arginine catabolism in aphanocapsa 6308
TRANSCRIPT

Arch. Microbiol. 118, 1 - 6 (1978)
Archives of
Hicrebiology �9 by Springer-Verlag 1978
Arginine Catabolism in Aphanocapsa 6308
Pamela J. Weathers*, Heng Leng Chee, and Mary Mennes Allen**
Department of Biological Sciences, Wellesley College, Wellesley, MA 02181, U.S.A.
Abstract. The catabolic products of arginine meta- bolism were observed in Aphanocapsa 6308, a uni- cellular cyanobacterium, by thin layer chromatog- raphy of growth media, by limiting growth conditions, and by enzymatic analysis. Of the organic, nitrogenous compounds examined, only arginine supported growth in CO2-free media. The excretion of ornithine at a concentration level greater than citrulline suggested the existence in Aphanocapsa 6308 of the arginine dihy- drolase pathway which produced ornithine, CO2, NH 4, + adenosine 5'=triphosphate. Its existence was con- firmed by enzymatic analysis. Although cells could not grow on urea as a sole carbon source a very active urease and subsequently an arginase were also dem- onstrated, indicating that Aphanocapsa can metabo- lize arginine via the arginase pathway. The level of enzymes for both pathways indicates a lack of genetic control. It is suggested that the arginase pathway provides only nitrogen for the cells whereas the arginine dihydrolase pathway provides not only nitrogen, but also COz and adenosine 5'-triphosphate.
Key words: Arginine catabolism - Arginine dihy- drolase pathway - Cyanobacteria - Aphanocapsa 6308 - Arginase - Urease.
The cyanobacterium Aphanocapsa6308 incorporates and metabolizes L-arginine when grown in air, in 5 CO 2 and air, or in CO2-free air. Various pathways of arginine catabolism exist in microorganisms (Fig. l).
Nonstandard Abbreviations. CCCP = carbonylcyanide m- chlorophenyl hydrazone; D C M U = 3-(3,4-dichlorophenyl)-l,1- dimethyl urea; CGP = cyanophycin granule protein; PSII = photosystem II; P S I = photosystem I; TLC = thin layer chroma-. tography; TCA = trichloroacetic acid; DPM = disintegrations per rain * Present address." Department of Biology, College of the Holy- cross, Worcester, MA01610, U.S.A. ** To whom offprint requests should be sent
The arginine dihydrolase pathway in which arginine is ultimately converted to CO 2, NH~, ATP and ornithine has been demonstrated in a nonfermenting Mycoplasma (Shimke et al., 1966) and in several anaerobes (Sperry and Wilkins, 1976; Blakemore and Canale-Parola, 1976; Townsend, 1976; Zolog and Ottow, 1973). Other possible catabolic routes for arginine metabolism include the decarboxylase path- way from arginine to putrescine and the arginase pathway from arginine to urea and ornithine.
This report demonstrates the existence of both the arginine dihydrolasepathway and the arginase pathway in Aphanocapsa 6308. The arginase pathway appears to exist only for nitrogen conservation, whereas the arginine dihydrolase pathway provides not only nit- rogen but also COz and energy for Aphanocapsa 6308.
Materials and Methods
Culture Conditions Aphanoeapsa 6308 (culture collection of Pasteur Institute) was grown routinely in medium No. 11 (Allen, 1968) at 25 ~ or 30 ~ C under cool white fluorescent bulbs at either high (10 800 lux) or low (3240 lux) light intensities. All experiments were initiated with axenic cells in logarithmic phase grown under optimal conditions. Standard growth conditions were in medium No. 11 for sidearm flasks and No. 11 medium supplemented with 2.4g/1 Na z CO3 for Roux flasks which were bubbled with 5 % C Q in air. In low carbonate media the initial pH of the medium was adjusted to 11.3 by the addition of 1 N NaOH. Low CO z cultures were grown in closed sidearm flasks containing small vials of 50 % KOH. Addition of L-arginine, L- aspartic acid, urea, [ -citrulline, L-ornithine, glycolate, succinate and L-ascorbate were in the 5 - 5 0 mM range. Growth inhibiting con- centration of 3-(3,4 dichloro phenyl)- 1,l-dimethyl urea (DCMU) and carbonylcyanide m-chlorophenyl hydrazone (CCCP) were 10 5 M and 10 -4 M, respectively.
