asparagine biosynthesis by cottonroots - plant physiology · analysis yielded 33 ninhydrin-positive...
TRANSCRIPT

Plant Physiol. (1970) 45, 429-434
Asparagine Biosynthesis by Cotton RootsCARBON DIOXIDE FIXATION AND CYANIDE INCORPORATION'
Received for publication October 17, 1969
IRWIN P. TING AND WILLIAM C. ZSCHOCHEUniversity of Califbrnia, Riverside, California 92502
ABSTRACT
Asparagine is the dominant amino acid in cotton root tips(Acala SJ-1). Two biosynthetic pathways may be operative.First, asparagine is an ultimate product of nonphotosyn-thetic CO2 fixation. Whereas short term '4CO2 labeling ex-periments indicate that malate is the predominant product,asparagine appears exponentially and does not appear to bein an active metabolic pool. Other products labeled with14CO2 are citrate, aspartate, and glutamate. No neutralcomponents are labeled. Secondly, asparagine is syn-thesized via a pathway starting with cyanide. Major aminoacid products labeled with '4CN- are 3-cyanoalanine andasparagine. Similarly to CO2 fixation, asparagine synthe-sized from cyanide is not in an active metabolic pool. Otherproducts labeled include anion and neutral components.The exact nature of the latter is not known.
The biosynthesis of asparagine by plant tissues has been ofinterest in recent years. Despite numerous investigations, theexact biosynthetic pathway is not clearly resolved. In 1955,Webster and Varner (20) reported asparagine biosynthesis inplant tissue by an enzymic mechanism similar to the glutaminesynthetase reaction in which aspartate is converted to asparaginein the presence of NH3, ATP, and magnesium ions. Somewhatlater, Oaks (12) reported the conversion of aspartate to aspara-gine in the presence of ATP, NH3, and magnesium in maizetissue. A specific enzyme catalyzing this reaction, however, hasnot been purified from plant tissue, and its existence (at least asasparagine synthetase, EC 6.3.1.1) remains somewhat ques-tionable at this time. Wilson, Krotkov, and Reed (21) demon-strated the biosynthesis of asparagine from CO2 in the light byLupinus angustifolius. In a similar experiment, Lees, Farnden, andElliott (9) demonstrated CO2 and cyanide incorporation intoasparagine in the light by the same species. They also reportedconversion in vivo, but not in vitro, of aspartate to asparagine.In 1963 Blumenthal-Goldschmidt, Butler, and Conn (4) reported
1 In part, this investigation was supported by the Agricultural Re-search Service, Department of Agriculture, Contract 12-14-100-9493(34), administered by the Crops Research Division, Beltsville,Maryland, and by United States Public Health Service Training GrantAP 00008. Mr. W. C. Zschoche was a graduate assistant supported byDepartment of Agriculture Contract 716-15-4.
2 Mention of a trademark name or a proprietary product does notconstitute a guarantee or warranty of the product by the United StatesDepartment of Agriculture and does not imply its approval to theexclusion of other products that may also be suitable.
that the amide carbon of asparagine could be derived from cy-anide. They clearly demonstrated asparagine synthesis fromcyanide in sorghum, barley, pea, flax, red clover, and white cloverafter 40 hr of incubation in the light. Since that time numerousinvestigators have demonstrated the incorporation of cyanideinto asparagine. Almost all experiments, however, have beenconducted under illumination with green tissue derived fromcyanogenic plants.
In this paper, we report that cotton root tissue can assimilateeither carbon dioxide or cyanide to form asparagine as anultimate product.
MATERIALS AND METHODS
Plant Material. Cotton seed (Acala SJ-1) was germinated onfilter paper at 25 C. Three-day old root tips 1 cm long were usedin all experiments.Amino Acid Analysis. Amino acid analyses were conducted with
an automatic analyzer with an 8-hr zinc ligand analysis (3). Tissuewas extracted by freezing in liquid nitrogen, grinding to a finepowder, and extracting with a sequential solvent system of chloro-form-methanol, water, and chloroform as previously described(17). The water-methanol fraction was passed through a Dowex502 (1- X 10-cm) column in the hydrogen form. Amino acids wereeluted from the column with 2 N ammonium hydroxide. Theammonium hydroxide eluate was freeze-dried, and the aminoacids were dissolved in a minimum volume of buffer.
Organic Acid Analysis. Organic acids were extracted from2.5-day-old cotton root tips and analyzed on a silicic acid columnaccording to the method described by Swim and Utter (14).Organic acid samples were prepared similarly to the amino acids.After extraction with the chloroform-methanol-water system,amino acids were removed with a Dowex 50 column (see above).The sample was subsequently placed on a Dowex 1 (1- X 10-cm)column prepared in the formate form. Organic acids were elutedwith 8 N formic acid, freeze-dried, and redissolved in 0.2 NH2SO4 prior to silicic acid column chromatography.
