autor: danny eugenio garcía marrero dr. félix ojeda … · origen, distribución y ecología...
TRANSCRIPT

1
UNIVERSIDAD DE MATANZAS “Camilo Cienfuegos” EEPF “Indio Hatuey”
EEFFEECCTTOO DDEE LLOOSS PPRRIINNCCIIPPAALLEESS FFAACCTTOORREESS QQUUEE IINNFFLLUUYYEENN EENN LLAA CCOOMMPPOOSSIICCIIÓÓNN FFIITTOOQQUUÍÍMMIICCAA
DDEE MMoorruuss aallbbaa ((LLiinnnn..))
AAuuttoorr:: Danny Eugenio García Marrero
TTuuttoorreess:: Dr. Félix Ojeda García Dra. Iraida Splengler Salabarría
Tesis presentada en opción al Título Académico de Master en Pastos y Forrajes
2003

i
A: Dios, ante todas las cosas Mis padres y abuelos Mi familia
No es la inteligencia, recibida y casual, lo que da al hombre honor. Sino el modo con que la usa y la salva,
No hay más que un modo de perdurar Y es servir.
José Martí

Tabla de contenido
ii
Tabla de contenido Introducción______________________________________________________________________________ 1 1. Revisión bibliográfica ____________________________________________________________________ 3
1.1. Morus alba (Linn.)______________________________________________________________ 3 1.1.1. Clasificación taxonómica y descripción botánica ________________________________________ 3 1.1.2. Origen, distribución y ecología ______________________________________________________ 3 1.1.3. Reserva de germoplasma__________________________________________________________ 4 1.1.4. Principales usos _________________________________________________________________ 5 1.1.4.1. Uso como árbol forrajero, consumo y respuesta animal _________________________________ 5 1.1.5. Aspectos agronómicos en el manejo de la especie ______________________________________ 6 1.1.5.1. Siembra y métodos de propagación ________________________________________________ 6 1.1.5.2. Distancia de siembra ____________________________________________________________ 7 1.1.5.3. Fertilización ___________________________________________________________________ 7 1.1.5.4. Frecuencia de defoliación ________________________________________________________ 7 1.1.5.5. Plagas y enfermedades__________________________________________________________ 7 1.1.5.6. Producción de biomasa __________________________________________________________ 7 1.1.6. Composición química y valor nutritivo ________________________________________________ 8 1.1.6.1. Carbohidratos solubles __________________________________________________________ 9 1.1.6.2. Fracción nitrogenada____________________________________________________________ 9 1.1.6.2.1. Proteína bruta_______________________________________________________________ 10 1.1.6.2.2. Proteína verdadera___________________________________________________________ 11 1.1.6.2.3. Fracción nitrogenada unida a la fibra detergente ____________________________________ 11
1.2. Los metabolitos secundarios _____________________________________________________ 11 1.2.1. Definición y Clasificación _________________________________________________________ 11 1.2.2. Distribución natural y función ______________________________________________________ 12 1.2.3. Biosíntesis ____________________________________________________________________ 13 1.2.4. Métodos de cuantificación ________________________________________________________ 15 1.2.5. Efecto de los metabolitos secundarios en los sistemas biológicos__________________________ 16 1.2.6. Metabolitos secundarios presentes en el género Morus. Actividad biológica__________________ 17
2. Metodología experimental________________________________________________________________ 19 2.1. Área experimental ________________________________________________________________ 19 2.2. Períodos de evaluación ____________________________________________________________ 19 2.3. Características climáticas de los períodos evaluados _____________________________________ 19 2.4. Características del suelo ___________________________________________________________ 19 2.5. Diseño experimental y tratamientos __________________________________________________ 20 2.6. Unidad experimental y manejo agronómico ____________________________________________ 20 2.6.1. Distribución de los cortes en los períodos de evaluación_________________________________ 20 2.7. Composición química del fertilizante orgánico __________________________________________ 21 2.8. Análisis cualitativo ________________________________________________________________ 21 2.8.1. Prueba exploratoria _____________________________________________________________ 21 2.8.2. Procedimiento de muestreo _______________________________________________________ 21 2.8.3. Pesquisaje fitoquímico ___________________________________________________________ 21 2.8.3.1. Metabolitos investigados ________________________________________________________ 21 2.8.3.2. Criterios tomados en la realización de las detecciones semicuantitativas __________________ 21 2.8.3.3. Control de reactivos____________________________________________________________ 22 2.9. Análisis cuantitativo _______________________________________________________________ 23 2.9.1. Procedimiento de muestreo, secado y conservación del material __________________________ 23 2.9.2. Instrumentación y mediciones analíticas _____________________________________________ 23 2.9.2.1. Soluciones patrones ___________________________________________________________ 23 2.10. Métodos estadísticos_____________________________________________________________ 23
3. Resultados y discusión __________________________________________________________________ 24 3.1. Experimento 1. “Detección y cuantificación de metabolitos secundarios en cuatro variedades de M. alba
con fertilización y corte”. ____________________________________________ 24 3.1.1. Consideraciones sobre el nivel cero de fertilización_____________________________________ 24 3.1.2. Análisis cualitativo ______________________________________________________________ 24 3.1.2.1. Fenoles _____________________________________________________________________ 24 3.1.2.2. Flavonoides __________________________________________________________________ 25

Tabla de contenido
iii
3.1.2.3. Cumarinas ___________________________________________________________________ 25 3.1.2.4. Carbohidratos reductores solubles ________________________________________________ 26 3.1.2.5. Esteroides ___________________________________________________________________ 26 3.1.2.6. Alcaloides ___________________________________________________________________ 27 3.1.2.7. Saponinas ___________________________________________________________________ 27 3.1.2.8. Metabolitos no detectados_______________________________________________________ 28 3.1.2.9. Análisis de agrupación en las pruebas cualitativas ____________________________________ 28 3.1.3. Análisis cuantitativo _____________________________________________________________ 29 3.1.3.1. Fenoles totales _______________________________________________________________ 29 3.1.3.2. Flavonoides __________________________________________________________________ 32 3.1.3.3. Cumarinas ___________________________________________________________________ 34 3.1.3.4. Carbohidratos solubles _________________________________________________________ 36 3.1.3.5. Esteroles totales ______________________________________________________________ 40 3.1.3.6. Influencia de los factores estudiados en el metabolismo secundario ______________________ 43
3.2. Experimento 2. “Efecto de la parte de la planta, la frecuencia de corte, la fertilización, las variedades y la época en el comportamiento de los compuestos nitrogenados en M. alba”.________ 44
3.2.1. Proteína bruta__________________________________________________________________ 44 3.2.2. Proteína verdadera______________________________________________________________ 50 3.2.3. Fracción unida a la FND__________________________________________________________ 56 3.2.4. Fracción unida a la FAD __________________________________________________________ 58 3.2.5. Dinámica de los compuestos nitrogenados ___________________________________________ 60 3.2.6. Consideraciones sobre el efecto de los factores evaluados_______________________________ 61
Conclusiones____________________________________________________________________________ 62 Recomendaciones________________________________________________________________________ 63 Bibliografía _____________________________________________________________________________ 64 Anexos ________________________________________________________________________________ 75

Indice de tablas
iv
Indice de tablas Tabla I.1. Distribución mundial de M. alba en algunas zonas representativas.___________________________ 4 Tabla I.2. Usos más novedosos de M. alba. _____________________________________________________ 5 Tabla I.3. Contenidos y rangos de CHS (%MS) en las hojas de algunas variedades de M. alba utilizadas en la
sericultura en Asia. Fuente: Yongkang (2002)_____________________________________________ 9 Tabla I.4. Principales factores que afectan el contenido de PB en M. alba. ____________________________ 10 Tabla I.5. Contenidos de PV y porcentaje de PV (%PB), en plantas investigadas para la alimentación animal y
humana en México. Fuente: Sotelo (1997) ______________________________________________ 11 Tabla I.6. Degradabilidad in sacco del N-FND a las 48 horas en algunas arbóreas, comparadas con M. alba.
Fuente: Singh y Makkar (2002) _______________________________________________________ 11 Tabla I.7. Ordenamiento de las especies fenólicas, basado en la cantidad de átomos de Carbono. Fuente:
Harborne (1990)___________________________________________________________________ 12 Tabla I.8. Distribución de los principales compuestos fenólicos en el reino vegetal según el Phylum. Fuente:
Harborne (1990)___________________________________________________________________ 13 Tabla II.1. Principales indicadores del clima en los períodos evaluados. ______________________________ 19 Tabla II.2. Composición química del suelo en las parcelas de cada tratamiento.________________________ 19 Tabla II.3. Distribución de los cortes en los meses del año. ________________________________________ 20 Tabla II.4. Composición química del fertilizante orgánico. _________________________________________ 21 Tabla II.5. Criterios seguidos en las detecciones cualitativas en el pesquizaje fito-químico________________ 22 Tabla II.6. Criterio tomado en dependencia de la altura de la espuma, en el ensayo de “índice de espuma”. __ 22 Tabla II.7. Soluciones utilizadas para el control de los reactivos. ____________________________________ 22 Tabla II.8. Patrones utilizados en el análisis espectrofotométrico. ___________________________________ 23 Tabla III.1. Comportamiento cualitativo de los fenoles en el PPLL. __________________________________ 24 Tabla III.2. Comportamiento cualitativo de los fenoles en el PLL.____________________________________ 24 Tabla III.3. Comportamiento cualitativo de los flavonoides en el PPLL. _______________________________ 25 Tabla III.4. Comportamiento cualitativo de los flavonoides en el PLL. ________________________________ 25 Tabla III.5. Comportamiento cualitativo de las cumarinas en el PPLL. ________________________________ 25 Tabla III.6. Comportamiento cualitativo de las cumarinas en el PLL. _________________________________ 26 Tabla III.7. Comportamiento cualitativo de los carbohidratos en el PPLL. _____________________________ 26 Tabla III.8. Comportamiento cualitativo de los carbohidratos en el PLL. ______________________________ 26 Tabla III.9. Comportamiento cualitativo de los esteroides en el PPLL. ________________________________ 27 Tabla III.10. Comportamiento cualitativo de los esteroides en el PLL. ________________________________ 27 Tabla III.11. Comportamiento cualitativo de los alcaloides en el PPLL. _______________________________ 27 Tabla III.12. Comportamiento cualitativo de los alcaloides en el PLL. ________________________________ 27 Tabla III.13. Agrupación en el análisis cualitativo en el PPLL. ______________________________________ 28 Tabla III.14. Agrupación en el análisis cualitativo en el PLL. _______________________________________ 29 Tabla III.15. Contenido de FT determinados mediante la reducción del ácido fosfomolíbdico en las hojas de
especies forrajeras. ________________________________________________________________ 30 Tabla III.16. Contenido de CHS en las hojas de algunos árboles, arbustos y plantas rastreras, comparados con
M. alba. _________________________________________________________________________ 38 Tabla III.17. Efecto de los grupos Variedad y Fertilización en el contenido de CHS en los tallos tiernos en el PLL.
________________________________________________________________________________ 39 Tabla III. 18. Contenido de ET en especies arbóreas (Galindo et al., 1989), comparadas con el de M. alba, en el
PPLL. ___________________________________________________________________________ 41

Indice de figuras
v
Indice de figuras Fig 1. Vías de síntesis de los principales grupos de metabolitos secundarios y su relación con la fotosíntesis.
Fuente: Azcón-Bieto y Talón (2000) ___________________________________________________ 14 Fig. 2. Comportamiento de los FT en las hojas de cuatro variedades de M. alba en el PPLL.______________ 30 Fig. 3. Comportamiento de los FT en las hojas de cuatro variedades de M. alba en el PLL. _______________ 30 Fig. 4. Comportamiento de los FT en los tallos tiernos de cuatro variedades de M. alba en el PPLL. ________ 31 Fig. 5. Comportamiento de los FT en los tallos tiernos de cuatro variedades de M. alba en el PLL. _________ 31 Fig. 6. Comportamiento de los FT con los factores parte de la fracción comestible y época del año empleando la
media Grand. _____________________________________________________________________ 32 Fig. 7. Comportamiento de los flavonoides con la fertilización en las hojas de M. alba ___________________ 32 Fig. 8. Comportamiento de los flavonoides en las hojas de cuatro variedades de M. alba. ________________ 33 Fig. 9. Comportamiento de los flavonoides con la fertilización en los tallos tiernos de cuatro variedades de M.
alba en el PPLL.___________________________________________________________________ 33 Fig. 10. Comportamiento de los flavonoides con la fertilización en los tallos tiernos de M. alba en el PLL.____ 33 Fig. 11. Comportamiento de los flavonoides en los tallos tiernos de cuatro variedades de M. alba en el PLL. _ 34 Fig. 12. Comportamiento de los flavonoides con los factores parte de la fracción comestible y época del año
empleando la media Grand.__________________________________________________________ 34 Fig. 13. Comportamiento de las cumarinas con la fertilización en las hojas de cuatro variedades de M. alba en el
PPLL. ___________________________________________________________________________ 34 Fig. 14. Comportamiento de las cumarinas con la fertilización en las hojas de cuatro variedades de M. alba en el
PLL. ____________________________________________________________________________ 35 Fig. 15. Comportamiento de las cumarinas con la fertilización en los tallos tiernos de cuatro variedades de M.
alba en el PPLL.___________________________________________________________________ 35 Fig. 16. Comportamiento de las cumarinas con la fertilización en los tallos tiernos de cuatro variedades de M.
alba en el PLL.____________________________________________________________________ 36 Fig. 17. Comportamiento de las cumarinas con los factores parte de la fracción comestible y época del año
empleando la media Grand.__________________________________________________________ 36 Fig. 18. Comportamiento de los CHS en las hojas de cuatro variedades de M. alba en el PPLL. ___________ 37 Fig. 19. Representación gráfica del comportamiento de la EB con la fertilización nitrogenada a los 90 días de
rebrote, en las partes comestibles de M. alba var. Tigriada durante ambas épocas del año. Fuente: González et al., 2002. ______________________________________________________________ 37
Fig. 20. Comportamiento de los CHS con la fertilización en las hojas de cuatro variedades de M. alba en el PLL.________________________________________________________________________________ 37
Fig. 21. Comportamiento de los CHS con la fertilización en los tallos tiernos de cuatro variedades de M. alba en el PPLL. _________________________________________________________________________ 38
Fig. 22. Comportamiento de los CHS en etanol con la fertilización en las hojas de cuatro variedades de M. alba en el PLL.________________________________________________________________________ 39
Fig. 23. Comportamiento de los CHS en alcohol con la fertilización en los tallos tiernos de cuatro variedades de M. alba. a) PPLL, b) PLL ____________________________________________________________ 40
Fig. 24. Comportamiento de los CHS con los factores parte de la fracción comestible y época del año empleando la media Grand.__________________________________________________________ 40
Fig. 25. Comportamiento de los ET con la fertilización en las hojas de cuatro variedades de M. alba en el PPLL.________________________________________________________________________________ 41
Fig. 26. Comportamiento de los ET con la fertilización en las hojas de cuatro variedades de M. alba en el PLL.41 Fig. 27. Comportamiento de los ET en las hojas de cuatro variedades de M. alba en el PLL. ______________ 42 Fig. 28. Comportamiento de los ET con la fertilización en los tallos tiernos de de M. alba en el PPLL. _______ 42 Fig. 29. Comportamiento de los ET con la fertilización en los tallos tiernos en el PLL. ___________________ 43 Fig. 30. Comportamiento de los ET con la fertilización en los tallos tiernos de cuatro variedades de M. alba en el
PLL. ____________________________________________________________________________ 43 Fig. 31. Comportamiento de los ET con los factores parte de la fracción comestible y época del año, empleando
la media Grand. ___________________________________________________________________ 43 Fig. 32. Comportamiento de la PB con la fertilización orgánica en las hojas de M. alba. __________________ 45 Fig. 33. Rendimiento de la PB con la fertilización orgánica en las hojas de M. alba. _____________________ 45 Fig. 34. Comportamiento de la PB con la frecuencia de corte en las hojas de M. alba. ___________________ 45 Fig. 35. Rendimiento de la PB con la frecuencia de corte en las hojas de M. alba. ______________________ 46 Fig. 36. Comportamiento de la PB en las hojas de cuatro variedades de M. alba._______________________ 46 Fig. 37. Rendimiento de la PB en las hojas de cuatro variedades de M. alba. __________________________ 47

Indice de figuras
vi
Fig. 38. Comportamiento de la PB con la fertilización en los tallos tiernos. ____________________________ 48 Fig. 39. Comportamiento de la PB/ha/corte con la fertilización en los tallos tiernos de M. alba. ____________ 48 Fig. 40. Comportamiento de la PB con la frecuencia de corte en los tallos tiernos de M. alba. _____________ 48 Fig. 41. Rendimiento de la PB con la frecuencia de corte en los tallos tiernos de M. alba. ________________ 49 Fig. 42. Comportamiento de la PB en los tallos tiernos de cuatro variedades de M. alba. _________________ 49 Fig. 43. Rendimiento de la PB en los tallos tiernos de cuatro variedades de M. alba. ____________________ 49 Fig. 44. Comportamiento de la PV con la fertilización en las hojas de cuatro variedades de M. alba en el PPLL.
________________________________________________________________________________ 50 Fig. 45. Comportamiento de la PV con la frecuencia de corte en las hojas de cuatro variedades de M. alba en el
PPLL. ___________________________________________________________________________ 51 Fig. 46. Comportamiento de la PV con la frecuencia de corte en las hojas de M. alba en el PPLL.__________ 51 Fig. 47. Comportamiento de la PV con la fertilización en las hojas de cuatro variedades de M. alba en el PLL. 52 Fig.48. Comportamiento de la PV con la frecuencia de corte en las hojas de cuatro variedades de M. alba en el
PLL. ____________________________________________________________________________ 52 Fig. 49. Comportamiento de la PV con la frecuencia de corte en las hojas de M. alba en el PLL. ___________ 53 Fig. 50. Comportamiento de la PV con la fertilización en los tallos tiernos en el PPLL. ___________________ 53 Fig. 51. Comportamiento de la PV con la frecuencia de corte en los tallos tiernos de M. alba en el PPLL. ____ 54 Fig. 52. Rendimiento de la PV y su relación con la frecuencia de corte en los tallos tiernos de M. alba en el PLL.
a) entre las variedades, b) con la fertilización ____________________________________________ 55 Fig. 53. Comportamiento de la PV (%PB) con la frecuencia de corte en las hojas en el PPLL. a) en las
variedades, b) con la fertilización______________________________________________________ 55 Fig. 54. Comportamiento de la PV (%PB) con la frecuencia de corte en los tallos tiernos en PPLL. a) en las
variedades, b) con la fertilización______________________________________________________ 56 Fig. 55. Comportamiento del N-FND en las hojas de M. alba en el PPLL. a) con la fertilización, b) entre las
variedades _______________________________________________________________________ 56 Fig. 56. Comportamiento del N-FND con la frecuencia de corte en las hojas de M. alba. _________________ 57 Fig. 57. Comportamiento del N-FND en los tallos tiernos de M. alba en el PPLL. a) con la fertilización, b) entre
las variedades ____________________________________________________________________ 58 Fig. 58. Comportamiento de N-FAD con la frecuencia de corte en las hojas de cuatro variedades de M. alba en
el PPLL. _________________________________________________________________________ 58 Fig. 59. Comportamiento del N-FAD con la fertilización en las hojas de M. alba.________________________ 59 Fig. 60. Comportamiento del N-FAD en las hojas de M. alba en el PLL. a) con la frecuencia de corte, b) entre las
variedades. ______________________________________________________________________ 59

Abreviaturas
vii
Abreviaturas
A Acorazonada AOAC Association of Official Agricultural Chemistry C Cubana CHS Carbohidratos solubles Cum Cumarinas Deg. Degradabilidad Est Esteroides ET Esteroides totales FAD Fibra ácido detergente FAO Food Agriculture Organization FD Fibra detergente Fig. Figura Flav Flavonoides FND Fibra neutro detergente FT Fenoles totales g Gramo ha Hectárea HPLC High Performance Liquid Cromatography I Indonesia K Potasio KDa Kilodalton kg Kilogramo mg Milígramo ml Mililitro MS Materia seca msnm Metros sobre el nivel del mar N Nitrógeno N-FAD Nitrógeno unido a la fibra ácido detergente N-FND Nitrógeno unido a la fibra neutro detergente NNP Nitrógeno no proteico NP Nitrógeno proteico Nt Nitrógeno total P Fósforo PB Proteína bruta PEG Polietilenglicol PEG-14C Polietilenglicol marcado con carbono radiactivo pH Potencial hidrogeniónico PLL Período lluvioso PPLL Período poco lluvioso Pt Fósforo total PV Proteína verdadera PV(%PB) PV expresada como el porcentaje de la PB t Tonelada T Tigriada Yb3+ Iterbio trivalente

Resumen
viii
Resumen
Con el objetivo de evaluar los principales factores que influyen en la composición fitoquímica de M. alba (Linn.) se efectuaron dos experimentos, en los cuales se caracterizó el metabolismo secundario y la fracción nitrogenada. Mediante el tamizaje fitoquímico, se investigaron 15 grupos de metabolitos de los cuales se detectaron: los fenoles, los flavonoides, las cumarinas, los carbohidratos solubles, los esteroides, los alcaloides y las saponinas. Mediante un análisis de conglomerados se pudo comprobar que la edad de rebrote fue la variable de mayor incidencia en el comportamiento de los metabolitos secundarios en ambas épocas. Los FT se encontraron en las hojas en un rango de 1,26-3,26 %MS y en los tallos tiernos entre 0,77-2,50 %MS; se observó un efecto marcado de las partes de la fracción comestible. Los flavonoides en las hojas estuvieron presentes en niveles de 1,50-1,76 %MS y en los tallos tiernos entre 0,69 y 1,72 %MS; no se observó un efecto significativo de los factores variedad, fertilización y época. Para las cumarinas en las hojas las concentraciones oscilaron entre 0,43 y 0,79 %MS y en los tallos tiernos entre 0,42 y 0,80 %MS; la combinación de los factores Variedad x Fertilización generó las mayores diferencias. Los CHS en las hojas se encontraron entre 10,40 y 18,78 %MS y en los tallos tiernos entre 3,86 y 10,92 %MS. El factor de mayor influencia en los resultados fue la parte de la planta analizada, seguido. Los niveles de ET presentaron grandes variaciones con la época, fundamentalmente en las hojas, donde los contenidos fluctuaron entre 4,08 y 16,65 mg/gMS; en los tallos tiernos las concentraciones oscilaron entre 0,90 y 3,76 mg/gMS. La variedad Cubana mostró diferencias en las tendencias de los compuestos fenólicos en el PPLL. En el segundo experimento la concentración de PB varió entre 17,02 y 22,40 %MS en las hojas y entre 7,72 y 9,68 %MS en los tallos tiernos. Las mayores diferencias en las concentraciones se encontraron entre las hojas y los tallos tiernos, seguido de la frecuencia de corte. La PV presentó valores entre 11,18 y 19,91 %MS en el caso de las hojas y entre 4,17 y 9,00 %MS en los tallos tiernos. De manera similar a la PB, las partes de la fracción comestible y la frecuencia de defoliación fueron los factores que más diferenciaron las concentraciones; la combinación de los factores variedad y fertilización proporcionaron las características más diferenciadas entre las variedades en hojas y tallos. La variedad Cubana también presentó diferencias en las tendencias observadas con el aumento de la fertilización.. Se constató que con el aumento de la edad de rebrote, tanto en las hojas como en los tallos tiernos, aumentó la calidad proteica expresada como PV (%PB).. Las investigaciones realizadas permiten afirmar que los factores fertilización, variedad y época crearon variaciones numéricas discretas entre los tratamientos, aunque existió una tendencia a diferenciarse las variedades cuando se combinaron con el factor fertilización. Las concentraciones de los metabolitos secundarios y la fracción nitrogenada de M. alba se vieron afectadas primariamente por la edad de rebrote y las diferencias encontradas entre las partes de la fracción comestible de la planta.

Introducción
1
Introducción
En el trópico Latinoamericano, la baja productividad del ganado está relacionada directamente con la poca disponibilidad en los pastizales y el pobre valor nutritivo que presentan los pastos, donde el comportamiento estacional de las especies pratenses determina un pobre suministro de biomasa en la época poco lluviosa y, por consiguiente, una deficiente respuesta animal (Sánchez, 2002).
Es conocido que la productividad ganadera mejora sustancialmente cuando se dispone de forraje suficiente de aceptable valor nutritivo para satisfacer los requerimientos de los animales. En este sentido, los pastos y forrajes constituyen la fuente basal alimenticia más económica para los sistemas de producción ganadera a nivel mundial.
Como producto de la situación económica actual, los países del tercer mundo están obligados a generar alternativas en el campo de la alimentación que permitan satisfacer las necesidades crecientes de la población. Recientemente están emergiendo modelos agropecuarios basados en el aumento de la producción, pero a su vez con la reducción de la dependencia de insumos externos, con vistas a disminuir los costos e incrementar los beneficios económicos por unidad de área en armonía con el ambiente.
En Cuba la alimentación animal se basa fundamentalmente, a través del consumo de pastos, forrajes, subproductos, y en menor medida a partir de alimentos conservados como ensilajes, henos y harinas.
La ganadería cubana está precisada a producir alimento durante todo el año, con la menor cantidad de recursos externos. Teniendo en cuenta que los pastos por sí solos no cubren los requerimientos nutricionales de los rumiantes, para una adecuada producción de leche y carne, los árboles son una buena fuente alternativa para su utilización como alimento suplementario. Estos se caracterizan por presentar elevados contenidos de proteínas y una alta digestibilidad comparada con la de los pastos (Simón, Hernández y Ojeda, 1998; Simón, 1998).
Según Hernández y Babbar (2001) los árboles desempeñan un importante papel para la sostenibilidad de los sistemas; aportan sombra; protegen los cultivos contra el viento; pueden ser usados como combustible y material de construcción; protegen de la escorrentía; no permiten el escape de Carbono y reciclan nutrientes de forma natural.
No obstante, la mayoría de estas plantas contienen elevados niveles de metabolitos secundarios, compuestos que bajo determinadas circunstancias pueden causar efectos diversos y hasta contrastantes en la fisiología animal (Ojeda, 1996).
El estudio del metabolismo secundario de forma integral, en las principales especies arbóreas utilizadas en la alimentación animal, resulta de vital importancia en aras de realizar un mejor manejo de la biomasa que aportan y un óptimo aprovechamiento de esta fuente de alimento.
Existen muchas especies de árboles y arbustos con buenas características forrajeras; en este sentido, la especie M. alba sobresale como fuente de forraje en Cuba por su excelente capacidad de producción de biomasa, composición química (Duke, 2001), alta degradabilidad (González, Delgado y Cáceres, 1998), adaptabilidad a diversas condiciones de clima y suelo (Datta, 2002), perennidad ante el corte (Martín, Reyes, Hernández y Milera, 2002) y disponibilidad (Benavides, 1994). Su forraje fresco o ensilado se utiliza como suplemento proteico para los rumiantes; puede estimular altos niveles de producción de leche y ganancias de peso (Benavides, 1999). Dadas estas características, esta planta se proyecta como una alternativa alimenticia con alto potencial en el futuro.
El uso principal de la morera a nivel mundial ha sido como alimento del gusano de seda; de ahí que la mayoría de las investigaciones realizadas en esta planta hayan sido orientadas a su producción en la sericultura. A partir de la década de los ochenta en América Central comenzó a evaluarse su potencial forrajero y se recomendó su uso en sistemas de corte y acarreo para ovinos, caprinos, bovinos y en la alimentación de monogástricos. No obstante, en la actualidad se carece de estudios encaminados a la dilucidación de su metabolismo secundario.
Si bien la composición química de esta planta ha sido ampliamente estudiada, no se conoce con profundidad aspectos relacionados con su fracción nitrogenada, y se ha investigado muy poco sobre los metabolitos secundarios presentes en la parte comestible, sus concentraciones, la repercusión nutricional, así como el efecto de los principales factores que influyen en sus variaciones.
Teniendo en cuenta los antecedentes anteriormente expuestos, la presente tesis tuvo como hipótesis de
trabajo: La frecuencia de corte, la fertilización, la variedad y la época, influyen en el metabolismo secundario y en la
concentración de los compuestos nitrogenados presentes en la fracción comestible de M. alba.

Introducción
2
Para dar respuesta a esta premisa, se propusieron los siguientes objetivos de trabajo. Objetivo general: Caracterizar la composición fitoquímica integral de la fracción comestible de M. alba Objetivos específicos: • Detectar y cuantificar los principales grupos de metabolitos secundarios presentes en la fracción
comestible de M. alba, teniendo en cuenta la frecuencia de corte, la fertilización, la variedad y la época del año.
• Determinar la concentración de los principales componentes nitrogenados en la fracción comestible de M. alba, teniendo en cuenta la frecuencia de corte, la fertilización, la variedad y la época del año.

Capítulo I-Revisión bibliográfica
3
1. Revisión bibliográfica 1.1. Morus alba (Linn.) 1.1.1. Clasificación taxonómica y descripción botánica
La especie M. alba pertenece al género Morus, familia Moraceae, orden Urticales, subclase Dicotiledónea, clase Angiosperma y división Spermatophyta. Dentro de esta familia también se encuentran otros géneros tales como Brossonetia, Ficus y Cudrania (Cifuentes y Kee-Wook, 1998).
M. alba es una planta leñosa perenne, de porte bajo a medio, semicaducifolia en las condiciones de Cuba, de rápido crecimiento, monoica o dioica, y con un sistema radical profundo. Presenta una copa aproximadamente redondeada y ramificada, con tronco de corteza grisácea que llega a medir 60 cm de diámetro. Hojas generalmente alternas, pecioladas, simples, íntegras, brillantes y estipuladas de uno a cinco lóbulos, con la haz lampiña y el envés ligeramente tomentoso en las axilas de los nervios principales; se pueden observar de anchamente ovadas a orbicular-ovadas, con ápice agudo o cortamente acuminado; base oblicua y semitruncada o subcordada. El borde es dentado o irregularmente lobulado de consistencia blanda. El pecíolo es grande, de 12 x 8 cm en las ramas con frutos, y 25 x 20 cm en las ramas sin frutos. La inflorescencia es simple, axilar, en amentos de color crema o verdosos; con pedúnculos pendientes o colgantes, en la que se encuentra las flores unisexuales en espigas densas de hasta 2 cm de largo, con cuatro sépalos. Las espigas masculinas se caen rápidamente, las flores están arregladas descuidadamente y después de dejar caer el polen la inflorescencia se seca. Las espigas femeninas son usualmente cortas y las flores están en forma compacta, éstas presentan cuatro lóbulos y cuatro estambres en yema, el estigma es bífido, el ovario unicelular madura en un fruto agregado (sincarpo) de drupéolas; de forma obloide a oblongo-cilíndrico de 1 a 5 cm de largo; este puede ser blanco, rosado o púrpura, aunque generalmente es negro violáceo.
El principal agente polinizador de la morera es el viento. La mayoría de las especies son diploides con 28 cromosomas; sin embargo, las triploides se cultivan
también extensivamente por su adaptabilidad, crecimiento, vigor y calidad de las hojas. La sistematización de M. alba ha sido engorrosa como producto de que las especies y variedades de morera
son llamadas con diferentes nombres locales, lo que no ayuda al ordenamiento taxonómico y dificulta la homogeneidad en la clasificación a nivel mundial (Cappellozza, 2002).
En los siglos XIX y XX se realizaron varias divisiones del género Morus, las que estuvieron fundamentalmente basadas en la presencia o ausencia de estilo en la flor, la protuberancia y vellosidad del estigma, la inflorescencia, la sorosis, la base del estilo bilobulado y la morfología de la hoja, principalmente la forma de la base. Recientemente, en la clasificación taxonómica propuesta por Yongkang (2002), M. alba se diferencia del resto de las especies del género por presentar pistilos con estilos largos distintivos, protuberancia dentro del estigma, hojas pequeñas desprovistas de vellos o con protuberancia en la etapa joven, venas en la superficie inferior y sorosis violácea obloide de 1 a 2,5 cm. 1.1.2. Origen, distribución y ecología
Por ser M. alba una especie cosmopolita, se ha hecho extremadamente difícil situar con seguridad el origen de esta planta; no obstante, varios autores señalan al Himalaya como el lugar más probable de origen (Benavides, 2000; Sánchez, 2001; Datta, 2002). Los dos reportes más ancestrales que incluyen a la morera en la historia de la humanidad provienen del año 1 123 antes de Cristo; 3 126 años atrás en Korea (Ho-Zoo y Won-Chu, 2001), y de la dinastía Ming en China (Xiangrui y Hongsheng, 2001). Por su parte Li (2001), clasificó los lugares de origen de M. alba en cinco regiones: 1) Este del continente asiático, 2) archipiélago de Malasia, 3) Suroeste de Asia, 4) Oeste de África y 5) Norte, Centro y Sur de América. La tabla I.1. muestra la distribución de las especies y variedades de morera en algunas de las zonas más representativas del mundo.
Desde tiempos ancestrales los árboles de morera han crecido de forma individual y silvestre en diferentes partes del mundo. Con el inicio de la sericultura, estas plantas fueron llevadas a diversos países para iniciar la producción del gusano de seda, por lo que existen evidencias de que la domesticación de la morera comenzó hace unos 5000 años.
El género Morus se ha distribuido en casi todo el mundo, tanto en áreas templadas como tropicales, donde solo la especie M. rubra es oriunda de América, y el continente australiano es el único que no cuenta con ninguna de las especies de morera en la actualidad (Sánchez, 2002).
China, la India y Brasil son los países más representativos en cantidades de este cultivo por hectárea (Sánchez, 2002), aunque en este último la mayor cantidad de variedades se han obtenido por cruzamientos genéticos (Almeida y Fonseca, 2002).

