are black flies of the subgenus wilhelmia (diptera: simuliidae) multiple ... · are black flies...
TRANSCRIPT

Are black flies of the subgenus Wilhelmia (Diptera:Simuliidae) multiple species or a single geographicalgeneralist? Insights from the macrogenome
PETER H. ADLER1*, ABDULLAH INCI2, ALPARSLAN YILDIRIM2, ONDER DUZLU2,JOHN W. MCCREADIE3, MATÚŠ KÚDELA4, ATEFEH KHAZENI5,TATIANA BRÚDEROVÁ4, GUNTHER SEITZ6, HIROYUKI TAKAOKA7,YASUSHI OTSUKA8 and JON BASS9
1Entomology Program, Clemson University, Clemson, SC 29634-0310, USA2Department of Parasitology, Faculty of Veterinary Medicine, Erciyes University, Kayseri, Turkey3Department of Biological Sciences, University of South Alabama, Mobile, AL 36688, USA4Department of Zoology, Comenius University, SK 842 15 Bratislava, Slovakia5Department of Parasitology and Mycology, Isfahan University of Medical Sciences, Isfahan73461-81746, Iran6District Government of Lower Bavaria, Regierungsplatz 540, 84028 Landshut, Germany7Institute of Biological Sciences, University of Malaya, Kuala Lumpur 50603, Malaysia8Research Center forPacific Islands,Kagoshima University,Korimoto 1-21-24,Kagoshima,890-8580,Japan9The Wessex Chalk Stream and Rivers Trust, Kimbridge, SO51 0LE, UK
Received 21 June 2014; revised 9 August 2014; accepted for publication 9 August 2014
Organisms with vast distributions often represent geographical mosaics of cryptic species. The black fly Simulium(Wilhelmia) lineatum is among the most widely distributed members of the family Simuliidae, ranging from theBritish Isles to eastern China. Rather than viewing S. lineatum as a possible aggregate of multiple species,taxonomists have suggested a more inclusive taxon with additional synonyms. Accordingly, S. lineatum is an idealcandidate for testing the hypothesis that a wide geographical distribution signals the presence of more than onespecies. A cytogenetic approach was used to probe the macrogenome of S. lineatum and other taxa proposed bytaxonomists as conspecific. The banding patterns in the polytene chromosomes of 480 larvae from 15 countries acrossthe Palearctic Region revealed 128 rearrangements of the complement. All rearrangements were autosomal and 89%were inversions nonrandomly distributed among species and among chromosome arms. The analyses clarifylong-standing confusion over previously proposed names and reveal a longitudinal succession of four speciessequentially replacing one another from west to east: Simulium lineatum s.s., Simulium balcanicum, Simuliumturgaicum, and Simulium takahasii. Thus, S. turgaicum is recalled from synonymy and the other three species arevalidated. Within the most-represented species, S. balcanicum, the frequency of inversions follows a longitudinalgradient with a north–south bias; as the distance between the sites increases along this north-west–south-east axis,the similarity of inversion frequencies between sites decreases. Validation of the concept that broadly distributedblack flies are composites of structurally similar species provides a framework for guiding discovery of additionalbiodiversity. © 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183.
ADDITIONAL KEYWORDS: biodiversity – pests – polytene chromosomes – Simulium lineatum.
INTRODUCTION
Much of ecology and applied biology is focused atthe species level: from understanding the effects of
specialization on metacommunity dynamics (Pandit,Kolasa & Cottenie, 2009) to controlling the vectors ofdisease agents and managing pests (Adler, 2009;Kamali et al., 2012). Species perceived as generalists,however, often involve cases of mistaken identity;putative species with broad host or geographical*Corresponding author. E-mail: [email protected]
bs_bs_banner
Biological Journal of the Linnean Society, 2015, 114, 163–183. With 14 figures
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183 163

