b lymphocyte development in rabbit: progenitor b cells ... anti-rabbit l chain ab (15); mouse...
TRANSCRIPT

of June 24, 2018.This information is current as
LymphopoiesisProgenitor B Cells and Waning of B B Lymphocyte Development in Rabbit:
Kingzette and Katherine L. KnightPaul J. Jasper, Shi-Kang Zhai, Susan L. Kalis, Mae
http://www.jimmunol.org/content/171/12/6372doi: 10.4049/jimmunol.171.12.6372
2003; 171:6372-6380; ;J Immunol
Referenceshttp://www.jimmunol.org/content/171/12/6372.full#ref-list-1
, 24 of which you can access for free at: cites 51 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2003 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 24, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

B Lymphocyte Development in Rabbit: Progenitor B Cells andWaning of B Lymphopoiesis1
Paul J. Jasper, Shi-Kang Zhai, Susan L. Kalis, Mae Kingzette, and Katherine L. Knight2
In mammals that use gut-associated lymphoid tissues for expansion and somatic diversification of the B cell repertoire, B lym-phopoiesis occurs early in ontogeny and does not appear to continue throughout life. In these species, including sheep, rabbit, andcattle, little is known about the pathway of B cell development and the time at which B lymphopoiesis wanes. We examined rabbitbone marrow by immunofluorescence with anti-CD79a and anti-� and identified both proB and preB cells. The proB cellsrepresent the vast majority of B-lineage cells in the bone marrow at birth and by incorporation of 5-bromo-2�-deoxyuridine, theyappear to be a dynamic population. PreB cells reach maximum levels in the bone marrow at 3 wk of age, and B cells begin toaccumulate at 7 wk of age. We cloned two VpreB and one �5 gene and demonstrated that they are expressed within B-lineage cellsin bone marrow. VpreB and �5 coimmunoprecipitated with the �-chain in lysates of 293T cells transfected with VpreB, �5, and�, indicating that VpreB, �5, and �-chains associate in a preB cell receptor-like complex. By 16 wk of age, essentially no proB orpreB cells are found in bone marrow and by PCR amplification, B cell recombination excision circles were reduced 200-fold. By18 mo of age, B cell recombination excision circles were reduced 500- to 1000-fold. We suggest that B cell development in the rabbitoccurs primarily through the classical, or ordered, pathway and show that B lymphopoiesis is reduced over 99% by 16 wk ofage. The Journal of Immunology, 2003, 171: 6372–6380.
D ifferent strategies have evolved to create and diversify aprimary Ab repertoire. In mice and humans, B lympho-poiesis occurs throughout life in bone marrow, and Ab
diversity is generated primarily through combinatorial joining ofVH, DH, and JH gene segments. In contrast, species such as chick-ens, sheep, and rabbits exhibit short-term B lymphopoiesis, whichoccurs early in development (1–6). In these species, the Ab rep-ertoire is expanded and diversified in gut-associated lymphoid tis-sue (GALT)3 thereby forming the primary or preimmune Ab rep-ertoire (7–9). If GALT is removed early in ontogeny, developmentof the B cell repertoire is severely impaired. In chickens, embry-onic bursectomy results in severe agammaglobulinemia (10–12),and in sheep and rabbits, neonatal removal of the ileal Peyer’spatch or GALT results in life-long B cell deficiency or a severelyreduced number of B cells (13–15).
The pathway of B cell development in mice and humans is well-described. In mice, B-lineage precursors progress through severalordered developmental stages which are defined by the status ofV(D)J gene rearrangement and expression of specific cell surfacemarkers. The ordered rearrangement of Ig genes begins in proBcells where D3J gene rearrangements occur. These cells progressto preB cells upon productive V3DJ gene rearrangement andmust express the preB cell receptor (preBCR) to continue devel-opment (16). The preBCR is a complex comprised of a�-chain in
association with a surrogate L chain, which is encoded byVpreBand �5. Signaling through the preBCR is required for the devel-opmental progression of early preB to late preB cells, which thenbecome immature B cells upon productive L chain gene rearrange-ment (16). In addition, a small percentage of B cells in mice de-velop through an alternative preBCR-independent pathway inwhich the L chain rearranges independently of the H chain (17–21). In rabbits, the only progenitor B cells described are preB cells,which have been identified in fetal liver, fetal spleen, and neonatalbone marrow (5, 22, 23). The presence of a�-chain and the ab-sence of the L chain in virtually all preB cells suggests that B celldevelopment in rabbit likely occurs through the classical, or or-dered, pathway, mediated by a preBCR. However, neither a sur-rogate L chain nor a preBCR has been identified.
In contrast to mice, in which preB cells are found in bone mar-row throughout life, in rabbits the number of preB cells is highestaround birth (22) or at 2–3 wk of age (5) and then steadily de-creases until they are undetectable in adults (5). To confirm thesedata, Crane et al. (6) examined levels of B cell recombinationexcision circles (BRECs) in bone marrow of young and adult rab-bits. The authors found high levels of BRECs in bone marrowDNA from young rabbits and almost none in adult rabbits, sug-gesting that by adulthood, lymphopoiesis had largely arrested. Al-though these data indicate that B lymphopoiesis largely arrestsearly in ontogeny, the timing of the decrease in B lymphopoiesisin rabbits remains unknown.
In the present study, we identified progenitor B cells, deter-mined the Ig gene rearrangements within these cells, and foundevidence of a preBCR. We also quantitated the decline in B lym-phopoiesis during the first several months of life by searching forB cell progenitors and BRECs.
Materials and MethodsImmunofluorescent staining of bone marrow cells
Antibodies. The Abs and indirect reagents used in this study were as fol-lows: mAb anti-rabbit�-chain (clone 367) (15); FITC-conjugated (24)goat anti-rabbit L chain Ab (15); mouse anti-human CD79a (Ig-�; mb-1),
Department of Microbiology and Immunology, Loyola University Chicago, May-wood, IL 60153
Received for publication July 25, 2003. Accepted for publication October 3, 2003.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by National Institutes of Health Grant AI50260.2 Address correspondence and reprint requests to Dr. Katherine L. Knight, Depart-ment of Microbiology and Immunology, Loyola University Chicago, Building 105,Room 3845, 2160 South 1st Avenue, Maywood, IL 60153. E-mail address:[email protected] Abbreviations used in this paper: GALT, gut-associated lymphoid tissue; preBCR,preB cell receptor; BREC, B cell recombination excision circle; RF, reading frame.
The Journal of Immunology
Copyright © 2003 by The American Association of Immunologists, Inc. 0022-1767/03/$02.00
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

which cross-reacts with rabbit CD79a (BD PharMingen clone HM47; SanDiego, CA); PE mouse anti-rabbit MHC class II (clone 2C4) (25); FITCgoat Fab anti-mouse IgG (Jackson ImmunoResearch Laboratories, WestGrove PA); streptavidin-PE (BD PharMingen); and streptavidin- allophy-cocyanin (BD PharMingen); FITC mouse anti-5-bromo-2�-deoxyuridine(BrdU) (eBioscience, San Diego CA).Immunofluorescence. To detect proB and preB cells, bone marrow cells(1 � 106) were fixed and permeabilized with Cytofix/Cytoperm (Promega,Madison, WI) and washed with Perm/Wash Buffer (Promega). Cells werestained with cross-reactive mouse anti-human CD79a/FITC goat Fab anti-mouse IgG and biotinylated mouse anti-rabbit �-chain/streptavidin-PE.Stained cells were analyzed at the FACS Core Facility (Loyola UniversityChicago Medical Center, Mayfield, IL) using a FACSCalibur flow cytom-eter (BD Biosciences, San Jose, CA). To distinguish surface �� and sur-face �� B-lineage cells, cells were first stained with mouse anti-rabbit�-chain/FITC goat Fab anti-mouse IgG. Subsequently, cells were fixed andpermeabilized and then stained with biotinylated mouse anti-rabbit�-chain/streptavidin-PE. Three-color immunofluorescent staining was per-formed first with mouse anti-rabbit �-chain/FITC goat Fab anti-mouseIgG. Cells were then fixed and permeabilized and stained with biotinylatedmouse anti-rabbit �-chain/streptavidin-allophycocyanin, and finally withPE mouse anti-human CD79a.