Measure of Growth
Growth was measured as cell scattering in either a Gilford240 spectrophotometer or a Klett-Summerson colorimeter at 750nm. Dry weight was measured in each experiment based on a standard curve.
0302-8933/78/0118/0001/$01.20

2 Arch. Microbiol., Vol. 118 (1978)
CO2, NH 3, ATP
CK
Glutamine, CO2, ATP ~ carbamyl phosphate
glutamate --* - , --, ornithine
; i
proline
!-
AL ! i I
arginino-succinate
~ASS J ] citrulline
_ phosphate
~CO1 ~ putresCinu;eae"~ AU
/ u urea
1
~ N H 3
AD
2 3
ADC AR
agmatine I
Fig. 1. Metabolic pathways in arginine biosynthesis and degradation [adapted from Stalon et al. (1967)]. Heavy lines indicate degradative pathways; light lines are biosynthetic pathways. 1 arginine dihydrolase pathway; 2 arginine decarboxylase pathway; 3 arginase pathway. AD arginine deiminase; ADC arginine decarboxylase; AL argininosuccinate lyase; AR arginase; ASS argininosuccinate synthetase; CK carbamate kinase; OCT ornithine carbamyl transferase; U urease; AU agmatine ureohydrolase
Enzyme Preparation and Assay
Cells were harvested, washed once with distilled water, and re- suspended in the buffer specific to the enzyme analysis in question (see below). The cell suspension was then disrupted by ultrasonics in ice for 3 min at 30 s intervals.
Urease activity was determined by the method of Berns et al. (1966) using phosphate buffer at pH7.0, estimated by measuring ammonia production using the Nessler technique and quantified in ~moles of urea hydrolyzed/h- mg protein.
Arginase was assayed by the method of Brown and Cohen (1959). Using sodium glycinate buffer at pH 9.5, activity was estimated and quantified by measuring ornithine production, rather than urea.
Carbamyl phosphokinase was measured by a coupled enzyme assay as described by Schimke et al. (] 966). The ATP generated by the enzyme reaction was used by yeast hexokinase to phosphorylate glucose to form glucose-6-phosphate which was dehydrogenated via glucose-6-phosphate dehydrogenase with the subsequent reduction of NADP. Increase in absorbance of reduced NADP was measured at 340 nm with a Gilford recording spectrophotometer. The reaction was initiated by the addition of carbamyl phosphate (dilithium salt). The initial rate without substrate was subtracted to compensate for auto-reduction of NADP. L-Arginine deiminase was assayed by substituting L-arginine for carbamyl phosphate in the presence of phosphate. Ornithine carbamyl transferase was assayed by substitut- ing L-citrulline for carbamyl phosphate in the presence of phosphate.
Arginine decarboxylase activity was measured in the crude extract suspended in Tris buffer pH 8.3, 0.01 M. The extract was incubated with 40 pmoles of L-arginine as substrate at 35 ~ C; 0.5 ml aliquots were removed and assayed for the presence of agmatine by the method of Goldschmidt and Lockhart (1971). Agmatine ureohydrolase was assayed by substituting agmatine sulphate for L- arginine. The appearance of putrescine was measured according to Goldschmidt et al. (1971), or the disappearance of agmatine was measured.
The release of 1~CO2 from arginine was measured in a crude extract using 14C guanido-labelled L-arginine (NEN; sp. act. 23.1 mCi/mmole). The procedure of Blakemore and Canale-Parola (1976) for ornithine carbamyl-transferase was used to incubate the
celt free extract, however, the reaction was carried out in air in a closed beaker. Small strips of filter paper saturated with equal amounts of 50 % KOH were hung from the rim of the beaker which was sealed with parafilm. At the appointed time, strips were removed and immersed in scintillation fluid and analyzed for radioactivity in a Beckman model LS150 liquid scintillation counter.
Protein was determined on the crude extract according to the method of Lowry et al. (1951).