ranges tend to be composites of structurally similar(cryptic) species, each occupying a discrete subset ofthe larger range (Williams et al., 2012; Hambäcket al., 2013).
The family Simuliidae, through study of the giantpolytene chromosomes in the larval silk glands, pro-vides classic examples of cryptic biodiversity in taxaonce considered single widespread species occupyingdiverse habitats (Rothfels, 1979; Adler, Cheke & Post,2010). Current examples of generalists in theSimuliidae are found in the Palearctic Simuliumsubgenus Wilhelmia. Among the 26 species in thesubgenus are some of the most widely distributedblack flies in the Old World (Adler & Crosskey,2014). Simulium lineatum (Meigen) as currently rec-ognized, for example, extends from southern Spain(Gallardo-Mayenco & Toja, 2002) and the British Islesacross a vast swath of the Palearctic through the Altaisouth to Iran and eastward to the coast of China, adistance of more than 9500 km (Adler & Crosskey,2014). Although wide distributions typically signal thepresence of cryptic species (Adler et al., 2004, 2010),S. lineatum has been viewed as broadly cohesive, andtwo former species, Simulium salopiense Edwards ofWestern Europe and Simulium turgaicum Rubtsovof Central Asia, were declared conspecific withS. lineatum (Crosskey & Davies, 1972; Crosskey &Howard, 1997). Populations of Wilhelmia in Japan andKorea were considered conspecific with S. lineatumuntil 1962, when they were described under the nameSimulium takahasii (Rubtsov), although this wasbased on questionable structural differences. Morerecently, Simulium balcanicum (Enderlein), whichdiffers structurally only in the pupal stage by havinga petiolate pair of gill filaments, was posited asconspecific with S. lineatum (Crosskey & Zwick, 2007).
The chromosomes of seven nominal species of thesubgenus Wilhelmia have been described (Grinchuk &Chubareva, 1974; Knoz, Grinchuk & Chubareva,1976; Petrova et al., 2003; Chubareva, Petrova &Reva, 2007; Chubareva & Petrova, 2008; Huang et al.,2012). Only one study, however, attempted a bandingcomparison between species [Simulium equinum (L.)and S. lineatum in south-western Germany (Weber &Grunewald, 1989)] and only one study, althoughlimited to Armenia, considered the possibility ofcryptic species (in Simulium paraequinum Puri)(Petrova et al., 2003). All chromosomally knownmembers of the subgenus Wilhelmia share a whole-arm interchange (IS + IIL, IL + IIS) possibly homolo-gous with that in the African subgenus Metomphalus(Vajime & Dunbar in Rothfels, 1979). The only otherexamples of a IS + IIL, IL + IIS interchange in theSimuliidae (Rothfels & Freeman, 1983; Procunier,1982) are of independent origin in distantly-relatedtaxa (i.e. different genera).
Our initial motivation for a cytogenetic study ofWilhelmia was sparked by a severe outbreak of blackflies that affected tourism and livestock productionalong the Kizilirmak River in Turkey’s CappadociaRegion during the middle of the first decade of 2000(Yilmaz et al., 2007; Sariözkan et al., 2014). To deter-mine the species involved in the Cappadocia outbreak,we conducted a cytogenetic analysis of the populationof Wilhelmia in the Kizilirmak River. To reconcile theoutbreak with the pest status of the relevant species inother regions and to place our results in an interpre-tive context, we adopted a collaborative approach andexpanded the geographical coverage to include theentire Palearctic Region. Our primary objective was totest the hypothesis that S. lineatum is a single species,as some taxonomists have claimed, versus a compositeof one or more of the variously recognized species orsynonyms: S. balcanicum, S. lineatum s.s., S. salopi-ense, S. takahasii, and S. turgaicum. We examinedmaterial from populations typically regarded as rep-resentative of each of these names.
MATERIAL AND METHODSCOLLECTION OF MATERIAL
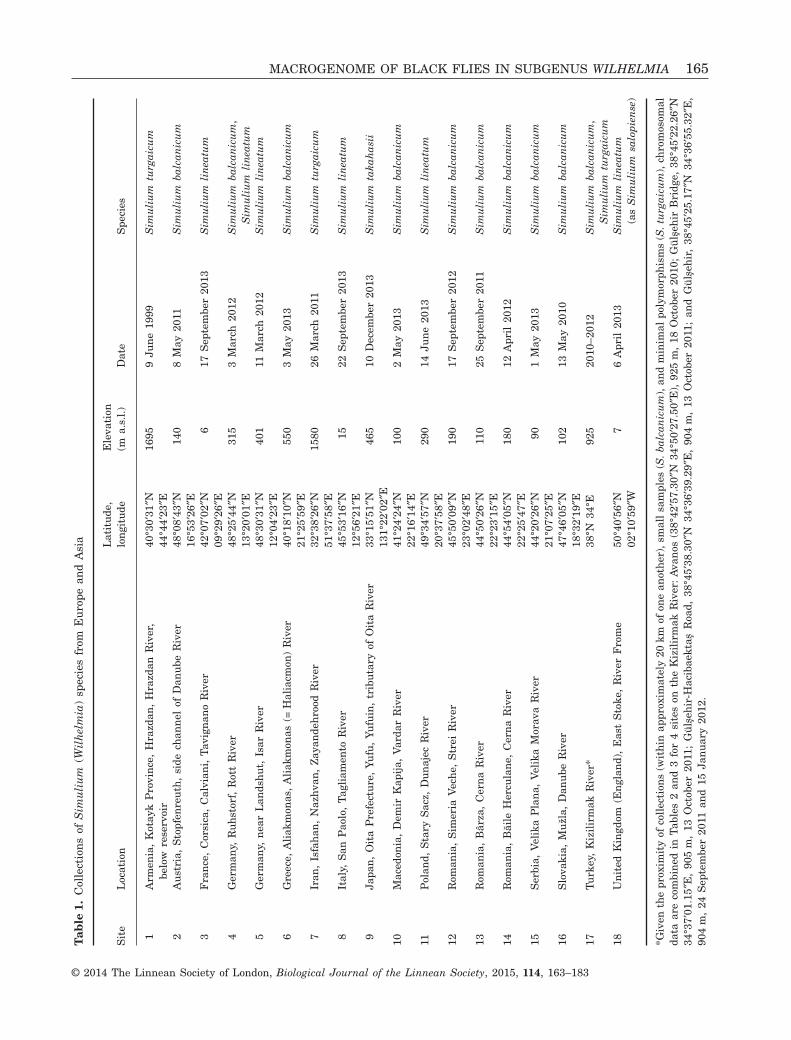
Larvae were hand collected from trailing vegetationat 18 sites in 15 countries (Table 1, Fig. 1) and fixedin two or three changes of modified Carnoy’s solution(1 : 3 acetic ethanol). Larvae patently infected withparasites (morphologically identified) were recorded(Tables 2, 3). Larval carcasses and photographic nega-tives of chromosomes were deposited in the ClemsonUniversity Arthropod Collection, Clemson, SC.
CHROMOSOME PREPARATION
Polytene chromosomes were prepared using theFeulgen technique of Rothfels & Dunbar (1953), inaccordance with the procedures outlined by Adler &Huang (2011), although modified by using acold, 30-min treatment in 5 N hydrochloric acid(Charalambous et al., 1996). The silk-gland contentsof larvae typically solidified into a gelatinous masswithin as little as a week of fixation; thus, afterFeulgen staining, the chromosomes were strippedfrom the mass with fine needles. Gender was deter-mined by gonadal shape. The ovoid to spherical malegonads of the subgenus Wilhelmia are among thelargest in the Simuliidae; their large size and melaninsheath made gender determination straightforward.Female gonads were elongated and ensheathed withmelanin only at the distal end.
CHROMOSOME INTERPRETATION
Selected chromosomes from larvae collected in Tur-key’s Kizilirmak River in September and October 2011
164 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183
Tab
le1.
Col
lect
ion
sof
Sim
uli
um
(Wil
hel
mia
)sp
ecie
sfr
omE
uro
pean
dA
sia
Sit
eL
ocat
ion
Lat
itu
de,
lon
gitu
deE
leva
tion
(ma.
s.l.)
Dat
eS
peci
es
1A
rmen
ia,
Kot
ayk
Pro
vin
ce,
Hra
zdan
,H
razd
anR
iver
,be
low
rese
rvoi
r40
°30′
31″N
44°4
4′23
″E16
959
Jun
e19
99S
imu
liu
mtu
rgai
cum
2A
ust
ria,
Sto
pfen
reu
th,
side
chan
nel
ofD
anu
beR
iver
48°0
8′43
″N16
°53′
26″E
140
8M
ay20
11S
imu
liu
mba
lcan
icu
m
3F
ran
ce,
Cor
sica
,C
alvi
ani,
Tavi
gnan
oR
iver
42°0
7′02
″N09
°29′
26″E
617
Sep
tem
ber
2013
Sim
uli
um
lin
eatu
m
4G
erm
any,
Ru
hst
orf,
Rot
tR
iver
48°2
5′44
″N13
°20′
01″E
315
3M
arch
2012
Sim
uli
um
balc
anic
um
,S
imu
liu
mli
nea
tum
5G
erm
any,
nea
rL
ands
hu
t,Is
arR
iver
48°3
0′31
″N12
°04′
23″E
401
11M
arch
2012
Sim
uli
um
lin
eatu
m
6G
reec
e,A
liak
mon
as,
Ali
akm
onas
(=H
alia
cmon
)R
iver
40°1
8′10
″N21
°25′
59″E
550
3M
ay20
13S
imu
liu
mba
lcan
icu
m
7Ir
an,
Isfa
han
,N
azh
van
,Z
ayan
deh
rood
Riv
er32
°38′
26″N
51°3
7′58
″E15
8026
Mar
ch20
11S
imu
liu
mtu
rgai
cum
8It
aly,
San
Pao
lo,
Tagl
iam
ento
Riv
er45
°53′
16″N
12°5
6′21
″E15
22S
epte
mbe
r20
13S
imu
liu
mli
nea
tum
9Ja
pan
,O
ita
Pre
fect
ure
,Y
ufu
,Y
ufu
in,
trib
uta
ryof
Oit
aR
iver
33°1
5′51
″N13
1°22
′02″
E46
510
Dec
embe
r20
13S
imu
liu
mta
kah
asii
10M
aced
onia
,D
emir
Kap
ija,
Var
dar
Riv
er41
°24′
24″N
22°1
6′14
″E10
02
May
2013
Sim
uli
um
balc
anic
um
11P
olan
d,S
tary
Sac
z,D
un
ajec
Riv
er49
°34′
57″N
20°3
7′58
″E29
014
Jun
e20
13S
imu
liu
mli
nea
tum
12R
oman
ia,
Sim
eria
Vec
he,
Str
eiR
iver
45°5
0′09
″N23
°02′
48″E
190
17S
epte
mbe
r20
12S
imu
liu
mba
lcan
icu
m
13R
oman
ia,
Bâr
za,
Cer
na
Riv
er44
°50′
26″N
22°2
3′15
″E11
025
Sep
tem
ber
2011
Sim
uli
um
balc
anic
um
14R
oman
ia,
Bai
leH
ercu
lan
e,C
ern
aR
iver
44°5
4′05
″N22
°25′
47″E
180
12A
pril
2012
Sim
uli
um
balc
anic
um
15S
erbi
a,V
elik
aP
lan
a,V
elik
aM
orav
aR
iver
44°2
0′26
″N21
°07′
25″E
901
May
2013
Sim
uli
um
balc
anic
um
16S
lova
kia,
Mu
žla,
Dan
ube
Riv
er47
°46′
05″N
18°3
2′19
″E10
213
May
2010
Sim
uli
um
balc
anic
um
17Tu
rkey
,K
izil
irm
akR
iver
*38
°N34
°E92
520
10–2
012
Sim
uli
um
balc
anic
um
,S
imu
liu
mtu
rgai
cum
18U
nit
edK
ingd
om(E
ngl
and)
,E
ast
Sto
ke,
Riv
erF
rom
e50
°40′
56″N
02°1
0′59
″W7
6A
pril
2013
Sim
uli
um
lin
eatu
m(a
sS
imu
liu
msa
lopi
ense
)
*Giv
enth
epr
oxim
ity
ofco
llec
tion
s(w
ith
inap
prox
imat
ely
20km
ofon
ean
oth
er),
smal
lsa
mpl
es(S
.bal
can
icu
m),
and
min
imal
poly
mor
phis
ms
(S.t
urg
aicu
m),
chro
mos
omal
data
are
com
bin
edin
Tabl
es2
and
3fo
r4
site
son
the
Kiz
ilir
mak
Riv
er:A
van
os(3
8°42
′57.
30″N
34°5
0′27
.50″
E),
925
m,
18O
ctob
er20
10;
Gü
lseh
irB
ridg
e,38
°45′
22.2
6″N
34°3
7′01
.15″
E,
905
m,
13O
ctob
er20
11;
Gü
lseh
ir-H
acib
aekt
asR
oad,
38°4
5′38
.30″
N34
°36′
39.2
9″E
,90
4m
,13
Oct
ober
2011
;an
dG
üls
ehir
,38
°45′
25.1
7″N
34°3
6′55
.32″
E,
904
m,
24S
epte
mbe
r20
11an
d15
Jan
uar
y20
12.
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 165
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