Capítulo I-Revisión bibliográfica
4
Esta especie puede crecer sobre los 4 000 msnm (FAO, 1990), un rango amplio de temperatura entre 13 y 38 ºC (Lim, Young-Taek y Sang-Poong, 1990), precipitaciones entre 600-2 500 mm y humedad relativa entre 65 y 80 % (Datta, 2002).
Tabla I.1. Distribución mundial de M. alba en algunas zonas representativas.
? no se conoce con exactitud
M. alba se adapta bien a diversos tipos de suelo, principalmente en aquellos que presentan mayor fertilidad (Cifuentes y Ham-Kim, 1998), con buen contenido de materia orgánica (Datta, 2002), bien drenados, de textura media de arcilloarenosas o arenoarcillosas y de topografía plana u ondulada con pendientes inferiores al 40 % (Cifuentes y Kee-Wook, 1998). Es tolerante a la salinidad y a la acidez, pero prefiere los suelos neutros con pH entre 6,5 y 6,8. 1.1.3. Reserva de germoplasma
La morera, por sus condiciones naturales de distribución, posee un amplio germoplasma, formado por una gran diversidad de especies y variedades, que constituye un valioso recurso genético. La sericultura ha sido la razón principal del impresionante proceso de selección y mejoramiento del género Morus a través del tiempo en los diferentes países, principalmente en Asia y Europa.
No obstante, Machii (2002) reporta que la mayor diversidad de especies se concentra en Asia, especialmente en China y Japón, por lo que se han propagado gran cantidad de variedades de excelente capacidad de producción de biomasa, calidad y alta resistencia a plagas y enfermedades, para un amplio rango de condiciones de clima y suelo. Las especies más importantes de este género son M. alba, M. nigra, M. indica, M. laevigata y M. bombycis, de las cuales se han originado innumerables clones, incluyendo muchas poliploides como producto de la intensiva selección y mejoramiento genético en muchos países (Sánchez, 2002).
En la India existen muchas especies, tanto en poblaciones naturales como bajo producción comercial; cuatro de ellas son las de mayor importancia: M. alba, M. indica, M. serrata y M. laevigata (Datta, 2002). Por otra parte, en China existen 15 variedades y se cultivan o se mantienen en colección cerca de 1 000 accesiones, la mayoría de estas originadas principalmente de M. multicaulis, M. bombycis y M. atropurpurea (Yongkang, 2002).
Machii, Koyama y Yamanouchi (2002) informan en Japón 24 especies y una subespecie, muchas de las cuales proceden de M. bombycis, M. alba y M. latifolia.
A pesar del creciente interés en esta planta, fuera de Asia la disponibilidad de germoplasma de morera es mucho más limitada.
En Italia, Cappelloza (2002) informa la existencia de una colección que cuenta con 51 accesiones conformada principalmente por M. alba y M. nigra. En Brasil se cuenta con tres colecciones con un total de 90 variedades, todas provenientes de M. alba (Almeida y Fonseca, 2002). En América Central, Benavides (2002) menciona cuatro variedades de M. alba, denominadas Criolla, Indonesia, Tigriada y Acorazonada, las cuales fueron introducidas a la región en el siglo pasado, cuando se intentó desarrollar la producción del gusano de seda. En Cuba, después de la primera introducción desde Costa Rica, la Estación Experimental de Pastos y Forrajes “Indio Hatuey” incrementó su germoplasma a un total de 10 variedades con la inclusión de algunas accesiones brasileñas (Martín G., comunicación personal).
Zona Geográfica País Especies del
género Morus Variedades de M. alba Referencia
China 5 10 Yongkang (2002) Indonesia 6 2 Viet Nam 3 100
Sánchez (2002)
Japón 6 19? Machii (2002) Asia
India 4 17? Ravindran y col (1997) América Brasil 1 90 Almedia y Fonseca (2002) Europa Italia 5 25 Cappellozza (2002) África Tanzania 1 ? Shayo (2002)

Capítulo I-Revisión bibliográfica
5
1.1.4. Principales usos
La morera es una verdadera planta multipropósito; aun cuando está distribuida ampliamente alrededor del mundo, ha recibido poca atención con respecto a su potencial de uso.
Su utilización en Cuba comenzó en la década del 40, cuando fue plantada por primera vez para el desarrollo de la sericultura (Martín et al., 2002).
El uso principal y más difundido a nivel mundial es en la sericultura como alimento del gusano de seda (Bombix mori), el cual consume las hojas de morera y las transforma en capullos, que luego son devanados y convertidos en finos hilos, con los cuales se producen telas para la elaboración de múltiples confecciones. Después de la decadencia de la industria de la seda, su uso se ha diversificado vertiginosamente. Dada su elevada adaptabilidad y grado de selección, se reportan más de una decena de usos en el mundo, y en la actualidad más de 42 países la utilizan de una u otra forma. Del total de naciones que cultivan la morera, el desglose según su uso corresponde a 60 % en actividades agrícolas; 48 % en la fabricación de la seda y como forrajera; 26 % en labores de jardinería, paisajismo y preparación de infusiones; 31 % como alimento y 14 % como frutal, además de emplearse para mejorar el ecosistema (Sánchez, 2002).
Independientemente de su utilización en la sericultura, se reconocen otros múltiples empleos y beneficios (Zepeda, 1991), los cuales demuestran el potencial de explotación desde el nivel familiar hasta la industria. En algunos países como México, Egipto, Turquía, Grecia, Japón y Korea, se utiliza como árbol frutal. La fruta, llamada mora, se consume fresca o procesada como jugo, mermelada, frutos secos y para fermentar y hacer vino (Arias y Sánchez, 2002; Gerasopoulos y Stavroulakis, 1997). En otros lugares como Argentina, Bolivia, Perú, Estados Unidos, Francia, Grecia, Italia y España se utiliza como planta ornamental y como árbol de sombra (Sánchez, 2002)
La madera de troncos y ramas se emplea como leña, en la elaboración de algunas piezas e implementos, ebanistería y construcción (Ye, 2002); en Japón la pulpa de la madera se utiliza para elaboración de papel. La tabla I.2. presenta un resumen de los usos más novedosos de M. alba en la actualidad.
Tabla I.2. Usos más novedosos de M. alba.
Su uso como medicina natural es milenario, en países como China y Japón le atribuyen propiedades
curativas a las hojas, los frutos y la corteza de las raíces, por la elevada actividad biológica de los metabolitos secundarios presentes.
Duke (2001) resume más de 60 propiedades terapéuticas en las diferentes partes de la especie. Se emplea en tratamientos para algunas enfermedades como la diabetes, la hipertensión arterial, la deposición de colesterol, la filiariasis y como laxante, antihelmíntico, expectorante y diurético (Xiangrui y Hongsheng, 2001), también las hojas deshidratadas son usadas en infusiones a manera de té (Yongkang, 2002) y el látex se utiliza con éxito en la industria farmacéutica.
M. alba presenta un gran potencial para el control de la erosión, especialmente en áreas con grandes pendientes (Pizarro, Ramos y Almeida, 1997). Su uso como forraje ha demostrado un gran potencial, por la calidad y producción de su follaje, características organolépticas y alto consumo animal (Benavides, 1996).
En algunas zonas de Tailandia, las hojas y brotes tiernos son consumidos como vegetales, su abundante fructificación permite mantener la biodiversidad animal, especialmente de aves y mamíferos. 1.1.4.1. Uso como árbol forrajero, consumo y respuesta animal
Las características nutricionales, la elevada producción de biomasa, la versatilidad agronómica, la palatabilidad, la tolerancia a la sequía y la disponibilidad mundial que posee M. alba, en comparación con otros
Usos Parte utilizada País Utilización Referencia
Construcción India Cabinas, muebles, decoraciones y carruajes Datta (2002)
Medio de cultivo
Tallo China Multiplicación de Ganoderma lucidum
y Auricularia auricula judae Yongkang (2002)
Materia prima Ramas y corteza
India y Japón Fabricación de papel Machii (2002)
Combustible Madera India Material energético Datta (2002) Alimento animal Forraje - Ganado, iguanas, caracoles, peces y
aves de corral Sánchez (2002)

Capítulo I-Revisión bibliográfica
6
forrajes utilizados tradicionalmente, hacen de esta planta una opción importante para la intensificación de los sistemas ganaderos.
El conocimiento de su potencial como forraje comenzó de forma empírica en América Central, al principio de la década del 80. No obstante, Sánchez (2002) hace referencia a que su uso como alimento animal ya era reconocido desde hace mucho tiempo, en varias regiones de Asia, India y Europa, donde estas plantas crecían de forma silvestre.
A partir de ese momento ha surgido un creciente interés por el estudio de su potencial alimenticio, manejo agronómico y formas de utilización en los sistemas de producción animal, no solo en Centroamérica y el Caribe, sino en otros países de Europa, África, India y Asia; incluyendo aquellos donde la morera ha sido tradicionalmente utilizada para la alimentación del gusano de seda, como en el caso de China y Japón.
En las áreas de producción en cultivos puros o asociados, así como en aquellos donde la morera crece en forma natural, el corte y acarreo es la manera más práctica de usarla como forraje para el ganado (Benavides, Esquivel y Lozano, 1995). La mayoría de los resultados obtenidos en este sistema han sido específicamente con cabras y corderos como suplemento de dietas basales y de baja calidad. Con respecto a su empleo en sistemas de producción con bovinos, las investigaciones al respecto son pocas (González, Benavides, Kass y Esperance, 2000).
En corderos en crecimiento, la sustitución parcial del concentrado por morera fresca, ofrecido como suplemento, mejoró significativamente las ganancias de peso (Ríos, Rondón y Combellas, 2000).
En cabras lecheras, Rojas y Benavides (1994) encontraron incrementos en la producción de leche de 2,0 a 2,5 kg/animal/día cuando la suplementación con morera pasó de 1,0 a 2,6 % del PV en base seca, con ligeros incrementos en los contenidos de grasa, proteína y sólidos totales de leche.
En vacas lecheras en pastoreo Oviedo (1995), al comparar el follaje de morera y concentrado como complemento, obtuvo un nivel de producción de leche similar (13,2 y 13,0 kg/animal/día, respectivamente) para cada suplemento a iguales niveles de consumo de MS (1,0 % del PV) y muy superior al obtenido con solo pastoreo (11,3 kg/animal/día). En terneras Jersey destetadas a los 90 días se alcanzaron pesos superiores a los 120 kg a la edad de 165 días, al ofertar ad libitum morera fresca y picada, en combinación con concentrado comercial a razón de 1,0 kg/animal/día, observándose un nivel de consumo máximo de 1,8 % del peso vivo (Jiménez, Aguirre, Ibrahim y Pezo, 1998).
En monogástricos, la posibilidad de utilizar la morera en la dieta base constituye un gran avance. En ensayos realizados con cerdos de preceba, la sustitución parcial del pienso convencional por harina de hojas de morera resultó ser una alternativa (Leiva, López y Quiñones, 2002).
En conejos, al sustituir entre el 25 y el 50 % de los concentrados comerciales por harina de morera, se lograron mantener ganancias de 20 g/animal/día, lo que implicó una disminución en los costos de alimentación (García, Mederos, Salinas y Reyes, 2002). También existe la posibilidad de empleo en otros animales domésticos como cuyes (Albert y Contreras, 2002) y aves de corral (Machii, 2002).
Recientemente se ha estado evaluando su potencial como planta de ramoneo en asociación con pastos; algunos resultados, aunque preliminares, son promisorios. Los primeros intentos de incorporar la morera dentro de sistemas de pastoreo, se iniciaron en Italia (Talamucci y Pardini, 1993, Talamucci, Pardini y Argenti, 2002), donde se estableció una asociación de morera con Trifolium subterraneum con el fin de evaluar y comparar el efecto del pastoreo de ovinos, bovinos y bovinos más ovinos en la producción y consumo de la asociación.
En Japón, además de los trabajos donde se ha evaluado el efecto de las variedades de morera y la época de pastoreo, en la digestibilidad de la MS (Ezenwa y Kitahara (2001), se han obtenido resultados interesantes relacionados con los sistemas asociados, en este sentido Kitahara y Kawano (2002) y Kitahara, Shibata y Nishida (2002), en estudios de más de tres años, han combinado a M. alba en pastoreo con cinco especies de pastos templados; los autores recomendaron este sistema como una alternativa en la obtención de cantidades apreciables de biomasa con buena calidad nutritiva para zonas templadas.
Aunque la mejor característica de esta combinación radica en los incrementos progresivos en las producción de MS, comparada con los sistemas puros (Kitahara, Shibata, Takahashi y Nishida, 2002), la disminución abrupta en los porcentajes de supervivencia (39-56 % después de cuatro años) con marcos estrechos de siembra, fue el principal inconveniente en el empleo de la especie al introducir bovinos (Kitahara, Shibata, Kawano, Takahashi y Nishida, 2002). 1.1.5. Aspectos agronómicos en el manejo de la especie 1.1.5.1. Siembra y métodos de propagación
La morera se puede propagar por semillas, estacas, acodos e injertos. Fuera de China, donde se realiza fundamentalmente por semilla botánica, el principal método de propagación es el vegetativo a través de estacas (Arias y Sánchez, 2002).

Capítulo I-Revisión bibliográfica
7
Cifuentes y Ham-Kim (1998) plantean que el medio de siembra debe servir de anclaje para que la estaca cuente con un buen balance de agua y aire, además que el terreno esté libre de organismos patógenos en el período de brotamiento. 1.1.5.2. Distancia de siembra
En cultivos de uso intensivo en la sericultura, los marcos de siembra propician densidades aproximadas de 25 000 plantas/ha (Cifuentes y Kee-Wook, 1998). Con la finalidad de estudiar la producción de biomasa para la alimentación animal, las distancias más empleadas entre plantas y surcos son de 60 x 60 cm, 90 x 90 cm y 120 x 120 cm (Boschini, Dormond y Castro, 1998; Boschini, Dormond y Castro, 1999).
En Cuba se recomienda para la siembra de este cultivo, distancias entre surcos de 1 m y 40-50 cm entre plantas; a partir de la experiencia práctica obtenida mediante los estudios agronómicos realizados por Martín et al. (2002).
1.1.5.3. Fertilización
La fertilización es la base del crecimiento de M. alba, siempre y cuando los demás factores como el clima y el suelo sean favorables (Cifuentes y Kee-Wook, 1998). La morera es muy exigente al N, P y K, por lo que su fertilización es imprescindible. En la sericultura siempre se ha empleado la fertilización química, donde se recomiendan dosis nitrogenadas de 350 kg/ha/año, entre 100 y 150 de P y 250 de K; independientemente de esto Lim et al. (1990), señala el incremento significativo en la producción de hojas con la combinación de abonos orgánico y químico.
Desde el punto de vista práctico, en cultivos destinados a estudios bioquímicos y agronómicos se han empleado dosis nitrogenadas a partir de gallinaza equivalentes a 100, 300 y 500 kg N/ha/año (García, Ojeda y Pérez, 2002). Por su parte Benavides, Lachaux y Fuentes (1994) utilizaron estiércol de cabra equivalentes a 240, 360, 480 kg N/ha/año y 480 kg a partir de NH4NO3. En otros estudios en América Central Rodríguez, Arias y Quiñones (1994) emplearon varias frecuencias de cortes y diversificaron las dosis nitrogenadas desde 160 hasta 640 kg N/ha/año a partir de urea. 1.1.5.4. Frecuencia de defoliación
Al igual que en los demás árboles forrajeros, el tiempo mínimo de recuperación de M. alba después de la poda es aproximadamente de 2 meses (Francisco, 2002; Rodríguez et al., 1994). Las principales investigaciones relacionadas con el manejo de la poda se han llevado a cabo utilizando 8, 12 y 16 semanas (Martín et al., 2002). Por su parte Rodríguez et al. (1994) emplearon 6, 9 y 12 semanas y Boschini (2002a) realizó un estudio de composición química empleando 56, 84 y 112 días. 1.1.5.5. Plagas y enfermedades
Como en la mayoría de las plantaciones, la morera es atacada por varias plagas y enfermedades. La mayor parte de estos organismos son reportados en países que producen seda de forma tradicional. En este sentido, se destaca la incidencia de Empoasca sp., Saissetia nigra y Tetranychus equitorius. Entre las principales enfermedades sobresalen la llaga de la raíz (Rosellinia sp.), las manchas foliares (Cercospora moricola y Septogloeum mori), la roya de la hoja (Aecidium mori) y las manchas de fuego (Ascochyta moricola y Homodendrum mori) (Cifuentes y Kee-Wook, 1998).
Según Lezcano (1999), el género Morus es afectado de forma general, por ocho enfermedades, cuyos agentes causales se clasifican en un micoplasma, una bacteria y siete hongos.
En un estudio reciente realizado en Cuba por Lezcano y Alonso (2002) se informa la incidencia de Cercosporella mori en las primeras variedades de morera introducidas en el país. 1.1.5.6. Producción de biomasa
Algunas de las características más sobresalientes de M. alba son su excelente producción de biomasa por unidad de área y su alta retención de hojas durante el período seco.
La información disponible acerca de la producción de biomasa está exclusivamente relacionada con las hojas, ya que es la parte utilizada para alimentar al gusano de seda.
En cuanto a la producción de forraje, algunos estudios demuestran que M. alba presenta altos rendimientos de MS a través de los ciclos anuales de producción.

Capítulo I-Revisión bibliográfica
8
El rendimiento de la morera es afectado por una serie de factores, de los cuales se destacan la densidad de siembra, la fertilización y la edad de la planta (Ye, 2002).
La mayoría de los resultados obtenidos señalan que los factores que influyen marcadamente en el rendimiento de la morera son la densidad de siembra y la frecuencia de corte (Boschini et al., 1998; Boschini, 2002b).
En el trópico seco de Guatemala se obtuvieron 19 t MS/ha (Blanco, 1992). Bajo las mismas condiciones Rodríguez et al. (1994), al utilizar tres intervalos de poda y tres niveles de fertilización nitrogenada, lograron un aumento significativo de producción de biomasa (6,5 t MS/ha) con la mayor frecuencia de poda (12 semanas) al incrementar el nivel de fertilización a 80 kg N/ha/corte.
Boschini et al. (1998) evaluaron la influencia de la distancia de siembra, la altura de corte (30 y 60 cm) y la frecuencia de defoliación en la producción de biomasa, la cual alcanzó 35 t MS/ha/año con la menor distancia de siembra (60 cm) y la mayor frecuencia de corte (112 días) La altura no tuvo una influencia significativa en los rendimientos alcanzados.
Por otra parte Martín et al. (2002), al utilizar dos alturas de corte y tres frecuencias, obtuvieron los mejores resultados para la frecuencia de 90 días (25 t MS/ha/año).
Con el uso de abonos orgánicos como fuente de nutrimento se han obtenido resultados alentadores en la producción de biomasa (Benavides et al., 1994), demostraron que la adición de cantidades crecientes de estiércol de cabra (expresado en kg N/ha/año) ejerció un efecto positivo importante en los rendimientos de materia seca de todos los componentes de la biomasa, ya que se obtuvo una producción de más de 35 t MS/ha/año.
La posibilidad de intercalar leguminosas herbáceas, arbustivas o arbóreas, así como otros cultivos para utilizar su follaje como abono verde, es una alternativa que puede estimular buenos niveles de producción de biomasa (Reyes, Milera y Matías, 2000).
Otros factores tales como la variedad y las condiciones edafoclimáticas también pueden influir en la producción de biomasa.
Martín, Yépes, Hernández y Benavides (1998) evaluaron cuatro variedades de morera fertilizadas con cachaza a razón de 40 t/ha. Con respecto a producción de biomasa, la variedad que más se destacó fue la Cubana, que alcanzó la mayor producción de biomasa total (8,2 t de MS/ha); sin embargo, la variedad Acorazonada obtuvo los mayores rendimientos (4,6 t MS/ha) en cuanto a biomasa comestible y tallos tiernos (2,4 t MS/ha).
En tres sitios de Costa Rica, Espinoza y Benavides (1996) reportaron rendimientos de MS total de 14,1, 22,3 y 25, 4 t/ha/año para las variedades Criolla, Indonesia y Tigreada, respectivamente. Este autor encontró diferencias en la producción, atribuibles a factores climáticos. En Paquera, donde ocurrió un largo período de sequía, la producción promedio de todas las variedades (31,2 t MS/ha/año) duplicó la de Coronado (15,5 t de MS/ha/año) a pesar de su mejor régimen de lluvia. Esto se atribuyó a la mayor luminosidad y las mayores temperaturas en Paquera, así como a la alta nubosidad y la menor temperatura en Coronado. 1.1.6. Composición química y valor nutritivo
M. alba es considerada una planta extremadamente peculiar; su composición química y su calidad desde el punto de vista nutricional se suman también a las características distintivas de la especie.
Presenta una composición aminoacídica similar a la de la harina de soya; definida como una gran fuente de aminoácidos, de los cuales la mitad son aminoácidos esenciales (Sánchez, 2002). Los contenidos de cenizas totales pueden llegar a ser superiores al 15 % en dependencia del grado de fertilización del suelo (Triguero y Villalta, 1997), aunque normalmente oscilan entre 10 y 15 % (Shayo, 1997; Shayo, 2002).
Las hojas contienen gran cantidad y diversidad de macro y micro elementos (Noda, 1998), llegándose a observar acumulaciones cuantitativas de Calcio en los idioblastos de las células (Sugimura, Mori, Nitta, Kotani, Furusawa, Tatsumi, Kusakari, Wada, Morita, 1999).
Esta planta presenta apreciables niveles de vitaminas, fundamentalmente de los grupos B y C, donde se destacan los ácidos nicotínico y pantoténico, la riboflavina (Ho-Zoo y Won- Chu, 2001) y el ácido ascórbico (0,3 %) (Singh y Makkar, 2002).
La fracción fibrosa de M. alba es baja, comparada con otros forrajes tales como Medicago sativa, Digitaria decumbens y el pasto elefante (Van Soest, 1992). Los tallos tiernos de mayor edad, contienen una fracción fibrosa similar, y en algunos casos inferior, a la de los pastos tropicales, además de presentar un grado de lignificación menor (Boschini, 2002a); en su conjunto, las hojas de la especie se caracterizan por presentar una composición química superior a la de los pastos tropicales y subtropicales, y los tallos no lignificados presentan mejor potencial nutricional que los encontrados en las hojas de los pastos característicos del trópico.
En la actualidad, su valor nutritivo ha sido estudiado mediante todas las técnicas de digestibilidad y degradabilidad disponibles en el mundo. Jegou, Waelput y Bronschwig (1994), en un experimento in vivo

Capítulo I-Revisión bibliográfica
9
utilizando cabras, demostraron que las hojas tuvieron una digestibilidad superior al 78 %, y mediante técnica in vitro se comprobó un porcentaje de desaparición entre 80 y 90 % (Rodríguez et al., 1994).
Por otra parte, en los estudios desarrollados por González et al. (1998) la degradabilidad ruminal de las hojas y los tallos tiernos, empleando bolsas de nailon, fue superior al 80 % a las 48 horas, lo que demuestra la mayor digestibilidad de estas porciones comparadas con otros forrajes tradicionales como Leucaena leucocephala (Tolera, Seyoum y Sundstol, 1998).
Adicionalmente, Schmidek, Takahashi, Nuñes de Medeiros y Resende (2002) observaron degradaciones de la MS, la PB y la FND de 93,3; 97,0 y 84,9; respectivamente.
Mediante la técnica de producción de gases, Bing, Jianxin y Jun (2001) llegaron a la conclusión que el estado de maduración de la hoja, así como el período del año, influían en la cantidad de gas producido; mientras que Makkar y Becker (1996) demostraron que las hojas jóvenes presentaban un potencial doblemente superior de producción de gases (60,6 ml/200mg) al compararlas con las hojas maduras.
1.1.6.1. Carbohidratos solubles
Uno de los aspectos más sobresalientes de M. alba lo constituye el equilibrio entre los niveles de carbohidratos solubles y proteínas que presentan tanto las hojas como los tallos tiernos, lo que la diferencia de la mayoría de las gramíneas y leguminosas (Colectivo de autores, 2000).
Son abundantes los reportes provenientes de la sericultura que describen las concentraciones de CHS de las especies de morera, variedades y clones; no obstante, la diversidad de factores climáticos, geográficos y fisiológicos que han influido en los experimentos, no esclarecen la influencia independiente de estos en las variaciones de los niveles energéticos de M. alba.
Las investigaciones más abarcadoras se han desarrollado en las variedades de morera oriundas del continente asiático (Yongkang, 2002), donde se observó una gran variabilidad en los contenidos de CHS relacionada fundamentalmente con las variaciones ambientales y genética.
La tabla I.3. muestra los contenidos de CHS en las hojas de alguna de las variedades más utilizadas en la sericultura en China.
Tabla I.3. Contenidos y rangos de CHS (%MS) en las hojas de algunas variedades de M. alba utilizadas en la
sericultura en Asia. Fuente: Yongkang (2002)
NS8 HYS DHS XGS Y2 YS2 DZS 23,5-26,1 22,5-31,0 27,7-29,4 18,8-22,0 22,8-26,2 21,5 22,0-24,6
1.1.6.2. Fracción nitrogenada
M. alba también se distingue de otros árboles multipropósitos por las características particulares de su fracción nitrogenada, que aunque es comparable con la que presentan la mayoría de las leguminosas forrajeras del trópico, esta tiene una calidad nutritiva y proteica superior (Benavides, 1999; González et al., 1998).
Desde el punto de vista cualitativo, la bibliografía recoge consensos divididos en cuanto a la principal proteína presente en las hojas. Sánchez (2002) señala a la Ribulosa-1,5-bisfosfato carboxilasa (RuBisCO) como la principal proteína en la especie, cuyo centro activo es responsable de la fijación del CO2 (Kellogg y Juliano, 1997). De forma independiente, Yamashita y Ohsawa (1990) demostraron que el 43 % del Nt en M. alba pertenece a este compuesto. Por otra parte, Singh y Makkar (2002) señalan a la Prolamina, aislada a partir del extracto alcohólico-alcalino de las hojas, como una proteína importante, la cual contiene el 12,6 % del Nt, distribuido fundamentalmente en N insoluble en HCl, amidas y ácidos mono y diaminados.
También han sido purificadas y caracterizadas las estructuras primarias y secundarias de dos glicoproteínas con actividad antidiabética, denominadas Moran A y Moran 20K con pesos moleculares de 7,50 y 21,86 KDa, respectivamente (Eun-Sun, Sung-Jean, Eun-Ju, Bak-Kwang, Hoon-Huh, Bong-Jin, 1999).
La solubilidad de la fracción nitrogenada, aunque no es elevada 17,3 (%Nt) en buffer de borato-fosfato y 15,7 (%Nt) en buffer de fosfato, es comparable con la de L. leucocephala y especies de los géneros Dendrocalamus, Artocarpus y Picus, lo que evidencia la naturaleza no proteica del N soluble. En cambio, otras especies de leguminosas tales como Acacia catechu, Albizia stipulata y Bauhinia variegata presentan una mayor solubilidad del Nt (Singh y Makkar, 2002). Otros autores empleando la misma técnica analítica, reportan una solubilidad del Nt inferior al 36 % (Sarma, Singh y Bhat, 2000).

Capítulo I-Revisión bibliográfica
10
1.1.6.2.1. Proteína bruta
La determinación de los niveles de PB en M. alba, mediante la utilización del análisis proximal, ha sido el método analítico más empleado por los autores que investigan la composición química de la especie (Deshmukh, Pathak y Takalikar, 1993; Trigueros y Villalta, 1997; Shayo, 1997; Boschini, 2002a). No obstante, este método si bien muestra una idea general de la dimensión cuantitativa de la fracción, no permite conocer las características cualitativas, y la calidad proteica del material nitrogenado presente. Las determinaciones simultáneas de PB y de PV aclaran con mayor solidez las propiedades y características propias del material nitrogenado (Liu, Zunyao, Yan, Shi, Wang y Yu, 2002).
Existen numerosas investigaciones en las cuales se ha determinado el contenido de PB en las partes comestibles de especies de morera, con interés en la alimentación animal y en la sericultura; en cultivos intensivos en la India se han obtenido tenores de hasta un 39 % (Singh y Makkar, 2002).
La parte de la planta es el factor que más diferencia las concentraciones de PB (Espìnosa et al, 1999). Otros factores tales como la variedad (Yongkang, 2002), la fertilización química u orgánica (Benavides et al., 1994), así como la fertilización basal del suelo y el tipo de abono (Ramos, Lara, Rivera y Sanginés, 2002), también influyen en los rangos de este indicador.
Al igual que la mayoría de las plantas arbóreas, los factores época (González y Cáceres, 2002), condiciones ambientales, altura de corte (Martín et al., 2002) y densidad de plantación (Boschini et al., 1998), afectan en menor medida los contenidos nitrogenados.
Con el cúmulo de material empírico, obtenido en las investigaciones sobre esta temática, ya se conocen patrones estables de comportamientos sobre la base de la fisiología vegetal en las plantas arbóreas perturbadas por el corte (Hernández, 2000; Francisco, 2002); en este sentido, la edad de rebrote es un factor determinante en la concentración de PB. Los mayores tenores de PB se observan en las frecuencias de defoliación más intensas, las cuales producen hojas de menor edad de rebrote. A partir de los 90 días después del corte, en la mayoría de las plantas arbóreas, incluyendo la morera, se obtienen contenidos similares de PB (Boschini, 2002a; Francisco, 2002).
La tabla I.4. muestra la incidencia de los principales factores que afectan el contenido de PB en M. alba.
Tabla I.4. Principales factores que afectan el contenido de PB en M. alba.
Factor PB (%MS) Referencia
Parte de la planta Hoja Tallo tierno
21,10 11,70 Espinosa et al. (1999)
Frecuencia de corte 56días 84 días 112 días
25,6 22,2 20,8
Boschini (2002a)
Fertilización química (urea)
0 kg N 240 kg N 480 kg N
17,1 16,6 17,4
Rodríguez et al. (1994)
Fertilización orgánica (Estiércol de cabra)
0 kg N 240 kg N 480 kg N
19,1 19,3 20,2
Benavides et al. (1994)
Época PPLL PLL
23,1 22,7 González y Cáceres (2002)
Variedad Indonesia Tigriada
21,06 20,19 Benavides (2002)
La influencia de la fertilización química en las concentraciones de PB en esta arbórea es polémica y
controvertida. La aplicación de 480 kg N/ha/año, a partir de NH4NO3, produce incrementos significativos en las concentraciones de PB en las hojas (Benavides et al., 1994); mientras que con la misma aplicación nitrogenada (480 kg/ha/año), a partir de urea, se observó poco efecto en los tenores de PB en las hojas y en la biomasa total (Rodríguez et al., 1994). Por otra parte, con la aplicación de fertilizante orgánico (0 a 480 kg N/ha/año) a partir de estiércol, tampoco se observaron incrementos considerables en los contenidos de PB, en las partes comestibles de la planta (Benavides et al., 1994).

Capítulo I-Revisión bibliográfica
11
1.1.6.2.2. Proteína verdadera
Existen escasos reportes sobre la determinación de PV en M. alba, así como la influencia de factores tales como la edad, el estado fisiológico, las variedad y la parte de la planta. No obstante, las investigaciones desarrolladas por Bing et al. (2001) y Liu et al. (2002) determinaron en M. alba que más del 80 % del Nt forma parte de la PV, sin diferencias apreciables en los contenidos encontrados en cuatro clones de morera tanto en el PPLL como el PLL, aunque se observaron ligeros incrementos en el porcentaje de PV (%PB) en el estado de mayor edad de la planta, comparado con el rebrote joven de la primavera. Adicionalmente, se encontraron ligeras diferencias en los contenidos de PV (%PB) entre los dos períodos evaluados (88,2 vs 85,8 %).
En un estudio realizado por Sotelo (1997) donde se determinó los contenidos de PV, así como el porcentaje de PV (%PB) en plantas silvestres y cultivadas, se obtuvieron concentraciones variables de estos tipos de compuestos relacionadas con la naturaleza y el hábitat de cada especie.
La tabla I.5. muestra los contenidos de PV y el porcentaje de PV (%PB) de algunas especies de la flora mexicana.
Tabla I.5. Contenidos de PV y porcentaje de PV (%PB), en plantas investigadas para la alimentación animal y
humana en México. Fuente: Sotelo (1997)
Especies PV (%MS) PV (%PB) Piper auritum 23,8 88,15 Crotalaria pumila 23,8 65,03 Cnidoscolus acutifolium (Chaya) 23,9 70,71
Silvestres
Portulaca oleracea (Verdolaga) 12,7 43,94 Spinacea olaracea (Espinaca) 23,2 64,99 Beta vulgaris 12,6 45,32 Cultivadas Medicago sativa (Alfalfa) 12,0 42,11 Erythina americana (Piñón) 13,7 59,05 Flores Cucurbita pepo (Calabaza) 14,7 55,47
1.1.6.2.3. Fracción nitrogenada unida a la fibra detergente
Las investigaciones relacionadas con la caracterización de la porción nitrogenada retenida en la fracción fibrosa de M. alba y con los factores que influyen en su comportamiento son escasas, como producto del poco interés que había tenido esta parte de la composición química en la sericultura. Estudios recientes realizados por Sarma et al. (2000) indican que M. alba, al igual que otras especies de los géneros Bauhinia, Cassia, Ficus, Melia y Quercus, presentan una considerable porción del N unido a la FND, entre 30 y 70 % del Nt.
Schmidek et al. (2002) encontraron diferencias significativas en los contenidos de N-FND entre tres clones de morera en Brasil, no así en las concentraciones de N-FAD.
Aunque en esta planta una parte considerable del Nt está unido a la porción fibrosa, además de una solubilidad baja del Nt en parte por esta unión, la principal diferencia de M. alba con el resto de las especies en este sentido, radica en la elevada degradabilidad de la fracción enlazada, quizás relacionada con las características químico-físicas de la fibra en esta especie, la distribución espacial de los componentes de la pared celular o la naturaleza de los enlaces que unen la fracción nitrogenada al esqueleto fibroso.
La tabla I.6. muestra la degradabilidad in sacco del N-FND a las 48 horas en algunas arbóreas comparadas con M. alba.
Tabla I.6. Degradabilidad in sacco del N-FND a las 48 horas en algunas arbóreas, comparadas con M. alba. Fuente: Singh y Makkar (2002)
Especies A. stipulata A. catechu B. variegata L. leucocephala M. alba %Deg. N-FND 0 11 27 56 79
1.2. Los metabolitos secundarios 1.2.1. Definición y Clasificación
El metabolismo secundario se puede definir como la biosíntesis, transformación y degradación de compuestos endógenos mediante proteínas de especialización, las cuales se han formado como resultado de

Capítulo I-Revisión bibliográfica
12
procesos de diferenciación y se clasifican según su significación biológica y función en la célula productora (Valdés y Balbín, 2000).
Según la definición en el marco ecológico propuesta por Stransburger, Noll, Schenk y Schimper (1994), estos compuestos son sustancias ecológicamente eficaces, frente a compuestos primarios que serían sustancias fisiológicamente eficaces.
Los compuestos que derivan de este tipo de metabolismo se pueden clasificar de varias formas. El ordenamiento estrictamente químico, basado en los principales grupos funcionales, es la forma más
secuenciada de organización (Ikan, 1991). Desde el punto de vista de su incidencia negativa en la nutrición, se pueden clasificar según el tipo de metabolito con que interactúa (Delgado, 1998), definiendo a los factores antinutricionales como aquellas sustancias generadas por el metabolismo natural de las especies vegetales y que, por diferentes mecanismos, ejercen efectos contrarios a la nutrición óptima de los animales por la disminución de los efectos digestivos y/o metabólicos (Ojeda, 1996).
La tabla I.7 muestra la complejidad ascendente de las cadenas carbonadas, basado en el fraccionamiento estructural para el caso particular de los compuestos fenólicos (Harborne, 1990).
Tabla I.7. Ordenamiento de las especies fenólicas, basado en la cantidad de átomos de Carbono. Fuente:
Harborne (1990)
Núm. de Átomos
Esqueleto carbonado básico Clases
6 C6 Fenoles simples y Benzoquinonas 7 C6-C1 Ácidos fenólicos 8 C6-C2 Acetofenonas y Acidos fenilacéticos
9 C6-C3 Acidos hidroxicinámicos, fenilpropanoides, Cumarinas, Isocumarinas y Cromonas
10 C6-C4 Naftoquinonas 13 C6-C1-C6 Xantonas 14 C6-C2-C6 Estilbenos y Antraquinonas 15 C6-C3-C6 Flavonoides e Isoflavonoides 18 (C6-C3)2 Lignanos y Neolignanos 30 (C6-C3-C6)2 Biflavonoides
N (C6-C3)n (C6)6, (C6-C3-C6)n
Ligninas,Melaninas de catecol y Flavolanos
Para un mismo grupo funcional, además de la clasificación por las cadenas carbonadas presentes, la
separación de los metabolitos secundarios también se realiza teniendo en cuenta la prioridad de organización de cada tipo de compuesto, la posición del grupo funcional, su configuración y la naturaleza de los residuos terminales (Thomsen, 1997). 1.2.2. Distribución natural y función
Una de las principales diferencias que presentan los metabolitos secundarios con relación a los primarios es su distribución limitada en el reino vegetal; mientras que los compuestos primarios se encuentran en todo el reino y las diferencias entre especies solo son de índole cuantitativa (Ramos, Frutos, Giráldes y Mantecón, 1998).
Estos compuestos químicos se encuentran típicamente en sólo una especie o un grupo de plantas taxonómicamente relacionadas (Gabriela-Anca, Dean y Wiley, 1997); por lo que muchos de ellos se consideran marcadores taxonómicos de familias y géneros (Garbarino, 2003).
Los metabolitos secundarios se encuentran principalmente en las plantas, pero cada estructura básica varía considerablemente acorde con el Phylum en consideración.
La tabla I.8. muestra la distribución de los principales compuestos fenólicos en el reino vegetal. Los compuestos fenólicos no son comunes en bacterias, hongos ni algas; donde las clases de esqueletos
hidroxilados son escasos, comparados con el resto de las plantas (Harborne, 1990); los flavonoides también están casi ausentes; sin embargo, los derivados clorinados solamente son producidos por el hongo Aspergillus candidus.
Los hongos y algas no sintetizan fenoles en asociación simbiótica, los líquenes producen especialmente sustancias fenólicas las que pueden ser coloreadas; estos organismos se caracterizan por tener dépsidos o

Capítulo I-Revisión bibliográfica
13
depsidonas, pero pueden ser sintetizados otras clases de mayor distribución como las Xantonas y Antraquinonas (Markham, 1988).
Tabla I.8. Distribución de los principales compuestos fenólicos en el reino vegetal según el Phylum. Fuente:
Harborne (1990)
Phylum Patrones estructurales Bacteria Fenoles derivados de policétidos y quinonas (ocasionalmente presentes). Hongos Fenoles simples, fenilpropanoides y quinonas (regularmente reportadas). Algas Fenoles iodados y bromados, derivados del floroglucinol en la pared celular. Líquenes Antraquinonas, dépsidos, depsidonas y xantonas
Bryofitas Fenoles en la pared celular, fenilpropanoides, estilbenos y algunos flavonoides.
Helechos, coníferas y plantas que florecen
Ligninas en la pared celular, amplio rango de fenoles de todo tipo.
En las plantas vasculares se encuentran la mayor cantidad de polifenoles (Kumar, 1992), los helechos,
Gymnospermas y Angiospermas contienen ligninas en la pared celular. Los Ácidos hidroxibenzóico y hidroxicinámico, además de los flavonoides, están universalmente presentes en todas las plantas; aunque otras clases tienen una distribución más discreta como el caso de los isoflavonoides, ampliamente reportados en la familia Leguminosae (Keh-Feng y Yuh-Fang, 1996), Antraquinonas (comunes en seis familias botánicas) y las Xantonas; abundantes en Gentianaceae, Guttiferae, Moraceae y Polygalaceae (Harborne, 1990).
Por otra parte, en cada planta, las yemas en crecimiento, las hojas jóvenes, los órganos reproductores y de dispersión, en general, todas las partes en crecimiento anual, muestran una mayor concentración, reactividad y diversidad de metabolitos secundarios (Makkar, Dawra y Singh, 1991).
Desde el punto de vista ultraestructural, los metabolitos secundarios se encuentran principalmente en las vacuolas, la periferia adyacente interna, o el centro de los orgánulos citoplasmáticos (Lees, Suttill y Gruber, 1993), además de que sus concentraciones se diferencian en las diferentes partes de la planta (Harborne, 1990).
El término “secundario” implicaba en el principio de las investigaciones que estas sustancias tenían un menor importancia y muchas veces se les atribuyó la propiedad de productos de desecho del metabolismo primario. Esta idea ha sido gradualmente cambiada, ya que los compuestos secundarios desempeñan un papel protagónico en la fisiología de la planta, la regulación del crecimiento, su desarrollo y la interacción con otros organismos (Raskin, 1992), por lo que a partir de 1960 se han realizado investigaciones que han hecho evidente la importante función ecológica de muchos de ellos (Valdés y Balbín, 2000).
Cada tipo de compuesto secundario está estrechamente relacionado con una o varias funciones específicas en la planta que lo contiene.
Son innumerables los reportes que describen el papel que desempeñan estos compuestos en el reino vegetal, aunque hasta la fecha, en la mayoría de ellos, no se conoce con exactitud cada función particular. En algunos casos intervienen en las relaciones de competencia con otras plantas, actuando como agentes alelopáticos y contra invasiones de hongos, bacterias y virus (Harborne, 1993), en relaciones de mutualismo en la atracción de polinizadores y dispersores de semillas (Ramos et al., 1998), en funciones defensivas causando toxicidad (Foo, Lu, Mc Nabb, Woghorn y Ulyatt, 1997), como protección contra la radiación ultravioleta y desecación (Ghasempour, Anderson, Gianello y Gaff, 1998), como reserva de material nitrogenado (Poulton, 1990), en la fijación del N2 atmosférico, la formación de nódulos y la relación simbiótica en las raíces de las leguminosas (Stafford, 1997).
Uno de los metabolitos secundarios mejor estudiado en cuanto a su distribución natural y función es el Ácido salicílico (Raskin, 1992). Este fenol simple está presente en las estructuras reproductivas y las hojas de 34 importantes especies botánicas empleadas en la agricultura, lo que confirma su distribución extensiva (Raskin, Skubatz, Tang y Meeuse, 1990). Este ácido induce la floración, participa en la regulación del potencial de las membranas celulares y la resistencia de enfermedades, induce cambios de temperatura en las plantas termogénicas, interviene en la expresión de algunos genes y actúa como atrayente de parásitos del género Striga. 1.2.3. Biosíntesis
Las rutas biosintéticas que forman parte del metabolismo secundario, es un tema que en los últimos años ha llamado poderosamente la atención. La síntesis específica de cada compuesto suele estar restringida a estados