Commercial enzymes and substrates were obtained from the Sigma Chemical Co., St. Louis, MO.
Thin Layer Chromatography
Cultures were grown under the various conditions previously de- scribed and harvested in log phase (or at a time corresponding to log phase in the case of dying cultures) by centrifuging at 10000 x g for 15 rain. The supernatants (2-5~1) were spotted on Anacil C plates (Analabs, NEN) which were developed and sprayed according to the method of Zolog and Ottow (1973).
Radioactive Incorporations
Determination of the differential localization in cell fractions of the guanido carbon compared to the remainder of the arginine molecule was performed using guanido 14C L-arginine (NEN; sp. act. 23.1mCi/mmole) and universal 14C L-arginine (NEN; sp. act. 277 mCi/mmole). Exponential, axenic cultures growing in medium No. 11 in air, and L-arginine, at 3240 lux, at 25 ~ C were injected with radioactively labeled L-arginine and allowed to grow for 2 h. Cells were then harvested and fractionated according to the method of Roberts et aL (1963). Aliquots of each cell fraction were measured for radioactivity in a Beckman model LS150 liquid scintillation counter.
R e s u l t s
G r o w t h yie lds o f Aphanocapsa6308 are s i gn i f i c an t l y
i n c r e a s e d a t 32401ux w h e n m e d i u m N o . 11 is sup-

P. J. Weathers et al.: Arginine Catabolism in Aphanocapsa 6308
Tablel. Growth yields of Aphanocapsa6308 under aerobic con- ditions in the presence of various organic substrates at 3240 lux, 25 ~ C
Culture conditions Growth yield" Amino acids in media ~
Air - C O 2 b
Medium No. 11-CO3 = 0.15 0 No R, C, O Arginine 5mM 0.17 0.01 R > O > C
20mM 0.18 0.05 R > O > C 50 mM 0.20 0.09 R > O > C
Aspartate 5 mM 0.18 0 n.d. d 20mM 0.21 0 red. 50 mM 0.22 0 n.d.
DCMU No. 11-CO3 = 0 0 n.d. DCMU 50mM arginine 0 0 n.d. CCCP No. 11-CO 3- 0 0 n.d. CCCP 50 mM arginine 0 0 n.d. Urea 5 mM 0.07 0 No R, O, C, U
20mM 0.01 0 No R, O, C, U 50 mM 0.005 0 No R, O, C, U
Ornithine 5raM 0.15 0 O, No R, C 50mM 0.16 0 O, No R, C
Citrulline 5mM 0.14 0 C > O, No R 50raM 0.14 0 C > O, No R
Granulated cells No. 1 I-CO 3 = n.d. 0 n.d. arginine 20mM n.d. 0.05 n.d.
a Growth expressed in terms of mg dry weight/ml at stationary phase u CO 2 free conditions maintained with a hanging center well of 50 ~ KOH in a sealed side arm flask ~ R = arginine; C = citrulline; O = ornithine; U = urea d n.d. = not determined
p lemented with L-arginine (Table 1). Aerob ica l ly grown cultures increase in dry weight p r o p o r t i o n a t e to in- creasing levels o f arginine (Fig. 2). Enhancemen t o f g rowth was also observed when cells were supplement - ed with L-aspar t ic acid, bu t ana logues o f arginine such as ci trul l ine and orn i th ine were unable to suppor t g rowth ; increased urea concent ra t ions p roduced toxicity.
The presence o f the in te rmedia ry p roduc ts o f the arginine ca tabol ic pa thways were assayed in the cul ture superna tan ts o f Aphanocapsa 6308 grown in the pres- ence o f arginine, urea, citrulline, and orni thine. The results o f thin layer c h r o m a t o g r a p h i c separa t ion o f the p roduc t s o f exponent ia l ly growing cultures are also shown in Table 1. Only cells grown in media sup- p lemented with L-arginine showed the expected pa t t e rn o f me tabo l i c in termedia tes co r respond ing to the ar- ginine d ihydro lase p a t h w a y ; arginine was present in levels greater than tha t for orni th ine which was greater than the level o fc i t ru l l ine as observed by the intensi ty of the n inhydr in reac t ion on the ch romatograms .