and in Japan in December 2013 were photographedunder oil immersion on an Olympus BX40 compoundmicroscope. Maps (Figs 2, 3, 4, 5, 6, 7, 8, 9, 10) wereprepared using PHOTOSHOP ELEMENTS, version 8(Adobe Inc.). The entire complement (chromosomes I,II, and III) was divided into 100 arbitrary sections ofapproximately equivalent length, beginning with theshort arm (S) of I and continuing through the long arm(L) of III. We maintained the section limits of Weber &Grunewald (1989) wherever possible, although ourattempts were stymied by the low resolution of theirmaps and the presence of heterozygous inversions(e.g. in the middle of IS) and geographically local,homozygous inversions (e.g. in the base of IIL) on theirmaps. For example, the IL, IIL, and IIIL photomaps forS. lineatum of Weber & Grunewald (1989) carry theIL-10, 11 and IIL-8, 9, 14, and IIIL-5 homozygoussequences, respectively, of our system, and IS in theirphotomap is heterozygous for our IS-4 sequence. Wetherefore established the standard map de novo, basedon the most common sequence in each arm across oursampled populations. Chromosomal rearrangements,chiefly inversions and heterobands, are indicated bybrackets or arrows on our maps. Inversions typicallywere numbered in order of their discovery. Eachheteroband (hb) was named for the section in which it
occurred (e.g. IIIL 97hb), as were supernumerarybands (e.g. IIIL 99i). Fixed inversions across all sites ofa species are italicized.
Frequencies of all polymorphisms, based on indi-vidual chromosomal homologues, were calculated(Tables 2, 3). Twenty-four (4.8%) of 504 chromosomallyprepared larvae could not be compared completelywith the standard map and were excluded from calcu-lations of polymorphism frequencies and all otherevaluations. Fifteen of these larvae were from Europe’slate-summer generation (September) of minusculemature larvae (length, mean ± SE = 5.3 ± 0.07 mm,N = 14) of S. lineatum s.s. The remaining nineindividuals (all S. balcanicum) with chromosomesnot amenable for analysis were pharate pupae.Polymorphisms of sufficient frequency were tested forHardy–Weinberg equilibrium. We used chi-squaredtests to evaluate independence of inversion sequencesand gender, and goodness of fit tests to determine if thedistribution of inversions was equal among species andamong chromosome arms.
GILL MORPHOLOGY
The lengths of all eight gill filaments (Fig. 11) weremeasured using IMAGEJ (NIH) for representative
Figure 1. Map of collecting sites for four species of Simulium (Wilhelmia) in the Palearctic Region; numbers correspondto sites in Table 1.
166 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183
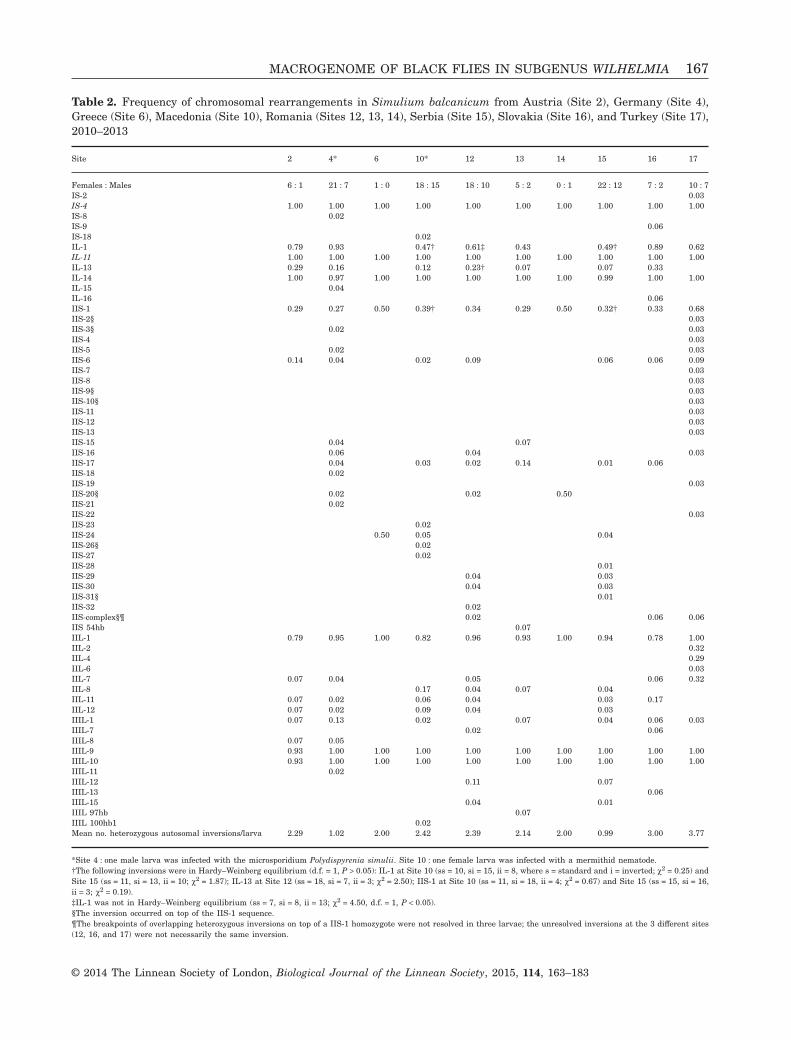
Table 2. Frequency of chromosomal rearrangements in Simulium balcanicum from Austria (Site 2), Germany (Site 4),Greece (Site 6), Macedonia (Site 10), Romania (Sites 12, 13, 14), Serbia (Site 15), Slovakia (Site 16), and Turkey (Site 17),2010–2013
Site 2 4* 6 10* 12 13 14 15 16 17
Females : Males 6 : 1 21 : 7 1 : 0 18 : 15 18 : 10 5 : 2 0 : 1 22 : 12 7 : 2 10 : 7IS-2 0.03IS-4 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00IS-8 0.02IS-9 0.06IS-18 0.02IL-1 0.79 0.93 0.47† 0.61‡ 0.43 0.49† 0.89 0.62IL-11 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00IL-13 0.29 0.16 0.12 0.23† 0.07 0.07 0.33IL-14 1.00 0.97 1.00 1.00 1.00 1.00 1.00 0.99 1.00 1.00IL-15 0.04IL-16 0.06IIS-1 0.29 0.27 0.50 0.39† 0.34 0.29 0.50 0.32† 0.33 0.68IIS-2§ 0.03IIS-3§ 0.02 0.03IIS-4 0.03IIS-5 0.02 0.03IIS-6 0.14 0.04 0.02 0.09 0.06 0.06 0.09IIS-7 0.03IIS-8 0.03IIS-9§ 0.03IIS-10§ 0.03IIS-11 0.03IIS-12 0.03IIS-13 0.03IIS-15 0.04 0.07IIS-16 0.06 0.04 0.03IIS-17 0.04 0.03 0.02 0.14 0.01 0.06IIS-18 0.02IIS-19 0.03IIS-20§ 0.02 0.02 0.50IIS-21 0.02IIS-22 0.03IIS-23 0.02IIS-24 0.50 0.05 0.04IIS-26§ 0.02IIS-27 0.02IIS-28 0.01IIS-29 0.04 0.03IIS-30 0.04 0.03IIS-31§ 0.01IIS-32 0.02IIS-complex§¶ 0.02 0.06 0.06IIS 54hb 0.07IIL-1 0.79 0.95 1.00 0.82 0.96 0.93 1.00 0.94 0.78 1.00IIL-2 0.32IIL-4 0.29IIL-6 0.03IIL-7 0.07 0.04 0.05 0.06 0.32IIL-8 0.17 0.04 0.07 0.04IIL-11 0.07 0.02 0.06 0.04 0.03 0.17IIL-12 0.07 0.02 0.09 0.04 0.03IIIL-1 0.07 0.13 0.02 0.07 0.04 0.06 0.03IIIL-7 0.02 0.06IIIL-8 0.07 0.05IIIL-9 0.93 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00IIIL-10 0.93 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00IIIL-11 0.02IIIL-12 0.11 0.07IIIL-13 0.06IIIL-15 0.04 0.01IIIL 97hb 0.07IIIL 100hb1 0.02Mean no. heterozygous autosomal inversions/larva 2.29 1.02 2.00 2.42 2.39 2.14 2.00 0.99 3.00 3.77
*Site 4 : one male larva was infected with the microsporidium Polydispyrenia simulii. Site 10 : one female larva was infected with a mermithid nematode.†The following inversions were in Hardy–Weinberg equilibrium (d.f. = 1, P > 0.05): IL-1 at Site 10 (ss = 10, si = 15, ii = 8, where s = standard and i = inverted; χ2 = 0.25) andSite 15 (ss = 11, si = 13, ii = 10; χ2 = 1.87); IL-13 at Site 12 (ss = 18, si = 7, ii = 3; χ2 = 2.50); IIS-1 at Site 10 (ss = 11, si = 18, ii = 4; χ2 = 0.67) and Site 15 (ss = 15, si = 16,ii = 3; χ2 = 0.19).‡IL-1 was not in Hardy–Weinberg equilibrium (ss = 7, si = 8, ii = 13; χ2 = 4.50, d.f. = 1, P < 0.05).§The inversion occurred on top of the IIS-1 sequence.¶The breakpoints of overlapping heterozygous inversions on top of a IIS-1 homozygote were not resolved in three larvae; the unresolved inversions at the 3 different sites(12, 16, and 17) were not necessarily the same inversion.
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 167
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

Table 3. Frequency of chromosomal rearrangements in three species of Simulium (Wilhelmia) from Armenia (Site 1),France (Site 3), Germany (Sites 4, 5), Iran (Site 7), Italy (Site 8), Japan (Site 9), Poland (Site 11), Turkey (Site 17), andEngland (Site 18), 1999 and 2010–2013
Site
Simulium lineatum Simulium takahasii Simulium turgaicum
3 4 5* 8 11 18* 9 1 7* 17*
Females : Males 5 : 7 0 : 1 9 : 10 4 : 2 1 : 0 31 : 18 19 : 30 1 : 0 22 : 12 82 : 61
IS-1 0.01IS-3 0.03IS-4 1.00 0.76 1.00 0.50 1.00IS-5 0.05 0.06IS-6 1.00 0.32 1.00 0.50 0.16†IS-7 0.03 0.08IS-10 0.04IS-11 0.02IS-12‡ 0.07IS-13‡ 0.07IS-14‡ 0.07IS-15‡ 0.07IS-16‡ 0.07IS-17 0.01IS N.O.§ 0.01IL-2 < 0.01IL-3a¶ < 0.01IL-3b¶ 0.32†IL-4 < 0.01IL-5 < 0.01IL-6 < 0.01IL-7 < 0.01IL-8 < 0.01IL-9 < 0.01IL-10 0.88 0.84 1.00 1.00IL-11 0.96 0.55 1.00 0.98IL-12 0.03IL-17 0.39†IL-18 0.24†IL-19 0.50IL-20 0.04IL-21 0.01IL 29hb 1.00IL 34hb 0.13IL 35hb 0.06IL 41hb < 0.01IL 29hb 1.00IIS-14 0.08 0.42IIS-25 0.50IIS 45i 0.01IIL-3 0.05IIL-5 < 0.01IIL-8 0.58 1.00 0.66† 1.00 0.39†IIL-9 1.00 0.50† 1.00 0.81IIL-10 0.21 0.08IIL-11 0.58 0.50 0.66 0.83 1.00 0.35†IIL-12 0.29IIL-13 0.03IIL-14 0.16 0.08 0.08IIL-15** 0.07IIL-16** 0.07IIL-17 0.01IIL-18 0.01
168 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