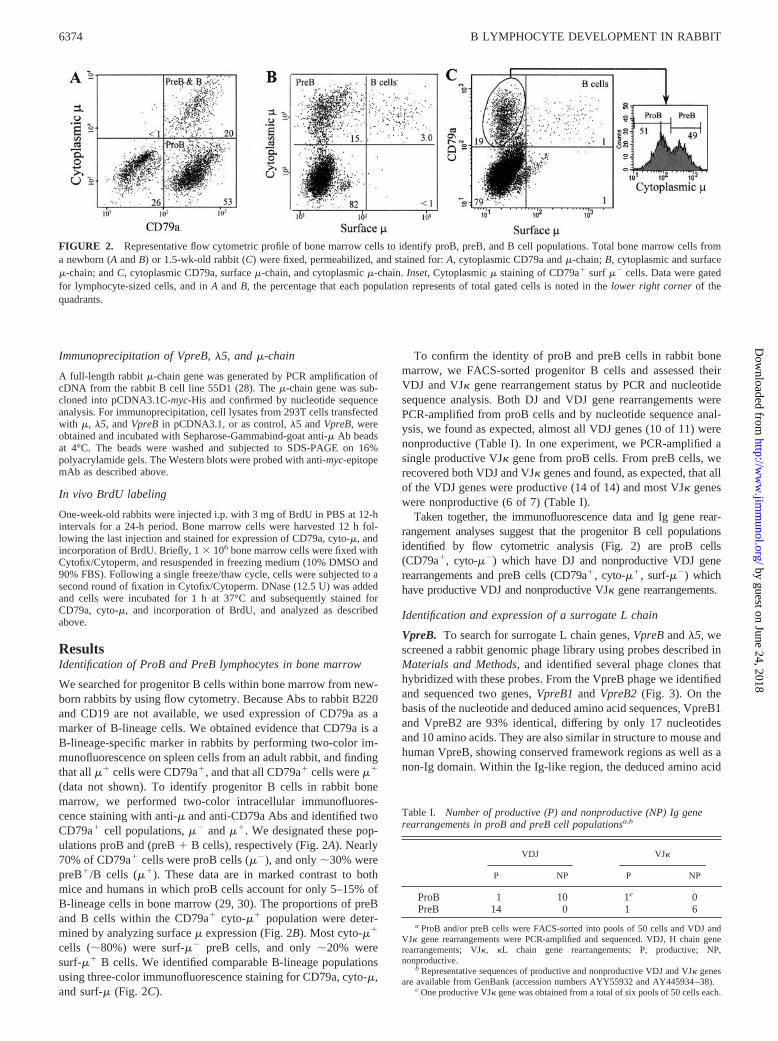
All flow cytometric data were gated for lymphocyte-sized cells on thebasis of forward and side scatter, and then analyzed using CellQuest soft-ware (BD Biosciences). ProB cells were defined as CD79a� cytoplasmic�� (cyto-��) surface �� (surf-��); preB cells were defined as CD79a�
cyto-�� surf-��; B cells were defined as CD79a� cyto-�� surf-��.
Detection of DJ, VDJ, and VJ� gene rearrangements by PCR
Pools of 50 cells were sorted into 96-well V-bottom plates containing 1�lysis buffer (100 �g/ml proteinase K in 1� PCR buffer (Promega)) usinga FACS-Star Plus (BD Biosciences) cell sorter. Cells were lysed at 37°Cfor 30 min and incubated at 97°C for 10 min to inactivate the proteinase K.The PCR mixture (200 pM dNTP; 100 pM each primer; 1� PCR buffer)was added to the samples, with 2.5 U Jumpstart Taq polymerase (Sigma-Aldrich, St. Louis, MO): the PCR was performed for 40 cycles of 1 min at95°C, 1 min at 60°C, and 1 min at 72°C. Aliquots (2 �l) of the first-roundPCR products were transferred to 48 �l of the second-round PCR mixture(containing nested primers), and 40 additional PCR cycles were performedunder the same conditions. PCR products were analyzed by PAGE. Primersused in nested PCR amplification of DJ, VDJ, and VJ� gene rearrange-ments are illustrated in Fig. 1).
Competitive PCR amplification for quantitation of BRECs
VD recombination signal sequence excision circles were quantified bycompetitive PCR using primers shown in Fig. 1. Briefly, BRECs from 2 �104 bone marrow cells (600 ng of total genomic DNA) were amplified with[32P]dCTP with varying numbers of copies of a competitor plasmid, which
results in a larger PCR product than the BREC target using the same prim-ers. To quantitate BRECs, the log of the number of copies of competitorplasmid in each reaction was plotted against the log of the ratio of cpmincorporated into target and competitor PCR products. To enhance thesensitivity of BREC detection in some experiments, we used circular DNAisolated from bone marrow (6) as template for PCR instead of totalgenomic DNA, and we added [32P]dCTP (0.5 �Ci) to the second round ofPCR amplification. To isolate circular DNA, bone marrow cells were lysedin 1% alkaline-SDS (pH 12.45) and the chromosomal DNA was sheared byvortexing, denatured in the alkaline-SDS solution, and removed from thecircular DNA by phenol extraction. The DNA circles were recovered byethanol precipitation and subjected to PCR amplification. PCR productswere separated by PAGE analysis and quantitated by phosphoimager anal-ysis on the Typhoon 8600 imager (Pharmacia, New York, NY). The iden-tity of BRECs was confirmed by nucleotide sequence analysis and is avail-able from GenBank (accession number AY455933).
Cloning, expression, and RT-PCR amplification of VpreB and �5
Rabbit VpreB and �5 probes were generated by PCR amplification of bonemarrow cDNA using primers to conserved regions of the mouse and humanVpreB and �5. The primers used for VpreB were 5�-CTACTGGTACCAGCAGAGGCC-3� (sense) and 5�-TCTTCCTCCCACTCCCTCTCC-3� (an-tisense); the primers used for �5 were 5�-CTGCTGCTGCTGGGTCT-3�(sense) and 5�-GTTGTTGCTCTGTTTGGAGGG-3� (antisense). PCRproducts were cloned into pGEMT-EZ (Promega), and their identities weredetermined by nucleotide sequence analysis using the BigDye terminationtechnique with an ABI Prism 310 Genetic Analyzer (Applied Biosystems,Foster City, CA). These clones were used to screen a genomic phage li-brary (�106 PFU) (26); VpreB� and �5� phage clones were purified, andthe nucleotide sequences of VpreB and �5 were determined. GenomicSouthern blot analysis was performed on HindIII- or XbaI-restrictedgenomic DNA according to standard protocols. The VpreB probe includedframework 2 through the non-Ig region and the �5 probe contained por-tions of exon I and exon II that were not expected to cross-hybridize withconventional C� L chain genes.
To identify �5 and VpreB genes expressed in bone marrow, cDNA wasPCR-amplified using two pairs of primers for each: for �5, A � (sense)5�-TTAAGCTTATGAGGCCCCGGAGTGGCCT-3�,(antisense)5�-AATCTAGAAGGACAGTGTGCAGGGGCCA-3� from the first and last 20 bpof the gene; and B � (sense) 5�-TATCTCAAGTCTGGAAGGA-3�, (an-tisense) 5�-GGGTGGGTGAGAGTTGTT-3� from the 5� and 3� untrans-lated regions. For VpreB, the primers were either pan-VpreB primers(sense) 5�-TTAAGCTTATGTCCTGGATTCCACTTT-3�, (antisense) 5�-GCACCGATCCTGGTGTCTCCA-3�; or the pan-VpreB sense primer andthe VpreB2-specific antisense primer, 5�-TCCTCCCCTCTCTTCCTCTCT-3�.