Capítulo I-Revisión bibliográfica
14
específicos del desarrollo respectivo de cada tipo de organismo, células especializadas, períodos de estrés causados por la deficiencia de nutrientes, o por el ataque de microorganismos. Este fenómeno se debe a la formación, dependiente de fase, de la correspondiente enzima, lo que significa que la expresión del metabolismo secundario se basa en un proceso de diferenciación (Azcón-Bieto y Talón, 2000).
Muchos factores afectan la síntesis de los metabolitos secundarios; la concentración de CHS, el estado acuífero y la temperatura ambiental son algunos de los más importantes.
Atendiendo a las vías biosintéticas que les dan origen, los compuestos secundarios se dividen en tres grandes grupos: los terpenos, las sustancias fenólicas y los compuestos que contienen nitrógeno en su estructura (Valdés y Balbín, 2000).
El metabolismo primario proporciona un gran número de moléculas simples, como el Ácido shiquímico, el acetato y los aminoácidos, los cuales constituyen los materiales de partida para las rutas biosintéticas del metabolismo secundario. El Ácido shiquímico; por la ruta metabólica que lleva su nombre, da origen a muchos compuestos aromáticos, entre ellos los aminoácidos, los ácidos cinámicos y algunas estructuras polifenólicas. El acetato es el precursor de los ácidos grasos y de los policétidos en la ruta del acetato-malonato, y los terpenos o isoprenoides son sintetizados en la ruta del acetato-mevalonato.
Los aminoácidos son precursores de los alcaloides y de los antibióticos peptídicos. Todos los terpenos naturales proceden de unidades de acetato activo (acetil-CoenzimaA), que se
condensan y transforman para originar Ácido mevalónico, unidad de 5 átomos de carbono, específica de la biosíntesis de terpenos.
La fig. 1. describe la interconexión de los principales grupos de metabolitos secundarios y su relación con la fotosíntesis. Vía de la Pentosa fosfato eritrosa 4P CHS Glicolisis Fosfoenol piruvato Vía del Shikimato Acetil CoA Vía del Acetato malato Ciclo de Krebs (ácido cítrico) Aminas alifáticas Aminas aromáticas Vía del Acetil-Mevalónico
Fig 1. Vías de síntesis de los principales grupos de metabolitos secundarios y su relación con la fotosíntesis. Fuente: Azcón-Bieto y Talón (2000)
La biosíntesis de las estructuras fenólicas se efectúa por dos rutas metabólicas esenciales; la del Ácido
shiquímico y la del Ácido malónico (conocida como la de los policétidos); la primera es mayoritaria en las plantas superiores y la segunda es favorecida en los microorganismos.
La Fenilalanina y la Tirosina son intermediarios metabólicos en la biosíntesis de numerosos compuestos fenólicos, y el Triptófano es el precursor de hormonas como el Ácido indolacético.
Fotosíntesis
Terpenoides Compuestos Nitrogenados
Compuestos Fenólicos

Capítulo I-Revisión bibliográfica
15
Los alcaloides se forman por rutas biosintéticas muy diversas, a partir de muchos aminoácidos como la L-arginina, la L-lisina, la L-fenilalanina, el L-triptófano y otros. Ocasionalmente derivan de la L-prolina, el ácido antranílico (precursor y producto de degradación del L-triptófano), el ácido nicotínico (formado a partir del ácido aspártico), raras veces del L-triptófano, restos de isopentenilo, isopentenil pirofosfato, acetilo proveniente del acetil-CoenzimaA y de grupos metilo de S-adenosilmetionina.
Según Valdés y Balbín (2000), en la biosíntesis de los alcaloides se han reconocido tres tipos generales de reacción que determinan su estructura principal, la formación de bases de Schiff, la reacción de Mannich, y el acoplamiento oxidativo de fenoles. 1.2.4. Métodos de cuantificación
Existen numerosos métodos para cuantificar los contenidos de metabolitos secundarios de interés en la nutrición animal. No obstante, cada técnica analítica presenta limitaciones intrínsecas, por lo que no son aplicables de manera general en todos los casos.
Las diferentes coloraciones y cantidad de color producidas por unidad de masa en los métodos colorimétricos (Schofield, Mbugua y Pell, 2001), la variabilidad en los resultados con la utilización de diferentes patrones y los diferentes mecanismos en las técnicas de precipitación (Krueger, Carter Dopke, Treichel, Folts y Reed, 2000), son las mayores limitantes en las marchas analíticas más utilizadas en la actualidad.
El grupo de metabolitos que más se ha cuantificado en el campo de la nutrición animal son los compuestos fenólicos, como producto de su amplia distribución en las plantas de interés agrícola y su repercusión en la fisiología digestiva de rumiantes y monogástricos (Mueller-Harvey, 2001).
Los métodos colorimétricos de Folin-Dennis y Folin-Ciocalteau, propuestos en los trabajos realizados por estos autores entre los años 1912 y 1927, se basan en la reducción del Ácido fosfomolíbdico hasta óxidos azules de Molibdeno con estados de oxidación inferiores a 7+, se han convertido en técnicas universales en la determinación de FT (Singleton, Orthofer y Lamuela-Raventós, 1999; Mueller-Harvey, 2001); sin embargo, Schofield et al. (2001) plantean que el método del Azul de Prusia (FeCl3/K3Fe(CN)6), en sentido general, propicia mejores lecturas.
Con el desarrollo vertiginoso de la química analítica ha sido validado un gran grupo de procedimientos modernos, que abarcan desde las más simples técnicas, hasta la utilización de los instrumentos más sofisticados (Harborne, 1998; Makkar, 1999).
Según la recopilación realizada por Schofield et al. (2001), los métodos para la cuantificación de compuestos fenólicos se pueden clasificar en: 1) Colorimétricos, 2) Cromatográficos, 3) Gravimétricos, 4) De inhibición enzimática, 5) Por precipitación, 6) Toxicológicos.
Las lecturas mediante el desarrollo de color son las más empleadas mundialmente, porque requieren menor complejidad de instrumentos y son más accesibles.
El ensayo del Butanol-HCl es específico para taninos condensados, pero requiere de patrones internos fidedignos a la naturaleza de la muestra analizada, ya que el color varía con la estructura tánica. Aunque es un método clásico, no es totalmente confiable para la determinación si no se conoce el tipo de tanino presente (Dallzell y Kerven, 1998).
La utilización de la Vainillina-HCl es específica para meta difenoles y se pueden cuantificar los monómeros de taninos condensados obtenidos después de la hidrólisis del material; no obstante, si están presentes unidades de flavonoides se pueden obtener sobreestimaciones de los contenidos de estos tipos de taninos (Sun, Ricardo da Silva y Spranger, 1998).
El método de Azul de Prusia es muy utilizado por presentar buena correlación con la actividad biológica. Aunque mediante este reactivo reaccionan todos los fenoles, las transformaciones dependen mucho de las condiciones del análisis y pueden interferir otros agentes reductores. Las cuantificaciones de diferentes estructuras hidroxiladas mediante el procedimiento de Tiólisis y el empleo de Floroglucinol, requieren de HPLC y son excelentes para determinaciones estructurales.
Mediante la Tiólisis se requieren taninos puros y con la utilización de Floroglucinol los rendimientos tienden a disminuir, además de que los bencil mercantanos formados presentan olores desagradables y las rupturas de los enlaces, en algunos casos, no son cuantitativas (Schofield et al., 2001).
La utilización de Yb3+ como agente precipitante de fenoles no requiere de patrones, pero los rendimientos pueden variar con la relación Yb/fenol, además de que la muestra necesita ser incinerada (Mueller-Harvey, 2001).
Los ensayos enzimáticos precisan de manera adicional otras técnicas biológicas y la susceptibilidad de la enzima puede ser variable (Adams y Harbertson, 1999), por lo que estas técnicas por sí solas no permiten llegar a conclusiones sólidas (Hagerman, 1998; Fickel, Pitra, Joest y Hofmann, 1999).

Capítulo I-Revisión bibliográfica
16
Los procedimientos de precipitación con proteínas reflejan una importante repercusión biológica, pero los resultados son diferentes en dependencia de la proteína seleccionada para el análisis (Hagerman, Zhao y Jonson, 1997).
El empleo de PEG para la obtención de resultados totalmente concluyentes requiere de Carbono marcado (14C-PEG).
Los procedimientos que emplean HPLC son factibles para determinaciones de estructuras complejas (Hedqvist, Mueller-Harvey, Reed, Krueger y Murphy, 2000), pero algunos solo son aplicables para taninos condensados de 7 a 8 unidades, y en ocasiones se observan uniones irreversibles con la matriz utilizada (Lazarus, Adamson, Hammerstone y Schmitz, 1999; Hammerstone, Lazarus, Mitchell, Rucker y Schmitz, 1999; Labarbe, Cheynier, Broussard, Bouguet y Moutounet, 1999).
Las pruebas de inhibición del crecimiento microbiano constituyen uno de los mejores ensayos biológicos en la actualidad, aunque la selección de la bacteria y el medio de cultivo pueden afectar los resultados. Por otra parte, se requiere de niveles relativamente elevados de polifenoles porque se presentan inconvenientes relacionados con la competencia del agente enlazante (Nelson, Pell, Doane, Giner-Chávez y Schofield, 1997).
Otros métodos significativamente importantes son el empleo de la Rodanina para la cuantificación de taninos hidrolizables del tipo Galotaninos (Mueller-Harvey, 2001), el empleo de KIO3 en las determinaciones de Galo y Elagitaninos (Willis y Allen, 1998), las determinaciones de peso molecular (Guyot, Doco, Bouguet, Moutounet y Driliau, 1997) y el apoyo que propicia la cromatografía de fase normal (Cheinier, Bouguet, Le Roux, Guyot y Rigaud, 1999) y de fase reversa en las separaciones cuantitativas de las diferentes especies fenólicas (Waterhouse, Price y McCord, 1999).
Los procedimientos analíticos más factibles para la determinación de cumarinas y flavonoides se basan en las propiedades estructurales de estos compuestos de tener patrones conformacionales rígidos, que le permiten una absorción muy intensa en el espectro ultravioleta, después de separaciones y fraccionamientos previos que evitan un gran número de interferencias analíticas (Mochiutti, 1995; Méndez, 1996; Gutiérrez, Miranda, Varona y Rodríguez, 2000); no obstante, se debe conocer aproximadamente las estructuras a cuantificar con vistas a realizar una óptima selección del patrón en el análisis (Mueller-Harvey, 2001)
Otros metabolitos secundarios, como los triterpenos y esteroles, se determinan fundamentalmente por diversos métodos cromatográficos (AOAC, 1990) o mediante la extensión de la prueba cualitativa de Liebermam y Buchard, basado en la formación de compuestos coloreados, como producto de la reacción en medio ácido del Anhídrido acético, con el doble enlace presente en los anillos B y C de los esteroides y triterpenos, respectivamente; instauraciones que resultan típicas en ambas agrupaciones (Galindo, Rosales, Murgueitio y Larrahondo, 1989). 1.2.5. Efecto de los metabolitos secundarios en los sistemas biológicos
A lo largo de la evolución la refinada especialización del metabolismo secundario ha constituido una de las adaptaciones más sorprendentes en las plantas superiores, con el objetivo de lograr mantener el equilibrio interespecífico en la naturaleza.
El propósito fundamental de esta diferenciación enzimática estuvo encaminado, en primer lugar, a que estos compuestos cumplieran funciones específicas en el metabolismo vegetal, además de actuar como defensas importantes frente a los herbívoros, por la imposibilidad de huir ante estos (Harborne, 1993; Cheecke, 1995; Ramos et al., 1998).
Es evidente que la amplia diversidad de estructuras generadas por estas proteínas especializadas produce efectos extremadamente diversos en la biología de los organismos vivos.
Indistintamente, en la literatura se reportan efectos negativos relacionados con problemas de toxicidad en insectos herbívoros, aves, pequeños mamíferos, cerdos, rumiantes y el hombre (Mueller-Harvey y Mc Allan, 1992); así como efectos beneficiosos en la producción industrial de fármacos (Mateos, 2003), la terapia contra el cáncer y la biotecnología (Anon, 2003a), en la domesticación y diversificación de plantas de interés agrícola (Anon, 2003b), en estudios biológicos y moleculares (Torpoco y Garbarino, 2003) y en las ciencias médicas (Anon, 2003c). Otro efecto polémico es la acción defaunante que causan en la microbiología ruminal (Galindo, Castillo, Aldama, Marrero, García y Martínez, 1998; Colectivo de autores, 2003).
La concentración de los metabolitos secundarios en el tejido vegetal es uno de los principales elementos que diferencian la acción positiva o detrimental en la nutrición animal (Aerts, Barry, Warren y Mc Nabb, 1999), aunque la variabilidad estructural dentro de un mismo grupo funcional, la isomería de posición y los alargamientos de los radicales carbonados, también diferencian la accción particular de cada compuesto en los diferentes animales (Baker, Medlock y Sheehan, 1998; Cassidy, Hanley y Lamuela-Raventos, 2000).
En el campo de la nutrición animal a nivel mundial, los taninos condensados han sido el grupo de compuestos mejor estudiados en cuanto a su repercusión fisiológica y su amplia distribución (Wolfgang y Shelton, 1995; Ben Salem, Nefzaoui, Ben Salem y Tisserand, 1999 a,b; Ben Salem, Nefzaoui, Ben Salem y

Capítulo I-Revisión bibliográfica
17
Tisserand, 2000); otros metabolitos inhiben la digestión, al afectar la actividad catalítica de algunas enzimas (Delgado, 1998), y pueden restringir la absorción de los alimentos (Liener, 1997). La tabla I.9. presenta algunos de los disímiles efectos que causan los metabolitos secundarios en los sistemas biológicos.
De las 1200 clases de compuestos secundarios que contienen las plantas (Kumar, 1992), por su elevada diversidad, no todos se encuentran bien estudiados; algunos grupos como los triterpenoides, las cumarinas, las anticarbohidratasas, las tioxalidonas, los tiocianatos e isotiocianatos, las antibiotinas, y las sustancias que aumentan de forma particular las pérdidas catabólicas, han recibido tratamientos más discretos en cuanto a su repercusión en el campo de la fisiología y la nutrición (Mitjavila, 1990). No obstante, existen algunas funciones químicas como los glucósidos cianogénicos y alcaloides (Sotelo, Contrera y Flores, 1995; Sotelo, Sousa y Sánchez, 1995), así como los fitohemoaglutinantes (Le Guen y Birk, 1993; Goodbole, Krishna y Brata, 1994), que se han investigado con mayor sistematicidad. 1.2.6. Metabolitos secundarios presentes en el género Morus. Actividad biológica
El reino vegetal es un reservorio extraordinario de metabolitos secundarios; sin embargo, solo un pequeño porcentaje de las especies estimadas en el globo terráqueo se han investigado fitoquímicamente (de 4 a 5 x 105), y cantidades menores han sido evaluadas en cuanto a su repercusión nutricional (Hostettmann y Wolfender, 1997).
El género Morus constituye una fuente importante de compuestos químicos con elevada actividad farmacológica (Peiyi y Xiusi, 1995; Kim, Park y Lee, 1998); no obstante, solo se conocen algunos metabolitos secundarios, los cuales han sido principalmente estudiados por líneas de investigación afines con la sericultura (Won-Chu y Yeou, 1997; Lee y Kim, 1997 Lee, Kim, Lee, Lee y Lee, 1999), y por la medicina tradicional china (Mingle, 1999).
Resultan asombrosos los reportes que provienen de la zona asiática que describen la elevada efectividad de los extractos de morera, en los tratamientos de innumerables aflecciones (Yun y Lee, 1995 a,b; Yun, Lee, Park y Kim, 1995; Kim, Lee, Kim, Kim y Kim; 1998). Independientemente de los estudios realizados a nivel de laboratorio para evaluar su potencial farmacológico, otras investigaciones desde el punto de vista alimentario se han llevado a cabo para conocer las propiedades curativas del té en la eliminación de metales pesados en el organismo humano (Weicheng, 1999; Kim, Lee, Kim, Lee y Bang, 1998).
Las tres propiedades hasta el momento más sobresalientes de M. alba en el campo médico, radican en la particularidad que presentan sus soluciones para contrarrestar la hipertensión arterial, la diabetes (Lemus, García, Del Villar y Knop, 1999) y los elevados niveles de colesterol (Ho-Zoo y Won-Chu, 2001). De estas tres características la más estudiada es la propiedad de planta antidiabética (Xiao, Xiangrong y Weiping, 1999; Yong y Jingzhi, 1999; Youye, y Fengging, 2000), donde los estudios han estado encaminados a la dilucidación de los mecanismos bioquímicos que lo propician y los compuestos causantes de esta acción (Lee, Cheng, Kim, Ruy y Lee, 1998; Kim, Ryu, Lee, Ku, Lee y Lee, 1999; Kim, Gao, Lee, Ryu, Lee y Kim, 1999).
Según el resumen realizado por Duke (2001), las hojas de M. alba contienen constituyentes volátiles tales como alcoholes (n-butanol y β-γ-hexenol), aldehídos (Metil-etil-acetaldehído, n-butil-aldehído, isobutil-aldehído, valeraldehído, hexaldehído, α-β-hexenal), cetonas alifáticas (Acetona, Metil-etil-cetona, Metil-hexil-cetona), butil-amina, y ácidos grasos volátiles (Acético, Propiónico y Butírico). Además contiene Malato de calcio, ácido succínico y tartárico, xantofilas, carotenoides, fitatos (forman el 18 % del Pt), isoflavonoides (Quercetina 3-glucósido), bases nitrogenadas (Adenina, Colina y Trigonellina), atrayentes del gusano de seda (Citral, Acetato de linalilo, Linalol, Acetato de terpenilo, Hexenol) y terpenoides, los cuales controlan las mordidas que realizan los gusanos a las hojas. La madera contiene Morina (0,3-0,4 %MS), Dihidromorina, Dihidrokaenferol, 2,4,4´,6´-Tetra- hidroxi- benzofenona, Maclurina y 2 %MS del estilbeno Hidroxi-resveratrol.
Ho-Zoo y Won-Chu (2001) afirman que las propiedades antioncogénicas y antisenecentes son atribuidas a los pigmentos (3%), neocarotenos, Isoquercetrina y Iuetina. La actividad diurética y la reacción antialérgica es causada por los flavonoides Rutina, Quercetina, Astragalina, Quercetina 3,7-diglucósido y Quercetina 3-glucósido, la actividad hipertensiva se debe en parte a la presencia de Ácido γ- amino butírico (GABA 0,62-1,11 %MS), la acción antibacteriana es causada por las moléculas de Kuwano, Mulberofurano, Moracenina, Moracina, Dimoracina, Chalcomoracina y Sanggenona. La Umbeliferona presenta propiedades antiflogísticas; los flavonoides pueden prevenir la alopecia, la arteriosclerosis e inhibir la obesidad; los fitosteroles contienen la asimilación del Colesterol en el intestino delgado; y los alcaloides Deoxinojimicina (peso molecular: 163,17 g/mol), N- Metil- Deoxinojimicina; así como la Amilosa (1,0 %MS), disminuyen los contenidos de CHS en el torrente sanguíneo.
Hornghuey, Jehjeng, Hsiencheng, Jinpyang y Chunnan (1999) aislaron de M. australis un nuevo compuesto denominado Australona B, el cual en plasma humano mostró una fuerte inhibición de agregación inducida por la Adrenalina, además de presentar efecto inhibidor de los aniones superóxidos en formación en ratas estimuladas

Capítulo I-Revisión bibliográfica
18
químicamente. El extracto de diclorometano-metanol obtenido de M. alba muestra actividad hipoglicémica como producto de un triterpeno y dos galactolípidos; uno de los últimos produjo el 16 % de la actividad (Chul-Young, Si-Hyun, Gi-Wha, Chun-Sik y Bakk-Wang, 2000).
Por su importancia farmacológica, algunos flavonoides de M. alba con importante actividad, han sido comparados después de su propagación in vitro (El-Kazzaz y Nazif; 2000) e in vivo (Ferrari, Monacelli y Messana, 1999). Los flavonoides (Quercetina-3-β-D-glucopiranosido y Quercetina-3,7-di-O-D-glucopiranósido) mostraron un significante efecto inhibidor en el crecimiento de la línea celular de la leucemia humana y una importante captura de radicales libres (Sun-Yeou, Jian-Jun y Hee-Kyoung, 2000).
Los extractos de morera presentan actividad antioxidante (Bokang,Huiyin y Gowchin, 1999), así como una elevada acción contra los microbios (Sung-Suk, Hak-Ju y DonHa, 2000; Kim, Gao et al., 1999). El Oxiresveratrol presenta un efecto relajante en la contracción de los músculos blandos de la tráquea en cuyes (Feng, Kano y Xinsheng, 1997), además de ser un potente inhibidor de la actividad dopa-oxidasa de la Tirosinasa en hongos (Nam-Ho y Shi-Yong, 1998).
Sun-Yeou, Kang-Sun, Won-Chu, Jun-Ok, Hoy-Seon y Kang-Ro (1999) observaron un decrecimiento en los niveles de glucosa en el plasma sanguíneo de ratas, en tratamiento anaeróbico, e incremento en los contenidos del alcaloide Deoxijirimicina.
La Fagomina, un seudo azúcar aislado de las hojas de morera, potencia la secreción de insulina (Sun-Yeou et al., 1999; Taniguchi, Asano, Tomino y Miwa, 1998), y las pectinas y hemicelulosas aisladas de M. alba presentaron una actividad hipoglicémica más fuerte que las aisladas a partir de M. nigra y M. rubra (Sanavova y Rakhimov, 1997). Por otra parte, el extracto de morera reduce la fracción de lípidos en la sangre (Doi, Kojima, Harada, Horiguchi y Fujimoto, 2000).
Un novedoso tipo de aducto “Diels-Alder” fue obtenido a partir de M. multicaulis por Ferrari, Monache, Suárez y Compagnone (2000).
A partir de M. alba ha sido aislado un terpenoide de tipo fitoalexina, el cual aparece cuando las hojas se contaminan con Phloeospora maculans; uno de los agentes causales de las manchas foliares (Kyunghee, Matsuyama y Kim, 1998). Además se han obtenido dos lectinas (MLL 1 y MLL 2); glicoproteínas con contenidos de azúcar de 8,8 y 40%, respectivamente. Estas se caracterizan por presentar el mismo peso molecular (51 KDa); con subunidades de 16,5 KDa, y una elevada especificidad por los Ácido N-glicolil-neuramínico y siálico (Ratanapo, Ngamjunyaporn y Chulavatnatol, 1999).

Capítulo II-Metodología experimental
19
2. Metodología experimental 2.1. Área experimental
El experimento se llevó a cabo en la Estación Experimental de Pastos y Forrajes “Indio Hatuey”, situada a los 22º 48’7’’ de latitud Norte y 81º 2’ de longitud Oeste, a 19 msnm, en el municipio de Perico, provincia de Matanzas, Cuba. 2.2. Períodos de evaluación
Los muestreos se realizaron en dos períodos correspondientes al año 2002, enmarcados entre los meses de enero, febrero, marzo, abril y mayo como PPLL; y mayo, junio, julio, agosto y septiembre como PLL., atendiendo al régimen pluviométrico que tuvieron las plantas desde el corte hasta su muestreo. 2.3. Características climáticas de los períodos evaluados
Los períodos evaluados presentaron características climáticas contrastantes. La tabla II.1. muestra los principales indicadores del clima en ambas etapas.
Tabla II.1. Principales indicadores del clima en los períodos evaluados.
Mes Temperatura máxima (ºC)
Temperatura mínima (ºC)
Temperatura media (ºC)
Precipitación acumulada (mm)
Enero 28,4 14,3 20,6 18,2
Febrero 28,7 14,8 21,3 44,7
Marzo 30,7 16,5 22,9 26,3
Abril 32,0 17,0 24,0 10,2
Mayo 31,3 20,3 25,8 249,2
Junio 32,1 22,3 26,1 309,4
Julio 33,9 21,9 26,8 205,5
Agosto 33,3 21,9 26,2 241,5
Septiembre 31,8 22,4 26,0 369,7 2.4. Características del suelo
El suelo donde se llevó a cabo la investigación presentó topografía plana; este se clasificó como Ferralítico Rojo lixiviado, según Hernández y col. (1999). La tabla II.2. muestra la composición inicial del suelo entre los 0-20 cm de profundidad, en las parcelas evaluadas para los diferentes tratamientos.
Tabla II.2. Composición química del suelo en las parcelas de cada tratamiento.
Frecuencia 60 días 90 días 120 días Fertilización 100* 300* 500* 100* 300* 500* 100* 300* 500*
PH (KCl) 6,3 6,4 6,4 5,9 6,3 6,4 6,2 6,3 6,6 P2O5 (mg/100g) 19,6 29,4 25,5 8,2 21,2 36,9 18,4 25,5 36,5 K2O (mg/100g) 16,8 19,3 18,5 10,8 20,8 14,5 11,8 14,6 18,9 Ca (cmol/kg) 19,8 17,6 20,6 19,4 20,6 22,2 16,6 20,5 20,0 Mg (cmol/kg) 3,3 2,8 3,2 1,1 2,3 2,2 2,9 4,3 4,2 K (cmol/kg) 0,54 0,48 0,60 0,35 0,65 0,47 0,30 0,42 0,63 Na (cmol/kg) 0,10 n.d 0,08 0,1 0,1 0,08 0,05 0,03 0,15 MO % 3,49 4,03 3,67 3,84 3,31 3,84 3,85 4,02 3,93
*N kg/ha/año (n.d) no detectado Los valores de cada indicador representan las medias obtenidas a partir de los análisis en las parcelas de cada tratamiento.

Capítulo II-Metodología experimental
20
2.5. Diseño experimental y tratamientos
En esta investigación se utilizó un diseño de bloques al azar con arreglo factorial 4 x 3 x 3 y tres repeticiones para las dos épocas, lo que hizo un total de 36 tratamientos/bloque.
Los factores estudiados fueron: Fertilización orgánica equivalente a: 100, 300 y 500 kg N/ha/año, además de los tratamientos donde no
se fertilizó (parcelas control) Frecuencia de corte (60, 90 y 120 días) Variedad (Cubana, Indonesia, Tigriada y Acorazonada) Época (PPLL y PLL) Parte de la fracción comestible (hojas y tallos tiernos)
La combinación de todos los factores generó 72 casos.
2.6. Unidad experimental y manejo agronómico
Las mediciones se realizaron en el tercer año de evaluación agronómica de una plantación de morera con 4 años de edad y una densidad de 25 000 plantas/ha.
El experimento se llevó a cabo en un área experimental que abarcó 108 parcelas de 7 x 3 m sin separación entre ellas, además de 12 parcelas control (anexos 1 y 2).
Cada parcela estuvo integrada por 64 plantas, separadas a 0,4 m y 1 m entre los surcos; los que se orientaron de Este a Oeste.
El corte de cada planta se realizó de manera manual con tijera de poda, a la altura fija de 0,5 m sobre el nivel del suelo.
La fertilización orgánica se aplicó directamente en el tronco de cada planta y el control de malezas se realizó de forma manual, ambos después de cada corte. 2.6.1. Distribución de los cortes en los períodos de evaluación
Los cortes correspondientes a las frecuencias de 60, 90 y 120 días para cada período muestreado se realizaron de forma tal que su distribución fuera equitativa en cada etapa, para lograr que se reflejara de forma directa el efecto de las condiciones ambientales en todo el proceso de evaluación .
La tabla II.3 muestra la distribución de la poda en los distintos meses del año 2002.
Tabla II.3. Distribución de los cortes en los meses del año.
Mes 60 días 90 días 120 días
Enero Ca Ca
Febrero Ca
Marzo Cm
Abril
Mayo Cm Ca Cm Ca
Junio
Julio Ca
Agosto Cm
Septiembre Cm CmCa corte anterior al muestreo Cm corte muestreado

Capítulo II-Metodología experimental
21
2.7. Composición química del fertilizante orgánico
Como fuente de fertilización nitrogenada se empleó gallinaza. La composición química del fertilizante orgánico utilizado en cada evaluación se muestra en la tabla II.4.
Tabla II.4. Composición química del fertilizante orgánico.
Período N(%MS) P(%MS) K(%MS) Ca(%MS)
PPLL 2,17 0,23 1,33 6,93
PLL 1,55 0,50 1,72 9,01 2.8. Análisis cualitativo 2.8.1. Prueba exploratoria
Con el objetivo de conocer si los grupos de metabolitos secundarios se encontraban presente de manera común en las dos partes de la planta, se realizó un análisis cualitativo inicial de manera independiente al conjunto hojas-pecíolo y a los tallos tiernos. Comprobándose la presencia de los mismos grupos en ambas partes. 2.8.2. Procedimiento de muestreo
El material vegetal formado por la fracción comestible de M. alba (hojas-pecíolos-tallos tiernos) fue recolectado de forma manual a partir de 10 plantas por parcela seleccionadas al azar, luego de ser eliminado el efecto borde de las unidades experimentales.
Las muestras provenientes de cada réplica se llevaron de forma inmediata al laboratorio, se pesaron 25 gramos, fueron trituradas hasta tamaño homogéneo y finalmente maceradas con 250 ml de etanol al 98 % durante 48 horas. 2.8.3. Pesquisaje fitoquímico
A los extractos alcohólicos provenientes de los diferentes tratamientos se les aplicó el tamizaje fitoquímico propuesto por Rondina y Cassio, descrito por Alfonso, Fernández, González y Avilés (2000); los análisis se realizaron por triplicado.
Las pruebas cualitativas para la detección de cumarinas se llevaron a cabo a partir de modificaciones realizadas al extracto crudo. La solución alcohólica fue tratada previamente con Pb(AcO)2 al 4 %, el que contenía 0,5 % de AcOH con el objetivo de eliminar las clorofilas, para después centrifugar por 10 minutos a 2 500 repeticiones por minuto, filtrar con papel y realizar una extracción con dos porciones de 20 ml de CHCl3 al filtrado. Del extracto clorofórmico se evaporaron 2 ml de la solución en una placa de porcelana y se le adicionó 1 ml del reactivo de Baljet. 2.8.3.1. Metabolitos investigados
Se investigaron un total de 15 grupos de metabolitos, estos fueron: los fenoles, los taninos, los grupos α-aminos, los triterpenos y los esteroides, las fitoquinonas, los flavonoides, las proantocianidinas y las catequinas, los carbohidratos reductores solubles, los cardenólidos, las saponinas, las cumarinas, los alcaloides y los cianógenos. 2.8.3.2. Criterios tomados en la realización de las detecciones semicuantitativas
Para la descripción de los ensayos se utilizó el sistema de cruces para especificar la presencia o ausencia de los metabolitos en los tratamientos.
En todos los análisis se siguieron los criterios que se muestran en la tabla II.5. En el caso del ensayo empleado para la detección de saponinas se partió del criterio que se muestra en la
tabla II.6.

Capítulo II-Metodología experimental
22
Tabla II.5. Criterios seguidos en las detecciones cualitativas en el pesquizaje fito-químico
Criterio Nomenclatura
Presencia cuantiosa +++
Presencia notable ++
Presencia leve +
Ausencia -
Tabla II.6. Criterio tomado en dependencia de la altura de la espuma, en el ensayo de “índice de espuma”.
Criterio Altura de la espuma (mm)
Contenido abundante >14
Contenido moderado 10-14
Contenido bajo <10 2.8.3.3. Control de reactivos
Previamente para el control de los reactivos se utilizaron soluciones de compuestos patrones en cada ensayo, con el fin de comprobar el estado de los reactivos a utilizar (tabla II.7.).
Tabla II.7. Soluciones utilizadas para el control de los reactivos.
Metabolito Ensayo Soluciones Control
Fenoles FeCl3 1-10 % Fenol 1%
Taninos Gelatina 1% Ácido tánico 1%
Grupos α-aminos Ninhidrina 0,2% L- Ácido aspártico 1%
Triterpenos/Esteroides Lieberman-Burchard Colesterol 2%
Fitoquinonas Borntrager Quinol 2%
Flavonoides Shinoda Quercetina 2%
Proantocianidinas/Catequinas Roseheim D(+) Catequina
Carbohidratos reductores Fehling Benedict
D(+)Glucosa 5%
Cardenólidos Kedde Digitalis 2% (masa/volumen)
Saponinas Prueba de espuma *
Cianógenos Guignard *
Cumarinas Baljet Cumarina 2%
Alcaloides Wagner, Hager y Dragendorff Gramina 2% Efedrina 2%
* No utilizadas

Capítulo II-Metodología experimental
23
2.9. Análisis cuantitativo 2.9.1. Procedimiento de muestreo, secado y conservación del material
El material vegetal, dividido en el conjunto hoja-pecíolo y tallos tiernos, fue tomado a partir de las muestras iniciales recolectadas para el análisis cualitativo. Estas fueron secadas a temperatura ambiente, en un local ventilado y oscuro por espacio de 12 días. Posteriormente fueron molinadas hasta un tamaño de partícula de 1 mm, y se almacenaron en frascos ámbar hasta el momento del análisis. 2.9.2. Instrumentación y mediciones analíticas
Las lecturas en el espectro visible fueron realizadas en un fotocolorímetro ERMA con filtros cambiables. Las mediciones en el espectro ultravioleta se llevaron a cabo en un espectrofotómetro U/V ULTROSPEC-
2000 de doble haz, con cubetas de cuarzo. La cuantificación de FT se realizó mediante el método de Folin-Dennis y los flavonoides por la metodología
desarrollada por Kostennikova; ambos descritos por Gutiérrez et al. (2000) La cuantificación de cumarinas se realizó mediante el método de Quirós descrito por Mochiutti (1995). La determinación de CHS se llevó a cabo mediante el método de la Antrona descrito por Lezcano y
González (2000). La determinación de ET se desarrolló mediante el método de Lieberman-Burchard, descrito por Galindo et
al. (1989). Las determinaciones de PB, PV, N-FND y N-FAD se llevaron a cabo mediante la metodología propuesta por
la AOAC (1990). Los cálculos de los aportes de los compuestos nitrogenados por unidad de área, fueron realizados a partir
de los datos de biomasa, correspondientes a los estudios agronómicos desarrollados de forma paralela a esta investigación. 2.9.2.1. Soluciones patrones
Los resultados del análisis espectrofotométrico, fueron expresados en las concentraciones de patrones específicos para cada grupo de metabolitos, los cuales coincidieron en buena medida con las estructuras reportadas en la literatura.
Para la obtención de las curvas de calibración de cada grupo, se prepararon soluciones madres a partir de los patrones siguientes (tabla II.8).
Tabla II.8. Patrones utilizados en el análisis espectrofotométrico.
Grupos de metabolitos Patrón (2000 ppm) Fenoles Ácido tánico Flavonoides Quercetina Cumarinas 4-metil Umbeliferona CHS D(+) Glucosa Esteroides Colesterol
2.10. Métodos estadísticos
Para el procesamiento de los datos se utilizó un análisis factorial, para lo cual se empleó la opción GLM (General Lineal Model) correspondiente al paquete estadístico SPSS versión 10.0. Fue usada la dócima de comparación múltiple de SNK (Student-Newman-Keuls) y las medias fueron comparadas según P<0,05.
Los datos obtenidos a partir de los ensayos cualitativo, se llevaron a una escala numérica (1-ausencia y 2, 3, 4-presencia). El análisis de conglomerados se realizó mediante el método de Ward, utilizando la distancia euclidiana como criterio de diferencia entre los casos.