pupae of S. balcanicum from Site 2 (N = 2); S. linea-tum from Sites 3 (N = 7), 8 (N = 3), and 18 (N = 1);S. takahasii from Beijing, China (N = 3); andS. turgaicum from Sites 7 (N = 3) and 17 (N = 5).Principal components analysis was used to displaydifferences in gill measurements including the ratiosof filaments 5 : 2 and 3 : 1 in multidimensional space.
STATISTICAL ANALYSIS OF INVERSION FREQUENCIES
Only S. balcanicum was collected in adequatenumbers (7–34 larvae/site) over a sufficient number ofsites (N = 8) for analysis of inversion frequencies.Samples were collected across north–south and east–west gradients (Table 1). These gradients represent
not only changes in location, but also measures ofdistance. Correlations were therefore used to comparethe relationships of distance to degrees north andeast. These correlations were based on all pairwisecomparisons of sites, rendering the data inherentlynon-independent; significance tests therefore werebased on 1000 random permutations (Lunneborg,2000). Regression analysis was used to determinewhether the distance gradients between sites haddirectionality. For example, are sites in more northernlocations also found in western, as opposed to eastern,locations? Because there is no predictor or responsevariable in this case, the regression equation wascomputed using reduced major-axis regression (Sokal& Rohlf, 1995), with degrees north arbitrarily assigned
Table 3. Continued
Site
Simulium lineatum Simulium takahasii Simulium turgaicum
3 4 5* 8 11 18* 9 1 7* 17*
Females : Males 5 : 7 0 : 1 9 : 10 4 : 2 1 : 0 31 : 18 19 : 30 1 : 0 22 : 12 82 : 61
IIL-19†† 0.02IIL-20 0.01IIL-21 0.01IIL 63hb 0.08IIIS-1 0.01IIIS-2 0.13 0.01IIIS-3 1.00IIIS-4 0.02IIIS 77hb < 0.01IIIL-2 < 0.01IIIL-3 < 0.01IIIL-4 < 0.01IIIL-5 1.00 0.50 0.96 1.00 1.00 1.00IIIL-6 0.50 0.03IIIL-10 1.00IIIL-14 0.07IIIL-complex 0.50 0.05IIIL 99i 1.00IIIL 100hb2 0.01Mean no. heterozygous autosomal inversions/larva 1.42 5.00 4.68 1.33 4.00 4.31 0.12 0.00 0.03 0.12
*Site 5 : 1 male larva was infected with the microsporidium Amblyospora sp., and 1 male larva was infected with a mermithid nematode. Site 18:one male larva was infected with Amblyospora varians. Site 7: one male larva was infected with Amblyospora bracteata. Site 17: one male larvawas infected with the microsporidium Polydispyrenia simulii; one male larva was infected with Amblyospora bracteata, and breakpoints of a mid-ISheterozygous inversion were not determined for this individual.†The following inversions were in Hardy-Weinberg equilibrium (d.f. = 1, P > 0.05): IS-6 at Site 18 (ss = 35, si = 12, ii = 2, where s = standard andi = inverted; χ2 = 0.53); IL-3b at Site 18 (ss = 24, si = 18, ii = 7; χ2 = 1.33); IL-17 at Site 18 (ss = 21, si = 18, ii = 10; χ2 = 2.51); IL-18 at Site 18 (ss = 31,si = 14, ii = 5; χ2 = 2.70); IIL-8 at Site 5 (ss = 4, si = 5, ii = 10; χ2 = 3.28) and Site 18 (ss = 17, si = 26, ii = 6; χ2 = 0.68); IIL-9 at Site 5 (ss = 5, si = 9,ii = 5; χ2 = 0.05); IIL-11 at Site 18 (ss = 21, si = 22, ii = 6; χ2 = 0.00).‡Five inversions (IS-12, IS-13, IS-14, IS-15, and IS-16) were completely linked and paired with the standard homologue in seven larvae (six females,one male).§Heterozygous for nucleolar organizer (N.O.) expression.¶Although IL-3a and IL-3b have the same macro-breakpoints, they must be independent inversions because IL-3b occurred only if IL-10 waspresent, whereas IL-3a occurred without IL-10.**IIL-15 and IIL-16 were completely linked and were found in all, and only, the same seven larvae with linked inversions IS-12, IS-13, IS-14, IS-15,and IS-16.††IIL-19 occurred only on top of IIL-9.
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 169
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

to the left side of the equation. A distance–decayrelationship (DDR) between site similarity, based oninversion-frequency profiles, and Euclidean distancebetween sites was examined using linear regression(Soininen, McDonald & Hillebrand, 2007). Similaritybetween sites was calculated as the Bray–Curtisresemblance coefficient of the square-root transformedfrequency of inversions (McCune & Grace, 2002). Weestablished a priori that the frequency of an inversionhad to be greater than zero for at least two of the eightsites to be included in the analysis. Significance testsof the DDR were based on 1000 random permutations(Lunneborg, 2000). Both similarity and distance wereexpressed on a log-log scale for this regression. Bydefinition, when a site is compared to itself, it will have100% similarity in the inversion profile and zerodistance, which could increase the overall strength ofthe DDR unfairly. Accordingly, we used a trimmed dataset in which these values were removed. Relationshipsbetween individual inversions and degrees north andeast were examined with simple linear regression(log-log scale).
RESULTSGENERALITIES
The characteristic Wilhelmia whole-arm interchange(IS + IIL, IL + IIS) was present in all larvae. Thecentromeres of the three metacentric chromosomeswere associated tightly in a persistent chromocentrein all nuclei. Although additional heterochromatinthat would suggest a genuine chromocentre (Fig. 2)was not evident in the majority of populations, it waspresent, although minimal, in some populations (e.g.
Site 10). Ectopic pairing of telomeres occurred insome populations of S. balcanicum (e.g. Site 15) andS. lineatum (e.g. Sites 3 and 11). The telomeres, espe-cially of IIS and, to a lesser extent IIIS (Fig. 8B), ofS. takahasii typically had extra heterochromatin.Sticky heterochromatin in IIIL, manifesting as bandsadhering to one another, particularly in section 92(Fig. 10B), was typical of populations at some sites(e.g. S. balcanicum at Site 10, S. turgaicum at Site17, and, less frequently, S. takahasii at Site 9).
Classic chromosomal markers that are widespreador universal in the Simuliidae (Rothfels, Feraday& Kaneps, 1978), such as the parabalbiani, werepresent in all material (Figs. 2, 3, 4, 5, 6, 7, 8, 9, 10).The doublet marker (dt; Fig. 7), a conspicuoussubterminal pair of bands in IIL (section 71),although taxonomically more restricted, minimally tothe subgenus Wilhelmia, provided utility as an addi-tional marker. The nucleolar organizer was in thebase of IS (Figs 2, 3). The bulge in IIS was cleaved bythe ring of Balbiani (Fig. 6). The end of IIIS wastypically flared, obscuring the banding in section 72.Clusters of chromatids could be discerned in theflared region at some sites (e.g. Site 17), producing abraided appearance. Flaring of the IIIS terminus wassuppressed in some nuclei of S. balcanicum (e.g. Sites10, 15, and 17), S. turgaicum (Site 17; Fig. 8A), andS. takahasii (Site 9).
More than 110 inversions were found among 480larvae, with 59 in S. balcanicum (N = 165 larvae), 41in S. lineatum (N = 88 larvae), six in S. takahasii(N = 49), and 14 in S. turgaicum (N = 178). Norearrangements were significantly linked (P > 0.05)to gender in any population. The sex chromosomes,therefore, were microscopically undifferentiated(X0Y0) in all populations. Additional rearrangements,particularly heterobands, were distributed among thefour species (Tables 2, 3).
SIMULIUM BALCANICUM
Ten sites had populations of S. balcanicum (N = 165).Two inversions, IS-4 and IL-11 (Fig. 5), were fixed inall individuals of S. balcanicum, and three additionalinversions (IL-14, IIIL-9, and IIIL-10) (Figs 4, 10A)were almost fixed (> 0.98), with only two larvaeheterozygous for IL-14 and one heterozygous forIIIL-9 and IIIL-10. Three additional inversions hadhigh representation in S. balcanicum across sites.IL-1 (Fig. 5B) was found in 61.8% of all homologues,and IIS-1 (Fig. 6) in 39.7%. The latter inversionserved as the base sequence on top of which at leasteight additional inversions were built (Table 2). IIL-1(Fig. 7) occurred in 91.2% of all homologues acrosssites, and was fixed in Turkish samples. InversionIIL-7 (Fig. 7), which occurred only on top of IIL-1,
Figure 2. Chromocentre of male larva of Simuliumturgaicum, showing bases of six chromosome arms (IS–IIIL), B chromosome (B), and nucleolar organizer (N.O.)
170 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183
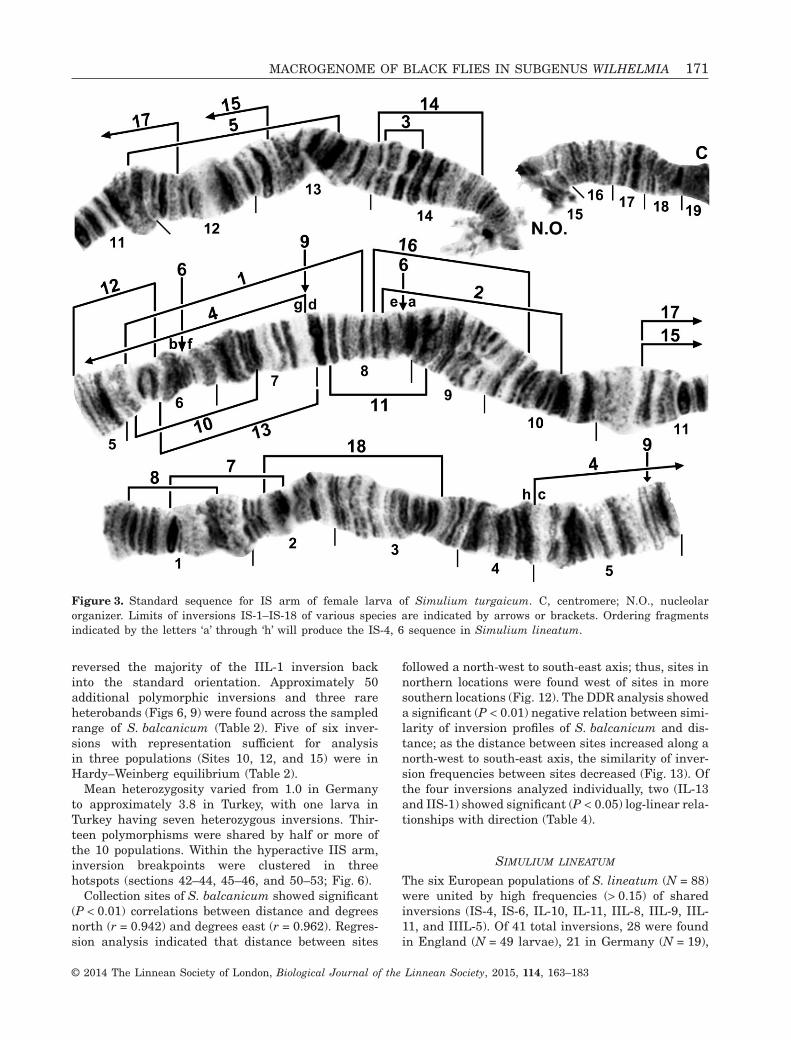
reversed the majority of the IIL-1 inversion backinto the standard orientation. Approximately 50additional polymorphic inversions and three rareheterobands (Figs 6, 9) were found across the sampledrange of S. balcanicum (Table 2). Five of six inver-sions with representation sufficient for analysisin three populations (Sites 10, 12, and 15) were inHardy–Weinberg equilibrium (Table 2).
Mean heterozygosity varied from 1.0 in Germanyto approximately 3.8 in Turkey, with one larva inTurkey having seven heterozygous inversions. Thir-teen polymorphisms were shared by half or more ofthe 10 populations. Within the hyperactive IIS arm,inversion breakpoints were clustered in threehotspots (sections 42–44, 45–46, and 50–53; Fig. 6).
Collection sites of S. balcanicum showed significant(P < 0.01) correlations between distance and degreesnorth (r = 0.942) and degrees east (r = 0.962). Regres-sion analysis indicated that distance between sites
followed a north-west to south-east axis; thus, sites innorthern locations were found west of sites in moresouthern locations (Fig. 12). The DDR analysis showeda significant (P < 0.01) negative relation between simi-larity of inversion profiles of S. balcanicum and dis-tance; as the distance between sites increased along anorth-west to south-east axis, the similarity of inver-sion frequencies between sites decreased (Fig. 13). Ofthe four inversions analyzed individually, two (IL-13and IIS-1) showed significant (P < 0.05) log-linear rela-tionships with direction (Table 4).
SIMULIUM LINEATUM
The six European populations of S. lineatum (N = 88)were united by high frequencies (> 0.15) of sharedinversions (IS-4, IS-6, IL-10, IL-11, IIL-8, IIL-9, IIL-11, and IIIL-5). Of 41 total inversions, 28 were foundin England (N = 49 larvae), 21 in Germany (N = 19),
Figure 3. Standard sequence for IS arm of female larva of Simulium turgaicum. C, centromere; N.O., nucleolarorganizer. Limits of inversions IS-1–IS-18 of various species are indicated by arrows or brackets. Ordering fragmentsindicated by the letters ‘a’ through ‘h’ will produce the IS-4, 6 sequence in Simulium lineatum.
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 171
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

