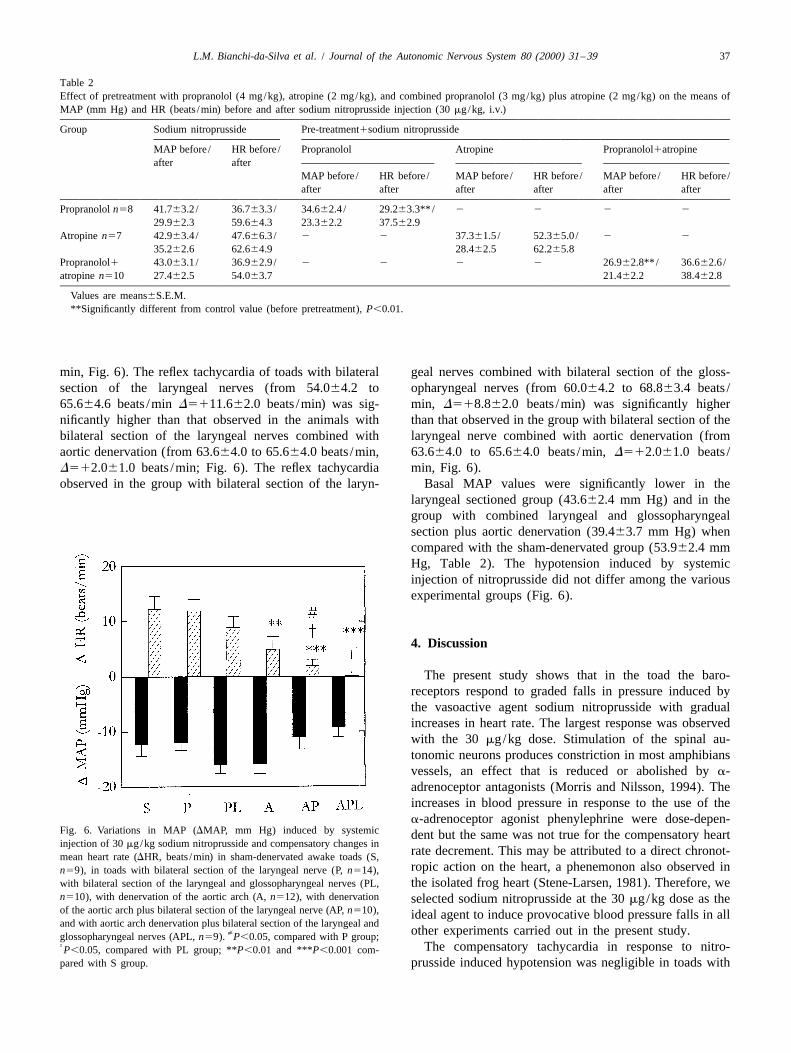
Fig. 4. Left side: Photomicrograph of the 4th ventricle region of a toad with bilateral electrolytic lesion (arrows) of the solitary tract nucleus. Right side:Schematic drawing of frontal sections (rostro-caudal) at representive levels of the 4th ventricle region of the toad. The shadowed area indicates therostro-caudal extension of the lesions in the animals of the group. Fsol: Fasciculus solitarius; Xm: Nucleus motorius nervi vagi; Ri: Nucleus reticularisinferior; Ra: nucleus raphe; XII: Nucleus nervi hypoglossi.