0 50 mM Arg [ ] 20 mM Arg
2.C - - +CO 2 , ~ A 3 M 3
_ _ CO 2
d
,c JY_2_ _ . . . . . . - - o . . . . . . . . . . -o
0.4
do ~o 6'0 8; ,;o ,~o ,~o ,~ TIME (hr s,)
Fig. 2. Growth of Aphanocapsa 6308 in arginine, in air, or CO2-free air. Growth was measured as culture density in side arm flasks at 25 ~ C and 3240 lux
Crude cell-free extracts o f Aphanocapsa 6308 were examined for the enzymat ic activities o f the three ma jo r pa thways o f arginine ca tabol i sm. Each ca tabol ic sys- tem was, in turn, tested in o rde r to determine if this cyanophy te was able in any way to cont ro l p roduc t ion o f any of these enzymes at the gene level. The presence o f the enzymes, L-arginine deiminase, orni th ine car- bamyl t ransferase, and ca rbamyl phosPhok inase were demons t r a t ed by observing the fo rma t ion o f one o f the end products , adenos ine t r iphospha te (ATP) (Table 2). The act ivi ty o f L-arginine deiminase was very high, whereas the act ivi ty o f ca rbamyl phosphok inase was much lower, and the act ivi ty o f orn i th ine ca rbamyl
t ransferase was bare ly detec table (Table 2). Orni th ine ca rbamyl t ransferase act ivi ty was no t detectable in arginine grown cells with citrull ine as subs t ra te al- t hough great act ivi ty was present with arginine sup- pl ied as subs t ra te (Table 2). A l t h o u g h no t shown in the table, exper iments were pe r fo rmed to enhance the act ivi ty o f orn i th ine ca rbamyl transferase. Only ad- d i t ion o f ca rbamyl phospha t e or L-arginine to the crude extract caused an increase in activity, again at the same level as ei ther o f these a d d e d const i tuents alone. As seen in Table 2 there was no signif icant change in act ivi ty o f any o f these enzymes whether grown in med ium No. 11 or with the add i t ion o f 0.01 M arginine.
Arg inase and urease act ivi ty were bo th detected in crude extracts of Apkanocapsa 6308 (Table 3). Arg inase act ivi ty is best de te rmined by measur ing the p roduc t ion o f bo th orn i th ine and urea. Since urease act ivi ty was greater than tha t for arginase (Table 3) arginase act ivi ty could only be indirect ly measured using the p roduc t ion o f orni thine, since urea was rap id ly conver ted to CO 2 and ammonia . Arg in ine de iminase activity, which also p roduces orni thine, can herein be d i scounted since adenos ine d iphospha te was no t a d d e d to the arginase
4 Arch. Microbiol., Vol. 118 (1978)
Table 2. Arginine dihydrolase enzyme activity in Aphanocapsa 6308 cell free extracts
Specific enzymes and pH a Specific activity b pmole NADPH formed per min per mg soluble protein
Growth media: 0.01 M No. 11 Arginine
L-Arg deiminase pH 8.3 33.7 49.4 pH 9.5 47.8 40.0
Ornithine carbamyl transferase pH 8.3 2.3 n.d. c pH9.5 1.0 n.d.
Carbamyl phosphokinase pHS.3 8.0 5.4 pH9.5 3.2 3.9
Complete + arginine, cell extract omitted n.d. n.d.