For in vitro expression of VpreB and �5, the genes were PCR-amplifiedusing primers 5� and 3� of the open reading frames: VpreB, (sense) 5�-TTAAGCTTATGTCCTGGATTCCACTTT-3�, (antisense) 5�-TTTCTAGAAGGCGCGCTGTCCTTGGGCA-3�; �5, primer pair A (above). The nu-cleotide sequences were confirmed and the genes were cloned into themammalian expression vector pCDNA3.1A-Myc-His (Invitrogen, Carls-bad, CA). Expression of each gene was confirmed by transfection into293T cells followed by Western blot analysis of lysates (1 � 106 cellequivalents) using 16% polyacrylamide gels. The blot was probed withanti-myc-epitope Ab (mAb 9E10) and HRP-conjugated goat anti-mouse Igand developed with the ECL chemiluminescent reagent (Amersham Phar-macia Biotech, Piscataway, NJ).
For RT-PCR amplification of VpreB and �5, cDNA was prepared asdescribed previously (27) from groups of FACS-sorted proB and preB cells(MHC class II�, surf-��). Two rounds of 40 cycles of PCR amplificationwere performed on the cDNA using the following sets of nested primers:VpreB primers were 5�-TTGTGCTGTGTGGCTAGGAA-3� (sense out-side primer), 5�-ATGTCCTGGATTCCACTTTG-3� (sense inside primer),5�-GCACCGATCCTGGTGTCTCCA-3� (antisense outside primer), and5�-CCTCAAGGCGCGCTGTCCTT-3� (antisense inside primer); �5 prim-ers were 5�-TATCTCAAGTCTGGAAGGA-3� (sense outside primer), 5�-ATGAGGCCCCGGAGTGGCCT-3� (sense inside primer), 5�-GGGTGGGTGAGAGTTGTT-3� (antisense outside primer), and 5�-GAGCTGGGTCCCACTGCCAA-3� (antisense inside primer), and VDJ primers were5�-CACTCACCATGGAGACT-3� (sense outside primer), 5�-GGACACGGCCACCTATTTCT-3� (sense inside primer), 5�-ACGGTCCGGCTGCTGATCTC-3� (antisense outside primer), and 5�-GTTGTTCTTGAAGCTCCAG-3� (antisense inside primer).
FIGURE 1. Illustration of PCR primers used to amplify DJ, VDJ, VJ�,and VD BREC gene rearrangements. The primer sequence used in eachPCR amplification is shown on the right. So, sense outside primer; Si, senseinside primer; Ao, antisense outside primer; Ai, antisense inside primer.The expected product size is: DJ � 100 bp; VDJ � 500 bp; VJ� � 400 bp;VD BREC � 150 bp. The percentage of rearrangements expected to bePCR-amplified with these primer sets is DJ � �50%; VDJ � �100%;VJ� � �90% (41); VD BREC � �50%.
6373The Journal of Immunology
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Immunoprecipitation of VpreB, �5, and �-chain
A full-length rabbit �-chain gene was generated by PCR amplification ofcDNA from the rabbit B cell line 55D1 (28). The �-chain gene was sub-cloned into pCDNA3.1C-myc-His and confirmed by nucleotide sequenceanalysis. For immunoprecipitation, cell lysates from 293T cells transfectedwith �, �5, and VpreB in pCDNA3.1, or as control, �5 and VpreB, wereobtained and incubated with Sepharose-Gammabind-goat anti-� Ab beadsat 4°C. The beads were washed and subjected to SDS-PAGE on 16%polyacrylamide gels. The Western blots were probed with anti-myc-epitopemAb as described above.
In vivo BrdU labeling
One-week-old rabbits were injected i.p. with 3 mg of BrdU in PBS at 12-hintervals for a 24-h period. Bone marrow cells were harvested 12 h fol-lowing the last injection and stained for expression of CD79a, cyto-�, andincorporation of BrdU. Briefly, 1 � 106 bone marrow cells were fixed withCytofix/Cytoperm, and resuspended in freezing medium (10% DMSO and90% FBS). Following a single freeze/thaw cycle, cells were subjected to asecond round of fixation in Cytofix/Cytoperm. DNase (12.5 U) was addedand cells were incubated for 1 h at 37°C and subsequently stained forCD79a, cyto-�, and incorporation of BrdU, and analyzed as describedabove.
ResultsIdentification of ProB and PreB lymphocytes in bone marrow
We searched for progenitor B cells within bone marrow from new-born rabbits by using flow cytometry. Because Abs to rabbit B220and CD19 are not available, we used expression of CD79a as amarker of B-lineage cells. We obtained evidence that CD79a is aB-lineage-specific marker in rabbits by performing two-color im-munofluorescence on spleen cells from an adult rabbit, and findingthat all �� cells were CD79a�, and that all CD79a� cells were ��
(data not shown). To identify progenitor B cells in rabbit bonemarrow, we performed two-color intracellular immunofluores-cence staining with anti-� and anti-CD79a Abs and identified twoCD79a� cell populations, �� and ��. We designated these pop-ulations proB and (preB � B cells), respectively (Fig. 2A). Nearly70% of CD79a� cells were proB cells (��), and only �30% werepreB�/B cells (��). These data are in marked contrast to bothmice and humans in which proB cells account for only 5–15% ofB-lineage cells in bone marrow (29, 30). The proportions of preBand B cells within the CD79a� cyto-�� population were deter-mined by analyzing surface � expression (Fig. 2B). Most cyto-��
cells (�80%) were surf-�� preB cells, and only �20% weresurf-�� B cells. We identified comparable B-lineage populationsusing three-color immunofluorescence staining for CD79a, cyto-�,and surf-� (Fig. 2C).
To confirm the identity of proB and preB cells in rabbit bonemarrow, we FACS-sorted progenitor B cells and assessed theirVDJ and VJ� gene rearrangement status by PCR and nucleotidesequence analysis. Both DJ and VDJ gene rearrangements werePCR-amplified from proB cells and by nucleotide sequence anal-ysis, we found as expected, almost all VDJ genes (10 of 11) werenonproductive (Table I). In one experiment, we PCR-amplified asingle productive VJ� gene from proB cells. From preB cells, werecovered both VDJ and VJ� genes and found, as expected, that allof the VDJ genes were productive (14 of 14) and most VJ� geneswere nonproductive (6 of 7) (Table I).
Taken together, the immunofluorescence data and Ig gene rear-rangement analyses suggest that the progenitor B cell populationsidentified by flow cytometric analysis (Fig. 2) are proB cells(CD79a�, cyto-��) which have DJ and nonproductive VDJ generearrangements and preB cells (CD79a�, cyto-��, surf-��) whichhave productive VDJ and nonproductive VJ� gene rearrangements.
Identification and expression of a surrogate L chain
VpreB. To search for surrogate L chain genes, VpreB and �5, wescreened a rabbit genomic phage library using probes described inMaterials and Methods, and identified several phage clones thathybridized with these probes. From the VpreB phage we identifiedand sequenced two genes, VpreB1 and VpreB2 (Fig. 3). On thebasis of the nucleotide and deduced amino acid sequences, VpreB1and VpreB2 are 93% identical, differing by only 17 nucleotidesand 10 amino acids. They are also similar in structure to mouse andhuman VpreB, showing conserved framework regions as well as anon-Ig domain. Within the Ig-like region, the deduced amino acid
Table I. Number of productive (P) and nonproductive (NP) Ig generearrangements in proB and preB cell populationsa,b
VDJ VJ�
P NP P NP
ProB 1 10 1c 0PreB 14 0 1 6
a ProB and/or preB cells were FACS-sorted into pools of 50 cells and VDJ andVJ� gene rearrangements were PCR-amplified and sequenced. VDJ, H chain generearrangements; VJ�, �L chain gene rearrangements; P, productive; NP,nonproductive.
b Representative sequences of productive and nonproductive VDJ and VJ� genesare available from GenBank (accession numbers AYY55932 and AY445934–38).
c One productive VJ� gene was obtained from a total of six pools of 50 cells each.