combined pretreatment (from 26.962.8 to 21.462.2 mm mm Hg) was significantly lower than before (43.063.1Hg; D 525.560.9 mm Hg, Fig. 5) than before (from mm Hg; Table 2).43.063.1 to 27.462.5 mm Hg; D 5215.661.2 mm Hg).After double pretreatment the basal MAP value (26.962.8 3.5. Variations in the reflex tachycardia induced by
sodium nitroprusside after selective elimination ofbaroreceptor afferents
Basal HR values did not differ among the variousexperimental groups in which baroreceptor fibers wereselectively eliminated (Table 1). The reflex tachycardiainduced by nitroprusside in animals with bilateral sectionof the laryngeal nerves (from 54.064.2 to 65.664.6 beats /min; D5111.662.0 beats /min) and in animals withcombined bilateral section of the laryngeal and glos-sopharyngeal nerves (from 60.064.2 to 68.863.4 beats /min; D518.862.0 beats /min) did not differ from thatobserved in sham-denervated animals (from 5566.1 to67.364.8 beats /min; D5112.362.3 beats /min; Table 1,Fig. 6). In contrast, the reflex tachycardia observed inanimals with aortic denervation (from 58.762.8 to63.563.6 beats /min; D514.862.2 beats /min) or withbilateral section of the laryngeal nerves combined withaortic denervation (from 63.664.0 to 65.664.0 beats /min;
Fig. 5. Variations in MAP (DMAP, mm Hg) induced by systemic D512.061.1 beats /min, Fig. 6) was significantly lowerinjection of 30 mg/kg sodium nitroprusside and compensatory changes in than in sham-denervated animals. The same occurred whenmean heart rate (DHR, beats /min) in toads before (left side) and after sham-denervated toads were compared to animals withpretreatment with the following drugs (right side): propranolol (PROP, 4
combined bilateral section of the laryngeal and glos-mg/kg, n58), atropine (ATRO, 2 mg/kg, n57) and propranolol1†† sopharyngeal nerves and with aortic denervation (fromatropine (PROP1ATRO, n510). *P,0.05 and P,0.01, compared with
respective pretreatment controls. 55.664.7 to 55.865.0 beats /min; D510.264.9 beats /
L.M. Bianchi-da-Silva et al. / Journal of the Autonomic Nervous System 80 (2000) 31 –39 37
Table 2Effect of pretreatment with propranolol (4 mg/kg), atropine (2 mg/kg), and combined propranolol (3 mg/kg) plus atropine (2 mg/kg) on the means ofMAP (mm Hg) and HR (beats /min) before and after sodium nitroprusside injection (30 mg/kg, i.v.)
Group Sodium nitroprusside Pre-treatment1sodium nitroprusside
MAP before / HR before / Propranolol Atropine Propranolol1atropineafter after
MAP before / HR before / MAP before / HR before / MAP before / HR before /after after after after after after
Propranolol n58 41.763.2 / 36.763.3 / 34.662.4 / 29.263.3**/ 2 2 2 2
29.962.3 59.664.3 23.362.2 37.562.9Atropine n57 42.963.4 / 47.666.3 / 2 2 37.361.5 / 52.365.0 / 2 2
35.262.6 62.664.9 28.462.5 62.265.8Propranolol1 43.063.1 / 36.962.9 / 2 2 2 2 26.962.8**/ 36.662.6 /atropine n510 27.462.5 54.063.7 21.462.2 38.462.8
Values are means6S.E.M.**Significantly different from control value (before pretreatment), P,0.01.
min, Fig. 6). The reflex tachycardia of toads with bilateral geal nerves combined with bilateral section of the gloss-section of the laryngeal nerves (from 54.064.2 to opharyngeal nerves (from 60.064.2 to 68.863.4 beats /65.664.6 beats /min D5111.662.0 beats /min) was sig- min, D518.862.0 beats /min) was significantly highernificantly higher than that observed in the animals with than that observed in the group with bilateral section of thebilateral section of the laryngeal nerves combined with laryngeal nerve combined with aortic denervation (fromaortic denervation (from 63.664.0 to 65.664.0 beats /min, 63.664.0 to 65.664.0 beats /min, D512.061.0 beats /D512.061.0 beats /min; Fig. 6). The reflex tachycardia min, Fig. 6).observed in the group with bilateral section of the laryn- Basal MAP values were significantly lower in the
laryngeal sectioned group (43.662.4 mm Hg) and in thegroup with combined laryngeal and glossopharyngealsection plus aortic denervation (39.463.7 mm Hg) whencompared with the sham-denervated group (53.962.4 mmHg, Table 2). The hypotension induced by systemicinjection of nitroprusside did not differ among the variousexperimental groups (Fig. 6).