" Assay conditions discussed in "Materials and Methods" b Average of several experiments ~ n.d. = not detectable
Table3. Arginase pathway activity in Aphanocapsa6308 cell free extract
Specific enzyme Specific activity a
Growth media 0.01 M No. 11 arginine
Arginase pH 9.5 0.140 0.118 Urease pH9.5 0.169 0.156
Specific activity for arginase is gmol Orn produced per h per mg soluble protein; for urease is pmol produced per h per mg soluble protein
Table4. Relative incorporation levels of the guanido and ornithine moieties of I=arginine into cell fractions of Aphanocapsa 6308 grown at 3280 lux, 25 ~ C, 0.05 M arginine
Guanido Universal 14C_L.arginin e 14C_L.arginin e
DPM a % DPM a %
Cold soluble TCA 830 75.0 940 43.5 Ethanol
ether soluble 90 8.1 210 9.7 Ethanol
soluble-ether insoluble 80 7.2 0 0
Hot TCA soluble i00 9.1 110 5.0 Hot TCA insoluble 0 0.0 900 41.6
a Corrected for quench and a background level of 35 DPM
XlO 3
o
o
XlO~
Q. ~3
XlO I o ,~ 2'0 ~o ~o 5~ M i n u t e s
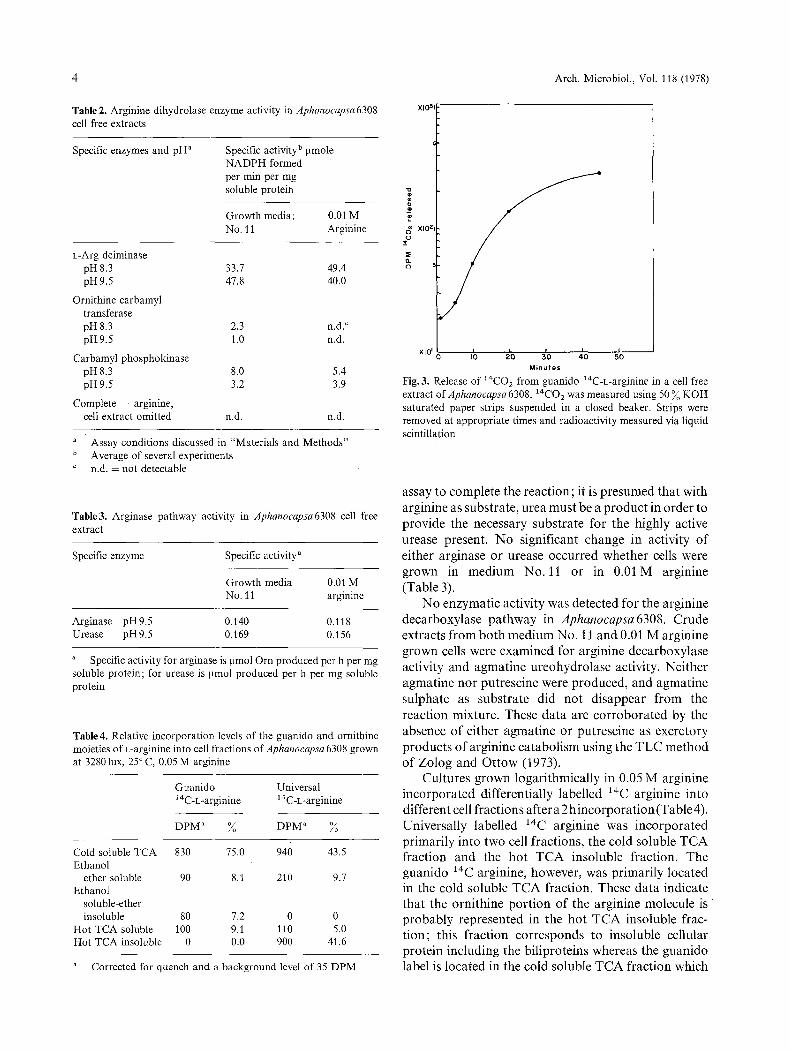
Fig.3. Release of 1~CO2 from guanido 14C-L-arginine in a cell free extract of Aphanocapsa 6308.14CO2 was measured using 50 ~o KOH saturated paper strips suspended in a closed beaker. Strips were removed at appropriate times and radioactivity measured via liquid scintillation
assay to comple te the reac t ion ; it is p resumed tha t with arginine as substrate , urea mus t be a p roduc t in o rder to provide the necessary subs t ra te for the highly active urease present. N o signif icant change in act ivi ty o f ei ther arginase or urease occur red whether cells were grown in m e d i u m No. l l or in 0 . 0 1 M arginine (Table 3).