Capítulo III-Resultados y Discusión
24
3. Resultados y discusión 3.1. Experimento 1. “Detección y cuantificación de metabolitos secundarios en cuatro variedades de M.
alba con fertilización y corte”. 3.1.1. Consideraciones sobre el nivel cero de fertilización
La biomasa proveniente de M. alba, que serviría como material vegetal de partida para los análisis de los experimentos que conforman esta tesis, fue nula para los tratamientos donde no se fertilizó, como producto de que ninguna de las variedades aportó cantidades considerables de hojas ni de tallos tiernos para poder ser incluidos en los resultados de esta investigación; este comportamiento fue común para los dos períodos evaluados, así como para las tres frecuencias de defoliación empleadas en este caso (60, 90 y 120 días).
La necesidad que tiene la morera de ser fertilizada, es un consenso generalizado al cual han arribado los principales autores que investigan sobre este tópico en diferentes partes del mundo, como Costa Rica (Benavides, 2002), Japón (Machii, 2001) y Cuba (Martín, et al., 2002). 3.1.2. Análisis cualitativo
Mediante el empleo del tamizaje fitoquímico de los 15 grupos de metabolitos en el conjunto hojas-pecíolos-tallos tiernos se detectaron los fenoles, los flavonoides, las cumarinas, los carbohidratos solubles, los esteroides, los alcaloides y las saponinas, estos aparecieron en todas las variedades, por lo que la presencia de los mismos grupos de compuestos es una de las evidencias de la marcada componente genética del metabolismo secundario en el género Morus (Ashok; Vincent y Nessler, 2000). 3.1.2.1. Fenoles
Las tablas III.1 y III.2 muestran el comportamiento de los compuestos fenólicos desde el punto de vista cualitativo mediante la utilización de FeCl3 al 1%. En ambas épocas, estos compuestos se detectaron de forma abundante.
Todos los ensayos se caracterizaron por presentar una coloración negra, la cual es una de las propiedades de los extractos que contienen una amplia diversidad de estructuras hidroxiladas (Sung-Suk et al., 2000).
En el PLL hubo una mayor variabilidad con respecto al PPLL, aunque fue poco marcada, también se observó una tendencia similar entre las variedades.
Tabla III.1. Comportamiento cualitativo de los fenoles en el PPLL.
60 días 90 días 120 días Variedad
100* 300* 500* 100* 300* 500* 100* 300* 500* Cubana +++ +++ +++ +++ +++ +++ +++ +++ +++
Indonesia +++ +++ +++ +++ +++ +++ +++ +++ +++ Tigriada +++ +++ +++ +++ +++ +++ +++ +++ +++
Acorazonada +++ +++ +++ +++ +++ +++ +++ +++ +++ *Kg N/ha/año
Tabla III.2. Comportamiento cualitativo de los fenoles en el PLL.
60 días 90 días 120 días Variedad 100* 300* 500* 100* 300* 500* 100* 300* 500*
Cubana +++ ++ ++ +++ +++ +++ +++ +++ +++ Indonesia +++ +++ ++ +++ +++ ++ +++ +++ ++ Tigriada +++ ++ +++ +++ +++ +++ +++ +++ +++
Acorazonada +++ ++ +++ +++ +++ +++ +++ +++ +++ *Kg N/ha/año
La presencia de fenoles en la especie ha sido reportada por Chunlian, Donsheng y Erjun (1999) y
Domínguez, Telles y Revilla (2001).

Capítulo III-Resultados y Discusión
25
Estos tipos de metabolitos se encuentran ampliamente distribuidos en el reino vegetal, formando parte de todas las plantas vasculares, y su presencia ha sido detectada en algunas de las principales plantas con interés forrajero en diferentes latitudes, como L. leucocephala (Pedraza, García y Pacheco, 1997), Calliandra calothyrsus (Salawu, Acamovic, Stewart y Maasdorp, 1997), Acacia cyanophylla (Ben Salem et al., 2000) y en Macroptilium atropurpureum y Lablab purpureus (Mupangwa, Acamovic, Topps, Ngongoni y Hamudikuwanda, 2000). 3.1.2.2. Flavonoides
Las tablas fitoquímicas III.3. y III.4. muestran el resultado correspondiente al análisis de los flavonoides. Las pruebas cualitativas mostraron una mayor variabilidad entre las variedades y las épocas.
Los ensayos oscilaron desde coloraciones rosadas tenues hasta tonos rojos intensos. Rangos de variabilidades en escalas numéricas han sido encontrados por Mengcheng, Zhishen y Xiangrui (1996) y Zhishen, Jianming y Mengsheng (1996).
Tabla III.3. Comportamiento cualitativo de los flavonoides en el PPLL.
60 días 90 días 120 días Variedad
100* 300* 500* 100* 300* 500* 100* 300* 500* Cubana +++ + + +++ +++ +++ +++ +++ +++
Indonesia +++ ++ + +++ ++ + + +++ +++ Tigriada + +++ + +++ +++ ++ +++ +++ +++
Acorazonada +++ ++ + +++ ++ ++ +++ +++ +++ *Kg N/ha/año
Tabla III.4. Comportamiento cualitativo de los flavonoides en el PLL.
60 días 90 días 120 días Variedad
100* 300* 500* 100* 300* 500* 100* 300* 500* Cubana + ++ ++ ++ ++ ++ +++ ++ ++
Indonesia ++ ++ ++ ++ ++ ++ +++ +++ ++ Tigriada ++ ++ ++ ++ ++ ++ +++ +++ ++
Acorazonada ++ ++ ++ ++ ++ ++ +++ +++ +++ *Kg N/ha/año
Ensayos positivos para el contenido de Rutina han sido efectuados por Yun y Lee (1995a) y detecciones
generales obtenidas por Chuanbu (2000). Detecciones similares se han realizadao en Gliricidia sepium, Albizia lebbeck y leguminosas rastreras
(Martínez, Hernández y Guevara, 1996). 3.1.2.3. Cumarinas Las cumarinas presentaron diferencias menos marcadas que en el caso de los flavonoides (tablas III.5. y III.6.). La coloración naranja desarrollada por el reactivo de Baljet fue común para todos los ensayos realizados; aunque con diversos grados de intensidad según la concentración inherente de cada tratamiento.
Tabla III.5. Comportamiento cualitativo de las cumarinas en el PPLL.
60 días 90 días 120 días Variedad 100* 300* 500* 100* 300* 500* 100* 300* 500*
Cubana +++ +++ +++ ++ ++ ++ +++ +++ +++ Indonesia + +++ +++ ++ ++ ++ +++ +++ +++ Tigriada +++ +++ +++ ++ ++ ++ +++ +++ +++
Acorazonada ++ +++ +++ ++ ++ ++ +++ +++ +++ * Kg N/ha/año

Capítulo III-Resultados y Discusión
26
Tabla III.6. Comportamiento cualitativo de las cumarinas en el PLL.
60 días 90 días 120 días Variedad 100* 300* 500* 100* 300* 500* 100* 300* 500*
Cubana ++ +++ +++ +++ +++ ++ +++ +++ +++ Indonesia ++ +++ ++ +++ +++ ++ +++ +++ +++ Tigriada ++ +++ ++ +++ +++ ++ +++ +++ +++
Acorazonada ++ +++ +++ +++ +++ +++ +++ +++ +++ * Kg N/ha/año
Las cumarinas en M. alba han sido detectadas por Domínguez et al. (2001), la Esculetina determinada por
Marles y Farnsworth (1995) y la Umbeliferona por Ho-Zoo y Won-Chu (2001). Este grupo de compuestos se ha encontrado en G. sepium (Ahn, Elliott y Norton, 1997) y en familia de especies templadas tales como Umbelíferas y Rutáceas (Berenbaum, 1991). 3.1.2.4. Carbohidratos reductores solubles Las tablas III.7. y III.8. muestran el comportamiento de los carbohidratos solubles en ambos períodos; estos intermediarios del metabolismo fueron detectados en forma cuantiosa de acuerdo con el contenido energético reportado en la especie y la elevada concentración de sacáridos en sus extractos alcohólicos (Fujun, Nakashima y Kimura, 1995; Jun, Rong y Hongzhang, 2000).
Tabla III.7. Comportamiento cualitativo de los carbohidratos en el PPLL.
60 días 90 días 120 días Variedad 100* 300* 500* 100* 300* 500* 100* 300* 500*
Cubana +++ +++ +++ +++ +++ +++ +++ +++ +++ Indonesia +++ +++ +++ +++ +++ ++ +++ +++ +++ Tigriada +++ +++ +++ +++ +++ +++ +++ +++ +++
Acorazonada +++ +++ +++ +++ +++ +++ +++ +++ +++ * Kg N/ha/año
Tabla III.8. Comportamiento cualitativo de los carbohidratos en el PLL.
60 días 90 días 120 días Variedad
100* 300* 500* 100* 300* 500* 100* 300* 500* Cubana + +++ +++ +++ +++ ++ +++ +++ +++
Indonesia +++ +++ + +++ +++ +++ +++ +++ +++ Tigriada +++ +++ +++ +++ +++ ++ +++ +++ +++
Acorazonada +++ +++ +++ +++ +++ ++ +++ +++ +++ * Kg N/ha/año
No se encontraron diferencias muy marcadas entre las épocas, las variedades y las frecuencias de
defoliación. 3.1.2.5. Esteroides El análisis cualitativo reveló una elevada similitud entre los tratamientos (tablas III.9. y III.10.). Los esteroides fueron detectados de manera abundante y el ensayo aplicado se caracterizó por una coloración azul verdosa intensa, lo que evidencia la presencia de varios esteroles en el tejido vegetal (Galindo et al., 1989). La presencia de β-Sitosterol y Estigmasterol ha sido señalada por Mengzhao (1989) y trazas de colesterina por Xiangrui y Hongsheng (2001).
Los resultados generales obtenidos para este grupo de metabolitos en la especie están acorde a lo reportado por Mengzhao (1989) en cuantificaciones realizadas en M. alba.

Capítulo III-Resultados y Discusión
27
Tabla III.9. Comportamiento cualitativo de los esteroides en el PPLL.
60 días 90 días 120 días Variedad 100* 300* 500* 100* 300* 500* 100* 300* 500*
Cubana +++ +++ +++ +++ +++ +++ +++ +++ +++ Indonesia +++ +++ +++ +++ +++ +++ +++ +++ +++ Tigriada +++ +++ +++ +++ +++ +++ +++ +++ +++
Acorazonada +++ +++ +++ +++ +++ +++ +++ +++ +++ * Kg N/ha/año
Tabla III.10. Comportamiento cualitativo de los esteroides en el PLL.
60 días 90 días 120 días Variedad
100* 300* 500* 100* 300* 500* 100* 300* 500* Cubana +++ +++ +++ +++ +++ ++ +++ +++ +++
Indonesia +++ +++ +++ +++ +++ +++ +++ +++ +++ Tigriada +++ +++ +++ +++ +++ +++ +++ +++ +++
Acorazonada +++ +++ +++ +++ +++ ++ +++ +++ +++ * Kg N/ha/año
3.1.2.6. Alcaloides
Los alcaloides fueron investigados mediante el empleo de tres reactivos de grupo. Las tablas III.11. y III.12. muestran algunas variaciones en el PLL cuando se compara con el PPLL.
Estos metabolitos conjuntamente con los flavonoides expresaron la mayor variabilidad en las pruebas de detección.
Tabla III.11. Comportamiento cualitativo de los alcaloides en el PPLL.
60 días 90 días 120 días Variedad 100* 300* 500* 100* 300* 500* 100* 300* 500*
Cubana +++ +++ +++ +++ ++ +++ +++ +++ +++ Indonesia +++ +++ +++ +++ +++ + +++ +++ +++ Tigriada +++ +++ +++ +++ +++ +++ +++ +++ +++
Acorazonada +++ +++ +++ +++ +++ +++ +++ +++ +++ * Kg N/ha/año
Tabla III.12. Comportamiento cualitativo de los alcaloides en el PLL.
60 días 90 días 120 días Variedad 100* 300* 500* 100* 300* 500* 100* 300* 500*
Cubana ++ ++ +++ + +++ +++ +++ +++ +++ Indonesia +++ +++ + ++ ++ +++ +++ +++ +++ Tigriada +++ +++ + + +++ +++ +++ +++ +++
Acorazonada ++ + + + ++ +++ +++ +++ +++ * Kg N/ha/año
La presencia de alcaloides y compuestos aminados en el tejido vegetal ha sido reportada por Ho-Zoo y Won-
Chu (2001). Los alcaloides también se han encontrado en árboles de uso forrajero, de forma general en plantas
dicotiledóneas (Ramos et al., 1998) y especialmente en leguminosas forrajeras del género Erythrina (Sotelo, Soto y Lucas, 1996). 3.1.2.7. Saponinas
Las saponinas se detectaron mediante la prueba de “índice de espuma”; todos los extractos ensayados mostraron alturas relativas de la espuma entre 10-14 mm, equivalente a contenidos moderados de estos metabolitos (Galindo et al., 1989).

Capítulo III-Resultados y Discusión
28
La prueba no resultó concluyente para poder aseverar la presencia de estos compuestos, ya que el principio del método consiste solamente en la disminución de la tensión superficial del medio, por lo que otros compuestos con propiedades estructurales similares en la planta, como mucílagos y glicósidos esteroidales, pudieron crear falsos positivos al respecto.
Detecciones positivas en M. alba han sido realizadas por Domínguez et al. (2001) y negativas por Maldonado, Grande, Aranda y Pérez-Gil (2000). 3.1.2.8. Metabolitos no detectados
El resto de los compuestos investigados mostraron resultados negativos en todas los tratamientos; el conjunto estuvo formado por: grupos α-aminos, taninos que precipitan proteínas, proantocianidinas/catequinas, cardenólidos, fitoquinonas y cianógenos.
La no presencia de grupos α-aminos, proantocianidinas/catequinas y fitoquinonas también ha sido reportada por Domínguez et al. (2000).
La ausencia de taninos que precipitan proteínas, corroborada en este experimento y que apoyan lo expresado por García et al. (2002) con la utilización de gelatina como proteína para inducir la precipitación, coincide con los resultados de pruebas realizadas por Makkar, Singh y Negi (1989) y Makkar y Becker (1998) al emplear Albúmina de Suero Bovino (BSA) como proteína precipitante.
Aunque las pruebas desarrolladas demuestran la inexistencia evidente de taninos condensados mediante los ensayos con el reactivo de Roseheim, no se puede expresar con certeza absoluta que no exista ningún tipo de tanoide en la especie, en primer lugar por lo controvertida que resulta la definición de estos metabolitos, y teniendo en cuenta también que en el ensayo de precipitación solo resultan positivos los taninos que presenten un peso molecular considerable, capaces de unirse a la proteína e insolubilizarse (Leinmueller, Steingass y Menke, 1993).
De todos los metabolitos detectados, los fenoles, los esteroides y los carbohidratos solubles presentaron menor variabilidad con los factores estudiados; en cambio, las cumarinas, los flavonoides y los alcaloides mostraron mayores diferencias entre los tratamientos. 3.1.2.9. Análisis de agrupación en las pruebas cualitativas
Para determinar dentro de los factores en estudio la tendencia de agrupación de los tratamientos, se empleó el análisis de conglomerados mediante el método de Ward (Rossi, Pereira y González 2000; Navarro, 2002).
Las tablas III. 13. y III.14. muestran la agrupación en el PPLL y PLL. En ambas épocas se observó un marcado efecto de la frecuencia de corte en comparación con los factores variedad y fertilización; la importancia de la edad de rebrote y su repercusión en el metabolismo secundario ha sido señalada por Brooks y Owen-Smith (1994).
Tabla III.13. Agrupación en el análisis cualitativo en el PPLL.
Conglomerados PPLL
1 2 3 Frec. Fert. C I T A C I T A C I T A 100 X X X X 300 X X X X 60 500 X X X X 100 X X X X 300 X X X X 90 500 X X X X 100 X X X X 300 X X X X 120 500 X X X X
El agrupamiento por edad de rebrote se hizo más marcado en el PPLL, excepto para la frecuencia de 60 días
en comparación con el PLL. En el PPLL el porcentaje de miembros de cada conglomerado, agrupados en la frecuencia más poblada fue de 57, 78 y 85% para los clusters 1, 2 y 3 respectivamente; en cambio, para el PLL fue de 67, 62 y 80% para los clusters formados.

Capítulo III-Resultados y Discusión
29
Tabla III.14. Agrupación en el análisis cualitativo en el PLL.
Conglomerados PLL 1 2 3 Frec. Fert. C I T A C I T A C I T A
100 X X X X 300 X X X X 60 500 X X X X 100 X X X X 300 X X X X 90 500 X X X X 100 X X X X 300 X X X X 120 500 X X X X
Basado en el análisis de las distancias euclidianas (anexo 3 y 4) se pudo comprobar que en el PPLL la
frecuencia de 60 días mostró particularidades con respecto a las de 90 y 120, este resultado coinciden con las diferencias encontradas por Mochiutti (1995) en las frecuencias de defoliación mayores (50 días), en comparación con las más bajas (75 y 100 días) para algunos metabolitos secundarios en G. sepium. En el PLL la frecuencia que mostró características diferenciadas al resto fue la de 120 días; la tendencia a la diferenciación de las frecuencias mayores, así como la menor repercusión fisiológica de los metabolitos secundarios en la nutrición animal para estas edades de rebrote, ha sido informada por Steward, Allison y Simons (1996).
Los dendogramas obtenidos mediante estos análisis (anexo 5 y 6) denotan que la frecuencia de corte que mostró un comportamiento intermedio en ambos períodos fue la de 90 días, por lo que las determinaciones cuantitativas de los metabolitos secundarios se basaron en dicha frecuencia; esta edad de rebrote muestra desde el punto de vista agronómico una mayor producción de biomasa total, comestible y de hojas (Martín et al., 2002). 3.1.3. Análisis cuantitativo 3.1.3.1. Fenoles totales -Hojas
El contenido de FT cuantificados en este experimento osciló entre (2,18-3,20) ±0,10 %MS en el PPLL, valores numéricos que concuerdan con los obtenidos en M. alba por Maldonado et al. (2000); estas plantas de morera fueron cultivadas en el trópico a 9 msnm y con una temperatura media anual de 25,9 0C, condiciones climáticas similares a las de este experimento.
El análisis estadístico mostró una interacción significativa entre los factores Variedad y Fertilización (fig. 2); la respuesta de los metabolitos fenólicos con variables tales como defoliación, estrés hídrico y fertilización ha sido demostrada por Anuraza, Duraza, Hill y Lovett (1993).
La tendencia de este efecto de grupo se caracterizó por presentar una disminución en la concentración de los FT con el incremento de la dosis de fertilización nitrogenada, para las variedades T, I y A. La disminución de los FT y los taninos condensados con el aumento de la fertilización ha sido probada sobre la base de la teoría del balance carbono-nitrógeno (Caygil y Mueller-Harvey, 1999) por Ruohomaki, Chapin, Haukioja, Neuvonen y Suomeka (1996), en estudios realizados en leguminosas templadas del género Lotus.
La variedad C tuvo un comportamiento diferente en comparación con el resto, además de caracterizarse por una mayor concentración de FT para los niveles de 300 y 500 kg N/ha/año. Para el nivel de 300 kg N/ha/año el contenido de FT exhibió las menores variaciones entre las variedades, que presentaron una tendencia similar.
Las concentraciones de FT obtenidas en las hojas, en las condiciones edafoclimáticas de este experimento, son superiores a la informada por Singh y Makkar (2002) en hojas de morera cultivadas en la India (1,80 %MS), y a la obtenida por Datta, Sarkar, Rama Mohan Rao y Singhui (2002), quienes refieren una concentración relativa de 0,85 %MS en la misma zona.
El conjunto de resultados recopilados por diferentes autores, en la cuantificación de FT por un método común de análisis en M. alba, denota variaciones probablemente ocasionadas por las diferentes variedades estudiadas, el estado fisiológico y de conservación del material de análisis, los diferentes climas, tipos de suelo y ubicaciones geográficas de cada experimento.

Capítulo III-Resultados y Discusión
30
Variedad x Fertilización(P<0,05)
Fertilización (N kg/ha/año)
500300100
FT (%
MS)
3,4
3,2
3,0
2,8
2,6
2,4
2,2
2,0
VARIEDAD
A
C
I
T
Fig. 2. Comportamiento de los FT en las hojas de cuatro variedades de M. alba en el PPLL.
Al compararla con otras especies forrajeras de interés (tabla III.15), los resultados sitúan a la morera como
una planta cuyos contenidos de FT son comparables con los de algunas de las principales especies de leguminosas arbóreas y rastreras más usadas en Cuba y en el trópico para la producción animal, y en algunos casos superiores a estas últimas (Pinto, Ramírez-Avilés, Ku-Vera, Hernández, Sánchez y Saucedo, 2000), aunque sus concentraciones son inferiores a la exhibida por la mayoría de las especies del género Acacia, típicas de climas áridos, con niveles de metabolitos secundarios muy elevados (Abdulrazak, Fujihara, Ondiek y Ørskov, 2000).
Tabla III.15. Contenido de FT determinados mediante la reducción del ácido fosfomolíbdico en las hojas de especies forrajeras.
Especie FT(%MS) Referencia
Acacia nilotica 16,20 A. tortilis 14,10 A. mellifera 10,40
Abdulrazak et al. (2000)
Bauhinia ungulata 4,20 A. milleriana 3,50
Pinto et al. (2000)
A. lebbeck 3,41 Maldonado et al. (2000) G. sepium 2,07 Martínez et al. (2000)
M. alba 1,26-3,26 (PLL) Arachis pintoi 2,40 Centrosema pubescens 1,70 Cratilia floribunda 1,60
Valerio (1994)
El contenido de FT en el PLL osciló entre (1,26-3,26) ±0,22 %MS; en esta época la interacción Variedad x
Fertilización fue significativa (fig. 3) y estuvo determinada por un comportamiento similar de las variedades A, C y T; en comparación con la variedad I, esta última se comportó de manera similar al PPLL.
Variedad xFertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
FT (%
MS)
3,5
3,0
2,5
2,0
1,5
1,0
VARIEDAD
A
C
I
T
Fig. 3. Comportamiento de los FT en las hojas de cuatro variedades de M. alba en el PLL.

Capítulo III-Resultados y Discusión
31
-Tallos tiernos
En el PPLL los contenidos de FT se encontraron en el rango de (0,77-2,50) ±0,14 %MS, con una tendencia no homogénea, aunque casi estable y cercana al 2 % entre las variedades (fig. 4).
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
FT (%
MS)
3,0
2,5
2,0
1,5
1,0
,5
VARIEDAD
A
C
I
T
Fig. 4. Comportamiento de los FT en los tallos tiernos de cuatro variedades de M. alba en el
PPLL.
En el caso del nivel intermedio de fertilización se observó la menor variabilidad dentro de las variedades, igualmente que para las hojas en el mismo período.
En el PLL el rango de concentraciones obtenidas fue más estrecho (0,69-1,43) ±0,08 %MS en comparación con el PPLL (fig. 5); la variedad T tuvo un comportamiento totalmente atípico al resto de las variedades.
En sentido general, en los tallos tiernos se observó una respuesta baja de los FT con la fertilización, con poca variabilidad en la dosis intermedia de N; al igual que en las hojas. Pocas afectaciones en el contenido de los FT en plantas de interés agrícola han sido encontradas por Caygil y Mueller-Harvey (1999).
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
FT (%
MS)
1,6
1,4
1,2
1,0
,8
,6
,4
VARIEDAD
A
C
I
T
Fig. 5. Comportamiento de los FT en los tallos tiernos de cuatro variedades de M. alba en el
PLL.
En las hojas, las concentraciones de FT entre épocas resultaron ser similares (2,18-3,20) ±0,10 %MS vs (1,26-3,26) ±0,22 %MS, aunque se observó una mayor variabilidad en el PLL, lo cual quedó evidenciado por el mayor valor del ES de la interacción. Estos resultados corroboran las conclusiones obtenidas sobre el efecto de la época, mediante los sistemas de agrupación en el análisis cualitativo.
Por su parte, los tallos tiernos en el PPLL presentaron una mayor concentración de FT que los correspondientes al PLL, aunque con una mayor dispersión en los valores obtenidos.
La mayor respuesta con la fertilización, descrita por las interacciones Variedad x Fertilización en las hojas para ambos períodos, evidencian la elevada actividad metabólica de los compuestos fenólicos en esta parte de la planta (Raskin, 1992).
Por otra parte, en algunos casos se observaron particularidades entre el comportamiento de las variedades con la fertilización. Aunque las diferencias numéricas fueron discretas, las tendencias se diferenciaron en al menos una variedad.
De todas las variables estudiadas, las diferencias encontradas entre las hojas y los tallos tiernos resultaron ser las más determinantes (fig 6). La combinación de los factores Fertilización y Variedad creó la mayor cantidad

Capítulo III-Resultados y Discusión
32
de elementos diferenciados en el comportamiento de las variedades. El factor época también produjo variaciones en las concentraciones. Fig. 6. Comportamiento de los FT con los factores parte de la fracción comestible y época del año empleando la
media Grand. 3.1.3.2. Flavonoides -Hojas El contenido de flavonoides en las hojas no mostró diferencias significativas con los niveles crecientes de nitrógeno aplicado al suelo para ninguno de los dos períodos evaluados, con valores muy similares entre épocas, cercanos a 1,60 %MS (fig. 7).
0
0,5
1
1,5
2
Flav (%MS)
100 300 500
N kg/ha/año
PPLLPLL
Fig. 7. Comportamiento de los flavonoides con la fertilización en las hojas de M. alba
La estabilidad en las concentraciones de las diferentes fracciones fenólicas ante factores externos, ha sido
señalada por Caygil y Mueller-Harvey (1999). En este sentido Zhishen et al. (1996) informan la concentración de algunos compuestos específicos dentro del contenido de flavonoides totales, como Rutina (0,34 %MS) y Quercetina (0,11 mg/g), además de las cuantificaciones particularmente realizadas para Rutósidos (0,44 %MS) (Chunlian et al., 1999).
Las variedades en estudio tampoco presentaron diferencias significativas entre ellas en ambos períodos (fig. 8).
Estos contenidos de flavonoides son superiores a los informados por Rusong, Jinyi y Yiping (2000) en la variedad “Lungiao” (1,17 %MS) e inferiores a los de la variedad “Tong Xiang Ping” (2,66 %MS).
Las interacciones Variedad x Fertilización en las hojas no fueron significativas; esto evidencia la poca diferenciación en el contenido de estos metabolitos con la fertilización para las variedades, al ser comparados con el comportamiento de los FT en las hojas, que sí presentaron interacciones significativas.
Variaciones entre los contenidos de FT y diferentes grupos de compuestos de esta familia en una misma parte de la planta, han sido descritas por Lindroth, Roth, Krueger, Volin y Koss (1997) en la explicación de fenómenos biológicos asociados a períodos de estrés.
00,5
11,5
22,5
3
FT (%MS)
PPLL PLL
hojastallos

Capítulo III-Resultados y Discusión
33
0
0,5
1
1,5
2
Flav(%MS)
C I T A
Variedad
PPLLPLL
Fig. 8. Comportamiento de los flavonoides en las hojas de cuatro variedades de M. alba.
-Tallos tiernos
La concentración de flavonoides en el PPLL osciló entre (0,81-1,72) ±0,07 %MS, caracterizados por una interacción significativa entre los factores Variedad y Fertilización (fig. 9).
Se observó un incremento en el contenido de flavonoides con el aumento de las dosis de fertilizante orgánico para las variedades A, C, y T; mientras que la variedad I mostró un comportamiento atípico en ese sentido, con el mayor valor para 300 kg N/ha/año.
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
Flav
(%M
S)
1,8
1,6
1,4
1,2
1,0
,8
,6
VARIEDAD
A
C
I
T
Fig. 9. Comportamiento de los flavonoides con la fertilización en los tallos tiernos de cuatro
variedades de M. alba en el PPLL.
Los resultados numéricos obtenidos coinciden con los informados en M. alba por Zhishen, Mencheng y Jianming (1999) al estudiar estos metabolitos en cuatro especies del género Morus.
Igualmente que para otros indicadores fitoquímicos, el nivel de fertilización que mostró la mayor homogeneidad en las variedades que tuvieron comportamientos similares fue el de 300 kg N/ha/año.
En el PLL no se observaron diferencias significativas en el contenido de flavonoides con el incremento de la fertilización, ni tampoco entre las variedades, de manera similar a lo obtenido en el caso de las hojas (fig. 10 y 11).
00,20,40,60,8
1
Flav (%MS)
100 300 500
N kg/ha/año
Fig. 10. Comportamiento de los flavonoides con la fertilización en los tallos tiernos de M. alba
en el PLL.

Capítulo III-Resultados y Discusión
34
00,20,40,60,8
1
Flav(%MS)
C I T A
Variedad
Fig. 11. Comportamiento de los flavonoides en los tallos tiernos de cuatro variedades de M.
alba en el PLL.
Las concentraciones encontradas en el PLL fueron inferiores a los niveles obtenidos en el PPLL, aunque las diferencias entre las concentraciones de las hojas y los tallos tiernos se diferenciaron más en el PLL.
El factor de mayor influencia en los niveles de flavonoides en M. alba fue la parte de la fracción comestible, seguido por la época (fig. 12).
Aunque los factores Fertilización y Variedad fueron menos relevantes de forma independiente, su combinación produjo diferenciaciones en cuanto a las tendencias observadas en los tallos tiernos del PPLL.
Fig. 12. Comportamiento de los flavonoides con los factores parte de la fracción comestible y época del año empleando la media Grand.
3.1.3.3. Cumarinas
-Hojas
En el período PPLL el contenido de cumarinas osciló entre (0,43-0,68) ±0,01 %MS y se observó una interacción significativa entre los factores Variedad y Fertilización, la cual estuvo descrita por las mayores concentraciones para el nivel de 100 kg N/ha/año; los tenores más elevados se observaron en la variedad C (fig. 13)
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
Cum
(%M
S)
,7
,6
,5
,4
,3
VARIEDAD
A
C
I
T
Fig. 13. Comportamiento de las cumarinas con la fertilización en las hojas de cuatro
variedades de M. alba en el PPLL.
00,5
11,5
2
Flav (%MS)
PPLL PLL
hojastallos

Capítulo III-Resultados y Discusión
35
A modo de comparación con otra especie que constituye una fuente clásica de cumarinas, los resultados obtenidos en este experimento son similares a las informados por Urriola (1994), el cual evaluó las concentraciones de estos compuestos en el follaje de cinco ecotipos de G. sepium de Costa Rica (0,55-0,66 %MS) mediante el mismo método analítico empleado en esta tesis.
En el PLL la interacción Variedad x Fertilización presentó valores entre (0,54-0,79) ±0,01 %MS, con una tendencia al incremento de las concentraciones para el nivel de 500 kg N/ha/año (fig 14).
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
Cum
(%M
S)
,9
,8
,7
,6
,5
,4
VARIEDAD
A
C
I
T
Fig. 14. Comportamiento de las cumarinas con la fertilización en las hojas de cuatro
variedades de M. alba en el PLL.
Estos valores numéricos también son similares a los obtenidos por López (1994), quien evaluó el contenido de cumarinas en dos procedencias de G. sepium de Guatemala establecidas en el trópico húmedo de Costa Rica (0,50-1,50 %MS), aunque este autor no comparó la concentración encontrada entre las procedencias. -Tallos tiernos
En el PPLL los valores oscilaron entre (0,43-0,80) ±0,01 %MS, con las mayores concentraciones para el nivel de 500 kg N/ha/año (fig. 15).
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
Cum
(%M
S)
,9
,8
,7
,6
,5
,4
VARIEDAD
A
C
I
T
Fig. 15. Comportamiento de las cumarinas con la fertilización en los tallos tiernos de cuatro
variedades de M. alba en el PPLL. El rango de valores obtenido en el PPLL coincide con el encontrado por Mochiutti (1995), al determinar las
concentraciones de cumarinas en G. sepium sometida a pastoreo en Costa Rica. En el PLL la interacción Variedad x Fertilización mostró tendencias variables entre las variedades; la
concentración más elevada la aportó la variedad A con 100 kg N/ha/año (0,65 %MS) y el menor nivel correspondió a la T con 300 kg N/ha/año (0,42 %MS).
La heterogeneidad en los contenidos de cumarinas totales ha sido señalada por López (1995) al evaluar cinco follajes provenientes de G. sepium, el cual obtuvo una diferencia de 1,90 % con un rango de 0,46 a 2,34 %MS. En el caso de las cumarinas, la combinación de los factores Variedad y Fertilización presentó el mayor efecto. Por otra parte las variables parte de la fracción comestible y período del año no crearon diferencias marcadas.

Capítulo III-Resultados y Discusión
36
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
Cum
(%M
S)
,7
,6
,5
,4
VARIEDAD
A
C
I
T
Fig. 16. Comportamiento de las cumarinas con la fertilización en los tallos tiernos de cuatro
variedades de M. alba en el PLL.
Fig. 17. Comportamiento de las cumarinas con los factores parte de la fracción comestible y época del año empleando la media Grand.
3.1.3.4. Carbohidratos solubles -Hojas
El contenido de CHS en el PPLL presentó una interacción Variedad x Fertilización significativa, descrita por valores que oscilaron entre (10,40-16,45) ±0,33 %MS; resultados similares fueron obtenidos por Ojeda, Montejo y Pérez (2000) en muestras de morera fresca, empleadas como control en la fabricación de ensilajes con aditivos.
Las cuatro variedades estudiadas tuvieron el mismo comportamiento en la interacción (fig. 18); la variedad I se caracterizó por exhibir las concentraciones de CHS más bajas para los tres niveles de fertilización, quizás por tener menor posibilidad de síntesis de CHS en el período seco, o por poseer una menor área foliar en comparación con el resto de las variedades, que sí presentan áreas foliares mayores y similares entre ellas. Los CHS aumentaron marcadamente con la aplicación de 300 kg N/ha/año. El incremento en el contenido energético con esta dosis de N, basado en mediciones de Energía Bruta (EB Kcal/kg MS) en el PPLL, fue también reportado en la misma plantación de estudio por González, Martín, Alvanell, Cajas y Rosas (2002) (fig. 19).
En este sentido, los autores también observaron mayores niveles energéticos en la época de lluvia comparada con el PPLL, así como un comportamiento diferente de las hojas y los tallos tiernos en el PLL; lo cual apoya lo obtenido en las presentes cuantificaciones.
Las concentraciones de CHS determinadas en este experimento fueron superiores en las hojas que en los tallos tiernos; estos resultados son inversos a los obtenidos por estos autores. Ambos resultados no necesariamente deben coincidir, ya que la EB fue determinada calorimétricamente, por lo que la energía desprendida en la combustión no solo provenía de los carbohidratos, sino también de otros compuestos como los componentes de la pared celular, lípidos y proteínas, que son también una fuente no despreciable de energía en los análisis por combustión (Ferrer, Higuera, Castro, García, López, Soto, Carrillo, Mogollón y Urdaneta, 1996).
0
0,2
0,4
0,6Cum
(%MS)
PPLL PLL
hojastallos
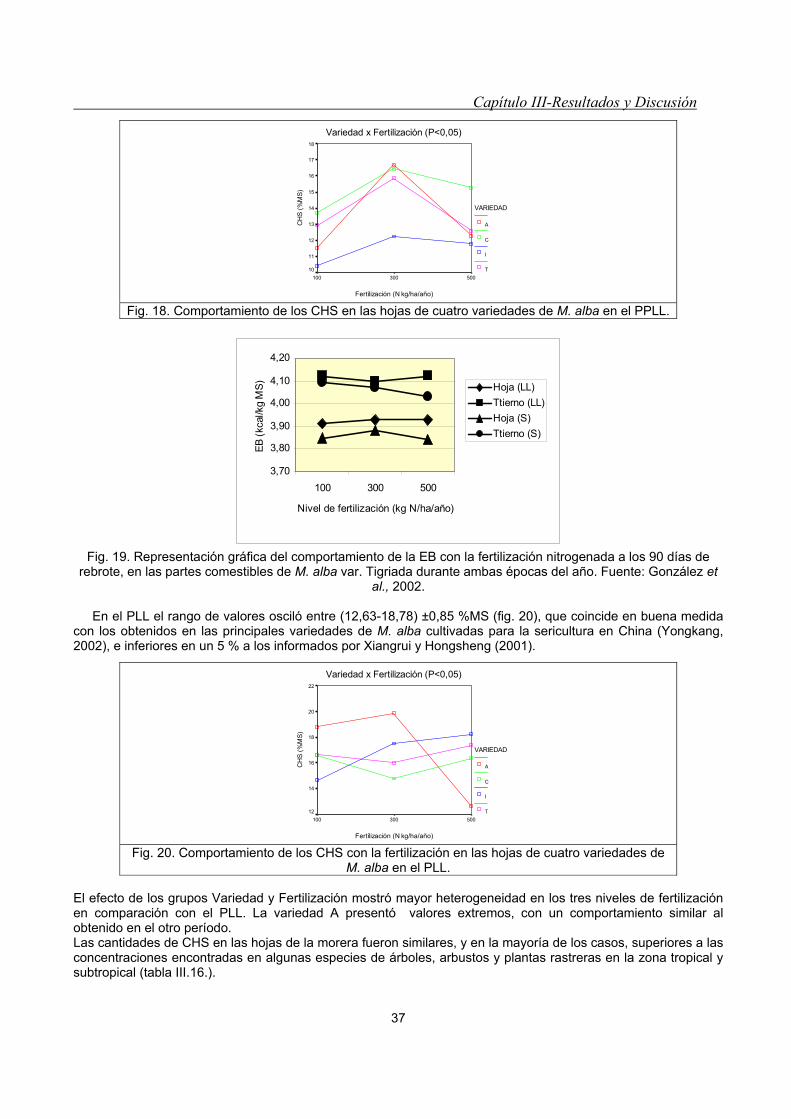
Capítulo III-Resultados y Discusión
37
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
CH
S (%
MS)
18
17
16
15
14
13
12
11
10
VARIEDAD
A
C
I
T
Fig. 18. Comportamiento de los CHS en las hojas de cuatro variedades de M. alba en el PPLL.
3,70
3,80
3,90
4,00
4,10
4,20
100 300 500
Nivel de fertilización (kg N/ha/año)
EB (k
cal/k
g M
S) Hoja (LL)Ttierno (LL)Hoja (S)Ttierno (S)
Fig. 19. Representación gráfica del comportamiento de la EB con la fertilización nitrogenada a los 90 días de
rebrote, en las partes comestibles de M. alba var. Tigriada durante ambas épocas del año. Fuente: González et al., 2002.
En el PLL el rango de valores osciló entre (12,63-18,78) ±0,85 %MS (fig. 20), que coincide en buena medida
con los obtenidos en las principales variedades de M. alba cultivadas para la sericultura en China (Yongkang, 2002), e inferiores en un 5 % a los informados por Xiangrui y Hongsheng (2001).
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
CH
S (%
MS)
22
20
18
16
14
12
VARIEDAD
A
C
I
T
Fig. 20. Comportamiento de los CHS con la fertilización en las hojas de cuatro variedades de
M. alba en el PLL. El efecto de los grupos Variedad y Fertilización mostró mayor heterogeneidad en los tres niveles de fertilización en comparación con el PLL. La variedad A presentó valores extremos, con un comportamiento similar al obtenido en el otro período. Las cantidades de CHS en las hojas de la morera fueron similares, y en la mayoría de los casos, superiores a las concentraciones encontradas en algunas especies de árboles, arbustos y plantas rastreras en la zona tropical y subtropical (tabla III.16.).