4. Discussion
The present study shows that in the toad the baro-receptors respond to graded falls in pressure induced bythe vasoactive agent sodium nitroprusside with gradualincreases in heart rate. The largest response was observedwith the 30 mg/kg dose. Stimulation of the spinal au-tonomic neurons produces constriction in most amphibiansvessels, an effect that is reduced or abolished by a-adrenoceptor antagonists (Morris and Nilsson, 1994). Theincreases in blood pressure in response to the use of thea-adrenoceptor agonist phenylephrine were dose-depen-
Fig. 6. Variations in MAP (DMAP, mm Hg) induced by systemic dent but the same was not true for the compensatory heartinjection of 30 mg/kg sodium nitroprusside and compensatory changes in
rate decrement. This may be attributed to a direct chronot-mean heart rate (DHR, beats /min) in sham-denervated awake toads (S,ropic action on the heart, a phenemonon also observed inn59), in toads with bilateral section of the laryngeal nerve (P, n514),
with bilateral section of the laryngeal and glossopharyngeal nerves (PL, the isolated frog heart (Stene-Larsen, 1981). Therefore, wen510), with denervation of the aortic arch (A, n512), with denervation selected sodium nitroprusside at the 30 mg/kg dose as theof the aortic arch plus bilateral section of the laryngeal nerve (AP, n510), ideal agent to induce provocative blood pressure falls in alland with aortic arch denervation plus bilateral section of the laryngeal and
[ other experiments carried out in the present study.glossopharyngeal nerves (APL, n59). P,0.05, compared with P group;† The compensatory tachycardia in response to nitro-P,0.05, compared with PL group; **P,0.01 and ***P,0.001 com-pared with S group. prusside induced hypotension was negligible in toads with

38 L.M. Bianchi-da-Silva et al. / Journal of the Autonomic Nervous System 80 (2000) 31 –39
high spinal section although the falls in pressure continued showing that they are active within the physiological rangeto be of the same magnitude as observed in control of pressures (Hoffmann and Cordeiro de Sousa, 1982; Ishiianimals. This proves the reflex nature of the tachycardia and Ishii, 1971; Van Vliet and West, 1987b; West and vanand also shows that it may be attributed to descending Vliet, 1983). Because of their ability to limit the range ofsympathetic activation. Basal heart rate values were sig- systemic arterial pressure and to contribute to the mainte-nificantly lower in spinal toads when compared to sham- nance of vasoconstrictive tone in the pulmonary artery theyoperated toads but since basal MAP values were not are important for the protection of the delicate pulmonarydifferent, the HR drop cannot be attributed to spinal shock. microvasculature (Smith et al., 1981; Smits et al., 1986;Therefore, this finding suggests that the spinal cord may Van Vliet and West, 1987b; West and van Vliet, 1983). Ourprovide a sympathetic vasomotor tone and that the cardiac data show that, in the absence of pulmocutaneous baro-tone depends on supraspinal structures. receptors, other areas may reflexively compensate for
The present results also show that in toads the baro- systemic changes in arterial pressure. It is possible that inreceptor afferents preferentially converge towards the this situation the pulmonary vasculature bed is unprotectedcaudal portion of the solitary tract nucleus since the reflex since pulmonary lesions suggestive of pulmonary hyper-tachycardia caused by the fall in arterial pressure induced tension occurred in B. marinus with laryngeal nerveby nitroprusside was significantly reduced after bilateral section after systemic injection of phenylephrine (Van Vlietelectrolytic lesion of this region. Studies thus far published and West, 1989). The magnitude of the compensatoryhave been limited to the evaluation of the peripheral cardiac response is also not affected by bilateral section ofaspects of the baroreflex at the receptor or effector level, the laryngeal nerve associated with bilateral section of thewithout considering the central, integrative part. A previ- glossopharyngeal nerve. This indicates that the glos-ous study from our laboratory indicated that glutamate sopharyngeal nerves do not harbour baroreceptor fibers or,(Faccio and Hoffmann, 1981) and carbachol (Cordeiro de if they do, these fibers may not play an important roleSouza and Hoffmann, 1982) microinjections into the within the physiological range, as also proposed by othersolitary tract nucleus induce both hypertensive and hypo- investigators (Ishii et al., 1966; Segura, 1979). Our resultstensive responses and those promoted by carbachol are also suggest that the most important barosensitive area forblocked by pretreatment with atropine (Cordeiro de Souza the compensation of baroreceptor unloading is the aorticand Hoffmann, 1982). arch, since in the absence of information from this region