14C Tracer Studies. Carbon dioxide fixation studies were con-ducted by floating 1 cm long, 3-day-old root tips on 5 ml of 0.05M, pH 7.5, tris buffer at 25 C for varying times up to 1 hr. Ap-proximately 20 roots were used per flask. The buffer contained50 ,uc of NaH'4CO3 (specific radioactivity = 50 ,uc/,umole).Incubations were conducted in a closed container. After the 1-hruptake period, a pulse experiment was conducted by quicklywashing and transferring the roots to a nonradioactive buffersolution. Samples were taken during a subsequent 90-min period.After the uptake period, the tissue was quickly washed withdistilled water, frozen in liquid nitrogen, and ground to a finepowder prior to extraction of water-soluble components. Extrac-tion and separation of amino and organic acids and subsequentchromatography was conducted as described previously (15, 17).
Radioactive cyanide uptake studies were conducted similarly429
www.plantphysiol.orgon January 31, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.
TING AND ZSCHOCHE
to the radioactive carbon dioxide experiments, except that tissuewas incubated in 0.05 M citrate-phosphate buffer at pH 4.6. Thebuffer contained 20 ,uc of K'4CN (specific radioactivity = 10.5,uc/,umole). Extraction and subsequent analysis was identical tothe above except that the amino acids were separated by thinlayer chromatography according to the method of Llosa et al.(10). The neutral fraction was chromatographed in methylethyl-ketone-acetone-water (30:10:7, v/v) according to the methoddescribed by Butler and Conn (6) for cyanogenic glucosides.
Cyanide Analysis. Aqueous extracts of cotton tissue were ana-lyzed for cyanide according to the method of Aldridge (2). Theextract was distilled and HCN was trapped in Na2CO3 prior tothe colorimetric analysis.
RESULTS
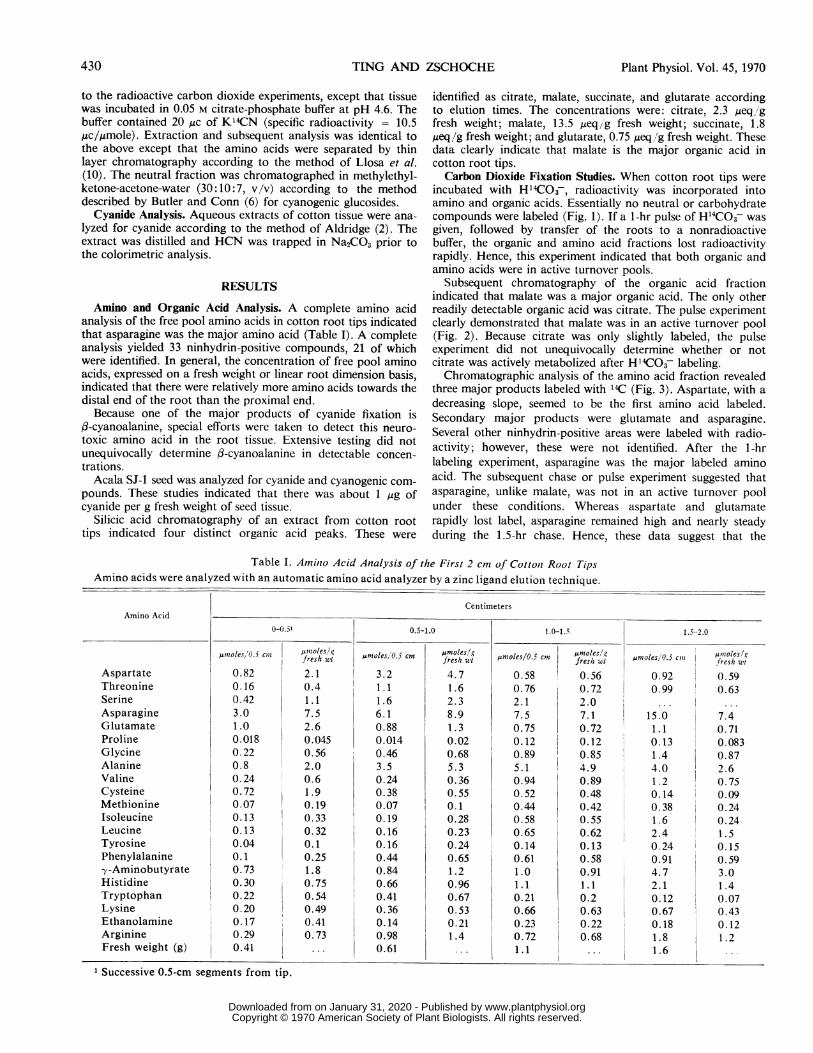
Amino and Organic Acid Analysis. A complete amino acidanalysis of the free pool amino acids in cotton root tips indicatedthat asparagine was the major amino acid (Table I). A completeanalysis yielded 33 ninhydrin-positive compounds, 21 of whichwere identified. In general, the concentration of free pool aminoacids, expressed on a fresh weight or linear root dimension basis,indicated that there were relatively more amino acids towards thedistal end of the root than the proximal end.