Capítulo III-Resultados y Discusión
38
Tabla III.16. Contenido de CHS en las hojas de algunos árboles, arbustos y plantas rastreras, comparados con M. alba.
Especie CHS (% MS) Referencia
Cajanus cajan 14, 95 (90 días) Ferrer et al. (1996) M. alba 12,63-18,78 (PLL)
Acacia mangium 7,16 Clavero (1998) Physalis peruviana 7,40 Fischer y Ludders (1997) Ramonda miconi 5,28 Xerophyta villosa 3,64
Ghasempour et al. (1998)
-Tallos tiernos
El contenido de CHS en los tallos tiernos en el PPLL fluctuó entre (3,86-10,92) ±0,45 %MS (fig. 21); se observó una tendencia homogénea en la interacción de todas las variedades, con un comportamiento similar a las hojas en ese mismo período. Sobresalió la variedad C por el contenido de CHS para la aplicación de 300 kg N/ha/año (igual que lo observado en las hojas).
Variedad Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
CH
S (%
MS)
12
10
8
6
4
2
VARIEDAD
A
C
I
T
Fig. 21. Comportamiento de los CHS con la fertilización en los tallos tiernos de cuatro
variedades de M. alba en el PPLL.
Los contenidos de CHS obtenidos en este experimento son superiores a los reportados por García, Nygren y Desfontaines (2001) para los tallos tiernos en G. sepium en el PPLL.
En el PLL se observó un efecto significativo de los grupos Fertilización y Variedad, en un intervalo menor que el obtenido para los tallos tiernos en el PPLL (tabla III.17.).
Rangos similares de CHS (<6,00 %MS) fueron obtenidos por García et al. (2001) en los tallos tiernos de la leguminosa G. sepium para la época lluviosa y concentraciones similares a las obtenidas por Clavero (1998) en A. mangium (5,01 %MS).
En el PPLL se encontró un rango de CHS similar al obtenido en el PLL en el caso de las hojas; comportamiento contrario con relación a la época mostraron los tallos tiernos, donde el rango más amplio se observó en el PPLL. Esto demuestra los contrastes que existen en las concentraciones de estas sustancias de reserva en los tallos de esta planta.
En el PPLL, donde las radiaciones solares fueron elevadas, pero la cantidad de agua disponible fue un factor limitante, es posible que M. alba haya utilizado más rápidamente los glúcidos ó haya sintetizado menor cantidad que en el PLL.
En este sentido Das, Sengupta, Chattopadhyay, Setua, Das y Saratchandra (2002) encontraron una reducción del crecimiento y el rendimiento foliar, unido al aumento sextuplicado de cantidades de Prolina, lo que evidentemente conllevó a una disminución energética considerable. Reducciones en las concentraciones de CHS y almidones con el aumento de cantidades apreciables de Prolina, han sido también reportadas por Singhal y Roopa (1998).
Por otra parte Ramanjulu, Screenivasalu, Kumar y Sudhakar (1998), al evaluar el efecto del estrés por supresión de agua en plantas adultas de morera, demostraron que el potencial hídrico de las hojas, los valores de área foliar, el contenido total de clorofila, la tasa fotosintética, la conductancia estomática y la transpiración fueron disminuyendo, lo que trajo consigo la disminución de los CHS disponibles para llevar a cabo las funciones metabólicas en la planta.

Capítulo III-Resultados y Discusión
39
Tabla III.17. Efecto de los grupos Variedad y Fertilización en el contenido de CHS en los tallos tiernos en el PLL.
Grupos Variedad Fertilización
CHS(%MS)
C 100 300 500
5,98 5,44 4,51
I 100 300 500
5,47 4,25 5,62
T 100 300 500
4,75 4,99 5,79
A 100 300 500
5,39 5,73 5,41
ES ± 0,19
Ramajulu y Sudhakar (2000) han encontrado acumulaciones apreciables de amonio cuaternario en las hojas de morera, como producto de la mayor actividad de la enzima-5-Carboxilato Reductasa relacionada con la síntesis de sales cuaternarias de aminoácidos nitrogenados, especialmente Prolina.
Por otra parte, del total de los CHS determinados en las hojas de las variedades, entre el 85 y 94 % fueron carbohidratos solubles en etanol al 95 % en el PPLL; de igual manera, en el PLL los valores oscilaron entre 74 y 87 %. Estos resultados ponen de manifiesto la elevada fracción de carbohidratos no estructurales complejos, al parecer en forma de polisacáridos solubles de tipo almidón. En sentido general, en las hojas los menores porcentajes de CHS en alcohol se observaron con la fertilización de 300 kg N/ha/año; excepto para la variedad C, la cual mostró un incremento con la fertilización (fig. 22).
Fertilización (N kg/ha/año)
500300100
CH
S Al
c(C
HS)
88
86
84
82
80
78
76
74
72
VARIEDAD
A
C
I
T
Fig. 22. Comportamiento de los CHS en etanol con la fertilización en las hojas de cuatro variedades de M. alba
en el PLL.
Los tallos tiernos en el PPLL se caracterizaron por presentar porcentajes de CHS en alcohol entre el 62 y 90 %, con una tendencia al incremento de estos tipos de carbohidratos con la fertilización, quizás por la mayor acumulación de estas sustancias en vías de ser almacenadas como reserva a nivel del tallo y/o raíz (fig. 23a).
En el PLL los tallos tiernos presentaron porcentajes entre 81 y 89 %, con una tendencia a la disminución de los CHS en etanol con el aumento fertilización (fig 23b); quizás por hacer un mayor uso de los CHS en la producción de tejido vegetativo a medida que la disponibilidad de nutrientes aumentó y las condiciones fueron más propicias para una mayor producción de biomasa. La dinámica inversa observada demuestra el protagonismo que tienen los tallos en la acumulación y traslocación de estas sustancias de reserva en la planta.
Cantidades apreciables de polisacáridos no estructurales solubles en alcohol, como Peptinas (12 %MS), Arabanos (7,4 %MS), Pentosanos (3,8 %MS) y Amilosa (1 %MS) han sido encontrados por Xiangrui y Hongsheng (2001).
En orden de efecto, las variables de mayor influencia en la concentración de CHS fueron, la parte de la fracción comestible, la combinación de los factores Variedad y la Fertilización y la época.

Capítulo III-Resultados y Discusión
40
Fertilización (N kg/ha/año)
500300100
CH
S Al
c(C
HS)
100
90
80
70
60
50
VARIEDAD
A
C
I
T
Fertilización (N kg/ha/año)
500300100
CH
SAlc
(CH
S)
90
88
86
84
82
80
VARIEDAD
A
C
I
T
a b
Fig. 23. Comportamiento de los CHS en alcohol con la fertilización en los tallos tiernos de cuatro variedades de M. alba. a) PPLL, b) PLL
Fig. 24. Comportamiento de los CHS con los factores parte de la fracción comestible y época del año empleando
la media Grand. 3.1.3.5. Esteroles totales -Hojas
El contenido de esteroides totales en el PPLL presentó una interacción Variedad x Fertilización significativa, descrita por valores que oscilaron entre (4,08-11,78) ±0,32 mg/gMS (fig. 25). Las cuatro variedades estudiadas mostraron el mismo comportamiento en la interacción, cuya tendencia fue el incremento de estos metabolitos para el nivel de fertilización intermedio, similar al obtenido para los CHS en el PPLL; la variedad A se caracterizó por exhibir los valores más elevados para los tres niveles de fertilización.
La conexión entre el contenido energético y la síntesis de compuestos esteroidales precursores de hormonas reguladoras del crecimiento vegetal ha sido señalada por Chappell, Vonlanken y Vogeli, 1991; Chappell, Waaf, Proulx, Cuellar y Saunders, 1995; Chappell, 1995.
Resultados inferiores han sido obtenidos por Duke (2001) en la determinación específica de β-sitosterol (2 mg/g) en las hojas de morera, aunque el autor también describe la presencia de otros esteroles los cuales no cuantificó, por lo que las determinaciones del contenido total de estos metabolitos en el presente experimento han sido superiores.
Los contenidos de ET expresados como colesterol, mediante la cuantificación por el mismo método analítico que el utilizado por Galindo et al. (1989), denotan resultados superiores en las hojas de morera analizadas en este experimento, en comparación con las arbóreas investigadas por estos autores (tabla III. 18.).
En el PLL la interacción Variedad x Fertilización no fue significativa; el comportamiento de los esteroides con la fertilización mostró diferencias significativas entre los niveles de 100 y 300 kg N/ha/año, con un ligero descenso en el contenido de estos compuestos para los niveles de fertilización más elevados.
0
5
10
15
20
CHS (% MS)
PPLL PLL
hojastallos

Capítulo III-Resultados y Discusión
41
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
ET (m
g/g)
14
12
10
8
6
4
2
VARIEDAD
A
C
I
T
Fig. 25. Comportamiento de los ET con la fertilización en las hojas de cuatro variedades de M.
alba en el PPLL. Tabla III. 18. Contenido de ET en especies arbóreas (Galindo et al., 1989), comparadas con el de M. alba, en el
PPLL.
Especie ET (mg/g) M. alba (hojas, PPLL) 4,08-11,78 Inga spectabilis (Guamo) 6,20 Trichantera gigantea (Nacedero) 6,20 G. sepium 1,70
ab ab
0
5
10
15
20
ET(mg/g)
100 300 500
N Kg/ha/año
Fig. 26. Comportamiento de los ET con la fertilización en las hojas de cuatro variedades de M. alba
en el PLL. a, b Valores con superíndices desiguales difieren estadísticamente mediante la dócima de Student-Newman-Keuls a P<0,05
Los contenidos en el PLL oscilaron entre 11,97 y 16,58 mg/gMS, resultados que triplican los obtenidos en el
PPLL; posiblemente por el aumento de la actividad hormonal y por el mayor protagonismo que ejercen estos compuestos en la regulación del metabolismo cuando el crecimiento vegetativo es más rápido.
Estos resultados fueron superiores quizás por la mayor disponibilidad de nutrientes y agua, unido a las favorables condiciones ambientales que contribuyeron al acelerado desarrollo de la planta en el PLL.
Los contenidos de ET en las hojas de las cuatro variedades en el PLL también mostraron diferencias significativas a favor de la variedad I (fig. 27).
Las variedades C y A mostraron concentraciones intermedias y similares, con un menor contenido en la variedad T.

Capítulo III-Resultados y Discusión
42
ab a
cbc
0
5
10
15
20
ET(mg/g)
C I T A
Variedad
Fig. 27. Comportamiento de los ET en las hojas de cuatro variedades de M. alba en el PLL. a, b, c Valores con superíndices desiguales difieren estadísticamente mediante la dócima de Student-Newman-Keuls a P<0,05
-Tallos tiernos
El contenido de ET en el PPLL fluctuó entre (1,60-3,76) ±0,17 mg/gMS (fig. 28) y se observó una tendencia homogénea en la interacción Variedad x Fertilización.
Un comportamiento diferente con la fertilización lo tuvo la variedad C, con las concentraciones más elevadas para las dosis de 100 y 300 kg N/ha/año.
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
ET (m
g/g)
4,0
3,5
3,0
2,5
2,0
1,5
1,0
VARIEDAD
A
C
I
T
Fig. 28. Comportamiento de los ET con la fertilización en los tallos tiernos de de M. alba en el
PPLL.
La cantidad de ET en los tallos tiernos de la morera fue superior a la obtenida por Mengzhao (1989), quien reportó contenidos de 0,03 y 0,04 mg/gMS de Estigmasterol y β-Sitosterol, respectivamente, así como trazas de colesterina.
En el PLL las concentraciones de ET se comportaron entre 0,94 y 1,38 mg/gMS para la fertilización, sin observarse diferencias significativas entre los valores de cada nivel (fig. 29).
En el caso de las variedades los valores oscilaron entre (0,90 y 1,25 mg/gMS) sin diferencias significativas entre ellas (fig. 30); el efecto de los grupos Fertilización y Variedad no fue significativo.
Los valores obtenidos son ligeramente superiores a los reportados por Ho-Zoo y Won-Chu (2001) para cuantificaciones de β-sitosterol solamente.
En sentido general, el PPLL mostró contenidos de ET inferiores a los del PLL en las hojas; en cambio se observó el comportamiento inverso en el caso de los tallos tiernos. Este resultado pone de manifiesto la mayor síntesis de esteroles en las hojas en el período lluvioso, relacionados quizás con una mayor actividad hormonal en la etapa de crecimiento de la planta. Por otra parte, se observó una marcada interacción Variedad x Fertilización en el PPLL en las hojas y los tallos tiernos, lo que evidencia la respuesta de las variedades con ese factor.

Capítulo III-Resultados y Discusión
43
0
0,5
1
1,5
2
ET(mg/g)
100 300 500
N Kg/ha/año
Fig. 29. Comportamiento de los ET con la fertilización en los tallos tiernos en el PLL.
0
0,5
1
1,5
2
ET(mg/g)
C I T A
Variedad
Fig. 30. Comportamiento de los ET con la fertilización en los tallos tiernos de cuatro
variedades de M. alba en el PLL.
En el PLL la interacción no fue significativa y aunque los contenidos de esteroles fueron marcadamente diferentes en las hojas y los tallos, se observó un comportamiento similar de las variedades ante la fertilización en el PPLL.
Los factores de mayor influencia en las concentraciones de los ET fueron la época y la parte de la fracción comestible (fig. 31), seguido por la variedad y la fertilización.
Fig. 31. Comportamiento de los ET con los factores parte de la fracción comestible y época del año, empleando
la media Grand. 3.1.3.6. Influencia de los factores estudiados en el metabolismo secundario
El efecto de los diferentes factores en los metabolitos secundarios estuvo estrechamente relacionado con la naturaleza de cada tipo de compuesto, con particularidades marcadas para cada grupo.
En el caso de los FT, los factores Variedad y Fertilización presentaron igual influencia en los resultados obtenidos; la época no afectó apreciablemente la concentración de estos compuestos y los cambios más drásticos se observaron entre las hojas y los tallos tiernos.
0
5
10
15
ET (mg/g)
PPLL PLL
hojastallos

Capítulo III-Resultados y Discusión
44
En los flavonoides, los factores Fertilización, Variedad y Época crearon pocas variaciones en los niveles de estas especies fenólicas; mientras que los tenores diferenciados entre las hojas y los tallos tiernos fueron los elementos más importante en cuanto a las mayores variaciones.
Estos resultados demuestran la estrecha relación que tienen los compuestos fenólicos con la actividad del metabolismo primario del nitrógeno.
Las cumarinas exhibieron las menores concentraciones tanto en las hojas como en los tallos tiernos, entre todos los fenólidos cuantificados.
Contrariamente al resto de los fenoles y demás grupos de metabolitos, la parte de la planta, no presentó una influencia marcada en las concentraciones, al igual que la época. Los factores Fertilización y Variedad tuvieron mayor importancia en cuanto a la diferenciación de las tendencias para cada caso.
Como producto de la prioridad relativa que han tenido los factores evaluados, las cumarinas presentes en M. alba quizás se relacionen con la posible acción de defensa frente a patógenos, insectos herbívoros, o como protección interespecífica.
En los CHS el factor de mayor incidencia fue la parte de la fracción comestible, seguido por la época. La Variedad y la Fertilización, aunque también presentaron un efecto significativo, fueron de menor cuantía.
Las diferencias encontradas entre las partes de la planta, así como la variabilidad que presentaron con la época, demuestran el papel protagónico que desempeñan estos compuestos como principal fuente energética en la especie.
En el caso de los ET los factores de mayor incidencia fueron la Época y la parte de la fracción comestible, seguido por la Fertilización y la Variedad.
Estos resultados, en buena medida, corroboran la función particular que desempeñan estos compuestos en las especies vegetales como activadores del crecimiento en la época lluviosa.
La variedad C se caracterizó por presentar atipicidades en las tendencias de los FT, las cumarinas y los CHS en el PPLL.
En el PLL el porcentaje de PV (%PB) en los tallos tiernos osciló entre 70,58-96,50 % en las variedades con la frecuencia. Los mayores valores se obtuvieron en la frecuencia de 120 días, al igual que ocurrió en las hojas. Los valores entre los niveles de fertilización y la frecuencia fluctuaron entre 68,00-88,62 %, con el aumento de la PV (%PB) a medida que disminuyó la frecuencia de corte.
El mayor cúmulo de interacciones en el caso de la PV con respecto a la PB en ambos períodos pone de manifiesto la mayor significación biológica de este indicador en el metabolismo del N en las hojas y los tallos tiernos en las variedades. 3.2. Experimento 2. “Efecto de la parte de la planta, la frecuencia de corte, la fertilización, las variedades
y la época en el comportamiento de los compuestos nitrogenados en M. alba”. 3.2.1. Proteína bruta -Hojas
Los contenidos de PB no presentaron diferencias significativas con el aumento de los niveles de fertilización en los dos períodos evaluados. Las concentraciones de PB en el PPLL fueron superiores a las obtenidas en el PLL (fig. 32); resultados similares han sido reportados por González y Cáceres (2002), al evaluar el contenido de PB en las hojas de M. alba, para ambas épocas. El aumento de los niveles de PB en el PPLL se debió fundamentalmente al menor porcentaje de humedad que contenía esta parte de la planta en el PPLL.
Mientras que los porcentajes de PB oscilaron alrededor del 18 %MS en los tratamientos, el rendimiento de PB/ha/corte tuvo un comportamiento definido con el incremento de la fertilización. Se observó una tendencia generalizada al aumento a medida que se incrementaron las dosis nitrogenadas (fig. 33).
Aunque las mayores concentraciones de PB se obtuvieron en el PPLL, los aportes más elevados de PB/ha/corte fueron elementos característicos del PLL, debido a los mayores rendimientos de hojas en esta etapa (fig. 33b).
Por otra parte, se observaron diferencias significativas entre los niveles de PB al aumentar los espaciamientos entre los cortes en el PPLL. La mayor concentración de PB se obtuvo en la frecuencia más intensa; en cambio, en el PLL aunque la tendencia fue similar, no se encontraron diferencias apreciables entre los tenores de PB con la disminución de los intervalos de corte.
Comportamiento típico similar de la PB en las hojas con el aumento de los espaciamientos entre cortes, han sido obtenidas por González et al. (2002) en la variedad T y por Francisco (2002) en el segundo año de evaluación en A. lebbeck.

Capítulo III-Resultados y Discusión
45
0
5
10
15
20
PB (%MS)
100 300 500
N Kg/ha/año
PPLLPLL
Fig. 32. Comportamiento de la PB con la fertilización orgánica en las hojas de M. alba.
0
100
200
300
400
kg PB/ha
100 300 500
N Kg/ha/año
PPLLPLL
Fig. 33. Rendimiento de la PB con la fertilización orgánica en las hojas de M. alba.
a
b b
0
5
10
15
20
25
PB (%MS)
60 90 120
Frecuencia
PPLLPLL
Fig. 34. Comportamiento de la PB con la frecuencia de corte en las hojas de M. alba. a, b Valores con superíndices desiguales difieren estadísticamente mediante la
dócima de Student-Newman-Keuls a P<0,05 Decrecimientos similares, aunque menos abruptos, en el comportamiento de la PB con el aumento de la
madurez de las hojas, ha sido reportados por Razz (1991) en L. leucocephala, y por Boschini (2002a) en una plantación de morera defoliada a los 56, 84 y 120 días, pero con fertilización química a razón de 150 kg de NH4NO3/ha/año.
Los rendimientos de PB/ha/corte aumentaron marcadamente con la disminución de la frecuencia de defoliación; esta tendencia fue común en ambos períodos, con rendimientos superiores a los 150 kg PB/ha/corte en el PLL (fig. 35). Los aportes de proteína de las hojas fueron similares a los obtenidos por Francisco (2002) en A. lebbeck, y similares a la tendencia descrita por Martín et al. (2002) en M. alba, la que fue manejada a 45, 60, 75 y 90 días entre cortes y donde las mayores producciones se obtuvieron en las defoliaciones cada 90 días.

Capítulo III-Resultados y Discusión
46
0
100
200
300
400
500
kg PB/ha
60 90 120
Frecuencia
PPLLPLL
Fig. 35. Rendimiento de la PB con la frecuencia de corte en las hojas de M. alba.
Aunque no se observaron variaciones considerables en los rendimientos a partir de los 90 días de rebrote en
el PLL, en la época de seca los incrementos fueron proporcionales con el aumento de los espaciamientos entre cortes.
Las variedades estudiadas en este experimento tampoco presentaron diferencias significativas en el contenido de PB en ambos períodos (fig. 36); las concentraciones en el PPLL fueron superiores en comparación con el PLL de manera homogénea para todas las variedades.
0
5
10
15
20
25
PB (%MS)
C I T A
Variedad
PPLLPLL
Fig. 36. Comportamiento de la PB en las hojas de cuatro variedades de M. alba.
Las concentraciones encontradas fueron inferiores a los obtenidas por González et al. (1998) para las
mismas variedades empleadas en esta investigación a los 90 días de rebrote. Para las plantas arbóreas forrajeras, como es el caso de M. alba, al igual que sus producciones de biomasa,
su contenido nutricional es elevado en los momentos iniciales o en los primeros años de defoliaciones continuas, cuando se compara con los cortes subsecuentes (Sánchez, Miquilena y Flores, 2000), donde se inicia un ciclo de variaciones hasta lograr estabilizarse y mostrar su verdadero potencial nutricional (Papanastasis, Platis y Dini-Papanastasi, 1998); quizás por eso, los tenores encontrados en estas determinaciones sean inferiores a los hallados tres años atrás.
Los rendimientos (expresados en kg PB/ha/corte) mostraron diferencias muy marcadas con la época, observándose la mayor variabilidad y los menores rendimientos entre las variedades en el PPLL (fig. 37). En esa etapa las variedades I y T sobresalieron con rendimientos superiores a los 100 kg PB/ha/corte, debido a las mayores producciones de hojas. La elevada producción de biomasa total de la variedad I ha sido reportada por Martín et al. (1998) en el período de establecimiento de las cuatro variedades en Cuba. La variedad C exhibió los resultados más bajos, con una producción inferior a 50 kg PB/ha/corte por su poca producción de hojas en el PPLL.
En el PLL las variedades produjeron rendimientos muy similares entre ellas, los cuales casi se triplicaron con respecto al PPLL.
Resumiendo lo ocurrido en las hojas, puede plantearse que la fertilización orgánica no produjo aumentos significativos en los contenidos de PB en ninguno de los dos períodos de evaluación, comportamiento similar al obtenido por Benavides et al. (1994) al utilizar estiércol de cabra como fuente nitrogenada.

Capítulo III-Resultados y Discusión
47
La frecuencia de corte produjo un efecto marcado en el PPLL. Con la disminución de la frecuencia de corte esta especie mostró el mismo patrón de comportamiento de las principales y más conocidas arbóreas en el trópico, sin observarse diferencias marcadas en las concentraciones en el PLL.
0
100
200
300
400
kg PB/ha
C I T A
Variedad
PPLLPLL
Fig. 37. Rendimiento de la PB en las hojas de cuatro variedades de M. alba.
Las concentraciones de PB en las variedades fueron superiores a las obtenidas en todas las especies de
gramíneas y comparables con las principales leguminosas del trópico (Jegou et al., 1994). Estas concentraciones son similares a las obtenidas por Shayo (1997) en M. alba cultivada en una de las partes semiáridas de Tanzania.
En las frecuencias evaluadas se pudieron constatar altos porcentajes de PB, similares a los obtenidos en L. leucocephala y superior a los de otras arbóreas forrajeras tales como Acacia spp. y Lisiloma latisiliquum, con 13,5 y 21,3 %MS, respectivamente (Arango, Adogla-Bessa, Omphile y Tshireletso, 2000). Los contenidos proteicos alcanzados en esta evaluación son aceptables, si se considera que acerca de algunas leguminosas arbóreas fijadoras de N, existen reportes desde 20 %MS (Russo y Botero, 1996) hasta 29 %MS (Simón, Cáceres, Santana y Hernández, Iglesias, Duquesne, Delgado y Docazal, 1992). Adicionalmente los tenores de PB son comparables a los de otras especies de considerable potencial forrajero, tales como G. sepium y otras especies del género Leucaena (Pinto, Ramírez-Avilés y Ku-Vera, 2001; Ibrahim, Franco, Pezo, Camero y Araya, 2000).
En las defoliaciones los tenores de proteínas superaron a los descritos por Colectivo de autores (2000) en todos los géneros de pastos comunmente usados en el trópico, lo que reafirma el potencial de la especie como una opción de suplemento importante en muchas dietas de baja calidad para rumiantes y monogástricos.
En todos los casos, los mayores y diferenciados rendimientos de PB/ha/corte se encontraron en el PLL, donde se triplicó la producción de hojas con respecto al PPLL. -Tallos tiernos
En el PPLL la concentración de PB presentó un aumento significativo con el incremento de los niveles de fertilizante orgánico, con diferencias a favor de las dos dosis superiores (fig. 38), de manera similar a lo obtenido por Boschini et al. (1999) al utilizar NH4NO3 como fertilizante químico.
Por otra parte, no se encontraron diferencias significativas en la concentración de PB con el aumento de la dosis de fertilización en el PLL. Estos resultados son similares a los obtenidos por Benavides et al. (1994).
Los rendimientos de PB/ha/corte presentaron la misma tendencia que la obtenida en unidades de concentración en el PPLL, con producciones superiores a los 10 kg/ha/corte (fig. 39).
Las producciones de PB/ha/corte en el PLL se incrementaron para el nivel de 300 kg N/ha/año, con respecto al resto de los niveles.
En sentido general, mediante las determinaciones realizadas se pudo comprobar que las dosis crecientes de fertilización trajeron consigo un aumento considerable en los rendimientos de PB/ha/corte; conjuntamente con el incremento de la concentración de PB en el PPLL y la poca variación en el PLL.
Los mayores rendimientos de PB/ha/corte en el PLL se debieron a la mayor producción de tallos tiernos para cada dosis de N. Resultados similares han sido obtenido por Boschini et al. (1999).
En el PPLL el contenido de PB para las diferentes frecuencias mostró diferencias significativas a favor de los 60 días; en cambio, no se encontraron diferencias entre las concentraciones en el PLL (fig. 40).

Capítulo III-Resultados y Discusión
48
b a a
02468
10
PB (%MS)
100 300 500
N Kg/ha/año
PPLLPLL
Fig. 38. Comportamiento de la PB con la fertilización en los tallos tiernos. a, b Valores con superíndices desiguales difieren estadísticamente
mediante la dócima de Student-Newman-Keuls a P<0,05
0
10
20
30
kg PB/ha
100 300 500
N Kg/ha/año
PPLLPLL
Fig. 39. Comportamiento de la PB/ha/corte con la fertilización en los tallos tiernos de M. alba.
ab b
0
5
10
PB (%MS)
60 90 120
Frecuencia
PPLLPLL
Fig. 40. Comportamiento de la PB con la frecuencia de corte en los tallos tiernos de M. alba. a, b Valores con superíndices desiguales difieren estadísticamente mediante la dócima de
Student-Newman-Keuls a P<0,05 Un decrecimiento en los contenidos de PB en G. sepium con la disminución de la frecuencia de corte fue
obtenido por Miquilena, Ferrer y Clavero (datos inéditos). Los rendimientos de PB/ha/corte aumentaron considerablemente con el espaciamiento entre los cortes
solamente hasta los 90 días; como producto de la menor fracción de tallos tiernos a los 120 días y la disminución en la relación hoja-tallo a partir de la frecuencia intermedia, similar a lo obtenido por Francisco (2002) en A. lebbeck.

Capítulo III-Resultados y Discusión
49
0
10
20
30
40
kg PB/ha
60 90 120
Frecuencia
PPLLPLL
Fig. 41. Rendimiento de la PB con la frecuencia de corte en los tallos tiernos de M. alba.
En ninguna de las dos épocas se observaron diferencias significativas en las concentraciones de PB en los
tallos tiernos de las variedades (fig. 42).
0
2
4
6
8
10
PB (%MS)
C I T A
Variedad
PPLLPLL
Fig. 42. Comportamiento de la PB en los tallos tiernos de cuatro variedades de M. alba.
Los contenidos de PB en los tallos tiernos fueron similares a los informados por Martín et al. (2002) en el
estudio del comportamiento agronómico de la especie en Cuba. Por otra parte, los rendimientos de PB/ha/corte en los tallos fueron superiores en el PLL, de igual manera que
en el caso de las hojas, los rendimientos entre las variedades fueron similares entre ellas, contrariamente a lo obtenido en el PPLL, donde la variedad C mostró los menores aportes (fig. 43). La menor producción de biomasa de la variedad C en el PPLL concuerda con lo expresado por Martín G. (comunicación personal).
0
10
20
30
kg PB/ha
C I T A
Variedad
PPLLPLL
Fig. 43. Rendimiento de la PB en los tallos tiernos de cuatro variedades de M. alba.
Los tallos tiernos mostraron un incremento significativo en la concentración de PB en el PPLL, además de no
producirse variaciones considerables en el PLL. Se observó un comportamiento paralelo en la relación de los niveles de PB con la frecuencia de corte para ambas partes de la planta, así como contenidos similares de PB en los tallos tiernos de las variedades, también se observaron los mayores rendimientos de PB/ha/corte en el PLL, así como una similitud en las producciones de las variedades en este período.

Capítulo III-Resultados y Discusión
50
De manera general, el contenido de PB en las hojas de la morera fue tres veces superior a los niveles encontrados en los tallos tiernos, lo que coincide con lo descrito por Boschini (2002a). 3.2.2. Proteína verdadera -Hojas
La PV se caracterizó por presentar interacciones significativas para todos los pares de factores en estudio. La interacción Variedad x Fertilización estuvo caracterizada por valores entre (14,61-16,32) ±0,17 %MS, con incrementos en las concentraciones de PV en los niveles de 300 y 500 con respecto a 100 kg N/ha/año en las variedades T, I y A. Las mayores concentraciones se obtuvieron para el nivel de 300 kg N/ha/año en las variedades I y A, las que se caracterizaron por presentar una menor capacidad de síntesis proteica a partir de esta dosis. La variedad C mostró un comportamiento atípico y la T se caracterizó por presentar los contenidos más bajos, aunque con una buena eficiencia de conversión nitrogenada (fig. 44a).
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
PV (%
MS)
16,5
16,0
15,5
15,0
14,5
VARIEDAD
A
C
I
T
Fertilización (N kg/ha/año)
500300100
kg P
V/ha
160
140
120
100
80
60
40
20
0
VARIEDAD
A
C
I
T
a b
Fig. 44. Comportamiento de la PV con la fertilización en las hojas de cuatro variedades de M. alba en el PPLL.
De forma diferente a lo obtenido en las variaciones de concentración con los niveles de fertilización, el
rendimiento de PV/ha/corte presentó un comportamiento homogéneo para todas las variedades, con incrementos definidos en las cantidades de PV/ha/corte con los aumentos en las dosis nitrogenadas aplicadas (fig. 44b).
La variedad I se caracterizó por presentar la mayor concentración de PV para los dos mayores niveles, así como el rendimiento por hectárea más elevado entre todas las variedades.
La interacción Variedad x Frecuencia presentó un rango entre (12,02-17,68) ±0,19 %MS; con una tendencia homogénea para todas las variedades, excepto para la A, la cual tuvo el mismo comportamiento que en el caso de la PB (fig. 45a).
Los mayores contenidos de PV se observaron en la frecuencias de 60 y 120 días, con una disminución a los 90 días. En esta frecuencia se observó un descenso en la producción de NP, y el resto del N asimilado por la planta se encontró en forma de NNP. La menor conversión metabólica del NNP a NP a los 90 días quizás fue el efecto de la incidencia de uno o varios factores, en todos los casos relacionados con la fisiología de la especie.
A los 90 días la planta crea tejido vegetativo joven, con la emisión de rebrotes y se encuentra en el período transitorio para la transformación de sus cadenas carbonadas en la síntesis de material leñoso, momento en el cual la relación hoja-tallo comienza a disminuir bruscamente; para dicha transformación se necesitó una elevada cantidad de compuestos endógenos y energía para la síntesis de los nuevos materiales.
Según Boschini et al. (1998) los 90 días de rebrote constituyen, aproximadamente, el punto de inflexión en los cambios fisiológicos en la especie.
El rendimiento de PV por unidad de área, para todas las variedades, presentó un comportamiento similar a lo obtenido en otros indicadores con el factor Frecuencia. La tendencia se caracterizó por el aumento de la productividad de PV/ha/corte para las frecuencias más espaciadas (fig. 45b).
Resultados similares en la concentración de PV en las hojas han sido reportados por Liu et al. (2002) en diferentes clones de morera en China.
La interacción Fertilización x Frecuencia presentó valores entre (12,38-17,33) ±0,65 %MS, con una tendencia a la disminución homogénea a los 90 días (fig. 46a).