FIGURE 2. Representative flow cytometric profile of bone marrow cells to identify proB, preB, and B cell populations. Total bone marrow cells froma newborn (A and B) or 1.5-wk-old rabbit (C) were fixed, permeabilized, and stained for: A, cytoplasmic CD79a and �-chain; B, cytoplasmic and surface�-chain; and C, cytoplasmic CD79a, surface �-chain, and cytoplasmic �-chain. Inset, Cytoplasmic � staining of CD79a� surf �� cells. Data were gatedfor lymphocyte-sized cells, and in A and B, the percentage that each population represents of total gated cells is noted in the lower right corner of thequadrants.
6374 B LYMPHOCYTE DEVELOPMENT IN RABBIT
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

sequence of rabbit VpreB1 is 70% identical (82% similar) to bothmouse and human VpreB1 (Fig. 4). The non-Ig region is much lessconserved, having only 29 and 43% amino acid identity to that ofmice and humans, respectively.
Southern blot analysis of genomic DNA using the VpreB proberesulted in three hybridizing bands (Fig. 5A) suggesting that therabbit genome contains at least three VpreB genes. To determinewhether one or more of these genes is expressed during B celldevelopment, we RT-PCR-amplified VpreB from newborn bonemarrow cDNA using pan-VpreB primers and determined their nu-cleotide sequences. We found that each of 10 PCR products en-coded VpreB1 (data not shown). To determine whether VpreB2 isexpressed, we PCR-amplified and cloned bone marrow cDNA us-ing VpreB2-specific primers and found that each of seven clonesencoded VpreB2 (data not shown). Although these data demon-
strate that both rabbit VpreB1 and VpreB2 are expressed in new-born bone marrow, VpreB1 is the predominantly expressed VpreBgene during B cell development. We found no evidence for a thirdVpreB molecule expressed in bone marrow even though we ob-served three VpreB-hybridizing bands by Southern blot analysis.
�5. The �5-hybridizing phage clones were restriction-mapped,and by Southern blot analysis, we found a phage (�2-1) that con-tained all three exons of a �5 gene (Fig. 6). From the deducedamino acid sequence, this gene appears functional and shares 57and 66% amino acid identity with mouse and human �5, respec-tively, within the Ig-like region (Fig. 6). Similar to the resultsfound with VpreB, the non-Ig region of �5 is significantly less-conserved across species, demonstrating 44 and 58% amino acididentity with mice and humans, respectively.
FIGURE 3. Restriction maps of VpreB1 and VpreB2 phage and comparison of deduced amino acid sequences. A, Restriction maps of phage 9 and phage 19;H, HindIII; S, SacI; B, BamHI; R, EcoRI; K, KpnI; X, XbaI; Z, XhoI. HindIII is not mapped in phage 19; XbaI is partially mapped in phage 19 and not mappedin phage 9 (B). The nucleotide sequence and amino acid translation of VpreB1 are compared with the deduced amino acid sequence of VpreB2. Positions of aminoacid identity are denoted by a dot (● ). To facilitate amino acid comparison, the leader was spliced to framework 1 on the basis of the splice site found in VpreBcDNA clones. The sequences of VpreB1 and VpreB2 are accessible at GenBank under accession numbers AY351269 and AY351268.
FIGURE 4. Comparison of deduced amino acid sequences of rabbit, human, and mouse VpreB1. The nucleotide sequence and amino acid translationof rabbit VpreB1 are shown on top. Positions of amino acid identity are denoted by a dot (●). Dashes (-) were introduced to maximize homology.
6375The Journal of Immunology
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

By Southern blot analysis, we found two hybridizing bands ineach of five different rabbits, indicating that the genome containstwo �5 genes (Fig. 5B). We PCR-amplified, cloned, and sequenced�5 genes from bone marrow cDNA of newborn rabbits using twodifferent pairs of �5-specific primers (see Material and Methods)to determine whether two different �5 molecules were expressed.We found that the nucleotide sequences of four clones obtainedfrom each set of primers were identical to that of the �5 genecontained on phage 2 (data not shown). Although these data sug-gest that only a single �5 gene is expressed in bone marrow ofnewborn rabbits, we cannot rule out the possibility that another �5gene is expressed but did not PCR amplify with either primer set.Alternatively, perhaps there is only a single �5 gene in rabbits, andthe second hybridizing band found by genomic Southern blot re-
sults from an exon I homologue upstream of the C�1 constantregion gene, as is found in humans (31).
PreBCR
To determine whether progenitor B cells express a preBCR, weperformed RT-PCR for VpreB and �5 on groups of FACS-sortedproB and preB cells. Because of the difficulty performing RT-PCRon fixed cells, we did not use the intracellular marker CD79a toisolate proB and preB cells, and instead searched for alternativesurface molecules that would allow us to obtain progenitor B cells.We found that all CD79a� bone marrow cells were also MHCclass II� and vice versa (Fig. 7A), suggesting that we could useMHC class II instead of CD79a to isolate B lineage cells. Westained total bone marrow cells with anti-MHC class II-PE andanti-�-FITC Abs, and FACS-sorted progenitor B cells (MHC classII�, surf-��). By RT-PCR amplification, we found that VpreB and�5 were expressed in these cells (25 cells/pool), but not in MHCclass II� cells (100 cells/pool) (Fig. 7B). We conclude that VpreBand �5 are expressed in proB and/or preB cells but not in other celllineages.
To determine whether VpreB and �5 can associate with the�-chain, as would be expected if they are part of a preBCR com-plex, VpreB and �5 proteins were myc-tagged and assayed for theirability to pair with the �-chain in 293T cells transfected withVpreB, �5, and �-chain genes. Immunoprecipitation of the�-chain from 293T cell lysates followed by Western blot showedthat two proteins of �18 and 22 kDa, as expected for VpreB and�5, respectively, were coimmunoprecipitated with the �-chain(Fig. 8). These two proteins were not observed in anti-� immu-noprecipitates of 293T cells transfected with VpreB and �5 in theabsence of �. These results demonstrate that the �-chain associateswith VpreB and �5 to form a preBCR-like protein complex.
FIGURE 6. Restriction map of �5 phage and comparison of deduced amino acid sequence encoded by rabbit, human, and mouse �5 genes. A, Therestriction map of phage 2-1 containing a �5 gene; H, HindIII; S, SacI; B, BamHI; R, EcoRI; K, KpnI; X, XbaI; Z � XhoI. Exons of �5 are designatedas f, I, II and III; B, the nucleotide sequence and amino acid translation of rabbit, human, and mouse �5. Positions of amino acid identity are denoted bya dot (●). Dashes (-) were introduced to maximize homology. The sequence of rabbit �5 is accessible at GenBank under accession number AY351267.
FIGURE 5. Genomic Southern blot analysis of rabbit VpreB (A) and �5(B). Genomic DNA was digested with HindIII or XbaI, and the blots wereprobed with (A) a fragment of VpreB including framework region 2through the non-Ig region, and (B) exon 1 of �5. No HindIII or XbaI sitesare present in VpreB or �5 genes. Identical results were obtained from eachof five rabbits. Sizes of �HindIII markers are shown at the left.
6376 B LYMPHOCYTE DEVELOPMENT IN RABBIT
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

One function of the preBCR is to select for preB cells using Din the preferred reading frame (RF), RF1 (32, 33). In rabbits, VDJgenes in proB cells show no bias for D RF1 (Table II), whereasVDJ genes in peripheral B cells are heavily biased toward RF1 (34,35). To determine whether D RF selection occurs at the preB cellstage, we PCR-amplified VDJ genes from FACS-sorted preB cellsand found that each of 14 productive VDJ genes used D in thepreferred RF (Table II). These data demonstrate that D RF selec-tion occurs between the proB and preB cell stages, and we suggestthat this selection is mediated by the preBCR.