Because one of the major products of cyanide fixation is,3-cyanoalanine, special efforts were taken to detect this neuro-toxic amino acid in the root tissue. Extensive testing did notunequivocally determine 3-cyanoalanine in detectable concen-trations.
Acala SJ-1 seed was analyzed for cyanide and cyanogenic com-pounds. These studies indicated that there was about 1 jig ofcyanide per g fresh weight of seed tissue.
Silicic acid chromatography of an extract from cotton roottips indicated four distinct organic acid peaks. These were
identified as citrate, malate, succinate, and glutarate accordingto elution times. The concentrations were: citrate, 2.3 geq,/gfresh weight; malate, 13.5 Aeq/lg fresh weight; succinate, 1.8j.eq/g fresh weight; and glutarate, 0.75 Aeq,/g fresh weight. Thesedata clearly indicate that malate is the major organic acid incotton root tips.Carbon Dioxide Fixation Studies. When cotton root tips were
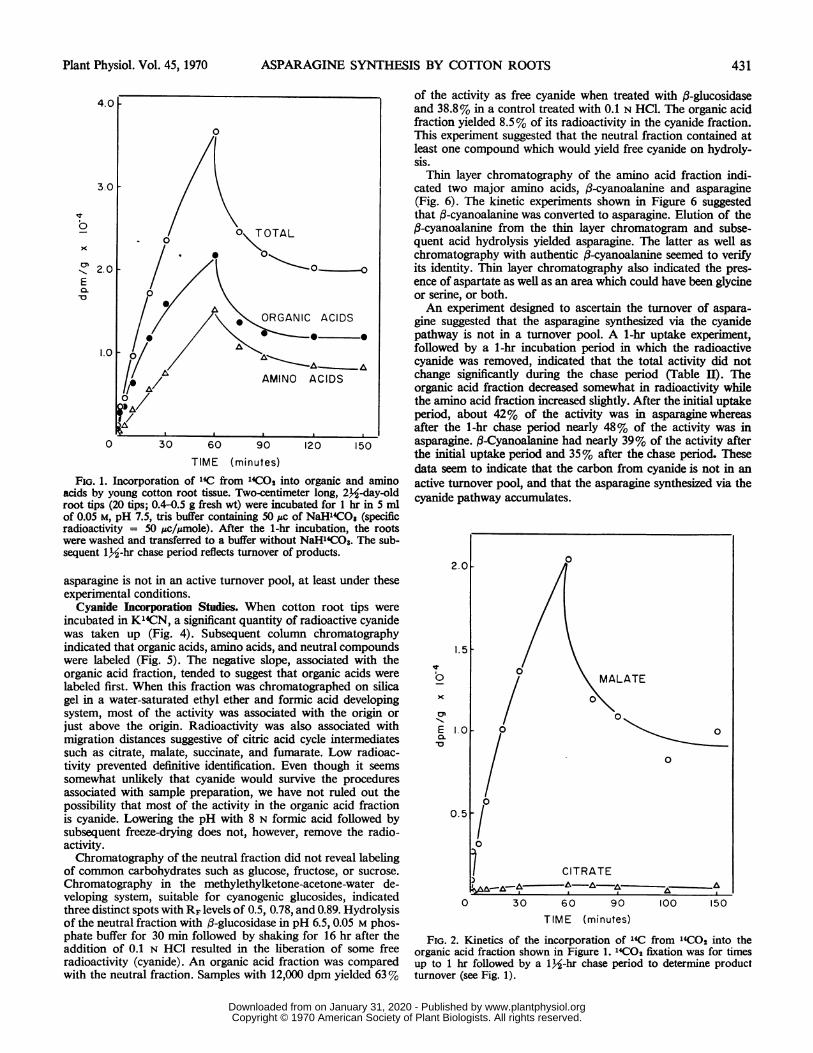
incubated with H'4CO3-, radioactivity was incorporated intoamino and organic acids. Essentially no neutral or carbohydratecompounds were labeled (Fig. 1). If a 1-hr pulse of H`C03- wasgiven, followed by transfer of the roots to a nonradioactivebuffer, the organic and amino acid fractions lost radioactivityrapidly. Hence, this experiment indicated that both organic andamino acids were in active turnover pools.Subsequent chromatography of the organic acid fraction
indicated that malate was a major organic acid. The only otherreadily detectable organic acid was citrate. The pulse experimentclearly demonstrated that malate was in an active turnover pool(Fig. 2). Because citrate was only slightly labeled, the pulseexperiment did not unequivocally determine whether or notcitrate was actively metabolized after H'4C03 labeling.Chromatographic analysis of the amino acid fraction revealed
three major products labeled with 14C (Fig. 3). Aspartate, with adecreasing slope, seemed to be the first amino acid labeled.Secondary major products were glutamate and asparagine.Several other ninhydrin-positive areas were labeled with radio-activity; however, these were not identified. After the 1-hrlabeling experiment, asparagine was the major labeled aminoacid. The subsequent chase or pulse experiment suggested thatasparagine, unlike malate, was not in an active turnover poolunder these conditions. Whereas aspartate and glutamaterapidly lost label, asparagine remained high and nearly steadyduring the 1.5-hr chase. Hence, these data suggest that the
Table I. Amilno Acid Analysis of the First 2 cm of Cottont Root TipsAmino acids were analyzed with an automatic amino acid analyzer by a zinc ligand elution technique.