Capítulo III-Resultados y Discusión
51
Variedad x Frecuencia (P<0,05)
Frecuencia
1209060
PV (%
MS)
18
17
16
15
14
13
12
11
VARIEDAD
A
C
I
T
Frecuencia
1209060
kg P
V/ha
200
100
0
VARIEDAD
A
C
I
T
a b
Fig. 45. Comportamiento de la PV con la frecuencia de corte en las hojas de cuatro variedades de M. alba en el PPLL.
Frecuencia
1209060
kg P
V/ha
160
140
120
100
80
60
40
20
Fertilización
100
300
500
a b Fig. 46. Comportamiento de la PV con la frecuencia de corte en las hojas de M. alba en el PPLL.
En cambio, los rendimientos de cada nivel de fertilización ascendieron a medida que se incrementó la dosis
de fertilizante (fig. 46b). En el PPLL los resultados indicaron que en las frecuencias de corte de 60 y 120 días se obtienen las
concentraciones de PV más elevadas y mínimos niveles de NNP; a los 90 días los tenores más bajos y máximos concentraciones de entidades no proteicas en las hojas.
En el PLL la PV también quedó regida por interacciones significativas entre todos los pares de factores en estudio.
En la interacción Variedad x Fertilización los valores oscilaron entre (12,18-19,43) ±0,67 %MS, con una tendencia compartida entre las variedades, resultado similar al obtenido en el PPLL (fig. 47a). La variedad C en este caso también presentó una tendencia diferenciada para la fertilización de 300 kg N/ha/año. La variedad A, en ambas épocas, mostró el mismo comportamiento de disminución a partir de la dosis intermedia, lo que evidencia el carácter distintivo de esta variedad en el caso del comportamiento de la PV con la fertilización.
Por otra parte, las variedades I y T presentaron tendencias diferentes entre las dos épocas. Los resultados expresados en kg de PV/ha/corte reflejaron un comportamiento similar al obtenido para las
concentraciones de PV en las variedades con la fertilización, donde también la variedad C mostró un comportamiento diferente al resto (fig. 47b). La variedad T, independientemente de presentar la mayor concentración de PV para los dos primeros niveles de fertilización, también fue la que aportó mejores rendimientos en dichos niveles; estos fueron aproximadamente tres veces superiores a los obtenidos en el PPLL, como en el caso de los rendimientos de la PB.
La interacción Variedad x Frecuencia presentó un rango entre (11,18-19,91) ±0,61 %MS, con una tendencia definida y homogénea dentro de las variedades, hacia una disminución inicial y posterior estabilización en el contenido de PV con la edad, e incrementos menos drásticos a los 120 días que el obtenido en el PPLL. Se encontraron contenidos máximos de PV a los 60 días de rebrote para todas las variedades en estudio (fig. 48a), resultado similar al obtenido en el PPLL.
Fertilización x Frecuencia (P<0,05)
Frecuencia
1209060
PV
(%M
S)
18
17
16
15
14
13
12
Fertización
100
300
500

Capítulo III-Resultados y Discusión
52
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
PV (%
MS)
20
18
16
14
12
10
VARIEDAD
A
C
I
T
Fertilización (N kg/ha/año)
500300100
kg P
V/ha
400
300
200
100
VARIEDAD
A
C
I
T
a b
Fig. 47. Comportamiento de la PV con la fertilización en las hojas de cuatro variedades de M. alba en el PLL.
Variedad x Frecuencia (P<0,05)
Frecuencia
1209060
PV (%
MS)
22
20
18
16
14
12
10
VARIEDAD
A
C
I
T
Frecuencia
1209060
kg P
V/ha
500
400
300
200
100
0
VARIEDAD
A
C
I
T
a b
Fig.48. Comportamiento de la PV con la frecuencia de corte en las hojas de cuatro variedades de M. alba en el PLL.
Concentraciones similares en el PLL han sido obtenidas por Bing et al. (2001) al estudiar el contenido de PV
de cuatro variedades de morera en zonas asiáticas. Los mayores niveles en el PLL se observaron, igualmente, para la frecuencia de 60 días, y contenidos
inferiores para los 90 y 120 días. La variedad T se caracterizó por presentar el mayor contenido de PV a los 60 días.
En sentido general, dentro de cada período de defoliación se observó un mayor rango de variabilidad entre las variedades, excepto a los 120 días, donde los rangos de concentraciones fueron más estrechos.
El crecimiento menos abrupto de los rendimientos a partir de los 90 días se debió a la estabilización en la producción de biomasa en ese período.
El comportamiento similar; aunque menos marcado, en la interacción Variedad x Frecuencia en el PLL, hace pensar que en esa etapa la utilización y transformación de compuestos endógenos a partir de la PV fue inferior, ya que desde el punto de vista funcional la planta pudo extraer del suelo una mayor cantidad de nutrientes en aras de los cambios fisiológicos que se imponían.
En la interacción Fertilización x Frecuencia los tenores oscilaron entre (12,83-19,08) ±1,53 %MS, con una tendencia a observarse, dentro de cada nivel de fertilización, los mayores contenidos para la frecuencia de corte de 60 días (19 %MS para 100 y 300 kg/ha/año y 15 %MS para 500 kg N/ha/año), en comparación con las frecuencias de 90 y 120 días.
Se obtuvo un incremento en el rendimiento de la PV/ha/corte, en cada nivel de fertilización, a medida que las frecuencias de defoliación fueron más espaciadas (fig.49a).
El PLL se caracterizó por presentar gran similitud, en sentido general, con las tendencias encontradas en el PPLL. Las concentraciones de PV en las variedades se comportaron similarmente en ambas épocas, pero se observaron contenidos y rangos ligeramente superiores en el PLL.
La significación de todas las interacciones de primer orden posible en las hojas, para las dos épocas, de manera comparativa con la PB, demostró el elevado nivel de respuesta que presentó la PV con los factores evaluados.

Capítulo III-Resultados y Discusión
53
Fertilización x Frecuencia (P<0,05)
Frecuencia
1209060
PV (%
MS)
20
19
18
17
16
15
14
13
12
Fertilización
100
300
500
Frecuencia
1209060
kg P
V/ha
500
400
300
200
100
Fertilización
100
300
500
a b
Fig. 49. Comportamiento de la PV con la frecuencia de corte en las hojas de M. alba en el PLL.
De forma general, los contenidos extremos de PV en las hojas de las variedades oscilaron entre 12,02 y 17,68 %MS en el PPLL; así como entre 11,18 y 19,91 %MS en el PLL, resultados que coinciden numéricamente con los valores de las interacciones Variedad x Frecuencia en ambos períodos, lo que demuestra la mayor variabilidad que crean estos factores en los contenidos de PV con respecto a la fertilización en las hojas.De todos los variables estudiadas en las hojas, la frecuencia de defoliación fue la que ocasionó los mayores cambios en los contenidos de PV.
Estos resultados evidencian la importancia, desde el punto de vista fisiológico y agronómico, que tiene la edad de rebrote, con vistas a la obtención de concentraciones de PV apreciables y diferenciadas a partir de las distintas variedades. -Tallos tiernos
En el PPLL la PV presentó una tendencia al aumento en los niveles de 300 y 500 con respecto a 100 kg N/ha/año, pero una disminución en los contenidos de 500 kg N/ha/año con respecto a la aplicación de 300 kg N; de manera muy similar a lo obtenido en el caso de las hojas en ambos períodos. Los resultados oscilaron entre (4,17-6,69) ±0,22 %MS y la variedad A se caracterizó por presentar los valores más elevados para las dosis de 100 y 300 kg N/ha/año (fig. 50a).
Variedad x Fertilización (P<0,05)
Fertilización (N kg/ha/año)
500300100
PV (%
MS)
7,0
6,5
6,0
5,5
5,0
4,5
4,0
VARIEDAD
A
C
I
T
Fertilización (N kg/ha/año)
500300100
kg P
V/ha
12
10
8
6
4
2
VARIEDAD
A
C
I
T
a b
Fig. 50. Comportamiento de la PV con la fertilización en los tallos tiernos en el PPLL.
Los rendimientos de PV, siempre fueron superiores a los 2 kg/ha/corte con promedio cercano a los 6 kg/ha/corte entre las variedades. Se observó una tendencia al aumento de los rendimientos de PV/ha/corte con el incremento de la fertilización, comportamiento similar al obtenido para todos los indicadores evaluados en este experimento. Los menores rendimientos se obtuvieron en la variedad C, con producciones inferiores a los 5 kg PV/ha/corte (fig. 50b).
La interacción Variedad x Frecuencia mostró valores entre (4,46-6,76) ±0,28 %MS; se observó un comportamientos diferente para las variedades A-I y T-C; estas últimas mostraron una tendencia similar a la observada en el caso de la PB en el mismo período. La producción de PV por unidad de área osciló entre 3 y 11 kg PV/ha/corte, con los mayores valores para la variedad I en la frecuencia de 120 días.

Capítulo III-Resultados y Discusión
54
La interacción Fertilización x Frecuencia se caracterizó por presentar rangos de concentración entre (4,59-6,67) ±0,23 %MS, con una tendencia homogénea para todos los niveles de fertilización (fig. 51a).
Fertilización x Frecuencia (P<0,05)
Frecuencia
1209060
PV (%
MS)
7,0
6,5
6,0
5,5
5,0
4,5
4,0
Fertilización
100
300
500
Frecuencia
1209060
kg P
V/ha
10
9
8
7
6
5
4
Fertilización
100
300
500
a b
Fig. 51. Comportamiento de la PV con la frecuencia de corte en los tallos tiernos de M. alba en el PPLL.
Las mayores producciones de PV/ha/corte en los tallos tiernos se obtuvieron, igualmente, en la frecuencia
más espaciada (fig. 51b). También en el PLL todas las interacciones fueron significativas, y la mayoría de ellas mostraron tendencias
no definidas en su totalidad para las variedades y los niveles de fertilización. El rango de concentración en el efecto de los grupos Variedad y Fertilización fue de (5,67-8,45) ±0,42 %MS,
en la interacción Variedad x Frecuencia entre (5,05-9,00) ±0,47 %MS; y en la combinación de los factores Fertilización y Frecuencia entre (5, 42-7,95) ±0,71 %MS.
Por otra parte los rendimientos de las variedades con la fertilización asociados al PLL oscilaron entre 20-30 kg PV/ha/corte para la aplicación de 100 y 500 kg de N y entre 20-37 kg PV/ha/corte para el nivel de 300 kg N/ha/año.
En cuanto al comportamiento de la frecuencia de corte en la producción de PV/ha/corte en las variedades, se observó un incremento considerable de los rendimientos promedio a los 90 días de rebrote en los tallos tiernos, resultados que coinciden con los mayores aportes y rendimientos/ha/corte de la fracción nitrogenada en los tallos tiernos de A. lebbeck (Francisco, 2002).
Las producciones de PV/ha/corte en cada frecuencia, así como su relación con los niveles de fertilización en los tallos tiernos, se muestra en la fig. 52. El nivel de 100 kg N/ha/año presentó los resultados más bajos (10-20 kg PV/ha/corte).
Al igual que en otros casos los rendimientos fueron inferiores en la menor frecuencia con respecto a la de 90 días, ya que a medida que aumentó el espaciamiento entre cortes las plantas se desarrollaron progresivamente y produjeron mayores cantidades de tallos lignificados, a medida que aumentó la maduración de los vástagos. Por ese motivo los aportes por unidad de área decrecieron de forma marcada.
La mayor producción de PV/ha/corte para el nivel de 300 kg N/ha/año apoya lo obtenido en el comportamiento de la PV con la fertilización, donde se obtuvieron rendimientos superiores para esta dosis de fertilizante.
En sentido general, los rendimientos de PV/ha/corte en las hojas y los tallos tiernos fueron muy superiores a los obtenidos en el PPLL; esto estuvo determinado por la mayor producción de biomasa en los meses de lluvia, independientemente de la frecuencia de corte empleada.
Luego de analizar particularmente y con profundidad los efectos de cada grupo en el comportamiento de la PV y la PB, como principales componentes de la fracción nitrogenada en esta especie, se llegó a la conclusión de que la frecuencia de corte fue el factor más determinante, dentro de cada parte de la fracción comestible de la planta, en las variaciones de estos compuestos, como producto de las diferencias marcadas que crearon los intervalos de corte en las edades de la fracción comestible.
En segundo lugar, la fertilización orgánica ocasionó menores variaciones, aunque también marcó diferencias discretas en las concentraciones de dichos compuestos. Los mayores cambios con este factor se observaron en la PV y la productividad de cada componente por unidad de área.
Por otra parte, la variedad C presentó un comportamiento atípico en el caso de la PV en la mayoría de los casos.
Al expresar la PV como el porcentaje de PB en las hojas en el PPLL, se observó que más del 70 % de la PB estuvo formado por PV, de acuerdo con lo reportado por Ojeda, Cáceres y Esperance (1991) en una valoración

Capítulo III-Resultados y Discusión
55
realizada sobre el contenido de la fracción proteica verdadera de los pastos y forrajes más utilizados para la conservación en el trópico.
Frecuencia
1209060
kg P
V/ha
40
30
20
10
0
VARIEDAD
A
C
I
T
Frecuencia
1209060
kg P
V/ha
40
30
20
10
0
Fertilización
100
300
500
a b
Fig. 52. Rendimiento de la PV y su relación con la frecuencia de corte en los tallos tiernos de M. alba en el PLL. a) entre las variedades, b) con la fertilización
También se observó un incremento de ésta en función de la disminución de la frecuencia de corte (fig. 53
a,b), tendencia inversa a la observada en la PB con el incremento de la edad de las hojas. Estos resultados indican que en el PPLL, mientras que los contenidos de PB de las variedades disminuyeron
marcadamente con la frecuencia de corte (comportamiento clásico de las arbóreas forrajeras), hubo una mayor fracción nitrogenada que formó parte de la PV.
Un disminución en el contenido de NNP (bases nitrogenadas) desde un 22 % del Nt hasta un 14 % con el aumento de la madurez de las hojas y, por ende, un incremento en la calidad proteica con el aumento de la edad, ha sido obtenido por Duke (2001).
Frecuencia
1209060
PV (P
B)
100
90
80
70
60
VARIEDAD
A
C
I
T
Frecuencia
1209060
PV (P
B)
100
90
80
70
Fertización
100
300
500
a b
Fig. 53. Comportamiento de la PV (%PB) con la frecuencia de corte en las hojas en el PPLL. a) en las variedades, b) con la fertilización
El aumento en las dosis de fertilización en las variedades no creó variaciones apreciables en los porcentajes
de PV (%PB) en este período. En el PLL el porcentaje de PV (%PB) estuvo por encima del 80 % en las variedades y la frecuencia. Los
resultados fueron inferiores a los 90 días y ligeramente superiores en los 60 y 120 días, aunque se observó una tendencia de incremento de la calidad proteica con el aumento de la edad de las hojas. Entre los niveles de fertilización y la frecuencia los valores oscilaron entre 74,79 y 94,01 %, y se observó un comportamiento de aumento menos marcado que en el caso del PPLL. En ninguno de los dos períodos evaluados se observó una variación considerable en el comportamiento de las variedades con el incremento de la fertilización en las hojas, sin fluctuaciones considerables en el porcentaje de PV (%PB).
Los porcentajes de PV (%PB) en los tallos comestibles en el PPLL también mostraron un incremento en función de la disminución de la frecuencia de corte. Estos resultados también evidencian que en el PPLL, mientras la concentración de PB en las variedades disminuyó marcadamente con la frecuencia de corte, existió una mayor fracción nitrogenada que estuvo constituida por PV.

Capítulo III-Resultados y Discusión
56
Frecuencia
1209060
PV (P
B)
90
80
70
60
50
40
VARIEDAD
A
C
I
T
Frecuencia
1209060
PV (P
B)
80
70
60
50
40
Fertización
100
300
500
a b
Fig. 54. Comportamiento de la PV (%PB) con la frecuencia de corte en los tallos tiernos en PPLL. a) en las variedades, b) con la fertilización
En el PLL el porcentaje de PV (%PB) en los tallos tiernos osciló entre 70,58-96,50 % en las variedades con la
frecuencia. Los mayores valores se obtuvieron en la frecuencia de 120 días, al igual que ocurrió en las hojas. Los valores entre los niveles de fertilización y la frecuencia fluctuaron entre 68,00-88,62 %, con el aumento de la PV (%PB) a medida que disminuyó la frecuencia de corte.
El mayor cúmulo de interacciones en el caso de la PV con respecto a la PB en ambos períodos pone de manifiesto la mayor significación biológica de este indicador en el metabolismo del N en las hojas y los tallos tiernos en las variedades. 3.2.3. Fracción unida a la FND -Hojas
Los contenidos de N-FND en el PPLL no presentaron variación significativa con el aumento de las dosis de fertilización (fig.55a); esto demuestra la particularidad de la fracción fijada a la pared celular y la relación estrecha que tuvo ésta con el comportamiento del Nt.
0
1
2
3
N-FND (%MS)
100 300 500
N kg/ha/año
abb b
a
0
1
2
3
N-FND (%MS)
C I T A
Variedad
a b Fig. 55. Comportamiento del N-FND en las hojas de M. alba en el PPLL. a) con la fertilización, b)
entre las variedades a, b Valores con superíndices desiguales difieren estadísticamente mediante la dócima de Student-
Newman-Keuls a P<0,05 Por su parte las variedades presentaron diferencias significativas en el contenido de N-FND, se destacaron
las variedades A y C por presentar los tenores más elevados de este indicador (fig. 55b). En cambio, se obtuvieron diferencias significativas, con la disminución de la frecuencia de defoliación, a favor
del intervalo más pequeño de corte para ambos períodos. En este sentido, se observó una disminución marcada en la concentración del N-FND con el incremento de la edad de las hojas (fig. 56).
El comportamiento análogo entre el N-FND y el Nt con la frecuencia de defoliación en los dos períodos, fue similar a la tendencia observada con el factor Fertilización en el PPLL.

Capítulo III-Resultados y Discusión
57
a
ac ab b b
0
1
2
3
N-FND (%MS)
60 90 120
Frecuencia
PPLL
PLL
Fig. 56. Comportamiento del N-FND con la frecuencia de corte en las hojas de M. alba. a, b, c Valores para una misma época con superíndices desiguales difieren
estadísticamente mediante la dócima de Student-Newman-Keuls a P<0,05
Por otra parte, la interacción Variedad x Fertilización en el PLL fue significativa, en este sentido, se observaron valores que oscilaron entre (1,53-2,34) ±0,10 %MS, sin una tendencia definida entre las variedades.
Contenidos similares de N-FND han sido informados por Schmidek et al. (2002), al estudiar la composición de las hojas de tres clones brasileños de morera: FMSM (3,14 %MS), Miura (3,01%MS) y FM-86 (2,83 %MS) a los 90 días de rebrote.
La mayor cantidad de N-FND se obtuvo en la frecuencia de 60 días, intervalo de corte que propicia un material vegetal más joven, con contenidos de Nt superiores al exhibido por las otras frecuencias; quizás por eso, el N-FND se haya concentrado, ya que la FND no presentó variaciones significativas con la disminución de la frecuencia de corte (datos inéditos). -Tallos tiernos
El contenido de N-FND estuvo regido por interacciones significativas entre todos los pares de factores estudiados. La interacción Variedad x Fertilización en el PPLL se caracterizó por no presentar una tendencia fija para las variedades; el rango osciló entre (0,48-0,77) ±0,05 %MS, con variabilidad para las tres dosis nitrogenadas.
La interacción Variedad x Frecuencia se caracterizó por presentar un comportamiento muy similar entre las variedades para los distintos tiempos de corte, el rango estuvo enmarcado entre (0,44-1,05) ±0,06 %MS (fig. 57a).
La mayor cantidad de N-FND se obtuvo en la frecuencia de 60 días, igual que en el caso de las hojas; el comportamiento del N-FND en los tallos tiernos se explica de manera análoga al caso anterior, solamente que los valores absolutos fueron inferiores y que los menores tenores se encontraron en la frecuencia de 120 días.
La interacción Fertilización x Frecuencia presentó valores entre (0,43-0,99) ±0,14 %MS, con un comportamiento similar al obtenido en las hojas, con la excepción de 100 kg N/ha/año, el cual tuvo una tendencia diferente a la de los niveles de 300 y 500 kg N/ha/año. En esta interacción, los valores más elevados se obtuvieron también para la frecuencia de 60 días (fig. 57b).
En el PLL las variedades presentaron un mismo comportamiento con la frecuencia de corte, caracterizado por la disminución de los contenidos de N-FND con el aumento de los espaciamientos entre cortes; la interacción Variedad x Frecuencia no fue significativa.
La interacción Variedad x Fertilización presentó pocas diferencias en los tenores de N-FND al aumentar las dosis de N, los valores oscilaron entre (0,5-1,16) ±0,09 %MS.
La interacción Fertilización x Frecuencia tampoco mostró variaciones considerables y el rango osciló entre (0,68-1,11) ±0,07 %MS con los menores valores a los 90 días; excepto para el caso de los 500 kg N/ha/año, que fue 120 días.
En sentido general, las hojas presentaron aproximadamente tres veces más N-FND que los tallos tiernos, teniendo en cuenta que en los foliolos se triplicó la cantidad de PB con respecto a los últimos.
El factor Frecuencia de corte fue decisivo en el comportamiento de las tendencias observadas en las variedades, unido a las diferencias encontradas entre las hojas y los tallos tiernos, así como las épocas.

Capítulo III-Resultados y Discusión
58
La Fertilización como factor presentó menor importancia, mostrando mayor heterogeneidad entre los niveles estudiados y tendencias menos definidas que en el caso de la Frecuencia.
Las variedades se comportaron de forma similar; aunque se observaron comportamientos atípicos al resto, por parte de la variedad A; en algunos casos.
Variedad x Frecuencia (P<0,05)
Frecuencia
1209060
% N
-FN
D
1,1
1,0
,9
,8
,7
,6
,5
,4
VARIEDAD
A
C
I
T
Fertilización x Frecuencia (P<0,05)
Frecuencia
1209060
% N
-FN
D
1,1
1,0
,9
,8
,7
,6
,5
,4
,3
Fertilización
100
300
500
a b
Fig. 57. Comportamiento del N-FND en los tallos tiernos de M. alba en el PPLL. a) con la fertilización, b) entre las variedades
3.2.4. Fracción unida a la FAD -Hojas
El análisis del N-FAD en el PPLL develó solamente un efecto de grupo significativo; este fue la interacción Variedad x Frecuencia, la cual se caracterizó por presentar una tendencia variable entre las variedades con el aumento en los intervalos de corte. Los valores oscilaron entre 1,10 %MS en la variedad A (90 días) y 2,20 %MS en la variedad C a los 60 días (fig. 58).
Variedad x Frecuencia (P<0,05)
Frecuencia
1209060
N-F
AD (%
MS)
2,4
2,2
2,0
1,8
1,6
1,4
1,2
1,0
VARIEDAD
A
C
I
T
Fig. 58. Comportamiento de N-FAD con la frecuencia de corte en las hojas de cuatro variedades de M. alba en el
PPLL. Por su parte, en este período se observó una disminución significativa en la concentración de N-FAD con el incremento de la dosis nitrogenada; en cambio, en el PPLL se observó un comportamiento significativamente inverso (fig. 59).
Por otro lado, en el PLL se encontraron diferencias significativas entre las concentraciones de N-FAD con la disminución de la frecuencia de defoliación, así como una gran similitud en los contenidos de este indicador entre las diferentes variedades (fig. 60).

Capítulo III-Resultados y Discusión
59
a
b
b a b ab
0
0,5
1
1,5
2
2,5
N-FAD (%MS)
100 300 500
N Kg/ha/año
PPLLPLL
Fig. 59. Comportamiento del N-FAD con la fertilización en las hojas de M. alba. a, b Valores con superíndices desiguales difieren estadísticamente mediante
la dócima de Student-Newman-Keuls a P<0,05
ba a
0
0,5
1
1,5
N-FAD (%MS)
60 90 120
Frecuencia
0
0,5
1
1,5
2
N-FAD (%MS)
C I T A
Variedad
a b Fig. 60. Comportamiento del N-FAD en las hojas de M. alba en el PLL. a) con la frecuencia de corte,
b) entre las variedades. a, b Valores con superíndices desiguales difieren estadísticamente mediante la dócima de Student-
Newman-Keuls a P<0,05 -Tallos tiernos
El análisis de la concentración de N-FAD reveló interacciones significativas para casi todos los pares de grupos estudiados en los dos períodos. En el PPLL la interacción Variedad x Fertilización se caracterizó por no presentar una tendencia homogénea entre las variedades. El rango de valores estuvo enmarcado entre 0,37 %MS en la variedad A con 500 kg N/ha/año y 0,69 %MS con 300 kg N/ha/año, en el caso de la variedad C.
La interacción Fertilización x Frecuencia no tuvo una tendencia fija y los valores extremos se encontraron en las frecuencias de 60 y 90 días.
El efecto de los grupos Variedad y Frecuencia sobre los contenidos de N-FAD en el PPLL no fue significativo. La interacción Variedad x Fertilización en el PLL mostró pocas variaciones en el comportamiento de las
variedades con el aumento de las dosis empleadas. Las mayores diferencias se encontraron en la variedad A en los niveles de 100 (0,86 %MS) y 500 kg N/ha/año (0,31 %MS). Al igual que en el PPLL, los valores límites de la interacción los aportó la variedad A, lo cual demostró que ésta presenta un nivel de respuesta más elevado en cuanto al N-FAD con la fertilización que el resto de las variedades.
El efecto de los grupos Variedad y Frecuencia sobre los contenidos de N-FAD en el PLL fue variable, con una tendencia no definida con el aumento de la frecuencia para ninguna de las variedades y mayor heterogeneidad a los 60 días. Los valores oscilaron entre 0,81 %MS para la variedad T (60 días) y 1,48 %MS en la variedad I a los 60 días.
La interacción Fertilización x Frecuencia no presentó tendencia homogénea, los valores extremos se observaron en la frecuencia de 120 días.

Capítulo III-Resultados y Discusión
60
De manera general, el contenido de N-FAD en los tallos tiernos no presentó un comportamiento definido con los factores estudiados y no se observaron variaciones considerables con el incremento de la fertilización. Por otra parte, la variedad A se destacó por su elevada variación en la fracción de N-FAD en ambos períodos.
La mayor variabilidad en los tallos tiernos en las interacciones con respecto a las hojas, evidencia el mayor estado de transformación estructural que tiene lugar en esa parte de la planta.
Se puede concluir expresando que la cantidad de N poco disponible en las hojas, desde el punto de vista nutricional, fue superior al 1 %MS, lo cual correspondió a más del 30 % del Nt.
Este indicador presentó pocas variaciones con la época y los comportamientos mejores definidos se observaron entre las variedades y la fertilización. 3.2.5. Dinámica de los compuestos nitrogenados
Resumiendo los resultados obtenidos, a partir de las determinaciones individuales de los componentes de la fracción nitrogenada, se pudo llegar a la conclusión que en las hojas se observó, en buena medida, una dinámica clara y coherente entre los componentes nitrogenados, aunque por su parte los tallos tiernos presentaron mayor heterogeneidad y tendencias menos homogéneas, particularmente en el PLL. -Parte de la fracción comestible
En todos los componentes nitrogenados se encontraron diferencias numéricas marcadas entre las hojas y los tallos tiernos, fundamentalmente en el caso de la PB y la PV. Este resultado es común en la mayoría de las plantas superiores, en las cuales estos compuestos, presentes en los foliolos, desempeñan funciones vitales para su desarrollo. -Frecuencia de corte
Analizando las transformaciones causadas por la frecuencia de defoliación, se debe señalar que al disminuir los intervalos de corte se observó un decrecimiento en los contenidos de Nt, tanto en las hojas como en los tallos tiernos, aunque en el PLL no se encontraron diferencias significativas.
El NP presentó los mayores valores a los 60 y 120 días en ambas épocas. Por su parte, el N-FND mostró diferencias significativas a favor de los 60 días. El N-FAD se incrementó al aumentar los intervalos en el PLL, con variaciones distintivas en los contenidos de las variedades en el PPLL.
Desde el punto de vista fenológico, a medida que aumentó la madurez de las hojas decreció la concentración de Nt y de N-FND en ambas época, pero hubo un aumento significativo a partir de los 90 días en la concentración de la PV y en los porcentajes de PV (%PB), por lo que aumentó la calidad de la fracción proteica.
También se observó una mayor disponibilidad de N desde el punto de vista nutricional, por su menor unión con la FND a medida que disminuyó la frecuencia de defoliación.
Los tallos tiernos en ambos períodos presentaron un comportamiento similar al de las hojas, aunque se observó una mayor variabilidad de las tendencias en el PLL.
En el PPLL los tallos mostraron una disminución marcada en la concentración de Nt con la frecuencia de corte.
Por otra parte, el aumento en los porcentajes de PV (%PB), así como la disminución en el contenido de N-FND, fueron las características fundamentales en los tallos de mayor edad.
En el PLL, si bien los resultados de las concentraciones de Nt y NP fueron similares al período anterior, los porcentajes de PV (%PB) tendieron a ser ligeramente superiores a los del PPLL. Los contenidos de N-FND tendieron a disminuir con la edad de los tallos no lignificados. -Fertilización
Las crecientes dosis de N no provocaron incrementos en las concentraciones de Nt en las hojas en ambas épocas; en cambio, en los tallos tiernos se observó un aumento significativo en el contenido de Nt en el PPLL.
Por otro lado, la concentración de NP, al menos en dos variedades por época luego de aumentar abruptamente hasta los 300 kg N/ha/año, disminuyó con la aplicación de 500 kg N/ha/año para ambos períodos en hojas y tallos; se observándose una estabilización en la calidad proteica de las hojas, al no variar considerablemente los porcentajes de PV (%PB) a medida que se incrementaron las dosis nitrogenadas.
Adicionalmente a lo obtenido en los indicadores anteriores, con el aumento de la fertilización se observaron pocas variaciones en el contenido de N-FND y un comportamiento inverso con la época en el caso del N-FAD.
Los tallos tiernos presentaron una dinámica muy parecida a la encontrada en las hojas.

Capítulo III-Resultados y Discusión
61
La disminución común en el contenido de NP en ambos períodos a partir de los 300 kg N/ha/año, en hojas y tallos tiernos, quizás se explique por la imposibilidad de continuar convirtiendo el NNP asimilado por las raíces en PV en las variedades que presentaron ese comportamiento. -Variedad
En el caso de la PB las variedades no presentaron diferencias tanto en las hojas como en los tallos tiernos. En cuanto a la PV, las variedades C y A exhibieron tendencias y niveles diferenciados en comparación con el resto; resultados análogos a los obtenidos en el caso del N-FND -Época
Este factor presentó poca influencia en los contenidos nitrogenados, principalmente en la PB y la PV. La fracción de N-FD tuvo ligeras variaciones con los períodos evaluados. 3.2.6. Consideraciones sobre el efecto de los factores evaluados
Globalmente, los contenidos de cada indicador presentaron diferencias sustanciales entre las hojas y los tallos tiernos, fundamentalmente en los compuestos que conformaron la fracción nitrogenada; en ese orden la edad de rebrote fue la variable con la mayor influencia en los resultados, creando tendencias definidas que caracterizaron las diferentes edades del material. Seguidamente, las épocas marcaron rangos de valores diferenciados, sobre todo en el caso de los metabolitos secundarios (a excepción de las cumarinas), los cuales presentan una mayor variación con los factores externos del medio; y en último lugar, con influencias numéricas más discretas, los factores variedad y fertilización, que de manera integrada definieron diferencias entre las variedades en el comportamiento de los metabolitos secundarios y de algunos indicadores nitrogenados como la PV y la fracción unida a la FD.

Conclusiones
62
Conclusiones
• Las variedades Cubana, Indonesia, Tigriada y Acorazonada contienen, en las hojas y en los tallos tiernos, cantidades apreciables de fenoles, flavonoides, esteroides, cumarinas y CHS, los que se encuentran presentes en las edades de rebrote de 60, 90 y 120 días, en los niveles de fertilización de 100, 300 y 500 kg N/ha/año y en las dos épocas del año.
• Cada grupo de metabolito secundario presentó un comportamiento particular y distintivo con los factores evaluados, lo que estuvo estrechamente relacionado con su actividad y con la posible función biológica que desempeñan en cada parte de la planta.
• En el nivel de 300 kg N/ha/año se observaron las menores diferencias en el metabolismo secundario entre las variedades.
• La variedad Cubana, desde el punto de vista fitoquímico, presentó en las hojas características distintivas en la concentración y las tendencias de los FT, las cumarinas, los CHS, la PV y los aportes de los indicadores evaluados por unidad de área, fundamentalmente en el PPLL.
• Las concentraciones de los metabolitos secundarios y la fracción nitrogenada en las partes de la fracción comestible de M. alba se afectan fundamentalmente por la edad de rebrote; los factores fertilización, variedad y época crearon variaciones numéricas más discretas entre los tratamientos.
• Las dosis crecientes de fertilizante orgánico no incrementan, de manera general, los contenidos nitrogenados totales, pero sí provocan variaciones en la concentración de la PV y la fracción enlazada a la porción fibrosa de la biomasa comestible.

Recomendaciones
63
Recomendaciones
• Aislar los compuestos mayoritarios de la fracción fenólica, con la finalidad de realizar evaluaciones nutricionales in vitro.
• Evaluar el impacto fisiológico y nutricional de los metabolitos de interés en animales monogástricos.
• Determinar la influencia de factores tales como fenología y fertilización química en el metabolismo secundario y en la fracción nitrogenada.
• Estudiar la dinámica de transformación de estos compuestos al emplear tecnologías de conservación, especialmente en la confección de harinas.
• Emplear esta metodología de investigación en el estudio de la composición fitoquímica de las plantas arbóreas forrajeras.

Bibliografía
64
Bibliografía
ADAMS, D.O. & HARBERTSON, J.F. 1999. Use of alkaline phosphatase for the analysis of tannins in grapes and red wine. Am. J. Enol. Vitic. 50:247-252
AERTS, R.J.; BARRY, T.N.; MC NABB, W.C. 1999. Polyphenols and agriculture: beneficial effect of proanthocyanidins in forages. Agriculture, Ecosystems and Environment. 75:1-12
AHN, J.; ELLIOTT, R. & NORTON, B. 1997. Oven drying improves the nutritional value of Calliandra calothyrsus and Gliricidia sepium as supplements for sheep given low. Journal of Science and Agriculture. 75:503-510
ALBERT, ANAYANSI & CONTRERAS, F. 2002. Utilización de la morera (Morus alba) en la alimentación de cuyes, en la localidad de Topes de Collantes. [cd room]. Memorias V Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical” y II Reunión Regional de Morera. EEPF “Indio Hatuey”. Matanzas, Cuba
ABDULRAZAK, S.A.; FUJIHARA, T.; ONDIEK, J.K. & ØRSKOV, E.R. 2000. Nutritive evaluation of some Acacia tree leaves from Kenya. Animal Feed Science and Technology. 85:89-98
ALFONSO, M., FERNÁNDEZ, L.; GONZÁLEZ, N. & AVILÉS, R. 2000. La Achira (Canna edulis Ker.) y su potencialidad en el control de plagas. Ponencia XII Forum de Ciencia y Técnica. INIFAT. Ciudad de La Habana, Cuba. 11 p.
ALMEIDA, J.E. DE & FONSECA, TAMARA. 2002. Mulberry germplasm and cultivation in Brazil. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 73-95
ANON. 2003a. Análisis de los ácidos nucleicos en la clasificación bacteriana. [en línea]. Disponible en: http://www.ffyb.uba.ar./Programa/P2000-2/MICROSUP.htlm. [Consulta: Agosto 2003].
ANON. 2003b. Domesticación y diversificación de Phaseolus en Mesoamérica y Los Andes. [en línea]. Disponible en: http://www.redpar-fpolar.info.ve/fagro/v24-2/m242a001.htlm. [Consulta: Agosto 2003].
ANON. 2003c. Efecto de los esteroles y antibióticos en membranas. [en línea]. Disponible en: http://www.fc.uaem.mx/PERSONAL/proftc/bioqui/nina/ninalin2.htlm. [Consulta: Agosto 2003].
ANURAGA, M.; DURAZA, P.; HILL, M.J. & LOVETT, J.V. 1993. Phenolic in nature. Australian Journal of Agricultural Research. 44:1667-1681
AOAC. 1990. Oficial methods of analysis. 15th ed. Association of Official Agricultural Chemistry. Washington, D. C., USA
ARANGO, A.A.; ADOGLA-BESSA, T.; OMPHILE, U.J. & TSHIRELETSO, A. 2000. Significance of browses in the nutrition of Tswana goats. Arch Zootecnia. 49:469-480
ARIAS, E. & SÁNCHEZ, M.D. 2002. La morera como frutal. [cd room]. Conferencia V Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical” y II Reunión Regional de Morera. EEPF “Indio Hatuey”. Matanzas, Cuba
ASHOK, K.J.; VINCENT, R.M. & NESSLER, C.L. 2000. Molecular characterization of a hydroxymethylglutaryl-CoA reductase gene from mulberry (Morus alba L.). Plant Molecular Biology. 42:559-569
AZCÓN-BIETO, J. & TALÓN, M. 2000. Fundamentos de fisiología vegetal. Interamericana Mc. Graw-Hill. 555 p.
BAKER, M.E.; MEDLOCK, K.L. & SHEEHAN, D.M. 1998. Flavonoids inhibit estrogen binding to rat alpha-fetoprotein. Proceedings of the Society for Experimental Biology and Medicine. 217:317-321
BEN SALEM, H.; NEFZAOUI, A.; BENT SALEM, L. & TISSERAND, J.L. 1999a. Intake digestibility, urinary excretion of purine derivates and growth by sheep given fresh, air-dried or polyethylene glycol-treated of foliage of Acacia cianophylla Lindl. Animal Feed Science and Technology. 98:297-311
BEN SALEM, H.; NEFZAOUI, A.; BENT SALEM, L. & TISSERAND, J.L. 1999b. Different means of administering polyethylene glycol to sheep: effect on the nutritive value of Acacia cianophylla Lindl. foliage. Animal Feed Science and Technology. 68:809-818
BEN SALEM, H.; NEFZAOUI, A.; BENT SALEM, L. & TISSERAND, J.L. 2000. Deactivation of condensed tannin in feed block. Effect on feed intake, diet digestibility, nitrogen balance, microbial synthesis and growth by sheep. Livestock Production Science. 64:51-60
BENAVIDES, J.E. 1994. La investigación en árboles forrajeros. En: Árboles y arbustos forrajeros en América Central. (Ed. Benavides, J.E). Serie técnica, Informe técnico No. 236. CATIE. Turrialba, Costa Rica. Vol. 1, p. 3-21
BENAVIDES, J.E. 1996. Manejo y utilización de la morera (Morus alba) como forraje. Agroforestería en las Américas. 2 (7):27-30