Decline in B lymphopoiesis after birth
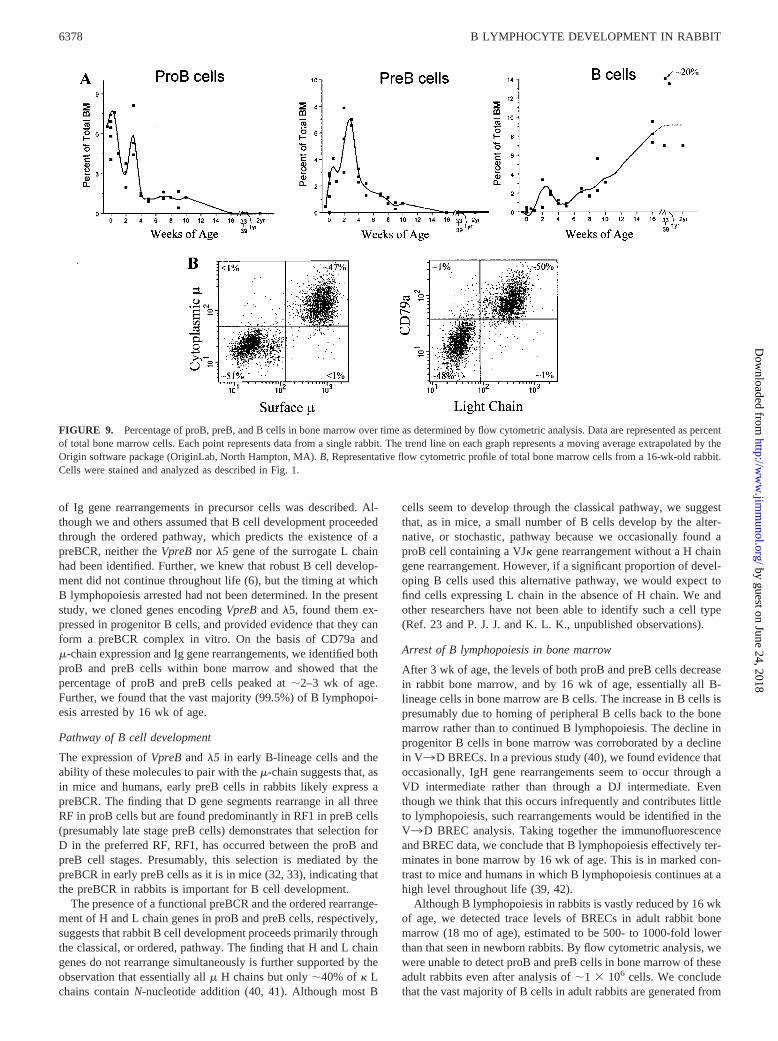
Previous studies suggested that B lymphopoiesis in rabbits is ro-bust early in ontogeny but is almost undetectable in adults (6).Using flow cytometric analysis and competitive PCR to quantify Bcell precursors and BRECs, we sought to define the time at whichB lymphopoiesis arrests. Bone marrow samples were taken fromrabbits at different ages, and the percentages of proB, preB, and Bcells were determined by flow cytometry analysis. We found highlevels of proB cells at birth (�7–8% of total bone marrow cells)and again at 3 wk of age (Fig. 9A). ProB cells rapidly declined by4 wk and continued to decline slowly until, by 16 wk of age, theywere undetectable. The percentage of preB cells was low at birth(�1.5%) and increased until �2–3 wk of age, at which time theycomprised �7% of total bone marrow cells. Similar to proB cells,the preB cells then rapidly declined during the next 2 wk andcontinued to decline until they were undetectable at 16 wk of age.
In contrast to proB and preB cells, essentially no B cells werefound in bone marrow at birth. Their numbers increased slowlyover time, until B cells constituted all B-lineage cells in the bonemarrow at 16 wk of age (Fig. 9B).
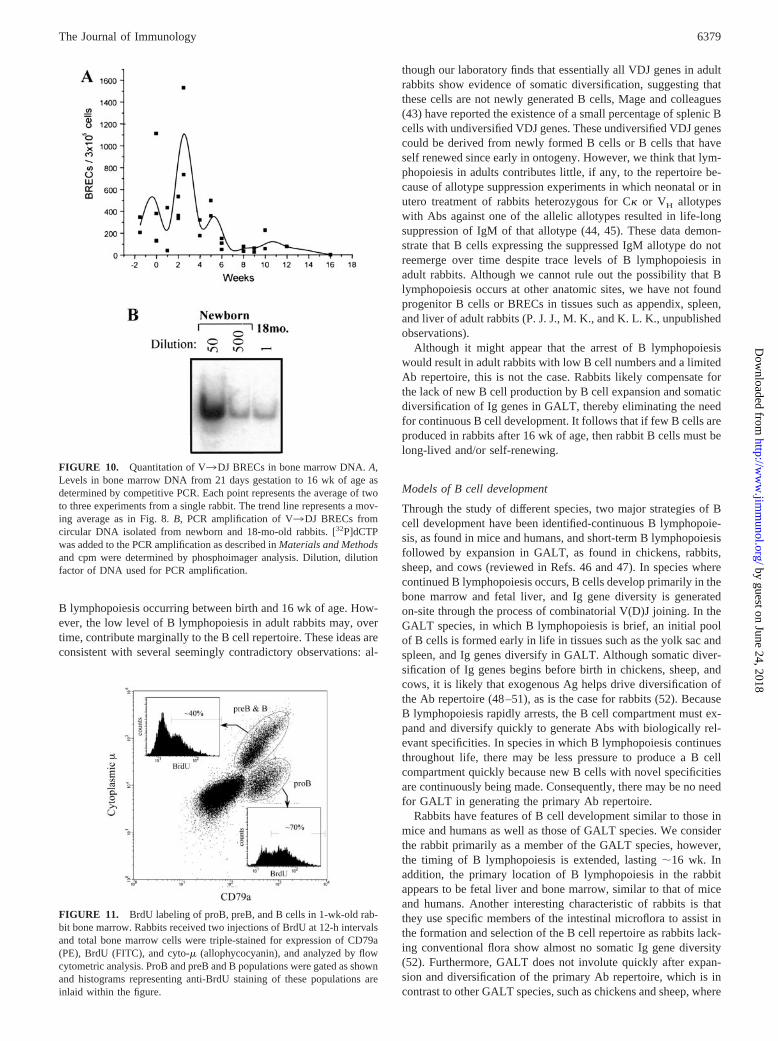
To confirm the decline in B lymphopoiesis observed by flowcytometric analyses, we quantitated the levels of V3D BRECs asa measure of V3DJ gene rearrangement leading to the formationof preB cells (Fig. 10A). We found that the number of BRECs atbirth was �300 per 3 � 105 cells and reached maximum levels(�1100 per 3 � 105 cells) at �3 wk of age, when we see thehighest percentage of preB cells in bone marrow (Fig. 9A). After3 wk, BRECs declined steadily and were undetectable by 16 wk ofage. Because in previous studies (6), it appeared that low levels ofBRECs were detectable in adult rabbits, we increased the sensi-tivity of the analysis by using circular DNA (enriched for BRECs)and by adding [32P]dCTP to the PCR amplification. In two adultrabbits (4 mo and 18 mo of age), we detected BRECs by phos-phoimager analysis but not by ethidium bromide staining. Wecompared the levels of BRECs in these adult rabbits with the levelsfound in bone marrow of newborn rabbits and found that BRECswere decreased �200-fold (99.5%) at 4 mo (data not shown) and500-1000 fold by 18 mo of age (Fig. 10B). Consistent with thegreatly decreased number of BRECs in bone marrow of adult rab-bits, we did not detect proB or preB cells by flow cytometric anal-ysis, even though we analyzed 10 times more cells than in previousexperiments (data not shown). Taken together, these data suggestthat by 16 wk of age, B lymphopoiesis in rabbit is decreased over99%, and by adulthood, is at a level ��0.2% of that in newbornrabbits.