CentimetersAmino Acid
0-0.5' 0.5-1.0 1.0-1.5 1.5-2.0
,uioles/0.5 cln jfreshut,moles/O.5 cm omstes/g 5cn umteswt oles 0.,Coes/g~smtes'O5c ~froeshgwtfes/i wI mle/ fresh wt -soesd5fresh wt
Aspartate 0.82 2.1 3.2 4.7 0.58 0.56 0.92 0.59Threonine 0.16 0.4 1.1 1.6 0.76 0.72 0.99 0.63Serine 0.42 1.1 1.6 2.3 2.1 2.0 ...
Asparagine 3.0 7.5 6.1 8.9 7.5 7. 1 15.0 7.4Glutamate 1.0 2.6 0.88 1.3 0.75 0.72 1.1 0.71Proline 0.018 0.045 0.014 0.02 0.12 0.12 0.13 0.083Glycine 0.22 0.56 0.46 0.68 0.89 0.85 1.4 0.87Alanine 0.8 2.0 3.5 5.3 5.1 4.9 4.0 2.6Valine 0.24 0.6 0.24 0.36 0.94 0.89 1.2 0.75Cysteine 0.72 1.9 0.38 0.55 0.52 0.48 0.14 0.09Methionine 0.07 0.19 0.07 0.1 0.44 0.42 0.38 0.24Isoleucine 0.13 0.33 0.19 0.28 0.58 0.55 1.6 0.24Leucine 0.13 0.32 0.16 0.23 0.65 0.62 2.4 1.5Tyrosine 0.04 0.1 0.16 0.24 0.14 0.13 0.24 0.15Phenylalanine 0.1 0.25 0.44 0.65 0.61 0.58 0.91 0.59-y-Aminobutyrate 0.73 1.8 0.84 1.2 1.0 0.91 4.7 3.0Histidine 0.30 0.75 0.66 0.96 1.1 1.1 2.1 1.4Tryptophan 0.22 0.54 0.41 0.67 0.21 0.2 0.12 0.07Lysine 0.20 0.49 0.36 0.53 0.66 0.63 0.67 0.43Ethanolamine 0.17 0.41 0.14 0.21 0.23 0.22 0.18 0.12Arginine 0.29 0.73 0.98 1.4 0.72 I 0.68 1.8 1.2Fresh weight (g) 0.41 ... 0.61 ... 1.1 ... 1.6
1 Successive 0.5-cm segments from tip.
430 Plant Physiol. Vol. 45, 1970
www.plantphysiol.orgon January 31, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.
ASPARAGINE SYNTHESIS BY COTTON ROOTS
4.0
3.0
0
x
-.2.0
E
1.0
0 30 60 90 120 150
TIME (minutes)
FIG. 1. Incorporation of 14C from 14CO2 into organic and aminoacids by young cotton root tissue. Two-centimeter long, 2k-day-oldroot tips (20 tips; 0.4-0.5 g fresh wt) were incubated for 1 hr in 5 mlof 0.05 M, pH 7.5, tris buffer containing 50 jAc of NaH14CO3 (specificradioactivity = 50 ,uc/Pmole). After the 1-hr incubation, the rootswere washed and transferred to a buffer without NaHl4C03. The sub-sequent 132-hr chase period reflects turnover of products.
asparagine is not in an active turnover pool, at least under theseexperimental conditions.