N o enzymat ic act ivi ty was detected for the arginine decarboxy lase p a t h w a y in Aphanocapsa6308. Crude extracts f rom bo th m e d i u m No. 11 and 0.01 M arginine grown cells were examined for arginine decarboxylase act ivi ty and agmat ine u reohydro la se activity. Ne i the r agmat ine nor put resc ine were p roduced , and agmat ine sulphate as subs t ra te d id no t d i sappea r f rom the reac t ion mixture. These da t a are c o r r o b o r a t e d by the absence o f ei ther agmat ine or putrescine as excre tory p roduc t s o f arginine ca tabo l i sm using the T L C m e t h o d o f Zo log and Ot tow (1973).
Cul tures g rown logar i thmica l ly in 0.05 M arginine i nco rpo ra t ed different ial ly label led t~C arginine into different cell f ract ions after a 2 h i n c o r p o r a t i o n (Table 4). Universa l ly label led 14C arginine was i nco rpo ra t ed p r imar i ly into two cell f ract ions, the cold soluble T C A fract ion and the ho t T C A insoluble fract ion. The guan ido 14C arginine, however , was p r imar i ly loca ted in the cold soluble T C A fract ion. These da ta indicate tha t the orn i th ine po r t i on o f the arginine molecule i s p r o b a b l y represented in the ho t T C A insoluble frac- t ion; this f ract ion co r re sponds to insoluble cellular p ro te in inc luding the b i l ipro te ins whereas the guan ido label is loca ted in the cold soluble T C A fract ion which

P. J. Weathers et al. : Arginine Catabolism in Aphanocapsa 6308 5
is considered to represent the soluble metabolic in- termediates of the cell. Evolution of 14CO2 from guanido 14C-L-arginine was also observed from an in vitro system of Aphanocapsa 6308 (Fig. 3) at the level of 1.87 x 10- 5 ~mol l~CO2/min - mg protein.
As mentioned previously, arginine may be a source of energy in the form of ATP for the cell. However, as shown in Tablel, in the presence of inhibitors of photosynthesis such as DCMU and CCCP, arginine was unable to sustain growth. Suspecting that a source of reduced NADP may be lacking, electron donors such as ascorbate, succinate and glycolate were added to cultures + arginine. In no case was arginine able to support growth in the presence of DCMU despite these added electron donors. It is uncertain whether or not the electron donors were actually taken up by the cells. Cyanophycin granules could not be used as a carbon source for growth in the absence of CO 2 since heavily granulated cells (obtained from late stationary phase cells) inoculated into fresh medium No. 11 did not grow, nor did arginine enhance the growth of granu- lated cells over that of non-granulated cells in either air or a CQ-free atmosphere.
Discussion
The data indicate that Aphanocapsa6308 converts L- arginine to citrulline, ornithine, CO 2, and ammonia with ATP as a by-product by employing the arginine dihydrolase pathway. The citrulline formed is metabo- lized to ornithine and carbamyl phosphate via ornithine carbamyl transferase. Carbamyl phosphate is further metabolized to CO 2, ammonia and ATP via carbamyl phosphokinase. L-Arginine is also converted to or- nithine, urea, CO 2 and ammonia via the arginase pathway. The ornithine and urea are formed, in this case, via arginase; urea is metabolized to CO2 and ammonia by an active urease. Agmatine and putrescine were never observed as products of cultures of Aphanocapsa 6308 ; this indicates that, under the expe- rimental conditions used, arginine decarboxylase and agmatine ureohydrolase activities were lacking.
Except for cells grown in citrulline no other known pathway of arginine catabolism could provide citrulline as an intermediate. Furthermore, although citrulline apparently was taken up by the cells and converted to ornithine (Table 1) it was unable to provide the nec- essary stimulus for growth in the absence of CO2. The absence of arginine in the media in citrulline and ornithine grown cells also indicates that arginine was not being resynthesized from either of these metabolic intermediates in a detectable level.