BrdU labeling of ProB cells
The above experiments demonstrate that at birth, most (�70%)B-lineage cells (CD79a�) are proB cells. PreB cell levels increaseslowly and reach their highest levels only after 3 wk of age. Thisresult is surprising because in mice, proB cells differentiate intopreB cells within a 24-h period (36–39). Two possible explana-tions for the apparent delay in preB cell accumulation are that: 1)the majority of proB cells are quiescent and require a few weeks todifferentiate into preB cells; or 2) ProB cells differentiate withkinetics similar to that of mice; however, the vast majority of proBcells die, thereby requiring a 3-wk time period for preB cells toaccumulate and/or proliferate. To distinguish between these pos-sibilities, we performed BrdU-labeling experiments. If the proBcell population is quiescent, we expected the majority of proB cellswould not be labeled within 24 h following BrdU injection. Wefound that following two injections of BrdU over a 24-h period, themajority of proB cells (�70%) labeled with BrdU (Fig. 11), im-plying that proB cells in rabbits are not quiescent. We concludethat the slow accumulation of preB cells is likely due to massivecell death within the proB and/or preB cell populations duringdifferentiation.
DiscussionBefore this study, rabbit preB cells had been identified by severalinvestigators (5, 22, 23); however, neither proB cells nor the status
Table II. D RF of VDJ genes cloned from proB and preB cellsa
RF1 RF2 RF3
ProB 2 6 3PreB 14 0 0
a D gene RF usage was determined from the deduced amino acid sequence; RF1encodes primarily G and Y; RF2 encodes primarily M, V, and L; RF3 encodes pri-marily W, L, and C, and/or a stop codon.
FIGURE 8. Western blot of anti-� immunoprecipitates of lysates of293T cells transfected with myc-tagged rabbit �5 and VpreB, with(�) orwithout(�) �. The blot was probed with anti-myc mAb.
FIGURE 7. Expression of surrogate L chain in proB and preB cells. A,Flow cytometric analysis of newborn bone marrow cells stained for CD79aand MHC-class II. B, RT-PCR amplification of VpreB and �5, and inFACS-sorted MHC class II�, surf-� � cells (proB and preB cells; pools of25 cells) or non B-lineage cells (MHC class II�; pools of 100 cells).
6377The Journal of Immunology
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
of Ig gene rearrangements in precursor cells was described. Al-though we and others assumed that B cell development proceededthrough the ordered pathway, which predicts the existence of apreBCR, neither the VpreB nor �5 gene of the surrogate L chainhad been identified. Further, we knew that robust B cell develop-ment did not continue throughout life (6), but the timing at whichB lymphopoiesis arrested had not been determined. In the presentstudy, we cloned genes encoding VpreB and �5, found them ex-pressed in progenitor B cells, and provided evidence that they canform a preBCR complex in vitro. On the basis of CD79a and�-chain expression and Ig gene rearrangements, we identified bothproB and preB cells within bone marrow and showed that thepercentage of proB and preB cells peaked at �2–3 wk of age.Further, we found that the vast majority (99.5%) of B lymphopoi-esis arrested by 16 wk of age.
Pathway of B cell development
The expression of VpreB and �5 in early B-lineage cells and theability of these molecules to pair with the �-chain suggests that, asin mice and humans, early preB cells in rabbits likely express apreBCR. The finding that D gene segments rearrange in all threeRF in proB cells but are found predominantly in RF1 in preB cells(presumably late stage preB cells) demonstrates that selection forD in the preferred RF, RF1, has occurred between the proB andpreB cell stages. Presumably, this selection is mediated by thepreBCR in early preB cells as it is in mice (32, 33), indicating thatthe preBCR in rabbits is important for B cell development.
The presence of a functional preBCR and the ordered rearrange-ment of H and L chain genes in proB and preB cells, respectively,suggests that rabbit B cell development proceeds primarily throughthe classical, or ordered, pathway. The finding that H and L chaingenes do not rearrange simultaneously is further supported by theobservation that essentially all � H chains but only �40% of � Lchains contain N-nucleotide addition (40, 41). Although most B
cells seem to develop through the classical pathway, we suggestthat, as in mice, a small number of B cells develop by the alter-native, or stochastic, pathway because we occasionally found aproB cell containing a VJ� gene rearrangement without a H chaingene rearrangement. However, if a significant proportion of devel-oping B cells used this alternative pathway, we would expect tofind cells expressing L chain in the absence of H chain. We andother researchers have not been able to identify such a cell type(Ref. 23 and P. J. J. and K. L. K., unpublished observations).
Arrest of B lymphopoiesis in bone marrow
After 3 wk of age, the levels of both proB and preB cells decreasein rabbit bone marrow, and by 16 wk of age, essentially all B-lineage cells in bone marrow are B cells. The increase in B cells ispresumably due to homing of peripheral B cells back to the bonemarrow rather than to continued B lymphopoiesis. The decline inprogenitor B cells in bone marrow was corroborated by a declinein V3D BRECs. In a previous study (40), we found evidence thatoccasionally, IgH gene rearrangements seem to occur through aVD intermediate rather than through a DJ intermediate. Eventhough we think that this occurs infrequently and contributes littleto lymphopoiesis, such rearrangements would be identified in theV3D BREC analysis. Taking together the immunofluorescenceand BREC data, we conclude that B lymphopoiesis effectively ter-minates in bone marrow by 16 wk of age. This is in marked con-trast to mice and humans in which B lymphopoiesis continues at ahigh level throughout life (39, 42).
Although B lymphopoiesis in rabbits is vastly reduced by 16 wkof age, we detected trace levels of BRECs in adult rabbit bonemarrow (18 mo of age), estimated to be 500- to 1000-fold lowerthan that seen in newborn rabbits. By flow cytometric analysis, wewere unable to detect proB and preB cells in bone marrow of theseadult rabbits even after analysis of �1 � 106 cells. We concludethat the vast majority of B cells in adult rabbits are generated from
FIGURE 9. Percentage of proB, preB, and B cells in bone marrow over time as determined by flow cytometric analysis. Data are represented as percentof total bone marrow cells. Each point represents data from a single rabbit. The trend line on each graph represents a moving average extrapolated by theOrigin software package (OriginLab, North Hampton, MA). B, Representative flow cytometric profile of total bone marrow cells from a 16-wk-old rabbit.Cells were stained and analyzed as described in Fig. 1.
6378 B LYMPHOCYTE DEVELOPMENT IN RABBIT
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
B lymphopoiesis occurring between birth and 16 wk of age. How-ever, the low level of B lymphopoiesis in adult rabbits may, overtime, contribute marginally to the B cell repertoire. These ideas areconsistent with several seemingly contradictory observations: al-
though our laboratory finds that essentially all VDJ genes in adultrabbits show evidence of somatic diversification, suggesting thatthese cells are not newly generated B cells, Mage and colleagues(43) have reported the existence of a small percentage of splenic Bcells with undiversified VDJ genes. These undiversified VDJ genescould be derived from newly formed B cells or B cells that haveself renewed since early in ontogeny. However, we think that lym-phopoiesis in adults contributes little, if any, to the repertoire be-cause of allotype suppression experiments in which neonatal or inutero treatment of rabbits heterozygous for C� or VH allotypeswith Abs against one of the allelic allotypes resulted in life-longsuppression of IgM of that allotype (44, 45). These data demon-strate that B cells expressing the suppressed IgM allotype do notreemerge over time despite trace levels of B lymphopoiesis inadult rabbits. Although we cannot rule out the possibility that Blymphopoiesis occurs at other anatomic sites, we have not foundprogenitor B cells or BRECs in tissues such as appendix, spleen,and liver of adult rabbits (P. J. J., M. K., and K. L. K., unpublishedobservations).
Although it might appear that the arrest of B lymphopoiesiswould result in adult rabbits with low B cell numbers and a limitedAb repertoire, this is not the case. Rabbits likely compensate forthe lack of new B cell production by B cell expansion and somaticdiversification of Ig genes in GALT, thereby eliminating the needfor continuous B cell development. It follows that if few B cells areproduced in rabbits after 16 wk of age, then rabbit B cells must belong-lived and/or self-renewing.