Cyanide Incorporation Studies. When cotton root tips were
incubated in K14CN, a significant quantity of radioactive cyanidewas taken up (Fig. 4). Subsequent column chromatographyindicated that organic acids, amino acids, and neutral compoundswere labeled (Fig. 5). The negative slope, associated with theorganic acid fraction, tended to suggest that organic acids werelabeled first. When this fraction was chromatographed on silicagel in a water-saturated ethyl ether and formic acid developingsystem, most of the activity was associated with the origin orjust above the origin. Radioactivity was also associated withmigration distances suggestive of citric acid cycle intermediatessuch as citrate, malate, succinate, and fumarate. Low radioac-tivity prevented definitive identification. Even though it seemssomewhat unlikely that cyanide would survive the proceduresassociated with sample preparation, we have not ruled out thepossibility that most of the activity in the organic acid fractionis cyanide. Lowering the pH with 8 N formic acid followed bysubsequent freeze-drying does not, however, remove the radio-activity.Chromatography of the neutral fraction did not reveal labeling
of common carbohydrates such as glucose, fructose, or sucrose.Chromatography in the methylethylketone-acetone-water de-veloping system, suitable for cyanogenic glucosides, indicatedthree distinct spots with RF levels of 0.5, 0.78, and 0.89. Hydrolysisof the neutral fraction with B-glucosidase in pH 6.5, 0.05 M phos-phate buffer for 30 min followed by shaking for 16 hr after theaddition of 0.1 N HCI resulted in the liberation of some freeradioactivity (cyanide). An organic acid fraction was comparedwith the neutral fraction. Samples with 12,000 dpm yielded 63%
of the activity as free cyanide when treated with ,-glucosidaseand 38.8% in a control treated with 0.1 N HCI. The organic acidfraction yielded 8.5% of its radioactivity in the cyanide fraction.This experiment suggested that the neutral fraction contained atleast one compound which would yield free cyanide on hydroly-sis.Thin layer chromatography of the amino acid fraction indi-
cated two major amino acids, ,B-cyanoalanine and asparagine(Fig. 6). The kinetic experiments shown in Figure 6 suggestedthat 3-cyanoalanine was converted to asparagine. Elution of the,B-cyanoalanine from the thin layer chromatogram and subse-quent acid hydrolysis yielded asparagine. The latter as well aschromatography with authentic ,B-cyanoalanine seemed to verifyits identity. Thin layer chromatography also indicated the pres-ence of aspartate as well as an area which could have been glycineor serine, or both.An experiment designed to ascertain the turnover of aspara-
gine suggested that the asparagine synthesized via the cyanidepathway is not in a turnover pool. A 1-hr uptake experiment,followed by a 1-hr incubation period in which the radioactivecyanide was removed, indicated that the total activity did notchange significantly during the chase period (Table II). Theorganic acid fraction decreased somewhat in radioactivity whilethe amino acid fraction increased slightly. After the initial uptakeperiod, about 42% of the activity was in asparagine whereasafter the 1-hr chase period nearly 48% of the activity was inasparagine. f3-Cyanoalanine had nearly 39% of the activity afterthe initial uptake period and 35% after the chase period. Thesedata seem to indicate that the carbon from cyanide is not in an
active turnover pool, and that the asparagine synthesized via thecyanide pathway accumulates.
2.0-
1.5 .v
b
x
EQ.
1.01
0.51
0 30 60 90 100 150
TIME (minutes)
FIG. 2. Kinetics of the incorporation of 14C from l4CO2 into theorganic acid fraction shown in Figure 1. 14CO2 fixation was for timesup to 1 hr followed by a 1k-hr chase period to determine productturnover (see Fig. 1).
MALATE
0
0
CITRATE
Wd- , . A.-
-P-lant Physiol. Vol. 45, 1970 431
www.plantphysiol.orgon January 31, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

TING AND ZSCHOCHE
0 30 60 90 120 150
TIME (minutes)
FIG. 3. Kinetics of the incorporation of 14C from 14CO2 into theamino acid fraction shown in Figure 1. 14CO2 fixation was from timesup to 1 hr followed by a 1k2-hr chase period to determine productturnover (see Fig. 1).
20
"C)
0
60r
0
0I-
0
0
50F
40F
30
20
10
Plant Physiol. Vol. 45, 1970
* .-t',- / * N
0. A.
* aXa
0
o A.A
I I I I I
0 1 2 3 4 5 6 7 8
Time (minutes)
FIG. 5. Incorporation of cyanide-'4C into neutral (N), amino acid(AA), and organic acid (OA) fractions of cotton root tips. Roots were
incubated in 5 ml of 0.05 M citrate-phosphate buffer, pH 4.6, contain-ing 20 ,uc of K14CN (specific radioactivity = 10.6 ,.c//mole). Anaqueous extract was separated into the three fractions by columnchromatography.