Bibliografía
65
BENAVIDES, J.E. 1999. Utilización de la morera en los sistemas de producción animal. En: Agroforestería para la producción animal en América Latina. Estudio FAO Producción y Sanidad Animal No. 143. FAO, Roma. p. 279
BENAVIDES, J.E. 2000. La morera, un forraje de alto valor nutricional para la alimentación animal en el trópico. Pastos y Forrajes. 23:1
BENAVIDES, J.E. 2002. Utilization of mulberry in animal production systems. In: Mulberry for animal production. Animal Production and Health Paper No. 147. FAO, Rome. p. 291-327
BENAVIDES, J.E.; ESQUIVEL, J. & LOZANO, E. 1995. Módulos agroforestales con cabras para la producción de leche. Guía técnica. Manual técnico No. 18. CATIE. Turrialba, Costa Rica. 56 p.
BENAVIDES, J.E.; LACHAUX, M. & FUENTES, M. 1994. Efecto de la aplicación de estiércol de cabra en el suelo sobre la calidad y producción de biomasa de morera (Morus sp.). En: Árboles y arbustos forrajeros en América Central. (Ed. Benavides, J.E.), CATIE. Turrialba, Costa Rica. Vol. 2, p. 495-514
BERENBAUM, M.R. 1991. Coumarins. In: Herbivores: Their interaction with secondary plant metabolites Vol. I. The chemical participant. (Eds. Rosenthal, G.A. & Berenbaum, M.R.). p. 221-249
BING, Y.; JIANXIN, L. & JUN, Y. 2001. Nutritional evaluation of mulberry leaves as feed for ruminants. In: Mulberry for animal feeding in China. (Eds. Jian, L.; Yuyin, C.; Sánchez, M. & Xingmeng, L.). Hangzhou, China. 75 p.
BLANCO, R. 1992. Distancia de siembra y altura de corte en la producción y calidad del forraje de morera (Morus sp.) en el parcelamiento cuyuta sscuintla, Guatemala. Universidad de Guatemala, Guatemala. 15 p.
BOKANG, L.; HUIYIN, C. & GOWCHIN, Y. 1999. Antioxidant activity of the methanolic extract from various traditional edible plant. Journal of the Chinese Agricultural Chemical Society. 37 (1):105-116
BOSCHINI, C. 2002a. Nutritional quality of mulberry cultivated for ruminant feeding. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 171-182
BOSCHINI, C. 2002b. Establishment and management of mulberry for intensive forage production. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 115-122
BOSCHINI, C.; DORMOND, H. & CASTRO, A. 1998. Producción de biomasa de la morera (Morus alba) en la Meseta Central de Costa Rica. Agronomía Mesoamericana. 9 (2):31-40
BOSCHINI, C.; DORMOND, H. & CASTRO, A. 1999. Respuesta de la morera (Morus alba) a la fertilización nitrogenada, dos distancias de siembra y a la defoliación. Agronomía Mesoamericana. 10 (2):7-16
BROOKS, R. & OWEN-SMITH, N. 1994. Plant defenses against mammalian herbivores: are juvenile Acacia more heavily defended than mature trees ?. Bothalia. 24 (2):211-215
CAPPELLOZZA, L. 2002. Mulberry germplasm resources in Italy. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 97-101
CASSIDY, A.; HANLEY, B. & LAMUELA-RAVENTOS, ROSA. 2000. Isoflavones lignans, and stilbenes-origins, metabolism and potential importance to human health. Journal of the Science of Food Agriculture. 80:1044-1062
CAYGIL, J.C. & MUELLER-HARVEY, IRENE. 1999. Tannins: Their nature and biological significance. In: Secondary plant products antinutritional and beneficial actions in animal feeding. Nottingham University Press. Nottingham, U. K. p. 17-37
CHAPPELL, J. 1995. The biochemistry and molecular biology of isoprenoid metabolism. Plant Physiol. 107:1-6 CHAPPELL, J.; VONLANKEN, C. & VOGELI, U. 1991. Elicitor inducible 3-hydroxy-3-methylglutaryl coenzyme
A reductase activity is required for sesquiterpene accumulation in tabacco cell suspension cultures. Plant Physiol. 97:693-698
CHAPPELL, J.; WAALF, F.; PROULX, J.; CUELLAR, R. & SAUNDERS, C. 1995. Is the reaction catalyzed by 3-hydroxy-3-methylglutaryl coenzyme A reductase gene in response to Phytophthora infestans and to its elicitor Arachidonic acid. Plant Cell. 4:1333-1344
CHEECKE, P. R. 1995. Endogenous toxins and mycotoxins in forage grasses and their effect on livestock. J. Anim. Sci. 73:909-918
CHEYNIER, V.; BOUGUET, J.M.; LEROUX, E.; GUYOT, S. & RIGAUD, J. 1999. Size separation of condensed tannins by normal-phase high-performance liquid chromatography. Meth Enzymol. 299:178-184
CHUANBU, Z. 2000. Determination of flavonol in mulberry leaves and its health drink. Food Science and Technology. 2:53
CHUL-YOUNG, K.; SI-HYUN, K.; GI-WHA, J.; CHUN-SIK, C. & BAKK-WANG, K. 2000. Galactolipids from Mori folium and their hypoglycemic effect. Korean Journal of Pharmacognosy. 31 (1):95-100
CHUNLIAN, L.; DONSHENG, L. & ERJUN, L. 1999. The content polyphenol and anthelmintic material in different mulberry leaves. Journal of Anhui Agriculture Science. 27 (4):356-402

Bibliografía
66
CIFUENTES, C. A. & HAM-KIM, M. 1998. Cartilla de Sericultura. C.D.T.S., Colombia. p. 9-25 CIFUENTES, C. A. & KEE-WOOK, S. 1998. Manual técnico de sericultura: Cultivo de la morera y cría del
gusano de seda en el trópico. Convenio SENA-CDTS, Colombia. 438 p. CLAVERO, T. 1998. Acacia mangium. Leguminosa promisoria para suelos ácidos. Cuadernos técnicos. Serie:
Árboles forrajeros. Facultad de Agronomía, Luz. Maracaibo, Venezuela. 6 p. COLECTIVO DE AUTORES. 2000. Tablas de valor nutritivo y requerimientos para el ganado bovino. Pastos y
Forrajes. 23 (2):105-122 COLECTIVO DE AUTORES. 2003. Impacto de los árboles, arbustos y otras leguminosas en la ecología ruminal
de animales que consumen dietas fibrosas de baja calidad. En: Boletín de divulgación de resultados y noticias del trabajo científico. MES. Ciudad de La Habana, Cuba. No. 1, p. 8-9
DALLZELL, S.A. & KERVEN, G.L. 1998. A rapid method for the measurement of Leucaena spp. proanthocyanidins by the proanthocyanidins (butanol/HCl) assay. J. Sci. Food Agric. 78:405-416
DAS, C.; SENGUPTA, T.; CHATTOPADHYAY, S.; SETUA, M.; DAS, N.K. & SARATCHANDRA, B. 2002. Involvement of kinetin and spermidine in controlling salinity stress in mulberry (Morus alba L. Cv. S1). [en línea]. Disponible en: http://www.zfr.pan.krakow.pl/ang/app/2002no1/4.html. [Consulta: Agosto 2003].
DATTA, R.K. 2002. Mulberry cultivation and utilization in India. In: Mulberry for animal production. Animal Production and Health Paper No. 147. FAO, Rome. p. 45-62
DATTA, R.K.; SARKA, A.; RAMA MOHAN RAO, P. & SINGHVI, W.R. 2002. Utilization of mulberry as animal fodder in India. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 183-188
DELGADO, E.J. 1998. Factores antinutricionales. Curso de Fisiología digestiva. ICA. La Habana, Cuba. p. 2-5
DESHMUKH, S.V.; PATHAK, N.V. & TAKALIKAR, D.A. 1993. Nutritional effect of mulberry (Morus alba) leaves and as sole ration of adult rabbits. World Rabbit Sci. 1 (2): 67-69
DOI, K.; KOJIMA, T.; HARADA, M.; HORIGUCHI, Y. & FUJIMOTO, Y. 2000. Effect of various fractions extracted from mulberry leaves on lipid metabolism in rabbit feed cholesterol diet. Journal of the Japanese Society of Nutrition and Food Science. 52 (2):85-90
DOMÍNGUEZ, A.; TELLES, ENIDIA & REVILLA, J. 2001. Comportamiento inicial de dos especies de morera en fase de establecimiento. Pastos y Forrajes 24 (2):147
DUKE, J.A. 2001. Morus alba (L.). [en línea]. Disponible en: http://newcrop.hort.purdue. edu/newcrop/duke-energy [Consulta: Diciembre 2001].
EL KAZZAZ, A.A.K. & NAZIF, N.M. 2000. Flavonoids content of in vitro micropropagation of mulberry (Morus alba L.). Bulletin of the National Research Center Cairo. 25 (1):13-22
ESPINOZA, E. & BENAVIDES, J.E. 1996. Efecto del sitio y la fertilización nitrogenada sobre la producción y calidad del forraje de tres variedades de morera (Morus alba L.). Agroforestería en las Américas. 3 (11):24-27
EUN-SUN, K.; SUNG-JEAN, P.; EUN-JU, L.; BAK-KWANG, K.; HOON, H. & BONG-JIN, L. 1999. Purification and characterization of Moran 20K from Morus alba. Arch. Pharm. Res. 22 (1): 9-12
EZENWA, I. & KITAHARA, N. 2001. Dry matter degradation of the leaves of eleven varieties in the rumen of cattle and changes during the grazing season. Grassland Science. 47 (3):245-250
FAO. 1990. Sericulture training manual. FAO. Agricultural Services Bulletin, No. 80, Rome. 117 p. FENG, Q.; KANO, Y. & XINSHENG, Y. 1997. Elucidation of the bioactive constituents in traditional
Chinese medicine “Mori Cortex”. Advances in plant glycosides, chemistry and biology. Proceedings of the International Symposium on Plant Glycosides. Kunming, China, p. 281
FERNÁNDEZ, N.; CHACÍN, E.; GARCÍA, C.; ALASTRE, N.; LEAL, F. & FORSTER, C.F. 1996. The use of seed pods from Albizia lebbeck for the removal of alkyl bencene sulphonates from aqueos solution. Process Biochemistry. 31 (4):383-387
FERRARI, E.; MONACELLI, B. & MESSANA, I. 1999. Comparison between in vitro and in vivo metabolite production of Morus nigra. Planta Medica. 65 (1):85-87
FERRARI, F.; MONACHE, F.D.; SUÁREZ, A.I. & COMPAGNONE, R.S. 2000. Multicaulisin, a new Diels-Alder type Adduct from Morus multicaulis. Phytoterapia. 71 (2):213-215
FERRER, F.; HIGUERA, A.; CASTRO, C.; GARCÍA, B.; LÓPEZ, Y.; SOTO, R.; CARRILLO, G.; MOGOLLÓN, M. & URDANETA, J. 1996. Efecto de la altura y tiempo de corte sobre la digestibilidad in vitro, el valor energético y el contenido de carbohidratos estructurales y no estructurales en hojas y tallos de las tres variedades forrajeras de quinchoncho (Cajanus cajan (L.) Mill sp.). Interciencia. 21 (1):42-45
FICKEL, J.; PITRA, C.; JOEST, B.A. & HOFMANN, P.R. 1999. A novel method to evaluate the relative tannin-binding capacities of salivary proteins. Comp. Biochem. Physiol. 122: 225-229
FISCHER, G. & LUDDERS, P. 1997. Developmental changes of carbohydrates in Cape Grooseberry (Physalis peruviana L.) fruits in relation to the calyx and the leaves. Agronomía Colombiana. 24 (2):95-107

Bibliografía
67
FOO, L.Y.; LU, Y.; MCNABB, W.C.; WAGHORN, G. & ULYATT, M.J. 1997. Proanthocyanidins from Lotus pedunculatus. Phytochemistry. 45:1689-1696
FRANCISCO, GERALDINE. 2002. Manejo de las defoliaciones de Albizia lebbeck para la producción de biomasa. Tesis presentada en opción al Título de Máster en Pastos y Forrajes. EEPF “Indio Hatuey”. Matanzas, Cuba. 80 p.
FUJUN, C.; NAKASHIMA, N. & KIMURA, I. 1995. Otentiating effect on pilocarpine-induced saliva secretion, by extracts and N-containing sugars derived from mulberry leaves, in streptozocin-diabetic mice. Biological and Pharmaceutical Bulletin. 18 (12):1676-1680
GABRIELA-ANCA, CAMELIA.; DEAN, K. & WILEY, D. 1997. Phytoestrogens and floral development in dioecious Maclura pomifera (Raf.) Schoneid. and Morus rubra (L.) (Moraceae). Plant Science. 130:27-40
GALINDO, J.; CASTILLO, E.; ALDAMA, A.M.; MARRERO, Y.; GARCÍA, C. & MARTÍNEZ, P. 1998. Efecto del pastoreo de Leucaena leucocephala en todo el área con gramíneas en la población microbiana ruminal. En: Memorias III Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería”. EEPF “Indio Hatuey”. Matanzas, Cuba. p. 237-239
GALINDO, W.; ROSALES, M., MURGUEITIO, E. & LARRAHONDO, J. 1989. Sustancias antinutricionales en las hojas de árboles forrajeros. Livestock Research for Rural Development. 1 (1):36-41
GARBARINO, J.A. 2003. Metabolismo secundario y quimiotaxonomía de especies chilenas de la familia Scrophylariaceae. [en línea]. Disponible en: http://www.conicyt.cl/cgi-bin/rut.cgi?3540 [Consulta: Agosto 2003].
GARCÍA, D.E.; OJEDA, F. & PÉREZ, GUADALUPE. 2002. Comportamiento fitoquímico de cuatro variedades de Morus alba en suelo Ferralítico Rojo con fertilización. [cd room]. En: Memorias V Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical” y II Reunión Regional de Morera. EEPF “Indio Hatuey”. Matanzas, Cuba
GARCÍA, F.; MEDEROS, M.L.; SALINAS, A. & REYES, J. 2002. Utilización de la harina de hojas de morera peletizadas como sustituto de concentrado en la ceba cunícula. [cd room]. Memorias V Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical” y II Reunión Regional de Morera. EEPF “Indio Hatuey”. Matanzas, Cuba
GARCIA, H.; NYGREN, P. & DESFONTAINES, L. 2001. Dynamics of non-structural carbohydrates and biomass yield in a fodder legume tree under different harvest intensities. Tree Physiol. 21:523
GERASOPOULOS, P. & STAVROULAKIS, G. 1997. Quality characteristcs of four mulberry (Morus sp.) cultivars in the area of Chania, Greece. J. Sci. Food Agric. 73:261-264
GHASEMPOUR, H.R.; ANDERSON, E.M.; GIANELLO, R.D. & GAFF, D.F. 1998. Growth inhibitor effects on protoplasmic drught tolerance and protein synthesis in leaf cells of the resurrection grass Sporobolus stapfianus. In: Plant growth regulation. Kluwer Academic Publishers. Netherlands. p. 179-183
GODBOLE, S.A.; KRISHNA, T.G. & BRATA, C.R. 1994. Changes in protease inhibitory activity from pigeon pea (Cajanus cajan, (L.) J. Mill. sp.) during seed development and germination. Journal of Science Food and Agriculture. 66:497-501
GÓMEZ, G.; QUESADA, S. & NANNE, C.I. 1998. The effect of peach palm (Bactris gasipaes) antinutritional factors in young mice metabolism. Agronomía Costarricense. 22 (2):185-189
GONZÁLEZ, E. & CÁCERES, O. 2002. Valor nutritivo de árboles, arbustos y otros plantas forrajeras para los rumiantes. Pastos y Forrajes. 25 (1):15-20
GONZÁLEZ, E.; DELGADO, DENIA & CÁCERES, O. 1998. Rendimiento, calidad y degradabilidad ruminal potencial de los principales nutrientes en el forraje de morera (Morus alba). En: Memorias III Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería”. EEPF “Indio Hatuey”. Matanzas, Cuba. p. 69-72
GONZÁLEZ, E.; MARTÍN, G.; ALVANELL, E.; CAJAS, G. & ROSAS, N. 2002. Composición nutritiva del forraje de morera (Morus alba var. Tigriada) ante diferentes frecuencias de corte y niveles de fertilización nitrogenada. I. Contenido celular. [cd room] En: Memorias V Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical” y II Reunión regional de Morera. EEPF “Indio Hatuey”. Matanzas, Cuba
GONZÁLEZ, J.; BENAVIDES, J.E.; KASS, MARÍA & ESPERANCE, M. 2000. Evaluación de la calidad nutritiva de la morera (Morus sp.) ensilada, con bovinos de engorde. [cd room]. Memorias I Taller Internacional de Morera “La morera (Morus alba) oportunidades y posibilidades de uso para la alimentación animal”. EEPF “Indio Hatuey”, Matanzas. Cuba
GUTIÉRREZ, Y.I.; MIRANDA, M.; VARONA, N. & RODRÍGUEZ, A.T. 2000. Validación de 2 métodos espectrofotométricos para la cuantificación de taninos y flavonoides (Quercetina) en Psidium guajava (L.). Rev. Cubana Farm. 34 (1):50-55
GUYOT, S.; DOCO, T.; BOUGUET, J.M.; MOUTOUNET, M. & DRILIAU, J.F. 1997. Characterization of highly polymerized procyanidins in cider apple (Malus sylvestris var. Kermerrien) skin and pulp. Phytochemistry. 44:351-357

Bibliografía
68
HAGERMAN, A.E. 1998. Tannin analysis. [en línea]. Disponible en: http://miavx1.edu/hagermae/. [Consulta: Diciembre 1998]
HAGERMAN, A.E.; ZHAO, Y. & JONSON, S. 1997. Methods for determination of condensed and hydrolizable tannins. In: Antinutrients and phytochemicals in food. (Ed. Shanidi, F.), Washington, USA. 230 p.
HAMMERSTONE, J.F.; LAZARUS, S.A.; MITCHELL, A.E.; RUCKER, R. & SCHMITZ, H.H. 1999. Identification of procyanidins in cocoa (Theobroma cacao) and chocolate using high-performance liquid chromatography/mass spectrometry. J. Agric. Food Chem. 47:490-496
HARBORNE, J. 1990. General procedure and measurement of total phenolics. In: Methods in plant Biochemistry I. Academic Press, USA. p. 1-28
HARBORNE, J. 1993. Introduction to ecological biochemistry. Academic Press, USA. 232 p. HARBORNE, J. 1998. Phytochemical methods: A guide to modern techniques of plant analysis.
Chapman & Hall. London, U. K. 200 p. HEDQVIST, HELENA.; MUELLER-HARVEY, IRENE.; REED, J.D.; KRUEGER, C.G. & MURPHY, M. 2000.
Characterization of tannins and in vitro protein digestibility of several Lotus corniculatus varieties. Animal Feed Science and Technology. 87:41-56
HERNÁNDEZ, A. Y COL. 1999. Clasificación genética de los suelos de Cuba. Instituto de Suelos. Ministerio de la Agricultura. AGRINFOR. Ciudad de La Habana, Cuba. 64 p.
HERNÁNDEZ, I. & BABBAR, LIANA. 2001. Sistemas de producción animal intensivos y el cuidado del ambiente: situación actual y oportunidades. Pastos y Forrajes. 24 (4):281
HERNÁNDEZ, I. 2000. Utilización de las leguminosa arbóreas Leucaena leucocephala, Albizia lebbeck y Bauhinia purpurea en sistemas silvopastoriles. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. EEPF “Indio Hatuey”. Matanzas, Cuba. 138 p.
HORNGHUEY, L.; JEHJENG, J.; HSIENCHENG, R.; JINPYANG, R. & CHUNNAM, K. 1999. Chemistry and biological activities of constituents from Morus australis. Biochimica et Biophysica Acta, General Subjects. 1428 (2): 293-299
HOSTETTMANN, K. & WOLFENDER, J.L. 1997. The search for biologically active secondary metabolites. Pestic. Sci. 51:471-482
HO-ZOO, L & WON-CHU, L. 2001. Utilization of mulberry leaf as animal feed: feasibility in Korea. In: Mulberry for animal feeding in China. (Eds. Jian, L.; Yuyin, C.; Sánchez, M. & Xingmeng, L.). Hangzhou, China. 75 p.
IBRAHIM, M.; FRANCO, F.; PEZO, D.; CAMERO, L.A. & ARAYA, J. 2000. Promoting intake of Cratylia argentea as a dry season supplement for cattle grazing Hyparrhenia rufa in the subhumid tropics. Agroforestry Systems. (In press)
IKAN, R. 1991. Natural products. A Laboratory guide. Academic Press, USA. 360 p. JEGOU, D.; WAELPUT, J.J. & BRONSCHWIG, G. 1994. Consumo y digestibilidad de la morera seca y del
nitrógeno del follaje de morera (Morus alba) y amapola (Malvaviscus arborea) en cabras lactantes. En: Árboles y arbustos forrajeros en América Central. (Ed. Benavides, J.E.). CATIE. Turrialba, Costa Rica, Vol 1, p. 155-162
JIMÉNEZ, MARIBEL.; AGUIRRE, J.; IBRAHIM, M. & PEZO, D. 1998. Efecto de la suplementación con morera (Morus alba) en la ganadería de peso posdestete de terneras de lechería. Agroforestería en las Américas. 5 (17):24-28
JUN, Z.; RONG, Z. & HONGZHANG, W. 2000. Technique selection for prepararing the total polysacharide of mulberry. Chinese Traditional and Herbal Drugs. 31 (5):347-348
KEH-FENG, H. & YUH-FANG, Y. 1996. Three phenylated isoflavones from Erythrina variegata. Journal of the Chinese Society. 43 (6):515-518
KELLOGG, E.A. & JULIANO, N.D. 1997. The structure and function of RuBisCO and their implication for systematic studies. America J. Botany. 84 (3):413-428
KIM, H.B.; LEE, W.C.; KIM, S.Y.; LEE, Y.K. & BANG, H.S. 1998. Effect of mulberry leaf tea for the removal on Cd and Pb in drinking water. Korean J. Seric. Sci. 40 (1):5
KIM, S.Y.; GAO, J.J.; LEE, W.C.; RYU, K.S.; LEE, K.R. & KIM, Y.C. 1999. Antioxidative flavonoids from the leaves of M. alba. Arch. Pharm. Res. 22 (1):81-85
KIM, S.Y.; LEE, W.C.; KIM, H.B.; KIM, A.J. & KIM, S.K. 1998. Antihy perlipidemic effects of methanol extracts from mulberry leaves in cholesterol-induced hyperlipidemia rats. J. Korean Soc. Food Sci. Nutr. 27 (6):1217-1222
KIM, S.Y.; PARK, K.J. & LEE, W.C. 1998. Antinflammatory and antioxidative effects of Morus spp. fruit extract. Korean J. Medicinal Crop Sci. 6 (3):204-209
KIM, S.Y.; RYU, K.S; LEE, W.C.; KU, M.O.; LEE, H.S. & LEE, K.R. 1999. Hypoglycemic effect of mulberry leave with anaerobic treatment in alloxan-induced diabetic mice. Kor. J. Pharmacogn. 30 (2):123-129
KITAHARA, N. & KAWANO, M. 2002. Use mulberry for animal feed. Farming Japan. 36:6

Bibliografía
69
KITAHARA, N.; SHIBATA, S. & NISHIDA, T. 2002. Management and utilization of mulberry for forages in Japan. 1. Productivity of mulberry-pasture association systems and nutritive value of mulberry. In: Mulberry for animal production. Animal Production and Health Paper No. 147. FAO, Rome. p. 231-236
KITAHARA, N.; SHIBATA, S.; KAWANO, M.; TAKAHASHI, S. & NISHIDA, T. 2002. Utilization and management of mulberry (Morus sp.) for forages. 2. Survivals of mulberry trees harvested by cattle browsing and clipping. Grassland Science. 48 (5):412-415
KITAHARA, N.; SHIBATA, S.; TAKAHASHI, S. & NISHIDA, T. 2002. Utilization and management of mulberry (Morus sp.) for forages. 1. Productivity of mulberry-pasture association systems. Grassland Science. 48 (1):1-6
KRUEGER, C.G.; CARTER-DOPKE, N.; TREICHEL, P.M.; FOLTS, J. & REED, J.D. 2000. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry of polygalloyl polyflavan-3-ols in grape seed extract. J. Agric. Food Chem. 48 (5):1663-1667
KUMAR, R. 1992. Antinutritional factors. The potential risks of toxicity and the methods to alleviate them. In: Legumes trees and other fodder trees as protein source for livestock. (Eds. Speedy, A. W. & Pugliese, P.L.). FAO Animal Production and Health Paper. No. 102. p. 145-160
KYUNGHEE, K.; MATSUYAMA, N. & KIM, K.M. 1998. A novel phytoalexin formed in mulberry leaves at the infection of Phloespora maculans causal agent of leaf spot. Journal of the Faculty of Agriculture Kyushu University. 43 (2):89-93
LABARBE, L.; CHEYNIER, V.; BROUSSARD, F.; BOUGUET, J.M. & MOUTOUNET, M. 1999. Quantitative fractionation of grape proanthocyanidins according to their degree of polymerization. J. Agric. Food Chem. 47:2719-2723
LAZARUS, S.A.; ADAMSON, G.E.; HAMMERSTONE, J.F. & SCHMITZ, H.H. 1999. High .performance liquid chromatography/mass spectrometry analysis of proanthocyanidins in food and beverages. J. Agric. Food Chem. 47:3693-3701
LE GUEN, M.P. & BIRK, Y. 1993. Protein protease inhibitor from legumes seeds: nutritional effect, mode of action and structure-function relationship. In: Recent advances of research in antinutritional factor in legumes seeds. Proceedings of the Second International Workshop on Antinutritional Factor (ANFs). (Eds. Vant der Poel, T.F.B.; Huisman, J. & Saini, H.S.). Wageningen Pers, Wageningen. p. 157-171
LEE, H.S.; CHENG, K.S.; KIM, S.Y.; RUY, K.S. & LEE, W.C. 1998. Effect of several sericultural products on block glucose lowering for alloxan-induced hyperglycemic mice. Korean J. Seric. Sci. 40 (1):7
LEE, H.S.; KIM, S.Y.; LEE, Y.K.; LEE, W.C. & LEE, S.D. 1999. Effect of silkworm powder, mulberry leaves and mulberry root bark administered to rat on gastrointestinal function. Korean J. Seric. Sci. 41 (1):29-35
LEE, W.C. & KIM, S.Y. 1997. Utilization and isolation of new active substances from sericulture related materials. National Sericulture and Entomology Research Institute RDA, Korea. 30 p.
LEES, G.L.; SUTTILL, N.H. & GRUBER, M.Y. 1993. Condensed tannins in sainfoin I. A histological and cytological survey of plant tissue. Can. J. Bot. 71:1147-1152
LEINMUELLER, E.; STEINGASS, H. & MENKE, K.H. 1993. Tannins in ruminant feedstuffs. In: Animal Research and Development Vol. 33. Institute for Scientific Cooperation, Germany. 350 p.
LEIVA, LILIAM; LÓPEZ, J.L. & QUIÑONES, Y. 2002. Digestibilidad y comportamiento de cerdos de preceba alimentados con harina de morera. [cd room]. Memorias V Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical” y II Reunión Regional de Morera. EEPF “Indio Hatuey”. Matanzas, Cuba
LEMUS, I.; GARCÍA, R.; DEL VILLAR, E. & KNOP, G. 1999. Hypoglycaemic activity of four plants used in Chilean popular medicine. Phytoterapy Research. 13 (2):91-94
LEZCANO, J.C. & ALONSO, O. 2002. Un patógeno foliar en variedades de morera introducidas en Cuba. Pastos y Forrajes. 25 (4):299
LEZCANO, J.C. 1999. Las enfermedades en plantas arbóreas de interés para la ganadería. Pastos y Forrajes. 22 (2):159
LEZCANO, S.Q. & GONZÁLEZ, R. 2000. Metodología para la evaluación de alimentos de consumo animal. Instituto de Ciencia Animal. La Habana, Cuba. 93 p.
LI, Y. 2001. An important topic to be discussed: Utilization of mulberry leaves and production of animal fibers. In: Mulberry for animal feeding in China (Eds. Jian, L.; Yuyin, C.; Sánchez, M. & Xingmeng, L.). Hangzhou, China. p. 1-7
LIENER, I.E. 1997. Plant lectins. Properties, nutritional significance and function. In: Antinutrients and phytochemicals in food. American Chemical Society, USA. p. 31-43

Bibliografía
70
LIM, S.; YOUNG-TAEK, K. & SANG-POONG, L. 1990. Sericulture training manual. FAO (Eds. Jun, R.; Jung-Sung, L. & Byung-Ho, L.). Agricultural Services Bulletin No. 80. 117 p.
LINDROTH, R.L.; ROTH, S.; KRUEGER, E.L.; VOLIN, J.C. & KOSS, P.A. 1997. Phenolic. Biology. 3:279-289 LIU, J.X.; ZUNYAO; YAN, B.J.; SHI, Z.Q.; WANG, X.Q. & YU, J.Q. 2002. Mulberry leaf supplement for sheep
fed ammoniated rice straw. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 189-201 LÓPEZ, O.S. 1994. Determinación de taninos y cumarina en dos procedencias de Gliricidia sepium. Tópico
especial, María Kass (Profr. Consejero). Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Turrialba, Costa Rica. (Mimeo).
LÓPEZ, SILVIA. 1995. Evaluación de compuestos secundarios y consumo voluntario de cinco procedencias de Gliricidia sepium (Jacq.) Walp, en dos épocas del año, en el trópico húmedo de Costa Rica. Tesis Mag. Sc. CATIE. Turrialba, Costa Rica. 76 p.
MACHII, H. 2002. Evaluation and utilization of mulberry for poultry production in Japan. In: Mulberry for animal Production. Animal Production and Health Paper No. 147. FAO, Rome. p. 237-243
MACHII, H.; KOYAMA, A. & YAMANOUCHI, H. 2002. Mulberry breeding, cultivation and utilization in Japan. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 63-71
MAKKAR, H.P.S. & BECKER, K. 1996. Nutritional value and antinutritional components of whole and ethanol extracted Moringa oleifera. Animal Feed Science and Technology. 63:211-228
MAKKAR, H.P.S. & BECKER, K. 1998. Do tannins in leaves of trees and shrubs from Africa and Himalayan regions differ in level and activity?. Agroforestry. 40 (1):59-68
MAKKAR, H.P.S. 1999. Quantification of tannins in trees foliage. A laboratory manual for the FAO/IAEA Co-ordinated Research Project on “Use of nuclear and related techniques to develop simple tannin assays for predicting and improving the safety and efficiency of feeding ruminants on tanniniferous trees foliage”. IAEA Working Document. IAEA, Vienna. p. 1-29
MAKKAR, H.P.S.; DAWRA, R.K. & SINGH, B. 1991. Tannin levels in leaves of some oak species at different stages of maturity. J. Sci. Food Agric. 54:513-519
MAKKAR, H.P.S.; SINGH, B. & NEGI, S.S. 1989. Relationship of rumen degradability with microbial colonization, cell wall constituents and tannin levels in some tree leaves. Animal Production. 49:299-303
MALDONADO, M.; GRANDE, D.; ARANDA, E. & PÉREZ-GIL, F. 2000. Evaluación de árboles forrajeros tropicales para la alimentación de rumiantes en Tabasco, México. En: Memorias IV Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical”. EEPF “Indio Hatuey”. Matanzas, Cuba. p. 135-142
MARKHAM, K.R. 1988. The flavonoids: Advances in research since 1980. (Ed. Harborne, J.B.) Chapman and Hall, London, U. K. p. 427-468
MARLES, R.J. & FARNSWORTH, N.R. 1995. Antidiabetic plants and their active constituents. Phytomedicine. 2 (2):137-189
MARTÍN, G.; REYES, F.; HERNÁNDEZ, I. & MILERA, MILAGROS. 2002. Agronomic studies with mulberry in Cuba. In: Mulberry for animal production. Animal Production and Health Paper No. 147. FAO, Rome. p. 103-112
MARTÍN, G.; YÉPES, I.; HERNÁNDEZ, I. & BENAVIDES, J.E. 1998. Evaluación del comportamiento de cuatro variedades de morera durante la fase de establecimiento. En: Memorias III Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería”. EEPF “Indio Hatuey”. Matanzas, Cuba. p. 92-96
MARTÍNEZ, S.J.; HERNÁNDEZ, YAUMARA & GUEVARA, R. 1996. Determinación cuantitativa de algunos factores antinutritivos en cinco leguminosas tropicales. En: Resúmenes Taller Internacional Silvopastoril “Los árboles en los sistemas de producción ganadera”. EEPF “Indio Hatuey”. Matanzas, Cuba. 121 p.
MATEOS, P.F. 2003. Metabolitos secundarios. Producción industrial de metabolitos secundarios. [en línea]. Disponible en: http://www.Metabolitos Secundarios.htlm [Consulta: Agosto 2003].
MÉNDEZ, G. 1996. Estudio farmacognóstico y fitoquímico preliminar de Cymbopogon citratus (DC.) stapf y sus extractos. Tesis de Maestría . IFAL. Ciudad de La Habana, Cuba. p. 36-38
MENGCHENG, T.; ZHISHEN, J. & XIANGRUI, Z. 1996. Study on flavonold content in mulberry leaves. Journal of Zhejiang Agricultural University. 22 (4):394-398
MENGZHAO, Y. 1989. Determination of sterol plant growing substance in mulberry leaves. Journal of Zhejiang Agricultural University. 15 (4):335-340
MIDJAVILA, S. 1990. Sustancias nocivas en los alimentos. En: Toxicología de los alimentos. (Ed. Derache, R.). Omega. Barcelona, España. p. 109-132
MIN, B.R.; MCNABB, W.C.; BARRY, T.N. & KEMP, P.D. 1999. The effect of condensed tannins in Lotus corniculatus upon reproductive efficiency wool production in sheep during late summer and autum. J. Agric. Sci. 32:141-145
MINGLE, Z. 1999. Chinese traditional medicinal prescription by mulberry leaves. Lishizhen Medicine and Materia Medica Research. 10 (4):280- 282

Bibliografía
71
MOCHIUTTI, S. 1995. Comportamiento agronómico y calidad nutritiva de Gliricidia sepium (Jacq.) Walp. bajo defoliación manual y pastoreo en el trópico húmedo. Tesis Mag. Sc. CATIE. Turrialba, Costa Rica. 144 p.
MUELLER-HARVEY, IRENE & MC ALLAN, A.B. 1992. Tannins. Their biochemistry and nutritional properties. In: Advances in plant cell biochemistry and biotechnology I. Morrison London IM:JAI Press Ltd, London, U. K. p. 151-217
MUELLER-HARVEY, IRENE. 2001. Analysis of hydrolysable tannins. Animal Feed Science and Technology. 91:3-20
MUPANGWA, J.F.; ACAMOVIC, T.; TOPPS, I.; NGONGONI, N.T. & HAMUDIKUWANDA, H. 2000. Content of soluble and bound condensed tannins of three tropical herbaceous forage legumes. Animal Feed Science and Technology. 83:139-144
NAM-HO, S. & SHI-YONG, R. 1998. Oxyresveratrol as the potent inhibitor on dopa oxidase activity of mushroom tyrosinase. Biochemical and Biophysical Research Communications. 243 (3):801-803
NAVARRO, MARLÉN. 2002. Evaluación del vigor de la semilla de Albizzia lebbeck (A. Benth.) durante la emergencia de plántula. Tesis presentada en opción al Título académico de Máster en Pastos y Forrajes. EEPF “Indio Hatuey”. Matanzas, Cuba. 89 p.
NELSON, K.E; PELL, A.N. ; DOANE, P.H.; GINER-CHÁVEZ, B. & SCHOFIELD, P. 1997. Chemical and biological assays to evaluate bacterial inhibition by tannins. J. Chem. Ecol. 23:1175-1194
NODA, N. 1998. Mulberry leaves, the new functionality food material. Food and Science. 40 (2):102-108 NWANNENNA, A.I.; MADEJ, A.; LUNDH, T.J.O. & FREDIKSSON, G. 1994. Effects of oestrogenic silage on
some clinical and endocrinological parameters in ovariectomized heifers. Acta Vet. Scand. 35 (2):173-183
OJEDA, F. 1996. Factores antinutricionales presentes en los árboles forrajeros. Diplomado en Silvopastoreo. EEPF “Indio Hatuey”. Matanzas, Cuba
OJEDA, F.; CÁCERES, O. & ESPERANCE, M. 1991. Conservación de forrajes. Editorial Pueblo y Educación. La Habana, Cuba. 80 p.
OJEDA, F.; MONTEJO, I. & PÉREZ, GUADALUPE. 2000. Estudio de la ensilabilidad de la morera. En: Memorias IV Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical”. EEPF “Indio Hatuey”. Matanzas, Cuba. p. 218-222
OVIEDO, J.F. 1995. Morera (Morus sp.) en asociación con Poró (Erythrina poeppigiana) como suplemento para vacas lecheras en pastoreo. Tesis Mag. Sc. CATIE. Turrialba, Costa Rica. 86 p.
PAPANASTASI, V.P.; PLATIS, P.D. & DINI-PAPANASTASI, O. 1998. Effects of age and frequency of cutting on productivity of Mediterranean deciduous fodder tree and shrubs plantation. Forest Ecology and Management. 110:283-292
PEDRAZA, R.M.; GARCÍA, A. & PACHECO, R. 1997. Nutrientes y factores antinutritivos en el follaje de Leucaena leucocephala cv. Perú a diferentes edades de rebrote. Pastos y Forrajes. 20:187-191
PEIYI, W. & XIUSI, H. 1995. Effect of mulberry leaves extraction capsule on treatment of chylaceous urine. Chinese Journal of Parasite Prevention and Cure. 8 (1):49-50
PELLISIER, F. 1994. Effect of phenolic compounds in humus on the natural regeneration of spruce. Phytochemistry. 36 (4):865-867
PERCHELLET, E.M.; GALI, H.U.; MAKKAR, H.P.S. & PERCHELLET, J.P. 1996. Ability of tannins extracted from the leaves of various trees and shrubs to inhibit the biomarkers of tumors promotion in mouse skin in vivo. International Journal of Oncology. 9:801-809
PINTO, R., RAMÍREZ-AVILÉS, L. & KU-VERA, J.C. 2001. Nutritional characteristics of forage species in the silvipastures of multiple strato in Chiapas, Mexico. In: International Symposium on Silvopastoral Systems. Second Congress and Livestock Production in Latin America. San José, Costa Rica. p. 394-398
PINTO, R.; RAMÍREZ-AVILÉS, L.; KU-VERA, J.C.; HERNÁNDEZ, A.; SÁNCHEZ, F. & SAUCEDO, H. 2000. Evaluación químico nutricional y degradabilidad ruminal de especies arbóreas del centro de Chiapas, México. Memorias IV Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical”. EEPF “Indio Hatuey”. Matanzas, Cuba. p. 47-50
PIZARRO, E.A.; RAMOS, A.K. & ALMEIDA, J.E. DE. 1997. Una nueva alternativa Morus spp. como arbustiva forrajera. Pasturas Tropicales. 19 (3):42-44
POLYA, G.M. & FOO, L.Y. 1994. Inhibition of eukaryote signal-regulated protein kinases by plant -derivated catechin-related compounds. Phytochemistry. 35 (6):1399-1405
POULTON, J.E. 1990. Cyanogenesis in plants. Plant Physiol. 94:401-405 RAMANJULU, S. & SUDHAKAR, C. 2000. Proline metabolism during dehydration in two mulberry genotypes
with contrasting drought tolerance. Journal of Plant Physiology. 157 (1):81-85 RAMANJULU, S.; SCREENIVASALU, N.; KUMAR, SG. & SUDHAKAR, C. 1998. Photosynthetic
characteristics in mulberry during water stress and rewatering. Photosynthetica. 35 (2):259-263