Models of B cell development
Through the study of different species, two major strategies of Bcell development have been identified-continuous B lymphopoie-sis, as found in mice and humans, and short-term B lymphopoiesisfollowed by expansion in GALT, as found in chickens, rabbits,sheep, and cows (reviewed in Refs. 46 and 47). In species wherecontinued B lymphopoiesis occurs, B cells develop primarily in thebone marrow and fetal liver, and Ig gene diversity is generatedon-site through the process of combinatorial V(D)J joining. In theGALT species, in which B lymphopoiesis is brief, an initial poolof B cells is formed early in life in tissues such as the yolk sac andspleen, and Ig genes diversify in GALT. Although somatic diver-sification of Ig genes begins before birth in chickens, sheep, andcows, it is likely that exogenous Ag helps drive diversification ofthe Ab repertoire (48–51), as is the case for rabbits (52). BecauseB lymphopoiesis rapidly arrests, the B cell compartment must ex-pand and diversify quickly to generate Abs with biologically rel-evant specificities. In species in which B lymphopoiesis continuesthroughout life, there may be less pressure to produce a B cellcompartment quickly because new B cells with novel specificitiesare continuously being made. Consequently, there may be no needfor GALT in generating the primary Ab repertoire.
Rabbits have features of B cell development similar to those inmice and humans as well as those of GALT species. We considerthe rabbit primarily as a member of the GALT species, however,the timing of B lymphopoiesis is extended, lasting �16 wk. Inaddition, the primary location of B lymphopoiesis in the rabbitappears to be fetal liver and bone marrow, similar to that of miceand humans. Another interesting characteristic of rabbits is thatthey use specific members of the intestinal microflora to assist inthe formation and selection of the B cell repertoire as rabbits lack-ing conventional flora show almost no somatic Ig gene diversity(52). Furthermore, GALT does not involute quickly after expan-sion and diversification of the primary Ab repertoire, which is incontrast to other GALT species, such as chickens and sheep, where
FIGURE 10. Quantitation of V3DJ BRECs in bone marrow DNA. A,Levels in bone marrow DNA from 21 days gestation to 16 wk of age asdetermined by competitive PCR. Each point represents the average of twoto three experiments from a single rabbit. The trend line represents a mov-ing average as in Fig. 8. B, PCR amplification of V3DJ BRECs fromcircular DNA isolated from newborn and 18-mo-old rabbits. [32P]dCTPwas added to the PCR amplification as described in Materials and Methodsand cpm were determined by phosphoimager analysis. Dilution, dilutionfactor of DNA used for PCR amplification.
FIGURE 11. BrdU labeling of proB, preB, and B cells in 1-wk-old rab-bit bone marrow. Rabbits received two injections of BrdU at 12-h intervalsand total bone marrow cells were triple-stained for expression of CD79a(PE), BrdU (FITC), and cyto-� (allophycocyanin), and analyzed by flowcytometric analysis. ProB and preB and B populations were gated as shownand histograms representing anti-BrdU staining of these populations areinlaid within the figure.
6379The Journal of Immunology
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

the bursa and ileal Peyer’s patch involute within weeks or monthsof age.
Through the study of B cell development in different species, itis clear that a variety of mechanisms are used in B cell develop-ment and generation of the Ab repertoire. The rabbit, in particular,provides an opportunity to study a mechanism by which B lym-phopoiesis occurs via the classical pathway in bone marrow, ter-minates during early development, and then the B cell repertoire isexpanded and diversified in GALT giving rise to a competent hu-moral immune system for the life of the animal.
References1. Weber, W. T., and R. Mausner. 1977. Migration patterns of avian embryonic
bone marrow cells and their differentiation to functional T and B cells. Adv. Exp.Med. Biol. 88:47.
2. Houssaint, E., A. Torano, and J. Ivanyi. 1983. Ontogenic restriction of coloni-zation of the bursa of Fabricius. Eur. J. Immunol. 13:590.
3. Reynolds, J. D. 1987. Peyer’s patches and the early development of B lympho-cytes. Curr. Top. Microbiol. Immunol. 135:43.
4. Press, C. M., J. D. Reynolds, S. J. McClure, M. W. Simpson-Morgan, andT. Landsverk. 1996. Fetal lambs are depleted of IgM� cells following a singleinjection of an anti-IgM antibody early in gestation. Immunology 88:28.
5. McElroy, P. J., N. Willcox, and D. Catty. 1981. Early precursors of B lympho-cytes. I. Rabbit/mouse species differences in the physical properties and surfacephenotype of pre-B cells, and in the maturation sequence of early B cells. Eur.J. Immunol. 11:76.
6. Crane, M. A., M. Kingzette, and K. L. Knight. 1996. Evidence for limited B-lymphopoiesis in adult rabbits. J. Exp. Med. 183:2119.
7. Reynaud, C. A., C. R. Mackay, R. G. Muller, and J. C. Weill. 1991. Somaticgeneration of diversity in a mammalian primary lymphoid organ: the sheep ilealPeyer’s patches. Cell 64:995.
8. Reynaud, C. A., V. Anquez, H. Grimal, and J. C. Weill. 1987. A hyperconversionmechanism generates the chicken light chain preimmune repertoire. Cell 48:379.
9. Knight, K. L., and M. A. Crane. 1994. Generating the antibody repertoire inrabbit. Adv. Immunol. 56:179.
10. Glick, G., T. S. Chang, and R. G. Jaap. 1956. The bursa of Fabricius and antibodyproduction. Poult. Sci. 35:224.
11. Mueller, A. P., H. R. Wolfe, and J. Meyer. 1959. Precipitin production in chick-ens. XXI. Antibody production in bursectomized chickens and in chickens in-jected with 19-nortestosterone on the fifth day of incubation. J. Immunol. 83:507.
12. Warner, N. L., J. W. Uhr, G. J. Thorbecke, and Z. Ovary. 1969. Immunoglobu-lins, antibodies and the bursa of Fabricus: induction of agammaglobulinemia andthe loss of all antibody-forming capacity by hormonal bursectomy. J. Immunol.103:1317.
13. Gerber, H. A., B. Morris, and W. Trevella. 1986. The role of gut-associatedlymphoid tissues in the generation of immunoglobulin-bearing lymphocytes insheep. Aust. J. Exp. Biol. Med Sci. 64:201.
14. Cooper, M. D., D. Y. Perey, A. E. Gabrielsen, D. E. Sutherland,M. F. McKneally, and R. A. Good. 1968. Production of an antibody deficiencysyndrome in rabbits by neonatal removal of organized intestinal lymphoid tissues.Int. Arch. Allergy Appl. Immunol. 33:65.
15. Vajdy, M., P. Sethupathi, and K. L. Knight. 1998. Dependence of antibody so-matic diversification on gut-associated lymphoid tissue in rabbits. J. Immunol.160:2725.
16. Kitamura, D., A. Kudo, S. Schaal, W. Muller, F. Melchers, and K. Rajewsky.1992. A critical role of �5 protein in B cell development. Cell 69:823.
17. Ehlich, A., S. Schaal, H. Gu, D. Kitamura, W. Muller, and K. Rajewsky. 1993.Immunoglobulin heavy and light chain genes rearrange independently at earlystages of B cell development. Cell 72:695.
18. Grawunder, U., D. Haasner, F. Melchers, and A. Rolink. 1993. Rearrangementand expression of � light chain genes can occur without � heavy chain expressionduring differentiation of pre-B cells. Int. Immunol. 5:1609.
19. Kubagawa, H., M. D. Cooper, A. J. Carroll, and P. D. Burrows. 1989. Light-chaingene expression before heavy-chain gene rearrangement in pre-B cells trans-formed by Epstein-Barr virus. Proc. Natl. Acad. Sci. USA 86:2356.
20. Novobrantseva, T. I., V. M. Martin, R. Pelanda, W. Muller, K. Rajewsky, andA. Ehlich. 1999. Rearrangement and expression of immunoglobulin light chaingenes can precede heavy chain expression during normal B cell development inmice. J. Exp. Med. 189:75.