100t .A
95 -
90
95 -0
0
150-
0-_
Asn 0
5 ~~~~~~0
<,oo -o I I0 1 2 3 4 5 6 7 8 9 l0
Time (minutes)
FIG. 6. Kinetics of 14CN incorporation into specific amino acids.See Figure 5. 3-CA: ,B-Cyanoalanine; Asn: asparagine.Table II. Incorporation and Turnover of Cyanide-'4C by Cotton
Root TipsThirty 1-cm long cotton root tips (0.45 g fresh wt) were incu-
bated in 3 ml of 0.05 M tris, pH 7.4, containing 12 ,uc of Na"4CN(specific radioactivity = 52,uc/tsmole) for 1 hr. After I hr, the rootswere washed and transferred to a nonradioactive buffer for an
additional 1 hr (chase period). The data are from two replications.
0 10 20 30 40 50 60Time (minutes)
FIG. 4. Time course of cyanide-14C uptake by 1-cm long 22-day-old cotton root tips (20 roots). Roots were incubated in 5 ml of 0.05 Mcitrate-phosphate buffer, pH 4.6, containing 20 uAc of K'4CN (specificradioactivity = 10.6 ,sc/hmole).
DISCUSSION
Despite much effort, the biosynthesis of asparagine by higherplants has remained enigmatic. In 1955, an enzyme was reportedto occur in lupine and wheat germ which could convert aspartateto asparagine (20). The latter enzyme was analogous to glutaminesynthetase and to the asparagine synthetases which are kno-n
Fraction Initial Uptake Chase
dpm/g fresh wt * hr
Neutral 7,000 3,400Organic acids 25,400 17,000Amino acids 62,300 76,500
Total 94,700 96,900
Amino acids'Aspartate 7.33% 6.8%Asparagine 41.8% 47.8%13-Cyanoalanine 38.5% 35.0%o
' Specific amino acid data are the percentage '4C in each com-pound using the amino acid fraction as 10070.
432
0bxI
E0l.-0
www.plantphysiol.orgon January 31, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

ASPARAGINE SYNTHESIS BY COTTON ROOTS
from nonhigher plant sources. Despite reports of the conversionin vivo of '4C-labeled aspartate to asparagine in various plantsources and numerous statements in the literature attributingan aspartate-asparagine relationship, documentation or purifica-tion of an asparagine synthetase enzyme has not been reportedin the recent literature (8). The photosynthetic biosynthesis ofasparagine from labeled carbon dioxide has been reported (12,21). Such experiments, however, do not yield specific evidenceconcerning biosynthetic pathways. Our data indicate that aspara-gine can be synthesized in the dark by cotton root tissue fromcyanide and bicarbonate. Important is the observation that anultimate product of both CO2 fixation and cyanide incorporationseems to be asparagine, the dominant amino acid in the cottonroot tissue.
In general most higher plants accumulate organic acids, usu-ally malate, after nonphotosynthetic carboxylation; however,certain halophytic plants may accumulate aspartate or otheramino acids in lieu of organic acids (19). The major organic acidproduct of dark CO2 fixation by cotton root tissue seemed to bemalate. In the experiments reported in this paper, after a 60-minincubation period there was four times the 'IC-carbon in malateas in asparagine. The kinetics of the experiment, however, sug-gest that the synthesis of malate occurs with a decreasing slopewhereas the synthesis of asparagine occurred with an increasingslope. Glutamate, a prominent amino acid in cotton root tips,was synthesized with a positive slope similarly to asparagine.Aspartate, however, appeared with a negative slope, suggestingthat it was a primary amino acid product. Three of the fourmajor products, malate, aspartate, and glutamate, appeared to bein metabolic turnover pools inasmuch as a 1-hr pulse labelingexperiment followed by a 90-min turnover period demonstratedrapid loss in activity. Asparagine, on the other hand, did not loseradioactivity during the turnover period following the pulselabeling. Hence, these data indicate quite clearly that asparaginesynthesized via the carbon dioxide fixation pathway is not in anactive metabolic pool, at least under these conditions. Thesedata further suggest that aspartate is converted to asparagine.The general pattern of CO2 fixation by cotton root tips is not
substantially different from CO2 fixation patterns observed inother plants. For example, in our own work the kinetics oflabeling and delabeling by corn root tips after a pulse showed anidentical pattern, except that in the corn root tips aconitateseems to be the compound that is not in an active turnover pool(17). It is worth emphasizing that in previously reported work,the malate synthesized via nonphotosynthetic CO2 fixation is ina relatively active metabolic pool (16, 17, 18).The kinetics of cyanide incorporation are considerably more
complex than those of the CO2 fixation pathway. Kinetic analysisof all the data (see Fig. 5) indicates that the organic fraction islabeled first. Unfortunately, we have not been able to rule outthe presence ofcyanide in this fraction. In the amino acid fraction,the kinetic studies seem to indicate clearly that f3-cyanoalanineis the primary acceptor, which is slowly converted to the ultimateproduct, asparagine (Fig. 6). A perplexing problem is that evenafter a 1-hr labeling period, nearly 40% of the radioactivity inthe amino acid fraction is associated with ,B-cyanoalanine andabout 40% is associated with asparagine. After the 90-min chaseperiod, 35%, of the radioactivity remained in 3-cyanoalanine.Because of the presence of substantial quantities of labeled3-cyanoalanine, we expected to be able to measure this amino
acid in the free pool amino acid extracts. A possible explanationfor the lack of detecting f-cyanoalanine is that very little aspara-gine is actually synthesized via the cyanide pathway in cottonroot tips. A further point of interest is that fl-cyanoalanine isnonenzymatically hydrolyzed to asparagine in acid solution,and hence much of the asparagine labeled from cyanide couldhave resulted from our analytical procedure.