Although both the arginase and the arginine dihy- drolase pathways produce CO 2 as an end product, only
the latter pathway provides a metabolic source of CO2 for Aphanocapsa 6308 since urea was unable to support growth (Table 1). This is especially significant since, when compared under similar growth conditions, the level of fixation of CO 2 vs that of arginine is equivalent. The availability of CO 2 from the arginine molecule suggests that it may be rapidly incorporated into products of photosynthesis or perhaps other carboxy- lation reactions in the cell. The level of incorporation of arginine labeled either universally or in the guanido carbon was equivalent at 2.47x 10-2mmol/mg dry weight and 3.48 x 10 -2 mmol/mg dry weight, respec- tively. Other studies in this laboratory (Allen et al., 1977) have shown, however, that both arginine and urea can serve as sources of nitrogen. Thus, the sole purpose of urease appears to be the production of nitrogen from arginine in the form of ammonia whereas the arginine dihydrolase pathway produces both CO2 and ammonia for growth.
The results indicate that arginine supplies a large, although not a major, portion of energy required for growth of Aphanocapsa6308. Apparently, either the energy produced via the arginine dihydrolase pathway is insufficient for growth of cells operating only on photosystem I, or lack of reduced NADP is critical as a result of the inhibition of DCMU. Provision of extra- cellular reductants as succinate and ascorbate in the presence of arginine did not enhance growth. CCCP is known to also be a potent inhibitor of many types of phosphorylation and, thus, is probably not solely an inhibitor of photosynthesis but also of the carbamyl phosphate-ATP reaction itself. It appears that any production of energy from the arginine molecule is at a level too low to overcome the depletion produced by DCMU. The exact role of the energy produced from arginine is unknown; however, the data using DCMU suggests that the energy produced from the arginine dihydrolase pathway can not compensate for the lack of reduced NADP.
Although citrulline was expelled into the growth media when cells were grown in arginine, the dose response curve showed increasing growth yield with increasing arginine concentrations. The growth rate, however, did not vary significantly despite increasing arginine concentrations. These data suggest that for the experimental conditions described, arginine is permeat- ing the cell faster than it can be fully utilized. The rate limiting step is probably at ornithine carbamyl trans- ferase. Activity of this enzyme is conspicuously low when provided with only citrulline as substrate. Low activity for this enzyme has also been observed in Eubacterium lentum (Sperry and Wilkins, 1976). Since the activity of ornithine carbamyl transferase increases significantly to the level of that for arginine deiminase when provided arginine as an additional substrate, it is

6 Arch. Microbiol., Vol. 118 (1978)
possible that ornithine carbamyl transferase is de- pendent upon arginine deiminase which probably pro- duces levels of citrulline in excess of the conversion ability of ornithine carbamyl transferase resulting in a preferred excretion rather than a pooling of citrulline. With the exception of ornithine carbamyl transferase, the apparent lack of control over either of these catabolic pathways is not unusual for cyanobacteria. The data suggesting repression of ornithine carbamyl transferase requires further investigation since use of arginine as a substrate apparently permits full oper- ation of the dihydrolase pathway in arginine-grown cells (Table2). Arginine synthesis was found to be feedback inhibited at N-acetyl glutamate kinase (Hoare and Hoare, 1966; Carr et al., 1969). No evidence of repression exists for the synthetic pathway (Hood and Carr, 1971).
Emphasis in this report has been on the utilization of the guanido group as a source of CO2 and nitrogen for the cells. However, the ornithine portion of the molecule is also of great importance, albeit the method of use is unknown. It probably is a carbon backbone for other amino acids since it becomes localized primarily in the insoluble protein fraction of the cell. Although citrulline and ornithine are both useful metabolic products of arginine, neither of these analogues alone provides a stimulus to growth, despite uptake and conversion of at least citrulline to ornithine.
Aphanocapsa 6308, by virtue of its ability to utilize arginine as an organic source of carbon thus permitting it to grow in a CO2-free atmosphere, is a facultative photoheterotroph. Stanier's definition of photohetero- trophy (1973) requires that a cell be capable of utilizing an organic substrate by replacing CO2 as the major cellular source of carbon. Usually this would be experimentally tested in a CO2-requiring organism by demonstrating that provision of an organic substrate reverses the inhibition of growth by DCMU in the light (DCMU being used to block CO 2 fixation by inhibition of PS II and the production of reduced NADP). Since Aphanocapsa6308 does not grow in the presence of DCMU even if provided arginine, it would appear that it is not a photoheterotroph. However, Aphano- capsa 6308 does grow in a CO2-free atmosphere when provided arginine, therefore the basic require- ment for photoheterotrophic growth must be more clearly defined and, as indicated above, subdivided in order to provide a clear description of a cell's metabolic capabilities.