The tachycardic response to hypotension is mainly due the cardiac compensation for the hypotension induced byto b-adrenoceptors for in the presence of nitroprusside nitroprusside was small and significantly different fromhypotension of the same magnitude, it is significantly that observed in control animals or in animals with sectionlower after pretreatment with propranolol. Studies by other of the laryngeal nerve. The importance of the aorticinvestigators have shown that the main neurotransmitter in receptors in baroreflex compensation suggested by our datathe heart of amphibians is adrenaline (Stene-Larsen, 1981; is a novel observation. Based on recordings of afferentWoods, 1970) and its effect on heart rate in Rana is impulses of the aortic nerve in B. vulgaris and also onblocked by propranolol (Burggren and Doyle, 1986; Lillo, electron microscopy observations of the aortic arch wall,1979). However, Morris et al. (1981) have suggested that Ishii et al. (1985) attributed a predominantly chemorecep-in toads a receptor different from alpha and beta adreno- tor role to these afferents. According to the authors, someceptors can mediate positive chronotropic and inotropic fibers may fire in synchrony with the fluctuations in arterialeffects. Unfortunately there are no later studies confirming pressure, but electrical stimulation of the central stump ofthese findings. In our toads the tachycardia was still the nerve or distension of the vascularly isolated aortapresent after pretreatment with atropine. This fact, taken were unable to provoke a baroreflex response. The authorstogether with the observations that tachycardia is residual attributed this fact to the presence of few baroreceptorin animals with spinal section, leads us to conclude that if fibers in the nerve. It should be considered, however, thatthe inhibition of cholinergic vagal tone is part of the reflex these investigators worked with anaesthetized or pithedresponse, it is of minor importance in response to baro- animals. Further studies are necessary to determine ifreceptor unloading. After combined pre-treatment with quantitative or qualitative differences in compensatoryatropine and propranolol there was a significant reduction responses may be expected from individual distensionin reflex bradycardia. The interpretation of this result is (loading) of the distinct baroreceptor areas. The same ishampered by the fact that the nitroprusside-induced hypo- true for the role of the cardiac mechanoreceptors. In ourtension was lower after combined pretreatment, with a experiments, MAP basal was lower in the laryngealconsequent significant reduction in basal MAP. sectioned toads. We attribute it to the fact that the two
The compensatory cardiac response to the arterial experimental groups (sham and sectioned) were tested inpressure fall induced by nitroprusside after bilateral section different times of year. In a former work (Bianchi-da-of the laryngeal nerve was not different from that of Silva, 1994) laryngeal section was performed after asham-operated animals. Several investigators studied the control recording period in the same animals and the MAPbaroreceptors of the pulmocutaneous artery in anurans, and HR basal values were followed step by step for 48 h.

L.M. Bianchi-da-Silva et al. / Journal of the Autonomic Nervous System 80 (2000) 31 –39 39
Herrick, C.J., 1944. The fasciculus solitarius and its connections inA rise in MAP basal occurred acutely, but after 1 h theamphibians and fishes. J. Comp. Neurol. 81, 307–331.values were identical to the pre-denervation condition and
Hoffmann, A., Cordeiro de Sousa, M.B., 1982. Cardiovascular reflexes inremained so over the following 2 days. The same was conscious toads. J. Auton. Nerv. System. 5, 345–355.observed by Van Vliet and West for the toad B. marinus Ishii, K., Ishii, K., 1971. A reflexogenic area for controlling the blood(Van Vliet and West, 1989). pressure in the toad (Bufo vulgaris formosa). Jap. J. Physiol. 28,
423–431.Thus, our data permit us to suggest that the reflexIshii, K., Honda, K., Ishii, K., 1966. The function of the carotid labyrinthchanges in heart rate in response to vascular unloading in
in the toad. Tohoku J. Exp. Med. 88, 103–116.awake toads are of a spinal autonomic nature and are Ishii, K., Ishii, K., Kusakabe, T., 1985. Chemo- and baroreceptormediated by catecholamines acting on b receptors. The innervation of the aortic trunk of the toad Bufo vulgaris, Respir.exclusion of baroreceptor information from the pulmocuta- Physiol. 60, 365–375.
Kusakabe, T., 1990. Comparative studies on the vascular organization ofneous artery has no significant effect on reflex cardiaccarotid labyrinths of anurans and caudates. J. Morphol. 204, 47–55.adjustments to unloading. Thus, it is possible that these
Kusakabe, T., 1992. Intimate apposition of the glomus and smoothreceptors are more sensitive to loading than to unloading. muscle cells (g-s connection) in the carotid labyrinth of juvenileFurthermore, the possible baroreceptor information of the bullfrogs. Anat. Embryol. 185, 39–44.carotid labyrinth does not have sufficient gain to promote Lillo, R.S., 1979. Autonomic cardiovascular control during submergence
and emergence in bullfrogs. Am. J. Physiol. 273, R210–R216.additional changes in heart rate. In the labyrinth of anuransMorris, J.L., Nilsson, S., 1994. The circulatory system. In: Nilsson, S.,there is a mechanical system of pressure dampening
Holmgren, S. (Eds), Comparative Physiology and Evolution of thetowards the head region, and falls in pressure are not so Autonomic Nervous System. Harwood Academic Publishers, Switzer-critical in view of the tolerance of the central nervous land, pp. 193–246.system of anurans to hypoxia. The most important cardiac Morris, J.L., Gibbins, L.G., Clevers, J., 1981. Resistance of adrenergic
neurotransmission in the toad heart to adrenoceptor blockade.compensation occurring in order to buffer a fall in pressureNaunyn-Schmiedeberg’s Arch. Pharmacol. 317, 331–338.is due to receptors in the aortic arch region.