moiety of cyanogenic glucosides does not arise directly from freecyanide. Our data indicate that the neutral fraction contains acompound or compounds which yield 14C (perhaps H14CN)after hydrolysis. Furthermore, the turnover experiments indicatethat the 'IC-compound or compounds are labile. Since it seemsimprobable that a cyanogenic glucoside accounts for the 14Cactivity (6), it may be that a cyanohydrin such as observed byStrobel (13) is present. Further research is necessary to elucidatethe nature of the neutral fraction.The observation that the asparagine synthesized via the CO2
fixation pathway and the cyanide incorporation pathway is notin an active metabolic pool is consistent with previous reportsconcerning asparagine. For example, Lees et al. (9) workingwith wheat shoots suggested that asparagine is a terminal metabo-lite except for some incorporation into protein.The question concerning the biological significance of the
cyanide incorporation pathway cannot be answered at this time.Floss et al. (7) considered the possibility that the incorporationof cyanide into asparagine may represent a detoxification proc-ess. In cyanogenic plants, breakdown of cyanogenic glucosidesyields cyanide which could be incorporated into asparagine. Ourdata suggest that cotton may have a cyanogenic compoundinasmuch as solutions obtained by acid hydrolysis of seed ex-tracts yield positive assays for cyanide. If the cyanide incorpora-tion pathway is significant in cotton root tips, a source of cya-nide is necessary. It seems somewhat unreasonable that cyanidewould be available normally from the environment. Cyanogeniccompounds, if present in cotton, may yield cyanide, which couldbe incorporated into asparagine. Further research is needed forclarification.
Lees et al. (9), during their studies with cyanide assimilationby green wheat shoots, compared the photosynthetic assimilationof CO2 with cyanide incorporation in the light. They observedthat in the presence of carbon dioxide there was a marked en-hancement of cyanide incorporation into asparagine. This ob-servation raised the possibility that there was a relationshipbetween CO2 fixation and cyanide incorporation. Their CO2 fixa-tion experiment, however, was photosynthetic whereas ours isnonphotosynthetic. The possibility that compounds such asserine and cysteine are synthesized photosynthetically fromcarbon dioxide and then further metabolized with cyanide toform asparagine seems to be quite high. Whether or not thereis a direct relationship between the nonphotosynthetic carbondioxide pathway for asparagine biosynthesis and the cyanidepathway is not clear.
In an attempt to clarify metabolic pathways of CO2 fixationand cyanide incorporation, we assayed for several known en-zymes which may be involved. Assays for CO2 fixation pathwayenzymes were conducted according to methods which have beenpreviously described (11). We obtained positive assays for P-enol-pyruvate carboxylase, malic dehydrogenase, malic enzyme, andglutamic-oxaloacetic transaminase (unpublished). With theseenzymes, metabolic pathways involving the carboxylation ofphosphoenolpyruvate and the ultimate formation of aspartatecan be visualized. Our extensive experimentation, with the use ofmethods reported by others in an attempt to assay an asparaginesynthetase enzyme, was negative. Hence, we were not able tofind an enzyme converting aspartate to asparagine. Although thekinetic data in vivo indicate that there is a pathway from carbondioxide to asparagine through aspartate, studies in vitro havenot substantiated this. Reactions converting cyanide to aspara-gine have been reported. Blumenthal et al. (5) described anenzyme (3-cyanoalanine synthetase) which will catalyze thereaction between cyanide and cysteine to form f-cyanoalanine.The isolation and purification of an enzyme hydrolyzing f3-cyano-alanine to asparagine has not as yet been reported.
In summary, cotton root tips synthesize asparagine via twopathways, one involving a nonphotosynthetic carboxylation
Plant Physiol. Vol. 45, 1970 433
Studies by Butler and Conn (6) have indicated that the cyanide www.plantphysiol.orgon January 31, 2020 - Published by Downloaded from
Copyright © 1970 American Society of Plant Biologists. All rights reserved.