11 in Italy (N = 6), nine in France (Corsica) (N = 12),and eight in the single larva from Poland. The meannumber of heterozygous inversions per larva rangedfrom 1.3 in Italy to more than 4.3 in England andGermany, with one larva in Germany having eightheterozygous inversions and two in England having13 each. Of the 11 inversions common to England andGermany, four [all in high frequency (> 0.50) inGermany] were fixed (IS-4, IL-10, and IIIL-5) or
almost fixed (0.98 for IL-11) in England. With fiveexceptions (IL-3b, IL-17, IL-18, IIL-10, and IIL-12),all inversions unique to either England or Germanywere in low (< 0.09) frequency. All inversions forwhich genotypic frequencies were adequate for statis-tical analysis (Site 5: IIL-8 and IIL-9; Site 18: IS-6,IL-3b, IL-17, IL-18, IIL-8, and IIL-11) were in Hardy–Weinberg equilibrium (Table 3). Six heterobands werefound (Figs 4, 7, 9; Table 3).
Figure 4. Standard sequence for IL arm of female larva of Simulium turgaicum. C, centromere; hb, location ofheteroband; m, marker (sensu Rothfels et al., 1978). Limits of inversions IL-2–IL-21 of various species are indicated bybrackets. The limits of inversion IL-14 of Simulium balcanicum are bracketed.
Table 4. Simple linear regression between the frequency of an inversion across sites (N = 8) for Simulium balcanicumand latitude and longitude; response and predictor variables were log10 transformed
Inversion Gradient Slope r2(adj) F1,6 P
IL-1 Latitude 3.67 0.104IL-1 Longitude 2.35 0.176IL-13 Latitude 0.0247 52.6% 8.76 0.025IL-13 Longitude 3.74 0.101IIS-1 Latitude –0.0321 73.1% 20.00 0.004IIS-1 Longitude 0.0198 78.6% 26.76 0.002IIL-1 Latitude 1.25 0.306IIL-1 Longitude 1.80 0.228
172 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

The English population demonstrated completeinversion linkage within chromosomes; the invertedsequences for IS-12, IS-13, IS-14, IS-15, and IS-16(Fig. 3) were traced on the same homologue (i.e. in cisconformation) in all seven larvae (six females, onemale). These same seven larvae also showed 100%linkage (cis conformation) of IIL-15 and IIL-16(Fig. 7). None of these IS or IIL inversions appearedindependently. We cannot be certain, however, thatthe IS and IIL inversions came from the same parent.The complement of these seven individuals was nomore loosely paired than that of typical individuals.
The following eight inversions in our study of S.lineatum were equivalent to those (in parentheses)found by Weber & Grunewald (1989): IS-4 (= IS-4),IS-5 (= IS-6), IIS-14 (= IIS-5), IIL-8 (= IIL-7), IIL-9
(= IIL-3), IIL-11 (= IIL-5), IIL-12 (= IIL-4), and IIL-14(= IIL-8).
SIMULIUM TAKAHASII
One sample of S. takahasii (N = 49) from Japan wasavailable for study (Table 1). All 49 larvae had thesame banding sequence, with two fixed inversions,IIIS-3 (Fig. 8B) and IIIL-10 (Fig. 10C). A darklystaining, glassy band of variable thickness washomozygously inserted in section 99 of all larvae;the chromosome typically was constricted at theband (Fig. 10C). Polymorphisms were minimal (fre-quencies < 0.02), with only four floating inversions,heterozygous expression of the nucleolar organizer,and a heterozygous band insert in section 45 of IIS
Figure 5. Chromosome I of female larva of Simulium balcanicum. A, IS (sections 4–9 only). The IS-4 sequence is present,with limits bracketed. Limits of inversions IS-6 and IS-9 are indicated by brackets. B, IL (sections 31–41 only). IL-1, IL-11,and IL-14 are present, with inversion limits indicated by brackets (IL-1, IL-14) or arrows (IL-11). Ordering fragmentsindicated by the letters ‘a’ through ‘h’ will produce the standard sequence. Limits of inversions IL-13, IL-15, and IL-16are indicated by brackets.
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 173
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

(Fig. 6, Table 3). The level of inversion heterozygo-sity in the population, consequently, was low(0.12).
SIMULIUM TURGAICUM
The three West Asian populations of S. turgaicum(N = 178) were characterized by a low level of poly-morphism. Of 14 total inversions, 13 were found incentral Turkey (N = 143 larvae) and one in Iran(N = 34), all with frequencies of 0.01 or less (Table 3).IIIS-1 in our system differed by one band from IIIS-1of Weber & Grunewald (1989). The mean number ofheterozygous inversions per larva was less than 0.15per site. Large supernumerary (B) chromosomes werepresent in one female and two male larvae fromTurkey. They paired at one end with the chromocentrein approximately 40% of the nuclei, suggesting anear-terminal centromere (Fig. 2). The puffed distalregion possibly signalled the presence of a nucleo-lar organizer (Procunier, 1975). Two heterobandsin low frequency (< 0.01) were discovered (Figs 4, 8;Table 3).
CHROMOSOMAL COMPARISONS
The distribution of inversions differed significantlyamong the four species (χ2 = 47.06, d.f. = 4, P < 0.001)(Table 5). Simulium balcanicum differed from allother species by two fixed inversions (IS-4 andIL-11), two almost fixed (i.e. frequencies ≥ 0.98)inversions (IL-14 and IIIL-9), and a unique set ofpolymorphisms. It shared IIIL-10 with S. takahasii,which had one unique fixed inversion (IIIS-3). Norearrangements were shared between S. balcanicumand S. turgaicum. However, five inversions wereshared between S. balcanicum and S. lineatum: IS-4,IL-11, IIL-8, IIL-11, and IIL-12. The number of inver-sions across chromosome arms was not independentof the two species (χ2 = 29.894, d.f. = 4, P < 0.001; IIISwas omitted from analysis because of only one inver-sion) (Table 5). Simulium lineatum, S. takahasii, andS. turgaicum shared no rearrangements. Simuliumlineatum could be distinguished from S. turgaicum byits high frequency of inversions (e.g. IS-4, IL-10,IL-11, IIL-9, and IIIL-5), which typically were presentin half or more of the chromosomal homologues ateach site. No hybrids were detected at any sites where
Figure 6. Standard sequence for IIS arm of female larva of Simulium turgaicum. Limits of inversions IIS-1–IIS-32 areindicated by arrows or brackets. Inversions above the chromosome are specific to Simulium balcanicum; those below thechromosome are specific to Simulium lineatum. Inversions IIS-2, IIS-3, IIS-9, IIS-10, IIS-20, IIS-26, and IIS-31 occur onlyon top of the IIS-1 inversion. Bu, bulge; C, centromere; hb, location of heteroband; i, location of band insert; RB, ring ofBalbiani; ss, shoestring marker; the two halves of the bulge are separated by the ring of Balbiani. Ordering fragmentsindicated by the letters ‘a’ through ‘h’ will produce the IIS-1, 2 sequence of S. balcanicum.
174 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

S. balcanicum occurred in the same rivers withS. lineatum (Germany) and S. turgaicum (Turkey).
MORPHOLOGICAL COMPARISONS
Only the gill of S. balcanicum, with its petiolatepair of filaments, was consistently distinguishablefrom the gills of the other three species. A principal
components analysis of gill lengths and selected ratiossuggested that S. takahasii was distinct but thatS. turgaicum was indistinguishable from S. lineatum,although sample sizes were small (Fig. 14). The firsttwo axes of the principal components analysisexplained 83.7% of the total variation in gill morphol-ogy; therefore, only these two were used to graph gilldimensions in multidimensional space.
Figure 7. Standard sequence for IIL arm of female larva of Simulium turgaicum. C, centromere; DNA, DNA puff; dt,doublet; gB, grey band; hb, location of heteroband; j, jagged marker; Pb, parabalbiani; pf, puffing band. Limits ofinversions IIL-1–IIL-21 of various species are indicated by brackets. Ordering fragments indicated by the letters ‘a’through ‘h’ will produce the IIL-9, 14 sequence of Simulium lineatum, and ordering the letters ‘a’ through ‘f ’ will producethe IIL-1, 7 sequence of Simulium balcanicum.
Table 5. Distribution of all inversions, fixed and floating, in each chromosome arm of 4 species in the subgenus Wilhelmiaacross 18 sites
Larvae (N) Inversions (N) IS IL IIS IIL IIIS IIIL
Simulium balcanicum 165 59 5 6 31 8 0 9Simulium lineatum 88 41 13 8 2 13 1 4Simulium takahasii 49 6 0 1 0 2 2 1Simulium turgaicum 178 14 1 8 0 1 1 3
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 175
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