Bibliografía
72
RAMOS, G.; FRUTOS, P.; GIRÁLDES, F.J. & MANTECÓN, A.R. 1998. Los compuestos secundarios de las plantas en la nutrición de los herbívoros. Arch. Zootec. 47 (180):597-620
RAMOS, T.O.; LARA, L.P.E.; RIVERA, L.J.A. & SANGINÉS, G.J.R. 2002. Mulberry production with swine lagoon effluent. In: Animal Production and Health Paper No. 147. FAO, Rome, p 261-269
RASKIN, I. 1992. Role of salicylic acid in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 43:439-463 RASKIN, I.; SKUBATZ, H.; TANG, W. & MEEUSE, B.J.D. 1990. Salicylic acid levels in thermogenic and
nonthermogenic plant. Ann. Bot. 66:376-373 RATANAPO, S; NGAMJUNYAPORN, W. & CHULAVATNATOL, M. 1999. Syalic acid binding lectins from
leaf of mulberry (Morus alba). Plant Science Limerick. 139 (2):141-148 RAVINDRAN, S. & COL. 1997. Distribution and variation in mulberry germplasm. Indian J. Plant Genetic
Resources. 10 (2):233-242 RAZZ, ROSA. 1991. Mata Ratón (Gliricidia sepium). Cuadernos técnicos. Facultad de Agronomía. Universidad
del Zulia, Venezuela. 5 p. REYES, F.; MILERA, MILAGROS & MATÍAS, C. 2000. Efecto del intercalamiento de leguminosas
temporales en el establecimiento de morera (Morus alba). Pastos y Forrajes. 23(3):219 RÍOS, LEILA; RONDÓN, ZORAIDA & COMBELLAS, JOSEFINA DE. 2000. Situación de concentrado de
morera (Morus sp.) y Gliricidia (Gliricidia sepium) fresca como suplemento de corderos en crecimiento. Memorias IV Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical”. EEPF “Indio Hatuey”. Matanzas, Cuba. p 348-351
RODRÍGUEZ, C.; ARIAS, R. & QUIÑÓNEZ, J. 1994. Efecto de la frecuencia de poda y el nivel de fertilización nitrogenada sobre el rendimiento y calidad de la biomasa de morera (Morus sp.). En: Árboles y arbustos forrajeros en América Central. (Ed. Benavides, J.E.). CATIE. Turrialba, Costa Rica. Vol. 1, p. 515-529
ROJAS, H. & BENAVIDES, J.E. 1994. Producción de leche de cabras alimentadas con pasto y suplementada con altos niveles de morera (Morus sp.). En: Árboles y arbustos forrajeros en América Central. (Ed. Benavides, J.E.). CATIE. Turrialba, Costa Rica. Vol. 2, p. 305-317
ROSSI, C.A.; PEREIRA, A.M. & GONZÁLEZ, G.L. 2000. Determinación de factores antinutricionales en leñosas forrajeras del Chaco árido argentino. Memorias IV Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical”. EEPF “Indio Hatuey”. Matanzas, Cuba p. 71-75
RUOHOMAKI, K.; CHAPING, F.S.; HAUKIOJA, E.; NEUVONEN, S. & SUOMEKA, J. 1996. Phenolic compounds, ecology and function. Ecology. 77:2302-2311
RUSONG, Z.; JINYI, X. & YIPING, Y. 2000. Effect on the content of flavonol in mulberry leaf by different growing area. The Chinese Journal of Modern Applied Pharmacy. 17 (1):11-13
RUSSO, R. & BOTERO, R. 1996. Nitrogen fixing tree for animal production on acid soils. In: Nitrogen fixing tree for acid soils. (Ed. Powell, M.). Winrock Internationals Nitrogen Fixing Tree Association and the Taiwan Forestry Research Institute. Taiwan, China. p. 67
SALAWU, M.B.; ACAMOVIC, T.; STEWART, C.S. & MAASDORP, B. 1997. Assesment of the nutritive value of Calliandra calothyrsus. its chemical composition and the influence of tannins, pipecolic acid and polyethyleneglycol on in vitro organic matter digestibility. Anim. Feed Sci. Technol. 69: 219-232
SANAVOVA, M.K. & RAKHIMOV, D.A. 1997. Plant polysaccharides. VII. Polysaccharides of Morus and their hypoglycaemic activity. Chemistry of Natural Compounds. 53 (6):617-619
SÁNCHEZ, A.; MIQUILENA, O. & FLORES, R. 2000. Efecto de corte en la arquitectura de la L. leucocephala regada por goteo artesanal. Memorias IV Taller Internacional Silvopastoril “Los árboles y arbustos en la ganadería tropical”. EEPF “Indio Hatuey”. Matanzas, Cuba. p. 235-239
SÁNCHEZ, M.D. 2001. Mulberry as animal feed in the world. In: Mulberry for animal feeding in China (Eds. Jian, L.; Yuyin, C.; Sánchez, M. & Xingmeng, L.). Hangzhou, China. p. 1-7
SÁNCHEZ, M.D. 2002. World distribution and utilization of mulberry and its potential for animal feeding. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 1-8
SÁNCHEZ, TANIA. 2002. Evaluación de un sistema silvopastoril con hembras Mambí de primera lactancia bajo condiciones comerciales. Tesis presentada en opción al Título académico de Máster en Pastos y Forrajes. EEPF “Indio Hatuey”. Matanzas, Cuba. 93 p.
SARMA, R.K.; SINGH, B. & BHAT, T.K. 2000. Nitrogen solubility, protein fractions, tannins and in sacco dry matter digestibility of tree fodders of Shiwalik range. Indian Journal of Animal Nutrition. 17 (1):1-7
SCHMIDEK, A.; TAKAHASHI, R.; NUÑES DE MEDEIROS, A. & RESENDE, K.T. 2002. Bromatological composition and degradation rate of mulberry in goats . In: Animal Production and Health Paper No. 147. FAO, Rome. p. 207-211
SCHOFIELD, P.; MBUGUA, D.M. & PELL, A.N. 2001. Analysis of condensed tannins: a review. Animal Feed Science and Technology. 91:21-40

Bibliografía
73
SHAYO, C.M. 1997. Uses, yield and nutritive value of mulberry (Morus alba) trees for ruminants in the semi-arid area of central Tanzania. Tropical Grasslands. 31 (6):599-604
SHAYO, C.M. 2002. The potential of mulberry as feed for ruminants in central Tanzania. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 131-138
SIMÓN, L. 1998. Del monocultivo de pastos al silvopastoreo. La experiencia de la EEPF IH. En: Los árboles en la ganadería. Tomo I. Silvopastoreo. (Ed. L. Simón). EEPF “Indio Hatuey”. Matanzas, Cuba. p. 9
SIMON, L.; CÁCERES, O.; SANTANA, H.; HERNÁNDEZ, I.; IGLESIAS, J.; DUQUESNE, P. DELGADO, R. & DOCAZAL, G. 1992. Resultados obtenidos en la alimentación de bovinos y ovinos con Albizia lebbeck Benth. VI Encuentro Técnico de la Filial Territorial de ACPA. Matanzas, Cuba. (Mimeo).
SIMÓN, L.; HERNÁNDEZ, I. & OJEDA, F. 1998. Protagonismo de los árboles en los sistemas silvopastoriles. En: Los árboles en la ganadería. Tomo I. Silvopastoreo. (Ed. L. Simón). EEPF “Indio Hatuey”. Matanzas, Cuba. p. 28
SINGH, B. & MAKKAR, H.P.S. 2002. The potential of mulberry foliage as a feed supplement in India. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 139-155
SINGHAL, B.K. & ROOPA, B. 1998. Changes in physiological and biochemical parameters of promising mulberry (Morus indica L.) under water stress condition. Philippine Journal of Science. 127 (4):277-286
SINGLETON, V.L.; ORTHOFER, R. & LAMUELA-RAVENTÓS, ROSA. 1999. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteau reagent. Meth. Enzymol. 299:152-178
SOTELO, ÁNGELA. 1997. Constituents of wild food plants. In: Functionality of food phytochemicals. Johns & Romeo. New York, USA. p. 89-111
SOTELO, ÁNGELA; CONTRERA, E. & FLORES, S. 1995. Nutritional value and content of antinutritional compounds and toxics in ten wild legumes of Yucatan Peninsula. Plant Food. 47:115-123
SOTELO, ÁNGELA; SOTO, M. & LUCAS, B. 1996. Comparative studies of the alkaloids composition of two Mexican Erythrina species and nutritive value of the detoxified seeds. Journal of Agricultural and Food Chemistry. 41:2340-2343
SOTELO, ÁNGELA; SOUSA, H. & SÁNCHEZ, M. 1995. Comparative study of the chemical composition of wild and cultivate beans (Phaseolus vulgaris). Plant Food Human Nutrition. 43:93-100
STAFFORD, HELEN. 1997. Roles of flavonoides in symbiotic and defense functions in legumes roots. The Botanical Review. 63 (1):27-39
STEWARD, J.L.; ALLISON, G.E. & SIMONS, A.J. 1996. Gliricidia sepium. Genetic resources for farmers. Oxford Forestry Institute, Oxford, U. K. 125 p.
STRANSBURGER, E.; NOLL, F.; SCHENK, H. & SCHIMPER, A.F.W. 1994. Tratado de Botánica. Marín. Barcelona, España. 520 p.
SUCK-JOON, P.; SANG-GIL, L.; SANG-CHEOL.; BOUM-YOUNG, L. & YOUNG-JOON, A. 1997. Larvicidal and antifeeding activities of oriental medicinal plant extracts against four species of forest insect pest. Appl. Entomol. Zool. 32 (4):601-608
SUGIMURA, Y.; MORI, T.; NITTA, I.; KOTANI, E.; FURUSAWA, T.; TATSUMI, M.; KUSAKARI, S.I.; WADA, M. & MORITA, Y. 1999. Calcium deposition in idioblasts of mulberry leaves. Annals of Botany. 83 (5):543-550
SUN, B.; RICARDO DA SILVA, J. M. & SPRANGER, I. 1998. Critical factors of vanillin assay for catechins and proanthocyanidins. J. Agric. Food Chem. 46:4267-4274
SUNG-SUK, L.; HAK-JU, L. & DON-HA, C. 2000. Studies of biological activity of wood extractives (III) on the phenolic compounds isolated from heatwood of M. bombycis. Journal of the Korean Wood Science and Technology. 28 (2):42-48
SUNG-YEOU, K.; KANG-SUN, R.; WON-CHU, L.; HYUN-OK, K.; HOI-SEON, L. & KANG-RO, L. 1999. Hypoglycaemic effect of mulberry leave with anaerobic treatment in alloxan-induce diabetical mice. Korean Journal of Pharmacognosy. 30 (2):123-129
SUN-YEOU, K.; JIAN-JUN, G. & HEE-KYOUNG, K. 2000. Two flavonoids from the leave of Morus alba induce differentiation of the human promyelocytic leukaemia (HL-60) cell line. Biological and Pharmaceutical Bulletin. 23 (4):451-455
TALAMUCCI, P. & PARDINI, A. & ARGENTI, G. 2002. Effects of grazing animals and cutting on the production and intake of a mulberry-subterranean clover association. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 223-230
TALAMUCCI, P. & PARDINI, A. 1993. Possibility of combined utilization of Morus alba and Trifolium subterraneum in the Tuscan Maremman (Italy). REUR Technical Series No. 28. p. 206-209
TANIGUCHI, S.; ASANO, N.; TOMINO, F. & MIWA, I. 1998. Potentiation of glucose induce insulin secretion by fagomine, a pseudo-sugar isoleaded from mulberry leaves. Hormone and Metabolic Research. 30 (4):619-683

Bibliografía
74
THOMSEN, K. 1997. Cyanogenic constituents in woody plants in natural lowland rain forest in Costa Rica. Botanical Journal of the Linnean Society. 124:273-294
TOLERA, A.; SEYOUM, M. & SUNDSTOL, F. 1998. Nutritive evaluation of Leucaena leucocephala, L. diversifolia and L. pallida in Awassa, Southern Ethiopia. In: Leucaena-adaptation, quality and farming systems. (Eds. Shelton, H. M.; Gutteridge, R. C.; Muller, B. F. & Bray, R. A.). ACIAR Proceedings No 86
TORPOCO, VIRGINIA & GARBARINO, J.A. 2003. Estudio de hongos chilenos I. Metabolitos en Geasprum triplex Jungh. [en línea]. Disponible en: http://www. TECNIA.(1998) 8(2).htlm. [Consulta: Agosto 2003].
TRIGUEROS, R.O. & VILLALTA, P. 1997. Evaluación del uso de follaje deshidratado de morera (Morus alba) en la alimentación de cerdos de la raza Landrace en etapa de engorde. En: Resultados de Investigación, CENTA, El Salvador. p. 150-155
URRIOLA, E.D.M. 1994. Efecto de la edad de rebrote sobre la composición química y digestibilidad in vitro de cinco procedencias de Gliricidia sepium (Jacq) y su aceptabilidad por cabras adultas. Tesis Mag. Sc. CATIE. Turrialba, Costa Rica. 92 p.
VALDÉS, R. & BALBÍN, MARÍA IRENE. 2000. Curso de fisiología y bioquímica vegetal. UNACH, La Habana. 89 p.
VALERIO, SILVIA. 1994. Contenido de taninos y digestibilidad in vitro de algunos forrajes tropicales. Agroforestería en las Américas. Julio-Septiembre:10-13
VANT SOEST, P.J. 1992. Nutritional ecology of the ruminant. Durhan and Downey Inc., Portland, USA. 345 p. WATERHOUSE, A.L.; PRICE, S.F. & MC CORD, J.D. 1999. Reversed-phase high- performance liquid
chromatography. Meth Enzymol. 299:113-121 WEICHENG, W. 1999. Development of health care tea. Bulletin of Sericulture. 30 (3):30-31 WILLIS, R.B. & ALLEN, P.R. 1998. Improved method for measuring hydrolysable tannins using potassium
iodate. Analyst. 123:435-439 WOLFGANG, G.C. & SHELTON, I.D. 1995. Effect of condensed tannins in Lotus pedunculatus on the
nutritive value of ryegrass (Lolium perenne) fed to sheep. J. Agric. Sci. 125:291-297 WON-CHU, L; & YEOU, K. 1997. Utilization and isolation of new active substances from sericulture
related materials. Progress and Future Development Sericulture Science & Technology. Korean Society of Sericultural Science. 4:119-131
XIANGRUI, Z. & HONGSHENG, L. 2001. Composition and medical value of mulberry leaves. In: Mulberry for animal feeding in China. (Eds. Jian, L.; Yuyin, C.; Sánchez, M. & Xingmeng, L.). Hangzhou, China. 75 p.
XIAO, F.; XIANGRONG, L. & WEIPING, C. 1999. Effect of mulberry leaves extract on reducing blood glucose in diabetic mice. Zhejiang Medicine. 21 (4):218-230
YAMASHITA, T. & OHSAWA, R. 1990. Quantitative investigation on nitrogen metabolism in mulberry leaves. Bull. Nat. Inst. Sericultural and Entomological Sci. March (1):27-44
YE, Z. 2002. Factor influencing mulberry leaf yield. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 123-130
YONG, L. & JINGZHI, M. 1999. Development of mulberry leaves functionality substances and its health food Chinese. Food and Nutritional. 3:25
YONGKANG, H. 2002. Mulberry cultivation and utilization in China. In: Animal Production and Health Paper No. 147. FAO, Rome. p. 11-43
YOUYE, L. & FENGGING, G. 2000. Mulberry leave, the new functionality food material. World Agriculture. 21 (4):44-45
YUN, S.J. & LEE, W.C. 1995a. Studies on the utilization of pharmacologically active constituents in mulberry. I. Varietal and seasonal variation of flavonold glycoside content in leaves. RDA. J. Agric. Sci. 37:201-205
YUN, S. J. & LEE, W.C. 1995b. Studies on the utilization of pharmacologically active constituents in mulberry. II. Gamma-aminobutyric acid in leaves and effects of an anaerobic condition on its content. RDA. J. Agric. Sci. 37:207-213
YUN, S.J. & LEE, W.C.; PARK, K.J. & KIM, S.M. 1995. Studies of utilization of pharmacologically active constituents in mulberry. III. Preparation of leaves conteining high level of Rutin and γ-aminobutyric acid and their use as tea material. RDA. J. Agric. Sci. 37:215-219
ZEPEDA, J. 1991. El árbol de oro. Los mil usos de la morera. Medio Ambiente. 47:28-29 ZHISHEN, J.; JIANMING, W. & MENGSHENG, T. 1996. Determination of Rutin and Quercetin from mulberry
leaves extract by reversed-phase high performance liquid chromatography. Chinese Journal of Chromatography. 14 (6):489-491
ZHISHEN, J.; MENCHENG, T. & JIANMING, W. 1999. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chemistry. 64 (4):555-559

Anexos
75
Anexos
Anexo 1. Distribución de las plantas en las unidades experimentales.
Anexo 2. Distribución de las parcelas en la plantación.

Anexos
76
Anexo 3. Matriz que relaciona las distancias euclidianas en el análisis de conglomerados en el PPLL.
Casos 2:C60300 3:C60500 4:C90100 5:C90300 6:C90500 7:C120100
1:C60100 2,000 2,000 1,000 1,000 1,000 0,000 2:C60300 0,000 2,236 2,236 2,236 2,000 3:C60500 2,236 2,236 2,236 2,000 4:C90100 0,000 0,000 1,000 5:C90300 0,000 1,000 6:C90500 1,000
Casos 8:C120300 9:C120500 10:I60100 11:I60300 12:I60500 13:I90100 1:C60100 0,000 0,000 2,000 1,000 2,000 1,000 2:C60300 2,000 2,000 2,828 1,000 0,000 2,236 3:C60500 2,000 2,000 2,828 1,000 0,000 2,236 4:C90100 1,000 1,000 1,000 1,414 2,236 0,000 5:C90300 1,000 1,000 1,000 1,414 2,236 0,000 6:C90500 1,000 1,000 1,000 1,414 2,236 0,000 7:C120100 0,000 0,000 2,000 1,000 2,000 1,000 8:C120300 0,000 2,000 1,000 2,000 1,000 9:C120500 2,000 1,000 2,000 1,000 10:I60100 2,236 2,828 1,000 11:I60300 1,000 1,414 12:I60500 2,236
Casos 14:I90300 15:I90500 16:I120100 17:I120300 18:I120500 19:T60100 1:C60100 1,414 2,449 2,000 0,000 0,000 2,000 2:C60300 1,414 1,414 0,000 2,000 2,000 0,000 3:C60500 1,414 1,414 0,000 2,000 2,000 0,000 4:C90100 1,000 2,236 2,236 1,000 1,000 2,236 5:C90300 1,000 2,236 2,236 1,000 1,000 2,236 6:C90500 1,000 2,236 2,236 1,000 1,000 2,236 7:C120100 1,414 2,449 2,000 0,000 0,000 2,000 8:C120300 1,414 2,449 2,000 0,000 0,000 2,000 9:C120500 1,414 2,449 2,000 0,000 0,000 2,000 10:I60100 1,414 2,449 2,828 2,000 2,000 2,828 11:I60300 1,000 1,732 1,000 1,000 1,000 1,000 12:I60500 1,414 1,414 0,000 2,000 2,000 0,000 13:I90100 1,000 2,236 2,236 1,000 1,000 2,236 14:I90300 1,414 1,414 1,414 1,414 1,414 15:I90500 1,414 2,449 2,449 1,414 16:I120100 2,000 2,000 0,000 17:I120300 0,000 2,000 18:I120500 2,000
Anexos
77
Casos 20:T60300 21:T60500 22:T90100 23:T90300 24:T90500 25:T120100 1:C60100 0,000 2,000 1,000 1,000 1,414 0,000 2:C60300 2,000 0,000 2,236 2,236 1,414 2,000 3:C60500 2,000 0,000 2,236 2,236 1,414 2,000 4:C90100 1,000 2,236 0,000 0,000 1,000 1,000 5:C90300 1,000 2,236 0,000 0,000 1,000 1,000 6:C90500 1,000 2,236 0,000 0,000 1,000 1,000 7:C120100 0,000 2,000 1,000 1,000 1,414 0,000 8:C120300 0,000 2,000 1,000 1,000 1,414 0,000 9:C120500 0,000 2,000 1,000 1,000 1,414 0,000 10:I60100 2,000 2,828 1,000 1,000 1,414 2,000 11:I60300 1,000 1,000 1,414 1,414 1,000 1,000 12:I60500 2,000 0,000 2,236 2,236 1,414 2,000 13:I90100 1,000 2,236 0,000 0,000 1,000 1,000 14:I90300 1,414 1,414 1,000 1,000 0,000 1,414 15:I90500 2,449 1,414 2,236 2,236 1,414 2,449 16:I120100 2,000 0,000 2,236 2,236 1,414 2,000 17:I120300 0,000 2,000 1,000 1,000 1,414 0,000 18:I120500 0,000 2,000 1,000 1,000 1,414 0,000 19:T60100 2,000 0,000 2,236 2,236 1,414 2,000 20:T60300 2,000 1,000 1,000 1,414 0,000 21:T60500 2,236 2,236 1,414 2,000 22:T90100 0,000 1,000 1,000 23:T90300 1,000 1,000 24:T90500 1,414
Casos 26:T120300 27:T120500 28:A60100 29:A60300 30:A60500 31:A90100 1:C60100 0,000 0,000 1,000 1,000 2,000 1,000 2:C60300 2,000 2,000 2,236 1,000 0,000 2,236 3:C60500 2,000 2,000 2,236 1,000 0,000 2,236 4:C90100 1,000 1,000 0,000 1,414 2,236 0,000 5:C90300 1,000 1,000 0,000 1,414 2,236 0,000 6:C90500 1,000 1,000 0,000 1,414 2,236 0,000 7:C120100 0,000 0,000 1,000 1,000 2,000 1,000 8:C120300 0,000 0,000 1,000 1,000 2,000 1,000 9:C120500 0,000 0,000 1,000 1,000 2,000 1,000 10:I60100 2,000 2,000 1,000 2,236 2,828 1,000 11:I60300 1,000 1,000 1,414 0,000 1,000 1,414 12:I60500 2,000 2,000 2,236 1,000 0,000 2,236 13:I90100 1,000 1,000 0,000 1,414 2,236 0,000 14:I90300 1,414 1,414 1,000 1,000 1,414 1,000 15:I90500 2,449 2,449 2,236 1,732 1,414 2,236 16:I120100 2,000 2,000 2,236 1,000 0,000 2,236 17:I120300 0,000 0,000 1,000 1,000 2,000 1,000 18:I120500 0,000 0,000 1,000 1,000 2,000 1,000 19:T60100 2,000 2,000 2,236 1,000 0,000 2,236 20:T60300 0,000 0,000 1,000 1,000 2,000 1,000 21:T60500 2,000 2,000 2,236 1,000 0,000 2,236 22:T90100 1,000 1,000 0,000 1,414 2,236 0,000 23:T90300 1,000 1,000 0,000 1,414 2,236 0,000 24:T90500 1,414 1,414 1,000 1,000 1,414 1,000 25:T120100 0,000 0,000 1,000 1,000 2,000 1,000 26:T120300 0,000 1,000 1,000 2,000 1,000 27:T120500 1,000 1,000 2,000 1,000 28:A60100 1,414 2,236 0,000 29:A60300 1,000 1,414 30:A60500 2,236

Anexos
78
Casos 32:A90300 33:A90500 34:A120100 35:A120300 36:A120500
1:C60100 1,414 1,414 0,000 0,000 0,000 2:C60300 1,414 1,414 2,000 2,000 2,000 3:C60500 1,414 1,414 2,000 2,000 2,000 4:C90100 1,000 1,000 1,000 1,000 1,000 5:C90300 1,000 1,000 1,000 1,000 1,000 6:C90500 1,000 1,000 1,000 1,000 1,000 7:C120100 1,414 1,414 0,000 0,000 0,000 8:C120300 1,414 1,414 0,000 0,000 0,000 9:C120500 1,414 1,414 0,000 0,000 0,000 10:I60100 1,414 1,414 2,000 2,000 2,000 11:I60300 1,000 1,000 1,000 1,000 1,000 12:I60500 1,414 1,414 2,000 2,000 2,000 13:I90100 1,000 1,000 1,000 1,000 1,000 14:I90300 0,000 0,000 1,414 1,414 1,414 15:I90500 1,414 1,414 2,449 2,449 2,449 16:I120100 1,414 1,414 2,000 2,000 2,000 17:I120300 1,414 1,414 0,000 0,000 0,000 18:I120500 1,414 1,414 0,000 0,000 0,000 19:T60100 1,414 1,414 2,000 2,000 2,000 20:T60300 1,414 1,414 0,000 0,000 0,000 21:T60500 1,414 1,414 2,000 2,000 2,000 22:T90100 1,000 1,000 1,000 1,000 1,000 23:T90300 1,000 1,000 1,000 1,000 1,000 24:T90500 0,000 0,000 1,414 1,414 1,414 25:T120100 1,414 1,414 0,000 0,000 0,000 26:T120300 1,414 1,414 0,000 0,000 0,000 27:T120500 1,414 1,414 0,000 0,000 0,000 28:A60100 1,000 1,000 1,000 1,000 1,000 29:A60300 1,000 1,000 1,000 1,000 1,000 30:A60500 1,414 1,414 2,000 2,000 2,000 31:A90100 1,000 1,000 1,000 1,000 1,000 32:A90300 0,000 1,414 1,414 1,414 33:A90500 1,414 1,414 1,414 34:A120100 0,000 0,000 35:A120300 0,000

Anexos
79
Anexo 4. Matriz que relaciona las distancias euclidianas en el análisis de conglomerados en el PLL.
Casos 2:C60300 3:C60500 4:C90100 5:C90300 6:C90500 7:C120100 1:C60100 2,646 2,646 2,449 2,449 1,732 3,000 2:C60300 0,000 1,000 1,000 2,000 1,414 3:C60500 1,000 1,000 2,000 1,414 4:C90100 0,000 1,732 1,000 5:C90300 1,732 1,000 6:C90500 2,000
Casos 8:C120300 9:C120500 10:I60100 11:I60300 12:I60500 13:I90100 1:C60100 2,449 2,449 2,236 2,449 1,414 2,449 2:C60300 1,000 1,000 1,414 1,000 2,236 1,000 3:C60500 1,000 1,000 1,414 1,000 2,236 1,000 4:C90100 0,000 0,000 1,000 0,000 2,449 0,000 5:C90300 0,000 0,000 1,000 0,000 2,449 0,000 6:C90500 1,732 1,732 1,414 1,732 1,732 1,732 7:C120100 1,000 1,000 1,414 1,000 2,646 1,000 8:C120300 0,000 1,000 0,000 2,449 0,000 9:C120500 1,000 0,000 2,449 0,000 10:I60100 1,000 2,236 1,000 11:I60300 2,449 0,000 12:I60500 2,449
Casos 14:I90300 15:I90500 16:I120100 17:I120300 18:I120500 19:T60100 1:C60100 2,449 2,449 3,000 3,000 2,646 2,236 2:C60300 1,000 1,000 1,414 1,414 0,000 1,414 3:C60500 1,000 1,000 1,414 1,414 0,000 1,414 4:C90100 0,000 1,414 1,000 1,000 1,000 1,000 5:C90300 0,000 1,414 1,000 1,000 1,000 1,000 6:C90500 1,732 1,732 2,000 2,000 2,000 1,414 7:C120100 1,000 1,732 0,000 0,000 1,414 1,414 8:C120300 0,000 1,414 1,000 1,000 1,000 1,000 9:C120500 0,000 1,414 1,000 1,000 1,000 1,000 10:I60100 1,000 1,000 1,414 1,414 1,414 0,000 11:I60300 0,000 1,414 1,000 1,000 1,000 1,000 12:I60500 2,449 2,000 2,646 2,646 2,236 2,236 13:I90100 0,000 1,414 1,000 1,000 1,000 1,000 14:I90300 1,414 1,000 1,000 1,000 1,000 15:I90500 1,732 1,732 1,000 1,000 16:I120100 0,000 1,414 1,414 17:I120300 1,414 1,414 18:I120500 1,414
Anexos
80
Casos 20:T60300 21:T60500 22:T90100 23:T90300 24:T90500 25:T120100 1:C60100 2,646 2,236 2,449 2,449 1,414 3,000 2:C60300 0,000 1,414 1,000 1,000 1,732 1,414 3:C60500 0,000 1,414 1,000 1,000 1,732 1,414 4:C90100 1,000 1,000 0,000 0,000 1,414 1,000 5:C90300 1,000 1,000 0,000 0,000 1,414 1,000 6:C90500 2,000 1,414 1,732 1,732 1,000 2,000 7:C120100 1,414 1,414 1,000 1,000 1,732 0,000 8:C120300 1,000 1,000 0,000 0,000 1,414 1,000 9:C120500 1,000 1,000 0,000 0,000 1,414 1,000 10:I60100 1,414 0,000 1,000 1,000 1,000 1,414 11:I60300 1,000 1,000 0,000 0,000 1,414 1,000 12:I60500 2,236 2,236 2,449 2,449 1,414 2,646 13:I90100 1,000 1,000 0,000 0,000 1,414 1,000 14:I90300 1,000 1,000 0,000 0,000 1,414 1,000 15:I90500 1,000 1,000 1,414 1,414 1,414 1,732 16:I120100 1,414 1,414 1,000 1,000 1,732 0,000 17:I120300 1,414 1,414 1,000 1,000 1,732 0,000 18:I120500 0,000 1,414 1,000 1,000 1,732 1,414 19:T60100 1,414 0,000 1,000 1,000 1,000 1,414 20:T60300 1,414 1,000 1,000 1,732 1,414 21:T60500 1,000 1,000 1,000 1,414 22:T90100 0,000 1,414 1,000 23:T90300 1,414 1,000 24:T90500 1,732
Casos 26:T120300 27:T120500 28:A60100 29:A60300 30:A60500 31:A90100 1:C60100 3,000 2,449 2,236 2,646 2,449 2,449 2:C60300 1,414 1,000 1,414 0,000 1,000 1,000 3:C60500 1,414 1,000 1,414 0,000 1,000 1,000 4:C90100 1,000 0,000 1,000 1,000 0,000 0,000 5:C90300 1,000 0,000 1,000 1,000 0,000 0,000 6:C90500 2,000 1,732 1,414 2,000 1,732 1,732 7:C120100 0,000 1,000 1,414 1,414 1,000 1,000 8:C120300 1,000 0,000 1,000 1,000 0,000 0,000 9:C120500 1,000 0,000 1,000 1,000 0,000 0,000 10:I60100 1,414 1,000 0,000 1,414 1,000 1,000 11:I60300 1,000 0,000 1,000 1,000 0,000 0,000 12:I60500 2,646 2,449 2,236 2,236 2,449 2,449 13:I90100 1,000 0,000 1,000 1,000 0,000 0,000 14:I90300 1,000 0,000 1,000 1,000 0,000 0,000 15:I90500 1,732 1,414 1,000 1,000 1,414 1,414 16:I120100 0,000 1,000 1,414 1,414 1,000 1,000 17:I120300 0,000 1,000 1,414 1,414 1,000 1,000 18:I120500 1,414 1,000 1,414 0,000 1,000 1,000 19:T60100 1,414 1,000 0,000 1,414 1,000 1,000 20:T60300 1,414 1,000 1,414 0,000 1,000 1,000 21:T60500 1,414 1,000 0,000 1,414 1,000 1,000 22:T90100 1,000 0,000 1,000 1,000 0,000 0,000 23:T90300 1,000 0,000 1,000 1,000 0,000 0,000 24:T90500 1,732 1,414 1,000 1,732 1,414 1,414 25:T120100 0,000 1,000 1,414 1,414 1,000 1,000 26:T120300 1,000 1,414 1,414 1,000 1,000 27:T120500 1,000 1,000 0,000 0,000 28:A60100 1,414 1,000 1,000 29:A60300 1,000 1,000 30:A60500 0,000

Anexos
81
Casos 32:A90300 33:A90500 34:A120100 35:A120300 36:A120500
1:C60100 2,449 2,000 3,000 3,000 3,000 2:C60300 1,000 1,732 1,414 1,414 1,414 3:C60500 1,000 1,732 1,414 1,414 1,414 4:C90100 0,000 1,414 1,000 1,000 1,000 5:C90300 0,000 1,414 1,000 1,000 1,000 6:C90500 1,732 1,000 2,000 2,000 2,000 7:C120100 1,000 1,732 0,000 0,000 0,000 8:C120300 0,000 1,414 1,000 1,000 1,000 9:C120500 0,000 1,414 1,000 1,000 1,000 10:I60100 1,000 1,732 1,414 1,414 1,414 11:I60300 0,000 1,414 1,000 1,000 1,000 12:I60500 2,449 2,000 2,646 2,646 2,646 13:I90100 0,000 1,414 1,000 1,000 1,000 14:I90300 0,000 1,414 1,000 1,000 1,000 15:I90500 1,414 2,000 1,732 1,732 1,732 16:I120100 1,000 1,732 0,000 0,000 0,000 17:I120300 1,000 1,732 0,000 0,000 0,000 18:I120500 1,000 1,732 1,414 1,414 1,414 19:T60100 1,000 1,732 1,414 1,414 1,414 20:T60300 1,000 1,732 1,414 1,414 1,414 21:T60500 1,000 1,732 1,414 1,414 1,414 22:T90100 0,000 1,414 1,000 1,000 1,000 23:T90300 0,000 1,414 1,000 1,000 1,000 24:T90500 1,414 1,414 1,732 1,732 1,732 25:T120100 1,000 1,732 0,000 0,000 0,000 26:T120300 1,000 1,732 0,000 0,000 0,000 27:T120500 0,000 1,414 1,000 1,000 1,000 28:A60100 1,000 1,732 1,414 1,414 1,414 29:A60300 1,000 1,732 1,414 1,414 1,414 30:A60500 0,000 1,414 1,000 1,000 1,000 31:A90100 0,000 1,414 1,000 1,000 1,000 32:A90300 1,414 1,000 1,000 1,000 33:A90500 1,732 1,732 1,732 34:A120100 0,000 0,000 35:A120300 0,000

Anexos
Anexo 5. Dendograma obtenido a partir de los resultados del análisis cualitativo de los metabolitos secundarios en el PPLL.
Distancia euclidiana CASOS 0 5 10 15 20 25 +---------+---------+---------+---------+---------+ A120300 35 A120500 36 C60100 1 T120500 27 A120100 34 T120100 25 T120300 26 I120500 18 T60300 20 C120500 9 I120300 17 C120100 7 C120300 8 A60100 28 A90100 31 C90100 4 T90100 22 T90300 23 C90500 6 I90100 13 C90300 5 I60100 10 T60500 21 A60500 30 C60300 2 I120100 16 T60100 19 C60500 3 I60500 12 I60300 11 A60300 29 A90300 32 A90500 33 I90300 14 T90500 24 I90500 15 Anexo 6. Dendograma obtenido a partir de los resulta
en el PLL. Distan CASOS 0 5 1 +---------+--------- A120300 35 A120500 36
82
dos del análisis cualitativo de los metabolitos secundarios
cia euclidiana 0 15 20 25 +---------+---------+---------+

Anexos
83
C120100 7 T120300 26 A120100 34 I120300 17
T120100 25 I120100 16 A90100 31 A90300 32 C90100 4 T120500 27 A60500 30 T90100 22 T90300 23 I90100 13 I90300 14 C120500 9
I60300 11 C90300 5 C120300 8 T60300 20 A60300 29 C60300 2 C60500 3 I120500 18 T60500 21 A60100 28 I60100 10 T60100 19 I90500 15 C90500 6 A90500 33 T90500 24 C60100 1 I60500 12