434 TING AND
reaction and the other involving the incorporation of cyanide.Asparagine, a main product of both pathways, does not seem tobe in a turnover pool and tends to accumulate.
Acknowledgments-We thank the United States Cotton Research Station,Shafter, California, for a gift of the cotton seed. We are also indebted to Dr. E. E.Conn for helpful discussions concerning cyanide metabolism. Dr. Bahman Khold-ebarin and Mr. Darryl Heustis assisted in some of the experiments.
LITERATURE CITED
1. ABROL, Y. P., E. E. CONN, AND J. R. STOKER. 1966. Studies on the identifica-tion, biosynthesis and metabolism of a cyanogenic glucoside in Nandina domesticaThumb. Phytochemistry 5: 1021-1027.
2. ALDRIDGE, W. N. 1944. A new method for the estimation of micro quantitiesof cyanide and thiocyanate. Analyst 69: 262-265.
3. ANONYMOUS. 1968. Ligand 2-sequential physiological fluid analysis. Hitachi,Perkin-Elmer Manual. N-6250E-1.
4. BLUMENTHAL-GOLDSCHMIDT, S., G. W. BUTLER, AND E. E. CONN. 1963. Incor-poration of hydrocyanic acid labelled with carbon-14 into asparagine in seedlings.Nature 197: 718-719.
5. BLUMENTHAL, S. G., H. R. HENDRICKSON, Y. P. ABROL, AND E. E. CONN. 1968.Cyanide metabolism in higher plants. III. The biosynthesis of 3-cyanoalanine.J. Biol. Chem. 243: 5302-5307.
6. BUTLER, G. W. AND E. E. CONN. 1964. Biosynthesis of the cyanogenic gluco-sides linamarin and lotaustralin. I. Labeling studies in vivo with Linum usitatissi-mum. J. Biol. Chem. 239: 1674-1679.
7. FLOSS, H. G., L. HADWIGER, AND E. E. CONN. 1965. Enzymatic formation offi-cyanoalanine from cyanide. Nature 208: 1207-1208.
ZSCHOCHE Plant Physiol. Vol. 45, 1970
8. FOWDEN, L. 1967. Aspects of amino acid metabolism in plants. Ann. Rev.Plant Physiol. 18: 85-106.
9. LEES, E. M., K. J. F. FARNDEN, AND W. H. ELLIOTr. 1968. Studies on asparaginesynthesis and utilization in seedlings. Arch. Biochem. Biophys. 126: 539-546.
10. LLOSA, P., C. TERTRIN, AND M. JUTISZ. 1964. Separation des acids amines parchromatographie en couche mince de diethylaminoethylcellulose. J. Chro-matogr. 14: 136-139.
11. MUKERJI, S. K. AND I. P. TING. 1968. Intracellular localization of CO2 metabo-lism enzymes in cactus phylloclades. Phytochemistry 7: 903-911.
12. OAKS, A. 1967. Asparagine synthesis in Zea nmays. Biochim. Biophys. Acta141: 436-439.
13. STROBEL, G. A. 1964. Hydrocyanic acid assimilation by a psychrophilic basi-diomycete. Can. J. Biochem. 42: 1637-1639.
14. SWIM, H. E. AND M. F. UTTER. 1957. Isotopic experimentation with inter-mediates of the tricarboxylic acid cycle. In: S. P. Colowick and N. 0. Kaplan,eds., Methods in Enzymology, Vol. IV. Academic Press, New York. pp. 584-609.
15. TING, 1. P. AND W. M. DUGGER. 1965. Separation and detection of organicacids on silica gel. Anal. Biochem. 12: 571-578.
16. TING, 1. P. AND W. M. DUGGER. 1966. CO2 fixation in Opuntia roots. PlantPhysiol. 41: 500-505.
17. TING, 1. P. AND W. M. DUGGER. 1967. CO2 metabolism in corn roots. I. Kineticsof carboxylation and decarboxylation. Plant Physiol. 42: 712-718.
18. TING, I. P. AND W. M. DUGGER. 1968. Non-autotrophic carbon dioxide metab-olism in cacti. Bot. Gaz. 129: 9-15.
19. WEBB, K. L. AND J. W. A. BURLEY. 1965. Dark fixation of C102 by obligateand facultative salt marsh halophytes. Can. J. Bot. 43: 281-285.
20. WEBSTER, G. C. AND J. E. VARNER. 1955. Aspartate metabolism and asparaginesynthesis in plant systems. J. Biol. Chem. 215: 91-99.
21. WILSON, D. G., G. KROTKOV, AND G. B. REED. 1951. Biosynthesis of radio-active asparagine from C402. Science 113: 695-696.
) I'd
www.plantphysiol.orgon January 31, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.