DISCUSSION
Our samples can be partitioned into four chromo-somal segregates, which we recognize as sepa-rate species (S. balcanicum, S. lineatum, S. taka-hasii, and S. turgaicum), each supported byunique cytogenetic characters and geographicaldistributions.
These four species can be distinguishedchromosomally at a glance from two sympatricmembers of the subgenus Wilhelmia, S. equinumand S. pseudequinum Séguy, by the presence ofa chromocentre (Weber & Grunewald, 1989;Chubareva & Petrova, 2008). Although ectopicpairing of centromeres occurs in some populations ofS. equinum, it is not present in all nuclei, and theexpanded centromere region of chromosome I inS. equinum contrasts markedly with the slender con-dition in S. balcanicum, S. lineatum, S. takahasii,and S. turgaicum. The positions of the bulge andring of Balbiani also are diagnostic. In our four taxa,they are in the distal one-third of the arm whenin the standard sequence, with the bulge proximalto the ring of Balbiani, whereas, in S. equinumand S. pseudequinum, they are subterminal, withthe bulge distal, unless displaced by floating inver-sions (Weber & Grunewald, 1989). Simuliumparaequinum, which also has a chromocentre, hasthe bulge and ring of Balbiani in the proximal one-third of the arm, nearer the centromere (Petrovaet al., 2003).
SPECIES STATUS
Our material of S. balcanicum from Germany andTurkey almost brackets the geographical range, whichextends from Italy (Rivosecchi, 1978) and our collec-tion site in Germany eastward to the Krasnodar Ter-ritory of Russia (Grinchuk & Chubareva, 1974). Oursample from Bârza, Romania, is within 200 km of thetype locality (Lakatnik) in the Balkan Mountains ofBulgaria. The chromosomes reveal unequivocally thatS. balcanicum is a distinct species, with no evidence ofcryptic species, across its range of approximately2000 km. Polymorphisms unique to one or a few popu-lations are in low frequency, typically less than 0.10per site. Common inversions exhibit a longitudinalgradient of frequencies, with a north–south tilt,although whether the frequencies are driven by selec-tion in specific environments or by restricted gene flow,or both, is unknown. Inversion-frequency gradientsare common in a number of simuliid species, typicallygrading toward loss or fixation at the distributionalperiphery (Rothfels & Featherston, 1981), a patternalso documented in Drosophila (Stocker, Foley &Hoffmann, 2004). We conclude that our samples ofS. balcanicum are conspecific with the type specimenand with the types representing the three synonyms(secundum, danubiense, and severinense). The petio-late pair of pupal gills, originally described by Rubtsov(1956), provides the only diagnostic structural featurefor S. balcanicum in any life stage (Crosskey & Zwick,2007).
Figure 8. Chromosome IIIS of Simulium (Wilhelmia) species. bl, blister; C, centromere; ca, capsule; hb, heteroband(shown as heterozygous). A, standard sequence for IIIS arm of female larva of Simulium turgaicum, showing suppressedterminal flaring (sections 72–73). Limits of inversions IIIS-1 and IIIS-2 and fixed inversion IIIS-3 are indicated bybrackets; autosomal inversion IIIS-4 of Simulium takahasii, indicated by arrows, occurs only on top of IIIS-3; the twoinversions share near-coincident proximal breakpoints such that IIIS-4 is based on the fixed IIIS-3 sequence. B, malelarva of S. takahasii; IIIS-3 and IIIS-4 as a heterozygote (inverted on lower homologue) are present.
176 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

As S. balcanicum increases in prevalence eastward,S. lineatum diminishes in prevalence and is replacedby S. turgaicum. Where S. balcanicum overlaps geo-graphically with S. lineatum, it typically occurs indifferent rivers. Our additional, albeit limited, find-ings of S. balcanicum in south-eastern Germany (G.Seitz, unpubl. data) suggest that it might be expand-ing its range westward.
Our German sample of S. lineatum probably repre-sents the type specimen of S. lineatum; the type local-ity in Stolberg, Germany, is approximately 490 kmnorth-east of our nearest sampling site (Landshut).Our English sample of S. lineatum is approximately215 km south of the type locality of S. salopiense andapproximately 1000 km west of our German sample.English and German populations differ only in thefrequency of rearrangements and the presence ofunique, rare inversions, which are not unexpected,
given the geographical distance. We, therefore, agreewith the synonymy of salopiense with lineatum origi-nally proposed by Crosskey & Davies (1972).Although similarly having a high frequency (> 0.15) ofthe same inversions (IS-4, IS-6, IL-10, IL-11, IIL-8,IIL-9, IIL-11, and IIIL-5) as in England and Germany,the Corsica and Italian populations have fewerpolymorphisms, perhaps reflecting their locations atthe periphery of the distribution.
Our analyses provide no concrete evidence forcryptic species in S. lineatum s.s., nor did the analysisby Weber & Grunewald (1989) of populations insouth-western Germany. Thus, the currently recog-nized synonymy (Adler & Crosskey, 2014) of falcula(Enderlein) with lineatum is valid. One caveat,however, is in order. Our English population ofS. lineatum has the highest number of heterozygousinversions (N = 13) and of linked inversions (≥ 5) ever
Figure 9. Standard sequence for IIIL arm of female larva of Simulium turgaicum. C, centromere; hb, location ofheteroband; 2 + 1, marker with two black bands and one grey band, possibly homologous with bands in section 93 of thestandard sequence for the subgenus Simulium. Limits of inversions IIIL-1–IIIL-6, IIIL-8, and IIIL-11–IIIL-15 areindicated by brackets. Positions of heterobands in section 97 of Simulium balcanicum and one each in section 100 ofS. balcanicum (hb1) and Simulium lineatum (hb2), and of a band insert in section 99 of Simulium takahasii (i) areindicated by arrows. Ordering fragments indicated by the letters ‘a’ through ‘h’ will produce the IIIL-9, 10 sequence ofS. balcanicum.
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 177
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183
recorded for a single simuliid larva (Adler et al.,2010). The seven individuals with linked inversions inIS and in IIL might represent products of introgres-sion, although we found no evidence of loose somatic
pairing of homologues typical of hybrid individuals(Rothfels & Nambiar, 1981). The only other knownspecies of the subgenus Wilhelmia in Britain areS. equinum and S. pseudequinum, and their chromo-somal features are markedly different from thoseof S. lineatum. The possibility of cryptic taxa inS. lineatum will require additional sampling, particu-larly in England.
We resurrect the name turgaicum from synonymywith lineatum and apply it to our West Asianpopulations (Armenia, Iran, and Turkey) formerlyknown as S. lineatum. Rubtsov (1956) describedS. turgaicum from Almaty, Kazakhstan, approxi-mately 2500 km east of our nearest sampling site(Iran), and reported that the larvae and pupae occur insemidesert, small, warm rivers and irrigation chan-nels, as well as cold rivers up to elevations of 2000 m inthe Transcaucasus and Central Asia. Former Sovietstudies (Grinchuk & Chubareva, 1974) consistentlyidentified Armenian populations as S. turgaicum.
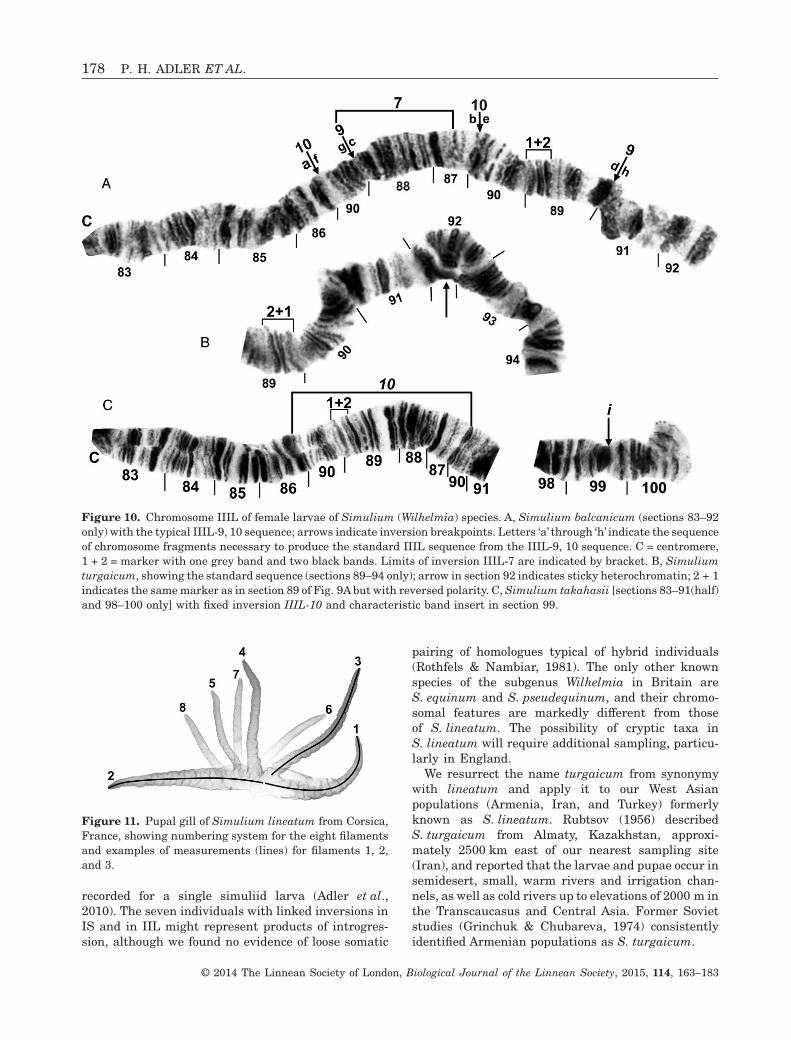
Figure 10. Chromosome IIIL of female larvae of Simulium (Wilhelmia) species. A, Simulium balcanicum (sections 83–92only) with the typical IIIL-9, 10 sequence; arrows indicate inversion breakpoints. Letters ‘a’ through ‘h’ indicate the sequenceof chromosome fragments necessary to produce the standard IIIL sequence from the IIIL-9, 10 sequence. C = centromere,1 + 2 = marker with one grey band and two black bands. Limits of inversion IIIL-7 are indicated by bracket. B, Simuliumturgaicum, showing the standard sequence (sections 89–94 only); arrow in section 92 indicates sticky heterochromatin; 2 + 1indicates the same marker as in section 89 of Fig. 9A but with reversed polarity. C, Simulium takahasii [sections 83–91(half)and 98–100 only] with fixed inversion IIIL-10 and characteristic band insert in section 99.
Figure 11. Pupal gill of Simulium lineatum from Corsica,France, showing numbering system for the eight filamentsand examples of measurements (lines) for filaments 1, 2,and 3.
178 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