The ability to grow in a CO2-free environment allows Aphanocapsa6308 a distinct advantage over other microorganisms by imparting great flexibility to this species. The potential to use both organic and inorganic forms of nitrogen compensates, in part, for the inability to fix nitrogen. Furthermore, the use of
both organic and inorganic forms of carbon permits ,them to grow in either organically rich refuse or in stagnant, alkaline waters.
Acknowledgements. This work was supported by NSF grant DEB 75- 17633 and by a Research Corporation Cottrell College Science Grant. The authors gratefully acknowledge the technical assistance of Frann Hutchison in this work.
References
Allen, M. M. : Simple conditions for growth of unicellular blue-green algae on plates. J. Phycol. 4, 1 - 4 (1968)
Allen, M. M., Hutchison, F., Weathers, P. J. : Nitrogen limitation and cyanophycin granule formation in Aphanocapsa 6308. American Society for Microbiology, Abstracts, p. 171 (1977)
Berns, D. S., Holohan, P., Scott, E.: Urease activity in blue-green algae. Science 152, 1077-1078 (1966)
Blakemore, R. P., Canale-Parola, E.: Arginine catabolism by Treponema denticola. J. Bacteriol. 128, 616-622 (1976)
Brown, G. W., Cohen, P. P.: Comparative biochemistry of urea synthesis. J. Biol. Chem. 234, 1769-1774 (1959)
Carr, N. G., Hood, W., Pearce, J.: Control and intermediary metabolism in blue-green algae. Prog. Photosyn. Res. 3, I565- 1569 (1969)
Goldschmidt, M. C., Lockhart, B. M. : Rapid methods for determin- ing decarboxylase activity: arginine decarboxylase. Appl. Microbiol. 22, 350-357 (1971)
Goldschmidt, M. C., Lockhart, B. M., Perry, K. : Rapid methods for determining decarboxylase activity: ornithine and lysine de- carboxylase. Appl. Microbiol. 22, 344-349 (197i)
Hoare, D. S., Hoare, L.: Feedback regulation of arginine biosyn- thesis in blue-green algae and photosynthetic bacteria. J. Bacteriol. 92, 375-379 (1966)
Hood, W., Carr, N. G. : Apparent lack of control by repression of arginine metabolism in blue-green algae. J. Bacteriol. 107, 365 - 367 (1971)
Lowry, O. H., Rosebrough, A. L., Farr, A. L., Randall, R. J. : Protein measurement with the Folin reagent. J. Biol. Chem. 193, 265- 275 (1951)
Roberts, R. B., Abelson, P. H., Cowie, D. B., Bolton, E. J., Britten, R. J. : Studies of biosynthesis in Escherichia coli. Carnegie Inst. Washington Publ. (1963)
Schimke, R. T., Berlin, C. M., Sweeney, E. W., Carroll, W. R. : The generation of energy by the arginine dihydrolase pathway in Mycoplasma hominis 07. J. Biol. Chem. 241, 2228-2236 (I966)
Sperry, J. F., Wilkins, T. D. : Arginine, a growth-limiting factor for Eubacterium lentum. J. Bacteriol. 127, 780-784 (1976)
Stalon, V., Ramos, F., Pierard, A., Wiame, J. M. : The occurrence of a catabolic and anabolic ornithine carbamyltransferase in Pseudomonas. Biochim. Biophys. Acta 139, 91-97 (1967)
Stanier, R.: Autotrophy and heterotrophy. In: The biology of the blue-green algae (N. G. Carr, B. A. Whitton, eds.), pp. 501 - 518. Berkeley-Los Angeles: Univ. of Calif. Press I973
Townsend, R. : Arginine Metabolism by Spiroplasma citri. J. Gen. Microbiol. 94, 417-420 (1976)
Zolog, W., Ottow, J. C. G.: Improved thin-layer technique for detection of arginine dihydrolase among the Pseudomonas species. Appl. Microbiol. 26, 1001-1003 (1973)
Received November 25, 1977