21. Shimizu, T., C. Mundt, S. Licence, F. Melchers, and I. L. Martensson. 2002.VpreB1/VpreB2/�5 triple-deficient mice show impaired B cell development butfunctional allelic exclusion of the IgH locus. J. Immunol. 168:6286.
22. Hayward, A. R., M. A. Simons, A. R. Lawton, R. G. Mage, and M. D. Cooper.1978. Pre-B and B cells in rabbits: ontogeny and allelic exclusion of � light chaingenes. J. Exp. Med. 148:1367.
23. Gathings, W. E., R. G. Mage, M. D. Cooper, and G. O. Young-Cooper. 1982. Asubpopulation of small pre-B cells in rabbit bone marrow expresses � light chainsand exhibits allelic exclusion of b locus allotypes. Eur. J. Immunol. 12:76.
24. Cebra, J. J., and G. Goldstein. 1965. Chromatographic purification of tetrameth-ylrhodamine-immune globulin conjugates and their use in the cellular localizationof rabbit gamma-globulin polypeptide chains. J. Immunol. 95:230.
25. Lobel, S. A., and K. L. Knight. 1984. The role of rabbit Ia molecules in immunefunctions as determined with the use of an anti-Ia monoclonal antibody. Immu-nology 51:35.
26. Gallarda, J. L., K. S. Gleason, and K. L. Knight. 1985. Organization of rabbitimmunoglobulin genes. I. Structure and multiplicity of germ-line VH genes: se-rologic and molecular genetic studies of rabbit Ig heavy chains: evidence foradditional C� and C� genes. J. Immunol. 135:4222.
27. Spieker-Polet, H., P. C. Yam, and K. L. Knight. 2002. Functional analysis of I�promoter regions of multiple IgA heavy chain genes. J. Immunol. 168:3360.
28. Sethupathi, P., P. C. Yam, and K. L. Knight. 1995. Lymphoid and non-lymphoidtumors in E �-myc transgenic rabbits. Proc. Natl. Acad. Sci. USA 92:9348.
29. Karasuyama, H., A. Rolink, Y. Shinkai, F. Young, F. W. Alt, and F. Melchers.1994. The expression of Vpre-B/�5 surrogate light chain in early bone marrowprecursor B cells of normal and B cell-deficient mutant mice. Cell 77:133.
30. Ghia, P., E. ten Boekel, E. Sanz, A. de la Hera, A. Rolink, and F. Melchers. 1996.Ordering of human bone marrow B lymphocyte precursors by single-cell poly-merase chain reaction analyses of the rearrangement status of the immunoglob-ulin H and L chain gene loci. J. Exp. Med. 184:2217.
31. Evans, R. J., and G. F. Hollis. 1991. Genomic structure of the human Ig �1 genesuggests that it may be expressed as an Ig� 14.1-like protein or as a canonical Bcell Ig� light chain: implications for Ig� gene evolution. J. Exp. Med. 173:305.
32. Loffert, D., A. Ehlich, W. Muller, and K. Rajewsky. 1996. Surrogate light chainexpression is required to establish immunoglobulin heavy chain allelic exclusionduring early B cell development. Immunity 4:133.
33. Haasner, D., U. Grawunder, C. Kalberer, H. Karasuyama, T. Winkler, andA. G. Rolink. 1994. Influence of surrogate L chain on DHJH-reading frame 2suppression in mouse precursor B cells. Annu. Rev. Immunol. 12:209.
34. Friedman, M. L., C. Tunyaplin, S. K. Zhai, and K. L. Knight. 1994. Neonatal VH,D, and JH gene usage in rabbit B lineage cells. J. Immunol. 152:632.
35. Lanning, D., P. Jasper, and K. Knight. 2002. IgH haplotype exclusion in rabbits.Semin. Immunol. 14:163.
36. Allman, D. M., S. E. Ferguson, V. M. Lentz, and M. P. Cancro. 1993. PeripheralB cell maturation. II. Heat-stable antigenhigh splenic B cells are an immaturedevelopmental intermediate in the production of long-lived marrow-derived Bcells. J. Immunol. 151:4431.
37. Forster, I., and K. Rajewsky. 1990. The bulk of the peripheral B-cell pool in miceis stable and not rapidly renewed from the bone marrow. Proc. Natl. Acad. Sci.USA 87:4781.
38. Fulcher, D. A., and A. Basten. 1997. Influences on the lifespan of B cell sub-populations defined by different phenotypes. Eur. J. Immunol. 27:1188.
39. Johnson, K. M., K. Owen, and P. L. Witte. 2002. Aging and developmentaltransitions in the B cell lineage. Int. Immunol. 14:1313.
40. Tunyaplin, C., and K. L. Knight. 1995. Fetal VDJ gene repertoire in rabbit:evidence for preferential rearrangement of VH1. Eur. J. Immunol. 25:2583.
41. Sehgal, D., G. Johnson, T. T. Wu, and R. G. Mage. 1999. Generation of theprimary antibody repertoire in rabbits: expression of a diverse set of Ig-� V genesmay compensate for limited combinatorial diversity at the heavy chain locus.Immunogenetics 50:31.
42. Nunez, C., N. Nishimoto, G. L. Gartland, L. G. Billips, P. D. Burrows,H. Kubagawa, and M. D. Cooper. 1996. B cells are generated throughout life inhumans. J. Immunol. 156:866.
43. Sehgal, D., E. Schiaffella, A. O. Anderson, and R. G. Mage. 1998. Analyses ofsingle B cells by polymerase chain reaction reveal rearranged VH with germlinesequences in spleens of immunized adult rabbits: implications for B cell reper-toire maintenance and renewal. J. Immunol. 161:5347.
44. Harrison, M. R., and R. G. Mage. 1973. Allotype suppression in the rabbit. I. Theontogeny of cells bearing immunoglobulin of paternal allotype and the fate ofthese cells after treatment with antiallotype antisera. J. Exp. Med. 138:764.
45. Eskinazi, D. P., K. L. Knight, and S. Dray. 1979. Kinetics of escape from sup-pression of Ig heavy chain allotypes in multiheterozygous rabbits. Eur. J. Immu-nol. 9:276.
46. Lanning, D., B. A. Osborne, and K. L. Knight. 2004. Immunoglobulin genes andgeneration of antibody repertoires in higher vertebrates: a key role for GALT. InMolecular Biology of B Cells. F. W. Alt, T. Honjo, and M. S. Neuberger, eds.Elsevier, London, p. 443.
47. Flajnik, M. F. 2002. Comparative analyses of immunoglobulin genes: surprisesand portents. Nat. Rev. Immunol. 2:688.
48. Maybaum, T. A., and J. D. Reynolds. 1996. B cells selected for apoptosis in thesheep ileal Peyer’s patch have enhanced mutational diversity in the Ig V� lightchain. J. Immunol. 157:1474.
49. Sayegh, C. E., and M. J. Ratcliffe. 2000. Perinatal deletion of B cells expressingsurface Ig molecules that lack V(D)J-encoded determinants in the bursa of Fab-ricius is not due to intrafollicular competition. J. Immunol. 164:5041.
50. Ekino, S., T. Urano, H Fujii, and M. Kotani. 1985. The bursa of Fabricius as atrapping mechanism for environmental antigens. Adv. Exp. Med. Biol. 186:487.
51. Arakawa, H., K. Kuma, M. Yasuda, S. Ekino, A. Shimizu, and H. Yamagishi.2002. Effect of environmental antigens on the Ig diversification and the selectionof productive V-J joints in the bursa. J. Immunol. 169:818.
52. Lanning, D., P. Sethupathi, K. J. Rhee, S. K. Zhai, and K. L. Knight. 2000.Intestinal microflora and diversification of the rabbit antibody repertoire. J. Im-munol. 165:2012.
6380 B LYMPHOCYTE DEVELOPMENT IN RABBIT
by guest on June 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from