The chromosomal distinction between S. lineatumand S. turgaicum cannot be evaluated in sympatry.Our nearest samples are approximately 2100 kmapart. Whether the distributions actually overlap isnot known. If S. lineatum and S. turgaicum aresympatric, the zone of overlap probably liesin southern Russia or in the Balkans, perhapsBulgaria, the easternmost reliable record forS. lineatum, based on the listing by Kovatschev(1979). One of the few putatively diagnostic featuresof S. turgaicum, the greater length of theanteriormost filaments of the pupal gill (Rubtsov,
1956), does not hold consistently. Some ecologicaldistinction, nonetheless, might exist. Simuliumlineatum is a lowland species, typically occupyinghabitats below 500 m a.s.l, whereas S. turgaicumis common at higher elevations (> 900 m). Thealtitudinal differences are reminiscent of siblingspecies in the S. ochraceum complex (Hirai et al.,1994). If the uniquely shared polymorphismsof S. balcanicum and S. lineatum are the result ofcommon ancestry, rather than introgression, thespecies status of S. turgaicum is further supportedby its relationship as sister to S. balcanicum plus
Figure 12. Reduced major-axis regression between latitude and longitude coordinates for eight sites from whichchromosomal inversion profiles of Simulium balcanicum were analyzed.
Figure 13. Distance–decay relationship between site similarity, based on chromosomal inversion profiles of Simuliumbalcanicum, and distance between sites.
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 179
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

S. lineatum, rather than to S. lineatum with whichit historically has been in synonymy.
The full geographical range of S. turgaicum isunknown but is considered here to extend from Turkeyeastward at least to the type locality in Kazakhstan. Itprobably extends well beyond Kazakhstan deep intoCentral Asia. Whether its range overlaps that ofS. takahasii is unknown. Simulium takahasii occupiesthe eastern perimeter of the Palearctic Region in east-ern China, Japan, and Korea (Adler & Crosskey, 2014);we consider the record of S. lineatum from Beijing(Adler & Crosskey, 2014) to apply to S. takahasii.
We continue to recognize S. takahasii as a validspecies, primarily on pragmatic grounds (to maintainthe current nomenclatural status) rather than onthe basis of the few chromosomal differences fromS. turgaicum. Whether S. takahasii is a valid speciesor conspecific with S. turgaicum suffers from thelack of an opportunity to test reproductive isolationin sympatry. Our collections of S. takahasii andS. turgaicum are separated by more than 7250 km.This geographical gap leaves open the question ofwhether some or all of the additional, putative speciesof S. (Wilhelmia), described from China and basedon questionable structural characters, might beconspecific with S. turgaicum or S. takahasii, includ-ing Simulium germuense Liu, Gong, Zhang, Luo &An; Simulium pekingense Sun; Simulium qinghaienseLiu, Gong, Zhang, Luo & An; Simuliumqingxilingense Cai & An; Simulium tachengenseMahe, An & Yan; Simulium tongbaishanense Chen &Luo; and Simulium xingyiense Chen & Zhang. Only
the polytenes of S. xingyiense have been published(Huang et al., 2012), although the low-magnificationresolution of the maps is not sufficient to allow com-parison with our populations.
CHROMOSOMAL RELATIONSHIPS
Our evaluation of the relationships of the four taxa isinfluenced by the choice of the standard bandingsequence and by the scrambled sequence of the com-plement, which currently does not invite comparisonwith sequences of known taxa that could serve asoutgroups. We can determine the polarity of onlyone relevant band sequence through outgroup com-parison. Our standard sequence for IL-14 (sections40–41) is equivalent to the standard sequence forthe Simulium vernum group (sections 42C–44C,Brockhouse, 1985), the subgenus Simulium (sections39c–41, Rothfels et al., 1978), and Simuliumerythrocephalum (De Geer) (P. H. Adler, unpubl.data). If these taxa are taken as outgroups for thissequence, IL-14 of S. balcanicum is derived.
The chromosomes indicate that S. balcanicumand S. lineatum are more closely related to oneanother than either is to S. turgaicum. Although therelationships are not rooted, the same autosomalpolymorphisms (e.g. IIL-8, IIL-11, and IIL-12) areuniquely shared regardless of the chromosomalsequence chosen as the standard; thus, the sisterrelationship of S. balcanicum and S. lineatum holds.The caveat, however, is that we do not know whetherthe shared rearrangements are primary (derived from
Figure 14. Principal components (PC) analysis of the lengths of all gill filaments and ratios of filaments 5 : 2 and 3 : 1of four species of Simulium (Wilhelmia). The first PC is largely a linear measure of filaments, and the second PC is largelya measure of proportions.
180 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

a common ancestor) or secondary (acquired by intro-gression). Simulium balcanicum and S. lineatum arechromosomally most similar in the area of overlap(Germany), where they share five inversions.
On the basis of one uniquely shared inversion(IIIL-10), S. takahasii is most closely related toS. balcanicum. The relationship, however, might bean artefact of our choice of standard. In other words,if IIIL-10 is actually ancestral, our standard sequencefor this inversion becomes the derived condition andthe relationship of S. takahasii with S. balcanicum isnot supported.
PEST PROBLEMS
The attacks on livestock and humans along Turkey’sKizilirmak River (Yilmaz et al., 2007; Sariözkan et al.,2014) are the most severe recorded for any member ofthe subgenus Wilhelmia. Citizens in Isfahan, Iran,where we have cytotyped S. turgaicum, have com-plained about biting insects, although the situation isobfuscated by a failure to distinguish black flies fromthe common Culex mosquitoes in the area. Simuliumlineatum is a pest of livestock and occasionally ofhumans in parts of Europe (Crosskey, 1990; Werner &Adler, 2005), and S. takahasii is a pest of cattle andhorses in Japan (Bentinck, 1955). No pest problemshave been reported for S. balcanicum, although theabsence of reports might be related to the inability todistinguish the isomorphic females of this speciesfrom those of S. lineatum and S. turgaicum. Peststatus of the group members is related to theirshared propensity to feed on large mammals and toproduce large populations in productive rivers andirrigation ditches near human and livestock foci. TheCappadocia outbreak probably represents an extremeexample of these factors.
GEOGRAPHICAL GENERALISTS AND SPECIALISTS
The existence of four discrete species, rather thanone, reinforces a general pattern in the Simuliidae:wide distributions of putatively single species repre-sent mosaics of geographically restricted crypticspecies. This is not to say that all species of black flieswith large geographical distributions are species com-plexes. Legitimate species with expansive ranges canreflect the ability to capitalize on anthropogenicallyaltered environments (Adler & Kim, 1984). The trend,nonetheless, offers a guide for screening putativespecies for additional biodiversity. Accordingly,S. turgaicum, with the largest range of our four sur-veyed taxa and which has been minimally sampled,might consist of yet additional species.
The discovery that widespread black flies are com-posites of geographical specialists largely reflects the
opportunity provided by the well-banded polytenechromosomes for whole-genome inspection of naturalpopulations. The same trend is apparent in othergroups of organisms that offer the polytene option,notably mosquitoes and drosophilids (Coluzzi et al.,2002; Rohde et al., 2006). This phenomenon, however,is not unique to black flies and other flies with high-quality polytenes. Variation in life-history aspectssuch as habitats, hosts, phenologies, mating behav-iours, and communication systems have long sig-nalled complexes of cryptic species (Mayr, 1963;Bickford et al., 2006). As genomic analyses becomemore powerful and routine, the opportunity fortesting hypotheses of generalist species versus com-plexes of specialists will become more democratized.
ACKNOWLEDGEMENTS
We thank C. E. Beard for excellent technical assis-tance and C. L. Brockhouse and three anonymousreviewers for their insightful comments on the manu-script. This work was funded, in part, by NationalScience Foundation awards DEB-0841636 (Discoveryand Prediction of Hidden Biodiversity in Black Flies(Diptera: Simuliidae)) under the American Recoveryand Reinvestment Act of 2009 and DEB-0933218(MIDGEPEET: A Collaborative Effort to IncreaseTaxonomic Expertise in Understudied Families ofNematocerous Diptera). Funding also came fromThe Scientific and Technological Research Council ofTurkey Research Project 111 O 426 ‘The MolecularClassification of Black Fly (Diptera: Simuliidae)Species Which Pose a Problem in Central KizilirmakBasin and the Investigation of Their Vector Potentialsby Real Time PCR’, VEGA grant no. 1/0561/14 of theMinistry of Education of the Slovak Republic, and theSlovak Research and Development Agency under con-tract No. APVV-0436-12. This is Technical Contribu-tion no. 6253 of the Clemson University ExperimentStation, and is based on work supported in part byNIFA/USDA under project number SC-1700433.
REFERENCES
Adler PH. 2009. Biodiversity of biting flies: implications forhumanity. In: Foottit RG, Adler PH, eds. Insect biodiversity:science and society. Chichester: Wiley-Blackwell Publishing,523–545.
Adler PH, Cheke RA, Post RJ. 2010. Evolution, epidemi-ology, and population genetics of black flies (Diptera:Simuliidae). Infection, Genetics and Evolution 10: 846–865.
Adler PH, Crosskey RW. 2014. World blackflies (Diptera:Simuliidae): a comprehensive revision of the taxonomicand geographical inventory [2014]. Available at: http://www.clemson.edu/cafls/biomia/pdfs/blackflyinventory.pdf
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 181
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