Nieuwenhuys, R., Opdam, P., 1976. Structure of the brain stem. In:´Llinas, R., Precht, W. (Eds.), Frog Neurobiology. Springer Verlag,
Berlin, pp. 811–847.Acknowledgements Poullet, M., 1977. A study of nerve endings in the carotid labyrinth of the
toad (Bufo bufo) after degeneration. Biol. Cell. 28, 75–80.Rogers, D.C., 1964. Possible baroreceptor nerve endings in the vicinity ofWe are deeply grateful to Aparecida Fim Pereira and
the amphibian carotid labyrinth. Acta Anat. 57, 253–266.Mariulza R. Brentegani for excellent technical assistance, Segura, E.T., 1979. Pressure and heart rate responses to raised carotidand to Rubens Mello for the histological preparations. This pressure in the toad. Am. J. Physiol. 237, H639–H643.
˜ `work was supported by grants from Fundacao de Amparo a Smith, D.G., Berger, P.J., Evans, B.K., 1981. Baroreceptor control ofheart rate in the conscious toad Bufo marinus. Am. J. Physiol. 242,˜Pesquisa do Estado de Sao Paulo (FAPESP), and Fun-H790–H796.˜ ˜dacao Coordenacao de Aperfeicoamento de Pessoal de
Smits, A.W., West, N.G., Burggren, W.W., 1986. Pulmonary fluid balance´Nıvel Superior (CAPES). following pulmocutaneous baroreceptor denervation in the toad. J.
Appl. Physiol. 61, 331–337.Stene-Larsen, G., 1981. Comparative aspects of cardiac adrenoceptors:
characterization of the b adrenoceptor as a common ‘‘adrenaline’’References 2
receptor in vertebrate hearts. Comp. Biochem. Physiol. 70C, 1–12.Toews, D., Shelton, G., Boutilier, R., 1982. The amphybian carotid
Adams, W.E., 1958. In: Thomas, C.C. (Ed.), The Comparative Morpholo- labyrinth: some anatomical and physiological relationships. Can. J.gy of the Carotid Body and Carotid Sinus. Springfield, IL, 202 pp. Zool. 60, 1153–1160.
Bagshaw, R.J., 1985. Evolution of cardiovascular control. Biol. Rev. 60, Van Vliet, B.N., West, N.H., 1987a. Responses to circulatory pressures,121–162. and conduction velocity, of pulmocutaneous baroreceptors in Bufo
Bianchi-da-Silva, L.M., 1994. Phasic and tonic function of the baro- marinus. J. Physiol. 388, 41–53.˜receptors in Bufo paracnemis. PhD thesis (in Portuguese), Sao Paulo Van Vliet, B.N., West, N.H., 1987b. Response characteristic of pulmocuta-
University. neous arterial baroreceptors in the toad, Bufo marinus. J. Physiol. 388,Burggren, W., Doyle, M., 1986. Ontogeny of heart rate regulation in the 55–70.
bullfrog, Rana catesbeiana. Am. J. Physiol. 251, R231–R239. Van Vliet, B.N., West, N.H., 1989. Cardiovascular responses to denerva-Carman, J.B., 1955. The carotid labyrinth in Hyla aurea, with a note on tion of pulmocutaneous baroreceptors in toads. Am. J. Physiol. 256,
that in Leiopelma Hochstetteri, J. Anat. 89, 503–525. R946–R954.Cordeiro de Souza, M.B., Hoffmann, A., 1982. Autonomic variations West, N.H., van Vliet, B.N., 1983. Open-loop analysis of the pulmocuta-
caused by stimulation of rhombencephalic visceral zone in anaesthet- neous baroreflex in the toad Bufo marinus. Am. J. Physiol. 245,ized toads. Comp. Biochem. Physiol. 70C, 123–127. R642–R650.
Faccio, A., Hoffmann, A., 1981. Regulation of cardiovascular function at Woods, R., 1970. The innervation of the frog heart: I. An examination ofthe bulbar level in anaesthetised toads: role of L-glutamic acid. the autonomic post-ganglionic nerve fibers and a comparison of the
´Proceedings of the 14th Congress of the Asociacion Latinoamericana autonomic and sensory ganglion cells. Proc. R. Soc. London, Ser. B´de Ciencias Fisiologicas. 176, 43–54.
Hanamori, T., Ishiko, N., 1987. Intraganglionic distribution of the primaryafferent neurons in the frog glossopharyngeal nerve studied by HRPmethod. Brain Res. 260, 191–196.