Adler PH, Currie DC, Wood DM. 2004. The black flies(Simuliidae) of North America. Ithaca, NY: Cornell Univer-sity Press.
Adler PH, Huang YT. 2011. Integrated systematics of theSimuliidae (Diptera): evolutionary relationships of the little-known Palearctic black fly Simulium acrotrichum. Cana-dian Entomologist 143: 612–628.
Adler PH, Kim KC. 1984. Ecological characterization of twosibling species, IIIL-l and IS-7, in the Simulium vittatumcomplex (Diptera: Simuliidae). Canadian Journal of Zoology62: 1308–1315.
Bentinck W. 1955. The black flies of Japan and Korea(Diptera: Simuliidae). Tokyo: Contribution from the 406thMedical General Laboratory.
Bickford D, Lohman DJ, Sodhi NS, Ng PKL, Meier R,Winker K, Ingram KK, Das I. 2006. Cryptic species as awindow on diversity and conservation. Trends in Ecologyand Evolution 22: 148–155.
Brockhouse C. 1985. Sibling species and sex chromosomesin Eusimulium vernum (Diptera: Simuliidae). CanadianJournal of Zoology 63: 2145–2161.
Charalambous M, Shelley AJ, Maia Herzog M,Luna Dias APA. 1996. Four new cytotypes of theonchocerciasis vector blackfly Simulium guianensein Brazil. Medical and Veterinary Entomology 10: 111–120.
Chubareva LA, Petrova NA. 2008. Tsitologicheskie kartypolitennykh khromosom i nekotorye morfologicheskieosobennosti krovososushchikh moshek Rossii i sopredel’nykhstran (Diptera: Simuliidae): atlas [Cytological maps ofpolytene chromosomes and some morphological featuresof bloodsucking black flies of Russia and adjacentcountries (Diptera: Simuliidae): atlas]. St Petersburg:Tovarishchestvo Nauchnykh Izdanii KMK, [In Russian].
Chubareva LA, Petrova NA, Reva MV. 2007.Karyotypic and morphological study of five species ofthe genus Wilhelmia Enderlein (Diptera: Simuliidae).Entomologicheskoe Obozrenie 86: 895–904. [In Russian withEnglish summary; full English translation in EntomologicalReview 87: 1290–1299 (2007).].
Coluzzi M, Sabatini A, della Torre A, Di Deco MA,Petrarca V. 2002. A polytene chromosome analysisof the Anopheles gambiae complex. Science 298: 1415–1418.
Crosskey RW. 1990. The natural history of blackflies. Chich-ester: John Wiley.
Crosskey RW, Davies L. 1972. The identities of Simuliumlineatum (Meigen), S. latipes (Meigen) and S. vernumMacquart (Diptera: Simuliidae). Entomologist’s Gazette 23:249–258.
Crosskey RW, Howard TM. 1997. A new taxonomic andgeographical inventory of world blackflies (Diptera:Simuliidae). London: The Natural History Museum.
Crosskey RW, Zwick H. 2007. New faunal records, withtaxonomic annotations, for the blackflies of Turkey (Diptera,Simuliidae). Aquatic Insects 29: 21–48.
Gallardo-Mayenco A, Toja J. 2002. Spatio-temporal distri-bution of simuliids (Diptera) and associated environmental
factors in two Mediterranean basins of southern Spain.Limnetica 21: 47–57.
Grinchuk TM, Chubareva LA. 1974. On intergroupkaryological differences in the genus Wilhelmia. Tsitologiya16: 1432–1435. [In Russian with English summary.].
Hambäck PA, Weingartner E, Ericson L, Fors L,Cassel-Lundhagen A, Stenberg JA, Bergsten J.2013. Bayesian species delimitation reveals generalistand specialist parasitic wasps on Galerucella beetles(Chrysomelidae): sorting by herbivore or plant host. BMCEvolutionary Biology 13: 92. Available at: http://www.biomedcentral.com/1471-2148/13/92.
Hirai H, Procunier WS, Ochoa JO, Uemoto K. 1994. Acytogenetic analysis of the Simulium ochraceum speciescomplex (Diptera: Simuliidae) in Central America. Genome37: 36–53.
Huang L, Zhang C-L, Jiang Y-H, Chen H-B. 2012. Polytenechromosomes of Simulium (Wilhelmia) xingyiense (Diptera:Simuliidae) from China. Acta Entomologica Sinica 55: 988–993.
Kamali M, Xia A, Tu S, Sharakhov IV. 2012. A newchromosomal phylogeny supports the repeated origin ofvectorial capacity in malaria mosquitoes of the Anophelesgambiae complex. PLoS Pathogens 8: e1002960.
Knoz J, Grinchuk TM, Chubareva LA. 1976. Chromo-somes of Wilhelmia. Scripta Facultatis ScientariumNaturalium Universita Jan Evangelista Purkyne Brunensis.Biologia 5: 91–98.
Kovatschev S. 1979. Bemerkungen über dieZusammensetzung, Verbreitung und Ökologie der Simuliiden(Diptera, Simuliidae) in der Bulgarischen Donauzuflüssen.XXI. Arbeitstagung IAD, 392–396.
Lunneborg CE. 2000. Data analysis by resampling: conceptsand applications. Pacific Grove, CA: Duxbury Press.
Mayr E. 1963. Animal species and evolution. Cambridge, MA:Harvard University Press.
McCune B, Grace JB. 2002. Analysis of ecological commu-nities. Gleneden Beach, OR: MJM Software Design.
Pandit SN, Kolasa J, Cottenie K. 2009. Contrasts betweenhabitat generalists and specialists: an empirical extensionto the basic metacommunity framework. Ecology 90: 2253–2262.
Petrova NA, Chubareva LA, Adler PH, Kachvoryan EA.2003. Cytogenetic features of the bloodsucking blackflyWilhelmia paraequina Puri (Diptera: Simuliidae) fromArmenia. Genetika 39: 41–50. [In Russian with Englishsummary; full English translation in Russian Journal ofGenetics 39: 32–40 (2003).].
Procunier WS. 1975. B-chromosomes of Cnephia dacotensisand C. ornithophilia (Diptera: Simuliidae). CanadianJournal of Zoology 53: 1638–1647.
Procunier WS. 1982. A cytological description of 10 taxa inMetacnephia (Diptera: Simuliidae). Canadian Journal ofZoology 60: 2852–2865.
Rivosecchi L. 1978. Simuliidae: Diptera Nematocera. Faunad’Italia 13: 1–533.
Rohde C, Garcia ACL, Valiati VH, VLcS V. 2006. Chro-mosomal evolution of sibling species of the Drosophila
182 P. H. ADLER ET AL.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183

willistoni group. I. Chromosomal arm IIR (Muller’s elementB). Genetica 126: 77–88.
Rothfels KH. 1979. Cytotaxonomy of black flies (Simuliidae).Annual Review of Entomology 24: 507–539.
Rothfels KH, Dunbar RW. 1953. The salivary gland chro-mosomes of the black fly Simulium vittatum Zett. CanadianJournal of Zoology 31: 226–241.
Rothfels KH, Featherston D. 1981. The population struc-ture of Simulium vittatum (Zett.): the IIIL-1 and IS-7sibling species. Canadian Journal of Zoology 59: 1857–1883.
Rothfels KH, Feraday R, Kaneps A. 1978. A cytologicaldescription of sibling species of Simulium venustum andS. verecundum with standard maps for the subgenusSimulium Davies [sic] (Diptera). Canadian Journal ofZoology 56: 1110–1128.
Rothfels K[H], Freeman DM. 1983. A new species ofProsimulium (Diptera: Simuliidae): an interchange as aprimary isolating mechanism? Canadian Journal of Zoology61: 2612–2617.
Rothfels K[H], Nambiar R. 1981. A cytological study ofnatural hybrids between Prosimulium multidentatumand P. magnum: with notes on sex determination in theSimuliidae (Diptera). Chromosoma 82: 673–691.
Rubtsov IA. 1956. Moshki (sem. Simuliidae) [Blackflies(fam. Simuliidae)]. Fauna of the USSR. New Series No. 64,Insects, Diptera 6 (6). Moscow & Leningrad [= StPetersburg]: Akademii Nauk SSSR. [In Russian; Englishtranslation: 1990. Blackflies (Simuliidae). 2nd edition.Fauna of the USSR. Diptera, 6 (6). Leiden: E. J. Brill].
Sariözkan S, Inci A, Yildirim A, Düzlü O, Gray EW,
Adler PH. 2014. Economic losses during an outbreak ofSimulium (Wilhelmia) species (Diptera: Simuliidae) in theCappadocia Region of Turkey. Turkiye Parazitolojii Dergisi38: 116–119.
Soininen J, McDonald R, Hillebrand H. 2007. Thedistance decay of similarity in ecological communities.Ecography 30: 3–12.
Sokal RR, Rohlf FJ. 1995. Biometry: the principles andpractices of statistics in biological research, 3rd edn. NewYork, NY: W H. Freeman.
Stocker AJ, Foley B, Hoffmann A. 2004. Inversion frequen-cies in Drosophila serrata along an eastern Australiantransect. Genome 47: 1144–1153.
Weber EA, Grunewald J. 1989. Cytotaxonomic differentia-tion of Wilhelmia equina (Linné, 1747) and Wilhelmialineata (Meigen, 1804) (Diptera: Simuliidae). Genome 32:589–595.
Werner D, Adler PH. 2005. A faunistic review of theblack flies (Simuliidae, Diptera) of the federal state ofSachsen-Anhalt, Germany. Abhandlungen und Berichte fürNaturkunde 27: 205–245.
Williams PH, An J, Brown MJF, Carolan JC, Goulson D,Huang J, Ito M. 2012. Cryptic bumblebee species: conse-quences for conservation and the trade in greenhouse pol-linators. PLoS ONE 7: e32992.
Yilmaz A, Inci A, Tunçbilek AS, Yesilöz H, Koçak Ö, SirinÜ, Iça A, Yildirim A, Demircioglu A, Düzlü Ö. 2007. OrtaKizilirmak Havzasinda Karasinek (Simulium (Wilhelmia)lineatum) (Diptera: Simuliidae) Istilasi. Erciyes ÜniversitesiVeteriner Fakültesi Dergisi [Journal of the Faculty of Veteri-nary Medicine, Erciyes University] 4: 91–95.
MACROGENOME OF BLACK FLIES IN SUBGENUS WILHELMIA 183
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2015, 114, 163–183